Supported by the National Natural Science Foundation(31870407) Supported by the National Natural Science Foundation(31370542) Special Fund Project for Basic Scientific Research Business Expenses of Central Universities(2020NZD03) Sichuan Science and Technology Plan to focus on Research and Development Project(2018SZ033)
Abstract Aims Our objectives were to investigate the soil phosphorus (P) stocks and distribution patterns in forests of Qinghai Province, and to determine the relationships between soil phosphorus stocks and environmental factors. Methods Phosphorus stocks in forest soils of Qinghai Province were estimated from measurement data of 240 standard sampling plots in combination with the forest type information in the Qinghai Forest Resources Inventory data. The distribution patterns were examined by spatial analysis. Important findings Forest soil P stocks in Qinghai Province is estimated at 1.74 Tg. The average soil P density to 1 m depth is about 4.65 Mg·hm -2, displaying of pattern of zonal distribution. Soil P density significantly decreases first and then increases with altitude, and is largest in Cinnamon forest soils and less in brown forest soils and dark cinnamon forest soils. Soil P content decreases significantly with altitude. The P content was highest in brown forest soils and lowest in dark cinnamon soils. Total P in the soil profile exhibited characteristics of surface accumulation. The structure equation model (SEM) shows that soil type, altitude, temperature, and soil moisture content have direct impacts on soil P content, with temperature and altitude being among the key factors. Soil P content, altitude, soil moisture content, soil depth, and soil bulk density all have significant effects on soil P density, with soil bulk density being the most prominent factor. Keywords:phosphorus stocks;forest soil;spatial distribution pattern;environmental factor;Qinghai
PDF (10308KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 唐立涛, 刘丹, 罗雪萍, 胡雷, 王长庭. 青海省森林土壤磷储量及其分布格局. 植物生态学报, 2019, 43(12): 1091-1103. DOI: 10.17521/cjpe.2019.0194 TANG Li-Tao, LIU Dan, LUO Xue-Ping, HU Lei, WANG Chang-Ting. Forest soil phosphorus stocks and distribution patterns in Qinghai, China. Chinese Journal of Plant Ecology, 2019, 43(12): 1091-1103. DOI: 10.17521/cjpe.2019.0194
Fig. 3Phosphorus density of different soil layers (mean ± SE). Different uppercase letters indicate significant differences in soil phosphorus density between soil layers (p < 0.05).
新窗口打开|下载原图ZIP|生成PPT 图4青海省森林土壤磷含量与海拔的关系。A, B, 0-10 cm; C, D, 10-20 cm; E, F, 20-30 cm; G, H, 30-50 cm; I, J, 50-100 cm。实线, p < 0.05; 虚线, p > 0.05。
Fig. 4Relationships between forest soil phosphorus content and altitude in Qinghai Province. A, B, 0-10 cm; C, D, 10-20 cm; E, F, 20-30 cm; G, H, 30-50 cm; I, J, 50-100 cm. Solid line, p < 0.05; dotted line, p > 0.05.
Fig. 6Soil phosphorus content in different soil types of forests in Qinghai Province (mean ± SE). Different uppercase letters indicate significant differences in soil phosphorus content between different soil types in the same soil layer (p < 0.05); different lowercase letters indicate significant differences in soil phosphorus content between different soil layers of the same soil type (p < 0.05).
Fig. 7Soil phosphorus density in the 0-100 cm soil layer of different soil types (mean ± SE). Different uppercase letters indicate significant differences in soil phosphorus density between different soil types (p < 0.05).
新窗口打开|下载原图ZIP|生成PPT 图8土壤磷与环境因子之间的关系。红线代表正相关, 蓝线代表负相关, 虚线代表相关性不显著。*, p < 0.05; **, p < 0.01; ***, p < 0.001。AGFI, 调整后适配度指数; CHI, 卡方值; DF, 自由度; GFI, 适配度指数; RMSEA, 近似误差均方根。
Fig. 8Relationships between soil phosphorus and environmental factors. The red line represents a positive correlation, the blue line a negative correlation, and the dotted line an insignificant correlation. *, p < 0.05; **, p < 0.01; ***, p < 0.001. AGFI, adjusted goodness-of-fit index; CHI, Chi-Square; DF, degrees of freedom; GFI, goodness-of-fit index; RMSEA, root mean square error of approximation.
AhernCR, MacNishSE ( 1983). Comparative study of phosphorus and potassium levels of basaltic soils associated with scrub and forest communities on the Darling Downs Soil Research, 21, 527-538. DOI:10.1071/SR9830527URL [本文引用: 1]
AllenSK, PlattnerGK, NauelsA, XiaY, StockerTF ( 2013). Climate change 2013: The physical science basis. An overview of the working group Ι contribution to the fifth assessment report of the Intergovernmental Panel on Climate Change (IPCC) Computational Geometry, 18, 95-123. [本文引用: 1]
AugustoL, AchatDL, JonardM, VidalD, RingevalB ( 2017). Soil parent material—A major driver of plant nutrient limitations in terrestrial ecosystems Global Change Biology, 23, 3808-3824. DOI:10.1111/gcb.13691URLPMID:28317232 [本文引用: 1] Because the capability of terrestrial ecosystems to fix carbon is constrained by nutrient availability, understanding how nutrients limit plant growth is a key contemporary question. However, what drives nutrient limitations at global scale remains to be clarified. Using global data on plant growth, plant nutritive status, and soil fertility, we investigated to which extent soil parent materials explain nutrient limitations. We found that N limitation was not linked to soil parent materials, but was best explained by climate: ecosystems under harsh (i.e., cold and or dry) climates were more N-limited than ecosystems under more favourable climates. Contrary to N limitation, P limitation was not driven by climate, but by soil parent materials. The influence of soil parent materials was the result of the tight link between actual P pools of soils and physical-chemical properties (acidity, P richness) of soil parent materials. Some other ground-related factors (i.e., soil weathering stage, landform) had a noticeable influence on P limitation, but their role appeared to be relatively smaller than that of geology. The relative importance of N limitation versus P limitation was explained by a combination of climate and soil parent material: at global scale, N limitation became prominent with increasing climatic constraints, but this global trend was modulated at lower scales by the effect of parent materials on P limitation, particularly under climates favourable to biological activity. As compared with soil parent materials, atmospheric deposition had only a weak influence on the global distribution of actual nutrient limitation. Our work advances our understanding of the distribution of nutrient limitation at global scale. In particular, it stresses the need to take soil parent materials into account when investigating plant growth response to environment changes.
BaetenL, VerstraetenG, de FrenneP, VanhellemontM, WuytsK, HermyM, VerheyenK ( 2011). Former land use affects the nitrogen and phosphorus concentrations and biomass of forest herbs Plant Ecology, 212, 901-909. DOI:10.1007/s11258-010-9876-9URL [本文引用: 1]
BaiJH, OuyangH, XiaoR, GaoJQ, GaoHF, CuiBS, HuangLB ( 2010). Spatial variability of soil carbon, nitrogen, and phosphorus content and storage in an alpine wetland in the Qinghai-Tibet Plateau, China Australian Journal of Soil Research, 48, 730-736. [本文引用: 1]
BarrowNJ ( 1983). A mechanistic model for describing the sorption and desorption of phosphate by soil Journal of Soil Science, 34, 733-750. DOI:10.1016/j.watres.2017.09.025URLPMID:28961494 [本文引用: 1] Abiotic and biotic reactions operate side by side in the cycling of phosphorus (P) in the environment, but the relative roles of these two reactions vary both spatially and temporally. In biotic reactions, the uptake and release of P are catalyzed by enzymes and thus change phosphate oxygen isotope ratios, while in abiotic reactions, the absence of hydrolysis-condensation reactions results in no apparent changes in isotope composition, except short-term kinetic isotope effect due solely to preferential ion exchange. Therefore, isotope method could be a promising tool to differentiate relative roles of these two reactions in the environment but the relationship of the dynamic concentration and isotope exchange at the biota-water interface is largely unknown. In this study, we aimed to develop a process-based isotope model underpinning the competition of abiotic (sorption, desorption, and ion exchange) and biotic (uptake, metabolism, and release) reactions during uptake and recycling of ferrihydrite-bound P by E.?coli. Our model comprises equations describing the partitioning relationship among different P pools and their corresponding oxygen isotope compositions and is based exclusively on oxygen isotope exchange at multiple sites including mineral surface, aqueous phase, and bacterial cells. The process-based model adequately reproduced the measured concentration and isotope compositions over time. Furthermore, parametric and sensitivity analyses using the model indicated that the rate of biological uptake of P was the major factor controlling the changes of phosphate isotope composition. In conclusion, our model provides new insights into a mechanistic aspect of isotope exchange and could be potentially useful for future efforts to understand the interplay of biotic and abiotic factors on phosphorus cycling in natural environments.
Carrino-KykerSR, KluberLA, PetersenSM, CoyleKP, HewinsCR, DeForestJL, SmemoKA, BurkeDJ ( 2016). Mycorrhizal fungal communities respond to experimental elevation of soil pH and P availability in temperate hardwood forests FEMS Microbiology Ecology, 92, fiw024. DOI: 10.1093/femsec/fiw024. DOI:10.1093/femsec/fiw024URLPMID:26850158 [本文引用: 1] Many forests are affected by chronic acid deposition, which can lower soil pH and limit the availability of nutrients such as phosphorus (P), but the response of mycorrhizal fungi to changes in soil pH and P availability and how this affects tree acquisition of nutrients is not well understood. Here, we describe an ecosystem-level manipulation in 72 plots, which increased pH and/or P availability across six forests in Ohio, USA. Two years after treatment initiation, mycorrhizal fungi on roots were examined with molecular techniques, including 454-pyrosequencing. Elevating pH significantly increased arbuscular mycorrhizal (AM) fungal colonization and total fungal biomass, and affected community structure of AM and ectomycorrhizal (EcM) fungi, suggesting that raising soil pH altered both mycorrhizal fungal communities and fungal growth. AM fungal taxa were generally negatively correlated with recalcitrant P pools and soil enzyme activity, whereas EcM fungal taxa displayed variable responses, suggesting that these groups respond differently to P availability. Additionally, the production of extracellular phosphatase enzymes in soil decreased under elevated pH, suggesting a shift in functional activity of soil microbes with pH alteration. Thus, our findings suggest that elevating pH increased soil P availability, which may partly underlie the mycorrhizal fungal responses we observed.
CassagneN, RemauryM, GauquelinT, FabreA ( 2000). Forms and profile distribution of soil phosphorus in alpine Inceptisols and Spodosols (Pyrenees, France) Geoderma, 95, 161-172. DOI:10.1016/S0016-7061(99)00093-2URL [本文引用: 1]
Chapin IIIFS, MatsonPA, MooneyHA ( 2002). Principles of Terrestrial Ecosystem Ecology Springer, New York. [本文引用: 1]
ChenML, ChenH, MaoQG, ZhuXM, MoJM ( 2016). Effect of nitrogen deposition on the soil phosphorus cycle in forest ecosystems: A review Acta Ecologica Sinica, 36, 4965-4976. [本文引用: 2]
ChengH, GongYB, WuQ, LiY, LiuY, ZhuDW ( 2018). Content and ecological stoichiometry characteristics of organic carbon, nitrogen and phosphorus of typical soils in sub-alpine/alpine mountain of western Sichuan Journal of Natural Resources, 33, 161-172. [本文引用: 1]
Compilation Group of Technical Specifications for Ecosystem Carbon Sequestration Project ( 2015). Technical Specifications for Observation and Investigation of Ecosystem Carbon Sequestration. Science Press, Beijing. [本文引用: 1]
CrossAF, SchlesingerWH ( 2001). Biological and geochemical controls on phosphorus fractions in semiarid soils Biogeochemistry, 52, 155-172. DOI:10.1023/A:1006437504494URL [本文引用: 1]
DambrineE, DupoueyJL, LaütL, HumbertL, ThinonM, BeaufilsT, RichardH ( 2007). Present forest biodiversity patterns in France related to former Roman agriculture Ecology, 88, 1430-1439. DOI:10.1890/05-1314URLPMID:17601136 [本文引用: 1] Combined archaeological and ecological investigations in a large ancient oak forest in Central France have revealed a dense network of ancient human settlements dating from the Roman period. We demonstrate a strong correlation between present-day forest plant diversity patterns and the location of Roman farm buildings. Plant species richness strongly increases toward the center of the settlements, and the frequency of neutrophilous and nitrogen-demanding species is higher. This pattern is paralleled by an increase in soil pH, available P, and delta(15)N, indicating the long-term impact of former agricultural practices on forest biogeochemical cycles. These extensive observations in a forested region on acid soils complement and confirm previous results from a single Roman settlement on limestone. Ancient Roman agricultural systems are increasingly being identified in contemporary French forests; the broad extent and long-lasting effects of previous cultivation shown in this study require that land-use history be considered as a primary control over biodiversity variations in many forest landscapes, even after millennia of abandonment.
Editorial Committee of Forest in Qinghai ( 1993). Forest in Qinghai. China Forestry Press, Beijing. [本文引用: 4]
FaheyTJ, WoodburyPB, BattlesJJ, GoodaleCL, HamburgSP, OllingerSV, WoodallCW ( 2010). Forest carbon storage: Ecology, management, and policy Frontiers in Ecology and the Environment, 8, 245-252. [本文引用: 1]
FAO ( 2005). Global forest resource assessment: Progress towards sustainable forest management In: FAO ed. FAO Forestry Paper 147. Food and Agriculture Organization of the United Nations, Rome. [本文引用: 2]
FukamiT, WardleDA ( 2005). Long-term ecological dynamics: Reciprocal insights from natural and anthropogenic gradients Proceedings of the Royal Society B: Biological Sciences, 272, 2105-2115. DOI:10.1098/rspb.2005.3277URLPMID:16191623 [本文引用: 1] Many ecological dynamics occur over time-scales that are well beyond the duration of conventional experiments or observations. One useful approach to overcome this problem is extrapolation of temporal dynamics from spatial variation. We review two complementary variants of this approach that have been of late increasingly employed: the use of natural gradients to infer anthropogenic effects and the use of anthropogenic gradients to infer natural dynamics. Recent studies have considered a variety of naturally occurring gradients associated with climate, CO2, disturbance and biodiversity gradients, as well as anthropogenic gradients such as those created by biological invasions, habitat fragmentation and land abandonment. These studies show that natural gradients are useful in predicting long-term consequences of human-induced environmental changes, whereas anthropogenic gradients are helpful in inferring the mechanisms behind natural dynamics because covarying factors are often more clearly understood in anthropogenic gradients than in natural gradients. We classify these studies into several categories, each with different strengths and weaknesses, and outline how the limitations can be overcome by combining the gradient-based approach with other approaches. Overall, studies reviewed here demonstrate that the development of basic ecological concepts and the application of these concepts to environmental problems can be more effective when conducted complementarily than when pursued separately.
GahooniaTS, RazaS, NielsenNE ( 1994). Phosphorus depletion in the rhizosphere as influenced by soil moisture Plant and Soil, 159, 213-218. DOI:10.1007/BF00009283URL [本文引用: 1]
GuoYP, YangX, MohhamotA, LiuHY, MaWH, YuSL, TangZY ( 2017). Storage of carbon, nitrogen and phosphorus in temperate shrubland ecosystems across Northern China Chinese Journal of Plant Ecology, 41, 14-21. [本文引用: 3]
HouEQ, ChenCR, LuoYQ, ZhouGY, KuangYW, ZhangYG, HeenanM, LuXK, WenDZ ( 2018). Effects of climate on soil phosphorus cycle and availability in natural terrestrial ecosystems Global Change Biology, 24, 3344-3356. DOI:10.1111/gcb.14093URLPMID:29450947 [本文引用: 1] Climate is predicted to change over the 21st century. However, little is known about how climate change can affect soil phosphorus (P) cycle and availability in global terrestrial ecosystems, where P is a key limiting nutrient. With a global database of Hedley P fractions and key-associated physiochemical properties of 760 (seminatural) natural soils compiled from 96 published studies, this study evaluated how climate pattern affected soil P cycle and availability in global terrestrial ecosystems. Overall, soil available P, indexed by Hedley labile inorganic P fraction, significantly decreased with increasing mean annual temperature (MAT) and precipitation (MAP). Hypothesis-oriented path model analysis suggests that MAT negatively affected soil available P mainly by decreasing soil organic P and primary mineral P and increasing soil sand content. MAP negatively affected soil available P both directly and indirectly through decreasing soil primary mineral P; however, these negative effects were offset by the positive effects of MAP on soil organic P and fine soil particles, resulting in a relatively minor total MAP effect on soil available P. As aridity degree was mainly determined by MAP, aridity also had a relatively minor total effect on soil available P. These global patterns generally hold true irrespective of soil depth (≤10?cm or &gt;10?cm) or site aridity index (≤1.0 or &gt;1.0), and were also true for the low-sand (≤50%) soils. In contrast, available P of the high-sand (&gt;50%) soils was positively affected by MAT and aridity and negatively affected by MAP. Our results suggest that temperature and precipitation have contrasting effects on soil P availability and can interact with soil particle size to control soil P availability.
HuL, XiangZY, WangGX, RafiqueR, LiuW, WangCT ( 2016). Changes in soil physicochemical and microbial properties along elevation gradients in two forest soils Scandinavian Journal of Forest Research, 31, 242-253. DOI:10.1080/02827581.2015.1125522URL [本文引用: 1]
IppolitoJA, BleckerSW, FreemanCL, McCulleyRL, BlairJM, KellyEF ( 2010). Phosphorus biogeochemistry across a precipitation gradient in grasslands of central North America Journal of Arid Environments, 74, 954-961. DOI:10.1016/j.jaridenv.2010.01.003URL
JobbágyEG, JacksonRB ( 2004). The uplift of soil nutrients by plants: Biogeochemical consequences across scales Ecology, 85, 2380-2389. DOI:10.1890/03-0245URL [本文引用: 2]
KhitrovNB, ZazovskayaEP, RogovnevaLV ( 2018). Morphology, radiocarbon age, and genesis of Vertisols of the Eisk Peninsula (the Kuban-Azov Lowland) Eurasian Soil Science, 51, 731-743. DOI:10.1134/S1064229318070050URL [本文引用: 1]
LiZW, CuiLT, GengSG, ZhangYP ( 2007). Environmental factors affecting soil phosphorus desorption Soil and Water Conservation in China, ( 6), 33-34. URL [本文引用: 1]
LiuQ, ZhengX, DengBL, ZhengLY, HuangLJ, WangSL, ZhangL, GuoXM ( 2017). Effect of altitude on the phosphorus contents of soil and plant litter deposits in a Wugong Mountain meadow Pratacultural Science, 34, 2183-2190. [本文引用: 1]
LiuY, VillalbaG, AyresRU, SchroderH ( 2008). Global phosphorus flows and environmental impacts from a consumption perspective Journal of Industrial Ecology, 12, 229-247. DOI:10.1111/jiec.2008.12.issue-2URL [本文引用: 1]
LuZQ, HuangQX, YangXB ( 2014). Research on hydrological effects of forest litters and soil of Pinus tabuliformis plantations in the different altitudes of Wuling Mountains in Hebei. Journal of Soil and Water Conservation, 28, 112-116. [本文引用: 1]
MaLM, RenaD, ZhangM, ZhaoJF ( 2010). Phosphorus fractions and soil release in alternately waterlogged and drained environments at the water-fluctuation-zone of the Three Gorges Reservoir Journal of Food Agriculture and Environment, 8, 1329-1335. [本文引用: 1]
MarkleinAR, HoultonBZ ( 2011). Nitrogen inputs accelerate phosphorus cycling rates across a wide variety of terrestrial ecosystems New Phytologist, 193, 696-704. DOI:10.1111/j.1469-8137.2011.03967.xURLPMID:22122515 [本文引用: 1] ? Biologically essential elements--especially nitrogen (N) and phosphorus (P)--constrain plant growth and microbial functioning; however, human activities are drastically altering the magnitude and pattern of such nutrient limitations on land. Here we examine interactions between N and P cycles of P mineralizing enzyme activities (phosphatase enzymes) across a wide variety of terrestrial biomes. ? We synthesized results from 34 separate studies and used meta-analysis to evaluate phosphatase activity with N, P, or N×P fertilization. ? Our results show that N fertilization enhances phosphatase activity, from the tropics to the extra-tropics, both on plant roots and in bulk soils. By contrast, P fertilization strongly suppresses rates of phosphatase activity. ? These results imply that phosphatase enzymes are strongly responsive to changes in local nutrient cycle conditions. We also show that plant phosphatases respond more strongly to fertilization than soil phosphatases. The tight coupling between N and P provides a mechanism for recent observations of N and P co-limitation on land. Moreover, our results suggest that terrestrial plants and microbes can allocate excess N to phosphatase enzymes, thus delaying the onset of single P limitation to plant productivity as can occur via human modifications to the global N cycle.
MoreiraA, MoraesLAC, ZaninettiRA, CanizellaBT ( 2013). Phosphorus dynamics in the conversion of a secondary forest into a rubber tree plantation in the Amazon rainforest Soil Science, 178, 618-625. DOI:10.1097/SS.0000000000000025URL [本文引用: 1]
?zgülM, GünesA, EsringüA, TuranM ( 2012). The effects of freeze-and-thaw cycles on phosphorus availability in highland soils in Turkey Journal of Plant Nutrition and Soil Science, 175, 827-839. DOI:10.1002/jpln.201100407URL Ongoing global warming may result in colder soil and thawing cycles and will increase the frequency of soil freezing-and-thawing-treated cycles (FTCs) during winter in the cool-temperate and high-latitude regions. The purpose of this study was to determine the effects of repeated freezethaw cycles on the solubility and adsorption of P in lab and field experiments on Pellustert, Argiustoll, Haplustept, Fluvaquent, and Calciorthid soils, the major soil groups in E Turkey. The results demonstrated that, depending on the soil type, the freezethaw cycle could increase the adsorption and desorption of P within a certain temperature range. Repeated freezing and thawing decreased equilibrium P concentration (EPC) and increased P adsorption. EPC and P adsorption were strongly correlated with the number of FTCs. The highest P adsorption and the lowest P desorption was found in Pellustert followed by Argiustoll, Calciorthid, Haplustept, Fluvaquent when refrozen at 10 degrees C for 15 d, then thawed at +2.5 degrees C for 18 h, and 9 times FTC. However, in the field study, the adsorption value was lower than the value obtained from the laboratory condition. It appears that increasing the frequency of freezethaw processes depending on increase in temperature that leads to decreased plant-available soil P pools, thus requires more P fertilizer in soil solution to supply adequate P during the plant-growth period.
PrescottCE, MaynardDG, LaihoR ( 2000b). Humus in northern forests: Friend or foe? Forest Ecology and Management, 133, 23-36. [本文引用: 1]
PrescottCE, ZabekLM, StaleyCL, KabzemsR ( 2000a). Decomposition of broadleaf and needle litter in forests of British Columbia: Influences of litter type, forest type, and litter mixtures Canadian Journal of Forest Research, 30, 1742-1750. [本文引用: 1]
Qinghai Provincial Agricultural Resources Regionalization Office ( 1997). The Soil in Qinghai Province. China Agriculture Press, Beijing. [本文引用: 5]
ReedSC, YangXJ, ThorntonPE ( 2015). Incorporating phosphorus cycling into global modeling efforts: A worthwhile, tractable endeavor New Phytologist, 208, 324-329. DOI:10.1111/nph.13521URLPMID:26115197 [本文引用: 1] 324 I. 324 II. 325 III. 326 IV. 327 328 References 328 SUMMARY: Myriad field, laboratory, and modeling studies show that nutrient availability plays a fundamental role in regulating CO2 exchange between the Earth's biosphere and atmosphere, and in determining how carbon pools and fluxes respond to climatic change. Accordingly, global models that incorporate coupled climate-carbon cycle feedbacks made a significant advance with the introduction of a prognostic nitrogen cycle. Here we propose that incorporating phosphorus cycling represents an important next step in coupled climate-carbon cycling model development, particularly for lowland tropical forests where phosphorus availability is often presumed to limit primary production. We highlight challenges to including phosphorus in modeling efforts and provide suggestions for how to move forward.
ReichPB, OleksynJ ( 2004). Global patterns of plant leaf N and P in relation to temperature and latitude Proceedings of the National Academy of Sciences of the United States of America, 101, 11001-11006. DOI:10.1073/pnas.0403588101URLPMID:15213326 [本文引用: 1] A global data set including 5,087 observations of leaf nitrogen (N) and phosphorus (P) for 1,280 plant species at 452 sites and of associated mean climate indices demonstrates broad biogeographic patterns. In general, leaf N and P decline and the N/P ratio increases toward the equator as average temperature and growing season length increase. These patterns are similar for five dominant plant groups, coniferous trees and four angiosperm groups (grasses, herbs, shrubs, and trees). These results support the hypotheses that (i) leaf N and P increase from the tropics to the cooler and drier midlatitudes because of temperature-related plant physiological stoichiometry and biogeographical gradients in soil substrate age and then plateau or decrease at high latitudes because of cold temperature effects on biogeochemistry and (ii) the N/P ratio increases with mean temperature and toward the equator, because P is a major limiting nutrient in older tropical soils and N is the major limiting nutrient in younger temperate and high-latitude soils.
RuttenbergKC ( 2003). The global phosphorus cycle Treatise on Geochemistry, 585-643. [本文引用: 1]
SiebersN, SumannM, KaiserK, AmelungW ( 2017). Climatic effects on phosphorus fractions of native and cultivated north American grassland soils Soil Science Society of America Journal, 81, 299-309. [本文引用: 1]
SmilV ( 2000). Phosphorus in the environment: Natural flows and human interferences Annual Review of Energy and the Environment, 25, 53-88. [本文引用: 2]
TangLT, ZiHB, HuL, AdeLJ, WangCT ( 2019). Forest biomass and its influencing factors in Qinghai Province Acta Ecologica Sinica, 39, 3677-3686. [本文引用: 1]
VincentAG, SchleucherJ, Gr?bnerG, VestergrenJ, PerssonP, JanssonM, GieslerR ( 2012). Changes in organic phosphorus composition in boreal forest humus soils: The role of iron and aluminium Biogeochemistry, 108, 485-499.
VincentAG, SundqvistMK, WardleDA, GieslerR ( 2014). Bioavailable soil phosphorus decreases with increasing elevation in a subarctic tundra landscape PLOS ONE, 9, e92942. DOI: 10.1371/journal.pone.0092942. [本文引用: 2]
Viscarra RosselRA, BuiEN ( 2016). A new detailed map of total phosphorus stocks in Australian soil Science of the Total Environment, 542, 1040-1049. [本文引用: 2]
VogtKA, GrierCC, VogtDJ ( 1986). Production, turnover, and nutrient dynamics of above-and belowground detritus of world forests Advances in Ecological Research, 15, 303-377.
WanD, LiangB, NieXG, YuW, ZhangB ( 2018). Research on vertical zonation of soil physical properties in Sygera Mountain, Tibet Acta Ecologica Sinica, 38, 1065-1074. [本文引用: 2]
WangT, YangYH, MaWH ( 2008). Storage, patterns and environmental controls of soil phosphorus in China Acta Scientiarum Naturalium Universitatis Pekinensis, 44, 945-952. [本文引用: 1]
WeiLL, ChenCR, XuZH ( 2010). Citric acid enhances the mobilization of organic phosphorus in subtropical and tropical forest soils Biology and Fertility of Soils, 46, 765-769. [本文引用: 1]
WrightRF ( 1998). Effect of increased carbon dioxide and temperature on runoff chemistry at a forested catchment in southern Norway (CLIMEX project) Ecosystems, 1, 216-225. [本文引用: 1]
YuanZY, JiaoF, ShiXR, SardansJ, MaestreFT, Delgado-BaquerizoM, ReichPB, Pe?uelasJ ( 2017). Experimental and observational studies find contrasting responses of soil nutrients to climate change eLife, 6, e23255. DOI: 10.7554/eLife.23255. [本文引用: 1]
YueK, YangWQ, PengY, PengCH, TanB, XuZF, ZhangL, NiXY, ZhouW, WuFZ ( 2018). Individual and combined effects of multiple global change drivers on terrestrial phosphorus pools: A meta-analysis Science of The Total Environment, 630, 181-188. [本文引用: 1]
ZhangC, TianHQ, LiuJY, WangSQ, LiuML, PanSF, ShiXZ ( 2005). Pools and distributions of soil phosphorus in China Global Biogeochemical Cycles, 19, GB1020. DOI: 10.1029/2004GB002296. [本文引用: 2]
ZhangTD, WangCK, ZhangQZ ( 2017). Vertical variation in stoichiometric relationships of soil carbon, nitrogen and phosphorus in five forest types in the Maoershan region, Northeast China Chinese Journal of Applied Ecology, 28, 3135-3143. [本文引用: 1]
ZhouJ, BingHJ, WuYH, SunSQ, LuoJ ( 2016). Variations in soil P biogeochemistry and its impact factors along an altitudinal gradient in the Yanzigou, eastern slope of the Gongga Mountain Mountain Research, 34, 385-392. [本文引用: 1]
ZiHB, XiangZY, WangGX, AdeLJ, WangCT ( 2017). Profile of soil microbial community under different stand types in Qinghai Province Scientia Silvae Sinicae, 53(3), 21-32.
Climate change 2013: The physical science basis. An overview of the working group Ι contribution to the fifth assessment report of the Intergovernmental Panel on Climate Change (IPCC) 1 2013
Phosphorus fractions and soil release in alternately waterlogged and drained environments at the water-fluctuation-zone of the Three Gorges Reservoir 1 2010
... 磷是生态系统中重要的生命元素, 是地球上所有生命化学组成的基础之一(Vincent et al., 2012).植物生长所必需的磷主要来源于土壤, 土壤是陆地生态系统中最大的磷库.土壤磷储量主要依靠矿石的风化和大气干湿沉降来维持(Ruttenberg, 2003).气候、地形地貌、土壤发育阶段、微生物动态等均在生态系统土壤磷库的输入与输出过程中扮演着重要角色(Ippolito et al., 2010).而土壤的生物化学过程以及植物自身的遗传特性则会影响土壤和植被之间的磷转移(Wei et al., 2010).磷的通量几乎是单向的, 从陆地生态系统逐渐向江河海洋移动(Liu et al., 2008).19世纪中叶以来, 全球变化显著地改变了陆地磷动态.陆地磷库对气候变暖响应敏感, 气候变暖使得土壤沙含量增加, 进而降低土壤原生矿物磷和有机磷库, 土壤总磷减少(Hou et al., 2018).高降水波动引发森林土壤磷减少(Yuan et al., 2017), 氮沉降加速了森林土壤磷循环致使土壤总磷相对减少(Marklein & Houlton, 2011). ...
Nitrogen inputs accelerate phosphorus cycling rates across a wide variety of terrestrial ecosystems 1 2011
... 高海拔区域50-100 cm土层磷含量随海拔升高有增加的趋势.因为一定范围内随着海拔升高风力变大, 有效积温降低, 植物为了应对高海拔严酷的生存环境, 调整生存对策即增加地下根系生物量(死根相对较多)来维持土壤局部小环境温度(唐立涛等, 2019), 加之根系分解是向土壤归还磷的重要途径(Vogt et al., 1986).此外, 高海拔区域冻融作用变大, 使得土壤的吸附位点增多, 土壤对磷的吸附能力增强, 进而土壤对磷的缓冲能力得到加强(?zgül et al., 2012).同时低温环境下土壤微生物为了维持自身的生存需要, 可能也会通过固定其生物量中的磷来增加土壤微生物中的磷储量(Moreira et al., 2013). ...
Phosphorus dynamics in the conversion of a secondary forest into a rubber tree plantation in the Amazon rainforest 1 2013
... 高海拔区域50-100 cm土层磷含量随海拔升高有增加的趋势.因为一定范围内随着海拔升高风力变大, 有效积温降低, 植物为了应对高海拔严酷的生存环境, 调整生存对策即增加地下根系生物量(死根相对较多)来维持土壤局部小环境温度(唐立涛等, 2019), 加之根系分解是向土壤归还磷的重要途径(Vogt et al., 1986).此外, 高海拔区域冻融作用变大, 使得土壤的吸附位点增多, 土壤对磷的吸附能力增强, 进而土壤对磷的缓冲能力得到加强(?zgül et al., 2012).同时低温环境下土壤微生物为了维持自身的生存需要, 可能也会通过固定其生物量中的磷来增加土壤微生物中的磷储量(Moreira et al., 2013). ...
The effects of freeze-and-thaw cycles on phosphorus availability in highland soils in Turkey 2012
 ,, 刘丹, 罗雪萍, 胡雷, 王长庭
,, 刘丹, 罗雪萍, 胡雷, 王长庭 ,*西南民族大学青藏高原研究院, 成都 610041
,*西南民族大学青藏高原研究院, 成都 610041
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT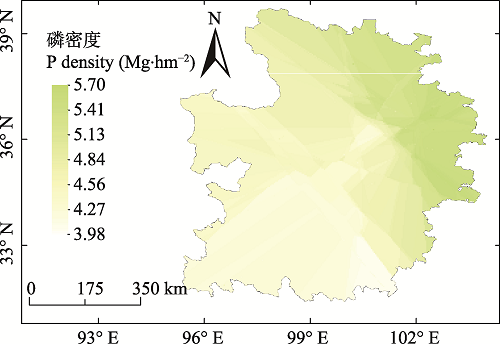 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT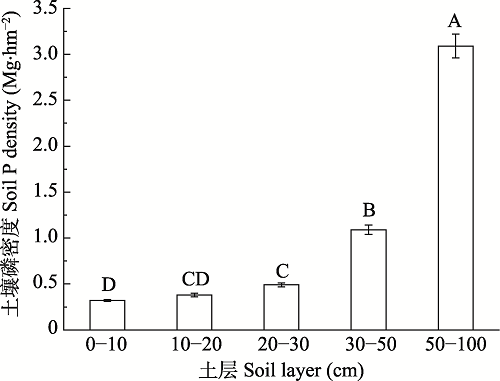 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT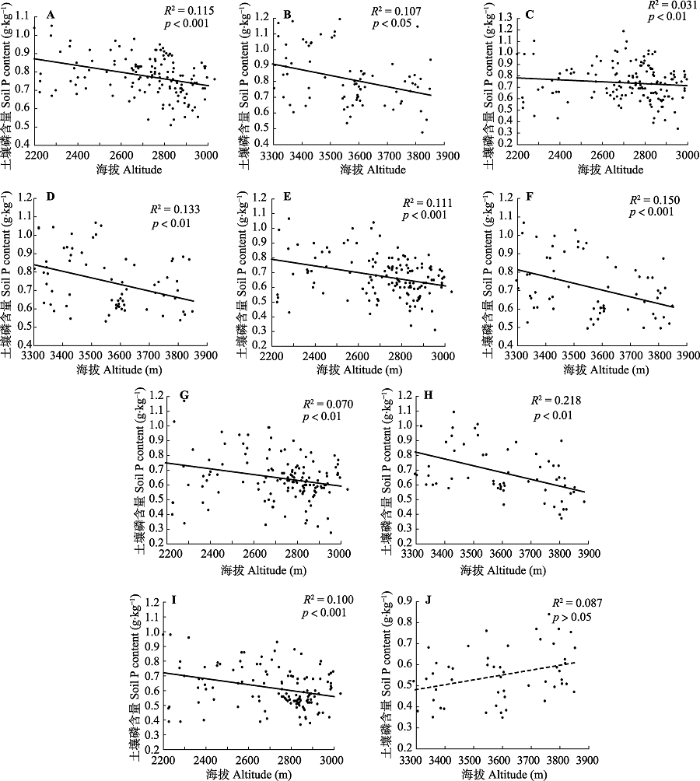 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT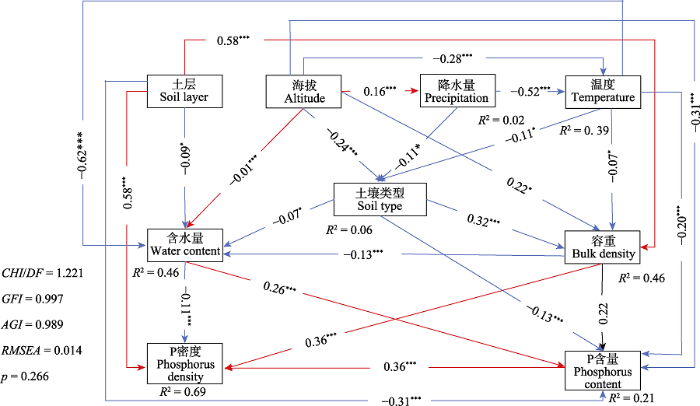 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT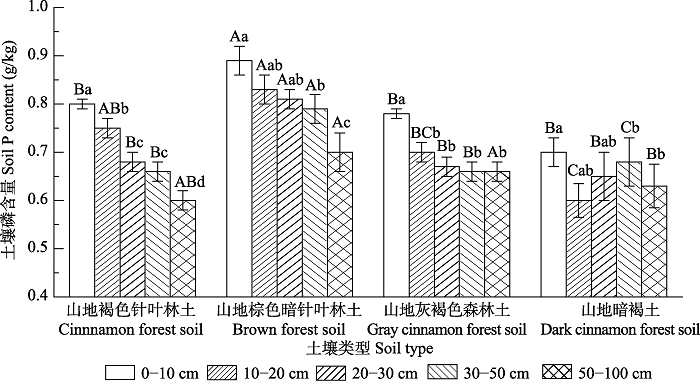 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}