Effects of different calcium concentrations on growth and physiology of Paspalum wettsteinii seedlings
Xin ZHAO,1, Wen-Juan WANG1, Pu-Chang WANG2,3, Li-Juan HUANG1, Li-Li ZHAO,1,2,*1Department of Grassland Science, College of Animal Science, Guizhou University, Guiyang 550025, China 2Key Laboratory of Mountain Plant Resources Protection and Germplasm Innovation Ministry of Education, Guiyang 550025, China 3Guizhou Institute of Prataculture, Guiyang 550006, China
Abstract Aims The study about the effects of different calcium concentrations on the growth and physiology of Paspalum wettsteinii seedlings is very important to reveal the adaptive mechanism of Paspalum wettsteinii to the environment with different calcium concentrations. Methods Potted sand culture was used to study the effects of different calcium concentrations (0, 5, 25, 50, 100 and 200 mmol��L-1 CaCl2) and different treatment times (7, 14, 21 and 28 d) on the growth, osmotic regulator content, antioxidant enzyme activity, chlorophyll content and photosynthetic parameters of Paspalum wettsteinii seedlings. Important findings Results showed that, with the increase of the CaCl2 concentration and the extension of treatment time, the morphological indexes, biomass, osmotic regulators content, antioxidant enzyme activity, chlorophyll content and photosynthetic parameters of Paspalum wettsteinii seedlings displayed a similar trend of first increasing and then decreasing. Under the low calcium concentrations (5-50 mmol��L-1), plant height, leaf length, leaf width, root length and biomass all increased. The contents of proline, soluble protein, soluble sugar and the activities of peroxidase, catalase, superoxide dismutase, chlorophyll content, net photosynthetic rate, transpiration rate and stomatal conductance increased as well, but, malondialdehyde content and intercellular CO2 concentration decreased. Under the high calcium concentrations (200 mmol��L-1), the contents of proline, soluble protein, soluble sugar and the activities of peroxidase, catalase and superoxide dismutase decreased. Malondialdehyde content and intercellular CO2 concentration increased as well, but chlorophyll content, net photosynthetic rate, transpiration rate and stomatal conductance decreased. Combined with the membership function analysis, the treatment of low calcium concentrations (5-50 mmol��L-1) had no inhibitory effect on the seedlings of Paspalum wettsteinii, indicating that Paspalum wettsteinii had certain tolerance to low calcium salt stress. Under the treatment of high calcium concentration (200 mmol��L-1), Paspalum wettsteinii seedlings could rapidly regulate the physiological and metabolic functions of plants by increasing the content of organic osmotic regulating substances, enhancing enzyme activity, chlorophyll content and photosynthesis, so as to adapt to high calcium concentration environment. Keywords��Paspalum wettsteinii;calcium concentrations;growth;physiological
PDF (1436KB)Ԫ������ά�����������������EndNote|Ris|Bibtex�ղر��� ���ñ��� ����, ���ľ�, ������, �����, ������. ��ͬ��Ũ�ȶԿ�Ҷȸ������������Ϳ���������Ӱ��. ֲ����̬ѧ��, 2019, 43(10): 909-920. DOI: 10.17521/cjpe.2019.0235 ZHAO Xin, WANG Wen-Juan, WANG Pu-Chang, HUANG Li-Juan, ZHAO Li-Li. Effects of different calcium concentrations on growth and physiology of Paspalum wettsteinii seedlings. Chinese Journal of Plant Ecology, 2019, 43(10): 909-920. DOI: 10.17521/cjpe.2019.0235
λ���ҹ����ϵĹ��ݸ�ԭ�������������ֲ���еĿ�˹�ص���������, Ҳ�ǿ�˹�ط�������͵�һ������(Zeng et al., 2017; Li et al., 2019)����ֲ��㷺��̼�����Ҳ��¶�����1.3 �� 105 km2, ռ����ȫʡ�������73% (��ï᪺��Լ�, 2013)�������������ȴ�ʪ������, ̼�����Ҳ��ܵ���ʴ��Ϊ����, �����������λ�(���ҳϵ�, 2014), ����̼�����ҷ�Χռ�������λ���Χ��1%-3%, ��ͬγ�ȹ����ε�����2-3��(��С��, 2006)��������ʴ����, ���������е����ס��ص����ִ�����ʧ(�����ܺ�������, 2007), ������в��˹�ص���ֲ�������, ���ѡ����ֲ�߸��ε�ֲ��Կ�˹�ص����ݵ�����ҵ�ķ�չ����̬�����ĸ��ƾ�����Ҫ���塣����ֲ�����������������Ԫ��(Liang et al., 2009), ����ͨ��ά��ϸ��Ĥͨ�ԡ��������䡢�ź�ת���ȷ�ʽֱ�Ӳ���ֲ��ֿ���в��(Ferit & F��sun, 2016)���ؽ���(Gabara et al., 1995)�ɺ�(Li et al., 2017)�ȷ������澳, ����ֲͬ��ԸƵ������Բ�ͬ, ȱ�ơ��ƻ��Ũ�ȹ��߶���Ӱ��ֲ������, ���˸�Ũ�Ȳ��ܴٽ�ֲ������, ������������������
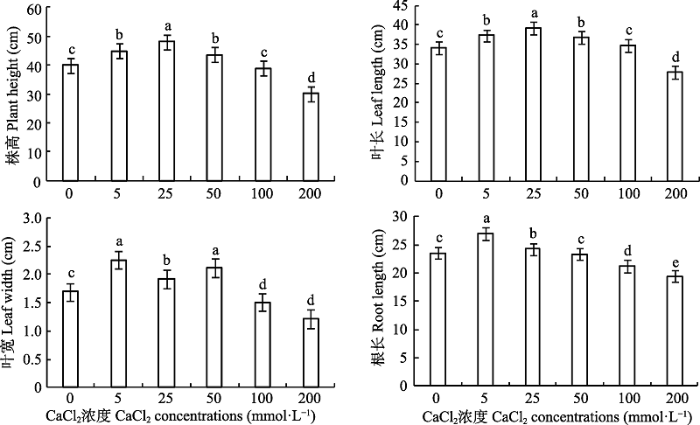
Fig. 1Effects of different CaCl2 concentrations on the growth indices of Paspalum wettsteinii seedlings (mean �� SE). Different lowercase letters indicate significant difference among different CaCl2 concentrations (p < 0.05).
Fig. 2Effects of different CaCl2 concentrations on the biomass of Paspalum wettsteinii seedlings (mean �� SE). Different lowercase letters indicate significant difference among different CaCl2 concentrations (p < 0.05).
Table 1 ��1 ��1��ͬCaCl2Ũ�ȶԿ�Ҷȸ�����縬���ᡢ�����Ե��Ϳ������Ǻ�����Ӱ��(ƽ��ֵ�������) Table 1Effects of different CaCl2 concentrations on proline content, soluble protein and soluble sugar content of Paspalum wettsteinii seedlings (mean �� SE)
����������Osmoregulation substance
CaCl2Ũ�� CaCl2 concentrations (mmol��L-1)
ʱ�䴦�� Time treatment (d)
7
14
21
28
������ Proline (��g��g-1)
0
16.13 �� 0.84Cc
27.24 �� 0.97Ac
23.87 �� 0.70Bc
10.08 �� 1.43Dc
5
23.44 �� 1.17Ca
30.91 �� 0.87Ab
27.64 �� 0.63Bb
13.25 �� 0.61Db
25
25.04 �� 1.41Ca
32.82 �� 0.68Aa
30.91 �� 0.52Ba
16.97 �� 0.65Da
50
20.51 �� 0.95Cb
24.46 �� 0.67Ad
22.68 �� 0.62Bc
12.94 �� 0.74Db
100
14.23 �� 0.57Cd
22.98 �� 0.62Ae
19.08 �� 0.38Bd
8.85 �� 0.62Dd
200
9.65 �� 0.32Be
14.66 �� 1.17Af
15.15 �� 0.32Ae
7.25 �� 0.68Bd
�����Ե��� Soluble protein (mg��g-1)
0
10.22 �� 0.70Bd
14.09 �� 0.41Ac
10.73 �� 0.43Bc
6.25 �� 0.36Cb
5
11.23 �� 0.50Bc
15.47 �� 0.31Ab
13.16 �� 0.73Ca
7.48 �� 0.23Db
25
12.50 �� 0.27Bb
17.38 �� 0.54Aa
14.11 �� 0.77Ba
8.99 �� 0.44Ca
50
13.67 �� 0.12Ba
18.03 �� 0.43Aa
11.84 �� 0.34Cb
6.13 �� 0.45Db
100
11.56 �� 0.49Bc
16.25 �� 0.64Ab
10.17 �� 0.57Cc
4.93 �� 0.25Dc
200
8.05 �� 0.54Ce
13.54 �� 0.35Ac
8.96 �� 0.07Bd
3.75 �� 0.11Dd
�������� Soluble sugar (mg��g-1)
0
17.04 �� 1.22Cd
27.07 �� 1.42Ad
22.54 �� 0.61Bc
12.26 �� 1.64Db
5
18.60 �� 0.64Cc
30.27 �� 0.70Ab
23.68 �� 0.85Bb
12.36 �� 0.80Db
25
21.25 �� 1.21Cb
32.00 �� 1.47Aa
25.40 �� 1.50Ba
14.36 �� 0.95Da
50
22.23 �� 0.51Ba
29.05 �� 1.36Ac
20.81 �� 1.10Cd
13.62 �� 0.49Da
100
15.82 �� 0.97Ce
27.23 �� 1.29Ad
19.09 �� 1.06Be
10.45 �� 0.43Dc
200
13.92 �� 0.87Cf
26.53 �� 0.80Ad
19.62 �� 0.44Be
10.50 �� 1.08Dc
Different uppercase letters indicate significant difference between treatments at different times of the same concentration (p < 0.05); different lowercase letters indicate significant difference between different concentrations at the same time (p < 0.05). ��ͬ��д��ĸ��ʾͬһŨ�Ȳ�ͬʱ�䴦�����������(p < 0.05); ��ͬСд��ĸ��ʾͬһʱ�䴦����ͬŨ�ȼ��������(p < 0.05)��
Table 2 ��2 ��2��ͬCaCl2Ũ�ȶԿ�Ҷȸ�������������ø(POD)����������ø(CAT)�ͳ��������绯ø(SOD)���Ժͱ���ȩ(MDA)������Ӱ��(ƽ��ֵ�������) Table 2Effects of different CaCl2 concentrations on peroxidase, catalase, superoxide dismutase activity and malondialdehyde content of Paspalum wettsteinii seedlings (mean �� SE)
������ø�ͱ���ȩ Antioxidative enzyme and malondialdehyde
CaCl2 Ũ�� CaCl2 concentrations (mmol��L-1)
ʱ�䴦�� Time treatment (d)
7
14
21
28
POD (U��g-1)
0
1 034.32 �� 65.54Bb
1 135.20 �� 29.29Ac
827.85 �� 37.30Cc
667.22 �� 38.99Db
5
1 043.20 �� 37.77Bb
1 240.38 �� 60.69Ab
967.84 �� 21.11Bb
825.57 �� 59.04Ca
25
1 173.26 �� 21.35Bb
1 381.73 �� 35.64Aa
1 111.83 �� 29.74Ba
883.05 �� 54.03Ca
50
1 185.94 �� 52.78Aa
1 158.08 �� 61.95Abc
802.64 �� 26.22Bc
639.95 �� 17.27Cb
100
1 048.06 �� 55.63Ab
998.21 �� 27.68Ad
661.87 �� 52.83Bd
458.34 �� 31.80Cc
200
890.13 �� 24.29Ac
726.07 �� 56.01Be
446.48 �� 63.14Ce
398.46 �� 13.29Cc
CAT (U��g-1)
0
49.58 �� 5.27Ccd
91.00 �� 4.84Ac
66.25 �� 2.84Bc
40.33 �� 2.02Dc
5
58.18 �� 3.69Cb
96.86 �� 3.75Abc
75.56 �� 3.61Bb
44.21 �� 0.98Db
25
66.18 �� 5.21Ca
106.90 �� 3.97Aa
81.33 �� 2.30Ba
50.22 �� 0.99Da
50
53.87 �� 2.95Cbc
102.75 �� 4.81Ab
71.13 �� 2.85Bb
37.60 �� 1.24Dd
100
44.38 �� 4.29Cd
78.95 �� 2.41Ad
52.96 �� 1.65Bd
32.82 �� 1.39De
200
42.96 �� 3.79Cd
63.27 �� 2.40Ae
49.20 �� 2.66Bd
30.58 �� 2.24De
SOD (U��g-1)
0
449.72 �� 15.86Ce
761.24 �� 19.78Ac
511.24 �� 1.00Bd
362.00 �� 4.92Dc
5
531.29 �� 17.48Cc
783.43 �� 7.93Ab
566.58 �� 10.39Bb
391.70 �� 1.45Db
25
598.90 �� 5.61Ca
878.34 �� 3.26Aa
626.77 �� 20.00Ba
416.30 �� 2.84Da
50
566.31 �� 1.98Bb
797.08 �� 3.70Ab
545.40 �� 5.00Cc
387.04 �� 5.43Db
100
516.22 �� 1.12Bc
660.98 �� 10.88Ad
431.00 �� 10.00Ce
333.14 �� 3.22Dd
200
468.37 �� 2.77Bd
629.66 �� 13.24Ae
404.64 �� 4.00Cf
219.90 �� 9.99De
MDA (��mol��g-1)
0
5.29 �� 0.37Dbc
8.83 �� 0.29Cab
9.49 �� 0.23Bbc
12.68 �� 0.20Abc
5
5.19 �� 0.17Dc
8.00 �� 0.62Cbc
9.04 �� 0.33Bcd
11.77 �� 0.16Acd
25
5.04 �� 0.41Dc
7.50 �� 0.75Cc
8.47 �� 0.14Bd
11.29 �� 0.44Ad
50
5.81 �� 0.53Dbc
8.59 �� 0.30Cab
10.40 �� 0.56Bb
13.66 �� 0.35Ab
100
6.00 �� 0.41Dab
8.94 �� 0.63Cab
11.12 �� 0.75Ba
15.22 �� 0.46Aa
200
6.60 �� 0.45Da
9.64 �� 0.61Ca
11.58 �� 0.27Ba
15.53 �� 0.34Aa
Different uppercase letters indicate significant difference among treatments at different times of the same concentration (p < 0.05); different lowercase letters indicate significant difference among different concentrations at the same time (p < 0.05). ��ͬ��д��ĸ��ʾͬһŨ�Ȳ�ͬʱ�䴦�����������(p < 0.05); ��ͬСд��ĸ��ʾͬһʱ�䴦����ͬŨ�ȼ��������(p < 0.05)��
Table 3 ��3 ��3��ͬCaCl2Ũ�ȶԿ�Ҷȸ������Ҷ���غ�����Ӱ��(ƽ��ֵ�������) Table 3Effects of different CaCl2 concentrations on the chlorophyll content of Paspalum wettsteinii seedlings (mean �� SE)
Ҷ���� Chlorophyll (chl) (mg��g-1)
CaCl2Ũ�� CaCl2 concentrations (mmol��L-1)
ʱ�䴦�� Time treatment (d)
7
14
21
28
Chl a (mg��g-1)
0
3.81 �� 0.34Aa
4.41 �� 0.19Aab
3.65 �� 0.32Aa
2.43 �� 0.06Aabc
5
4.38 �� 0.34ABa
4.57 �� 0.20Aab
3.75 �� 0.13ABa
2.74 �� 0.31Bab
25
5.23 �� 0.15Aa
5.00 �� 0.19Aa
4.34 �� 0.11Aa
3.20 �� 0.19Aa
50
4.08 �� 0.16Aa
4.03 �� 0.13Aab
2.89 �� 0.12ABab
2.47 �� 0.13Bab
100
3.46 �� 0.13Aa
3.33 �� 0.11Aab
2.48 �� 0.08Aab
2.20 �� 0.07Abc
200
2.26 �� 0.04Aa
2.06 �� 0.14Ab
1.94 �� 0.24Ab
1.72 �� 0.11Ac
Chl b (mg��g-1)
0
1.06 �� 0.08Aab
1.24 �� 0.03Aa
1.41 �� 0.06Aab
1.26 �� 0.04Aa
5
1.33 �� 0.05Aa
1.71 �� 0.09Aa
2.05 �� 0.07Aa
1.67 �� 0.02Aa
25
1.19 �� 0.06Aa
1.57 �� 0.03Aa
1.75 �� 0.03Aab
1.59 �� 0.02Aa
50
1.12 �� 0.10Aab
1.31 �� 0.06Aa
1.74 �� 0.02Aab
1.45 �� 0.06Aa
100
0.87 �� 0.06Aab
1.20 �� 0.06Aa
1.38 �� 0.09Ab
1.23 �� 0.12Aa
200
0.65 �� 0.04Bb
1.13 �� 0.09Aa
1.20 �� 0.06Ab
0.84 �� 0.13ABa
Chl a+b (mg��g-1)
0
4.74 �� 0.47Aabc
5.92 �� 0.15Aab
6.48 �� 0.24Aab
5.00 �� 0.30Aab
5
5.95 �� 0.09Aa
7.09 �� 0.08Aa
7.85 �� 0.31Aa
6.51 �� 0.17Aa
25
5.44 �� 0.17Aab
6.23 �� 0.37Aab
7.23 �� 0.17Aab
5.77 �� 0.32Aab
50
4.65 �� 0.17Aabc
5.55 �� 0.35Aab
6.19 �� 0.59Aab
5.04 �� 0.33Aab
100
3.69 �� 0.38Bbc
5.05 �� 0.46Abc
5.37 �� 0.18Aab
4.74 �� 0.15Ab
200
3.08 �� 0.65Bc
3.78 �� 0.77ABc
4.24 �� 0.18Ab
3.15 �� 0.15Bc
Different uppercase letters indicate significant difference among treatments at different times of the same concentration (p < 0.05); Different lowercase letters indicate significant difference among different concentrations at the same time (p < 0.05). ��ͬ��д��ĸ��ʾͬһŨ�Ȳ�ͬʱ�䴦�����������(p < 0.05); ��ͬСд��ĸ��ʾͬһʱ�䴦����ͬŨ�ȼ��������(p < 0.05)��
Fig. 3Effects of different CaCl2 concentrations on photosynthetic parameters of Paspalum wettsteinii seedlings (mean �� SE). Ci, intercellular CO2 concentration; Gs, stomatal conductance; Pn, net photosynthetic rate; Tr, transpiration rate. Different uppercase letters indicate significant difference between treatments at different times of the same concentration (p < 0.05); different lowercase letters indicate significant difference between different concentrations at the same time (p < 0.05).
Table 5 ��5 ��5��ͬCaCl2Ũ���¿�Ҷȸ��������̬������ָ�����������ֵ Table 5Sobordinative function among all growth and physiological indices of Paspalum wettsteinii seedlings under different CaCl2 concentrations
.BaiXS (2019). Adaptive responses of soybean leaves to osmotic retion under drought and salt stress .Modern Agricultural Sciences and Technology, (10), 5-6. [��������: 1]
.BorM, ?zdemirF, T��rkanI (2003). The effect of salt stress on lipid peroxidation and antioxidants in leaves of sugar beet Beta vulgaris L. and wild beet Beta maritima L. Plant Science, 164, 77-84. [��������: 1]
.Celik?, AtakC (2012). The effect of salt stress on antioxidative enzymes and proline content of two Turkish tobacco varieties .Turkish Journal of Biology, 36, 339-356. DOI:10.3906/biy-1108-11URL [��������: 1] The aim of this study was to compare the salinity tolerances of 2 oriental tobacco varieties (Izmir Ozbas and Akhisar 97). Salinity stress experiments were performed under both in vitro and in vivo conditions. Seedlings of each variety were subjected to 0, 50, 100, 150, 200, 250, 300, and 350 mM NaCl. Photosynthetic pigment levels, lipid peroxidation rate, total protein content, antioxidant enzyme activities, and proline concentrations were determined for seedlings treated with salt for 14 days. The Akhisar 97 variety was found to be more sensitive to salinity stress than the Izmir Ozbas variety. Although proline is thought to accumulate in salt-tolerant plants, we found a negative correlation between salinity tolerance and proline accumulation in the plants. According to biochemical analyses, there were no differences in SOD, APX, GPX, or CAT activity levels between the 2 varieties, either in vivo or in vitro. However, differences in glutathione reductase (GR) activity between control plants and plants under NaCl stress were statistically significant in both varieties, both in vitro and in vivo. Our results support the hypothesis that GR is a key element in the evaluation of salinity tolerance of tobacco varieties.
.ChenSY, ZhaoYM, LiZX, HanH, HouXL, CaiLP (2018). Effects of Pb, Cd and acid stress on seed germination, seedling growth and antioxidant enzyme activities of Paspalum wettsteinii. Acta Agrestia Sinica, 26, 1173-1180. [��������: 1]
.DoganlarZB, DemirK, BasakH, GulIH (2010). Effects of salt stress on pigment and total soluble protein contents of three different tomato cultivars .African Journal of Agricultural Research, 15, 2056-2065.
.Farhangi-AbrizS, TorabianS (2017). Antioxidant enzyme and osmotic adjustment changes in bean seedlings as affected by biochar under salt stress .Ecotoxicology and Environmental Safety, 137, 64-70. DOI:10.1016/j.ecoenv.2016.11.029URLPMID:27915144 [��������: 1] Salinity damaged cellular membranes through overproduction of reactive oxygen species (ROS), while osmolytes and antioxidant capacities play a vital role in protecting plants from salinity caused oxidative damages. Biochar also could alleviate the negative impacts of salt stress in crops. The pot experiment was conducted to investigate the effects of biochar on some antioxidant enzyme activities and osmolyte adjustments of common bean (Phaseolus vulgaris L. cv. Derakhshan) under salinity stress. Bean plants were subjected to three salinity levels (non-saline, 6 and 12 dSm-1 of NaCl) and biochar treatments (non-biochar, 10% and 20% total pot mass). Shoot and root dry weights of bean were decreased at two salt stress treatments. Salinity increased the activity of catalase (CAT), ascorbate peroxidase (APX), peroxidase (POD), polyphenol oxidase (PPO) and superoxide dismutase (SOD), and the content of malondialdehyde (MDA), oxygen radicals (O2?-), and hydrogen peroxide (H2O2) in leaf and root compared to control. Additionally, increased magnitudes of proline, glycine betaine, soluble sugar and soluble protein contents were more pronounced under 12 dSm-1 NaCl than those under 6 dSm-1 NaCl. In contrast, biochar applied to soil enhanced the shoot and root dry weight in comparison with the non-biochar treatment. Furthermore, all of the antioxidant activities of seedlings in soil treated with biochar, particularly at 20% biochar, declined. With the addition of biochar, the contents of MDA, O2?- and H2O2 displayed remarkable decrease, and the osmotic substances accumulation in leaves and roots also reduced. The presented results supported the view that biochar can contribute to protect common bean seedlings against NaCl stress by alleviating the oxidative stress.
.FeritS?nmez, F��sunG��lser (2016). Effects of humic acid and CaNO32 on nutrient contents in pepper ( Apsicum annuum) seedling under salt stress. Acta Agriculturae Scandinavica, 66, 1-6. [��������: 1]
.GabaraB, KrajewskaM, SteckaE (1995). Calcium effect on number, dimension and activity of nucleoli in cortex cells of pea ( Pisum sativum L.) roots after treatment with heavy metals. Plant Science, 111, 153-161. [��������: 1]
.GrotkoppE, Rejm��nekM, RostTL (2002). Toward a causal explanation of plant invasiveness: Seedling growth and life-history strategies of 29 pine ( Pinus) species. The American Naturalist, 159, 396-419. DOI:10.1086/338995URLPMID:18707424 [��������: 1] We studied 29 pine (Pinus) species to test the hypothesis that invasive species in disturbed habitats have distinct attributes. Seedling relative growth rate (RGR) and measures of invasiveness were positively associated across species as well as within phylogenetically independent contrasts. High RGR, small seed masses, and short generation times characterize pine species that are successful invaders in disturbed habitats. Discriminant analysis and logistic regression revealed that RGR was the most significant factor among these life-history traits separating invasive and noninvasive species. We also explored the causes of differences in RGR among invasive and noninvasive species. While net assimilation rate, leaf mass ratio, and specific leaf area (SLA) were all found to be contributing positively to RGR, SLA was found to be the main component responsible for differences in RGR between invasive and noninvasive pines. We investigated differences in SLA further by studying leaf anatomy, leaf density, and leaf thickness. We also evaluated relative leaf production rate as an important aspect of SLA. We proposed a hypothetical causal network of all relevant variables.
.GuoYY, YuHY, YangMM, KongDS, ZhangYJ (2018). Effect of drought stress on lipid peroxidation, osmotic adjustment and antioxidant enzyme activity of leaves and roots of Lycium ruthenicum Murr. seedling. Russian Journal of Plant Physiology, 65, 244-250. [��������: 1]
.JiaPY, TianFP, LiuYF, LiuY, WuGL, HuY, LuY (2017). Response of leaf physiological indexes to short-term salinity stress for seedlings of Sonchus oleraceus L. Acta Botanica Boreali-Occidentalia Sinica, 37, 1303-1311. [��������: 1]
.JiaXP, DengYM, SunXB, LiangLJ (2015). Impacts of salt stress on the growth and physiological characteristics of Paspalum vaginatum. Acta Prataculturae Sinica, 24, 204-212. [��������: 1]
.JiangZC, LuoWQ, DengY, CaoJH, QinXM, LiYQ, YangQY (2014). The leakage of water and soil in the karst peak cluster depression and its prevention and treatment .Acta Geoscientica Sinica, 35, 535-542. [��������: 1]
.KilliD, HaworthM (2017). Diffusive and metabolic constraints to photosynthesis in quinoa during drought and salt stress .Plants, 6, 49. DOI: 10.3390/plants6040049. DOI:10.3390/plants6040049URLPMID:29039809 [��������: 1] Quinoa (Chenopodium quinoa Willd.) has been proposed as a hardy alternative to traditional grain crops in areas with warm-to-hot climates that are likely to experience increased drought and salt stress in the future. We characterised the diffusive and metabolic limitations to photosynthesis in quinoa exposed to drought and salt stress in isolation and combination. Drought-induced pronounced stomatal and mesophyll limitations to CO? transport, but quinoa retained photosynthetic capacity and photosystem II (PSII) performance. Saline water (300 mmol NaCl-equivalent to 60% of the salinity of sea-water) supplied in identical volumes to the irrigation received by the control and drought treatments induced similar reductions in stomatal and mesophyll conductance, but also reduced carboxylation of ribulose-1,5-bisphosphate carboxylase/oxygenase, regeneration of ribulose-1,5-bisphosphate, increased non-photochemical dissipation of energy as heat and impaired PSII electron transport. This suggests that ion toxicity reduced PN via interference with photosynthetic enzymes and degradation of pigment-protein complexes within the thylakoid membranes. The results of this study demonstrate that the photosynthetic physiology of quinoa is resistant to the effects of drought, but quinoa may not be a suitable crop for areas subject to strong salt stress or irrigation with a concentration of saline water equivalent to a 300 mmol NaCl solution.
.LaiZQ (1989). Studies on tropical and subtropical fine grass Paspalum wettsteiini. Grassland of China, 11, 60-63. [��������: 1]
.LiXF (2006). Speciation of Calcium in Soil and Plants�� Leaves in Karst Ecosystem and Its Ecological Significance. Master degree dissertation, Guangxi Normal University, Guilin Guangxi. [��������: 1]
.LiXG, MengQW, JiangGQ, ZouQ (2003). The susceptibility of cucumber and sweet pepper to chilling under low irradiance is related to energy dissipation and water-water cycle .Photosynthetica, 41, 259-265. DOI:10.1023/B:PHOT.0000011959.30746.c0URL [��������: 1] The photoprotection of energy dissipation and water-water cycle were investigated by comparing chilling sensitivity of photosystems 2 (PS2) and 1 (PS1) in two chilling-sensitive plants, cucumber and sweet pepper, upon exposure to 4 ��C under low irradiance (100 μmol m−2 s−1) for 6 h. During chilling stress, the maximum photochemical efficiency of PS2 (Fv/Fm) decreased only slightly in both plants, but the oxidisable P700 decreased markedly, which indicated that PS1 was more sensitive to chilling treatment under low irradiance than PS2. Sweet pepper leaves had lower Fv/Fm, higher non-photochemical quenching (NPQ), and higher oxidisable P700 during chilling stress. Activity of superoxide dismutase (SOD) and ascorbate peroxidase (APX) in cucumber leaves was higher, but APX activity decreased apparently compared to that at room temperature. The productions of active oxygen species (H2O2, O2−) increased in both plants, faster in cucumber leaves than in sweet pepper leaves. In sweet pepper leaves, a stronger de-epoxidation of the xanthophyll cycle pigments, a higher NPQ could act as a major protective mechanism to reduce the formation of active oxygen species during stress. Thus sensitivity of both plants to chilling under low irradiance was dominated by the protective mechanisms between PS1 and PS2, especially the energy dissipation and the water-water cycle.
.LiZ, TanXF, LuK, LiuZM, WuLL (2017). The effect of CaCl2 on calcium content, photosynthesis, and chlorophyll fluorescence of tung tree seedlings under drought conditions .Photosynthetica, 55, 553-560. DOI:10.1007/s11099-016-0676-xURL [��������: 1]
.LiZW, XuXL, ZhuJX, XuCH, WangKL (2019). Effects of lithology and geomorphology on sediment yield in karst mountainous catchments . Geomorphology, 343, 119-128. DOI:10.1016/j.geomorph.2019.07.001URL [��������: 1]
.LiangWJ, WangML, AiXZ (2009). The role of calcium in regulating photosynthesis and related physiological indexes of cucumber seedlings under low light intensity and suboptimal temperature stress .Scientia Horticulturae, 123, 34-38. DOI:10.1016/j.scienta.2009.07.015URL [��������: 1]
.LongJM, LiXG (2016). Effects of different sowing date on the growth and agronomic traits of Paspalum auriculatum. Journal of Anhui Agricultural Sciences, 44, 124-126. [��������: 1]
.LuoD, ShiYJ, SongFH, LiJC (2019). Effects of salt stress on growth, photosynthetic and fluorescence characteristics, and root architecture of Corylus heterophylla �� C. avellan seedlings. Journal of Applied Ecology, 30, 3376-3384. [��������: 1]
.MiYW, WangGX, GongCW, CaiZP, WuWG (2018). Effects of salt stress on growth and physiology of Isatis indigotica seedlings. Acta Prataculturae Sinica, 27, 43-51. [��������: 1]
.MittlerR, VanderauweraS, GolleryM, van BreusegemF (2004). Reactive oxygen gene network of plants .Trends in Plant Science, 9, 490-498. DOI:10.1016/j.tplants.2004.08.009URL [��������: 1] Reactive oxygen species (ROS) control many different processes in plants. However, being toxic molecules, they are also capable of injuring cells. How this conflict is resolved in plants is largely unknown. Nonetheless, it is clear that the steady-state level of ROS in cells needs to be tightly regulated. In Arabidopsis, a network of at least 152 genes is involved in managing the level of ROS. This network is highly dynamic and redundant, and encodes ROS-scavenging and ROS-producing proteins. Although recent studies have unraveled some of the key players in the network, many questions related to its mode of regulation, its protective roles and its modulation of signaling networks that control growth, development and stress response remain unanswered.
.NingMQ, ZhaoJ (2013). The dynamic evolution of rocky desertification in Guizhou during 2005-2010 .Guizhou Agricultural Science, (9), 75-78. [��������: 1]
.SnehaS, RishiA, ChandraS (2014). Effect of short term salt stress on chlorophyll content, protein and activities of catalase and ascorbate peroxidase enzymes in pearl millet .American Journal of Plant Physiology, 9, 32-37. DOI:10.3923/ajpp.2014.32.37URL [��������: 1]
.SunCC, ZhaoHY, ZhengCX (2017). Effects of NaCl stress on osmolyte and proline metabolism in Ginkgo biloba seedling. Plant Physiology Journal, 53, 470-476. [��������: 1]
.WangB, YuMK, SunHJ, ChengXR, ShanQH, FangYM (2009). Photosynthetic characters of Quercus acutissima from different provenances under effects of salt stress. Journal of Applied Ecology, 20, 1817-1824. [��������: 1]
.WangSJ, LiYB (2007). Problems and development trends about researches on karst rocky desertification .Advances in Earth Science, 22, 573-582. URL [��������: 1] Karst rocky desertification is a unique kind of land desertification which happens in humid climate area in China. There has not been enough knowledge about the karst ecosystem stability and the forming mechanisms of karst rocky desertification, and there has been the lack of perfect prevention and cure techniques. Therefore, the total extending trend of rocky desertification has not been controlled yet effectively. This paper discusses the existing problems of present basic researches on the processes of karst rocky desertification and adaptive recovery of karst ecosystems, including the temporal��spatial changes and driving mechanisms of karst rocky desertification, the soil erosion processes and its driving forces as well as risk evaluation in karst mountainous regions, the relationship of biogeochemical processes in karst rocky desertification with degradation processes of karst ecosystems, adaptive recovery of the degraded karst ecosystems, the optimization of service function and comprehensive adjusting and controlling measures of karst ecosystem, etc. It is emphasized that karst rocky desertification is not induced only by natural processes that happen in the geographical zones, and is the comprehensive ecological problems concerning the naturally, socially and economically driving factors. Multidisciplinary integration and comprehensive research methods based on karst science should be adopted in the researches on karst rocky desertification. The development trends on the above research fields have also been predicted also in the last decades. �� [������, ������ (2007). ��˹��ʯĮ���о����ڵ������뷢չ���� �����ѧ��չ, 22, 573-582.] URL [��������: 1] Karst rocky desertification is a unique kind of land desertification which happens in humid climate area in China. There has not been enough knowledge about the karst ecosystem stability and the forming mechanisms of karst rocky desertification, and there has been the lack of perfect prevention and cure techniques. Therefore, the total extending trend of rocky desertification has not been controlled yet effectively. This paper discusses the existing problems of present basic researches on the processes of karst rocky desertification and adaptive recovery of karst ecosystems, including the temporal��spatial changes and driving mechanisms of karst rocky desertification, the soil erosion processes and its driving forces as well as risk evaluation in karst mountainous regions, the relationship of biogeochemical processes in karst rocky desertification with degradation processes of karst ecosystems, adaptive recovery of the degraded karst ecosystems, the optimization of service function and comprehensive adjusting and controlling measures of karst ecosystem, etc. It is emphasized that karst rocky desertification is not induced only by natural processes that happen in the geographical zones, and is the comprehensive ecological problems concerning the naturally, socially and economically driving factors. Multidisciplinary integration and comprehensive research methods based on karst science should be adopted in the researches on karst rocky desertification. The development trends on the above research fields have also been predicted also in the last decades. ��
.WangWJ, ZhaoLL, WangPC, ChenC, YuQQ, ZhangYJ (2019). Effect of different nitrogen levels on the physiology and ecology of Paspalum wettsteinii. Pratacultural Science, 36, 744-753. [��������: 1]
.XuDH, WangWY, GaoTP, FangXW, GaoXG, LiJH, BuHY, MuJ (2017). Calcium alleviates decreases in photosynthesis under salt stress by enhancing antioxidant metabolism and adjusting solute accumulation in Calligonum mongolicum Conservation Physiology, 5, cox060. DOI:10.1093/conphys/cox060. [��������: 1]
.YangFR, LiuWY, HuangJ, WeiYM, JinQ (2017). Physiological responses of different quinoa varieties to salt stress and evaluation of salt tolerance .Acta Prataculturae Sinica, 26, 77-88. [��������: 1]
.YangYQ, GuoY (2018). Elucidating the molecular mechanisms mediating plant salt-stress responses .New Phytologist, 217, 523-539. DOI:10.1111/nph.14920URLPMID:29205383 [��������: 1] Contents Summary 523 I. Introduction 523 II. Sensing salt stress 524 III. Ion homeostasis regulation 524 IV. Metabolite and cell activity responses to salt stress 527 V. Conclusions and perspectives 532 Acknowledgements 533 References 533 SUMMARY: Excess soluble salts in soil (saline soils) are harmful to most plants. Salt imposes osmotic, ionic, and secondary stresses on plants. Over the past two decades, many determinants of salt tolerance and their regulatory mechanisms have been identified and characterized using molecular genetics and genomics approaches. This review describes recent progress in deciphering the mechanisms controlling ion homeostasis, cell activity responses, and epigenetic regulation in plants under salt stress. Finally, we highlight research areas that require further research to reveal new determinants of salt tolerance in plants.
.ZengC, WangSJ, BaiXY, LiYB, TianYC, LiY, WuLH, LuoGJ (2017). Soil erosion evolution and spatial correlation analysis in a typical karst geomorphology using RUSLE with GIS .Solid Earth, 8, 721-736. DOI:10.5194/se-8-721-2017URL [��������: 1]
.ZhangHH, ZhangXL, LiX, DingJN, ZhuWX, QiF, ZhangT, TianY, SunGY (2012). Effects of NaCl and Na2CO3 stresses on the growth and photosynthesis characteristics of Morus alba seedlings. Journal of Applied Ecology, 23, 625-631. URLPMID:22720603 [��������: 1] Taking 1-year old Morus alba variety 'Qinglong' seedlings as test materials, this paper studied their growth and photosynthetic characteristics under the stresses of different concentration neutral salt NaCl and alkali salt Na2CO3. Salt stresses decreased the plant height and the leaf number, biomass, and photosynthetic capacity of the seedlings markedly. With increasing concentration Na+, the leaf stomatal conductance, transpiration rate, net photosynthetic rate, actual photochemical efficiency, electron transport rate, and photochemical quenching (qP) decreased obviously, the energy dissipation rate increased, and the light use efficiency and photosynthetic capacity dropped down. At low concentrations Na+ (&lt; 150 mmol x L(-1)), the seedlings growth and leaf photosynthetic capacity were slightly inhibited, and the adaptability of the seedlings to the salt stresses increased via the increase of root/shoot ratio. However, this protection mechanism was impaired by increasing salt concentration. Na2CO3 stress (Na+ concentration &gt; 50 mmol x L(-)) had stronger inhibitory effects on the seedlings growth and leaf photosynthetic capacity, and the effect increased with increasing Na+ concentration. It was concluded that at Na+ concentration &lt; 150 mmol x L(-1), the photosynthetic adaptability of M. alba to neutral salt stress was mainly dependent on the plant morphology and photosynthetic metabolism, but at Na+ concentration &gt; 150 mmol x L(-1), the photosynthetic adaptability of M. alba to alkali salt stress was mainly dependent on the photosynthetic metabolism. [�Ż��, ������, ����, ������, ������, ���, ����, ��Ұ, ����� (2012). NaCl��Na2CO3в�ȶ�ɣ����������������Ե�Ӱ�� Ӧ����̬ѧ��, 23, 625-631.] URLPMID:22720603 [��������: 1] Taking 1-year old Morus alba variety 'Qinglong' seedlings as test materials, this paper studied their growth and photosynthetic characteristics under the stresses of different concentration neutral salt NaCl and alkali salt Na2CO3. Salt stresses decreased the plant height and the leaf number, biomass, and photosynthetic capacity of the seedlings markedly. With increasing concentration Na+, the leaf stomatal conductance, transpiration rate, net photosynthetic rate, actual photochemical efficiency, electron transport rate, and photochemical quenching (qP) decreased obviously, the energy dissipation rate increased, and the light use efficiency and photosynthetic capacity dropped down. At low concentrations Na+ (&lt; 150 mmol x L(-1)), the seedlings growth and leaf photosynthetic capacity were slightly inhibited, and the adaptability of the seedlings to the salt stresses increased via the increase of root/shoot ratio. However, this protection mechanism was impaired by increasing salt concentration. Na2CO3 stress (Na+ concentration &gt; 50 mmol x L(-)) had stronger inhibitory effects on the seedlings growth and leaf photosynthetic capacity, and the effect increased with increasing Na+ concentration. It was concluded that at Na+ concentration &lt; 150 mmol x L(-1), the photosynthetic adaptability of M. alba to neutral salt stress was mainly dependent on the plant morphology and photosynthetic metabolism, but at Na+ concentration &gt; 150 mmol x L(-1), the photosynthetic adaptability of M. alba to alkali salt stress was mainly dependent on the photosynthetic metabolism.
.ZhangSR (1999). A discussion on chlorophyll fluorescence kinetics parameters and their significance .Chinese Bulletin of Botany, 34, 444-448. [��������: 1]
The effect of CaCl2 on calcium content, photosynthesis, and chlorophyll fluorescence of tung tree seedlings under drought conditions 1 2017
... λ���ҹ����ϵĹ��ݸ�ԭ�������������ֲ���еĿ�˹�ص���������, Ҳ�ǿ�˹�ط�������͵�һ������(Zeng et al., 2017; Li et al., 2019).��ֲ��㷺��̼�����Ҳ��¶�����1.3 �� 105 km2, ռ����ȫʡ�������73% (��ï᪺��Լ�, 2013).�����������ȴ�ʪ������, ̼�����Ҳ��ܵ���ʴ��Ϊ����, �����������λ�(���ҳϵ�, 2014), ����̼�����ҷ�Χռ�������λ���Χ��1%-3%, ��ͬγ�ȹ����ε�����2-3��(��С��, 2006).������ʴ����, ���������е����ס��ص����ִ�����ʧ(�����ܺ�������, 2007), ������в��˹�ص���ֲ�������, ���ѡ����ֲ�߸��ε�ֲ��Կ�˹�ص����ݵ�����ҵ�ķ�չ����̬�����ĸ��ƾ�����Ҫ����.����ֲ�����������������Ԫ��(Liang et al., 2009), ����ͨ��ά��ϸ��Ĥͨ�ԡ��������䡢�ź�ת���ȷ�ʽֱ�Ӳ���ֲ��ֿ���в��(Ferit & F��sun, 2016)���ؽ���(Gabara et al., 1995)�ɺ�(Li et al., 2017)�ȷ������澳, ����ֲͬ��ԸƵ������Բ�ͬ, ȱ�ơ��ƻ��Ũ�ȹ��߶���Ӱ��ֲ������, ���˸�Ũ�Ȳ��ܴٽ�ֲ������, ����������������. ...
Effects of lithology and geomorphology on sediment yield in karst mountainous catchments 1 2019
... λ���ҹ����ϵĹ��ݸ�ԭ�������������ֲ���еĿ�˹�ص���������, Ҳ�ǿ�˹�ط�������͵�һ������(Zeng et al., 2017; Li et al., 2019).��ֲ��㷺��̼�����Ҳ��¶�����1.3 �� 105 km2, ռ����ȫʡ�������73% (��ï᪺��Լ�, 2013).�����������ȴ�ʪ������, ̼�����Ҳ��ܵ���ʴ��Ϊ����, �����������λ�(���ҳϵ�, 2014), ����̼�����ҷ�Χռ�������λ���Χ��1%-3%, ��ͬγ�ȹ����ε�����2-3��(��С��, 2006).������ʴ����, ���������е����ס��ص����ִ�����ʧ(�����ܺ�������, 2007), ������в��˹�ص���ֲ�������, ���ѡ����ֲ�߸��ε�ֲ��Կ�˹�ص����ݵ�����ҵ�ķ�չ����̬�����ĸ��ƾ�����Ҫ����.����ֲ�����������������Ԫ��(Liang et al., 2009), ����ͨ��ά��ϸ��Ĥͨ�ԡ��������䡢�ź�ת���ȷ�ʽֱ�Ӳ���ֲ��ֿ���в��(Ferit & F��sun, 2016)���ؽ���(Gabara et al., 1995)�ɺ�(Li et al., 2017)�ȷ������澳, ����ֲͬ��ԸƵ������Բ�ͬ, ȱ�ơ��ƻ��Ũ�ȹ��߶���Ӱ��ֲ������, ���˸�Ũ�Ȳ��ܴٽ�ֲ������, ����������������. ...
The role of calcium in regulating photosynthesis and related physiological indexes of cucumber seedlings under low light intensity and suboptimal temperature stress 1 2009
... λ���ҹ����ϵĹ��ݸ�ԭ�������������ֲ���еĿ�˹�ص���������, Ҳ�ǿ�˹�ط�������͵�һ������(Zeng et al., 2017; Li et al., 2019).��ֲ��㷺��̼�����Ҳ��¶�����1.3 �� 105 km2, ռ����ȫʡ�������73% (��ï᪺��Լ�, 2013).�����������ȴ�ʪ������, ̼�����Ҳ��ܵ���ʴ��Ϊ����, �����������λ�(���ҳϵ�, 2014), ����̼�����ҷ�Χռ�������λ���Χ��1%-3%, ��ͬγ�ȹ����ε�����2-3��(��С��, 2006).������ʴ����, ���������е����ס��ص����ִ�����ʧ(�����ܺ�������, 2007), ������в��˹�ص���ֲ�������, ���ѡ����ֲ�߸��ε�ֲ��Կ�˹�ص����ݵ�����ҵ�ķ�չ����̬�����ĸ��ƾ�����Ҫ����.����ֲ�����������������Ԫ��(Liang et al., 2009), ����ͨ��ά��ϸ��Ĥͨ�ԡ��������䡢�ź�ת���ȷ�ʽֱ�Ӳ���ֲ��ֿ���в��(Ferit & F��sun, 2016)���ؽ���(Gabara et al., 1995)�ɺ�(Li et al., 2017)�ȷ������澳, ����ֲͬ��ԸƵ������Բ�ͬ, ȱ�ơ��ƻ��Ũ�ȹ��߶���Ӱ��ֲ������, ���˸�Ũ�Ȳ��ܴٽ�ֲ������, ����������������. ...
Calcium alleviates decreases in photosynthesis under salt stress by enhancing antioxidant metabolism and adjusting solute accumulation in Calligonum mongolicum 1 2017
 ,1, ���ľ�1, ������2,3, �����1, ������
,1, ���ľ�1, ������2,3, �����1, ������ ,1,2,*
,1,2,*
 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPT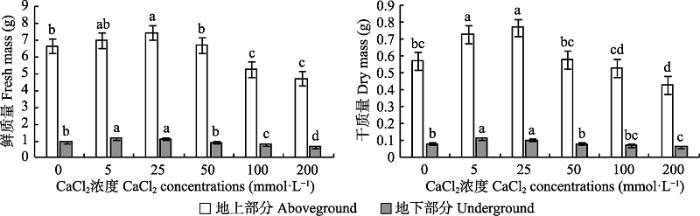 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPT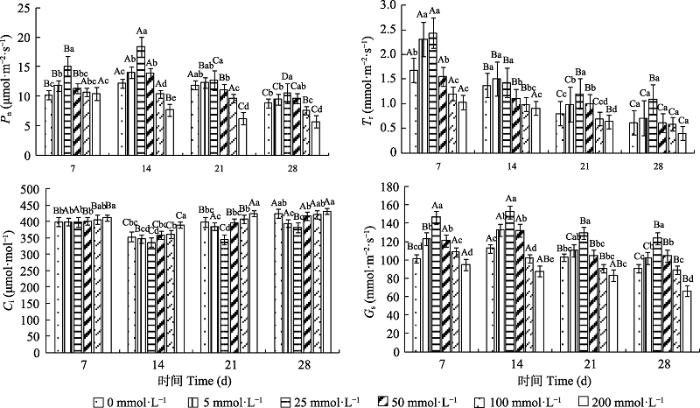 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}