Community assembly processes in fragmented forests and its testing methods
Jin-Liang LIU,1,2, Ming-Jian YU,,2,*1College of Life and Environmental Science, Wenzhou University, Wenzhou, Zhejiang 325035, China 2College of Life Sciences, Zhejiang University, Hangzhou 310058, China
Supported by the National Natural Science Foundation of China(31870401) Supported by the National Natural Science Foundation of China(31901104) Supported by the National Natural Science Foundation of China(31570524) the Natural Science Foundation of Zhejiang Province(LD19C030001) the National Key R&D Program(2018YFE0112800)
Abstract Nowadays, almost all forests in the world are fragmented, and thus, it is necessary to understand how forest fragmentation influence assembly of forest communities. This review summarized the main community assembly processes in the field of community ecology, namely ecological drift, dispersal, selection and speciation, and summarized the relative roles of these community assembly processes in fragmented forests. Due to differences in formation trajectory of different forests in fragmented region, the relative effects of the above four ecological processes are different for different types of forest communities: reassembled community (i.e., forest communities re-assembled through secondary succession in fragmented habitats) and disassembled community (i.e., continuous forest disassembled into fragmented forests). The effects of ecological processes can be effectively tested by analyzing short-term observed species distribution pattern within and among communities (e.g., species abundance distribution analyses, null model combined with beta diversity analyses, and convergence and divergence of functional trait distribution, etc.), controlled experiments, and long-term community monitoring (e.g., community dynamics analysis). Yet, it is insufficient to assess ecological processes undergoing habitat fragmentation by controlled experiments. In the future, studies should focus on developing and testing theoretical models, designing controlled experiments to investigate varied ecological processes undergoing habitat fragmentation, and combining ecological theory with practical biodiversity conservation. Keywords:community assembly;habitat fragmentation;dispersal limitation;neutral theory;niche theory
片段化森林形成后, 群落间物种的扩散(含果实/种子的传播和动物的迁移等)将受到限制(扩散限制), 并且斑块内生境条件和物种间关系的改变将使得能够适应该环境的物种存活(选择过程), 森林斑块面积的减少将增加物种的随机死亡概率(生态漂变), 斑块间隔离度的增加会降低基因交流的频率, 可能促使新物种的产生(物种形成)(Lasky & Keitt, 2013; Vellend, 2016)。而在已有片段化森林群落的研究中, 由于局域群落的定义不明确, 且物种组成受救援效应和源-库动态(附件I)的影响(Leibold et al., 2004), 使得选择、扩散、生态漂变和物种形成等生态过程在片段化森林中的作用有时会被混淆(Das et al., 2017)。
随着森林植被的恢复, 虽然森林总覆盖面积将处于增加趋势, 但森林的片段化程度仍可能逐年增加(Kozak et al., 2018; Liu et al., 2019a)。通过对片段化森林群落构建过程的研究和理论整合, 我们可以更好地理解不同生态过程对群落内物种组成和多样性的贡献, 从而更加有效地指导片段化生境中森林群落的恢复和对生物多样性的保护, 对于后续的森林管理、政策制定、生态系统服务功能提升和物种保护等均具有非常重要的意义。
Fig. 1Timeline of key events in fragmentation theory, landscape experiments, and discourse on application for conservation. Figure was redrawn from Resasco et al., (2017). Major experiments and projects and the corresponding references are as following: Biological Dynamics of Forest Fragments Project (BDFFP; Brazil)(Laurance et al., 2011); Kansas Fragmentation Experiment (USA)(Holt et al., 1995); Wog-Wog Habitat Fragmentation Experiment (Wog-Wog; Australia)(Margules, 1992); Savannah River Site Corridor Experiment (SRS Corridor Experiment; USA)(Haddad & Baum, 1999; Tewksbury et al., 2002); Moss Fragmentation Experiments (UK: Gonzalez et al. 1998; Canada: Lindo et al., 2012); The Thousand Island Lake Experiments (China)(Yu et al., 2012; Liu et al., 2020); The Stability of Altered Forest Ecosystems (SAFE) Project (Malaysia)(Ewers et al., 2011); Metatron experiment (France)(Legrand et al., 2012).
Fig. 2Conceptual cartoon illustrating processes in forest community assembly in fragmented habitats. Redrawn from HilleRisLambers et al. (2012) and Vellend (2016). Different letters denote different species; circles of different sizes denote different size patches; different fan-shaped areas in circles denote the relative abundance of each species in one community.
1.1 扩散
扩散是有机体或其繁殖体离开出生地后的迁移或传播, 该过程对群落构建的影响主要依赖于扩散者产生群落(源群落)和抵达群落(汇群落)的大小和物种组成(Holyoak et al., 2005)。与连续森林相比, 存在隔离的森林斑块将限制物种在斑块间的自由扩散(如种子传播), 即群落间物种的扩散限制。根据源群落和汇群落的相对大小, 片段化生境中物种的扩散可以分为两种情形: 一是大陆-岛屿模型, 该模型中将扩散假定为单向扩散, 由无限大的源群落(大陆或大面积斑块)向一个或多个小的、隔离的汇群落(岛屿或小面积斑块)中扩散, 源群落的物种组成相对恒定, 而汇群落的物种组成变化则高于源群落; 二是岛屿模型, 该模型中的群落(岛屿或小面积斑块)通过扩散相互联系在一起, 形成集合群落(Holyoak et al., 2005)。一般而言, 扩散过程对群落构建的作用是受其他生态过程影响的, 尤其是选择过程和生态漂变(Ron et al., 2018)。
AckerlyDD, CornwellWK ( 2007). A trait-based approach to community assembly: Partitioning of species trait values into within- and among-community components Ecology Letters, 10, 135-145. [本文引用: 1]
AlexanderHM, FosterBL, Ford BallantyneIV, CollinsCD, AntonovicsJ, HoltRD ( 2012). Metapopulations and metacommunities: Combining spatial and temporal perspectives in plant ecology Journal of Ecology, 100, 88-103. [本文引用: 1]
AltermattF, FronhoferEA, GarnierA, GiomettoA, HammesF, KleckaJ, LegrandD, M?chlerE, MassieTM, PennekampF, PlebaniM, PontarpM, SchtickzelleN, ThuillierV, PetcheyOL ( 2015). Big answers from small worlds: A user?s guide for protist microcosms as a model system in ecology and evolution Methods in Ecology and Evolution, 6, 218-231. [本文引用: 1]
BaiB, LiN, LuCH ( 2011). Influence of habitat fragmentation on frugivores and their seed dispersal Chinese Journal of Ecology, 30, 2613-2620. [本文引用: 1]
BaynesJ, HerbohnJ, ChazdonRL, NguyenH, FirnJ, GregorioN, LambD ( 2016). Effects of fragmentation and landscape variation on tree diversity in post-logging regrowth forests of the Southern Philippines Biodiversity and Conservation, 25, 923-941. [本文引用: 1]
BenchimolM, PeresCA ( 2015). Edge-mediated compositional and functional decay of tree assemblages in Amazonian forest islands after 26 years of isolation Journal of Ecology, 103, 408-420. [本文引用: 2]
ChaseJM, LeiboldMA ( 2003). Ecological Niches: Linking Classical and Contemporary Approaches. University of Chicago Press, Chicago. [本文引用: 1]
ChaseJM, MyersJA ( 2011). Disentangling the importance of ecological niches from stochastic processes across scales Philosophical Transactions of the Royal Society B: Biological Sciences, 366, 2351-2363. [本文引用: 2]
ChenL, MiXC, MaKP ( 2014). Niche differentiation and its consequence on biodiversity maintenance in forest communities Chinese Bulletin of Life Sciences, 26, 112-117. [本文引用: 1]
ChessonP ( 2000). Mechanisms of maintenance of species diversity Annual Review of Ecology and Systematics, 31, 343-366. [本文引用: 3]
ChessonP ( 2018). Updates on mechanisms of maintenance of species diversity Journal of Ecology, 106, 1773-1794. [本文引用: 3]
ChessonPL, WarnerRR ( 1981). Environmental variability promotes coexistence in lottery competitive systems The American Naturalist, 117, 923-943. [本文引用: 1]
CirtwillAR, StoufferDB ( 2016). Knowledge of predator-prey interactions improves predictions of immigration and extinction in island biogeography Global Ecology and Biogeography, 25, 900-911. [本文引用: 1]
CollinsCD, Banks-LeiteC, BrudvigLA, FosterBL, CookWM, DamschenEI, AndradeA, AustinM, CamargoJL, DriscollDA, HoltRD, LauranceWF, NichollsAO, OrrockJL ( 2017). Fragmentation affects plant community composition over time Ecography, 40, 119-130. [本文引用: 3]
ConnellJH, TraceyJG, WebbLJ ( 1984). Compensatory recruitment, growth, and mortality as factors maintaining rain forest tree diversity Ecological Monographs?, 54, 141-164. [本文引用: 1]
CottenieK ( 2005). Integrating environmental and spatial processes in ecological community dynamics Ecology Letters, 8, 1175-1182. [本文引用: 1]
DamschenEI, BrudvigLA, BurtMA, Fletcher JrRJ, HaddadNM, LeveyDJ, OrrockJL, ResascoJ, TewksburyJJ ( 2019). Ongoing accumulation of plant diversity through habitat connectivity in an 18-year experiment Science, 365, 1478-1480. [本文引用: 1]
DasAA, JohnR, AnandM ( 2017). Does structural connectivity influence tree species distributions and abundance in a naturally discontinuous tropical forest formation? Journal of Vegetation Science, 28, 7-18. [本文引用: 2]
DaviesKF, HolyoakM, PrestonKA, OffemanVA, LumQ ( 2009). Factors controlling community structure in heterogeneous metacommunities Journal of Animal Ecology, 78, 937-944. [本文引用: 1]
DebinskiDM, HoltRD ( 2000). A survey and overview of habitat fragmentation experiments Conservation Biology, 14, 342-355. [本文引用: 2]
ErnestSKM, BrownJH, ThibaultKM, WhiteEP, GoheenJR ( 2008). Zero sum, the niche, and metacommunities: Long-term dynamics of community assembly The American Naturalist, 172, E257-E269. DOI: 10.1086/592402. [本文引用: 1]
EwersRM, AndradeA, LauranceSG, CamargoJL, LovejoyTE, LauranceWF ( 2017). Predicted trajectories of tree community change in Amazonian rainforest fragments Ecography, 40, 26-35. [本文引用: 3]
EwersRM, DidhamRK, FahrigL, FerrazG, HectorA, HoltRD, KaposV, ReynoldsG, SinunW, SnaddonJL, TurnerEC ( 2011). A large-scale forest fragmentation experiment: The stability of altered forest ecosystems project Philosophical Transactions of the Royal Society B: Biological Sciences, 366, 3292-3302. [本文引用: 2]
FahrigL ( 2003). Effects of habitat fragmentation on biodiversity Annual Review of Ecology, Evolution, and Systematics, 34, 487-515. [本文引用: 2]
FahrigL ( 2013). Rethinking patch size and isolation effects: The habitat amount hypothesis Journal of Biogeography, 40, 1649-1663. [本文引用: 2]
FahrigL ( 2017). Ecological responses to habitat fragmentation per se Annual Review of Ecology, Evolution, and Systematics, 48, 1-23. [本文引用: 1]
FahrigL ( 2019). Habitat fragmentation: A long and tangled tale Global Ecology and Biogeography, 28, 33-41. [本文引用: 1]
FattoriniS, RigalF, CardosoP, BorgesPAV ( 2016). Using species abundance distribution models and diversity indices for biogeographical analyses Acta Oecologica, 70, 21-28. [本文引用: 1]
Favre-BacL, Lamberti-RaverotB, PuijalonS, ErnoultA, BurelF, GuillardL, MonyC ( 2017). Plant dispersal traits determine hydrochorous species tolerance to connectivity loss at the landscape scale Journal of Vegetation Science, 28, 605-615. [本文引用: 1]
FukamiT ( 2010). Community assembly dynamics in space In: Verhoef HA, Morin PJ eds. Community Ecology: Processes, Models, and Applications. Oxford University Press, Oxford, USA. 45-54. [本文引用: 2]
FukamiT ( 2015). Historical contingency in community assembly: Integrating niches, species pools, and priority effects Annual Review of Ecology, Evolution, and Systematics, 46, 1-23. [本文引用: 3]
GastonKJ, BlackburnTM ( 2008). Pattern and Process in Macroecology John Wiley & Sons, Oxford. [本文引用: 1]
GenuaL, StartD, GilbertB ( 2017). Fragment size affects plant herbivory via predator loss Oikos, 126, 1357-1365. [本文引用: 1]
GonzalezA, LawtonJH, GilbertFS, BlackburnTM, Evans-?FrekeI ( 1998). Metapopulation dynamics, abundance, and distribution in a microecosystem Science, 281, 2045-2047. [本文引用: 2]
GravelD, MassolF, CanardE, MouillotD, MouquetN ( 2011). Trophic theory of island biogeography Ecology Letters, 14, 1010-1016. [本文引用: 1]
HaddadNM, BaumKA ( 1999). An experimental test of corridor effects on butterfly densities Ecological Applications, 9, 623-633. [本文引用: 4]
HanskiI ( 1994). A practical model of metapopulation dynamics Journal of Animal Ecology, 63, 151-162. [本文引用: 1]
HarrisonS, BrunaE ( 1999). Habitat fragmentation and largescale conservation: What do we know for sure? Ecography, 22, 225-232. [本文引用: 1]
HilleRisLambersJ, AdlerPB, HarpoleWS, LevineJM, MayfieldMM ( 2012). Rethinking community assembly through the lens of coexistence theory Annual Review of Ecology, Evolution, and Systematics, 43, 227-248. [本文引用: 3]
HoltRD, RobinsonGR, GainesMS ( 1995). Vegetation dynamics in an experimentally fragmented landscape Ecology, 76, 1610-1624. [本文引用: 3]
HolyoakM, LeiboldMA, HoltRD ( 2005). Metacommunities: Spatial Dynamics and Ecological Communities. University of Chicago Press, Chicago. [本文引用: 6]
HubbellSP ( 2001). The Unified Neutral Theory of Biodiversity and Biogeography. Princeton University Press, Princeton. [本文引用: 7]
HubbellSP, ConditR, FosterRB, GrubbP, ThomasC ( 1990). Presence and absence of density dependence in a neotropical tree community Philosophical Transactions of the Royal Society of London B: Biological Sciences, 330, 269-281. [本文引用: 1]
HyattLA, RosenbergMS, HowardTG, BoleG, FangW, AnastasiaJ, BrownK, GrellaR, HinmanK, KurdzielJP, GurevitchJ ( 2003). The distance dependence prediction of the Janzen-Connell hypothesis: A meta-analysis Oikos, 103, 590-602. [本文引用: 1]
JacquetC, MouillotD, KulbickiM, GravelD ( 2017). Extensions of Island Biogeography Theory predict the scaling of functional trait composition with habitat area and isolation Ecology Letters, 20, 135-146. [本文引用: 1]
JanzenDH ( 1970). Herbivores and the number of tree species in tropical forests The American Naturalist, 104, 501-528. [本文引用: 1]
JiangF, XunYH, CaiHY, JinGZ ( 2018). Functional traits can improve our understanding of niche- and dispersal-based processes Oecologia, 186, 783-792. [本文引用: 1]
JiangL, PatelSN ( 2008). Community assembly in the presence of disturbance: A microcosm experiment Ecology, 89, 1931-1940. [本文引用: 1]
KadmonR, PulliamHR ( 1995). Effects of isolation, logging and dispersal on woody-species richness of islands Vegetatio, 116, 63-68. [本文引用: 1]
KenkelNC ( 1988). Pattern of self-thinning in jack pine: Testing the random mortality hypothesis Ecology, 69, 1017-1024. [本文引用: 1]
KozakJ, Zió?kowskaE, VogtP, DoboszM, KaimD, KoleckaN, OstafinK ( 2018). Forest-cover increase does not trigger forest-fragmentation decrease: Case study from the Polish carpathians Sustainability, 10, 1472. DOI: 10.3390/su10051472. [本文引用: 1]
KraftNJB, AckerlyDD ( 2010). Functional trait and phylogenetic tests of community assembly across spatial scales in an Amazonian forest Ecological Monographs, 80, 401-422. [本文引用: 1]
KraftNJB, ComitaLS, ChaseJM, SandersNJ, SwensonNG, CristTO, StegenJC, VellendM, BoyleB, AndersonMJ, CornellHV, DaviesKF, FreestoneAL, InouyeBD, HarrisonSP, MyersJA ( 2011). Disentangling the drivers of diversity along latitudinal and elevational gradients Science, 333, 1755-1758. [本文引用: 1]
KraftNJB, ValenciaR, AckerlyDD ( 2008). Functional traits and niche-based tree community assembly in an Amazonian forest Science, 322, 580-582. [本文引用: 2]
KrishnadasM, BagchiR, SridharaS, ComitaLS ( 2018). Weaker plant-enemy interactions decrease tree seedling diversity with edge-effects in a fragmented tropical forest Nature Communications, 9, 4523. [本文引用: 3]
LariosL, PearsonDE, MaronJL ( 2017). Incorporating the effects of generalist seed predators into plant community theory Functional Ecology, 31, 1856-1867. [本文引用: 1]
LaskyJR, KeittTH ( 2013). Reserve size and fragmentation alter community assembly, diversity, and dynamics The American Naturalist, 182, E142-E160. DOI: 10.1086/673205. [本文引用: 2]
LauranceWF ( 2008). Theory meets reality: How habitat fragmentation research has transcended island biogeographic theory Biological Conservation, 141, 1731-1744. [本文引用: 1]
LauranceWF, CamargoJLC, Luiz?oRCC, LauranceSG, PimmSL, BrunaEM, StoufferPC, Bruce WilliamsonG, Benítez-MalvidoJ, VasconcelosHL, Van HoutanKS, ZartmanCE, BoyleSA, DidhamRK, AndradeA, LovejoyTE ( 2011). The fate of Amazonian forest fragments: A 32-year investigation Biological Conservation, 144, 56-67. [本文引用: 5]
LauranceWF, Delam?nicaP, LauranceSG, VasconcelosHL, LovejoyTE ( 2000). Rainforest fragmentation kills big trees Nature, 404, 836. [本文引用: 1]
LauranceWF, NascimentoHEM, LauranceSG, AndradeA, RibeiroJELS, GiraldoJP, LovejoyTE, ConditR, ChaveJ, HarmsKE, D’AngeloS ( 2006). Rapid decay of tree-community composition in Amazonian forest fragments Proceedings of the National Academy of Sciences of the United States of America, 103, 19010-19014. [本文引用: 1]
LegendreP, de CáceresM ( 2013). Beta diversity as the variance of community data: Dissimilarity coefficients and partitioning Ecology Letters, 16, 951-963. [本文引用: 1]
LegendreP, MiXC, RenHB, MaKP, YuMJ, SunIF, HeFL ( 2009). Partitioning beta diversity in a subtropical broad-leaved forest of China Ecology, 90, 663-674. [本文引用: 2]
LegrandD, GuillaumeO, BaguetteM, CoteJ, TrochetA, CalvezO, ZajitschekS, ZajitschekF, LecomteJ, Be?nardQ, Le GalliardJF, ClobertJ ( 2012). The Metatron: An experimental system to study dispersal and metaecosystems for terrestrial organisms Nature Methods, 9, 828-833. [本文引用: 2]
LeiboldMA, HolyoakM, MouquetN, AmarasekareP, ChaseJM, HoopesMF, HoltRD, ShurinJB, LawR, TilmanD, LoreauM, GonzalezA ( 2004). The metacommunity concept: A framework for multi-scale community ecology Ecology Letters, 7, 601-613. [本文引用: 2]
LettenAD, KePJ, FukamiT ( 2017). Linking modern coexistence theory and contemporary niche theory Ecological Monographs, 87, 161-177. [本文引用: 1]
LevinsR ( 1969). Some demographic and genetic consequences of environmental heterogeneity for biological control Bulletin of the Entomological Society of America, 15, 237-240. [本文引用: 1]
LindoZ, WhiteleyJ, GonzalezA ( 2012). Traits explain community disassembly and trophic contraction following experimental environmental change Global Change Biology, 18, 2448-2457. [本文引用: 2]
LiuJJ, CoomesDA, GibsonL, HuG, LiuJL, LuoYQ, WuCP, YuMJ ( 2019a). Forest fragmentation in China and its effect on biodiversity Biological Reviews, 94, 1636-1657. [本文引用: 2]
LiuJJ, CoomesDA, HuG, LiuJL, YuJJ, LuoYQ, YuMJ ( 2019b). Larger fragments have more late-successional species of woody plants than smaller fragments after 50 years of secondary succession Journal of Ecology, 107, 582-594. [本文引用: 4]
LiuJJ, SlikF, CoomesDA, CorlettRT, WangYP, WilsonM, HuG, DingP, YuMJ ( 2019c). The distribution of plants and seed dispersers in response to habitat fragmentation in an artificial island archipelago Journal of Biogeography, 46, 1152-1162. [本文引用: 1]
LiuJJ, SlikJWF ( 2014). Forest fragment spatial distribution matters for tropical tree conservation Biological Conservation, 171, 99-106. [本文引用: 2]
LiuJL, MatthewsTJ, ZhongL, LiuJJ, WuDH, YuMJ ( 2020). Environmental filtering underpins the island species-area relationship in a subtropical anthropogenic archipelago Journal of Ecology, 108, 424-432. [本文引用: 3]
LiuJL, QianH, JinY, WuCP, ChenJH, YuSQ, WeiXL, JinXF, LiuJJ, YuMJ ( 2016). Disentangling the drivers of taxonomic and phylogenetic beta diversities in disturbed and undisturbed subtropical forests Scientific Reports, 6, 35926. DOI: 10.1038/srep35926. [本文引用: 3]
LiuJL, VellendM, WangZH, YuMJ ( 2018). High beta diversity among small islands is due to environmental heterogeneity rather than ecological drift Journal of Biogeography, 45, 2252-2261. [本文引用: 4]
LiuXY, ZhaoCL, XuS, LiangQM, ZhuXT, LiL, YanER ( 2019). Beta diversity of vascular plants and its drivers in sea-islands of eastern China Biodiversity Science, 27, 380-387. [本文引用: 2]
LopezBE, BurgioKR, CarlucciMB, PalmquistKA, ParadaA, WeinbergerVP, HurlbertAH ( 2016). A new framework for inferring community assembly processes using phylogenetic information, relevant traits and environmental gradients One Ecosystem, 1, e9501. DOI: 10.3897/oneeco.1.e9501. [本文引用: 1]
LuoYQ, YuMJ, YuJJ, ZhengSL, LiuJJ, YuMJ ( 2017). Effects of plant traits and the relative abundance of common woody species on seedling herbivory in the Thousand Island Lake region Chinese Journal of Plant Ecology, 41, 1033-1040. [本文引用: 1]
MariniL, BruunHH, HeikkinenRK, HelmA, HonnayO, KraussJ, KühnI, LindborgR, P?rtelM, BommarcoR ( 2012). Traits related to species persistence and dispersal explain changes in plant communities subjected to habitat loss Diversity and Distributions, 18, 898-908. [本文引用: 1]
MatthewsTJ, BorgesPAV, de AzevedoEB, WhittakerRJ ( 2017). A biogeographical perspective on species abundance distributions: Recent advances and opportunities for future research Journal of Biogeography, 44, 1705-1710. [本文引用: 1]
MatthewsTJ, Cottee-JonesHEW, WhittakerRJ ( 2015a). Quantifying and interpreting nestedness in habitat islands: A synthetic analysis of multiple datasets Diversity and Distributions, 21, 392-404. [本文引用: 1]
MatthewsTJ, WhittakerRJ ( 2015b). On the species abundance distribution in applied ecology and biodiversity management Journal of Applied Ecology, 52, 443-454. [本文引用: 1]
McGarigalK, CushmanSA ( 2002). Comparative evaluation of experimental approaches to the study of habitat fragmentation effects Ecological Applications, 12, 335-345. [本文引用: 1]
McGillBJ, EtienneRS, GrayJS, AlonsoD, AndersonMJ, BenechaHK, DornelasM, EnquistBJ, GreenJL, HeF, HurlbertAH, MagurranAE, MarquetPA, MaurerBA, OstlingA, SoykanCU, UglandKI, WhiteEP ( 2007). Species abundance distributions: Moving beyond single prediction theories to integration within an ecological framework Ecology Letters, 10, 995-1015. [本文引用: 2]
McPeekMA ( 2007). The macroevolutionary consequences of ecological differences among species Palaeontology, 50, 111-129. [本文引用: 1]
MittelbachGG, SchemskeDW ( 2015). Ecological and evolutionary perspectives on community assembly Trends in Ecology & Evolution, 30, 241-247. [本文引用: 1]
MoriAS, IsbellF, SeidlR ( 2018). β-diversity, community assembly, and ecosystem functioning Trends in Ecology & Evolution, 33, 549-564. [本文引用: 1]
MouchetMA, VillégerS, MasonNWH, MouillotD ( 2010). Functional diversity measures: An overview of their redundancy and their ability to discriminate community assembly rules Functional Ecology, 24, 867-876. [本文引用: 1]
MouquetN, LoreauM ( 2003). Community patterns in source-sink metacommunities The American Naturalist, 162, 544-557. [本文引用: 1]
MutshindaCM, O’HaraRB, WoiwodIP ( 2009). What drives community dynamics? Proceedings of the Royal Society B: Biological Sciences, 276, 2923-2929. [本文引用: 1]
MyersJA, LaMannaJA ( 2016). The promise and pitfalls of β-diversity in ecology and conservation Journal of Vegetation Science, 27, 1081-1083. [本文引用: 1]
NiuKC, LiuYN, ShenZH, HeFL, FangJY ( 2009). Community assembly: The relative importance of neutral theory and niche theory Biodiversity Science, 17, 579-593. [本文引用: 1]
?ckingerE, SchweigerO, CristTO, DebinskiDM, KraussJ, KuussaariM, PetersenJD, P?yryJ, SetteleJ, SummervilleKS, BommarcoR ( 2010). Life-history traits predict species responses to habitat area and isolation: A cross-continental synthesis Ecology Letters, 13, 969-979. [本文引用: 1]
OjimaMN, JiangL ( 2017). Interactive effects of disturbance and dispersal on community assembly Oikos, 126, 682-691. [本文引用: 1]
PearseWD, BarbosaAM, FritzSA, KeithSA, HarmonLJ, HarteJ, SilvestroD, XiaoX, DaviesTJ ( 2018). Building up biogeography: Pattern to process Journal of Biogeography, 45, 1223-1230. [本文引用: 1]
PengSY, HuG, YuMJ ( 2014). Beta diversity of vascular plants and its influencing factors on islands in the Thousand Island Lake Acta Ecologica Sinica, 34, 3866-3872. [本文引用: 1]
PerronneR, MunozF, BorgyB, ReboudX, GabaS ( 2017). How to design trait-based analyses of community assembly mechanisms: Insights and guidelines from a literature review Perspectives in Plant Ecology, Evolution and Systematics, 25, 29-44. [本文引用: 1]
QiaoXJ, JabotF, TangZY, JiangMX, FangJY ( 2015a). A latitudinal gradient in tree community assembly processes evidenced in Chinese forests Global Ecology and Biogeography, 24, 314-323. [本文引用: 1]
QiaoXJ, LiQX, JiangQH, LuJM, FranklinS, TangZY, WangQG, ZhangJX, LuZJ, BaoDC, GuoYL, LiuHB, XuYZ, JiangMX ( 2015b). Beta diversity determinants in Badagongshan, a subtropical forest in central China Scientific Reports, 5, 17043. DOI: 10.1038/srep17043. [本文引用: 1]
ResascoJ, BrunaEM, HaddadNM, Banks-LeiteC, MargulesCR ( 2017). The contribution of theory and experiments to conservation in fragmented landscapes Ecography, 40, 109-118. [本文引用: 5]
RicklefsRE ( 2004). A comprehensive framework for global patterns in biodiversity Ecology Letters, 7, 1-15. [本文引用: 1]
RicklefsRE, SchluterD ( 1993). Species Diversity in Ecological Communities: Historical and Geographical Perspectives. University of Chicago Press, Chicago. [本文引用: 1]
RomingerAJ, GoodmanKR, LimJY, ArmstrongEE, BeckingLE, BennettGM, BrewerMS, CotorasDD, EwingCP, HarteJ, MartinezND, O’GradyPM, PercyDM, PriceDK, RoderickGK, ShawKL, ValdovinosFS, GrunerDS, GillespieRG ( 2016). Community assembly on isolated islands: Macroecology meets evolution Global Ecology and Biogeography, 25, 769-780. [本文引用: 5]
RonR, Fragman-SapirO, KadmonR ( 2018). Dispersal increases ecological selection by increasing effective community size Proceedings of the National Academy of Sciences of the United States of America, 115, 11280-11285. [本文引用: 5]
R?schV, TscharntkeT, ScherberC, BatáryP ( 2015). Biodiversity conservation across taxa and landscapes requires many small as well as single large habitat fragments Oecologia, 179, 209-222. [本文引用: 1]
RuffellJ, Banks-LeiteC, DidhamRK ( 2016). Accounting for the causal basis of collinearity when measuring the effects of habitat loss versus habitat fragmentation Oikos, 125, 117-125. [本文引用: 1]
RyserR, Ha?usslerJ, StarkM, BroseU, RallBC, GuillC ( 2019). The biggest losers: Habitat isolation deconstructs complex food webs from top to bottom Proceedings of the Royal Society B: Biological Sciences, 286, 20191177. DOI: 10.1098/rspb.2019.1177. [本文引用: 1]
SantosAMC, FieldR, RicklefsRE ( 2016). New directions in island biogeography Global Ecology and Biogeography, 25, 751-768. [本文引用: 4]
SaundersDA, HobbsRJ, MargulesCR ( 1991). Biological consequences of ecosystem fragmentation: A review Conservation Biology, 5, 18-32. [本文引用: 1]
SeiboldS, CadotteMW, MacIvorJS, ThornS, Mu?llerJ ( 2018). The necessity of multitrophic approaches in community ecology Trends in Ecology & Evoluation, 33, 754-764. [本文引用: 1]
TerborghJ, Alvarez-LoayzaP, DexterK, CornejoF, CarrascoC ( 2011). Decomposing dispersal limitation: Limits on fecundity or seed distribution? Journal of Ecology, 99, 935-944. [本文引用: 1]
TewksburyJJ, LeveyDJ, HaddadNM, SargentS, OrrockJL, WeldonA, DanielsonBJ, BrinkerhoffJ, DamschenEI, TownsendP ( 2002). Corridors affect plants, animals, and their interactions in fragmented landscapes Proceedings of the National Academy of Sciences of the United States of America, 99, 12923-12926. [本文引用: 3]
TilmanD ( 1982). Resource competition and community structure Monographs in Population Biology, 17, 1-296 [本文引用: 1]
TscharntkeT, Steffan-DewenterI, KruessA, ThiesC ( 2002). Contribution of small habitat fragments to conservation of insect communities of grassland-cropland landscapes Ecological Applications, 12, 354-363. [本文引用: 1]
UlrichW, KusumotoB, ShionoT, KubotaY ( 2016). Climatic and geographic correlates of global forest tree species-?abundance distributions and community evenness Journal of Vegetation Science, 27, 295-305. [本文引用: 1]
UlrichW, OllikM, UglandKI ( 2010). A meta-analysis of ?species- abundance distributions Oikos, 119, 1149-1155. [本文引用: 1]
VellendM ( 2010). Conceptual synthesis in community ecology The Quarterly Review of Biology, 85, 183-206. [本文引用: 5]
VellendM ( 2016). The Theory of Ecological Communities. Princeton University Press, Princeton. [本文引用: 15]
ViolleC, NavasML, VileD, KazakouE, FortunelC, HummelI, GarnierE ( 2007). Let the concept of trait be functional! Oikos, 116, 882-892. [本文引用: 1]
WalkerLR, WardleDA, BardgettRD, ClarksonBD ( 2010). The use of chronosequences in studies of ecological succession and soil development Journal of Ecology, 98, 725-736. [本文引用: 1]
WallerDM, MudrakEL, RogersDA ( 2018). Do metacommunity mass effects predict changes in species incidence and abundance? Ecography, 41, 11-23. [本文引用: 2]
WardleDA, H?rnbergG, ZackrissonO, Kalela-BrundinM, CoomesDA ( 2003). Long-term effects of wildfire on ecosystem properties across an island area gradient Science, 300, 972-975. [本文引用: 1]
WardleDA, JonssonM, BansalS, BardgettRD, GundaleMJ, MetcalfeDB ( 2012). Linking vegetation change, carbon sequestration and biodiversity: Insights from island ecosystems in a long-term natural experiment Journal of Ecology, 100, 16-30. [本文引用: 1]
WarrenBH, SimberloffD, RicklefsRE, AguiléeR, CondamineFL, GravelD, MorlonH, MouquetN, RosindellJ, CasquetJ, ContiE, CornuaultJ, Fernández-PalaciosJM, HenglT, NorderSJ, RijsdijkKF, SanmartínI, StrasbergD, TriantisKA, ValenteLM, WhittakerRJ, GillespieRG, EmersonBC, ThébaudC ( 2015). Islands as model systems in ecology and evolution: Prospects fifty years after Macarthur- Wilson Ecology Letters, 18, 200-217. [本文引用: 1]
WebbCO, AckerlyDD, McPeekMA, DonoghueMJ ( 2002). Phylogenies and community ecology Annual Review of Ecology and Systematics, 33, 475-505. [本文引用: 1]
WhittakerRJ, JonesSH, PartomihardjoT ( 1997). The rebuilding of an isolated rain forestassemblage: How disharmonicis the flora of Krakatau? Biodiversity and Conservation, 6, 1671-1696. [本文引用: 1]
WiensJJ, DonoghueMJ ( 2004). Historical biogeography, ecology and species richness Trends in Ecology & Evolution, 19, 639-644. [本文引用: 2]
WilcoveDS, McLellanCH, DobsonAP ( 1986). Habitat fragmentation in the temperate zone Conservation Biology, 6, 237-256. [本文引用: 1]
WilsonEO, WillisEO ( 1975). Applied biogeography In: Cody ML, Diamond JM eds. Ecology and Evolution of Communities. Harvard University Press, Cambridge, USA. 522-534. [本文引用: 1]
WintleBA, KujalaH, WhiteheadA, CameronA, VelozS, KukkalaA, MoilanenA, GordonA, LentiniPE, CadenheadNCR, BekessySA ( 2019). Global synthesis of conservation studies reveals the importance of small habitat patches for biodiversity Proceedings of the National Academy of Sciences of the United States of America, 116, 909-914. [本文引用: 1]
WrightS ( 1940). Breeding structure of populations in relation to speciation The American Naturalist, 74, 232-248. [本文引用: 1]
WuCP, VellendM, YuanWG, JiangB, LiuJJ, ShenAH, LiuJL, ZhuJR, YuMJ ( 2017a). Patterns and determinants of plant biodiversity in non-commercial forests of eastern China PLOS ONE, 12, e0188409. DOI: 10.1371/journal.pone.0188409. [本文引用: 1]
WuH, FranklinSB, LiuJM, LuZJ ( 2017b). Relative importance of density dependence and topography on tree mortality in a subtropical mountain forest Forest Ecology and Management, 384, 169-179. [本文引用: 1]
YuMJ, HuG, FeeleyKJ, WuJG, DingP ( 2012). Richness and composition of plants and birds on land-bridge islands: Effects of island attributes and differential responses of species groups Journal of Biogeography, 39, 1124-1133. [本文引用: 3]
ZambranoJ, Garzon-LopezCX, YeagerL, FortunelC, CordeiroNJ, BeckmanNG ( 2019). The effects of habitat loss and fragmentation on plant functional traits and functional diversity: What do we know so far? Oecologia, 191, 505-518. [本文引用: 3]
ZavaletaE, PasariJ, MooreJ, HernándezD, SuttleKB, WilmersCC ( 2009). Ecosystem responses to community disassembly Annals of the New York Academy of Sciences, 1162, 311-333. [本文引用: 1]
ZhuY, MiXC, MaKP ( 2009). A mechanism of plant species coexistence: The negative density-dependent hypothesis Biodiversity Science, 17, 594-604. [本文引用: 2]
ZhuY, QueenboroughSA, ConditR, HubbellSP, MaKP, ComitaLS ( 2018). Density-dependent survival varies with species life-history strategy in a tropical forest Ecology Letters, 21, 506-515. [本文引用: 1]
A trait-based approach to community assembly: Partitioning of species trait values into within- and among-community components 1 2007
... 植物功能特征/功能性状指植物的生理、结构和行为特征, 能够指示物种对环境的适应能力, 影响物种在特定生境中的定殖和种间关系(Violle et al., 2007; Lindo et al., 2012).在过去近20年的研究中, 越来越多的生态学家将物种的功能特征与选择过程结合起来, 通过分析群落内物种功能特征的收敛或者发散, 探讨环境过滤或竞争作用引起的限制相似性在群落构建过程中的相对作用(Perronne et al., 2017).当环境过滤影响群落内的物种组成时, 如果具有某一类功能特征的物种更能适应环境而被保留, 此时群落内功能特征的分布表现为收敛(群落内物种间的功能特征更为相似).在小尺度上, 物种间功能特征属性越相似, 种间竞争将越激烈, 竞争排斥作用可能仅保留具有强竞争力的性状, 使得功能特征仍表现为收敛; 但当物种能够共存时, 此时限制相似性的作用(如群落内物种间的竞争, 天敌介导的密度依赖等过程)会致使功能特征值具有更宽的范围, 引起性状发散(非相似性功能特征), 即生态位分化(Kraft et al., 2008).目前, 已有多种量度群落功能特征的α多样性和β多样性的指数, 可以反映群落内物种功能性状的分布和多样性(Ackerly & Cornwell, 2007; Mouchet et al., 2010).因此, 基于群落内实际观察到的性状分布格局, 依赖于性状分布的零模型(随机过程, 即所有个体功能相同且物种随机共存), 可以衡量群落内物种功能特征是收敛的还是发散的, 进而探讨选择过程(环境过滤和种间竞争等)在群落构建中的作用(Kraft et al., 2008). ...
Metapopulations and metacommunities: Combining spatial and temporal perspectives in plant ecology 1 2012
... 野外控制实验也是检验片段化生境中群落构建过程非常有效的手段(Debinski & Holt, 2000; Resasco et al., 2017).Haddad等(2015)综述了目前世界上已有的针对片段化生境的控制实验(图1), 通过设置控制实验和长期观察, 对片段化生境中或岛屿上群落构建的过程进行了有效的检验(Wilson et al., 2016; Resasco et al., 2017).例如, 巴西的片段化森林动态实验是目前最大和时间最长的野外控制实验, 通过设置不同大小的森林斑块且控制斑块之间的隔离度差异, 分析森林斑块面积的大小对群落构建过程的影响(Laurance et al., 2011); 美国的Savannah河流廊道实验通过增加斑块间的隔离度, 且控制斑块的面积差异, 分析廊道效应(扩散)在群落构建过程中的作用(Haddad & Baum, 1999; Tewksbury et al., 2002); Kansas生境片段化实验则通过设置不同面积大小和隔离度的片段化斑块, 长期监测斑块内群落的动态变化, 研究生境片段化对群落构建和演替过程的影响(Holt et al., 1995); 另外, Alexander等(2012)利用此实验在种群水平上分析了种子扩散与斑块间连通性之间的关系, 在群落水平上分析了环境过滤和扩散限制的共同作用对群落演替的作用.然而, 除上述主要的实验外, 基于野外控制实验检验片段化生境中森林群落构建过程的研究仍然不足. ...
Big answers from small worlds: A user?s guide for protist microcosms as a model system in ecology and evolution 1 2015
... 此外, 结合微宇宙的实验方法, 也可以构建模拟群落, 通过直接观察不同实验情景下的群落动态, 能够验证某一生态过程在群落构建中的作用(Davies et al., 2009; Altermatt et al., 2015; Ron et al., 2018).通常利用微生物群落, 通过配置菌落的大小(生态漂变)、菌群间隔离度(扩散)、环境胁迫或干扰(选择)、起始菌落的物种组成(优先效应)等, 验证生态群落的构建过程(Jiang & Patel, 2008; Ojima & Jiang, 2017).然而, 对于森林群落而言, 很难利用微宇宙的实验方法对群落构建过程进行检验, 由微宇宙实验获得的结论是否适用于片段化生境中的森林群落, 仍值得商榷. ...
生境破碎化对食果动物及种子传播的影响 1 2011
... 关于选择过程对植物群落构建的影响, 主要集中于局域尺度上基于生态位理论的研究, 即非生物环境因子的筛选和种内/种间的竞争作用(Tilman, 1982; Chase & Leibold, 2003; Letten et al., 2017).植物群落中, 选择过程产生于群落内不同物种个体间适合度的差异(Vellend, 2010).群落构建过程中, 一般存在3种形式的选择过程: (1)固定选择效应; (2)密度依赖效应; (3)时空选择效应.固定选择效应是指在特定的环境条件下, 如果群落内物种的相对适合度在时间和空间上固定不变, 那么群落内具有最高适合度的物种将会排除其他所有的物种(Vellend, 2010).密度依赖效应取决于群落内物种间定性(例如竞争、捕食、互利共生和疾病等)和定量的生态关系, 即所研究物种的适合度依赖于群落内同种和异种的个体密度, 包括负密度依赖效应(或被称为负密度制约效应)(祝燕等, 2009)和正密度依赖效应两个方面(Vellend, 2016).负密度依赖效应的研究内容主要包括: (1)密度依赖的自疏过程(随机死亡假说), 即同种个体的密度与死亡率呈现正相关关系(Kenkel, 1988); (2)距离依赖的死亡过程(Janzen-Connell假说), 即母树周围同种个体具有较高的密度时, 越靠近母树的同种个体死亡率越高(Janzen, 1970; Hubbell et al., 1990; Hyatt et al., 2003); (3)群落补偿趋势假说, 即群落越大, 负密度依赖作用越强, 越有利于稀有种在群落内的存活(Connell et al., 1984).负密度依赖效应对群落内的低密度物种更加有利, 故而可以使群落内物种维持稳定的共存状态(祝燕等, 2009).在片段化生境中, 不管是固定选择效应还是密度依赖效应都可能在时间和空间上存在差异, 即时空选择效应, 该效应也会潜在地影响群落动态.此外, 生境片段化限制了植物种子传播/扩散(Terborgh et al., 2011)以及特异性植食动物组成和数量的改变(Valladares et al., 2006; Emer et al., 2018), 进而影响种子或幼苗被植食的概率(白冰等, 2011; Genua et al., 2017; 骆杨青等, 2017).生境片段化所产生的边缘效应也能改变植物与天敌之间的关系(Krishnadas et al., 2018).生境片段化所引起的种间相互作用的改变, 最终会导致植物个体在空间分布格局上的差异, 影响密度效应的选择强度. ...
生境破碎化对食果动物及种子传播的影响 1 2011
... 关于选择过程对植物群落构建的影响, 主要集中于局域尺度上基于生态位理论的研究, 即非生物环境因子的筛选和种内/种间的竞争作用(Tilman, 1982; Chase & Leibold, 2003; Letten et al., 2017).植物群落中, 选择过程产生于群落内不同物种个体间适合度的差异(Vellend, 2010).群落构建过程中, 一般存在3种形式的选择过程: (1)固定选择效应; (2)密度依赖效应; (3)时空选择效应.固定选择效应是指在特定的环境条件下, 如果群落内物种的相对适合度在时间和空间上固定不变, 那么群落内具有最高适合度的物种将会排除其他所有的物种(Vellend, 2010).密度依赖效应取决于群落内物种间定性(例如竞争、捕食、互利共生和疾病等)和定量的生态关系, 即所研究物种的适合度依赖于群落内同种和异种的个体密度, 包括负密度依赖效应(或被称为负密度制约效应)(祝燕等, 2009)和正密度依赖效应两个方面(Vellend, 2016).负密度依赖效应的研究内容主要包括: (1)密度依赖的自疏过程(随机死亡假说), 即同种个体的密度与死亡率呈现正相关关系(Kenkel, 1988); (2)距离依赖的死亡过程(Janzen-Connell假说), 即母树周围同种个体具有较高的密度时, 越靠近母树的同种个体死亡率越高(Janzen, 1970; Hubbell et al., 1990; Hyatt et al., 2003); (3)群落补偿趋势假说, 即群落越大, 负密度依赖作用越强, 越有利于稀有种在群落内的存活(Connell et al., 1984).负密度依赖效应对群落内的低密度物种更加有利, 故而可以使群落内物种维持稳定的共存状态(祝燕等, 2009).在片段化生境中, 不管是固定选择效应还是密度依赖效应都可能在时间和空间上存在差异, 即时空选择效应, 该效应也会潜在地影响群落动态.此外, 生境片段化限制了植物种子传播/扩散(Terborgh et al., 2011)以及特异性植食动物组成和数量的改变(Valladares et al., 2006; Emer et al., 2018), 进而影响种子或幼苗被植食的概率(白冰等, 2011; Genua et al., 2017; 骆杨青等, 2017).生境片段化所产生的边缘效应也能改变植物与天敌之间的关系(Krishnadas et al., 2018).生境片段化所引起的种间相互作用的改变, 最终会导致植物个体在空间分布格局上的差异, 影响密度效应的选择强度. ...
Phylogenetic turnover along local environmental gradients in tropical forest communities 4 2016
... 不论控制实验还是野外观察实验, 根据被片段化前后森林所处的状态以及片段化后森林的演替状态, 可以把片段化森林分为两大群落类型: 解构群落和重构群落(Wilson et al., 2016; Collins et al., 2017).解构群落指处于演替后期的老龄林或成熟林被片段化后形成的森林群落, 出现逆行演替现象, 在演替过程中伴随着物种减少, 如中国西双版纳和巴西亚马孙的热带雨林中形成的片段化森林(Liu & Slik, 2014).重构群落是指在片段化生境中通过进展演替而重新构建形成的次生林群落, 处于进展演替过程之中, 一般处于演替的前期阶段, 群落内的优势种主要为一些阳性、生长快、扩散能力强的先锋树种(Baldeck et al., 2016; Liu et al., 2019b).解构群落和重构群落中存在的定殖和灭绝过程将促使群落内的物种组成和多样性发生改变, 然而受选择、生态漂变等过程的影响, 重构群落中定殖的物种组成和数量与解构群落中易于灭绝的物种组成和数量存在差异(Zavaleta et al., 2009).已有研究证实, 片段化后植物群落的长期动态变化, 如物种β多样性的变化, 将受到群落起始状态的影响(Collins et al., 2017), 即优先效应的作用(Fukami, 2015).因此, 依赖于森林群落起始状态的不同, 生境片段化对群落构建过程的影响可能也会不同(图3), 但以往的研究通常忽视了群落的起始状态.已有研究主要关注原来连续的老龄林或成熟林被片段化后形成的解构群落, 对片段生境中重构群落的构建过程关注较少(Debinski & Holt, 2000; Wilson et al., 2016; Liu et al., 2019b). ...
... 迄今关于片段化生境中重构森林群落的研究仍然十分缺乏.我们对片段化生境中森林的重新构建过程可以作如下预测: 1)重构群落演替的起始阶段, 空间的隔离作用将比斑块面积的大小更加重要, 因短期的干扰或生境质量的改变, 空间的隔离将阻止物种占用适合的生境, 导致种子传播或者强风等随机事件决定了物种的抵达和在斑块上的定殖(Walker et al., 2010).例如, Kadmon和Pulliam (1995)发现在新近皆伐的湖泊岛屿上空间的隔离增加了风力传播物种的比例, Whittaker等(1997)发现在新近形成的火山岛屿上, 物种早期阶段的定殖可以根据其扩散性状进行预测, 不过在具有近60年片段化历史的千岛湖, 岛屿面积是影响植物群落物种组成的主要因素, 而非空间的隔离(Yu et al., 2012; Baldeck et al., 2016).2)当某一物种扩散到某一斑块后, 斑块内已有的种子/幼苗库, 会对后续扩散来的物种的定殖产生影响, 即优先效应, 该效应与决定性过程和随机过程会共同影响片段化生境中群落的构建过程(Fukami, 2015).3)物种抵达相应的斑块之后, 斑块内的生境异质性、种间/种内关系和生态漂变等过程将共同决定物种定殖成功的概率和物种组成, 如筛选个体生长快、木质密度低以及喜阳的先锋物种(Baldeck et al., 2016; Liu et al., 2019b, 2020), 或具有菌根的植物物种等(Delavaux et al., 2019).Baynes等(2016)通过比较砍伐后重构的片段化森林群落与连续森林群落内的物种多样性, 发现重构的森林群落中反而具有更多的稀有种以及较高的β多样性, 生态漂变过程也会影响片段化森林群落的重构. ...
... ).3)物种抵达相应的斑块之后, 斑块内的生境异质性、种间/种内关系和生态漂变等过程将共同决定物种定殖成功的概率和物种组成, 如筛选个体生长快、木质密度低以及喜阳的先锋物种(Baldeck et al., 2016; Liu et al., 2019b, 2020), 或具有菌根的植物物种等(Delavaux et al., 2019).Baynes等(2016)通过比较砍伐后重构的片段化森林群落与连续森林群落内的物种多样性, 发现重构的森林群落中反而具有更多的稀有种以及较高的β多样性, 生态漂变过程也会影响片段化森林群落的重构. ...
... 此外, 利用空间代替时间的方法(Wardle et al., 2012), 如选择不同年龄的岛屿群、不同群落演替历史岛屿、不同干扰梯度的森林斑块等, 通过分析不同群落间物种组成、功能特征和谱系结构的变化, 也是检验群落构建过程的方法之一(Wardle et al., 2003; Rominger et al., 2016).对于森林群落而言, 群落内幼苗和成树个体的多度和物种组成的变化, 也能够反映生态漂变或者选择过程在由幼苗发展到成树阶段的相对作用(Baldeck et al., 2016). ...
Effects of fragmentation and landscape variation on tree diversity in post-logging regrowth forests of the Southern Philippines 1 2016
... 迄今关于片段化生境中重构森林群落的研究仍然十分缺乏.我们对片段化生境中森林的重新构建过程可以作如下预测: 1)重构群落演替的起始阶段, 空间的隔离作用将比斑块面积的大小更加重要, 因短期的干扰或生境质量的改变, 空间的隔离将阻止物种占用适合的生境, 导致种子传播或者强风等随机事件决定了物种的抵达和在斑块上的定殖(Walker et al., 2010).例如, Kadmon和Pulliam (1995)发现在新近皆伐的湖泊岛屿上空间的隔离增加了风力传播物种的比例, Whittaker等(1997)发现在新近形成的火山岛屿上, 物种早期阶段的定殖可以根据其扩散性状进行预测, 不过在具有近60年片段化历史的千岛湖, 岛屿面积是影响植物群落物种组成的主要因素, 而非空间的隔离(Yu et al., 2012; Baldeck et al., 2016).2)当某一物种扩散到某一斑块后, 斑块内已有的种子/幼苗库, 会对后续扩散来的物种的定殖产生影响, 即优先效应, 该效应与决定性过程和随机过程会共同影响片段化生境中群落的构建过程(Fukami, 2015).3)物种抵达相应的斑块之后, 斑块内的生境异质性、种间/种内关系和生态漂变等过程将共同决定物种定殖成功的概率和物种组成, 如筛选个体生长快、木质密度低以及喜阳的先锋物种(Baldeck et al., 2016; Liu et al., 2019b, 2020), 或具有菌根的植物物种等(Delavaux et al., 2019).Baynes等(2016)通过比较砍伐后重构的片段化森林群落与连续森林群落内的物种多样性, 发现重构的森林群落中反而具有更多的稀有种以及较高的β多样性, 生态漂变过程也会影响片段化森林群落的重构. ...
Edge-mediated compositional and functional decay of tree assemblages in Amazonian forest islands after 26 years of isolation 2 2015
... 生境片段化(或破碎化, habitat fragmentation)是指原来连续的生境受自然和人为干扰的影响, 逐渐转变为隔离的且面积较小的斑块(Wilcove et al., 1986; Fahrig, 2003).人类干扰(如择伐、都市化、公路和水库建设等)对自然生态系统的破坏以及自然因素(如森林火灾、岛屿等)的影响, 造成全球森林植被呈片段化分布(Fahrig, 2003; Didham, 2010; Haddad et al., 2015; Taubert et al., 2018; Liu et al., 2019a).从全球森林的分布来看, 世界上大部分森林斑块的面积小于10 hm2 (Haddad et al., 2015).森林面积的丧失、隔离度的增加和边缘效应的加剧将显著影响片段化生境中森林群落的结构(Laurance et al., 2006)和演替动态(Benchimol & Peres, 2015; Liu et al., 2019b). ...
... 关于对片段化生境中解构森林群落的研究, 主要集中在生物多样性丰富的热带地区, 比如亚马孙热带雨林(Ewers et al., 2017)和西双版纳热带雨林(Liu & Slik, 2014)等地区.这些森林之前均为连续森林, 随着人类活动, 如砍伐、农业种植、都市化等因素的影响, 逐渐被分割成具有不同隔离度和面积的陆地“森林生境岛屿”.受到边缘效应的影响, 面积较小的“生境岛屿”出现光照强度增加、水分限制加剧和随机干扰事件(如火干扰)频率增加等现象, 将影响不同斑块中物种的组成(Saunders et al., 1991; Ewers et al., 2017).环境过滤所引起的非生物选择过程是影响解构群落的主要生态过程, 该过程所引起的物种功能属性的改变和群落结构的变化在片段化后的10-15年尤为明显(Laurance et al., 2014).Das等(2017)对解构森林群落结构和物种组成变化的进一步研究表明, 集合群落中的物种筛选模型(即生物和非生物因子的选择过程)主要影响群落的结构和周转, 而空间连通性(即扩散限制)对群落构建的影响较小.在亚马孙和亚洲东南部片段化森林的研究中也发现, 边缘效应对森林群落动态具有重要的影响(Laurance, 2008; Laurance et al., 2011; Ruffell et al., 2016), 该过程更易导致大树的死亡从而令更多的先锋物种占据空出的生态位(Laurance et al., 2000; Benchimol & Peres, 2015), 故而可以认为这些成熟林群落片段化后所经历的演替过程为“逆行演替”.虽然经历逆行演替, 但是这些森林群落, 尤其是处于边缘的森林群落, 受不同干扰条件的影响, 在经过长期的演替后并不一定会处于演替早期时的群落状态(Ewers et al., 2017). ...
1 2003
... 关于选择过程对植物群落构建的影响, 主要集中于局域尺度上基于生态位理论的研究, 即非生物环境因子的筛选和种内/种间的竞争作用(Tilman, 1982; Chase & Leibold, 2003; Letten et al., 2017).植物群落中, 选择过程产生于群落内不同物种个体间适合度的差异(Vellend, 2010).群落构建过程中, 一般存在3种形式的选择过程: (1)固定选择效应; (2)密度依赖效应; (3)时空选择效应.固定选择效应是指在特定的环境条件下, 如果群落内物种的相对适合度在时间和空间上固定不变, 那么群落内具有最高适合度的物种将会排除其他所有的物种(Vellend, 2010).密度依赖效应取决于群落内物种间定性(例如竞争、捕食、互利共生和疾病等)和定量的生态关系, 即所研究物种的适合度依赖于群落内同种和异种的个体密度, 包括负密度依赖效应(或被称为负密度制约效应)(祝燕等, 2009)和正密度依赖效应两个方面(Vellend, 2016).负密度依赖效应的研究内容主要包括: (1)密度依赖的自疏过程(随机死亡假说), 即同种个体的密度与死亡率呈现正相关关系(Kenkel, 1988); (2)距离依赖的死亡过程(Janzen-Connell假说), 即母树周围同种个体具有较高的密度时, 越靠近母树的同种个体死亡率越高(Janzen, 1970; Hubbell et al., 1990; Hyatt et al., 2003); (3)群落补偿趋势假说, 即群落越大, 负密度依赖作用越强, 越有利于稀有种在群落内的存活(Connell et al., 1984).负密度依赖效应对群落内的低密度物种更加有利, 故而可以使群落内物种维持稳定的共存状态(祝燕等, 2009).在片段化生境中, 不管是固定选择效应还是密度依赖效应都可能在时间和空间上存在差异, 即时空选择效应, 该效应也会潜在地影响群落动态.此外, 生境片段化限制了植物种子传播/扩散(Terborgh et al., 2011)以及特异性植食动物组成和数量的改变(Valladares et al., 2006; Emer et al., 2018), 进而影响种子或幼苗被植食的概率(白冰等, 2011; Genua et al., 2017; 骆杨青等, 2017).生境片段化所产生的边缘效应也能改变植物与天敌之间的关系(Krishnadas et al., 2018).生境片段化所引起的种间相互作用的改变, 最终会导致植物个体在空间分布格局上的差异, 影响密度效应的选择强度. ...
Disentangling the importance of ecological niches from stochastic processes across scales 2 2011
Ecological Consequences of Habitat Fragmentation 1 2010
... 生境片段化(或破碎化, habitat fragmentation)是指原来连续的生境受自然和人为干扰的影响, 逐渐转变为隔离的且面积较小的斑块(Wilcove et al., 1986; Fahrig, 2003).人类干扰(如择伐、都市化、公路和水库建设等)对自然生态系统的破坏以及自然因素(如森林火灾、岛屿等)的影响, 造成全球森林植被呈片段化分布(Fahrig, 2003; Didham, 2010; Haddad et al., 2015; Taubert et al., 2018; Liu et al., 2019a).从全球森林的分布来看, 世界上大部分森林斑块的面积小于10 hm2 (Haddad et al., 2015).森林面积的丧失、隔离度的增加和边缘效应的加剧将显著影响片段化生境中森林群落的结构(Laurance et al., 2006)和演替动态(Benchimol & Peres, 2015; Liu et al., 2019b). ...
Seed-dispersal interactions in fragmented landscapes—A metanetwork approach 3 2018
... 关于选择过程对植物群落构建的影响, 主要集中于局域尺度上基于生态位理论的研究, 即非生物环境因子的筛选和种内/种间的竞争作用(Tilman, 1982; Chase & Leibold, 2003; Letten et al., 2017).植物群落中, 选择过程产生于群落内不同物种个体间适合度的差异(Vellend, 2010).群落构建过程中, 一般存在3种形式的选择过程: (1)固定选择效应; (2)密度依赖效应; (3)时空选择效应.固定选择效应是指在特定的环境条件下, 如果群落内物种的相对适合度在时间和空间上固定不变, 那么群落内具有最高适合度的物种将会排除其他所有的物种(Vellend, 2010).密度依赖效应取决于群落内物种间定性(例如竞争、捕食、互利共生和疾病等)和定量的生态关系, 即所研究物种的适合度依赖于群落内同种和异种的个体密度, 包括负密度依赖效应(或被称为负密度制约效应)(祝燕等, 2009)和正密度依赖效应两个方面(Vellend, 2016).负密度依赖效应的研究内容主要包括: (1)密度依赖的自疏过程(随机死亡假说), 即同种个体的密度与死亡率呈现正相关关系(Kenkel, 1988); (2)距离依赖的死亡过程(Janzen-Connell假说), 即母树周围同种个体具有较高的密度时, 越靠近母树的同种个体死亡率越高(Janzen, 1970; Hubbell et al., 1990; Hyatt et al., 2003); (3)群落补偿趋势假说, 即群落越大, 负密度依赖作用越强, 越有利于稀有种在群落内的存活(Connell et al., 1984).负密度依赖效应对群落内的低密度物种更加有利, 故而可以使群落内物种维持稳定的共存状态(祝燕等, 2009).在片段化生境中, 不管是固定选择效应还是密度依赖效应都可能在时间和空间上存在差异, 即时空选择效应, 该效应也会潜在地影响群落动态.此外, 生境片段化限制了植物种子传播/扩散(Terborgh et al., 2011)以及特异性植食动物组成和数量的改变(Valladares et al., 2006; Emer et al., 2018), 进而影响种子或幼苗被植食的概率(白冰等, 2011; Genua et al., 2017; 骆杨青等, 2017).生境片段化所产生的边缘效应也能改变植物与天敌之间的关系(Krishnadas et al., 2018).生境片段化所引起的种间相互作用的改变, 最终会导致植物个体在空间分布格局上的差异, 影响密度效应的选择强度. ...
... 以往的理论/假说(如ETIB、集合群落理论和群落生态学理论)主要针对单一营养级或两个营养级上的生态过程进行分析, 如单一营养级上的种间相互作用及其与非生物因子间的关系(选择过程).近年来, 在ETIB理论的基础上, 许多研究开始逐渐关注岛屿生物地理学的营养理论, 即将营养级间物种的相互作用对定殖率和灭绝率的影响考虑其中(Gravel et al., 2011).在片段化生境中, 食物网各营养级间关联物种的次生灭绝是非常普遍的, 越顶级的消费者越容易消失(Ryser et al., 2019), 消费者缺失所引发的下行调控丧失效应通过影响传粉、种子传播和植食等与扩散和选择相关的生态过程, 从而影响片段化森林群落的构建过程(Valladares et al., 2006; Larios et al., 2017; Emer et al., 2018).Cirtwill和Stouffer (2016)发现, 当考虑岛屿上的营养级结构时, 如果包含上行效应的影响, 能够改进ETIB中对迁移率和灭绝率的预测.但在片段化森林景观中, 针对多营养级上的种间相互作用(如下行效应)在森林群落构建过程的作用研究, 仍鲜有报道. ...
... 随着群落大小的变小, 生态漂变作用的增大, 会引起物种在局域群落内的消失(Fukami, 2010).扩散过程可以通过增加有效群落的大小, 来减少生态漂变的作用.因此, 在片段化生境中, 促进斑块间物种的扩散过程, 可以增加有效群落的大小, 减少生态漂变的作用(Ron et al., 2018).此外, 增加斑块间的连通性, 也能促进植食性昆虫等天敌和种子扩散者在片段化森林之间的扩散, 稳定食物链结构、避免物种之间级联效应的灭绝过程, 增加同种负密度制约效应的强度等, 进而提高生物多样性(Emer et al., 2018; Seibold et al., 2018).故而, 增加小面积森林斑块之间的连通性, 降低生态漂变的影响, 也是维持生物多样性的有效手段. ...
Zero sum, the niche, and metacommunities: Long-term dynamics of community assembly 1 2008
... 所有自然种群多度的波动, 均受到环境的随机波动、种内和种间竞争以及随机过程等因子影响(Mutshinda et al., 2009).而群落的构建依赖于群落内各种群的动态变化, 即物种多度、物种定殖和灭绝的动态变化, 通过分析群落内物种组成或种群多度随时间的动态变化, 可以为群落的构建过程提供最直接的证据(McGarigal & Cushman, 2002; Ernest et al., 2008).Laurance等(2014)通过对比片段化和连续森林中年际间树木的死亡率和更新率, 发现环境过滤(如大气中CO2浓度)是影响群落构建的主要因子; Damschen等(2019)通过长期监测植物群落的定殖率和死亡率, 发现连接度高的片段化群落间能够通过提高定殖率和降低灭绝率而维持更高的物种多样性.另外, 通过比较分析一定时间内片段化森林与连续森林内物种组成/功能特征的周转、群落功能特征/谱系结构的收敛或发散, 也能揭示各生态过程在森林演替不同阶段的相对作用(Collins et al., 2017).利用不同大小或隔离度的群落内各种群的多度在时间上的波动趋势, 也可以分析生态漂变和选择过程的相对作用(Vellend, 2016). ...
Predicted trajectories of tree community change in Amazonian rainforest fragments 3 2017
... 关于对片段化生境中解构森林群落的研究, 主要集中在生物多样性丰富的热带地区, 比如亚马孙热带雨林(Ewers et al., 2017)和西双版纳热带雨林(Liu & Slik, 2014)等地区.这些森林之前均为连续森林, 随着人类活动, 如砍伐、农业种植、都市化等因素的影响, 逐渐被分割成具有不同隔离度和面积的陆地“森林生境岛屿”.受到边缘效应的影响, 面积较小的“生境岛屿”出现光照强度增加、水分限制加剧和随机干扰事件(如火干扰)频率增加等现象, 将影响不同斑块中物种的组成(Saunders et al., 1991; Ewers et al., 2017).环境过滤所引起的非生物选择过程是影响解构群落的主要生态过程, 该过程所引起的物种功能属性的改变和群落结构的变化在片段化后的10-15年尤为明显(Laurance et al., 2014).Das等(2017)对解构森林群落结构和物种组成变化的进一步研究表明, 集合群落中的物种筛选模型(即生物和非生物因子的选择过程)主要影响群落的结构和周转, 而空间连通性(即扩散限制)对群落构建的影响较小.在亚马孙和亚洲东南部片段化森林的研究中也发现, 边缘效应对森林群落动态具有重要的影响(Laurance, 2008; Laurance et al., 2011; Ruffell et al., 2016), 该过程更易导致大树的死亡从而令更多的先锋物种占据空出的生态位(Laurance et al., 2000; Benchimol & Peres, 2015), 故而可以认为这些成熟林群落片段化后所经历的演替过程为“逆行演替”.虽然经历逆行演替, 但是这些森林群落, 尤其是处于边缘的森林群落, 受不同干扰条件的影响, 在经过长期的演替后并不一定会处于演替早期时的群落状态(Ewers et al., 2017). ...
... ; Ewers et al., 2017).环境过滤所引起的非生物选择过程是影响解构群落的主要生态过程, 该过程所引起的物种功能属性的改变和群落结构的变化在片段化后的10-15年尤为明显(Laurance et al., 2014).Das等(2017)对解构森林群落结构和物种组成变化的进一步研究表明, 集合群落中的物种筛选模型(即生物和非生物因子的选择过程)主要影响群落的结构和周转, 而空间连通性(即扩散限制)对群落构建的影响较小.在亚马孙和亚洲东南部片段化森林的研究中也发现, 边缘效应对森林群落动态具有重要的影响(Laurance, 2008; Laurance et al., 2011; Ruffell et al., 2016), 该过程更易导致大树的死亡从而令更多的先锋物种占据空出的生态位(Laurance et al., 2000; Benchimol & Peres, 2015), 故而可以认为这些成熟林群落片段化后所经历的演替过程为“逆行演替”.虽然经历逆行演替, 但是这些森林群落, 尤其是处于边缘的森林群落, 受不同干扰条件的影响, 在经过长期的演替后并不一定会处于演替早期时的群落状态(Ewers et al., 2017). ...
Conceptual cartoon illustrating processes in forest community assembly in fragmented habitats. Redrawn from <xref ref-type="bibr" rid="b46">HilleRisLambers <i>et al</i>. (2012)</xref> and <xref ref-type="bibr" rid="b133">Vellend (2016)</xref>. Different letters denote different species; circles of different sizes denote different size patches; different fan-shaped areas in circles denote the relative abundance of each species in one community.Fig. 21.1 扩散 扩散是有机体或其繁殖体离开出生地后的迁移或传播, 该过程对群落构建的影响主要依赖于扩散者产生群落(源群落)和抵达群落(汇群落)的大小和物种组成(Holyoak et al., 2005).与连续森林相比, 存在隔离的森林斑块将限制物种在斑块间的自由扩散(如种子传播), 即群落间物种的扩散限制.根据源群落和汇群落的相对大小, 片段化生境中物种的扩散可以分为两种情形: 一是大陆-岛屿模型, 该模型中将扩散假定为单向扩散, 由无限大的源群落(大陆或大面积斑块)向一个或多个小的、隔离的汇群落(岛屿或小面积斑块)中扩散, 源群落的物种组成相对恒定, 而汇群落的物种组成变化则高于源群落; 二是岛屿模型, 该模型中的群落(岛屿或小面积斑块)通过扩散相互联系在一起, 形成集合群落(Holyoak et al., 2005).一般而言, 扩散过程对群落构建的作用是受其他生态过程影响的, 尤其是选择过程和生态漂变(Ron et al., 2018). ...
... ); The Stability of Altered Forest Ecosystems (SAFE) Project (Malaysia)(Ewers et al., 2011); Metatron experiment (France)(Legrand et al., 2012). Fig. 1 10.17521/cjpe.2019.0155.F002
Conceptual cartoon illustrating processes in forest community assembly in fragmented habitats. Redrawn from <xref ref-type="bibr" rid="b46">HilleRisLambers <i>et al</i>. (2012)</xref> and <xref ref-type="bibr" rid="b133">Vellend (2016)</xref>. Different letters denote different species; circles of different sizes denote different size patches; different fan-shaped areas in circles denote the relative abundance of each species in one community.Fig. 21.1 扩散 扩散是有机体或其繁殖体离开出生地后的迁移或传播, 该过程对群落构建的影响主要依赖于扩散者产生群落(源群落)和抵达群落(汇群落)的大小和物种组成(Holyoak et al., 2005).与连续森林相比, 存在隔离的森林斑块将限制物种在斑块间的自由扩散(如种子传播), 即群落间物种的扩散限制.根据源群落和汇群落的相对大小, 片段化生境中物种的扩散可以分为两种情形: 一是大陆-岛屿模型, 该模型中将扩散假定为单向扩散, 由无限大的源群落(大陆或大面积斑块)向一个或多个小的、隔离的汇群落(岛屿或小面积斑块)中扩散, 源群落的物种组成相对恒定, 而汇群落的物种组成变化则高于源群落; 二是岛屿模型, 该模型中的群落(岛屿或小面积斑块)通过扩散相互联系在一起, 形成集合群落(Holyoak et al., 2005).一般而言, 扩散过程对群落构建的作用是受其他生态过程影响的, 尤其是选择过程和生态漂变(Ron et al., 2018). ...
Effects of habitat fragmentation on biodiversity 2 2003
... 生境片段化(或破碎化, habitat fragmentation)是指原来连续的生境受自然和人为干扰的影响, 逐渐转变为隔离的且面积较小的斑块(Wilcove et al., 1986; Fahrig, 2003).人类干扰(如择伐、都市化、公路和水库建设等)对自然生态系统的破坏以及自然因素(如森林火灾、岛屿等)的影响, 造成全球森林植被呈片段化分布(Fahrig, 2003; Didham, 2010; Haddad et al., 2015; Taubert et al., 2018; Liu et al., 2019a).从全球森林的分布来看, 世界上大部分森林斑块的面积小于10 hm2 (Haddad et al., 2015).森林面积的丧失、隔离度的增加和边缘效应的加剧将显著影响片段化生境中森林群落的结构(Laurance et al., 2006)和演替动态(Benchimol & Peres, 2015; Liu et al., 2019b). ...
... ).人类干扰(如择伐、都市化、公路和水库建设等)对自然生态系统的破坏以及自然因素(如森林火灾、岛屿等)的影响, 造成全球森林植被呈片段化分布(Fahrig, 2003; Didham, 2010; Haddad et al., 2015; Taubert et al., 2018; Liu et al., 2019a).从全球森林的分布来看, 世界上大部分森林斑块的面积小于10 hm2 (Haddad et al., 2015).森林面积的丧失、隔离度的增加和边缘效应的加剧将显著影响片段化生境中森林群落的结构(Laurance et al., 2006)和演替动态(Benchimol & Peres, 2015; Liu et al., 2019b). ...
Rethinking patch size and isolation effects: The habitat amount hypothesis 2 2013
Using species abundance distribution models and diversity indices for biogeographical analyses 1 2016
... 物种多度分布(SAD)描述了一个取样单元或群落内所有物种的个体多度分布状况, 是一个群落最基本的描述(McGill et al., 2007).通常, SAD通过物种序列多度图(RAD)进行可视化(Ulrich et al., 2010).已有大量的研究对群落内RAD的形状进行模型拟合, 如常见的模型有: 对数级数模型、对数正态模型等近30个模型(McGill et al., 2007).但是, 如何根据RAD曲线所拟合的模型反映群落的构建过程仍是研究的热点和难点问题.虽然仅通过对不同SAD模型的拟合尚无法准确地推测群落的构建过程, 但在地理尺度上, 通过分析不同环境因子和空间因子等对RAD形状的影响, 可以探讨扩散过程对群落构建的影响(Fattorini et al., 2016).在全球尺度上, Ulrich等(2016)通过对数级数模型和对数正态模型的拟合, 在纬度梯度上分析了扩散限制和环境过滤对群落构建的影响, 发现处于不同纬度的群落内扩散限制所产生的相对作用不同, 导致了低纬度地区群落内的RAD符合对数级数模型而高纬度地区群落内RAD更加符合对数正态模型.相似地, 在纬度梯度上利用非中性模型对森林群落的RAD进行拟合, 发现随纬度梯度的增加,环境过滤作用导致实际的SAD模型与中性模型的分化逐渐增加, 即中性过程对群落构建作用的影响减弱, 而非中性过程增加(Qiao et al., 2015a).Matthews等(2017)发现岛屿间的隔离程度可以改变岛屿上节肢动物的RAD的形状, 但关于岛屿参数如何影响森林群落的RAD尚缺乏研究.SAD不仅可以了解片段化生境中群落构建的过程, 也能够反映群落内稀有种和常见种的分布格局, 通过探讨片段化生境内物种多度的改变, 可应用于生物多样性的管理和应用生态学等方面(Matthews & Whittaker, 2015b).但在不同面积和隔离度的片段化森林中, 结合RAD形状的改变探讨群落构建过程的研究仍然不足. ...
Plant dispersal traits determine hydrochorous species tolerance to connectivity loss at the landscape scale 1 2017
... 在片段化生境中, 根据群落内物种的某一类性状, 如与种群扩散、建立和维持等相关的植物生活史性状(Zambrano et al., 2019), 结合不同大小斑块内的生境条件和隔离度等对群落内功能特征收敛或发散程度的影响, 也能有助于我们理解选择过程或扩散过程在群落构建中的作用(?ckinger et al., 2010; Marini et al., 2012; Jiang et al., 2018).例如, 结合种子的功能特征, 可以分析扩散限制作用在群落构建过程中的作用(Negoita et al., 2016; Favre-Bac et al., 2017).有研究发现在越大的岛屿上小种子植物比例更高(种子特征的收敛), 可能是由于大岛上小种子扩散者(鸟类)的比例更高所引起的(Liu et al., 2019c). ...
Conceptual cartoon illustrating processes in forest community assembly in fragmented habitats. Redrawn from <xref ref-type="bibr" rid="b46">HilleRisLambers <i>et al</i>. (2012)</xref> and <xref ref-type="bibr" rid="b133">Vellend (2016)</xref>. Different letters denote different species; circles of different sizes denote different size patches; different fan-shaped areas in circles denote the relative abundance of each species in one community.Fig. 21.1 扩散 扩散是有机体或其繁殖体离开出生地后的迁移或传播, 该过程对群落构建的影响主要依赖于扩散者产生群落(源群落)和抵达群落(汇群落)的大小和物种组成(Holyoak et al., 2005).与连续森林相比, 存在隔离的森林斑块将限制物种在斑块间的自由扩散(如种子传播), 即群落间物种的扩散限制.根据源群落和汇群落的相对大小, 片段化生境中物种的扩散可以分为两种情形: 一是大陆-岛屿模型, 该模型中将扩散假定为单向扩散, 由无限大的源群落(大陆或大面积斑块)向一个或多个小的、隔离的汇群落(岛屿或小面积斑块)中扩散, 源群落的物种组成相对恒定, 而汇群落的物种组成变化则高于源群落; 二是岛屿模型, 该模型中的群落(岛屿或小面积斑块)通过扩散相互联系在一起, 形成集合群落(Holyoak et al., 2005).一般而言, 扩散过程对群落构建的作用是受其他生态过程影响的, 尤其是选择过程和生态漂变(Ron et al., 2018). ...
... ); Moss Fragmentation Experiments (UK: Gonzalez et al. 1998; Canada: Lindo et al., 2012); The Thousand Island Lake Experiments (China)(Yu et al., 2012; Liu et al., 2020); The Stability of Altered Forest Ecosystems (SAFE) Project (Malaysia)(Ewers et al., 2011); Metatron experiment (France)(Legrand et al., 2012). Fig. 1 10.17521/cjpe.2019.0155.F002
Conceptual cartoon illustrating processes in forest community assembly in fragmented habitats. Redrawn from <xref ref-type="bibr" rid="b46">HilleRisLambers <i>et al</i>. (2012)</xref> and <xref ref-type="bibr" rid="b133">Vellend (2016)</xref>. Different letters denote different species; circles of different sizes denote different size patches; different fan-shaped areas in circles denote the relative abundance of each species in one community.Fig. 21.1 扩散 扩散是有机体或其繁殖体离开出生地后的迁移或传播, 该过程对群落构建的影响主要依赖于扩散者产生群落(源群落)和抵达群落(汇群落)的大小和物种组成(Holyoak et al., 2005).与连续森林相比, 存在隔离的森林斑块将限制物种在斑块间的自由扩散(如种子传播), 即群落间物种的扩散限制.根据源群落和汇群落的相对大小, 片段化生境中物种的扩散可以分为两种情形: 一是大陆-岛屿模型, 该模型中将扩散假定为单向扩散, 由无限大的源群落(大陆或大面积斑块)向一个或多个小的、隔离的汇群落(岛屿或小面积斑块)中扩散, 源群落的物种组成相对恒定, 而汇群落的物种组成变化则高于源群落; 二是岛屿模型, 该模型中的群落(岛屿或小面积斑块)通过扩散相互联系在一起, 形成集合群落(Holyoak et al., 2005).一般而言, 扩散过程对群落构建的作用是受其他生态过程影响的, 尤其是选择过程和生态漂变(Ron et al., 2018). ...
Trophic theory of island biogeography 1 2011
... 以往的理论/假说(如ETIB、集合群落理论和群落生态学理论)主要针对单一营养级或两个营养级上的生态过程进行分析, 如单一营养级上的种间相互作用及其与非生物因子间的关系(选择过程).近年来, 在ETIB理论的基础上, 许多研究开始逐渐关注岛屿生物地理学的营养理论, 即将营养级间物种的相互作用对定殖率和灭绝率的影响考虑其中(Gravel et al., 2011).在片段化生境中, 食物网各营养级间关联物种的次生灭绝是非常普遍的, 越顶级的消费者越容易消失(Ryser et al., 2019), 消费者缺失所引发的下行调控丧失效应通过影响传粉、种子传播和植食等与扩散和选择相关的生态过程, 从而影响片段化森林群落的构建过程(Valladares et al., 2006; Larios et al., 2017; Emer et al., 2018).Cirtwill和Stouffer (2016)发现, 当考虑岛屿上的营养级结构时, 如果包含上行效应的影响, 能够改进ETIB中对迁移率和灭绝率的预测.但在片段化森林景观中, 针对多营养级上的种间相互作用(如下行效应)在森林群落构建过程的作用研究, 仍鲜有报道. ...
An experimental test of corridor effects on butterfly densities 4 1999
Conceptual cartoon illustrating processes in forest community assembly in fragmented habitats. Redrawn from <xref ref-type="bibr" rid="b46">HilleRisLambers <i>et al</i>. (2012)</xref> and <xref ref-type="bibr" rid="b133">Vellend (2016)</xref>. Different letters denote different species; circles of different sizes denote different size patches; different fan-shaped areas in circles denote the relative abundance of each species in one community.Fig. 21.1 扩散 扩散是有机体或其繁殖体离开出生地后的迁移或传播, 该过程对群落构建的影响主要依赖于扩散者产生群落(源群落)和抵达群落(汇群落)的大小和物种组成(Holyoak et al., 2005).与连续森林相比, 存在隔离的森林斑块将限制物种在斑块间的自由扩散(如种子传播), 即群落间物种的扩散限制.根据源群落和汇群落的相对大小, 片段化生境中物种的扩散可以分为两种情形: 一是大陆-岛屿模型, 该模型中将扩散假定为单向扩散, 由无限大的源群落(大陆或大面积斑块)向一个或多个小的、隔离的汇群落(岛屿或小面积斑块)中扩散, 源群落的物种组成相对恒定, 而汇群落的物种组成变化则高于源群落; 二是岛屿模型, 该模型中的群落(岛屿或小面积斑块)通过扩散相互联系在一起, 形成集合群落(Holyoak et al., 2005).一般而言, 扩散过程对群落构建的作用是受其他生态过程影响的, 尤其是选择过程和生态漂变(Ron et al., 2018). ...
... ); Savannah River Site Corridor Experiment (SRS Corridor Experiment; USA)(Haddad & Baum, 1999; Tewksbury et al., 2002); Moss Fragmentation Experiments (UK: Gonzalez et al. 1998; Canada: Lindo et al., 2012); The Thousand Island Lake Experiments (China)(Yu et al., 2012; Liu et al., 2020); The Stability of Altered Forest Ecosystems (SAFE) Project (Malaysia)(Ewers et al., 2011); Metatron experiment (France)(Legrand et al., 2012). Fig. 1 10.17521/cjpe.2019.0155.F002
Conceptual cartoon illustrating processes in forest community assembly in fragmented habitats. Redrawn from <xref ref-type="bibr" rid="b46">HilleRisLambers <i>et al</i>. (2012)</xref> and <xref ref-type="bibr" rid="b133">Vellend (2016)</xref>. Different letters denote different species; circles of different sizes denote different size patches; different fan-shaped areas in circles denote the relative abundance of each species in one community.Fig. 21.1 扩散 扩散是有机体或其繁殖体离开出生地后的迁移或传播, 该过程对群落构建的影响主要依赖于扩散者产生群落(源群落)和抵达群落(汇群落)的大小和物种组成(Holyoak et al., 2005).与连续森林相比, 存在隔离的森林斑块将限制物种在斑块间的自由扩散(如种子传播), 即群落间物种的扩散限制.根据源群落和汇群落的相对大小, 片段化生境中物种的扩散可以分为两种情形: 一是大陆-岛屿模型, 该模型中将扩散假定为单向扩散, 由无限大的源群落(大陆或大面积斑块)向一个或多个小的、隔离的汇群落(岛屿或小面积斑块)中扩散, 源群落的物种组成相对恒定, 而汇群落的物种组成变化则高于源群落; 二是岛屿模型, 该模型中的群落(岛屿或小面积斑块)通过扩散相互联系在一起, 形成集合群落(Holyoak et al., 2005).一般而言, 扩散过程对群落构建的作用是受其他生态过程影响的, 尤其是选择过程和生态漂变(Ron et al., 2018). ...
... 野外控制实验也是检验片段化生境中群落构建过程非常有效的手段(Debinski & Holt, 2000; Resasco et al., 2017).Haddad等(2015)综述了目前世界上已有的针对片段化生境的控制实验(图1), 通过设置控制实验和长期观察, 对片段化生境中或岛屿上群落构建的过程进行了有效的检验(Wilson et al., 2016; Resasco et al., 2017).例如, 巴西的片段化森林动态实验是目前最大和时间最长的野外控制实验, 通过设置不同大小的森林斑块且控制斑块之间的隔离度差异, 分析森林斑块面积的大小对群落构建过程的影响(Laurance et al., 2011); 美国的Savannah河流廊道实验通过增加斑块间的隔离度, 且控制斑块的面积差异, 分析廊道效应(扩散)在群落构建过程中的作用(Haddad & Baum, 1999; Tewksbury et al., 2002); Kansas生境片段化实验则通过设置不同面积大小和隔离度的片段化斑块, 长期监测斑块内群落的动态变化, 研究生境片段化对群落构建和演替过程的影响(Holt et al., 1995); 另外, Alexander等(2012)利用此实验在种群水平上分析了种子扩散与斑块间连通性之间的关系, 在群落水平上分析了环境过滤和扩散限制的共同作用对群落演替的作用.然而, 除上述主要的实验外, 基于野外控制实验检验片段化生境中森林群落构建过程的研究仍然不足. ...
Habitat fragmentation and its lasting impact on Earth?s ecosystems 3 2015
... 生境片段化(或破碎化, habitat fragmentation)是指原来连续的生境受自然和人为干扰的影响, 逐渐转变为隔离的且面积较小的斑块(Wilcove et al., 1986; Fahrig, 2003).人类干扰(如择伐、都市化、公路和水库建设等)对自然生态系统的破坏以及自然因素(如森林火灾、岛屿等)的影响, 造成全球森林植被呈片段化分布(Fahrig, 2003; Didham, 2010; Haddad et al., 2015; Taubert et al., 2018; Liu et al., 2019a).从全球森林的分布来看, 世界上大部分森林斑块的面积小于10 hm2 (Haddad et al., 2015).森林面积的丧失、隔离度的增加和边缘效应的加剧将显著影响片段化生境中森林群落的结构(Laurance et al., 2006)和演替动态(Benchimol & Peres, 2015; Liu et al., 2019b). ...
... (Haddad et al., 2015).森林面积的丧失、隔离度的增加和边缘效应的加剧将显著影响片段化生境中森林群落的结构(Laurance et al., 2006)和演替动态(Benchimol & Peres, 2015; Liu et al., 2019b). ...
... 野外控制实验也是检验片段化生境中群落构建过程非常有效的手段(Debinski & Holt, 2000; Resasco et al., 2017).Haddad等(2015)综述了目前世界上已有的针对片段化生境的控制实验(图1), 通过设置控制实验和长期观察, 对片段化生境中或岛屿上群落构建的过程进行了有效的检验(Wilson et al., 2016; Resasco et al., 2017).例如, 巴西的片段化森林动态实验是目前最大和时间最长的野外控制实验, 通过设置不同大小的森林斑块且控制斑块之间的隔离度差异, 分析森林斑块面积的大小对群落构建过程的影响(Laurance et al., 2011); 美国的Savannah河流廊道实验通过增加斑块间的隔离度, 且控制斑块的面积差异, 分析廊道效应(扩散)在群落构建过程中的作用(Haddad & Baum, 1999; Tewksbury et al., 2002); Kansas生境片段化实验则通过设置不同面积大小和隔离度的片段化斑块, 长期监测斑块内群落的动态变化, 研究生境片段化对群落构建和演替过程的影响(Holt et al., 1995); 另外, Alexander等(2012)利用此实验在种群水平上分析了种子扩散与斑块间连通性之间的关系, 在群落水平上分析了环境过滤和扩散限制的共同作用对群落演替的作用.然而, 除上述主要的实验外, 基于野外控制实验检验片段化生境中森林群落构建过程的研究仍然不足. ...
A practical model of metapopulation dynamics 1 1994
... 片段化生境中森林群落构建过程的概念图.参考自HilleRisLambers等(2012)和Vellend (2016).不同的英文字母表示不同的物种; 不同大小的圆圈表示不同面积的斑块, 斑块内不同的扇形面积表示各物种的相对多度. Conceptual cartoon illustrating processes in forest community assembly in fragmented habitats. Redrawn from <xref ref-type="bibr" rid="b46">HilleRisLambers <i>et al</i>. (2012)</xref> and <xref ref-type="bibr" rid="b133">Vellend (2016)</xref>. Different letters denote different species; circles of different sizes denote different size patches; different fan-shaped areas in circles denote the relative abundance of each species in one community.Fig. 21.1 扩散 扩散是有机体或其繁殖体离开出生地后的迁移或传播, 该过程对群落构建的影响主要依赖于扩散者产生群落(源群落)和抵达群落(汇群落)的大小和物种组成(Holyoak et al., 2005).与连续森林相比, 存在隔离的森林斑块将限制物种在斑块间的自由扩散(如种子传播), 即群落间物种的扩散限制.根据源群落和汇群落的相对大小, 片段化生境中物种的扩散可以分为两种情形: 一是大陆-岛屿模型, 该模型中将扩散假定为单向扩散, 由无限大的源群落(大陆或大面积斑块)向一个或多个小的、隔离的汇群落(岛屿或小面积斑块)中扩散, 源群落的物种组成相对恒定, 而汇群落的物种组成变化则高于源群落; 二是岛屿模型, 该模型中的群落(岛屿或小面积斑块)通过扩散相互联系在一起, 形成集合群落(Holyoak et al., 2005).一般而言, 扩散过程对群落构建的作用是受其他生态过程影响的, 尤其是选择过程和生态漂变(Ron et al., 2018). ...
... Conceptual cartoon illustrating processes in forest community assembly in fragmented habitats. Redrawn from HilleRisLambers et al. (2012) and Vellend (2016). Different letters denote different species; circles of different sizes denote different size patches; different fan-shaped areas in circles denote the relative abundance of each species in one community. Fig. 21.1 扩散 扩散是有机体或其繁殖体离开出生地后的迁移或传播, 该过程对群落构建的影响主要依赖于扩散者产生群落(源群落)和抵达群落(汇群落)的大小和物种组成(Holyoak et al., 2005).与连续森林相比, 存在隔离的森林斑块将限制物种在斑块间的自由扩散(如种子传播), 即群落间物种的扩散限制.根据源群落和汇群落的相对大小, 片段化生境中物种的扩散可以分为两种情形: 一是大陆-岛屿模型, 该模型中将扩散假定为单向扩散, 由无限大的源群落(大陆或大面积斑块)向一个或多个小的、隔离的汇群落(岛屿或小面积斑块)中扩散, 源群落的物种组成相对恒定, 而汇群落的物种组成变化则高于源群落; 二是岛屿模型, 该模型中的群落(岛屿或小面积斑块)通过扩散相互联系在一起, 形成集合群落(Holyoak et al., 2005).一般而言, 扩散过程对群落构建的作用是受其他生态过程影响的, 尤其是选择过程和生态漂变(Ron et al., 2018). ...
Vegetation dynamics in an experimentally fragmented landscape 3 1995
Conceptual cartoon illustrating processes in forest community assembly in fragmented habitats. Redrawn from <xref ref-type="bibr" rid="b46">HilleRisLambers <i>et al</i>. (2012)</xref> and <xref ref-type="bibr" rid="b133">Vellend (2016)</xref>. Different letters denote different species; circles of different sizes denote different size patches; different fan-shaped areas in circles denote the relative abundance of each species in one community.Fig. 21.1 扩散 扩散是有机体或其繁殖体离开出生地后的迁移或传播, 该过程对群落构建的影响主要依赖于扩散者产生群落(源群落)和抵达群落(汇群落)的大小和物种组成(Holyoak et al., 2005).与连续森林相比, 存在隔离的森林斑块将限制物种在斑块间的自由扩散(如种子传播), 即群落间物种的扩散限制.根据源群落和汇群落的相对大小, 片段化生境中物种的扩散可以分为两种情形: 一是大陆-岛屿模型, 该模型中将扩散假定为单向扩散, 由无限大的源群落(大陆或大面积斑块)向一个或多个小的、隔离的汇群落(岛屿或小面积斑块)中扩散, 源群落的物种组成相对恒定, 而汇群落的物种组成变化则高于源群落; 二是岛屿模型, 该模型中的群落(岛屿或小面积斑块)通过扩散相互联系在一起, 形成集合群落(Holyoak et al., 2005).一般而言, 扩散过程对群落构建的作用是受其他生态过程影响的, 尤其是选择过程和生态漂变(Ron et al., 2018). ...
... ); Kansas Fragmentation Experiment (USA)(Holt et al., 1995); Wog-Wog Habitat Fragmentation Experiment (Wog-Wog; Australia)(Margules, 1992); Savannah River Site Corridor Experiment (SRS Corridor Experiment; USA)(Haddad & Baum, 1999; Tewksbury et al., 2002); Moss Fragmentation Experiments (UK: Gonzalez et al. 1998; Canada: Lindo et al., 2012); The Thousand Island Lake Experiments (China)(Yu et al., 2012; Liu et al., 2020); The Stability of Altered Forest Ecosystems (SAFE) Project (Malaysia)(Ewers et al., 2011); Metatron experiment (France)(Legrand et al., 2012). Fig. 1 10.17521/cjpe.2019.0155.F002
Conceptual cartoon illustrating processes in forest community assembly in fragmented habitats. Redrawn from <xref ref-type="bibr" rid="b46">HilleRisLambers <i>et al</i>. (2012)</xref> and <xref ref-type="bibr" rid="b133">Vellend (2016)</xref>. Different letters denote different species; circles of different sizes denote different size patches; different fan-shaped areas in circles denote the relative abundance of each species in one community.Fig. 21.1 扩散 扩散是有机体或其繁殖体离开出生地后的迁移或传播, 该过程对群落构建的影响主要依赖于扩散者产生群落(源群落)和抵达群落(汇群落)的大小和物种组成(Holyoak et al., 2005).与连续森林相比, 存在隔离的森林斑块将限制物种在斑块间的自由扩散(如种子传播), 即群落间物种的扩散限制.根据源群落和汇群落的相对大小, 片段化生境中物种的扩散可以分为两种情形: 一是大陆-岛屿模型, 该模型中将扩散假定为单向扩散, 由无限大的源群落(大陆或大面积斑块)向一个或多个小的、隔离的汇群落(岛屿或小面积斑块)中扩散, 源群落的物种组成相对恒定, 而汇群落的物种组成变化则高于源群落; 二是岛屿模型, 该模型中的群落(岛屿或小面积斑块)通过扩散相互联系在一起, 形成集合群落(Holyoak et al., 2005).一般而言, 扩散过程对群落构建的作用是受其他生态过程影响的, 尤其是选择过程和生态漂变(Ron et al., 2018). ...
... 野外控制实验也是检验片段化生境中群落构建过程非常有效的手段(Debinski & Holt, 2000; Resasco et al., 2017).Haddad等(2015)综述了目前世界上已有的针对片段化生境的控制实验(图1), 通过设置控制实验和长期观察, 对片段化生境中或岛屿上群落构建的过程进行了有效的检验(Wilson et al., 2016; Resasco et al., 2017).例如, 巴西的片段化森林动态实验是目前最大和时间最长的野外控制实验, 通过设置不同大小的森林斑块且控制斑块之间的隔离度差异, 分析森林斑块面积的大小对群落构建过程的影响(Laurance et al., 2011); 美国的Savannah河流廊道实验通过增加斑块间的隔离度, 且控制斑块的面积差异, 分析廊道效应(扩散)在群落构建过程中的作用(Haddad & Baum, 1999; Tewksbury et al., 2002); Kansas生境片段化实验则通过设置不同面积大小和隔离度的片段化斑块, 长期监测斑块内群落的动态变化, 研究生境片段化对群落构建和演替过程的影响(Holt et al., 1995); 另外, Alexander等(2012)利用此实验在种群水平上分析了种子扩散与斑块间连通性之间的关系, 在群落水平上分析了环境过滤和扩散限制的共同作用对群落演替的作用.然而, 除上述主要的实验外, 基于野外控制实验检验片段化生境中森林群落构建过程的研究仍然不足. ...
Presence and absence of density dependence in a neotropical tree community 1 1990
... 关于选择过程对植物群落构建的影响, 主要集中于局域尺度上基于生态位理论的研究, 即非生物环境因子的筛选和种内/种间的竞争作用(Tilman, 1982; Chase & Leibold, 2003; Letten et al., 2017).植物群落中, 选择过程产生于群落内不同物种个体间适合度的差异(Vellend, 2010).群落构建过程中, 一般存在3种形式的选择过程: (1)固定选择效应; (2)密度依赖效应; (3)时空选择效应.固定选择效应是指在特定的环境条件下, 如果群落内物种的相对适合度在时间和空间上固定不变, 那么群落内具有最高适合度的物种将会排除其他所有的物种(Vellend, 2010).密度依赖效应取决于群落内物种间定性(例如竞争、捕食、互利共生和疾病等)和定量的生态关系, 即所研究物种的适合度依赖于群落内同种和异种的个体密度, 包括负密度依赖效应(或被称为负密度制约效应)(祝燕等, 2009)和正密度依赖效应两个方面(Vellend, 2016).负密度依赖效应的研究内容主要包括: (1)密度依赖的自疏过程(随机死亡假说), 即同种个体的密度与死亡率呈现正相关关系(Kenkel, 1988); (2)距离依赖的死亡过程(Janzen-Connell假说), 即母树周围同种个体具有较高的密度时, 越靠近母树的同种个体死亡率越高(Janzen, 1970; Hubbell et al., 1990; Hyatt et al., 2003); (3)群落补偿趋势假说, 即群落越大, 负密度依赖作用越强, 越有利于稀有种在群落内的存活(Connell et al., 1984).负密度依赖效应对群落内的低密度物种更加有利, 故而可以使群落内物种维持稳定的共存状态(祝燕等, 2009).在片段化生境中, 不管是固定选择效应还是密度依赖效应都可能在时间和空间上存在差异, 即时空选择效应, 该效应也会潜在地影响群落动态.此外, 生境片段化限制了植物种子传播/扩散(Terborgh et al., 2011)以及特异性植食动物组成和数量的改变(Valladares et al., 2006; Emer et al., 2018), 进而影响种子或幼苗被植食的概率(白冰等, 2011; Genua et al., 2017; 骆杨青等, 2017).生境片段化所产生的边缘效应也能改变植物与天敌之间的关系(Krishnadas et al., 2018).生境片段化所引起的种间相互作用的改变, 最终会导致植物个体在空间分布格局上的差异, 影响密度效应的选择强度. ...
The distance dependence prediction of the Janzen-Connell hypothesis: A meta-analysis 1 2003
... 关于选择过程对植物群落构建的影响, 主要集中于局域尺度上基于生态位理论的研究, 即非生物环境因子的筛选和种内/种间的竞争作用(Tilman, 1982; Chase & Leibold, 2003; Letten et al., 2017).植物群落中, 选择过程产生于群落内不同物种个体间适合度的差异(Vellend, 2010).群落构建过程中, 一般存在3种形式的选择过程: (1)固定选择效应; (2)密度依赖效应; (3)时空选择效应.固定选择效应是指在特定的环境条件下, 如果群落内物种的相对适合度在时间和空间上固定不变, 那么群落内具有最高适合度的物种将会排除其他所有的物种(Vellend, 2010).密度依赖效应取决于群落内物种间定性(例如竞争、捕食、互利共生和疾病等)和定量的生态关系, 即所研究物种的适合度依赖于群落内同种和异种的个体密度, 包括负密度依赖效应(或被称为负密度制约效应)(祝燕等, 2009)和正密度依赖效应两个方面(Vellend, 2016).负密度依赖效应的研究内容主要包括: (1)密度依赖的自疏过程(随机死亡假说), 即同种个体的密度与死亡率呈现正相关关系(Kenkel, 1988); (2)距离依赖的死亡过程(Janzen-Connell假说), 即母树周围同种个体具有较高的密度时, 越靠近母树的同种个体死亡率越高(Janzen, 1970; Hubbell et al., 1990; Hyatt et al., 2003); (3)群落补偿趋势假说, 即群落越大, 负密度依赖作用越强, 越有利于稀有种在群落内的存活(Connell et al., 1984).负密度依赖效应对群落内的低密度物种更加有利, 故而可以使群落内物种维持稳定的共存状态(祝燕等, 2009).在片段化生境中, 不管是固定选择效应还是密度依赖效应都可能在时间和空间上存在差异, 即时空选择效应, 该效应也会潜在地影响群落动态.此外, 生境片段化限制了植物种子传播/扩散(Terborgh et al., 2011)以及特异性植食动物组成和数量的改变(Valladares et al., 2006; Emer et al., 2018), 进而影响种子或幼苗被植食的概率(白冰等, 2011; Genua et al., 2017; 骆杨青等, 2017).生境片段化所产生的边缘效应也能改变植物与天敌之间的关系(Krishnadas et al., 2018).生境片段化所引起的种间相互作用的改变, 最终会导致植物个体在空间分布格局上的差异, 影响密度效应的选择强度. ...
Extensions of Island Biogeography Theory predict the scaling of functional trait composition with habitat area and isolation 1 2017
... 集合群落理论包含中性过程、斑块动态、物种筛选和质量效应4种理论框架(Leibold et al., 2004; Holyoak et al., 2005)(附件I), 在ETIB和中性理论的基础上, 将选择过程考虑在内, 阐述了片段化生境中群落构建的基本过程.Vellend (2016)所提出的生态群落构建的4个高阶过程中的生态漂变类似于集合群落模型中的中性过程, 扩散和选择过程分别类似于斑块动态和物种筛选.虽然物种形成在集合群落理论中没有被明确排除在外, 但在多数情况下, 集合群落理论中并没有考虑该过程.在Vellend (2016)群落生态学理论提出后, 已有部分研究在片段化/岛屿生境中检验不同的生态过程在群落构建中的相对作用, 如选择过程和生态漂变过程的相对作用(Liu et al., 2018)、扩散对选择和生态漂变过程相对作用的影响(Ron et al., 2018)等.另外, 通过扩展ETIB理论, 考虑物种间功能特征的差异及其所受面积和隔离度的影响(Jacquet et al., 2017), 以及物种功能性状特征分布与环境因子间的关系, 逐渐成为检验片段化生境中群落构建过程的重要手段(Zambrano et al., 2019).然而, 针对不同生态过程如何影响片段化森林群落构建的研究仍然十分缺乏, 尤其是对于扩散和物种形成过程的相关检验. ...
Herbivores and the number of tree species in tropical forests 1 1970
... 关于选择过程对植物群落构建的影响, 主要集中于局域尺度上基于生态位理论的研究, 即非生物环境因子的筛选和种内/种间的竞争作用(Tilman, 1982; Chase & Leibold, 2003; Letten et al., 2017).植物群落中, 选择过程产生于群落内不同物种个体间适合度的差异(Vellend, 2010).群落构建过程中, 一般存在3种形式的选择过程: (1)固定选择效应; (2)密度依赖效应; (3)时空选择效应.固定选择效应是指在特定的环境条件下, 如果群落内物种的相对适合度在时间和空间上固定不变, 那么群落内具有最高适合度的物种将会排除其他所有的物种(Vellend, 2010).密度依赖效应取决于群落内物种间定性(例如竞争、捕食、互利共生和疾病等)和定量的生态关系, 即所研究物种的适合度依赖于群落内同种和异种的个体密度, 包括负密度依赖效应(或被称为负密度制约效应)(祝燕等, 2009)和正密度依赖效应两个方面(Vellend, 2016).负密度依赖效应的研究内容主要包括: (1)密度依赖的自疏过程(随机死亡假说), 即同种个体的密度与死亡率呈现正相关关系(Kenkel, 1988); (2)距离依赖的死亡过程(Janzen-Connell假说), 即母树周围同种个体具有较高的密度时, 越靠近母树的同种个体死亡率越高(Janzen, 1970; Hubbell et al., 1990; Hyatt et al., 2003); (3)群落补偿趋势假说, 即群落越大, 负密度依赖作用越强, 越有利于稀有种在群落内的存活(Connell et al., 1984).负密度依赖效应对群落内的低密度物种更加有利, 故而可以使群落内物种维持稳定的共存状态(祝燕等, 2009).在片段化生境中, 不管是固定选择效应还是密度依赖效应都可能在时间和空间上存在差异, 即时空选择效应, 该效应也会潜在地影响群落动态.此外, 生境片段化限制了植物种子传播/扩散(Terborgh et al., 2011)以及特异性植食动物组成和数量的改变(Valladares et al., 2006; Emer et al., 2018), 进而影响种子或幼苗被植食的概率(白冰等, 2011; Genua et al., 2017; 骆杨青等, 2017).生境片段化所产生的边缘效应也能改变植物与天敌之间的关系(Krishnadas et al., 2018).生境片段化所引起的种间相互作用的改变, 最终会导致植物个体在空间分布格局上的差异, 影响密度效应的选择强度. ...
Functional traits can improve our understanding of niche- and dispersal-based processes 1 2018
... 在片段化生境中, 根据群落内物种的某一类性状, 如与种群扩散、建立和维持等相关的植物生活史性状(Zambrano et al., 2019), 结合不同大小斑块内的生境条件和隔离度等对群落内功能特征收敛或发散程度的影响, 也能有助于我们理解选择过程或扩散过程在群落构建中的作用(?ckinger et al., 2010; Marini et al., 2012; Jiang et al., 2018).例如, 结合种子的功能特征, 可以分析扩散限制作用在群落构建过程中的作用(Negoita et al., 2016; Favre-Bac et al., 2017).有研究发现在越大的岛屿上小种子植物比例更高(种子特征的收敛), 可能是由于大岛上小种子扩散者(鸟类)的比例更高所引起的(Liu et al., 2019c). ...
Community assembly in the presence of disturbance: A microcosm experiment 1 2008
... 此外, 结合微宇宙的实验方法, 也可以构建模拟群落, 通过直接观察不同实验情景下的群落动态, 能够验证某一生态过程在群落构建中的作用(Davies et al., 2009; Altermatt et al., 2015; Ron et al., 2018).通常利用微生物群落, 通过配置菌落的大小(生态漂变)、菌群间隔离度(扩散)、环境胁迫或干扰(选择)、起始菌落的物种组成(优先效应)等, 验证生态群落的构建过程(Jiang & Patel, 2008; Ojima & Jiang, 2017).然而, 对于森林群落而言, 很难利用微宇宙的实验方法对群落构建过程进行检验, 由微宇宙实验获得的结论是否适用于片段化生境中的森林群落, 仍值得商榷. ...
Effects of isolation, logging and dispersal on woody-species richness of islands 1 1995
... 迄今关于片段化生境中重构森林群落的研究仍然十分缺乏.我们对片段化生境中森林的重新构建过程可以作如下预测: 1)重构群落演替的起始阶段, 空间的隔离作用将比斑块面积的大小更加重要, 因短期的干扰或生境质量的改变, 空间的隔离将阻止物种占用适合的生境, 导致种子传播或者强风等随机事件决定了物种的抵达和在斑块上的定殖(Walker et al., 2010).例如, Kadmon和Pulliam (1995)发现在新近皆伐的湖泊岛屿上空间的隔离增加了风力传播物种的比例, Whittaker等(1997)发现在新近形成的火山岛屿上, 物种早期阶段的定殖可以根据其扩散性状进行预测, 不过在具有近60年片段化历史的千岛湖, 岛屿面积是影响植物群落物种组成的主要因素, 而非空间的隔离(Yu et al., 2012; Baldeck et al., 2016).2)当某一物种扩散到某一斑块后, 斑块内已有的种子/幼苗库, 会对后续扩散来的物种的定殖产生影响, 即优先效应, 该效应与决定性过程和随机过程会共同影响片段化生境中群落的构建过程(Fukami, 2015).3)物种抵达相应的斑块之后, 斑块内的生境异质性、种间/种内关系和生态漂变等过程将共同决定物种定殖成功的概率和物种组成, 如筛选个体生长快、木质密度低以及喜阳的先锋物种(Baldeck et al., 2016; Liu et al., 2019b, 2020), 或具有菌根的植物物种等(Delavaux et al., 2019).Baynes等(2016)通过比较砍伐后重构的片段化森林群落与连续森林群落内的物种多样性, 发现重构的森林群落中反而具有更多的稀有种以及较高的β多样性, 生态漂变过程也会影响片段化森林群落的重构. ...
Pattern of self-thinning in jack pine: Testing the random mortality hypothesis 1 1988
... 关于选择过程对植物群落构建的影响, 主要集中于局域尺度上基于生态位理论的研究, 即非生物环境因子的筛选和种内/种间的竞争作用(Tilman, 1982; Chase & Leibold, 2003; Letten et al., 2017).植物群落中, 选择过程产生于群落内不同物种个体间适合度的差异(Vellend, 2010).群落构建过程中, 一般存在3种形式的选择过程: (1)固定选择效应; (2)密度依赖效应; (3)时空选择效应.固定选择效应是指在特定的环境条件下, 如果群落内物种的相对适合度在时间和空间上固定不变, 那么群落内具有最高适合度的物种将会排除其他所有的物种(Vellend, 2010).密度依赖效应取决于群落内物种间定性(例如竞争、捕食、互利共生和疾病等)和定量的生态关系, 即所研究物种的适合度依赖于群落内同种和异种的个体密度, 包括负密度依赖效应(或被称为负密度制约效应)(祝燕等, 2009)和正密度依赖效应两个方面(Vellend, 2016).负密度依赖效应的研究内容主要包括: (1)密度依赖的自疏过程(随机死亡假说), 即同种个体的密度与死亡率呈现正相关关系(Kenkel, 1988); (2)距离依赖的死亡过程(Janzen-Connell假说), 即母树周围同种个体具有较高的密度时, 越靠近母树的同种个体死亡率越高(Janzen, 1970; Hubbell et al., 1990; Hyatt et al., 2003); (3)群落补偿趋势假说, 即群落越大, 负密度依赖作用越强, 越有利于稀有种在群落内的存活(Connell et al., 1984).负密度依赖效应对群落内的低密度物种更加有利, 故而可以使群落内物种维持稳定的共存状态(祝燕等, 2009).在片段化生境中, 不管是固定选择效应还是密度依赖效应都可能在时间和空间上存在差异, 即时空选择效应, 该效应也会潜在地影响群落动态.此外, 生境片段化限制了植物种子传播/扩散(Terborgh et al., 2011)以及特异性植食动物组成和数量的改变(Valladares et al., 2006; Emer et al., 2018), 进而影响种子或幼苗被植食的概率(白冰等, 2011; Genua et al., 2017; 骆杨青等, 2017).生境片段化所产生的边缘效应也能改变植物与天敌之间的关系(Krishnadas et al., 2018).生境片段化所引起的种间相互作用的改变, 最终会导致植物个体在空间分布格局上的差异, 影响密度效应的选择强度. ...
Forest-cover increase does not trigger forest-fragmentation decrease: Case study from the Polish carpathians 1 2018
... 随着森林植被的恢复, 虽然森林总覆盖面积将处于增加趋势, 但森林的片段化程度仍可能逐年增加(Kozak et al., 2018; Liu et al., 2019a).通过对片段化森林群落构建过程的研究和理论整合, 我们可以更好地理解不同生态过程对群落内物种组成和多样性的贡献, 从而更加有效地指导片段化生境中森林群落的恢复和对生物多样性的保护, 对于后续的森林管理、政策制定、生态系统服务功能提升和物种保护等均具有非常重要的意义. ...
Functional trait and phylogenetic tests of community assembly across spatial scales in an Amazonian forest 1 2010
Conceptual cartoon illustrating processes in forest community assembly in fragmented habitats. Redrawn from <xref ref-type="bibr" rid="b46">HilleRisLambers <i>et al</i>. (2012)</xref> and <xref ref-type="bibr" rid="b133">Vellend (2016)</xref>. Different letters denote different species; circles of different sizes denote different size patches; different fan-shaped areas in circles denote the relative abundance of each species in one community.Fig. 21.1 扩散 扩散是有机体或其繁殖体离开出生地后的迁移或传播, 该过程对群落构建的影响主要依赖于扩散者产生群落(源群落)和抵达群落(汇群落)的大小和物种组成(Holyoak et al., 2005).与连续森林相比, 存在隔离的森林斑块将限制物种在斑块间的自由扩散(如种子传播), 即群落间物种的扩散限制.根据源群落和汇群落的相对大小, 片段化生境中物种的扩散可以分为两种情形: 一是大陆-岛屿模型, 该模型中将扩散假定为单向扩散, 由无限大的源群落(大陆或大面积斑块)向一个或多个小的、隔离的汇群落(岛屿或小面积斑块)中扩散, 源群落的物种组成相对恒定, 而汇群落的物种组成变化则高于源群落; 二是岛屿模型, 该模型中的群落(岛屿或小面积斑块)通过扩散相互联系在一起, 形成集合群落(Holyoak et al., 2005).一般而言, 扩散过程对群落构建的作用是受其他生态过程影响的, 尤其是选择过程和生态漂变(Ron et al., 2018). ...
... . Major experiments and projects and the corresponding references are as following: Biological Dynamics of Forest Fragments Project (BDFFP; Brazil)(Laurance et al., 2011); Kansas Fragmentation Experiment (USA)(Holt et al., 1995); Wog-Wog Habitat Fragmentation Experiment (Wog-Wog; Australia)(Margules, 1992); Savannah River Site Corridor Experiment (SRS Corridor Experiment; USA)(Haddad & Baum, 1999; Tewksbury et al., 2002); Moss Fragmentation Experiments (UK: Gonzalez et al. 1998; Canada: Lindo et al., 2012); The Thousand Island Lake Experiments (China)(Yu et al., 2012; Liu et al., 2020); The Stability of Altered Forest Ecosystems (SAFE) Project (Malaysia)(Ewers et al., 2011); Metatron experiment (France)(Legrand et al., 2012). Fig. 1 10.17521/cjpe.2019.0155.F002
Conceptual cartoon illustrating processes in forest community assembly in fragmented habitats. Redrawn from <xref ref-type="bibr" rid="b46">HilleRisLambers <i>et al</i>. (2012)</xref> and <xref ref-type="bibr" rid="b133">Vellend (2016)</xref>. Different letters denote different species; circles of different sizes denote different size patches; different fan-shaped areas in circles denote the relative abundance of each species in one community.Fig. 21.1 扩散 扩散是有机体或其繁殖体离开出生地后的迁移或传播, 该过程对群落构建的影响主要依赖于扩散者产生群落(源群落)和抵达群落(汇群落)的大小和物种组成(Holyoak et al., 2005).与连续森林相比, 存在隔离的森林斑块将限制物种在斑块间的自由扩散(如种子传播), 即群落间物种的扩散限制.根据源群落和汇群落的相对大小, 片段化生境中物种的扩散可以分为两种情形: 一是大陆-岛屿模型, 该模型中将扩散假定为单向扩散, 由无限大的源群落(大陆或大面积斑块)向一个或多个小的、隔离的汇群落(岛屿或小面积斑块)中扩散, 源群落的物种组成相对恒定, 而汇群落的物种组成变化则高于源群落; 二是岛屿模型, 该模型中的群落(岛屿或小面积斑块)通过扩散相互联系在一起, 形成集合群落(Holyoak et al., 2005).一般而言, 扩散过程对群落构建的作用是受其他生态过程影响的, 尤其是选择过程和生态漂变(Ron et al., 2018). ...
... 关于对片段化生境中解构森林群落的研究, 主要集中在生物多样性丰富的热带地区, 比如亚马孙热带雨林(Ewers et al., 2017)和西双版纳热带雨林(Liu & Slik, 2014)等地区.这些森林之前均为连续森林, 随着人类活动, 如砍伐、农业种植、都市化等因素的影响, 逐渐被分割成具有不同隔离度和面积的陆地“森林生境岛屿”.受到边缘效应的影响, 面积较小的“生境岛屿”出现光照强度增加、水分限制加剧和随机干扰事件(如火干扰)频率增加等现象, 将影响不同斑块中物种的组成(Saunders et al., 1991; Ewers et al., 2017).环境过滤所引起的非生物选择过程是影响解构群落的主要生态过程, 该过程所引起的物种功能属性的改变和群落结构的变化在片段化后的10-15年尤为明显(Laurance et al., 2014).Das等(2017)对解构森林群落结构和物种组成变化的进一步研究表明, 集合群落中的物种筛选模型(即生物和非生物因子的选择过程)主要影响群落的结构和周转, 而空间连通性(即扩散限制)对群落构建的影响较小.在亚马孙和亚洲东南部片段化森林的研究中也发现, 边缘效应对森林群落动态具有重要的影响(Laurance, 2008; Laurance et al., 2011; Ruffell et al., 2016), 该过程更易导致大树的死亡从而令更多的先锋物种占据空出的生态位(Laurance et al., 2000; Benchimol & Peres, 2015), 故而可以认为这些成熟林群落片段化后所经历的演替过程为“逆行演替”.虽然经历逆行演替, 但是这些森林群落, 尤其是处于边缘的森林群落, 受不同干扰条件的影响, 在经过长期的演替后并不一定会处于演替早期时的群落状态(Ewers et al., 2017). ...
... 野外控制实验也是检验片段化生境中群落构建过程非常有效的手段(Debinski & Holt, 2000; Resasco et al., 2017).Haddad等(2015)综述了目前世界上已有的针对片段化生境的控制实验(图1), 通过设置控制实验和长期观察, 对片段化生境中或岛屿上群落构建的过程进行了有效的检验(Wilson et al., 2016; Resasco et al., 2017).例如, 巴西的片段化森林动态实验是目前最大和时间最长的野外控制实验, 通过设置不同大小的森林斑块且控制斑块之间的隔离度差异, 分析森林斑块面积的大小对群落构建过程的影响(Laurance et al., 2011); 美国的Savannah河流廊道实验通过增加斑块间的隔离度, 且控制斑块的面积差异, 分析廊道效应(扩散)在群落构建过程中的作用(Haddad & Baum, 1999; Tewksbury et al., 2002); Kansas生境片段化实验则通过设置不同面积大小和隔离度的片段化斑块, 长期监测斑块内群落的动态变化, 研究生境片段化对群落构建和演替过程的影响(Holt et al., 1995); 另外, Alexander等(2012)利用此实验在种群水平上分析了种子扩散与斑块间连通性之间的关系, 在群落水平上分析了环境过滤和扩散限制的共同作用对群落演替的作用.然而, 除上述主要的实验外, 基于野外控制实验检验片段化生境中森林群落构建过程的研究仍然不足. ...
Rainforest fragmentation kills big trees 1 2000
... 关于对片段化生境中解构森林群落的研究, 主要集中在生物多样性丰富的热带地区, 比如亚马孙热带雨林(Ewers et al., 2017)和西双版纳热带雨林(Liu & Slik, 2014)等地区.这些森林之前均为连续森林, 随着人类活动, 如砍伐、农业种植、都市化等因素的影响, 逐渐被分割成具有不同隔离度和面积的陆地“森林生境岛屿”.受到边缘效应的影响, 面积较小的“生境岛屿”出现光照强度增加、水分限制加剧和随机干扰事件(如火干扰)频率增加等现象, 将影响不同斑块中物种的组成(Saunders et al., 1991; Ewers et al., 2017).环境过滤所引起的非生物选择过程是影响解构群落的主要生态过程, 该过程所引起的物种功能属性的改变和群落结构的变化在片段化后的10-15年尤为明显(Laurance et al., 2014).Das等(2017)对解构森林群落结构和物种组成变化的进一步研究表明, 集合群落中的物种筛选模型(即生物和非生物因子的选择过程)主要影响群落的结构和周转, 而空间连通性(即扩散限制)对群落构建的影响较小.在亚马孙和亚洲东南部片段化森林的研究中也发现, 边缘效应对森林群落动态具有重要的影响(Laurance, 2008; Laurance et al., 2011; Ruffell et al., 2016), 该过程更易导致大树的死亡从而令更多的先锋物种占据空出的生态位(Laurance et al., 2000; Benchimol & Peres, 2015), 故而可以认为这些成熟林群落片段化后所经历的演替过程为“逆行演替”.虽然经历逆行演替, 但是这些森林群落, 尤其是处于边缘的森林群落, 受不同干扰条件的影响, 在经过长期的演替后并不一定会处于演替早期时的群落状态(Ewers et al., 2017). ...
Rapid decay of tree-community composition in Amazonian forest fragments 1 2006
... 生境片段化(或破碎化, habitat fragmentation)是指原来连续的生境受自然和人为干扰的影响, 逐渐转变为隔离的且面积较小的斑块(Wilcove et al., 1986; Fahrig, 2003).人类干扰(如择伐、都市化、公路和水库建设等)对自然生态系统的破坏以及自然因素(如森林火灾、岛屿等)的影响, 造成全球森林植被呈片段化分布(Fahrig, 2003; Didham, 2010; Haddad et al., 2015; Taubert et al., 2018; Liu et al., 2019a).从全球森林的分布来看, 世界上大部分森林斑块的面积小于10 hm2 (Haddad et al., 2015).森林面积的丧失、隔离度的增加和边缘效应的加剧将显著影响片段化生境中森林群落的结构(Laurance et al., 2006)和演替动态(Benchimol & Peres, 2015; Liu et al., 2019b). ...
Beta diversity as the variance of community data: Dissimilarity coefficients and partitioning 1 2013
... 影响群落构建的生态过程可以单独或共同作用于局域群落内物种的组成和个体多度, 进而影响群落间的β多样性(Myers & LaManna, 2016).通常使用方差分解的方法探讨空间因子(扩散限制)、环境因子(选择过程)和随机过程(生态漂变)对群落间β多样性的相对作用(Legendre & de Cáceres, 2013; 彭思羿等, 2014; Liu et al., 2018; Mori et al., 2018; 刘翔宇等, 2019).例如, 刘翔宇等(2019)发现中国东部海洋岛屿间的距离和岛屿距大陆的距离是影响群落间物种组成的主要因子, 即扩散限制过程可能是影响海洋岛屿上物种组成的主要因子. ...
Partitioning beta diversity in a subtropical broad-leaved forest of China 2 2009
Conceptual cartoon illustrating processes in forest community assembly in fragmented habitats. Redrawn from <xref ref-type="bibr" rid="b46">HilleRisLambers <i>et al</i>. (2012)</xref> and <xref ref-type="bibr" rid="b133">Vellend (2016)</xref>. Different letters denote different species; circles of different sizes denote different size patches; different fan-shaped areas in circles denote the relative abundance of each species in one community.Fig. 21.1 扩散 扩散是有机体或其繁殖体离开出生地后的迁移或传播, 该过程对群落构建的影响主要依赖于扩散者产生群落(源群落)和抵达群落(汇群落)的大小和物种组成(Holyoak et al., 2005).与连续森林相比, 存在隔离的森林斑块将限制物种在斑块间的自由扩散(如种子传播), 即群落间物种的扩散限制.根据源群落和汇群落的相对大小, 片段化生境中物种的扩散可以分为两种情形: 一是大陆-岛屿模型, 该模型中将扩散假定为单向扩散, 由无限大的源群落(大陆或大面积斑块)向一个或多个小的、隔离的汇群落(岛屿或小面积斑块)中扩散, 源群落的物种组成相对恒定, 而汇群落的物种组成变化则高于源群落; 二是岛屿模型, 该模型中的群落(岛屿或小面积斑块)通过扩散相互联系在一起, 形成集合群落(Holyoak et al., 2005).一般而言, 扩散过程对群落构建的作用是受其他生态过程影响的, 尤其是选择过程和生态漂变(Ron et al., 2018). ...
Conceptual cartoon illustrating processes in forest community assembly in fragmented habitats. Redrawn from <xref ref-type="bibr" rid="b46">HilleRisLambers <i>et al</i>. (2012)</xref> and <xref ref-type="bibr" rid="b133">Vellend (2016)</xref>. Different letters denote different species; circles of different sizes denote different size patches; different fan-shaped areas in circles denote the relative abundance of each species in one community.Fig. 21.1 扩散 扩散是有机体或其繁殖体离开出生地后的迁移或传播, 该过程对群落构建的影响主要依赖于扩散者产生群落(源群落)和抵达群落(汇群落)的大小和物种组成(Holyoak et al., 2005).与连续森林相比, 存在隔离的森林斑块将限制物种在斑块间的自由扩散(如种子传播), 即群落间物种的扩散限制.根据源群落和汇群落的相对大小, 片段化生境中物种的扩散可以分为两种情形: 一是大陆-岛屿模型, 该模型中将扩散假定为单向扩散, 由无限大的源群落(大陆或大面积斑块)向一个或多个小的、隔离的汇群落(岛屿或小面积斑块)中扩散, 源群落的物种组成相对恒定, 而汇群落的物种组成变化则高于源群落; 二是岛屿模型, 该模型中的群落(岛屿或小面积斑块)通过扩散相互联系在一起, 形成集合群落(Holyoak et al., 2005).一般而言, 扩散过程对群落构建的作用是受其他生态过程影响的, 尤其是选择过程和生态漂变(Ron et al., 2018). ...
The metacommunity concept: A framework for multi-scale community ecology 2 2004
Conceptual cartoon illustrating processes in forest community assembly in fragmented habitats. Redrawn from <xref ref-type="bibr" rid="b46">HilleRisLambers <i>et al</i>. (2012)</xref> and <xref ref-type="bibr" rid="b133">Vellend (2016)</xref>. Different letters denote different species; circles of different sizes denote different size patches; different fan-shaped areas in circles denote the relative abundance of each species in one community.Fig. 21.1 扩散 扩散是有机体或其繁殖体离开出生地后的迁移或传播, 该过程对群落构建的影响主要依赖于扩散者产生群落(源群落)和抵达群落(汇群落)的大小和物种组成(Holyoak et al., 2005).与连续森林相比, 存在隔离的森林斑块将限制物种在斑块间的自由扩散(如种子传播), 即群落间物种的扩散限制.根据源群落和汇群落的相对大小, 片段化生境中物种的扩散可以分为两种情形: 一是大陆-岛屿模型, 该模型中将扩散假定为单向扩散, 由无限大的源群落(大陆或大面积斑块)向一个或多个小的、隔离的汇群落(岛屿或小面积斑块)中扩散, 源群落的物种组成相对恒定, 而汇群落的物种组成变化则高于源群落; 二是岛屿模型, 该模型中的群落(岛屿或小面积斑块)通过扩散相互联系在一起, 形成集合群落(Holyoak et al., 2005).一般而言, 扩散过程对群落构建的作用是受其他生态过程影响的, 尤其是选择过程和生态漂变(Ron et al., 2018). ...
... ; Canada: Lindo et al., 2012); The Thousand Island Lake Experiments (China)(Yu et al., 2012; Liu et al., 2020); The Stability of Altered Forest Ecosystems (SAFE) Project (Malaysia)(Ewers et al., 2011); Metatron experiment (France)(Legrand et al., 2012). Fig. 1 10.17521/cjpe.2019.0155.F002
Conceptual cartoon illustrating processes in forest community assembly in fragmented habitats. Redrawn from <xref ref-type="bibr" rid="b46">HilleRisLambers <i>et al</i>. (2012)</xref> and <xref ref-type="bibr" rid="b133">Vellend (2016)</xref>. Different letters denote different species; circles of different sizes denote different size patches; different fan-shaped areas in circles denote the relative abundance of each species in one community.Fig. 21.1 扩散 扩散是有机体或其繁殖体离开出生地后的迁移或传播, 该过程对群落构建的影响主要依赖于扩散者产生群落(源群落)和抵达群落(汇群落)的大小和物种组成(Holyoak et al., 2005).与连续森林相比, 存在隔离的森林斑块将限制物种在斑块间的自由扩散(如种子传播), 即群落间物种的扩散限制.根据源群落和汇群落的相对大小, 片段化生境中物种的扩散可以分为两种情形: 一是大陆-岛屿模型, 该模型中将扩散假定为单向扩散, 由无限大的源群落(大陆或大面积斑块)向一个或多个小的、隔离的汇群落(岛屿或小面积斑块)中扩散, 源群落的物种组成相对恒定, 而汇群落的物种组成变化则高于源群落; 二是岛屿模型, 该模型中的群落(岛屿或小面积斑块)通过扩散相互联系在一起, 形成集合群落(Holyoak et al., 2005).一般而言, 扩散过程对群落构建的作用是受其他生态过程影响的, 尤其是选择过程和生态漂变(Ron et al., 2018). ...
Forest fragmentation in China and its effect on biodiversity 2 2019
... 生境片段化(或破碎化, habitat fragmentation)是指原来连续的生境受自然和人为干扰的影响, 逐渐转变为隔离的且面积较小的斑块(Wilcove et al., 1986; Fahrig, 2003).人类干扰(如择伐、都市化、公路和水库建设等)对自然生态系统的破坏以及自然因素(如森林火灾、岛屿等)的影响, 造成全球森林植被呈片段化分布(Fahrig, 2003; Didham, 2010; Haddad et al., 2015; Taubert et al., 2018; Liu et al., 2019a).从全球森林的分布来看, 世界上大部分森林斑块的面积小于10 hm2 (Haddad et al., 2015).森林面积的丧失、隔离度的增加和边缘效应的加剧将显著影响片段化生境中森林群落的结构(Laurance et al., 2006)和演替动态(Benchimol & Peres, 2015; Liu et al., 2019b). ...
... 随着森林植被的恢复, 虽然森林总覆盖面积将处于增加趋势, 但森林的片段化程度仍可能逐年增加(Kozak et al., 2018; Liu et al., 2019a).通过对片段化森林群落构建过程的研究和理论整合, 我们可以更好地理解不同生态过程对群落内物种组成和多样性的贡献, 从而更加有效地指导片段化生境中森林群落的恢复和对生物多样性的保护, 对于后续的森林管理、政策制定、生态系统服务功能提升和物种保护等均具有非常重要的意义. ...
Larger fragments have more late-successional species of woody plants than smaller fragments after 50 years of secondary succession 4 2019
... 生境片段化(或破碎化, habitat fragmentation)是指原来连续的生境受自然和人为干扰的影响, 逐渐转变为隔离的且面积较小的斑块(Wilcove et al., 1986; Fahrig, 2003).人类干扰(如择伐、都市化、公路和水库建设等)对自然生态系统的破坏以及自然因素(如森林火灾、岛屿等)的影响, 造成全球森林植被呈片段化分布(Fahrig, 2003; Didham, 2010; Haddad et al., 2015; Taubert et al., 2018; Liu et al., 2019a).从全球森林的分布来看, 世界上大部分森林斑块的面积小于10 hm2 (Haddad et al., 2015).森林面积的丧失、隔离度的增加和边缘效应的加剧将显著影响片段化生境中森林群落的结构(Laurance et al., 2006)和演替动态(Benchimol & Peres, 2015; Liu et al., 2019b). ...
... 不论控制实验还是野外观察实验, 根据被片段化前后森林所处的状态以及片段化后森林的演替状态, 可以把片段化森林分为两大群落类型: 解构群落和重构群落(Wilson et al., 2016; Collins et al., 2017).解构群落指处于演替后期的老龄林或成熟林被片段化后形成的森林群落, 出现逆行演替现象, 在演替过程中伴随着物种减少, 如中国西双版纳和巴西亚马孙的热带雨林中形成的片段化森林(Liu & Slik, 2014).重构群落是指在片段化生境中通过进展演替而重新构建形成的次生林群落, 处于进展演替过程之中, 一般处于演替的前期阶段, 群落内的优势种主要为一些阳性、生长快、扩散能力强的先锋树种(Baldeck et al., 2016; Liu et al., 2019b).解构群落和重构群落中存在的定殖和灭绝过程将促使群落内的物种组成和多样性发生改变, 然而受选择、生态漂变等过程的影响, 重构群落中定殖的物种组成和数量与解构群落中易于灭绝的物种组成和数量存在差异(Zavaleta et al., 2009).已有研究证实, 片段化后植物群落的长期动态变化, 如物种β多样性的变化, 将受到群落起始状态的影响(Collins et al., 2017), 即优先效应的作用(Fukami, 2015).因此, 依赖于森林群落起始状态的不同, 生境片段化对群落构建过程的影响可能也会不同(图3), 但以往的研究通常忽视了群落的起始状态.已有研究主要关注原来连续的老龄林或成熟林被片段化后形成的解构群落, 对片段生境中重构群落的构建过程关注较少(Debinski & Holt, 2000; Wilson et al., 2016; Liu et al., 2019b). ...
... ; Liu et al., 2019b). ...
... 迄今关于片段化生境中重构森林群落的研究仍然十分缺乏.我们对片段化生境中森林的重新构建过程可以作如下预测: 1)重构群落演替的起始阶段, 空间的隔离作用将比斑块面积的大小更加重要, 因短期的干扰或生境质量的改变, 空间的隔离将阻止物种占用适合的生境, 导致种子传播或者强风等随机事件决定了物种的抵达和在斑块上的定殖(Walker et al., 2010).例如, Kadmon和Pulliam (1995)发现在新近皆伐的湖泊岛屿上空间的隔离增加了风力传播物种的比例, Whittaker等(1997)发现在新近形成的火山岛屿上, 物种早期阶段的定殖可以根据其扩散性状进行预测, 不过在具有近60年片段化历史的千岛湖, 岛屿面积是影响植物群落物种组成的主要因素, 而非空间的隔离(Yu et al., 2012; Baldeck et al., 2016).2)当某一物种扩散到某一斑块后, 斑块内已有的种子/幼苗库, 会对后续扩散来的物种的定殖产生影响, 即优先效应, 该效应与决定性过程和随机过程会共同影响片段化生境中群落的构建过程(Fukami, 2015).3)物种抵达相应的斑块之后, 斑块内的生境异质性、种间/种内关系和生态漂变等过程将共同决定物种定殖成功的概率和物种组成, 如筛选个体生长快、木质密度低以及喜阳的先锋物种(Baldeck et al., 2016; Liu et al., 2019b, 2020), 或具有菌根的植物物种等(Delavaux et al., 2019).Baynes等(2016)通过比较砍伐后重构的片段化森林群落与连续森林群落内的物种多样性, 发现重构的森林群落中反而具有更多的稀有种以及较高的β多样性, 生态漂变过程也会影响片段化森林群落的重构. ...
The distribution of plants and seed dispersers in response to habitat fragmentation in an artificial island archipelago 1 2019
... 在片段化生境中, 根据群落内物种的某一类性状, 如与种群扩散、建立和维持等相关的植物生活史性状(Zambrano et al., 2019), 结合不同大小斑块内的生境条件和隔离度等对群落内功能特征收敛或发散程度的影响, 也能有助于我们理解选择过程或扩散过程在群落构建中的作用(?ckinger et al., 2010; Marini et al., 2012; Jiang et al., 2018).例如, 结合种子的功能特征, 可以分析扩散限制作用在群落构建过程中的作用(Negoita et al., 2016; Favre-Bac et al., 2017).有研究发现在越大的岛屿上小种子植物比例更高(种子特征的收敛), 可能是由于大岛上小种子扩散者(鸟类)的比例更高所引起的(Liu et al., 2019c). ...
Forest fragment spatial distribution matters for tropical tree conservation 2 2014
... 不论控制实验还是野外观察实验, 根据被片段化前后森林所处的状态以及片段化后森林的演替状态, 可以把片段化森林分为两大群落类型: 解构群落和重构群落(Wilson et al., 2016; Collins et al., 2017).解构群落指处于演替后期的老龄林或成熟林被片段化后形成的森林群落, 出现逆行演替现象, 在演替过程中伴随着物种减少, 如中国西双版纳和巴西亚马孙的热带雨林中形成的片段化森林(Liu & Slik, 2014).重构群落是指在片段化生境中通过进展演替而重新构建形成的次生林群落, 处于进展演替过程之中, 一般处于演替的前期阶段, 群落内的优势种主要为一些阳性、生长快、扩散能力强的先锋树种(Baldeck et al., 2016; Liu et al., 2019b).解构群落和重构群落中存在的定殖和灭绝过程将促使群落内的物种组成和多样性发生改变, 然而受选择、生态漂变等过程的影响, 重构群落中定殖的物种组成和数量与解构群落中易于灭绝的物种组成和数量存在差异(Zavaleta et al., 2009).已有研究证实, 片段化后植物群落的长期动态变化, 如物种β多样性的变化, 将受到群落起始状态的影响(Collins et al., 2017), 即优先效应的作用(Fukami, 2015).因此, 依赖于森林群落起始状态的不同, 生境片段化对群落构建过程的影响可能也会不同(图3), 但以往的研究通常忽视了群落的起始状态.已有研究主要关注原来连续的老龄林或成熟林被片段化后形成的解构群落, 对片段生境中重构群落的构建过程关注较少(Debinski & Holt, 2000; Wilson et al., 2016; Liu et al., 2019b). ...
... 关于对片段化生境中解构森林群落的研究, 主要集中在生物多样性丰富的热带地区, 比如亚马孙热带雨林(Ewers et al., 2017)和西双版纳热带雨林(Liu & Slik, 2014)等地区.这些森林之前均为连续森林, 随着人类活动, 如砍伐、农业种植、都市化等因素的影响, 逐渐被分割成具有不同隔离度和面积的陆地“森林生境岛屿”.受到边缘效应的影响, 面积较小的“生境岛屿”出现光照强度增加、水分限制加剧和随机干扰事件(如火干扰)频率增加等现象, 将影响不同斑块中物种的组成(Saunders et al., 1991; Ewers et al., 2017).环境过滤所引起的非生物选择过程是影响解构群落的主要生态过程, 该过程所引起的物种功能属性的改变和群落结构的变化在片段化后的10-15年尤为明显(Laurance et al., 2014).Das等(2017)对解构森林群落结构和物种组成变化的进一步研究表明, 集合群落中的物种筛选模型(即生物和非生物因子的选择过程)主要影响群落的结构和周转, 而空间连通性(即扩散限制)对群落构建的影响较小.在亚马孙和亚洲东南部片段化森林的研究中也发现, 边缘效应对森林群落动态具有重要的影响(Laurance, 2008; Laurance et al., 2011; Ruffell et al., 2016), 该过程更易导致大树的死亡从而令更多的先锋物种占据空出的生态位(Laurance et al., 2000; Benchimol & Peres, 2015), 故而可以认为这些成熟林群落片段化后所经历的演替过程为“逆行演替”.虽然经历逆行演替, 但是这些森林群落, 尤其是处于边缘的森林群落, 受不同干扰条件的影响, 在经过长期的演替后并不一定会处于演替早期时的群落状态(Ewers et al., 2017). ...
Environmental filtering underpins the island species-area relationship in a subtropical anthropogenic archipelago 3 2020
Conceptual cartoon illustrating processes in forest community assembly in fragmented habitats. Redrawn from <xref ref-type="bibr" rid="b46">HilleRisLambers <i>et al</i>. (2012)</xref> and <xref ref-type="bibr" rid="b133">Vellend (2016)</xref>. Different letters denote different species; circles of different sizes denote different size patches; different fan-shaped areas in circles denote the relative abundance of each species in one community.Fig. 21.1 扩散 扩散是有机体或其繁殖体离开出生地后的迁移或传播, 该过程对群落构建的影响主要依赖于扩散者产生群落(源群落)和抵达群落(汇群落)的大小和物种组成(Holyoak et al., 2005).与连续森林相比, 存在隔离的森林斑块将限制物种在斑块间的自由扩散(如种子传播), 即群落间物种的扩散限制.根据源群落和汇群落的相对大小, 片段化生境中物种的扩散可以分为两种情形: 一是大陆-岛屿模型, 该模型中将扩散假定为单向扩散, 由无限大的源群落(大陆或大面积斑块)向一个或多个小的、隔离的汇群落(岛屿或小面积斑块)中扩散, 源群落的物种组成相对恒定, 而汇群落的物种组成变化则高于源群落; 二是岛屿模型, 该模型中的群落(岛屿或小面积斑块)通过扩散相互联系在一起, 形成集合群落(Holyoak et al., 2005).一般而言, 扩散过程对群落构建的作用是受其他生态过程影响的, 尤其是选择过程和生态漂变(Ron et al., 2018). ...
... ; Liu et al., 2020); The Stability of Altered Forest Ecosystems (SAFE) Project (Malaysia)(Ewers et al., 2011); Metatron experiment (France)(Legrand et al., 2012). Fig. 1 10.17521/cjpe.2019.0155.F002
Conceptual cartoon illustrating processes in forest community assembly in fragmented habitats. Redrawn from <xref ref-type="bibr" rid="b46">HilleRisLambers <i>et al</i>. (2012)</xref> and <xref ref-type="bibr" rid="b133">Vellend (2016)</xref>. Different letters denote different species; circles of different sizes denote different size patches; different fan-shaped areas in circles denote the relative abundance of each species in one community.Fig. 21.1 扩散 扩散是有机体或其繁殖体离开出生地后的迁移或传播, 该过程对群落构建的影响主要依赖于扩散者产生群落(源群落)和抵达群落(汇群落)的大小和物种组成(Holyoak et al., 2005).与连续森林相比, 存在隔离的森林斑块将限制物种在斑块间的自由扩散(如种子传播), 即群落间物种的扩散限制.根据源群落和汇群落的相对大小, 片段化生境中物种的扩散可以分为两种情形: 一是大陆-岛屿模型, 该模型中将扩散假定为单向扩散, 由无限大的源群落(大陆或大面积斑块)向一个或多个小的、隔离的汇群落(岛屿或小面积斑块)中扩散, 源群落的物种组成相对恒定, 而汇群落的物种组成变化则高于源群落; 二是岛屿模型, 该模型中的群落(岛屿或小面积斑块)通过扩散相互联系在一起, 形成集合群落(Holyoak et al., 2005).一般而言, 扩散过程对群落构建的作用是受其他生态过程影响的, 尤其是选择过程和生态漂变(Ron et al., 2018). ...
... 迄今关于片段化生境中重构森林群落的研究仍然十分缺乏.我们对片段化生境中森林的重新构建过程可以作如下预测: 1)重构群落演替的起始阶段, 空间的隔离作用将比斑块面积的大小更加重要, 因短期的干扰或生境质量的改变, 空间的隔离将阻止物种占用适合的生境, 导致种子传播或者强风等随机事件决定了物种的抵达和在斑块上的定殖(Walker et al., 2010).例如, Kadmon和Pulliam (1995)发现在新近皆伐的湖泊岛屿上空间的隔离增加了风力传播物种的比例, Whittaker等(1997)发现在新近形成的火山岛屿上, 物种早期阶段的定殖可以根据其扩散性状进行预测, 不过在具有近60年片段化历史的千岛湖, 岛屿面积是影响植物群落物种组成的主要因素, 而非空间的隔离(Yu et al., 2012; Baldeck et al., 2016).2)当某一物种扩散到某一斑块后, 斑块内已有的种子/幼苗库, 会对后续扩散来的物种的定殖产生影响, 即优先效应, 该效应与决定性过程和随机过程会共同影响片段化生境中群落的构建过程(Fukami, 2015).3)物种抵达相应的斑块之后, 斑块内的生境异质性、种间/种内关系和生态漂变等过程将共同决定物种定殖成功的概率和物种组成, 如筛选个体生长快、木质密度低以及喜阳的先锋物种(Baldeck et al., 2016; Liu et al., 2019b, 2020), 或具有菌根的植物物种等(Delavaux et al., 2019).Baynes等(2016)通过比较砍伐后重构的片段化森林群落与连续森林群落内的物种多样性, 发现重构的森林群落中反而具有更多的稀有种以及较高的β多样性, 生态漂变过程也会影响片段化森林群落的重构. ...
Disentangling the drivers of taxonomic and phylogenetic beta diversities in disturbed and undisturbed subtropical forests 3 2016
High beta diversity among small islands is due to environmental heterogeneity rather than ecological drift 4 2018
... 集合群落理论包含中性过程、斑块动态、物种筛选和质量效应4种理论框架(Leibold et al., 2004; Holyoak et al., 2005)(附件I), 在ETIB和中性理论的基础上, 将选择过程考虑在内, 阐述了片段化生境中群落构建的基本过程.Vellend (2016)所提出的生态群落构建的4个高阶过程中的生态漂变类似于集合群落模型中的中性过程, 扩散和选择过程分别类似于斑块动态和物种筛选.虽然物种形成在集合群落理论中没有被明确排除在外, 但在多数情况下, 集合群落理论中并没有考虑该过程.在Vellend (2016)群落生态学理论提出后, 已有部分研究在片段化/岛屿生境中检验不同的生态过程在群落构建中的相对作用, 如选择过程和生态漂变过程的相对作用(Liu et al., 2018)、扩散对选择和生态漂变过程相对作用的影响(Ron et al., 2018)等.另外, 通过扩展ETIB理论, 考虑物种间功能特征的差异及其所受面积和隔离度的影响(Jacquet et al., 2017), 以及物种功能性状特征分布与环境因子间的关系, 逐渐成为检验片段化生境中群落构建过程的重要手段(Zambrano et al., 2019).然而, 针对不同生态过程如何影响片段化森林群落构建的研究仍然十分缺乏, 尤其是对于扩散和物种形成过程的相关检验. ...
... 影响群落构建的生态过程可以单独或共同作用于局域群落内物种的组成和个体多度, 进而影响群落间的β多样性(Myers & LaManna, 2016).通常使用方差分解的方法探讨空间因子(扩散限制)、环境因子(选择过程)和随机过程(生态漂变)对群落间β多样性的相对作用(Legendre & de Cáceres, 2013; 彭思羿等, 2014; Liu et al., 2018; Mori et al., 2018; 刘翔宇等, 2019).例如, 刘翔宇等(2019)发现中国东部海洋岛屿间的距离和岛屿距大陆的距离是影响群落间物种组成的主要因子, 即扩散限制过程可能是影响海洋岛屿上物种组成的主要因子. ...
... 根据片段化森林构建的主要生态过程(扩散限制、选择过程和生态漂变), 如果所有小斑块中的物种, 随机来源于大斑块中的物种库(源-库动态), 且斑块间物种呈嵌套分布, 即生态漂变是形成森林斑块间物种组成差异的主要过程, 此时保护一个大的保护区将变得更为有意义.然而, 已发现显著的嵌套式分布格局仅在9%的系统中出现, 而反嵌套的物种分布格局在16%的系统中出现(Matthews et al., 2015a).如果小斑块之间存在较高的生境异质性, 不同的斑块上选择不同的物种, 即小面积森林斑块之间存在较高的β多样性(Liu et al., 2018), 那么小面积的斑块也具有保护价值, 不能被忽略(Tscharntke et al., 2002; Rösch et al., 2015).基于全球尺度的分析, 也发现小面积的斑块具有一定的保护价值(Wintle et al., 2019).因此, 在植被恢复和管理中, 如果仅能在多个小面积的斑块上进行植被恢复, 应尽可能地增加斑块之间的生境异质性以及物种组成的差异性, 以维持更高的区域物种多样性. ...
中国东部海岛维管植物的beta多样性及其驱动因素 2 2019
... 影响群落构建的生态过程可以单独或共同作用于局域群落内物种的组成和个体多度, 进而影响群落间的β多样性(Myers & LaManna, 2016).通常使用方差分解的方法探讨空间因子(扩散限制)、环境因子(选择过程)和随机过程(生态漂变)对群落间β多样性的相对作用(Legendre & de Cáceres, 2013; 彭思羿等, 2014; Liu et al., 2018; Mori et al., 2018; 刘翔宇等, 2019).例如, 刘翔宇等(2019)发现中国东部海洋岛屿间的距离和岛屿距大陆的距离是影响群落间物种组成的主要因子, 即扩散限制过程可能是影响海洋岛屿上物种组成的主要因子. ...
... 影响群落构建的生态过程可以单独或共同作用于局域群落内物种的组成和个体多度, 进而影响群落间的β多样性(Myers & LaManna, 2016).通常使用方差分解的方法探讨空间因子(扩散限制)、环境因子(选择过程)和随机过程(生态漂变)对群落间β多样性的相对作用(Legendre & de Cáceres, 2013; 彭思羿等, 2014; Liu et al., 2018; Mori et al., 2018; 刘翔宇等, 2019).例如, 刘翔宇等(2019)发现中国东部海洋岛屿间的距离和岛屿距大陆的距离是影响群落间物种组成的主要因子, 即扩散限制过程可能是影响海洋岛屿上物种组成的主要因子. ...
Conceptual cartoon illustrating processes in forest community assembly in fragmented habitats. Redrawn from <xref ref-type="bibr" rid="b46">HilleRisLambers <i>et al</i>. (2012)</xref> and <xref ref-type="bibr" rid="b133">Vellend (2016)</xref>. Different letters denote different species; circles of different sizes denote different size patches; different fan-shaped areas in circles denote the relative abundance of each species in one community.Fig. 21.1 扩散 扩散是有机体或其繁殖体离开出生地后的迁移或传播, 该过程对群落构建的影响主要依赖于扩散者产生群落(源群落)和抵达群落(汇群落)的大小和物种组成(Holyoak et al., 2005).与连续森林相比, 存在隔离的森林斑块将限制物种在斑块间的自由扩散(如种子传播), 即群落间物种的扩散限制.根据源群落和汇群落的相对大小, 片段化生境中物种的扩散可以分为两种情形: 一是大陆-岛屿模型, 该模型中将扩散假定为单向扩散, 由无限大的源群落(大陆或大面积斑块)向一个或多个小的、隔离的汇群落(岛屿或小面积斑块)中扩散, 源群落的物种组成相对恒定, 而汇群落的物种组成变化则高于源群落; 二是岛屿模型, 该模型中的群落(岛屿或小面积斑块)通过扩散相互联系在一起, 形成集合群落(Holyoak et al., 2005).一般而言, 扩散过程对群落构建的作用是受其他生态过程影响的, 尤其是选择过程和生态漂变(Ron et al., 2018). ...
... ); Wog-Wog Habitat Fragmentation Experiment (Wog-Wog; Australia)(Margules, 1992); Savannah River Site Corridor Experiment (SRS Corridor Experiment; USA)(Haddad & Baum, 1999; Tewksbury et al., 2002); Moss Fragmentation Experiments (UK: Gonzalez et al. 1998; Canada: Lindo et al., 2012); The Thousand Island Lake Experiments (China)(Yu et al., 2012; Liu et al., 2020); The Stability of Altered Forest Ecosystems (SAFE) Project (Malaysia)(Ewers et al., 2011); Metatron experiment (France)(Legrand et al., 2012). Fig. 1 10.17521/cjpe.2019.0155.F002
Conceptual cartoon illustrating processes in forest community assembly in fragmented habitats. Redrawn from <xref ref-type="bibr" rid="b46">HilleRisLambers <i>et al</i>. (2012)</xref> and <xref ref-type="bibr" rid="b133">Vellend (2016)</xref>. Different letters denote different species; circles of different sizes denote different size patches; different fan-shaped areas in circles denote the relative abundance of each species in one community.Fig. 21.1 扩散 扩散是有机体或其繁殖体离开出生地后的迁移或传播, 该过程对群落构建的影响主要依赖于扩散者产生群落(源群落)和抵达群落(汇群落)的大小和物种组成(Holyoak et al., 2005).与连续森林相比, 存在隔离的森林斑块将限制物种在斑块间的自由扩散(如种子传播), 即群落间物种的扩散限制.根据源群落和汇群落的相对大小, 片段化生境中物种的扩散可以分为两种情形: 一是大陆-岛屿模型, 该模型中将扩散假定为单向扩散, 由无限大的源群落(大陆或大面积斑块)向一个或多个小的、隔离的汇群落(岛屿或小面积斑块)中扩散, 源群落的物种组成相对恒定, 而汇群落的物种组成变化则高于源群落; 二是岛屿模型, 该模型中的群落(岛屿或小面积斑块)通过扩散相互联系在一起, 形成集合群落(Holyoak et al., 2005).一般而言, 扩散过程对群落构建的作用是受其他生态过程影响的, 尤其是选择过程和生态漂变(Ron et al., 2018). ...
Traits related to species persistence and dispersal explain changes in plant communities subjected to habitat loss 1 2012
... 在片段化生境中, 根据群落内物种的某一类性状, 如与种群扩散、建立和维持等相关的植物生活史性状(Zambrano et al., 2019), 结合不同大小斑块内的生境条件和隔离度等对群落内功能特征收敛或发散程度的影响, 也能有助于我们理解选择过程或扩散过程在群落构建中的作用(?ckinger et al., 2010; Marini et al., 2012; Jiang et al., 2018).例如, 结合种子的功能特征, 可以分析扩散限制作用在群落构建过程中的作用(Negoita et al., 2016; Favre-Bac et al., 2017).有研究发现在越大的岛屿上小种子植物比例更高(种子特征的收敛), 可能是由于大岛上小种子扩散者(鸟类)的比例更高所引起的(Liu et al., 2019c). ...
A biogeographical perspective on species abundance distributions: Recent advances and opportunities for future research 1 2017
... 物种多度分布(SAD)描述了一个取样单元或群落内所有物种的个体多度分布状况, 是一个群落最基本的描述(McGill et al., 2007).通常, SAD通过物种序列多度图(RAD)进行可视化(Ulrich et al., 2010).已有大量的研究对群落内RAD的形状进行模型拟合, 如常见的模型有: 对数级数模型、对数正态模型等近30个模型(McGill et al., 2007).但是, 如何根据RAD曲线所拟合的模型反映群落的构建过程仍是研究的热点和难点问题.虽然仅通过对不同SAD模型的拟合尚无法准确地推测群落的构建过程, 但在地理尺度上, 通过分析不同环境因子和空间因子等对RAD形状的影响, 可以探讨扩散过程对群落构建的影响(Fattorini et al., 2016).在全球尺度上, Ulrich等(2016)通过对数级数模型和对数正态模型的拟合, 在纬度梯度上分析了扩散限制和环境过滤对群落构建的影响, 发现处于不同纬度的群落内扩散限制所产生的相对作用不同, 导致了低纬度地区群落内的RAD符合对数级数模型而高纬度地区群落内RAD更加符合对数正态模型.相似地, 在纬度梯度上利用非中性模型对森林群落的RAD进行拟合, 发现随纬度梯度的增加,环境过滤作用导致实际的SAD模型与中性模型的分化逐渐增加, 即中性过程对群落构建作用的影响减弱, 而非中性过程增加(Qiao et al., 2015a).Matthews等(2017)发现岛屿间的隔离程度可以改变岛屿上节肢动物的RAD的形状, 但关于岛屿参数如何影响森林群落的RAD尚缺乏研究.SAD不仅可以了解片段化生境中群落构建的过程, 也能够反映群落内稀有种和常见种的分布格局, 通过探讨片段化生境内物种多度的改变, 可应用于生物多样性的管理和应用生态学等方面(Matthews & Whittaker, 2015b).但在不同面积和隔离度的片段化森林中, 结合RAD形状的改变探讨群落构建过程的研究仍然不足. ...
Quantifying and interpreting nestedness in habitat islands: A synthetic analysis of multiple datasets 1 2015
... 根据片段化森林构建的主要生态过程(扩散限制、选择过程和生态漂变), 如果所有小斑块中的物种, 随机来源于大斑块中的物种库(源-库动态), 且斑块间物种呈嵌套分布, 即生态漂变是形成森林斑块间物种组成差异的主要过程, 此时保护一个大的保护区将变得更为有意义.然而, 已发现显著的嵌套式分布格局仅在9%的系统中出现, 而反嵌套的物种分布格局在16%的系统中出现(Matthews et al., 2015a).如果小斑块之间存在较高的生境异质性, 不同的斑块上选择不同的物种, 即小面积森林斑块之间存在较高的β多样性(Liu et al., 2018), 那么小面积的斑块也具有保护价值, 不能被忽略(Tscharntke et al., 2002; Rösch et al., 2015).基于全球尺度的分析, 也发现小面积的斑块具有一定的保护价值(Wintle et al., 2019).因此, 在植被恢复和管理中, 如果仅能在多个小面积的斑块上进行植被恢复, 应尽可能地增加斑块之间的生境异质性以及物种组成的差异性, 以维持更高的区域物种多样性. ...
On the species abundance distribution in applied ecology and biodiversity management 1 2015
... 物种多度分布(SAD)描述了一个取样单元或群落内所有物种的个体多度分布状况, 是一个群落最基本的描述(McGill et al., 2007).通常, SAD通过物种序列多度图(RAD)进行可视化(Ulrich et al., 2010).已有大量的研究对群落内RAD的形状进行模型拟合, 如常见的模型有: 对数级数模型、对数正态模型等近30个模型(McGill et al., 2007).但是, 如何根据RAD曲线所拟合的模型反映群落的构建过程仍是研究的热点和难点问题.虽然仅通过对不同SAD模型的拟合尚无法准确地推测群落的构建过程, 但在地理尺度上, 通过分析不同环境因子和空间因子等对RAD形状的影响, 可以探讨扩散过程对群落构建的影响(Fattorini et al., 2016).在全球尺度上, Ulrich等(2016)通过对数级数模型和对数正态模型的拟合, 在纬度梯度上分析了扩散限制和环境过滤对群落构建的影响, 发现处于不同纬度的群落内扩散限制所产生的相对作用不同, 导致了低纬度地区群落内的RAD符合对数级数模型而高纬度地区群落内RAD更加符合对数正态模型.相似地, 在纬度梯度上利用非中性模型对森林群落的RAD进行拟合, 发现随纬度梯度的增加,环境过滤作用导致实际的SAD模型与中性模型的分化逐渐增加, 即中性过程对群落构建作用的影响减弱, 而非中性过程增加(Qiao et al., 2015a).Matthews等(2017)发现岛屿间的隔离程度可以改变岛屿上节肢动物的RAD的形状, 但关于岛屿参数如何影响森林群落的RAD尚缺乏研究.SAD不仅可以了解片段化生境中群落构建的过程, 也能够反映群落内稀有种和常见种的分布格局, 通过探讨片段化生境内物种多度的改变, 可应用于生物多样性的管理和应用生态学等方面(Matthews & Whittaker, 2015b).但在不同面积和隔离度的片段化森林中, 结合RAD形状的改变探讨群落构建过程的研究仍然不足. ...
Comparative evaluation of experimental approaches to the study of habitat fragmentation effects 1 2002
... 所有自然种群多度的波动, 均受到环境的随机波动、种内和种间竞争以及随机过程等因子影响(Mutshinda et al., 2009).而群落的构建依赖于群落内各种群的动态变化, 即物种多度、物种定殖和灭绝的动态变化, 通过分析群落内物种组成或种群多度随时间的动态变化, 可以为群落的构建过程提供最直接的证据(McGarigal & Cushman, 2002; Ernest et al., 2008).Laurance等(2014)通过对比片段化和连续森林中年际间树木的死亡率和更新率, 发现环境过滤(如大气中CO2浓度)是影响群落构建的主要因子; Damschen等(2019)通过长期监测植物群落的定殖率和死亡率, 发现连接度高的片段化群落间能够通过提高定殖率和降低灭绝率而维持更高的物种多样性.另外, 通过比较分析一定时间内片段化森林与连续森林内物种组成/功能特征的周转、群落功能特征/谱系结构的收敛或发散, 也能揭示各生态过程在森林演替不同阶段的相对作用(Collins et al., 2017).利用不同大小或隔离度的群落内各种群的多度在时间上的波动趋势, 也可以分析生态漂变和选择过程的相对作用(Vellend, 2016). ...
Species abundance distributions: Moving beyond single prediction theories to integration within an ecological framework 2 2007
... 物种多度分布(SAD)描述了一个取样单元或群落内所有物种的个体多度分布状况, 是一个群落最基本的描述(McGill et al., 2007).通常, SAD通过物种序列多度图(RAD)进行可视化(Ulrich et al., 2010).已有大量的研究对群落内RAD的形状进行模型拟合, 如常见的模型有: 对数级数模型、对数正态模型等近30个模型(McGill et al., 2007).但是, 如何根据RAD曲线所拟合的模型反映群落的构建过程仍是研究的热点和难点问题.虽然仅通过对不同SAD模型的拟合尚无法准确地推测群落的构建过程, 但在地理尺度上, 通过分析不同环境因子和空间因子等对RAD形状的影响, 可以探讨扩散过程对群落构建的影响(Fattorini et al., 2016).在全球尺度上, Ulrich等(2016)通过对数级数模型和对数正态模型的拟合, 在纬度梯度上分析了扩散限制和环境过滤对群落构建的影响, 发现处于不同纬度的群落内扩散限制所产生的相对作用不同, 导致了低纬度地区群落内的RAD符合对数级数模型而高纬度地区群落内RAD更加符合对数正态模型.相似地, 在纬度梯度上利用非中性模型对森林群落的RAD进行拟合, 发现随纬度梯度的增加,环境过滤作用导致实际的SAD模型与中性模型的分化逐渐增加, 即中性过程对群落构建作用的影响减弱, 而非中性过程增加(Qiao et al., 2015a).Matthews等(2017)发现岛屿间的隔离程度可以改变岛屿上节肢动物的RAD的形状, 但关于岛屿参数如何影响森林群落的RAD尚缺乏研究.SAD不仅可以了解片段化生境中群落构建的过程, 也能够反映群落内稀有种和常见种的分布格局, 通过探讨片段化生境内物种多度的改变, 可应用于生物多样性的管理和应用生态学等方面(Matthews & Whittaker, 2015b).但在不同面积和隔离度的片段化森林中, 结合RAD形状的改变探讨群落构建过程的研究仍然不足. ...
β-diversity, community assembly, and ecosystem functioning 1 2018
... 影响群落构建的生态过程可以单独或共同作用于局域群落内物种的组成和个体多度, 进而影响群落间的β多样性(Myers & LaManna, 2016).通常使用方差分解的方法探讨空间因子(扩散限制)、环境因子(选择过程)和随机过程(生态漂变)对群落间β多样性的相对作用(Legendre & de Cáceres, 2013; 彭思羿等, 2014; Liu et al., 2018; Mori et al., 2018; 刘翔宇等, 2019).例如, 刘翔宇等(2019)发现中国东部海洋岛屿间的距离和岛屿距大陆的距离是影响群落间物种组成的主要因子, 即扩散限制过程可能是影响海洋岛屿上物种组成的主要因子. ...
Functional diversity measures: An overview of their redundancy and their ability to discriminate community assembly rules 1 2010
... 植物功能特征/功能性状指植物的生理、结构和行为特征, 能够指示物种对环境的适应能力, 影响物种在特定生境中的定殖和种间关系(Violle et al., 2007; Lindo et al., 2012).在过去近20年的研究中, 越来越多的生态学家将物种的功能特征与选择过程结合起来, 通过分析群落内物种功能特征的收敛或者发散, 探讨环境过滤或竞争作用引起的限制相似性在群落构建过程中的相对作用(Perronne et al., 2017).当环境过滤影响群落内的物种组成时, 如果具有某一类功能特征的物种更能适应环境而被保留, 此时群落内功能特征的分布表现为收敛(群落内物种间的功能特征更为相似).在小尺度上, 物种间功能特征属性越相似, 种间竞争将越激烈, 竞争排斥作用可能仅保留具有强竞争力的性状, 使得功能特征仍表现为收敛; 但当物种能够共存时, 此时限制相似性的作用(如群落内物种间的竞争, 天敌介导的密度依赖等过程)会致使功能特征值具有更宽的范围, 引起性状发散(非相似性功能特征), 即生态位分化(Kraft et al., 2008).目前, 已有多种量度群落功能特征的α多样性和β多样性的指数, 可以反映群落内物种功能性状的分布和多样性(Ackerly & Cornwell, 2007; Mouchet et al., 2010).因此, 基于群落内实际观察到的性状分布格局, 依赖于性状分布的零模型(随机过程, 即所有个体功能相同且物种随机共存), 可以衡量群落内物种功能特征是收敛的还是发散的, 进而探讨选择过程(环境过滤和种间竞争等)在群落构建中的作用(Kraft et al., 2008). ...
Community patterns in source-sink metacommunities 1 2003
... 所有自然种群多度的波动, 均受到环境的随机波动、种内和种间竞争以及随机过程等因子影响(Mutshinda et al., 2009).而群落的构建依赖于群落内各种群的动态变化, 即物种多度、物种定殖和灭绝的动态变化, 通过分析群落内物种组成或种群多度随时间的动态变化, 可以为群落的构建过程提供最直接的证据(McGarigal & Cushman, 2002; Ernest et al., 2008).Laurance等(2014)通过对比片段化和连续森林中年际间树木的死亡率和更新率, 发现环境过滤(如大气中CO2浓度)是影响群落构建的主要因子; Damschen等(2019)通过长期监测植物群落的定殖率和死亡率, 发现连接度高的片段化群落间能够通过提高定殖率和降低灭绝率而维持更高的物种多样性.另外, 通过比较分析一定时间内片段化森林与连续森林内物种组成/功能特征的周转、群落功能特征/谱系结构的收敛或发散, 也能揭示各生态过程在森林演替不同阶段的相对作用(Collins et al., 2017).利用不同大小或隔离度的群落内各种群的多度在时间上的波动趋势, 也可以分析生态漂变和选择过程的相对作用(Vellend, 2016). ...
The promise and pitfalls of β-diversity in ecology and conservation 1 2016
... 影响群落构建的生态过程可以单独或共同作用于局域群落内物种的组成和个体多度, 进而影响群落间的β多样性(Myers & LaManna, 2016).通常使用方差分解的方法探讨空间因子(扩散限制)、环境因子(选择过程)和随机过程(生态漂变)对群落间β多样性的相对作用(Legendre & de Cáceres, 2013; 彭思羿等, 2014; Liu et al., 2018; Mori et al., 2018; 刘翔宇等, 2019).例如, 刘翔宇等(2019)发现中国东部海洋岛屿间的距离和岛屿距大陆的距离是影响群落间物种组成的主要因子, 即扩散限制过程可能是影响海洋岛屿上物种组成的主要因子. ...
Isolation-driven functional assembly of plant communities on islands 1 2016
... 在片段化生境中, 根据群落内物种的某一类性状, 如与种群扩散、建立和维持等相关的植物生活史性状(Zambrano et al., 2019), 结合不同大小斑块内的生境条件和隔离度等对群落内功能特征收敛或发散程度的影响, 也能有助于我们理解选择过程或扩散过程在群落构建中的作用(?ckinger et al., 2010; Marini et al., 2012; Jiang et al., 2018).例如, 结合种子的功能特征, 可以分析扩散限制作用在群落构建过程中的作用(Negoita et al., 2016; Favre-Bac et al., 2017).有研究发现在越大的岛屿上小种子植物比例更高(种子特征的收敛), 可能是由于大岛上小种子扩散者(鸟类)的比例更高所引起的(Liu et al., 2019c). ...
群落构建的中性理论和生态位理论 1 2009
... 片段化生境中的森林群落, 并不是由某个生态过程单独作用形成的, 而是4个生态过程共同作用的结果.在探讨选择过程(或生态位过程, niche-based process)和中性过程(扩散限制+生态漂变)在群落构建中的相对作用时, 通常将未被环境因子(反映选择过程)和空间因子(反映扩散过程)所解释的部分归因于随机过程(生态漂变)和其他未测量的环境因子(Legendre et al., 2009; 牛克昌等, 2009; 陈磊等, 2014; Liu et al., 2016).部分研究也发现所测量的环境因子通常仅能解释亚热带森林群落物种组成差异的20%左右(Liu et al., 2016; Wu et al., 2017a).另外, Cottenie (2005)通过meta分析发现, 几乎50%的集合群落中物种组成的差异需要用环境和空间因子的共同作用所解释.对于片段化森林而言, 群落间的扩散可以增加有效群落的大小, 从而降低生态漂变的作用, 进而增加选择过程的相对作用(Ron et al., 2018).在岛屿模型中, 如果仅有生态漂变过程, 而没有选择和物种形成过程, 扩散将通过补偿生态漂变所产生的物种局域灭绝而增加局域群落内的物种多样性, 并且导致群落间的物种相似性增加(Wright, 1940); 但如果没有新物种形成或者扩散者迁入该集合群落, 该集合群落内的物种除保留一个物种外, 其余所有的物种将会由于生态漂变作用而从局域群落中灭绝; 如果同时存在生态漂变、物种形成和扩散过程, 但未受到选择过程的影响, 那么群落内物种的分布格局将决定于群落的大小、整个集合群落的大小和物种形成及扩散的速率(Hubbell, 2001).在大陆-岛屿模型中, 如果存在生态漂变过程, 而没有选择和物种形成过程, 扩散将促使岛屿上的物种组成与大陆趋同(Holyoak et al., 2005).另外, 竞争或者天敌侵害等选择过程可导致局域群落内的部分物种消失, 但如果这些物种具有较强的扩散能力, 可扩散到缺乏天敌或竞争者的群落中, 则会与其天敌或者竞争者共存于集合群落中.当物种在不同的局域群落中具有不同的适合度时, 扩散能力强的物种将会排除局域群落内其他的物种(Fukami, 2015).因此, 针对具有不同面积和隔离度的片段化森林群落, 受群落内物种属性(如扩散能力、竞争能力等)差异和种间相互作用等的影响, 在不同的空间和时间尺度上, 不同生态过程所起的相对作用是不同的. ...
群落构建的中性理论和生态位理论 1 2009
... 片段化生境中的森林群落, 并不是由某个生态过程单独作用形成的, 而是4个生态过程共同作用的结果.在探讨选择过程(或生态位过程, niche-based process)和中性过程(扩散限制+生态漂变)在群落构建中的相对作用时, 通常将未被环境因子(反映选择过程)和空间因子(反映扩散过程)所解释的部分归因于随机过程(生态漂变)和其他未测量的环境因子(Legendre et al., 2009; 牛克昌等, 2009; 陈磊等, 2014; Liu et al., 2016).部分研究也发现所测量的环境因子通常仅能解释亚热带森林群落物种组成差异的20%左右(Liu et al., 2016; Wu et al., 2017a).另外, Cottenie (2005)通过meta分析发现, 几乎50%的集合群落中物种组成的差异需要用环境和空间因子的共同作用所解释.对于片段化森林而言, 群落间的扩散可以增加有效群落的大小, 从而降低生态漂变的作用, 进而增加选择过程的相对作用(Ron et al., 2018).在岛屿模型中, 如果仅有生态漂变过程, 而没有选择和物种形成过程, 扩散将通过补偿生态漂变所产生的物种局域灭绝而增加局域群落内的物种多样性, 并且导致群落间的物种相似性增加(Wright, 1940); 但如果没有新物种形成或者扩散者迁入该集合群落, 该集合群落内的物种除保留一个物种外, 其余所有的物种将会由于生态漂变作用而从局域群落中灭绝; 如果同时存在生态漂变、物种形成和扩散过程, 但未受到选择过程的影响, 那么群落内物种的分布格局将决定于群落的大小、整个集合群落的大小和物种形成及扩散的速率(Hubbell, 2001).在大陆-岛屿模型中, 如果存在生态漂变过程, 而没有选择和物种形成过程, 扩散将促使岛屿上的物种组成与大陆趋同(Holyoak et al., 2005).另外, 竞争或者天敌侵害等选择过程可导致局域群落内的部分物种消失, 但如果这些物种具有较强的扩散能力, 可扩散到缺乏天敌或竞争者的群落中, 则会与其天敌或者竞争者共存于集合群落中.当物种在不同的局域群落中具有不同的适合度时, 扩散能力强的物种将会排除局域群落内其他的物种(Fukami, 2015).因此, 针对具有不同面积和隔离度的片段化森林群落, 受群落内物种属性(如扩散能力、竞争能力等)差异和种间相互作用等的影响, 在不同的空间和时间尺度上, 不同生态过程所起的相对作用是不同的. ...
Life-history traits predict species responses to habitat area and isolation: A cross-continental synthesis 1 2010
... 在片段化生境中, 根据群落内物种的某一类性状, 如与种群扩散、建立和维持等相关的植物生活史性状(Zambrano et al., 2019), 结合不同大小斑块内的生境条件和隔离度等对群落内功能特征收敛或发散程度的影响, 也能有助于我们理解选择过程或扩散过程在群落构建中的作用(?ckinger et al., 2010; Marini et al., 2012; Jiang et al., 2018).例如, 结合种子的功能特征, 可以分析扩散限制作用在群落构建过程中的作用(Negoita et al., 2016; Favre-Bac et al., 2017).有研究发现在越大的岛屿上小种子植物比例更高(种子特征的收敛), 可能是由于大岛上小种子扩散者(鸟类)的比例更高所引起的(Liu et al., 2019c). ...
Interactive effects of disturbance and dispersal on community assembly 1 2017
... 此外, 结合微宇宙的实验方法, 也可以构建模拟群落, 通过直接观察不同实验情景下的群落动态, 能够验证某一生态过程在群落构建中的作用(Davies et al., 2009; Altermatt et al., 2015; Ron et al., 2018).通常利用微生物群落, 通过配置菌落的大小(生态漂变)、菌群间隔离度(扩散)、环境胁迫或干扰(选择)、起始菌落的物种组成(优先效应)等, 验证生态群落的构建过程(Jiang & Patel, 2008; Ojima & Jiang, 2017).然而, 对于森林群落而言, 很难利用微宇宙的实验方法对群落构建过程进行检验, 由微宇宙实验获得的结论是否适用于片段化生境中的森林群落, 仍值得商榷. ...
Building up biogeography: Pattern to process 1 2018
... 影响群落构建的生态过程可以单独或共同作用于局域群落内物种的组成和个体多度, 进而影响群落间的β多样性(Myers & LaManna, 2016).通常使用方差分解的方法探讨空间因子(扩散限制)、环境因子(选择过程)和随机过程(生态漂变)对群落间β多样性的相对作用(Legendre & de Cáceres, 2013; 彭思羿等, 2014; Liu et al., 2018; Mori et al., 2018; 刘翔宇等, 2019).例如, 刘翔宇等(2019)发现中国东部海洋岛屿间的距离和岛屿距大陆的距离是影响群落间物种组成的主要因子, 即扩散限制过程可能是影响海洋岛屿上物种组成的主要因子. ...
千岛湖岛屿维管植物β多样性及其影响因素 1 2014
... 影响群落构建的生态过程可以单独或共同作用于局域群落内物种的组成和个体多度, 进而影响群落间的β多样性(Myers & LaManna, 2016).通常使用方差分解的方法探讨空间因子(扩散限制)、环境因子(选择过程)和随机过程(生态漂变)对群落间β多样性的相对作用(Legendre & de Cáceres, 2013; 彭思羿等, 2014; Liu et al., 2018; Mori et al., 2018; 刘翔宇等, 2019).例如, 刘翔宇等(2019)发现中国东部海洋岛屿间的距离和岛屿距大陆的距离是影响群落间物种组成的主要因子, 即扩散限制过程可能是影响海洋岛屿上物种组成的主要因子. ...
How to design trait-based analyses of community assembly mechanisms: Insights and guidelines from a literature review 1 2017
... 植物功能特征/功能性状指植物的生理、结构和行为特征, 能够指示物种对环境的适应能力, 影响物种在特定生境中的定殖和种间关系(Violle et al., 2007; Lindo et al., 2012).在过去近20年的研究中, 越来越多的生态学家将物种的功能特征与选择过程结合起来, 通过分析群落内物种功能特征的收敛或者发散, 探讨环境过滤或竞争作用引起的限制相似性在群落构建过程中的相对作用(Perronne et al., 2017).当环境过滤影响群落内的物种组成时, 如果具有某一类功能特征的物种更能适应环境而被保留, 此时群落内功能特征的分布表现为收敛(群落内物种间的功能特征更为相似).在小尺度上, 物种间功能特征属性越相似, 种间竞争将越激烈, 竞争排斥作用可能仅保留具有强竞争力的性状, 使得功能特征仍表现为收敛; 但当物种能够共存时, 此时限制相似性的作用(如群落内物种间的竞争, 天敌介导的密度依赖等过程)会致使功能特征值具有更宽的范围, 引起性状发散(非相似性功能特征), 即生态位分化(Kraft et al., 2008).目前, 已有多种量度群落功能特征的α多样性和β多样性的指数, 可以反映群落内物种功能性状的分布和多样性(Ackerly & Cornwell, 2007; Mouchet et al., 2010).因此, 基于群落内实际观察到的性状分布格局, 依赖于性状分布的零模型(随机过程, 即所有个体功能相同且物种随机共存), 可以衡量群落内物种功能特征是收敛的还是发散的, 进而探讨选择过程(环境过滤和种间竞争等)在群落构建中的作用(Kraft et al., 2008). ...
A latitudinal gradient in tree community assembly processes evidenced in Chinese forests 1 2015
... 物种多度分布(SAD)描述了一个取样单元或群落内所有物种的个体多度分布状况, 是一个群落最基本的描述(McGill et al., 2007).通常, SAD通过物种序列多度图(RAD)进行可视化(Ulrich et al., 2010).已有大量的研究对群落内RAD的形状进行模型拟合, 如常见的模型有: 对数级数模型、对数正态模型等近30个模型(McGill et al., 2007).但是, 如何根据RAD曲线所拟合的模型反映群落的构建过程仍是研究的热点和难点问题.虽然仅通过对不同SAD模型的拟合尚无法准确地推测群落的构建过程, 但在地理尺度上, 通过分析不同环境因子和空间因子等对RAD形状的影响, 可以探讨扩散过程对群落构建的影响(Fattorini et al., 2016).在全球尺度上, Ulrich等(2016)通过对数级数模型和对数正态模型的拟合, 在纬度梯度上分析了扩散限制和环境过滤对群落构建的影响, 发现处于不同纬度的群落内扩散限制所产生的相对作用不同, 导致了低纬度地区群落内的RAD符合对数级数模型而高纬度地区群落内RAD更加符合对数正态模型.相似地, 在纬度梯度上利用非中性模型对森林群落的RAD进行拟合, 发现随纬度梯度的增加,环境过滤作用导致实际的SAD模型与中性模型的分化逐渐增加, 即中性过程对群落构建作用的影响减弱, 而非中性过程增加(Qiao et al., 2015a).Matthews等(2017)发现岛屿间的隔离程度可以改变岛屿上节肢动物的RAD的形状, 但关于岛屿参数如何影响森林群落的RAD尚缺乏研究.SAD不仅可以了解片段化生境中群落构建的过程, 也能够反映群落内稀有种和常见种的分布格局, 通过探讨片段化生境内物种多度的改变, 可应用于生物多样性的管理和应用生态学等方面(Matthews & Whittaker, 2015b).但在不同面积和隔离度的片段化森林中, 结合RAD形状的改变探讨群落构建过程的研究仍然不足. ...
Beta diversity determinants in Badagongshan, a subtropical forest in central China 1 2015
Conceptual cartoon illustrating processes in forest community assembly in fragmented habitats. Redrawn from <xref ref-type="bibr" rid="b46">HilleRisLambers <i>et al</i>. (2012)</xref> and <xref ref-type="bibr" rid="b133">Vellend (2016)</xref>. Different letters denote different species; circles of different sizes denote different size patches; different fan-shaped areas in circles denote the relative abundance of each species in one community.Fig. 21.1 扩散 扩散是有机体或其繁殖体离开出生地后的迁移或传播, 该过程对群落构建的影响主要依赖于扩散者产生群落(源群落)和抵达群落(汇群落)的大小和物种组成(Holyoak et al., 2005).与连续森林相比, 存在隔离的森林斑块将限制物种在斑块间的自由扩散(如种子传播), 即群落间物种的扩散限制.根据源群落和汇群落的相对大小, 片段化生境中物种的扩散可以分为两种情形: 一是大陆-岛屿模型, 该模型中将扩散假定为单向扩散, 由无限大的源群落(大陆或大面积斑块)向一个或多个小的、隔离的汇群落(岛屿或小面积斑块)中扩散, 源群落的物种组成相对恒定, 而汇群落的物种组成变化则高于源群落; 二是岛屿模型, 该模型中的群落(岛屿或小面积斑块)通过扩散相互联系在一起, 形成集合群落(Holyoak et al., 2005).一般而言, 扩散过程对群落构建的作用是受其他生态过程影响的, 尤其是选择过程和生态漂变(Ron et al., 2018). ...
... Timeline of key events in fragmentation theory, landscape experiments, and discourse on application for conservation. Figure was redrawn from Resasco et al., (2017). Major experiments and projects and the corresponding references are as following: Biological Dynamics of Forest Fragments Project (BDFFP; Brazil)(Laurance et al., 2011); Kansas Fragmentation Experiment (USA)(Holt et al., 1995); Wog-Wog Habitat Fragmentation Experiment (Wog-Wog; Australia)(Margules, 1992); Savannah River Site Corridor Experiment (SRS Corridor Experiment; USA)(Haddad & Baum, 1999; Tewksbury et al., 2002); Moss Fragmentation Experiments (UK: Gonzalez et al. 1998; Canada: Lindo et al., 2012); The Thousand Island Lake Experiments (China)(Yu et al., 2012; Liu et al., 2020); The Stability of Altered Forest Ecosystems (SAFE) Project (Malaysia)(Ewers et al., 2011); Metatron experiment (France)(Legrand et al., 2012). Fig. 1 10.17521/cjpe.2019.0155.F002
Conceptual cartoon illustrating processes in forest community assembly in fragmented habitats. Redrawn from <xref ref-type="bibr" rid="b46">HilleRisLambers <i>et al</i>. (2012)</xref> and <xref ref-type="bibr" rid="b133">Vellend (2016)</xref>. Different letters denote different species; circles of different sizes denote different size patches; different fan-shaped areas in circles denote the relative abundance of each species in one community.Fig. 21.1 扩散 扩散是有机体或其繁殖体离开出生地后的迁移或传播, 该过程对群落构建的影响主要依赖于扩散者产生群落(源群落)和抵达群落(汇群落)的大小和物种组成(Holyoak et al., 2005).与连续森林相比, 存在隔离的森林斑块将限制物种在斑块间的自由扩散(如种子传播), 即群落间物种的扩散限制.根据源群落和汇群落的相对大小, 片段化生境中物种的扩散可以分为两种情形: 一是大陆-岛屿模型, 该模型中将扩散假定为单向扩散, 由无限大的源群落(大陆或大面积斑块)向一个或多个小的、隔离的汇群落(岛屿或小面积斑块)中扩散, 源群落的物种组成相对恒定, 而汇群落的物种组成变化则高于源群落; 二是岛屿模型, 该模型中的群落(岛屿或小面积斑块)通过扩散相互联系在一起, 形成集合群落(Holyoak et al., 2005).一般而言, 扩散过程对群落构建的作用是受其他生态过程影响的, 尤其是选择过程和生态漂变(Ron et al., 2018). ...
... 野外控制实验也是检验片段化生境中群落构建过程非常有效的手段(Debinski & Holt, 2000; Resasco et al., 2017).Haddad等(2015)综述了目前世界上已有的针对片段化生境的控制实验(图1), 通过设置控制实验和长期观察, 对片段化生境中或岛屿上群落构建的过程进行了有效的检验(Wilson et al., 2016; Resasco et al., 2017).例如, 巴西的片段化森林动态实验是目前最大和时间最长的野外控制实验, 通过设置不同大小的森林斑块且控制斑块之间的隔离度差异, 分析森林斑块面积的大小对群落构建过程的影响(Laurance et al., 2011); 美国的Savannah河流廊道实验通过增加斑块间的隔离度, 且控制斑块的面积差异, 分析廊道效应(扩散)在群落构建过程中的作用(Haddad & Baum, 1999; Tewksbury et al., 2002); Kansas生境片段化实验则通过设置不同面积大小和隔离度的片段化斑块, 长期监测斑块内群落的动态变化, 研究生境片段化对群落构建和演替过程的影响(Holt et al., 1995); 另外, Alexander等(2012)利用此实验在种群水平上分析了种子扩散与斑块间连通性之间的关系, 在群落水平上分析了环境过滤和扩散限制的共同作用对群落演替的作用.然而, 除上述主要的实验外, 基于野外控制实验检验片段化生境中森林群落构建过程的研究仍然不足. ...
... ; Resasco et al., 2017).例如, 巴西的片段化森林动态实验是目前最大和时间最长的野外控制实验, 通过设置不同大小的森林斑块且控制斑块之间的隔离度差异, 分析森林斑块面积的大小对群落构建过程的影响(Laurance et al., 2011); 美国的Savannah河流廊道实验通过增加斑块间的隔离度, 且控制斑块的面积差异, 分析廊道效应(扩散)在群落构建过程中的作用(Haddad & Baum, 1999; Tewksbury et al., 2002); Kansas生境片段化实验则通过设置不同面积大小和隔离度的片段化斑块, 长期监测斑块内群落的动态变化, 研究生境片段化对群落构建和演替过程的影响(Holt et al., 1995); 另外, Alexander等(2012)利用此实验在种群水平上分析了种子扩散与斑块间连通性之间的关系, 在群落水平上分析了环境过滤和扩散限制的共同作用对群落演替的作用.然而, 除上述主要的实验外, 基于野外控制实验检验片段化生境中森林群落构建过程的研究仍然不足. ...
Biological consequences of ecosystem fragmentation: A review 1 1991
... 关于对片段化生境中解构森林群落的研究, 主要集中在生物多样性丰富的热带地区, 比如亚马孙热带雨林(Ewers et al., 2017)和西双版纳热带雨林(Liu & Slik, 2014)等地区.这些森林之前均为连续森林, 随着人类活动, 如砍伐、农业种植、都市化等因素的影响, 逐渐被分割成具有不同隔离度和面积的陆地“森林生境岛屿”.受到边缘效应的影响, 面积较小的“生境岛屿”出现光照强度增加、水分限制加剧和随机干扰事件(如火干扰)频率增加等现象, 将影响不同斑块中物种的组成(Saunders et al., 1991; Ewers et al., 2017).环境过滤所引起的非生物选择过程是影响解构群落的主要生态过程, 该过程所引起的物种功能属性的改变和群落结构的变化在片段化后的10-15年尤为明显(Laurance et al., 2014).Das等(2017)对解构森林群落结构和物种组成变化的进一步研究表明, 集合群落中的物种筛选模型(即生物和非生物因子的选择过程)主要影响群落的结构和周转, 而空间连通性(即扩散限制)对群落构建的影响较小.在亚马孙和亚洲东南部片段化森林的研究中也发现, 边缘效应对森林群落动态具有重要的影响(Laurance, 2008; Laurance et al., 2011; Ruffell et al., 2016), 该过程更易导致大树的死亡从而令更多的先锋物种占据空出的生态位(Laurance et al., 2000; Benchimol & Peres, 2015), 故而可以认为这些成熟林群落片段化后所经历的演替过程为“逆行演替”.虽然经历逆行演替, 但是这些森林群落, 尤其是处于边缘的森林群落, 受不同干扰条件的影响, 在经过长期的演替后并不一定会处于演替早期时的群落状态(Ewers et al., 2017). ...
The necessity of multitrophic approaches in community ecology 1 2018
... 随着群落大小的变小, 生态漂变作用的增大, 会引起物种在局域群落内的消失(Fukami, 2010).扩散过程可以通过增加有效群落的大小, 来减少生态漂变的作用.因此, 在片段化生境中, 促进斑块间物种的扩散过程, 可以增加有效群落的大小, 减少生态漂变的作用(Ron et al., 2018).此外, 增加斑块间的连通性, 也能促进植食性昆虫等天敌和种子扩散者在片段化森林之间的扩散, 稳定食物链结构、避免物种之间级联效应的灭绝过程, 增加同种负密度制约效应的强度等, 进而提高生物多样性(Emer et al., 2018; Seibold et al., 2018).故而, 增加小面积森林斑块之间的连通性, 降低生态漂变的影响, 也是维持生物多样性的有效手段. ...
Global patterns of tropical forest fragmentation 1 2018
... 生境片段化(或破碎化, habitat fragmentation)是指原来连续的生境受自然和人为干扰的影响, 逐渐转变为隔离的且面积较小的斑块(Wilcove et al., 1986; Fahrig, 2003).人类干扰(如择伐、都市化、公路和水库建设等)对自然生态系统的破坏以及自然因素(如森林火灾、岛屿等)的影响, 造成全球森林植被呈片段化分布(Fahrig, 2003; Didham, 2010; Haddad et al., 2015; Taubert et al., 2018; Liu et al., 2019a).从全球森林的分布来看, 世界上大部分森林斑块的面积小于10 hm2 (Haddad et al., 2015).森林面积的丧失、隔离度的增加和边缘效应的加剧将显著影响片段化生境中森林群落的结构(Laurance et al., 2006)和演替动态(Benchimol & Peres, 2015; Liu et al., 2019b). ...
Decomposing dispersal limitation: Limits on fecundity or seed distribution? 1 2011
... 关于选择过程对植物群落构建的影响, 主要集中于局域尺度上基于生态位理论的研究, 即非生物环境因子的筛选和种内/种间的竞争作用(Tilman, 1982; Chase & Leibold, 2003; Letten et al., 2017).植物群落中, 选择过程产生于群落内不同物种个体间适合度的差异(Vellend, 2010).群落构建过程中, 一般存在3种形式的选择过程: (1)固定选择效应; (2)密度依赖效应; (3)时空选择效应.固定选择效应是指在特定的环境条件下, 如果群落内物种的相对适合度在时间和空间上固定不变, 那么群落内具有最高适合度的物种将会排除其他所有的物种(Vellend, 2010).密度依赖效应取决于群落内物种间定性(例如竞争、捕食、互利共生和疾病等)和定量的生态关系, 即所研究物种的适合度依赖于群落内同种和异种的个体密度, 包括负密度依赖效应(或被称为负密度制约效应)(祝燕等, 2009)和正密度依赖效应两个方面(Vellend, 2016).负密度依赖效应的研究内容主要包括: (1)密度依赖的自疏过程(随机死亡假说), 即同种个体的密度与死亡率呈现正相关关系(Kenkel, 1988); (2)距离依赖的死亡过程(Janzen-Connell假说), 即母树周围同种个体具有较高的密度时, 越靠近母树的同种个体死亡率越高(Janzen, 1970; Hubbell et al., 1990; Hyatt et al., 2003); (3)群落补偿趋势假说, 即群落越大, 负密度依赖作用越强, 越有利于稀有种在群落内的存活(Connell et al., 1984).负密度依赖效应对群落内的低密度物种更加有利, 故而可以使群落内物种维持稳定的共存状态(祝燕等, 2009).在片段化生境中, 不管是固定选择效应还是密度依赖效应都可能在时间和空间上存在差异, 即时空选择效应, 该效应也会潜在地影响群落动态.此外, 生境片段化限制了植物种子传播/扩散(Terborgh et al., 2011)以及特异性植食动物组成和数量的改变(Valladares et al., 2006; Emer et al., 2018), 进而影响种子或幼苗被植食的概率(白冰等, 2011; Genua et al., 2017; 骆杨青等, 2017).生境片段化所产生的边缘效应也能改变植物与天敌之间的关系(Krishnadas et al., 2018).生境片段化所引起的种间相互作用的改变, 最终会导致植物个体在空间分布格局上的差异, 影响密度效应的选择强度. ...
Corridors affect plants, animals, and their interactions in fragmented landscapes 3 2002
Conceptual cartoon illustrating processes in forest community assembly in fragmented habitats. Redrawn from <xref ref-type="bibr" rid="b46">HilleRisLambers <i>et al</i>. (2012)</xref> and <xref ref-type="bibr" rid="b133">Vellend (2016)</xref>. Different letters denote different species; circles of different sizes denote different size patches; different fan-shaped areas in circles denote the relative abundance of each species in one community.Fig. 21.1 扩散 扩散是有机体或其繁殖体离开出生地后的迁移或传播, 该过程对群落构建的影响主要依赖于扩散者产生群落(源群落)和抵达群落(汇群落)的大小和物种组成(Holyoak et al., 2005).与连续森林相比, 存在隔离的森林斑块将限制物种在斑块间的自由扩散(如种子传播), 即群落间物种的扩散限制.根据源群落和汇群落的相对大小, 片段化生境中物种的扩散可以分为两种情形: 一是大陆-岛屿模型, 该模型中将扩散假定为单向扩散, 由无限大的源群落(大陆或大面积斑块)向一个或多个小的、隔离的汇群落(岛屿或小面积斑块)中扩散, 源群落的物种组成相对恒定, 而汇群落的物种组成变化则高于源群落; 二是岛屿模型, 该模型中的群落(岛屿或小面积斑块)通过扩散相互联系在一起, 形成集合群落(Holyoak et al., 2005).一般而言, 扩散过程对群落构建的作用是受其他生态过程影响的, 尤其是选择过程和生态漂变(Ron et al., 2018). ...
... ; Tewksbury et al., 2002); Moss Fragmentation Experiments (UK: Gonzalez et al. 1998; Canada: Lindo et al., 2012); The Thousand Island Lake Experiments (China)(Yu et al., 2012; Liu et al., 2020); The Stability of Altered Forest Ecosystems (SAFE) Project (Malaysia)(Ewers et al., 2011); Metatron experiment (France)(Legrand et al., 2012). Fig. 1 10.17521/cjpe.2019.0155.F002
Conceptual cartoon illustrating processes in forest community assembly in fragmented habitats. Redrawn from <xref ref-type="bibr" rid="b46">HilleRisLambers <i>et al</i>. (2012)</xref> and <xref ref-type="bibr" rid="b133">Vellend (2016)</xref>. Different letters denote different species; circles of different sizes denote different size patches; different fan-shaped areas in circles denote the relative abundance of each species in one community.Fig. 21.1 扩散 扩散是有机体或其繁殖体离开出生地后的迁移或传播, 该过程对群落构建的影响主要依赖于扩散者产生群落(源群落)和抵达群落(汇群落)的大小和物种组成(Holyoak et al., 2005).与连续森林相比, 存在隔离的森林斑块将限制物种在斑块间的自由扩散(如种子传播), 即群落间物种的扩散限制.根据源群落和汇群落的相对大小, 片段化生境中物种的扩散可以分为两种情形: 一是大陆-岛屿模型, 该模型中将扩散假定为单向扩散, 由无限大的源群落(大陆或大面积斑块)向一个或多个小的、隔离的汇群落(岛屿或小面积斑块)中扩散, 源群落的物种组成相对恒定, 而汇群落的物种组成变化则高于源群落; 二是岛屿模型, 该模型中的群落(岛屿或小面积斑块)通过扩散相互联系在一起, 形成集合群落(Holyoak et al., 2005).一般而言, 扩散过程对群落构建的作用是受其他生态过程影响的, 尤其是选择过程和生态漂变(Ron et al., 2018). ...
... 野外控制实验也是检验片段化生境中群落构建过程非常有效的手段(Debinski & Holt, 2000; Resasco et al., 2017).Haddad等(2015)综述了目前世界上已有的针对片段化生境的控制实验(图1), 通过设置控制实验和长期观察, 对片段化生境中或岛屿上群落构建的过程进行了有效的检验(Wilson et al., 2016; Resasco et al., 2017).例如, 巴西的片段化森林动态实验是目前最大和时间最长的野外控制实验, 通过设置不同大小的森林斑块且控制斑块之间的隔离度差异, 分析森林斑块面积的大小对群落构建过程的影响(Laurance et al., 2011); 美国的Savannah河流廊道实验通过增加斑块间的隔离度, 且控制斑块的面积差异, 分析廊道效应(扩散)在群落构建过程中的作用(Haddad & Baum, 1999; Tewksbury et al., 2002); Kansas生境片段化实验则通过设置不同面积大小和隔离度的片段化斑块, 长期监测斑块内群落的动态变化, 研究生境片段化对群落构建和演替过程的影响(Holt et al., 1995); 另外, Alexander等(2012)利用此实验在种群水平上分析了种子扩散与斑块间连通性之间的关系, 在群落水平上分析了环境过滤和扩散限制的共同作用对群落演替的作用.然而, 除上述主要的实验外, 基于野外控制实验检验片段化生境中森林群落构建过程的研究仍然不足. ...
Resource competition and community structure 1 1982
... 关于选择过程对植物群落构建的影响, 主要集中于局域尺度上基于生态位理论的研究, 即非生物环境因子的筛选和种内/种间的竞争作用(Tilman, 1982; Chase & Leibold, 2003; Letten et al., 2017).植物群落中, 选择过程产生于群落内不同物种个体间适合度的差异(Vellend, 2010).群落构建过程中, 一般存在3种形式的选择过程: (1)固定选择效应; (2)密度依赖效应; (3)时空选择效应.固定选择效应是指在特定的环境条件下, 如果群落内物种的相对适合度在时间和空间上固定不变, 那么群落内具有最高适合度的物种将会排除其他所有的物种(Vellend, 2010).密度依赖效应取决于群落内物种间定性(例如竞争、捕食、互利共生和疾病等)和定量的生态关系, 即所研究物种的适合度依赖于群落内同种和异种的个体密度, 包括负密度依赖效应(或被称为负密度制约效应)(祝燕等, 2009)和正密度依赖效应两个方面(Vellend, 2016).负密度依赖效应的研究内容主要包括: (1)密度依赖的自疏过程(随机死亡假说), 即同种个体的密度与死亡率呈现正相关关系(Kenkel, 1988); (2)距离依赖的死亡过程(Janzen-Connell假说), 即母树周围同种个体具有较高的密度时, 越靠近母树的同种个体死亡率越高(Janzen, 1970; Hubbell et al., 1990; Hyatt et al., 2003); (3)群落补偿趋势假说, 即群落越大, 负密度依赖作用越强, 越有利于稀有种在群落内的存活(Connell et al., 1984).负密度依赖效应对群落内的低密度物种更加有利, 故而可以使群落内物种维持稳定的共存状态(祝燕等, 2009).在片段化生境中, 不管是固定选择效应还是密度依赖效应都可能在时间和空间上存在差异, 即时空选择效应, 该效应也会潜在地影响群落动态.此外, 生境片段化限制了植物种子传播/扩散(Terborgh et al., 2011)以及特异性植食动物组成和数量的改变(Valladares et al., 2006; Emer et al., 2018), 进而影响种子或幼苗被植食的概率(白冰等, 2011; Genua et al., 2017; 骆杨青等, 2017).生境片段化所产生的边缘效应也能改变植物与天敌之间的关系(Krishnadas et al., 2018).生境片段化所引起的种间相互作用的改变, 最终会导致植物个体在空间分布格局上的差异, 影响密度效应的选择强度. ...
Contribution of small habitat fragments to conservation of insect communities of grassland-cropland landscapes 1 2002
... 根据片段化森林构建的主要生态过程(扩散限制、选择过程和生态漂变), 如果所有小斑块中的物种, 随机来源于大斑块中的物种库(源-库动态), 且斑块间物种呈嵌套分布, 即生态漂变是形成森林斑块间物种组成差异的主要过程, 此时保护一个大的保护区将变得更为有意义.然而, 已发现显著的嵌套式分布格局仅在9%的系统中出现, 而反嵌套的物种分布格局在16%的系统中出现(Matthews et al., 2015a).如果小斑块之间存在较高的生境异质性, 不同的斑块上选择不同的物种, 即小面积森林斑块之间存在较高的β多样性(Liu et al., 2018), 那么小面积的斑块也具有保护价值, 不能被忽略(Tscharntke et al., 2002; Rösch et al., 2015).基于全球尺度的分析, 也发现小面积的斑块具有一定的保护价值(Wintle et al., 2019).因此, 在植被恢复和管理中, 如果仅能在多个小面积的斑块上进行植被恢复, 应尽可能地增加斑块之间的生境异质性以及物种组成的差异性, 以维持更高的区域物种多样性. ...
Climatic and geographic correlates of global forest tree species-?abundance distributions and community evenness 1 2016
... 物种多度分布(SAD)描述了一个取样单元或群落内所有物种的个体多度分布状况, 是一个群落最基本的描述(McGill et al., 2007).通常, SAD通过物种序列多度图(RAD)进行可视化(Ulrich et al., 2010).已有大量的研究对群落内RAD的形状进行模型拟合, 如常见的模型有: 对数级数模型、对数正态模型等近30个模型(McGill et al., 2007).但是, 如何根据RAD曲线所拟合的模型反映群落的构建过程仍是研究的热点和难点问题.虽然仅通过对不同SAD模型的拟合尚无法准确地推测群落的构建过程, 但在地理尺度上, 通过分析不同环境因子和空间因子等对RAD形状的影响, 可以探讨扩散过程对群落构建的影响(Fattorini et al., 2016).在全球尺度上, Ulrich等(2016)通过对数级数模型和对数正态模型的拟合, 在纬度梯度上分析了扩散限制和环境过滤对群落构建的影响, 发现处于不同纬度的群落内扩散限制所产生的相对作用不同, 导致了低纬度地区群落内的RAD符合对数级数模型而高纬度地区群落内RAD更加符合对数正态模型.相似地, 在纬度梯度上利用非中性模型对森林群落的RAD进行拟合, 发现随纬度梯度的增加,环境过滤作用导致实际的SAD模型与中性模型的分化逐渐增加, 即中性过程对群落构建作用的影响减弱, 而非中性过程增加(Qiao et al., 2015a).Matthews等(2017)发现岛屿间的隔离程度可以改变岛屿上节肢动物的RAD的形状, 但关于岛屿参数如何影响森林群落的RAD尚缺乏研究.SAD不仅可以了解片段化生境中群落构建的过程, 也能够反映群落内稀有种和常见种的分布格局, 通过探讨片段化生境内物种多度的改变, 可应用于生物多样性的管理和应用生态学等方面(Matthews & Whittaker, 2015b).但在不同面积和隔离度的片段化森林中, 结合RAD形状的改变探讨群落构建过程的研究仍然不足. ...
A meta-analysis of ?species- abundance distributions 1 2010
... 物种多度分布(SAD)描述了一个取样单元或群落内所有物种的个体多度分布状况, 是一个群落最基本的描述(McGill et al., 2007).通常, SAD通过物种序列多度图(RAD)进行可视化(Ulrich et al., 2010).已有大量的研究对群落内RAD的形状进行模型拟合, 如常见的模型有: 对数级数模型、对数正态模型等近30个模型(McGill et al., 2007).但是, 如何根据RAD曲线所拟合的模型反映群落的构建过程仍是研究的热点和难点问题.虽然仅通过对不同SAD模型的拟合尚无法准确地推测群落的构建过程, 但在地理尺度上, 通过分析不同环境因子和空间因子等对RAD形状的影响, 可以探讨扩散过程对群落构建的影响(Fattorini et al., 2016).在全球尺度上, Ulrich等(2016)通过对数级数模型和对数正态模型的拟合, 在纬度梯度上分析了扩散限制和环境过滤对群落构建的影响, 发现处于不同纬度的群落内扩散限制所产生的相对作用不同, 导致了低纬度地区群落内的RAD符合对数级数模型而高纬度地区群落内RAD更加符合对数正态模型.相似地, 在纬度梯度上利用非中性模型对森林群落的RAD进行拟合, 发现随纬度梯度的增加,环境过滤作用导致实际的SAD模型与中性模型的分化逐渐增加, 即中性过程对群落构建作用的影响减弱, 而非中性过程增加(Qiao et al., 2015a).Matthews等(2017)发现岛屿间的隔离程度可以改变岛屿上节肢动物的RAD的形状, 但关于岛屿参数如何影响森林群落的RAD尚缺乏研究.SAD不仅可以了解片段化生境中群落构建的过程, 也能够反映群落内稀有种和常见种的分布格局, 通过探讨片段化生境内物种多度的改变, 可应用于生物多样性的管理和应用生态学等方面(Matthews & Whittaker, 2015b).但在不同面积和隔离度的片段化森林中, 结合RAD形状的改变探讨群落构建过程的研究仍然不足. ...
Habitat fragmentation effects on trophic processes of insect-plant food webs 2 2006
... 关于选择过程对植物群落构建的影响, 主要集中于局域尺度上基于生态位理论的研究, 即非生物环境因子的筛选和种内/种间的竞争作用(Tilman, 1982; Chase & Leibold, 2003; Letten et al., 2017).植物群落中, 选择过程产生于群落内不同物种个体间适合度的差异(Vellend, 2010).群落构建过程中, 一般存在3种形式的选择过程: (1)固定选择效应; (2)密度依赖效应; (3)时空选择效应.固定选择效应是指在特定的环境条件下, 如果群落内物种的相对适合度在时间和空间上固定不变, 那么群落内具有最高适合度的物种将会排除其他所有的物种(Vellend, 2010).密度依赖效应取决于群落内物种间定性(例如竞争、捕食、互利共生和疾病等)和定量的生态关系, 即所研究物种的适合度依赖于群落内同种和异种的个体密度, 包括负密度依赖效应(或被称为负密度制约效应)(祝燕等, 2009)和正密度依赖效应两个方面(Vellend, 2016).负密度依赖效应的研究内容主要包括: (1)密度依赖的自疏过程(随机死亡假说), 即同种个体的密度与死亡率呈现正相关关系(Kenkel, 1988); (2)距离依赖的死亡过程(Janzen-Connell假说), 即母树周围同种个体具有较高的密度时, 越靠近母树的同种个体死亡率越高(Janzen, 1970; Hubbell et al., 1990; Hyatt et al., 2003); (3)群落补偿趋势假说, 即群落越大, 负密度依赖作用越强, 越有利于稀有种在群落内的存活(Connell et al., 1984).负密度依赖效应对群落内的低密度物种更加有利, 故而可以使群落内物种维持稳定的共存状态(祝燕等, 2009).在片段化生境中, 不管是固定选择效应还是密度依赖效应都可能在时间和空间上存在差异, 即时空选择效应, 该效应也会潜在地影响群落动态.此外, 生境片段化限制了植物种子传播/扩散(Terborgh et al., 2011)以及特异性植食动物组成和数量的改变(Valladares et al., 2006; Emer et al., 2018), 进而影响种子或幼苗被植食的概率(白冰等, 2011; Genua et al., 2017; 骆杨青等, 2017).生境片段化所产生的边缘效应也能改变植物与天敌之间的关系(Krishnadas et al., 2018).生境片段化所引起的种间相互作用的改变, 最终会导致植物个体在空间分布格局上的差异, 影响密度效应的选择强度. ...
... 以往的理论/假说(如ETIB、集合群落理论和群落生态学理论)主要针对单一营养级或两个营养级上的生态过程进行分析, 如单一营养级上的种间相互作用及其与非生物因子间的关系(选择过程).近年来, 在ETIB理论的基础上, 许多研究开始逐渐关注岛屿生物地理学的营养理论, 即将营养级间物种的相互作用对定殖率和灭绝率的影响考虑其中(Gravel et al., 2011).在片段化生境中, 食物网各营养级间关联物种的次生灭绝是非常普遍的, 越顶级的消费者越容易消失(Ryser et al., 2019), 消费者缺失所引发的下行调控丧失效应通过影响传粉、种子传播和植食等与扩散和选择相关的生态过程, 从而影响片段化森林群落的构建过程(Valladares et al., 2006; Larios et al., 2017; Emer et al., 2018).Cirtwill和Stouffer (2016)发现, 当考虑岛屿上的营养级结构时, 如果包含上行效应的影响, 能够改进ETIB中对迁移率和灭绝率的预测.但在片段化森林景观中, 针对多营养级上的种间相互作用(如下行效应)在森林群落构建过程的作用研究, 仍鲜有报道. ...
... 和Vellend (2016).不同的英文字母表示不同的物种; 不同大小的圆圈表示不同面积的斑块, 斑块内不同的扇形面积表示各物种的相对多度. Conceptual cartoon illustrating processes in forest community assembly in fragmented habitats. Redrawn from <xref ref-type="bibr" rid="b46">HilleRisLambers <i>et al</i>. (2012)</xref> and <xref ref-type="bibr" rid="b133">Vellend (2016)</xref>. Different letters denote different species; circles of different sizes denote different size patches; different fan-shaped areas in circles denote the relative abundance of each species in one community.Fig. 21.1 扩散 扩散是有机体或其繁殖体离开出生地后的迁移或传播, 该过程对群落构建的影响主要依赖于扩散者产生群落(源群落)和抵达群落(汇群落)的大小和物种组成(Holyoak et al., 2005).与连续森林相比, 存在隔离的森林斑块将限制物种在斑块间的自由扩散(如种子传播), 即群落间物种的扩散限制.根据源群落和汇群落的相对大小, 片段化生境中物种的扩散可以分为两种情形: 一是大陆-岛屿模型, 该模型中将扩散假定为单向扩散, 由无限大的源群落(大陆或大面积斑块)向一个或多个小的、隔离的汇群落(岛屿或小面积斑块)中扩散, 源群落的物种组成相对恒定, 而汇群落的物种组成变化则高于源群落; 二是岛屿模型, 该模型中的群落(岛屿或小面积斑块)通过扩散相互联系在一起, 形成集合群落(Holyoak et al., 2005).一般而言, 扩散过程对群落构建的作用是受其他生态过程影响的, 尤其是选择过程和生态漂变(Ron et al., 2018). ...
... and Vellend (2016). Different letters denote different species; circles of different sizes denote different size patches; different fan-shaped areas in circles denote the relative abundance of each species in one community. Fig. 21.1 扩散 扩散是有机体或其繁殖体离开出生地后的迁移或传播, 该过程对群落构建的影响主要依赖于扩散者产生群落(源群落)和抵达群落(汇群落)的大小和物种组成(Holyoak et al., 2005).与连续森林相比, 存在隔离的森林斑块将限制物种在斑块间的自由扩散(如种子传播), 即群落间物种的扩散限制.根据源群落和汇群落的相对大小, 片段化生境中物种的扩散可以分为两种情形: 一是大陆-岛屿模型, 该模型中将扩散假定为单向扩散, 由无限大的源群落(大陆或大面积斑块)向一个或多个小的、隔离的汇群落(岛屿或小面积斑块)中扩散, 源群落的物种组成相对恒定, 而汇群落的物种组成变化则高于源群落; 二是岛屿模型, 该模型中的群落(岛屿或小面积斑块)通过扩散相互联系在一起, 形成集合群落(Holyoak et al., 2005).一般而言, 扩散过程对群落构建的作用是受其他生态过程影响的, 尤其是选择过程和生态漂变(Ron et al., 2018). ...
Habitat fragmentation in the temperate zone 1 1986
... 生境片段化(或破碎化, habitat fragmentation)是指原来连续的生境受自然和人为干扰的影响, 逐渐转变为隔离的且面积较小的斑块(Wilcove et al., 1986; Fahrig, 2003).人类干扰(如择伐、都市化、公路和水库建设等)对自然生态系统的破坏以及自然因素(如森林火灾、岛屿等)的影响, 造成全球森林植被呈片段化分布(Fahrig, 2003; Didham, 2010; Haddad et al., 2015; Taubert et al., 2018; Liu et al., 2019a).从全球森林的分布来看, 世界上大部分森林斑块的面积小于10 hm2 (Haddad et al., 2015).森林面积的丧失、隔离度的增加和边缘效应的加剧将显著影响片段化生境中森林群落的结构(Laurance et al., 2006)和演替动态(Benchimol & Peres, 2015; Liu et al., 2019b). ...
Conceptual cartoon illustrating processes in forest community assembly in fragmented habitats. Redrawn from <xref ref-type="bibr" rid="b46">HilleRisLambers <i>et al</i>. (2012)</xref> and <xref ref-type="bibr" rid="b133">Vellend (2016)</xref>. Different letters denote different species; circles of different sizes denote different size patches; different fan-shaped areas in circles denote the relative abundance of each species in one community.Fig. 21.1 扩散 扩散是有机体或其繁殖体离开出生地后的迁移或传播, 该过程对群落构建的影响主要依赖于扩散者产生群落(源群落)和抵达群落(汇群落)的大小和物种组成(Holyoak et al., 2005).与连续森林相比, 存在隔离的森林斑块将限制物种在斑块间的自由扩散(如种子传播), 即群落间物种的扩散限制.根据源群落和汇群落的相对大小, 片段化生境中物种的扩散可以分为两种情形: 一是大陆-岛屿模型, 该模型中将扩散假定为单向扩散, 由无限大的源群落(大陆或大面积斑块)向一个或多个小的、隔离的汇群落(岛屿或小面积斑块)中扩散, 源群落的物种组成相对恒定, 而汇群落的物种组成变化则高于源群落; 二是岛屿模型, 该模型中的群落(岛屿或小面积斑块)通过扩散相互联系在一起, 形成集合群落(Holyoak et al., 2005).一般而言, 扩散过程对群落构建的作用是受其他生态过程影响的, 尤其是选择过程和生态漂变(Ron et al., 2018). ...
... ); The Thousand Island Lake Experiments (China)(Yu et al., 2012; Liu et al., 2020); The Stability of Altered Forest Ecosystems (SAFE) Project (Malaysia)(Ewers et al., 2011); Metatron experiment (France)(Legrand et al., 2012). Fig. 1 10.17521/cjpe.2019.0155.F002
Conceptual cartoon illustrating processes in forest community assembly in fragmented habitats. Redrawn from <xref ref-type="bibr" rid="b46">HilleRisLambers <i>et al</i>. (2012)</xref> and <xref ref-type="bibr" rid="b133">Vellend (2016)</xref>. Different letters denote different species; circles of different sizes denote different size patches; different fan-shaped areas in circles denote the relative abundance of each species in one community.Fig. 21.1 扩散 扩散是有机体或其繁殖体离开出生地后的迁移或传播, 该过程对群落构建的影响主要依赖于扩散者产生群落(源群落)和抵达群落(汇群落)的大小和物种组成(Holyoak et al., 2005).与连续森林相比, 存在隔离的森林斑块将限制物种在斑块间的自由扩散(如种子传播), 即群落间物种的扩散限制.根据源群落和汇群落的相对大小, 片段化生境中物种的扩散可以分为两种情形: 一是大陆-岛屿模型, 该模型中将扩散假定为单向扩散, 由无限大的源群落(大陆或大面积斑块)向一个或多个小的、隔离的汇群落(岛屿或小面积斑块)中扩散, 源群落的物种组成相对恒定, 而汇群落的物种组成变化则高于源群落; 二是岛屿模型, 该模型中的群落(岛屿或小面积斑块)通过扩散相互联系在一起, 形成集合群落(Holyoak et al., 2005).一般而言, 扩散过程对群落构建的作用是受其他生态过程影响的, 尤其是选择过程和生态漂变(Ron et al., 2018). ...
... 迄今关于片段化生境中重构森林群落的研究仍然十分缺乏.我们对片段化生境中森林的重新构建过程可以作如下预测: 1)重构群落演替的起始阶段, 空间的隔离作用将比斑块面积的大小更加重要, 因短期的干扰或生境质量的改变, 空间的隔离将阻止物种占用适合的生境, 导致种子传播或者强风等随机事件决定了物种的抵达和在斑块上的定殖(Walker et al., 2010).例如, Kadmon和Pulliam (1995)发现在新近皆伐的湖泊岛屿上空间的隔离增加了风力传播物种的比例, Whittaker等(1997)发现在新近形成的火山岛屿上, 物种早期阶段的定殖可以根据其扩散性状进行预测, 不过在具有近60年片段化历史的千岛湖, 岛屿面积是影响植物群落物种组成的主要因素, 而非空间的隔离(Yu et al., 2012; Baldeck et al., 2016).2)当某一物种扩散到某一斑块后, 斑块内已有的种子/幼苗库, 会对后续扩散来的物种的定殖产生影响, 即优先效应, 该效应与决定性过程和随机过程会共同影响片段化生境中群落的构建过程(Fukami, 2015).3)物种抵达相应的斑块之后, 斑块内的生境异质性、种间/种内关系和生态漂变等过程将共同决定物种定殖成功的概率和物种组成, 如筛选个体生长快、木质密度低以及喜阳的先锋物种(Baldeck et al., 2016; Liu et al., 2019b, 2020), 或具有菌根的植物物种等(Delavaux et al., 2019).Baynes等(2016)通过比较砍伐后重构的片段化森林群落与连续森林群落内的物种多样性, 发现重构的森林群落中反而具有更多的稀有种以及较高的β多样性, 生态漂变过程也会影响片段化森林群落的重构. ...
The effects of habitat loss and fragmentation on plant functional traits and functional diversity: What do we know so far? 3 2019
... 集合群落理论包含中性过程、斑块动态、物种筛选和质量效应4种理论框架(Leibold et al., 2004; Holyoak et al., 2005)(附件I), 在ETIB和中性理论的基础上, 将选择过程考虑在内, 阐述了片段化生境中群落构建的基本过程.Vellend (2016)所提出的生态群落构建的4个高阶过程中的生态漂变类似于集合群落模型中的中性过程, 扩散和选择过程分别类似于斑块动态和物种筛选.虽然物种形成在集合群落理论中没有被明确排除在外, 但在多数情况下, 集合群落理论中并没有考虑该过程.在Vellend (2016)群落生态学理论提出后, 已有部分研究在片段化/岛屿生境中检验不同的生态过程在群落构建中的相对作用, 如选择过程和生态漂变过程的相对作用(Liu et al., 2018)、扩散对选择和生态漂变过程相对作用的影响(Ron et al., 2018)等.另外, 通过扩展ETIB理论, 考虑物种间功能特征的差异及其所受面积和隔离度的影响(Jacquet et al., 2017), 以及物种功能性状特征分布与环境因子间的关系, 逐渐成为检验片段化生境中群落构建过程的重要手段(Zambrano et al., 2019).然而, 针对不同生态过程如何影响片段化森林群落构建的研究仍然十分缺乏, 尤其是对于扩散和物种形成过程的相关检验. ...
... 在片段化生境中, 根据群落内物种的某一类性状, 如与种群扩散、建立和维持等相关的植物生活史性状(Zambrano et al., 2019), 结合不同大小斑块内的生境条件和隔离度等对群落内功能特征收敛或发散程度的影响, 也能有助于我们理解选择过程或扩散过程在群落构建中的作用(?ckinger et al., 2010; Marini et al., 2012; Jiang et al., 2018).例如, 结合种子的功能特征, 可以分析扩散限制作用在群落构建过程中的作用(Negoita et al., 2016; Favre-Bac et al., 2017).有研究发现在越大的岛屿上小种子植物比例更高(种子特征的收敛), 可能是由于大岛上小种子扩散者(鸟类)的比例更高所引起的(Liu et al., 2019c). ...
 ,1,2, 于明坚
,1,2, 于明坚 ,2,*
,2,*
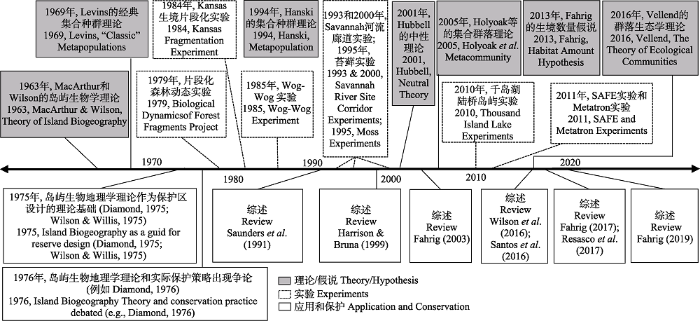 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT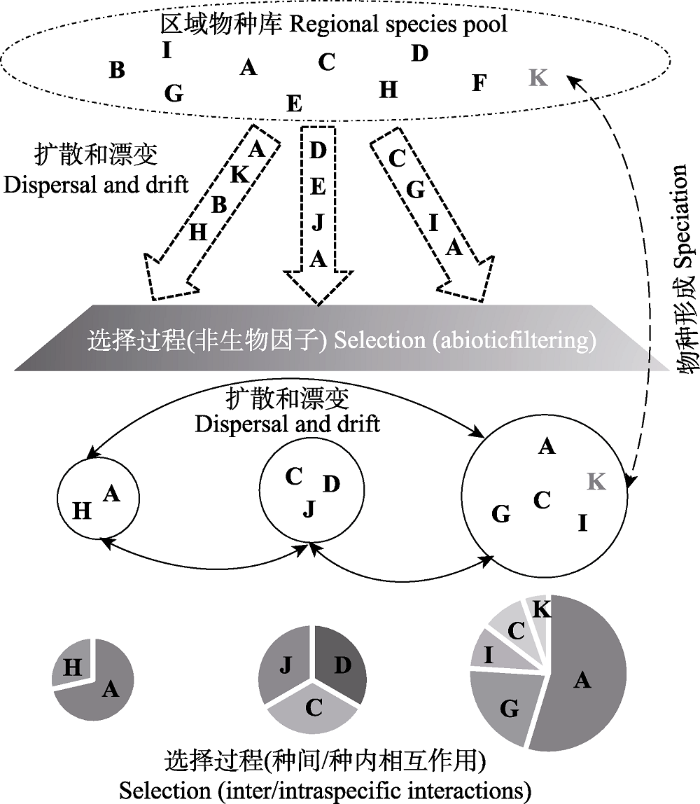 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT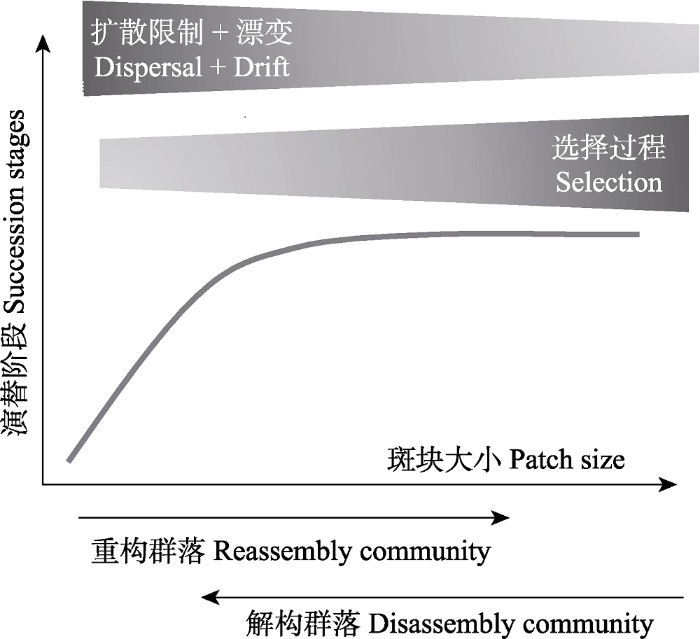 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}