唐丽丽
1 , 张梅
1 , 赵香林
1 , 康慕谊
2 , 3 , 刘鸿雁
4 , 高贤明
5 , 杨彤
1 , 郑璞帆
1 , 石福臣
, 1 , * 1 南开大学生命科学学院, 天津 300071 2 北京师范大学地表过程与资源生态国家重点实验室, 北京 100875 3 北京师范大学地理科学学部自然资源学院, 北京 100875 4 北京大学城市与环境学院, 北京大学生态研究中心, 地表过程分析与模拟教育部重点实验室, 北京 100871 5 中国科学院植物研究所植被与环境变化国家重点试验室, 北京 100093 Species distribution and community assembly rules of Juglans mandshurica in North China Li-Li TANG
1 , Mei ZHANG
1 , Xiang-Lin ZHAO
1 , Mu-Yi KANG
2 , 3 , Hong-Yan LIU
4 , Xian-Ming GAO
5 , Tong YANG
1 , Pu-Fan ZHENG
1 , Fu-Chen SHI
, 1 , * 1 College of Life Sciences, Nankai University, Tianjin 300071, China 2 State Key Laboratory of Earth Surface Processes and Resource Ecology, Beijing Normal University, Beijing 100875, China 3 College of Resources Science & Technology, Faculty of Geographical Science, Beijing Normal University, Beijing 100875, China 4 Institute of Ecology, College of Urban and Environmental Sciences, Laboratory for Earth Surface Processes of the Ministry of Education, Peking University, Beijing 100871, China 5 State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China 通讯作者: * fcshi@nankai.edu.cn 编委: 唐志尧
责任编辑: 李敏, 赵航(实习)
收稿日期: 2018-07-07
接受日期: 2018-12-13
网络出版日期: 2019-09-20
基金资助: 国家科技基础性工作专项 (2011FY110300 )
Corresponding authors: * fcshi@nankai.edu.cn Received: 2018-07-07
Accepted: 2018-12-13
Online: 2019-09-20
Fund supported: Supported by the National Basic Work of Science and Technology of China (2011FY110300 )
摘要 探究植物分布规律和群落构建机制是揭示植物群落空间分布、群落物种多样性的形成、发展及其影响因素的重要途径。该文以华北地区胡桃楸(
Juglans mandshurica )林为研究对象, 基于野外84个样方的调查数据, 通过径级分析、典范对应分析等方法研究了胡桃楸林的空间分布规律, 并通过亲缘关系指数计算、植物功能性状等方法研究了胡桃楸林的物种共存机制。结果表明, 华北地区胡桃楸的胸径相对较小(平均5.36 cm), 种群年龄较低; 海拔、坡度、坡位和人为干扰程度是影响华北地区胡桃楸分布的主要因子。研究区域内的胡桃楸大部分生长于山体中下部海拔较低的缓坡, 且人为干扰相对较少的区域, 各分布区域中胡桃楸林的分布规律各异。胡桃楸林物种的构建由生态位机制主导, 其中河北、陕西、天津的胡桃楸林物种共存过程主要受负密度制约的影响, 北京、山西的胡桃楸林的物种共存过程主要受环境选择驱动。
关键词: 胡桃楸 ;
分布规律 ;
谱系发育 ;
群落构建机制 ;
植物群落 Abstract Aims Explore the distribution pattern and the processes controlling the assembly of the
Juglans mandshurica dominated forests in North China.
Methods We investigated 84 plots, each with an area of 20 m × 30 m, for the
J. mandshurica forests in the North China. We analyzed demography of the
J. mandshurica based on its diameters, and applied canonical correspondence analysis (CCA) to explore the relationship between environment and species composition of different formations of the
J. mandshurica forests. We then calculated phylogenetic index, i.e., net relatedness index (
NRI ) and nearest taxonomic index (
NTI ) and Gaussian Kernel Density Estimation (Gaussian KDE), for each plot to explore the rules controlling community assembly of these forests
.Important findings The population of
J. mandshurica is relatively young in the Northern China, with a mean diameters at breast height (
DBH ) of 5.36 cm. Distribution of
J. mandshurica in North China varied significantly with altitude, slope, slope position and human disturbance. Most of the
J. mandshurica located at the low part of the massif with lower altitude and less human disturbance. In North China, niche related processes regulated species coexistence of Form.
J. mandshurica, with competition controlled the community assembly in Hebei, Shaanxi and Tianjin, while environment filtering dominated in Beijing and Shanxi.
Keywords: Juglans mandshurica ;
distribution pattern ;
phylogeny ;
community assembly rules ;
plant community PDF (2112KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 唐丽丽, 张梅, 赵香林, 康慕谊, 刘鸿雁, 高贤明, 杨彤, 郑璞帆, 石福臣. 华北地区胡桃楸林分布规律及群落构建机制分析. 植物生态学报, 2019, 43(9): 753-761. DOI:
10.17521/cjpe.2018.0161 TANG Li-Li, ZHANG Mei, ZHAO Xiang-Lin, KANG Mu-Yi, LIU Hong-Yan, GAO Xian-Ming, YANG Tong, ZHENG Pu-Fan, SHI Fu-Chen.
Species distribution and community assembly rules of Juglans mandshurica in North China .
Chinese Journal of Plant Ecology , 2019, 43(9): 753-761. DOI:
10.17521/cjpe.2018.0161 群落构建机制即群落物种多样性形成及其维持机制, 主要研究植物群落如何构建, 什么驱使植物的多样性及群落功能的变化(
Cavender-Bares et al ., 2009 )。目前对群落构建机制的分析主要基于谱系发育和植物功能性状(
Webb et al ., 2002 ;
Wright, 2004 ;
Kraft et al ., 2008 ;
Swenson, 2011 ;
马克平, 2016 ;
D’Amen, 2017 ), 对群落构建机制的解释包括生态位机制(
Diamond, 1975 )和中性理论(
Hubbell, 2001 )。中性理论认为群落内物种的出生、死亡、迁入、迁出处于一个动态的平衡, 这种动态平衡维持着群落内物种数量的稳定(
Hubbell, 2001 )。生态位机制则认为群落内植物的生长繁殖、迁入迁出源自于群落的生境、资源及生物间相互作用, 生态位机制是由环境过滤和负密度制约(
Bazzaz, 1991 ;
Weiher & Keddy, 1999 )两个过程主导群落构建。
随着相关学科领域的快速发展,
Webb等(2002) 提出用植物谱系发育理念来理解群落物种共存过程, 并提出净亲缘指数(
NRI )和最近亲缘指数(
NTI )这两种亲缘关系指数, 以此来计算群落内物种间的系统发育距离, 并在植物功能性状生态位保守和趋同两种情况下讨论群落的构建机制(
Kraft et al ., 2007 ;
Vamosi et al ., 2009 )。若亲缘关系指数大于0, 则物种系统发育距离较小, 群落由亲缘关系较近的物种构成; 若亲缘关系指数小于0, 说明物种系统发育距离较大, 群落由亲缘关系较远的物种构成; 若亲缘关系指数为0, 说明物种在系统发育结构上呈现随机状态(
Webb et al ., 2002 )。若群落物种生态位保守, 环境过滤作用会导致群落谱系结构的聚集, 竞争排斥(负密度制约)作用会导致群落谱系结构的发散; 若群落物种生态位趋同, 环境过滤作用会导致群落谱系结构发散, 负密度制约作用会导致群落谱系结构随机或聚集(
Webb et al ., 2002 ;
Kraft et al ., 2007 ;
Vamosi et al ., 2009 )。在中性作用下, 群落谱系结构均呈随机状态(
McPeek, 2007 )。群落生态学与植物谱系发育概念的结合, 为解释群落的构建提供了新的理念。
胡桃楸(
Juglans mandshurica )又名核桃楸, 隶属于胡桃科胡桃属(
Juglans ), 属第三纪孑遗植物, 在我国多分布于东北的东部山地。同时, 胡桃楸的分布区域也在向南延伸, 扩展到我国华北地区, 主要为北京、天津等地的山区, 且颇具规模。目前针对东北地区分布的胡桃楸种群的研究较为丰富全面(
马万里等, 2001 ;
梁淑娟等, 2005 ;
王东娜等, 2011 ;
王东亮, 2014 ;
苏金娟和王晓春, 2017 ), 而华北地区胡桃楸种群的研究相对较少, 且多集中在胡桃楸生理学特性及群落结构等方面(
闫娜等, 2014 ;
高张莹等, 2017 ,
Song et al ., 2017 )。华北地区胡桃楸种群的分布规律、影响其分布的环境因子、胡桃楸林的系统发育结构, 以及群落构建机制都尚未清楚。本文利用在“华北地区自然植物群落资源综合考察”中获得的大量胡桃楸样方数据, 对华北地区胡桃楸种群分布规律及其影响因素进行分析, 并综合谱系发育和功能性状探讨胡桃楸林的物种共存过程, 为胡桃楸林及其生境的保护提供科学依据, 也为进一步探究华北地区胡桃楸林物种组成及群落发展方向奠定基础。
1 材料和方法 1.1 研究区域 研究区域以华北地区为主, 包括北京、天津、河北、山西、陕西、河南6个省市(
图1 )。该区域背依中国西北腹地, 南接长江中下游平原, 东临渤海。研究区内地势西北高, 东南低, 海拔1-3 691 m, 地形复杂多样, 包括平原、山地、高原、丘陵、盆地等。气候以温带大陆性气候为主, 夏季高温多雨, 冬季寒冷干旱, 年平均气温8-13 ℃, 年降水量400- 1 000 mm。研究区内生态系统类型丰富多样, 涵盖山地森林、灌丛、草原、湿地等。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1华北地区胡桃楸样地分布。 Fig. 1Sampling sites of Juglans mandshurica in North China. 1.2 研究方法 1.2.1 野外调查及径级划分
华北地区胡桃楸林主要为天然次生林, 本文采用样方法对其进行野外调查, 样方设置方法参照
方精云等(2009) 的植物群落清查的技术规范, 样方尺度主要为20 m × 30 m, 记录样方内所有胸径(
DBH ) ≥1 cm的木本植物和所有草本植物的相关信息, 其中木本植物主要记录种类、数量、树高、胸径, 草本植物主要记录种类、多度、盖度, 同时记录样地生境信息, 如样方坐标、海拔、坡度、坡向等信息。根据各地胡桃楸种群分布及规模, 在研究区域内共设12个样地, 其中北京3个、天津1个、河北2个、山西2个、陕西3个、河南1个, 12个样地中共设有84个样方(附录I), 样地具体分布位置如
图1 所示。根据相关技术规范, 本研究采用2 cm径距将胡桃楸胸径划分为9个径级, 并结合其分布位点, 利用ArcGIS 10.2分析不同径级胡桃楸在华北地区的分布。
1.2.2 胡桃楸空间分布及其影响因子分析
根据样地调查结果及华北地区数字高程图(来源于中国科学院资源环境科学数据中心), 利用ArcGIS 10.2, 分析华北地区胡桃楸的空间分布规律。使用典范对应分析(CCA)方法, 定量分析海拔、坡度、坡向、坡位、人为干扰对胡桃楸在华北地区分布的影响。其中坡向按照其度数分为北、东北、西北、东、西、东南、西南、南, 分别用编号1-8代表。坡位分为谷地、下、中下、中、中上和上6个坡位, 编号1-6。人为干扰主要为伐木, 根据国家标准生物多样性观测技术导则(HJ710.12-2016)中对人为干扰活动的分类, 将人为干扰分为4个程度: 无干扰指生境及植物群落没有受到人为干扰, 植被保持原始状态, 对野生植物的生存繁衍没有任何影响, 在计算中赋值为1; 轻微干扰指生境及植物群落受到一定程度的影响, 但植被基本保持原始状态, 对野生植物生存繁衍影响不大, 在计算中赋值为2; 中度干扰指生境及植物群落受到干扰, 部分植被消失, 但干扰消失后, 植被仍可恢复, 野生植物生存繁衍受到一定程度的影响, 但仍然可以栖息繁衍, 在计算中赋值为3; 重度干扰指生境及植物群落受到严重干扰, 植被基本消失, 野生植物难以生存繁衍, 在计算中赋值为4。此过程在R 3.3.1中的“vegan”程序包(
Jari et al ., 2016 )中进行。
1.2.3 亲缘关系指数分析
本文参考
Webb等(2002) 的实验方法构建系统发育树。将样方调查中得到的种子植物, 按照科/属/种的格式, 输入到phylomatic平台(
Webb & Dongoghue, 2005 ;
Webb et al ., 2008 ), 得到基于APG III分类系统的具有进化枝长的系统发育树。根据各物种在系统发育树上的枝长和所在样方中的数量, 计算得出各样方的亲缘关系指数(
NRI 和
NTI )。此过程在R 3.3.1的“picante”程序包(
Kembel et al ., 2010 )中进行。因河南的胡桃楸数量较少, 仅在落叶栎林和华山松林中零散分布, 因此并未对河南的胡桃楸数据进行群落构建机制分析。
本文采用
唐丽丽等(2017) 对燕山东麓植物群落构建机制的研究中提出的高斯核密度估计的方法对亲缘关系指数的正负趋势进行分析。高斯核密度估计方法就是利用高斯核密度估计来绘制出亲缘关系指数的密度分布图, 并估算
NRI 、
NTI 在正、负区域内的面积, 若指数在正值区域面积大于其在负值区域的面积, 则该指数趋近于正值, 反之则该指数 趋近于负值, 进而探究各群落谱系结构聚集、发散还是随机。可视化及面积计算过程在R 3.3.1的“ggplot2”和“zoo”程序包(Achim & Gabor, 2005;
Wickham, 2016 )中完成。
1.2.4 系统发育信号检验
本文采用
Blomberg等(2003) 提出的
K 值法, 以植物最大树高作为功能性状, 检测胡桃楸林内的植物系统发育信号。
K 值为实际的
MSE0 /
MSE 相对于其期望值的比值,
MSE0 是系统发育树末端物种性状数据的均方误差,
MSE 是基于系统发育树的方差-协方差矩阵计算的均方误差, 期望值
MSE0 /
MSE 是基于现有系统发育树的拓扑结构和枝长, 并假设性状以布朗模型方式进化计算得出的。此外, 通过置换检验来判断功能性状的系统发育信号是否显著(
p < 0.05), 随机置换次数为999次。在R的“ape”和“picante”程序包(
Kembel et al ., 2010 ;
Paradis & Schliep, 2019 )中完成
K 值的计算。若
K < 1, 物种表现出趋同性进化趋势;
K = 1, 物种随机进化;
K > 1, 物种进化与系统发育关系紧密, 功能性状保守。
2 结果和分析 2.1 华北地区胡桃楸分布 根据调查结果, 胡桃楸在华北地区分布范围较广, 其分布海拔自320 m至1 700 m, 纬度上跨越10°左右, 经度上跨越13°左右(
图1 )。根据调查结果, 胡桃楸种群密度在河北最大, 河南最小(
表1 )。由
图2 可知, 调查样方内胡桃楸的胸径普遍较小, 胸径为3-5 cm的胡桃楸最多, 胸径大于11 cm的胡桃楸数量较少, 而胸径为15-17 cm的胡桃楸只有2株, 17 cm以上的仅有1株。由
表1 可知, 天津的胡桃楸胸径多样化程度最高, 其中胸径为1-3 cm和7-9 cm的胡桃楸较多, 而且调查样方内胸径最大(17.13 cm)的胡桃楸也生长于天津; 河北胡桃楸的胸径多样性最小, 大部分胸径集中在3-5 cm。山西和北京胡桃楸的胸径多集中于3-7 cm; 陕西胡桃楸的数量较少, 但胸径多样化程度相对较高, 其中胸径为1-3 cm的胡桃楸较多, 但胸径为15-17 cm的2株胡桃楸分布于此。河南胡桃楸的数量最少, 其胸径多样化程度较低, 胸径多为1-3 cm。
Table 1 表1 表1 华北地区各地核桃楸种群密度及各径级胡桃楸数量
Table 1
Juglans mandshurica’ s population density and the numbers of each diameter classes in North China径级 天津 北京 河北 山西 河南 陕西 1-3 109 12 17 7 11 18 3-5 58 38 110 44 1 3 5-7 90 30 0 46 1 1 7-9 104 10 0 22 0 1 9-11 56 5 0 2 0 3 11-13 11 5 0 1 0 0 13-15 5 0 0 0 0 3 15-17 0 0 0 0 0 2 >17 1 0 0 0 0 0 种群密度-2 ) 14 25 53 23 6 9
新窗口打开 |
下载CSV 图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2华北地区胡桃楸胸径分级及其数量。 Fig. 2Diameter classification and quantity of Juglans mandshurica in North China. 2.2 华北地区胡桃楸分布格局及其影响因子 本文将胡桃楸种群分布情况与环境因子进行CCA分析, 模型变量的方差膨胀因子(
VIF )值为7.56, 小于10, 说明模型中共线性不高, 模型可用。根据CCA结果(
表2 )可知, 前两轴与环境因子极显著相关(
p < 0.001), 相关系数分别为0.59、0.31, 前两轴的方差贡献率共为59.52%。
Table 2 表2 表2 华北地区胡桃楸分布与环境因子在前四排序轴上的相关系数
Table 2
Correlation coefficients between distribution of Juglans mandshurica and environmental factors at the first fourth canonical correspondence analysis (CCA) ordination axes in North China 环境因子 CCA排序轴 CCA 1 CCA 2 CCA 3 CCA 4 海拔 Elevation -0.95*** -0.06 -0.27* -0.02 坡度 Slop 0.57*** -0.54*** -0.39*** 0.28* 坡向 Aspect 0.13 0.30 0.57*** -0.49*** 坡位 Slop position 0.45** -0.48*** -0.37*** -0.64*** 人为干扰 Disturbance -0.17** -0.65*** 0.68*** -0.23 特征值 Eigenvalue 0.59*** 0.31*** 0.28*** 0.20*** 累计贡献率 39.08% 59.52% 77.93% 91.39%
*, p < 0.05; **, p < 0.01; ***, p < 0.001. 新窗口打开 |
下载CSV 根据前两轴种群分布-环境因子二维空间排序(
表2 ;
图3 )可知, 在CCA第一轴上, 胡桃楸分布与海拔极显著负相关, 相关系数为0.95; 与坡度极显著正相关, 相关系数为0.57; 与坡位显著正相关, 相关系数为0.45; 与人为干扰显著负相关, 相关系数为0.17, 即沿着第一轴从左到右, 海拔逐渐降低, 坡度增大, 坡位升高, 人为干扰减弱。而对于CCA第二轴上, 坡度、坡位、人为干扰均分别与胡桃楸分布呈显著负相关关系, 沿着第二轴从下到上坡度减小、坡位降低, 人为干扰减弱。坡向在第一、第二轴上相关性不显著, 但在第三轴上呈极显著正相关关系, 在第四轴上呈极显著负相关关系, 因此模型中也保留了坡向这一环境因子。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3华北地区胡桃楸分布与环境因子典范对应分析(CCA)的二维排序图。Ele, 海拔; Asp, 坡向; Slo, 坡度; Slp, 坡位; Dis, 人为干扰。´, 北京; °, 天津; ·, 河北; Fig. 3Two-dimensional ordination diagram of canonical correspondence analysis (CCA) between distribution of Juglans mandshurica and environmental factors in North China. Ele, elevation; Asp, aspect; Slo, slope; Slp, slop position; Dis, disturbance. ´, Beijing; °, Tianjin; ·, Hebei; 根据前两轴种群分布-环境因子二维空间排序中胡桃楸的分布(
图3 )可知, 北京、天津、河北地区的胡桃楸普遍分布在海拔较低、坡度较缓、人为干扰较少的区域; 河南的胡桃楸则集中分布在高海拔、人为干扰程度强的缓坡上; 山西、陕西地区的胡桃楸分布在海拔上呈现出低、中、高3个水平。
2.3 不同分布区域胡桃楸群落构建机制 通过高斯核密度估计结果(
表3 ;
图4 )可知, 北京的胡桃楸林
NRI 、
NTI 趋近于负值, 河北的
NRI 、
NTI 趋近于正值, 山西的
NRI 、
NTI 趋近于负值, 陕西和天津的
NRI 、
NTI 均趋近于正值。山西只有一个胡桃楸样方, 其
NRI 为-2.19,
NTI 为-4.10, 无法估计其整体趋势。
Table 3 表3 表3 5个省市净亲缘关系指数(
NRI )、最近亲缘关系指数(
NTI )在正负范围内的面积
Table 3
Positive and negative area under curve of the net relatedness index (NRI ) and net nearest taxa index (NTI ) in five provinces 北京 Beijing 河北 Hebei 山西 Shanxi 陕西 Shaanxi 天津 NRI <0 0.72 0.08 - 0.00 0.26 >0 0.28 0.92 - 1.00 0.74 NTI <0 0.86 0.08 - 0.00 0.20 >0 0.13 0.92 - 1.00 0.80
新窗口打开 |
下载CSV 图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4亲缘关系指数高斯核密度估计曲线。NRI, 净亲缘关系指数; NTI, 最近亲缘关系指数. Fig. 4Results of Gaussian kernel density estimation of phylogenetic index. NRI, net relatedness index; NTI, nearest taxonomic index. 根据功能性状-最大树高的系统发育信号检测结果(
K = 0.82,
p = 0.001)可知, 这一功能性状检测到了显著的系统发育信号(
p < 0.05), 群落物种的功能性状表现出趋同性进化趋势。根据
NRI 、
NTI 值可知, 河北、陕西地区胡桃楸林物种的系统发育结构聚集(
NRI > 0,
NTI > 0)(
图4 ), 群落由亲缘关系比较密切的物种构成; 天津、北京地区的胡桃楸林物种的系统发育结构有些复杂, 整体上天津呈现聚集状态, 北京呈现发散状态, 但在天津也有部分胡桃楸林的系统发育结构是发散的, 在北京也有部分胡桃楸林的发育结构是聚集的(
NRI 、
NTI 正负都有)(
图4 )。
基于功能性状系统发育信号和亲缘关系指数, 可以认为整体上河北、陕西、天津地区胡桃楸林物种共存是负密度制约作用驱动的, 而北京、山西胡桃楸林物种共存是环境选择作用驱动的。但对于天津地区胡桃楸林也有部分可能受环境选择驱动物种共存, 且在北京也有部分胡桃楸林受负密度制约驱动物种共存。
3 讨论 3.1 胡桃楸种群分布规律 胡桃楸种群在我国主要分布在东北和华北地区, 东北地区主要集中在长白山和小兴安岭海拔200- 1 000 m的阔叶林带和针阔混交林带(
马万里等, 2008 )。在东北长白山地区的胡桃楸种群呈现出衰退型的年龄结构, 幼苗和幼树比例较小(
马万里等, 2008 )。从华北地区的胡桃楸的胸径来看, 华北地区的胡桃楸种群年龄较小, 幼苗、幼树较多(胸径集中在11 cm以下)。这与胡桃楸林在东北、华北地区所处的生境及森林发展历史的差异联系密切(
赵光仪等, 1991 ;
马万里等, 2005 ;
王建文, 2006 )。胡桃楸属第三纪孑遗植物, 起源古老, 东北长白山和小兴安岭地区是胡桃楸的最适生长区, 相对而言, 华北是我国胡桃楸种群分布的南部边缘地区,
苏金娟和王晓春(2017) 的研究表明胡桃楸的生长与气温呈现负相关关系, 因此在生境, 尤其是生物气候方面, 气温较低的东北地区更适合胡桃楸生长, 其种群生长速度比华北地区胡桃楸种群相对要快。另外, 在森林发展历史方面, 东北地区森林历史悠久, 虽遭砍伐, 但仍保有一定面积的原始森林, 其中不乏大量的高龄树木(
王建文, 2006 )。而华北地区因人口众多, 迫于经济等压力, 在新中国成立前此地区的原始森林均被破坏, 当前所看到的森林是新中国成立后通过植树造林、森林保育等措施培育的人工林和衍生的次生林, 因此华北地区成林历史较短, 其树木年龄相对较小(
王建文, 2006 )。
3.2 影响胡桃楸种群分布的因素 根据本研究结果, 华北地区的胡桃楸种群主要分布在燕山山脉海拔较低, 且人为干扰相对较小的山体中下部缓坡区域。海拔、坡度、坡位及人为干扰是影响华北地区胡桃楸种群分布的主要因子。CCA分析结果中环境因子前两轴的方差贡献率为59.52%, 说明还存在其他影响因子, 如小气候和土壤等都是影响植物分布的重要因素(
Condit et al ., 2013 ;
Jones, 2013 )。在华北地区森林群落和环境因子关系的研究中曾发现土壤的相关指标是影响森林群落分布的重要因素(
余敏等, 2013 ;
白晓航等, 2017 )。小气候和土壤因子对华北地区胡桃楸种群分布的影响有待进一步研究。
3.3 不同分布区域胡桃楸林物种共存过程的差异 本研究中华北地区各分布区域的胡桃楸物种共存过程有所不同, 这与各区域内胡桃楸林所处的环境及其自身生长阶段有一定的关系。河北、陕西、天津地区胡桃楸多分布于山体中部、人为干扰程度较小、坡度较缓、海拔较低的生境, 这3个地区的生境适宜植物生长, 环境条件并不是限制物种生存的最大因素, 因此环境选择作用并不是胡桃楸林物种共存的主要驱动因子; 而且这3个地区胡桃楸种群密度较大, 且胸径在1-3 cm的胡桃楸幼苗极多, 植物群落中种子和幼苗的生长易受其母体成树的限制(
Hubbell, 1980 ;
Connell et al ., 1984 ;
Gilbert et al ., 1994 ;
Hyatt et al ., 2003 ;
祝燕等, 2009 ), 因此在这3个地区胡桃楸林的物种共存中起主要驱动作用的是负密度制约作用。相对而言, 北京、山西胡桃楸所处生境的山体坡度、坡位、人为干扰的梯度较大, 生境异质性高, 环境在植物生长共存过程中发挥更大作用。而且北京、山西的胡桃楸林多为胸径3-7 cm的幼树, 在此生长阶段, 母体成树对其的限制作用会有所减弱(
祝燕等, 2009 )。环境条件和自身生长阶段决定了北京、山西的胡桃楸林物种共存过程主要受环境选择作用驱动。
另外, 天津部分胡桃楸林物种共存过程由环境选择作用主导, 北京也有部分群落物种共存由负密度制约作用主导。其原因在于, 首先天津也有部分胸径较大的胡桃楸林存在, 很有可能随着幼苗的长大, 母体成树对其的限制作用在减弱(
Connell et al ., 1984 ;
Condit et al ., 2000 ;
Wright, 2002 ), 负密度制约作用也随之降低; 其次天津地区部分胡桃楸林的人为干扰程度较强, 且处在坡度较陡的山体中上部, 生存环境较为恶劣, 因此环境选择作用主导这部分群落中的物种共存过程。北京的胡桃楸林中也有部分是胸径为1-3 cm的幼苗, 且北京地区部分胡桃楸林所处环境海拔相对较低, 人为干扰较少, 在这部分胡桃楸林中环境可能不是其主要限制因子, 而是负密度制约作用驱动着物种共存。
4 结论 本文对华北地区胡桃楸林的分布规律及其影响因素、群落物种共存过程进行了系统分析。研究发现受森林发展历史等因素的影响, 华北地区的胡桃楸林种群年龄结构较低, 其分布也受众多生物、非生物因子的影响, 低山、缓坡、人为干扰较少的生境是大多胡桃楸种群的分布区。生态位机制主导华北地区胡桃楸林中群落结构及物种组成, 受所处的环境及胡桃楸林自身发展阶段的影响, 环境选择和负密度制约作用在各地发挥不同作用。经比较发现, 在样本量较大, 且群落环境较为复杂的研究中, 高斯核密度估计方法能较为直接地揭示出亲缘关系指数的正负趋势, 能够更为全面地判断群落中物种的亲缘关系。
附录 附录? 华北地区胡桃楸样方信息 Supplement I The plots of
Juglans mandshurica in North China
http://www.plant-ecology.com/fileup/1005-264X/PDF/cjpe.2018.0161-S1.pdf 致谢 感谢承担“华北地区自然植物群落资源综合考察”项目的北京师范大学康慕谊、北京大学刘鸿雁、中国科学院植物研究所高贤明、西北大学岳明、山东大学王仁卿等课题组的项目参与人员, 在获取本研究样方数据中付出的辛勤劳动。感谢天津市农业农村委员会项目(ITTFPRS2018001)和天津市科学技术委员会项目(18ZXZYNC00120)资助。
[1] Bai XH Zhang JT Cao K Wang YQ Sadia S Cao G 2017 ). Relationship between forest communities and the environment in the Xiaowutai Mountain National Nature Reserve, HebeiActa Ecologica Sinica ,3683 -3696 . [本文引用: 1] [ 白晓航 , 张金屯 , 曹科 , 王云泉 , Sehrish Sadia 曹格 ( 2017 ). 河北小五台山国家级自然保护区森林群落与环境的关系生态学报 , 37, 3683 -3696 .] [本文引用: 1] [2] Bazzaz FA 1991 ). Habitat selection in plantsThe American Naturalist ,S116 -S130 . [本文引用: 1] [3] Blomberg SP Garland T Ives AR 2003 ). Testing for phylogenetic signal in comparative data: Behavioral traits are more labileEvolution ,717 -745 . [本文引用: 1] [4] Cavender-Bares J Kozak KH Fine PVA Kembel SW 2009 ). The merging of community ecology and phylogenetic biologyEcology Letters ,693 -715 . [本文引用: 1] [5] Condit R Ashton PS Baker P Bunyavejchewin S Gunatilleke S Gunatilleke N Hubbell SP Foster RB Itoh A LaFrankie JV Lee HS Losos E Manokaran N Sukumar R Yamakura T 2000 ). Spatial patterns in the distribution of tropical tree speciesScience ,1414 -1418 . [本文引用: 1] [6] Condit R Engelbrecht BMJ Pino D Pérez R Turner BL 2013 ). Species distributions in response to individual soil nutrients and seasonal drought across a community of tropical treesProceedings of the National Academy of Sciences of the United States of America ,5064 -5068 . [本文引用: 1] [7] Connell JH Tracey JG Webb LJ 1984 ). Compensatory recruitment, growth, and mortality as factors maintaining rain forest tree diversityEcological Monographs ,141 -164 . [本文引用: 2] [8] D’Amen M Rahbek C Zimmermann NE Guisan A 2017 ). Spatial predictions at the community level: From current approaches to future frameworksBiological Reviews ,169 -187 . [本文引用: 1] [9] Diamond JM 1975 ). Assembly of Species Communities . Harvard University Press, Boston. 342 -344 . [本文引用: 1] [10] Fang JY Wang XP Shen ZH Tang ZY He JS Yu D Jiang Y Wang ZH Zheng CY Zhu JL Guo ZD 2009 ). Methods and protocols for plant community inventoryBiodiversity Science ,533 -548 . [本文引用: 1] [ 方精云 , 王襄平 , 沈泽昊 , 唐志尧 , 贺金生 , 于丹 , 江源 , 王志恒 , 郑成洋 , 朱江玲 , 郭兆迪 ( 2009 ). 植物群落清查的主要内容、方法和技术规范生物多样性 , 17, 533 -548 .] [本文引用: 1] [11] Gao ZY Zhang HF Chen GP Feng XM Zhao TJ Gao X Shi FC 2017 ). Fruit stone morphology and geographic variation in Juglans mandshurica populationsChinese Journal of Applied and Environmental Biology, 609 -615 . [本文引用: 1] [ 高张莹 , 张海峰 , 陈国平 , 冯小梅 , 赵铁建 , 高鑫 , 石福臣 ( 2017 ). 核桃楸种群果核形态及地理变异应用与环境生物学报 , 23, 609 -615 .] [本文引用: 1] [12] Gilbert GS Foster RB Hubbell SP 1994 ). Density and distance?-?to-adult effects of a canker disease of trees in a moist tropical forestOecologia ,100 -108 . [本文引用: 1] [13] Hubbell SP 1980 ). Seed predation and the coexistence of tree species in tropical forestsOikos ,214 -229 . [本文引用: 1] [14] Hubbell SP 2001 ). The Unified Neutral Theory of Biodiversity and Biogeography . Princeton University Press, Princeton. [本文引用: 2] [15] Hyatt LA Rosenberg MS Howard TG Bole G Fang W Anastasia J Brown K Grella R Hinman K Kurdziel JP Gurevitch J 2003 ). The distance dependence prediction of the Janzen-Connell hypothesis: A meta-analysisOikos ,590 -602 . [本文引用: 1] [16] Jari O Blanchet FG Roeland K Pierre L Peter RM O’Hara RB Gavin LS Peter S Stevens MHH Helene W 2016 ). Vegan: Community Ecology Package. R package version: 2.4-1. URL [本文引用: 1] [17] Jones HG 2013 ). Plants and Microclimate: A Quantitative Approach to Environmental Plant Physiology . Cambridge University Press, Cambridge, UK. [本文引用: 1] [18] Kembel SW Cowan PD Helmus MR Cornwell WK Morlon H Ackerly DD Blomberg SP Webb CO 2010 ). Picante: R tools for integrating phylogenies and ecologyBioinformatics ,1463 -1464 . [本文引用: 2] [19] Kraft NJB Cornwell WK Webb CO Ackerly DD 2007 ). Trait evolution, community assembly, and the phylogenetic structure of ecological communitiesThe American Naturalist ,271 -283 . [本文引用: 2] [20] Kraft NJB Valencia R Ackerly DD 2008 ). Functional traits and niche-based tree community assembly in an Amazonian forestScience ,580 -582 . [本文引用: 1] [21] Liang SJ Pan P Sun ZH Wang QC 2005 ). Influence of slope on the growth of Fraxinus mandshurica Rup. and Juglans manshurica Maxim. plantationsJournal of Northeast Forestry University, 3 ), 18 -19 . [本文引用: 1] [ 梁淑娟 , 潘攀 , 孙志虎 , 王庆成 ( 2005 ). 坡位对水曲柳及胡桃楸生长的影响东北林业大学学报 , 33(3 ), 18 -19 .] [本文引用: 1] [22] Ma KP 2016 ). Hot topics for biodiversity scienceBiodiversity Science , 24, 1 -2 . [本文引用: 1] [ 马克平 ( 2016 ). 生物多样性科学的热点问题生物多样性 , 24, 1 -2 .] [本文引用: 1] [23] Ma WL Jing T Kujansuu J Luo JC Sun B Wang GF 2001 ). The dynamics of seed rain and seed bank of Juglans mandshurica population in the Changbai MountainJournal of Beijing Forestry University, 3 ), 70 -72 . [本文引用: 1] [ 马万里 , 荆涛 , Kujansuu J 罗菊春 , 孙波 , 王广发 ( 2001 ). 长白山地区胡桃楸种群的种子雨和种子库动态北京林业大学学报 , 23(3 ), 70 -72 .] [本文引用: 1] [24] Ma WL Luo JC Jing T Kujansuu J 2005 ). Ecological studies and prospect of cultivation on Juglans mandshurica populationJournal of Inner Mongolia Normal University (Natural Science Edition )489 -492 . [本文引用: 1] [ 马万里 , 罗菊春 , 荆涛 , Kujansuu J 2005 ). 珍贵树种核桃楸的生态学问题及培育前景内蒙古师范大学学报(自然科学汉文版) , 34, 489 -492 .] [本文引用: 1] [25] Ma WL Luo JC Jing T Kujansuu J 2008 ). Study on dynamics of Juglans mandshurica population from Changbai MountainBulletin of Botanical Research, 249 -253 . [本文引用: 2] [ 马万里 , 罗菊春 , 荆涛 , Kujansuu J 2008 ). 长白山林区核桃楸种群数量动态变化的研究植物研究 , 28, 249 -253 .] [本文引用: 2] [26] McPeek MA 2007 ). The macroevolutionary consequences of ecological differences among speciesPalaeontology ,111 -129 . [本文引用: 1] [27] Paradis E Schliep K 201P ). ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R.Bioinformatics, 35, 526 -528 . [本文引用: 1] [28] Song NQ Zhang JT Zhao FG 2017 ). The PCA index for measuring functional diversity and its application to Juglans mandshurica communities in the Beijing mountains, ChinaInternational Journal of Biomathematics, 1750097 . DOI: 10.1142/S1793524517500978 . [本文引用: 1] [29] Su JJ Wang XC 2017 ). Spatio-temporal variations in climate-?growth relationships of three hardwood tree species across the north Zhangguangcai Mountains, northeast ChinaActa Ecologica Sinica ,1484 -1495 . [本文引用: 2] [ 苏金娟 , 王晓春 ( 2017 ). 张广才岭北部三大硬阔树木生长-气候关系的时空变异生态学报 , 37, 1484 -1495 .] [本文引用: 2] [30] Swenson NG 2011 ). The role of evolutionary processes in producing biodiversity patterns, and the interrelationships between taxonomic, functional and phylogenetic biodiversityAmerican Journal of Botany ,472 -480 . [本文引用: 1] [31] Tang LL Chen GP Feng XM Zhao TJ Shi FC 2017 ). Community assembly rules of the east of Yanshan Mountain based on phylogenyBulletin of Botanical Research ,807 -815 . [本文引用: 1] [ 唐丽丽 , 陈国平 , 冯小梅 , 赵铁建 , 石福臣 ( 2017 ). 基于系统发育的燕山东麓植物群落的构建机制植物研究 , 37, 807 -815 .] [本文引用: 1] [32] Vamosi SM Heard SB Vamosi JC Webb CO 2009 ). Emerging patterns in the comparative analysis of phylogenetic community structureMolecular Ecology ,572 -592 . [本文引用: 2] [33] Wang DL 2014 ). The Research on the Interspecies Allelopathic Effect of Juglans mandshurica and Larix gmelinii Forest Plantation. Master degree dissertation, Northeast Forestry University, Harbin. [本文引用: 1] [ 王东亮 ( 2014 ). 胡桃楸-落叶松林地种间化感效应的研究硕士学位论文, 东北林业大学, 哈尔滨 .] [本文引用: 1] [34] Wang DN Mu CC Gao Z Feng FJ 2011 ). ISSR analysis of genetic diversity of Juglans mandshurica Maxim. populationsNonwood Forest Research, 2 ), 22 -29 . [本文引用: 1] [ 王东娜 , 牟长城 , 高卓 , 冯福娟 ( 2011 ). 胡桃楸天然种群遗传多样性的ISSR分析经济林研究 , 29(2 ), 22 -29 .] [本文引用: 1] [35] Wang JW 2006 ). A Study on the Historical Change of Foreset\Grassland and Eco-catastrophes in North China. PhD dissertation, Beijing Forestry University, Beijing. [本文引用: 3] [ 王建文 ( 2006 ). 中国北方地区森林、草原变迁和生态灾害的历史研究博士学位论文, 北京林业大学, 北京 .] [本文引用: 3] [36] Webb CO Ackerly DD Kembel SW 2008 ). Phylocom: Software for the analysis of phylogenetic community structure and trait evolutionBioinformatics ,2098 -2100 . [本文引用: 1] [37] Webb CO Ackerly DD McPeek MA Donoghue MJ 2002 ). Phylogenies and community ecologyAnnual Review of Ecology and Systematics ,475 -505 . [本文引用: 5] [38] Webb CO Donoghue MJ 2005 ). Phylomatic: Tree assembly for applied phylogeneticsMolecular Ecology Notes ,181 -183 . [本文引用: 1] [39] Weiher E Keddy P 1999 ). Ecological Assembly Rules: Perspectives, Advances, Retreats . Cambridge University Press, Cambridge, UK. [本文引用: 1] [40] Wickham H 2016 ). ggplot2: Elegant Graphics for Data Analysis. Springer, New York. [本文引用: 1] [41] Wright IJ Reich PB Westoby M Ackerly DD Baruch Z Bongers F Cavender-Bares J Chapin T Cornelissen JHC Diemer M Flexas J Garnier E Groom PK Gulias J Hikosaka K Lamont BB Lee T Lee W Lusk C Midgley JJ Navas ML Niinemets ü Oleksyn J Osada N Poorter H Poot P Prior L Pyankov VI Roumet C Thomas SC Tjoelker MG Veneklaas EJ Villar R 2004 ). The worldwide leaf economics spectrum. Nature ,821 -827 . [本文引用: 1] [42] Wright SJ 2002 ). Plant diversity in tropical forests: A review of mechanisms of species coexistenceOecologia ,1 -14 . [本文引用: 1] [43] Yan N Chen T Chen YL Wu XL Wu LJ Dawa CM Basang DJ Xu JY 2014 ). Characteristics of population structure Juglans mandshurica from the Wuling Mountain Nature ReserveJournal of Capital Normal University (Natural Sciences Edition )6 ), 64 -67, 81 . [本文引用: 1] [ 闫娜 , 陈彤 , 陈云丽 , 吴晓丽 , 吴丽娟 , 达瓦措姆 , 巴桑多吉 , 徐建英 ( 2014 ). 雾灵山自然保护区胡桃楸种群结构特征分析首都师范大学学报(自然科学版) , 35(6 ), 64 -67, 81 .] [本文引用: 1] [44] Yu M Zhou ZY Kang FF Ouyang S Mi XC Sun JX 2013 ). Gradient analysis and environmental interpretation of understory herb-layer communities in Xiaoshegou of Lingkong Mountain, Shanxi, ChinaChinese Journal of Plant Ecology ,373 -383 . [本文引用: 1] [ 余敏 , 周志勇 , 康峰峰 , 欧阳帅 , 米湘成 , 孙建新 ( 2013 ). 山西灵空山小蛇沟林下草本层植物群落梯度分析及环境解释植物生态学报 , 37, 373 -383 .] [本文引用: 1] [45] Zeileis A Grothendieck G 2005 ). Zoo: S3 infrastructure for regular and irregular time seriesJournal of Statistical Software ,1 -27 . [46] Zhao GY Tian XJ Wu ZH 1991 ). Analysis and discussion on the north distributing limitation of amur corktree, manchurian walnut and manchurian ashJournal of Northest Forestry University ,Suppl. 1 ), 290 -295 . [本文引用: 1] [ 赵光仪 , 田兴军 , 吴振河 ( 1991 ). 黄波罗、胡桃楸、水曲柳分布北限论析东北林业大学学报 , 19(Suppl. 1 ), 290 -295 .] [本文引用: 1] [47] Zhu Y Mi XC Ma KP 2009 ). A mechanism of plant species coexistence: The negative density-dependent hypothesisBiodiversity Science ,594 -604 . [本文引用: 2] [ 祝燕 , 米湘成 , 马克平 ( 2009 ). 植物群落物种共存机制: 负密度制约假说生物多样性 , 17, 594 -604 .] [本文引用: 2] 河北小五台山国家级自然保护区森林群落与环境的关系
1
2017
... 根据本研究结果, 华北地区的胡桃楸种群主要分布在燕山山脉海拔较低, 且人为干扰相对较小的山体中下部缓坡区域.海拔、坡度、坡位及人为干扰是影响华北地区胡桃楸种群分布的主要因子.CCA分析结果中环境因子前两轴的方差贡献率为59.52%, 说明还存在其他影响因子, 如小气候和土壤等都是影响植物分布的重要因素(
Condit et al ., 2013 ;
Jones, 2013 ).在华北地区森林群落和环境因子关系的研究中曾发现土壤的相关指标是影响森林群落分布的重要因素(
余敏等, 2013 ;
白晓航等, 2017 ).小气候和土壤因子对华北地区胡桃楸种群分布的影响有待进一步研究. ...
河北小五台山国家级自然保护区森林群落与环境的关系
1
2017
... 根据本研究结果, 华北地区的胡桃楸种群主要分布在燕山山脉海拔较低, 且人为干扰相对较小的山体中下部缓坡区域.海拔、坡度、坡位及人为干扰是影响华北地区胡桃楸种群分布的主要因子.CCA分析结果中环境因子前两轴的方差贡献率为59.52%, 说明还存在其他影响因子, 如小气候和土壤等都是影响植物分布的重要因素(
Condit et al ., 2013 ;
Jones, 2013 ).在华北地区森林群落和环境因子关系的研究中曾发现土壤的相关指标是影响森林群落分布的重要因素(
余敏等, 2013 ;
白晓航等, 2017 ).小气候和土壤因子对华北地区胡桃楸种群分布的影响有待进一步研究. ...
Habitat selection in plants
1
1991
... 群落构建机制即群落物种多样性形成及其维持机制, 主要研究植物群落如何构建, 什么驱使植物的多样性及群落功能的变化(
Cavender-Bares et al ., 2009 ).目前对群落构建机制的分析主要基于谱系发育和植物功能性状(
Webb et al ., 2002 ;
Wright, 2004 ;
Kraft et al ., 2008 ;
Swenson, 2011 ;
马克平, 2016 ;
D’Amen, 2017 ), 对群落构建机制的解释包括生态位机制(
Diamond, 1975 )和中性理论(
Hubbell, 2001 ).中性理论认为群落内物种的出生、死亡、迁入、迁出处于一个动态的平衡, 这种动态平衡维持着群落内物种数量的稳定(
Hubbell, 2001 ).生态位机制则认为群落内植物的生长繁殖、迁入迁出源自于群落的生境、资源及生物间相互作用, 生态位机制是由环境过滤和负密度制约(
Bazzaz, 1991 ;
Weiher & Keddy, 1999 )两个过程主导群落构建. ...
Testing for phylogenetic signal in comparative data: Behavioral traits are more labile
1
2003
... 本文采用
Blomberg等(2003) 提出的
K 值法, 以植物最大树高作为功能性状, 检测胡桃楸林内的植物系统发育信号.
K 值为实际的
MSE0 /
MSE 相对于其期望值的比值,
MSE0 是系统发育树末端物种性状数据的均方误差,
MSE 是基于系统发育树的方差-协方差矩阵计算的均方误差, 期望值
MSE0 /
MSE 是基于现有系统发育树的拓扑结构和枝长, 并假设性状以布朗模型方式进化计算得出的.此外, 通过置换检验来判断功能性状的系统发育信号是否显著(
p < 0.05), 随机置换次数为999次.在R的“ape”和“picante”程序包(
Kembel et al ., 2010 ;
Paradis & Schliep, 2019 )中完成
K 值的计算.若
K < 1, 物种表现出趋同性进化趋势;
K = 1, 物种随机进化;
K > 1, 物种进化与系统发育关系紧密, 功能性状保守. ...
The merging of community ecology and phylogenetic biology
1
2009
... 群落构建机制即群落物种多样性形成及其维持机制, 主要研究植物群落如何构建, 什么驱使植物的多样性及群落功能的变化(
Cavender-Bares et al ., 2009 ).目前对群落构建机制的分析主要基于谱系发育和植物功能性状(
Webb et al ., 2002 ;
Wright, 2004 ;
Kraft et al ., 2008 ;
Swenson, 2011 ;
马克平, 2016 ;
D’Amen, 2017 ), 对群落构建机制的解释包括生态位机制(
Diamond, 1975 )和中性理论(
Hubbell, 2001 ).中性理论认为群落内物种的出生、死亡、迁入、迁出处于一个动态的平衡, 这种动态平衡维持着群落内物种数量的稳定(
Hubbell, 2001 ).生态位机制则认为群落内植物的生长繁殖、迁入迁出源自于群落的生境、资源及生物间相互作用, 生态位机制是由环境过滤和负密度制约(
Bazzaz, 1991 ;
Weiher & Keddy, 1999 )两个过程主导群落构建. ...
Spatial patterns in the distribution of tropical tree species
1
2000
... 另外, 天津部分胡桃楸林物种共存过程由环境选择作用主导, 北京也有部分群落物种共存由负密度制约作用主导.其原因在于, 首先天津也有部分胸径较大的胡桃楸林存在, 很有可能随着幼苗的长大, 母体成树对其的限制作用在减弱(
Connell et al ., 1984 ;
Condit et al ., 2000 ;
Wright, 2002 ), 负密度制约作用也随之降低; 其次天津地区部分胡桃楸林的人为干扰程度较强, 且处在坡度较陡的山体中上部, 生存环境较为恶劣, 因此环境选择作用主导这部分群落中的物种共存过程.北京的胡桃楸林中也有部分是胸径为1-3 cm的幼苗, 且北京地区部分胡桃楸林所处环境海拔相对较低, 人为干扰较少, 在这部分胡桃楸林中环境可能不是其主要限制因子, 而是负密度制约作用驱动着物种共存. ...
Species distributions in response to individual soil nutrients and seasonal drought across a community of tropical trees
1
2013
... 根据本研究结果, 华北地区的胡桃楸种群主要分布在燕山山脉海拔较低, 且人为干扰相对较小的山体中下部缓坡区域.海拔、坡度、坡位及人为干扰是影响华北地区胡桃楸种群分布的主要因子.CCA分析结果中环境因子前两轴的方差贡献率为59.52%, 说明还存在其他影响因子, 如小气候和土壤等都是影响植物分布的重要因素(
Condit et al ., 2013 ;
Jones, 2013 ).在华北地区森林群落和环境因子关系的研究中曾发现土壤的相关指标是影响森林群落分布的重要因素(
余敏等, 2013 ;
白晓航等, 2017 ).小气候和土壤因子对华北地区胡桃楸种群分布的影响有待进一步研究. ...
Compensatory recruitment, growth, and mortality as factors maintaining rain forest tree diversity
2
1984
... 本研究中华北地区各分布区域的胡桃楸物种共存过程有所不同, 这与各区域内胡桃楸林所处的环境及其自身生长阶段有一定的关系.河北、陕西、天津地区胡桃楸多分布于山体中部、人为干扰程度较小、坡度较缓、海拔较低的生境, 这3个地区的生境适宜植物生长, 环境条件并不是限制物种生存的最大因素, 因此环境选择作用并不是胡桃楸林物种共存的主要驱动因子; 而且这3个地区胡桃楸种群密度较大, 且胸径在1-3 cm的胡桃楸幼苗极多, 植物群落中种子和幼苗的生长易受其母体成树的限制(
Hubbell, 1980 ;
Connell et al ., 1984 ;
Gilbert et al ., 1994 ;
Hyatt et al ., 2003 ;
祝燕等, 2009 ), 因此在这3个地区胡桃楸林的物种共存中起主要驱动作用的是负密度制约作用.相对而言, 北京、山西胡桃楸所处生境的山体坡度、坡位、人为干扰的梯度较大, 生境异质性高, 环境在植物生长共存过程中发挥更大作用.而且北京、山西的胡桃楸林多为胸径3-7 cm的幼树, 在此生长阶段, 母体成树对其的限制作用会有所减弱(
祝燕等, 2009 ).环境条件和自身生长阶段决定了北京、山西的胡桃楸林物种共存过程主要受环境选择作用驱动. ...
... 另外, 天津部分胡桃楸林物种共存过程由环境选择作用主导, 北京也有部分群落物种共存由负密度制约作用主导.其原因在于, 首先天津也有部分胸径较大的胡桃楸林存在, 很有可能随着幼苗的长大, 母体成树对其的限制作用在减弱(
Connell et al ., 1984 ;
Condit et al ., 2000 ;
Wright, 2002 ), 负密度制约作用也随之降低; 其次天津地区部分胡桃楸林的人为干扰程度较强, 且处在坡度较陡的山体中上部, 生存环境较为恶劣, 因此环境选择作用主导这部分群落中的物种共存过程.北京的胡桃楸林中也有部分是胸径为1-3 cm的幼苗, 且北京地区部分胡桃楸林所处环境海拔相对较低, 人为干扰较少, 在这部分胡桃楸林中环境可能不是其主要限制因子, 而是负密度制约作用驱动着物种共存. ...
Spatial predictions at the community level: From current approaches to future frameworks
1
2017
... 群落构建机制即群落物种多样性形成及其维持机制, 主要研究植物群落如何构建, 什么驱使植物的多样性及群落功能的变化(
Cavender-Bares et al ., 2009 ).目前对群落构建机制的分析主要基于谱系发育和植物功能性状(
Webb et al ., 2002 ;
Wright, 2004 ;
Kraft et al ., 2008 ;
Swenson, 2011 ;
马克平, 2016 ;
D’Amen, 2017 ), 对群落构建机制的解释包括生态位机制(
Diamond, 1975 )和中性理论(
Hubbell, 2001 ).中性理论认为群落内物种的出生、死亡、迁入、迁出处于一个动态的平衡, 这种动态平衡维持着群落内物种数量的稳定(
Hubbell, 2001 ).生态位机制则认为群落内植物的生长繁殖、迁入迁出源自于群落的生境、资源及生物间相互作用, 生态位机制是由环境过滤和负密度制约(
Bazzaz, 1991 ;
Weiher & Keddy, 1999 )两个过程主导群落构建. ...
1
1975
... 群落构建机制即群落物种多样性形成及其维持机制, 主要研究植物群落如何构建, 什么驱使植物的多样性及群落功能的变化(
Cavender-Bares et al ., 2009 ).目前对群落构建机制的分析主要基于谱系发育和植物功能性状(
Webb et al ., 2002 ;
Wright, 2004 ;
Kraft et al ., 2008 ;
Swenson, 2011 ;
马克平, 2016 ;
D’Amen, 2017 ), 对群落构建机制的解释包括生态位机制(
Diamond, 1975 )和中性理论(
Hubbell, 2001 ).中性理论认为群落内物种的出生、死亡、迁入、迁出处于一个动态的平衡, 这种动态平衡维持着群落内物种数量的稳定(
Hubbell, 2001 ).生态位机制则认为群落内植物的生长繁殖、迁入迁出源自于群落的生境、资源及生物间相互作用, 生态位机制是由环境过滤和负密度制约(
Bazzaz, 1991 ;
Weiher & Keddy, 1999 )两个过程主导群落构建. ...
植物群落清查的主要内容、方法和技术规范
1
2009
... 华北地区胡桃楸林主要为天然次生林, 本文采用样方法对其进行野外调查, 样方设置方法参照
方精云等(2009) 的植物群落清查的技术规范, 样方尺度主要为20 m × 30 m, 记录样方内所有胸径(
DBH ) ≥1 cm的木本植物和所有草本植物的相关信息, 其中木本植物主要记录种类、数量、树高、胸径, 草本植物主要记录种类、多度、盖度, 同时记录样地生境信息, 如样方坐标、海拔、坡度、坡向等信息.根据各地胡桃楸种群分布及规模, 在研究区域内共设12个样地, 其中北京3个、天津1个、河北2个、山西2个、陕西3个、河南1个, 12个样地中共设有84个样方(附录I), 样地具体分布位置如
图1 所示.根据相关技术规范, 本研究采用2 cm径距将胡桃楸胸径划分为9个径级, 并结合其分布位点, 利用ArcGIS 10.2分析不同径级胡桃楸在华北地区的分布. ...
植物群落清查的主要内容、方法和技术规范
1
2009
... 华北地区胡桃楸林主要为天然次生林, 本文采用样方法对其进行野外调查, 样方设置方法参照
方精云等(2009) 的植物群落清查的技术规范, 样方尺度主要为20 m × 30 m, 记录样方内所有胸径(
DBH ) ≥1 cm的木本植物和所有草本植物的相关信息, 其中木本植物主要记录种类、数量、树高、胸径, 草本植物主要记录种类、多度、盖度, 同时记录样地生境信息, 如样方坐标、海拔、坡度、坡向等信息.根据各地胡桃楸种群分布及规模, 在研究区域内共设12个样地, 其中北京3个、天津1个、河北2个、山西2个、陕西3个、河南1个, 12个样地中共设有84个样方(附录I), 样地具体分布位置如
图1 所示.根据相关技术规范, 本研究采用2 cm径距将胡桃楸胸径划分为9个径级, 并结合其分布位点, 利用ArcGIS 10.2分析不同径级胡桃楸在华北地区的分布. ...
核桃楸种群果核形态及地理变异
1
2017
... 胡桃楸(
Juglans mandshurica )又名核桃楸, 隶属于胡桃科胡桃属(
Juglans ), 属第三纪孑遗植物, 在我国多分布于东北的东部山地.同时, 胡桃楸的分布区域也在向南延伸, 扩展到我国华北地区, 主要为北京、天津等地的山区, 且颇具规模.目前针对东北地区分布的胡桃楸种群的研究较为丰富全面(
马万里等, 2001 ;
梁淑娟等, 2005 ;
王东娜等, 2011 ;
王东亮, 2014 ;
苏金娟和王晓春, 2017 ), 而华北地区胡桃楸种群的研究相对较少, 且多集中在胡桃楸生理学特性及群落结构等方面(
闫娜等, 2014 ;
高张莹等, 2017 ,
Song et al ., 2017 ).华北地区胡桃楸种群的分布规律、影响其分布的环境因子、胡桃楸林的系统发育结构, 以及群落构建机制都尚未清楚.本文利用在“华北地区自然植物群落资源综合考察”中获得的大量胡桃楸样方数据, 对华北地区胡桃楸种群分布规律及其影响因素进行分析, 并综合谱系发育和功能性状探讨胡桃楸林的物种共存过程, 为胡桃楸林及其生境的保护提供科学依据, 也为进一步探究华北地区胡桃楸林物种组成及群落发展方向奠定基础. ...
核桃楸种群果核形态及地理变异
1
2017
... 胡桃楸(
Juglans mandshurica )又名核桃楸, 隶属于胡桃科胡桃属(
Juglans ), 属第三纪孑遗植物, 在我国多分布于东北的东部山地.同时, 胡桃楸的分布区域也在向南延伸, 扩展到我国华北地区, 主要为北京、天津等地的山区, 且颇具规模.目前针对东北地区分布的胡桃楸种群的研究较为丰富全面(
马万里等, 2001 ;
梁淑娟等, 2005 ;
王东娜等, 2011 ;
王东亮, 2014 ;
苏金娟和王晓春, 2017 ), 而华北地区胡桃楸种群的研究相对较少, 且多集中在胡桃楸生理学特性及群落结构等方面(
闫娜等, 2014 ;
高张莹等, 2017 ,
Song et al ., 2017 ).华北地区胡桃楸种群的分布规律、影响其分布的环境因子、胡桃楸林的系统发育结构, 以及群落构建机制都尚未清楚.本文利用在“华北地区自然植物群落资源综合考察”中获得的大量胡桃楸样方数据, 对华北地区胡桃楸种群分布规律及其影响因素进行分析, 并综合谱系发育和功能性状探讨胡桃楸林的物种共存过程, 为胡桃楸林及其生境的保护提供科学依据, 也为进一步探究华北地区胡桃楸林物种组成及群落发展方向奠定基础. ...
Density and distance?-?to-adult effects of a canker disease of trees in a moist tropical forest
1
1994
... 本研究中华北地区各分布区域的胡桃楸物种共存过程有所不同, 这与各区域内胡桃楸林所处的环境及其自身生长阶段有一定的关系.河北、陕西、天津地区胡桃楸多分布于山体中部、人为干扰程度较小、坡度较缓、海拔较低的生境, 这3个地区的生境适宜植物生长, 环境条件并不是限制物种生存的最大因素, 因此环境选择作用并不是胡桃楸林物种共存的主要驱动因子; 而且这3个地区胡桃楸种群密度较大, 且胸径在1-3 cm的胡桃楸幼苗极多, 植物群落中种子和幼苗的生长易受其母体成树的限制(
Hubbell, 1980 ;
Connell et al ., 1984 ;
Gilbert et al ., 1994 ;
Hyatt et al ., 2003 ;
祝燕等, 2009 ), 因此在这3个地区胡桃楸林的物种共存中起主要驱动作用的是负密度制约作用.相对而言, 北京、山西胡桃楸所处生境的山体坡度、坡位、人为干扰的梯度较大, 生境异质性高, 环境在植物生长共存过程中发挥更大作用.而且北京、山西的胡桃楸林多为胸径3-7 cm的幼树, 在此生长阶段, 母体成树对其的限制作用会有所减弱(
祝燕等, 2009 ).环境条件和自身生长阶段决定了北京、山西的胡桃楸林物种共存过程主要受环境选择作用驱动. ...
Seed predation and the coexistence of tree species in tropical forests
1
1980
... 本研究中华北地区各分布区域的胡桃楸物种共存过程有所不同, 这与各区域内胡桃楸林所处的环境及其自身生长阶段有一定的关系.河北、陕西、天津地区胡桃楸多分布于山体中部、人为干扰程度较小、坡度较缓、海拔较低的生境, 这3个地区的生境适宜植物生长, 环境条件并不是限制物种生存的最大因素, 因此环境选择作用并不是胡桃楸林物种共存的主要驱动因子; 而且这3个地区胡桃楸种群密度较大, 且胸径在1-3 cm的胡桃楸幼苗极多, 植物群落中种子和幼苗的生长易受其母体成树的限制(
Hubbell, 1980 ;
Connell et al ., 1984 ;
Gilbert et al ., 1994 ;
Hyatt et al ., 2003 ;
祝燕等, 2009 ), 因此在这3个地区胡桃楸林的物种共存中起主要驱动作用的是负密度制约作用.相对而言, 北京、山西胡桃楸所处生境的山体坡度、坡位、人为干扰的梯度较大, 生境异质性高, 环境在植物生长共存过程中发挥更大作用.而且北京、山西的胡桃楸林多为胸径3-7 cm的幼树, 在此生长阶段, 母体成树对其的限制作用会有所减弱(
祝燕等, 2009 ).环境条件和自身生长阶段决定了北京、山西的胡桃楸林物种共存过程主要受环境选择作用驱动. ...
2
2001
... 群落构建机制即群落物种多样性形成及其维持机制, 主要研究植物群落如何构建, 什么驱使植物的多样性及群落功能的变化(
Cavender-Bares et al ., 2009 ).目前对群落构建机制的分析主要基于谱系发育和植物功能性状(
Webb et al ., 2002 ;
Wright, 2004 ;
Kraft et al ., 2008 ;
Swenson, 2011 ;
马克平, 2016 ;
D’Amen, 2017 ), 对群落构建机制的解释包括生态位机制(
Diamond, 1975 )和中性理论(
Hubbell, 2001 ).中性理论认为群落内物种的出生、死亡、迁入、迁出处于一个动态的平衡, 这种动态平衡维持着群落内物种数量的稳定(
Hubbell, 2001 ).生态位机制则认为群落内植物的生长繁殖、迁入迁出源自于群落的生境、资源及生物间相互作用, 生态位机制是由环境过滤和负密度制约(
Bazzaz, 1991 ;
Weiher & Keddy, 1999 )两个过程主导群落构建. ...
... ).中性理论认为群落内物种的出生、死亡、迁入、迁出处于一个动态的平衡, 这种动态平衡维持着群落内物种数量的稳定(
Hubbell, 2001 ).生态位机制则认为群落内植物的生长繁殖、迁入迁出源自于群落的生境、资源及生物间相互作用, 生态位机制是由环境过滤和负密度制约(
Bazzaz, 1991 ;
Weiher & Keddy, 1999 )两个过程主导群落构建. ...
The distance dependence prediction of the Janzen-Connell hypothesis: A meta-analysis
1
2003
... 本研究中华北地区各分布区域的胡桃楸物种共存过程有所不同, 这与各区域内胡桃楸林所处的环境及其自身生长阶段有一定的关系.河北、陕西、天津地区胡桃楸多分布于山体中部、人为干扰程度较小、坡度较缓、海拔较低的生境, 这3个地区的生境适宜植物生长, 环境条件并不是限制物种生存的最大因素, 因此环境选择作用并不是胡桃楸林物种共存的主要驱动因子; 而且这3个地区胡桃楸种群密度较大, 且胸径在1-3 cm的胡桃楸幼苗极多, 植物群落中种子和幼苗的生长易受其母体成树的限制(
Hubbell, 1980 ;
Connell et al ., 1984 ;
Gilbert et al ., 1994 ;
Hyatt et al ., 2003 ;
祝燕等, 2009 ), 因此在这3个地区胡桃楸林的物种共存中起主要驱动作用的是负密度制约作用.相对而言, 北京、山西胡桃楸所处生境的山体坡度、坡位、人为干扰的梯度较大, 生境异质性高, 环境在植物生长共存过程中发挥更大作用.而且北京、山西的胡桃楸林多为胸径3-7 cm的幼树, 在此生长阶段, 母体成树对其的限制作用会有所减弱(
祝燕等, 2009 ).环境条件和自身生长阶段决定了北京、山西的胡桃楸林物种共存过程主要受环境选择作用驱动. ...
1
2016
... 根据样地调查结果及华北地区数字高程图(来源于中国科学院资源环境科学数据中心), 利用ArcGIS 10.2, 分析华北地区胡桃楸的空间分布规律.使用典范对应分析(CCA)方法, 定量分析海拔、坡度、坡向、坡位、人为干扰对胡桃楸在华北地区分布的影响.其中坡向按照其度数分为北、东北、西北、东、西、东南、西南、南, 分别用编号1-8代表.坡位分为谷地、下、中下、中、中上和上6个坡位, 编号1-6.人为干扰主要为伐木, 根据国家标准生物多样性观测技术导则(HJ710.12-2016)中对人为干扰活动的分类, 将人为干扰分为4个程度: 无干扰指生境及植物群落没有受到人为干扰, 植被保持原始状态, 对野生植物的生存繁衍没有任何影响, 在计算中赋值为1; 轻微干扰指生境及植物群落受到一定程度的影响, 但植被基本保持原始状态, 对野生植物生存繁衍影响不大, 在计算中赋值为2; 中度干扰指生境及植物群落受到干扰, 部分植被消失, 但干扰消失后, 植被仍可恢复, 野生植物生存繁衍受到一定程度的影响, 但仍然可以栖息繁衍, 在计算中赋值为3; 重度干扰指生境及植物群落受到严重干扰, 植被基本消失, 野生植物难以生存繁衍, 在计算中赋值为4.此过程在R 3.3.1中的“vegan”程序包(
Jari et al ., 2016 )中进行. ...
1
2013
... 根据本研究结果, 华北地区的胡桃楸种群主要分布在燕山山脉海拔较低, 且人为干扰相对较小的山体中下部缓坡区域.海拔、坡度、坡位及人为干扰是影响华北地区胡桃楸种群分布的主要因子.CCA分析结果中环境因子前两轴的方差贡献率为59.52%, 说明还存在其他影响因子, 如小气候和土壤等都是影响植物分布的重要因素(
Condit et al ., 2013 ;
Jones, 2013 ).在华北地区森林群落和环境因子关系的研究中曾发现土壤的相关指标是影响森林群落分布的重要因素(
余敏等, 2013 ;
白晓航等, 2017 ).小气候和土壤因子对华北地区胡桃楸种群分布的影响有待进一步研究. ...
Picante: R tools for integrating phylogenies and ecology
2
2010
... 本文参考
Webb等(2002) 的实验方法构建系统发育树.将样方调查中得到的种子植物, 按照科/属/种的格式, 输入到phylomatic平台(
Webb & Dongoghue, 2005 ;
Webb et al ., 2008 ), 得到基于APG III分类系统的具有进化枝长的系统发育树.根据各物种在系统发育树上的枝长和所在样方中的数量, 计算得出各样方的亲缘关系指数(
NRI 和
NTI ).此过程在R 3.3.1的“picante”程序包(
Kembel et al ., 2010 )中进行.因河南的胡桃楸数量较少, 仅在落叶栎林和华山松林中零散分布, 因此并未对河南的胡桃楸数据进行群落构建机制分析. ...
... 本文采用
Blomberg等(2003) 提出的
K 值法, 以植物最大树高作为功能性状, 检测胡桃楸林内的植物系统发育信号.
K 值为实际的
MSE0 /
MSE 相对于其期望值的比值,
MSE0 是系统发育树末端物种性状数据的均方误差,
MSE 是基于系统发育树的方差-协方差矩阵计算的均方误差, 期望值
MSE0 /
MSE 是基于现有系统发育树的拓扑结构和枝长, 并假设性状以布朗模型方式进化计算得出的.此外, 通过置换检验来判断功能性状的系统发育信号是否显著(
p < 0.05), 随机置换次数为999次.在R的“ape”和“picante”程序包(
Kembel et al ., 2010 ;
Paradis & Schliep, 2019 )中完成
K 值的计算.若
K < 1, 物种表现出趋同性进化趋势;
K = 1, 物种随机进化;
K > 1, 物种进化与系统发育关系紧密, 功能性状保守. ...
Trait evolution, community assembly, and the phylogenetic structure of ecological communities
2
2007
... 随着相关学科领域的快速发展,
Webb等(2002) 提出用植物谱系发育理念来理解群落物种共存过程, 并提出净亲缘指数(
NRI )和最近亲缘指数(
NTI )这两种亲缘关系指数, 以此来计算群落内物种间的系统发育距离, 并在植物功能性状生态位保守和趋同两种情况下讨论群落的构建机制(
Kraft et al ., 2007 ;
Vamosi et al ., 2009 ).若亲缘关系指数大于0, 则物种系统发育距离较小, 群落由亲缘关系较近的物种构成; 若亲缘关系指数小于0, 说明物种系统发育距离较大, 群落由亲缘关系较远的物种构成; 若亲缘关系指数为0, 说明物种在系统发育结构上呈现随机状态(
Webb et al ., 2002 ).若群落物种生态位保守, 环境过滤作用会导致群落谱系结构的聚集, 竞争排斥(负密度制约)作用会导致群落谱系结构的发散; 若群落物种生态位趋同, 环境过滤作用会导致群落谱系结构发散, 负密度制约作用会导致群落谱系结构随机或聚集(
Webb et al ., 2002 ;
Kraft et al ., 2007 ;
Vamosi et al ., 2009 ).在中性作用下, 群落谱系结构均呈随机状态(
McPeek, 2007 ).群落生态学与植物谱系发育概念的结合, 为解释群落的构建提供了新的理念. ...
... ;
Kraft et al ., 2007 ;
Vamosi et al ., 2009 ).在中性作用下, 群落谱系结构均呈随机状态(
McPeek, 2007 ).群落生态学与植物谱系发育概念的结合, 为解释群落的构建提供了新的理念. ...
Functional traits and niche-based tree community assembly in an Amazonian forest
1
2008
... 群落构建机制即群落物种多样性形成及其维持机制, 主要研究植物群落如何构建, 什么驱使植物的多样性及群落功能的变化(
Cavender-Bares et al ., 2009 ).目前对群落构建机制的分析主要基于谱系发育和植物功能性状(
Webb et al ., 2002 ;
Wright, 2004 ;
Kraft et al ., 2008 ;
Swenson, 2011 ;
马克平, 2016 ;
D’Amen, 2017 ), 对群落构建机制的解释包括生态位机制(
Diamond, 1975 )和中性理论(
Hubbell, 2001 ).中性理论认为群落内物种的出生、死亡、迁入、迁出处于一个动态的平衡, 这种动态平衡维持着群落内物种数量的稳定(
Hubbell, 2001 ).生态位机制则认为群落内植物的生长繁殖、迁入迁出源自于群落的生境、资源及生物间相互作用, 生态位机制是由环境过滤和负密度制约(
Bazzaz, 1991 ;
Weiher & Keddy, 1999 )两个过程主导群落构建. ...
坡位对水曲柳及胡桃楸生长的影响
1
2005
... 胡桃楸(
Juglans mandshurica )又名核桃楸, 隶属于胡桃科胡桃属(
Juglans ), 属第三纪孑遗植物, 在我国多分布于东北的东部山地.同时, 胡桃楸的分布区域也在向南延伸, 扩展到我国华北地区, 主要为北京、天津等地的山区, 且颇具规模.目前针对东北地区分布的胡桃楸种群的研究较为丰富全面(
马万里等, 2001 ;
梁淑娟等, 2005 ;
王东娜等, 2011 ;
王东亮, 2014 ;
苏金娟和王晓春, 2017 ), 而华北地区胡桃楸种群的研究相对较少, 且多集中在胡桃楸生理学特性及群落结构等方面(
闫娜等, 2014 ;
高张莹等, 2017 ,
Song et al ., 2017 ).华北地区胡桃楸种群的分布规律、影响其分布的环境因子、胡桃楸林的系统发育结构, 以及群落构建机制都尚未清楚.本文利用在“华北地区自然植物群落资源综合考察”中获得的大量胡桃楸样方数据, 对华北地区胡桃楸种群分布规律及其影响因素进行分析, 并综合谱系发育和功能性状探讨胡桃楸林的物种共存过程, 为胡桃楸林及其生境的保护提供科学依据, 也为进一步探究华北地区胡桃楸林物种组成及群落发展方向奠定基础. ...
坡位对水曲柳及胡桃楸生长的影响
1
2005
... 胡桃楸(
Juglans mandshurica )又名核桃楸, 隶属于胡桃科胡桃属(
Juglans ), 属第三纪孑遗植物, 在我国多分布于东北的东部山地.同时, 胡桃楸的分布区域也在向南延伸, 扩展到我国华北地区, 主要为北京、天津等地的山区, 且颇具规模.目前针对东北地区分布的胡桃楸种群的研究较为丰富全面(
马万里等, 2001 ;
梁淑娟等, 2005 ;
王东娜等, 2011 ;
王东亮, 2014 ;
苏金娟和王晓春, 2017 ), 而华北地区胡桃楸种群的研究相对较少, 且多集中在胡桃楸生理学特性及群落结构等方面(
闫娜等, 2014 ;
高张莹等, 2017 ,
Song et al ., 2017 ).华北地区胡桃楸种群的分布规律、影响其分布的环境因子、胡桃楸林的系统发育结构, 以及群落构建机制都尚未清楚.本文利用在“华北地区自然植物群落资源综合考察”中获得的大量胡桃楸样方数据, 对华北地区胡桃楸种群分布规律及其影响因素进行分析, 并综合谱系发育和功能性状探讨胡桃楸林的物种共存过程, 为胡桃楸林及其生境的保护提供科学依据, 也为进一步探究华北地区胡桃楸林物种组成及群落发展方向奠定基础. ...
生物多样性科学的热点问题
1
2016
... 群落构建机制即群落物种多样性形成及其维持机制, 主要研究植物群落如何构建, 什么驱使植物的多样性及群落功能的变化(
Cavender-Bares et al ., 2009 ).目前对群落构建机制的分析主要基于谱系发育和植物功能性状(
Webb et al ., 2002 ;
Wright, 2004 ;
Kraft et al ., 2008 ;
Swenson, 2011 ;
马克平, 2016 ;
D’Amen, 2017 ), 对群落构建机制的解释包括生态位机制(
Diamond, 1975 )和中性理论(
Hubbell, 2001 ).中性理论认为群落内物种的出生、死亡、迁入、迁出处于一个动态的平衡, 这种动态平衡维持着群落内物种数量的稳定(
Hubbell, 2001 ).生态位机制则认为群落内植物的生长繁殖、迁入迁出源自于群落的生境、资源及生物间相互作用, 生态位机制是由环境过滤和负密度制约(
Bazzaz, 1991 ;
Weiher & Keddy, 1999 )两个过程主导群落构建. ...
生物多样性科学的热点问题
1
2016
... 群落构建机制即群落物种多样性形成及其维持机制, 主要研究植物群落如何构建, 什么驱使植物的多样性及群落功能的变化(
Cavender-Bares et al ., 2009 ).目前对群落构建机制的分析主要基于谱系发育和植物功能性状(
Webb et al ., 2002 ;
Wright, 2004 ;
Kraft et al ., 2008 ;
Swenson, 2011 ;
马克平, 2016 ;
D’Amen, 2017 ), 对群落构建机制的解释包括生态位机制(
Diamond, 1975 )和中性理论(
Hubbell, 2001 ).中性理论认为群落内物种的出生、死亡、迁入、迁出处于一个动态的平衡, 这种动态平衡维持着群落内物种数量的稳定(
Hubbell, 2001 ).生态位机制则认为群落内植物的生长繁殖、迁入迁出源自于群落的生境、资源及生物间相互作用, 生态位机制是由环境过滤和负密度制约(
Bazzaz, 1991 ;
Weiher & Keddy, 1999 )两个过程主导群落构建. ...
长白山地区胡桃楸种群的种子雨和种子库动态
1
2001
... 胡桃楸(
Juglans mandshurica )又名核桃楸, 隶属于胡桃科胡桃属(
Juglans ), 属第三纪孑遗植物, 在我国多分布于东北的东部山地.同时, 胡桃楸的分布区域也在向南延伸, 扩展到我国华北地区, 主要为北京、天津等地的山区, 且颇具规模.目前针对东北地区分布的胡桃楸种群的研究较为丰富全面(
马万里等, 2001 ;
梁淑娟等, 2005 ;
王东娜等, 2011 ;
王东亮, 2014 ;
苏金娟和王晓春, 2017 ), 而华北地区胡桃楸种群的研究相对较少, 且多集中在胡桃楸生理学特性及群落结构等方面(
闫娜等, 2014 ;
高张莹等, 2017 ,
Song et al ., 2017 ).华北地区胡桃楸种群的分布规律、影响其分布的环境因子、胡桃楸林的系统发育结构, 以及群落构建机制都尚未清楚.本文利用在“华北地区自然植物群落资源综合考察”中获得的大量胡桃楸样方数据, 对华北地区胡桃楸种群分布规律及其影响因素进行分析, 并综合谱系发育和功能性状探讨胡桃楸林的物种共存过程, 为胡桃楸林及其生境的保护提供科学依据, 也为进一步探究华北地区胡桃楸林物种组成及群落发展方向奠定基础. ...
长白山地区胡桃楸种群的种子雨和种子库动态
1
2001
... 胡桃楸(
Juglans mandshurica )又名核桃楸, 隶属于胡桃科胡桃属(
Juglans ), 属第三纪孑遗植物, 在我国多分布于东北的东部山地.同时, 胡桃楸的分布区域也在向南延伸, 扩展到我国华北地区, 主要为北京、天津等地的山区, 且颇具规模.目前针对东北地区分布的胡桃楸种群的研究较为丰富全面(
马万里等, 2001 ;
梁淑娟等, 2005 ;
王东娜等, 2011 ;
王东亮, 2014 ;
苏金娟和王晓春, 2017 ), 而华北地区胡桃楸种群的研究相对较少, 且多集中在胡桃楸生理学特性及群落结构等方面(
闫娜等, 2014 ;
高张莹等, 2017 ,
Song et al ., 2017 ).华北地区胡桃楸种群的分布规律、影响其分布的环境因子、胡桃楸林的系统发育结构, 以及群落构建机制都尚未清楚.本文利用在“华北地区自然植物群落资源综合考察”中获得的大量胡桃楸样方数据, 对华北地区胡桃楸种群分布规律及其影响因素进行分析, 并综合谱系发育和功能性状探讨胡桃楸林的物种共存过程, 为胡桃楸林及其生境的保护提供科学依据, 也为进一步探究华北地区胡桃楸林物种组成及群落发展方向奠定基础. ...
珍贵树种核桃楸的生态学问题及培育前景
1
2005
... 胡桃楸种群在我国主要分布在东北和华北地区, 东北地区主要集中在长白山和小兴安岭海拔200- 1 000 m的阔叶林带和针阔混交林带(
马万里等, 2008 ).在东北长白山地区的胡桃楸种群呈现出衰退型的年龄结构, 幼苗和幼树比例较小(
马万里等, 2008 ).从华北地区的胡桃楸的胸径来看, 华北地区的胡桃楸种群年龄较小, 幼苗、幼树较多(胸径集中在11 cm以下).这与胡桃楸林在东北、华北地区所处的生境及森林发展历史的差异联系密切(
赵光仪等, 1991 ;
马万里等, 2005 ;
王建文, 2006 ).胡桃楸属第三纪孑遗植物, 起源古老, 东北长白山和小兴安岭地区是胡桃楸的最适生长区, 相对而言, 华北是我国胡桃楸种群分布的南部边缘地区,
苏金娟和王晓春(2017) 的研究表明胡桃楸的生长与气温呈现负相关关系, 因此在生境, 尤其是生物气候方面, 气温较低的东北地区更适合胡桃楸生长, 其种群生长速度比华北地区胡桃楸种群相对要快.另外, 在森林发展历史方面, 东北地区森林历史悠久, 虽遭砍伐, 但仍保有一定面积的原始森林, 其中不乏大量的高龄树木(
王建文, 2006 ).而华北地区因人口众多, 迫于经济等压力, 在新中国成立前此地区的原始森林均被破坏, 当前所看到的森林是新中国成立后通过植树造林、森林保育等措施培育的人工林和衍生的次生林, 因此华北地区成林历史较短, 其树木年龄相对较小(
王建文, 2006 ). ...
珍贵树种核桃楸的生态学问题及培育前景
1
2005
... 胡桃楸种群在我国主要分布在东北和华北地区, 东北地区主要集中在长白山和小兴安岭海拔200- 1 000 m的阔叶林带和针阔混交林带(
马万里等, 2008 ).在东北长白山地区的胡桃楸种群呈现出衰退型的年龄结构, 幼苗和幼树比例较小(
马万里等, 2008 ).从华北地区的胡桃楸的胸径来看, 华北地区的胡桃楸种群年龄较小, 幼苗、幼树较多(胸径集中在11 cm以下).这与胡桃楸林在东北、华北地区所处的生境及森林发展历史的差异联系密切(
赵光仪等, 1991 ;
马万里等, 2005 ;
王建文, 2006 ).胡桃楸属第三纪孑遗植物, 起源古老, 东北长白山和小兴安岭地区是胡桃楸的最适生长区, 相对而言, 华北是我国胡桃楸种群分布的南部边缘地区,
苏金娟和王晓春(2017) 的研究表明胡桃楸的生长与气温呈现负相关关系, 因此在生境, 尤其是生物气候方面, 气温较低的东北地区更适合胡桃楸生长, 其种群生长速度比华北地区胡桃楸种群相对要快.另外, 在森林发展历史方面, 东北地区森林历史悠久, 虽遭砍伐, 但仍保有一定面积的原始森林, 其中不乏大量的高龄树木(
王建文, 2006 ).而华北地区因人口众多, 迫于经济等压力, 在新中国成立前此地区的原始森林均被破坏, 当前所看到的森林是新中国成立后通过植树造林、森林保育等措施培育的人工林和衍生的次生林, 因此华北地区成林历史较短, 其树木年龄相对较小(
王建文, 2006 ). ...
长白山林区核桃楸种群数量动态变化的研究
2
2008
... 胡桃楸种群在我国主要分布在东北和华北地区, 东北地区主要集中在长白山和小兴安岭海拔200- 1 000 m的阔叶林带和针阔混交林带(
马万里等, 2008 ).在东北长白山地区的胡桃楸种群呈现出衰退型的年龄结构, 幼苗和幼树比例较小(
马万里等, 2008 ).从华北地区的胡桃楸的胸径来看, 华北地区的胡桃楸种群年龄较小, 幼苗、幼树较多(胸径集中在11 cm以下).这与胡桃楸林在东北、华北地区所处的生境及森林发展历史的差异联系密切(
赵光仪等, 1991 ;
马万里等, 2005 ;
王建文, 2006 ).胡桃楸属第三纪孑遗植物, 起源古老, 东北长白山和小兴安岭地区是胡桃楸的最适生长区, 相对而言, 华北是我国胡桃楸种群分布的南部边缘地区,
苏金娟和王晓春(2017) 的研究表明胡桃楸的生长与气温呈现负相关关系, 因此在生境, 尤其是生物气候方面, 气温较低的东北地区更适合胡桃楸生长, 其种群生长速度比华北地区胡桃楸种群相对要快.另外, 在森林发展历史方面, 东北地区森林历史悠久, 虽遭砍伐, 但仍保有一定面积的原始森林, 其中不乏大量的高龄树木(
王建文, 2006 ).而华北地区因人口众多, 迫于经济等压力, 在新中国成立前此地区的原始森林均被破坏, 当前所看到的森林是新中国成立后通过植树造林、森林保育等措施培育的人工林和衍生的次生林, 因此华北地区成林历史较短, 其树木年龄相对较小(
王建文, 2006 ). ...
... ).在东北长白山地区的胡桃楸种群呈现出衰退型的年龄结构, 幼苗和幼树比例较小(
马万里等, 2008 ).从华北地区的胡桃楸的胸径来看, 华北地区的胡桃楸种群年龄较小, 幼苗、幼树较多(胸径集中在11 cm以下).这与胡桃楸林在东北、华北地区所处的生境及森林发展历史的差异联系密切(
赵光仪等, 1991 ;
马万里等, 2005 ;
王建文, 2006 ).胡桃楸属第三纪孑遗植物, 起源古老, 东北长白山和小兴安岭地区是胡桃楸的最适生长区, 相对而言, 华北是我国胡桃楸种群分布的南部边缘地区,
苏金娟和王晓春(2017) 的研究表明胡桃楸的生长与气温呈现负相关关系, 因此在生境, 尤其是生物气候方面, 气温较低的东北地区更适合胡桃楸生长, 其种群生长速度比华北地区胡桃楸种群相对要快.另外, 在森林发展历史方面, 东北地区森林历史悠久, 虽遭砍伐, 但仍保有一定面积的原始森林, 其中不乏大量的高龄树木(
王建文, 2006 ).而华北地区因人口众多, 迫于经济等压力, 在新中国成立前此地区的原始森林均被破坏, 当前所看到的森林是新中国成立后通过植树造林、森林保育等措施培育的人工林和衍生的次生林, 因此华北地区成林历史较短, 其树木年龄相对较小(
王建文, 2006 ). ...
长白山林区核桃楸种群数量动态变化的研究
2
2008
... 胡桃楸种群在我国主要分布在东北和华北地区, 东北地区主要集中在长白山和小兴安岭海拔200- 1 000 m的阔叶林带和针阔混交林带(
马万里等, 2008 ).在东北长白山地区的胡桃楸种群呈现出衰退型的年龄结构, 幼苗和幼树比例较小(
马万里等, 2008 ).从华北地区的胡桃楸的胸径来看, 华北地区的胡桃楸种群年龄较小, 幼苗、幼树较多(胸径集中在11 cm以下).这与胡桃楸林在东北、华北地区所处的生境及森林发展历史的差异联系密切(
赵光仪等, 1991 ;
马万里等, 2005 ;
王建文, 2006 ).胡桃楸属第三纪孑遗植物, 起源古老, 东北长白山和小兴安岭地区是胡桃楸的最适生长区, 相对而言, 华北是我国胡桃楸种群分布的南部边缘地区,
苏金娟和王晓春(2017) 的研究表明胡桃楸的生长与气温呈现负相关关系, 因此在生境, 尤其是生物气候方面, 气温较低的东北地区更适合胡桃楸生长, 其种群生长速度比华北地区胡桃楸种群相对要快.另外, 在森林发展历史方面, 东北地区森林历史悠久, 虽遭砍伐, 但仍保有一定面积的原始森林, 其中不乏大量的高龄树木(
王建文, 2006 ).而华北地区因人口众多, 迫于经济等压力, 在新中国成立前此地区的原始森林均被破坏, 当前所看到的森林是新中国成立后通过植树造林、森林保育等措施培育的人工林和衍生的次生林, 因此华北地区成林历史较短, 其树木年龄相对较小(
王建文, 2006 ). ...
... ).在东北长白山地区的胡桃楸种群呈现出衰退型的年龄结构, 幼苗和幼树比例较小(
马万里等, 2008 ).从华北地区的胡桃楸的胸径来看, 华北地区的胡桃楸种群年龄较小, 幼苗、幼树较多(胸径集中在11 cm以下).这与胡桃楸林在东北、华北地区所处的生境及森林发展历史的差异联系密切(
赵光仪等, 1991 ;
马万里等, 2005 ;
王建文, 2006 ).胡桃楸属第三纪孑遗植物, 起源古老, 东北长白山和小兴安岭地区是胡桃楸的最适生长区, 相对而言, 华北是我国胡桃楸种群分布的南部边缘地区,
苏金娟和王晓春(2017) 的研究表明胡桃楸的生长与气温呈现负相关关系, 因此在生境, 尤其是生物气候方面, 气温较低的东北地区更适合胡桃楸生长, 其种群生长速度比华北地区胡桃楸种群相对要快.另外, 在森林发展历史方面, 东北地区森林历史悠久, 虽遭砍伐, 但仍保有一定面积的原始森林, 其中不乏大量的高龄树木(
王建文, 2006 ).而华北地区因人口众多, 迫于经济等压力, 在新中国成立前此地区的原始森林均被破坏, 当前所看到的森林是新中国成立后通过植树造林、森林保育等措施培育的人工林和衍生的次生林, 因此华北地区成林历史较短, 其树木年龄相对较小(
王建文, 2006 ). ...
The macroevolutionary consequences of ecological differences among species
1
2007
... 随着相关学科领域的快速发展,
Webb等(2002) 提出用植物谱系发育理念来理解群落物种共存过程, 并提出净亲缘指数(
NRI )和最近亲缘指数(
NTI )这两种亲缘关系指数, 以此来计算群落内物种间的系统发育距离, 并在植物功能性状生态位保守和趋同两种情况下讨论群落的构建机制(
Kraft et al ., 2007 ;
Vamosi et al ., 2009 ).若亲缘关系指数大于0, 则物种系统发育距离较小, 群落由亲缘关系较近的物种构成; 若亲缘关系指数小于0, 说明物种系统发育距离较大, 群落由亲缘关系较远的物种构成; 若亲缘关系指数为0, 说明物种在系统发育结构上呈现随机状态(
Webb et al ., 2002 ).若群落物种生态位保守, 环境过滤作用会导致群落谱系结构的聚集, 竞争排斥(负密度制约)作用会导致群落谱系结构的发散; 若群落物种生态位趋同, 环境过滤作用会导致群落谱系结构发散, 负密度制约作用会导致群落谱系结构随机或聚集(
Webb et al ., 2002 ;
Kraft et al ., 2007 ;
Vamosi et al ., 2009 ).在中性作用下, 群落谱系结构均呈随机状态(
McPeek, 2007 ).群落生态学与植物谱系发育概念的结合, 为解释群落的构建提供了新的理念. ...
1
201P
... 本文采用
Blomberg等(2003) 提出的
K 值法, 以植物最大树高作为功能性状, 检测胡桃楸林内的植物系统发育信号.
K 值为实际的
MSE0 /
MSE 相对于其期望值的比值,
MSE0 是系统发育树末端物种性状数据的均方误差,
MSE 是基于系统发育树的方差-协方差矩阵计算的均方误差, 期望值
MSE0 /
MSE 是基于现有系统发育树的拓扑结构和枝长, 并假设性状以布朗模型方式进化计算得出的.此外, 通过置换检验来判断功能性状的系统发育信号是否显著(
p < 0.05), 随机置换次数为999次.在R的“ape”和“picante”程序包(
Kembel et al ., 2010 ;
Paradis & Schliep, 2019 )中完成
K 值的计算.若
K < 1, 物种表现出趋同性进化趋势;
K = 1, 物种随机进化;
K > 1, 物种进化与系统发育关系紧密, 功能性状保守. ...
The PCA index for measuring functional diversity and its application to
Juglans mandshurica communities in the Beijing mountains, China
1
2017
... 胡桃楸(
Juglans mandshurica )又名核桃楸, 隶属于胡桃科胡桃属(
Juglans ), 属第三纪孑遗植物, 在我国多分布于东北的东部山地.同时, 胡桃楸的分布区域也在向南延伸, 扩展到我国华北地区, 主要为北京、天津等地的山区, 且颇具规模.目前针对东北地区分布的胡桃楸种群的研究较为丰富全面(
马万里等, 2001 ;
梁淑娟等, 2005 ;
王东娜等, 2011 ;
王东亮, 2014 ;
苏金娟和王晓春, 2017 ), 而华北地区胡桃楸种群的研究相对较少, 且多集中在胡桃楸生理学特性及群落结构等方面(
闫娜等, 2014 ;
高张莹等, 2017 ,
Song et al ., 2017 ).华北地区胡桃楸种群的分布规律、影响其分布的环境因子、胡桃楸林的系统发育结构, 以及群落构建机制都尚未清楚.本文利用在“华北地区自然植物群落资源综合考察”中获得的大量胡桃楸样方数据, 对华北地区胡桃楸种群分布规律及其影响因素进行分析, 并综合谱系发育和功能性状探讨胡桃楸林的物种共存过程, 为胡桃楸林及其生境的保护提供科学依据, 也为进一步探究华北地区胡桃楸林物种组成及群落发展方向奠定基础. ...
张广才岭北部三大硬阔树木生长-气候关系的时空变异
2
2017
... 胡桃楸(
Juglans mandshurica )又名核桃楸, 隶属于胡桃科胡桃属(
Juglans ), 属第三纪孑遗植物, 在我国多分布于东北的东部山地.同时, 胡桃楸的分布区域也在向南延伸, 扩展到我国华北地区, 主要为北京、天津等地的山区, 且颇具规模.目前针对东北地区分布的胡桃楸种群的研究较为丰富全面(
马万里等, 2001 ;
梁淑娟等, 2005 ;
王东娜等, 2011 ;
王东亮, 2014 ;
苏金娟和王晓春, 2017 ), 而华北地区胡桃楸种群的研究相对较少, 且多集中在胡桃楸生理学特性及群落结构等方面(
闫娜等, 2014 ;
高张莹等, 2017 ,
Song et al ., 2017 ).华北地区胡桃楸种群的分布规律、影响其分布的环境因子、胡桃楸林的系统发育结构, 以及群落构建机制都尚未清楚.本文利用在“华北地区自然植物群落资源综合考察”中获得的大量胡桃楸样方数据, 对华北地区胡桃楸种群分布规律及其影响因素进行分析, 并综合谱系发育和功能性状探讨胡桃楸林的物种共存过程, 为胡桃楸林及其生境的保护提供科学依据, 也为进一步探究华北地区胡桃楸林物种组成及群落发展方向奠定基础. ...
... 胡桃楸种群在我国主要分布在东北和华北地区, 东北地区主要集中在长白山和小兴安岭海拔200- 1 000 m的阔叶林带和针阔混交林带(
马万里等, 2008 ).在东北长白山地区的胡桃楸种群呈现出衰退型的年龄结构, 幼苗和幼树比例较小(
马万里等, 2008 ).从华北地区的胡桃楸的胸径来看, 华北地区的胡桃楸种群年龄较小, 幼苗、幼树较多(胸径集中在11 cm以下).这与胡桃楸林在东北、华北地区所处的生境及森林发展历史的差异联系密切(
赵光仪等, 1991 ;
马万里等, 2005 ;
王建文, 2006 ).胡桃楸属第三纪孑遗植物, 起源古老, 东北长白山和小兴安岭地区是胡桃楸的最适生长区, 相对而言, 华北是我国胡桃楸种群分布的南部边缘地区,
苏金娟和王晓春(2017) 的研究表明胡桃楸的生长与气温呈现负相关关系, 因此在生境, 尤其是生物气候方面, 气温较低的东北地区更适合胡桃楸生长, 其种群生长速度比华北地区胡桃楸种群相对要快.另外, 在森林发展历史方面, 东北地区森林历史悠久, 虽遭砍伐, 但仍保有一定面积的原始森林, 其中不乏大量的高龄树木(
王建文, 2006 ).而华北地区因人口众多, 迫于经济等压力, 在新中国成立前此地区的原始森林均被破坏, 当前所看到的森林是新中国成立后通过植树造林、森林保育等措施培育的人工林和衍生的次生林, 因此华北地区成林历史较短, 其树木年龄相对较小(
王建文, 2006 ). ...
张广才岭北部三大硬阔树木生长-气候关系的时空变异
2
2017
... 胡桃楸(
Juglans mandshurica )又名核桃楸, 隶属于胡桃科胡桃属(
Juglans ), 属第三纪孑遗植物, 在我国多分布于东北的东部山地.同时, 胡桃楸的分布区域也在向南延伸, 扩展到我国华北地区, 主要为北京、天津等地的山区, 且颇具规模.目前针对东北地区分布的胡桃楸种群的研究较为丰富全面(
马万里等, 2001 ;
梁淑娟等, 2005 ;
王东娜等, 2011 ;
王东亮, 2014 ;
苏金娟和王晓春, 2017 ), 而华北地区胡桃楸种群的研究相对较少, 且多集中在胡桃楸生理学特性及群落结构等方面(
闫娜等, 2014 ;
高张莹等, 2017 ,
Song et al ., 2017 ).华北地区胡桃楸种群的分布规律、影响其分布的环境因子、胡桃楸林的系统发育结构, 以及群落构建机制都尚未清楚.本文利用在“华北地区自然植物群落资源综合考察”中获得的大量胡桃楸样方数据, 对华北地区胡桃楸种群分布规律及其影响因素进行分析, 并综合谱系发育和功能性状探讨胡桃楸林的物种共存过程, 为胡桃楸林及其生境的保护提供科学依据, 也为进一步探究华北地区胡桃楸林物种组成及群落发展方向奠定基础. ...
... 胡桃楸种群在我国主要分布在东北和华北地区, 东北地区主要集中在长白山和小兴安岭海拔200- 1 000 m的阔叶林带和针阔混交林带(
马万里等, 2008 ).在东北长白山地区的胡桃楸种群呈现出衰退型的年龄结构, 幼苗和幼树比例较小(
马万里等, 2008 ).从华北地区的胡桃楸的胸径来看, 华北地区的胡桃楸种群年龄较小, 幼苗、幼树较多(胸径集中在11 cm以下).这与胡桃楸林在东北、华北地区所处的生境及森林发展历史的差异联系密切(
赵光仪等, 1991 ;
马万里等, 2005 ;
王建文, 2006 ).胡桃楸属第三纪孑遗植物, 起源古老, 东北长白山和小兴安岭地区是胡桃楸的最适生长区, 相对而言, 华北是我国胡桃楸种群分布的南部边缘地区,
苏金娟和王晓春(2017) 的研究表明胡桃楸的生长与气温呈现负相关关系, 因此在生境, 尤其是生物气候方面, 气温较低的东北地区更适合胡桃楸生长, 其种群生长速度比华北地区胡桃楸种群相对要快.另外, 在森林发展历史方面, 东北地区森林历史悠久, 虽遭砍伐, 但仍保有一定面积的原始森林, 其中不乏大量的高龄树木(
王建文, 2006 ).而华北地区因人口众多, 迫于经济等压力, 在新中国成立前此地区的原始森林均被破坏, 当前所看到的森林是新中国成立后通过植树造林、森林保育等措施培育的人工林和衍生的次生林, 因此华北地区成林历史较短, 其树木年龄相对较小(
王建文, 2006 ). ...
The role of evolutionary processes in producing biodiversity patterns, and the interrelationships between taxonomic, functional and phylogenetic biodiversity
1
2011
... 群落构建机制即群落物种多样性形成及其维持机制, 主要研究植物群落如何构建, 什么驱使植物的多样性及群落功能的变化(
Cavender-Bares et al ., 2009 ).目前对群落构建机制的分析主要基于谱系发育和植物功能性状(
Webb et al ., 2002 ;
Wright, 2004 ;
Kraft et al ., 2008 ;
Swenson, 2011 ;
马克平, 2016 ;
D’Amen, 2017 ), 对群落构建机制的解释包括生态位机制(
Diamond, 1975 )和中性理论(
Hubbell, 2001 ).中性理论认为群落内物种的出生、死亡、迁入、迁出处于一个动态的平衡, 这种动态平衡维持着群落内物种数量的稳定(
Hubbell, 2001 ).生态位机制则认为群落内植物的生长繁殖、迁入迁出源自于群落的生境、资源及生物间相互作用, 生态位机制是由环境过滤和负密度制约(
Bazzaz, 1991 ;
Weiher & Keddy, 1999 )两个过程主导群落构建. ...
基于系统发育的燕山东麓植物群落的构建机制
1
2017
... 本文采用
唐丽丽等(2017) 对燕山东麓植物群落构建机制的研究中提出的高斯核密度估计的方法对亲缘关系指数的正负趋势进行分析.高斯核密度估计方法就是利用高斯核密度估计来绘制出亲缘关系指数的密度分布图, 并估算
NRI 、
NTI 在正、负区域内的面积, 若指数在正值区域面积大于其在负值区域的面积, 则该指数趋近于正值, 反之则该指数 趋近于负值, 进而探究各群落谱系结构聚集、发散还是随机.可视化及面积计算过程在R 3.3.1的“ggplot2”和“zoo”程序包(Achim & Gabor, 2005;
Wickham, 2016 )中完成. ...
基于系统发育的燕山东麓植物群落的构建机制
1
2017
... 本文采用
唐丽丽等(2017) 对燕山东麓植物群落构建机制的研究中提出的高斯核密度估计的方法对亲缘关系指数的正负趋势进行分析.高斯核密度估计方法就是利用高斯核密度估计来绘制出亲缘关系指数的密度分布图, 并估算
NRI 、
NTI 在正、负区域内的面积, 若指数在正值区域面积大于其在负值区域的面积, 则该指数趋近于正值, 反之则该指数 趋近于负值, 进而探究各群落谱系结构聚集、发散还是随机.可视化及面积计算过程在R 3.3.1的“ggplot2”和“zoo”程序包(Achim & Gabor, 2005;
Wickham, 2016 )中完成. ...
Emerging patterns in the comparative analysis of phylogenetic community structure
2
2009
... 随着相关学科领域的快速发展,
Webb等(2002) 提出用植物谱系发育理念来理解群落物种共存过程, 并提出净亲缘指数(
NRI )和最近亲缘指数(
NTI )这两种亲缘关系指数, 以此来计算群落内物种间的系统发育距离, 并在植物功能性状生态位保守和趋同两种情况下讨论群落的构建机制(
Kraft et al ., 2007 ;
Vamosi et al ., 2009 ).若亲缘关系指数大于0, 则物种系统发育距离较小, 群落由亲缘关系较近的物种构成; 若亲缘关系指数小于0, 说明物种系统发育距离较大, 群落由亲缘关系较远的物种构成; 若亲缘关系指数为0, 说明物种在系统发育结构上呈现随机状态(
Webb et al ., 2002 ).若群落物种生态位保守, 环境过滤作用会导致群落谱系结构的聚集, 竞争排斥(负密度制约)作用会导致群落谱系结构的发散; 若群落物种生态位趋同, 环境过滤作用会导致群落谱系结构发散, 负密度制约作用会导致群落谱系结构随机或聚集(
Webb et al ., 2002 ;
Kraft et al ., 2007 ;
Vamosi et al ., 2009 ).在中性作用下, 群落谱系结构均呈随机状态(
McPeek, 2007 ).群落生态学与植物谱系发育概念的结合, 为解释群落的构建提供了新的理念. ...
... ;
Vamosi et al ., 2009 ).在中性作用下, 群落谱系结构均呈随机状态(
McPeek, 2007 ).群落生态学与植物谱系发育概念的结合, 为解释群落的构建提供了新的理念. ...
胡桃楸-落叶松林地种间化感效应的研究
1
2014
... 胡桃楸(
Juglans mandshurica )又名核桃楸, 隶属于胡桃科胡桃属(
Juglans ), 属第三纪孑遗植物, 在我国多分布于东北的东部山地.同时, 胡桃楸的分布区域也在向南延伸, 扩展到我国华北地区, 主要为北京、天津等地的山区, 且颇具规模.目前针对东北地区分布的胡桃楸种群的研究较为丰富全面(
马万里等, 2001 ;
梁淑娟等, 2005 ;
王东娜等, 2011 ;
王东亮, 2014 ;
苏金娟和王晓春, 2017 ), 而华北地区胡桃楸种群的研究相对较少, 且多集中在胡桃楸生理学特性及群落结构等方面(
闫娜等, 2014 ;
高张莹等, 2017 ,
Song et al ., 2017 ).华北地区胡桃楸种群的分布规律、影响其分布的环境因子、胡桃楸林的系统发育结构, 以及群落构建机制都尚未清楚.本文利用在“华北地区自然植物群落资源综合考察”中获得的大量胡桃楸样方数据, 对华北地区胡桃楸种群分布规律及其影响因素进行分析, 并综合谱系发育和功能性状探讨胡桃楸林的物种共存过程, 为胡桃楸林及其生境的保护提供科学依据, 也为进一步探究华北地区胡桃楸林物种组成及群落发展方向奠定基础. ...
胡桃楸-落叶松林地种间化感效应的研究
1
2014
... 胡桃楸(
Juglans mandshurica )又名核桃楸, 隶属于胡桃科胡桃属(
Juglans ), 属第三纪孑遗植物, 在我国多分布于东北的东部山地.同时, 胡桃楸的分布区域也在向南延伸, 扩展到我国华北地区, 主要为北京、天津等地的山区, 且颇具规模.目前针对东北地区分布的胡桃楸种群的研究较为丰富全面(
马万里等, 2001 ;
梁淑娟等, 2005 ;
王东娜等, 2011 ;
王东亮, 2014 ;
苏金娟和王晓春, 2017 ), 而华北地区胡桃楸种群的研究相对较少, 且多集中在胡桃楸生理学特性及群落结构等方面(
闫娜等, 2014 ;
高张莹等, 2017 ,
Song et al ., 2017 ).华北地区胡桃楸种群的分布规律、影响其分布的环境因子、胡桃楸林的系统发育结构, 以及群落构建机制都尚未清楚.本文利用在“华北地区自然植物群落资源综合考察”中获得的大量胡桃楸样方数据, 对华北地区胡桃楸种群分布规律及其影响因素进行分析, 并综合谱系发育和功能性状探讨胡桃楸林的物种共存过程, 为胡桃楸林及其生境的保护提供科学依据, 也为进一步探究华北地区胡桃楸林物种组成及群落发展方向奠定基础. ...
胡桃楸天然种群遗传多样性的ISSR分析
1
2011
... 胡桃楸(
Juglans mandshurica )又名核桃楸, 隶属于胡桃科胡桃属(
Juglans ), 属第三纪孑遗植物, 在我国多分布于东北的东部山地.同时, 胡桃楸的分布区域也在向南延伸, 扩展到我国华北地区, 主要为北京、天津等地的山区, 且颇具规模.目前针对东北地区分布的胡桃楸种群的研究较为丰富全面(
马万里等, 2001 ;
梁淑娟等, 2005 ;
王东娜等, 2011 ;
王东亮, 2014 ;
苏金娟和王晓春, 2017 ), 而华北地区胡桃楸种群的研究相对较少, 且多集中在胡桃楸生理学特性及群落结构等方面(
闫娜等, 2014 ;
高张莹等, 2017 ,
Song et al ., 2017 ).华北地区胡桃楸种群的分布规律、影响其分布的环境因子、胡桃楸林的系统发育结构, 以及群落构建机制都尚未清楚.本文利用在“华北地区自然植物群落资源综合考察”中获得的大量胡桃楸样方数据, 对华北地区胡桃楸种群分布规律及其影响因素进行分析, 并综合谱系发育和功能性状探讨胡桃楸林的物种共存过程, 为胡桃楸林及其生境的保护提供科学依据, 也为进一步探究华北地区胡桃楸林物种组成及群落发展方向奠定基础. ...
胡桃楸天然种群遗传多样性的ISSR分析
1
2011
... 胡桃楸(
Juglans mandshurica )又名核桃楸, 隶属于胡桃科胡桃属(
Juglans ), 属第三纪孑遗植物, 在我国多分布于东北的东部山地.同时, 胡桃楸的分布区域也在向南延伸, 扩展到我国华北地区, 主要为北京、天津等地的山区, 且颇具规模.目前针对东北地区分布的胡桃楸种群的研究较为丰富全面(
马万里等, 2001 ;
梁淑娟等, 2005 ;
王东娜等, 2011 ;
王东亮, 2014 ;
苏金娟和王晓春, 2017 ), 而华北地区胡桃楸种群的研究相对较少, 且多集中在胡桃楸生理学特性及群落结构等方面(
闫娜等, 2014 ;
高张莹等, 2017 ,
Song et al ., 2017 ).华北地区胡桃楸种群的分布规律、影响其分布的环境因子、胡桃楸林的系统发育结构, 以及群落构建机制都尚未清楚.本文利用在“华北地区自然植物群落资源综合考察”中获得的大量胡桃楸样方数据, 对华北地区胡桃楸种群分布规律及其影响因素进行分析, 并综合谱系发育和功能性状探讨胡桃楸林的物种共存过程, 为胡桃楸林及其生境的保护提供科学依据, 也为进一步探究华北地区胡桃楸林物种组成及群落发展方向奠定基础. ...
中国北方地区森林、草原变迁和生态灾害的历史研究
3
2006
... 胡桃楸种群在我国主要分布在东北和华北地区, 东北地区主要集中在长白山和小兴安岭海拔200- 1 000 m的阔叶林带和针阔混交林带(
马万里等, 2008 ).在东北长白山地区的胡桃楸种群呈现出衰退型的年龄结构, 幼苗和幼树比例较小(
马万里等, 2008 ).从华北地区的胡桃楸的胸径来看, 华北地区的胡桃楸种群年龄较小, 幼苗、幼树较多(胸径集中在11 cm以下).这与胡桃楸林在东北、华北地区所处的生境及森林发展历史的差异联系密切(
赵光仪等, 1991 ;
马万里等, 2005 ;
王建文, 2006 ).胡桃楸属第三纪孑遗植物, 起源古老, 东北长白山和小兴安岭地区是胡桃楸的最适生长区, 相对而言, 华北是我国胡桃楸种群分布的南部边缘地区,
苏金娟和王晓春(2017) 的研究表明胡桃楸的生长与气温呈现负相关关系, 因此在生境, 尤其是生物气候方面, 气温较低的东北地区更适合胡桃楸生长, 其种群生长速度比华北地区胡桃楸种群相对要快.另外, 在森林发展历史方面, 东北地区森林历史悠久, 虽遭砍伐, 但仍保有一定面积的原始森林, 其中不乏大量的高龄树木(
王建文, 2006 ).而华北地区因人口众多, 迫于经济等压力, 在新中国成立前此地区的原始森林均被破坏, 当前所看到的森林是新中国成立后通过植树造林、森林保育等措施培育的人工林和衍生的次生林, 因此华北地区成林历史较短, 其树木年龄相对较小(
王建文, 2006 ). ...
... 的研究表明胡桃楸的生长与气温呈现负相关关系, 因此在生境, 尤其是生物气候方面, 气温较低的东北地区更适合胡桃楸生长, 其种群生长速度比华北地区胡桃楸种群相对要快.另外, 在森林发展历史方面, 东北地区森林历史悠久, 虽遭砍伐, 但仍保有一定面积的原始森林, 其中不乏大量的高龄树木(
王建文, 2006 ).而华北地区因人口众多, 迫于经济等压力, 在新中国成立前此地区的原始森林均被破坏, 当前所看到的森林是新中国成立后通过植树造林、森林保育等措施培育的人工林和衍生的次生林, 因此华北地区成林历史较短, 其树木年龄相对较小(
王建文, 2006 ). ...
... ).而华北地区因人口众多, 迫于经济等压力, 在新中国成立前此地区的原始森林均被破坏, 当前所看到的森林是新中国成立后通过植树造林、森林保育等措施培育的人工林和衍生的次生林, 因此华北地区成林历史较短, 其树木年龄相对较小(
王建文, 2006 ). ...
中国北方地区森林、草原变迁和生态灾害的历史研究
3
2006
... 胡桃楸种群在我国主要分布在东北和华北地区, 东北地区主要集中在长白山和小兴安岭海拔200- 1 000 m的阔叶林带和针阔混交林带(
马万里等, 2008 ).在东北长白山地区的胡桃楸种群呈现出衰退型的年龄结构, 幼苗和幼树比例较小(
马万里等, 2008 ).从华北地区的胡桃楸的胸径来看, 华北地区的胡桃楸种群年龄较小, 幼苗、幼树较多(胸径集中在11 cm以下).这与胡桃楸林在东北、华北地区所处的生境及森林发展历史的差异联系密切(
赵光仪等, 1991 ;
马万里等, 2005 ;
王建文, 2006 ).胡桃楸属第三纪孑遗植物, 起源古老, 东北长白山和小兴安岭地区是胡桃楸的最适生长区, 相对而言, 华北是我国胡桃楸种群分布的南部边缘地区,
苏金娟和王晓春(2017) 的研究表明胡桃楸的生长与气温呈现负相关关系, 因此在生境, 尤其是生物气候方面, 气温较低的东北地区更适合胡桃楸生长, 其种群生长速度比华北地区胡桃楸种群相对要快.另外, 在森林发展历史方面, 东北地区森林历史悠久, 虽遭砍伐, 但仍保有一定面积的原始森林, 其中不乏大量的高龄树木(
王建文, 2006 ).而华北地区因人口众多, 迫于经济等压力, 在新中国成立前此地区的原始森林均被破坏, 当前所看到的森林是新中国成立后通过植树造林、森林保育等措施培育的人工林和衍生的次生林, 因此华北地区成林历史较短, 其树木年龄相对较小(
王建文, 2006 ). ...
... 的研究表明胡桃楸的生长与气温呈现负相关关系, 因此在生境, 尤其是生物气候方面, 气温较低的东北地区更适合胡桃楸生长, 其种群生长速度比华北地区胡桃楸种群相对要快.另外, 在森林发展历史方面, 东北地区森林历史悠久, 虽遭砍伐, 但仍保有一定面积的原始森林, 其中不乏大量的高龄树木(
王建文, 2006 ).而华北地区因人口众多, 迫于经济等压力, 在新中国成立前此地区的原始森林均被破坏, 当前所看到的森林是新中国成立后通过植树造林、森林保育等措施培育的人工林和衍生的次生林, 因此华北地区成林历史较短, 其树木年龄相对较小(
王建文, 2006 ). ...
... ).而华北地区因人口众多, 迫于经济等压力, 在新中国成立前此地区的原始森林均被破坏, 当前所看到的森林是新中国成立后通过植树造林、森林保育等措施培育的人工林和衍生的次生林, 因此华北地区成林历史较短, 其树木年龄相对较小(
王建文, 2006 ). ...
Phylocom: Software for the analysis of phylogenetic community structure and trait evolution
1
2008
... 本文参考
Webb等(2002) 的实验方法构建系统发育树.将样方调查中得到的种子植物, 按照科/属/种的格式, 输入到phylomatic平台(
Webb & Dongoghue, 2005 ;
Webb et al ., 2008 ), 得到基于APG III分类系统的具有进化枝长的系统发育树.根据各物种在系统发育树上的枝长和所在样方中的数量, 计算得出各样方的亲缘关系指数(
NRI 和
NTI ).此过程在R 3.3.1的“picante”程序包(
Kembel et al ., 2010 )中进行.因河南的胡桃楸数量较少, 仅在落叶栎林和华山松林中零散分布, 因此并未对河南的胡桃楸数据进行群落构建机制分析. ...
Phylogenies and community ecology
5
2002
... 群落构建机制即群落物种多样性形成及其维持机制, 主要研究植物群落如何构建, 什么驱使植物的多样性及群落功能的变化(
Cavender-Bares et al ., 2009 ).目前对群落构建机制的分析主要基于谱系发育和植物功能性状(
Webb et al ., 2002 ;
Wright, 2004 ;
Kraft et al ., 2008 ;
Swenson, 2011 ;
马克平, 2016 ;
D’Amen, 2017 ), 对群落构建机制的解释包括生态位机制(
Diamond, 1975 )和中性理论(
Hubbell, 2001 ).中性理论认为群落内物种的出生、死亡、迁入、迁出处于一个动态的平衡, 这种动态平衡维持着群落内物种数量的稳定(
Hubbell, 2001 ).生态位机制则认为群落内植物的生长繁殖、迁入迁出源自于群落的生境、资源及生物间相互作用, 生态位机制是由环境过滤和负密度制约(
Bazzaz, 1991 ;
Weiher & Keddy, 1999 )两个过程主导群落构建. ...
... 随着相关学科领域的快速发展,
Webb等(2002) 提出用植物谱系发育理念来理解群落物种共存过程, 并提出净亲缘指数(
NRI )和最近亲缘指数(
NTI )这两种亲缘关系指数, 以此来计算群落内物种间的系统发育距离, 并在植物功能性状生态位保守和趋同两种情况下讨论群落的构建机制(
Kraft et al ., 2007 ;
Vamosi et al ., 2009 ).若亲缘关系指数大于0, 则物种系统发育距离较小, 群落由亲缘关系较近的物种构成; 若亲缘关系指数小于0, 说明物种系统发育距离较大, 群落由亲缘关系较远的物种构成; 若亲缘关系指数为0, 说明物种在系统发育结构上呈现随机状态(
Webb et al ., 2002 ).若群落物种生态位保守, 环境过滤作用会导致群落谱系结构的聚集, 竞争排斥(负密度制约)作用会导致群落谱系结构的发散; 若群落物种生态位趋同, 环境过滤作用会导致群落谱系结构发散, 负密度制约作用会导致群落谱系结构随机或聚集(
Webb et al ., 2002 ;
Kraft et al ., 2007 ;
Vamosi et al ., 2009 ).在中性作用下, 群落谱系结构均呈随机状态(
McPeek, 2007 ).群落生态学与植物谱系发育概念的结合, 为解释群落的构建提供了新的理念. ...
... ).若亲缘关系指数大于0, 则物种系统发育距离较小, 群落由亲缘关系较近的物种构成; 若亲缘关系指数小于0, 说明物种系统发育距离较大, 群落由亲缘关系较远的物种构成; 若亲缘关系指数为0, 说明物种在系统发育结构上呈现随机状态(
Webb et al ., 2002 ).若群落物种生态位保守, 环境过滤作用会导致群落谱系结构的聚集, 竞争排斥(负密度制约)作用会导致群落谱系结构的发散; 若群落物种生态位趋同, 环境过滤作用会导致群落谱系结构发散, 负密度制约作用会导致群落谱系结构随机或聚集(
Webb et al ., 2002 ;
Kraft et al ., 2007 ;
Vamosi et al ., 2009 ).在中性作用下, 群落谱系结构均呈随机状态(
McPeek, 2007 ).群落生态学与植物谱系发育概念的结合, 为解释群落的构建提供了新的理念. ...
... ).若群落物种生态位保守, 环境过滤作用会导致群落谱系结构的聚集, 竞争排斥(负密度制约)作用会导致群落谱系结构的发散; 若群落物种生态位趋同, 环境过滤作用会导致群落谱系结构发散, 负密度制约作用会导致群落谱系结构随机或聚集(
Webb et al ., 2002 ;
Kraft et al ., 2007 ;
Vamosi et al ., 2009 ).在中性作用下, 群落谱系结构均呈随机状态(
McPeek, 2007 ).群落生态学与植物谱系发育概念的结合, 为解释群落的构建提供了新的理念. ...
... 本文参考
Webb等(2002) 的实验方法构建系统发育树.将样方调查中得到的种子植物, 按照科/属/种的格式, 输入到phylomatic平台(
Webb & Dongoghue, 2005 ;
Webb et al ., 2008 ), 得到基于APG III分类系统的具有进化枝长的系统发育树.根据各物种在系统发育树上的枝长和所在样方中的数量, 计算得出各样方的亲缘关系指数(
NRI 和
NTI ).此过程在R 3.3.1的“picante”程序包(
Kembel et al ., 2010 )中进行.因河南的胡桃楸数量较少, 仅在落叶栎林和华山松林中零散分布, 因此并未对河南的胡桃楸数据进行群落构建机制分析. ...
Phylomatic: Tree assembly for applied phylogenetics
1
2005
... 本文参考
Webb等(2002) 的实验方法构建系统发育树.将样方调查中得到的种子植物, 按照科/属/种的格式, 输入到phylomatic平台(
Webb & Dongoghue, 2005 ;
Webb et al ., 2008 ), 得到基于APG III分类系统的具有进化枝长的系统发育树.根据各物种在系统发育树上的枝长和所在样方中的数量, 计算得出各样方的亲缘关系指数(
NRI 和
NTI ).此过程在R 3.3.1的“picante”程序包(
Kembel et al ., 2010 )中进行.因河南的胡桃楸数量较少, 仅在落叶栎林和华山松林中零散分布, 因此并未对河南的胡桃楸数据进行群落构建机制分析. ...
1
1999
... 群落构建机制即群落物种多样性形成及其维持机制, 主要研究植物群落如何构建, 什么驱使植物的多样性及群落功能的变化(
Cavender-Bares et al ., 2009 ).目前对群落构建机制的分析主要基于谱系发育和植物功能性状(
Webb et al ., 2002 ;
Wright, 2004 ;
Kraft et al ., 2008 ;
Swenson, 2011 ;
马克平, 2016 ;
D’Amen, 2017 ), 对群落构建机制的解释包括生态位机制(
Diamond, 1975 )和中性理论(
Hubbell, 2001 ).中性理论认为群落内物种的出生、死亡、迁入、迁出处于一个动态的平衡, 这种动态平衡维持着群落内物种数量的稳定(
Hubbell, 2001 ).生态位机制则认为群落内植物的生长繁殖、迁入迁出源自于群落的生境、资源及生物间相互作用, 生态位机制是由环境过滤和负密度制约(
Bazzaz, 1991 ;
Weiher & Keddy, 1999 )两个过程主导群落构建. ...
ggplot2: Elegant Graphics for Data Analysis. 1
2016
... 本文采用
唐丽丽等(2017) 对燕山东麓植物群落构建机制的研究中提出的高斯核密度估计的方法对亲缘关系指数的正负趋势进行分析.高斯核密度估计方法就是利用高斯核密度估计来绘制出亲缘关系指数的密度分布图, 并估算
NRI 、
NTI 在正、负区域内的面积, 若指数在正值区域面积大于其在负值区域的面积, 则该指数趋近于正值, 反之则该指数 趋近于负值, 进而探究各群落谱系结构聚集、发散还是随机.可视化及面积计算过程在R 3.3.1的“ggplot2”和“zoo”程序包(Achim & Gabor, 2005;
Wickham, 2016 )中完成. ...
The worldwide leaf economics spectrum
1
2004
... 群落构建机制即群落物种多样性形成及其维持机制, 主要研究植物群落如何构建, 什么驱使植物的多样性及群落功能的变化(
Cavender-Bares et al ., 2009 ).目前对群落构建机制的分析主要基于谱系发育和植物功能性状(
Webb et al ., 2002 ;
Wright, 2004 ;
Kraft et al ., 2008 ;
Swenson, 2011 ;
马克平, 2016 ;
D’Amen, 2017 ), 对群落构建机制的解释包括生态位机制(
Diamond, 1975 )和中性理论(
Hubbell, 2001 ).中性理论认为群落内物种的出生、死亡、迁入、迁出处于一个动态的平衡, 这种动态平衡维持着群落内物种数量的稳定(
Hubbell, 2001 ).生态位机制则认为群落内植物的生长繁殖、迁入迁出源自于群落的生境、资源及生物间相互作用, 生态位机制是由环境过滤和负密度制约(
Bazzaz, 1991 ;
Weiher & Keddy, 1999 )两个过程主导群落构建. ...
Plant diversity in tropical forests: A review of mechanisms of species coexistence
1
2002
... 另外, 天津部分胡桃楸林物种共存过程由环境选择作用主导, 北京也有部分群落物种共存由负密度制约作用主导.其原因在于, 首先天津也有部分胸径较大的胡桃楸林存在, 很有可能随着幼苗的长大, 母体成树对其的限制作用在减弱(
Connell et al ., 1984 ;
Condit et al ., 2000 ;
Wright, 2002 ), 负密度制约作用也随之降低; 其次天津地区部分胡桃楸林的人为干扰程度较强, 且处在坡度较陡的山体中上部, 生存环境较为恶劣, 因此环境选择作用主导这部分群落中的物种共存过程.北京的胡桃楸林中也有部分是胸径为1-3 cm的幼苗, 且北京地区部分胡桃楸林所处环境海拔相对较低, 人为干扰较少, 在这部分胡桃楸林中环境可能不是其主要限制因子, 而是负密度制约作用驱动着物种共存. ...
雾灵山自然保护区胡桃楸种群结构特征分析
1
2014
... 胡桃楸(
Juglans mandshurica )又名核桃楸, 隶属于胡桃科胡桃属(
Juglans ), 属第三纪孑遗植物, 在我国多分布于东北的东部山地.同时, 胡桃楸的分布区域也在向南延伸, 扩展到我国华北地区, 主要为北京、天津等地的山区, 且颇具规模.目前针对东北地区分布的胡桃楸种群的研究较为丰富全面(
马万里等, 2001 ;
梁淑娟等, 2005 ;
王东娜等, 2011 ;
王东亮, 2014 ;
苏金娟和王晓春, 2017 ), 而华北地区胡桃楸种群的研究相对较少, 且多集中在胡桃楸生理学特性及群落结构等方面(
闫娜等, 2014 ;
高张莹等, 2017 ,
Song et al ., 2017 ).华北地区胡桃楸种群的分布规律、影响其分布的环境因子、胡桃楸林的系统发育结构, 以及群落构建机制都尚未清楚.本文利用在“华北地区自然植物群落资源综合考察”中获得的大量胡桃楸样方数据, 对华北地区胡桃楸种群分布规律及其影响因素进行分析, 并综合谱系发育和功能性状探讨胡桃楸林的物种共存过程, 为胡桃楸林及其生境的保护提供科学依据, 也为进一步探究华北地区胡桃楸林物种组成及群落发展方向奠定基础. ...
雾灵山自然保护区胡桃楸种群结构特征分析
1
2014
... 胡桃楸(
Juglans mandshurica )又名核桃楸, 隶属于胡桃科胡桃属(
Juglans ), 属第三纪孑遗植物, 在我国多分布于东北的东部山地.同时, 胡桃楸的分布区域也在向南延伸, 扩展到我国华北地区, 主要为北京、天津等地的山区, 且颇具规模.目前针对东北地区分布的胡桃楸种群的研究较为丰富全面(
马万里等, 2001 ;
梁淑娟等, 2005 ;
王东娜等, 2011 ;
王东亮, 2014 ;
苏金娟和王晓春, 2017 ), 而华北地区胡桃楸种群的研究相对较少, 且多集中在胡桃楸生理学特性及群落结构等方面(
闫娜等, 2014 ;
高张莹等, 2017 ,
Song et al ., 2017 ).华北地区胡桃楸种群的分布规律、影响其分布的环境因子、胡桃楸林的系统发育结构, 以及群落构建机制都尚未清楚.本文利用在“华北地区自然植物群落资源综合考察”中获得的大量胡桃楸样方数据, 对华北地区胡桃楸种群分布规律及其影响因素进行分析, 并综合谱系发育和功能性状探讨胡桃楸林的物种共存过程, 为胡桃楸林及其生境的保护提供科学依据, 也为进一步探究华北地区胡桃楸林物种组成及群落发展方向奠定基础. ...
山西灵空山小蛇沟林下草本层植物群落梯度分析及环境解释
1
2013
... 根据本研究结果, 华北地区的胡桃楸种群主要分布在燕山山脉海拔较低, 且人为干扰相对较小的山体中下部缓坡区域.海拔、坡度、坡位及人为干扰是影响华北地区胡桃楸种群分布的主要因子.CCA分析结果中环境因子前两轴的方差贡献率为59.52%, 说明还存在其他影响因子, 如小气候和土壤等都是影响植物分布的重要因素(
Condit et al ., 2013 ;
Jones, 2013 ).在华北地区森林群落和环境因子关系的研究中曾发现土壤的相关指标是影响森林群落分布的重要因素(
余敏等, 2013 ;
白晓航等, 2017 ).小气候和土壤因子对华北地区胡桃楸种群分布的影响有待进一步研究. ...
山西灵空山小蛇沟林下草本层植物群落梯度分析及环境解释
1
2013
... 根据本研究结果, 华北地区的胡桃楸种群主要分布在燕山山脉海拔较低, 且人为干扰相对较小的山体中下部缓坡区域.海拔、坡度、坡位及人为干扰是影响华北地区胡桃楸种群分布的主要因子.CCA分析结果中环境因子前两轴的方差贡献率为59.52%, 说明还存在其他影响因子, 如小气候和土壤等都是影响植物分布的重要因素(
Condit et al ., 2013 ;
Jones, 2013 ).在华北地区森林群落和环境因子关系的研究中曾发现土壤的相关指标是影响森林群落分布的重要因素(
余敏等, 2013 ;
白晓航等, 2017 ).小气候和土壤因子对华北地区胡桃楸种群分布的影响有待进一步研究. ...
Zoo: S3 infrastructure for regular and irregular time series
2005
黄波罗、胡桃楸、水曲柳分布北限论析
1
1991
... 胡桃楸种群在我国主要分布在东北和华北地区, 东北地区主要集中在长白山和小兴安岭海拔200- 1 000 m的阔叶林带和针阔混交林带(
马万里等, 2008 ).在东北长白山地区的胡桃楸种群呈现出衰退型的年龄结构, 幼苗和幼树比例较小(
马万里等, 2008 ).从华北地区的胡桃楸的胸径来看, 华北地区的胡桃楸种群年龄较小, 幼苗、幼树较多(胸径集中在11 cm以下).这与胡桃楸林在东北、华北地区所处的生境及森林发展历史的差异联系密切(
赵光仪等, 1991 ;
马万里等, 2005 ;
王建文, 2006 ).胡桃楸属第三纪孑遗植物, 起源古老, 东北长白山和小兴安岭地区是胡桃楸的最适生长区, 相对而言, 华北是我国胡桃楸种群分布的南部边缘地区,
苏金娟和王晓春(2017) 的研究表明胡桃楸的生长与气温呈现负相关关系, 因此在生境, 尤其是生物气候方面, 气温较低的东北地区更适合胡桃楸生长, 其种群生长速度比华北地区胡桃楸种群相对要快.另外, 在森林发展历史方面, 东北地区森林历史悠久, 虽遭砍伐, 但仍保有一定面积的原始森林, 其中不乏大量的高龄树木(
王建文, 2006 ).而华北地区因人口众多, 迫于经济等压力, 在新中国成立前此地区的原始森林均被破坏, 当前所看到的森林是新中国成立后通过植树造林、森林保育等措施培育的人工林和衍生的次生林, 因此华北地区成林历史较短, 其树木年龄相对较小(
王建文, 2006 ). ...
黄波罗、胡桃楸、水曲柳分布北限论析
1
1991
... 胡桃楸种群在我国主要分布在东北和华北地区, 东北地区主要集中在长白山和小兴安岭海拔200- 1 000 m的阔叶林带和针阔混交林带(
马万里等, 2008 ).在东北长白山地区的胡桃楸种群呈现出衰退型的年龄结构, 幼苗和幼树比例较小(
马万里等, 2008 ).从华北地区的胡桃楸的胸径来看, 华北地区的胡桃楸种群年龄较小, 幼苗、幼树较多(胸径集中在11 cm以下).这与胡桃楸林在东北、华北地区所处的生境及森林发展历史的差异联系密切(
赵光仪等, 1991 ;
马万里等, 2005 ;
王建文, 2006 ).胡桃楸属第三纪孑遗植物, 起源古老, 东北长白山和小兴安岭地区是胡桃楸的最适生长区, 相对而言, 华北是我国胡桃楸种群分布的南部边缘地区,
苏金娟和王晓春(2017) 的研究表明胡桃楸的生长与气温呈现负相关关系, 因此在生境, 尤其是生物气候方面, 气温较低的东北地区更适合胡桃楸生长, 其种群生长速度比华北地区胡桃楸种群相对要快.另外, 在森林发展历史方面, 东北地区森林历史悠久, 虽遭砍伐, 但仍保有一定面积的原始森林, 其中不乏大量的高龄树木(
王建文, 2006 ).而华北地区因人口众多, 迫于经济等压力, 在新中国成立前此地区的原始森林均被破坏, 当前所看到的森林是新中国成立后通过植树造林、森林保育等措施培育的人工林和衍生的次生林, 因此华北地区成林历史较短, 其树木年龄相对较小(
王建文, 2006 ). ...
植物群落物种共存机制: 负密度制约假说
2
2009
... 本研究中华北地区各分布区域的胡桃楸物种共存过程有所不同, 这与各区域内胡桃楸林所处的环境及其自身生长阶段有一定的关系.河北、陕西、天津地区胡桃楸多分布于山体中部、人为干扰程度较小、坡度较缓、海拔较低的生境, 这3个地区的生境适宜植物生长, 环境条件并不是限制物种生存的最大因素, 因此环境选择作用并不是胡桃楸林物种共存的主要驱动因子; 而且这3个地区胡桃楸种群密度较大, 且胸径在1-3 cm的胡桃楸幼苗极多, 植物群落中种子和幼苗的生长易受其母体成树的限制(
Hubbell, 1980 ;
Connell et al ., 1984 ;
Gilbert et al ., 1994 ;
Hyatt et al ., 2003 ;
祝燕等, 2009 ), 因此在这3个地区胡桃楸林的物种共存中起主要驱动作用的是负密度制约作用.相对而言, 北京、山西胡桃楸所处生境的山体坡度、坡位、人为干扰的梯度较大, 生境异质性高, 环境在植物生长共存过程中发挥更大作用.而且北京、山西的胡桃楸林多为胸径3-7 cm的幼树, 在此生长阶段, 母体成树对其的限制作用会有所减弱(
祝燕等, 2009 ).环境条件和自身生长阶段决定了北京、山西的胡桃楸林物种共存过程主要受环境选择作用驱动. ...
... ), 因此在这3个地区胡桃楸林的物种共存中起主要驱动作用的是负密度制约作用.相对而言, 北京、山西胡桃楸所处生境的山体坡度、坡位、人为干扰的梯度较大, 生境异质性高, 环境在植物生长共存过程中发挥更大作用.而且北京、山西的胡桃楸林多为胸径3-7 cm的幼树, 在此生长阶段, 母体成树对其的限制作用会有所减弱(
祝燕等, 2009 ).环境条件和自身生长阶段决定了北京、山西的胡桃楸林物种共存过程主要受环境选择作用驱动. ...
植物群落物种共存机制: 负密度制约假说
2
2009
... 本研究中华北地区各分布区域的胡桃楸物种共存过程有所不同, 这与各区域内胡桃楸林所处的环境及其自身生长阶段有一定的关系.河北、陕西、天津地区胡桃楸多分布于山体中部、人为干扰程度较小、坡度较缓、海拔较低的生境, 这3个地区的生境适宜植物生长, 环境条件并不是限制物种生存的最大因素, 因此环境选择作用并不是胡桃楸林物种共存的主要驱动因子; 而且这3个地区胡桃楸种群密度较大, 且胸径在1-3 cm的胡桃楸幼苗极多, 植物群落中种子和幼苗的生长易受其母体成树的限制(
Hubbell, 1980 ;
Connell et al ., 1984 ;
Gilbert et al ., 1994 ;
Hyatt et al ., 2003 ;
祝燕等, 2009 ), 因此在这3个地区胡桃楸林的物种共存中起主要驱动作用的是负密度制约作用.相对而言, 北京、山西胡桃楸所处生境的山体坡度、坡位、人为干扰的梯度较大, 生境异质性高, 环境在植物生长共存过程中发挥更大作用.而且北京、山西的胡桃楸林多为胸径3-7 cm的幼树, 在此生长阶段, 母体成树对其的限制作用会有所减弱(
祝燕等, 2009 ).环境条件和自身生长阶段决定了北京、山西的胡桃楸林物种共存过程主要受环境选择作用驱动. ...
... ), 因此在这3个地区胡桃楸林的物种共存中起主要驱动作用的是负密度制约作用.相对而言, 北京、山西胡桃楸所处生境的山体坡度、坡位、人为干扰的梯度较大, 生境异质性高, 环境在植物生长共存过程中发挥更大作用.而且北京、山西的胡桃楸林多为胸径3-7 cm的幼树, 在此生长阶段, 母体成树对其的限制作用会有所减弱(
祝燕等, 2009 ).环境条件和自身生长阶段决定了北京、山西的胡桃楸林物种共存过程主要受环境选择作用驱动. ...
 ,1,*
,1,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT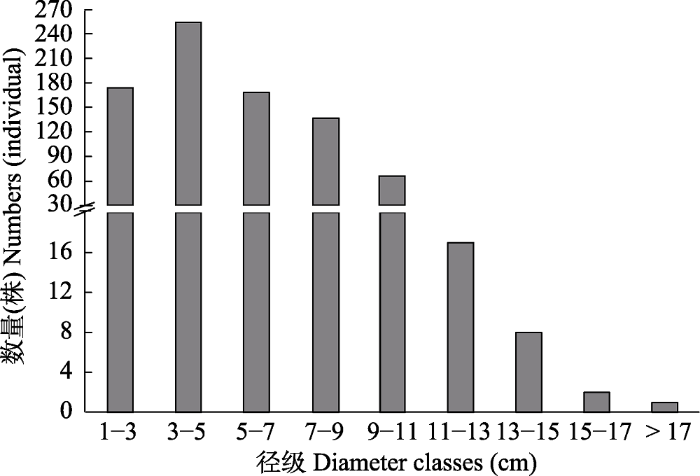 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT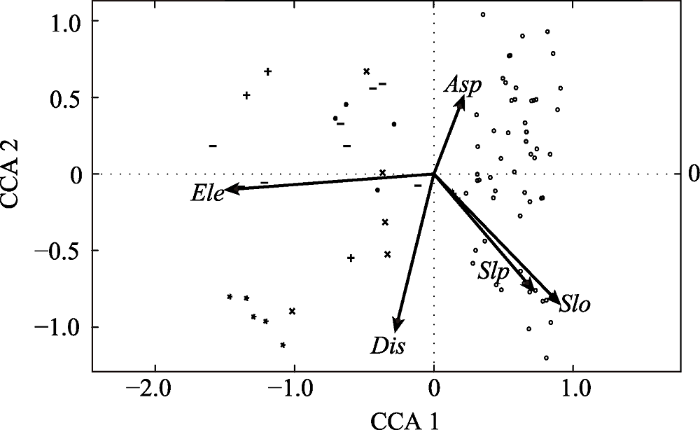 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT , 河南; +, 山西; -, 陕西。
, 河南; +, 山西; -, 陕西。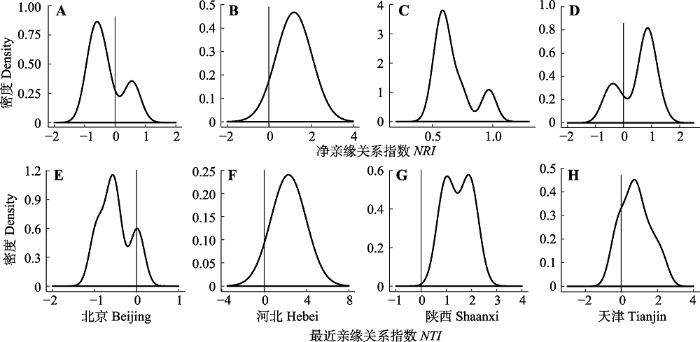 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}