秦浩
1 , 2 , 张殷波
3 , 董刚
4 , 张峰
, 2 , * 1 山西财经大学统计学院, 太原 030006 2 山西大学黄土高原研究所, 太原 030006 3 山西大学环境与资源学院, 太原 030006 4 山西大学生命科学学院, 太原 030006 Altitudinal patterns of taxonomic, phylogenetic and functional diversity of forest communities in Mount Guandi, Shanxi, China Hao QIN
1 , 2 , Yin-Bo ZHANG
3 , Gang DONG
4 , Feng ZHANG
, 2 , * 1 School of Statistics, Shanxi University of Finance and Economics, Taiyuan 030006, China 2 Institute of Loess Plateau, Shanxi University, Taiyuan 030006, China 3 School of Environmental and Resource Sciences, Shanxi University, Taiyuan 030006, China 4 School of Life Science, Shanxi University, Taiyuan 030006, China 通讯作者: * fzhang@sxu.edu.cn 编委: 唐志尧
责任编辑: 李敏, 赵航(实习)
收稿日期: 2018-04-16
接受日期: 2018-09-6
网络出版日期: 2019-09-20
基金资助: 国家科技基础性工作专项 (2011FY110300 )国家科技基础性工作专项 (2015FY110300 )
Corresponding authors: * fzhang@sxu.edu.cn Received: 2018-04-16
Accepted: 2018-09-6
Online: 2019-09-20
Fund supported: Supported by the National Basic Work of Science and Technology of China (2011FY110300 )Supported by the National Basic Work of Science and Technology of China (2015FY110300 )
摘要 探索和揭示生物多样性的空间格局和维持机制是生态学和生物地理学研究的热点内容, 但综合物种、系统进化和功能属性等方面的多样性海拔格局研究很少。该文以关帝山森林群落为研究对象, 综合物种、谱系和功能α和β多样性指数, 旨在初步探讨关帝山森林群落多样性海拔格局及其维持机制。研究结果表明: 随着海拔的升高(1 409-2 150 m), 关帝山森林群落物种丰富度指数(
S )、谱系多样性指数(
PD )和功能丰富度指数(
FRic )整体上表现出上升的趋势, 特别是海拔1 800 m以上区域。随着海拔的升高, 总β多样性(
β total )和更替(
β repl )上升趋势明显, 而丰富度差异(
β rich )则逐渐下降。不同生活型植物的物种、谱系和功能多样性海拔格局差异较大。随着海拔的升高, 草本植物
S 和物种多样性指数(
H′ )上升趋势高于木本植物。影响草本植物
S 分布的主要因素是地形因子, 而影响木本植物
S 分布的主要因素是历史过程。随着海拔的升高, 木本植物
β total 上升趋势要比草本植物明显。随着海拔的升高, 木本植物
β repl 和
β rich 分别表现出单峰格局和“U”形格局, 而草本植物
β repl 和
β rich 则分别表现出单调递增和单调递减的格局。随着环境差异和地理距离的增加, 群落间物种、谱系和功能β多样性显著增加。环境差异(环境过滤)对木本植物的β多样性具有相对较强的作用; 而环境差异(环境过滤)和地理距离(扩散限制)共同作用于草本植物的β多样性。
关键词: 物种多样性 ;
谱系多样性 ;
功能多样性 ;
海拔格局 ;
森林群落 ;
关帝山 Abstract Aims Understanding the spatial patterns and maintenance mechanisms of biodiversity is one of the central issues in ecology and biogeography. Specifically, altitudinal patterns of biodiversity have been widely explored to represent to mimic the latitudinal patterns. However, previous studies on altitudinal patterns of plant diversity have focused mainly on the taxonomic diversity, with less attention paid to the comprehensive information of taxonomic, phylogenetic evolution and functional traits. In this study, we explored maintenance mechanisms of diversity of forest communities in Mount Guandi, by comparing the altitudinal patterns of taxonomic, phylogenetic and functional diversity, based on the systematic investigation of forest communities.
Methods 52 forest plots, each with an area of 30 m × 20 m and divided into six subplots (10 m × 10 m), were investigated along the altitudinal gradient (1 409-2 150 m) in Mount Guandi, Shanxi. Name, diameter at breast height (
DBH ) and height were identified and measured for each tree stem with
DBH ≥3 cm at each plot; name, average height, coverage and basal diameter for each shrub species were identified and measured in two of the six subplots; name, abundance, coverage and average height for each herbaceous species were identified and measured for one quadrat of 1 m × 1 m in each subplot. The indices of taxonomic, phylogenetic and functional α and β diversity were then calculated.
Important findings Species richness (
S ), phylogenetic diversity (
PD ) and functional richness (
FRic ) increased along the altitudinal gradient, especially in altitudes above 1 800 m. Total β diversity (
β total ) and replacement β diversity (
β repl ) increased, while the richness difference (
β rich ) decreased, along the altitudinal gradient. The patterns of taxonomic, phylogenetic and functional diversity were significantly different between woody and herbaceous plants.
S and Shannon-Wiener diversity (
H′ ) of herbaceous plants increased more obvious along the altitude than those of woody plants. Topographic factors regulated the pattern of the herbaceous plant richness, while historical process regulated the woody plant richness. Along the altitude,
β total increased more obvious for the woody plants than for the herbaceous plants.
β repl and
β rich of woody plant showed a unimodal pattern and U-shaped pattern, while those of herbaceous plants increased or decreased, respectively. Taxonomic, phylogenetic and functional β diversity of both woody and herbaceous plants among communities increased with the environmental and geographical distances. Environmental distance had a relatively stronger effects than geographical distance on β diversity of woody plants, while environmental distance and geographical distance jointly influenced β diversity of the herbaceous plants.
Keywords: taxonomic diversity ;
phylogenetic diversity ;
functional diversity ;
altitudinal pattern ;
forest community ;
Mount Guandi PDF (2833KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 秦浩, 张殷波, 董刚, 张峰. 山西关帝山森林群落物种、谱系和功能多样性海拔格局. 植物生态学报, 2019, 43(9): 762-773. DOI:
10.17521/cjpe.2018.0088 QIN Hao, ZHANG Yin-Bo, DONG Gang, ZHANG Feng.
Altitudinal patterns of taxonomic, phylogenetic and functional diversity of forest communities in Mount Guandi, Shanxi, China .
Chinese Journal of Plant Ecology , 2019, 43(9): 762-773. DOI:
10.17521/cjpe.2018.0088 揭示和阐明生物多样性空间格局及其维持机制是生态学研究的热点问题(
Cavender-Bares et al ., 2009 )。由于水热条件变化急剧, 山地一直是生态学家研究多样性海拔格局的理想场所(
Jump et al ., 2009 )。受水热条件、生物类群、微地形、土壤质地和人为干扰等因素的影响, 物种多样性表现出不同的海拔格局, 主要为递减格局和单峰格局(
唐志尧等, 2004 ;
朱珣之和张金屯, 2005 )。Ohsawa (
1991 ,
1995 )研究表明亚洲热带山地的物种多样性随海拔升高而降低, 而温带山地的物种多样性随海拔变化不明显。
朱珣之和张金屯(2005) 总结了我国不同气候带的19座山峰物种多样性海拔格局, 发现低纬度地区山地的物种多样性主要表现出单峰格局, 而高纬度地区山地的物种多样性主要以递减格局为主。
Begon等(1996) 认为物种多样性随海拔增加而递减的格局主要受热量因素影响, 随着海拔升高, 温度的快速下降导致多样性的下降。
Whittaker和Niering(1975) 认为物种多样性的单峰格局受水热条件的综合影响, 低海拔地区限制群落组成和多样性的关键因子主要是降水, 而在高海拔地区主要是温度。
以往多样性海拔格局的研究主要集中于物种多样性, 随着生物信息和功能属性等方面的研究愈加深入, 关于谱系多样性和功能多样性海拔格局的研究越来越多(
Butterfield & Suding, 2013 )。已有大量研究表明谱系和功能多样性随海拔梯度的变化主要表现出单峰格局, 即在中海拔地区出现最大值(
Austrheim, 2002; Watkins Jr et al ., 2006 ), 这主要与水热条件随海拔的变化有关(
Zhang et al ., 2015 )。在环境压力较大的条件下(低海拔的缺水和高海拔的低温), 环境过滤导致亲缘关系较近的或者功能属性相似的物种聚集在一起, 表现出相对较低的谱系和功能多样性(
Kluge & Kessler, 2011 )。另外, 不同生活型的植物对环境响应不同, 导致不同的多样性海拔格局。对于木本植物来说, 受温度的影响较大; 随着海拔的升高, 木本植物多样性逐渐降低(
Pausas & Austin, 2001 )。对于草本植物来说, 受光照、水分和土壤理化性质等多种因素影响, 具有多样的海拔格局(
Qian et al ., 2014 )。
山西关帝山保存了华北地区较为完整的森林群落类型, 但目前关于该地区森林群落谱系和功能多样性海拔格局的研究相对较少。本文基于野外植被调查, 选取物种、谱系和功能α和β多样性指数, 通过方差分解和相关性分析等方法研究了多样性的海拔格局, 分析了多样性与环境因子、群落结构的关系, 探讨了环境差异和地理距离对群落组成的影响, 旨在探讨: (1)物种、谱系和功能多样性沿海拔梯度是如何变化的? (2)环境因子和历史进程是如何影响物种丰富度的? (3)环境差异和地理距离对群落组成(β多样性)的相对作用如何? (4)木本植物和草本植物多样性格局是否相同? 限制因素是否相同? 通过对以上问题的研究, 希望能初步揭示山西关帝山森林群落多样性格局和维持机制, 为关帝山森林保护与管理提供科学依据。
1 材料和方法 1.1 研究区概况 山西关帝山地处吕梁山脉中段, 地理位置: 37.33°-38.33° N, 110.30°-111.30° E。气候属暖温带大陆性季风气候, 年平均气温3.5 ℃, 最冷月(1月)平均气温-10.6 ℃, 最暖月(7月)平均气温16.1 ℃; 年降水量830 mm。区域土壤类型主要包括山地棕壤、山地褐土、亚高山草甸土和河漫滩草甸土等(
张峰等, 2007 ;
秦浩等, 2013 )。
森林群落主要包括云杉林(Form.
Picea spp.)、华北落叶松林(Form.
Larix principis-rupprechtii )、桦木林(Form.
Betula spp.)、山杨林(Form.
Populus davidiana )和油松林(From.
Pinus tabulaeformis )等; 灌丛群落主要包括黄刺玫灌丛(Form.
Rosa xanthina )、沙棘灌丛(Form.
Hippophae rhamnoides subsp.
sinensis )和荆条灌丛(Form.
Vitex negundo var.
heterophylla )等; 草本群落主要包括蒿类草丛(Form.
Artemisia spp.)和薹草草甸(Form.
Carex spp.)等(
赵小娜等, 2014 )。
1.2 野外调查 2011-2013年, 采用典型取样的方法沿海拔梯度在孝文山、云顶山、庞泉沟、双家寨和西葫芦等地共调查52块森林样方, 海拔1 409-2 150 m (
图1 )。本文主要关注关帝山中低海拔段森林群落的多样性海拔格局, 并未涉及灌丛和草地类型。森林样方面积为20 m × 30 m, 将其划分为6个10 m × 10 m的样格, 对样格内胸径(
DBH ) ≥3 cm的乔木进行每木调查, 记录乔木的种名、胸径、高度和健康状况。在样方对角线处选择2个10 m × 10 m的样格进行灌木层调查, 记录灌木的平均基径、平均高度、盖度和丛数。在6个10 m × 10 m的样格中分别布设1个1 m × 1 m的小样方, 进行草本层调查, 记录草本的多度、盖度和平均高度。同时, 以记名方式记录样格外草本植物名称。另外, 记录每个样地的地理信息(经度、纬度、海拔、地形、坡度、坡向和坡位)、干扰程度、土壤类型以及人为活动等情况(
方精云等, 2009 )。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1关帝山森林群落分布图。 Fig. 1Distribution of plots of forest communities in Mount Guandi. 1.3 环境数据 1.3.1 地形数据
选取海拔、坡度、坡向和坡位作为地形变量, 其中将坡向和坡位数据转化为等级制数据。坡向数据以正北方向为0°, 45°夹角为一个等级, 划分为8个坡向等级, 1表示北坡(0°-22.5°, 337.5°-360°), 2表示东北坡(22.5°-67.5°), 3表示西北坡(292.5°-337.5°), 4表示东坡(67.5°-112.5°), 5表示西坡(247.5°-292.5°), 6表示东南坡(112.5°-157.5°), 7表示西南坡(202.5°- 247.5°), 8表示南坡(157.5°-202.5°)(
赵小娜等,2014 ); 坡位共分为5个等级, 即: 1 (下坡), 2 (中下坡), 3 (中坡), 4 (中上坡)和5 (上坡)(
赵小娜等, 2014 )。
1.3.2 群落结构数据
选取森林类型、森林郁闭度、平均树高、平均胸径、密度和干扰程度作为群落结构变量。将其中的属性变量依次转化为等级制数据。如将森林类型按乔木层建群种叶的生活型依次划分为1 (常绿针叶林)、2 (落叶针叶林)和3 (落叶阔叶林) 3个等级; 将干扰程度变量按照干扰强度大小依次转化为1 (无干扰)、2 (轻微干扰)、3 (中度干扰)和4 (重度干扰) 4个等级。
1.4 α多样性指数 1.4.1 物种α多样性指数
分别选取物种丰富度指数(
S )、物种多样性指数(
H′ )和物种均匀度指数(
E )来度量群落的物种多样性。同时, 分别计算木本植物和草本植物的物种α多样性。
1.4.2 谱系α多样性指数
按照被子植物分类系统III (APGIII)整理所有被子植物的科属种信息, 通过Phylomatic软件 (http://www.phylodiversity.net)获得样地所有被子植物的树状结构, 建立相关的系统进化树。另外, 分别建立木本植物和草本植物的系统进化树。
选择Faith’s谱系多样性指数(
PD )和平均谱系距离指数(
MPD )作为度量谱系α多样性指数。
PD 指数被定义为群落内所有物种进化分枝长度之和(
Faith, 1992 )。
MPD 指数是指群落内所有物种对间的平均谱系距离(
Webb et al ., 2008 )。
选取净关联指数(
NRI )来表征群落的谱系结构, 反映群落内物种对间的亲缘关系(
Webb et al ., 2008 )。计算公式如下:
(1) $NRI=-1\times \frac{(MPD_{observed}-MPD_{randomize})}{SRMPD_{randomized}}$ 式中,
MPD observed 是指群落实际观测到的所有物种对间的平均谱系距离,
MPD randomize 和
SRMPD randomized 是指在随机零模型模拟下所产生的谱系距离的平均值和标准误差。
当
NRI > 0时, 群落谱系结构是聚集的, 即亲缘关系较近的物种聚集在一起; 当
NRI < 0时, 群落谱系结构是发散的, 即群落中存在较多亲缘关系远的物种; 而当
NRI 在0上下波动时, 群落中的物种组成是随机的(
Webb et al ., 2008 )。
以上所有多样性指数的计算用Phylocom 4.2软件完成。另外, 分别计算木本植物和草本植物的谱系α多样性和群落谱系结构指数。
1.4.3 功能α多样性指数
选取植物的生长型、生活型、传粉方式、开花时间、花期、果实类型、结果时间、果期、种子传播方式、光合途径、光耐受性和固氮类型12个功能属性, 这些功能属性特征能很好地反映植物的生理和物候等特征(
Zhang et al ., 2015 )。
基于选取的植物功能属性, 应用R 3.4.3版本的“FD”软件包计算功能丰富度指数(
FRic )、功能均匀度指数(
FEve )和功能离散度指数(
FDiv )(
Mason et al ., 2005 ;
Casanoves et al ., 2011 )。另外, 分别计算木本植物和草本植物的功能α多样性指数。
1.5 β多样性指数 基于物种、系统发育和功能属性数据, 采用Jaccard相异性指数分别计算物种(
Tβ )、谱系(
Pβ )和功能(
Fβ ) β多样性指数。同时将总β多样性(
β total )进一步分解为更替(
β repl )和丰富度差异(
β rich )两部分之和(
Cardoso et al ., 2015 )。
β repl 指的是由于物种组成或者系统进化(功能属性)发生一对一替换所导致的群落间差异; 而
β rich 则强调由于物种组成或者系统进化(功能属性)在丰富度上的不同所导致的群落间差异。计算公式如下:
(2) $\beta_{total}=\beta_{repl}+\beta_{rich}$ (3) $\beta_{total}=\frac{b+c}{a+b+c}$ (4) $\beta_{repl}=2 \times \frac{min(b,c)}{a+b+c}$ (5) $\beta_{rich}= \frac{|b-c|}{a+b+c}$ 式中,
a 表示两个群落共有的物种数或者系统进化(功能属性)枝长;
b 和
c 分别表示两个群落各自所特有的物种数或者系统进化(功能属性)枝长。另外, 分别计算木本植物和草本植物的物种、谱系和功能β多样性。物种、谱系和功能β多样性计算在R语言的“BAT”包下完成(
Cardoso et al ., 2015 )。
1.6 统计分析 对所有的环境数据进行主成分分析(PCA), 选取前四个轴(解释总变量的85.5%)作为环境因子变量。同样, 对地形因子和群落结构数据也分别进行了PCA分析, 分别选取前四个轴(解释总变量的100%和93.5%)作为地形因子和群落结构变量。
运用SAM软件对群落多样性的空间自相关进行了检验, 结果表明群落多样性并没有表现出显著的空间自相关(
p > 0.05)。
对于α多样性指数, 首先将物种、谱系和功能α多样性指数与海拔进行回归分析, 研究α多样性的海拔格局。其次, 基于R语言的“Vegan”软件包, 采用方差分解法分析了地形因子、群落结构和历史过程对物种丰富度的相对影响。历史过程选用
MPD 和
NRI 来间接反映,
MPD 和
NRI 指数可以反映群落的系统进化背景和物种间的谱系关系(
Qian et al ., 2016 )。
对于β多样性, 首先将样地按照海拔梯度每隔100 m划分为7个区间(样地分布海拔1 409-2 150 m), 然后对β多样性与海拔进行了回归分析, 研究β多样性的海拔格局。其次, 基于R语言的“ecodist”软件包对环境距离、地理距离与β多样性的关系进行了基于距离矩阵的多元回归分析(MRM)。最后, 基于R语言的“Vegan”软件包, 采用方差分解法分析了环境距离和地理距离对β多样性的相对影响。环境距离和地理距离均采用欧氏距离来度量。
2 研究结果 2.1 物种、谱系和功能α多样性海拔格局 随着海拔的升高(1 409-2 150 m),
S 和
H′ 整体上表现出上升的趋势, 特别是在海拔1 800 m以上时, 这种上升趋势更加明显(
图2A 、2B)。草本植物
S 和
H′ 的上升趋势要高于木本植物。均匀度指数
E 整体上表现出单峰格局, 即随着海拔的升高,
E 先增加后减少, 峰值出现在海拔1 700-1 800 m之间(
图2C )。木本植物和草本植物沿海拔梯度表现出不同的均匀度格局。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2关帝山森林群落物种、谱系和功能α多样性海拔格局。 Fig. 2Altitudinal patterns of taxonomic, phylogenetic and functional α diversity of forest communities in Mount Guandi. 随着海拔的升高, 特别是在海拔1 800 m以上,
PD 整体上表现出明显的上升趋势, 这主要是由于草本植物
PD 的增加所致。随着海拔的升高,
MPD 整体上上升趋势不明显(
图2E )。随着海拔的升高,
NRI 整体上是逐渐下降的, 表明群落谱系结构由聚集向随机或者发散过渡。木本植物的
NRI 大多为正值, 而草本植物的
NRI 大多为负值。
FRic 与
S 和
PD 表现出较为一致的海拔格局, 即随着海拔的升高,
FRic 逐渐升高, 特别是草本植物(
图2F )。
FEve 整体上表现出与
E 类似的海拔格局, 木本植物表现出单峰格局, 而草本植物呈现逐渐上升的趋势(
图2H )。
2.2 地形因子、群落结构和历史进程对物种丰富度的影响 方差分解结果表明: 地形因子、群落结构和历史过程对群落整体的物种丰富度
S 解释力为60.58% (
图3A ), 其中地形因子独立解释力最高(21.37%); 其次为地形因子和历史过程共有解释力(12.08%)、群落结构(11.54%), 历史过程独立解释了5.36%。地形因子、群落结构和历史过程对木本植物
S 总的解释力为42.11% (
图3B ), 其中历史过程独立解释力最高(14.6%), 其次为地形因子(8.8%), 因子之间存在一定的共有解释力。地形因子、群落结构和历史过程对草本植物
S 总的解释力为50.40% (
图3C ), 其中地形因子独立解释了绝大部分(31.28%), 其次为群落结构(8.07%), 历史过程独立解释力较低, 仅有1.19%。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3地形因子、群落结构和历史过程对关帝山森林群落物种丰富度的解释。 Fig. 3Variance in species richness of forest communities explained by topography, community structure and historical process factors in Mount Guandi. 2.3 物种、谱系和功能β多样性海拔格局 物种、谱系和功能β多样性表现出较为一致的海拔格局(
图4 ), 本文以物种β多样性海拔格局为例阐述研究结果。
图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4关帝山森林群落所有植物(A、D、G)、木本植物(B、E、H)和草本植物(C、F、I)物种、谱系和功能β多样性海拔格局。 Fig. 4Altitudinal patterns of taxonomic (Tβ ), phylogenetic (Pβ ) and functional (Fβ ) β diversity for all plants (A, D, G), woody plants (B, E, H) and herbaceous plants (C, F, I) of forest communities in Mount Guandi. 整体上来说, 随着海拔的升高,
β total 和
β repl 表现出上升的趋势, 而
β rich 表现出下降的趋势(
图4A 、4D、4G)。在低海拔(1 400-1 500 m)地区,
β repl 和
β rich 共同影响了
β total , 分别解释
β total 的62.84%和37.16%。随着海拔的升高, 群落间物种组成差异主要受物种更替的影响,
β repl 占的比重越来越大; 在高海拔(海拔> 2 000 m)地区,
β repl 占
β total 的88.62%, 仅有11.38%由
β rich 解释。
对于木本植物而言, 随着海拔的升高,
β total 逐渐增加,
β repl 和
β rich 分别表现出单峰格局和U型格局, 即
β repl 先上升后下降, 而
β rich 先下降后上升(
图4B 、4E、4H)。在低海拔(1 400-1 500 m)地区, 群落间
β rich 相对较高, 占
β total 的78.54%。海拔1 500-1 600 m之间,
β repl 和β
rich 共同影响了
β total 。随着海拔继续升高, 在海拔1 700-1 900 m之间,
β repl 达到最大, 然后开始下降,
β rich 达到最小值后开始上升。
对于草本植物而言, 随着海拔的升高,
β total 上升趋势不明显(
图4C 、4F、4I)。在海拔低于1 800 m的区间,
β repl 和
β rich 随海拔的变化不明显, 两者共同影响着
β total 。在海拔高于1 800 m的区间,
β repl 上升趋势明显, 而
β rich 下降趋势明显。整体上
β repl 和
β rich 分别表现出单调递增和单调递减的格局。海拔2 000 m以上的区域,
β repl 对
β total 起主要作用, 占
β total 的78.01%。
2.4 环境距离和地理距离对β多样性的影响 随着环境距离和地理距离的增加, 关帝山森林群落间物种、谱系和功能β多样性均增加显著(
图5 ,
图6 ;
p < 0.001), 即群落间物种的组成、亲缘关系和功能属性差异性增加。对所有植物来说(
图5 ,
图6 A、6D、6G), 群落间物种、谱系和功能β多样性与环境距离的线性关系要强于地理距离。
图5 新窗口打开 |
下载原图ZIP |
生成PPT 图5关帝山森林群落所有植物(A、D、G)、木本植物(B、E、H)和草本植物(C、F、I)物种、谱系和功能β多样性随环境距离的变化趋势。 Fig. 5Variance in taxonomic (Tβ ), phylogenetic (Pβ ) and functional (Fβ ) β diversity along the environmental distance for all plants (A, D, G), woody plants (B, E, H) and herbaceous plants (C, F, I) of forest communities in Mount Guandi. 图6 新窗口打开 |
下载原图ZIP |
生成PPT 图6关帝山森林群落所有植物(A、D、G)、木本植物(B、E、H)和草本植物(C、F、I)物种、谱系和功能β多样性随地理距离的变化趋势。 Fig. 6Variance in taxonomic (Tβ ), phylogenetic (Pβ ) and functional (Fβ ) β diversity along the geographical distance for all plants (A, D, G), woody plants (B, E, H) and herbaceous plants (C, F, I) of forest communities in Mount Guandi. 对所有植物来说, 方差分解结果表明: 环境距离对整体的β多样性影响较为明显, 地理距离的影响相对较小(
图7 )。环境距离独立对物种、谱系和功能β多样性的解释力分别为16.42%、12.09%和15.78%; 地理距离独立解释分别为6.20%、5.07%和9.05%; 环境距离和地理距离的共有解释力分别为6.23%、4.05%和7.24%。
图7 新窗口打开 |
下载原图ZIP |
生成PPT 图7环境距离和地理距离对关帝山森林群落物种(Tβ )、谱系(Pβ )和功能(Fβ ) β多样性的解释。AS, 所有物种; HS, 草本植物; WS, 木本植物。 Fig. 7Variance in taxonomic (Tβ ), phylogenetic (Pβ ) and functional (Fβ ) β diversity of forest communities explained by environmental distance and geographical distance in Mount Guandi. AS, all species; HS, herbaceous species; WS, woody species. 对木本植物而言, 环境距离对物种、谱系和功能β多样性的影响起决定性作用, 分别解释了13.84%、8.08%和15.35%。对草本植物来说, 环境距离和地理距离决定了草本植物的β多样性。环境距离独立对物种、谱系和功能β多样性的解释力分别为13.20%、8.21%和11.02%; 地理距离独立解释力分别为9.24%、5.60%和8.13%。
3 讨论 3.1 物种、谱系和功能α多样性海拔格局及其影响因素 随着海拔的升高,
S 、
PD 和
FRic 整体上表现出上升的趋势, 特别是在海拔1 800 m以上(
图2 )。这主要与我们研究的海拔范围(1 409-2 150 m)有关, 在中低海拔区段, 降水是影响该区域多样性最主要的因素(
赵小娜等, 2014 )。随着海拔的升高, 降水量逐渐增加,
S 、
PD 和
FRic 整体上呈现上升的趋势。不同气候带下, 山地α多样性海拔格局的差异主要与气候类型、地形地貌、植被类型和人为活动等因素密切相关, 但水热条件是影响多样性海拔格局的最主要因素(
朱珣之和张金屯, 2005 )。在水分条件较好的区域, 随着海拔升高, 温度快速下降是导致多样性降低的主要原因; 而受水分条件限制的区域, 水热条件的制约和平衡是影响多样性海拔格局的关键。在低海拔区域, 降水是影响群落组成和多样性的关键因子; 而在高海拔区域, 低温成为限制群落组成和多样性的关键因子(
唐志尧等, 2004 ;
Zhang et al ., 2015 )。随着海拔的升高, 群落的
FEve 和
E 指数表现出单峰格局, 最大值大致出现在海拔1 800 m处。该海拔段是温性林向寒温性林逐渐过渡的区域, 群落组成兼有耐旱性和耐寒性物种, 表现出较高的均匀性。随着海拔的升高,
NRI 是逐渐下降的, 群落谱系结构由聚集向随机或者发散过渡。在低海拔区域, 环境过滤(较少的降水量和较强的人为干扰)导致群落内亲缘关系较近的物种聚集在一起, 表现出较强的谱系聚集。而在海拔相对较高的区域, 环境过滤和种间关系可能共同作用于群落组成, 进而表现出随机或者发散的谱系结构。
NRI 指数结果表明群落内木本植物大多为亲缘关系较近的物种(
NRI > 0), 而草本植物大多为亲缘关系较远的物种(
NRI < 0)。
陈廷贵等(2000) 研究表明关帝山物种丰富度表现出不明显的海拔格局, 与本研究结果的差异主要表现在: (1)本研究仅涉及关帝山森林群落, 而陈廷贵等的研究不仅包括森林群落, 还包括灌丛群落和草本群落。(2)研究区海拔范围不同。本研究区域海拔较低(1 409-2 150 m), 该区域降水是多样性最主要的影响因素;
陈廷贵等(2000) 的研究区海拔较高(1 700-2 700 m), 涉及水热限制因子的转变, 因此多样性格局不明显。本研究与
Gao和Zhang (2006) 等在相同海拔段(1 400-2 000 m)的研究具有一致的多样性格局, 降水是影响低海拔地区多样性的重要因素, 随着海拔的升高, 降水的增加有利于多样性增加。本文的不足在于缺乏该区域高海拔地段的森林群落样方, 研究结果无法表明该区域整个海拔范围的森林群落多样性格局, 只是反映了中低海拔段的森林群落多样性格局。在以后的研究中, 我们希望能够补充高海拔地段的森林群落样方, 完善该区域整个海拔梯度上的多样性格局。
3.2 物种、谱系和功能β多样性海拔格局及其影响因素 物种、谱系和功能β多样性表现出较为一致的海拔格局(
图4 )。整体上来说, 随着海拔的升高,
β total 和
β repl 表现出上升的趋势, 而
β rich 表现出下降的趋势(
图4 A、4D、4G), 这与常见的β多样性随海拔单调递减的格局不一致(
Wang et al ., 2002 )。单调递减格局认为低海拔地区较高的温度和干扰, 导致种间竞争加剧, 造成较高的β多样性(
Antonio Vázquez & Givnish, 1998 )。根据Rapoport法则, 高海拔地区由于温度较低往往导致该地区的物种具有较宽的生态位, 分布范围相对较广, 因此相邻海拔区间的群落组成变化相对较小(
Rapoport, 1982 )。本研究涉及的海拔相对较低, 温度不是主要的决定因子。低海拔地区森林群落以油松林和辽东栎林为主, 物种组成大多为菊科喜阳耐旱的物种, 而β多样性较高的地区为落叶阔叶林向寒温性针叶林的过渡带, 物种组成具有较大的变化。
Itow (1991) 研究表明森林过渡带往往具有较高的β多样性。在不同海拔区段,
β repl 和
β rich 的相对作用不同。在低海拔地区, 森林管理、干扰强度和微生境等导致
β repl 和
β rich 共同影响了
β total ; 而在高海拔地区, 由于森林类型的不同导致群落间呈现较高的更替速率。
3.3 环境差异和地理距离对β多样性的影响 随着环境差异和地理距离的增加, 群落间物种、谱系和功能β多样性均增加显著(
p < 0.001)。这与以往的研究是一致的, 即随着环境差异和地理距离的增加, 群落间物种组成、亲缘关系和功能属性差异性增加(
Chi et al ., 2014 )。基于生态位理论的生态位分化假说认为不论群落间地理距离多大, 环境差异越大, 群落组成差异越大(
Fernandez-Going et al ., 2013 )。而基于中性理论的距离衰减假说认为不论群落间环境差异多大, 地理距离越大, 群落组成差异越大(
Qian, 2009 )。我们的研究表明, 对所有植物来说, 群落间物种、谱系和功能β多样性与环境距离的线性关系要强于地理距离(
图5 ,
图6A 、6D、6G)。这表明在较小的地理单元上, 由于海拔梯度上存在较大的环境差异, 导致环境差异(环境过滤)在该区域的群落组成上发挥着重要作用, 而地理距离(扩散限制)的作用相对较小。
另外, 环境差异和地理距离对不同生活型植物的β多样性影响不同。环境差异(环境过滤)对木本植物的β多样性具有相对较强的作用; 而环境差异(环境过滤)和地理距离(扩散限制)共同作用于草本植物的β多样性。
Qian等(2014) 对长白山谱系多样性的海拔格局研究也表明, 环境差异对木本植物β多样性的影响要高于草本植物。环境差异和地理距离对群落β多样性总的解释力整体上不超过30%, 表明还有其他因素(如土壤质地、土壤动物、微生物等)影响了群落的β多样性。
3.4 木本植物和草本植物多样性海拔格局差异 木本植物和草本植物多样性海拔格局表现出较大的差异, 这主要与它们对环境响应和生活史策略不同有关。木本植物受温度影响较大(
Pausas & Austin, 2001 ), 草本植物受光照、水分、微地形、土壤理化性质和人为活动等影响更大, 表现出多样的海拔格局(
Qian et al ., 2014 )。随着海拔的升高, 草本植物
S 和
H′ 的上升趋势要高于木本植物。在关帝山低海拔地区, 降水是影响草本植物多样性格局最主要的因子。随着海拔升高, 降水量逐渐增加, 草本植物多样性明显增加。而对于木本植物来说, 影响其分布的主要因素是历史过程(
MPD 和
NRI )。这表明木本植物在系统进化方面更加保守, 对环境的适应性和生活史策略与草本植物有所差异。
NRI 指数表明关帝山森林群落的木本植物大多表现出谱系聚集的结构, 即亲缘关系较近的物种共存; 而草本植物大多表现出谱系发散的结构, 即亲缘关系较远的物种 共存。
随着海拔的升高, 木本植物
β total 上升趋势要比草本植物明显, 这说明随着海拔升高, 群落间木本植物的组成、亲缘关系和功能属性差异越来越大。在低海拔地区, 油松林是主要的森林类型, 群落间木本植物的差异相对较小。随着海拔的升高, 森林类型逐渐由温性针叶林、落叶阔叶林过渡到寒温性针叶林, 森林类型的转变导致群落间木本植物组成上的显著变化, 造成木本植物较高的
β total , 在这一过程中物种间的更替(
β repl )起主导作用。在高海拔地区, 森林群落以华北落叶松林、云杉林为主, 群落组成相对稳定, 群落间木本植物的更替速率下降, 而由微地形、群落结构和土壤性质等微生境引起的丰富度差异(
β rich )开始增加。对于草本植物来说, 在海拔1 800 m以下, 森林类型以油松林和辽东栎林为主, 群落生境相对一致, 群落间的差异主要是由微生境的差异所致,
β repl 和
β rich 共同影响了
β total 。海拔1 800 m以上地区, 由于森林类型的转变导致林下草本植物组成发生明显的改变, 造成群落间草本植物较高的
β repl 。
[1] Antonio Vázquez GJ Givnish TJ 1998 ). Altitudinal gradients in tropical forest composition, structure, and diversity in the Sierra de ManantlánJournal of Ecology ,999 -1020 . [本文引用: 1] [2] Austrheim G 2002 ). Plant diversity patterns in semi-natural grasslands along an elevational gradient in southern NorwayPlant Ecology ,193 -205 . [本文引用: 1] [3] Begon M Harper JL Townsend CR 1996 ). Ecology: Individuals, Populations and Communities. Blackwell Science, Oxford, UK. [本文引用: 1] [4] Butterfield BJ Suding KN 2013 ). Single-trait functional indices outperform multi-trait indices in linking environmental gradients and ecosystem services in a complex landscapeJournal of Ecology ,9 -17 . [本文引用: 1] [5] Cardoso P Rigal F Carvalho JC 2015 ). BAT-Biodiversity Assessment Tools, an R package for the measurement and estimation of alpha and beta taxon, phylogenetic and functional diversityMethods in Ecology and Evolution ,232 -236 . [本文引用: 2] [6] Casanoves F Pla L di Rienzo JA Díaz S 2011 ). FDiversity: A software package for the integrated analysis of functional diversityMethods in Ecology and Evolution ,233 -237 . [本文引用: 1] [7] Cavender-Bares J Kozak KH Fine PVA Kembel SW 2009 ). The merging of community ecology and phylogenetic biologyEcology Letters ,693 -715 . [本文引用: 1] [8] Chen TG Zhang JT Shangguan TL Zhang F Wang QX 2000 ). The study of diversity in Shenweigou of Guandi Mountain, Shanxi ProvinceActa Botanica Boreali-Occidentalia Sinica ,638 -646 . [本文引用: 2] [ 陈廷贵 , 张金屯 , 上官铁梁 , 张峰 , 王巧仙 ( 2000 ). 山西关帝山神尾沟植物群落多样性研究西北植物学报 , 20, 638 -646 .] [本文引用: 2] [9] Chi XL Tang ZY Fang JY 2014 ). Patterns of phylogenetic beta diversity in China’s grasslands in relation to geographical and environmental distanceBasic and Applied Ecology ,416 -425 . [本文引用: 1] [10] Faith DP 1992 ). Conservation evaluation and phylogenetic diversityBiological Conservation ,1 -10 . [本文引用: 1] [11] Fang JY Wang XP Shen ZH Tang ZY He JS Yu D Jiang Y Wang ZH Zheng CY Zhu JL Guo ZD 2009 ). Methods and protocols for plant community inventoryBiodiversity Science ,533 -548 . [本文引用: 1] [ 方精云 , 王襄平 , 沈泽昊 , 唐志尧 , 贺金生 , 于丹 , 江源 , 王志恒 , 郑成洋 , 朱江玲 , 郭兆迪 ( 2009 ). 植物群落清查的主要内容、方法和技术规范生物多样性 , 17, 533 -548 .] [本文引用: 1] [12] Fernandez-Going BM Harrison SP Anacker BL Safford HD 2013 ). Climate interacts with soil to produce beta diversity in Californian plant communitiesEcology ,2007 -2018 . [本文引用: 1] [13] Gao JF Zhang YX 2006 ). Distributional patterns of species diversity of main plant communities along altitudinal gradient in secondary forest region, Guandi Mountain, ChinaJournal of Forestry Research ,111 -115 . [本文引用: 1] [14] Itow S 1991 ). Species turnover and diversity patterns along an evergreen broad-leaved forest coenoclineJournal of Vegetation Science ,477 -484 . [本文引用: 1] [15] Jump AS Mátyás C Pe?uelas J 2009 ). The altitude-for- latitude disparity in the range retractions of woody speciesTrends in Ecology & Evolution ,694 -701 . [本文引用: 1] [16] Kluge J Kessler M 2011 ). Phylogenetic diversity, trait diversity and niches: Species assembly of ferns along a tropical elevational gradientJournal of Biogeography ,394 -405 . [本文引用: 1] [17] Mason NWH Mouillot D Lee WG Wilson JB 2005 ). Functional richness, functional evenness and functional divergence: The primary components of functional diversityOikos ,112 -118 . [本文引用: 1] [18] Ohsawa M 1991 ). Structural comparison of tropical montane rain forests along latitudinal and altitudinal gradients in south and east AsiaVegetatio ,1 -10 . [本文引用: 1] [19] Ohsawa M 1995 ). Latitudinal comparison of altitudinal changes in forest structure, leaf-type, and species richness in humid monsoon AsiaVegetatio ,3 -10 . [本文引用: 1] [20] Pausas JG Austin MP 2001 ). Patterns of plant species richness in relation to different environments: An appraisalJournal of Vegetation Science ,153 -166 . [本文引用: 2] [21] Qian H 2009 ). Beta diversity in relation to dispersal ability for vascular plants in North AmericaGlobal Ecology and Biogeography ,327 -332 . [本文引用: 1] [22] Qian H Field R Zhang JL Zhang J Chen SB 2016 ). Phylogenetic structure and ecological and evolutionary determinants of species richness for angiosperm trees in forest communities in ChinaJournal of Biogeography ,603 -615 . [本文引用: 1] [23] Qian H Hao ZQ Zhang J 2014 ). Phylogenetic structure and phylogenetic diversity of angiosperm assemblages in forests along an elevational gradient in Changbaishan, ChinaJournal of Plant Ecology ,154 -165 . [本文引用: 3] [24] Qin H Dong G Zhang F 2013 ). Distribution patterns of dominant populations of forest communities in Pangquangou National Nature Reserve, ShanxiBulletin of Botanical Research ,605 -609 . [本文引用: 1] [ 秦浩 , 董刚 , 张峰 ( 2013 ). 庞泉沟自然保护区森林群落优势种群分布格局研究植物研究 , 33, 605 -609 .] [本文引用: 1] [25] Rapoport EH 1982 ). Areography, Geographical Strategies of Species . Pergamon Press, New York. [本文引用: 1] [26] Tang ZY Fang JY Zhang L 2004 ). Patterns of woody plant species diversity along environmental gradients on Mt. Taibai, Qinling MountainsBiodiversity Science ,115 -122 . [本文引用: 2] [ 唐志尧 , 方精云 , 张玲 ( 2004 ). 秦岭太白山木本植物物种多样性的梯度格局及环境解释生物多样性 , 12, 115 -122 .] [本文引用: 2] [27] Wang GH Zhou GS Yang LZ Li ZQ 2002 ). Distribution, species diversity and life-form spectra of plant communities along an altitudinal gradient in the northern slopes of Qilianshan Mountains, Gansu, ChinaPlant Ecology ,169 -181 . [本文引用: 1] [28] Watkins Jr JE Cardelús C Colwell RK Moran RC 2006 ). Species richness and distribution of ferns along an elevational gradient in Costa RicaAmerican Journal of Botany ,73 -83 . [29] Webb CO Ackerly DD Kembel SW 2008 ). Phylocom: Software for the analysis of phylogenetic community structure and trait evolutionBioinformatics ,2098 -2100 . [本文引用: 3] [30] Whittaker RH Niering WA 1975 ). Vegetation of the Santa Catalina Mountains, Arizona. V. Biomass, production, and diversity along the elevation gradientEcology ,771 -790 . [本文引用: 1] [31] Zhang F Qiao LP Zhang GP Wu YZ Han H 2007 ). Quantitative analysis on interspecific relationships among the dominant of plant communities populations of abandoned land in Guandi Mountains, ShanxiJournal of Shanxi University (Natural Science Edition )290 -294 . [本文引用: 1] [ 张峰 , 乔利鹏 , 张桂萍 , 武玉珍 , 韩虹 ( 2007 ). 关帝山撂荒地植物群落种间关系数量分析山西大学学报(自然科学版) , 30, 290 -294 .] [本文引用: 1] [32] Zhang JT Zhang BY Qian ZY 2015 ). Functional diversity of Cercidiphyllum japonicum, communities in the Shennongjia Reserve, central ChinaJournal of Forestry Research, 171 -177 . [本文引用: 3] [33] Zhao XN Qin XJ Dong G Zhang F 2014 ). Taxonomic diversity of the plant communities in Pangquangou Nature Reserve, Shanxi Province, ChinaChinese Journal of Applied Ecolog y, 3437 -3442 . [本文引用: 4] [ 赵小娜 , 秦晓娟 , 董刚 , 张峰 ( 2014 ). 庞泉沟自然保护区植物群落分类学多样性应用生态学报 , 25, 3437 -3442 .] [本文引用: 4] [34] Zhu XZ Zhang JT 2005 ). Altitudinal patterns of plant diversity of China MountainsActa Botanica Boreali- Occidentalia Sinica ,1480 -1486 . [本文引用: 3] [ 朱珣之 , 张金屯 ( 2005 ). 中国山地植物多样性的垂直变化格局西北植物学报 , 25, 1480 -1486 .] [本文引用: 3] Altitudinal gradients in tropical forest composition, structure, and diversity in the Sierra de Manantlán
1
1998
... 物种、谱系和功能β多样性表现出较为一致的海拔格局(
图4 ).整体上来说, 随着海拔的升高,
β total 和
β repl 表现出上升的趋势, 而
β rich 表现出下降的趋势(
图4 A、4D、4G), 这与常见的β多样性随海拔单调递减的格局不一致(
Wang et al ., 2002 ).单调递减格局认为低海拔地区较高的温度和干扰, 导致种间竞争加剧, 造成较高的β多样性(
Antonio Vázquez & Givnish, 1998 ).根据Rapoport法则, 高海拔地区由于温度较低往往导致该地区的物种具有较宽的生态位, 分布范围相对较广, 因此相邻海拔区间的群落组成变化相对较小(
Rapoport, 1982 ).本研究涉及的海拔相对较低, 温度不是主要的决定因子.低海拔地区森林群落以油松林和辽东栎林为主, 物种组成大多为菊科喜阳耐旱的物种, 而β多样性较高的地区为落叶阔叶林向寒温性针叶林的过渡带, 物种组成具有较大的变化.
Itow (1991) 研究表明森林过渡带往往具有较高的β多样性.在不同海拔区段,
β repl 和
β rich 的相对作用不同.在低海拔地区, 森林管理、干扰强度和微生境等导致
β repl 和
β rich 共同影响了
β total ; 而在高海拔地区, 由于森林类型的不同导致群落间呈现较高的更替速率. ...
Plant diversity patterns in semi-natural grasslands along an elevational gradient in southern Norway
1
2002
... 以往多样性海拔格局的研究主要集中于物种多样性, 随着生物信息和功能属性等方面的研究愈加深入, 关于谱系多样性和功能多样性海拔格局的研究越来越多(
Butterfield & Suding, 2013 ).已有大量研究表明谱系和功能多样性随海拔梯度的变化主要表现出单峰格局, 即在中海拔地区出现最大值(
Austrheim, 2002; Watkins Jr et al ., 2006 ), 这主要与水热条件随海拔的变化有关(
Zhang et al ., 2015 ).在环境压力较大的条件下(低海拔的缺水和高海拔的低温), 环境过滤导致亲缘关系较近的或者功能属性相似的物种聚集在一起, 表现出相对较低的谱系和功能多样性(
Kluge & Kessler, 2011 ).另外, 不同生活型的植物对环境响应不同, 导致不同的多样性海拔格局.对于木本植物来说, 受温度的影响较大; 随着海拔的升高, 木本植物多样性逐渐降低(
Pausas & Austin, 2001 ).对于草本植物来说, 受光照、水分和土壤理化性质等多种因素影响, 具有多样的海拔格局(
Qian et al ., 2014 ). ...
Ecology: Individuals, Populations and Communities. 1
1996
... 揭示和阐明生物多样性空间格局及其维持机制是生态学研究的热点问题(
Cavender-Bares et al ., 2009 ).由于水热条件变化急剧, 山地一直是生态学家研究多样性海拔格局的理想场所(
Jump et al ., 2009 ).受水热条件、生物类群、微地形、土壤质地和人为干扰等因素的影响, 物种多样性表现出不同的海拔格局, 主要为递减格局和单峰格局(
唐志尧等, 2004 ;
朱珣之和张金屯, 2005 ).Ohsawa (
1991 ,
1995 )研究表明亚洲热带山地的物种多样性随海拔升高而降低, 而温带山地的物种多样性随海拔变化不明显.
朱珣之和张金屯(2005) 总结了我国不同气候带的19座山峰物种多样性海拔格局, 发现低纬度地区山地的物种多样性主要表现出单峰格局, 而高纬度地区山地的物种多样性主要以递减格局为主.
Begon等(1996) 认为物种多样性随海拔增加而递减的格局主要受热量因素影响, 随着海拔升高, 温度的快速下降导致多样性的下降.
Whittaker和Niering(1975) 认为物种多样性的单峰格局受水热条件的综合影响, 低海拔地区限制群落组成和多样性的关键因子主要是降水, 而在高海拔地区主要是温度. ...
Single-trait functional indices outperform multi-trait indices in linking environmental gradients and ecosystem services in a complex landscape
1
2013
... 以往多样性海拔格局的研究主要集中于物种多样性, 随着生物信息和功能属性等方面的研究愈加深入, 关于谱系多样性和功能多样性海拔格局的研究越来越多(
Butterfield & Suding, 2013 ).已有大量研究表明谱系和功能多样性随海拔梯度的变化主要表现出单峰格局, 即在中海拔地区出现最大值(
Austrheim, 2002; Watkins Jr et al ., 2006 ), 这主要与水热条件随海拔的变化有关(
Zhang et al ., 2015 ).在环境压力较大的条件下(低海拔的缺水和高海拔的低温), 环境过滤导致亲缘关系较近的或者功能属性相似的物种聚集在一起, 表现出相对较低的谱系和功能多样性(
Kluge & Kessler, 2011 ).另外, 不同生活型的植物对环境响应不同, 导致不同的多样性海拔格局.对于木本植物来说, 受温度的影响较大; 随着海拔的升高, 木本植物多样性逐渐降低(
Pausas & Austin, 2001 ).对于草本植物来说, 受光照、水分和土壤理化性质等多种因素影响, 具有多样的海拔格局(
Qian et al ., 2014 ). ...
BAT-Biodiversity Assessment Tools, an R package for the measurement and estimation of alpha and beta taxon, phylogenetic and functional diversity
2
2015
... 基于物种、系统发育和功能属性数据, 采用Jaccard相异性指数分别计算物种(
Tβ )、谱系(
Pβ )和功能(
Fβ ) β多样性指数.同时将总β多样性(
β total )进一步分解为更替(
β repl )和丰富度差异(
β rich )两部分之和(
Cardoso et al ., 2015 ).
β repl 指的是由于物种组成或者系统进化(功能属性)发生一对一替换所导致的群落间差异; 而
β rich 则强调由于物种组成或者系统进化(功能属性)在丰富度上的不同所导致的群落间差异.计算公式如下: ...
... 式中,
a 表示两个群落共有的物种数或者系统进化(功能属性)枝长;
b 和
c 分别表示两个群落各自所特有的物种数或者系统进化(功能属性)枝长.另外, 分别计算木本植物和草本植物的物种、谱系和功能β多样性.物种、谱系和功能β多样性计算在R语言的“BAT”包下完成(
Cardoso et al ., 2015 ). ...
FDiversity: A software package for the integrated analysis of functional diversity
1
2011
... 基于选取的植物功能属性, 应用R 3.4.3版本的“FD”软件包计算功能丰富度指数(
FRic )、功能均匀度指数(
FEve )和功能离散度指数(
FDiv )(
Mason et al ., 2005 ;
Casanoves et al ., 2011 ).另外, 分别计算木本植物和草本植物的功能α多样性指数. ...
The merging of community ecology and phylogenetic biology
1
2009
... 揭示和阐明生物多样性空间格局及其维持机制是生态学研究的热点问题(
Cavender-Bares et al ., 2009 ).由于水热条件变化急剧, 山地一直是生态学家研究多样性海拔格局的理想场所(
Jump et al ., 2009 ).受水热条件、生物类群、微地形、土壤质地和人为干扰等因素的影响, 物种多样性表现出不同的海拔格局, 主要为递减格局和单峰格局(
唐志尧等, 2004 ;
朱珣之和张金屯, 2005 ).Ohsawa (
1991 ,
1995 )研究表明亚洲热带山地的物种多样性随海拔升高而降低, 而温带山地的物种多样性随海拔变化不明显.
朱珣之和张金屯(2005) 总结了我国不同气候带的19座山峰物种多样性海拔格局, 发现低纬度地区山地的物种多样性主要表现出单峰格局, 而高纬度地区山地的物种多样性主要以递减格局为主.
Begon等(1996) 认为物种多样性随海拔增加而递减的格局主要受热量因素影响, 随着海拔升高, 温度的快速下降导致多样性的下降.
Whittaker和Niering(1975) 认为物种多样性的单峰格局受水热条件的综合影响, 低海拔地区限制群落组成和多样性的关键因子主要是降水, 而在高海拔地区主要是温度. ...
山西关帝山神尾沟植物群落多样性研究
2
2000
...
陈廷贵等(2000) 研究表明关帝山物种丰富度表现出不明显的海拔格局, 与本研究结果的差异主要表现在: (1)本研究仅涉及关帝山森林群落, 而陈廷贵等的研究不仅包括森林群落, 还包括灌丛群落和草本群落.(2)研究区海拔范围不同.本研究区域海拔较低(1 409-2 150 m), 该区域降水是多样性最主要的影响因素;
陈廷贵等(2000) 的研究区海拔较高(1 700-2 700 m), 涉及水热限制因子的转变, 因此多样性格局不明显.本研究与
Gao和Zhang (2006) 等在相同海拔段(1 400-2 000 m)的研究具有一致的多样性格局, 降水是影响低海拔地区多样性的重要因素, 随着海拔的升高, 降水的增加有利于多样性增加.本文的不足在于缺乏该区域高海拔地段的森林群落样方, 研究结果无法表明该区域整个海拔范围的森林群落多样性格局, 只是反映了中低海拔段的森林群落多样性格局.在以后的研究中, 我们希望能够补充高海拔地段的森林群落样方, 完善该区域整个海拔梯度上的多样性格局. ...
... 研究表明关帝山物种丰富度表现出不明显的海拔格局, 与本研究结果的差异主要表现在: (1)本研究仅涉及关帝山森林群落, 而陈廷贵等的研究不仅包括森林群落, 还包括灌丛群落和草本群落.(2)研究区海拔范围不同.本研究区域海拔较低(1 409-2 150 m), 该区域降水是多样性最主要的影响因素;
陈廷贵等(2000) 的研究区海拔较高(1 700-2 700 m), 涉及水热限制因子的转变, 因此多样性格局不明显.本研究与
Gao和Zhang (2006) 等在相同海拔段(1 400-2 000 m)的研究具有一致的多样性格局, 降水是影响低海拔地区多样性的重要因素, 随着海拔的升高, 降水的增加有利于多样性增加.本文的不足在于缺乏该区域高海拔地段的森林群落样方, 研究结果无法表明该区域整个海拔范围的森林群落多样性格局, 只是反映了中低海拔段的森林群落多样性格局.在以后的研究中, 我们希望能够补充高海拔地段的森林群落样方, 完善该区域整个海拔梯度上的多样性格局. ...
山西关帝山神尾沟植物群落多样性研究
2
2000
...
陈廷贵等(2000) 研究表明关帝山物种丰富度表现出不明显的海拔格局, 与本研究结果的差异主要表现在: (1)本研究仅涉及关帝山森林群落, 而陈廷贵等的研究不仅包括森林群落, 还包括灌丛群落和草本群落.(2)研究区海拔范围不同.本研究区域海拔较低(1 409-2 150 m), 该区域降水是多样性最主要的影响因素;
陈廷贵等(2000) 的研究区海拔较高(1 700-2 700 m), 涉及水热限制因子的转变, 因此多样性格局不明显.本研究与
Gao和Zhang (2006) 等在相同海拔段(1 400-2 000 m)的研究具有一致的多样性格局, 降水是影响低海拔地区多样性的重要因素, 随着海拔的升高, 降水的增加有利于多样性增加.本文的不足在于缺乏该区域高海拔地段的森林群落样方, 研究结果无法表明该区域整个海拔范围的森林群落多样性格局, 只是反映了中低海拔段的森林群落多样性格局.在以后的研究中, 我们希望能够补充高海拔地段的森林群落样方, 完善该区域整个海拔梯度上的多样性格局. ...
... 研究表明关帝山物种丰富度表现出不明显的海拔格局, 与本研究结果的差异主要表现在: (1)本研究仅涉及关帝山森林群落, 而陈廷贵等的研究不仅包括森林群落, 还包括灌丛群落和草本群落.(2)研究区海拔范围不同.本研究区域海拔较低(1 409-2 150 m), 该区域降水是多样性最主要的影响因素;
陈廷贵等(2000) 的研究区海拔较高(1 700-2 700 m), 涉及水热限制因子的转变, 因此多样性格局不明显.本研究与
Gao和Zhang (2006) 等在相同海拔段(1 400-2 000 m)的研究具有一致的多样性格局, 降水是影响低海拔地区多样性的重要因素, 随着海拔的升高, 降水的增加有利于多样性增加.本文的不足在于缺乏该区域高海拔地段的森林群落样方, 研究结果无法表明该区域整个海拔范围的森林群落多样性格局, 只是反映了中低海拔段的森林群落多样性格局.在以后的研究中, 我们希望能够补充高海拔地段的森林群落样方, 完善该区域整个海拔梯度上的多样性格局. ...
Patterns of phylogenetic beta diversity in China’s grasslands in relation to geographical and environmental distance
1
2014
... 随着环境差异和地理距离的增加, 群落间物种、谱系和功能β多样性均增加显著(
p < 0.001).这与以往的研究是一致的, 即随着环境差异和地理距离的增加, 群落间物种组成、亲缘关系和功能属性差异性增加(
Chi et al ., 2014 ).基于生态位理论的生态位分化假说认为不论群落间地理距离多大, 环境差异越大, 群落组成差异越大(
Fernandez-Going et al ., 2013 ).而基于中性理论的距离衰减假说认为不论群落间环境差异多大, 地理距离越大, 群落组成差异越大(
Qian, 2009 ).我们的研究表明, 对所有植物来说, 群落间物种、谱系和功能β多样性与环境距离的线性关系要强于地理距离(
图5 ,
图6A 、6D、6G).这表明在较小的地理单元上, 由于海拔梯度上存在较大的环境差异, 导致环境差异(环境过滤)在该区域的群落组成上发挥着重要作用, 而地理距离(扩散限制)的作用相对较小. ...
Conservation evaluation and phylogenetic diversity
1
1992
... 选择Faith’s谱系多样性指数(
PD )和平均谱系距离指数(
MPD )作为度量谱系α多样性指数.
PD 指数被定义为群落内所有物种进化分枝长度之和(
Faith, 1992 ).
MPD 指数是指群落内所有物种对间的平均谱系距离(
Webb et al ., 2008 ). ...
植物群落清查的主要内容、方法和技术规范
1
2009
... 2011-2013年, 采用典型取样的方法沿海拔梯度在孝文山、云顶山、庞泉沟、双家寨和西葫芦等地共调查52块森林样方, 海拔1 409-2 150 m (
图1 ).本文主要关注关帝山中低海拔段森林群落的多样性海拔格局, 并未涉及灌丛和草地类型.森林样方面积为20 m × 30 m, 将其划分为6个10 m × 10 m的样格, 对样格内胸径(
DBH ) ≥3 cm的乔木进行每木调查, 记录乔木的种名、胸径、高度和健康状况.在样方对角线处选择2个10 m × 10 m的样格进行灌木层调查, 记录灌木的平均基径、平均高度、盖度和丛数.在6个10 m × 10 m的样格中分别布设1个1 m × 1 m的小样方, 进行草本层调查, 记录草本的多度、盖度和平均高度.同时, 以记名方式记录样格外草本植物名称.另外, 记录每个样地的地理信息(经度、纬度、海拔、地形、坡度、坡向和坡位)、干扰程度、土壤类型以及人为活动等情况(
方精云等, 2009 ). ...
植物群落清查的主要内容、方法和技术规范
1
2009
... 2011-2013年, 采用典型取样的方法沿海拔梯度在孝文山、云顶山、庞泉沟、双家寨和西葫芦等地共调查52块森林样方, 海拔1 409-2 150 m (
图1 ).本文主要关注关帝山中低海拔段森林群落的多样性海拔格局, 并未涉及灌丛和草地类型.森林样方面积为20 m × 30 m, 将其划分为6个10 m × 10 m的样格, 对样格内胸径(
DBH ) ≥3 cm的乔木进行每木调查, 记录乔木的种名、胸径、高度和健康状况.在样方对角线处选择2个10 m × 10 m的样格进行灌木层调查, 记录灌木的平均基径、平均高度、盖度和丛数.在6个10 m × 10 m的样格中分别布设1个1 m × 1 m的小样方, 进行草本层调查, 记录草本的多度、盖度和平均高度.同时, 以记名方式记录样格外草本植物名称.另外, 记录每个样地的地理信息(经度、纬度、海拔、地形、坡度、坡向和坡位)、干扰程度、土壤类型以及人为活动等情况(
方精云等, 2009 ). ...
Climate interacts with soil to produce beta diversity in Californian plant communities
1
2013
... 随着环境差异和地理距离的增加, 群落间物种、谱系和功能β多样性均增加显著(
p < 0.001).这与以往的研究是一致的, 即随着环境差异和地理距离的增加, 群落间物种组成、亲缘关系和功能属性差异性增加(
Chi et al ., 2014 ).基于生态位理论的生态位分化假说认为不论群落间地理距离多大, 环境差异越大, 群落组成差异越大(
Fernandez-Going et al ., 2013 ).而基于中性理论的距离衰减假说认为不论群落间环境差异多大, 地理距离越大, 群落组成差异越大(
Qian, 2009 ).我们的研究表明, 对所有植物来说, 群落间物种、谱系和功能β多样性与环境距离的线性关系要强于地理距离(
图5 ,
图6A 、6D、6G).这表明在较小的地理单元上, 由于海拔梯度上存在较大的环境差异, 导致环境差异(环境过滤)在该区域的群落组成上发挥着重要作用, 而地理距离(扩散限制)的作用相对较小. ...
Distributional patterns of species diversity of main plant communities along altitudinal gradient in secondary forest region, Guandi Mountain, China
1
2006
...
陈廷贵等(2000) 研究表明关帝山物种丰富度表现出不明显的海拔格局, 与本研究结果的差异主要表现在: (1)本研究仅涉及关帝山森林群落, 而陈廷贵等的研究不仅包括森林群落, 还包括灌丛群落和草本群落.(2)研究区海拔范围不同.本研究区域海拔较低(1 409-2 150 m), 该区域降水是多样性最主要的影响因素;
陈廷贵等(2000) 的研究区海拔较高(1 700-2 700 m), 涉及水热限制因子的转变, 因此多样性格局不明显.本研究与
Gao和Zhang (2006) 等在相同海拔段(1 400-2 000 m)的研究具有一致的多样性格局, 降水是影响低海拔地区多样性的重要因素, 随着海拔的升高, 降水的增加有利于多样性增加.本文的不足在于缺乏该区域高海拔地段的森林群落样方, 研究结果无法表明该区域整个海拔范围的森林群落多样性格局, 只是反映了中低海拔段的森林群落多样性格局.在以后的研究中, 我们希望能够补充高海拔地段的森林群落样方, 完善该区域整个海拔梯度上的多样性格局. ...
Species turnover and diversity patterns along an evergreen broad-leaved forest coenocline
1
1991
... 物种、谱系和功能β多样性表现出较为一致的海拔格局(
图4 ).整体上来说, 随着海拔的升高,
β total 和
β repl 表现出上升的趋势, 而
β rich 表现出下降的趋势(
图4 A、4D、4G), 这与常见的β多样性随海拔单调递减的格局不一致(
Wang et al ., 2002 ).单调递减格局认为低海拔地区较高的温度和干扰, 导致种间竞争加剧, 造成较高的β多样性(
Antonio Vázquez & Givnish, 1998 ).根据Rapoport法则, 高海拔地区由于温度较低往往导致该地区的物种具有较宽的生态位, 分布范围相对较广, 因此相邻海拔区间的群落组成变化相对较小(
Rapoport, 1982 ).本研究涉及的海拔相对较低, 温度不是主要的决定因子.低海拔地区森林群落以油松林和辽东栎林为主, 物种组成大多为菊科喜阳耐旱的物种, 而β多样性较高的地区为落叶阔叶林向寒温性针叶林的过渡带, 物种组成具有较大的变化.
Itow (1991) 研究表明森林过渡带往往具有较高的β多样性.在不同海拔区段,
β repl 和
β rich 的相对作用不同.在低海拔地区, 森林管理、干扰强度和微生境等导致
β repl 和
β rich 共同影响了
β total ; 而在高海拔地区, 由于森林类型的不同导致群落间呈现较高的更替速率. ...
The altitude-for- latitude disparity in the range retractions of woody species
1
2009
... 揭示和阐明生物多样性空间格局及其维持机制是生态学研究的热点问题(
Cavender-Bares et al ., 2009 ).由于水热条件变化急剧, 山地一直是生态学家研究多样性海拔格局的理想场所(
Jump et al ., 2009 ).受水热条件、生物类群、微地形、土壤质地和人为干扰等因素的影响, 物种多样性表现出不同的海拔格局, 主要为递减格局和单峰格局(
唐志尧等, 2004 ;
朱珣之和张金屯, 2005 ).Ohsawa (
1991 ,
1995 )研究表明亚洲热带山地的物种多样性随海拔升高而降低, 而温带山地的物种多样性随海拔变化不明显.
朱珣之和张金屯(2005) 总结了我国不同气候带的19座山峰物种多样性海拔格局, 发现低纬度地区山地的物种多样性主要表现出单峰格局, 而高纬度地区山地的物种多样性主要以递减格局为主.
Begon等(1996) 认为物种多样性随海拔增加而递减的格局主要受热量因素影响, 随着海拔升高, 温度的快速下降导致多样性的下降.
Whittaker和Niering(1975) 认为物种多样性的单峰格局受水热条件的综合影响, 低海拔地区限制群落组成和多样性的关键因子主要是降水, 而在高海拔地区主要是温度. ...
Phylogenetic diversity, trait diversity and niches: Species assembly of ferns along a tropical elevational gradient
1
2011
... 以往多样性海拔格局的研究主要集中于物种多样性, 随着生物信息和功能属性等方面的研究愈加深入, 关于谱系多样性和功能多样性海拔格局的研究越来越多(
Butterfield & Suding, 2013 ).已有大量研究表明谱系和功能多样性随海拔梯度的变化主要表现出单峰格局, 即在中海拔地区出现最大值(
Austrheim, 2002; Watkins Jr et al ., 2006 ), 这主要与水热条件随海拔的变化有关(
Zhang et al ., 2015 ).在环境压力较大的条件下(低海拔的缺水和高海拔的低温), 环境过滤导致亲缘关系较近的或者功能属性相似的物种聚集在一起, 表现出相对较低的谱系和功能多样性(
Kluge & Kessler, 2011 ).另外, 不同生活型的植物对环境响应不同, 导致不同的多样性海拔格局.对于木本植物来说, 受温度的影响较大; 随着海拔的升高, 木本植物多样性逐渐降低(
Pausas & Austin, 2001 ).对于草本植物来说, 受光照、水分和土壤理化性质等多种因素影响, 具有多样的海拔格局(
Qian et al ., 2014 ). ...
Functional richness, functional evenness and functional divergence: The primary components of functional diversity
1
2005
... 基于选取的植物功能属性, 应用R 3.4.3版本的“FD”软件包计算功能丰富度指数(
FRic )、功能均匀度指数(
FEve )和功能离散度指数(
FDiv )(
Mason et al ., 2005 ;
Casanoves et al ., 2011 ).另外, 分别计算木本植物和草本植物的功能α多样性指数. ...
Structural comparison of tropical montane rain forests along latitudinal and altitudinal gradients in south and east Asia
1
1991
... 揭示和阐明生物多样性空间格局及其维持机制是生态学研究的热点问题(
Cavender-Bares et al ., 2009 ).由于水热条件变化急剧, 山地一直是生态学家研究多样性海拔格局的理想场所(
Jump et al ., 2009 ).受水热条件、生物类群、微地形、土壤质地和人为干扰等因素的影响, 物种多样性表现出不同的海拔格局, 主要为递减格局和单峰格局(
唐志尧等, 2004 ;
朱珣之和张金屯, 2005 ).Ohsawa (
1991 ,
1995 )研究表明亚洲热带山地的物种多样性随海拔升高而降低, 而温带山地的物种多样性随海拔变化不明显.
朱珣之和张金屯(2005) 总结了我国不同气候带的19座山峰物种多样性海拔格局, 发现低纬度地区山地的物种多样性主要表现出单峰格局, 而高纬度地区山地的物种多样性主要以递减格局为主.
Begon等(1996) 认为物种多样性随海拔增加而递减的格局主要受热量因素影响, 随着海拔升高, 温度的快速下降导致多样性的下降.
Whittaker和Niering(1975) 认为物种多样性的单峰格局受水热条件的综合影响, 低海拔地区限制群落组成和多样性的关键因子主要是降水, 而在高海拔地区主要是温度. ...
Latitudinal comparison of altitudinal changes in forest structure, leaf-type, and species richness in humid monsoon Asia
1
1995
... 揭示和阐明生物多样性空间格局及其维持机制是生态学研究的热点问题(
Cavender-Bares et al ., 2009 ).由于水热条件变化急剧, 山地一直是生态学家研究多样性海拔格局的理想场所(
Jump et al ., 2009 ).受水热条件、生物类群、微地形、土壤质地和人为干扰等因素的影响, 物种多样性表现出不同的海拔格局, 主要为递减格局和单峰格局(
唐志尧等, 2004 ;
朱珣之和张金屯, 2005 ).Ohsawa (
1991 ,
1995 )研究表明亚洲热带山地的物种多样性随海拔升高而降低, 而温带山地的物种多样性随海拔变化不明显.
朱珣之和张金屯(2005) 总结了我国不同气候带的19座山峰物种多样性海拔格局, 发现低纬度地区山地的物种多样性主要表现出单峰格局, 而高纬度地区山地的物种多样性主要以递减格局为主.
Begon等(1996) 认为物种多样性随海拔增加而递减的格局主要受热量因素影响, 随着海拔升高, 温度的快速下降导致多样性的下降.
Whittaker和Niering(1975) 认为物种多样性的单峰格局受水热条件的综合影响, 低海拔地区限制群落组成和多样性的关键因子主要是降水, 而在高海拔地区主要是温度. ...
Patterns of plant species richness in relation to different environments: An appraisal
2
2001
... 以往多样性海拔格局的研究主要集中于物种多样性, 随着生物信息和功能属性等方面的研究愈加深入, 关于谱系多样性和功能多样性海拔格局的研究越来越多(
Butterfield & Suding, 2013 ).已有大量研究表明谱系和功能多样性随海拔梯度的变化主要表现出单峰格局, 即在中海拔地区出现最大值(
Austrheim, 2002; Watkins Jr et al ., 2006 ), 这主要与水热条件随海拔的变化有关(
Zhang et al ., 2015 ).在环境压力较大的条件下(低海拔的缺水和高海拔的低温), 环境过滤导致亲缘关系较近的或者功能属性相似的物种聚集在一起, 表现出相对较低的谱系和功能多样性(
Kluge & Kessler, 2011 ).另外, 不同生活型的植物对环境响应不同, 导致不同的多样性海拔格局.对于木本植物来说, 受温度的影响较大; 随着海拔的升高, 木本植物多样性逐渐降低(
Pausas & Austin, 2001 ).对于草本植物来说, 受光照、水分和土壤理化性质等多种因素影响, 具有多样的海拔格局(
Qian et al ., 2014 ). ...
... 木本植物和草本植物多样性海拔格局表现出较大的差异, 这主要与它们对环境响应和生活史策略不同有关.木本植物受温度影响较大(
Pausas & Austin, 2001 ), 草本植物受光照、水分、微地形、土壤理化性质和人为活动等影响更大, 表现出多样的海拔格局(
Qian et al ., 2014 ).随着海拔的升高, 草本植物
S 和
H′ 的上升趋势要高于木本植物.在关帝山低海拔地区, 降水是影响草本植物多样性格局最主要的因子.随着海拔升高, 降水量逐渐增加, 草本植物多样性明显增加.而对于木本植物来说, 影响其分布的主要因素是历史过程(
MPD 和
NRI ).这表明木本植物在系统进化方面更加保守, 对环境的适应性和生活史策略与草本植物有所差异.
NRI 指数表明关帝山森林群落的木本植物大多表现出谱系聚集的结构, 即亲缘关系较近的物种共存; 而草本植物大多表现出谱系发散的结构, 即亲缘关系较远的物种 共存. ...
Beta diversity in relation to dispersal ability for vascular plants in North America
1
2009
... 随着环境差异和地理距离的增加, 群落间物种、谱系和功能β多样性均增加显著(
p < 0.001).这与以往的研究是一致的, 即随着环境差异和地理距离的增加, 群落间物种组成、亲缘关系和功能属性差异性增加(
Chi et al ., 2014 ).基于生态位理论的生态位分化假说认为不论群落间地理距离多大, 环境差异越大, 群落组成差异越大(
Fernandez-Going et al ., 2013 ).而基于中性理论的距离衰减假说认为不论群落间环境差异多大, 地理距离越大, 群落组成差异越大(
Qian, 2009 ).我们的研究表明, 对所有植物来说, 群落间物种、谱系和功能β多样性与环境距离的线性关系要强于地理距离(
图5 ,
图6A 、6D、6G).这表明在较小的地理单元上, 由于海拔梯度上存在较大的环境差异, 导致环境差异(环境过滤)在该区域的群落组成上发挥着重要作用, 而地理距离(扩散限制)的作用相对较小. ...
Phylogenetic structure and ecological and evolutionary determinants of species richness for angiosperm trees in forest communities in China
1
2016
... 对于α多样性指数, 首先将物种、谱系和功能α多样性指数与海拔进行回归分析, 研究α多样性的海拔格局.其次, 基于R语言的“Vegan”软件包, 采用方差分解法分析了地形因子、群落结构和历史过程对物种丰富度的相对影响.历史过程选用
MPD 和
NRI 来间接反映,
MPD 和
NRI 指数可以反映群落的系统进化背景和物种间的谱系关系(
Qian et al ., 2016 ). ...
Phylogenetic structure and phylogenetic diversity of angiosperm assemblages in forests along an elevational gradient in Changbaishan, China
3
2014
... 以往多样性海拔格局的研究主要集中于物种多样性, 随着生物信息和功能属性等方面的研究愈加深入, 关于谱系多样性和功能多样性海拔格局的研究越来越多(
Butterfield & Suding, 2013 ).已有大量研究表明谱系和功能多样性随海拔梯度的变化主要表现出单峰格局, 即在中海拔地区出现最大值(
Austrheim, 2002; Watkins Jr et al ., 2006 ), 这主要与水热条件随海拔的变化有关(
Zhang et al ., 2015 ).在环境压力较大的条件下(低海拔的缺水和高海拔的低温), 环境过滤导致亲缘关系较近的或者功能属性相似的物种聚集在一起, 表现出相对较低的谱系和功能多样性(
Kluge & Kessler, 2011 ).另外, 不同生活型的植物对环境响应不同, 导致不同的多样性海拔格局.对于木本植物来说, 受温度的影响较大; 随着海拔的升高, 木本植物多样性逐渐降低(
Pausas & Austin, 2001 ).对于草本植物来说, 受光照、水分和土壤理化性质等多种因素影响, 具有多样的海拔格局(
Qian et al ., 2014 ). ...
... 另外, 环境差异和地理距离对不同生活型植物的β多样性影响不同.环境差异(环境过滤)对木本植物的β多样性具有相对较强的作用; 而环境差异(环境过滤)和地理距离(扩散限制)共同作用于草本植物的β多样性.
Qian等(2014) 对长白山谱系多样性的海拔格局研究也表明, 环境差异对木本植物β多样性的影响要高于草本植物.环境差异和地理距离对群落β多样性总的解释力整体上不超过30%, 表明还有其他因素(如土壤质地、土壤动物、微生物等)影响了群落的β多样性. ...
... 木本植物和草本植物多样性海拔格局表现出较大的差异, 这主要与它们对环境响应和生活史策略不同有关.木本植物受温度影响较大(
Pausas & Austin, 2001 ), 草本植物受光照、水分、微地形、土壤理化性质和人为活动等影响更大, 表现出多样的海拔格局(
Qian et al ., 2014 ).随着海拔的升高, 草本植物
S 和
H′ 的上升趋势要高于木本植物.在关帝山低海拔地区, 降水是影响草本植物多样性格局最主要的因子.随着海拔升高, 降水量逐渐增加, 草本植物多样性明显增加.而对于木本植物来说, 影响其分布的主要因素是历史过程(
MPD 和
NRI ).这表明木本植物在系统进化方面更加保守, 对环境的适应性和生活史策略与草本植物有所差异.
NRI 指数表明关帝山森林群落的木本植物大多表现出谱系聚集的结构, 即亲缘关系较近的物种共存; 而草本植物大多表现出谱系发散的结构, 即亲缘关系较远的物种 共存. ...
庞泉沟自然保护区森林群落优势种群分布格局研究
1
2013
... 山西关帝山地处吕梁山脉中段, 地理位置: 37.33°-38.33° N, 110.30°-111.30° E.气候属暖温带大陆性季风气候, 年平均气温3.5 ℃, 最冷月(1月)平均气温-10.6 ℃, 最暖月(7月)平均气温16.1 ℃; 年降水量830 mm.区域土壤类型主要包括山地棕壤、山地褐土、亚高山草甸土和河漫滩草甸土等(
张峰等, 2007 ;
秦浩等, 2013 ). ...
庞泉沟自然保护区森林群落优势种群分布格局研究
1
2013
... 山西关帝山地处吕梁山脉中段, 地理位置: 37.33°-38.33° N, 110.30°-111.30° E.气候属暖温带大陆性季风气候, 年平均气温3.5 ℃, 最冷月(1月)平均气温-10.6 ℃, 最暖月(7月)平均气温16.1 ℃; 年降水量830 mm.区域土壤类型主要包括山地棕壤、山地褐土、亚高山草甸土和河漫滩草甸土等(
张峰等, 2007 ;
秦浩等, 2013 ). ...
1
1982
... 物种、谱系和功能β多样性表现出较为一致的海拔格局(
图4 ).整体上来说, 随着海拔的升高,
β total 和
β repl 表现出上升的趋势, 而
β rich 表现出下降的趋势(
图4 A、4D、4G), 这与常见的β多样性随海拔单调递减的格局不一致(
Wang et al ., 2002 ).单调递减格局认为低海拔地区较高的温度和干扰, 导致种间竞争加剧, 造成较高的β多样性(
Antonio Vázquez & Givnish, 1998 ).根据Rapoport法则, 高海拔地区由于温度较低往往导致该地区的物种具有较宽的生态位, 分布范围相对较广, 因此相邻海拔区间的群落组成变化相对较小(
Rapoport, 1982 ).本研究涉及的海拔相对较低, 温度不是主要的决定因子.低海拔地区森林群落以油松林和辽东栎林为主, 物种组成大多为菊科喜阳耐旱的物种, 而β多样性较高的地区为落叶阔叶林向寒温性针叶林的过渡带, 物种组成具有较大的变化.
Itow (1991) 研究表明森林过渡带往往具有较高的β多样性.在不同海拔区段,
β repl 和
β rich 的相对作用不同.在低海拔地区, 森林管理、干扰强度和微生境等导致
β repl 和
β rich 共同影响了
β total ; 而在高海拔地区, 由于森林类型的不同导致群落间呈现较高的更替速率. ...
秦岭太白山木本植物物种多样性的梯度格局及环境解释
2
2004
... 揭示和阐明生物多样性空间格局及其维持机制是生态学研究的热点问题(
Cavender-Bares et al ., 2009 ).由于水热条件变化急剧, 山地一直是生态学家研究多样性海拔格局的理想场所(
Jump et al ., 2009 ).受水热条件、生物类群、微地形、土壤质地和人为干扰等因素的影响, 物种多样性表现出不同的海拔格局, 主要为递减格局和单峰格局(
唐志尧等, 2004 ;
朱珣之和张金屯, 2005 ).Ohsawa (
1991 ,
1995 )研究表明亚洲热带山地的物种多样性随海拔升高而降低, 而温带山地的物种多样性随海拔变化不明显.
朱珣之和张金屯(2005) 总结了我国不同气候带的19座山峰物种多样性海拔格局, 发现低纬度地区山地的物种多样性主要表现出单峰格局, 而高纬度地区山地的物种多样性主要以递减格局为主.
Begon等(1996) 认为物种多样性随海拔增加而递减的格局主要受热量因素影响, 随着海拔升高, 温度的快速下降导致多样性的下降.
Whittaker和Niering(1975) 认为物种多样性的单峰格局受水热条件的综合影响, 低海拔地区限制群落组成和多样性的关键因子主要是降水, 而在高海拔地区主要是温度. ...
... 随着海拔的升高,
S 、
PD 和
FRic 整体上表现出上升的趋势, 特别是在海拔1 800 m以上(
图2 ).这主要与我们研究的海拔范围(1 409-2 150 m)有关, 在中低海拔区段, 降水是影响该区域多样性最主要的因素(
赵小娜等, 2014 ).随着海拔的升高, 降水量逐渐增加,
S 、
PD 和
FRic 整体上呈现上升的趋势.不同气候带下, 山地α多样性海拔格局的差异主要与气候类型、地形地貌、植被类型和人为活动等因素密切相关, 但水热条件是影响多样性海拔格局的最主要因素(
朱珣之和张金屯, 2005 ).在水分条件较好的区域, 随着海拔升高, 温度快速下降是导致多样性降低的主要原因; 而受水分条件限制的区域, 水热条件的制约和平衡是影响多样性海拔格局的关键.在低海拔区域, 降水是影响群落组成和多样性的关键因子; 而在高海拔区域, 低温成为限制群落组成和多样性的关键因子(
唐志尧等, 2004 ;
Zhang et al ., 2015 ).随着海拔的升高, 群落的
FEve 和
E 指数表现出单峰格局, 最大值大致出现在海拔1 800 m处.该海拔段是温性林向寒温性林逐渐过渡的区域, 群落组成兼有耐旱性和耐寒性物种, 表现出较高的均匀性.随着海拔的升高,
NRI 是逐渐下降的, 群落谱系结构由聚集向随机或者发散过渡.在低海拔区域, 环境过滤(较少的降水量和较强的人为干扰)导致群落内亲缘关系较近的物种聚集在一起, 表现出较强的谱系聚集.而在海拔相对较高的区域, 环境过滤和种间关系可能共同作用于群落组成, 进而表现出随机或者发散的谱系结构.
NRI 指数结果表明群落内木本植物大多为亲缘关系较近的物种(
NRI > 0), 而草本植物大多为亲缘关系较远的物种(
NRI < 0). ...
秦岭太白山木本植物物种多样性的梯度格局及环境解释
2
2004
... 揭示和阐明生物多样性空间格局及其维持机制是生态学研究的热点问题(
Cavender-Bares et al ., 2009 ).由于水热条件变化急剧, 山地一直是生态学家研究多样性海拔格局的理想场所(
Jump et al ., 2009 ).受水热条件、生物类群、微地形、土壤质地和人为干扰等因素的影响, 物种多样性表现出不同的海拔格局, 主要为递减格局和单峰格局(
唐志尧等, 2004 ;
朱珣之和张金屯, 2005 ).Ohsawa (
1991 ,
1995 )研究表明亚洲热带山地的物种多样性随海拔升高而降低, 而温带山地的物种多样性随海拔变化不明显.
朱珣之和张金屯(2005) 总结了我国不同气候带的19座山峰物种多样性海拔格局, 发现低纬度地区山地的物种多样性主要表现出单峰格局, 而高纬度地区山地的物种多样性主要以递减格局为主.
Begon等(1996) 认为物种多样性随海拔增加而递减的格局主要受热量因素影响, 随着海拔升高, 温度的快速下降导致多样性的下降.
Whittaker和Niering(1975) 认为物种多样性的单峰格局受水热条件的综合影响, 低海拔地区限制群落组成和多样性的关键因子主要是降水, 而在高海拔地区主要是温度. ...
... 随着海拔的升高,
S 、
PD 和
FRic 整体上表现出上升的趋势, 特别是在海拔1 800 m以上(
图2 ).这主要与我们研究的海拔范围(1 409-2 150 m)有关, 在中低海拔区段, 降水是影响该区域多样性最主要的因素(
赵小娜等, 2014 ).随着海拔的升高, 降水量逐渐增加,
S 、
PD 和
FRic 整体上呈现上升的趋势.不同气候带下, 山地α多样性海拔格局的差异主要与气候类型、地形地貌、植被类型和人为活动等因素密切相关, 但水热条件是影响多样性海拔格局的最主要因素(
朱珣之和张金屯, 2005 ).在水分条件较好的区域, 随着海拔升高, 温度快速下降是导致多样性降低的主要原因; 而受水分条件限制的区域, 水热条件的制约和平衡是影响多样性海拔格局的关键.在低海拔区域, 降水是影响群落组成和多样性的关键因子; 而在高海拔区域, 低温成为限制群落组成和多样性的关键因子(
唐志尧等, 2004 ;
Zhang et al ., 2015 ).随着海拔的升高, 群落的
FEve 和
E 指数表现出单峰格局, 最大值大致出现在海拔1 800 m处.该海拔段是温性林向寒温性林逐渐过渡的区域, 群落组成兼有耐旱性和耐寒性物种, 表现出较高的均匀性.随着海拔的升高,
NRI 是逐渐下降的, 群落谱系结构由聚集向随机或者发散过渡.在低海拔区域, 环境过滤(较少的降水量和较强的人为干扰)导致群落内亲缘关系较近的物种聚集在一起, 表现出较强的谱系聚集.而在海拔相对较高的区域, 环境过滤和种间关系可能共同作用于群落组成, 进而表现出随机或者发散的谱系结构.
NRI 指数结果表明群落内木本植物大多为亲缘关系较近的物种(
NRI > 0), 而草本植物大多为亲缘关系较远的物种(
NRI < 0). ...
Distribution, species diversity and life-form spectra of plant communities along an altitudinal gradient in the northern slopes of Qilianshan Mountains, Gansu, China
1
2002
... 物种、谱系和功能β多样性表现出较为一致的海拔格局(
图4 ).整体上来说, 随着海拔的升高,
β total 和
β repl 表现出上升的趋势, 而
β rich 表现出下降的趋势(
图4 A、4D、4G), 这与常见的β多样性随海拔单调递减的格局不一致(
Wang et al ., 2002 ).单调递减格局认为低海拔地区较高的温度和干扰, 导致种间竞争加剧, 造成较高的β多样性(
Antonio Vázquez & Givnish, 1998 ).根据Rapoport法则, 高海拔地区由于温度较低往往导致该地区的物种具有较宽的生态位, 分布范围相对较广, 因此相邻海拔区间的群落组成变化相对较小(
Rapoport, 1982 ).本研究涉及的海拔相对较低, 温度不是主要的决定因子.低海拔地区森林群落以油松林和辽东栎林为主, 物种组成大多为菊科喜阳耐旱的物种, 而β多样性较高的地区为落叶阔叶林向寒温性针叶林的过渡带, 物种组成具有较大的变化.
Itow (1991) 研究表明森林过渡带往往具有较高的β多样性.在不同海拔区段,
β repl 和
β rich 的相对作用不同.在低海拔地区, 森林管理、干扰强度和微生境等导致
β repl 和
β rich 共同影响了
β total ; 而在高海拔地区, 由于森林类型的不同导致群落间呈现较高的更替速率. ...
Species richness and distribution of ferns along an elevational gradient in Costa Rica
2006
Phylocom: Software for the analysis of phylogenetic community structure and trait evolution
3
2008
... 选择Faith’s谱系多样性指数(
PD )和平均谱系距离指数(
MPD )作为度量谱系α多样性指数.
PD 指数被定义为群落内所有物种进化分枝长度之和(
Faith, 1992 ).
MPD 指数是指群落内所有物种对间的平均谱系距离(
Webb et al ., 2008 ). ...
... 选取净关联指数(
NRI )来表征群落的谱系结构, 反映群落内物种对间的亲缘关系(
Webb et al ., 2008 ).计算公式如下: ...
... 当
NRI > 0时, 群落谱系结构是聚集的, 即亲缘关系较近的物种聚集在一起; 当
NRI < 0时, 群落谱系结构是发散的, 即群落中存在较多亲缘关系远的物种; 而当
NRI 在0上下波动时, 群落中的物种组成是随机的(
Webb et al ., 2008 ). ...
Vegetation of the Santa Catalina Mountains, Arizona. V. Biomass, production, and diversity along the elevation gradient
1
1975
... 揭示和阐明生物多样性空间格局及其维持机制是生态学研究的热点问题(
Cavender-Bares et al ., 2009 ).由于水热条件变化急剧, 山地一直是生态学家研究多样性海拔格局的理想场所(
Jump et al ., 2009 ).受水热条件、生物类群、微地形、土壤质地和人为干扰等因素的影响, 物种多样性表现出不同的海拔格局, 主要为递减格局和单峰格局(
唐志尧等, 2004 ;
朱珣之和张金屯, 2005 ).Ohsawa (
1991 ,
1995 )研究表明亚洲热带山地的物种多样性随海拔升高而降低, 而温带山地的物种多样性随海拔变化不明显.
朱珣之和张金屯(2005) 总结了我国不同气候带的19座山峰物种多样性海拔格局, 发现低纬度地区山地的物种多样性主要表现出单峰格局, 而高纬度地区山地的物种多样性主要以递减格局为主.
Begon等(1996) 认为物种多样性随海拔增加而递减的格局主要受热量因素影响, 随着海拔升高, 温度的快速下降导致多样性的下降.
Whittaker和Niering(1975) 认为物种多样性的单峰格局受水热条件的综合影响, 低海拔地区限制群落组成和多样性的关键因子主要是降水, 而在高海拔地区主要是温度. ...
关帝山撂荒地植物群落种间关系数量分析
1
2007
... 山西关帝山地处吕梁山脉中段, 地理位置: 37.33°-38.33° N, 110.30°-111.30° E.气候属暖温带大陆性季风气候, 年平均气温3.5 ℃, 最冷月(1月)平均气温-10.6 ℃, 最暖月(7月)平均气温16.1 ℃; 年降水量830 mm.区域土壤类型主要包括山地棕壤、山地褐土、亚高山草甸土和河漫滩草甸土等(
张峰等, 2007 ;
秦浩等, 2013 ). ...
关帝山撂荒地植物群落种间关系数量分析
1
2007
... 山西关帝山地处吕梁山脉中段, 地理位置: 37.33°-38.33° N, 110.30°-111.30° E.气候属暖温带大陆性季风气候, 年平均气温3.5 ℃, 最冷月(1月)平均气温-10.6 ℃, 最暖月(7月)平均气温16.1 ℃; 年降水量830 mm.区域土壤类型主要包括山地棕壤、山地褐土、亚高山草甸土和河漫滩草甸土等(
张峰等, 2007 ;
秦浩等, 2013 ). ...
Functional diversity of
Cercidiphyllum japonicum, communities in the Shennongjia Reserve, central China
3
2015
... 以往多样性海拔格局的研究主要集中于物种多样性, 随着生物信息和功能属性等方面的研究愈加深入, 关于谱系多样性和功能多样性海拔格局的研究越来越多(
Butterfield & Suding, 2013 ).已有大量研究表明谱系和功能多样性随海拔梯度的变化主要表现出单峰格局, 即在中海拔地区出现最大值(
Austrheim, 2002; Watkins Jr et al ., 2006 ), 这主要与水热条件随海拔的变化有关(
Zhang et al ., 2015 ).在环境压力较大的条件下(低海拔的缺水和高海拔的低温), 环境过滤导致亲缘关系较近的或者功能属性相似的物种聚集在一起, 表现出相对较低的谱系和功能多样性(
Kluge & Kessler, 2011 ).另外, 不同生活型的植物对环境响应不同, 导致不同的多样性海拔格局.对于木本植物来说, 受温度的影响较大; 随着海拔的升高, 木本植物多样性逐渐降低(
Pausas & Austin, 2001 ).对于草本植物来说, 受光照、水分和土壤理化性质等多种因素影响, 具有多样的海拔格局(
Qian et al ., 2014 ). ...
... 选取植物的生长型、生活型、传粉方式、开花时间、花期、果实类型、结果时间、果期、种子传播方式、光合途径、光耐受性和固氮类型12个功能属性, 这些功能属性特征能很好地反映植物的生理和物候等特征(
Zhang et al ., 2015 ). ...
... 随着海拔的升高,
S 、
PD 和
FRic 整体上表现出上升的趋势, 特别是在海拔1 800 m以上(
图2 ).这主要与我们研究的海拔范围(1 409-2 150 m)有关, 在中低海拔区段, 降水是影响该区域多样性最主要的因素(
赵小娜等, 2014 ).随着海拔的升高, 降水量逐渐增加,
S 、
PD 和
FRic 整体上呈现上升的趋势.不同气候带下, 山地α多样性海拔格局的差异主要与气候类型、地形地貌、植被类型和人为活动等因素密切相关, 但水热条件是影响多样性海拔格局的最主要因素(
朱珣之和张金屯, 2005 ).在水分条件较好的区域, 随着海拔升高, 温度快速下降是导致多样性降低的主要原因; 而受水分条件限制的区域, 水热条件的制约和平衡是影响多样性海拔格局的关键.在低海拔区域, 降水是影响群落组成和多样性的关键因子; 而在高海拔区域, 低温成为限制群落组成和多样性的关键因子(
唐志尧等, 2004 ;
Zhang et al ., 2015 ).随着海拔的升高, 群落的
FEve 和
E 指数表现出单峰格局, 最大值大致出现在海拔1 800 m处.该海拔段是温性林向寒温性林逐渐过渡的区域, 群落组成兼有耐旱性和耐寒性物种, 表现出较高的均匀性.随着海拔的升高,
NRI 是逐渐下降的, 群落谱系结构由聚集向随机或者发散过渡.在低海拔区域, 环境过滤(较少的降水量和较强的人为干扰)导致群落内亲缘关系较近的物种聚集在一起, 表现出较强的谱系聚集.而在海拔相对较高的区域, 环境过滤和种间关系可能共同作用于群落组成, 进而表现出随机或者发散的谱系结构.
NRI 指数结果表明群落内木本植物大多为亲缘关系较近的物种(
NRI > 0), 而草本植物大多为亲缘关系较远的物种(
NRI < 0). ...
庞泉沟自然保护区植物群落分类学多样性
4
2014
... 森林群落主要包括云杉林(Form.
Picea spp.)、华北落叶松林(Form.
Larix principis-rupprechtii )、桦木林(Form.
Betula spp.)、山杨林(Form.
Populus davidiana )和油松林(From.
Pinus tabulaeformis )等; 灌丛群落主要包括黄刺玫灌丛(Form.
Rosa xanthina )、沙棘灌丛(Form.
Hippophae rhamnoides subsp.
sinensis )和荆条灌丛(Form.
Vitex negundo var.
heterophylla )等; 草本群落主要包括蒿类草丛(Form.
Artemisia spp.)和薹草草甸(Form.
Carex spp.)等(
赵小娜等, 2014 ). ...
... 选取海拔、坡度、坡向和坡位作为地形变量, 其中将坡向和坡位数据转化为等级制数据.坡向数据以正北方向为0°, 45°夹角为一个等级, 划分为8个坡向等级, 1表示北坡(0°-22.5°, 337.5°-360°), 2表示东北坡(22.5°-67.5°), 3表示西北坡(292.5°-337.5°), 4表示东坡(67.5°-112.5°), 5表示西坡(247.5°-292.5°), 6表示东南坡(112.5°-157.5°), 7表示西南坡(202.5°- 247.5°), 8表示南坡(157.5°-202.5°)(
赵小娜等,2014 ); 坡位共分为5个等级, 即: 1 (下坡), 2 (中下坡), 3 (中坡), 4 (中上坡)和5 (上坡)(
赵小娜等, 2014 ). ...
... ); 坡位共分为5个等级, 即: 1 (下坡), 2 (中下坡), 3 (中坡), 4 (中上坡)和5 (上坡)(
赵小娜等, 2014 ). ...
... 随着海拔的升高,
S 、
PD 和
FRic 整体上表现出上升的趋势, 特别是在海拔1 800 m以上(
图2 ).这主要与我们研究的海拔范围(1 409-2 150 m)有关, 在中低海拔区段, 降水是影响该区域多样性最主要的因素(
赵小娜等, 2014 ).随着海拔的升高, 降水量逐渐增加,
S 、
PD 和
FRic 整体上呈现上升的趋势.不同气候带下, 山地α多样性海拔格局的差异主要与气候类型、地形地貌、植被类型和人为活动等因素密切相关, 但水热条件是影响多样性海拔格局的最主要因素(
朱珣之和张金屯, 2005 ).在水分条件较好的区域, 随着海拔升高, 温度快速下降是导致多样性降低的主要原因; 而受水分条件限制的区域, 水热条件的制约和平衡是影响多样性海拔格局的关键.在低海拔区域, 降水是影响群落组成和多样性的关键因子; 而在高海拔区域, 低温成为限制群落组成和多样性的关键因子(
唐志尧等, 2004 ;
Zhang et al ., 2015 ).随着海拔的升高, 群落的
FEve 和
E 指数表现出单峰格局, 最大值大致出现在海拔1 800 m处.该海拔段是温性林向寒温性林逐渐过渡的区域, 群落组成兼有耐旱性和耐寒性物种, 表现出较高的均匀性.随着海拔的升高,
NRI 是逐渐下降的, 群落谱系结构由聚集向随机或者发散过渡.在低海拔区域, 环境过滤(较少的降水量和较强的人为干扰)导致群落内亲缘关系较近的物种聚集在一起, 表现出较强的谱系聚集.而在海拔相对较高的区域, 环境过滤和种间关系可能共同作用于群落组成, 进而表现出随机或者发散的谱系结构.
NRI 指数结果表明群落内木本植物大多为亲缘关系较近的物种(
NRI > 0), 而草本植物大多为亲缘关系较远的物种(
NRI < 0). ...
庞泉沟自然保护区植物群落分类学多样性
4
2014
... 森林群落主要包括云杉林(Form.
Picea spp.)、华北落叶松林(Form.
Larix principis-rupprechtii )、桦木林(Form.
Betula spp.)、山杨林(Form.
Populus davidiana )和油松林(From.
Pinus tabulaeformis )等; 灌丛群落主要包括黄刺玫灌丛(Form.
Rosa xanthina )、沙棘灌丛(Form.
Hippophae rhamnoides subsp.
sinensis )和荆条灌丛(Form.
Vitex negundo var.
heterophylla )等; 草本群落主要包括蒿类草丛(Form.
Artemisia spp.)和薹草草甸(Form.
Carex spp.)等(
赵小娜等, 2014 ). ...
... 选取海拔、坡度、坡向和坡位作为地形变量, 其中将坡向和坡位数据转化为等级制数据.坡向数据以正北方向为0°, 45°夹角为一个等级, 划分为8个坡向等级, 1表示北坡(0°-22.5°, 337.5°-360°), 2表示东北坡(22.5°-67.5°), 3表示西北坡(292.5°-337.5°), 4表示东坡(67.5°-112.5°), 5表示西坡(247.5°-292.5°), 6表示东南坡(112.5°-157.5°), 7表示西南坡(202.5°- 247.5°), 8表示南坡(157.5°-202.5°)(
赵小娜等,2014 ); 坡位共分为5个等级, 即: 1 (下坡), 2 (中下坡), 3 (中坡), 4 (中上坡)和5 (上坡)(
赵小娜等, 2014 ). ...
... ); 坡位共分为5个等级, 即: 1 (下坡), 2 (中下坡), 3 (中坡), 4 (中上坡)和5 (上坡)(
赵小娜等, 2014 ). ...
... 随着海拔的升高,
S 、
PD 和
FRic 整体上表现出上升的趋势, 特别是在海拔1 800 m以上(
图2 ).这主要与我们研究的海拔范围(1 409-2 150 m)有关, 在中低海拔区段, 降水是影响该区域多样性最主要的因素(
赵小娜等, 2014 ).随着海拔的升高, 降水量逐渐增加,
S 、
PD 和
FRic 整体上呈现上升的趋势.不同气候带下, 山地α多样性海拔格局的差异主要与气候类型、地形地貌、植被类型和人为活动等因素密切相关, 但水热条件是影响多样性海拔格局的最主要因素(
朱珣之和张金屯, 2005 ).在水分条件较好的区域, 随着海拔升高, 温度快速下降是导致多样性降低的主要原因; 而受水分条件限制的区域, 水热条件的制约和平衡是影响多样性海拔格局的关键.在低海拔区域, 降水是影响群落组成和多样性的关键因子; 而在高海拔区域, 低温成为限制群落组成和多样性的关键因子(
唐志尧等, 2004 ;
Zhang et al ., 2015 ).随着海拔的升高, 群落的
FEve 和
E 指数表现出单峰格局, 最大值大致出现在海拔1 800 m处.该海拔段是温性林向寒温性林逐渐过渡的区域, 群落组成兼有耐旱性和耐寒性物种, 表现出较高的均匀性.随着海拔的升高,
NRI 是逐渐下降的, 群落谱系结构由聚集向随机或者发散过渡.在低海拔区域, 环境过滤(较少的降水量和较强的人为干扰)导致群落内亲缘关系较近的物种聚集在一起, 表现出较强的谱系聚集.而在海拔相对较高的区域, 环境过滤和种间关系可能共同作用于群落组成, 进而表现出随机或者发散的谱系结构.
NRI 指数结果表明群落内木本植物大多为亲缘关系较近的物种(
NRI > 0), 而草本植物大多为亲缘关系较远的物种(
NRI < 0). ...
中国山地植物多样性的垂直变化格局
3
2005
... 揭示和阐明生物多样性空间格局及其维持机制是生态学研究的热点问题(
Cavender-Bares et al ., 2009 ).由于水热条件变化急剧, 山地一直是生态学家研究多样性海拔格局的理想场所(
Jump et al ., 2009 ).受水热条件、生物类群、微地形、土壤质地和人为干扰等因素的影响, 物种多样性表现出不同的海拔格局, 主要为递减格局和单峰格局(
唐志尧等, 2004 ;
朱珣之和张金屯, 2005 ).Ohsawa (
1991 ,
1995 )研究表明亚洲热带山地的物种多样性随海拔升高而降低, 而温带山地的物种多样性随海拔变化不明显.
朱珣之和张金屯(2005) 总结了我国不同气候带的19座山峰物种多样性海拔格局, 发现低纬度地区山地的物种多样性主要表现出单峰格局, 而高纬度地区山地的物种多样性主要以递减格局为主.
Begon等(1996) 认为物种多样性随海拔增加而递减的格局主要受热量因素影响, 随着海拔升高, 温度的快速下降导致多样性的下降.
Whittaker和Niering(1975) 认为物种多样性的单峰格局受水热条件的综合影响, 低海拔地区限制群落组成和多样性的关键因子主要是降水, 而在高海拔地区主要是温度. ...
... )研究表明亚洲热带山地的物种多样性随海拔升高而降低, 而温带山地的物种多样性随海拔变化不明显.
朱珣之和张金屯(2005) 总结了我国不同气候带的19座山峰物种多样性海拔格局, 发现低纬度地区山地的物种多样性主要表现出单峰格局, 而高纬度地区山地的物种多样性主要以递减格局为主.
Begon等(1996) 认为物种多样性随海拔增加而递减的格局主要受热量因素影响, 随着海拔升高, 温度的快速下降导致多样性的下降.
Whittaker和Niering(1975) 认为物种多样性的单峰格局受水热条件的综合影响, 低海拔地区限制群落组成和多样性的关键因子主要是降水, 而在高海拔地区主要是温度. ...
... 随着海拔的升高,
S 、
PD 和
FRic 整体上表现出上升的趋势, 特别是在海拔1 800 m以上(
图2 ).这主要与我们研究的海拔范围(1 409-2 150 m)有关, 在中低海拔区段, 降水是影响该区域多样性最主要的因素(
赵小娜等, 2014 ).随着海拔的升高, 降水量逐渐增加,
S 、
PD 和
FRic 整体上呈现上升的趋势.不同气候带下, 山地α多样性海拔格局的差异主要与气候类型、地形地貌、植被类型和人为活动等因素密切相关, 但水热条件是影响多样性海拔格局的最主要因素(
朱珣之和张金屯, 2005 ).在水分条件较好的区域, 随着海拔升高, 温度快速下降是导致多样性降低的主要原因; 而受水分条件限制的区域, 水热条件的制约和平衡是影响多样性海拔格局的关键.在低海拔区域, 降水是影响群落组成和多样性的关键因子; 而在高海拔区域, 低温成为限制群落组成和多样性的关键因子(
唐志尧等, 2004 ;
Zhang et al ., 2015 ).随着海拔的升高, 群落的
FEve 和
E 指数表现出单峰格局, 最大值大致出现在海拔1 800 m处.该海拔段是温性林向寒温性林逐渐过渡的区域, 群落组成兼有耐旱性和耐寒性物种, 表现出较高的均匀性.随着海拔的升高,
NRI 是逐渐下降的, 群落谱系结构由聚集向随机或者发散过渡.在低海拔区域, 环境过滤(较少的降水量和较强的人为干扰)导致群落内亲缘关系较近的物种聚集在一起, 表现出较强的谱系聚集.而在海拔相对较高的区域, 环境过滤和种间关系可能共同作用于群落组成, 进而表现出随机或者发散的谱系结构.
NRI 指数结果表明群落内木本植物大多为亲缘关系较近的物种(
NRI > 0), 而草本植物大多为亲缘关系较远的物种(
NRI < 0). ...
中国山地植物多样性的垂直变化格局
3
2005
... 揭示和阐明生物多样性空间格局及其维持机制是生态学研究的热点问题(
Cavender-Bares et al ., 2009 ).由于水热条件变化急剧, 山地一直是生态学家研究多样性海拔格局的理想场所(
Jump et al ., 2009 ).受水热条件、生物类群、微地形、土壤质地和人为干扰等因素的影响, 物种多样性表现出不同的海拔格局, 主要为递减格局和单峰格局(
唐志尧等, 2004 ;
朱珣之和张金屯, 2005 ).Ohsawa (
1991 ,
1995 )研究表明亚洲热带山地的物种多样性随海拔升高而降低, 而温带山地的物种多样性随海拔变化不明显.
朱珣之和张金屯(2005) 总结了我国不同气候带的19座山峰物种多样性海拔格局, 发现低纬度地区山地的物种多样性主要表现出单峰格局, 而高纬度地区山地的物种多样性主要以递减格局为主.
Begon等(1996) 认为物种多样性随海拔增加而递减的格局主要受热量因素影响, 随着海拔升高, 温度的快速下降导致多样性的下降.
Whittaker和Niering(1975) 认为物种多样性的单峰格局受水热条件的综合影响, 低海拔地区限制群落组成和多样性的关键因子主要是降水, 而在高海拔地区主要是温度. ...
... )研究表明亚洲热带山地的物种多样性随海拔升高而降低, 而温带山地的物种多样性随海拔变化不明显.
朱珣之和张金屯(2005) 总结了我国不同气候带的19座山峰物种多样性海拔格局, 发现低纬度地区山地的物种多样性主要表现出单峰格局, 而高纬度地区山地的物种多样性主要以递减格局为主.
Begon等(1996) 认为物种多样性随海拔增加而递减的格局主要受热量因素影响, 随着海拔升高, 温度的快速下降导致多样性的下降.
Whittaker和Niering(1975) 认为物种多样性的单峰格局受水热条件的综合影响, 低海拔地区限制群落组成和多样性的关键因子主要是降水, 而在高海拔地区主要是温度. ...
... 随着海拔的升高,
S 、
PD 和
FRic 整体上表现出上升的趋势, 特别是在海拔1 800 m以上(
图2 ).这主要与我们研究的海拔范围(1 409-2 150 m)有关, 在中低海拔区段, 降水是影响该区域多样性最主要的因素(
赵小娜等, 2014 ).随着海拔的升高, 降水量逐渐增加,
S 、
PD 和
FRic 整体上呈现上升的趋势.不同气候带下, 山地α多样性海拔格局的差异主要与气候类型、地形地貌、植被类型和人为活动等因素密切相关, 但水热条件是影响多样性海拔格局的最主要因素(
朱珣之和张金屯, 2005 ).在水分条件较好的区域, 随着海拔升高, 温度快速下降是导致多样性降低的主要原因; 而受水分条件限制的区域, 水热条件的制约和平衡是影响多样性海拔格局的关键.在低海拔区域, 降水是影响群落组成和多样性的关键因子; 而在高海拔区域, 低温成为限制群落组成和多样性的关键因子(
唐志尧等, 2004 ;
Zhang et al ., 2015 ).随着海拔的升高, 群落的
FEve 和
E 指数表现出单峰格局, 最大值大致出现在海拔1 800 m处.该海拔段是温性林向寒温性林逐渐过渡的区域, 群落组成兼有耐旱性和耐寒性物种, 表现出较高的均匀性.随着海拔的升高,
NRI 是逐渐下降的, 群落谱系结构由聚集向随机或者发散过渡.在低海拔区域, 环境过滤(较少的降水量和较强的人为干扰)导致群落内亲缘关系较近的物种聚集在一起, 表现出较强的谱系聚集.而在海拔相对较高的区域, 环境过滤和种间关系可能共同作用于群落组成, 进而表现出随机或者发散的谱系结构.
NRI 指数结果表明群落内木本植物大多为亲缘关系较近的物种(
NRI > 0), 而草本植物大多为亲缘关系较远的物种(
NRI < 0). ...
 ,2,*
,2,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT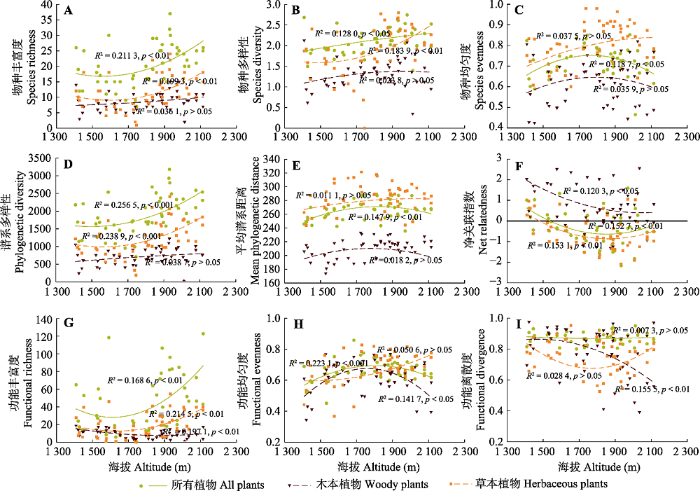 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT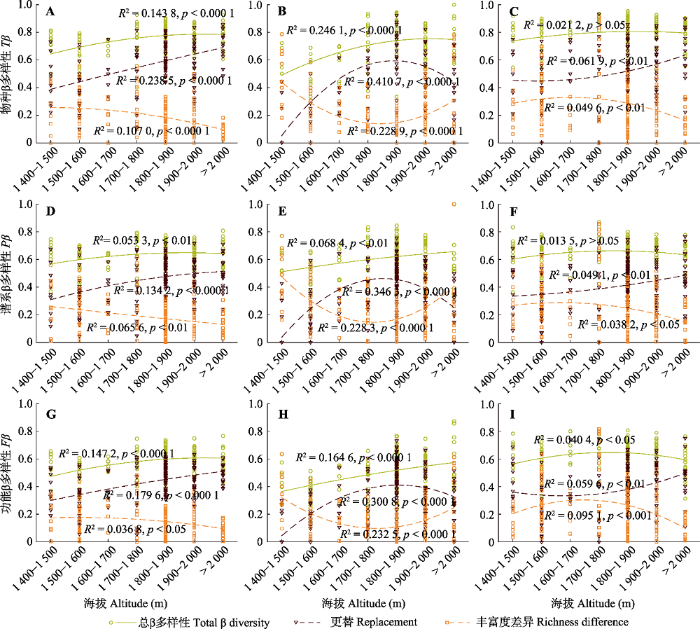 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT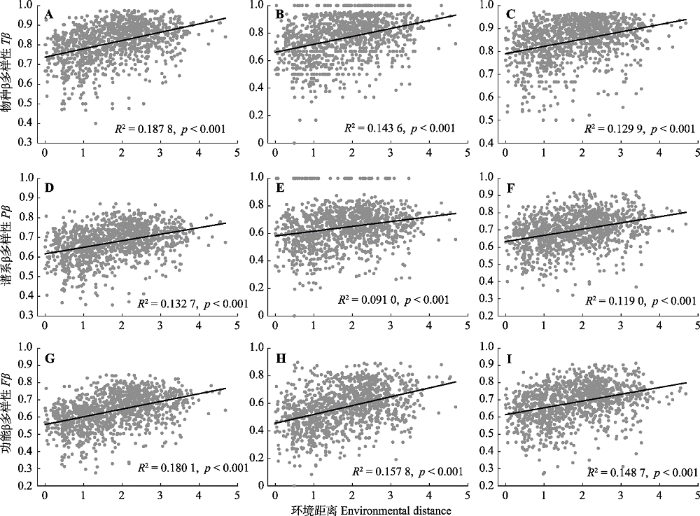 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT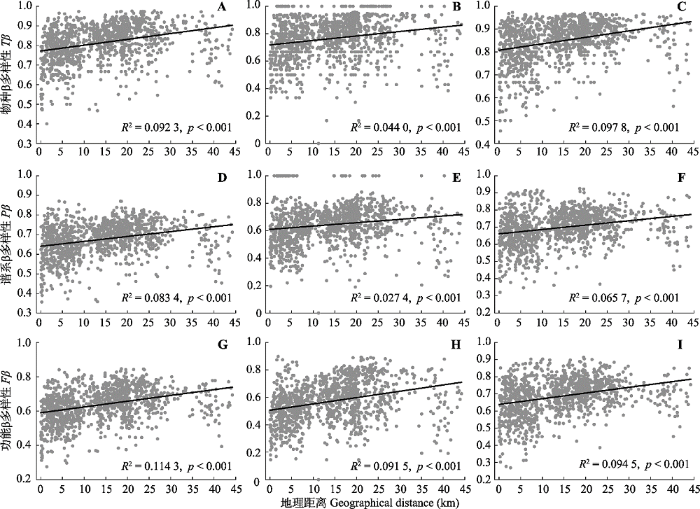 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT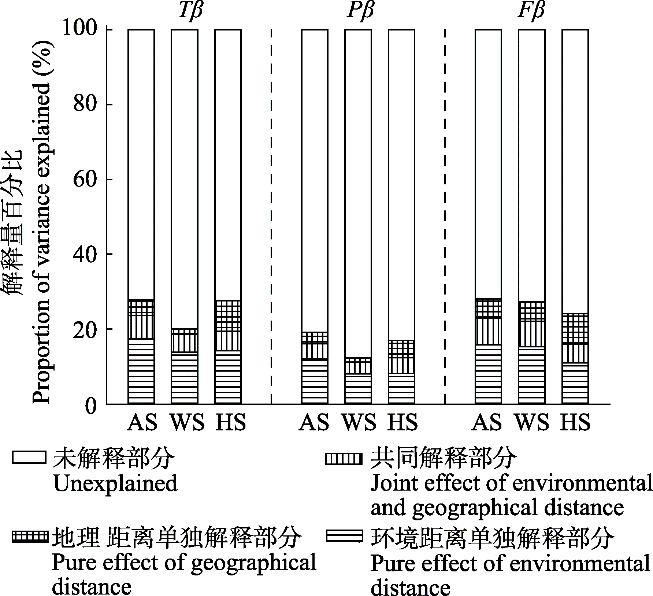 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}