Supported by the National Natural Science Foundation of China(31901293) the National Key Technology Research and Development Program of China(2011BAD37B01) the Program for Changjiang Scholars and Innovative Research Team of Ministry of Education of China(IRT_15R09) the Young Elite Scientists Sponsorship Program by China Association for Science and Technology(2018QNRC001)
Abstract Aims Altitude-induced changes in temperature, moisture, vegetation types and other conditions would significantly affect soil carbon (Csoil), nitrogen (Nsoil), phosphorus (Psoil) concentrations and their stoichiometry. How soil microorganisms adapt to the variability of soil resource stoichiometry by regulating their biomass and extracellular enzymatic stoichiometry remains uncertain. The objective of this study was to quantify the altitudinal trends of soil-microbe-exoenzyme C:N:P stoichiometry and to explore the correlations among soil-microbe- exoenzyme stoichiometry.Methods In the present study, we investigated the Csoil, Nsoil, Psoil concentrations, microbial biomass C (Cmic), N (Nmic), P (Pmic) concentrations, and the activities of C (β-1,4-glucosidase, BG), N (N-acetyl-β-glucosaminidase, NAG), and P (acid phosphatase) acquiring extracellular enzymes for microorganisms in four ecosystems along an altitudinal gradient on Mt. Datudingzi, Northeast China. These four ecosystems are a mixed broadleaf-coniferous forest at 800 m, a coniferous forest at 1 100 m, a Betula ermanii forest at 1 600 m and a grassland at 1 700 m.Important findings The results showed that: (1) altitude had no significant effect on Csoil and Cmic concentrations but had significant effects on soil and microbial biomass N and P concentrations. (2) The activities of BG and NAG decreased significantly with increasing altitude, likely due to the high elevation induced low temperature that inhibits microbial activities. (3) Altitude had significant effects on soil C:N, microbe C:N:P, and exoenzyme C:N:P; exoenzyme C:N:P decreased with the increasing stoichiometric imbalances between microorganisms and soils (ratios of soil C:N:P to microbe C:N:P, respectively). Overall, these results suggested that microorganisms can adapt to the variability of soil C:N:P by regulating their biomass C:N:P and exoenzyme C:N:P, and supported the microbial resource allocation theory. Keywords:stoichiometry;microbial activity;C:N:P;altitude;extracellular enzyme
PDF (1501KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 殷爽, 王传宽, 金鹰, 周正虎. 东北地区大秃顶子山土壤-微生物-胞外酶C:N:P 化学计量特征沿海拔梯度的变化. 植物生态学报, 2019, 43(11): 999-1009. DOI: 10.17521/cjpe.2019.0141 YIN Shuang, WANG Chuan-Kuan, JIN Ying, ZHOU Zheng-Hu. Changes in soil-microbe-exoenzyme C:N:P stoichiometry along an altitudinal gradient in Mt. Datudingzi, Northeast China. Chinese Journal of Plant Ecology, 2019, 43(11): 999-1009. DOI: 10.17521/cjpe.2019.0141
土壤微生物是陆地生态系统的重要组分, 驱动着生态系统的物质循环和能量流动, 如有机质分解、有机碳固持、氮循环等(Leff et al., 2015), 影响生态系统的功能(Chapin et al., 2003; Mooshammer et al., 2014)。海拔梯度变化会引起小气候和植被的变化, 改变土壤微生物群落结构和活性(Fierer et al., 2011)。温度是限制微生物生长和活性的重要生态因子(Margesin et al., 2009; Looby & Treseder, 2018)。随海拔的升高, 温度逐渐降低, 这可能会降低微生物生物量, 抑制微生物的活性。许多研究表明土壤微生物生物量沿海拔的分布格局不相一致。例如, 赵盼盼等(2019a)在中亚热带戴云山不同海拔高度的台湾松(Pinus taiwanensis)林中的研究发现土壤微生物生物量碳(C)、氮(N)、磷(P)含量随着海拔升高均呈现降低的趋势; 曹瑞等(2016)在川西发现随着海拔的不断升高, 土壤微生物生物量C、N含量呈现先增加后减少再增加的趋势。不同研究中土壤微生物生物量的不同垂直分布格局可能是由于调控土壤微生物生物量的主导因子不同所导致。赵盼盼等(2019a)研究认为温度是影响不同海拔土壤微生物生物量分布的主要因素, 曹瑞等(2016)研究认为土壤微生物生物量沿海拔分布格局的主要调控因子是土壤有机碳、全氮含量和土壤含水量。可见, 土壤微生物生物量沿着海拔的垂直分布格局及其潜在驱动机制还有待进一步深入研究。
微生物分解者只有通过产生胞外酶将环境中复杂有机质分解成可溶性小分子物质才能被自身吸收利用(Schimel & Bennett, 2004), 微生物胞外酶活性的大小直接影响土壤C、N、P循环过程。例如: Morrissey等(2014)研究表明土壤C矿化速率与β-1,4-葡萄糖苷酶(BG)活性正相关。Chen等(2017)整合分析发现C获取相关的胞外酶对N添加的响应与土壤呼吸对N添加的响应之间存在显著正相关关系, 也就是说N添加会通过影响胞外酶活性来影响土壤呼吸, 最终影响土壤C储量。还有研究发现土壤磷酸酶活性与土壤有效磷含量之间呈负相关关系(Allison et al., 2007)。通过量化胞外酶活性有助于理解土壤的生物地球化学循环过程(Sinsabaugh et al., 2009; Buchkowski et al., 2015)。目前, 生态化学计量学相关理论已经被应用到微生物胞外酶研究中, 探索微生物胞外酶C:N:P对不同土壤C:N:P的适应机制。Xu等(2017)分析我国东部南北样带森林生态系统土壤酶活性及其化学计量比时发现, 土壤胞外酶C:P和N:P分别与土壤C:P和土壤N:P呈显著负相关关系。Peng和Wang (2016)研究发现土壤C、N、P是导致土壤胞外酶及其化学计量变异的主要因素。海拔梯度引起气候、植被、土壤特征的变化会进一步导致土壤C、N、P化学计量特征的变化(李相楹等, 2016), 然而土壤微生物是否会通过调整自身生物量C:N:P和胞外酶C:N:P进行适应仍不明确。通过研究微生物生物量和胞外酶C:N:P的环境可塑性反过来有助于我们对土壤C、N、P循环过程的理解。
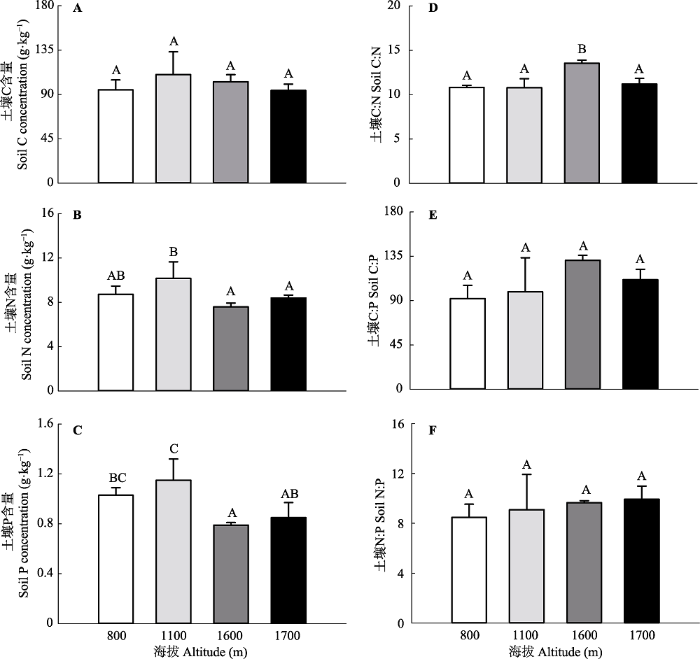
新窗口打开|下载原图ZIP|生成PPT 图1大秃顶子山不同海拔土壤碳(C)、氮(N)、磷(P)含量及其化学计量比(平均值+标准误差, n = 3)。不同大写字母表示不同海拔间差异显著(p < 0.05)。
Fig. 1Soil carbon (C), nitrogen (N), and phosphorus (P) concentrations and their stoichiometric ratios under different altitudes in Mt. Datudingzi (mean + SE, n = 3). Different uppercase letters represent significant differences at 0.05 level among different altitudes.
新窗口打开|下载原图ZIP|生成PPT 图2大秃顶子山不同海拔土壤微生物生物量碳(C)、氮(N)、磷(P)含量及其化学计量比(平均值+标准误差, n = 3)。不同大写字母表示不同海拔间差异显著(p < 0.05)。
Fig. 2Microbial biomass carbon (C), nitrogen (N), and phosphorus (P) concentrations and their stoichiometric ratios under different altitudes in Mt. Datudingzi (mean + SE, n = 3). Different uppercase letters represent significant differences at 0.05 level among different altitudes.
Fig. 3Exoenzyme carbon (C), nitrogen (N), and phosphorus (P) activities and their stoichiometric ratios under different altitudes in Mt. Datudingzi (mean + SE, n = 3). Different uppercase letters represent significant differences at 0.05 level among different altitudes. AP, acid phosphomonoesterase; BG, β-1,4-glucosidase; NAG, N-acetyl-β- glucosaminidase.
2.4 土壤-微生物-胞外酶之间碳氮磷化学计量特征间的相关性
胞外酶化学计量比受土壤和微生物生物量化学计量比影响(图4)。回归分析显示胞外酶C:N与土壤C:N无显著相关性(p > 0.05)(图4A), 与微生物生物量C:N正相关(R2 = 0.36, p = 0.038, 图4D)。胞外酶N:P与土壤N:P负相关(R2 = 0.35, p = 0.045, 图4C), 但与微生物生物量N:P无显著相关性(p > 0.05)(图4F)。胞外酶C:P与土壤C:P和微生物生物量C:P相关性均不显著(p > 0.05)(图4B、4E)。尽管仅发现胞外酶C:P和C:Pimb (R2= 0.37, p=0.036)以及胞外酶N:P和N:Pimb间(R2 = 0.53, p = 0.007)显著相关(图4H、4I), 胞外酶C:N、C:P、N:P分别随着C:Nimb、C:Pimb、N:Pimb的增加均呈现出减小的趋势(图4G、4H、4I)。
Fig. 5Relationship between mean annual temperature and the slope of linear relationship between soil C storage and altitude. The data is from the global meta-analysis of Tashi et al. (2016). The open triangle is the slope from the current study. The open circle is the outlier. The slope stands for the change in soil C storage (kg·m-2) per km increase in elevation.
AllisonSD, VitousekPM ( 2005). Responses of extracellular enzymes to simple and complex nutrient inputs Soil Biology & Biochemistry, 37, 937-944. [本文引用: 1]
AllisonSD, WeintraubMN, GartnerTB, WaldropMP ( 2010). Evolutionary-economic principles as regulators of soil enzyme production and ecosystem function In: Shukla G, Varma A eds. Soil Enzymology. Springer, Berlin. 229-243. [本文引用: 1]
AllisonVJ, CondronLM, PeltzerDA, RichardsonSJ, TurnerBL ( 2007). Changes in enzyme activities and soil microbial community composition along carbon and nutrient gradients at the Franz Josef chronosequence, New Zealand Soil Biology & Biochemistry, 39, 1770-1781. [本文引用: 1]
BrookesPC, LandmanA, PrudenG, JenkinsonDS ( 1985). Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil Soil Biology & Biochemistry, 17, 837-842. [本文引用: 1]
BuchkowskiRW, SchmitzOJ, BradfordMA ( 2015). Microbial stoichiometry overrides biomass as a regulator of soil carbon and nitrogen cycling Ecology, 96, 1139-1149. [本文引用: 2]
CaoR, WuFZ, YangWQ, XuZF, TanB, WangB, LiJ, ChangCH ( 2016). Effects of altitudes on soil microbial biomass and enzyme activity in alpine-gorge regions Chinese Journal of Applied Ecology, 27, 1257-1264. [本文引用: 2]
ChapinCT, BridghamSD, PastorJ, UpdegraffK ( 2003). Nitrogen, phosphorus and carbon mineralization in response to nutrient and lime additions in peatlands Soil Science, 168, 409-420. [本文引用: 1]
ChenJ, LuoYQ, LIJW, ZhouXH, CaoJJ, WangRW, WangYQ, SheltonS, JinZ, WalkerLM, FengZZ, NiuSL, FengWT, JianSY, ZhouLY ( 2017). Costimulation of soil glycosidase activity and soil respiration by nitrogen addition Global Change Biology, 23, 1328-1337. [本文引用: 1]
ClevelandCC, LiptzinD ( 2007). C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass? Biogeochemistry, 85, 235-252. [本文引用: 1]
DavidsonECA, BelkE, BooneRD ( 1998). Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest Global Change Biology, 4, 217-227. [本文引用: 1]
EnsslinA, RuttenG, PommerU, ZimmermannR, HempA, FischerM ( 2015). Effects of elevation and land use on the biomass of trees, shrubs and herbs at Mount Kilimanjaro Ecosphere, 6, art45. DOI: 10.1890/ES14-00492.1. [本文引用: 1]
FiererN, MccainCM, MeirP, ZimmermannM, RappJM, SilmanMR, KnightR ( 2011). Microbes do not follow the elevational diversity patterns of plants and animals Ecology, 92, 797-804. [本文引用: 1]
Garten JrCT, HansonPJ ( 2006). Measured forest soil C stocks and estimated turnover times along an elevation gradient Geoderma, 136, 342-352. [本文引用: 2]
GuXN, HeHS, TaoY, JinYH, ZhangXY, XuZW, WangYT, SongXX ( 2017). Soil microbial community structure, enzyme activities, and their influencing factors along different altitudes of Changbai Mountain Acta Ecologica Sinica, 37, 8374-8384. [本文引用: 2]
HofmannK, LamprechtA, PauliH, IllmerP ( 2016). Distribution of prokaryotic abundance and microbial nutrient cycling across a high-alpine altitudinal gradient in the Austrian Central Alps is affected by vegetation, temperature, and soil nutrients Microbial Ecology, 72, 704-716. [本文引用: 3]
HuZD, LiuSR, ShiZM, LiuXL, HeF ( 2012). Variations of soil nitrogen and microbial biomass carbon and nitrogen of Quercus aquifolioides forest at different altitudes in Balangshan, Sichuan Forest Research, 25, 261-268. [本文引用: 1]
KangHZ, ZhuangHL, WuLL, LiuQL, ShenGR, BergB, ManRZ, LiuCJ ( 2011). Variation in leaf nitrogen and phosphorus stoichiometry in Picea abies across Europe: An analysis based on local observations Forest Ecology & Management, 261,195-202. [本文引用: 1]
LeffJW, JonesSE, ProberSM, BarberánA, BorerET, FirnJL, HarpoleWS, HobbieSE, HofmockelKS, KnopsJMH, McCulleyRL, La PierreK, RischAC, SeabloomEW, SchützM, SteenbockC, StevensCJ, FiererN ( 2015). Consistent responses of soil microbial communities to elevated nutrient inputs in grasslands across the globe Proceedings of the National Academy of Sciences of the United States of America, 112, 10967-10972. [本文引用: 1]
LiHJ, LiuJW, YangL, ZhengHF, LiuY, YangWQ, ZhangJ ( 2016). Effects of simulated climate warming on soil microbial biomass carbon, nitrogen and phosphorus of alpine forest Chinese Journal of Applied and Environmental Biology, 22, 599-605. [本文引用: 1]
LiXY, ZhangWY, LiuF, ZhangZM, HeTB, LinCH ( 2016). The distribution characteristics of soil carbon, nitrogen and phosphorus at different altitudes in Fanjingshan Mountain Research of Soil & Water Conservation, 23(3), 19-24. [本文引用: 1]
LiY, WuJS, LiuSL, ShenJL, HuangDY, SuYR, WeiWX, SyersJK ( 2012). Is the C:N:P stoichiometry in soil and soil microbial biomass related to the landscape and land use in southern subtropical China? Global Biogeochemical Cycles, 26, GB4002. DOI: 10.1029/2012GB004399. [本文引用: 1]
LiuBR ( 2010). Changes in soil microbial biomass carbon and nitrogen under typical plant communities along an altitudinal gradient in east side of Helan Mountain Ecology and Environmental Sciences, 19, 883-888. [本文引用: 2]
LoobyCI, TresederKK ( 2018). Shifts in soil fungi and extracellular enzyme activity with simulated climate change in a tropical montane cloud forest Soil Biology & Biochemistry, 117, 87-96. [本文引用: 1]
MargesinR, JudM, TscherkoD, SchinnerF ( 2009). Microbial communities and activities in alpine and subalpine soils FEMS Microbiology Ecology, 67, 208-218. [本文引用: 1]
MartensDA, JohansonJB, Frankenberger JrWT ( 1992). Production and persistence of soil enzymes with repeated addition of organic residues Soil Science, 153, 53-61. [本文引用: 1]
MooshammerM, WanekW, Zechmeister-BoltensternS, RichterA ( 2014). Stoichiometric imbalances between terrestrial decomposer communities and their resources: Mechanisms and implications of microbial adaptations to their resources Frontiers in Microbiology, 5, 22. DOI: 10.3389/?fmicb.2014.00022. [本文引用: 3]
MorrisseyEM, BerrierDJ, NeubauerSC, FranklinRB ( 2014). Using microbial communities and extracellular enzymes to link soil organic matter characteristics to greenhouse gas production in a tidal freshwater wetland Biogeochemistry, 117, 473-490. [本文引用: 1]
PengXQ, WangW ( 2016). Stoichiometry of soil extracellular enzyme activity along a climatic transect in temperate grasslands of northern China Soil Biology & Biochemistry, 98, 74-84. [本文引用: 2]
SchimelJP, BennettJ ( 2004). Nitrogen mineralization: Challenges of a changing paradigm Ecology, 85, 591-602. [本文引用: 1]
SinsabaughRL, HillBH, Follstad ShahJJ ( 2009). Ecoenzymatic stoichiometry of microbial organic nutrient acquisition in soil and sediment Nature, 462, 795-798. [本文引用: 2]
?tursováM, BaldrianP ( 2011). Effects of soil properties and management on the activity of soil organic matter transforming enzymes and the quantification of soil-bound and free activity Plant and Soil, 338, 99-110. [本文引用: 1]
TashiS, SinghB, KeitelC, AdamsM ( 2016). Soil carbon and nitrogen stocks in forests along an altitudinal gradient in the eastern Himalayas and a meta-analysis of global data Global Change Biology, 22, 2255-2268. [本文引用: 5]
WalleniusK, RitaH, MikkonenA, LappiK, Lindstr?mK, HartikainenH, RaatelandA, NiemiRM ( 2011). Effects of land use on the level, variation and spatial structure of soil enzyme activities and bacterial communities Soil Biology & Biochemistry, 43, 1464-1473. [本文引用: 1]
WangXC ( 2004). Response of Subalpine Timberline in Northeast China to Global Climate Change PhD dissertation. Northeast Forestry University, Harbin. [本文引用: 1]
WardleDA ( 1992). A comparative assessment of factors which influence microbial biomass carbon and nitrogen levels in soil Biological Reviews, 67, 321-358. [本文引用: 1]
WaringBG, WeintraubSR, SinsabaughRL ( 2014). Ecoenzymatic stoichiometry of microbial nutrient acquisition in tropical soils Biogeochemistry, 117, 101-113. [本文引用: 1]
WeiJ, WuG, WangH, HaoYJ, ShangWY ( 2005). Phosphorus and sulphur bio-cycling in alpine tundra ecosystem of Changbai Mountains Chinese Journal of Applied Ecology, 16, 1230-1234. [本文引用: 1]
XuQF, JiangPK, ShenQ ( 2005). Comparison of organic carbon pool of soil in bush and broad-leaved forests Journal of Beijing Forestry University, 27(2), 18-22. [本文引用: 2]
XuXF, ThorntonPE, PostWM ( 2013). A global analysis of soil microbial biomass carbon, nitrogen and phosphorus in terrestrial ecosystems Global Ecology and Biogeography, 22, 737-749. [本文引用: 1]
XuZW, YuGR, ZhangXY, GeJP, HeNP, WangQF, WangD ( 2015). The variations in soil microbial communities, enzyme activities and their relationships with soil organic matter decomposition along the northern slope of Changbai Mountain Applied Soil Ecology, 86, 19-29. [本文引用: 1]
XuZW, YuGR, ZhangXY, HeNP, WangQF, WangSZ, WangRL, ZhaoN, JiaYL, WangCY ( 2017). Soil enzyme activity and stoichiometry in forest ecosystems along the North-South Transect in Eastern China (NSTEC) Soil Biology & Biochemistry, 104, 152-163. [本文引用: 2]
ZhangP, ZhangGQ, ZhaoYP, PengSZ, ChenYM, CaoY ( 2018). Ecological stoichiometry characteristics of leaf-litter-soil interactions in different forest types in the Loess hilly-gully region of China Acta Ecologica Sinica, 38, 5087-5098. [本文引用: 1]
ZhaoPP, ZhouJC, LinKM, ZhangQF, YuanP, ZengXM, SuY, XuJG, ChenYM, YangYS ( 2019b). Effect of different altitudes on soil microbial biomass and community structure of Pinus taiwanensis forest in mid-subtropical zone Acta Ecologica Sinica, 39, 2215-2225. [本文引用: 1]
ZhengMH, ZhouZH, LuoYQ, ZhaoP, MoJM ( 2019). Global pattern and controls of biological nitrogen fixation under nutrient enrichment: A meta-analysis Global Change Biology, 25, 3018-3030. [本文引用: 1]
ZhouZH, WangCK ( 2017). Soil-microbe-mineralization carbon and nitrogen stoichiometry under different land-uses in the Maoershan region Acta Ecologica Sinica, 37, 2428-2436. [本文引用: 3]
ZhouZH, WangCK, JinY ( 2017). Stoichiometric responses of soil microflora to nutrient additions for two temperate forest soils Biology and Fertility of Soils, 53, 397-406.
Responses of extracellular enzymes to simple and complex nutrient inputs 1 2005
Changes in enzyme activities and soil microbial community composition along carbon and nutrient gradients at the Franz Josef chronosequence, New Zealand 1 2007
... 微生物分解者只有通过产生胞外酶将环境中复杂有机质分解成可溶性小分子物质才能被自身吸收利用(Schimel & Bennett, 2004), 微生物胞外酶活性的大小直接影响土壤C、N、P循环过程.例如: Morrissey等(2014)研究表明土壤C矿化速率与β-1,4-葡萄糖苷酶(BG)活性正相关.Chen等(2017)整合分析发现C获取相关的胞外酶对N添加的响应与土壤呼吸对N添加的响应之间存在显著正相关关系, 也就是说N添加会通过影响胞外酶活性来影响土壤呼吸, 最终影响土壤C储量.还有研究发现土壤磷酸酶活性与土壤有效磷含量之间呈负相关关系(Allison et al., 2007).通过量化胞外酶活性有助于理解土壤的生物地球化学循环过程(Sinsabaugh et al., 2009; Buchkowski et al., 2015).目前, 生态化学计量学相关理论已经被应用到微生物胞外酶研究中, 探索微生物胞外酶C:N:P对不同土壤C:N:P的适应机制.Xu等(2017)分析我国东部南北样带森林生态系统土壤酶活性及其化学计量比时发现, 土壤胞外酶C:P和N:P分别与土壤C:P和土壤N:P呈显著负相关关系.Peng和Wang (2016)研究发现土壤C、N、P是导致土壤胞外酶及其化学计量变异的主要因素.海拔梯度引起气候、植被、土壤特征的变化会进一步导致土壤C、N、P化学计量特征的变化(李相楹等, 2016), 然而土壤微生物是否会通过调整自身生物量C:N:P和胞外酶C:N:P进行适应仍不明确.通过研究微生物生物量和胞外酶C:N:P的环境可塑性反过来有助于我们对土壤C、N、P循环过程的理解. ...
Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil 1 1985
... 微生物生物量C、N、P含量的测定采用氯仿熏蒸浸提法(Brookes et al., 1985).称取新鲜土壤样品5 g置于100 mL小烧杯中, 将小烧杯放于真空干燥器中, 并在干燥器中加入NaOH溶液和氯仿, 25 ℃真空条件下处理24 h (熏蒸组).培养结束后用0.5 mol·L-1 K2SO4 (土液比为1:2.5)浸提C、N, 或用0.5 mol·L-1 NaHCO3溶液浸提P.对照组不熏蒸, 浸提过程同熏蒸组.用multi N/C 3000分析仪测定浸提液的C、N浓度, 用钼蓝比色法测定浸提液的P浓度.然后用熏蒸组测定元素浓度与未熏蒸组测定元素浓度的差值乘以相应的转换系数来计算微生物生物量C、N、P含量.其中C、N和P转换系数分别为0.45、0.45和0.40 (Jenkinson et al., 2004). ...
Microbial stoichiometry overrides biomass as a regulator of soil carbon and nitrogen cycling 2 2015
... 微生物分解者只有通过产生胞外酶将环境中复杂有机质分解成可溶性小分子物质才能被自身吸收利用(Schimel & Bennett, 2004), 微生物胞外酶活性的大小直接影响土壤C、N、P循环过程.例如: Morrissey等(2014)研究表明土壤C矿化速率与β-1,4-葡萄糖苷酶(BG)活性正相关.Chen等(2017)整合分析发现C获取相关的胞外酶对N添加的响应与土壤呼吸对N添加的响应之间存在显著正相关关系, 也就是说N添加会通过影响胞外酶活性来影响土壤呼吸, 最终影响土壤C储量.还有研究发现土壤磷酸酶活性与土壤有效磷含量之间呈负相关关系(Allison et al., 2007).通过量化胞外酶活性有助于理解土壤的生物地球化学循环过程(Sinsabaugh et al., 2009; Buchkowski et al., 2015).目前, 生态化学计量学相关理论已经被应用到微生物胞外酶研究中, 探索微生物胞外酶C:N:P对不同土壤C:N:P的适应机制.Xu等(2017)分析我国东部南北样带森林生态系统土壤酶活性及其化学计量比时发现, 土壤胞外酶C:P和N:P分别与土壤C:P和土壤N:P呈显著负相关关系.Peng和Wang (2016)研究发现土壤C、N、P是导致土壤胞外酶及其化学计量变异的主要因素.海拔梯度引起气候、植被、土壤特征的变化会进一步导致土壤C、N、P化学计量特征的变化(李相楹等, 2016), 然而土壤微生物是否会通过调整自身生物量C:N:P和胞外酶C:N:P进行适应仍不明确.通过研究微生物生物量和胞外酶C:N:P的环境可塑性反过来有助于我们对土壤C、N、P循环过程的理解. ...
... 尽管胞外酶C:N与土壤C:N之间不存在显著相关性, 本研究却发现胞外酶C:N与微生物C:N之间存在显著正相关关系(p < 0.05)(图4).此外, 胞外酶C:P和N:P也分别和微生物C:P和N:P存在正相关趋势.这些结果可能意味着微生物生物量C、N、P之间的相对比例大小与微生物对C、N、P 3种元素的获取能力相关.长久以来, 微生物生物量C:N:P的环境可塑性得到了广泛关注(Cleveland & Liptzin, 2007; Li et al., 2012; Xu et al., 2013), 而现在关注更多的是微生物生物量C:N:P对土壤功能过程的影响.周正虎和王传宽(2017)对帽儿山地区不同土地利用方式下土壤-微生物-矿化碳氮化学计量特征的研究中发现单位微生物生物量的碳矿化速率随着微生物C:N增加而降低, 单位微生物生物量的氮矿化速率随着微生物C:N的增加而增加.Buchkowski等(2015)甚至发现微生物生物量C:N:P对C、N矿化速率的影响程度和微生物生物量对C、N矿化速率的影响程度相当.微生物与其土壤资源之间的化学计量不平衡性综合考虑了土壤和微生物两者C:N:P的变异性, 能够更加准确地来衡量微生物化学组成与资源化学组成的差异性.本研究也发现胞外酶C:N:P和C:N:P化学计量不平衡性间存在负相关关系, 进一步证明了土壤与微生物之间C、N、P的差异会对胞外酶化学计量比产生影响(Mooshammer et al., 2014).前期研究已经证实土壤和微生物间的C:N化学计量不平衡性会对土壤C、N矿化速率产生影响(周正虎和王传宽, 2017), 综合考虑土壤和微生物C:N:P的变异性有助于我们对土壤C、N、P循环的理解. ...
海拔对高山峡谷区土壤微生物生物量和酶活性的影响 2 2016
... 土壤微生物是陆地生态系统的重要组分, 驱动着生态系统的物质循环和能量流动, 如有机质分解、有机碳固持、氮循环等(Leff et al., 2015), 影响生态系统的功能(Chapin et al., 2003; Mooshammer et al., 2014).海拔梯度变化会引起小气候和植被的变化, 改变土壤微生物群落结构和活性(Fierer et al., 2011).温度是限制微生物生长和活性的重要生态因子(Margesin et al., 2009; Looby & Treseder, 2018).随海拔的升高, 温度逐渐降低, 这可能会降低微生物生物量, 抑制微生物的活性.许多研究表明土壤微生物生物量沿海拔的分布格局不相一致.例如, 赵盼盼等(2019a)在中亚热带戴云山不同海拔高度的台湾松(Pinus taiwanensis)林中的研究发现土壤微生物生物量碳(C)、氮(N)、磷(P)含量随着海拔升高均呈现降低的趋势; 曹瑞等(2016)在川西发现随着海拔的不断升高, 土壤微生物生物量C、N含量呈现先增加后减少再增加的趋势.不同研究中土壤微生物生物量的不同垂直分布格局可能是由于调控土壤微生物生物量的主导因子不同所导致.赵盼盼等(2019a)研究认为温度是影响不同海拔土壤微生物生物量分布的主要因素, 曹瑞等(2016)研究认为土壤微生物生物量沿海拔分布格局的主要调控因子是土壤有机碳、全氮含量和土壤含水量.可见, 土壤微生物生物量沿着海拔的垂直分布格局及其潜在驱动机制还有待进一步深入研究. ...
... ; 谷晓楠等, 2017).为了阐明不同研究间土壤C沿海拔分布格局的差异性, 我们重新分析了Tashi等(2016)的全球数据.结果发现, 土壤C与海拔之间线性关系的斜率会随着最低海拔处的年平均气温的增加而显著增加(R2 = 0.26, p = 0.021, 图5), 说明温暖地区土壤C含量随着海拔升高的增加速度要高于寒冷地区土壤C含量随着海拔升高的增加速度.一般来说, 海拔每增加100 m, 温度会降低0.6 ℃.由于土壤呼吸与土壤温度之间普遍存在着指数函数关系(Davidson et al., 1998), 因此, 寒冷地区相比, 温暖地区相同程度的温度降低(即, 相同程度的海拔增加导致相同程度的温度降低)会导致更大程度的土壤呼吸速率降低, 这就解释了其中土壤C含量与海拔之间线性关系的斜率会随着年平均气温的增加而显著增加(图5).本研究地位于高纬度地区, 年平均气温较低, 海拔升高导致的土壤呼吸速率降低程度也较低, 导致本研究中不同海拔间土壤C含量不存在显著的差异(图1A). ...
Distribution of prokaryotic abundance and microbial nutrient cycling across a high-alpine altitudinal gradient in the Austrian Central Alps is affected by vegetation, temperature, and soil nutrients 3 2016
Shifts in soil fungi and extracellular enzyme activity with simulated climate change in a tropical montane cloud forest 1 2018
... 土壤微生物是陆地生态系统的重要组分, 驱动着生态系统的物质循环和能量流动, 如有机质分解、有机碳固持、氮循环等(Leff et al., 2015), 影响生态系统的功能(Chapin et al., 2003; Mooshammer et al., 2014).海拔梯度变化会引起小气候和植被的变化, 改变土壤微生物群落结构和活性(Fierer et al., 2011).温度是限制微生物生长和活性的重要生态因子(Margesin et al., 2009; Looby & Treseder, 2018).随海拔的升高, 温度逐渐降低, 这可能会降低微生物生物量, 抑制微生物的活性.许多研究表明土壤微生物生物量沿海拔的分布格局不相一致.例如, 赵盼盼等(2019a)在中亚热带戴云山不同海拔高度的台湾松(Pinus taiwanensis)林中的研究发现土壤微生物生物量碳(C)、氮(N)、磷(P)含量随着海拔升高均呈现降低的趋势; 曹瑞等(2016)在川西发现随着海拔的不断升高, 土壤微生物生物量C、N含量呈现先增加后减少再增加的趋势.不同研究中土壤微生物生物量的不同垂直分布格局可能是由于调控土壤微生物生物量的主导因子不同所导致.赵盼盼等(2019a)研究认为温度是影响不同海拔土壤微生物生物量分布的主要因素, 曹瑞等(2016)研究认为土壤微生物生物量沿海拔分布格局的主要调控因子是土壤有机碳、全氮含量和土壤含水量.可见, 土壤微生物生物量沿着海拔的垂直分布格局及其潜在驱动机制还有待进一步深入研究. ...
Microbial communities and activities in alpine and subalpine soils 1 2009
... 土壤微生物是陆地生态系统的重要组分, 驱动着生态系统的物质循环和能量流动, 如有机质分解、有机碳固持、氮循环等(Leff et al., 2015), 影响生态系统的功能(Chapin et al., 2003; Mooshammer et al., 2014).海拔梯度变化会引起小气候和植被的变化, 改变土壤微生物群落结构和活性(Fierer et al., 2011).温度是限制微生物生长和活性的重要生态因子(Margesin et al., 2009; Looby & Treseder, 2018).随海拔的升高, 温度逐渐降低, 这可能会降低微生物生物量, 抑制微生物的活性.许多研究表明土壤微生物生物量沿海拔的分布格局不相一致.例如, 赵盼盼等(2019a)在中亚热带戴云山不同海拔高度的台湾松(Pinus taiwanensis)林中的研究发现土壤微生物生物量碳(C)、氮(N)、磷(P)含量随着海拔升高均呈现降低的趋势; 曹瑞等(2016)在川西发现随着海拔的不断升高, 土壤微生物生物量C、N含量呈现先增加后减少再增加的趋势.不同研究中土壤微生物生物量的不同垂直分布格局可能是由于调控土壤微生物生物量的主导因子不同所导致.赵盼盼等(2019a)研究认为温度是影响不同海拔土壤微生物生物量分布的主要因素, 曹瑞等(2016)研究认为土壤微生物生物量沿海拔分布格局的主要调控因子是土壤有机碳、全氮含量和土壤含水量.可见, 土壤微生物生物量沿着海拔的垂直分布格局及其潜在驱动机制还有待进一步深入研究. ...
Production and persistence of soil enzymes with repeated addition of organic residues 1 1992
... 土壤酶活性主要测定BG、β-1, 4-N-乙酰葡糖氨糖苷酶(NAG)、酸性磷酸酶(AP) 3种水解酶的活性.其中, BG主要是微生物获取C的胞外酶, NAG是微生物获取N的胞外酶, 而AP是微生物获取P的胞外酶(Sinsabaugh et al., 2009; Waring et al., 2014).酶活性的测定参考Martens等(1992)的方法.BG或NAG活性的测定方法为: 称取新鲜土壤样品1 g, 加入4 mL 0.1 mol·L-1醋酸缓冲液(pH = 5.0)和对硝基苯-β-D吡喃葡萄糖苷(BG底物, 0.025 mol·L-1)或β-N-乙酰氨基葡萄糖苷(NAG底物, 0.01 mol·L-1), 振荡混匀, 然后将土壤浆在35 ℃下培养1 h.培养结束后加入1 mL 0.5 mol·L-1 CaCl2和4 mL NaOH, 振荡过滤, 分光光度计410 nm下测量吸光值.AP活性的测定, 缓冲液使用改进的通用缓冲液(pH = 6.0), 底物为对硝基苯磷酸二钠(0.025 mol·L-1), 其他实验条件与程序同BG和NAG的测定方法. ...
Stoichiometric imbalances between terrestrial decomposer communities and their resources: Mechanisms and implications of microbial adaptations to their resources 3 2014
... 土壤微生物是陆地生态系统的重要组分, 驱动着生态系统的物质循环和能量流动, 如有机质分解、有机碳固持、氮循环等(Leff et al., 2015), 影响生态系统的功能(Chapin et al., 2003; Mooshammer et al., 2014).海拔梯度变化会引起小气候和植被的变化, 改变土壤微生物群落结构和活性(Fierer et al., 2011).温度是限制微生物生长和活性的重要生态因子(Margesin et al., 2009; Looby & Treseder, 2018).随海拔的升高, 温度逐渐降低, 这可能会降低微生物生物量, 抑制微生物的活性.许多研究表明土壤微生物生物量沿海拔的分布格局不相一致.例如, 赵盼盼等(2019a)在中亚热带戴云山不同海拔高度的台湾松(Pinus taiwanensis)林中的研究发现土壤微生物生物量碳(C)、氮(N)、磷(P)含量随着海拔升高均呈现降低的趋势; 曹瑞等(2016)在川西发现随着海拔的不断升高, 土壤微生物生物量C、N含量呈现先增加后减少再增加的趋势.不同研究中土壤微生物生物量的不同垂直分布格局可能是由于调控土壤微生物生物量的主导因子不同所导致.赵盼盼等(2019a)研究认为温度是影响不同海拔土壤微生物生物量分布的主要因素, 曹瑞等(2016)研究认为土壤微生物生物量沿海拔分布格局的主要调控因子是土壤有机碳、全氮含量和土壤含水量.可见, 土壤微生物生物量沿着海拔的垂直分布格局及其潜在驱动机制还有待进一步深入研究. ...
... 尽管胞外酶C:N与土壤C:N之间不存在显著相关性, 本研究却发现胞外酶C:N与微生物C:N之间存在显著正相关关系(p < 0.05)(图4).此外, 胞外酶C:P和N:P也分别和微生物C:P和N:P存在正相关趋势.这些结果可能意味着微生物生物量C、N、P之间的相对比例大小与微生物对C、N、P 3种元素的获取能力相关.长久以来, 微生物生物量C:N:P的环境可塑性得到了广泛关注(Cleveland & Liptzin, 2007; Li et al., 2012; Xu et al., 2013), 而现在关注更多的是微生物生物量C:N:P对土壤功能过程的影响.周正虎和王传宽(2017)对帽儿山地区不同土地利用方式下土壤-微生物-矿化碳氮化学计量特征的研究中发现单位微生物生物量的碳矿化速率随着微生物C:N增加而降低, 单位微生物生物量的氮矿化速率随着微生物C:N的增加而增加.Buchkowski等(2015)甚至发现微生物生物量C:N:P对C、N矿化速率的影响程度和微生物生物量对C、N矿化速率的影响程度相当.微生物与其土壤资源之间的化学计量不平衡性综合考虑了土壤和微生物两者C:N:P的变异性, 能够更加准确地来衡量微生物化学组成与资源化学组成的差异性.本研究也发现胞外酶C:N:P和C:N:P化学计量不平衡性间存在负相关关系, 进一步证明了土壤与微生物之间C、N、P的差异会对胞外酶化学计量比产生影响(Mooshammer et al., 2014).前期研究已经证实土壤和微生物间的C:N化学计量不平衡性会对土壤C、N矿化速率产生影响(周正虎和王传宽, 2017), 综合考虑土壤和微生物C:N:P的变异性有助于我们对土壤C、N、P循环的理解. ...
Using microbial communities and extracellular enzymes to link soil organic matter characteristics to greenhouse gas production in a tidal freshwater wetland 1 2014
... 微生物分解者只有通过产生胞外酶将环境中复杂有机质分解成可溶性小分子物质才能被自身吸收利用(Schimel & Bennett, 2004), 微生物胞外酶活性的大小直接影响土壤C、N、P循环过程.例如: Morrissey等(2014)研究表明土壤C矿化速率与β-1,4-葡萄糖苷酶(BG)活性正相关.Chen等(2017)整合分析发现C获取相关的胞外酶对N添加的响应与土壤呼吸对N添加的响应之间存在显著正相关关系, 也就是说N添加会通过影响胞外酶活性来影响土壤呼吸, 最终影响土壤C储量.还有研究发现土壤磷酸酶活性与土壤有效磷含量之间呈负相关关系(Allison et al., 2007).通过量化胞外酶活性有助于理解土壤的生物地球化学循环过程(Sinsabaugh et al., 2009; Buchkowski et al., 2015).目前, 生态化学计量学相关理论已经被应用到微生物胞外酶研究中, 探索微生物胞外酶C:N:P对不同土壤C:N:P的适应机制.Xu等(2017)分析我国东部南北样带森林生态系统土壤酶活性及其化学计量比时发现, 土壤胞外酶C:P和N:P分别与土壤C:P和土壤N:P呈显著负相关关系.Peng和Wang (2016)研究发现土壤C、N、P是导致土壤胞外酶及其化学计量变异的主要因素.海拔梯度引起气候、植被、土壤特征的变化会进一步导致土壤C、N、P化学计量特征的变化(李相楹等, 2016), 然而土壤微生物是否会通过调整自身生物量C:N:P和胞外酶C:N:P进行适应仍不明确.通过研究微生物生物量和胞外酶C:N:P的环境可塑性反过来有助于我们对土壤C、N、P循环过程的理解. ...
Stoichiometry of soil extracellular enzyme activity along a climatic transect in temperate grasslands of northern China 2 2016
... 微生物分解者只有通过产生胞外酶将环境中复杂有机质分解成可溶性小分子物质才能被自身吸收利用(Schimel & Bennett, 2004), 微生物胞外酶活性的大小直接影响土壤C、N、P循环过程.例如: Morrissey等(2014)研究表明土壤C矿化速率与β-1,4-葡萄糖苷酶(BG)活性正相关.Chen等(2017)整合分析发现C获取相关的胞外酶对N添加的响应与土壤呼吸对N添加的响应之间存在显著正相关关系, 也就是说N添加会通过影响胞外酶活性来影响土壤呼吸, 最终影响土壤C储量.还有研究发现土壤磷酸酶活性与土壤有效磷含量之间呈负相关关系(Allison et al., 2007).通过量化胞外酶活性有助于理解土壤的生物地球化学循环过程(Sinsabaugh et al., 2009; Buchkowski et al., 2015).目前, 生态化学计量学相关理论已经被应用到微生物胞外酶研究中, 探索微生物胞外酶C:N:P对不同土壤C:N:P的适应机制.Xu等(2017)分析我国东部南北样带森林生态系统土壤酶活性及其化学计量比时发现, 土壤胞外酶C:P和N:P分别与土壤C:P和土壤N:P呈显著负相关关系.Peng和Wang (2016)研究发现土壤C、N、P是导致土壤胞外酶及其化学计量变异的主要因素.海拔梯度引起气候、植被、土壤特征的变化会进一步导致土壤C、N、P化学计量特征的变化(李相楹等, 2016), 然而土壤微生物是否会通过调整自身生物量C:N:P和胞外酶C:N:P进行适应仍不明确.通过研究微生物生物量和胞外酶C:N:P的环境可塑性反过来有助于我们对土壤C、N、P循环过程的理解. ...
... 随着海拔的升高, 微生物C、N获取相关胞外酶的活性呈现显著降低趋势(p < 0.05)(图3).这与Xu等(2015)在长白山的研究相符.由于土壤有机质或者土壤C、N含量会显著影响胞外酶的活性(Štursová & Baldrian, 2011; Wallenius et al., 2011), 以往研究认为土壤胞外酶活性随海拔升高而降低是由于土壤有机质含量降低所导致(Xu et al., 2015; Hofmann et al., 2016).然而在本研究中, 尽管针阔混交林的土壤C、N含量和微生物生物量C、N含量并不是最高, 但针阔混交林的BG和NAG活性却最高.这些结果表明海拔升高导致的温度降低可能抑制了微生物胞外酶的活性.与BG和NAG活性不同, 不同海拔间AP酶活性不存在显著差异(p > 0.05).AP与BG和NAG不同的海拔分布格局导致胞外酶C:P和N:P随着海拔的升高而显著降低.胞外酶化学计量比的这种变异性还可能与微生物对土壤化学计量比的适应策略相关(Peng & Wang, 2016; Zhou et al., 2017). ...
Nitrogen mineralization: Challenges of a changing paradigm 1 2004
... 微生物分解者只有通过产生胞外酶将环境中复杂有机质分解成可溶性小分子物质才能被自身吸收利用(Schimel & Bennett, 2004), 微生物胞外酶活性的大小直接影响土壤C、N、P循环过程.例如: Morrissey等(2014)研究表明土壤C矿化速率与β-1,4-葡萄糖苷酶(BG)活性正相关.Chen等(2017)整合分析发现C获取相关的胞外酶对N添加的响应与土壤呼吸对N添加的响应之间存在显著正相关关系, 也就是说N添加会通过影响胞外酶活性来影响土壤呼吸, 最终影响土壤C储量.还有研究发现土壤磷酸酶活性与土壤有效磷含量之间呈负相关关系(Allison et al., 2007).通过量化胞外酶活性有助于理解土壤的生物地球化学循环过程(Sinsabaugh et al., 2009; Buchkowski et al., 2015).目前, 生态化学计量学相关理论已经被应用到微生物胞外酶研究中, 探索微生物胞外酶C:N:P对不同土壤C:N:P的适应机制.Xu等(2017)分析我国东部南北样带森林生态系统土壤酶活性及其化学计量比时发现, 土壤胞外酶C:P和N:P分别与土壤C:P和土壤N:P呈显著负相关关系.Peng和Wang (2016)研究发现土壤C、N、P是导致土壤胞外酶及其化学计量变异的主要因素.海拔梯度引起气候、植被、土壤特征的变化会进一步导致土壤C、N、P化学计量特征的变化(李相楹等, 2016), 然而土壤微生物是否会通过调整自身生物量C:N:P和胞外酶C:N:P进行适应仍不明确.通过研究微生物生物量和胞外酶C:N:P的环境可塑性反过来有助于我们对土壤C、N、P循环过程的理解. ...
Ecoenzymatic stoichiometry of microbial organic nutrient acquisition in soil and sediment 2 2009
... 微生物分解者只有通过产生胞外酶将环境中复杂有机质分解成可溶性小分子物质才能被自身吸收利用(Schimel & Bennett, 2004), 微生物胞外酶活性的大小直接影响土壤C、N、P循环过程.例如: Morrissey等(2014)研究表明土壤C矿化速率与β-1,4-葡萄糖苷酶(BG)活性正相关.Chen等(2017)整合分析发现C获取相关的胞外酶对N添加的响应与土壤呼吸对N添加的响应之间存在显著正相关关系, 也就是说N添加会通过影响胞外酶活性来影响土壤呼吸, 最终影响土壤C储量.还有研究发现土壤磷酸酶活性与土壤有效磷含量之间呈负相关关系(Allison et al., 2007).通过量化胞外酶活性有助于理解土壤的生物地球化学循环过程(Sinsabaugh et al., 2009; Buchkowski et al., 2015).目前, 生态化学计量学相关理论已经被应用到微生物胞外酶研究中, 探索微生物胞外酶C:N:P对不同土壤C:N:P的适应机制.Xu等(2017)分析我国东部南北样带森林生态系统土壤酶活性及其化学计量比时发现, 土壤胞外酶C:P和N:P分别与土壤C:P和土壤N:P呈显著负相关关系.Peng和Wang (2016)研究发现土壤C、N、P是导致土壤胞外酶及其化学计量变异的主要因素.海拔梯度引起气候、植被、土壤特征的变化会进一步导致土壤C、N、P化学计量特征的变化(李相楹等, 2016), 然而土壤微生物是否会通过调整自身生物量C:N:P和胞外酶C:N:P进行适应仍不明确.通过研究微生物生物量和胞外酶C:N:P的环境可塑性反过来有助于我们对土壤C、N、P循环过程的理解. ...
... 土壤酶活性主要测定BG、β-1, 4-N-乙酰葡糖氨糖苷酶(NAG)、酸性磷酸酶(AP) 3种水解酶的活性.其中, BG主要是微生物获取C的胞外酶, NAG是微生物获取N的胞外酶, 而AP是微生物获取P的胞外酶(Sinsabaugh et al., 2009; Waring et al., 2014).酶活性的测定参考Martens等(1992)的方法.BG或NAG活性的测定方法为: 称取新鲜土壤样品1 g, 加入4 mL 0.1 mol·L-1醋酸缓冲液(pH = 5.0)和对硝基苯-β-D吡喃葡萄糖苷(BG底物, 0.025 mol·L-1)或β-N-乙酰氨基葡萄糖苷(NAG底物, 0.01 mol·L-1), 振荡混匀, 然后将土壤浆在35 ℃下培养1 h.培养结束后加入1 mL 0.5 mol·L-1 CaCl2和4 mL NaOH, 振荡过滤, 分光光度计410 nm下测量吸光值.AP活性的测定, 缓冲液使用改进的通用缓冲液(pH = 6.0), 底物为对硝基苯磷酸二钠(0.025 mol·L-1), 其他实验条件与程序同BG和NAG的测定方法. ...
Effects of soil properties and management on the activity of soil organic matter transforming enzymes and the quantification of soil-bound and free activity 1 2011
... 随着海拔的升高, 微生物C、N获取相关胞外酶的活性呈现显著降低趋势(p < 0.05)(图3).这与Xu等(2015)在长白山的研究相符.由于土壤有机质或者土壤C、N含量会显著影响胞外酶的活性(Štursová & Baldrian, 2011; Wallenius et al., 2011), 以往研究认为土壤胞外酶活性随海拔升高而降低是由于土壤有机质含量降低所导致(Xu et al., 2015; Hofmann et al., 2016).然而在本研究中, 尽管针阔混交林的土壤C、N含量和微生物生物量C、N含量并不是最高, 但针阔混交林的BG和NAG活性却最高.这些结果表明海拔升高导致的温度降低可能抑制了微生物胞外酶的活性.与BG和NAG活性不同, 不同海拔间AP酶活性不存在显著差异(p > 0.05).AP与BG和NAG不同的海拔分布格局导致胞外酶C:P和N:P随着海拔的升高而显著降低.胞外酶化学计量比的这种变异性还可能与微生物对土壤化学计量比的适应策略相关(Peng & Wang, 2016; Zhou et al., 2017). ...
Soil carbon and nitrogen stocks in forests along an altitudinal gradient in the eastern Himalayas and a meta-analysis of global data 5 2016
... ; Tashi et al., 2016; 谷晓楠等, 2017).为了阐明不同研究间土壤C沿海拔分布格局的差异性, 我们重新分析了Tashi等(2016)的全球数据.结果发现, 土壤C与海拔之间线性关系的斜率会随着最低海拔处的年平均气温的增加而显著增加(R2 = 0.26, p = 0.021, 图5), 说明温暖地区土壤C含量随着海拔升高的增加速度要高于寒冷地区土壤C含量随着海拔升高的增加速度.一般来说, 海拔每增加100 m, 温度会降低0.6 ℃.由于土壤呼吸与土壤温度之间普遍存在着指数函数关系(Davidson et al., 1998), 因此, 寒冷地区相比, 温暖地区相同程度的温度降低(即, 相同程度的海拔增加导致相同程度的温度降低)会导致更大程度的土壤呼吸速率降低, 这就解释了其中土壤C含量与海拔之间线性关系的斜率会随着年平均气温的增加而显著增加(图5).本研究地位于高纬度地区, 年平均气温较低, 海拔升高导致的土壤呼吸速率降低程度也较低, 导致本研究中不同海拔间土壤C含量不存在显著的差异(图1A). ...
... ).为了阐明不同研究间土壤C沿海拔分布格局的差异性, 我们重新分析了Tashi等(2016)的全球数据.结果发现, 土壤C与海拔之间线性关系的斜率会随着最低海拔处的年平均气温的增加而显著增加(R2 = 0.26, p = 0.021, 图5), 说明温暖地区土壤C含量随着海拔升高的增加速度要高于寒冷地区土壤C含量随着海拔升高的增加速度.一般来说, 海拔每增加100 m, 温度会降低0.6 ℃.由于土壤呼吸与土壤温度之间普遍存在着指数函数关系(Davidson et al., 1998), 因此, 寒冷地区相比, 温暖地区相同程度的温度降低(即, 相同程度的海拔增加导致相同程度的温度降低)会导致更大程度的土壤呼吸速率降低, 这就解释了其中土壤C含量与海拔之间线性关系的斜率会随着年平均气温的增加而显著增加(图5).本研究地位于高纬度地区, 年平均气温较低, 海拔升高导致的土壤呼吸速率降低程度也较低, 导致本研究中不同海拔间土壤C含量不存在显著的差异(图1A). ...
... 年平均气温和土壤C储量与海拔之间线性关系斜率的相关性.数据来源于Tashi等(2016)的全球整合分析.空心三角为本研究中的斜率.空心圆为异常值.图中斜率代表海拔每升高1 km土壤C储量(kg·m-2)的变化程度. Relationship between mean annual temperature and the slope of linear relationship between soil C storage and altitude. The data is from the global meta-analysis of <xref ref-type="bibr" rid="b33">Tashi <i>et al</i>. (2016)</xref>. The open triangle is the slope from the current study. The open circle is the outlier. The slope stands for the change in soil C storage (kg·m<sup>-2</sup>) per km increase in elevation.Fig. 5 与土壤C含量不同, 不同海拔间土壤N、P含量存在显著差异(p < 0.05).与低海拔的针阔混交林和针叶林相比, 高海拔的岳桦林和草地具有显著低的土壤N、P含量(图1B、1C).Kang等(2011)研究发现: 随着海拔的升高, 植物叶片的N含量会显著降低, 这可能在一定程度上解释了为什么高海拔的岳桦林和草地具有较低的土壤N含量.此外, 海拔升高导致的温度降低会抑制生物固氮过程(Zheng et al., 2019), 较低的生物固氮作用也可能是岳桦林和草地土壤N含量较低的另一个原因.对于P而言, 主要来源是矿物岩石的缓慢风化作用, 绝大部分存储在土壤中.魏晶等(2005)在长白山高山冻原系统的研究中发现土壤P库约占生态系统P库的99.2%.加之P又具有较低的迁移率, 因此, 生态系统总的P库大小基本保持不变.由于低海拔的生态系统生产力要显著高于高海拔的生态系统生产力, 这必然会将更多土壤深层的P通过凋落物的形式转运到土壤表层, 最终导致针阔混交林和针叶林0-10 cm土壤P含量显著高于岳桦林和草地.一般来说, 中海拔地区的地上生物量最高(Ensslin et al., 2015) , 因此, 本研究中土壤N、P含量随海拔升高呈现先增后降的垂直分布格局.尽管针阔混交林和针叶林之间土壤N、P含量差异不显著, 但是前者的土壤N、P含量要略低于后者, 这可能和针阔混交林受到人为干扰严重有关. ...
... Relationship between mean annual temperature and the slope of linear relationship between soil C storage and altitude. The data is from the global meta-analysis of Tashi et al. (2016). The open triangle is the slope from the current study. The open circle is the outlier. The slope stands for the change in soil C storage (kg·m-2) per km increase in elevation. Fig. 5 与土壤C含量不同, 不同海拔间土壤N、P含量存在显著差异(p < 0.05).与低海拔的针阔混交林和针叶林相比, 高海拔的岳桦林和草地具有显著低的土壤N、P含量(图1B、1C).Kang等(2011)研究发现: 随着海拔的升高, 植物叶片的N含量会显著降低, 这可能在一定程度上解释了为什么高海拔的岳桦林和草地具有较低的土壤N含量.此外, 海拔升高导致的温度降低会抑制生物固氮过程(Zheng et al., 2019), 较低的生物固氮作用也可能是岳桦林和草地土壤N含量较低的另一个原因.对于P而言, 主要来源是矿物岩石的缓慢风化作用, 绝大部分存储在土壤中.魏晶等(2005)在长白山高山冻原系统的研究中发现土壤P库约占生态系统P库的99.2%.加之P又具有较低的迁移率, 因此, 生态系统总的P库大小基本保持不变.由于低海拔的生态系统生产力要显著高于高海拔的生态系统生产力, 这必然会将更多土壤深层的P通过凋落物的形式转运到土壤表层, 最终导致针阔混交林和针叶林0-10 cm土壤P含量显著高于岳桦林和草地.一般来说, 中海拔地区的地上生物量最高(Ensslin et al., 2015) , 因此, 本研究中土壤N、P含量随海拔升高呈现先增后降的垂直分布格局.尽管针阔混交林和针叶林之间土壤N、P含量差异不显著, 但是前者的土壤N、P含量要略低于后者, 这可能和针阔混交林受到人为干扰严重有关. ...
Effects of land use on the level, variation and spatial structure of soil enzyme activities and bacterial communities 1 2011
... 随着海拔的升高, 微生物C、N获取相关胞外酶的活性呈现显著降低趋势(p < 0.05)(图3).这与Xu等(2015)在长白山的研究相符.由于土壤有机质或者土壤C、N含量会显著影响胞外酶的活性(Štursová & Baldrian, 2011; Wallenius et al., 2011), 以往研究认为土壤胞外酶活性随海拔升高而降低是由于土壤有机质含量降低所导致(Xu et al., 2015; Hofmann et al., 2016).然而在本研究中, 尽管针阔混交林的土壤C、N含量和微生物生物量C、N含量并不是最高, 但针阔混交林的BG和NAG活性却最高.这些结果表明海拔升高导致的温度降低可能抑制了微生物胞外酶的活性.与BG和NAG活性不同, 不同海拔间AP酶活性不存在显著差异(p > 0.05).AP与BG和NAG不同的海拔分布格局导致胞外酶C:P和N:P随着海拔的升高而显著降低.胞外酶化学计量比的这种变异性还可能与微生物对土壤化学计量比的适应策略相关(Peng & Wang, 2016; Zhou et al., 2017). ...
... 随着海拔的升高, 微生物C、N获取相关胞外酶的活性呈现显著降低趋势(p < 0.05)(图3).这与Xu等(2015)在长白山的研究相符.由于土壤有机质或者土壤C、N含量会显著影响胞外酶的活性(Štursová & Baldrian, 2011; Wallenius et al., 2011), 以往研究认为土壤胞外酶活性随海拔升高而降低是由于土壤有机质含量降低所导致(Xu et al., 2015; Hofmann et al., 2016).然而在本研究中, 尽管针阔混交林的土壤C、N含量和微生物生物量C、N含量并不是最高, 但针阔混交林的BG和NAG活性却最高.这些结果表明海拔升高导致的温度降低可能抑制了微生物胞外酶的活性.与BG和NAG活性不同, 不同海拔间AP酶活性不存在显著差异(p > 0.05).AP与BG和NAG不同的海拔分布格局导致胞外酶C:P和N:P随着海拔的升高而显著降低.胞外酶化学计量比的这种变异性还可能与微生物对土壤化学计量比的适应策略相关(Peng & Wang, 2016; Zhou et al., 2017). ...
A global analysis of soil microbial biomass carbon, nitrogen and phosphorus in terrestrial ecosystems 1 2013
... 尽管胞外酶C:N与土壤C:N之间不存在显著相关性, 本研究却发现胞外酶C:N与微生物C:N之间存在显著正相关关系(p < 0.05)(图4).此外, 胞外酶C:P和N:P也分别和微生物C:P和N:P存在正相关趋势.这些结果可能意味着微生物生物量C、N、P之间的相对比例大小与微生物对C、N、P 3种元素的获取能力相关.长久以来, 微生物生物量C:N:P的环境可塑性得到了广泛关注(Cleveland & Liptzin, 2007; Li et al., 2012; Xu et al., 2013), 而现在关注更多的是微生物生物量C:N:P对土壤功能过程的影响.周正虎和王传宽(2017)对帽儿山地区不同土地利用方式下土壤-微生物-矿化碳氮化学计量特征的研究中发现单位微生物生物量的碳矿化速率随着微生物C:N增加而降低, 单位微生物生物量的氮矿化速率随着微生物C:N的增加而增加.Buchkowski等(2015)甚至发现微生物生物量C:N:P对C、N矿化速率的影响程度和微生物生物量对C、N矿化速率的影响程度相当.微生物与其土壤资源之间的化学计量不平衡性综合考虑了土壤和微生物两者C:N:P的变异性, 能够更加准确地来衡量微生物化学组成与资源化学组成的差异性.本研究也发现胞外酶C:N:P和C:N:P化学计量不平衡性间存在负相关关系, 进一步证明了土壤与微生物之间C、N、P的差异会对胞外酶化学计量比产生影响(Mooshammer et al., 2014).前期研究已经证实土壤和微生物间的C:N化学计量不平衡性会对土壤C、N矿化速率产生影响(周正虎和王传宽, 2017), 综合考虑土壤和微生物C:N:P的变异性有助于我们对土壤C、N、P循环的理解. ...
The variations in soil microbial communities, enzyme activities and their relationships with soil organic matter decomposition along the northern slope of Changbai Mountain 1 2015
... 随着海拔的升高, 微生物C、N获取相关胞外酶的活性呈现显著降低趋势(p < 0.05)(图3).这与Xu等(2015)在长白山的研究相符.由于土壤有机质或者土壤C、N含量会显著影响胞外酶的活性(Štursová & Baldrian, 2011; Wallenius et al., 2011), 以往研究认为土壤胞外酶活性随海拔升高而降低是由于土壤有机质含量降低所导致(Xu et al., 2015; Hofmann et al., 2016).然而在本研究中, 尽管针阔混交林的土壤C、N含量和微生物生物量C、N含量并不是最高, 但针阔混交林的BG和NAG活性却最高.这些结果表明海拔升高导致的温度降低可能抑制了微生物胞外酶的活性.与BG和NAG活性不同, 不同海拔间AP酶活性不存在显著差异(p > 0.05).AP与BG和NAG不同的海拔分布格局导致胞外酶C:P和N:P随着海拔的升高而显著降低.胞外酶化学计量比的这种变异性还可能与微生物对土壤化学计量比的适应策略相关(Peng & Wang, 2016; Zhou et al., 2017). ...
Soil enzyme activity and stoichiometry in forest ecosystems along the North-South Transect in Eastern China (NSTEC) 2 2017
... 微生物分解者只有通过产生胞外酶将环境中复杂有机质分解成可溶性小分子物质才能被自身吸收利用(Schimel & Bennett, 2004), 微生物胞外酶活性的大小直接影响土壤C、N、P循环过程.例如: Morrissey等(2014)研究表明土壤C矿化速率与β-1,4-葡萄糖苷酶(BG)活性正相关.Chen等(2017)整合分析发现C获取相关的胞外酶对N添加的响应与土壤呼吸对N添加的响应之间存在显著正相关关系, 也就是说N添加会通过影响胞外酶活性来影响土壤呼吸, 最终影响土壤C储量.还有研究发现土壤磷酸酶活性与土壤有效磷含量之间呈负相关关系(Allison et al., 2007).通过量化胞外酶活性有助于理解土壤的生物地球化学循环过程(Sinsabaugh et al., 2009; Buchkowski et al., 2015).目前, 生态化学计量学相关理论已经被应用到微生物胞外酶研究中, 探索微生物胞外酶C:N:P对不同土壤C:N:P的适应机制.Xu等(2017)分析我国东部南北样带森林生态系统土壤酶活性及其化学计量比时发现, 土壤胞外酶C:P和N:P分别与土壤C:P和土壤N:P呈显著负相关关系.Peng和Wang (2016)研究发现土壤C、N、P是导致土壤胞外酶及其化学计量变异的主要因素.海拔梯度引起气候、植被、土壤特征的变化会进一步导致土壤C、N、P化学计量特征的变化(李相楹等, 2016), 然而土壤微生物是否会通过调整自身生物量C:N:P和胞外酶C:N:P进行适应仍不明确.通过研究微生物生物量和胞外酶C:N:P的环境可塑性反过来有助于我们对土壤C、N、P循环过程的理解. ...
 ,*东北林业大学生态研究中心, 哈尔滨 150040; 东北林业大学森林生态系统可持续经营教育部重点实验室, 哈尔滨 150040
,*东北林业大学生态研究中心, 哈尔滨 150040; 东北林业大学森林生态系统可持续经营教育部重点实验室, 哈尔滨 150040
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT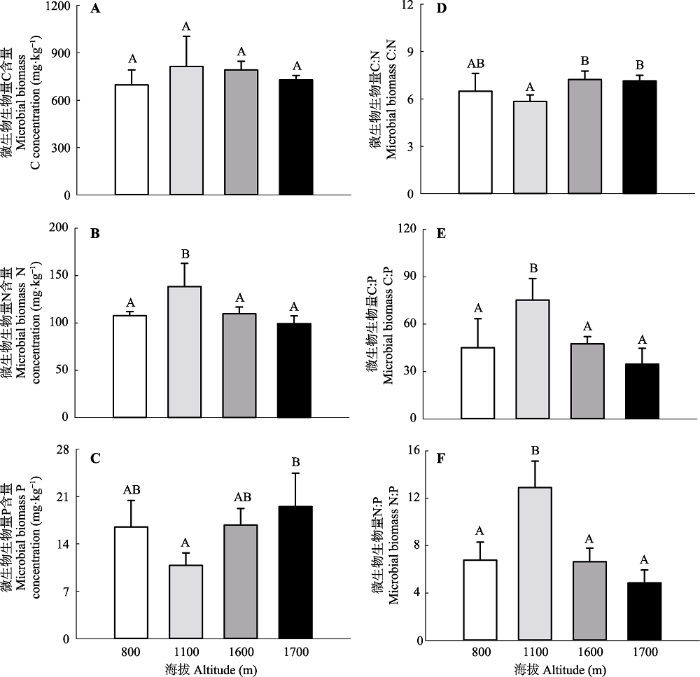 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT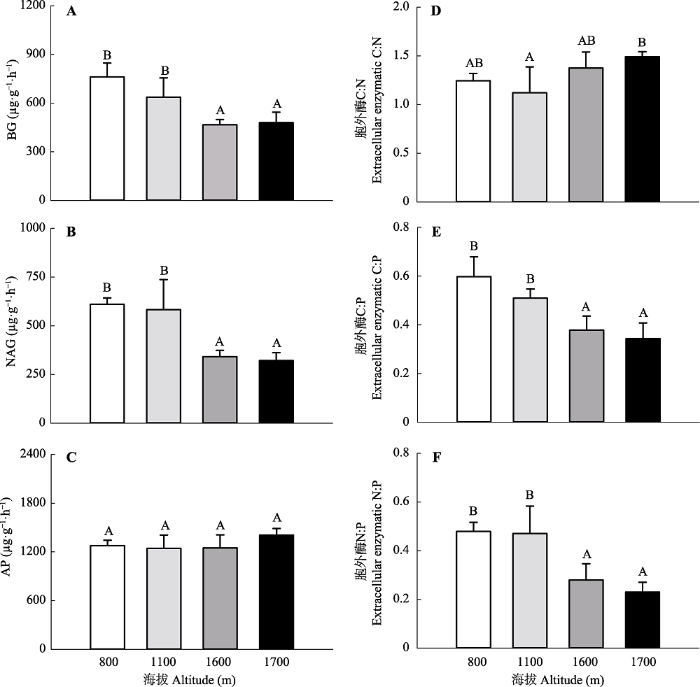 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT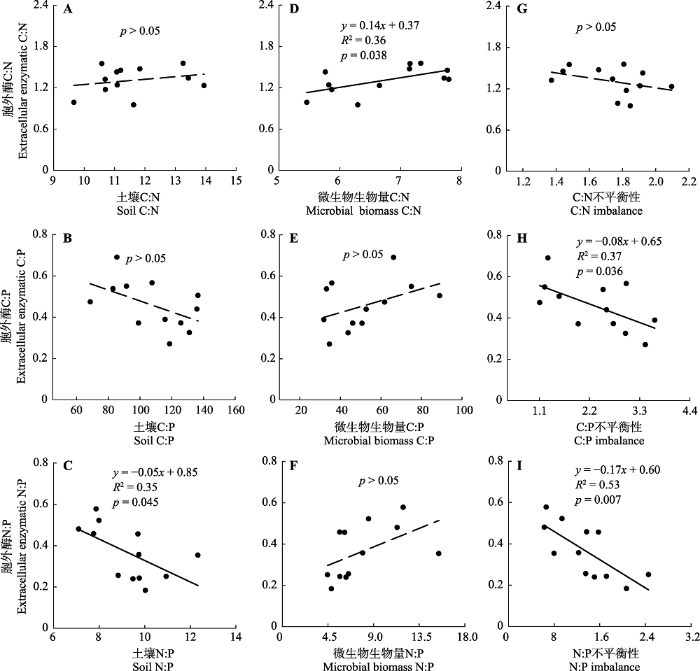 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT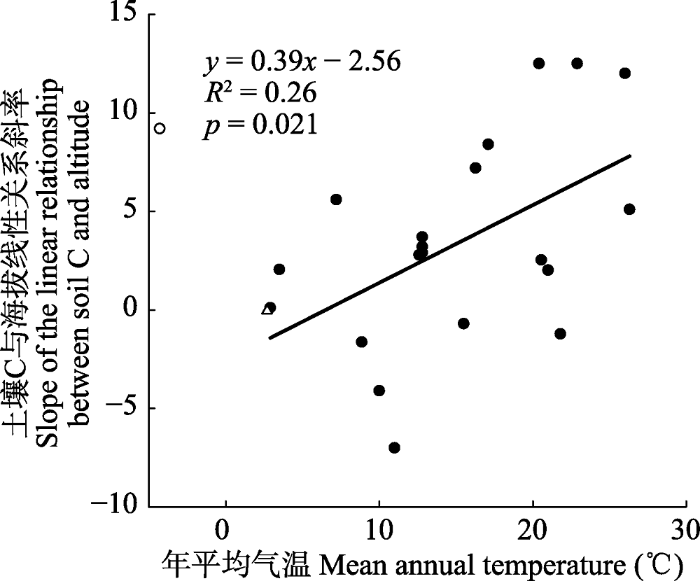 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}