Effects of nitrogen and water addition on soil respiration in a Nei Mongol desert steppe with different intensities of grazing history
Chao WEN,1,2,*, Yu-Mei SHAN1,2, Ru-Han YE1,2, Pu-Jin ZHANG1,2, Lan MU1,2, Hong CHANG1,2, Ting-Ting REN3,4, Shi-Ping CHEN3,4, Yong-Fei BAI3,4, Jian-Hui HUANG3,4, Hai-Lian SUN1,21Inner Mongolia Research Center for Prataculture, Chinese Academy of Sciences, Hohhot 010031, China 2Inner Mongolia Academy of Agricultural and Animal Husbandry Sciences, Hohhot 010031, China 3State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China 4Inner Mongolia Research Center for Prataculture, Chinese Academy of Sciences, Beijing 100093, China
National Natural Science Foundation of China(31860138) Natural Science Foundation of Nei Mongol of China(2019MS03029) National Key R&D Program of China(2016YFC0500705) Science and Technology Innovation Foundation of Nei Mongol Agriculture and Animal Husbandry(2017CXJJM07)
Abstract Aims Soil respiration is an important indicator for evaluation of ecosystem health in the grazing grasslands of arid regions, and thus can be used to assess dynamics of ecosystem functioning during the restoration of degraded grasslands from enduring intensive grazing. Methods This study was carried out in a Nei Mongol desert grassland with four grazing intensity treatments, i.e., control, light, moderate, and heavy grazing intensity designated as CK, LG, MG, and HG, respectively. Our objectives of this study were to explore the responses of soil respiration in these treatments with additional nitrogen (N) and water (W) addition. The plant community was dominated by a grass species, Stipa breviflora. Important findings Our results showed that: (1) previous grazing intensity had significant impacts on soil respiration in 2016 and 2017, but not in 2015. Grazing increased soil respiration. Moreover, both nitrogen and water addition significantly enhanced soil respiration in MG plots, while only combined addition of nitrogen and water significantly increased soil respiration in HG plots. (2) Neither grazing intensity nor addition of nitrogen and water changed the seasonal dynamics of growing season soil respiration in this desert grassland. Soil respiration showed a single-peak curve model, and the peak occurred in July with both rain and heat. (3) The effects of nitrogen and water addition varied in different growing seasons. Nitrogen addition had no significant effects in the first two years (2015 and 2016), while showed significant effects in the third year (2017). Water addition had significant effects in years with normal precipitation (2015 and 2017), while had insignificant effect in the year with high precipitation (2016). Combined addition of nitrogen and water showed stronger effects than only addition of water in CK, LG, and HG plots, indicating that the synergistic effects of nitrogen and water addition on soil respiration. (4) The sensitivity of soil respiration to soil temperature at 10 cm depth (i.e., the Q10 value) ranged between 1.13 and 2.41, with an average value of 1.71. Without addition of nitrogen and water, Q10 values in grazing plots were all lower than in CK plots, with the lowest value occurring in HG plots. With the addition of water and combined addition of water and nitrogen, the Q10 value increased significantly by 100%. Taken together, our results indicated that soil moisture was the leading environmental factor affecting soil respiration in this desert grassland, while nitrogen played an effective role only after the minimum requirement of water availability was met. Results from this study will provide important helpful information for the restoration and rational utilization of the degraded desert steppe. Keywords:desert steppe;grazing intensity;soil respiration;precipitation increase;nitrogen addition
PDF (1364KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 温超, 单玉梅, 晔薷罕, 张璞进, 木兰, 常虹, 任婷婷, 陈世苹, 白永飞, 黄建辉, 孙海莲. 氮和水分添加对内蒙古荒漠草原放牧生态系统土壤呼吸的影响. 植物生态学报, 2020, 44(1): 80-92. DOI: 10.17521/cjpe.2018.0177 WEN Chao, SHAN Yu-Mei, YE Ru-Han, ZHANG Pu-Jin, MU Lan, CHANG Hong, REN Ting-Ting, CHEN Shi-Ping, BAI Yong-Fei, HUANG Jian-Hui, SUN Hai-Lian. Effects of nitrogen and water addition on soil respiration in a Nei Mongol desert steppe with different intensities of grazing history. Chinese Journal of Plant Ecology, 2020, 44(1): 80-92. DOI: 10.17521/cjpe.2018.0177
Table 1 表1 表12015-2017年不同月份(M)、放牧强度(GI)、加水(W)、加氮(N)对内蒙古荒漠草原生长季土壤温度(ST)、土壤含水量(SM)、土壤呼吸速率(SR)影响的四因素方差分析结果 Table 1Results (F-value) of four-way ANOVA on the effects of month (M), grazing intensity (GI), water (W), nitrogen addition (N) and their interactions on growing season soil temperature (ST), soil moisture (SM) and soil respiration (SR), 2015 to 2017, in a Nei Mongol desert steppe
Fig. 2Changes in growing season soil moisture content and soil temperature, 2015 to 2017, in a Nei Mongol desert steppe (mean + SE). CKs, N, W, NW represent no addition of water and nitrogen, addition of nitrogen, addition of water, and combined addition of water and nitrogen, respectively. Different lowercase letters indicate significant difference between treatments in the same year at the 0.05 level; different uppercase letters indicate a significant difference between years at the 0.05 level.
Fig. 3Average soil respiration rate for 3 years under different grazing intensities under different treatments in a Nei Mongol desert steppe (mean + SE). CK, LG, MG, HG represent non-grazing, light grazing, moderate grazing and heavy grazing, respectively. CKs, N, W, NW represent no addition of water and nitrogen, addition of nitrogen, addition of water, and combined addition of water and nitrogen, respectively. Different lowercase letters indicate significant difference between treatments in the same year at the 0.05 level.
新窗口打开|下载原图ZIP|生成PPT 图4内蒙古荒漠草原土壤呼吸速率对氮水添加的响应(平均值±标准误差)。CK、LG、MG、HG分别表示不放牧、轻牧、中牧和重牧。CKs、N、W、NW分别表示不加水和氮、加氮、加水、加氮加水。*, p < 0.05; ns, p > 0.05。
Fig. 4Response of soil respiration to nitrogen and water addition in a Nei Mongol desert steppe (mean ± SE). CK, LG, MG, HG represent non-grazing, light grazing, moderate grazing and heavy grazing, respectively. CKs, N, W, NW represent no addition of water and nitrogen, addition of nitrogen, addition of water, and combined addition of water and nitrogen, respectively. *, p < 0.05; ns, p > 0.05。
Fig. 5Effects of different grazing intensities on the average soil respiration rate during the three years (2015-2017) under different water and nitrogen treatments in a Nei Mongol desert steppe (mean + SE). CK, LG, MG, HG represent non-grazing, light grazing, moderate grazing and heavy grazing, respectively. CKs, N, W, NW represent no addition of water and nitrogen, addition of nitrogen, addition of water, and combined addition of water and nitrogen, respectively. Different lowercase letters indicate significant difference between treatments in the same year at the 0.05 level.
Table 2 表2 表2内蒙古荒漠草原土壤呼吸速率与10 cm土壤温度的关系 Table 2Relationship between soil respiration rate and soil temperature at 10 cm depth in a Nei Mongol desert steppe
放牧强度 Grazing intensity
处理 Treatment
n
方程 Equation
R2
Q10
p
CK
CKs
24
Rs = 0.35e0.036t
0.026
1.43
0.477
N
24
Rs = 0.46e0.029t
0.023
1.33
0.554
W
24
Rs = 0.27e0.062t
0.102
1.86
0.243
NW
24
Rs = 0.17e0.085t
0.222
2.35
0.064
LG
CKs
24
Rs = 0.60e0.020t
0.008
1.22
0.560
N
24
Rs = 0.54e0.026t
0.020
1.30
0.566
W
24
Rs = 0.23e0.069t
0.141
2.00
0.204
NW
24
Rs = 0.24e0.075t
0.193
2.11
0.097
MG
CKs
24
Rs = 0.51e0.026t
0.012
1.29
0.511
N
24
Rs = 0.38e0.041t
0.051
1.51
0.369
W
24
Rs = 0.25e0.069t
0.153
2.00
0.170
NW
24
Rs = 0.17e0.088t
0.244
2.41
0.062
HG
CKs
24
Rs = 0.71e0.012t
0.003
1.13
0.546
N
24
Rs = 0.36e0.039t
0.045
1.47
0.412
W
24
Rs = 0.29e0.060t
0.135
1.83
0.147
NW
24
Rs = 0.25e0.074t
0.213
2.09
0.072
CK, LG, MG, HG represent non-grazing, light grazing, moderate grazing and heavy grazing, respectively. CKs, N, W, NW represent no addition of water and nitrogen, addition of nitrogen, addition of water, and combined addition of water and nitrogen, respectively. Bold text indicates significant edges (p < 0.10). CK、LG、MG、HG分别表示不放牧、轻牧、中牧和重牧。CKs、N、W、NW分别表示不加水和氮、加氮、加水、加氮加水。加粗字体表示边缘显著(p < 0.10)。
Table 3 表3 表3内蒙古荒漠草原土壤呼吸速率(Rs)与土壤含水量(Ws)的关系 Table 3Relationship between soil respiration rate (Rs) and soil moisture content Ws in a Nei Mongol desert steppe
放牧强度 Grazing intensity
处理 Treatment
n
方程 Equation
R2
p
CK
CKs
24
Rs = 0.12Ws - 0.35
0.44
<0.001
N
24
Rs = 0.09Ws - 0.02
0.30
<0.001
W
24
Rs = 0.08Ws + 0.19
0.28
<0.001
NW
24
Rs = 0.08Ws + 0.10
0.34
<0.001
LG
CKs
24
Rs = 0.14Ws - 0.46
0.46
<0.001
N
24
Rs = 0.09Ws + 0.12
0.29
<0.001
W
24
Rs = 0.08Ws + 0.29
0.24
<0.001
NW
24
Rs = 0.09Ws + 0.23
0.34
<0.001
MG
CKs
24
Rs = 0.13Ws - 0.34
0.40
<0.001
N
24
Rs = 0.11Ws - 0.08
0.36
<0.001
W
24
Rs = 0.11Ws - 0.04
0.32
<0.001
NW
24
Rs = 0.10Ws - 0.15
0.44
<0.001
HG
CKs
24
Rs = 0.10Ws - 0.05
0.34
<0.001
N
24
Rs = 0.08Ws + 0.18
0.25
<0.05
W
24
Rs = 0.09Ws + 0.01
0.31
<0.001
NW
24
Rs = 0.08Ws + 0.31
0.28
<0.001
CK, LG, MG, HG represent non-grazing, light grazing, moderate grazing and heavy grazing, respectively. CKs, N, W, NW represent no addition of water and nitrogen, addition of nitrogen, addition of water, and combined addition of water and nitrogen, respectively. CK、LG、MG、HG分别表示不放牧、轻牧、中牧和重牧。CKs、N、W、NW分别表示不加水和氮、加氮、加水、加氮加水。
AanderudZT, SchoolmasterDR, LennonJT (2011). Plants mediate the sensitivity of soil respiration to rainfall variability Ecosystems, 14, 156-167. DOI:10.1007/s10021-010-9401-yURL [本文引用: 1]
AmmannC, FlechardCR, LeifeldJ, NeftelA, FuhrerJ (2007). The carbon budget of newly established temperate grassland depends on management intensity Agriculture, Ecosystems & Environment, 121, 5-20. [本文引用: 1]
Amurijirigala, HongM, HanGD, GaoFG, ZhaoB, LiuCC (2013). Effects of different grazing intensity on soil respiration in the Stipa breviflora desert steppe Chinese Journal of Soil Science, 44, 321-327. [本文引用: 2]
BirchHF (1958). The effect of soil drying on humus decomposition and nitrogen availability Plant and Soil, 10, 9-31. [本文引用: 1]
Bond-LambertyB, ThomsonA (2010). Temperature-associated increases in the global soil respiration record Nature, 464, 579-582. [本文引用: 1]
BremerDJ, HamJM, OwensbyCE, KnappAK (1998). Responses of soil respiration to clipping and grazing in a tallgrass prairie Journal of Environmental Quality, 27, 1539-1548. [本文引用: 2]
CaoGM, TangYH, MoWH, WangYS, LiYN, ZhaoXQ (2004). Grazing intensity alters soil respiration in an alpine meadow on the Tibetan Plateau Soil Biology & Biochemistry, 36, 237-243. [本文引用: 3]
CasalsP, RomanyaJ, CortinaJ, BottnerP, CouteauxMM, VallejoVR (2000). CO2 efflux from a Mediterranean semi-arid forest soil. I. Seasonality and effects of stoniness Biogeochemistry, 48, 261-281. DOI:10.1023/A:1006289905991URL [本文引用: 1]
ChenJ, ZhouXH, WangJF, HruskaT, ShiWY, CaoJJ, ZhangBC, XuGX, ChenYZ, LuoYQ (2016). Grazing exclusion reduced soil respiration but increased its temperature sensitivity in a meadow grassland on the Tibetan Plateau Ecology & Evolution, 6, 675-687. [本文引用: 3]
ChenJB, HouFJ, ChenXJ, WanXL, MillnerJ (2015). Stocking rate and grazing season modify soil respiration on the Loess Plateau, China Rangeland Ecology & Management, 68, 48-53. [本文引用: 3]
ConantRT, Dalla-BettaP, KlopatekCC, KlopatekJM (2004). Controls on soil respiration in semiarid soils Soil Biology & Biochemistry, 36, 945-951. [本文引用: 1]
ConantRT, KlopatekJM, KlopatekCC (2000). Environmental factors controlling soil respiration in three semiarid ecosysterms Soil Science Society of America Journal, 64, 383-390. [本文引用: 1]
DavidsonEA, BelkE, BooneRD (1998). Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest Global Change Biology, 4, 217-227. DOI:10.1046/j.1365-2486.1998.00128.xURL [本文引用: 1]
DengY, LiuXN, YanRR, WangX, YangGX, RenZC, XinXP (2013). Soil respiration of Hulunber meadow steppe and response of its controlling factors to different grazing intensities Acta Prataculturae Sinica, 22(2), 22-29. [本文引用: 2]
DiaoLW, LiP, LiuWX, XuS, QiaoCL, ZengH, LiuLL (2018). Response of plant biomass to nitrogen addition and precipitation increasing under different climate conditions and time scales in grassland Chinese Journal of Plant Ecology, 42, 818-830. [本文引用: 1]
DiemerM (1997). Effects of elevated CO2 on gas exchange characteristics of alpine grassland Acta Oecologica, 18, 177-182. [本文引用: 1]
DongYS, QiYC, LiuJY, GengYB, ManfredD, YangXH, LiuLX (2005). Variation characteristics of soil respiration fluxes in four types of grassland communities under different precipitation intensity Chinese Science Bulletin, 50, 583-591. DOI:10.1007/BF02897484URL [本文引用: 1]
FuG, ZhangXZ, YuCQ, ShiPL, ZhouYT, LiYL, YangPW, ShenZX (2014). Response of soil respiration to grazing in an alpine meadow at three elevations in Tibet The Scientific World Journal, 20, 265142. DOI: 10.1155/2014/265142. [本文引用: 3]
GaoXF, WuCY, HanGD (2010). Effect of grazing on several ecological factors of the soil in typical steppe Journal of Arid Land Resources and Environment, 24(4), 130-133. [本文引用: 1]
GaoYZ, GieseM, LinS, SattelmacherB, ZhaoY, BrueckH (2008). Belowground net primary productivity and biomass allocation of a grassland in Inner Mongolia is affected by grazing intensity Plant and Soil, 307, 41-50. [本文引用: 2]
GrandyAS, SinsabaughRL, NeffJC, StursovaM, ZakDR (2008). Nitrogen deposition effects on soil organic matter chemistry are linked to variation in enzymes, ecosystems and size fractions Biogeochemistry, 91, 37-49. [本文引用: 1]
GuoMY, WeiZJ, XuLJ, YangGX, LiuHM, WuYL, XinXP (2011). Soil respiration of different mowing types of meadows Acta Agrestia Sinica, 19, 51-57. [本文引用: 1]
HagedornF, JoosO (2014). Experimental summer drought reduces soil CO2 effluxes and DOC leaching in Swiss grassland soils along an elevational gradient Biogeochemistry, 117, 395-412. [本文引用: 1]
HarrisonAF, TaylorK, ScottA, PoskittJ, BenhamD, GraceJ, ChaplowJ, RowlandP (2008). Potential effects of climate change on DOC release from three different soil types on the Northern Pennines UK: Examination using field manipulation experiments Global Change Biology, 14, 687-702. DOI:10.1111/gcb.2008.14.issue-3URL [本文引用: 1]
HooperDU, JohnsonL (1999). Nitrogen limitation in dryland ecosystems: Responses to geographical and temporal variation in precipitation Biogeochemistry, 46, 247-293. [本文引用: 2]
HouXY (2005). Grassland Ecology and Environment Construction Strategy in China. China Agriculture Press, Beijing. 38-49. [本文引用: 1]
HouXY, WangZ, MichaelSP, JiL, YunXJ (2014). The response of grassland productivity, soil carbon content and soil respiration rates to different grazing regimes in a desert steppe in northern China The Rangeland Journal, 36, 573-582. [本文引用: 2]
JiaBR, ZhouGS, WangFY, WangYH (2005). Soil respiration and its influencing factors at grazing and fenced typical Leymus chinensis steppe, Nei Mongol Environmental Science, 26, 3-9. [本文引用: 1]
KlumppK, FontaineS, AttardE, AttardE, Le RouxX, GleixnerG, SoussanaJF (2009). Grazing triggers soil carbon loss by altering plant roots and their control on soil microbial community Journal of Ecology, 97, 876-885. [本文引用: 2]
KuteschWL, KappenL (1997). Aspects of carbon and nitrogen cycling in soils of Bornh?ved Lake district. II. Modelling the influence of temperature increase on soil respiration and organic carbon content in arable soils under different managements Biogeochemistry, 39, 207-224. [本文引用: 1]
LiJZ, LinS, TaubeF, PanQM, DittertK (2011). Above and belowground net primary productivity of grassland influenced by supplemental water and nitrogen in Inner Mongolia Plant and Soil, 340, 253-264. [本文引用: 1]
LiLH, WangQB, BaiYF, ZhouGS, XingXR (2000). Soil respiration of a Leymus chinensis grassland stand in the Xilin river basin as affected by over-grazing and climate Acta Phytoecologica Sinica, 24, 680-686. [本文引用: 1]
LiMF, DongYS, QiYC, GengYB, LüY (2003). The analysis of diurnal variation of CO2 flux in Leymus chinensis grassland of Xilin River Basin Grassland of China, 25, 9-14. [本文引用: 1]
LiYL, HongM, BaiWM, HanGD, WangHM, ZhouM (2015). The responses of soil respiration to water and nitrogen in Stipa breviflora steppe Acta Ecologica Sinica, 35, 1727-1733. [本文引用: 3]
LinXW, ZhangZH, WangSP, HuYG, XuGP, LuoCY, ChangXF, DuanJC, LinQY, XuB, WangYF, ZhaoXQ, XieZB (2011). Response of ecosystem respiration to warming and grazing during the growing seasons in the alpine meadow on the Tibetan Plateau Agricultural and Forest Meteorology, 151, 792-802. [本文引用: 2]
LiuLL, WangX, LajeunesseMJ, MiaoGF, PiaoSL, WanSQ, WuYX, WangZH, YangS, LiP, DengMF (2016). A cross-biome synthesis of soil respiration and its determinants under simulated precipitation changes Global Change Biology, 22, 1394-1405. [本文引用: 2]
LiuWX, ZhangZ, WanSQ (2009). Predominant role of water in regulating soil and microbial respiration and their responses to climate change in a semiarid grassland Global Change Biology, 15, 184-195. DOI:10.1111/gcb.2009.15.issue-1URL [本文引用: 2]
LiuY, ZhouLR, MiaoSJ (2010). Effects of long-term fertilization on mollisoil enzyme activities and microbial respiration Soil and Fertilizer Sciences in China, (1), 7-10. [本文引用: 1]
MoJM, ZhangW, ZhuWX, GundersenP, FangYT, LiDJ, WangH (2007). Nitrogen addition reduces soil respiration in a mature tropical forest in southern China Global Change Biology, 14, 403-412. [本文引用: 2]
PanQM, BaiYF, HanXG, YangJC (2005). Effects of nitrogen additions on a Leymus chinensis population in typical steppe of Inner Mongolia Acta Phytoecologica Sinica, 29, 311-317. [本文引用: 1]
PengQ, DongYS, QiYC, XiaoSS, HeYT, MaT (2011). Effects of nitrogen fertilization on soil respiration in temperate grassland in Inner Mongolia, China Environmental Earth Sciences, 62, 1163-1171. [本文引用: 2]
PrietoLH, BertillerMB, CarreraAL, OliveraNL (2011). Soil enzyme and microbial activities in a grazing ecosystem of Patagonian Monte, Argentina Geoderma, 162, 281-287. [本文引用: 2]
QiYC, LiuXC, DongYS, PengQ, HeYT, SunLJ, JiaJQ, CaoCC (2014). Differential responses of short-term soil respiration dynamics to the experimental addition of nitrogen and water in the temperate semi-arid steppe of Inner Mongolia, China Journal of Environmental Sciences, 26, 834-845. [本文引用: 1]
RaichJW, PotterCS, BhagawatiD (2002). Interannual variability in global soil respiration, 1980-94 Global Change Biology, 8, 800-812. [本文引用: 1]
RaichJW, SchlesingerWH (1992). The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate Tellus B: Chemical & Physical Meteorology, 44(2), 81-99. [本文引用: 2]
ReichmannLG, SalaOE, PetersDPC (2013). Water controls on nitrogen transformations and stocks in an arid ecosystem Ecosphere, 4, 1-17. [本文引用: 1]
SánchezML, OzoresMI, LópezMJ, CollcR, de TorreB, GarcíaMA, PérezI (2003). Soil CO2 fluxes beneath barley on the central Spanish plateau Agricultural and Forest Meteorology, 118, 85-95. DOI:10.1016/S0168-1923(03)00066-2URL [本文引用: 1]
ShanD, HanGD, ZhaoML, WangZ, HanX, GaoFG (2009). The effects of experimental warming and nitrogen addition on soil respiration in desert Steppe Journal of Arid Land Resources and Environment, 23, 106-112. [本文引用: 1]
SongCC, LiuDY, YangGS, SongYY, MaoR (2011). Effect of nitrogen addition on decomposition of Calamagrostis angustifolia litters from freshwater marshes of Northeast China Ecological Engineering, 37, 1578-1582. [本文引用: 2]
SowerbyA, EmmettBA, TietemaA, BeierC (2008). Contrasting effects of repeated summer drought on soil carbon efflux in hydric and mesic heathland soils Global Change Biology, 14, 2388-2404. [本文引用: 1]
SowerbyA, EmmettBA, WilliamsD, BeierC, EvansCD (2010). The response of dissolved organic carbon (DOC) and the ecosystem carbon balance to experimental drought in a temperate shrubland European Journal of Soil Science, 61, 697-709. DOI:10.1111/j.1365-2389.2010.01276.xURL [本文引用: 1]
SponsellerRA (2007). Precipitation pulses and soil CO2 flux in a Sonoran desert ecosystem Global Change Biology, 13, 426-436. [本文引用: 1]
StarkS, M?nnist?MK, GanzertL, TiirolaM, H?ggblomMM (2015). Grazing intensity in subarctic tundra affects the temperature adaptation of soil microbial communities Soil Biology & Biochemistry, 84, 147-157. DOI:10.1016/j.soilbio.2015.02.023URL [本文引用: 3]
StedutoP, ?etink?kü?, AlbrizioR, KanberR (2002). Automated closed-system canopy-chamber for continuous field-crop monitoring of CO2 and H2O fluxes Agricultural and Forest Meteorology, 111, 171-186. [本文引用: 1]
WangYH, SongXH, WangZW, KangJ, HanGD, WangZW (2018). Response of plant community diversity and productivity on different stocking rates Chinese Journal of Grassland, 40(6), 107-112. [本文引用: 2]
WangYS, JiBM, HuangY, HuYQ, WangYF (2001). Effects of grazing and cultivating on emission of nitrous oxide, carbon dioxide and uptake of methane from grasslands Environmental Science, 22(6), 7-13. [本文引用: 1]
WeltzinJF, LoikME, SchwinningS, WilliamsDG, FayPA, HaddadBM, HarteJ, HuxmanTE, KnappAK, LinGH, PockmanWT, ShawMR, SmallEE, SmithMD, SmithSD, TissueDT, ZakJC (2003). Assessing the response of terrestrial ecosystems to potential changes in precipitation BioScience, 53, 941-952. DOI:10.1641/0006-3568(2003)053[0941:ATROTE]2.0.CO;2URL [本文引用: 1]
XuLK, BaldocchiDD, TangJW (2004). How soil moisture, rain pulses, and growth alter the response of ecosystem respiration to temperature Global Biogeochemical Cycles, 18(4), 1-10. [本文引用: 1]
YanLM, ChenSP, HuangJH, LinGH (2010). Differential responses of auto- and heterotrophic soil respiration to water and nitrogen addition in a semiarid temperate steppe Global Change Biology, 16, 2345-2357. [本文引用: 2]
YanLM, ChenSP, HuangJH, LinGH (2011). Water regulated effects of photosynthetic substrate supply on soil respiration in a semiarid steppe Global Change Biology, 17, 1990-2001. [本文引用: 2]
ZhangL, HuangJH, BaiYF, HanXG (2009). Effects of nitrogen additions on net nitrogen mineralization in Leymus chinensis grassland, Inner Mongolia, China Chinese Journal of Plant Ecology, 33, 563-569. [本文引用: 1]
ZhouLY, ZhouXH, ZhangBC, LuM, LuoYQ, LiuLL, LiB (2014). Different responses of soil respiration and its components to nitrogen addition among biomes: A meta- analysis Global Change Biology, 20, 2332-2343. DOI:10.1111/gcb.2014.20.issue-7URL [本文引用: 1]
Plants mediate the sensitivity of soil respiration to rainfall variability 1 2011
Responses of soil respiration to clipping and grazing in a tallgrass prairie 2 1998
... 放牧是我国北方草地生态系统主要的利用方式.关于放牧对草地生态系统碳交换过程的影响一直以来受到学术界的广泛关注(Lin et al., 2011; Fu et al., 2014; Hou et al., 2014; Chen et al., 2015, 2016; Stark et al., 2015).当前的研究结果表明, 放牧对土壤呼吸的影响并没有一致的规律性, 放牧可能导致土壤呼吸速率升高、降低, 或者对土壤呼吸没有显著的影响(Chen et al., 2015, 2016; Stark et al., 2015).事实上, 放牧对土壤呼吸的影响可能是多种因素综合作用的结果.一方面, 放牧牲畜的啃食作用直接降低了植被冠层的光合作用(Bremer et al., 1998), 使得分配于根系的光合同化产物减少, 降低根系生物量(Cao et al., 2004; Gao et al., 2008; Klumpp et al., 2009), 减少了地表凋落物和根际分泌物(Bremer et al., 1998), 降低土壤微生物生物量和土壤酶活性(Prieto et al., 2011; Li et al., 2013), 最终导致土壤呼吸速率降低.另一方面, 放牧牲畜以粪尿形式归还有机物, 大量的活性有机质和氮输入为微生物呼吸作用提供更加丰富的底物, 激发微生物的活性和增加养分的循环速度(Fu et al., 2014), 加速根系生长, 增加根系分泌物, 进而促进土壤呼吸.上述因素在不同的时间和研究区域, 对土壤呼吸的贡献大小并不一致, 进而导致土壤呼吸速率对放牧响应的不确定性. ...
... ), 减少了地表凋落物和根际分泌物(Bremer et al., 1998), 降低土壤微生物生物量和土壤酶活性(Prieto et al., 2011; Li et al., 2013), 最终导致土壤呼吸速率降低.另一方面, 放牧牲畜以粪尿形式归还有机物, 大量的活性有机质和氮输入为微生物呼吸作用提供更加丰富的底物, 激发微生物的活性和增加养分的循环速度(Fu et al., 2014), 加速根系生长, 增加根系分泌物, 进而促进土壤呼吸.上述因素在不同的时间和研究区域, 对土壤呼吸的贡献大小并不一致, 进而导致土壤呼吸速率对放牧响应的不确定性. ...
Grazing intensity alters soil respiration in an alpine meadow on the Tibetan Plateau 3 2004
... 放牧是我国北方草地生态系统主要的利用方式.关于放牧对草地生态系统碳交换过程的影响一直以来受到学术界的广泛关注(Lin et al., 2011; Fu et al., 2014; Hou et al., 2014; Chen et al., 2015, 2016; Stark et al., 2015).当前的研究结果表明, 放牧对土壤呼吸的影响并没有一致的规律性, 放牧可能导致土壤呼吸速率升高、降低, 或者对土壤呼吸没有显著的影响(Chen et al., 2015, 2016; Stark et al., 2015).事实上, 放牧对土壤呼吸的影响可能是多种因素综合作用的结果.一方面, 放牧牲畜的啃食作用直接降低了植被冠层的光合作用(Bremer et al., 1998), 使得分配于根系的光合同化产物减少, 降低根系生物量(Cao et al., 2004; Gao et al., 2008; Klumpp et al., 2009), 减少了地表凋落物和根际分泌物(Bremer et al., 1998), 降低土壤微生物生物量和土壤酶活性(Prieto et al., 2011; Li et al., 2013), 最终导致土壤呼吸速率降低.另一方面, 放牧牲畜以粪尿形式归还有机物, 大量的活性有机质和氮输入为微生物呼吸作用提供更加丰富的底物, 激发微生物的活性和增加养分的循环速度(Fu et al., 2014), 加速根系生长, 增加根系分泌物, 进而促进土壤呼吸.上述因素在不同的时间和研究区域, 对土壤呼吸的贡献大小并不一致, 进而导致土壤呼吸速率对放牧响应的不确定性. ...
... 放牧对草地生态系统的影响是复杂、多方面且持久的, 放牧对草原植物(Cao et al., 2004; Gao et al., 2008)、微生物(Li et al., 2013)、凋落物(Klumpp et al., 2009)、土壤理化特性(Prieto et al., 2011; Li et al., 2013)等都存在着不同程度的影响, 而且放牧强度及其持续时间的不同也强烈影响着草地生态系统结构和功能.不同放牧强度的干扰将会迅速引起草地植被群落特征(王悦骅等, 2018)、土壤微生物数量、土壤酶和土壤理化性质如含水量、土壤有机质含量等方面的变化(高雪峰等, 2010), 而且在一定程度上影响水、热因子的作用效应, 进而导致土壤呼吸作用改变.已有的研究结果显示, 放牧不仅可以降低土壤呼吸速率(邓钰等, 2013; Hou et al., 2014), 也可以促进土壤呼吸(Chen et al., 2015, 2016; Stark et al., 2015), 也有影响不显著的(Lin et al., 2011; Fu et al., 2014).有研究发现放牧对土壤呼吸影响的大小与方向和放牧强度有关, 轻度放牧促进土壤呼吸, 中度和重度放牧则降低土壤呼吸速率(李玉强等, 2006).Wang等(2001)在内蒙古温带草原的研究表明, 自由放牧可增加土壤呼吸的年排放量, 且土壤呼吸与放牧强度呈正相关关系.然而, 也有研究认为, 荒漠草原放牧草地的土壤呼吸速率显著低于未放牧草地(阿木日吉日嘎拉等, 2013).本研究中, 过去不同的放牧强度对土壤呼吸速率的影响具有明显的年际变化, 表现为无显著影响和促进两种.放牧在总体上促进了土壤呼吸, 表现为放牧(LG、MG、HG)区3年平均土壤呼吸速率都高于禁牧区.两者研究结果截然不同, 这里存在两种可能, 一种是放牧增加土壤呼吸, 即使在停止放牧后, 放牧区的土壤呼吸速率仍然较高; 还有一种可能是放牧可能抑制土壤呼吸, 但停止放牧后, 土壤呼吸产生了超补偿, 使得土壤呼吸迅速恢复, 并超过对照区的, 而我们的研究结果属于哪种情况, 我们并不清楚.另外, 本研究是在多年放牧的草地上进行了小区围封, 实际上变成了围封草地, 围封后放牧的扰动已经去除, 草地的生物量和水分条件得到了一定程度的恢复, 从而引起土壤呼吸变化.已有研究也表明, 围封草地的土壤呼吸速率要明显高于放牧草地(贾丙瑞等, 2005), 而且放牧会导致全年的土壤呼吸速率比围封草地降低1/3 (马涛等, 2009).另外, 本研究持续3年测定土壤呼吸速率, 研究年限的不同对草地土壤呼吸的影响也存在差异(郭明英等, 2011). ...
Grazing exclusion reduced soil respiration but increased its temperature sensitivity in a meadow grassland on the Tibetan Plateau 3 2016
... 放牧是我国北方草地生态系统主要的利用方式.关于放牧对草地生态系统碳交换过程的影响一直以来受到学术界的广泛关注(Lin et al., 2011; Fu et al., 2014; Hou et al., 2014; Chen et al., 2015, 2016; Stark et al., 2015).当前的研究结果表明, 放牧对土壤呼吸的影响并没有一致的规律性, 放牧可能导致土壤呼吸速率升高、降低, 或者对土壤呼吸没有显著的影响(Chen et al., 2015, 2016; Stark et al., 2015).事实上, 放牧对土壤呼吸的影响可能是多种因素综合作用的结果.一方面, 放牧牲畜的啃食作用直接降低了植被冠层的光合作用(Bremer et al., 1998), 使得分配于根系的光合同化产物减少, 降低根系生物量(Cao et al., 2004; Gao et al., 2008; Klumpp et al., 2009), 减少了地表凋落物和根际分泌物(Bremer et al., 1998), 降低土壤微生物生物量和土壤酶活性(Prieto et al., 2011; Li et al., 2013), 最终导致土壤呼吸速率降低.另一方面, 放牧牲畜以粪尿形式归还有机物, 大量的活性有机质和氮输入为微生物呼吸作用提供更加丰富的底物, 激发微生物的活性和增加养分的循环速度(Fu et al., 2014), 加速根系生长, 增加根系分泌物, 进而促进土壤呼吸.上述因素在不同的时间和研究区域, 对土壤呼吸的贡献大小并不一致, 进而导致土壤呼吸速率对放牧响应的不确定性. ...
... , 2016; Stark et al., 2015).事实上, 放牧对土壤呼吸的影响可能是多种因素综合作用的结果.一方面, 放牧牲畜的啃食作用直接降低了植被冠层的光合作用(Bremer et al., 1998), 使得分配于根系的光合同化产物减少, 降低根系生物量(Cao et al., 2004; Gao et al., 2008; Klumpp et al., 2009), 减少了地表凋落物和根际分泌物(Bremer et al., 1998), 降低土壤微生物生物量和土壤酶活性(Prieto et al., 2011; Li et al., 2013), 最终导致土壤呼吸速率降低.另一方面, 放牧牲畜以粪尿形式归还有机物, 大量的活性有机质和氮输入为微生物呼吸作用提供更加丰富的底物, 激发微生物的活性和增加养分的循环速度(Fu et al., 2014), 加速根系生长, 增加根系分泌物, 进而促进土壤呼吸.上述因素在不同的时间和研究区域, 对土壤呼吸的贡献大小并不一致, 进而导致土壤呼吸速率对放牧响应的不确定性. ...
... 放牧对草地生态系统的影响是复杂、多方面且持久的, 放牧对草原植物(Cao et al., 2004; Gao et al., 2008)、微生物(Li et al., 2013)、凋落物(Klumpp et al., 2009)、土壤理化特性(Prieto et al., 2011; Li et al., 2013)等都存在着不同程度的影响, 而且放牧强度及其持续时间的不同也强烈影响着草地生态系统结构和功能.不同放牧强度的干扰将会迅速引起草地植被群落特征(王悦骅等, 2018)、土壤微生物数量、土壤酶和土壤理化性质如含水量、土壤有机质含量等方面的变化(高雪峰等, 2010), 而且在一定程度上影响水、热因子的作用效应, 进而导致土壤呼吸作用改变.已有的研究结果显示, 放牧不仅可以降低土壤呼吸速率(邓钰等, 2013; Hou et al., 2014), 也可以促进土壤呼吸(Chen et al., 2015, 2016; Stark et al., 2015), 也有影响不显著的(Lin et al., 2011; Fu et al., 2014).有研究发现放牧对土壤呼吸影响的大小与方向和放牧强度有关, 轻度放牧促进土壤呼吸, 中度和重度放牧则降低土壤呼吸速率(李玉强等, 2006).Wang等(2001)在内蒙古温带草原的研究表明, 自由放牧可增加土壤呼吸的年排放量, 且土壤呼吸与放牧强度呈正相关关系.然而, 也有研究认为, 荒漠草原放牧草地的土壤呼吸速率显著低于未放牧草地(阿木日吉日嘎拉等, 2013).本研究中, 过去不同的放牧强度对土壤呼吸速率的影响具有明显的年际变化, 表现为无显著影响和促进两种.放牧在总体上促进了土壤呼吸, 表现为放牧(LG、MG、HG)区3年平均土壤呼吸速率都高于禁牧区.两者研究结果截然不同, 这里存在两种可能, 一种是放牧增加土壤呼吸, 即使在停止放牧后, 放牧区的土壤呼吸速率仍然较高; 还有一种可能是放牧可能抑制土壤呼吸, 但停止放牧后, 土壤呼吸产生了超补偿, 使得土壤呼吸迅速恢复, 并超过对照区的, 而我们的研究结果属于哪种情况, 我们并不清楚.另外, 本研究是在多年放牧的草地上进行了小区围封, 实际上变成了围封草地, 围封后放牧的扰动已经去除, 草地的生物量和水分条件得到了一定程度的恢复, 从而引起土壤呼吸变化.已有研究也表明, 围封草地的土壤呼吸速率要明显高于放牧草地(贾丙瑞等, 2005), 而且放牧会导致全年的土壤呼吸速率比围封草地降低1/3 (马涛等, 2009).另外, 本研究持续3年测定土壤呼吸速率, 研究年限的不同对草地土壤呼吸的影响也存在差异(郭明英等, 2011). ...
Stocking rate and grazing season modify soil respiration on the Loess Plateau, China 3 2015
... 放牧是我国北方草地生态系统主要的利用方式.关于放牧对草地生态系统碳交换过程的影响一直以来受到学术界的广泛关注(Lin et al., 2011; Fu et al., 2014; Hou et al., 2014; Chen et al., 2015, 2016; Stark et al., 2015).当前的研究结果表明, 放牧对土壤呼吸的影响并没有一致的规律性, 放牧可能导致土壤呼吸速率升高、降低, 或者对土壤呼吸没有显著的影响(Chen et al., 2015, 2016; Stark et al., 2015).事实上, 放牧对土壤呼吸的影响可能是多种因素综合作用的结果.一方面, 放牧牲畜的啃食作用直接降低了植被冠层的光合作用(Bremer et al., 1998), 使得分配于根系的光合同化产物减少, 降低根系生物量(Cao et al., 2004; Gao et al., 2008; Klumpp et al., 2009), 减少了地表凋落物和根际分泌物(Bremer et al., 1998), 降低土壤微生物生物量和土壤酶活性(Prieto et al., 2011; Li et al., 2013), 最终导致土壤呼吸速率降低.另一方面, 放牧牲畜以粪尿形式归还有机物, 大量的活性有机质和氮输入为微生物呼吸作用提供更加丰富的底物, 激发微生物的活性和增加养分的循环速度(Fu et al., 2014), 加速根系生长, 增加根系分泌物, 进而促进土壤呼吸.上述因素在不同的时间和研究区域, 对土壤呼吸的贡献大小并不一致, 进而导致土壤呼吸速率对放牧响应的不确定性. ...
... ).当前的研究结果表明, 放牧对土壤呼吸的影响并没有一致的规律性, 放牧可能导致土壤呼吸速率升高、降低, 或者对土壤呼吸没有显著的影响(Chen et al., 2015, 2016; Stark et al., 2015).事实上, 放牧对土壤呼吸的影响可能是多种因素综合作用的结果.一方面, 放牧牲畜的啃食作用直接降低了植被冠层的光合作用(Bremer et al., 1998), 使得分配于根系的光合同化产物减少, 降低根系生物量(Cao et al., 2004; Gao et al., 2008; Klumpp et al., 2009), 减少了地表凋落物和根际分泌物(Bremer et al., 1998), 降低土壤微生物生物量和土壤酶活性(Prieto et al., 2011; Li et al., 2013), 最终导致土壤呼吸速率降低.另一方面, 放牧牲畜以粪尿形式归还有机物, 大量的活性有机质和氮输入为微生物呼吸作用提供更加丰富的底物, 激发微生物的活性和增加养分的循环速度(Fu et al., 2014), 加速根系生长, 增加根系分泌物, 进而促进土壤呼吸.上述因素在不同的时间和研究区域, 对土壤呼吸的贡献大小并不一致, 进而导致土壤呼吸速率对放牧响应的不确定性. ...
... 放牧对草地生态系统的影响是复杂、多方面且持久的, 放牧对草原植物(Cao et al., 2004; Gao et al., 2008)、微生物(Li et al., 2013)、凋落物(Klumpp et al., 2009)、土壤理化特性(Prieto et al., 2011; Li et al., 2013)等都存在着不同程度的影响, 而且放牧强度及其持续时间的不同也强烈影响着草地生态系统结构和功能.不同放牧强度的干扰将会迅速引起草地植被群落特征(王悦骅等, 2018)、土壤微生物数量、土壤酶和土壤理化性质如含水量、土壤有机质含量等方面的变化(高雪峰等, 2010), 而且在一定程度上影响水、热因子的作用效应, 进而导致土壤呼吸作用改变.已有的研究结果显示, 放牧不仅可以降低土壤呼吸速率(邓钰等, 2013; Hou et al., 2014), 也可以促进土壤呼吸(Chen et al., 2015, 2016; Stark et al., 2015), 也有影响不显著的(Lin et al., 2011; Fu et al., 2014).有研究发现放牧对土壤呼吸影响的大小与方向和放牧强度有关, 轻度放牧促进土壤呼吸, 中度和重度放牧则降低土壤呼吸速率(李玉强等, 2006).Wang等(2001)在内蒙古温带草原的研究表明, 自由放牧可增加土壤呼吸的年排放量, 且土壤呼吸与放牧强度呈正相关关系.然而, 也有研究认为, 荒漠草原放牧草地的土壤呼吸速率显著低于未放牧草地(阿木日吉日嘎拉等, 2013).本研究中, 过去不同的放牧强度对土壤呼吸速率的影响具有明显的年际变化, 表现为无显著影响和促进两种.放牧在总体上促进了土壤呼吸, 表现为放牧(LG、MG、HG)区3年平均土壤呼吸速率都高于禁牧区.两者研究结果截然不同, 这里存在两种可能, 一种是放牧增加土壤呼吸, 即使在停止放牧后, 放牧区的土壤呼吸速率仍然较高; 还有一种可能是放牧可能抑制土壤呼吸, 但停止放牧后, 土壤呼吸产生了超补偿, 使得土壤呼吸迅速恢复, 并超过对照区的, 而我们的研究结果属于哪种情况, 我们并不清楚.另外, 本研究是在多年放牧的草地上进行了小区围封, 实际上变成了围封草地, 围封后放牧的扰动已经去除, 草地的生物量和水分条件得到了一定程度的恢复, 从而引起土壤呼吸变化.已有研究也表明, 围封草地的土壤呼吸速率要明显高于放牧草地(贾丙瑞等, 2005), 而且放牧会导致全年的土壤呼吸速率比围封草地降低1/3 (马涛等, 2009).另外, 本研究持续3年测定土壤呼吸速率, 研究年限的不同对草地土壤呼吸的影响也存在差异(郭明英等, 2011). ...
Controls on soil respiration in semiarid soils 1 2004
... 氮和水分是草地生态系统生产力的两个主要限制性因子(Hooper & Johnson, 1999; Yan et al., 2011).鉴于草地退化常导致土壤养分缺乏, 氮添加作为一种重要手段被用于对退化草地的恢复和管理中(潘庆民等, 2005; 张璐等, 2009).氮添加可能影响生态系统光合作用和植物生长, 导致植物生产力的不同响应(Ren et al., 2017), 进而对土壤呼吸产生促进(Song et al., 2011)、抑制(Mo et al., 2007)或无显著影响(Peng et al., 2011; Qi et al., 2014; 李寅龙等, 2015) 3种不同的作用.水分作为干旱区草地生态系统过程重要的限制因子, 通常降水增加对土壤呼吸有显著促进作用(Sowerby et al., 2008; Yan et al., 2010, 2011), 而氮添加的效果常常会依赖于水分条件.刁励玮等(2018)统计分析了1990-2017年间发表的有关氮添加的研究论文结果, 发现单独施氮对地下生物量无显著影响, 氮水同时添加能显著提高地下生物量, 两者对生态系统生产力存在显著的交互作用(Li et al., 2011), 因此氮和水分两者可能共同影响生态系统的碳交换(Hooper & Johnson, 1999). ...
草地生态系统生物量在不同气候及多时间尺度上对氮添加和增雨处理的响应 1 2018
... 氮和水分是草地生态系统生产力的两个主要限制性因子(Hooper & Johnson, 1999; Yan et al., 2011).鉴于草地退化常导致土壤养分缺乏, 氮添加作为一种重要手段被用于对退化草地的恢复和管理中(潘庆民等, 2005; 张璐等, 2009).氮添加可能影响生态系统光合作用和植物生长, 导致植物生产力的不同响应(Ren et al., 2017), 进而对土壤呼吸产生促进(Song et al., 2011)、抑制(Mo et al., 2007)或无显著影响(Peng et al., 2011; Qi et al., 2014; 李寅龙等, 2015) 3种不同的作用.水分作为干旱区草地生态系统过程重要的限制因子, 通常降水增加对土壤呼吸有显著促进作用(Sowerby et al., 2008; Yan et al., 2010, 2011), 而氮添加的效果常常会依赖于水分条件.刁励玮等(2018)统计分析了1990-2017年间发表的有关氮添加的研究论文结果, 发现单独施氮对地下生物量无显著影响, 氮水同时添加能显著提高地下生物量, 两者对生态系统生产力存在显著的交互作用(Li et al., 2011), 因此氮和水分两者可能共同影响生态系统的碳交换(Hooper & Johnson, 1999). ...
Effects of elevated CO2 on gas exchange characteristics of alpine grassland 1 1997
Response of soil respiration to grazing in an alpine meadow at three elevations in Tibet 3 2014
... 放牧是我国北方草地生态系统主要的利用方式.关于放牧对草地生态系统碳交换过程的影响一直以来受到学术界的广泛关注(Lin et al., 2011; Fu et al., 2014; Hou et al., 2014; Chen et al., 2015, 2016; Stark et al., 2015).当前的研究结果表明, 放牧对土壤呼吸的影响并没有一致的规律性, 放牧可能导致土壤呼吸速率升高、降低, 或者对土壤呼吸没有显著的影响(Chen et al., 2015, 2016; Stark et al., 2015).事实上, 放牧对土壤呼吸的影响可能是多种因素综合作用的结果.一方面, 放牧牲畜的啃食作用直接降低了植被冠层的光合作用(Bremer et al., 1998), 使得分配于根系的光合同化产物减少, 降低根系生物量(Cao et al., 2004; Gao et al., 2008; Klumpp et al., 2009), 减少了地表凋落物和根际分泌物(Bremer et al., 1998), 降低土壤微生物生物量和土壤酶活性(Prieto et al., 2011; Li et al., 2013), 最终导致土壤呼吸速率降低.另一方面, 放牧牲畜以粪尿形式归还有机物, 大量的活性有机质和氮输入为微生物呼吸作用提供更加丰富的底物, 激发微生物的活性和增加养分的循环速度(Fu et al., 2014), 加速根系生长, 增加根系分泌物, 进而促进土壤呼吸.上述因素在不同的时间和研究区域, 对土壤呼吸的贡献大小并不一致, 进而导致土壤呼吸速率对放牧响应的不确定性. ...
... 放牧对草地生态系统的影响是复杂、多方面且持久的, 放牧对草原植物(Cao et al., 2004; Gao et al., 2008)、微生物(Li et al., 2013)、凋落物(Klumpp et al., 2009)、土壤理化特性(Prieto et al., 2011; Li et al., 2013)等都存在着不同程度的影响, 而且放牧强度及其持续时间的不同也强烈影响着草地生态系统结构和功能.不同放牧强度的干扰将会迅速引起草地植被群落特征(王悦骅等, 2018)、土壤微生物数量、土壤酶和土壤理化性质如含水量、土壤有机质含量等方面的变化(高雪峰等, 2010), 而且在一定程度上影响水、热因子的作用效应, 进而导致土壤呼吸作用改变.已有的研究结果显示, 放牧不仅可以降低土壤呼吸速率(邓钰等, 2013; Hou et al., 2014), 也可以促进土壤呼吸(Chen et al., 2015, 2016; Stark et al., 2015), 也有影响不显著的(Lin et al., 2011; Fu et al., 2014).有研究发现放牧对土壤呼吸影响的大小与方向和放牧强度有关, 轻度放牧促进土壤呼吸, 中度和重度放牧则降低土壤呼吸速率(李玉强等, 2006).Wang等(2001)在内蒙古温带草原的研究表明, 自由放牧可增加土壤呼吸的年排放量, 且土壤呼吸与放牧强度呈正相关关系.然而, 也有研究认为, 荒漠草原放牧草地的土壤呼吸速率显著低于未放牧草地(阿木日吉日嘎拉等, 2013).本研究中, 过去不同的放牧强度对土壤呼吸速率的影响具有明显的年际变化, 表现为无显著影响和促进两种.放牧在总体上促进了土壤呼吸, 表现为放牧(LG、MG、HG)区3年平均土壤呼吸速率都高于禁牧区.两者研究结果截然不同, 这里存在两种可能, 一种是放牧增加土壤呼吸, 即使在停止放牧后, 放牧区的土壤呼吸速率仍然较高; 还有一种可能是放牧可能抑制土壤呼吸, 但停止放牧后, 土壤呼吸产生了超补偿, 使得土壤呼吸迅速恢复, 并超过对照区的, 而我们的研究结果属于哪种情况, 我们并不清楚.另外, 本研究是在多年放牧的草地上进行了小区围封, 实际上变成了围封草地, 围封后放牧的扰动已经去除, 草地的生物量和水分条件得到了一定程度的恢复, 从而引起土壤呼吸变化.已有研究也表明, 围封草地的土壤呼吸速率要明显高于放牧草地(贾丙瑞等, 2005), 而且放牧会导致全年的土壤呼吸速率比围封草地降低1/3 (马涛等, 2009).另外, 本研究持续3年测定土壤呼吸速率, 研究年限的不同对草地土壤呼吸的影响也存在差异(郭明英等, 2011). ...
放牧对典型草原土壤中几种生态因子影响的研究 1 2010
... 放牧对草地生态系统的影响是复杂、多方面且持久的, 放牧对草原植物(Cao et al., 2004; Gao et al., 2008)、微生物(Li et al., 2013)、凋落物(Klumpp et al., 2009)、土壤理化特性(Prieto et al., 2011; Li et al., 2013)等都存在着不同程度的影响, 而且放牧强度及其持续时间的不同也强烈影响着草地生态系统结构和功能.不同放牧强度的干扰将会迅速引起草地植被群落特征(王悦骅等, 2018)、土壤微生物数量、土壤酶和土壤理化性质如含水量、土壤有机质含量等方面的变化(高雪峰等, 2010), 而且在一定程度上影响水、热因子的作用效应, 进而导致土壤呼吸作用改变.已有的研究结果显示, 放牧不仅可以降低土壤呼吸速率(邓钰等, 2013; Hou et al., 2014), 也可以促进土壤呼吸(Chen et al., 2015, 2016; Stark et al., 2015), 也有影响不显著的(Lin et al., 2011; Fu et al., 2014).有研究发现放牧对土壤呼吸影响的大小与方向和放牧强度有关, 轻度放牧促进土壤呼吸, 中度和重度放牧则降低土壤呼吸速率(李玉强等, 2006).Wang等(2001)在内蒙古温带草原的研究表明, 自由放牧可增加土壤呼吸的年排放量, 且土壤呼吸与放牧强度呈正相关关系.然而, 也有研究认为, 荒漠草原放牧草地的土壤呼吸速率显著低于未放牧草地(阿木日吉日嘎拉等, 2013).本研究中, 过去不同的放牧强度对土壤呼吸速率的影响具有明显的年际变化, 表现为无显著影响和促进两种.放牧在总体上促进了土壤呼吸, 表现为放牧(LG、MG、HG)区3年平均土壤呼吸速率都高于禁牧区.两者研究结果截然不同, 这里存在两种可能, 一种是放牧增加土壤呼吸, 即使在停止放牧后, 放牧区的土壤呼吸速率仍然较高; 还有一种可能是放牧可能抑制土壤呼吸, 但停止放牧后, 土壤呼吸产生了超补偿, 使得土壤呼吸迅速恢复, 并超过对照区的, 而我们的研究结果属于哪种情况, 我们并不清楚.另外, 本研究是在多年放牧的草地上进行了小区围封, 实际上变成了围封草地, 围封后放牧的扰动已经去除, 草地的生物量和水分条件得到了一定程度的恢复, 从而引起土壤呼吸变化.已有研究也表明, 围封草地的土壤呼吸速率要明显高于放牧草地(贾丙瑞等, 2005), 而且放牧会导致全年的土壤呼吸速率比围封草地降低1/3 (马涛等, 2009).另外, 本研究持续3年测定土壤呼吸速率, 研究年限的不同对草地土壤呼吸的影响也存在差异(郭明英等, 2011). ...
放牧对典型草原土壤中几种生态因子影响的研究 1 2010
... 放牧对草地生态系统的影响是复杂、多方面且持久的, 放牧对草原植物(Cao et al., 2004; Gao et al., 2008)、微生物(Li et al., 2013)、凋落物(Klumpp et al., 2009)、土壤理化特性(Prieto et al., 2011; Li et al., 2013)等都存在着不同程度的影响, 而且放牧强度及其持续时间的不同也强烈影响着草地生态系统结构和功能.不同放牧强度的干扰将会迅速引起草地植被群落特征(王悦骅等, 2018)、土壤微生物数量、土壤酶和土壤理化性质如含水量、土壤有机质含量等方面的变化(高雪峰等, 2010), 而且在一定程度上影响水、热因子的作用效应, 进而导致土壤呼吸作用改变.已有的研究结果显示, 放牧不仅可以降低土壤呼吸速率(邓钰等, 2013; Hou et al., 2014), 也可以促进土壤呼吸(Chen et al., 2015, 2016; Stark et al., 2015), 也有影响不显著的(Lin et al., 2011; Fu et al., 2014).有研究发现放牧对土壤呼吸影响的大小与方向和放牧强度有关, 轻度放牧促进土壤呼吸, 中度和重度放牧则降低土壤呼吸速率(李玉强等, 2006).Wang等(2001)在内蒙古温带草原的研究表明, 自由放牧可增加土壤呼吸的年排放量, 且土壤呼吸与放牧强度呈正相关关系.然而, 也有研究认为, 荒漠草原放牧草地的土壤呼吸速率显著低于未放牧草地(阿木日吉日嘎拉等, 2013).本研究中, 过去不同的放牧强度对土壤呼吸速率的影响具有明显的年际变化, 表现为无显著影响和促进两种.放牧在总体上促进了土壤呼吸, 表现为放牧(LG、MG、HG)区3年平均土壤呼吸速率都高于禁牧区.两者研究结果截然不同, 这里存在两种可能, 一种是放牧增加土壤呼吸, 即使在停止放牧后, 放牧区的土壤呼吸速率仍然较高; 还有一种可能是放牧可能抑制土壤呼吸, 但停止放牧后, 土壤呼吸产生了超补偿, 使得土壤呼吸迅速恢复, 并超过对照区的, 而我们的研究结果属于哪种情况, 我们并不清楚.另外, 本研究是在多年放牧的草地上进行了小区围封, 实际上变成了围封草地, 围封后放牧的扰动已经去除, 草地的生物量和水分条件得到了一定程度的恢复, 从而引起土壤呼吸变化.已有研究也表明, 围封草地的土壤呼吸速率要明显高于放牧草地(贾丙瑞等, 2005), 而且放牧会导致全年的土壤呼吸速率比围封草地降低1/3 (马涛等, 2009).另外, 本研究持续3年测定土壤呼吸速率, 研究年限的不同对草地土壤呼吸的影响也存在差异(郭明英等, 2011). ...
Belowground net primary productivity and biomass allocation of a grassland in Inner Mongolia is affected by grazing intensity 2 2008
... 放牧是我国北方草地生态系统主要的利用方式.关于放牧对草地生态系统碳交换过程的影响一直以来受到学术界的广泛关注(Lin et al., 2011; Fu et al., 2014; Hou et al., 2014; Chen et al., 2015, 2016; Stark et al., 2015).当前的研究结果表明, 放牧对土壤呼吸的影响并没有一致的规律性, 放牧可能导致土壤呼吸速率升高、降低, 或者对土壤呼吸没有显著的影响(Chen et al., 2015, 2016; Stark et al., 2015).事实上, 放牧对土壤呼吸的影响可能是多种因素综合作用的结果.一方面, 放牧牲畜的啃食作用直接降低了植被冠层的光合作用(Bremer et al., 1998), 使得分配于根系的光合同化产物减少, 降低根系生物量(Cao et al., 2004; Gao et al., 2008; Klumpp et al., 2009), 减少了地表凋落物和根际分泌物(Bremer et al., 1998), 降低土壤微生物生物量和土壤酶活性(Prieto et al., 2011; Li et al., 2013), 最终导致土壤呼吸速率降低.另一方面, 放牧牲畜以粪尿形式归还有机物, 大量的活性有机质和氮输入为微生物呼吸作用提供更加丰富的底物, 激发微生物的活性和增加养分的循环速度(Fu et al., 2014), 加速根系生长, 增加根系分泌物, 进而促进土壤呼吸.上述因素在不同的时间和研究区域, 对土壤呼吸的贡献大小并不一致, 进而导致土壤呼吸速率对放牧响应的不确定性. ...
... 放牧对草地生态系统的影响是复杂、多方面且持久的, 放牧对草原植物(Cao et al., 2004; Gao et al., 2008)、微生物(Li et al., 2013)、凋落物(Klumpp et al., 2009)、土壤理化特性(Prieto et al., 2011; Li et al., 2013)等都存在着不同程度的影响, 而且放牧强度及其持续时间的不同也强烈影响着草地生态系统结构和功能.不同放牧强度的干扰将会迅速引起草地植被群落特征(王悦骅等, 2018)、土壤微生物数量、土壤酶和土壤理化性质如含水量、土壤有机质含量等方面的变化(高雪峰等, 2010), 而且在一定程度上影响水、热因子的作用效应, 进而导致土壤呼吸作用改变.已有的研究结果显示, 放牧不仅可以降低土壤呼吸速率(邓钰等, 2013; Hou et al., 2014), 也可以促进土壤呼吸(Chen et al., 2015, 2016; Stark et al., 2015), 也有影响不显著的(Lin et al., 2011; Fu et al., 2014).有研究发现放牧对土壤呼吸影响的大小与方向和放牧强度有关, 轻度放牧促进土壤呼吸, 中度和重度放牧则降低土壤呼吸速率(李玉强等, 2006).Wang等(2001)在内蒙古温带草原的研究表明, 自由放牧可增加土壤呼吸的年排放量, 且土壤呼吸与放牧强度呈正相关关系.然而, 也有研究认为, 荒漠草原放牧草地的土壤呼吸速率显著低于未放牧草地(阿木日吉日嘎拉等, 2013).本研究中, 过去不同的放牧强度对土壤呼吸速率的影响具有明显的年际变化, 表现为无显著影响和促进两种.放牧在总体上促进了土壤呼吸, 表现为放牧(LG、MG、HG)区3年平均土壤呼吸速率都高于禁牧区.两者研究结果截然不同, 这里存在两种可能, 一种是放牧增加土壤呼吸, 即使在停止放牧后, 放牧区的土壤呼吸速率仍然较高; 还有一种可能是放牧可能抑制土壤呼吸, 但停止放牧后, 土壤呼吸产生了超补偿, 使得土壤呼吸迅速恢复, 并超过对照区的, 而我们的研究结果属于哪种情况, 我们并不清楚.另外, 本研究是在多年放牧的草地上进行了小区围封, 实际上变成了围封草地, 围封后放牧的扰动已经去除, 草地的生物量和水分条件得到了一定程度的恢复, 从而引起土壤呼吸变化.已有研究也表明, 围封草地的土壤呼吸速率要明显高于放牧草地(贾丙瑞等, 2005), 而且放牧会导致全年的土壤呼吸速率比围封草地降低1/3 (马涛等, 2009).另外, 本研究持续3年测定土壤呼吸速率, 研究年限的不同对草地土壤呼吸的影响也存在差异(郭明英等, 2011). ...
Nitrogen deposition effects on soil organic matter chemistry are linked to variation in enzymes, ecosystems and size fractions 1 2008
Potential effects of climate change on DOC release from three different soil types on the Northern Pennines UK: Examination using field manipulation experiments 1 2008
... 荒漠草原地处温带干旱半干旱区域, 无论是植物生长, 还是土壤生态学过程, 都更加受到水分条件的限制.许多研究发现水分添加会显著增加土壤微生物生物量(Liu et al., 2009; Yan et al., 2010), 增强土壤有机质的矿化作用(Harrison et al., 2008; Liu et al., 2009), 这主要是由于土壤水分供给的增加导致营养元素扩散能力的提高, 使其更容易被植物和微生物利用(Sowerby et al., 2010; Reichmann et al., 2013).因此, 水分变化能够影响草原植物根系生长、土壤代谢活力、根系呼吸、土壤微生物群落构成以及微生物活性(Xu et al., 2004), 进而引起土壤呼吸强度的改变.在全球尺度上, 土壤呼吸强度与降水量呈正相关关系(Raich & Schlesinger, 1992; Zhou et al., 2014; Liu et al., 2016).而对于干旱和半干旱生态系统, 土壤湿度的影响尤其显著(Conant et al., 2000), 水分是导致土壤呼吸速率季节波动的重要因素和年际变化的主要驱动因子(Raich et al., 2002).降水可以在短时间内引起土壤呼吸作用的急剧增强(Birch, 1958), 而且这种对土壤呼吸的激发作用在干旱和半干旱地区变得更加显著(Sponseller, 2007).由降水格局改变引起的土壤呼吸变化效应可能远高于其他全球变化因子(增温、氮沉降、CO2升高)(Weltzin et al., 2003).本实验结果表明, 连续3年的加水处理(W和NW)使得土壤含水量明显增加, 对土壤呼吸产生了显著的促进作用, 而氮添加的有效性依赖于水分条件, 两者表现为协同作用.另外, 加水对土壤呼吸的促进作用大小也取决于自然降水的多少, 如在丰水年(2016年)影响不显著, 说明水分对该地区土壤呼吸的影响程度与总体上降水量高低有关, 在降水量较低的年份(平水或欠水年)能够促进土壤呼吸, 反之, 在降水量充沛的年份, 增加降水对土壤呼吸的促进作用有限, 甚至可能抑制土壤呼吸, 因为过量的水分添加会降低土壤通透性, 导致土壤氧气供应不足, 制约土壤呼吸.Liu等(2016)也发现土壤呼吸对增雨的响应随着环境湿润度的增加而减弱. ...
Nitrogen limitation in dryland ecosystems: Responses to geographical and temporal variation in precipitation 2 1999
... 氮和水分是草地生态系统生产力的两个主要限制性因子(Hooper & Johnson, 1999; Yan et al., 2011).鉴于草地退化常导致土壤养分缺乏, 氮添加作为一种重要手段被用于对退化草地的恢复和管理中(潘庆民等, 2005; 张璐等, 2009).氮添加可能影响生态系统光合作用和植物生长, 导致植物生产力的不同响应(Ren et al., 2017), 进而对土壤呼吸产生促进(Song et al., 2011)、抑制(Mo et al., 2007)或无显著影响(Peng et al., 2011; Qi et al., 2014; 李寅龙等, 2015) 3种不同的作用.水分作为干旱区草地生态系统过程重要的限制因子, 通常降水增加对土壤呼吸有显著促进作用(Sowerby et al., 2008; Yan et al., 2010, 2011), 而氮添加的效果常常会依赖于水分条件.刁励玮等(2018)统计分析了1990-2017年间发表的有关氮添加的研究论文结果, 发现单独施氮对地下生物量无显著影响, 氮水同时添加能显著提高地下生物量, 两者对生态系统生产力存在显著的交互作用(Li et al., 2011), 因此氮和水分两者可能共同影响生态系统的碳交换(Hooper & Johnson, 1999). ...
The response of grassland productivity, soil carbon content and soil respiration rates to different grazing regimes in a desert steppe in northern China 2 2014
... 放牧是我国北方草地生态系统主要的利用方式.关于放牧对草地生态系统碳交换过程的影响一直以来受到学术界的广泛关注(Lin et al., 2011; Fu et al., 2014; Hou et al., 2014; Chen et al., 2015, 2016; Stark et al., 2015).当前的研究结果表明, 放牧对土壤呼吸的影响并没有一致的规律性, 放牧可能导致土壤呼吸速率升高、降低, 或者对土壤呼吸没有显著的影响(Chen et al., 2015, 2016; Stark et al., 2015).事实上, 放牧对土壤呼吸的影响可能是多种因素综合作用的结果.一方面, 放牧牲畜的啃食作用直接降低了植被冠层的光合作用(Bremer et al., 1998), 使得分配于根系的光合同化产物减少, 降低根系生物量(Cao et al., 2004; Gao et al., 2008; Klumpp et al., 2009), 减少了地表凋落物和根际分泌物(Bremer et al., 1998), 降低土壤微生物生物量和土壤酶活性(Prieto et al., 2011; Li et al., 2013), 最终导致土壤呼吸速率降低.另一方面, 放牧牲畜以粪尿形式归还有机物, 大量的活性有机质和氮输入为微生物呼吸作用提供更加丰富的底物, 激发微生物的活性和增加养分的循环速度(Fu et al., 2014), 加速根系生长, 增加根系分泌物, 进而促进土壤呼吸.上述因素在不同的时间和研究区域, 对土壤呼吸的贡献大小并不一致, 进而导致土壤呼吸速率对放牧响应的不确定性. ...
... 放牧对草地生态系统的影响是复杂、多方面且持久的, 放牧对草原植物(Cao et al., 2004; Gao et al., 2008)、微生物(Li et al., 2013)、凋落物(Klumpp et al., 2009)、土壤理化特性(Prieto et al., 2011; Li et al., 2013)等都存在着不同程度的影响, 而且放牧强度及其持续时间的不同也强烈影响着草地生态系统结构和功能.不同放牧强度的干扰将会迅速引起草地植被群落特征(王悦骅等, 2018)、土壤微生物数量、土壤酶和土壤理化性质如含水量、土壤有机质含量等方面的变化(高雪峰等, 2010), 而且在一定程度上影响水、热因子的作用效应, 进而导致土壤呼吸作用改变.已有的研究结果显示, 放牧不仅可以降低土壤呼吸速率(邓钰等, 2013; Hou et al., 2014), 也可以促进土壤呼吸(Chen et al., 2015, 2016; Stark et al., 2015), 也有影响不显著的(Lin et al., 2011; Fu et al., 2014).有研究发现放牧对土壤呼吸影响的大小与方向和放牧强度有关, 轻度放牧促进土壤呼吸, 中度和重度放牧则降低土壤呼吸速率(李玉强等, 2006).Wang等(2001)在内蒙古温带草原的研究表明, 自由放牧可增加土壤呼吸的年排放量, 且土壤呼吸与放牧强度呈正相关关系.然而, 也有研究认为, 荒漠草原放牧草地的土壤呼吸速率显著低于未放牧草地(阿木日吉日嘎拉等, 2013).本研究中, 过去不同的放牧强度对土壤呼吸速率的影响具有明显的年际变化, 表现为无显著影响和促进两种.放牧在总体上促进了土壤呼吸, 表现为放牧(LG、MG、HG)区3年平均土壤呼吸速率都高于禁牧区.两者研究结果截然不同, 这里存在两种可能, 一种是放牧增加土壤呼吸, 即使在停止放牧后, 放牧区的土壤呼吸速率仍然较高; 还有一种可能是放牧可能抑制土壤呼吸, 但停止放牧后, 土壤呼吸产生了超补偿, 使得土壤呼吸迅速恢复, 并超过对照区的, 而我们的研究结果属于哪种情况, 我们并不清楚.另外, 本研究是在多年放牧的草地上进行了小区围封, 实际上变成了围封草地, 围封后放牧的扰动已经去除, 草地的生物量和水分条件得到了一定程度的恢复, 从而引起土壤呼吸变化.已有研究也表明, 围封草地的土壤呼吸速率要明显高于放牧草地(贾丙瑞等, 2005), 而且放牧会导致全年的土壤呼吸速率比围封草地降低1/3 (马涛等, 2009).另外, 本研究持续3年测定土壤呼吸速率, 研究年限的不同对草地土壤呼吸的影响也存在差异(郭明英等, 2011). ...
放牧与围栏羊草草原土壤呼吸作用及其影响因子 1 2005
... 放牧对草地生态系统的影响是复杂、多方面且持久的, 放牧对草原植物(Cao et al., 2004; Gao et al., 2008)、微生物(Li et al., 2013)、凋落物(Klumpp et al., 2009)、土壤理化特性(Prieto et al., 2011; Li et al., 2013)等都存在着不同程度的影响, 而且放牧强度及其持续时间的不同也强烈影响着草地生态系统结构和功能.不同放牧强度的干扰将会迅速引起草地植被群落特征(王悦骅等, 2018)、土壤微生物数量、土壤酶和土壤理化性质如含水量、土壤有机质含量等方面的变化(高雪峰等, 2010), 而且在一定程度上影响水、热因子的作用效应, 进而导致土壤呼吸作用改变.已有的研究结果显示, 放牧不仅可以降低土壤呼吸速率(邓钰等, 2013; Hou et al., 2014), 也可以促进土壤呼吸(Chen et al., 2015, 2016; Stark et al., 2015), 也有影响不显著的(Lin et al., 2011; Fu et al., 2014).有研究发现放牧对土壤呼吸影响的大小与方向和放牧强度有关, 轻度放牧促进土壤呼吸, 中度和重度放牧则降低土壤呼吸速率(李玉强等, 2006).Wang等(2001)在内蒙古温带草原的研究表明, 自由放牧可增加土壤呼吸的年排放量, 且土壤呼吸与放牧强度呈正相关关系.然而, 也有研究认为, 荒漠草原放牧草地的土壤呼吸速率显著低于未放牧草地(阿木日吉日嘎拉等, 2013).本研究中, 过去不同的放牧强度对土壤呼吸速率的影响具有明显的年际变化, 表现为无显著影响和促进两种.放牧在总体上促进了土壤呼吸, 表现为放牧(LG、MG、HG)区3年平均土壤呼吸速率都高于禁牧区.两者研究结果截然不同, 这里存在两种可能, 一种是放牧增加土壤呼吸, 即使在停止放牧后, 放牧区的土壤呼吸速率仍然较高; 还有一种可能是放牧可能抑制土壤呼吸, 但停止放牧后, 土壤呼吸产生了超补偿, 使得土壤呼吸迅速恢复, 并超过对照区的, 而我们的研究结果属于哪种情况, 我们并不清楚.另外, 本研究是在多年放牧的草地上进行了小区围封, 实际上变成了围封草地, 围封后放牧的扰动已经去除, 草地的生物量和水分条件得到了一定程度的恢复, 从而引起土壤呼吸变化.已有研究也表明, 围封草地的土壤呼吸速率要明显高于放牧草地(贾丙瑞等, 2005), 而且放牧会导致全年的土壤呼吸速率比围封草地降低1/3 (马涛等, 2009).另外, 本研究持续3年测定土壤呼吸速率, 研究年限的不同对草地土壤呼吸的影响也存在差异(郭明英等, 2011). ...
放牧与围栏羊草草原土壤呼吸作用及其影响因子 1 2005
... 放牧对草地生态系统的影响是复杂、多方面且持久的, 放牧对草原植物(Cao et al., 2004; Gao et al., 2008)、微生物(Li et al., 2013)、凋落物(Klumpp et al., 2009)、土壤理化特性(Prieto et al., 2011; Li et al., 2013)等都存在着不同程度的影响, 而且放牧强度及其持续时间的不同也强烈影响着草地生态系统结构和功能.不同放牧强度的干扰将会迅速引起草地植被群落特征(王悦骅等, 2018)、土壤微生物数量、土壤酶和土壤理化性质如含水量、土壤有机质含量等方面的变化(高雪峰等, 2010), 而且在一定程度上影响水、热因子的作用效应, 进而导致土壤呼吸作用改变.已有的研究结果显示, 放牧不仅可以降低土壤呼吸速率(邓钰等, 2013; Hou et al., 2014), 也可以促进土壤呼吸(Chen et al., 2015, 2016; Stark et al., 2015), 也有影响不显著的(Lin et al., 2011; Fu et al., 2014).有研究发现放牧对土壤呼吸影响的大小与方向和放牧强度有关, 轻度放牧促进土壤呼吸, 中度和重度放牧则降低土壤呼吸速率(李玉强等, 2006).Wang等(2001)在内蒙古温带草原的研究表明, 自由放牧可增加土壤呼吸的年排放量, 且土壤呼吸与放牧强度呈正相关关系.然而, 也有研究认为, 荒漠草原放牧草地的土壤呼吸速率显著低于未放牧草地(阿木日吉日嘎拉等, 2013).本研究中, 过去不同的放牧强度对土壤呼吸速率的影响具有明显的年际变化, 表现为无显著影响和促进两种.放牧在总体上促进了土壤呼吸, 表现为放牧(LG、MG、HG)区3年平均土壤呼吸速率都高于禁牧区.两者研究结果截然不同, 这里存在两种可能, 一种是放牧增加土壤呼吸, 即使在停止放牧后, 放牧区的土壤呼吸速率仍然较高; 还有一种可能是放牧可能抑制土壤呼吸, 但停止放牧后, 土壤呼吸产生了超补偿, 使得土壤呼吸迅速恢复, 并超过对照区的, 而我们的研究结果属于哪种情况, 我们并不清楚.另外, 本研究是在多年放牧的草地上进行了小区围封, 实际上变成了围封草地, 围封后放牧的扰动已经去除, 草地的生物量和水分条件得到了一定程度的恢复, 从而引起土壤呼吸变化.已有研究也表明, 围封草地的土壤呼吸速率要明显高于放牧草地(贾丙瑞等, 2005), 而且放牧会导致全年的土壤呼吸速率比围封草地降低1/3 (马涛等, 2009).另外, 本研究持续3年测定土壤呼吸速率, 研究年限的不同对草地土壤呼吸的影响也存在差异(郭明英等, 2011). ...
Grazing triggers soil carbon loss by altering plant roots and their control on soil microbial community 2 2009
... 放牧是我国北方草地生态系统主要的利用方式.关于放牧对草地生态系统碳交换过程的影响一直以来受到学术界的广泛关注(Lin et al., 2011; Fu et al., 2014; Hou et al., 2014; Chen et al., 2015, 2016; Stark et al., 2015).当前的研究结果表明, 放牧对土壤呼吸的影响并没有一致的规律性, 放牧可能导致土壤呼吸速率升高、降低, 或者对土壤呼吸没有显著的影响(Chen et al., 2015, 2016; Stark et al., 2015).事实上, 放牧对土壤呼吸的影响可能是多种因素综合作用的结果.一方面, 放牧牲畜的啃食作用直接降低了植被冠层的光合作用(Bremer et al., 1998), 使得分配于根系的光合同化产物减少, 降低根系生物量(Cao et al., 2004; Gao et al., 2008; Klumpp et al., 2009), 减少了地表凋落物和根际分泌物(Bremer et al., 1998), 降低土壤微生物生物量和土壤酶活性(Prieto et al., 2011; Li et al., 2013), 最终导致土壤呼吸速率降低.另一方面, 放牧牲畜以粪尿形式归还有机物, 大量的活性有机质和氮输入为微生物呼吸作用提供更加丰富的底物, 激发微生物的活性和增加养分的循环速度(Fu et al., 2014), 加速根系生长, 增加根系分泌物, 进而促进土壤呼吸.上述因素在不同的时间和研究区域, 对土壤呼吸的贡献大小并不一致, 进而导致土壤呼吸速率对放牧响应的不确定性. ...
... 放牧对草地生态系统的影响是复杂、多方面且持久的, 放牧对草原植物(Cao et al., 2004; Gao et al., 2008)、微生物(Li et al., 2013)、凋落物(Klumpp et al., 2009)、土壤理化特性(Prieto et al., 2011; Li et al., 2013)等都存在着不同程度的影响, 而且放牧强度及其持续时间的不同也强烈影响着草地生态系统结构和功能.不同放牧强度的干扰将会迅速引起草地植被群落特征(王悦骅等, 2018)、土壤微生物数量、土壤酶和土壤理化性质如含水量、土壤有机质含量等方面的变化(高雪峰等, 2010), 而且在一定程度上影响水、热因子的作用效应, 进而导致土壤呼吸作用改变.已有的研究结果显示, 放牧不仅可以降低土壤呼吸速率(邓钰等, 2013; Hou et al., 2014), 也可以促进土壤呼吸(Chen et al., 2015, 2016; Stark et al., 2015), 也有影响不显著的(Lin et al., 2011; Fu et al., 2014).有研究发现放牧对土壤呼吸影响的大小与方向和放牧强度有关, 轻度放牧促进土壤呼吸, 中度和重度放牧则降低土壤呼吸速率(李玉强等, 2006).Wang等(2001)在内蒙古温带草原的研究表明, 自由放牧可增加土壤呼吸的年排放量, 且土壤呼吸与放牧强度呈正相关关系.然而, 也有研究认为, 荒漠草原放牧草地的土壤呼吸速率显著低于未放牧草地(阿木日吉日嘎拉等, 2013).本研究中, 过去不同的放牧强度对土壤呼吸速率的影响具有明显的年际变化, 表现为无显著影响和促进两种.放牧在总体上促进了土壤呼吸, 表现为放牧(LG、MG、HG)区3年平均土壤呼吸速率都高于禁牧区.两者研究结果截然不同, 这里存在两种可能, 一种是放牧增加土壤呼吸, 即使在停止放牧后, 放牧区的土壤呼吸速率仍然较高; 还有一种可能是放牧可能抑制土壤呼吸, 但停止放牧后, 土壤呼吸产生了超补偿, 使得土壤呼吸迅速恢复, 并超过对照区的, 而我们的研究结果属于哪种情况, 我们并不清楚.另外, 本研究是在多年放牧的草地上进行了小区围封, 实际上变成了围封草地, 围封后放牧的扰动已经去除, 草地的生物量和水分条件得到了一定程度的恢复, 从而引起土壤呼吸变化.已有研究也表明, 围封草地的土壤呼吸速率要明显高于放牧草地(贾丙瑞等, 2005), 而且放牧会导致全年的土壤呼吸速率比围封草地降低1/3 (马涛等, 2009).另外, 本研究持续3年测定土壤呼吸速率, 研究年限的不同对草地土壤呼吸的影响也存在差异(郭明英等, 2011). ...
Aspects of carbon and nitrogen cycling in soils of Bornh?ved Lake district. II. Modelling the influence of temperature increase on soil respiration and organic carbon content in arable soils under different managements 1 1997
Above and belowground net primary productivity of grassland influenced by supplemental water and nitrogen in Inner Mongolia 1 2011
... 氮和水分是草地生态系统生产力的两个主要限制性因子(Hooper & Johnson, 1999; Yan et al., 2011).鉴于草地退化常导致土壤养分缺乏, 氮添加作为一种重要手段被用于对退化草地的恢复和管理中(潘庆民等, 2005; 张璐等, 2009).氮添加可能影响生态系统光合作用和植物生长, 导致植物生产力的不同响应(Ren et al., 2017), 进而对土壤呼吸产生促进(Song et al., 2011)、抑制(Mo et al., 2007)或无显著影响(Peng et al., 2011; Qi et al., 2014; 李寅龙等, 2015) 3种不同的作用.水分作为干旱区草地生态系统过程重要的限制因子, 通常降水增加对土壤呼吸有显著促进作用(Sowerby et al., 2008; Yan et al., 2010, 2011), 而氮添加的效果常常会依赖于水分条件.刁励玮等(2018)统计分析了1990-2017年间发表的有关氮添加的研究论文结果, 发现单独施氮对地下生物量无显著影响, 氮水同时添加能显著提高地下生物量, 两者对生态系统生产力存在显著的交互作用(Li et al., 2011), 因此氮和水分两者可能共同影响生态系统的碳交换(Hooper & Johnson, 1999). ...
Grazing exclusion alters soil microbial respiration, root respiration and the soil carbon balance in grasslands of the Loess Plateau, northern China 3 2013
... 放牧是我国北方草地生态系统主要的利用方式.关于放牧对草地生态系统碳交换过程的影响一直以来受到学术界的广泛关注(Lin et al., 2011; Fu et al., 2014; Hou et al., 2014; Chen et al., 2015, 2016; Stark et al., 2015).当前的研究结果表明, 放牧对土壤呼吸的影响并没有一致的规律性, 放牧可能导致土壤呼吸速率升高、降低, 或者对土壤呼吸没有显著的影响(Chen et al., 2015, 2016; Stark et al., 2015).事实上, 放牧对土壤呼吸的影响可能是多种因素综合作用的结果.一方面, 放牧牲畜的啃食作用直接降低了植被冠层的光合作用(Bremer et al., 1998), 使得分配于根系的光合同化产物减少, 降低根系生物量(Cao et al., 2004; Gao et al., 2008; Klumpp et al., 2009), 减少了地表凋落物和根际分泌物(Bremer et al., 1998), 降低土壤微生物生物量和土壤酶活性(Prieto et al., 2011; Li et al., 2013), 最终导致土壤呼吸速率降低.另一方面, 放牧牲畜以粪尿形式归还有机物, 大量的活性有机质和氮输入为微生物呼吸作用提供更加丰富的底物, 激发微生物的活性和增加养分的循环速度(Fu et al., 2014), 加速根系生长, 增加根系分泌物, 进而促进土壤呼吸.上述因素在不同的时间和研究区域, 对土壤呼吸的贡献大小并不一致, 进而导致土壤呼吸速率对放牧响应的不确定性. ...
... 放牧对草地生态系统的影响是复杂、多方面且持久的, 放牧对草原植物(Cao et al., 2004; Gao et al., 2008)、微生物(Li et al., 2013)、凋落物(Klumpp et al., 2009)、土壤理化特性(Prieto et al., 2011; Li et al., 2013)等都存在着不同程度的影响, 而且放牧强度及其持续时间的不同也强烈影响着草地生态系统结构和功能.不同放牧强度的干扰将会迅速引起草地植被群落特征(王悦骅等, 2018)、土壤微生物数量、土壤酶和土壤理化性质如含水量、土壤有机质含量等方面的变化(高雪峰等, 2010), 而且在一定程度上影响水、热因子的作用效应, 进而导致土壤呼吸作用改变.已有的研究结果显示, 放牧不仅可以降低土壤呼吸速率(邓钰等, 2013; Hou et al., 2014), 也可以促进土壤呼吸(Chen et al., 2015, 2016; Stark et al., 2015), 也有影响不显著的(Lin et al., 2011; Fu et al., 2014).有研究发现放牧对土壤呼吸影响的大小与方向和放牧强度有关, 轻度放牧促进土壤呼吸, 中度和重度放牧则降低土壤呼吸速率(李玉强等, 2006).Wang等(2001)在内蒙古温带草原的研究表明, 自由放牧可增加土壤呼吸的年排放量, 且土壤呼吸与放牧强度呈正相关关系.然而, 也有研究认为, 荒漠草原放牧草地的土壤呼吸速率显著低于未放牧草地(阿木日吉日嘎拉等, 2013).本研究中, 过去不同的放牧强度对土壤呼吸速率的影响具有明显的年际变化, 表现为无显著影响和促进两种.放牧在总体上促进了土壤呼吸, 表现为放牧(LG、MG、HG)区3年平均土壤呼吸速率都高于禁牧区.两者研究结果截然不同, 这里存在两种可能, 一种是放牧增加土壤呼吸, 即使在停止放牧后, 放牧区的土壤呼吸速率仍然较高; 还有一种可能是放牧可能抑制土壤呼吸, 但停止放牧后, 土壤呼吸产生了超补偿, 使得土壤呼吸迅速恢复, 并超过对照区的, 而我们的研究结果属于哪种情况, 我们并不清楚.另外, 本研究是在多年放牧的草地上进行了小区围封, 实际上变成了围封草地, 围封后放牧的扰动已经去除, 草地的生物量和水分条件得到了一定程度的恢复, 从而引起土壤呼吸变化.已有研究也表明, 围封草地的土壤呼吸速率要明显高于放牧草地(贾丙瑞等, 2005), 而且放牧会导致全年的土壤呼吸速率比围封草地降低1/3 (马涛等, 2009).另外, 本研究持续3年测定土壤呼吸速率, 研究年限的不同对草地土壤呼吸的影响也存在差异(郭明英等, 2011). ...
... ; Li et al., 2013)等都存在着不同程度的影响, 而且放牧强度及其持续时间的不同也强烈影响着草地生态系统结构和功能.不同放牧强度的干扰将会迅速引起草地植被群落特征(王悦骅等, 2018)、土壤微生物数量、土壤酶和土壤理化性质如含水量、土壤有机质含量等方面的变化(高雪峰等, 2010), 而且在一定程度上影响水、热因子的作用效应, 进而导致土壤呼吸作用改变.已有的研究结果显示, 放牧不仅可以降低土壤呼吸速率(邓钰等, 2013; Hou et al., 2014), 也可以促进土壤呼吸(Chen et al., 2015, 2016; Stark et al., 2015), 也有影响不显著的(Lin et al., 2011; Fu et al., 2014).有研究发现放牧对土壤呼吸影响的大小与方向和放牧强度有关, 轻度放牧促进土壤呼吸, 中度和重度放牧则降低土壤呼吸速率(李玉强等, 2006).Wang等(2001)在内蒙古温带草原的研究表明, 自由放牧可增加土壤呼吸的年排放量, 且土壤呼吸与放牧强度呈正相关关系.然而, 也有研究认为, 荒漠草原放牧草地的土壤呼吸速率显著低于未放牧草地(阿木日吉日嘎拉等, 2013).本研究中, 过去不同的放牧强度对土壤呼吸速率的影响具有明显的年际变化, 表现为无显著影响和促进两种.放牧在总体上促进了土壤呼吸, 表现为放牧(LG、MG、HG)区3年平均土壤呼吸速率都高于禁牧区.两者研究结果截然不同, 这里存在两种可能, 一种是放牧增加土壤呼吸, 即使在停止放牧后, 放牧区的土壤呼吸速率仍然较高; 还有一种可能是放牧可能抑制土壤呼吸, 但停止放牧后, 土壤呼吸产生了超补偿, 使得土壤呼吸迅速恢复, 并超过对照区的, 而我们的研究结果属于哪种情况, 我们并不清楚.另外, 本研究是在多年放牧的草地上进行了小区围封, 实际上变成了围封草地, 围封后放牧的扰动已经去除, 草地的生物量和水分条件得到了一定程度的恢复, 从而引起土壤呼吸变化.已有研究也表明, 围封草地的土壤呼吸速率要明显高于放牧草地(贾丙瑞等, 2005), 而且放牧会导致全年的土壤呼吸速率比围封草地降低1/3 (马涛等, 2009).另外, 本研究持续3年测定土壤呼吸速率, 研究年限的不同对草地土壤呼吸的影响也存在差异(郭明英等, 2011). ...
... 氮和水分是草地生态系统生产力的两个主要限制性因子(Hooper & Johnson, 1999; Yan et al., 2011).鉴于草地退化常导致土壤养分缺乏, 氮添加作为一种重要手段被用于对退化草地的恢复和管理中(潘庆民等, 2005; 张璐等, 2009).氮添加可能影响生态系统光合作用和植物生长, 导致植物生产力的不同响应(Ren et al., 2017), 进而对土壤呼吸产生促进(Song et al., 2011)、抑制(Mo et al., 2007)或无显著影响(Peng et al., 2011; Qi et al., 2014; 李寅龙等, 2015) 3种不同的作用.水分作为干旱区草地生态系统过程重要的限制因子, 通常降水增加对土壤呼吸有显著促进作用(Sowerby et al., 2008; Yan et al., 2010, 2011), 而氮添加的效果常常会依赖于水分条件.刁励玮等(2018)统计分析了1990-2017年间发表的有关氮添加的研究论文结果, 发现单独施氮对地下生物量无显著影响, 氮水同时添加能显著提高地下生物量, 两者对生态系统生产力存在显著的交互作用(Li et al., 2011), 因此氮和水分两者可能共同影响生态系统的碳交换(Hooper & Johnson, 1999). ...
... 氮和水分是草地生态系统生产力的两个主要限制性因子(Hooper & Johnson, 1999; Yan et al., 2011).鉴于草地退化常导致土壤养分缺乏, 氮添加作为一种重要手段被用于对退化草地的恢复和管理中(潘庆民等, 2005; 张璐等, 2009).氮添加可能影响生态系统光合作用和植物生长, 导致植物生产力的不同响应(Ren et al., 2017), 进而对土壤呼吸产生促进(Song et al., 2011)、抑制(Mo et al., 2007)或无显著影响(Peng et al., 2011; Qi et al., 2014; 李寅龙等, 2015) 3种不同的作用.水分作为干旱区草地生态系统过程重要的限制因子, 通常降水增加对土壤呼吸有显著促进作用(Sowerby et al., 2008; Yan et al., 2010, 2011), 而氮添加的效果常常会依赖于水分条件.刁励玮等(2018)统计分析了1990-2017年间发表的有关氮添加的研究论文结果, 发现单独施氮对地下生物量无显著影响, 氮水同时添加能显著提高地下生物量, 两者对生态系统生产力存在显著的交互作用(Li et al., 2011), 因此氮和水分两者可能共同影响生态系统的碳交换(Hooper & Johnson, 1999). ...
... 放牧对草地生态系统的影响是复杂、多方面且持久的, 放牧对草原植物(Cao et al., 2004; Gao et al., 2008)、微生物(Li et al., 2013)、凋落物(Klumpp et al., 2009)、土壤理化特性(Prieto et al., 2011; Li et al., 2013)等都存在着不同程度的影响, 而且放牧强度及其持续时间的不同也强烈影响着草地生态系统结构和功能.不同放牧强度的干扰将会迅速引起草地植被群落特征(王悦骅等, 2018)、土壤微生物数量、土壤酶和土壤理化性质如含水量、土壤有机质含量等方面的变化(高雪峰等, 2010), 而且在一定程度上影响水、热因子的作用效应, 进而导致土壤呼吸作用改变.已有的研究结果显示, 放牧不仅可以降低土壤呼吸速率(邓钰等, 2013; Hou et al., 2014), 也可以促进土壤呼吸(Chen et al., 2015, 2016; Stark et al., 2015), 也有影响不显著的(Lin et al., 2011; Fu et al., 2014).有研究发现放牧对土壤呼吸影响的大小与方向和放牧强度有关, 轻度放牧促进土壤呼吸, 中度和重度放牧则降低土壤呼吸速率(李玉强等, 2006).Wang等(2001)在内蒙古温带草原的研究表明, 自由放牧可增加土壤呼吸的年排放量, 且土壤呼吸与放牧强度呈正相关关系.然而, 也有研究认为, 荒漠草原放牧草地的土壤呼吸速率显著低于未放牧草地(阿木日吉日嘎拉等, 2013).本研究中, 过去不同的放牧强度对土壤呼吸速率的影响具有明显的年际变化, 表现为无显著影响和促进两种.放牧在总体上促进了土壤呼吸, 表现为放牧(LG、MG、HG)区3年平均土壤呼吸速率都高于禁牧区.两者研究结果截然不同, 这里存在两种可能, 一种是放牧增加土壤呼吸, 即使在停止放牧后, 放牧区的土壤呼吸速率仍然较高; 还有一种可能是放牧可能抑制土壤呼吸, 但停止放牧后, 土壤呼吸产生了超补偿, 使得土壤呼吸迅速恢复, 并超过对照区的, 而我们的研究结果属于哪种情况, 我们并不清楚.另外, 本研究是在多年放牧的草地上进行了小区围封, 实际上变成了围封草地, 围封后放牧的扰动已经去除, 草地的生物量和水分条件得到了一定程度的恢复, 从而引起土壤呼吸变化.已有研究也表明, 围封草地的土壤呼吸速率要明显高于放牧草地(贾丙瑞等, 2005), 而且放牧会导致全年的土壤呼吸速率比围封草地降低1/3 (马涛等, 2009).另外, 本研究持续3年测定土壤呼吸速率, 研究年限的不同对草地土壤呼吸的影响也存在差异(郭明英等, 2011). ...
不同强度放牧后自然恢复的沙质草地土壤呼吸、碳平衡与碳储量 1 2006
... 放牧对草地生态系统的影响是复杂、多方面且持久的, 放牧对草原植物(Cao et al., 2004; Gao et al., 2008)、微生物(Li et al., 2013)、凋落物(Klumpp et al., 2009)、土壤理化特性(Prieto et al., 2011; Li et al., 2013)等都存在着不同程度的影响, 而且放牧强度及其持续时间的不同也强烈影响着草地生态系统结构和功能.不同放牧强度的干扰将会迅速引起草地植被群落特征(王悦骅等, 2018)、土壤微生物数量、土壤酶和土壤理化性质如含水量、土壤有机质含量等方面的变化(高雪峰等, 2010), 而且在一定程度上影响水、热因子的作用效应, 进而导致土壤呼吸作用改变.已有的研究结果显示, 放牧不仅可以降低土壤呼吸速率(邓钰等, 2013; Hou et al., 2014), 也可以促进土壤呼吸(Chen et al., 2015, 2016; Stark et al., 2015), 也有影响不显著的(Lin et al., 2011; Fu et al., 2014).有研究发现放牧对土壤呼吸影响的大小与方向和放牧强度有关, 轻度放牧促进土壤呼吸, 中度和重度放牧则降低土壤呼吸速率(李玉强等, 2006).Wang等(2001)在内蒙古温带草原的研究表明, 自由放牧可增加土壤呼吸的年排放量, 且土壤呼吸与放牧强度呈正相关关系.然而, 也有研究认为, 荒漠草原放牧草地的土壤呼吸速率显著低于未放牧草地(阿木日吉日嘎拉等, 2013).本研究中, 过去不同的放牧强度对土壤呼吸速率的影响具有明显的年际变化, 表现为无显著影响和促进两种.放牧在总体上促进了土壤呼吸, 表现为放牧(LG、MG、HG)区3年平均土壤呼吸速率都高于禁牧区.两者研究结果截然不同, 这里存在两种可能, 一种是放牧增加土壤呼吸, 即使在停止放牧后, 放牧区的土壤呼吸速率仍然较高; 还有一种可能是放牧可能抑制土壤呼吸, 但停止放牧后, 土壤呼吸产生了超补偿, 使得土壤呼吸迅速恢复, 并超过对照区的, 而我们的研究结果属于哪种情况, 我们并不清楚.另外, 本研究是在多年放牧的草地上进行了小区围封, 实际上变成了围封草地, 围封后放牧的扰动已经去除, 草地的生物量和水分条件得到了一定程度的恢复, 从而引起土壤呼吸变化.已有研究也表明, 围封草地的土壤呼吸速率要明显高于放牧草地(贾丙瑞等, 2005), 而且放牧会导致全年的土壤呼吸速率比围封草地降低1/3 (马涛等, 2009).另外, 本研究持续3年测定土壤呼吸速率, 研究年限的不同对草地土壤呼吸的影响也存在差异(郭明英等, 2011). ...
Response of ecosystem respiration to warming and grazing during the growing seasons in the alpine meadow on the Tibetan Plateau 2 2011
... 放牧是我国北方草地生态系统主要的利用方式.关于放牧对草地生态系统碳交换过程的影响一直以来受到学术界的广泛关注(Lin et al., 2011; Fu et al., 2014; Hou et al., 2014; Chen et al., 2015, 2016; Stark et al., 2015).当前的研究结果表明, 放牧对土壤呼吸的影响并没有一致的规律性, 放牧可能导致土壤呼吸速率升高、降低, 或者对土壤呼吸没有显著的影响(Chen et al., 2015, 2016; Stark et al., 2015).事实上, 放牧对土壤呼吸的影响可能是多种因素综合作用的结果.一方面, 放牧牲畜的啃食作用直接降低了植被冠层的光合作用(Bremer et al., 1998), 使得分配于根系的光合同化产物减少, 降低根系生物量(Cao et al., 2004; Gao et al., 2008; Klumpp et al., 2009), 减少了地表凋落物和根际分泌物(Bremer et al., 1998), 降低土壤微生物生物量和土壤酶活性(Prieto et al., 2011; Li et al., 2013), 最终导致土壤呼吸速率降低.另一方面, 放牧牲畜以粪尿形式归还有机物, 大量的活性有机质和氮输入为微生物呼吸作用提供更加丰富的底物, 激发微生物的活性和增加养分的循环速度(Fu et al., 2014), 加速根系生长, 增加根系分泌物, 进而促进土壤呼吸.上述因素在不同的时间和研究区域, 对土壤呼吸的贡献大小并不一致, 进而导致土壤呼吸速率对放牧响应的不确定性. ...
... 放牧对草地生态系统的影响是复杂、多方面且持久的, 放牧对草原植物(Cao et al., 2004; Gao et al., 2008)、微生物(Li et al., 2013)、凋落物(Klumpp et al., 2009)、土壤理化特性(Prieto et al., 2011; Li et al., 2013)等都存在着不同程度的影响, 而且放牧强度及其持续时间的不同也强烈影响着草地生态系统结构和功能.不同放牧强度的干扰将会迅速引起草地植被群落特征(王悦骅等, 2018)、土壤微生物数量、土壤酶和土壤理化性质如含水量、土壤有机质含量等方面的变化(高雪峰等, 2010), 而且在一定程度上影响水、热因子的作用效应, 进而导致土壤呼吸作用改变.已有的研究结果显示, 放牧不仅可以降低土壤呼吸速率(邓钰等, 2013; Hou et al., 2014), 也可以促进土壤呼吸(Chen et al., 2015, 2016; Stark et al., 2015), 也有影响不显著的(Lin et al., 2011; Fu et al., 2014).有研究发现放牧对土壤呼吸影响的大小与方向和放牧强度有关, 轻度放牧促进土壤呼吸, 中度和重度放牧则降低土壤呼吸速率(李玉强等, 2006).Wang等(2001)在内蒙古温带草原的研究表明, 自由放牧可增加土壤呼吸的年排放量, 且土壤呼吸与放牧强度呈正相关关系.然而, 也有研究认为, 荒漠草原放牧草地的土壤呼吸速率显著低于未放牧草地(阿木日吉日嘎拉等, 2013).本研究中, 过去不同的放牧强度对土壤呼吸速率的影响具有明显的年际变化, 表现为无显著影响和促进两种.放牧在总体上促进了土壤呼吸, 表现为放牧(LG、MG、HG)区3年平均土壤呼吸速率都高于禁牧区.两者研究结果截然不同, 这里存在两种可能, 一种是放牧增加土壤呼吸, 即使在停止放牧后, 放牧区的土壤呼吸速率仍然较高; 还有一种可能是放牧可能抑制土壤呼吸, 但停止放牧后, 土壤呼吸产生了超补偿, 使得土壤呼吸迅速恢复, 并超过对照区的, 而我们的研究结果属于哪种情况, 我们并不清楚.另外, 本研究是在多年放牧的草地上进行了小区围封, 实际上变成了围封草地, 围封后放牧的扰动已经去除, 草地的生物量和水分条件得到了一定程度的恢复, 从而引起土壤呼吸变化.已有研究也表明, 围封草地的土壤呼吸速率要明显高于放牧草地(贾丙瑞等, 2005), 而且放牧会导致全年的土壤呼吸速率比围封草地降低1/3 (马涛等, 2009).另外, 本研究持续3年测定土壤呼吸速率, 研究年限的不同对草地土壤呼吸的影响也存在差异(郭明英等, 2011). ...
A cross-biome synthesis of soil respiration and its determinants under simulated precipitation changes 2 2016
... 荒漠草原地处温带干旱半干旱区域, 无论是植物生长, 还是土壤生态学过程, 都更加受到水分条件的限制.许多研究发现水分添加会显著增加土壤微生物生物量(Liu et al., 2009; Yan et al., 2010), 增强土壤有机质的矿化作用(Harrison et al., 2008; Liu et al., 2009), 这主要是由于土壤水分供给的增加导致营养元素扩散能力的提高, 使其更容易被植物和微生物利用(Sowerby et al., 2010; Reichmann et al., 2013).因此, 水分变化能够影响草原植物根系生长、土壤代谢活力、根系呼吸、土壤微生物群落构成以及微生物活性(Xu et al., 2004), 进而引起土壤呼吸强度的改变.在全球尺度上, 土壤呼吸强度与降水量呈正相关关系(Raich & Schlesinger, 1992; Zhou et al., 2014; Liu et al., 2016).而对于干旱和半干旱生态系统, 土壤湿度的影响尤其显著(Conant et al., 2000), 水分是导致土壤呼吸速率季节波动的重要因素和年际变化的主要驱动因子(Raich et al., 2002).降水可以在短时间内引起土壤呼吸作用的急剧增强(Birch, 1958), 而且这种对土壤呼吸的激发作用在干旱和半干旱地区变得更加显著(Sponseller, 2007).由降水格局改变引起的土壤呼吸变化效应可能远高于其他全球变化因子(增温、氮沉降、CO2升高)(Weltzin et al., 2003).本实验结果表明, 连续3年的加水处理(W和NW)使得土壤含水量明显增加, 对土壤呼吸产生了显著的促进作用, 而氮添加的有效性依赖于水分条件, 两者表现为协同作用.另外, 加水对土壤呼吸的促进作用大小也取决于自然降水的多少, 如在丰水年(2016年)影响不显著, 说明水分对该地区土壤呼吸的影响程度与总体上降水量高低有关, 在降水量较低的年份(平水或欠水年)能够促进土壤呼吸, 反之, 在降水量充沛的年份, 增加降水对土壤呼吸的促进作用有限, 甚至可能抑制土壤呼吸, 因为过量的水分添加会降低土壤通透性, 导致土壤氧气供应不足, 制约土壤呼吸.Liu等(2016)也发现土壤呼吸对增雨的响应随着环境湿润度的增加而减弱. ...
... 放牧对草地生态系统的影响是复杂、多方面且持久的, 放牧对草原植物(Cao et al., 2004; Gao et al., 2008)、微生物(Li et al., 2013)、凋落物(Klumpp et al., 2009)、土壤理化特性(Prieto et al., 2011; Li et al., 2013)等都存在着不同程度的影响, 而且放牧强度及其持续时间的不同也强烈影响着草地生态系统结构和功能.不同放牧强度的干扰将会迅速引起草地植被群落特征(王悦骅等, 2018)、土壤微生物数量、土壤酶和土壤理化性质如含水量、土壤有机质含量等方面的变化(高雪峰等, 2010), 而且在一定程度上影响水、热因子的作用效应, 进而导致土壤呼吸作用改变.已有的研究结果显示, 放牧不仅可以降低土壤呼吸速率(邓钰等, 2013; Hou et al., 2014), 也可以促进土壤呼吸(Chen et al., 2015, 2016; Stark et al., 2015), 也有影响不显著的(Lin et al., 2011; Fu et al., 2014).有研究发现放牧对土壤呼吸影响的大小与方向和放牧强度有关, 轻度放牧促进土壤呼吸, 中度和重度放牧则降低土壤呼吸速率(李玉强等, 2006).Wang等(2001)在内蒙古温带草原的研究表明, 自由放牧可增加土壤呼吸的年排放量, 且土壤呼吸与放牧强度呈正相关关系.然而, 也有研究认为, 荒漠草原放牧草地的土壤呼吸速率显著低于未放牧草地(阿木日吉日嘎拉等, 2013).本研究中, 过去不同的放牧强度对土壤呼吸速率的影响具有明显的年际变化, 表现为无显著影响和促进两种.放牧在总体上促进了土壤呼吸, 表现为放牧(LG、MG、HG)区3年平均土壤呼吸速率都高于禁牧区.两者研究结果截然不同, 这里存在两种可能, 一种是放牧增加土壤呼吸, 即使在停止放牧后, 放牧区的土壤呼吸速率仍然较高; 还有一种可能是放牧可能抑制土壤呼吸, 但停止放牧后, 土壤呼吸产生了超补偿, 使得土壤呼吸迅速恢复, 并超过对照区的, 而我们的研究结果属于哪种情况, 我们并不清楚.另外, 本研究是在多年放牧的草地上进行了小区围封, 实际上变成了围封草地, 围封后放牧的扰动已经去除, 草地的生物量和水分条件得到了一定程度的恢复, 从而引起土壤呼吸变化.已有研究也表明, 围封草地的土壤呼吸速率要明显高于放牧草地(贾丙瑞等, 2005), 而且放牧会导致全年的土壤呼吸速率比围封草地降低1/3 (马涛等, 2009).另外, 本研究持续3年测定土壤呼吸速率, 研究年限的不同对草地土壤呼吸的影响也存在差异(郭明英等, 2011). ...
放牧对内蒙古羊草群落土壤呼吸的影响 1 2009
... 放牧对草地生态系统的影响是复杂、多方面且持久的, 放牧对草原植物(Cao et al., 2004; Gao et al., 2008)、微生物(Li et al., 2013)、凋落物(Klumpp et al., 2009)、土壤理化特性(Prieto et al., 2011; Li et al., 2013)等都存在着不同程度的影响, 而且放牧强度及其持续时间的不同也强烈影响着草地生态系统结构和功能.不同放牧强度的干扰将会迅速引起草地植被群落特征(王悦骅等, 2018)、土壤微生物数量、土壤酶和土壤理化性质如含水量、土壤有机质含量等方面的变化(高雪峰等, 2010), 而且在一定程度上影响水、热因子的作用效应, 进而导致土壤呼吸作用改变.已有的研究结果显示, 放牧不仅可以降低土壤呼吸速率(邓钰等, 2013; Hou et al., 2014), 也可以促进土壤呼吸(Chen et al., 2015, 2016; Stark et al., 2015), 也有影响不显著的(Lin et al., 2011; Fu et al., 2014).有研究发现放牧对土壤呼吸影响的大小与方向和放牧强度有关, 轻度放牧促进土壤呼吸, 中度和重度放牧则降低土壤呼吸速率(李玉强等, 2006).Wang等(2001)在内蒙古温带草原的研究表明, 自由放牧可增加土壤呼吸的年排放量, 且土壤呼吸与放牧强度呈正相关关系.然而, 也有研究认为, 荒漠草原放牧草地的土壤呼吸速率显著低于未放牧草地(阿木日吉日嘎拉等, 2013).本研究中, 过去不同的放牧强度对土壤呼吸速率的影响具有明显的年际变化, 表现为无显著影响和促进两种.放牧在总体上促进了土壤呼吸, 表现为放牧(LG、MG、HG)区3年平均土壤呼吸速率都高于禁牧区.两者研究结果截然不同, 这里存在两种可能, 一种是放牧增加土壤呼吸, 即使在停止放牧后, 放牧区的土壤呼吸速率仍然较高; 还有一种可能是放牧可能抑制土壤呼吸, 但停止放牧后, 土壤呼吸产生了超补偿, 使得土壤呼吸迅速恢复, 并超过对照区的, 而我们的研究结果属于哪种情况, 我们并不清楚.另外, 本研究是在多年放牧的草地上进行了小区围封, 实际上变成了围封草地, 围封后放牧的扰动已经去除, 草地的生物量和水分条件得到了一定程度的恢复, 从而引起土壤呼吸变化.已有研究也表明, 围封草地的土壤呼吸速率要明显高于放牧草地(贾丙瑞等, 2005), 而且放牧会导致全年的土壤呼吸速率比围封草地降低1/3 (马涛等, 2009).另外, 本研究持续3年测定土壤呼吸速率, 研究年限的不同对草地土壤呼吸的影响也存在差异(郭明英等, 2011). ...
Nitrogen addition reduces soil respiration in a mature tropical forest in southern China 2 2007
... 氮和水分是草地生态系统生产力的两个主要限制性因子(Hooper & Johnson, 1999; Yan et al., 2011).鉴于草地退化常导致土壤养分缺乏, 氮添加作为一种重要手段被用于对退化草地的恢复和管理中(潘庆民等, 2005; 张璐等, 2009).氮添加可能影响生态系统光合作用和植物生长, 导致植物生产力的不同响应(Ren et al., 2017), 进而对土壤呼吸产生促进(Song et al., 2011)、抑制(Mo et al., 2007)或无显著影响(Peng et al., 2011; Qi et al., 2014; 李寅龙等, 2015) 3种不同的作用.水分作为干旱区草地生态系统过程重要的限制因子, 通常降水增加对土壤呼吸有显著促进作用(Sowerby et al., 2008; Yan et al., 2010, 2011), 而氮添加的效果常常会依赖于水分条件.刁励玮等(2018)统计分析了1990-2017年间发表的有关氮添加的研究论文结果, 发现单独施氮对地下生物量无显著影响, 氮水同时添加能显著提高地下生物量, 两者对生态系统生产力存在显著的交互作用(Li et al., 2011), 因此氮和水分两者可能共同影响生态系统的碳交换(Hooper & Johnson, 1999). ...
... 氮和水分是草地生态系统生产力的两个主要限制性因子(Hooper & Johnson, 1999; Yan et al., 2011).鉴于草地退化常导致土壤养分缺乏, 氮添加作为一种重要手段被用于对退化草地的恢复和管理中(潘庆民等, 2005; 张璐等, 2009).氮添加可能影响生态系统光合作用和植物生长, 导致植物生产力的不同响应(Ren et al., 2017), 进而对土壤呼吸产生促进(Song et al., 2011)、抑制(Mo et al., 2007)或无显著影响(Peng et al., 2011; Qi et al., 2014; 李寅龙等, 2015) 3种不同的作用.水分作为干旱区草地生态系统过程重要的限制因子, 通常降水增加对土壤呼吸有显著促进作用(Sowerby et al., 2008; Yan et al., 2010, 2011), 而氮添加的效果常常会依赖于水分条件.刁励玮等(2018)统计分析了1990-2017年间发表的有关氮添加的研究论文结果, 发现单独施氮对地下生物量无显著影响, 氮水同时添加能显著提高地下生物量, 两者对生态系统生产力存在显著的交互作用(Li et al., 2011), 因此氮和水分两者可能共同影响生态系统的碳交换(Hooper & Johnson, 1999). ...
氮素对内蒙古典型草原羊草种群的影响 1 2005
... 氮和水分是草地生态系统生产力的两个主要限制性因子(Hooper & Johnson, 1999; Yan et al., 2011).鉴于草地退化常导致土壤养分缺乏, 氮添加作为一种重要手段被用于对退化草地的恢复和管理中(潘庆民等, 2005; 张璐等, 2009).氮添加可能影响生态系统光合作用和植物生长, 导致植物生产力的不同响应(Ren et al., 2017), 进而对土壤呼吸产生促进(Song et al., 2011)、抑制(Mo et al., 2007)或无显著影响(Peng et al., 2011; Qi et al., 2014; 李寅龙等, 2015) 3种不同的作用.水分作为干旱区草地生态系统过程重要的限制因子, 通常降水增加对土壤呼吸有显著促进作用(Sowerby et al., 2008; Yan et al., 2010, 2011), 而氮添加的效果常常会依赖于水分条件.刁励玮等(2018)统计分析了1990-2017年间发表的有关氮添加的研究论文结果, 发现单独施氮对地下生物量无显著影响, 氮水同时添加能显著提高地下生物量, 两者对生态系统生产力存在显著的交互作用(Li et al., 2011), 因此氮和水分两者可能共同影响生态系统的碳交换(Hooper & Johnson, 1999). ...
Effects of nitrogen fertilization on soil respiration in temperate grassland in Inner Mongolia, China 2 2011
... 氮和水分是草地生态系统生产力的两个主要限制性因子(Hooper & Johnson, 1999; Yan et al., 2011).鉴于草地退化常导致土壤养分缺乏, 氮添加作为一种重要手段被用于对退化草地的恢复和管理中(潘庆民等, 2005; 张璐等, 2009).氮添加可能影响生态系统光合作用和植物生长, 导致植物生产力的不同响应(Ren et al., 2017), 进而对土壤呼吸产生促进(Song et al., 2011)、抑制(Mo et al., 2007)或无显著影响(Peng et al., 2011; Qi et al., 2014; 李寅龙等, 2015) 3种不同的作用.水分作为干旱区草地生态系统过程重要的限制因子, 通常降水增加对土壤呼吸有显著促进作用(Sowerby et al., 2008; Yan et al., 2010, 2011), 而氮添加的效果常常会依赖于水分条件.刁励玮等(2018)统计分析了1990-2017年间发表的有关氮添加的研究论文结果, 发现单独施氮对地下生物量无显著影响, 氮水同时添加能显著提高地下生物量, 两者对生态系统生产力存在显著的交互作用(Li et al., 2011), 因此氮和水分两者可能共同影响生态系统的碳交换(Hooper & Johnson, 1999). ...
Soil enzyme and microbial activities in a grazing ecosystem of Patagonian Monte, Argentina 2 2011
... 放牧是我国北方草地生态系统主要的利用方式.关于放牧对草地生态系统碳交换过程的影响一直以来受到学术界的广泛关注(Lin et al., 2011; Fu et al., 2014; Hou et al., 2014; Chen et al., 2015, 2016; Stark et al., 2015).当前的研究结果表明, 放牧对土壤呼吸的影响并没有一致的规律性, 放牧可能导致土壤呼吸速率升高、降低, 或者对土壤呼吸没有显著的影响(Chen et al., 2015, 2016; Stark et al., 2015).事实上, 放牧对土壤呼吸的影响可能是多种因素综合作用的结果.一方面, 放牧牲畜的啃食作用直接降低了植被冠层的光合作用(Bremer et al., 1998), 使得分配于根系的光合同化产物减少, 降低根系生物量(Cao et al., 2004; Gao et al., 2008; Klumpp et al., 2009), 减少了地表凋落物和根际分泌物(Bremer et al., 1998), 降低土壤微生物生物量和土壤酶活性(Prieto et al., 2011; Li et al., 2013), 最终导致土壤呼吸速率降低.另一方面, 放牧牲畜以粪尿形式归还有机物, 大量的活性有机质和氮输入为微生物呼吸作用提供更加丰富的底物, 激发微生物的活性和增加养分的循环速度(Fu et al., 2014), 加速根系生长, 增加根系分泌物, 进而促进土壤呼吸.上述因素在不同的时间和研究区域, 对土壤呼吸的贡献大小并不一致, 进而导致土壤呼吸速率对放牧响应的不确定性. ...
... 放牧对草地生态系统的影响是复杂、多方面且持久的, 放牧对草原植物(Cao et al., 2004; Gao et al., 2008)、微生物(Li et al., 2013)、凋落物(Klumpp et al., 2009)、土壤理化特性(Prieto et al., 2011; Li et al., 2013)等都存在着不同程度的影响, 而且放牧强度及其持续时间的不同也强烈影响着草地生态系统结构和功能.不同放牧强度的干扰将会迅速引起草地植被群落特征(王悦骅等, 2018)、土壤微生物数量、土壤酶和土壤理化性质如含水量、土壤有机质含量等方面的变化(高雪峰等, 2010), 而且在一定程度上影响水、热因子的作用效应, 进而导致土壤呼吸作用改变.已有的研究结果显示, 放牧不仅可以降低土壤呼吸速率(邓钰等, 2013; Hou et al., 2014), 也可以促进土壤呼吸(Chen et al., 2015, 2016; Stark et al., 2015), 也有影响不显著的(Lin et al., 2011; Fu et al., 2014).有研究发现放牧对土壤呼吸影响的大小与方向和放牧强度有关, 轻度放牧促进土壤呼吸, 中度和重度放牧则降低土壤呼吸速率(李玉强等, 2006).Wang等(2001)在内蒙古温带草原的研究表明, 自由放牧可增加土壤呼吸的年排放量, 且土壤呼吸与放牧强度呈正相关关系.然而, 也有研究认为, 荒漠草原放牧草地的土壤呼吸速率显著低于未放牧草地(阿木日吉日嘎拉等, 2013).本研究中, 过去不同的放牧强度对土壤呼吸速率的影响具有明显的年际变化, 表现为无显著影响和促进两种.放牧在总体上促进了土壤呼吸, 表现为放牧(LG、MG、HG)区3年平均土壤呼吸速率都高于禁牧区.两者研究结果截然不同, 这里存在两种可能, 一种是放牧增加土壤呼吸, 即使在停止放牧后, 放牧区的土壤呼吸速率仍然较高; 还有一种可能是放牧可能抑制土壤呼吸, 但停止放牧后, 土壤呼吸产生了超补偿, 使得土壤呼吸迅速恢复, 并超过对照区的, 而我们的研究结果属于哪种情况, 我们并不清楚.另外, 本研究是在多年放牧的草地上进行了小区围封, 实际上变成了围封草地, 围封后放牧的扰动已经去除, 草地的生物量和水分条件得到了一定程度的恢复, 从而引起土壤呼吸变化.已有研究也表明, 围封草地的土壤呼吸速率要明显高于放牧草地(贾丙瑞等, 2005), 而且放牧会导致全年的土壤呼吸速率比围封草地降低1/3 (马涛等, 2009).另外, 本研究持续3年测定土壤呼吸速率, 研究年限的不同对草地土壤呼吸的影响也存在差异(郭明英等, 2011). ...
Differential responses of short-term soil respiration dynamics to the experimental addition of nitrogen and water in the temperate semi-arid steppe of Inner Mongolia, China 1 2014
... 氮和水分是草地生态系统生产力的两个主要限制性因子(Hooper & Johnson, 1999; Yan et al., 2011).鉴于草地退化常导致土壤养分缺乏, 氮添加作为一种重要手段被用于对退化草地的恢复和管理中(潘庆民等, 2005; 张璐等, 2009).氮添加可能影响生态系统光合作用和植物生长, 导致植物生产力的不同响应(Ren et al., 2017), 进而对土壤呼吸产生促进(Song et al., 2011)、抑制(Mo et al., 2007)或无显著影响(Peng et al., 2011; Qi et al., 2014; 李寅龙等, 2015) 3种不同的作用.水分作为干旱区草地生态系统过程重要的限制因子, 通常降水增加对土壤呼吸有显著促进作用(Sowerby et al., 2008; Yan et al., 2010, 2011), 而氮添加的效果常常会依赖于水分条件.刁励玮等(2018)统计分析了1990-2017年间发表的有关氮添加的研究论文结果, 发现单独施氮对地下生物量无显著影响, 氮水同时添加能显著提高地下生物量, 两者对生态系统生产力存在显著的交互作用(Li et al., 2011), 因此氮和水分两者可能共同影响生态系统的碳交换(Hooper & Johnson, 1999). ...
Interannual variability in global soil respiration, 1980-94 1 2002
... 荒漠草原地处温带干旱半干旱区域, 无论是植物生长, 还是土壤生态学过程, 都更加受到水分条件的限制.许多研究发现水分添加会显著增加土壤微生物生物量(Liu et al., 2009; Yan et al., 2010), 增强土壤有机质的矿化作用(Harrison et al., 2008; Liu et al., 2009), 这主要是由于土壤水分供给的增加导致营养元素扩散能力的提高, 使其更容易被植物和微生物利用(Sowerby et al., 2010; Reichmann et al., 2013).因此, 水分变化能够影响草原植物根系生长、土壤代谢活力、根系呼吸、土壤微生物群落构成以及微生物活性(Xu et al., 2004), 进而引起土壤呼吸强度的改变.在全球尺度上, 土壤呼吸强度与降水量呈正相关关系(Raich & Schlesinger, 1992; Zhou et al., 2014; Liu et al., 2016).而对于干旱和半干旱生态系统, 土壤湿度的影响尤其显著(Conant et al., 2000), 水分是导致土壤呼吸速率季节波动的重要因素和年际变化的主要驱动因子(Raich et al., 2002).降水可以在短时间内引起土壤呼吸作用的急剧增强(Birch, 1958), 而且这种对土壤呼吸的激发作用在干旱和半干旱地区变得更加显著(Sponseller, 2007).由降水格局改变引起的土壤呼吸变化效应可能远高于其他全球变化因子(增温、氮沉降、CO2升高)(Weltzin et al., 2003).本实验结果表明, 连续3年的加水处理(W和NW)使得土壤含水量明显增加, 对土壤呼吸产生了显著的促进作用, 而氮添加的有效性依赖于水分条件, 两者表现为协同作用.另外, 加水对土壤呼吸的促进作用大小也取决于自然降水的多少, 如在丰水年(2016年)影响不显著, 说明水分对该地区土壤呼吸的影响程度与总体上降水量高低有关, 在降水量较低的年份(平水或欠水年)能够促进土壤呼吸, 反之, 在降水量充沛的年份, 增加降水对土壤呼吸的促进作用有限, 甚至可能抑制土壤呼吸, 因为过量的水分添加会降低土壤通透性, 导致土壤氧气供应不足, 制约土壤呼吸.Liu等(2016)也发现土壤呼吸对增雨的响应随着环境湿润度的增加而减弱. ...
The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate 2 1992
... 荒漠草原地处温带干旱半干旱区域, 无论是植物生长, 还是土壤生态学过程, 都更加受到水分条件的限制.许多研究发现水分添加会显著增加土壤微生物生物量(Liu et al., 2009; Yan et al., 2010), 增强土壤有机质的矿化作用(Harrison et al., 2008; Liu et al., 2009), 这主要是由于土壤水分供给的增加导致营养元素扩散能力的提高, 使其更容易被植物和微生物利用(Sowerby et al., 2010; Reichmann et al., 2013).因此, 水分变化能够影响草原植物根系生长、土壤代谢活力、根系呼吸、土壤微生物群落构成以及微生物活性(Xu et al., 2004), 进而引起土壤呼吸强度的改变.在全球尺度上, 土壤呼吸强度与降水量呈正相关关系(Raich & Schlesinger, 1992; Zhou et al., 2014; Liu et al., 2016).而对于干旱和半干旱生态系统, 土壤湿度的影响尤其显著(Conant et al., 2000), 水分是导致土壤呼吸速率季节波动的重要因素和年际变化的主要驱动因子(Raich et al., 2002).降水可以在短时间内引起土壤呼吸作用的急剧增强(Birch, 1958), 而且这种对土壤呼吸的激发作用在干旱和半干旱地区变得更加显著(Sponseller, 2007).由降水格局改变引起的土壤呼吸变化效应可能远高于其他全球变化因子(增温、氮沉降、CO2升高)(Weltzin et al., 2003).本实验结果表明, 连续3年的加水处理(W和NW)使得土壤含水量明显增加, 对土壤呼吸产生了显著的促进作用, 而氮添加的有效性依赖于水分条件, 两者表现为协同作用.另外, 加水对土壤呼吸的促进作用大小也取决于自然降水的多少, 如在丰水年(2016年)影响不显著, 说明水分对该地区土壤呼吸的影响程度与总体上降水量高低有关, 在降水量较低的年份(平水或欠水年)能够促进土壤呼吸, 反之, 在降水量充沛的年份, 增加降水对土壤呼吸的促进作用有限, 甚至可能抑制土壤呼吸, 因为过量的水分添加会降低土壤通透性, 导致土壤氧气供应不足, 制约土壤呼吸.Liu等(2016)也发现土壤呼吸对增雨的响应随着环境湿润度的增加而减弱. ...
Water controls on nitrogen transformations and stocks in an arid ecosystem 1 2013
... 荒漠草原地处温带干旱半干旱区域, 无论是植物生长, 还是土壤生态学过程, 都更加受到水分条件的限制.许多研究发现水分添加会显著增加土壤微生物生物量(Liu et al., 2009; Yan et al., 2010), 增强土壤有机质的矿化作用(Harrison et al., 2008; Liu et al., 2009), 这主要是由于土壤水分供给的增加导致营养元素扩散能力的提高, 使其更容易被植物和微生物利用(Sowerby et al., 2010; Reichmann et al., 2013).因此, 水分变化能够影响草原植物根系生长、土壤代谢活力、根系呼吸、土壤微生物群落构成以及微生物活性(Xu et al., 2004), 进而引起土壤呼吸强度的改变.在全球尺度上, 土壤呼吸强度与降水量呈正相关关系(Raich & Schlesinger, 1992; Zhou et al., 2014; Liu et al., 2016).而对于干旱和半干旱生态系统, 土壤湿度的影响尤其显著(Conant et al., 2000), 水分是导致土壤呼吸速率季节波动的重要因素和年际变化的主要驱动因子(Raich et al., 2002).降水可以在短时间内引起土壤呼吸作用的急剧增强(Birch, 1958), 而且这种对土壤呼吸的激发作用在干旱和半干旱地区变得更加显著(Sponseller, 2007).由降水格局改变引起的土壤呼吸变化效应可能远高于其他全球变化因子(增温、氮沉降、CO2升高)(Weltzin et al., 2003).本实验结果表明, 连续3年的加水处理(W和NW)使得土壤含水量明显增加, 对土壤呼吸产生了显著的促进作用, 而氮添加的有效性依赖于水分条件, 两者表现为协同作用.另外, 加水对土壤呼吸的促进作用大小也取决于自然降水的多少, 如在丰水年(2016年)影响不显著, 说明水分对该地区土壤呼吸的影响程度与总体上降水量高低有关, 在降水量较低的年份(平水或欠水年)能够促进土壤呼吸, 反之, 在降水量充沛的年份, 增加降水对土壤呼吸的促进作用有限, 甚至可能抑制土壤呼吸, 因为过量的水分添加会降低土壤通透性, 导致土壤氧气供应不足, 制约土壤呼吸.Liu等(2016)也发现土壤呼吸对增雨的响应随着环境湿润度的增加而减弱. ...
Exacerbated nitrogen limitation ends transient stimulation of grassland productivity by increased precipitation 1 2017
... 氮和水分是草地生态系统生产力的两个主要限制性因子(Hooper & Johnson, 1999; Yan et al., 2011).鉴于草地退化常导致土壤养分缺乏, 氮添加作为一种重要手段被用于对退化草地的恢复和管理中(潘庆民等, 2005; 张璐等, 2009).氮添加可能影响生态系统光合作用和植物生长, 导致植物生产力的不同响应(Ren et al., 2017), 进而对土壤呼吸产生促进(Song et al., 2011)、抑制(Mo et al., 2007)或无显著影响(Peng et al., 2011; Qi et al., 2014; 李寅龙等, 2015) 3种不同的作用.水分作为干旱区草地生态系统过程重要的限制因子, 通常降水增加对土壤呼吸有显著促进作用(Sowerby et al., 2008; Yan et al., 2010, 2011), 而氮添加的效果常常会依赖于水分条件.刁励玮等(2018)统计分析了1990-2017年间发表的有关氮添加的研究论文结果, 发现单独施氮对地下生物量无显著影响, 氮水同时添加能显著提高地下生物量, 两者对生态系统生产力存在显著的交互作用(Li et al., 2011), 因此氮和水分两者可能共同影响生态系统的碳交换(Hooper & Johnson, 1999). ...
Soil CO2 fluxes beneath barley on the central Spanish plateau 1 2003
... 采用指数模型拟合土壤呼吸速率与土壤温度之间的关系(Sánchez et al., 2003), 采用线性方程对土壤呼吸速率与土壤含水量间进行回归分析(Davidson et al., 1998).方程如下: ...
Effect of nitrogen addition on decomposition of Calamagrostis angustifolia litters from freshwater marshes of Northeast China 2 2011
... 氮和水分是草地生态系统生产力的两个主要限制性因子(Hooper & Johnson, 1999; Yan et al., 2011).鉴于草地退化常导致土壤养分缺乏, 氮添加作为一种重要手段被用于对退化草地的恢复和管理中(潘庆民等, 2005; 张璐等, 2009).氮添加可能影响生态系统光合作用和植物生长, 导致植物生产力的不同响应(Ren et al., 2017), 进而对土壤呼吸产生促进(Song et al., 2011)、抑制(Mo et al., 2007)或无显著影响(Peng et al., 2011; Qi et al., 2014; 李寅龙等, 2015) 3种不同的作用.水分作为干旱区草地生态系统过程重要的限制因子, 通常降水增加对土壤呼吸有显著促进作用(Sowerby et al., 2008; Yan et al., 2010, 2011), 而氮添加的效果常常会依赖于水分条件.刁励玮等(2018)统计分析了1990-2017年间发表的有关氮添加的研究论文结果, 发现单独施氮对地下生物量无显著影响, 氮水同时添加能显著提高地下生物量, 两者对生态系统生产力存在显著的交互作用(Li et al., 2011), 因此氮和水分两者可能共同影响生态系统的碳交换(Hooper & Johnson, 1999). ...
Contrasting effects of repeated summer drought on soil carbon efflux in hydric and mesic heathland soils 1 2008
... 氮和水分是草地生态系统生产力的两个主要限制性因子(Hooper & Johnson, 1999; Yan et al., 2011).鉴于草地退化常导致土壤养分缺乏, 氮添加作为一种重要手段被用于对退化草地的恢复和管理中(潘庆民等, 2005; 张璐等, 2009).氮添加可能影响生态系统光合作用和植物生长, 导致植物生产力的不同响应(Ren et al., 2017), 进而对土壤呼吸产生促进(Song et al., 2011)、抑制(Mo et al., 2007)或无显著影响(Peng et al., 2011; Qi et al., 2014; 李寅龙等, 2015) 3种不同的作用.水分作为干旱区草地生态系统过程重要的限制因子, 通常降水增加对土壤呼吸有显著促进作用(Sowerby et al., 2008; Yan et al., 2010, 2011), 而氮添加的效果常常会依赖于水分条件.刁励玮等(2018)统计分析了1990-2017年间发表的有关氮添加的研究论文结果, 发现单独施氮对地下生物量无显著影响, 氮水同时添加能显著提高地下生物量, 两者对生态系统生产力存在显著的交互作用(Li et al., 2011), 因此氮和水分两者可能共同影响生态系统的碳交换(Hooper & Johnson, 1999). ...
The response of dissolved organic carbon (DOC) and the ecosystem carbon balance to experimental drought in a temperate shrubland 1 2010
... 荒漠草原地处温带干旱半干旱区域, 无论是植物生长, 还是土壤生态学过程, 都更加受到水分条件的限制.许多研究发现水分添加会显著增加土壤微生物生物量(Liu et al., 2009; Yan et al., 2010), 增强土壤有机质的矿化作用(Harrison et al., 2008; Liu et al., 2009), 这主要是由于土壤水分供给的增加导致营养元素扩散能力的提高, 使其更容易被植物和微生物利用(Sowerby et al., 2010; Reichmann et al., 2013).因此, 水分变化能够影响草原植物根系生长、土壤代谢活力、根系呼吸、土壤微生物群落构成以及微生物活性(Xu et al., 2004), 进而引起土壤呼吸强度的改变.在全球尺度上, 土壤呼吸强度与降水量呈正相关关系(Raich & Schlesinger, 1992; Zhou et al., 2014; Liu et al., 2016).而对于干旱和半干旱生态系统, 土壤湿度的影响尤其显著(Conant et al., 2000), 水分是导致土壤呼吸速率季节波动的重要因素和年际变化的主要驱动因子(Raich et al., 2002).降水可以在短时间内引起土壤呼吸作用的急剧增强(Birch, 1958), 而且这种对土壤呼吸的激发作用在干旱和半干旱地区变得更加显著(Sponseller, 2007).由降水格局改变引起的土壤呼吸变化效应可能远高于其他全球变化因子(增温、氮沉降、CO2升高)(Weltzin et al., 2003).本实验结果表明, 连续3年的加水处理(W和NW)使得土壤含水量明显增加, 对土壤呼吸产生了显著的促进作用, 而氮添加的有效性依赖于水分条件, 两者表现为协同作用.另外, 加水对土壤呼吸的促进作用大小也取决于自然降水的多少, 如在丰水年(2016年)影响不显著, 说明水分对该地区土壤呼吸的影响程度与总体上降水量高低有关, 在降水量较低的年份(平水或欠水年)能够促进土壤呼吸, 反之, 在降水量充沛的年份, 增加降水对土壤呼吸的促进作用有限, 甚至可能抑制土壤呼吸, 因为过量的水分添加会降低土壤通透性, 导致土壤氧气供应不足, 制约土壤呼吸.Liu等(2016)也发现土壤呼吸对增雨的响应随着环境湿润度的增加而减弱. ...
Precipitation pulses and soil CO2 flux in a Sonoran desert ecosystem 1 2007
... 荒漠草原地处温带干旱半干旱区域, 无论是植物生长, 还是土壤生态学过程, 都更加受到水分条件的限制.许多研究发现水分添加会显著增加土壤微生物生物量(Liu et al., 2009; Yan et al., 2010), 增强土壤有机质的矿化作用(Harrison et al., 2008; Liu et al., 2009), 这主要是由于土壤水分供给的增加导致营养元素扩散能力的提高, 使其更容易被植物和微生物利用(Sowerby et al., 2010; Reichmann et al., 2013).因此, 水分变化能够影响草原植物根系生长、土壤代谢活力、根系呼吸、土壤微生物群落构成以及微生物活性(Xu et al., 2004), 进而引起土壤呼吸强度的改变.在全球尺度上, 土壤呼吸强度与降水量呈正相关关系(Raich & Schlesinger, 1992; Zhou et al., 2014; Liu et al., 2016).而对于干旱和半干旱生态系统, 土壤湿度的影响尤其显著(Conant et al., 2000), 水分是导致土壤呼吸速率季节波动的重要因素和年际变化的主要驱动因子(Raich et al., 2002).降水可以在短时间内引起土壤呼吸作用的急剧增强(Birch, 1958), 而且这种对土壤呼吸的激发作用在干旱和半干旱地区变得更加显著(Sponseller, 2007).由降水格局改变引起的土壤呼吸变化效应可能远高于其他全球变化因子(增温、氮沉降、CO2升高)(Weltzin et al., 2003).本实验结果表明, 连续3年的加水处理(W和NW)使得土壤含水量明显增加, 对土壤呼吸产生了显著的促进作用, 而氮添加的有效性依赖于水分条件, 两者表现为协同作用.另外, 加水对土壤呼吸的促进作用大小也取决于自然降水的多少, 如在丰水年(2016年)影响不显著, 说明水分对该地区土壤呼吸的影响程度与总体上降水量高低有关, 在降水量较低的年份(平水或欠水年)能够促进土壤呼吸, 反之, 在降水量充沛的年份, 增加降水对土壤呼吸的促进作用有限, 甚至可能抑制土壤呼吸, 因为过量的水分添加会降低土壤通透性, 导致土壤氧气供应不足, 制约土壤呼吸.Liu等(2016)也发现土壤呼吸对增雨的响应随着环境湿润度的增加而减弱. ...
Grazing intensity in subarctic tundra affects the temperature adaptation of soil microbial communities 3 2015
... 放牧是我国北方草地生态系统主要的利用方式.关于放牧对草地生态系统碳交换过程的影响一直以来受到学术界的广泛关注(Lin et al., 2011; Fu et al., 2014; Hou et al., 2014; Chen et al., 2015, 2016; Stark et al., 2015).当前的研究结果表明, 放牧对土壤呼吸的影响并没有一致的规律性, 放牧可能导致土壤呼吸速率升高、降低, 或者对土壤呼吸没有显著的影响(Chen et al., 2015, 2016; Stark et al., 2015).事实上, 放牧对土壤呼吸的影响可能是多种因素综合作用的结果.一方面, 放牧牲畜的啃食作用直接降低了植被冠层的光合作用(Bremer et al., 1998), 使得分配于根系的光合同化产物减少, 降低根系生物量(Cao et al., 2004; Gao et al., 2008; Klumpp et al., 2009), 减少了地表凋落物和根际分泌物(Bremer et al., 1998), 降低土壤微生物生物量和土壤酶活性(Prieto et al., 2011; Li et al., 2013), 最终导致土壤呼吸速率降低.另一方面, 放牧牲畜以粪尿形式归还有机物, 大量的活性有机质和氮输入为微生物呼吸作用提供更加丰富的底物, 激发微生物的活性和增加养分的循环速度(Fu et al., 2014), 加速根系生长, 增加根系分泌物, 进而促进土壤呼吸.上述因素在不同的时间和研究区域, 对土壤呼吸的贡献大小并不一致, 进而导致土壤呼吸速率对放牧响应的不确定性. ...
... ; Stark et al., 2015).事实上, 放牧对土壤呼吸的影响可能是多种因素综合作用的结果.一方面, 放牧牲畜的啃食作用直接降低了植被冠层的光合作用(Bremer et al., 1998), 使得分配于根系的光合同化产物减少, 降低根系生物量(Cao et al., 2004; Gao et al., 2008; Klumpp et al., 2009), 减少了地表凋落物和根际分泌物(Bremer et al., 1998), 降低土壤微生物生物量和土壤酶活性(Prieto et al., 2011; Li et al., 2013), 最终导致土壤呼吸速率降低.另一方面, 放牧牲畜以粪尿形式归还有机物, 大量的活性有机质和氮输入为微生物呼吸作用提供更加丰富的底物, 激发微生物的活性和增加养分的循环速度(Fu et al., 2014), 加速根系生长, 增加根系分泌物, 进而促进土壤呼吸.上述因素在不同的时间和研究区域, 对土壤呼吸的贡献大小并不一致, 进而导致土壤呼吸速率对放牧响应的不确定性. ...
... 放牧对草地生态系统的影响是复杂、多方面且持久的, 放牧对草原植物(Cao et al., 2004; Gao et al., 2008)、微生物(Li et al., 2013)、凋落物(Klumpp et al., 2009)、土壤理化特性(Prieto et al., 2011; Li et al., 2013)等都存在着不同程度的影响, 而且放牧强度及其持续时间的不同也强烈影响着草地生态系统结构和功能.不同放牧强度的干扰将会迅速引起草地植被群落特征(王悦骅等, 2018)、土壤微生物数量、土壤酶和土壤理化性质如含水量、土壤有机质含量等方面的变化(高雪峰等, 2010), 而且在一定程度上影响水、热因子的作用效应, 进而导致土壤呼吸作用改变.已有的研究结果显示, 放牧不仅可以降低土壤呼吸速率(邓钰等, 2013; Hou et al., 2014), 也可以促进土壤呼吸(Chen et al., 2015, 2016; Stark et al., 2015), 也有影响不显著的(Lin et al., 2011; Fu et al., 2014).有研究发现放牧对土壤呼吸影响的大小与方向和放牧强度有关, 轻度放牧促进土壤呼吸, 中度和重度放牧则降低土壤呼吸速率(李玉强等, 2006).Wang等(2001)在内蒙古温带草原的研究表明, 自由放牧可增加土壤呼吸的年排放量, 且土壤呼吸与放牧强度呈正相关关系.然而, 也有研究认为, 荒漠草原放牧草地的土壤呼吸速率显著低于未放牧草地(阿木日吉日嘎拉等, 2013).本研究中, 过去不同的放牧强度对土壤呼吸速率的影响具有明显的年际变化, 表现为无显著影响和促进两种.放牧在总体上促进了土壤呼吸, 表现为放牧(LG、MG、HG)区3年平均土壤呼吸速率都高于禁牧区.两者研究结果截然不同, 这里存在两种可能, 一种是放牧增加土壤呼吸, 即使在停止放牧后, 放牧区的土壤呼吸速率仍然较高; 还有一种可能是放牧可能抑制土壤呼吸, 但停止放牧后, 土壤呼吸产生了超补偿, 使得土壤呼吸迅速恢复, 并超过对照区的, 而我们的研究结果属于哪种情况, 我们并不清楚.另外, 本研究是在多年放牧的草地上进行了小区围封, 实际上变成了围封草地, 围封后放牧的扰动已经去除, 草地的生物量和水分条件得到了一定程度的恢复, 从而引起土壤呼吸变化.已有研究也表明, 围封草地的土壤呼吸速率要明显高于放牧草地(贾丙瑞等, 2005), 而且放牧会导致全年的土壤呼吸速率比围封草地降低1/3 (马涛等, 2009).另外, 本研究持续3年测定土壤呼吸速率, 研究年限的不同对草地土壤呼吸的影响也存在差异(郭明英等, 2011). ...
Automated closed-system canopy-chamber for continuous field-crop monitoring of CO2 and H2O fluxes 1 2002
... 为了避免操作的影响, 计算时将数据记录中的开始后和结束前10 s的数据去除.采用线性拟合的方法计算CO2浓度变化速率(dc/dt), 即斜率.CO2交换速率(Rs, μmol·m-2·s-1)的计算采用以下公式(Steduto et al., 2002): ...
植物群落多样性和生产力对不同载畜率的响应 2 2018
... 研究在内蒙古农牧业科学院综合试验示范中心的长期放牧控制实验平台上开展, 该平台放牧实验始于2003年6月, 每年放牧期从6月1日到11月30日.放牧区采取随机区组设计, 设4个放牧强度, 分别为对照样地(CK)、轻度放牧(LG)、中度放牧(MG)和重度放牧(HG), 放牧强度分别为0、0.93、1.82和2.71 sheep·hm-2, 每个强度设3个重复, 共计12个实验小区, 每个实验小区占地面积基本相等, 整个实验区占地面积约为50 hm2.多年连续放牧, 尤其是重度放牧总体上降低了群落的物种多样性和生产力(王悦骅等, 2018).2014年在每个放牧实验小区设置一个面积为8 m × 8 m的处理围栏, 每个处理围栏内设有4个处理亚区, 分别设置对照(CKs)、加氮(N)、加水(W)、氮水共同添加(NW)处理, 每个处理亚区的面积2 m × 2 m (亚区边缘用镀锌铁皮围起来, 埋入土壤20 cm, 露出地表5 cm, 以防止养分和水分侧渗), 中间留1 m的缓冲区, 共计48个亚区(4放牧强度× 3个重复× 4个处理围栏).2015年开始进行氮、水添加处理, 加水处理每年增水108 mm, 约占年降水量的40%, 在生长季6、7、8月进行人工增水处理, 每月4次, 共12次, 每次增水量为9 mm, 即每次每小区增加36 kg人工降水.选用硝酸铵钙进行氮添加, 施氮量为10.0 g·m-2·a-1, 分3次施入, 每年6月1日、 7月1日和8月1日手工均匀撒入样方, 即每次每小区施入85.68 g硝酸铵钙. ...
... 放牧对草地生态系统的影响是复杂、多方面且持久的, 放牧对草原植物(Cao et al., 2004; Gao et al., 2008)、微生物(Li et al., 2013)、凋落物(Klumpp et al., 2009)、土壤理化特性(Prieto et al., 2011; Li et al., 2013)等都存在着不同程度的影响, 而且放牧强度及其持续时间的不同也强烈影响着草地生态系统结构和功能.不同放牧强度的干扰将会迅速引起草地植被群落特征(王悦骅等, 2018)、土壤微生物数量、土壤酶和土壤理化性质如含水量、土壤有机质含量等方面的变化(高雪峰等, 2010), 而且在一定程度上影响水、热因子的作用效应, 进而导致土壤呼吸作用改变.已有的研究结果显示, 放牧不仅可以降低土壤呼吸速率(邓钰等, 2013; Hou et al., 2014), 也可以促进土壤呼吸(Chen et al., 2015, 2016; Stark et al., 2015), 也有影响不显著的(Lin et al., 2011; Fu et al., 2014).有研究发现放牧对土壤呼吸影响的大小与方向和放牧强度有关, 轻度放牧促进土壤呼吸, 中度和重度放牧则降低土壤呼吸速率(李玉强等, 2006).Wang等(2001)在内蒙古温带草原的研究表明, 自由放牧可增加土壤呼吸的年排放量, 且土壤呼吸与放牧强度呈正相关关系.然而, 也有研究认为, 荒漠草原放牧草地的土壤呼吸速率显著低于未放牧草地(阿木日吉日嘎拉等, 2013).本研究中, 过去不同的放牧强度对土壤呼吸速率的影响具有明显的年际变化, 表现为无显著影响和促进两种.放牧在总体上促进了土壤呼吸, 表现为放牧(LG、MG、HG)区3年平均土壤呼吸速率都高于禁牧区.两者研究结果截然不同, 这里存在两种可能, 一种是放牧增加土壤呼吸, 即使在停止放牧后, 放牧区的土壤呼吸速率仍然较高; 还有一种可能是放牧可能抑制土壤呼吸, 但停止放牧后, 土壤呼吸产生了超补偿, 使得土壤呼吸迅速恢复, 并超过对照区的, 而我们的研究结果属于哪种情况, 我们并不清楚.另外, 本研究是在多年放牧的草地上进行了小区围封, 实际上变成了围封草地, 围封后放牧的扰动已经去除, 草地的生物量和水分条件得到了一定程度的恢复, 从而引起土壤呼吸变化.已有研究也表明, 围封草地的土壤呼吸速率要明显高于放牧草地(贾丙瑞等, 2005), 而且放牧会导致全年的土壤呼吸速率比围封草地降低1/3 (马涛等, 2009).另外, 本研究持续3年测定土壤呼吸速率, 研究年限的不同对草地土壤呼吸的影响也存在差异(郭明英等, 2011). ...
植物群落多样性和生产力对不同载畜率的响应 2 2018
... 研究在内蒙古农牧业科学院综合试验示范中心的长期放牧控制实验平台上开展, 该平台放牧实验始于2003年6月, 每年放牧期从6月1日到11月30日.放牧区采取随机区组设计, 设4个放牧强度, 分别为对照样地(CK)、轻度放牧(LG)、中度放牧(MG)和重度放牧(HG), 放牧强度分别为0、0.93、1.82和2.71 sheep·hm-2, 每个强度设3个重复, 共计12个实验小区, 每个实验小区占地面积基本相等, 整个实验区占地面积约为50 hm2.多年连续放牧, 尤其是重度放牧总体上降低了群落的物种多样性和生产力(王悦骅等, 2018).2014年在每个放牧实验小区设置一个面积为8 m × 8 m的处理围栏, 每个处理围栏内设有4个处理亚区, 分别设置对照(CKs)、加氮(N)、加水(W)、氮水共同添加(NW)处理, 每个处理亚区的面积2 m × 2 m (亚区边缘用镀锌铁皮围起来, 埋入土壤20 cm, 露出地表5 cm, 以防止养分和水分侧渗), 中间留1 m的缓冲区, 共计48个亚区(4放牧强度× 3个重复× 4个处理围栏).2015年开始进行氮、水添加处理, 加水处理每年增水108 mm, 约占年降水量的40%, 在生长季6、7、8月进行人工增水处理, 每月4次, 共12次, 每次增水量为9 mm, 即每次每小区增加36 kg人工降水.选用硝酸铵钙进行氮添加, 施氮量为10.0 g·m-2·a-1, 分3次施入, 每年6月1日、 7月1日和8月1日手工均匀撒入样方, 即每次每小区施入85.68 g硝酸铵钙. ...
... 放牧对草地生态系统的影响是复杂、多方面且持久的, 放牧对草原植物(Cao et al., 2004; Gao et al., 2008)、微生物(Li et al., 2013)、凋落物(Klumpp et al., 2009)、土壤理化特性(Prieto et al., 2011; Li et al., 2013)等都存在着不同程度的影响, 而且放牧强度及其持续时间的不同也强烈影响着草地生态系统结构和功能.不同放牧强度的干扰将会迅速引起草地植被群落特征(王悦骅等, 2018)、土壤微生物数量、土壤酶和土壤理化性质如含水量、土壤有机质含量等方面的变化(高雪峰等, 2010), 而且在一定程度上影响水、热因子的作用效应, 进而导致土壤呼吸作用改变.已有的研究结果显示, 放牧不仅可以降低土壤呼吸速率(邓钰等, 2013; Hou et al., 2014), 也可以促进土壤呼吸(Chen et al., 2015, 2016; Stark et al., 2015), 也有影响不显著的(Lin et al., 2011; Fu et al., 2014).有研究发现放牧对土壤呼吸影响的大小与方向和放牧强度有关, 轻度放牧促进土壤呼吸, 中度和重度放牧则降低土壤呼吸速率(李玉强等, 2006).Wang等(2001)在内蒙古温带草原的研究表明, 自由放牧可增加土壤呼吸的年排放量, 且土壤呼吸与放牧强度呈正相关关系.然而, 也有研究认为, 荒漠草原放牧草地的土壤呼吸速率显著低于未放牧草地(阿木日吉日嘎拉等, 2013).本研究中, 过去不同的放牧强度对土壤呼吸速率的影响具有明显的年际变化, 表现为无显著影响和促进两种.放牧在总体上促进了土壤呼吸, 表现为放牧(LG、MG、HG)区3年平均土壤呼吸速率都高于禁牧区.两者研究结果截然不同, 这里存在两种可能, 一种是放牧增加土壤呼吸, 即使在停止放牧后, 放牧区的土壤呼吸速率仍然较高; 还有一种可能是放牧可能抑制土壤呼吸, 但停止放牧后, 土壤呼吸产生了超补偿, 使得土壤呼吸迅速恢复, 并超过对照区的, 而我们的研究结果属于哪种情况, 我们并不清楚.另外, 本研究是在多年放牧的草地上进行了小区围封, 实际上变成了围封草地, 围封后放牧的扰动已经去除, 草地的生物量和水分条件得到了一定程度的恢复, 从而引起土壤呼吸变化.已有研究也表明, 围封草地的土壤呼吸速率要明显高于放牧草地(贾丙瑞等, 2005), 而且放牧会导致全年的土壤呼吸速率比围封草地降低1/3 (马涛等, 2009).另外, 本研究持续3年测定土壤呼吸速率, 研究年限的不同对草地土壤呼吸的影响也存在差异(郭明英等, 2011). ...
Effects of grazing and cultivating on emission of nitrous oxide, carbon dioxide and uptake of methane from grasslands 1 2001
... 放牧对草地生态系统的影响是复杂、多方面且持久的, 放牧对草原植物(Cao et al., 2004; Gao et al., 2008)、微生物(Li et al., 2013)、凋落物(Klumpp et al., 2009)、土壤理化特性(Prieto et al., 2011; Li et al., 2013)等都存在着不同程度的影响, 而且放牧强度及其持续时间的不同也强烈影响着草地生态系统结构和功能.不同放牧强度的干扰将会迅速引起草地植被群落特征(王悦骅等, 2018)、土壤微生物数量、土壤酶和土壤理化性质如含水量、土壤有机质含量等方面的变化(高雪峰等, 2010), 而且在一定程度上影响水、热因子的作用效应, 进而导致土壤呼吸作用改变.已有的研究结果显示, 放牧不仅可以降低土壤呼吸速率(邓钰等, 2013; Hou et al., 2014), 也可以促进土壤呼吸(Chen et al., 2015, 2016; Stark et al., 2015), 也有影响不显著的(Lin et al., 2011; Fu et al., 2014).有研究发现放牧对土壤呼吸影响的大小与方向和放牧强度有关, 轻度放牧促进土壤呼吸, 中度和重度放牧则降低土壤呼吸速率(李玉强等, 2006).Wang等(2001)在内蒙古温带草原的研究表明, 自由放牧可增加土壤呼吸的年排放量, 且土壤呼吸与放牧强度呈正相关关系.然而, 也有研究认为, 荒漠草原放牧草地的土壤呼吸速率显著低于未放牧草地(阿木日吉日嘎拉等, 2013).本研究中, 过去不同的放牧强度对土壤呼吸速率的影响具有明显的年际变化, 表现为无显著影响和促进两种.放牧在总体上促进了土壤呼吸, 表现为放牧(LG、MG、HG)区3年平均土壤呼吸速率都高于禁牧区.两者研究结果截然不同, 这里存在两种可能, 一种是放牧增加土壤呼吸, 即使在停止放牧后, 放牧区的土壤呼吸速率仍然较高; 还有一种可能是放牧可能抑制土壤呼吸, 但停止放牧后, 土壤呼吸产生了超补偿, 使得土壤呼吸迅速恢复, 并超过对照区的, 而我们的研究结果属于哪种情况, 我们并不清楚.另外, 本研究是在多年放牧的草地上进行了小区围封, 实际上变成了围封草地, 围封后放牧的扰动已经去除, 草地的生物量和水分条件得到了一定程度的恢复, 从而引起土壤呼吸变化.已有研究也表明, 围封草地的土壤呼吸速率要明显高于放牧草地(贾丙瑞等, 2005), 而且放牧会导致全年的土壤呼吸速率比围封草地降低1/3 (马涛等, 2009).另外, 本研究持续3年测定土壤呼吸速率, 研究年限的不同对草地土壤呼吸的影响也存在差异(郭明英等, 2011). ...
Assessing the response of terrestrial ecosystems to potential changes in precipitation 1 2003
... 荒漠草原地处温带干旱半干旱区域, 无论是植物生长, 还是土壤生态学过程, 都更加受到水分条件的限制.许多研究发现水分添加会显著增加土壤微生物生物量(Liu et al., 2009; Yan et al., 2010), 增强土壤有机质的矿化作用(Harrison et al., 2008; Liu et al., 2009), 这主要是由于土壤水分供给的增加导致营养元素扩散能力的提高, 使其更容易被植物和微生物利用(Sowerby et al., 2010; Reichmann et al., 2013).因此, 水分变化能够影响草原植物根系生长、土壤代谢活力、根系呼吸、土壤微生物群落构成以及微生物活性(Xu et al., 2004), 进而引起土壤呼吸强度的改变.在全球尺度上, 土壤呼吸强度与降水量呈正相关关系(Raich & Schlesinger, 1992; Zhou et al., 2014; Liu et al., 2016).而对于干旱和半干旱生态系统, 土壤湿度的影响尤其显著(Conant et al., 2000), 水分是导致土壤呼吸速率季节波动的重要因素和年际变化的主要驱动因子(Raich et al., 2002).降水可以在短时间内引起土壤呼吸作用的急剧增强(Birch, 1958), 而且这种对土壤呼吸的激发作用在干旱和半干旱地区变得更加显著(Sponseller, 2007).由降水格局改变引起的土壤呼吸变化效应可能远高于其他全球变化因子(增温、氮沉降、CO2升高)(Weltzin et al., 2003).本实验结果表明, 连续3年的加水处理(W和NW)使得土壤含水量明显增加, 对土壤呼吸产生了显著的促进作用, 而氮添加的有效性依赖于水分条件, 两者表现为协同作用.另外, 加水对土壤呼吸的促进作用大小也取决于自然降水的多少, 如在丰水年(2016年)影响不显著, 说明水分对该地区土壤呼吸的影响程度与总体上降水量高低有关, 在降水量较低的年份(平水或欠水年)能够促进土壤呼吸, 反之, 在降水量充沛的年份, 增加降水对土壤呼吸的促进作用有限, 甚至可能抑制土壤呼吸, 因为过量的水分添加会降低土壤通透性, 导致土壤氧气供应不足, 制约土壤呼吸.Liu等(2016)也发现土壤呼吸对增雨的响应随着环境湿润度的增加而减弱. ...
How soil moisture, rain pulses, and growth alter the response of ecosystem respiration to temperature 1 2004
... 荒漠草原地处温带干旱半干旱区域, 无论是植物生长, 还是土壤生态学过程, 都更加受到水分条件的限制.许多研究发现水分添加会显著增加土壤微生物生物量(Liu et al., 2009; Yan et al., 2010), 增强土壤有机质的矿化作用(Harrison et al., 2008; Liu et al., 2009), 这主要是由于土壤水分供给的增加导致营养元素扩散能力的提高, 使其更容易被植物和微生物利用(Sowerby et al., 2010; Reichmann et al., 2013).因此, 水分变化能够影响草原植物根系生长、土壤代谢活力、根系呼吸、土壤微生物群落构成以及微生物活性(Xu et al., 2004), 进而引起土壤呼吸强度的改变.在全球尺度上, 土壤呼吸强度与降水量呈正相关关系(Raich & Schlesinger, 1992; Zhou et al., 2014; Liu et al., 2016).而对于干旱和半干旱生态系统, 土壤湿度的影响尤其显著(Conant et al., 2000), 水分是导致土壤呼吸速率季节波动的重要因素和年际变化的主要驱动因子(Raich et al., 2002).降水可以在短时间内引起土壤呼吸作用的急剧增强(Birch, 1958), 而且这种对土壤呼吸的激发作用在干旱和半干旱地区变得更加显著(Sponseller, 2007).由降水格局改变引起的土壤呼吸变化效应可能远高于其他全球变化因子(增温、氮沉降、CO2升高)(Weltzin et al., 2003).本实验结果表明, 连续3年的加水处理(W和NW)使得土壤含水量明显增加, 对土壤呼吸产生了显著的促进作用, 而氮添加的有效性依赖于水分条件, 两者表现为协同作用.另外, 加水对土壤呼吸的促进作用大小也取决于自然降水的多少, 如在丰水年(2016年)影响不显著, 说明水分对该地区土壤呼吸的影响程度与总体上降水量高低有关, 在降水量较低的年份(平水或欠水年)能够促进土壤呼吸, 反之, 在降水量充沛的年份, 增加降水对土壤呼吸的促进作用有限, 甚至可能抑制土壤呼吸, 因为过量的水分添加会降低土壤通透性, 导致土壤氧气供应不足, 制约土壤呼吸.Liu等(2016)也发现土壤呼吸对增雨的响应随着环境湿润度的增加而减弱. ...
Differential responses of auto- and heterotrophic soil respiration to water and nitrogen addition in a semiarid temperate steppe 2 2010
... 氮和水分是草地生态系统生产力的两个主要限制性因子(Hooper & Johnson, 1999; Yan et al., 2011).鉴于草地退化常导致土壤养分缺乏, 氮添加作为一种重要手段被用于对退化草地的恢复和管理中(潘庆民等, 2005; 张璐等, 2009).氮添加可能影响生态系统光合作用和植物生长, 导致植物生产力的不同响应(Ren et al., 2017), 进而对土壤呼吸产生促进(Song et al., 2011)、抑制(Mo et al., 2007)或无显著影响(Peng et al., 2011; Qi et al., 2014; 李寅龙等, 2015) 3种不同的作用.水分作为干旱区草地生态系统过程重要的限制因子, 通常降水增加对土壤呼吸有显著促进作用(Sowerby et al., 2008; Yan et al., 2010, 2011), 而氮添加的效果常常会依赖于水分条件.刁励玮等(2018)统计分析了1990-2017年间发表的有关氮添加的研究论文结果, 发现单独施氮对地下生物量无显著影响, 氮水同时添加能显著提高地下生物量, 两者对生态系统生产力存在显著的交互作用(Li et al., 2011), 因此氮和水分两者可能共同影响生态系统的碳交换(Hooper & Johnson, 1999). ...
... 荒漠草原地处温带干旱半干旱区域, 无论是植物生长, 还是土壤生态学过程, 都更加受到水分条件的限制.许多研究发现水分添加会显著增加土壤微生物生物量(Liu et al., 2009; Yan et al., 2010), 增强土壤有机质的矿化作用(Harrison et al., 2008; Liu et al., 2009), 这主要是由于土壤水分供给的增加导致营养元素扩散能力的提高, 使其更容易被植物和微生物利用(Sowerby et al., 2010; Reichmann et al., 2013).因此, 水分变化能够影响草原植物根系生长、土壤代谢活力、根系呼吸、土壤微生物群落构成以及微生物活性(Xu et al., 2004), 进而引起土壤呼吸强度的改变.在全球尺度上, 土壤呼吸强度与降水量呈正相关关系(Raich & Schlesinger, 1992; Zhou et al., 2014; Liu et al., 2016).而对于干旱和半干旱生态系统, 土壤湿度的影响尤其显著(Conant et al., 2000), 水分是导致土壤呼吸速率季节波动的重要因素和年际变化的主要驱动因子(Raich et al., 2002).降水可以在短时间内引起土壤呼吸作用的急剧增强(Birch, 1958), 而且这种对土壤呼吸的激发作用在干旱和半干旱地区变得更加显著(Sponseller, 2007).由降水格局改变引起的土壤呼吸变化效应可能远高于其他全球变化因子(增温、氮沉降、CO2升高)(Weltzin et al., 2003).本实验结果表明, 连续3年的加水处理(W和NW)使得土壤含水量明显增加, 对土壤呼吸产生了显著的促进作用, 而氮添加的有效性依赖于水分条件, 两者表现为协同作用.另外, 加水对土壤呼吸的促进作用大小也取决于自然降水的多少, 如在丰水年(2016年)影响不显著, 说明水分对该地区土壤呼吸的影响程度与总体上降水量高低有关, 在降水量较低的年份(平水或欠水年)能够促进土壤呼吸, 反之, 在降水量充沛的年份, 增加降水对土壤呼吸的促进作用有限, 甚至可能抑制土壤呼吸, 因为过量的水分添加会降低土壤通透性, 导致土壤氧气供应不足, 制约土壤呼吸.Liu等(2016)也发现土壤呼吸对增雨的响应随着环境湿润度的增加而减弱. ...
Water regulated effects of photosynthetic substrate supply on soil respiration in a semiarid steppe 2 2011
... 氮和水分是草地生态系统生产力的两个主要限制性因子(Hooper & Johnson, 1999; Yan et al., 2011).鉴于草地退化常导致土壤养分缺乏, 氮添加作为一种重要手段被用于对退化草地的恢复和管理中(潘庆民等, 2005; 张璐等, 2009).氮添加可能影响生态系统光合作用和植物生长, 导致植物生产力的不同响应(Ren et al., 2017), 进而对土壤呼吸产生促进(Song et al., 2011)、抑制(Mo et al., 2007)或无显著影响(Peng et al., 2011; Qi et al., 2014; 李寅龙等, 2015) 3种不同的作用.水分作为干旱区草地生态系统过程重要的限制因子, 通常降水增加对土壤呼吸有显著促进作用(Sowerby et al., 2008; Yan et al., 2010, 2011), 而氮添加的效果常常会依赖于水分条件.刁励玮等(2018)统计分析了1990-2017年间发表的有关氮添加的研究论文结果, 发现单独施氮对地下生物量无显著影响, 氮水同时添加能显著提高地下生物量, 两者对生态系统生产力存在显著的交互作用(Li et al., 2011), 因此氮和水分两者可能共同影响生态系统的碳交换(Hooper & Johnson, 1999). ...
... 氮和水分是草地生态系统生产力的两个主要限制性因子(Hooper & Johnson, 1999; Yan et al., 2011).鉴于草地退化常导致土壤养分缺乏, 氮添加作为一种重要手段被用于对退化草地的恢复和管理中(潘庆民等, 2005; 张璐等, 2009).氮添加可能影响生态系统光合作用和植物生长, 导致植物生产力的不同响应(Ren et al., 2017), 进而对土壤呼吸产生促进(Song et al., 2011)、抑制(Mo et al., 2007)或无显著影响(Peng et al., 2011; Qi et al., 2014; 李寅龙等, 2015) 3种不同的作用.水分作为干旱区草地生态系统过程重要的限制因子, 通常降水增加对土壤呼吸有显著促进作用(Sowerby et al., 2008; Yan et al., 2010, 2011), 而氮添加的效果常常会依赖于水分条件.刁励玮等(2018)统计分析了1990-2017年间发表的有关氮添加的研究论文结果, 发现单独施氮对地下生物量无显著影响, 氮水同时添加能显著提高地下生物量, 两者对生态系统生产力存在显著的交互作用(Li et al., 2011), 因此氮和水分两者可能共同影响生态系统的碳交换(Hooper & Johnson, 1999). ...
氮素添加对内蒙古羊草草原净氮矿化的影响 1 2009
... 氮和水分是草地生态系统生产力的两个主要限制性因子(Hooper & Johnson, 1999; Yan et al., 2011).鉴于草地退化常导致土壤养分缺乏, 氮添加作为一种重要手段被用于对退化草地的恢复和管理中(潘庆民等, 2005; 张璐等, 2009).氮添加可能影响生态系统光合作用和植物生长, 导致植物生产力的不同响应(Ren et al., 2017), 进而对土壤呼吸产生促进(Song et al., 2011)、抑制(Mo et al., 2007)或无显著影响(Peng et al., 2011; Qi et al., 2014; 李寅龙等, 2015) 3种不同的作用.水分作为干旱区草地生态系统过程重要的限制因子, 通常降水增加对土壤呼吸有显著促进作用(Sowerby et al., 2008; Yan et al., 2010, 2011), 而氮添加的效果常常会依赖于水分条件.刁励玮等(2018)统计分析了1990-2017年间发表的有关氮添加的研究论文结果, 发现单独施氮对地下生物量无显著影响, 氮水同时添加能显著提高地下生物量, 两者对生态系统生产力存在显著的交互作用(Li et al., 2011), 因此氮和水分两者可能共同影响生态系统的碳交换(Hooper & Johnson, 1999). ...
Different responses of soil respiration and its components to nitrogen addition among biomes: A meta- analysis 1 2014
... 荒漠草原地处温带干旱半干旱区域, 无论是植物生长, 还是土壤生态学过程, 都更加受到水分条件的限制.许多研究发现水分添加会显著增加土壤微生物生物量(Liu et al., 2009; Yan et al., 2010), 增强土壤有机质的矿化作用(Harrison et al., 2008; Liu et al., 2009), 这主要是由于土壤水分供给的增加导致营养元素扩散能力的提高, 使其更容易被植物和微生物利用(Sowerby et al., 2010; Reichmann et al., 2013).因此, 水分变化能够影响草原植物根系生长、土壤代谢活力、根系呼吸、土壤微生物群落构成以及微生物活性(Xu et al., 2004), 进而引起土壤呼吸强度的改变.在全球尺度上, 土壤呼吸强度与降水量呈正相关关系(Raich & Schlesinger, 1992; Zhou et al., 2014; Liu et al., 2016).而对于干旱和半干旱生态系统, 土壤湿度的影响尤其显著(Conant et al., 2000), 水分是导致土壤呼吸速率季节波动的重要因素和年际变化的主要驱动因子(Raich et al., 2002).降水可以在短时间内引起土壤呼吸作用的急剧增强(Birch, 1958), 而且这种对土壤呼吸的激发作用在干旱和半干旱地区变得更加显著(Sponseller, 2007).由降水格局改变引起的土壤呼吸变化效应可能远高于其他全球变化因子(增温、氮沉降、CO2升高)(Weltzin et al., 2003).本实验结果表明, 连续3年的加水处理(W和NW)使得土壤含水量明显增加, 对土壤呼吸产生了显著的促进作用, 而氮添加的有效性依赖于水分条件, 两者表现为协同作用.另外, 加水对土壤呼吸的促进作用大小也取决于自然降水的多少, 如在丰水年(2016年)影响不显著, 说明水分对该地区土壤呼吸的影响程度与总体上降水量高低有关, 在降水量较低的年份(平水或欠水年)能够促进土壤呼吸, 反之, 在降水量充沛的年份, 增加降水对土壤呼吸的促进作用有限, 甚至可能抑制土壤呼吸, 因为过量的水分添加会降低土壤通透性, 导致土壤氧气供应不足, 制约土壤呼吸.Liu等(2016)也发现土壤呼吸对增雨的响应随着环境湿润度的增加而减弱. ...
 ,1,2,*, 单玉梅1,2, 晔薷罕1,2, 张璞进1,2, 木兰1,2, 常虹1,2, 任婷婷3,4, 陈世苹3,4, 白永飞3,4, 黄建辉3,4, 孙海莲1,2
,1,2,*, 单玉梅1,2, 晔薷罕1,2, 张璞进1,2, 木兰1,2, 常虹1,2, 任婷婷3,4, 陈世苹3,4, 白永飞3,4, 黄建辉3,4, 孙海莲1,2
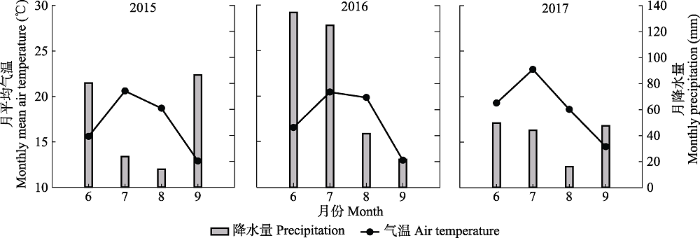 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT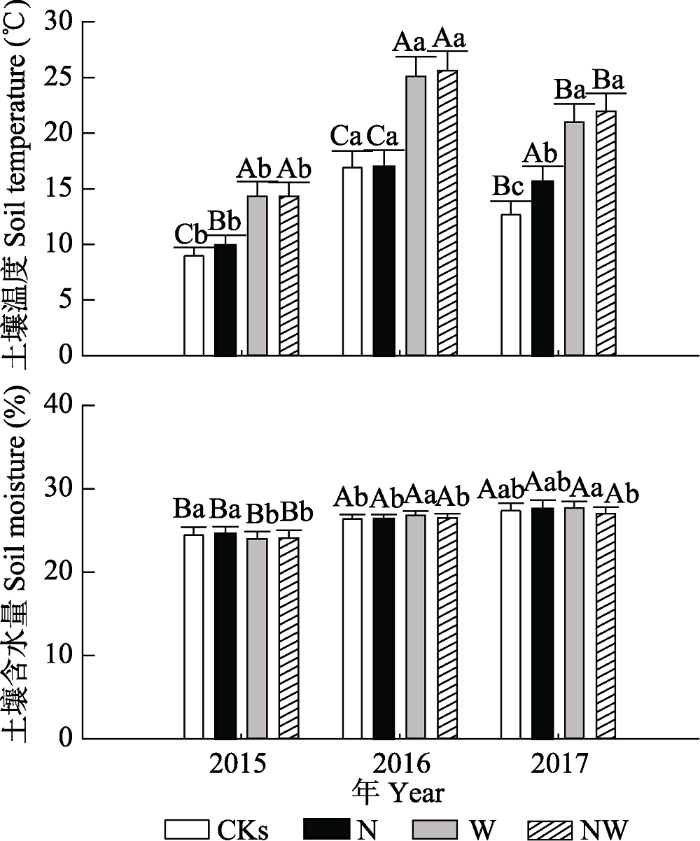 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT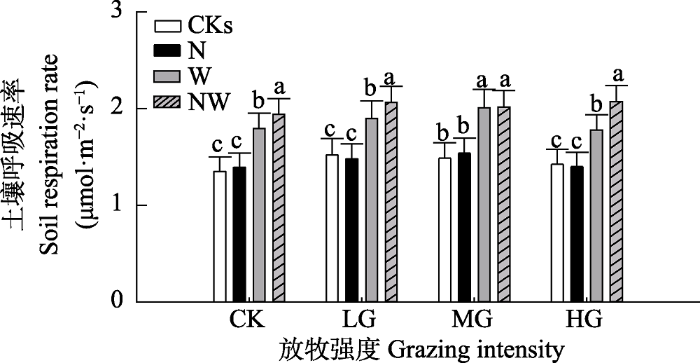 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}