Distribution patterns and driving factors of leaf C, N and P stoichiometry of coniferous species on the eastern Qinghai-Xizang Plateau, China
Qin CAI1,2, Jun-Xiang DING1,2, Zi-Liang ZHANG1, Jun HU1,2, Qi-Tong WANG1,2, Ming-Zhen YIN1,2, Qing LIU1, Hua-Jun YIN,,1,*1 Chengdu Institute of Biology, Chinese Academy of Sciences, Key Laboratory of Mountain Ecological Restoration and Bioresource Utilization of Chinese Academy of Sciences, and Ecological Restoration and Biodiversity Conservation Key Laboratory of Sichuan Province, Chengdu 610041, China 2 University of Chinese Academy of Sciences, Beijing 100049, China
Supported by the Second Qinghai-Xizang Plateau Scientific Expedition and Research Program(STEP2019QZKK0301) Frontier Science Key Research Programs of Chinese Academy of Science(QYZDB-SSW-SMC023) National Natural Science Foundation of China(31872700) National Natural Science Foundation of China(31700387) Key Research Programs of Sichuan Province(18ZDYF1821)
Abstract Aims The leaf stoichiometry and potential driving factors play a vital role in understanding the distribution patterns of plant community and predicting the plant responses to environmental changes. In this study, we aimed to investigate the spatial distribution patterns and driving factors of leaf carbon (C), nitrogen (N) and phosphorus (P) stoichiometry of coniferous species on the eastern Qinghai-Xizang Plateau, China. Methods We collected leaf and soil samples from 29 coniferous tree species at 84 sampling sites on the eastern Qinghai-Xizang Plateau. Linear fitting was used to analyze the variation patterns of leaf stoichiometry along geographical and climatic gradients. Partial redundancy analysis was used to characterize the relative contributions of climate and soil factors to leaf stoichiometry variation patterns. Important findings (1) At the level of family and genus, C and N concentrations as well as C:N of leaves were significantly different across distinct conifer species. The leaf N:P was less than 14, indicating that conifer species in the study region were mainly N-limited. (2) Leaf N and P concentrations showed a consistent distribution pattern along environmental gradients. Specifically, N and P concentrations of leaves were significantly decreased with elevated latitude and altitude, while remarkably increased with the increase of mean annual temperature (MAT) and mean annual precipitation (MAP). In comparison, leaf C concentration had no significant correlation with latitude, altitude, MAT or MAP. (3) The leaf C:N and C:P showed an opposite distribution pattern with leaf N and P concentrations, which significantly increased with elevated latitude and altitude, while markedly declined with the increase of MAT and MAP. Leaf N:P had no significant correlation with altitude, MAT or MAP. (4) The main driving factors of leaf C, N, P concentrations and their stoichiometric characteristics were different. Specifically, soil properties were the main driving factors accounting for the variations of leaf C concentration and N:P. The variations of leaf N and P concentrations as well as ratios of C:N and C:P were primarily explained by climatic factors. Collectively, variations of leaf stoichiometry of coniferous species along environmental gradients in the study region provided a compelling support for the Temperature Biogeochemistry Hypothesis. These findings largely improved the understanding of the distribution patterns and driving mechanism of leaf stoichiometry under changing environments. Keywords:leaf stoichiometry;distribution pattern;driving factor;the eastern Qinghai-Xizang Plateau;subalpine coniferous species
PDF (4278KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 蔡琴, 丁俊祥, 张子良, 胡君, 汪其同, 尹明珍, 刘庆, 尹华军. 青藏高原东缘主要针叶树种叶片碳氮磷化学计量分布格局及其驱动因素. 植物生态学报, 2019, 43(12): 1048-1060. DOI: 10.17521/cjpe.2019.0221 CAI Qin, DING Jun-Xiang, ZHANG Zi-Liang, HU Jun, WANG Qi-Tong, YIN Ming-Zhen, LIU Qing, YIN Hua-Jun. Distribution patterns and driving factors of leaf C, N and P stoichiometry of coniferous species on the eastern Qinghai-Xizang Plateau, China. Chinese Journal of Plant Ecology, 2019, 43(12): 1048-1060. DOI: 10.17521/cjpe.2019.0221
生态化学计量学主要关注生态过程中多种化学元素平衡, 它将生态学不同层次(个体、种群、群落、生态系统以及全球尺度)的研究有机地统一起来, 目前广泛应用于种群动态、物种共存和竞争以及碳(C)、氮(N)、磷(P)生物地球化学循环等研究中, 已成为当前生态学研究的前沿领域(He et al., 2019)。叶片是植物进行光合作用的主要器官, 研究植物叶片C、N、P化学计量对理解植物生长发育过程、物种组成和功能以及养分利用策略等具有重要意义(田地等, 2018)。
Fig. 2Distribution patterns of leaf C, N and P stoichiometry of coniferous species on the Qinghai-Xizang Plateau. Solid red line represents significant correlation (p < 0.05), while the dotted line represents insignificant correlation (p > 0.05).
Fig. 3Relationships between leaf C, N and P stoichiometry and climate factors in coniferous species on the eastern Qinghai- Xizang Plateau. MAP, mean annual precipitation; MAT, mean annual temperature. Solid red line represents significant correlation (p < 0.05), while the dotted line represents insignificant correlation (p > 0.05).
Fig. 4Relationships between leaf C, N and P stoichiometry and soil stoichiometry and pH in coniferous species on the eastern Qinghai-Xizang Plateau. Solid red line represents significant correlation (p < 0.05), while the dotted line represents insignificant correlation (p > 0.05).
Fig. 5Contribution rates of climate and soil nutrients to the leaf C (A)、N (B)、P (C)、C:N (D)、C:P (E)和N:P (F) stoichiometry of coniferous species on the eastern Qinghai-Xizang Plateau (R2%). c and s represent climate and soil nutrients, respectively; cs represents the common interpretation between climate and soil nutrients.
AertsR, Chapin ⅢFS ( 1999). The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns Advances in Ecological Research, 30, 1-67. [本文引用: 1]
BaiYF, WuJG, ClarkCM, PanQM, ZhangLX, ChenSP, WangQB, HanXG ( 2012). Grazing alters ecosystem functioning and C: N: P stoichiometry of grasslands along a regional precipitation gradient Journal of Applied Ecology, 49, 1204-1215. [本文引用: 1]
BaoSD (2000). Soil and Agricultural Chemistry Analysis. 3rd edn. China Agriculture Press, Beijing.
[ 鲍士旦 (2000). 土壤农化分析. 第三版. 中国农业出版社, 北京.]
ChenYH, HanWX, TangLY, TangZY, FangJY ( 2013). Leaf nitrogen and phosphorus concentrations of woody plants differ in responses to climate, soil and plant growth form Ecography, 36, 178-184. [本文引用: 6]
ElserJJ, FaganWF, KerkhoffAJ, SwensonNG, EnquistBJ ( 2010). Biological stoichiometry of plant production: Metabolism, scaling and ecological response to global change New Phytologist, 186, 593-608. DOI:10.1111/j.1469-8137.2010.03214.xURLPMID:20298486 [本文引用: 1] Biological stoichiometry theory considers the balance of multiple chemical elements in living systems, whereas metabolic scaling theory considers how size affects metabolic properties from cells to ecosystems. We review recent developments integrating biological stoichiometry and metabolic scaling theories in the context of plant ecology and global change. Although vascular plants exhibit wide variation in foliar carbon:nitrogen:phosphorus ratios, they exhibit a higher degree of 'stoichiometric homeostasis' than previously appreciated. Thus, terrestrial carbon:nitrogen:phosphorus stoichiometry will reflect the effects of adjustment to local growth conditions as well as species' replacements. Plant stoichiometry exhibits size scaling, as foliar nutrient concentration decreases with increasing plant size, especially for phosphorus. Thus, small plants have lower nitrogen:phosphorus ratios. Furthermore, foliar nutrient concentration is reflected in other tissues (root, reproductive, support), permitting the development of empirical models of production that scale from tissue to whole-plant levels. Plant stoichiometry exhibits large-scale macroecological patterns, including stronger latitudinal trends and environmental correlations for phosphorus concentration (relative to nitrogen) and a positive correlation between nutrient concentrations and geographic range size. Given this emerging knowledge of how plant nutrients respond to environmental variables and are connected to size, the effects of global change factors (such as carbon dioxide, temperature, nitrogen deposition) can be better understood.
FangZ, LiDD, JiaoF, YaoJ, DuHT ( 2019). The latitudinal patterns of leaf and soil C: N: P stoichiometry in the Loess Plateau of China Frontiers in Plant Science, 10, 85. DOI: 10.3389/fpls.2019.00085. DOI:10.3389/fpls.2019.00085URLPMID:30949183 [本文引用: 1] Understanding the spatial patterns and the driving factors of plant leaf and soil stoichiometry are critical for improving the parameterization of future ecological models and to predict the responses of ecosystems to environmental changes. This study aimed to determine how the latitudinal patterns of leaf and soil C:N:P stoichiometry are affected by climate and vegetation types in the dryland ecosystems. The concentrations of leaf C, N, and P in herb community as well as soil nutrient concentrations along a 500-km-long latitudinal gradient in Northern Shaanxi of the Loess Plateau, were measured. The results showed that the soil C, N, P and C:N:P ratios at all three depths (0-10, 10-20, and 20-40 cm) showed significant latitudinal trends (except for soil C:N ratios) (P < 0.01). In general, the soil C, N and C:N:P ratios decreased exponentially while soil P increased first and then decreased with the latitude. The soil C, N, C:P, and N:P ratios at all three depths (0-10, 10-20 and 20-40 cm) were positively correlated with MAT and MAP (P < 0.05), while soil P and C:N ratios at all three depths were weakly correlated with MAT and MAP (P > 0.05). In addition, leaf C:N:P stoichiometry was significantly correlated with the latitude, MAT, and MAP (except for N:P ratios) (P < 0.01), such that, leaf C, C:N, and C:P ratios decreased as the latitude increased and MAT and MAP decreased, and leaf N, P concentrations increased as the latitude increased and MAT and MAP decreased, while leaf N:P ratios were weakly correlated with the latitude, MAT, and MAP (P > 0.05). Furthermore, the leaf C:N:P stoichiometry of herbaceous communities was related to the soil properties (except for soil P), and we found that the C:P ratios between the soil and leaves were strongly correlated. Compared with the global scale, the relatively high N:P ratios indicated that the vegetation growth of the herb community in the dryland of the Loess Plateau was more susceptible to P limitation.
FisherJB, MalhiY, TorresIC, MetcalfeDB, van de WegMJ, MeirP, Silva-EspejoJE, HuascoWH ( 2013). Nutrient limitation in rainforests and cloud forests along a 3,000-m elevation gradient in the Peruvian Andes Oecologia, 172, 889-902. DOI:10.1007/s00442-012-2522-6URLPMID:23180422 [本文引用: 1] We report results from a large-scale nutrient fertilization experiment along a &quot;megadiverse&quot; (154 unique species were included in the study) 3,000-m elevation transect in the Peruvian Andes and adjacent lowland Amazonia. Our objectives were to test if nitrogen (N) and phosphorus (P) limitation shift along this elevation gradient, and to determine how an alleviation of nutrient limitation would manifest in ecosystem changes. Tree height decreased with increasing elevation, but leaf area index (LAI) and diameter at breast height (DBH) did not vary with elevation. Leaf N:P decreased with increasing elevation (from 24 at 200 m to 11 at 3,000 m), suggesting increased N limitation and decreased P limitation with increasing elevation. After 4 years of fertilization (N, P, N + P), plots at the lowland site (200 m) fertilized with N + P showed greater relative growth rates in DBH than did the control plots; no significant differences were evident at the 1,000 m site, and plots fertilized with N at the highest elevation sites (1,500, 3,000 m) showed greater relative growth rates in DBH than did the control plots, again suggesting increased N constraint with elevation. Across elevations in general N fertilization led to an increase in microbial respiration, while P and N + P addition led to an increase in root respiration and corresponding decrease in hyphal respiration. There was no significant canopy response (LAI, leaf nutrients) to fertilization, suggesting that photosynthetic capacity was not N or P limited in these ecosystems. In sum, our study significantly advances ecological understanding of nutrient cycling and ecosystem response in a region where our collective knowledge and data are sparse: we demonstrate N limitation in high elevation tropical montane forests, N and P co-limitation in lowland Amazonia, and a nutrient limitation response manifested not in canopy changes, but rather in stem and belowground changes.
HanWX, FangJY, GuoDL, ZhangY ( 2005). Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China New Phytologist, 168, 377-385. DOI:10.1111/j.1469-8137.2005.01530.xURLPMID:16219077 [本文引用: 5] Leaf nitrogen and phosphorus stoichiometry of Chinese terrestrial plants was studied based on a national data set including 753 species across the country. Geometric means were calculated for functional groups based on life form, phylogeny and photosynthetic pathway, as well as for all 753 species. The relationships between leaf N and P stoichiometric traits and latitude (and temperature) were analysed. The geometric means of leaf N, P, and N : P ratio for the 753 species were 18.6 and 1.21 mg g(-1) and 14.4, respectively. With increasing latitude (decreasing mean annual temperature, MAT), leaf N and P increased, but the N : P ratio did not show significant changes. Although patterns of leaf N, P and N : P ratios across the functional groups were generally consistent with those reported previously, the overall N : P ratio of China's flora was considerably higher than the global averages, probably caused by a greater shortage of soil P in China than elsewhere. The relationships between leaf N, P and N : P ratio and latitude (and MAT) also suggested the existence of broad biogeographical patterns of these leaf traits in Chinese flora.
HeHL, YangXC, LiDD, YinCY, LiYX, ZhouGY, ZhangL, LiuQ ( 2017). Stoichiometric characteristics of carbon, nitrogen and phosphorus of Sibiraea angustata shrub on the eastern Tibetan Plateau. Chinese Journal of Plant Ecology, 41, 126-135. [本文引用: 1]
HeJS, FangJY, WangZH, GuoDL, FlynnDFB, GengZ ( 2006). Stoichiometry and large-scale patterns of leaf carbon and nitrogen in the grassland biomes of China Oecologia, 149, 115-122. DOI:10.1007/s00442-006-0425-0URLPMID:16639565 [本文引用: 4] Nitrogen (N) and carbon-nitrogen (C:N) ratio are key foliar traits with great ecological importance, but their patterns across biomes have only recently been explored. We conducted a systematic census of foliar C, N and C:N ratio for 213 species, from 41 families over 199 research sites across the grassland biomes of China following the same protocol, to explore how different environmental conditions and species composition affect leaf N and C:N stoichiometry. Leaf C:N stoichiometry is stable in three distinct climatic regions in Inner Mongolia, the Tibetan Plateau, and Xinjiang Autonomous Region, despite considerable variations among co-existing species and among different vegetation types. Our results also show that life form and genus identity explain more than 70% of total variations of foliar N and C:N ratio, while mean growing season temperature and growing season precipitation explained only less than 3%. This suggests that, at the biome scale, temperature affects leaf N mainly through a change in plant species composition rather than via temperature itself. When our data were pooled with a global dataset, the previously observed positive correlation between leaf N and mean annual temperature (MAT) at very low MATs, disappeared. Thus, our data do not support the previously proposed biogeochemical hypothesis that low temperature limitations on mineralization of organic matter and N availability in soils lead to low leaf N in cold environments.
HeJS, WangL, FlynnDFB, WangXP, MaWH, FangJY ( 2008). Leaf nitrogen:phosphorus stoichiometry across Chinese grassland biomes Oecologia, 155, 301-310. DOI:10.1007/s00442-007-0912-yURLPMID:18278518 [本文引用: 3] Leaf N and P stoichiometry covaries with many aspects of plant biology, yet the drivers of this trait at biogeographic scales remain uncertain. Recently we reported the patterns of leaf C and N based on systematic census of 213 species over 199 research sites in the grassland biomes of China. With the expanded analysis of leaf P, here we report patterns of leaf P and N:P ratios, and analyze the relative contribution of climatic variables and phylogeny in structuring patterns of leaf N:P stoichiometry. Average values of leaf P and N:P ratio were 1.9 mg g(-1) and 15.3 (mass ratio), respectively, consistent with the previous observation of a higher N:P ratio in China's flora than the global averages (ca. 13.8), resulting from a lower leaf P. Climatic variables had very little direct correlation with leaf P and N:P ratios, with growing season precipitation and temperature together explaining less than 2% of the variation, while inter-site differences and within-site phylogenetic variation explained 55 and 26% of the total variation in leaf P and N:P ratios. Across all sites and species, leaf N and P were highly positively correlated at all levels. However, the within-site, within-species covariations of leaf N and P were weaker than those across sites and across species. Leaf N and P relationships are driven by both variation between sites at the landscape scale (explaining 58% of the variance) and within sites at the local scale (explaining 24%), while the climatic factors exerted limited influence (explaining less than 3%). In addition, leaf N:P ratios in two dominant genera Kobresia and Stipa had different responses to precipitation. This study suggests that geographic variation and between-species variation, rather than climatic variation, are the major determinants of grassland foliar stoichiometry at the biome level.
HeNP, LiuCC, PiaoSL, SackL, XuL, LuoYQ, HeJS, HanXG, ZhouGS, ZhouXH, LinY, YuQ, LiuSR, SunW, NiuSL, LiSG, ZhangJH, YuGR ( 2019). Ecosystem traits linking functional traits to macroecology Trends in Ecology & Evolution, 34, 200-210. DOI:10.1016/j.tree.2018.11.004URLPMID:30527959 [本文引用: 1] As the range of studies on macroecology and functional traits expands, integration of traits into higher-level approaches offers new opportunities to improve clarification of larger-scale patterns and their mechanisms and predictions using models. Here, we propose a framework for quantifying 'ecosystem traits' and means to address the challenges of broadening the applicability of functional traits to macroecology. Ecosystem traits are traits or quantitative characteristics of organisms (plants, animals, and microbes) at the community level expressed as the intensity (or density) normalized per unit land area. Ecosystem traits can inter-relate and integrate data from field trait surveys, eddy-flux observation, remote sensing, and ecological models, and thereby provide new resolution of the responses and feedback at regional to global scale.
KoerselmanW, MeulemanAFM ( 1996). The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation Journal of Applied Ecology, 33, 1441-1450. [本文引用: 1]
LaiJS, MiXC (2012). Ordination analysis of ecological data using vegan package in R. In: Chinese National Committee for DIVERSITAS eds. Advances in Biodiversity Conservation and Research in China Ⅸ. China Meteorological Press , Beijing. 332-343.
MindenV, KleyerM ( 2014). Internal and external regulation of plant organ stoichiometry Plant Biology, 16, 897-907. DOI:10.1111/plb.12155URLPMID:24552639 [本文引用: 1] Internal differences between plant organs are caused by the functional differentiation of plant tissue, whereas external supply rates of elements constrain nutrient uptake. Previous studies have concentrated on foliar or whole-plant stoichiometric response to the environment, whereas investigation of organ-specific comparisons is still pending. We explore C:N:P ratios of stems, leaves, diaspores and belowground organs in marsh plants, and evaluate the influence of environmental constraints using standardised major axis regression (SMA). For a pooled dataset, SMA resulted in distinct patterns of isometric and anisometric slopes between plant organs. Bivariate line-fitting for a split dataset of four ecological groups revealed that species of the frequently inundated marsh had higher N:C ratios than those of the infrequently inundated marsh. The influence of nutrient availability was detectable in decreased P:C and increased N:P ratios in P-poor sites. Across ecological groups, leaves and diaspores showed higher elemental homeostasis than stems and belowground organs. Any change in N:C ratios of belowground organs and diaspores in response to the environment was accompanied by an even stronger internal change in stem N:C ratios, indicating a pivotal role of stems of herbaceous plants in ecosystem processes. We found distinct patterns of C:N:P ratios in plant organs related to their internal function and external environmental constraints. Leaves and diaspores showed a higher degree of homeostasis than stems and belowground organs. We detected a clear external signal in element:element ratios of plant organs, with low soil P translating into lower tissue P:C ratio and stronger N retention in leaves as a response to salt stress.
ReichPB, OleksynJ ( 2004). Global patterns of plant leaf N and P in relation to temperature and latitude Proceedings of the National Academy of Sciences of the United States of America, 101, 11001-11006. DOI:10.1073/pnas.0403588101URLPMID:15213326 [本文引用: 8] A global data set including 5,087 observations of leaf nitrogen (N) and phosphorus (P) for 1,280 plant species at 452 sites and of associated mean climate indices demonstrates broad biogeographic patterns. In general, leaf N and P decline and the N/P ratio increases toward the equator as average temperature and growing season length increase. These patterns are similar for five dominant plant groups, coniferous trees and four angiosperm groups (grasses, herbs, shrubs, and trees). These results support the hypotheses that (i) leaf N and P increase from the tropics to the cooler and drier midlatitudes because of temperature-related plant physiological stoichiometry and biogeographical gradients in soil substrate age and then plateau or decrease at high latitudes because of cold temperature effects on biogeochemistry and (ii) the N/P ratio increases with mean temperature and toward the equator, because P is a major limiting nutrient in older tropical soils and N is the major limiting nutrient in younger temperate and high-latitude soils.
RenSJ, YuGR, TaoB, WangSQ ( 2007). Leaf nitrogen and phosphorus stoichiometry across 654 terrestrial plant species in NSTEC Chinese Journal of Environmental Science, 28, 2665-2673. [本文引用: 1]
SardansJ, Pe?uelasJ ( 2013). Tree growth changes with climate and forest type are associated with relative allocation of nutrients, especially phosphorus, to leaves and wood Global Ecology and Biogeography, 22, 494-507. [本文引用: 1]
ShiWQ, WangGA, HanWX ( 2012). Altitudinal variation in leaf nitrogen concentration on the eastern slope of Mount Gongga on the Tibetan Plateau, China PLOS ONE, 7, e44628. DOI: 10.1371/journal.pone.0044628. DOI:10.1371/journal.pone.0044628URLPMID:23028570 [本文引用: 3] Mount Gongga spans 6500 m in elevation and has intact and continuous vertical vegetation belts, ranging from subtropical evergreen broad-leaved vegetation to an alpine frigid sparse grass and desert zone. Investigating the altitudinal trends in leaf nitrogen (N) on Mount Gongga can increase our understanding of the global biogeography of foliar N. In this study, 460 leaf samples from mosses, ferns, and seed plants were collected along an altitudinal gradient on the eastern slope of Mount Gongga, and the variation in leaf N concentration (mass basis) with elevation was analyzed. There are considerable differences in leaf N between mosses and ferns, mosses and seed plants, C(4) and C(3) plants, and evergreen and deciduous woody plants. The general altitudial pattern of leaf N in Mount Gongga plants was that leaf N kept increasing until an elevation of about 2200 m above sea level, with a corresponding mean annual temperature (MAT) of 8.5°C, and then decreased with increasing elevation. However, the evergreen woody plants displayed a decline trend in leaf N across the altitude gradient. Our findings provide an insight into the altitudinal variation in leaf N.
SistlaSA, SchimelJP ( 2012). Stoichiometric flexibility as a regulator of carbon and nutrient cycling in terrestrial ecosystems under change New Phytologist, 196, 68-78. DOI:10.1111/j.1469-8137.2012.04234.xURLPMID:22924404 [本文引用: 1] Ecosystems across the biosphere are subject to rapid changes in elemental balance and climatic regimes. A major force structuring ecological responses to these perturbations lies in the stoichiometric flexibility of systems - the ability to adjust their elemental balance whilst maintaining function. The potential for stoichiometric flexibility underscores the utility of the application of a framework highlighting the constraints and consequences of elemental mass balance and energy cycling in biological systems to address global change phenomena. Improvement in the modeling of ecological responses to disturbance requires the consideration of the stoichiometric flexibility of systems within and across relevant scales. Although a multitude of global change studies over various spatial and temporal scales exist, the explicit consideration of the role played by stoichiometric flexibility in linking micro-scale to macro-scale biogeochemical processes in terrestrial ecosystems remains relatively unexplored. Focusing on terrestrial systems under change, we discuss the mechanisms by which stoichiometric flexibility might be expressed and connected from organisms to ecosystems. We suggest that the transition from the expression of stoichiometric flexibility within individuals to the community and ecosystem scales is a key mechanism regulating the extent to which environmental perturbation may alter ecosystem carbon and nutrient cycling dynamics.
TangZY, XuWT, ZhouGY, BaiYF, LiJX, TangXL, ChenDM, LiuQ, MaWH, XiongGM, HeHL, HeNP, GuoYP, GuoQ, ZhuJL, HanWX, HuHF, FangJY, XieZQ ( 2018). Patterns of plant carbon, nitrogen, and phosphorus concentration in relation to productivity in China’s terrestrial ecosystems Proceedings of the National Academy of Sciences of the United States of America, 115, 4033-4038. DOI:10.1073/pnas.1700295114URLPMID:29666316 [本文引用: 2] Plant nitrogen (N) and phosphorus (P) content regulate productivity and carbon (C) sequestration in terrestrial ecosystems. Estimates of the allocation of N and P content in plant tissues and the relationship between nutrient content and photosynthetic capacity are critical to predicting future ecosystem C sequestration under global change. In this study, by investigating the nutrient concentrations of plant leaves, stems, and roots across China's terrestrial biomes, we document large-scale patterns of community-level concentrations of C, N, and P. We also examine the possible correlation between nutrient content and plant production as indicated by vegetation gross primary productivity (GPP). The nationally averaged community concentrations of C, N, and P were 436.8, 14.14, and 1.11 mg·g-1 for leaves; 448.3, 3.04 and 0.31 mg·g-1 for stems; and 418.2, 4.85, and 0.47 mg·g-1 for roots, respectively. The nationally averaged leaf N and P productivity was 249.5 g C GPP·g-1 N·y-1 and 3,157.9 g C GPP·g-1 P·y-1, respectively. The N and P concentrations in stems and roots were generally more sensitive to the abiotic environment than those in leaves. There were strong power-law relationships between N (or P) content in different tissues for all biomes, which were closely coupled with vegetation GPP. These findings not only provide key parameters to develop empirical models to scale the responses of plants to global change from a single tissue to the whole community but also offer large-scale evidence of biome-dependent regulation of C sequestration by nutrients.
TianD, YanZB, FangJY ( 2018). Plant stoichiometry: A research frontier in ecology Chinese Journal of Nature, 40, 235-241. [本文引用: 2]
TianD, YanZB, MaSH, DingYH, LuoYK, ChenYH, DuEZ, HanWX, KovacsED, ShenHH, HuHF, KattgeJ, SchmidB, FangJY ( 2019). Family-level leaf nitrogen and phosphorus stoichiometry of global terrestrial plants Science China Life Sciences, 62, 1047-1057. DOI:10.1007/s11427-019-9584-1URLPMID:31290101 [本文引用: 1] Leaf nitrogen (N) and phosphorus (P) concentrations are critical for photosynthesis, growth, reproduction and other ecological processes of plants. Previous studies on large-scale biogeographic patterns of leaf N and P stoichiometric relationships were mostly conducted using data pooled across taxa, while family/genus-level analyses are rarely reported. Here, we examined global patterns of family-specific leaf N and P stoichiometry using a global data set of 12,716 paired leaf N and P records which includes 204 families, 1,305 genera, and 3,420 species. After determining the minimum size of samples (i.e., 35 records), we analyzed leaf N and P concentrations, N:P ratios and N~P scaling relationships of plants for 62 families with 11,440 records. The numeric values of leaf N and P stoichiometry varied significantly across families and showed diverse trends along gradients of mean annual temperature (MAT) and mean annual precipitation (MAP). The leaf N and P concentrations and N:P ratios of 62 families ranged from 6.11 to 30.30 mg g-1, 0.27 to 2.17 mg g-1, and 10.20 to 35.40, respectively. Approximately 1/3-1/2 of the families (22-35 of 62) showed a decrease in leaf N and P concentrations and N:P ratios with increasing MAT or MAP, while the remainder either did not show a significant trend or presented the opposite pattern. Family-specific leaf N~P scaling exponents did not converge to a certain empirical value, with a range of 0.307-0.991 for 54 out of 62 families which indicated a significant N~P scaling relationship. Our results for the first time revealed large variation in the family-level leaf N and P stoichiometry of global terrestrial plants and that the stoichiometric relationships for at least one-third of the families were not consistent with the global trends reported previously. The numeric values of the family-specific leaf N and P stoichiometry documented in the current study provide critical synthetic parameters for biogeographic modeling and for further studies on the physiological and ecological mechanisms underlying the nutrient use strategies of plants from different phylogenetic taxa.
TianD, YanZB, NiklasKJ, HanWX, KattgeJ, ReichPB, LuoYK, ChenYH, TangZY, HuHF, WrightIJ, SchmidB, FangJY ( 2017). Global leaf nitrogen and phosphorus stoichiometry and their scaling exponent National Science Review, 5, 728-739. DOI:10.1002/ecy.2812URLPMID:31291467 [本文引用: 3] Nitrogen (N) and phosphorus (P) are essential components of the basic cell structure of plants. In particular, leaf N and P concentrations and their stoichiometric relationship largely determine the photosynthesis, growth, reproduction, and ecophysiological processes of plants. As important leaf functional traits, leaf N and P concentrations and their stoichiometric relationship play vital roles in indicating plant nutrient-use strategies and their evolution in terrestrial ecosystems. They also influence physiological and ecological processes in leaves (e.g., growth rate and energy metabolism) and productivity (e.g., net primary production and net ecosystem production) at ecosystem level. However, the lack of a comprehensive data set containing paired leaf N and P concentration records has distinctly limited research on nutrient stoichiometry and leaf functional traits. Here, we provide a global database of paired records of leaf N and P concentrations. A total of 11,354 individual records were acquired spanning 1,291 sites worldwide, including 201 families, 1,265 genera, and 3,227 species. The records span a latitudinal range of 45.28 °S to 68.35 °N and a longitudinal range of 155.5 °W to 168.0 °E. The variables provided for each individual record are (1) geographical location (longitude, latitude, and altitude); (2) matched leaf N and P concentrations and N:P ratio; (3) taxonomic information (family, genera, and species); (4) life form (angiosperm/gymnosperm, monocotyledonous/dicotyledonous and woody plants/herbaceous plants; note that woody plants were further divided into coniferous, deciduous broad-leaved, and evergreen broad-leaved woody species and that herbaceous plants were further divided into annual and perennial species); (5) mean annual temperature (MAT) and mean annual precipitation (MAP); and (6) soil N and P concentrations and pH value in some records. To date, this database is the world's largest database of paired leaf N and P concentrations, which contains matched information of geographical location, environmental factors, and taxa. We believe that the database will play a fundamental and crucial part of ecological stoichiometric studies. There are no copyright restrictions. When using this database, we kindly request that you cite this article, respecting all the authors' hard work during sample collection and data compilation.
TylerG, OlssonT ( 2001). Plant uptake of major and minor mineral elements as influenced by soil acidity and liming Plant and Soil, 230, 307-321. [本文引用: 1]
WangZ, XiaCX, YuD, WuZG ( 2015). Low-temperature induced leaf elements accumulation in aquatic macrophytes across Tibetan Plateau Ecological Engineering, 75, 1-8. DOI:10.1016/j.ecoleng.2014.11.015URL [本文引用: 1]
XiaCX, YuD, WangZ, XieD ( 2014). Stoichiometry patterns of leaf carbon, nitrogen and phosphorous in aquatic macrophytes in eastern China Ecological Engineering, 70, 406-413. DOI:10.3389/fpls.2018.01398URLPMID:30298083 [本文引用: 1] Ecological stoichiometry is a powerful indicator for understanding the adaptation of plants to environment. However, understanding of stoichiometric characteristics of leaf carbon (C%), nitrogen (N%), and phosphorus (P%) for aquatic macrophytes remains limited. In this study, 707 samples from 146 sites were collected to study the variations in leaf C%, N%, and P%, and tried to explore how different environmental conditions affect leaf C, N, and P stoichiometry. Results showed that the mean values of leaf C%, N%, P%, and N:P ratios were 39.95%, 2.12%, 0.14%, and 16.60% of macrophytes across the arid zone of northwestern China, respectively. And the mean values of leaf P% were lower than those from the Tibetan Plateau and eastern China, which maybe due to an adaptation strategy of the plants to the unique conditions in the arid zone in the long-term evolutionary process. The higher N:P ratios suggested that P was established as the limiting factor of the macrophytes communities in the arid zone of northwestern China. There were significant differences in leaf C%, N%, P%, and their ratios among different life forms. Our results also showed strong relationships between leaf N% and N:P ratios and longitude, leaf N%, P%, and N:P ratios and latitude, and leaf N% and P% and altitude, respectively. In addition, the results showed that pH can significantly influence leaf C%. Our results supported the temperature-plant physiology hypothesis owing to a negative relationship between leaf N% and P% of macrophytes and mean annual temperature in the arid zone of northwestern China. The different patterns of leaf stoichiometry between the arid zone of northwestern China and eastern China indicated that there were different physiological and ecological adaptability of macrophytes to environmental gradients in different climatic zones.
YangH, YinCY, ZhengDH, TangB, ZhaoWQ, LiN, PuXZ, LiuQ ( 2017). Leaf C:N:P stoichiometry in a growing season and nongrowing season of Picea asperata and Abies faxoniana, dominant tree species in subalpine coniferous forests of western Sichuan. Chinese Journal of Applied and Environmental Biology, 23, 1089-1095. [本文引用: 2]
YangL, SunH, FanYW, HanW, ZengLB, LiuC, WangXP ( 2017). Changes in leaf nitrogen and phosphorus stoichiometry of woody plants along an altitudinal gradient in Changbai Mountain, China Chinese Journal of Plant Ecology, 41, 1228-1238. [本文引用: 1]
YuQ, ElserJJ, HeNP, WuHH, ChenQS, ZhangGM, HanXG ( 2011). Stoichiometric homeostasis of vascular plants in the Inner Mongolia grassland Oecologia, 166, 1-10. DOI:10.1007/s00442-010-1902-zURLPMID:21221646 [本文引用: 1] Stoichiometric homeostasis, the degree to which an organism maintains its C:N:P ratios around a given species- or stage-specific value despite variation in the relative availabilities of elements in its resource supplies, is a key parameter in ecological stoichiometry. However, its regulation and role in affecting organismal and ecosystem processes is still poorly understood in vascular plants. We performed a sand culture experiment and a field nitrogen (N) and phosphorus (P) addition experiment to evaluate the strength of N, P and N:P homeostasis in higher plants in the Inner Mongolia grassland. Our results showed that homeostatic regulation coefficients (H) of vascular plants ranged from 1.93 to 14.5. H varied according to plant species, aboveground and belowground compartments, plant developmental stage, and overall plant nutrient content and N:P ratio. H for belowground and for foliage were inversely related, while H increased with plant developmental stage. H for N (H(N)) was consistently greater than H for P (H(P)) while H for N:P (H(N:P)) was consistently greater than H(N) and H(P). Furthermore, species with greater N and P contents and lower N:P were less homeostatic, suggesting that more homeostatic plants are more conservative nutrient users. The results demonstrate that H of plants encompasses a considerable range but is stronger than that of algae and fungi and weaker than that of animals. This is the first comprehensive evaluation of factors influencing stoichiometric homeostasis in vascular plants.
YuanZY, ChenHYH ( 2009). Global trends in senesced-leaf nitrogen and phosphorus Global Ecology and Biogeography, 18, 532-542. DOI:10.1111/geb.2009.18.issue-5URL [本文引用: 2]
ZhangGQ, ZhangP, PengSZ, ChenYM, CaoY ( 2017). The coupling of leaf, litter, and soil nutrients in warm temperate forests in northwestern China Scientific Reports, 7, 11754. DOI: 10.1038/s41598-017-12199-5. DOI:10.1038/s41598-017-12199-5URLPMID:28924160 [本文引用: 2] The nutrient ecological stoichiometry of plants and soil is important for the growth and dynamics of species, but the stoichiometric relationships among leaf, litter, and soil remain poorly understood. We analyzed the carbon (C), nitrogen (N), and phosphorus (P) stoichiometry of the leaves, litter, and soil for 31 species at 140 sites in warm temperate forests in northwestern China to document the patterns of nutrient traits and their relationships with climatic factors. The average concentrations of C, N, and P in the combined forests were 462.97, 18.04, and 1.32?g?kg-1 for leaves, 365.12, 12.34, and 0.87?g?kg-1 for litter, and 15.72, 1.29, and 0.54?g?kg-1 for soil, respectively. The concentrations differed significantly among the leaves, litter, and soil. Leaf and soil nutrients were not significantly correlated, whereas leaf and litter nutrients and litter and soil nutrients were significantly correlated, indicating that litter provided a link between leaves and soil and demonstrating the nutrient associations among leaves, litter, and soil. Soil nutrients were strongly correlated with climatic factors, and precipitation had a larger impact than temperature on the plants and soil. This study will help to predict the growth and dynamics of species under environmental changes.
ZhangLX, BaiYF, HanXG ( 2004). Differential responses of N:P stoichiometry of Leymus chinensis and Carex korshinskyi to N additions in a steppe ecosystem in Nei Mongol. Acta Botanica Sinica, 46, 259-270. [本文引用: 1]
ZhangMM, GaoRX ( 2012). Research review on comparative anatomy and ecological anatomy of conifers blade Forest Engineering, 28, 9-13. [本文引用: 1]
ZhangY, LiC, WangML ( 2019). Linkages of C:N:P stoichiometry between soil and leaf and their response to climatic factors along altitudinal gradients Journal of Soils and Sediments, 19, 1820-1829. [本文引用: 2]
ZhaoN, HeNP, WangQF, ZhangXY, WangRL, XuZW, YuGR ( 2014). The altitudinal patterns of leaf C:N:P stoichiometry are regulated by plant growth form, climate and soil on Changbai Mountain, China PLOS ONE, 9, e95196. DOI: 10.1371//journal.pone.0095196. DOI:10.1371/journal.pone.0095196URLPMID:24743878 [本文引用: 3] Understanding the geographic patterns and potential drivers of leaf stoichiometry is critical for modelling the nutrient fluxes of ecosystems and to predict the responses of ecosystems to global changes. This study aimed to explore the altitudinal patterns and potential drivers of leaf C∶N∶P stoichiometry. We measured the concentrations of leaf C, N and P in 175 plant species as well as soil nutrient concentrations along an altitudinal transect (500-2300 m) on the northern slope of Changbai Mountain, China to explore the response of leaf C∶N∶P stoichiometry to plant growth form (PGF), climate and soil. Leaf C, N, P and C∶N∶P ratios showed significant altitudinal trends. In general, leaf C and C∶N∶P ratios increased while leaf N and P decreased with elevation. Woody and herbaceous species showed different responses to altitudinal gradients. Trees had the largest variation in leaf C, C∶N and C∶P ratios, while herbs showed the largest variation in leaf N, P and N∶P ratio. PGF, climate and soil jointly regulated leaf stoichiometry, explaining 17.6% to 52.1% of the variation in the six leaf stoichiometric traits. PGF was more important in explaining leaf stoichiometry variation than soil and climate. Our findings will help to elucidate the altitudinal patterns of leaf stoichiometry and to model ecosystem nutrient cycling.
ZhaoWQ, ReichPB, YuQN, ZhaoN, YinCY, ZhaoCZ, LiDD, HuJ, LiT, YinHJ, LiuQ ( 2018). Shrub type dominates the vertical distribution of leaf C:N:P stoichiometry across an extensive altitudinal gradient Biogeosciences, 15, 2033-2053. [本文引用: 6]
The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns 1 1999
Leaf nitrogen and phosphorus concentrations of woody plants differ in responses to climate, soil and plant growth form 6 2013
... 近年来, 围绕植物叶片化学计量特征在区域(Chen et al., 2013; Fang et al., 2019)、国家(Tang et al., 2018; Liu et al., 2019)以及全球(Tian et al., 2017, 2019)尺度上开展了大量研究, 极大地提升和丰富了人们对叶片化学计量特征及相关理论的认识和理解.然而, 目前关于植物叶片化学计量分布规律与空间变异格局仍缺乏统一性结论.例如, 植物叶片元素含量随海拔增加呈现出无显著变化, 或显著增加, 或显著降低, 甚至先增加后减少的差异化变异规律(Fisher et al., 2013; Zhao et al., 2014, 2018).此外, 前人研究也表明植物叶片N:P随纬度增加表现出无显著变化或显著降低等多种变异趋势(Reich & Oleksyn, 2004; Han et al., 2005).基于此, 相关****提出了众多理论假说来解释植物叶片化学计量特征多样化的空间变化规律, 如温度生物地球化学假说(TBH)和温度植物生理假说(TPPH).前者主要认为低温会抑制微生物活性和土壤有机质分解与矿化, 降低土壤N、P养分可利用性, 从而导致低温环境下植物叶片N、P含量较低.而后一种假说则认为低温降低了植物的生物化学反应速率, 导致植物富N的酶及富P的RNA活性下降.相应地, 低温地区植物主要通过提高叶片N、P含量以增强其新陈代谢水平(Reich & Oleksyn, 2004; Han et al., 2005).以上两种假说分别诠释了植物叶片N、P含量随温度变化而呈现出的两种截然相反的变异规律.目前上述两种假说在不同研究中得到了相应地验证.例如, 一些研究结果支持TBH (Xia et al., 2014; Zhao et al., 2014), 而另外一些研究结果则支持TPPH (Wang et al., 2015; Tian et al., 2017), 甚至部分研究结果同时支持上述两种理论假说(Reich & Oleksyn, 2004; Shi et al., 2012).因此, 亟需选择不同典型植物, 进一步加强植物叶片化学计量空间分布格局的研究, 以验证和丰富多变环境条件下植物叶片化学计量特征变化理论假说和植物养分利用策略的科学认知. ...
... 此外, 影响植物叶片化学计量特征和空间分布格局的因素众多, 主要包括气候因子(Reich & Oleksyn, 2004; Yuan & Chen, 2009)、土壤特性(Chen et al., 2013; Zhang et al., 2019)和生物因素(如植物种类、生活型等)(He et al., 2006, 2008; Zhao et al., 2018).研究表明, 当研究对象涉及的植物生活型较多时, 生活型往往成为植物叶片化学计量特征变异的关键驱动因素, 并很大程度上弱化了气候因子和土壤特性的影响效应(He et al., 2006, 2008; Zhao et al., 2018).此外, 不同生活型植物叶片化学计量特征及其空间分布规律也存在较大差异.例如, 草本植物叶片N、P含量通常显著高于乔木, 而落叶树种叶片元素含量显著高于常绿树种, 且两者的叶片化学计量海拔分布格局也存在较明显的差异(Han et al., 2005; Shi et al., 2012).总之, 当生活型与气候因子、土壤特性等多种因素叠加后, 势必会加剧植物叶片化学计量特征分布规律及其驱动因素认知的复杂性和不确定性.相应地, 通过降低或者排除不同生活型对植物叶片化学计量特征的潜在影响, 专一性地聚焦生活型相同或相近的植物叶片化学计量空间分布规律及其驱动因素的深入研究, 将在很大程度上丰富不同环境因子对叶片化学计量特征与空间分布变异相对贡献的认知水平. ...
... 本研究发现: (1)该区域叶片C含量平均值为504.83 g·kg-1, 高于其他****对中国西北温带森林针叶树种(Zhang et al., 2017)、中国陆地针叶树种(Tang et al., 2018)叶片C含量的研究结果.推测其原因可能与植物自身的生长特性以及该区域的气候特点有关.C是植物的结构性元素, 是植物生长发育和繁殖等生理活动的物质基础和能量来源.有研究表明, 较高的叶片C含量有助于延长植物叶片寿命并能提高其对温度、水分和养分贫瘠等胁迫环境的适应力(张明明和高瑞馨, 2012; 贺合亮等, 2017).此外, 本研究区域位于高寒的青藏高原, 温度较低且生长季较短.因此, 本研究区域针叶树种叶片C含量相对较高.(2)该区域针叶树种叶片N含量平均值为13.03 g·kg-1,叶片P含量平均值为1.30 g·kg-1.低于其他****关于贡嘎山针叶树种(Shi et al., 2012)、中国东部针叶树种(Chen et al., 2013)和长白山针叶树种(杨蕾等, 2017)等叶片N、P含量的研究结果, 而高于一些****对中国陆地针叶树种(Han et al., 2005)、中国东部针叶树种(任书杰等, 2007)、全球陆地针叶树种(Tian et al., 2017)和中国西北温带森林针叶树种(Zhang et al., 2017)等叶片N、P含量的研究结果.总体而言, 青藏高原主要针叶树种叶片N、P含量与其他研究结果相比无明显规律, 我们推测其原因可能源于不同植物叶片N、P含量变异系数较大, 比如不同水分、光照和养分等环境条件下植物在生长发育过程的N、P养分需求存在较大差异, 致使不同植物叶片N、P含量在不同生境中具有差别(田地等, 2018).此外, 该区域主要针叶树种叶片N:P均值为10.01, 满足Koerselman和Meuleman (1996)和Zhang等(2004)对植物生长N限制的评定标准(N:P < 14和N:P < 21), 进一步佐证了青藏高原东缘针叶林生长发育主要受N限制(杨欢等, 2017).另外, 该区域各针叶树种叶片C、N含量和C:N在科和属水平上差异显著, 表明物种类型对植物叶片C、N含量和C:N影响较大. ...
... 前人众多研究已清楚地表明, 气候因子(Reich & Oleksyn 2004; Yuan & Chen, 2009)、土壤特性(Chen et al., 2013; Zhang et al., 2019)和植物生活型(He et al., 2006, 2008; Zhao et al., 2018)对植物叶片化学计量特征有重要影响.对中国草地(He et al., 2006)和西南灌丛(Zhao et al., 2018)植物叶片C、N、P化学计量主要驱动因素的研究结果显示, 生活型是驱动叶片化学计量变异的主要关键因子, 并一定程度上弱化了环境因子对植物叶片化学计量特征变异的相对贡献率.在本研究中, 我们通过选择青藏高原东缘单一生活型的针叶树种叶片为研究对象, 尽可能去除或弱化生活型的潜在影响, 专一性地辨识了单一生活型植物叶片空间变异格局以及气候因子和土壤特性对叶片化学计量的相对贡献. ...
The latitudinal patterns of leaf and soil C: N: P stoichiometry in the Loess Plateau of China 1 2019
... 近年来, 围绕植物叶片化学计量特征在区域(Chen et al., 2013; Fang et al., 2019)、国家(Tang et al., 2018; Liu et al., 2019)以及全球(Tian et al., 2017, 2019)尺度上开展了大量研究, 极大地提升和丰富了人们对叶片化学计量特征及相关理论的认识和理解.然而, 目前关于植物叶片化学计量分布规律与空间变异格局仍缺乏统一性结论.例如, 植物叶片元素含量随海拔增加呈现出无显著变化, 或显著增加, 或显著降低, 甚至先增加后减少的差异化变异规律(Fisher et al., 2013; Zhao et al., 2014, 2018).此外, 前人研究也表明植物叶片N:P随纬度增加表现出无显著变化或显著降低等多种变异趋势(Reich & Oleksyn, 2004; Han et al., 2005).基于此, 相关****提出了众多理论假说来解释植物叶片化学计量特征多样化的空间变化规律, 如温度生物地球化学假说(TBH)和温度植物生理假说(TPPH).前者主要认为低温会抑制微生物活性和土壤有机质分解与矿化, 降低土壤N、P养分可利用性, 从而导致低温环境下植物叶片N、P含量较低.而后一种假说则认为低温降低了植物的生物化学反应速率, 导致植物富N的酶及富P的RNA活性下降.相应地, 低温地区植物主要通过提高叶片N、P含量以增强其新陈代谢水平(Reich & Oleksyn, 2004; Han et al., 2005).以上两种假说分别诠释了植物叶片N、P含量随温度变化而呈现出的两种截然相反的变异规律.目前上述两种假说在不同研究中得到了相应地验证.例如, 一些研究结果支持TBH (Xia et al., 2014; Zhao et al., 2014), 而另外一些研究结果则支持TPPH (Wang et al., 2015; Tian et al., 2017), 甚至部分研究结果同时支持上述两种理论假说(Reich & Oleksyn, 2004; Shi et al., 2012).因此, 亟需选择不同典型植物, 进一步加强植物叶片化学计量空间分布格局的研究, 以验证和丰富多变环境条件下植物叶片化学计量特征变化理论假说和植物养分利用策略的科学认知. ...
Nutrient limitation in rainforests and cloud forests along a 3,000-m elevation gradient in the Peruvian Andes 1 2013
... 近年来, 围绕植物叶片化学计量特征在区域(Chen et al., 2013; Fang et al., 2019)、国家(Tang et al., 2018; Liu et al., 2019)以及全球(Tian et al., 2017, 2019)尺度上开展了大量研究, 极大地提升和丰富了人们对叶片化学计量特征及相关理论的认识和理解.然而, 目前关于植物叶片化学计量分布规律与空间变异格局仍缺乏统一性结论.例如, 植物叶片元素含量随海拔增加呈现出无显著变化, 或显著增加, 或显著降低, 甚至先增加后减少的差异化变异规律(Fisher et al., 2013; Zhao et al., 2014, 2018).此外, 前人研究也表明植物叶片N:P随纬度增加表现出无显著变化或显著降低等多种变异趋势(Reich & Oleksyn, 2004; Han et al., 2005).基于此, 相关****提出了众多理论假说来解释植物叶片化学计量特征多样化的空间变化规律, 如温度生物地球化学假说(TBH)和温度植物生理假说(TPPH).前者主要认为低温会抑制微生物活性和土壤有机质分解与矿化, 降低土壤N、P养分可利用性, 从而导致低温环境下植物叶片N、P含量较低.而后一种假说则认为低温降低了植物的生物化学反应速率, 导致植物富N的酶及富P的RNA活性下降.相应地, 低温地区植物主要通过提高叶片N、P含量以增强其新陈代谢水平(Reich & Oleksyn, 2004; Han et al., 2005).以上两种假说分别诠释了植物叶片N、P含量随温度变化而呈现出的两种截然相反的变异规律.目前上述两种假说在不同研究中得到了相应地验证.例如, 一些研究结果支持TBH (Xia et al., 2014; Zhao et al., 2014), 而另外一些研究结果则支持TPPH (Wang et al., 2015; Tian et al., 2017), 甚至部分研究结果同时支持上述两种理论假说(Reich & Oleksyn, 2004; Shi et al., 2012).因此, 亟需选择不同典型植物, 进一步加强植物叶片化学计量空间分布格局的研究, 以验证和丰富多变环境条件下植物叶片化学计量特征变化理论假说和植物养分利用策略的科学认知. ...
Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China 5 2005
... 近年来, 围绕植物叶片化学计量特征在区域(Chen et al., 2013; Fang et al., 2019)、国家(Tang et al., 2018; Liu et al., 2019)以及全球(Tian et al., 2017, 2019)尺度上开展了大量研究, 极大地提升和丰富了人们对叶片化学计量特征及相关理论的认识和理解.然而, 目前关于植物叶片化学计量分布规律与空间变异格局仍缺乏统一性结论.例如, 植物叶片元素含量随海拔增加呈现出无显著变化, 或显著增加, 或显著降低, 甚至先增加后减少的差异化变异规律(Fisher et al., 2013; Zhao et al., 2014, 2018).此外, 前人研究也表明植物叶片N:P随纬度增加表现出无显著变化或显著降低等多种变异趋势(Reich & Oleksyn, 2004; Han et al., 2005).基于此, 相关****提出了众多理论假说来解释植物叶片化学计量特征多样化的空间变化规律, 如温度生物地球化学假说(TBH)和温度植物生理假说(TPPH).前者主要认为低温会抑制微生物活性和土壤有机质分解与矿化, 降低土壤N、P养分可利用性, 从而导致低温环境下植物叶片N、P含量较低.而后一种假说则认为低温降低了植物的生物化学反应速率, 导致植物富N的酶及富P的RNA活性下降.相应地, 低温地区植物主要通过提高叶片N、P含量以增强其新陈代谢水平(Reich & Oleksyn, 2004; Han et al., 2005).以上两种假说分别诠释了植物叶片N、P含量随温度变化而呈现出的两种截然相反的变异规律.目前上述两种假说在不同研究中得到了相应地验证.例如, 一些研究结果支持TBH (Xia et al., 2014; Zhao et al., 2014), 而另外一些研究结果则支持TPPH (Wang et al., 2015; Tian et al., 2017), 甚至部分研究结果同时支持上述两种理论假说(Reich & Oleksyn, 2004; Shi et al., 2012).因此, 亟需选择不同典型植物, 进一步加强植物叶片化学计量空间分布格局的研究, 以验证和丰富多变环境条件下植物叶片化学计量特征变化理论假说和植物养分利用策略的科学认知. ...
... ; Han et al., 2005).以上两种假说分别诠释了植物叶片N、P含量随温度变化而呈现出的两种截然相反的变异规律.目前上述两种假说在不同研究中得到了相应地验证.例如, 一些研究结果支持TBH (Xia et al., 2014; Zhao et al., 2014), 而另外一些研究结果则支持TPPH (Wang et al., 2015; Tian et al., 2017), 甚至部分研究结果同时支持上述两种理论假说(Reich & Oleksyn, 2004; Shi et al., 2012).因此, 亟需选择不同典型植物, 进一步加强植物叶片化学计量空间分布格局的研究, 以验证和丰富多变环境条件下植物叶片化学计量特征变化理论假说和植物养分利用策略的科学认知. ...
... 此外, 影响植物叶片化学计量特征和空间分布格局的因素众多, 主要包括气候因子(Reich & Oleksyn, 2004; Yuan & Chen, 2009)、土壤特性(Chen et al., 2013; Zhang et al., 2019)和生物因素(如植物种类、生活型等)(He et al., 2006, 2008; Zhao et al., 2018).研究表明, 当研究对象涉及的植物生活型较多时, 生活型往往成为植物叶片化学计量特征变异的关键驱动因素, 并很大程度上弱化了气候因子和土壤特性的影响效应(He et al., 2006, 2008; Zhao et al., 2018).此外, 不同生活型植物叶片化学计量特征及其空间分布规律也存在较大差异.例如, 草本植物叶片N、P含量通常显著高于乔木, 而落叶树种叶片元素含量显著高于常绿树种, 且两者的叶片化学计量海拔分布格局也存在较明显的差异(Han et al., 2005; Shi et al., 2012).总之, 当生活型与气候因子、土壤特性等多种因素叠加后, 势必会加剧植物叶片化学计量特征分布规律及其驱动因素认知的复杂性和不确定性.相应地, 通过降低或者排除不同生活型对植物叶片化学计量特征的潜在影响, 专一性地聚焦生活型相同或相近的植物叶片化学计量空间分布规律及其驱动因素的深入研究, 将在很大程度上丰富不同环境因子对叶片化学计量特征与空间分布变异相对贡献的认知水平. ...
... 本研究发现: (1)该区域叶片C含量平均值为504.83 g·kg-1, 高于其他****对中国西北温带森林针叶树种(Zhang et al., 2017)、中国陆地针叶树种(Tang et al., 2018)叶片C含量的研究结果.推测其原因可能与植物自身的生长特性以及该区域的气候特点有关.C是植物的结构性元素, 是植物生长发育和繁殖等生理活动的物质基础和能量来源.有研究表明, 较高的叶片C含量有助于延长植物叶片寿命并能提高其对温度、水分和养分贫瘠等胁迫环境的适应力(张明明和高瑞馨, 2012; 贺合亮等, 2017).此外, 本研究区域位于高寒的青藏高原, 温度较低且生长季较短.因此, 本研究区域针叶树种叶片C含量相对较高.(2)该区域针叶树种叶片N含量平均值为13.03 g·kg-1,叶片P含量平均值为1.30 g·kg-1.低于其他****关于贡嘎山针叶树种(Shi et al., 2012)、中国东部针叶树种(Chen et al., 2013)和长白山针叶树种(杨蕾等, 2017)等叶片N、P含量的研究结果, 而高于一些****对中国陆地针叶树种(Han et al., 2005)、中国东部针叶树种(任书杰等, 2007)、全球陆地针叶树种(Tian et al., 2017)和中国西北温带森林针叶树种(Zhang et al., 2017)等叶片N、P含量的研究结果.总体而言, 青藏高原主要针叶树种叶片N、P含量与其他研究结果相比无明显规律, 我们推测其原因可能源于不同植物叶片N、P含量变异系数较大, 比如不同水分、光照和养分等环境条件下植物在生长发育过程的N、P养分需求存在较大差异, 致使不同植物叶片N、P含量在不同生境中具有差别(田地等, 2018).此外, 该区域主要针叶树种叶片N:P均值为10.01, 满足Koerselman和Meuleman (1996)和Zhang等(2004)对植物生长N限制的评定标准(N:P < 14和N:P < 21), 进一步佐证了青藏高原东缘针叶林生长发育主要受N限制(杨欢等, 2017).另外, 该区域各针叶树种叶片C、N含量和C:N在科和属水平上差异显著, 表明物种类型对植物叶片C、N含量和C:N影响较大. ...
The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation 1 1996
... 本研究发现: (1)该区域叶片C含量平均值为504.83 g·kg-1, 高于其他****对中国西北温带森林针叶树种(Zhang et al., 2017)、中国陆地针叶树种(Tang et al., 2018)叶片C含量的研究结果.推测其原因可能与植物自身的生长特性以及该区域的气候特点有关.C是植物的结构性元素, 是植物生长发育和繁殖等生理活动的物质基础和能量来源.有研究表明, 较高的叶片C含量有助于延长植物叶片寿命并能提高其对温度、水分和养分贫瘠等胁迫环境的适应力(张明明和高瑞馨, 2012; 贺合亮等, 2017).此外, 本研究区域位于高寒的青藏高原, 温度较低且生长季较短.因此, 本研究区域针叶树种叶片C含量相对较高.(2)该区域针叶树种叶片N含量平均值为13.03 g·kg-1,叶片P含量平均值为1.30 g·kg-1.低于其他****关于贡嘎山针叶树种(Shi et al., 2012)、中国东部针叶树种(Chen et al., 2013)和长白山针叶树种(杨蕾等, 2017)等叶片N、P含量的研究结果, 而高于一些****对中国陆地针叶树种(Han et al., 2005)、中国东部针叶树种(任书杰等, 2007)、全球陆地针叶树种(Tian et al., 2017)和中国西北温带森林针叶树种(Zhang et al., 2017)等叶片N、P含量的研究结果.总体而言, 青藏高原主要针叶树种叶片N、P含量与其他研究结果相比无明显规律, 我们推测其原因可能源于不同植物叶片N、P含量变异系数较大, 比如不同水分、光照和养分等环境条件下植物在生长发育过程的N、P养分需求存在较大差异, 致使不同植物叶片N、P含量在不同生境中具有差别(田地等, 2018).此外, 该区域主要针叶树种叶片N:P均值为10.01, 满足Koerselman和Meuleman (1996)和Zhang等(2004)对植物生长N限制的评定标准(N:P < 14和N:P < 21), 进一步佐证了青藏高原东缘针叶林生长发育主要受N限制(杨欢等, 2017).另外, 该区域各针叶树种叶片C、N含量和C:N在科和属水平上差异显著, 表明物种类型对植物叶片C、N含量和C:N影响较大. ...
2010
2010
Patterns and controlling factors of plant nitrogen and phosphorus stoichiometry across China’s forests 1 2019
... 近年来, 围绕植物叶片化学计量特征在区域(Chen et al., 2013; Fang et al., 2019)、国家(Tang et al., 2018; Liu et al., 2019)以及全球(Tian et al., 2017, 2019)尺度上开展了大量研究, 极大地提升和丰富了人们对叶片化学计量特征及相关理论的认识和理解.然而, 目前关于植物叶片化学计量分布规律与空间变异格局仍缺乏统一性结论.例如, 植物叶片元素含量随海拔增加呈现出无显著变化, 或显著增加, 或显著降低, 甚至先增加后减少的差异化变异规律(Fisher et al., 2013; Zhao et al., 2014, 2018).此外, 前人研究也表明植物叶片N:P随纬度增加表现出无显著变化或显著降低等多种变异趋势(Reich & Oleksyn, 2004; Han et al., 2005).基于此, 相关****提出了众多理论假说来解释植物叶片化学计量特征多样化的空间变化规律, 如温度生物地球化学假说(TBH)和温度植物生理假说(TPPH).前者主要认为低温会抑制微生物活性和土壤有机质分解与矿化, 降低土壤N、P养分可利用性, 从而导致低温环境下植物叶片N、P含量较低.而后一种假说则认为低温降低了植物的生物化学反应速率, 导致植物富N的酶及富P的RNA活性下降.相应地, 低温地区植物主要通过提高叶片N、P含量以增强其新陈代谢水平(Reich & Oleksyn, 2004; Han et al., 2005).以上两种假说分别诠释了植物叶片N、P含量随温度变化而呈现出的两种截然相反的变异规律.目前上述两种假说在不同研究中得到了相应地验证.例如, 一些研究结果支持TBH (Xia et al., 2014; Zhao et al., 2014), 而另外一些研究结果则支持TPPH (Wang et al., 2015; Tian et al., 2017), 甚至部分研究结果同时支持上述两种理论假说(Reich & Oleksyn, 2004; Shi et al., 2012).因此, 亟需选择不同典型植物, 进一步加强植物叶片化学计量空间分布格局的研究, 以验证和丰富多变环境条件下植物叶片化学计量特征变化理论假说和植物养分利用策略的科学认知. ...
2002
2002
Variation partitioning by partial redundancy analysis (RDA) 2 1997
Global patterns of plant leaf N and P in relation to temperature and latitude 8 2004
... 近年来, 围绕植物叶片化学计量特征在区域(Chen et al., 2013; Fang et al., 2019)、国家(Tang et al., 2018; Liu et al., 2019)以及全球(Tian et al., 2017, 2019)尺度上开展了大量研究, 极大地提升和丰富了人们对叶片化学计量特征及相关理论的认识和理解.然而, 目前关于植物叶片化学计量分布规律与空间变异格局仍缺乏统一性结论.例如, 植物叶片元素含量随海拔增加呈现出无显著变化, 或显著增加, 或显著降低, 甚至先增加后减少的差异化变异规律(Fisher et al., 2013; Zhao et al., 2014, 2018).此外, 前人研究也表明植物叶片N:P随纬度增加表现出无显著变化或显著降低等多种变异趋势(Reich & Oleksyn, 2004; Han et al., 2005).基于此, 相关****提出了众多理论假说来解释植物叶片化学计量特征多样化的空间变化规律, 如温度生物地球化学假说(TBH)和温度植物生理假说(TPPH).前者主要认为低温会抑制微生物活性和土壤有机质分解与矿化, 降低土壤N、P养分可利用性, 从而导致低温环境下植物叶片N、P含量较低.而后一种假说则认为低温降低了植物的生物化学反应速率, 导致植物富N的酶及富P的RNA活性下降.相应地, 低温地区植物主要通过提高叶片N、P含量以增强其新陈代谢水平(Reich & Oleksyn, 2004; Han et al., 2005).以上两种假说分别诠释了植物叶片N、P含量随温度变化而呈现出的两种截然相反的变异规律.目前上述两种假说在不同研究中得到了相应地验证.例如, 一些研究结果支持TBH (Xia et al., 2014; Zhao et al., 2014), 而另外一些研究结果则支持TPPH (Wang et al., 2015; Tian et al., 2017), 甚至部分研究结果同时支持上述两种理论假说(Reich & Oleksyn, 2004; Shi et al., 2012).因此, 亟需选择不同典型植物, 进一步加强植物叶片化学计量空间分布格局的研究, 以验证和丰富多变环境条件下植物叶片化学计量特征变化理论假说和植物养分利用策略的科学认知. ...
... ).基于此, 相关****提出了众多理论假说来解释植物叶片化学计量特征多样化的空间变化规律, 如温度生物地球化学假说(TBH)和温度植物生理假说(TPPH).前者主要认为低温会抑制微生物活性和土壤有机质分解与矿化, 降低土壤N、P养分可利用性, 从而导致低温环境下植物叶片N、P含量较低.而后一种假说则认为低温降低了植物的生物化学反应速率, 导致植物富N的酶及富P的RNA活性下降.相应地, 低温地区植物主要通过提高叶片N、P含量以增强其新陈代谢水平(Reich & Oleksyn, 2004; Han et al., 2005).以上两种假说分别诠释了植物叶片N、P含量随温度变化而呈现出的两种截然相反的变异规律.目前上述两种假说在不同研究中得到了相应地验证.例如, 一些研究结果支持TBH (Xia et al., 2014; Zhao et al., 2014), 而另外一些研究结果则支持TPPH (Wang et al., 2015; Tian et al., 2017), 甚至部分研究结果同时支持上述两种理论假说(Reich & Oleksyn, 2004; Shi et al., 2012).因此, 亟需选择不同典型植物, 进一步加强植物叶片化学计量空间分布格局的研究, 以验证和丰富多变环境条件下植物叶片化学计量特征变化理论假说和植物养分利用策略的科学认知. ...
... ), 甚至部分研究结果同时支持上述两种理论假说(Reich & Oleksyn, 2004; Shi et al., 2012).因此, 亟需选择不同典型植物, 进一步加强植物叶片化学计量空间分布格局的研究, 以验证和丰富多变环境条件下植物叶片化学计量特征变化理论假说和植物养分利用策略的科学认知. ...
... 此外, 影响植物叶片化学计量特征和空间分布格局的因素众多, 主要包括气候因子(Reich & Oleksyn, 2004; Yuan & Chen, 2009)、土壤特性(Chen et al., 2013; Zhang et al., 2019)和生物因素(如植物种类、生活型等)(He et al., 2006, 2008; Zhao et al., 2018).研究表明, 当研究对象涉及的植物生活型较多时, 生活型往往成为植物叶片化学计量特征变异的关键驱动因素, 并很大程度上弱化了气候因子和土壤特性的影响效应(He et al., 2006, 2008; Zhao et al., 2018).此外, 不同生活型植物叶片化学计量特征及其空间分布规律也存在较大差异.例如, 草本植物叶片N、P含量通常显著高于乔木, 而落叶树种叶片元素含量显著高于常绿树种, 且两者的叶片化学计量海拔分布格局也存在较明显的差异(Han et al., 2005; Shi et al., 2012).总之, 当生活型与气候因子、土壤特性等多种因素叠加后, 势必会加剧植物叶片化学计量特征分布规律及其驱动因素认知的复杂性和不确定性.相应地, 通过降低或者排除不同生活型对植物叶片化学计量特征的潜在影响, 专一性地聚焦生活型相同或相近的植物叶片化学计量空间分布规律及其驱动因素的深入研究, 将在很大程度上丰富不同环境因子对叶片化学计量特征与空间分布变异相对贡献的认知水平. ...
... 前人众多研究已清楚地表明, 气候因子(Reich & Oleksyn 2004; Yuan & Chen, 2009)、土壤特性(Chen et al., 2013; Zhang et al., 2019)和植物生活型(He et al., 2006, 2008; Zhao et al., 2018)对植物叶片化学计量特征有重要影响.对中国草地(He et al., 2006)和西南灌丛(Zhao et al., 2018)植物叶片C、N、P化学计量主要驱动因素的研究结果显示, 生活型是驱动叶片化学计量变异的主要关键因子, 并一定程度上弱化了环境因子对植物叶片化学计量特征变异的相对贡献率.在本研究中, 我们通过选择青藏高原东缘单一生活型的针叶树种叶片为研究对象, 尽可能去除或弱化生活型的潜在影响, 专一性地辨识了单一生活型植物叶片空间变异格局以及气候因子和土壤特性对叶片化学计量的相对贡献. ...
... 本研究发现: (1)该区域叶片C含量平均值为504.83 g·kg-1, 高于其他****对中国西北温带森林针叶树种(Zhang et al., 2017)、中国陆地针叶树种(Tang et al., 2018)叶片C含量的研究结果.推测其原因可能与植物自身的生长特性以及该区域的气候特点有关.C是植物的结构性元素, 是植物生长发育和繁殖等生理活动的物质基础和能量来源.有研究表明, 较高的叶片C含量有助于延长植物叶片寿命并能提高其对温度、水分和养分贫瘠等胁迫环境的适应力(张明明和高瑞馨, 2012; 贺合亮等, 2017).此外, 本研究区域位于高寒的青藏高原, 温度较低且生长季较短.因此, 本研究区域针叶树种叶片C含量相对较高.(2)该区域针叶树种叶片N含量平均值为13.03 g·kg-1,叶片P含量平均值为1.30 g·kg-1.低于其他****关于贡嘎山针叶树种(Shi et al., 2012)、中国东部针叶树种(Chen et al., 2013)和长白山针叶树种(杨蕾等, 2017)等叶片N、P含量的研究结果, 而高于一些****对中国陆地针叶树种(Han et al., 2005)、中国东部针叶树种(任书杰等, 2007)、全球陆地针叶树种(Tian et al., 2017)和中国西北温带森林针叶树种(Zhang et al., 2017)等叶片N、P含量的研究结果.总体而言, 青藏高原主要针叶树种叶片N、P含量与其他研究结果相比无明显规律, 我们推测其原因可能源于不同植物叶片N、P含量变异系数较大, 比如不同水分、光照和养分等环境条件下植物在生长发育过程的N、P养分需求存在较大差异, 致使不同植物叶片N、P含量在不同生境中具有差别(田地等, 2018).此外, 该区域主要针叶树种叶片N:P均值为10.01, 满足Koerselman和Meuleman (1996)和Zhang等(2004)对植物生长N限制的评定标准(N:P < 14和N:P < 21), 进一步佐证了青藏高原东缘针叶林生长发育主要受N限制(杨欢等, 2017).另外, 该区域各针叶树种叶片C、N含量和C:N在科和属水平上差异显著, 表明物种类型对植物叶片C、N含量和C:N影响较大. ...
中国东部南北样带654种植物叶片氮和磷的化学计量学特征研究 1 2007
... 本研究发现: (1)该区域叶片C含量平均值为504.83 g·kg-1, 高于其他****对中国西北温带森林针叶树种(Zhang et al., 2017)、中国陆地针叶树种(Tang et al., 2018)叶片C含量的研究结果.推测其原因可能与植物自身的生长特性以及该区域的气候特点有关.C是植物的结构性元素, 是植物生长发育和繁殖等生理活动的物质基础和能量来源.有研究表明, 较高的叶片C含量有助于延长植物叶片寿命并能提高其对温度、水分和养分贫瘠等胁迫环境的适应力(张明明和高瑞馨, 2012; 贺合亮等, 2017).此外, 本研究区域位于高寒的青藏高原, 温度较低且生长季较短.因此, 本研究区域针叶树种叶片C含量相对较高.(2)该区域针叶树种叶片N含量平均值为13.03 g·kg-1,叶片P含量平均值为1.30 g·kg-1.低于其他****关于贡嘎山针叶树种(Shi et al., 2012)、中国东部针叶树种(Chen et al., 2013)和长白山针叶树种(杨蕾等, 2017)等叶片N、P含量的研究结果, 而高于一些****对中国陆地针叶树种(Han et al., 2005)、中国东部针叶树种(任书杰等, 2007)、全球陆地针叶树种(Tian et al., 2017)和中国西北温带森林针叶树种(Zhang et al., 2017)等叶片N、P含量的研究结果.总体而言, 青藏高原主要针叶树种叶片N、P含量与其他研究结果相比无明显规律, 我们推测其原因可能源于不同植物叶片N、P含量变异系数较大, 比如不同水分、光照和养分等环境条件下植物在生长发育过程的N、P养分需求存在较大差异, 致使不同植物叶片N、P含量在不同生境中具有差别(田地等, 2018).此外, 该区域主要针叶树种叶片N:P均值为10.01, 满足Koerselman和Meuleman (1996)和Zhang等(2004)对植物生长N限制的评定标准(N:P < 14和N:P < 21), 进一步佐证了青藏高原东缘针叶林生长发育主要受N限制(杨欢等, 2017).另外, 该区域各针叶树种叶片C、N含量和C:N在科和属水平上差异显著, 表明物种类型对植物叶片C、N含量和C:N影响较大. ...
Tree growth changes with climate and forest type are associated with relative allocation of nutrients, especially phosphorus, to leaves and wood 1 2013
Altitudinal variation in leaf nitrogen concentration on the eastern slope of Mount Gongga on the Tibetan Plateau, China 3 2012
... 近年来, 围绕植物叶片化学计量特征在区域(Chen et al., 2013; Fang et al., 2019)、国家(Tang et al., 2018; Liu et al., 2019)以及全球(Tian et al., 2017, 2019)尺度上开展了大量研究, 极大地提升和丰富了人们对叶片化学计量特征及相关理论的认识和理解.然而, 目前关于植物叶片化学计量分布规律与空间变异格局仍缺乏统一性结论.例如, 植物叶片元素含量随海拔增加呈现出无显著变化, 或显著增加, 或显著降低, 甚至先增加后减少的差异化变异规律(Fisher et al., 2013; Zhao et al., 2014, 2018).此外, 前人研究也表明植物叶片N:P随纬度增加表现出无显著变化或显著降低等多种变异趋势(Reich & Oleksyn, 2004; Han et al., 2005).基于此, 相关****提出了众多理论假说来解释植物叶片化学计量特征多样化的空间变化规律, 如温度生物地球化学假说(TBH)和温度植物生理假说(TPPH).前者主要认为低温会抑制微生物活性和土壤有机质分解与矿化, 降低土壤N、P养分可利用性, 从而导致低温环境下植物叶片N、P含量较低.而后一种假说则认为低温降低了植物的生物化学反应速率, 导致植物富N的酶及富P的RNA活性下降.相应地, 低温地区植物主要通过提高叶片N、P含量以增强其新陈代谢水平(Reich & Oleksyn, 2004; Han et al., 2005).以上两种假说分别诠释了植物叶片N、P含量随温度变化而呈现出的两种截然相反的变异规律.目前上述两种假说在不同研究中得到了相应地验证.例如, 一些研究结果支持TBH (Xia et al., 2014; Zhao et al., 2014), 而另外一些研究结果则支持TPPH (Wang et al., 2015; Tian et al., 2017), 甚至部分研究结果同时支持上述两种理论假说(Reich & Oleksyn, 2004; Shi et al., 2012).因此, 亟需选择不同典型植物, 进一步加强植物叶片化学计量空间分布格局的研究, 以验证和丰富多变环境条件下植物叶片化学计量特征变化理论假说和植物养分利用策略的科学认知. ...
... 此外, 影响植物叶片化学计量特征和空间分布格局的因素众多, 主要包括气候因子(Reich & Oleksyn, 2004; Yuan & Chen, 2009)、土壤特性(Chen et al., 2013; Zhang et al., 2019)和生物因素(如植物种类、生活型等)(He et al., 2006, 2008; Zhao et al., 2018).研究表明, 当研究对象涉及的植物生活型较多时, 生活型往往成为植物叶片化学计量特征变异的关键驱动因素, 并很大程度上弱化了气候因子和土壤特性的影响效应(He et al., 2006, 2008; Zhao et al., 2018).此外, 不同生活型植物叶片化学计量特征及其空间分布规律也存在较大差异.例如, 草本植物叶片N、P含量通常显著高于乔木, 而落叶树种叶片元素含量显著高于常绿树种, 且两者的叶片化学计量海拔分布格局也存在较明显的差异(Han et al., 2005; Shi et al., 2012).总之, 当生活型与气候因子、土壤特性等多种因素叠加后, 势必会加剧植物叶片化学计量特征分布规律及其驱动因素认知的复杂性和不确定性.相应地, 通过降低或者排除不同生活型对植物叶片化学计量特征的潜在影响, 专一性地聚焦生活型相同或相近的植物叶片化学计量空间分布规律及其驱动因素的深入研究, 将在很大程度上丰富不同环境因子对叶片化学计量特征与空间分布变异相对贡献的认知水平. ...
... 本研究发现: (1)该区域叶片C含量平均值为504.83 g·kg-1, 高于其他****对中国西北温带森林针叶树种(Zhang et al., 2017)、中国陆地针叶树种(Tang et al., 2018)叶片C含量的研究结果.推测其原因可能与植物自身的生长特性以及该区域的气候特点有关.C是植物的结构性元素, 是植物生长发育和繁殖等生理活动的物质基础和能量来源.有研究表明, 较高的叶片C含量有助于延长植物叶片寿命并能提高其对温度、水分和养分贫瘠等胁迫环境的适应力(张明明和高瑞馨, 2012; 贺合亮等, 2017).此外, 本研究区域位于高寒的青藏高原, 温度较低且生长季较短.因此, 本研究区域针叶树种叶片C含量相对较高.(2)该区域针叶树种叶片N含量平均值为13.03 g·kg-1,叶片P含量平均值为1.30 g·kg-1.低于其他****关于贡嘎山针叶树种(Shi et al., 2012)、中国东部针叶树种(Chen et al., 2013)和长白山针叶树种(杨蕾等, 2017)等叶片N、P含量的研究结果, 而高于一些****对中国陆地针叶树种(Han et al., 2005)、中国东部针叶树种(任书杰等, 2007)、全球陆地针叶树种(Tian et al., 2017)和中国西北温带森林针叶树种(Zhang et al., 2017)等叶片N、P含量的研究结果.总体而言, 青藏高原主要针叶树种叶片N、P含量与其他研究结果相比无明显规律, 我们推测其原因可能源于不同植物叶片N、P含量变异系数较大, 比如不同水分、光照和养分等环境条件下植物在生长发育过程的N、P养分需求存在较大差异, 致使不同植物叶片N、P含量在不同生境中具有差别(田地等, 2018).此外, 该区域主要针叶树种叶片N:P均值为10.01, 满足Koerselman和Meuleman (1996)和Zhang等(2004)对植物生长N限制的评定标准(N:P < 14和N:P < 21), 进一步佐证了青藏高原东缘针叶林生长发育主要受N限制(杨欢等, 2017).另外, 该区域各针叶树种叶片C、N含量和C:N在科和属水平上差异显著, 表明物种类型对植物叶片C、N含量和C:N影响较大. ...
Stoichiometric flexibility as a regulator of carbon and nutrient cycling in terrestrial ecosystems under change 1 2012
Patterns of plant carbon, nitrogen, and phosphorus concentration in relation to productivity in China’s terrestrial ecosystems 2 2018
... 近年来, 围绕植物叶片化学计量特征在区域(Chen et al., 2013; Fang et al., 2019)、国家(Tang et al., 2018; Liu et al., 2019)以及全球(Tian et al., 2017, 2019)尺度上开展了大量研究, 极大地提升和丰富了人们对叶片化学计量特征及相关理论的认识和理解.然而, 目前关于植物叶片化学计量分布规律与空间变异格局仍缺乏统一性结论.例如, 植物叶片元素含量随海拔增加呈现出无显著变化, 或显著增加, 或显著降低, 甚至先增加后减少的差异化变异规律(Fisher et al., 2013; Zhao et al., 2014, 2018).此外, 前人研究也表明植物叶片N:P随纬度增加表现出无显著变化或显著降低等多种变异趋势(Reich & Oleksyn, 2004; Han et al., 2005).基于此, 相关****提出了众多理论假说来解释植物叶片化学计量特征多样化的空间变化规律, 如温度生物地球化学假说(TBH)和温度植物生理假说(TPPH).前者主要认为低温会抑制微生物活性和土壤有机质分解与矿化, 降低土壤N、P养分可利用性, 从而导致低温环境下植物叶片N、P含量较低.而后一种假说则认为低温降低了植物的生物化学反应速率, 导致植物富N的酶及富P的RNA活性下降.相应地, 低温地区植物主要通过提高叶片N、P含量以增强其新陈代谢水平(Reich & Oleksyn, 2004; Han et al., 2005).以上两种假说分别诠释了植物叶片N、P含量随温度变化而呈现出的两种截然相反的变异规律.目前上述两种假说在不同研究中得到了相应地验证.例如, 一些研究结果支持TBH (Xia et al., 2014; Zhao et al., 2014), 而另外一些研究结果则支持TPPH (Wang et al., 2015; Tian et al., 2017), 甚至部分研究结果同时支持上述两种理论假说(Reich & Oleksyn, 2004; Shi et al., 2012).因此, 亟需选择不同典型植物, 进一步加强植物叶片化学计量空间分布格局的研究, 以验证和丰富多变环境条件下植物叶片化学计量特征变化理论假说和植物养分利用策略的科学认知. ...
... 本研究发现: (1)该区域叶片C含量平均值为504.83 g·kg-1, 高于其他****对中国西北温带森林针叶树种(Zhang et al., 2017)、中国陆地针叶树种(Tang et al., 2018)叶片C含量的研究结果.推测其原因可能与植物自身的生长特性以及该区域的气候特点有关.C是植物的结构性元素, 是植物生长发育和繁殖等生理活动的物质基础和能量来源.有研究表明, 较高的叶片C含量有助于延长植物叶片寿命并能提高其对温度、水分和养分贫瘠等胁迫环境的适应力(张明明和高瑞馨, 2012; 贺合亮等, 2017).此外, 本研究区域位于高寒的青藏高原, 温度较低且生长季较短.因此, 本研究区域针叶树种叶片C含量相对较高.(2)该区域针叶树种叶片N含量平均值为13.03 g·kg-1,叶片P含量平均值为1.30 g·kg-1.低于其他****关于贡嘎山针叶树种(Shi et al., 2012)、中国东部针叶树种(Chen et al., 2013)和长白山针叶树种(杨蕾等, 2017)等叶片N、P含量的研究结果, 而高于一些****对中国陆地针叶树种(Han et al., 2005)、中国东部针叶树种(任书杰等, 2007)、全球陆地针叶树种(Tian et al., 2017)和中国西北温带森林针叶树种(Zhang et al., 2017)等叶片N、P含量的研究结果.总体而言, 青藏高原主要针叶树种叶片N、P含量与其他研究结果相比无明显规律, 我们推测其原因可能源于不同植物叶片N、P含量变异系数较大, 比如不同水分、光照和养分等环境条件下植物在生长发育过程的N、P养分需求存在较大差异, 致使不同植物叶片N、P含量在不同生境中具有差别(田地等, 2018).此外, 该区域主要针叶树种叶片N:P均值为10.01, 满足Koerselman和Meuleman (1996)和Zhang等(2004)对植物生长N限制的评定标准(N:P < 14和N:P < 21), 进一步佐证了青藏高原东缘针叶林生长发育主要受N限制(杨欢等, 2017).另外, 该区域各针叶树种叶片C、N含量和C:N在科和属水平上差异显著, 表明物种类型对植物叶片C、N含量和C:N影响较大. ...
... 本研究发现: (1)该区域叶片C含量平均值为504.83 g·kg-1, 高于其他****对中国西北温带森林针叶树种(Zhang et al., 2017)、中国陆地针叶树种(Tang et al., 2018)叶片C含量的研究结果.推测其原因可能与植物自身的生长特性以及该区域的气候特点有关.C是植物的结构性元素, 是植物生长发育和繁殖等生理活动的物质基础和能量来源.有研究表明, 较高的叶片C含量有助于延长植物叶片寿命并能提高其对温度、水分和养分贫瘠等胁迫环境的适应力(张明明和高瑞馨, 2012; 贺合亮等, 2017).此外, 本研究区域位于高寒的青藏高原, 温度较低且生长季较短.因此, 本研究区域针叶树种叶片C含量相对较高.(2)该区域针叶树种叶片N含量平均值为13.03 g·kg-1,叶片P含量平均值为1.30 g·kg-1.低于其他****关于贡嘎山针叶树种(Shi et al., 2012)、中国东部针叶树种(Chen et al., 2013)和长白山针叶树种(杨蕾等, 2017)等叶片N、P含量的研究结果, 而高于一些****对中国陆地针叶树种(Han et al., 2005)、中国东部针叶树种(任书杰等, 2007)、全球陆地针叶树种(Tian et al., 2017)和中国西北温带森林针叶树种(Zhang et al., 2017)等叶片N、P含量的研究结果.总体而言, 青藏高原主要针叶树种叶片N、P含量与其他研究结果相比无明显规律, 我们推测其原因可能源于不同植物叶片N、P含量变异系数较大, 比如不同水分、光照和养分等环境条件下植物在生长发育过程的N、P养分需求存在较大差异, 致使不同植物叶片N、P含量在不同生境中具有差别(田地等, 2018).此外, 该区域主要针叶树种叶片N:P均值为10.01, 满足Koerselman和Meuleman (1996)和Zhang等(2004)对植物生长N限制的评定标准(N:P < 14和N:P < 21), 进一步佐证了青藏高原东缘针叶林生长发育主要受N限制(杨欢等, 2017).另外, 该区域各针叶树种叶片C、N含量和C:N在科和属水平上差异显著, 表明物种类型对植物叶片C、N含量和C:N影响较大. ...
Family-level leaf nitrogen and phosphorus stoichiometry of global terrestrial plants 1 2019
... 近年来, 围绕植物叶片化学计量特征在区域(Chen et al., 2013; Fang et al., 2019)、国家(Tang et al., 2018; Liu et al., 2019)以及全球(Tian et al., 2017, 2019)尺度上开展了大量研究, 极大地提升和丰富了人们对叶片化学计量特征及相关理论的认识和理解.然而, 目前关于植物叶片化学计量分布规律与空间变异格局仍缺乏统一性结论.例如, 植物叶片元素含量随海拔增加呈现出无显著变化, 或显著增加, 或显著降低, 甚至先增加后减少的差异化变异规律(Fisher et al., 2013; Zhao et al., 2014, 2018).此外, 前人研究也表明植物叶片N:P随纬度增加表现出无显著变化或显著降低等多种变异趋势(Reich & Oleksyn, 2004; Han et al., 2005).基于此, 相关****提出了众多理论假说来解释植物叶片化学计量特征多样化的空间变化规律, 如温度生物地球化学假说(TBH)和温度植物生理假说(TPPH).前者主要认为低温会抑制微生物活性和土壤有机质分解与矿化, 降低土壤N、P养分可利用性, 从而导致低温环境下植物叶片N、P含量较低.而后一种假说则认为低温降低了植物的生物化学反应速率, 导致植物富N的酶及富P的RNA活性下降.相应地, 低温地区植物主要通过提高叶片N、P含量以增强其新陈代谢水平(Reich & Oleksyn, 2004; Han et al., 2005).以上两种假说分别诠释了植物叶片N、P含量随温度变化而呈现出的两种截然相反的变异规律.目前上述两种假说在不同研究中得到了相应地验证.例如, 一些研究结果支持TBH (Xia et al., 2014; Zhao et al., 2014), 而另外一些研究结果则支持TPPH (Wang et al., 2015; Tian et al., 2017), 甚至部分研究结果同时支持上述两种理论假说(Reich & Oleksyn, 2004; Shi et al., 2012).因此, 亟需选择不同典型植物, 进一步加强植物叶片化学计量空间分布格局的研究, 以验证和丰富多变环境条件下植物叶片化学计量特征变化理论假说和植物养分利用策略的科学认知. ...
Global leaf nitrogen and phosphorus stoichiometry and their scaling exponent 3 2017
... 近年来, 围绕植物叶片化学计量特征在区域(Chen et al., 2013; Fang et al., 2019)、国家(Tang et al., 2018; Liu et al., 2019)以及全球(Tian et al., 2017, 2019)尺度上开展了大量研究, 极大地提升和丰富了人们对叶片化学计量特征及相关理论的认识和理解.然而, 目前关于植物叶片化学计量分布规律与空间变异格局仍缺乏统一性结论.例如, 植物叶片元素含量随海拔增加呈现出无显著变化, 或显著增加, 或显著降低, 甚至先增加后减少的差异化变异规律(Fisher et al., 2013; Zhao et al., 2014, 2018).此外, 前人研究也表明植物叶片N:P随纬度增加表现出无显著变化或显著降低等多种变异趋势(Reich & Oleksyn, 2004; Han et al., 2005).基于此, 相关****提出了众多理论假说来解释植物叶片化学计量特征多样化的空间变化规律, 如温度生物地球化学假说(TBH)和温度植物生理假说(TPPH).前者主要认为低温会抑制微生物活性和土壤有机质分解与矿化, 降低土壤N、P养分可利用性, 从而导致低温环境下植物叶片N、P含量较低.而后一种假说则认为低温降低了植物的生物化学反应速率, 导致植物富N的酶及富P的RNA活性下降.相应地, 低温地区植物主要通过提高叶片N、P含量以增强其新陈代谢水平(Reich & Oleksyn, 2004; Han et al., 2005).以上两种假说分别诠释了植物叶片N、P含量随温度变化而呈现出的两种截然相反的变异规律.目前上述两种假说在不同研究中得到了相应地验证.例如, 一些研究结果支持TBH (Xia et al., 2014; Zhao et al., 2014), 而另外一些研究结果则支持TPPH (Wang et al., 2015; Tian et al., 2017), 甚至部分研究结果同时支持上述两种理论假说(Reich & Oleksyn, 2004; Shi et al., 2012).因此, 亟需选择不同典型植物, 进一步加强植物叶片化学计量空间分布格局的研究, 以验证和丰富多变环境条件下植物叶片化学计量特征变化理论假说和植物养分利用策略的科学认知. ...
... ; Tian et al., 2017), 甚至部分研究结果同时支持上述两种理论假说(Reich & Oleksyn, 2004; Shi et al., 2012).因此, 亟需选择不同典型植物, 进一步加强植物叶片化学计量空间分布格局的研究, 以验证和丰富多变环境条件下植物叶片化学计量特征变化理论假说和植物养分利用策略的科学认知. ...
... 本研究发现: (1)该区域叶片C含量平均值为504.83 g·kg-1, 高于其他****对中国西北温带森林针叶树种(Zhang et al., 2017)、中国陆地针叶树种(Tang et al., 2018)叶片C含量的研究结果.推测其原因可能与植物自身的生长特性以及该区域的气候特点有关.C是植物的结构性元素, 是植物生长发育和繁殖等生理活动的物质基础和能量来源.有研究表明, 较高的叶片C含量有助于延长植物叶片寿命并能提高其对温度、水分和养分贫瘠等胁迫环境的适应力(张明明和高瑞馨, 2012; 贺合亮等, 2017).此外, 本研究区域位于高寒的青藏高原, 温度较低且生长季较短.因此, 本研究区域针叶树种叶片C含量相对较高.(2)该区域针叶树种叶片N含量平均值为13.03 g·kg-1,叶片P含量平均值为1.30 g·kg-1.低于其他****关于贡嘎山针叶树种(Shi et al., 2012)、中国东部针叶树种(Chen et al., 2013)和长白山针叶树种(杨蕾等, 2017)等叶片N、P含量的研究结果, 而高于一些****对中国陆地针叶树种(Han et al., 2005)、中国东部针叶树种(任书杰等, 2007)、全球陆地针叶树种(Tian et al., 2017)和中国西北温带森林针叶树种(Zhang et al., 2017)等叶片N、P含量的研究结果.总体而言, 青藏高原主要针叶树种叶片N、P含量与其他研究结果相比无明显规律, 我们推测其原因可能源于不同植物叶片N、P含量变异系数较大, 比如不同水分、光照和养分等环境条件下植物在生长发育过程的N、P养分需求存在较大差异, 致使不同植物叶片N、P含量在不同生境中具有差别(田地等, 2018).此外, 该区域主要针叶树种叶片N:P均值为10.01, 满足Koerselman和Meuleman (1996)和Zhang等(2004)对植物生长N限制的评定标准(N:P < 14和N:P < 21), 进一步佐证了青藏高原东缘针叶林生长发育主要受N限制(杨欢等, 2017).另外, 该区域各针叶树种叶片C、N含量和C:N在科和属水平上差异显著, 表明物种类型对植物叶片C、N含量和C:N影响较大. ...
Plant uptake of major and minor mineral elements as influenced by soil acidity and liming 1 2001
Low-temperature induced leaf elements accumulation in aquatic macrophytes across Tibetan Plateau 1 2015
... 近年来, 围绕植物叶片化学计量特征在区域(Chen et al., 2013; Fang et al., 2019)、国家(Tang et al., 2018; Liu et al., 2019)以及全球(Tian et al., 2017, 2019)尺度上开展了大量研究, 极大地提升和丰富了人们对叶片化学计量特征及相关理论的认识和理解.然而, 目前关于植物叶片化学计量分布规律与空间变异格局仍缺乏统一性结论.例如, 植物叶片元素含量随海拔增加呈现出无显著变化, 或显著增加, 或显著降低, 甚至先增加后减少的差异化变异规律(Fisher et al., 2013; Zhao et al., 2014, 2018).此外, 前人研究也表明植物叶片N:P随纬度增加表现出无显著变化或显著降低等多种变异趋势(Reich & Oleksyn, 2004; Han et al., 2005).基于此, 相关****提出了众多理论假说来解释植物叶片化学计量特征多样化的空间变化规律, 如温度生物地球化学假说(TBH)和温度植物生理假说(TPPH).前者主要认为低温会抑制微生物活性和土壤有机质分解与矿化, 降低土壤N、P养分可利用性, 从而导致低温环境下植物叶片N、P含量较低.而后一种假说则认为低温降低了植物的生物化学反应速率, 导致植物富N的酶及富P的RNA活性下降.相应地, 低温地区植物主要通过提高叶片N、P含量以增强其新陈代谢水平(Reich & Oleksyn, 2004; Han et al., 2005).以上两种假说分别诠释了植物叶片N、P含量随温度变化而呈现出的两种截然相反的变异规律.目前上述两种假说在不同研究中得到了相应地验证.例如, 一些研究结果支持TBH (Xia et al., 2014; Zhao et al., 2014), 而另外一些研究结果则支持TPPH (Wang et al., 2015; Tian et al., 2017), 甚至部分研究结果同时支持上述两种理论假说(Reich & Oleksyn, 2004; Shi et al., 2012).因此, 亟需选择不同典型植物, 进一步加强植物叶片化学计量空间分布格局的研究, 以验证和丰富多变环境条件下植物叶片化学计量特征变化理论假说和植物养分利用策略的科学认知. ...
Stoichiometry patterns of leaf carbon, nitrogen and phosphorous in aquatic macrophytes in eastern China 1 2014
... 近年来, 围绕植物叶片化学计量特征在区域(Chen et al., 2013; Fang et al., 2019)、国家(Tang et al., 2018; Liu et al., 2019)以及全球(Tian et al., 2017, 2019)尺度上开展了大量研究, 极大地提升和丰富了人们对叶片化学计量特征及相关理论的认识和理解.然而, 目前关于植物叶片化学计量分布规律与空间变异格局仍缺乏统一性结论.例如, 植物叶片元素含量随海拔增加呈现出无显著变化, 或显著增加, 或显著降低, 甚至先增加后减少的差异化变异规律(Fisher et al., 2013; Zhao et al., 2014, 2018).此外, 前人研究也表明植物叶片N:P随纬度增加表现出无显著变化或显著降低等多种变异趋势(Reich & Oleksyn, 2004; Han et al., 2005).基于此, 相关****提出了众多理论假说来解释植物叶片化学计量特征多样化的空间变化规律, 如温度生物地球化学假说(TBH)和温度植物生理假说(TPPH).前者主要认为低温会抑制微生物活性和土壤有机质分解与矿化, 降低土壤N、P养分可利用性, 从而导致低温环境下植物叶片N、P含量较低.而后一种假说则认为低温降低了植物的生物化学反应速率, 导致植物富N的酶及富P的RNA活性下降.相应地, 低温地区植物主要通过提高叶片N、P含量以增强其新陈代谢水平(Reich & Oleksyn, 2004; Han et al., 2005).以上两种假说分别诠释了植物叶片N、P含量随温度变化而呈现出的两种截然相反的变异规律.目前上述两种假说在不同研究中得到了相应地验证.例如, 一些研究结果支持TBH (Xia et al., 2014; Zhao et al., 2014), 而另外一些研究结果则支持TPPH (Wang et al., 2015; Tian et al., 2017), 甚至部分研究结果同时支持上述两种理论假说(Reich & Oleksyn, 2004; Shi et al., 2012).因此, 亟需选择不同典型植物, 进一步加强植物叶片化学计量空间分布格局的研究, 以验证和丰富多变环境条件下植物叶片化学计量特征变化理论假说和植物养分利用策略的科学认知. ...
Differential responses of N:P stoichiometry of Leymus chinensis and Carex korshinskyi to N additions in a steppe ecosystem in Nei Mongol. 1 2004
... 本研究发现: (1)该区域叶片C含量平均值为504.83 g·kg-1, 高于其他****对中国西北温带森林针叶树种(Zhang et al., 2017)、中国陆地针叶树种(Tang et al., 2018)叶片C含量的研究结果.推测其原因可能与植物自身的生长特性以及该区域的气候特点有关.C是植物的结构性元素, 是植物生长发育和繁殖等生理活动的物质基础和能量来源.有研究表明, 较高的叶片C含量有助于延长植物叶片寿命并能提高其对温度、水分和养分贫瘠等胁迫环境的适应力(张明明和高瑞馨, 2012; 贺合亮等, 2017).此外, 本研究区域位于高寒的青藏高原, 温度较低且生长季较短.因此, 本研究区域针叶树种叶片C含量相对较高.(2)该区域针叶树种叶片N含量平均值为13.03 g·kg-1,叶片P含量平均值为1.30 g·kg-1.低于其他****关于贡嘎山针叶树种(Shi et al., 2012)、中国东部针叶树种(Chen et al., 2013)和长白山针叶树种(杨蕾等, 2017)等叶片N、P含量的研究结果, 而高于一些****对中国陆地针叶树种(Han et al., 2005)、中国东部针叶树种(任书杰等, 2007)、全球陆地针叶树种(Tian et al., 2017)和中国西北温带森林针叶树种(Zhang et al., 2017)等叶片N、P含量的研究结果.总体而言, 青藏高原主要针叶树种叶片N、P含量与其他研究结果相比无明显规律, 我们推测其原因可能源于不同植物叶片N、P含量变异系数较大, 比如不同水分、光照和养分等环境条件下植物在生长发育过程的N、P养分需求存在较大差异, 致使不同植物叶片N、P含量在不同生境中具有差别(田地等, 2018).此外, 该区域主要针叶树种叶片N:P均值为10.01, 满足Koerselman和Meuleman (1996)和Zhang等(2004)对植物生长N限制的评定标准(N:P < 14和N:P < 21), 进一步佐证了青藏高原东缘针叶林生长发育主要受N限制(杨欢等, 2017).另外, 该区域各针叶树种叶片C、N含量和C:N在科和属水平上差异显著, 表明物种类型对植物叶片C、N含量和C:N影响较大. ...
针叶植物叶片比较解剖及生态解剖研究综述 1 2012
... 本研究发现: (1)该区域叶片C含量平均值为504.83 g·kg-1, 高于其他****对中国西北温带森林针叶树种(Zhang et al., 2017)、中国陆地针叶树种(Tang et al., 2018)叶片C含量的研究结果.推测其原因可能与植物自身的生长特性以及该区域的气候特点有关.C是植物的结构性元素, 是植物生长发育和繁殖等生理活动的物质基础和能量来源.有研究表明, 较高的叶片C含量有助于延长植物叶片寿命并能提高其对温度、水分和养分贫瘠等胁迫环境的适应力(张明明和高瑞馨, 2012; 贺合亮等, 2017).此外, 本研究区域位于高寒的青藏高原, 温度较低且生长季较短.因此, 本研究区域针叶树种叶片C含量相对较高.(2)该区域针叶树种叶片N含量平均值为13.03 g·kg-1,叶片P含量平均值为1.30 g·kg-1.低于其他****关于贡嘎山针叶树种(Shi et al., 2012)、中国东部针叶树种(Chen et al., 2013)和长白山针叶树种(杨蕾等, 2017)等叶片N、P含量的研究结果, 而高于一些****对中国陆地针叶树种(Han et al., 2005)、中国东部针叶树种(任书杰等, 2007)、全球陆地针叶树种(Tian et al., 2017)和中国西北温带森林针叶树种(Zhang et al., 2017)等叶片N、P含量的研究结果.总体而言, 青藏高原主要针叶树种叶片N、P含量与其他研究结果相比无明显规律, 我们推测其原因可能源于不同植物叶片N、P含量变异系数较大, 比如不同水分、光照和养分等环境条件下植物在生长发育过程的N、P养分需求存在较大差异, 致使不同植物叶片N、P含量在不同生境中具有差别(田地等, 2018).此外, 该区域主要针叶树种叶片N:P均值为10.01, 满足Koerselman和Meuleman (1996)和Zhang等(2004)对植物生长N限制的评定标准(N:P < 14和N:P < 21), 进一步佐证了青藏高原东缘针叶林生长发育主要受N限制(杨欢等, 2017).另外, 该区域各针叶树种叶片C、N含量和C:N在科和属水平上差异显著, 表明物种类型对植物叶片C、N含量和C:N影响较大. ...
针叶植物叶片比较解剖及生态解剖研究综述 1 2012
... 本研究发现: (1)该区域叶片C含量平均值为504.83 g·kg-1, 高于其他****对中国西北温带森林针叶树种(Zhang et al., 2017)、中国陆地针叶树种(Tang et al., 2018)叶片C含量的研究结果.推测其原因可能与植物自身的生长特性以及该区域的气候特点有关.C是植物的结构性元素, 是植物生长发育和繁殖等生理活动的物质基础和能量来源.有研究表明, 较高的叶片C含量有助于延长植物叶片寿命并能提高其对温度、水分和养分贫瘠等胁迫环境的适应力(张明明和高瑞馨, 2012; 贺合亮等, 2017).此外, 本研究区域位于高寒的青藏高原, 温度较低且生长季较短.因此, 本研究区域针叶树种叶片C含量相对较高.(2)该区域针叶树种叶片N含量平均值为13.03 g·kg-1,叶片P含量平均值为1.30 g·kg-1.低于其他****关于贡嘎山针叶树种(Shi et al., 2012)、中国东部针叶树种(Chen et al., 2013)和长白山针叶树种(杨蕾等, 2017)等叶片N、P含量的研究结果, 而高于一些****对中国陆地针叶树种(Han et al., 2005)、中国东部针叶树种(任书杰等, 2007)、全球陆地针叶树种(Tian et al., 2017)和中国西北温带森林针叶树种(Zhang et al., 2017)等叶片N、P含量的研究结果.总体而言, 青藏高原主要针叶树种叶片N、P含量与其他研究结果相比无明显规律, 我们推测其原因可能源于不同植物叶片N、P含量变异系数较大, 比如不同水分、光照和养分等环境条件下植物在生长发育过程的N、P养分需求存在较大差异, 致使不同植物叶片N、P含量在不同生境中具有差别(田地等, 2018).此外, 该区域主要针叶树种叶片N:P均值为10.01, 满足Koerselman和Meuleman (1996)和Zhang等(2004)对植物生长N限制的评定标准(N:P < 14和N:P < 21), 进一步佐证了青藏高原东缘针叶林生长发育主要受N限制(杨欢等, 2017).另外, 该区域各针叶树种叶片C、N含量和C:N在科和属水平上差异显著, 表明物种类型对植物叶片C、N含量和C:N影响较大. ...
Linkages of C:N:P stoichiometry between soil and leaf and their response to climatic factors along altitudinal gradients 2 2019
... 此外, 影响植物叶片化学计量特征和空间分布格局的因素众多, 主要包括气候因子(Reich & Oleksyn, 2004; Yuan & Chen, 2009)、土壤特性(Chen et al., 2013; Zhang et al., 2019)和生物因素(如植物种类、生活型等)(He et al., 2006, 2008; Zhao et al., 2018).研究表明, 当研究对象涉及的植物生活型较多时, 生活型往往成为植物叶片化学计量特征变异的关键驱动因素, 并很大程度上弱化了气候因子和土壤特性的影响效应(He et al., 2006, 2008; Zhao et al., 2018).此外, 不同生活型植物叶片化学计量特征及其空间分布规律也存在较大差异.例如, 草本植物叶片N、P含量通常显著高于乔木, 而落叶树种叶片元素含量显著高于常绿树种, 且两者的叶片化学计量海拔分布格局也存在较明显的差异(Han et al., 2005; Shi et al., 2012).总之, 当生活型与气候因子、土壤特性等多种因素叠加后, 势必会加剧植物叶片化学计量特征分布规律及其驱动因素认知的复杂性和不确定性.相应地, 通过降低或者排除不同生活型对植物叶片化学计量特征的潜在影响, 专一性地聚焦生活型相同或相近的植物叶片化学计量空间分布规律及其驱动因素的深入研究, 将在很大程度上丰富不同环境因子对叶片化学计量特征与空间分布变异相对贡献的认知水平. ...
... 前人众多研究已清楚地表明, 气候因子(Reich & Oleksyn 2004; Yuan & Chen, 2009)、土壤特性(Chen et al., 2013; Zhang et al., 2019)和植物生活型(He et al., 2006, 2008; Zhao et al., 2018)对植物叶片化学计量特征有重要影响.对中国草地(He et al., 2006)和西南灌丛(Zhao et al., 2018)植物叶片C、N、P化学计量主要驱动因素的研究结果显示, 生活型是驱动叶片化学计量变异的主要关键因子, 并一定程度上弱化了环境因子对植物叶片化学计量特征变异的相对贡献率.在本研究中, 我们通过选择青藏高原东缘单一生活型的针叶树种叶片为研究对象, 尽可能去除或弱化生活型的潜在影响, 专一性地辨识了单一生活型植物叶片空间变异格局以及气候因子和土壤特性对叶片化学计量的相对贡献. ...
The altitudinal patterns of leaf C:N:P stoichiometry are regulated by plant growth form, climate and soil on Changbai Mountain, China 3 2014
... 近年来, 围绕植物叶片化学计量特征在区域(Chen et al., 2013; Fang et al., 2019)、国家(Tang et al., 2018; Liu et al., 2019)以及全球(Tian et al., 2017, 2019)尺度上开展了大量研究, 极大地提升和丰富了人们对叶片化学计量特征及相关理论的认识和理解.然而, 目前关于植物叶片化学计量分布规律与空间变异格局仍缺乏统一性结论.例如, 植物叶片元素含量随海拔增加呈现出无显著变化, 或显著增加, 或显著降低, 甚至先增加后减少的差异化变异规律(Fisher et al., 2013; Zhao et al., 2014, 2018).此外, 前人研究也表明植物叶片N:P随纬度增加表现出无显著变化或显著降低等多种变异趋势(Reich & Oleksyn, 2004; Han et al., 2005).基于此, 相关****提出了众多理论假说来解释植物叶片化学计量特征多样化的空间变化规律, 如温度生物地球化学假说(TBH)和温度植物生理假说(TPPH).前者主要认为低温会抑制微生物活性和土壤有机质分解与矿化, 降低土壤N、P养分可利用性, 从而导致低温环境下植物叶片N、P含量较低.而后一种假说则认为低温降低了植物的生物化学反应速率, 导致植物富N的酶及富P的RNA活性下降.相应地, 低温地区植物主要通过提高叶片N、P含量以增强其新陈代谢水平(Reich & Oleksyn, 2004; Han et al., 2005).以上两种假说分别诠释了植物叶片N、P含量随温度变化而呈现出的两种截然相反的变异规律.目前上述两种假说在不同研究中得到了相应地验证.例如, 一些研究结果支持TBH (Xia et al., 2014; Zhao et al., 2014), 而另外一些研究结果则支持TPPH (Wang et al., 2015; Tian et al., 2017), 甚至部分研究结果同时支持上述两种理论假说(Reich & Oleksyn, 2004; Shi et al., 2012).因此, 亟需选择不同典型植物, 进一步加强植物叶片化学计量空间分布格局的研究, 以验证和丰富多变环境条件下植物叶片化学计量特征变化理论假说和植物养分利用策略的科学认知. ...
... ; Zhao et al., 2014), 而另外一些研究结果则支持TPPH (Wang et al., 2015; Tian et al., 2017), 甚至部分研究结果同时支持上述两种理论假说(Reich & Oleksyn, 2004; Shi et al., 2012).因此, 亟需选择不同典型植物, 进一步加强植物叶片化学计量空间分布格局的研究, 以验证和丰富多变环境条件下植物叶片化学计量特征变化理论假说和植物养分利用策略的科学认知. ...
Shrub type dominates the vertical distribution of leaf C:N:P stoichiometry across an extensive altitudinal gradient 6 2018
... 近年来, 围绕植物叶片化学计量特征在区域(Chen et al., 2013; Fang et al., 2019)、国家(Tang et al., 2018; Liu et al., 2019)以及全球(Tian et al., 2017, 2019)尺度上开展了大量研究, 极大地提升和丰富了人们对叶片化学计量特征及相关理论的认识和理解.然而, 目前关于植物叶片化学计量分布规律与空间变异格局仍缺乏统一性结论.例如, 植物叶片元素含量随海拔增加呈现出无显著变化, 或显著增加, 或显著降低, 甚至先增加后减少的差异化变异规律(Fisher et al., 2013; Zhao et al., 2014, 2018).此外, 前人研究也表明植物叶片N:P随纬度增加表现出无显著变化或显著降低等多种变异趋势(Reich & Oleksyn, 2004; Han et al., 2005).基于此, 相关****提出了众多理论假说来解释植物叶片化学计量特征多样化的空间变化规律, 如温度生物地球化学假说(TBH)和温度植物生理假说(TPPH).前者主要认为低温会抑制微生物活性和土壤有机质分解与矿化, 降低土壤N、P养分可利用性, 从而导致低温环境下植物叶片N、P含量较低.而后一种假说则认为低温降低了植物的生物化学反应速率, 导致植物富N的酶及富P的RNA活性下降.相应地, 低温地区植物主要通过提高叶片N、P含量以增强其新陈代谢水平(Reich & Oleksyn, 2004; Han et al., 2005).以上两种假说分别诠释了植物叶片N、P含量随温度变化而呈现出的两种截然相反的变异规律.目前上述两种假说在不同研究中得到了相应地验证.例如, 一些研究结果支持TBH (Xia et al., 2014; Zhao et al., 2014), 而另外一些研究结果则支持TPPH (Wang et al., 2015; Tian et al., 2017), 甚至部分研究结果同时支持上述两种理论假说(Reich & Oleksyn, 2004; Shi et al., 2012).因此, 亟需选择不同典型植物, 进一步加强植物叶片化学计量空间分布格局的研究, 以验证和丰富多变环境条件下植物叶片化学计量特征变化理论假说和植物养分利用策略的科学认知. ...
... 此外, 影响植物叶片化学计量特征和空间分布格局的因素众多, 主要包括气候因子(Reich & Oleksyn, 2004; Yuan & Chen, 2009)、土壤特性(Chen et al., 2013; Zhang et al., 2019)和生物因素(如植物种类、生活型等)(He et al., 2006, 2008; Zhao et al., 2018).研究表明, 当研究对象涉及的植物生活型较多时, 生活型往往成为植物叶片化学计量特征变异的关键驱动因素, 并很大程度上弱化了气候因子和土壤特性的影响效应(He et al., 2006, 2008; Zhao et al., 2018).此外, 不同生活型植物叶片化学计量特征及其空间分布规律也存在较大差异.例如, 草本植物叶片N、P含量通常显著高于乔木, 而落叶树种叶片元素含量显著高于常绿树种, 且两者的叶片化学计量海拔分布格局也存在较明显的差异(Han et al., 2005; Shi et al., 2012).总之, 当生活型与气候因子、土壤特性等多种因素叠加后, 势必会加剧植物叶片化学计量特征分布规律及其驱动因素认知的复杂性和不确定性.相应地, 通过降低或者排除不同生活型对植物叶片化学计量特征的潜在影响, 专一性地聚焦生活型相同或相近的植物叶片化学计量空间分布规律及其驱动因素的深入研究, 将在很大程度上丰富不同环境因子对叶片化学计量特征与空间分布变异相对贡献的认知水平. ...
... ; Zhao et al., 2018).此外, 不同生活型植物叶片化学计量特征及其空间分布规律也存在较大差异.例如, 草本植物叶片N、P含量通常显著高于乔木, 而落叶树种叶片元素含量显著高于常绿树种, 且两者的叶片化学计量海拔分布格局也存在较明显的差异(Han et al., 2005; Shi et al., 2012).总之, 当生活型与气候因子、土壤特性等多种因素叠加后, 势必会加剧植物叶片化学计量特征分布规律及其驱动因素认知的复杂性和不确定性.相应地, 通过降低或者排除不同生活型对植物叶片化学计量特征的潜在影响, 专一性地聚焦生活型相同或相近的植物叶片化学计量空间分布规律及其驱动因素的深入研究, 将在很大程度上丰富不同环境因子对叶片化学计量特征与空间分布变异相对贡献的认知水平. ...
... 前人众多研究已清楚地表明, 气候因子(Reich & Oleksyn 2004; Yuan & Chen, 2009)、土壤特性(Chen et al., 2013; Zhang et al., 2019)和植物生活型(He et al., 2006, 2008; Zhao et al., 2018)对植物叶片化学计量特征有重要影响.对中国草地(He et al., 2006)和西南灌丛(Zhao et al., 2018)植物叶片C、N、P化学计量主要驱动因素的研究结果显示, 生活型是驱动叶片化学计量变异的主要关键因子, 并一定程度上弱化了环境因子对植物叶片化学计量特征变异的相对贡献率.在本研究中, 我们通过选择青藏高原东缘单一生活型的针叶树种叶片为研究对象, 尽可能去除或弱化生活型的潜在影响, 专一性地辨识了单一生活型植物叶片空间变异格局以及气候因子和土壤特性对叶片化学计量的相对贡献. ...
 ,
, ,1,*
,1,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT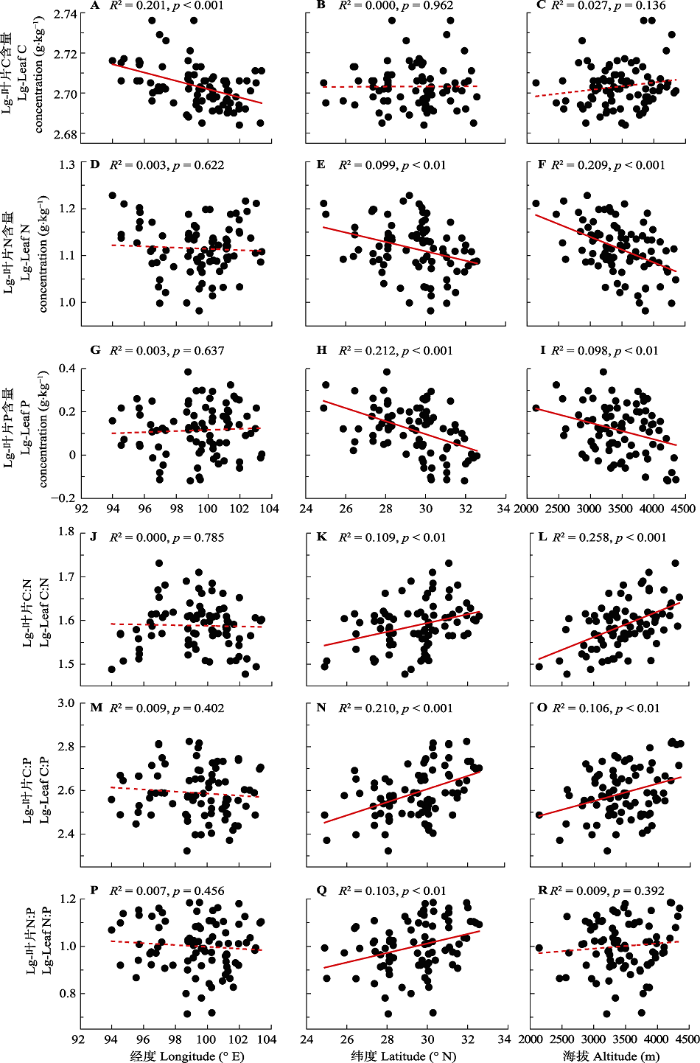 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT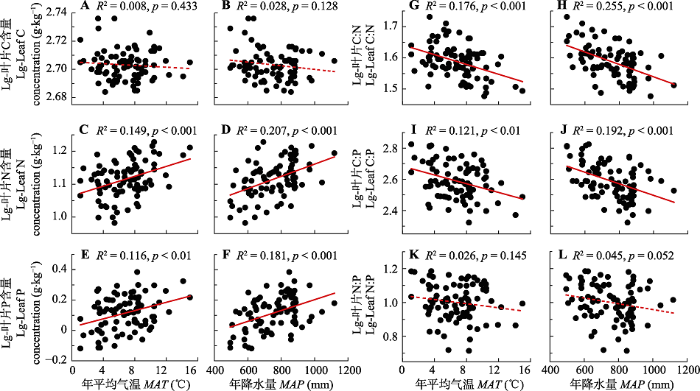 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT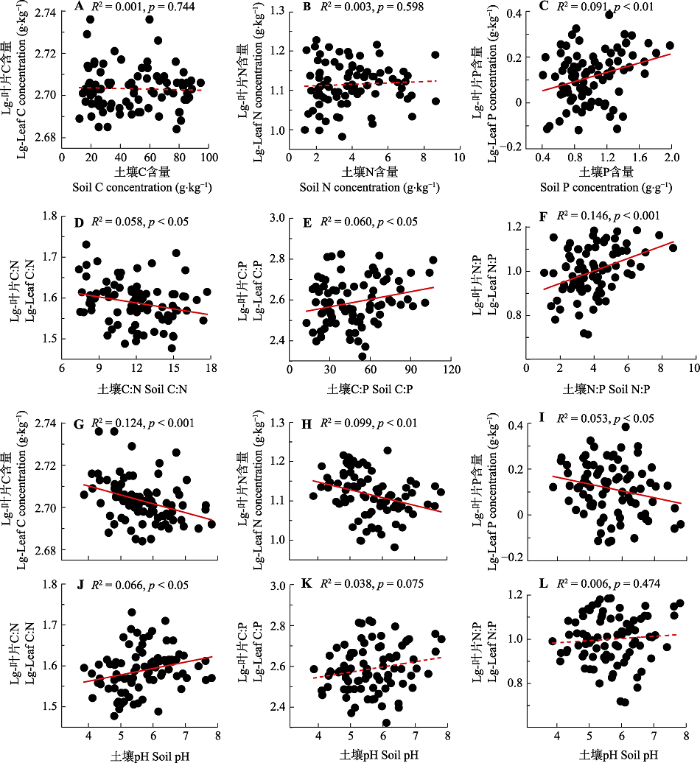 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT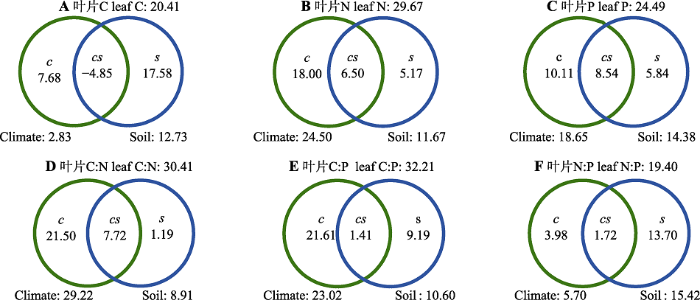 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}