Predicting phenology shifts of herbaceous plants on the Qinghai-Xizang Plateau under climate warming with the space-for-time method
Xue-Ying LI, Wen-Quan ZHU,*, Pei-Xian LI, Zhi-Ying XIE, Cen-Liang ZHAOState Key Laboratory of Earth Surface Processes and Resource Ecology, Beijing Normal University, Beijing 100875, China; and Beijing Engineering Research Center for Global Land Remote Sensing Products, Institute of Remote Sensing Science and Engineering, Faculty of Geographical Science, Beijing Normal University, Beijing 100875, China
Abstract Aims To analyse the feasibility of space-for-time method in predicting phenology shifts of Plantago asiatica and Taraxacum mongolicum on the Qinghai-Xizang Plateau, as well as revealing the phenological changes of the two herbaceous plants under climate warming. Methods The observed phenological data for Plantago asiatica and Taraxacum mongolicum from 10 sites on the Qinghai-Xizang Plateau during 2002-2011, as well as the meteorological data (i.e., daily mean air temperature) were collected. First, multiple linear regression models were bulit between geographic factors (longitude, latitude and altitude) and phenological events/annual mean temperature at different altitude gradients. Then, the longitude and latitude were kept to be unchanged, and the unary linear regression models between phenological events/annual mean temperature and altitude were built. Finally, the altitude was used as the “bridge” to indicate the relationship between the change of phenological events and the change of annual mean temperature. Important findings The temperature decreased with the increasing altitude (R2 > 0.89, p < 0.05), illustrating that changes of altitude gradients can be used to substitute for changes of annual mean temperature. The change in the simulated phenological events of the two herbaceous plants all showed a strong dependence on the change of altitude (R2 > 0.70, p < 0.05), which contributed the most among the geographic factors. Strong dependences were observed between the simulated phenological events and the simulated annual mean temperature (R2 > 0.93, p < 0.05), showing that phenological events could be predicted by the annual mean temperature with the space-for- time method. For Plantago asiatica, the first leaf date (FLD) and the first flowering date (FFD) occurred earlier with increasing annual mean temperature as 5.1 and 5.4 days per ℃, respectively, while the common leaf coloring date (LCD) occurred later as 4.8 days per ℃. The FLD and FFD of Taraxacum mongolicum advanced by 6.5 days and 7.8 days per ℃ of increase in the mean annual temperature while the LCD delayed by 6.7 days per ℃. Keywords:Qinghai-Xizang Plateau;climate warming;Plantago asiatica;Taraxacum mongolicum;space-for-time method;phenology
PDF (1438KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 李雪莹, 朱文泉, 李培先, 谢志英, 赵涔良. 气候变暖背景下青藏高原草本植物物候变化空间换时间预测. 植物生态学报, 2020, 44(7): 742-751. DOI: 10.17521/cjpe.2019.0308 LI Xue-Ying, ZHU Wen-Quan, LI Pei-Xian, XIE Zhi-Ying, ZHAO Cen-Liang. Predicting phenology shifts of herbaceous plants on the Qinghai-Xizang Plateau under climate warming with the space-for-time method. Chinese Journal of Plant Ecology, 2020, 44(7): 742-751. DOI: 10.17521/cjpe.2019.0308
2002-2011年各年份的年平均气温空间差异均与海拔梯度关系显著(p < 0.05)(表2), R2均大于0.89。海拔每升高1 000 m, 年平均气温降低6.3-6.7 ℃。控制经纬度不变, 根据海拔梯度拟合的年平均气温与各站点的海拔关系显著(R2 = 0.98): 海拔每升高1 000 m, 年平均气温降低6.5 ℃ (图2)。
Table 2 表2 表2青藏高原年平均气温与海拔逐年回归分析建模结果 Table 2Regression modeling results between mean annual air temperature and altitude across the Qinghai-Xizang Plateau
年份 Year
R2
a (℃·°-1)
b (℃·°-1)
c (℃·m-1)
d
2002
0.90*
-0.3
-1.1*
-0.006 7*
88.4
2003
0.90*
-0.2
-1.1*
-0.006 3*
83.8
2004
0.91*
-0.3
-1.1*
-0.006 3*
88.9
2005
0.91*
-0.3
-1.1*
-0.006 6*
91.3
2006
0.90*
-0.3
-1.1*
-0.006 4*
89.8
2007
0.91*
-0.3
-1.1*
-0.006 7*
95.8
2008
0.90*
-0.3
-1.1*
-0.006 7*
91.6
2009
0.90*
-0.3
-1.2*
-0.006 4*
92.8
2010
0.91*
-0.3
-1.1*
-0.006 7*
93.6
2011
0.89*
-0.3
-1.1
-0.006 3*
87.1
*表示p < 0.05。a、b、c和d分别表示经度、纬度、海拔的回归系数和截距。 * indicates p < 0.05. a, b, c and d indicate the regression coefficients of longitude, latitude, altitude and intercept, respectively.
Table 3 表3 表3青藏高原车前和蒲公英各物候事件和地理因子回归分析建模结果 Table 3Regression modeling results between each phenological event of Plantago asiatica and Taraxacum mongolicum and geographic factors across the Qinghai-Xizang Plateau
年份 Year
车前展叶始期 FLD of P. asiatica
蒲公英展叶始期 FLD of T. mongolicum
R2
a (d·°-1)
b (d·°-1)
c (d·m-1)
d
R2
a (d·°-1)
b (d·°-1)
c (d·m-1)
d
2002
0.95*
1.1
6.3*
0.033 8*
-342.3
0.95*
1.2
6.2*
0.043 8*
-382.8
2003
0.98*
0.7
6.8*
0.036 2*
-318.3
1.00*
0.9*
6.3*
0.045 2*
-362.8
2004
0.95*
1.0
6.9*
0.036 7*
-355.0
0.92*
1.3
6.1*
0.041 2*
-383.3
2005
0.90*
1.0
5.9*
0.032 9*
-320.1
0.92*
1.7
6.1*
0.043 2*
-428.5
2006
0.96*
1.5*
6.7*
0.037 3*
-404.9
0.92*
2.0*
6.6*
0.044 2*
-470.6
2007
0.91*
1.6
5.9*
0.040 0*
-398.6
0.93*
1.9*
5.7*
0.047 8*
-448.9
2008
0.85*
1.5
5.0*
0.032 7*
-327.8
0.99*
1.4*
5.7*
0.045 0*
-380.9
2009
0.93*
1.6*
5.0*
0.032 9*
-340.4
0.91*
2.0*
4.3*
0.039 4*
-385.8
2010
0.83*
0.6
4.5*
0.030 5*
-212.2
0.85*
0.9
4.3*
0.041 5*
-274.2
2011
0.67
0.7
4.5*
0.025 1
-199.5
0.70*
0.9
4.3
0.037 3*
-254.7
年份 Year
车前开花始期 FFD of P. asiatica
蒲公英开花始期 FFD of T. mongolicum
R2
a (d·°-1)
b (d·°-1)
c (d·m-1)
d
R2
a (d·°-1)
b (d·°-1)
c (d·m-1)
d
2002
0.76*
-0.1
3.9*
0.023 2*
-45.1
0.88*
0.5
6.9*
0.047 9*
-306.5
2003
0.96*
0.9
5.3*
0.032 2*
-226.1
0.89*
0.9
6.5*
0.048 8*
-334.9
2004
0.95*
0.8
5.1*
0.029 8*
-198.4
0.88*
2.0
6.5*
-0.059 6*
-480.5
2005
0.92*
1.4*
3.9*
0.030 8*
-215.4
0.91*
1.7
6.2*
0.051 7*
-418.4
2006
0.95*
1.5*
4.7*
0.030 2*
-248.2
0.89*
2.3
5.7*
0.046 7*
-439.9
2007
0.96*
2.8*
2.8*
0.034 7*
-330.0
0.91*
2.1*
5.5*
0.054 8*
-440.4
2008
0.93*
2.2*
6.7*
0.054 7*
-475.7
0.98*
3.0*
4.9*
0.058 8*
-523.0
2009
0.83*
1.9
5.5*
0.044 6*
-367.8
0.94*
2.9*
3.7*
0.057 5*
-470.9
2010
0.73*
2.7
4.0
0.040 2*
-378.8
0.89*
1.6
4.8*
0.040 2*
-313.3
2011
0.77*
2.0
4.8*
0.037 5*
-328.6
0.83*
1.2
6.5*
0.049 6*
-360.7
年份 Year
车前黄枯普遍期 LCD of P. asiatica
蒲公英黄枯普遍期 LCD of T. mongolicum
R2
a (d·°-1)
b (d·°-1)
c (d·m-1)
d
R2
a (d·°-1)
b (d·°-1)
c (d·m-1)
d
2002
0.82*
-2.7*
-4.0*
-0.035 8*
803.2
0.80*
-2.3
-4.7*
-0.048 1*
825.3
2003
0.55
-1.7
-2.9
-0.028 4
646.4
0.74*
-3.2
-2.9
-0.041 7*
847.7
2004
0.79*
-1.3
-3.1*
-0.020 5*
593.3
0.71*
-2.7
-3.2
-0.035 3*
781.8
2005
0.87*
-1.2
-4.4*
-0.031 6*
666.8
0.86*
-1.9*
-4.4*
-0.042 9*
769.7
2006
0.84*
-1.0
-3.8*
-0.026 6*
596.2
0.91*
-2.0*
-3.9*
-0.043 6*
762.4
2007
0.90*
-1.9*
-4.1*
-0.034 8*
729.8
0.89*
-2.4*
-4.4*
-0.048 7*
836.1
2008
0.79*
-2.5*
-1.8
-0.029 5*
693.2
0.78*
-3.0*
-1.7
-0.039 6*
775.7
2009
0.96*
-2.7*
-3.1*
-0.031 2*
763.4
0.90*
-2.6*
-4.2*
-0.044 4*
831.6
2010
0.91*
-3.6*
-2.6
-0.040 3*
874.7
0.93*
-3.8*
-3.0*
-0.048 3*
935.5
2011
0.91*
-2.5*
-4.0*
-0.038 6*
801.5
0.95*
-3.4*
-5.2*
-0.047 4*
961.1
*表示p < 0.05。a、b、c和d分别表示经度、纬度、海拔的回归系数和截距。 * indicates p < 0.05. a, b, c and d indicate the regression coefficients of longitude, latitude, altitude and intercept, respectively. FFD, first flowering date; FLD, first leaf date; LCD, the common leaf coloring date.
Fig. 3Regression results between fitted values of the first leaf date (FLD), first flowering date (FFD) and the common leaf coloring date (LCD) of Plantago asiatica (A-C) and Taraxacum mongolicum (D-F) and altitudes across the Qinghai-Xizang Plateau.
Table 4 表4 表4青藏高原车前和蒲公英所有站点及年份各物候事件拟合值与观测值的均方根误差 Table 4Root Mean Square Error (RMSE) values of each phenological event in all sites and years of Plantago asiatica and Taraxacum mongolicum across the Qinghai-Xizang Plateau
物候事件 Phenological event
车前均方根误差 RMSE value of P. asiatica (d)
蒲公英均方根误差 RMSE value of T. mongolicum (d)
展叶始期 FLD
5.7
6.8
开花始期 FFD
6.7
7.0
黄枯普遍期 LCD
5.4
7.0
FFD, first flowering date; FLD, first leaf date; LCD, the common leaf coloring date.
Fig. 4Regression results between fitted values of the first leaf date (FLD), first flowering date (FFD) and the common leaf coloring date (LCD) of Plantago asiatica (A-C) and Taraxacum mongolicum (D-F) and fitted values of mean annual air temperature across the Qinghai-Xizang Plateau.
Appendix I Multi-year mean values and standard deviations of each phenological event for Plantago asiatica at each site across the Qinghai-Xizang Plateau
Appendix II Multi-year mean values and standard deviations of each phenological event for Taraxacum mongolicum at each site across the Qinghai-Xizang Plateau
Appendix IV Regression modeling results between the end of fruit ripening dates (FRD) and geographic factors for Plantago asiatica and Taraxacum mongolicum across the Qinghai-Xizang Plateau
Appendix V Regression modeling results between the end of fruit ripening dates and the first flowering dates for Plantago asiatica and Taraxacum mongolicum across the Qinghai-Xizang Plateau
AhasR, AasaA, MenzelA, FedotovaVG, ScheifingerH (2002). Changes in European spring phenology International Journal of Climatology, 22, 1727-1738. [本文引用: 1]
BenistonM (2003). Climatic change in mountain regions: a review of possible impacts Climatic Change, 59, 5-31. [本文引用: 1]
BloisJL, WilliamsJW, FitzpatrickMC, JacksonST, FerrierS (2013). Space can substitute for time in predicting climate- change effects on biodiversity Proceedings of the National Academy of Sciences of the United States of America, 110, 9374-9379. URLPMID:23690569 [本文引用: 3]
BolmgrenK, VanhoenackerD, Miller-RushingAJ (2013). One man, 73 years, and 25 species. Evaluating phenological responses using a lifelong study of first flowering dates International Journal of Biometeorology, 57, 367-375. URLPMID:22744801 [本文引用: 1]
CaoYF, WuRF, YanWX (2008). Variation characteristics of Plantago phenology and its response to climate change Meteorology Journal of Inner Mongolia, (6), 8-9,31. [本文引用: 1]
ChenHL, LiuJ (2019). Response of phenological period of Plantago to climate change in grassland henanin county in Qinghai Province Qinghai Prataculture, 28(3), 39-43. [本文引用: 1]
ChenXQ, ZhangFC (2001). Spring phonological change in Beijing in the last 50 years and its response to the climatic changes Chinese Journal of Agrometeorology, 22(1), 2-6. [本文引用: 1]
ChmielewskiFM, R?tzerT (2001). Response of tree phenology to climate change across Europe Agricultural and Forest Meteorology, 108, 101-112. [本文引用: 1]
CuiS, MengF, SuonanJ, WangQ, LiB, LiuP, RenzengW, LvW, JiangL, ZhangL, LiX, LiY, ZhangZ, LuoC, TsechoeD, WangS (2017). Responses of phenology and seed production of annual Koenigia islandica to warming in a desertified alpine meadow Agricultural and Forest Meteorology, 247, 376-384. [本文引用: 1]
DaiJH, WangHJ, GeQS (2013). Multiple phenological responses to climate change among 42 plant species in Xi?an, China International Journal of Biometeorology, 57, 749-758. URLPMID:23114575 [本文引用: 1]
DaiJH, WangHJ, GeQS (2014). The spatial pattern of leaf phenology and its response to climate change in China International Journal of Biometeorology, 58, 521-528. URLPMID:23732443 [本文引用: 1]
DingMJ, ZhangYL, LiuLS, WangZF (2011). Spatiotemporal changes of commencement of vegetation regreening and its response to climate change on Tibetan Plateau Advances in Climate Change Research, 7, 317-323. [本文引用: 2]
FitzpatrickMC, SandersNJ, FerrierS, LonginoJT, WeiserMD, DunnR (2011). Forecasting the future of biodiversity: a test of single- and multi-species models for ants in North America Ecography, 34, 836-847. [本文引用: 1]
GuoSL (2002). Advances in ecology and evolutionary biology of the genus Plantago Chinese Bulletin of Botany, 19, 567-574. [本文引用: 1]
HuangWJ, GeQS, DaiJH, WangHJ (2017). Sensitivity of first flowering dates to temperature change for typical woody plants in Guiyang City, China Progress in Geography, 36, 1015-1024. [本文引用: 1]
IPCC (2013). Summary for policymakers//Stocker TF, Qin D, Plattner GK, Tignor M, Allen SK, Boschung J, Nauels A, Xia Y, Bex V, Midgley PM Climate Change 2013: the Physical Science Basis. Cambridge University Press, Cambridge, UK. 3-29. [本文引用: 2]
JiangLL, WangSP, MengFD, DuanJC, NiuHS, XuGP, ZhuXX, ZhangZH, LuoCY, CuiSJ, LiYM, LiXE, WangQ, ZhouY, BaoXY, LiYN, DorjiT, PiaoSL, CiaisP, Pe?uelasJ, DuMY, ZhaoXQ, ZhaoL, ZhangFW, WangGJ (2016). Relatively stable response of fruiting stage to warming and cooling relative to other phenological events Ecology, 97, 1961-1969. URLPMID:27859198 [本文引用: 1]
JohnsonEA, MiyanishiK (2008). Testing the assumptions of chronosequences in succession Ecology Letters, 11, 419-431. URLPMID:18341585 [本文引用: 1]
KarlTR, ArguezA, HuangB, LawrimoreJH, McMahonJR, MenneMJ, PetersonTC, VoseRS, ZhangHM (2015). Possible artifacts of data biases in the recent global surface warming hiatus Science, 348, 1469-1472. URLPMID:26044301 [本文引用: 1]
KoenigU, AbeggB (1997). Impacts of climate change on winter tourism in the Swiss Alps Journal of Sustainable Tourism, 5, 46-58. [本文引用: 1]
KongDD, ZhangQ, HuangWL, GuXH (2017). Vegetation phenology change in Tibetan Plateau from 1982 to 2013 and its related meteorological factors Acta Geographica Sinica, 72, 39-52. [本文引用: 1]
MalhiY, SilmanM, SalinasN, BushM, MeirP, SaatchiS (2010). Introduction: elevation gradients in the tropics: laboratories for ecosystem ecology and global change research Global Change Biology, 16, 3171-3175. [本文引用: 1]
MenzelA, FabianP (1999). Growing season extended in Europe Nature, 397, 659. DOI:10.1038/17709URL [本文引用: 1]
PeiSX, GuoQS, XinXB, HongM, KangY (2009). Research on plant phenological responses to climate change abroad World Forestry Research, 22(6), 31-37. [本文引用: 1]
Pe?uelasJ, SardansJ, EstiarteM, OgayaR, CarnicerJ, CollM, BarbetaA, Rivas-UbachA, LlusiàJ, GarbulskyM, FilellaI, JumpAS (2013). Evidence of current impact of climate change on life: a walk from genes to the biosphere Global Change Biology, 19, 2303-2338. URLPMID:23505157 [本文引用: 1]
PiaoSL, FangJY, ZhouLM, CiaisP, ZhuB (2006). Variations in satellite-derived phenology in China?s temperate vegetation Global Change Biology, 12, 672-685. [本文引用: 1]
QiRY, ZhaoLX (2013). Response of plantain phenology to climate change in Qinghai Journal of Anhui Agricultural Sciences, 41, 3025-3026, 3028. [本文引用: 1]
SchaberJ, BadeckFW (2002). Evaluation of methods for the combination of phenological time series and outlier detection Tree Physiology, 22, 973-982. URLPMID:12359524 [本文引用: 1]
SherryRA, ZhouX, GuS, ArnoneJA, SchimelDS, VerburgPS, WallaceLL, LuoYQ (2007). Divergence of reproductive phenology under climate warming Proceedings of the National Academy of Sciences of the United States of America, 104, 198-202. URLPMID:17182748 [本文引用: 1]
ThomasCD, CameronA, GreenRE, BakkenesM, BeaumontLJ, CollinghamYC, ErasmusBFN, deSiqueira MF, GraingerA, HannahL, HughesL, HuntleyB, vanJaarsveld AS, MidgleyGF, MilesL, Ortega-HuertaMA, TownsendPeterson A, PhillipsOL, WilliamsSE (2004). Extinction risk from climate change Nature, 427, 145-148. DOI:10.1038/nature02121URLPMID:14712274 [本文引用: 1] Climate change over the past approximately 30 years has produced numerous shifts in the distributions and abundances of species and has been implicated in one species-level extinction. Using projections of species' distributions for future climate scenarios, we assess extinction risks for sample regions that cover some 20% of the Earth's terrestrial surface. Exploring three approaches in which the estimated probability of extinction shows a power-law relationship with geographical range size, we predict, on the basis of mid-range climate-warming scenarios for 2050, that 15-37% of species in our sample of regions and taxa will be 'committed to extinction'. When the average of the three methods and two dispersal scenarios is taken, minimal climate-warming scenarios produce lower projections of species committed to extinction ( approximately 18%) than mid-range ( approximately 24%) and maximum-change ( approximately 35%) scenarios. These estimates show the importance of rapid implementation of technologies to decrease greenhouse gas emissions and strategies for carbon sequestration.
TierneyJE, RussellJM, EggermontH, HopmansEC, VerschurenD, SinningheDamsté JS (2010). Environmental controls on branched tetraether lipid distributions in tropical East African lake sediments Geochimica et Cosmochimica Acta, 74, 4902-4918. DOI:10.1016/j.gca.2010.06.002URL [本文引用: 1]
WangHJ, DaiJH, ZhengJY, GeQS (2014a). Temperature sensitivity of plant phenology in temperate and subtropical regions of China from 1850 to 2009 International Journal of Climatology, 35, 913-922. DOI:10.1002/joc.2015.35.issue-6URL [本文引用: 2]
WangSP, WangCS, DuanJC, ZhuXX, XuGP, LuoCY, ZhangZH, MengFD, LiYN, DuMY (2014b). Timing and duration of phenological sequences of alpine plants along an elevation gradient on the Tibetan Plateau Agricultural and Forest Meteorology, 189-190, 220-228. [本文引用: 3]
WuRJ, ZhengYF, ZhaoGQ, WangM (2009). Spring phenophase changes of dominant plants in Zhengzhou and their responses to air temperature change Chinese Journal of Ecology, 28, 1049-1054. [本文引用: 1]
XuGX, LuoSX, GuoQS, PeiSX, ShiZM, ZhuL, ZhuNN (2014). Responses of leaf unfolding and flowering to climate change in 12 tropical evergreen broadleaf tree species in Jianfengling, Hainan Island Chinese Journal of Plant Ecology, 38, 585-598. [本文引用: 1]
XuYJ, ZhongSY, DaiJH, TaoZX, WangHJ (2017). Changes in flowering phenology of plants and their model simulation in Mudanjiang, China Geographical Research, 36, 779-789. [本文引用: 2]
ZhangXX, GeQS, ZhengJY, ZhangFC (2005). Responses of spring phenology to climate changes in Beijing in last 150 years Chinese Journal of Agrometeorology, 26, 263-267. [本文引用: 2]
ZhaoJB, ZhangYP, SongFQ, XuZF, XiaoYL (2009). A comparison of the phenological characteristics of introduced plant species in the Xishuangbanna Tropical Botanical Garden Bulletin of Botany, 44, 464-472. [本文引用: 1]
ZhengZT (2018). Responses of Net Primary Productivity to Green-Up Dynamics in the Alpine Grassland on the Qinghai-Tibetan Plateau PhD dissertation, Beijing Normal University, Beijing. 7-29. [本文引用: 1]
ZhengZT, ZhuWQ, ChenGS, JiangN, FanDQ, ZhangDH (2016). Continuous but diverse advancement of spring- summer phenology in response to climate warming across the Qinghai-Tibetan Plateau Agricultural and Forest Meteorology, 223, 194-202. [本文引用: 1]
ZhuWQ, ZhengZT, JiangN, ZhangDH (2018). A comparative analysis of the spatio-temporal variation in the phenologies of two herbaceous species and associated climatic driving factors on the Tibetan Plateau Agricultural and Forest Meteorology, 248, 177-184. [本文引用: 1]
Changes in European spring phenology 1 2002
... 自工业革命以来, 人类活动排放温室气体导致全球陆地和海洋表面温度上升约0.7-1.1 ℃ (IPCC, 2013).21世纪温度上升速率比20世纪提高了约一倍(Karl et al., 2015), 且高纬度及高海拔地区的气温升幅更加明显(李林等, 2002; Thomas et al., 2004; Pe?uelas et al., 2013).预计在21世纪末, 全球表面温度将在21世纪初温度基础上升高0.3-4.8 ℃ (IPCC, 2013).许多研究证实气候变暖提前了植物的生物学春夏季(Menzel & Fabian, 1999; Ahas et al., 2002; 徐雨晴等, 2005; Wolkovich et al., 2012), 延迟了生物学秋冬季(张福春, 1995; Chmielewski & R?tzer, 2001; Steltzer & Post, 2009), 延长了整个植物生长季(方修琦和余卫红, 2002; Piao et al., 2006). ...
Climatic change in mountain regions: a review of possible impacts 1 2003
A comparative analysis of the spatio-temporal variation in the phenologies of two herbaceous species and associated climatic driving factors on the Tibetan Plateau 1 2018
 ,*, 李培先, 谢志英, 赵涔良北京师范大学地表过程与资源生态国家重点实验室, 北京 100875; 北京师范大学地理科学学部, 遥感科学与工程研究院, 北京市陆表遥感数据产品工程技术研究中心, 北京 100875
,*, 李培先, 谢志英, 赵涔良北京师范大学地表过程与资源生态国家重点实验室, 北京 100875; 北京师范大学地理科学学部, 遥感科学与工程研究院, 北京市陆表遥感数据产品工程技术研究中心, 北京 100875
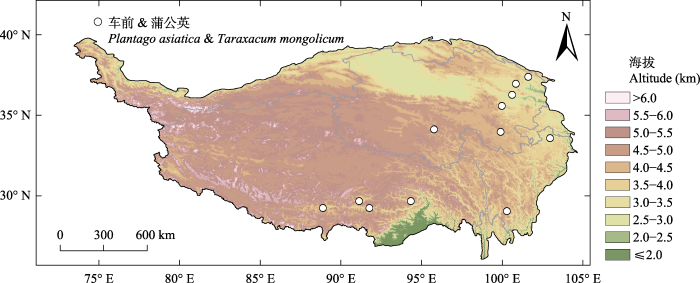 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT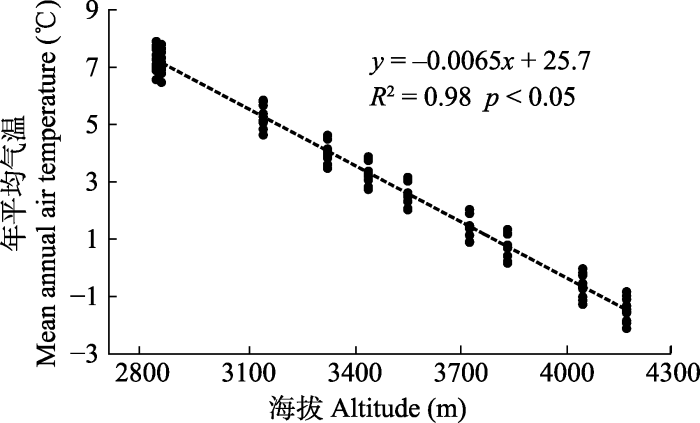 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT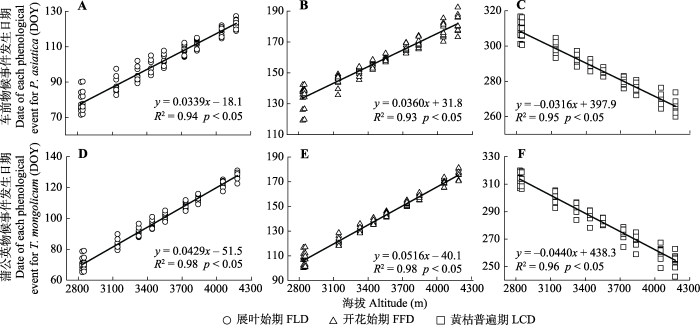 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT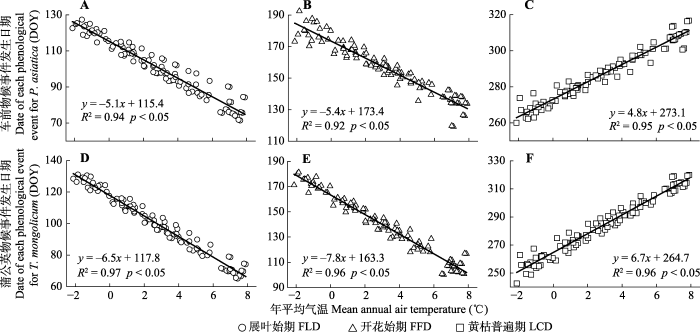 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}