Defense strategies of dominant plants under different grazing intensity in the typical temperate steppe of Nei Mongol, China
Ying LI1, Ji-Rui GONG,,1,*, Min LIU1, Xiang-Yang HOU2, Yong DING2, Bo YANG1, Zi-He ZHANG1, Biao WANG1, Chen-Chen ZHU11Laboratory of Earth Surface Processes and Resource Ecology, Beijing Key Laboratory of Traditional Chinese Medicine Protection and Utilization, School of Natural Resources, Faculty of Geographical Science, Beijing Normal University, Beijing 100875, China 2Grassland Research Institute of Chinese Academy of Agricultural Science, Hohhot 010021, China
National Natural Science Foundation of China(41571048) National Key R&D Program of China(2016YFC0500502) State Key Basic R&D Plan of China(2014CB138803)
Abstract Aims Grazing is one of the main grassland using modes, which has caused certain biotic stress on pasture. In order to survive, pasture initiates defense mechanisms by regulating the primary and secondary metabolic processes. This article aims to 1) figure out the distribution of defensive substances in different organs produced by the dominant plants Stipa grandis and Leymus chinensis; 2) explore the similarities and differences of the defense mechanisms and carbon-nitrogen trade-off strategies in S. grandis and L. chinensis. Methods We conducted a different grazing intensity experiment in the typical temperate steppe of Nei Mongol. The lignin and secondary metabolites, such as tannins, flavonoids, phenols and alkaloids, as well as the ratio of carbon and nitrogen in different organs of the dominant plants S. grandis and L. chinensis were investigated. Important findings Our results showed that S. grandis and L. chinensis produced a large number of secondary metabolites such as tannins, flavonoids, phenols and alkaloids in the face of grazing stress. The leaves were the main synthesis and storage organs. In addition, the chemical defense mechanism of L. chinensis was well induced and expressed under moderate grazing. There was no significant increase in the lignin content in the aboveground parts of S. grandis and L. chinensis. Therefore, both of them were more inclined to chemical defense than mechanical defense in the mechanism of avoiding grazing. Because of the higher nitrogen use efficiency of L. chinensis, it could be protected by both carbon and nitrogen metabolic pathways. However, Stipa grandis did not efficiently distribute nitrogen into chemical defense in the early stages of growth. More resources still used for primary metabolism in the early stage of S. grandis and L. chinensis growth, under light grazing stress, which increased their grazing resistance. Therefore, light grazing is conducive to improving the carbon and nitrogen use efficiency of pasture, and improving ecosystem productivity and stability. Keywords:secondary metabolism;defense mechanism;carbon-nitrogen trade-off;dominant species;grazing intensity;Stipa grandis;Leymus chinensis
PDF (1343KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 李颖, 龚吉蕊, 刘敏, 侯向阳, 丁勇, 杨波, 张子荷, 王彪, 朱趁趁. 不同放牧强度下内蒙古温带典型草原优势种植物防御策略. 植物生态学报, 2020, 44(6): 642-653. DOI: 10.17521/cjpe.2019.0329 LI Ying, GONG Ji-Rui, LIU Min, HOU Xiang-Yang, DING Yong, YANG Bo, ZHANG Zi-He, WANG Biao, ZHU Chen-Chen. Defense strategies of dominant plants under different grazing intensity in the typical temperate steppe of Nei Mongol, China. Chinese Journal of Plant Ecology, 2020, 44(6): 642-653. DOI: 10.17521/cjpe.2019.0329
CK, 围封; LG, 轻度放牧; MG, 中度放牧; HG, 重度放牧; EHG, 极重度放牧。不同小写字母表示处理间差异显著(p < 0.05)。 Fig. 2Lignin content of Stipa grandis and Leymus chinensis in the typical temperate steppe of Nei Mongol in June (A, B) and August (C, D) under different grazing levels (mean ± SE).
CK, grazing exclusion; LG, light grazing; MG, moderate grazing; HG, heavy grazing; EHG, extremely heavy grazing. Different lowercase letters indicate significant differences among treatments (p < 0.05).
2.2 不同放牧强度下优势种植物单宁含量的变化
在各营养器官中, 大针茅和羊草体内的单宁含量均表现为叶>茎>根。在不同放牧强度下, 大针茅和羊草叶片中的单宁含量差异显著, 而根、茎中的单宁含量差异不显著(图3, p < 0.05)。大针茅叶片中的单宁含量随放牧强度的增强而增多(图3A、3C, p < 0.05), 羊草叶片中的单宁含量在中度放牧下达到最大值(图3B、3D, p < 0.05); 随着放牧的持续, 大针茅和羊草叶片中的单宁含量增多, 而根、茎中的单宁含量减少(图3C、3D, p < 0.05)。
CK, 围封; LG, 轻度放牧; MG, 中度放牧; HG, 重度放牧; EHG, 极重度放牧。不同小写字母表示处理间差异显著(p < 0.05)。 Fig. 3Tannins content of Stipa grandis and Leymus chinensis in the typical temperate steppe of Nei Mongol in June (A, B) and August (C, D) under different grazing levels (mean ± SE).
CK, grazing exclusion; LG, light grazing; MG, moderate grazing; HG, heavy grazing; EHG, extremely heavy grazing. Different lowercase letters indicate significant differences among treatments (p < 0.05).
2.3 不同放牧强度下优势种植物总黄酮含量的变化
在各营养器官中, 大针茅和羊草体内的总黄酮含量均表现为叶>茎>根。在不同放牧强度下, 大针茅和羊草叶中的总黄酮含量差异显著, 而根、茎中总黄酮含量差异不显著(图4, p < 0.05)。在6月生长初期, 大针茅叶片中的总黄酮含量在重度放牧下达到最大值, 羊草叶片中的总黄酮含量在中度放牧下最高(图4A、4B, p < 0.05); 在8月生长末期, 羊草叶片中的总黄酮含量在轻度放牧下最低, 在重度放牧下最高(图4D)。
CK, 围封; LG, 轻度放牧; MG, 中度放牧; HG, 重度放牧; EHG, 极重度放牧。不同小写字母表示处理间差异显著(p < 0.05)。 Fig. 4Total flavonoids contents of Stipa grandis and Leymus chinensis in the typical temperate steppe of Nei Mongol in June (A, B) and August (C, D) under different grazing levels (mean ± SE).
CK, grazing exclusion; LG, light grazing; MG, moderate grazing; HG, heavy grazing; EHG, extremely heavy grazing. Different lowercase letters indicate significant differences among treatments (p < 0.05).
2.4 不同放牧强度下优势种植物总酚含量的变化
在各营养器官中, 大针茅和羊草体内的总酚含量均表现为叶>茎>根。在不同放牧强度下, 大针茅和羊草茎、叶中的总酚含量差异显著, 而根中总酚含量差异不显著(图5, p < 0.05)。在6月生长初期, 大针茅叶中的总酚含量在轻度放牧下最低, 在极重度放牧下最高; 羊草叶中的总酚含量在轻度放牧下最低, 在中度放牧下最高(图5A、5B, p < 0.05)。随着放牧的持续, 大针茅和羊草叶中的总酚含量增多。在8月生长末期, 大针茅在轻度放牧下叶中总酚含量最高, 而羊草在轻度放牧下含量最低, 在重度放牧下含量最高(图5C、5D, p < 0.05)。
CK, 围封; LG, 轻度放牧; MG, 中度放牧; HG, 重度放牧; EHG, 极重度放牧。不同小写字母表示处理间差异显著(p < 0.05)。 Fig. 5Total phenols contents of Stipa grandis and Leymus chinensis in the typical temperate steppe of Nei Mongol in June (A, B) and August (C, D) under different grazing levels (mean ± SE).
CK, grazing exclusion; LG, light grazing; MG, moderate grazing; HG, heavy grazing; EHG, extremely heavy grazing. Different lowercase letters indicate significant differences among treatments (p < 0.05).
2.5 不同放牧强度下优势种植物总生物碱含量的变化
不同放牧强度下, 大针茅和羊草叶片中的生物碱含量差异显著(图6, p < 0.05)。在6月生长初期, 大针茅叶片中的生物碱含量在轻度和极重度放牧下显著降低, 羊草叶片中的生物碱含量在中度放牧下显著提高(图6A, p < 0.05); 在8月生长末期, 大针茅叶片中的生物碱含量在轻度放牧下显著降低, 在中度和重度放牧下显著提高; 羊草叶片中的生物碱含量在轻度和中度放牧下显著提高(图6B, p < 0.05)。
CK, 围封; LG, 轻度放牧; MG, 中度放牧; HG, 重度放牧; EHG, 极重度放牧。不同小写字母表示处理间差异显著(p < 0.05)。 Fig. 6Alkaloid contents of Stipa grandis and Leymus chinensis in the typical temperate steppe of Nei Mongol in June and August under different grazing levels (mean ± SE).
CK, grazing exclusion; LG, light grazing; MG, moderate grazing; HG, heavy grazing; EHG, extremely heavy grazing. Different lowercase letters indicate significant differences among treatments (p < 0.05).
2.6 在不同生长时期次级代谢产物与碳氮的关系
大针茅和羊草在生长初期与生长末期均表现为地上部分C:N小于地下部分, 但碳氮权衡策略不同(图7; 表1)。在6月生长初期, 大针茅地上部分C:N在放牧胁迫下显著降低(图7A, p < 0.05); 含碳次级代谢产物与C:N无相关关系, 生物碱与C:N呈不显著的正相关关系。羊草地下部分C:N在放牧胁迫下显著提高(图7B, p < 0.05); 含碳次级代谢产物与C:N呈不显著的正相关关系, 生物碱与C:N呈不显著的负相关关系(表1)。在8月生长末期, 大针茅和羊草地下部分C:N在放牧胁迫下均显著降低(图7C、7D, p < 0.05), 大针茅的总酚与C:N呈显著正相关关系, 羊草的总酚与C:N呈显著负相关关系(表1)。
CK, 围封; LG, 轻度放牧; MG, 中度放牧; HG, 重度放牧; EHG, 极重度放牧。不同小写字母表示处理间差异显著(p < 0.05)。 Fig. 7The C:N in leafs of Stipa grandis and Leymus chinensis in the typical temperate steppe of Nei Mongol in June (A, B) and August (C, D) under different grazing levels (mean ± SE).
CK, grazing exclusion; LG, light grazing; MG, moderate grazing; HG, heavy grazing; EHG, extremely heavy grazing. Different lowercase letters indicate significant differences among treatments (p < 0.05).
Table 1 表1 表16月及8月内蒙古温带典型草原大针茅与羊草叶片中防御代谢产物与C:N的相关性分析 Table 1Correlation analysis of defense metabolites and C:N in leaves of Stipa grandis and Leymus chinensis in the typical temperate steppe of Nei Mongol in June and August
Al-SaeediAH, HossainMA (2015). Total phenols, total flavonoids contents and free radical scavenging activity of seeds crude extracts of pigeon pea traditionally used in Oman for the treatment of several chronic diseases Asian Pacific Journal of Tropical Disease, 5, 316-321. DOI:10.1016/S2222-1808(14)60790-8URL [本文引用: 2]
AppelHM, ArnoldTM, SchultzJC (2012). Effects of jasmonic acid, branching and girdling on carbon and nitrogen transport in poplar New Phytologist, 195, 419-426. DOI:10.1111/j.1469-8137.2012.04171.xURL [本文引用: 2] Here, we examined the impact of jasmonate (JA) treatment, branching and phloem girdling on 13C and 15N import, invertase activity and polyphenol accumulation in juvenile tissues of unbranched and branched hybrid poplar saplings (Populus nigra x P.deltoides). The import of 13C to juvenile tissues was positively correlated with invertase activity at the treatment site and enhanced by JA. Both invertase activity and 13C import were greater in shorter, younger branches and smaller, younger leaves. By contrast, JA treatments, branching and girdling had little or no impact on 15N import. In poplar saplings with multiple lateral branches, we observed almost no 13C movement from subtending source leaves into lateral branches above them, with or without JA treatment. The presence of potentially competing branches, treated with JA or not, girdled or not, had no impact on carbohydrate (CHO) import or polyphenol accumulation in target branches. We conclude that poplar branches comprise modules that are relatively independent from each other and from the stem below in terms of CHO movement, carbon-based defence production and response to elicitors. By contrast, branches are closely linked modules in terms of nitrogen movement. This should produce trees that are highly heterogeneous in quality for herbivores.
BrandoliniA, CastoldiP, PlizzariL, HidalgoA (2013). Phenolic acids composition, total polyphenols content and antioxidant activity of Triticum monococcum, Triticum turgidum and Triticum aestivum: a two-years evaluation Journal of Cereal Science, 58, 123-131. DOI:10.1016/j.jcs.2013.03.011URL [本文引用: 2]
BryantJP, ChapinFS, KleinDR (1983). Carbon/nutrient balance of boreal plants in relation to vertebrate herbivory Oikos, 40, 357-368. DOI:10.2307/3544308URL [本文引用: 1]
ChenHY, ZhangJH, HuangYM, GongJR (2014). Traits related to carbon sequestration of common plant species in a Stipa grandis steppe in Nei Mongol under different land- uses Chinese Journal of Plant Ecology, 38, 821-832. DOI:10.3724/SP.J.1258.2014.00077URL [本文引用: 4]
DongBC, FuT, LuoFL, YuFH (2017). Herbivory-induced maternal effects on growth and defense traits in the clonal species Alternanthera philoxeroides Science of the Total Environment, 605-606, 114-123. DOI:10.1016/j.scitotenv.2017.06.141URL [本文引用: 2]
ErbM (2018). Volatiles as inducers and suppressors of plant defense and immunity—Origins, specificity, perception and signaling Current Opinion in Plant Biology, 44, 117-121. DOI:10.1016/j.pbi.2018.03.008URLPMID:29674130 [本文引用: 1] Volatiles from attacked plants, microbes and herbivores can enhance plant defenses. However, the absence of volatiles rather than their presence has sometimes been associated with enhanced defense, suggesting that volatiles may also act as defense suppressors. Recent work provides a potential mechanistic explanation for these observations by showing that volatile cues can modulate different hormonal pathways, including jasmonate (JA), salicylic acid (SA) and auxin (IAA) signaling. Many of these pathways interact with each other through crosstalk. Thus, volatiles may suppress plant defenses through negative hormonal crosstalk. Hormonal crosstalk may also allow plants to integrate different volatile cues to respond specifically and appropriately to environmental change.
FuchsB, KrischkeM, MuellerMJ, KraussJ (2017a). Herbivore- specific induction of defence metabolites in a grass-endophyte association Functional Ecology, 31, 318-324. DOI:10.1111/fec.2017.31.issue-2URL [本文引用: 3]
FuchsB, KrischkeM, MuellerMJ, KraussJ (2017b). Plant age and seasonal timing determine endophyte growth and alkaloid biosynthesis Fungal Ecology, 29, 52-58. DOI:10.1016/j.funeco.2017.06.003URL [本文引用: 1]
Garland JrT (2014). Trade-offs Current Biology, 24, 60-61. [本文引用: 1]
GuoQ, MajorIT, HoweGA (2018). Resolution of growth- defense conflict: mechanistic insights from jasmonate signaling Current Opinion in Plant Biology, 44, 72-81. DOI:10.1016/j.pbi.2018.02.009URLPMID:29555489 [本文引用: 4] Induced plant resistance depends on the production of specialized metabolites that repel attack by biotic aggressors and is often associated with reduced growth of vegetative tissues. Despite progress in understanding the signal transduction networks that control growth-defense tradeoffs, much remains to be learned about how growth rate is coordinated with changes in metabolism during growth-to-defense transitions. Here, we highlight recent advances in jasmonate research to suggest how a major branch of plant immunity is dynamically regulated to calibrate growth-defense balance with shifts in carbon availability. We review evidence that diminished growth, as an integral facet of induced resistance, may optimize the temporal and spatial expression of defense compounds without compromising other critical roles of central metabolism. New insights into the evolution of jasmonate signaling further suggest that opposing selective pressures associated with too much or too little defense may have shaped the emergence of a modular jasmonate pathway that integrates primary and specialized metabolism through the control of repressor-transcription factor complexes. A better understanding of the mechanistic basis of growth-defense balance has important implications for boosting plant productivity, including insights into how these tradeoffs may be uncoupled for agricultural improvement.
HermsDA, MattsonWJ (1992). The dilemma of plants: to grow or defend The Quarterly Review of Biology, 67, 283-335. DOI:10.1086/417659URL [本文引用: 1]
Herrero-JáureguiC, OesterheldM (2018). Effects of grazing intensity on plant richness and diversity: a meta-analysis Oikos, 127, 757-766. DOI:10.1111/oik.2017.v127.i6URL [本文引用: 1]
HuJ, HouXY, SaRL, LiXL, WangZ, DingY (2016). Regulatory effects of stip grandis on above-ground biomass of plant community in grazing ecosystem Acta Agrestia Sinica, 24, 1-11. [本文引用: 2]
IsahT (2019). Stress and defense responses in plant secondary metabolites production Biological Research, 52, 39. DOI: 10.1186/s40659-019-0246-3. DOI:10.1186/s40659-019-0246-3URLPMID:31358053 [本文引用: 6] In the growth condition(s) of plants, numerous secondary metabolites (SMs) are produced by them to serve variety of cellular functions essential for physiological processes, and recent increasing evidences have implicated stress and defense response signaling in their production. The type and concentration(s) of secondary molecule(s) produced by a plant are determined by the species, genotype, physiology, developmental stage and environmental factors during growth. This suggests the physiological adaptive responses employed by various plant taxonomic groups in coping with the stress and defensive stimuli. The past recent decades had witnessed renewed interest to study abiotic factors that influence secondary metabolism during in vitro and in vivo growth of plants. Application of molecular biology tools and techniques are facilitating understanding the signaling processes and pathways involved in the SMs production at subcellular, cellular, organ and whole plant systems during in vivo and in vitro growth, with application in metabolic engineering of biosynthetic pathways intermediates.
KabirMSH, AhmadS, MahamoudMS, MasumMAA, KamalATMM, HoqueMA, ChakrabartyN, HasanatA, AdnanM (2015). Evaluation of Total condensed tannin content and anthelmintic activities of organic extracts of four Bangladeshi plants on Tubifex tubifex worm using in vitro method International Journal of Pharmacy, 5, 903-910. [本文引用: 1]
LiJM, JinZX, ZhouY (2007). Endangered species long Ye Torreya jackii secondary metabolic content analysis Journal of Fujian Forestry Science and Technology, 34, 29-32. [本文引用: 2]
LiSY (2013). Study on Determination of Fifteen Alkaloids and Their Distribution in Grass and Derived Products by Liquid Chromatography Tandem Mass Spectrometry Chinese Academy of Agricultural Sciences, Beijing. [本文引用: 1]
LiXL, LiuZY, HouXY, WuXH, WangZ, HuJ, WuZN (2015). Plant functional traits and their trade-offs in response to grazing: a review Chinese Bulletin of Botany, 50, 159-170. DOI:10.3724/SP.J.1259.2015.00159URL [本文引用: 2]
LinKM, YeFM, LinY, LiQS (2010). Research advances of phenolic functional mechanisms in soils and plants Chinese Journal of Eco-Agriculture, 18, 1130-1137. URL [本文引用: 2] Phenol is an important element of secondary metabolism. It affects plant growth, nutrient absorption, physiological characteristics, enzyme activity, soil and micro-organism in growth environment, etc. This paper describes phenolic functional mechanisms in soils and plants. The content and distribution of phenol in plant ecosystem, its effect on physiological and biochemical characteristics of plants, and on physical and chemical properties of soils were discussed too. The paper prospects the research trend of phenolic substances in practical agriculture and forestry to provide the basis for solving phenol-related production decline in agriculture and forestry. [ 林开敏, 叶发茂, 林艳, 李卿叁 (2010). 酚类物质对土壤和植物的作用机制研究进展 中国生态农业学报, 18, 1130-1137.] [本文引用: 2]
LiuM, GongJR, LiY, LiXB, YangB, ZhangZH, YangLL, HouXY (2019a). Growth-defense trade-off regulated by hormones in grass plants growing under different grazing intensities Physiologia Plantarum, 166, 553-569. DOI:10.1111/ppl.12802URLPMID:30091152 [本文引用: 5] Herbivory creates conflicts between a plant's need to allocate resources for growth and defense. It is not yet clear how plants rebalance resource utilization between growth and defense in response to increasing grazing intensity. We measured characteristics of the primary and secondary metabolism of Leymus chinensis at five levels of grazing intensity (control, light, moderate, heavy and extremely heavy). Furthermore, we evaluated hormone signaling by quantifying the impact of key hormones on plant growth and defense. Under light grazing intensity, indole-3-acetic acid and jasmonates appeared to promote the growth of L. chinensis through a high photosynthetic rate, high water-use efficiency and high soluble protein contents, whereas abscisic acid decreased these properties. Under moderate grazing intensity, L. chinensis had a low photosynthetic capacity but greater production of secondary metabolites (tannins, total flavonoids and total phenols), possibly induced by salicylic acid. When the grazing pressure further intensified, L. chinensis translocated more carbohydrates to its roots in order to survive and regrow. Leymus chinensis therefore exhibited a trade-off between growth and defense in order to survive and reproduce under herbivory. Plants developed different mechanisms to enhance their grazing tolerance by means of hormonal regulation.
LiuMM, JiaL, ZhangHQ, ZangXL, ZhangRM, GaoY (2015). Mechanical damage on secondary metabolites from Artemisia frigida leaves Journal of Zhejiang A & F University, 32, 845-852. [本文引用: 1]
LiuYY, WangQ, ZhangZY, TongLJ, WangZQ, LiJL (2019b). Grassland dynamics in responses to climate variation and human activities in China from 2000 to 2013 Science of the Total Environment, 690, 27-39. DOI:10.1016/j.scitotenv.2019.06.503URL [本文引用: 1]
LouaultF, PillarVD, AufrèreJ, GarnierE, SoussanaJF (2005). Plant traits and functional types in response to reduced disturbance in a semi-natural grassland Journal of Vegetation Science, 16, 151-160. DOI:10.1111/jvs.2005.16.issue-2URL [本文引用: 2]
LuH, McKnightTD (1999). Tissue-specific expression of the β-subunit of tryptophan synthase in Camptotheca acuminata, an indole alkaloid-producing plant Plant Physiology, 120, 43-52. DOI:10.1104/pp.120.1.43URL [本文引用: 1]
MathesiusU (2018). Flavonoid functions in plants and their interactions with other organisms Plants, 7, 30. DOI: 10.3390/plants7020030. DOI:10.3390/plants7020030URL [本文引用: 2]
MundimFM, PringleEG (2018). Whole-plant metabolic allocation under water stress Frontiers in Plant Science, 9, 852. DOI: 10.3389/fpls.2018.00852. DOI:10.3389/fpls.2018.00852URLPMID:29988542 [本文引用: 2] Trade-offs between plant growth and defense depend on environmental resource availability. Plants are predicted to prioritize growth when environmental resources are abundant and defense when environmental resources are scarce. Nevertheless, such predictions lack a whole-plant perspective-they do not account for potential differences in plant allocation above- and belowground. Such accounting is important because leaves and roots, though both critical to plant survival and fitness, differ in their resource-uptake roles and, often, in their vulnerability to herbivores. Here we aimed to determine how water availability affects plant allocation to multiple metabolic components of growth and defense in both leaves and roots. To do this, we conducted a meta-analysis of data from experimental studies in the literature. We assessed plant metabolic responses to experimentally reduced water availability, including changes in growth, nutrients, physical defenses, primary metabolites, hormones, and other secondary metabolites. Both above- and belowground, reduced water availability reduced plant biomass but increased the concentrations of primary metabolites and hormones. Importantly, however, reduced water had opposite effects in different organs on the concentrations of other secondary metabolites: reduced water increased carbon-based secondary metabolites in leaves but reduced them in roots. In addition, plants suffering from co-occurring drought and herbivory stresses exhibited dampened metabolic responses, suggesting a metabolic cost of multiple stresses. Our study highlights the needs for additional empirical studies of whole-plant metabolic responses under multiple stresses and for refinement of existing plant growth-defense theory in the context of whole plants.
Olmo-GarcíaL, KesslerN, NeuwegerH, WendtK, Olmo- PeinadoJ, Fernández-GutiérrezA, BaessmannC, Carrasco- PancorboA (2018). Unravelling the distribution of secondary metabolites in Olea europaea L.: exhaustive characterization of eight olive-tree derived matrices by complementary platforms (LC-ESI/APCI-MS and GC-APCI-MS) Molecules, 23, 2419. DOI: 10.3390/molecules23102419. DOI:10.3390/molecules23102419URL [本文引用: 1]
PuJS (2016). Determination, Extraction and Concentration Analysis of Alkaloid in Mulberry Leaves Master degree dissertation, Southwest University, Chongqing. [本文引用: 1]
QinHY, LiW, DaiLY (2015). Research progress of plant metabolites function on resistant response Chinese Agricultural Science Bulletin, 31, 256-259. [本文引用: 1]
QinJ, HanGD, QiaoJ, WuQ, JinYX (2016). Response of Leymus chinensis above-ground biomass to grazing intensity in different grasslands of Inner Mongolia Chinese Journal of Grassland, 38, 76-82. [本文引用: 1]
SasakiT, LauenrothWK (2011). Dominant species, rather than diversity, regulates temporal stability of plant communities Oecologia, 166, 761-768. DOI:10.1007/s00442-011-1916-1URL [本文引用: 1] Bouteloua gracilis, in stabilizing plant communities. We also examined the relationships between the variables of diversity and dominance hierarchy and the statistical components of temporal stability. We found a significant negative relationship between temporal stability and species richness, number of rare species, and relative abundance of rare species, whereas a significant positive relationship existed between temporal stability and relative abundance of the dominant species. Variances and covariances summed over all species significantly increased with increasing species richness, whereas they significantly decreased with increasing relative abundance of dominant species. We showed that temporal stability in a shortgrass steppe plant community was controlled by dominant species rather than by diversity itself. The generality of diversity–stability relationships might be restricted by the dynamics of dominant species, especially when they have characteristics that contribute to stability in highly stochastic systems. A clear implication is that dominance hierarchies and their changes might be among the most important ecological components to consider in managing communities to maintain ecosystem functioning.]]>
SrinivasaYB (2018). Breaching plant defence theories: growth rates of plants directly impact the evolution of consumption rates of herbivorous insects Current Science, 114, 258-260. DOI:10.18520/cs/v114/i02/258-260URL [本文引用: 3]
SwemmerAM, KnappAK (2008). Defoliation synchronizes aboveground growth of co-occurring C4 grass species Ecology, 89, 2860-2867. DOI:10.1890/07-1434.1URLPMID:18959323 [本文引用: 1] The aboveground net primary productivity (ANPP) of grass communities in grasslands and savannas is primarily determined by precipitation quantity. Recent research, motivated by predictions of changes in the distribution of rainfall events by global climate change models, indicates that ANPP may be affected by rainfall distribution as much as by annual totals. Grazing and community composition are also known to affect grassland ANPP. The manner in which interactions between rainfall distribution, grazing, and community composition affect the relationship between precipitation and ANPP represents a critical knowledge gap. The effects of community composition and grazing on aboveground growth responses to intraseasonal variation in water availability were investigated at seven grassland sites with a nonselective clipping experiment. The aboveground growth of the dominant C4 species at each site was measured at regular intervals for 2-3 growing seasons in the presence or absence of regular defoliation. In the absence of defoliation, there was a general lack of synchrony of intraseasonal growth among co-occurring species. Variation in growth rates was high and was only partially explained by variation in rainfall. Regular defoliation increased growth synchrony at all sites, but changes in growth responses to rainfall varied between sites. These results suggest that community composition will be important in determining ANPP-precipitation relationships under conditions of altered rainfall distribution. However this effect appears to be a result of species responding differently to soil water or other resources rather than to rainfall per se. Grazing may override the effects of community composition by reducing differences in growth patterns between species and has the potential to weaken precipitation controls on ANPP.
van SoestPJ (1967). Development of a comprehensive system of feed analyses and its application to forages Journal of Animal Science, 26, 119-128. DOI:10.2527/jas1967.261119xURL [本文引用: 1]
XuR, NanZB, ZhouYF, LiCJ (2012). Distribution and seasonal dynamics of ergot alkaloids in Elymus dahuricus- endophytic fungus symbiont Acta Prataculturae Sinica, 21, 84-92. [本文引用: 2]
XuS, GongJR, ZhangZY, LiuM, WangYH, LuoQP (2014). The ecological stoichiometry of dominant species in different land uses type of grassland Acta Prataculturae Sinica, 23, 45-53. DOI:10.11686/cyxb20140606URL [本文引用: 1] Stipa grandis, Artemisia frigida and Leymus chinensis were measured in different land use (grazing-exclusion, grazing and mowing) in the temperate grassland in Inner Mongolia. The seasonal variations of stoichiometric characteristic of plants and soil under different grassland uses were discussed. The stoichiometric characteristic response mechanisms of the plants and the nutrient limiting conditions were analyzed. The results show that obvious seasonal variations in C, N, P concentrations and their mass ratio of the three species exist in each grassland use type. Grassland use type, species and their interactions have significant impacts on N, P concentrations and the ratios C∶N, C∶P and N∶P, among which C concentration is relatively stable in the whole growing season and different uses show no significant impact on it. N concentration reduces overall. N and P concentrations in leaves are higher in grazing plot than those in grazing-exclusion and mowing plots. It shows that grazing can increased N and P concentrations. On the whole, plant growth is limited by N. Characteristics of soil nutrients among different grassland uses have certain differences, where comparably more serious N deficiency happens in the enclosed plot and P deficiency in the grazing plot. Generally the organic matter, C, N and P concentrations in the grazing plot are higher than those in the mowing and enclosed plots. It shows that soil nutrients can be increased which can promote plants growth in grazing plot.]]> [ 徐沙, 龚吉蕊, 张梓榆, 刘敏, 王忆慧, 罗亲普 (2014). 不同利用方式下草地优势植物的生态化学计量特征 草业学报, 23, 45-53.] [本文引用: 1]
YanXF, WangY, LiYM (2007). Plant secondary metabolism and its response to environment Acta Ecologica Sinica, 27, 2554-2562. [本文引用: 2]
ZhaiZW, GongJR, LuoQP, PanY, BaoY, XuS, LiuM, YangLL (2017). Effects of nitrogen addition on photosynthetic characteristics of Leymus chinensis in the temperate grassland of Nei Mongol, China Chinese Journal of Plant Ecology, 41, 196-208. DOI:10.17521/cjpe.2016.0128URL [本文引用: 3]
ZhaoQ, DixonRA (2011). Transcriptional networks for lignin biosynthesis: More complex than we thought? Trends in Plant Science, 16, 227-233. DOI:10.1016/j.tplants.2010.12.005URL [本文引用: 2] Lignin is an aromatic heteropolymer and the second most abundant plant biopolymer after cellulose. It is deposited mostly in the secondary cell walls of vascular plants and is essential for water transport, mechanical support and for plant pathogen defense. Lignin biosynthesis is a highly energy-consuming and irreversible process that responds to many developmental and environmental cues, including light, sugar content, circadian clock, plant hormones and wounding. During the past decade, many transcription factors involved in lignin biosynthesis have been identified and characterized. In this review, we assess how these transcriptional activators and repressors modulate lignin biosynthesis, and discuss crosstalk between the lignin biosynthesis pathway and other physiological processes.
ZhouGY, ZhouXH, HeYH, ShaoJJ, HuZH, LiuRQ, ZhouHM, HosseinibaiS (2017). Grazing intensity significantly affects belowground carbon and nitrogen cycling in grassland ecosystems: a meta-analysis Global Change Biology, 23, 1167-1179. DOI:10.1111/gcb.13431URLPMID:27416555 [本文引用: 2] Livestock grazing activities potentially alter ecosystem carbon (C) and nitrogen (N) cycles in grassland ecosystems. Despite the fact that numerous individual studies and a few meta-analyses had been conducted, how grazing, especially its intensity, affects belowground C and N cycling in grasslands remains unclear. In this study, we performed a comprehensive meta-analysis of 115 published studies to examine the responses of 19 variables associated with belowground C and N cycling to livestock grazing in global grasslands. Our results showed that, on average, grazing significantly decreased belowground C and N pools in grassland ecosystems, with the largest decreases in microbial biomass C and N (21.62% and 24.40%, respectively). In contrast, belowground fluxes, including soil respiration, soil net N mineralization and soil N nitrification increased by 4.25%, 34.67% and 25.87%, respectively, in grazed grasslands compared to ungrazed ones. More importantly, grazing intensity significantly affected the magnitude (even direction) of changes in the majority of the assessed belowground C and N pools and fluxes, and C : N ratio as well as soil moisture. Specifically,light grazing contributed to soil C and N sequestration whereas moderate and heavy grazing significantly increased C and N losses. In addition, soil depth, livestock type and climatic conditions influenced the responses of selected variables to livestock grazing to some degree. Our findings highlight the importance of the effects of grazing intensity on belowground C and N cycling, which may need to be incorporated into regional and global models for predicting effects of human disturbance on global grasslands and assessing the climate-biosphere feedbacks.
ZhouSQ, LouYR, TzinV, JanderG (2015). Alteration of plant primary metabolism in response to insect herbivory Plant Physiology, 169, 1488-1498. DOI:10.1104/pp.15.01405URLPMID:26378101 [本文引用: 7] Plants in nature, which are continuously challenged by diverse insect herbivores, produce constitutive and inducible defenses to reduce insect damage and preserve their own fitness. In addition to inducing pathways that are directly responsible for the production of toxic and deterrent compounds, insect herbivory causes numerous changes in plant primary metabolism. Whereas the functions of defensive metabolites such as alkaloids, terpenes, and glucosinolates have been studied extensively, the fitness benefits of changes in photosynthesis, carbon transport, and nitrogen allocation remain less well understood. Adding to the complexity of the observed responses, the feeding habits of different insect herbivores can significantly influence the induced changes in plant primary metabolism. In this review, we summarize experimental data addressing the significance of insect feeding habits, as related to herbivore-induced changes in plant primary metabolism. Where possible, we link these physiological changes with current understanding of their underlying molecular mechanisms. Finally, we discuss the potential fitness benefits that host plants receive from altering their primary metabolism in response to insect herbivory. Total phenols, total flavonoids contents and free radical scavenging activity of seeds crude extracts of pigeon pea traditionally used in Oman for the treatment of several chronic diseases 2 2015
Effects of jasmonic acid, branching and girdling on carbon and nitrogen transport in poplar 2 2012
... 食草动物的啃食胁迫会诱导牧草产生耐牧和避牧两种防御机制(赵钢和崔泽仁, 1999).牧草一方面会调控体内代谢途径, 调配初级代谢产物以修复损伤, 通过补偿生长达到耐牧的目的(Appel et al., 2012; Zhou et al., 2015; Guo et al., 2018); 另一方面, 牧草个体趋向于矮小化、减缓生长速度, 伴随着体内木质素等物质含量的增多, 以提高自身机械防御能力, 并诱导化学防御机制的表达, 产生各类次级代谢产物以提高抗性(李西良等, 2015; Srinivasa, 2018).已有研究表明, 在受到严重放牧胁迫时, 牧草更倾向于避牧防御而非补偿生长(Dong et al., 2017; Liu et al., 2019a).但目前很多与牧草次级代谢相关的研究主要集中在模拟啃食实验, 不能完全反映真实的放牧胁迫(刘盟盟等, 2015; Zhou et al., 2015; Fuchs et al., 2017b).羊草是无性繁殖的根茎型禾草, 繁育过程需要大量的资源, 且由于对食草动物的适口性较高, 更易被采食; 大针茅为叶呈披针状的丛生型禾草, 较羊草有更高的木质素含量, 这些生理差异可能使得二者的防御策略有所不同(Louault et al., 2005; 陈慧颖等, 2014; 胡静等, 2016).因此, 大针茅与羊草面对实际放牧胁迫时更倾向于机械防御还是化学防御仍需进一步探讨. ...
... 牧草在生长过程中, 尤其在生长初期, 需要与外界生物环境竞争有限的资源, 次级代谢过程比初级代谢对碳氮资源的消耗更大, 因此, 牧草面对食草动物的啃食胁迫时需要权衡利弊, 在生长与防御间做出选择(Srinivasa, 2018; Liu et al., 2019a).在6月生长初期, 大针茅和羊草的含碳次级代谢产物含量均在轻度放牧下最低, 这可能是由于生长竞争压力大于啃食胁迫, 所以化学防御机制并没有得到充分的诱导及表达, 更多的资源可能用于补偿生长, 提高了耐牧性(Appel et al., 2012; Zhou et al., 2015; Guo et al., 2018).在8月生长末期, 羊草在重度放牧胁迫下具有较低的C:N, 把更多的碳资源用于次级代谢产物的合成.说明胁迫时间的延长、胁迫强度的增加, 到生长竞争压力不及放牧胁迫时, 化学防御机制才被逐步诱导激活.这一结果与Herms提出的生长-分化平衡假说(growth-differentiation balance hypothesis)相一致(Herms & Mattson, 1992). ...
Phenolic acids composition, total polyphenols content and antioxidant activity of Triticum monococcum, Triticum turgidum and Triticum aestivum: a two-years evaluation 2 2013
... 植物在生长过程中所需的资源, 比如能量、空间、水分及养分通常都是有限的(Garland Jr, 2014).随着放牧强度的增强, 植物所能获取的资源会更加有限(Zhou et al., 2017).在面对啃食胁迫时, 植物会把原本用于生长繁殖的部分资源用来提高自身的抗性, 而植物的防御又是一个对碳氮资源消耗较大的过程, 所以, 植物需要权衡生长和防御的利弊, 以保证自己能在有限的资源下健康地生长繁殖(Srinivasa, 2018; Yang et al., 2018; Liu et al., 2019a).由于牛羊等脊椎动物没有饮食特异性, 因此它们可以诱导牧草产生更多种类而非单一性质的化学防御物质(Zhou et al., 2015; Isah, 2019).萜烯类及酚类等次级代谢产物是以碳为基础的物质, 而近年来引起了广泛关注的生物碱主要利用氮作为原料(Brandolini et al., 2013; Fuchs et al., 2017a).因此, 植物在产生各类化学防御物质时也应该存在某种权衡(Dong et al., 2017; Yang et al., 2018; Isah, 2019).在养分不足的环境条件下, 植物主要生成以碳为基础的物质; 而在养分充足或者低碳环境条件下, 主要生成以氮为基础的物质(Bryant et al., 1983; 阎秀峰等, 2007).羊草较大针茅有更高的全碳及全氮含量(陈慧颖等, 2014).因此, 二者对碳氮资源的权衡策略可能有所不同, 在不同放牧强度下, 大针茅与羊草的化学防御策略会发生怎样的变化仍需深入探讨. ...
Carbon/nutrient balance of boreal plants in relation to vertebrate herbivory 1 1983
... 植物在生长过程中所需的资源, 比如能量、空间、水分及养分通常都是有限的(Garland Jr, 2014).随着放牧强度的增强, 植物所能获取的资源会更加有限(Zhou et al., 2017).在面对啃食胁迫时, 植物会把原本用于生长繁殖的部分资源用来提高自身的抗性, 而植物的防御又是一个对碳氮资源消耗较大的过程, 所以, 植物需要权衡生长和防御的利弊, 以保证自己能在有限的资源下健康地生长繁殖(Srinivasa, 2018; Yang et al., 2018; Liu et al., 2019a).由于牛羊等脊椎动物没有饮食特异性, 因此它们可以诱导牧草产生更多种类而非单一性质的化学防御物质(Zhou et al., 2015; Isah, 2019).萜烯类及酚类等次级代谢产物是以碳为基础的物质, 而近年来引起了广泛关注的生物碱主要利用氮作为原料(Brandolini et al., 2013; Fuchs et al., 2017a).因此, 植物在产生各类化学防御物质时也应该存在某种权衡(Dong et al., 2017; Yang et al., 2018; Isah, 2019).在养分不足的环境条件下, 植物主要生成以碳为基础的物质; 而在养分充足或者低碳环境条件下, 主要生成以氮为基础的物质(Bryant et al., 1983; 阎秀峰等, 2007).羊草较大针茅有更高的全碳及全氮含量(陈慧颖等, 2014).因此, 二者对碳氮资源的权衡策略可能有所不同, 在不同放牧强度下, 大针茅与羊草的化学防御策略会发生怎样的变化仍需深入探讨. ...
内蒙古大针茅草原常见植物在不同土地利用方式下的固碳相关属性 4 2014
... 食草动物的啃食胁迫会诱导牧草产生耐牧和避牧两种防御机制(赵钢和崔泽仁, 1999).牧草一方面会调控体内代谢途径, 调配初级代谢产物以修复损伤, 通过补偿生长达到耐牧的目的(Appel et al., 2012; Zhou et al., 2015; Guo et al., 2018); 另一方面, 牧草个体趋向于矮小化、减缓生长速度, 伴随着体内木质素等物质含量的增多, 以提高自身机械防御能力, 并诱导化学防御机制的表达, 产生各类次级代谢产物以提高抗性(李西良等, 2015; Srinivasa, 2018).已有研究表明, 在受到严重放牧胁迫时, 牧草更倾向于避牧防御而非补偿生长(Dong et al., 2017; Liu et al., 2019a).但目前很多与牧草次级代谢相关的研究主要集中在模拟啃食实验, 不能完全反映真实的放牧胁迫(刘盟盟等, 2015; Zhou et al., 2015; Fuchs et al., 2017b).羊草是无性繁殖的根茎型禾草, 繁育过程需要大量的资源, 且由于对食草动物的适口性较高, 更易被采食; 大针茅为叶呈披针状的丛生型禾草, 较羊草有更高的木质素含量, 这些生理差异可能使得二者的防御策略有所不同(Louault et al., 2005; 陈慧颖等, 2014; 胡静等, 2016).因此, 大针茅与羊草面对实际放牧胁迫时更倾向于机械防御还是化学防御仍需进一步探讨. ...
... 植物在生长过程中所需的资源, 比如能量、空间、水分及养分通常都是有限的(Garland Jr, 2014).随着放牧强度的增强, 植物所能获取的资源会更加有限(Zhou et al., 2017).在面对啃食胁迫时, 植物会把原本用于生长繁殖的部分资源用来提高自身的抗性, 而植物的防御又是一个对碳氮资源消耗较大的过程, 所以, 植物需要权衡生长和防御的利弊, 以保证自己能在有限的资源下健康地生长繁殖(Srinivasa, 2018; Yang et al., 2018; Liu et al., 2019a).由于牛羊等脊椎动物没有饮食特异性, 因此它们可以诱导牧草产生更多种类而非单一性质的化学防御物质(Zhou et al., 2015; Isah, 2019).萜烯类及酚类等次级代谢产物是以碳为基础的物质, 而近年来引起了广泛关注的生物碱主要利用氮作为原料(Brandolini et al., 2013; Fuchs et al., 2017a).因此, 植物在产生各类化学防御物质时也应该存在某种权衡(Dong et al., 2017; Yang et al., 2018; Isah, 2019).在养分不足的环境条件下, 植物主要生成以碳为基础的物质; 而在养分充足或者低碳环境条件下, 主要生成以氮为基础的物质(Bryant et al., 1983; 阎秀峰等, 2007).羊草较大针茅有更高的全碳及全氮含量(陈慧颖等, 2014).因此, 二者对碳氮资源的权衡策略可能有所不同, 在不同放牧强度下, 大针茅与羊草的化学防御策略会发生怎样的变化仍需深入探讨. ...
... 食草动物的啃食胁迫会诱导牧草产生耐牧和避牧两种防御机制(赵钢和崔泽仁, 1999).牧草一方面会调控体内代谢途径, 调配初级代谢产物以修复损伤, 通过补偿生长达到耐牧的目的(Appel et al., 2012; Zhou et al., 2015; Guo et al., 2018); 另一方面, 牧草个体趋向于矮小化、减缓生长速度, 伴随着体内木质素等物质含量的增多, 以提高自身机械防御能力, 并诱导化学防御机制的表达, 产生各类次级代谢产物以提高抗性(李西良等, 2015; Srinivasa, 2018).已有研究表明, 在受到严重放牧胁迫时, 牧草更倾向于避牧防御而非补偿生长(Dong et al., 2017; Liu et al., 2019a).但目前很多与牧草次级代谢相关的研究主要集中在模拟啃食实验, 不能完全反映真实的放牧胁迫(刘盟盟等, 2015; Zhou et al., 2015; Fuchs et al., 2017b).羊草是无性繁殖的根茎型禾草, 繁育过程需要大量的资源, 且由于对食草动物的适口性较高, 更易被采食; 大针茅为叶呈披针状的丛生型禾草, 较羊草有更高的木质素含量, 这些生理差异可能使得二者的防御策略有所不同(Louault et al., 2005; 陈慧颖等, 2014; 胡静等, 2016).因此, 大针茅与羊草面对实际放牧胁迫时更倾向于机械防御还是化学防御仍需进一步探讨. ...
... 植物在生长过程中所需的资源, 比如能量、空间、水分及养分通常都是有限的(Garland Jr, 2014).随着放牧强度的增强, 植物所能获取的资源会更加有限(Zhou et al., 2017).在面对啃食胁迫时, 植物会把原本用于生长繁殖的部分资源用来提高自身的抗性, 而植物的防御又是一个对碳氮资源消耗较大的过程, 所以, 植物需要权衡生长和防御的利弊, 以保证自己能在有限的资源下健康地生长繁殖(Srinivasa, 2018; Yang et al., 2018; Liu et al., 2019a).由于牛羊等脊椎动物没有饮食特异性, 因此它们可以诱导牧草产生更多种类而非单一性质的化学防御物质(Zhou et al., 2015; Isah, 2019).萜烯类及酚类等次级代谢产物是以碳为基础的物质, 而近年来引起了广泛关注的生物碱主要利用氮作为原料(Brandolini et al., 2013; Fuchs et al., 2017a).因此, 植物在产生各类化学防御物质时也应该存在某种权衡(Dong et al., 2017; Yang et al., 2018; Isah, 2019).在养分不足的环境条件下, 植物主要生成以碳为基础的物质; 而在养分充足或者低碳环境条件下, 主要生成以氮为基础的物质(Bryant et al., 1983; 阎秀峰等, 2007).羊草较大针茅有更高的全碳及全氮含量(陈慧颖等, 2014).因此, 二者对碳氮资源的权衡策略可能有所不同, 在不同放牧强度下, 大针茅与羊草的化学防御策略会发生怎样的变化仍需深入探讨. ...
Herbivory-induced maternal effects on growth and defense traits in the clonal species Alternanthera philoxeroides 2 2017
... 食草动物的啃食胁迫会诱导牧草产生耐牧和避牧两种防御机制(赵钢和崔泽仁, 1999).牧草一方面会调控体内代谢途径, 调配初级代谢产物以修复损伤, 通过补偿生长达到耐牧的目的(Appel et al., 2012; Zhou et al., 2015; Guo et al., 2018); 另一方面, 牧草个体趋向于矮小化、减缓生长速度, 伴随着体内木质素等物质含量的增多, 以提高自身机械防御能力, 并诱导化学防御机制的表达, 产生各类次级代谢产物以提高抗性(李西良等, 2015; Srinivasa, 2018).已有研究表明, 在受到严重放牧胁迫时, 牧草更倾向于避牧防御而非补偿生长(Dong et al., 2017; Liu et al., 2019a).但目前很多与牧草次级代谢相关的研究主要集中在模拟啃食实验, 不能完全反映真实的放牧胁迫(刘盟盟等, 2015; Zhou et al., 2015; Fuchs et al., 2017b).羊草是无性繁殖的根茎型禾草, 繁育过程需要大量的资源, 且由于对食草动物的适口性较高, 更易被采食; 大针茅为叶呈披针状的丛生型禾草, 较羊草有更高的木质素含量, 这些生理差异可能使得二者的防御策略有所不同(Louault et al., 2005; 陈慧颖等, 2014; 胡静等, 2016).因此, 大针茅与羊草面对实际放牧胁迫时更倾向于机械防御还是化学防御仍需进一步探讨. ...
... 植物在生长过程中所需的资源, 比如能量、空间、水分及养分通常都是有限的(Garland Jr, 2014).随着放牧强度的增强, 植物所能获取的资源会更加有限(Zhou et al., 2017).在面对啃食胁迫时, 植物会把原本用于生长繁殖的部分资源用来提高自身的抗性, 而植物的防御又是一个对碳氮资源消耗较大的过程, 所以, 植物需要权衡生长和防御的利弊, 以保证自己能在有限的资源下健康地生长繁殖(Srinivasa, 2018; Yang et al., 2018; Liu et al., 2019a).由于牛羊等脊椎动物没有饮食特异性, 因此它们可以诱导牧草产生更多种类而非单一性质的化学防御物质(Zhou et al., 2015; Isah, 2019).萜烯类及酚类等次级代谢产物是以碳为基础的物质, 而近年来引起了广泛关注的生物碱主要利用氮作为原料(Brandolini et al., 2013; Fuchs et al., 2017a).因此, 植物在产生各类化学防御物质时也应该存在某种权衡(Dong et al., 2017; Yang et al., 2018; Isah, 2019).在养分不足的环境条件下, 植物主要生成以碳为基础的物质; 而在养分充足或者低碳环境条件下, 主要生成以氮为基础的物质(Bryant et al., 1983; 阎秀峰等, 2007).羊草较大针茅有更高的全碳及全氮含量(陈慧颖等, 2014).因此, 二者对碳氮资源的权衡策略可能有所不同, 在不同放牧强度下, 大针茅与羊草的化学防御策略会发生怎样的变化仍需深入探讨. ...
Volatiles as inducers and suppressors of plant defense and immunity—Origins, specificity, perception and signaling 1 2018
... 植物在生长过程中所需的资源, 比如能量、空间、水分及养分通常都是有限的(Garland Jr, 2014).随着放牧强度的增强, 植物所能获取的资源会更加有限(Zhou et al., 2017).在面对啃食胁迫时, 植物会把原本用于生长繁殖的部分资源用来提高自身的抗性, 而植物的防御又是一个对碳氮资源消耗较大的过程, 所以, 植物需要权衡生长和防御的利弊, 以保证自己能在有限的资源下健康地生长繁殖(Srinivasa, 2018; Yang et al., 2018; Liu et al., 2019a).由于牛羊等脊椎动物没有饮食特异性, 因此它们可以诱导牧草产生更多种类而非单一性质的化学防御物质(Zhou et al., 2015; Isah, 2019).萜烯类及酚类等次级代谢产物是以碳为基础的物质, 而近年来引起了广泛关注的生物碱主要利用氮作为原料(Brandolini et al., 2013; Fuchs et al., 2017a).因此, 植物在产生各类化学防御物质时也应该存在某种权衡(Dong et al., 2017; Yang et al., 2018; Isah, 2019).在养分不足的环境条件下, 植物主要生成以碳为基础的物质; 而在养分充足或者低碳环境条件下, 主要生成以氮为基础的物质(Bryant et al., 1983; 阎秀峰等, 2007).羊草较大针茅有更高的全碳及全氮含量(陈慧颖等, 2014).因此, 二者对碳氮资源的权衡策略可能有所不同, 在不同放牧强度下, 大针茅与羊草的化学防御策略会发生怎样的变化仍需深入探讨. ...
... 由于大针茅与羊草的生理基础存在差异, 因而二者的碳氮资源利用策略有所不同.在6月生长初期, 大针茅面对轻度和极重度放牧胁迫, 叶片中的生物碱含量显著降低, 且与C:N呈较弱的正相关关系, 说明在生长初期大针茅虽然面对啃食胁迫提高了叶片内氮含量, 但并没有将氮用于合成生物碱增强防御能力, 而用于维持生长发育(Zhou et al., 2015; Guo et al., 2018).在8月生长末期, 中度及重度放牧可以促使大针茅叶片中的生物碱含量显著提高, 说明大针茅防御机制中的氮代谢途径可能在长时间胁迫条件下才会表达, 也可能是对季节变化的响应(徐瑞等, 2012; Fuchs et al., 2017a; Isah, 2019).在6月生长初期, 羊草面对中度放牧胁迫, 叶片中各种次级代谢产物含量均增多, 说明其化学防御机制中的碳氮代谢途径都得到了较充分的诱导和表达, 叶片中的氮被高效利用(翟占伟等, 2017).在8月生长末期, 羊草叶片中的生物碱含量基本都较6月时有所降低, 这可能是由于牧草将部分生物碱转移到了果穗及种子中, 以提高种子抗性; 也可能是由于牧草将氮用于合成氨基酸等其他物质以便繁殖(Zhou et al., 2015; Liu et al., 2019a). ...
Plant age and seasonal timing determine endophyte growth and alkaloid biosynthesis 1 2017b
... 食草动物的啃食胁迫会诱导牧草产生耐牧和避牧两种防御机制(赵钢和崔泽仁, 1999).牧草一方面会调控体内代谢途径, 调配初级代谢产物以修复损伤, 通过补偿生长达到耐牧的目的(Appel et al., 2012; Zhou et al., 2015; Guo et al., 2018); 另一方面, 牧草个体趋向于矮小化、减缓生长速度, 伴随着体内木质素等物质含量的增多, 以提高自身机械防御能力, 并诱导化学防御机制的表达, 产生各类次级代谢产物以提高抗性(李西良等, 2015; Srinivasa, 2018).已有研究表明, 在受到严重放牧胁迫时, 牧草更倾向于避牧防御而非补偿生长(Dong et al., 2017; Liu et al., 2019a).但目前很多与牧草次级代谢相关的研究主要集中在模拟啃食实验, 不能完全反映真实的放牧胁迫(刘盟盟等, 2015; Zhou et al., 2015; Fuchs et al., 2017b).羊草是无性繁殖的根茎型禾草, 繁育过程需要大量的资源, 且由于对食草动物的适口性较高, 更易被采食; 大针茅为叶呈披针状的丛生型禾草, 较羊草有更高的木质素含量, 这些生理差异可能使得二者的防御策略有所不同(Louault et al., 2005; 陈慧颖等, 2014; 胡静等, 2016).因此, 大针茅与羊草面对实际放牧胁迫时更倾向于机械防御还是化学防御仍需进一步探讨. ...
Trade-offs 1 2014
... 植物在生长过程中所需的资源, 比如能量、空间、水分及养分通常都是有限的(Garland Jr, 2014).随着放牧强度的增强, 植物所能获取的资源会更加有限(Zhou et al., 2017).在面对啃食胁迫时, 植物会把原本用于生长繁殖的部分资源用来提高自身的抗性, 而植物的防御又是一个对碳氮资源消耗较大的过程, 所以, 植物需要权衡生长和防御的利弊, 以保证自己能在有限的资源下健康地生长繁殖(Srinivasa, 2018; Yang et al., 2018; Liu et al., 2019a).由于牛羊等脊椎动物没有饮食特异性, 因此它们可以诱导牧草产生更多种类而非单一性质的化学防御物质(Zhou et al., 2015; Isah, 2019).萜烯类及酚类等次级代谢产物是以碳为基础的物质, 而近年来引起了广泛关注的生物碱主要利用氮作为原料(Brandolini et al., 2013; Fuchs et al., 2017a).因此, 植物在产生各类化学防御物质时也应该存在某种权衡(Dong et al., 2017; Yang et al., 2018; Isah, 2019).在养分不足的环境条件下, 植物主要生成以碳为基础的物质; 而在养分充足或者低碳环境条件下, 主要生成以氮为基础的物质(Bryant et al., 1983; 阎秀峰等, 2007).羊草较大针茅有更高的全碳及全氮含量(陈慧颖等, 2014).因此, 二者对碳氮资源的权衡策略可能有所不同, 在不同放牧强度下, 大针茅与羊草的化学防御策略会发生怎样的变化仍需深入探讨. ...
Resolution of growth- defense conflict: mechanistic insights from jasmonate signaling 4 2018
... 植物在生长过程中所需的资源, 比如能量、空间、水分及养分通常都是有限的(Garland Jr, 2014).随着放牧强度的增强, 植物所能获取的资源会更加有限(Zhou et al., 2017).在面对啃食胁迫时, 植物会把原本用于生长繁殖的部分资源用来提高自身的抗性, 而植物的防御又是一个对碳氮资源消耗较大的过程, 所以, 植物需要权衡生长和防御的利弊, 以保证自己能在有限的资源下健康地生长繁殖(Srinivasa, 2018; Yang et al., 2018; Liu et al., 2019a).由于牛羊等脊椎动物没有饮食特异性, 因此它们可以诱导牧草产生更多种类而非单一性质的化学防御物质(Zhou et al., 2015; Isah, 2019).萜烯类及酚类等次级代谢产物是以碳为基础的物质, 而近年来引起了广泛关注的生物碱主要利用氮作为原料(Brandolini et al., 2013; Fuchs et al., 2017a).因此, 植物在产生各类化学防御物质时也应该存在某种权衡(Dong et al., 2017; Yang et al., 2018; Isah, 2019).在养分不足的环境条件下, 植物主要生成以碳为基础的物质; 而在养分充足或者低碳环境条件下, 主要生成以氮为基础的物质(Bryant et al., 1983; 阎秀峰等, 2007).羊草较大针茅有更高的全碳及全氮含量(陈慧颖等, 2014).因此, 二者对碳氮资源的权衡策略可能有所不同, 在不同放牧强度下, 大针茅与羊草的化学防御策略会发生怎样的变化仍需深入探讨. ...
... 由于大针茅与羊草的生理基础存在差异, 因而二者的碳氮资源利用策略有所不同.在6月生长初期, 大针茅面对轻度和极重度放牧胁迫, 叶片中的生物碱含量显著降低, 且与C:N呈较弱的正相关关系, 说明在生长初期大针茅虽然面对啃食胁迫提高了叶片内氮含量, 但并没有将氮用于合成生物碱增强防御能力, 而用于维持生长发育(Zhou et al., 2015; Guo et al., 2018).在8月生长末期, 中度及重度放牧可以促使大针茅叶片中的生物碱含量显著提高, 说明大针茅防御机制中的氮代谢途径可能在长时间胁迫条件下才会表达, 也可能是对季节变化的响应(徐瑞等, 2012; Fuchs et al., 2017a; Isah, 2019).在6月生长初期, 羊草面对中度放牧胁迫, 叶片中各种次级代谢产物含量均增多, 说明其化学防御机制中的碳氮代谢途径都得到了较充分的诱导和表达, 叶片中的氮被高效利用(翟占伟等, 2017).在8月生长末期, 羊草叶片中的生物碱含量基本都较6月时有所降低, 这可能是由于牧草将部分生物碱转移到了果穗及种子中, 以提高种子抗性; 也可能是由于牧草将氮用于合成氨基酸等其他物质以便繁殖(Zhou et al., 2015; Liu et al., 2019a). ...
Evaluation of Total condensed tannin content and anthelmintic activities of organic extracts of four Bangladeshi plants on Tubifex tubifex worm using in vitro method 1 2015
... 植物在生长过程中所需的资源, 比如能量、空间、水分及养分通常都是有限的(Garland Jr, 2014).随着放牧强度的增强, 植物所能获取的资源会更加有限(Zhou et al., 2017).在面对啃食胁迫时, 植物会把原本用于生长繁殖的部分资源用来提高自身的抗性, 而植物的防御又是一个对碳氮资源消耗较大的过程, 所以, 植物需要权衡生长和防御的利弊, 以保证自己能在有限的资源下健康地生长繁殖(Srinivasa, 2018; Yang et al., 2018; Liu et al., 2019a).由于牛羊等脊椎动物没有饮食特异性, 因此它们可以诱导牧草产生更多种类而非单一性质的化学防御物质(Zhou et al., 2015; Isah, 2019).萜烯类及酚类等次级代谢产物是以碳为基础的物质, 而近年来引起了广泛关注的生物碱主要利用氮作为原料(Brandolini et al., 2013; Fuchs et al., 2017a).因此, 植物在产生各类化学防御物质时也应该存在某种权衡(Dong et al., 2017; Yang et al., 2018; Isah, 2019).在养分不足的环境条件下, 植物主要生成以碳为基础的物质; 而在养分充足或者低碳环境条件下, 主要生成以氮为基础的物质(Bryant et al., 1983; 阎秀峰等, 2007).羊草较大针茅有更高的全碳及全氮含量(陈慧颖等, 2014).因此, 二者对碳氮资源的权衡策略可能有所不同, 在不同放牧强度下, 大针茅与羊草的化学防御策略会发生怎样的变化仍需深入探讨. ...
... 牧草在生长过程中, 尤其在生长初期, 需要与外界生物环境竞争有限的资源, 次级代谢过程比初级代谢对碳氮资源的消耗更大, 因此, 牧草面对食草动物的啃食胁迫时需要权衡利弊, 在生长与防御间做出选择(Srinivasa, 2018; Liu et al., 2019a).在6月生长初期, 大针茅和羊草的含碳次级代谢产物含量均在轻度放牧下最低, 这可能是由于生长竞争压力大于啃食胁迫, 所以化学防御机制并没有得到充分的诱导及表达, 更多的资源可能用于补偿生长, 提高了耐牧性(Appel et al., 2012; Zhou et al., 2015; Guo et al., 2018).在8月生长末期, 羊草在重度放牧胁迫下具有较低的C:N, 把更多的碳资源用于次级代谢产物的合成.说明胁迫时间的延长、胁迫强度的增加, 到生长竞争压力不及放牧胁迫时, 化学防御机制才被逐步诱导激活.这一结果与Herms提出的生长-分化平衡假说(growth-differentiation balance hypothesis)相一致(Herms & Mattson, 1992). ...
... 由于大针茅与羊草的生理基础存在差异, 因而二者的碳氮资源利用策略有所不同.在6月生长初期, 大针茅面对轻度和极重度放牧胁迫, 叶片中的生物碱含量显著降低, 且与C:N呈较弱的正相关关系, 说明在生长初期大针茅虽然面对啃食胁迫提高了叶片内氮含量, 但并没有将氮用于合成生物碱增强防御能力, 而用于维持生长发育(Zhou et al., 2015; Guo et al., 2018).在8月生长末期, 中度及重度放牧可以促使大针茅叶片中的生物碱含量显著提高, 说明大针茅防御机制中的氮代谢途径可能在长时间胁迫条件下才会表达, 也可能是对季节变化的响应(徐瑞等, 2012; Fuchs et al., 2017a; Isah, 2019).在6月生长初期, 羊草面对中度放牧胁迫, 叶片中各种次级代谢产物含量均增多, 说明其化学防御机制中的碳氮代谢途径都得到了较充分的诱导和表达, 叶片中的氮被高效利用(翟占伟等, 2017).在8月生长末期, 羊草叶片中的生物碱含量基本都较6月时有所降低, 这可能是由于牧草将部分生物碱转移到了果穗及种子中, 以提高种子抗性; 也可能是由于牧草将氮用于合成氨基酸等其他物质以便繁殖(Zhou et al., 2015; Liu et al., 2019a). ...
机械损伤对冷蒿叶片次生代谢产物的影响 1 2015
... 食草动物的啃食胁迫会诱导牧草产生耐牧和避牧两种防御机制(赵钢和崔泽仁, 1999).牧草一方面会调控体内代谢途径, 调配初级代谢产物以修复损伤, 通过补偿生长达到耐牧的目的(Appel et al., 2012; Zhou et al., 2015; Guo et al., 2018); 另一方面, 牧草个体趋向于矮小化、减缓生长速度, 伴随着体内木质素等物质含量的增多, 以提高自身机械防御能力, 并诱导化学防御机制的表达, 产生各类次级代谢产物以提高抗性(李西良等, 2015; Srinivasa, 2018).已有研究表明, 在受到严重放牧胁迫时, 牧草更倾向于避牧防御而非补偿生长(Dong et al., 2017; Liu et al., 2019a).但目前很多与牧草次级代谢相关的研究主要集中在模拟啃食实验, 不能完全反映真实的放牧胁迫(刘盟盟等, 2015; Zhou et al., 2015; Fuchs et al., 2017b).羊草是无性繁殖的根茎型禾草, 繁育过程需要大量的资源, 且由于对食草动物的适口性较高, 更易被采食; 大针茅为叶呈披针状的丛生型禾草, 较羊草有更高的木质素含量, 这些生理差异可能使得二者的防御策略有所不同(Louault et al., 2005; 陈慧颖等, 2014; 胡静等, 2016).因此, 大针茅与羊草面对实际放牧胁迫时更倾向于机械防御还是化学防御仍需进一步探讨. ...
机械损伤对冷蒿叶片次生代谢产物的影响 1 2015
... 食草动物的啃食胁迫会诱导牧草产生耐牧和避牧两种防御机制(赵钢和崔泽仁, 1999).牧草一方面会调控体内代谢途径, 调配初级代谢产物以修复损伤, 通过补偿生长达到耐牧的目的(Appel et al., 2012; Zhou et al., 2015; Guo et al., 2018); 另一方面, 牧草个体趋向于矮小化、减缓生长速度, 伴随着体内木质素等物质含量的增多, 以提高自身机械防御能力, 并诱导化学防御机制的表达, 产生各类次级代谢产物以提高抗性(李西良等, 2015; Srinivasa, 2018).已有研究表明, 在受到严重放牧胁迫时, 牧草更倾向于避牧防御而非补偿生长(Dong et al., 2017; Liu et al., 2019a).但目前很多与牧草次级代谢相关的研究主要集中在模拟啃食实验, 不能完全反映真实的放牧胁迫(刘盟盟等, 2015; Zhou et al., 2015; Fuchs et al., 2017b).羊草是无性繁殖的根茎型禾草, 繁育过程需要大量的资源, 且由于对食草动物的适口性较高, 更易被采食; 大针茅为叶呈披针状的丛生型禾草, 较羊草有更高的木质素含量, 这些生理差异可能使得二者的防御策略有所不同(Louault et al., 2005; 陈慧颖等, 2014; 胡静等, 2016).因此, 大针茅与羊草面对实际放牧胁迫时更倾向于机械防御还是化学防御仍需进一步探讨. ...
Grassland dynamics in responses to climate variation and human activities in China from 2000 to 2013 1 2019b
Unravelling the distribution of secondary metabolites in Olea europaea L.: exhaustive characterization of eight olive-tree derived matrices by complementary platforms (LC-ESI/APCI-MS and GC-APCI-MS) 1 2018
Breaching plant defence theories: growth rates of plants directly impact the evolution of consumption rates of herbivorous insects 3 2018
... 食草动物的啃食胁迫会诱导牧草产生耐牧和避牧两种防御机制(赵钢和崔泽仁, 1999).牧草一方面会调控体内代谢途径, 调配初级代谢产物以修复损伤, 通过补偿生长达到耐牧的目的(Appel et al., 2012; Zhou et al., 2015; Guo et al., 2018); 另一方面, 牧草个体趋向于矮小化、减缓生长速度, 伴随着体内木质素等物质含量的增多, 以提高自身机械防御能力, 并诱导化学防御机制的表达, 产生各类次级代谢产物以提高抗性(李西良等, 2015; Srinivasa, 2018).已有研究表明, 在受到严重放牧胁迫时, 牧草更倾向于避牧防御而非补偿生长(Dong et al., 2017; Liu et al., 2019a).但目前很多与牧草次级代谢相关的研究主要集中在模拟啃食实验, 不能完全反映真实的放牧胁迫(刘盟盟等, 2015; Zhou et al., 2015; Fuchs et al., 2017b).羊草是无性繁殖的根茎型禾草, 繁育过程需要大量的资源, 且由于对食草动物的适口性较高, 更易被采食; 大针茅为叶呈披针状的丛生型禾草, 较羊草有更高的木质素含量, 这些生理差异可能使得二者的防御策略有所不同(Louault et al., 2005; 陈慧颖等, 2014; 胡静等, 2016).因此, 大针茅与羊草面对实际放牧胁迫时更倾向于机械防御还是化学防御仍需进一步探讨. ...
... 植物在生长过程中所需的资源, 比如能量、空间、水分及养分通常都是有限的(Garland Jr, 2014).随着放牧强度的增强, 植物所能获取的资源会更加有限(Zhou et al., 2017).在面对啃食胁迫时, 植物会把原本用于生长繁殖的部分资源用来提高自身的抗性, 而植物的防御又是一个对碳氮资源消耗较大的过程, 所以, 植物需要权衡生长和防御的利弊, 以保证自己能在有限的资源下健康地生长繁殖(Srinivasa, 2018; Yang et al., 2018; Liu et al., 2019a).由于牛羊等脊椎动物没有饮食特异性, 因此它们可以诱导牧草产生更多种类而非单一性质的化学防御物质(Zhou et al., 2015; Isah, 2019).萜烯类及酚类等次级代谢产物是以碳为基础的物质, 而近年来引起了广泛关注的生物碱主要利用氮作为原料(Brandolini et al., 2013; Fuchs et al., 2017a).因此, 植物在产生各类化学防御物质时也应该存在某种权衡(Dong et al., 2017; Yang et al., 2018; Isah, 2019).在养分不足的环境条件下, 植物主要生成以碳为基础的物质; 而在养分充足或者低碳环境条件下, 主要生成以氮为基础的物质(Bryant et al., 1983; 阎秀峰等, 2007).羊草较大针茅有更高的全碳及全氮含量(陈慧颖等, 2014).因此, 二者对碳氮资源的权衡策略可能有所不同, 在不同放牧强度下, 大针茅与羊草的化学防御策略会发生怎样的变化仍需深入探讨. ...
... 牧草在生长过程中, 尤其在生长初期, 需要与外界生物环境竞争有限的资源, 次级代谢过程比初级代谢对碳氮资源的消耗更大, 因此, 牧草面对食草动物的啃食胁迫时需要权衡利弊, 在生长与防御间做出选择(Srinivasa, 2018; Liu et al., 2019a).在6月生长初期, 大针茅和羊草的含碳次级代谢产物含量均在轻度放牧下最低, 这可能是由于生长竞争压力大于啃食胁迫, 所以化学防御机制并没有得到充分的诱导及表达, 更多的资源可能用于补偿生长, 提高了耐牧性(Appel et al., 2012; Zhou et al., 2015; Guo et al., 2018).在8月生长末期, 羊草在重度放牧胁迫下具有较低的C:N, 把更多的碳资源用于次级代谢产物的合成.说明胁迫时间的延长、胁迫强度的增加, 到生长竞争压力不及放牧胁迫时, 化学防御机制才被逐步诱导激活.这一结果与Herms提出的生长-分化平衡假说(growth-differentiation balance hypothesis)相一致(Herms & Mattson, 1992). ...
Defoliation synchronizes aboveground growth of co-occurring C4 grass species 1 2008
... 植物在生长过程中所需的资源, 比如能量、空间、水分及养分通常都是有限的(Garland Jr, 2014).随着放牧强度的增强, 植物所能获取的资源会更加有限(Zhou et al., 2017).在面对啃食胁迫时, 植物会把原本用于生长繁殖的部分资源用来提高自身的抗性, 而植物的防御又是一个对碳氮资源消耗较大的过程, 所以, 植物需要权衡生长和防御的利弊, 以保证自己能在有限的资源下健康地生长繁殖(Srinivasa, 2018; Yang et al., 2018; Liu et al., 2019a).由于牛羊等脊椎动物没有饮食特异性, 因此它们可以诱导牧草产生更多种类而非单一性质的化学防御物质(Zhou et al., 2015; Isah, 2019).萜烯类及酚类等次级代谢产物是以碳为基础的物质, 而近年来引起了广泛关注的生物碱主要利用氮作为原料(Brandolini et al., 2013; Fuchs et al., 2017a).因此, 植物在产生各类化学防御物质时也应该存在某种权衡(Dong et al., 2017; Yang et al., 2018; Isah, 2019).在养分不足的环境条件下, 植物主要生成以碳为基础的物质; 而在养分充足或者低碳环境条件下, 主要生成以氮为基础的物质(Bryant et al., 1983; 阎秀峰等, 2007).羊草较大针茅有更高的全碳及全氮含量(陈慧颖等, 2014).因此, 二者对碳氮资源的权衡策略可能有所不同, 在不同放牧强度下, 大针茅与羊草的化学防御策略会发生怎样的变化仍需深入探讨. ...
... 植物在生长过程中所需的资源, 比如能量、空间、水分及养分通常都是有限的(Garland Jr, 2014).随着放牧强度的增强, 植物所能获取的资源会更加有限(Zhou et al., 2017).在面对啃食胁迫时, 植物会把原本用于生长繁殖的部分资源用来提高自身的抗性, 而植物的防御又是一个对碳氮资源消耗较大的过程, 所以, 植物需要权衡生长和防御的利弊, 以保证自己能在有限的资源下健康地生长繁殖(Srinivasa, 2018; Yang et al., 2018; Liu et al., 2019a).由于牛羊等脊椎动物没有饮食特异性, 因此它们可以诱导牧草产生更多种类而非单一性质的化学防御物质(Zhou et al., 2015; Isah, 2019).萜烯类及酚类等次级代谢产物是以碳为基础的物质, 而近年来引起了广泛关注的生物碱主要利用氮作为原料(Brandolini et al., 2013; Fuchs et al., 2017a).因此, 植物在产生各类化学防御物质时也应该存在某种权衡(Dong et al., 2017; Yang et al., 2018; Isah, 2019).在养分不足的环境条件下, 植物主要生成以碳为基础的物质; 而在养分充足或者低碳环境条件下, 主要生成以氮为基础的物质(Bryant et al., 1983; 阎秀峰等, 2007).羊草较大针茅有更高的全碳及全氮含量(陈慧颖等, 2014).因此, 二者对碳氮资源的权衡策略可能有所不同, 在不同放牧强度下, 大针茅与羊草的化学防御策略会发生怎样的变化仍需深入探讨. ...
Response of plant secondary metabolites to environmental factors 2 2018
... 植物在生长过程中所需的资源, 比如能量、空间、水分及养分通常都是有限的(Garland Jr, 2014).随着放牧强度的增强, 植物所能获取的资源会更加有限(Zhou et al., 2017).在面对啃食胁迫时, 植物会把原本用于生长繁殖的部分资源用来提高自身的抗性, 而植物的防御又是一个对碳氮资源消耗较大的过程, 所以, 植物需要权衡生长和防御的利弊, 以保证自己能在有限的资源下健康地生长繁殖(Srinivasa, 2018; Yang et al., 2018; Liu et al., 2019a).由于牛羊等脊椎动物没有饮食特异性, 因此它们可以诱导牧草产生更多种类而非单一性质的化学防御物质(Zhou et al., 2015; Isah, 2019).萜烯类及酚类等次级代谢产物是以碳为基础的物质, 而近年来引起了广泛关注的生物碱主要利用氮作为原料(Brandolini et al., 2013; Fuchs et al., 2017a).因此, 植物在产生各类化学防御物质时也应该存在某种权衡(Dong et al., 2017; Yang et al., 2018; Isah, 2019).在养分不足的环境条件下, 植物主要生成以碳为基础的物质; 而在养分充足或者低碳环境条件下, 主要生成以氮为基础的物质(Bryant et al., 1983; 阎秀峰等, 2007).羊草较大针茅有更高的全碳及全氮含量(陈慧颖等, 2014).因此, 二者对碳氮资源的权衡策略可能有所不同, 在不同放牧强度下, 大针茅与羊草的化学防御策略会发生怎样的变化仍需深入探讨. ...
... ; Yang et al., 2018; Isah, 2019).在养分不足的环境条件下, 植物主要生成以碳为基础的物质; 而在养分充足或者低碳环境条件下, 主要生成以氮为基础的物质(Bryant et al., 1983; 阎秀峰等, 2007).羊草较大针茅有更高的全碳及全氮含量(陈慧颖等, 2014).因此, 二者对碳氮资源的权衡策略可能有所不同, 在不同放牧强度下, 大针茅与羊草的化学防御策略会发生怎样的变化仍需深入探讨. ...
... 植物在生长过程中所需的资源, 比如能量、空间、水分及养分通常都是有限的(Garland Jr, 2014).随着放牧强度的增强, 植物所能获取的资源会更加有限(Zhou et al., 2017).在面对啃食胁迫时, 植物会把原本用于生长繁殖的部分资源用来提高自身的抗性, 而植物的防御又是一个对碳氮资源消耗较大的过程, 所以, 植物需要权衡生长和防御的利弊, 以保证自己能在有限的资源下健康地生长繁殖(Srinivasa, 2018; Yang et al., 2018; Liu et al., 2019a).由于牛羊等脊椎动物没有饮食特异性, 因此它们可以诱导牧草产生更多种类而非单一性质的化学防御物质(Zhou et al., 2015; Isah, 2019).萜烯类及酚类等次级代谢产物是以碳为基础的物质, 而近年来引起了广泛关注的生物碱主要利用氮作为原料(Brandolini et al., 2013; Fuchs et al., 2017a).因此, 植物在产生各类化学防御物质时也应该存在某种权衡(Dong et al., 2017; Yang et al., 2018; Isah, 2019).在养分不足的环境条件下, 植物主要生成以碳为基础的物质; 而在养分充足或者低碳环境条件下, 主要生成以氮为基础的物质(Bryant et al., 1983; 阎秀峰等, 2007).羊草较大针茅有更高的全碳及全氮含量(陈慧颖等, 2014).因此, 二者对碳氮资源的权衡策略可能有所不同, 在不同放牧强度下, 大针茅与羊草的化学防御策略会发生怎样的变化仍需深入探讨. ...
Alteration of plant primary metabolism in response to insect herbivory 7 2015
... 食草动物的啃食胁迫会诱导牧草产生耐牧和避牧两种防御机制(赵钢和崔泽仁, 1999).牧草一方面会调控体内代谢途径, 调配初级代谢产物以修复损伤, 通过补偿生长达到耐牧的目的(Appel et al., 2012; Zhou et al., 2015; Guo et al., 2018); 另一方面, 牧草个体趋向于矮小化、减缓生长速度, 伴随着体内木质素等物质含量的增多, 以提高自身机械防御能力, 并诱导化学防御机制的表达, 产生各类次级代谢产物以提高抗性(李西良等, 2015; Srinivasa, 2018).已有研究表明, 在受到严重放牧胁迫时, 牧草更倾向于避牧防御而非补偿生长(Dong et al., 2017; Liu et al., 2019a).但目前很多与牧草次级代谢相关的研究主要集中在模拟啃食实验, 不能完全反映真实的放牧胁迫(刘盟盟等, 2015; Zhou et al., 2015; Fuchs et al., 2017b).羊草是无性繁殖的根茎型禾草, 繁育过程需要大量的资源, 且由于对食草动物的适口性较高, 更易被采食; 大针茅为叶呈披针状的丛生型禾草, 较羊草有更高的木质素含量, 这些生理差异可能使得二者的防御策略有所不同(Louault et al., 2005; 陈慧颖等, 2014; 胡静等, 2016).因此, 大针茅与羊草面对实际放牧胁迫时更倾向于机械防御还是化学防御仍需进一步探讨. ...
... ; Zhou et al., 2015; Fuchs et al., 2017b).羊草是无性繁殖的根茎型禾草, 繁育过程需要大量的资源, 且由于对食草动物的适口性较高, 更易被采食; 大针茅为叶呈披针状的丛生型禾草, 较羊草有更高的木质素含量, 这些生理差异可能使得二者的防御策略有所不同(Louault et al., 2005; 陈慧颖等, 2014; 胡静等, 2016).因此, 大针茅与羊草面对实际放牧胁迫时更倾向于机械防御还是化学防御仍需进一步探讨. ...
... 植物在生长过程中所需的资源, 比如能量、空间、水分及养分通常都是有限的(Garland Jr, 2014).随着放牧强度的增强, 植物所能获取的资源会更加有限(Zhou et al., 2017).在面对啃食胁迫时, 植物会把原本用于生长繁殖的部分资源用来提高自身的抗性, 而植物的防御又是一个对碳氮资源消耗较大的过程, 所以, 植物需要权衡生长和防御的利弊, 以保证自己能在有限的资源下健康地生长繁殖(Srinivasa, 2018; Yang et al., 2018; Liu et al., 2019a).由于牛羊等脊椎动物没有饮食特异性, 因此它们可以诱导牧草产生更多种类而非单一性质的化学防御物质(Zhou et al., 2015; Isah, 2019).萜烯类及酚类等次级代谢产物是以碳为基础的物质, 而近年来引起了广泛关注的生物碱主要利用氮作为原料(Brandolini et al., 2013; Fuchs et al., 2017a).因此, 植物在产生各类化学防御物质时也应该存在某种权衡(Dong et al., 2017; Yang et al., 2018; Isah, 2019).在养分不足的环境条件下, 植物主要生成以碳为基础的物质; 而在养分充足或者低碳环境条件下, 主要生成以氮为基础的物质(Bryant et al., 1983; 阎秀峰等, 2007).羊草较大针茅有更高的全碳及全氮含量(陈慧颖等, 2014).因此, 二者对碳氮资源的权衡策略可能有所不同, 在不同放牧强度下, 大针茅与羊草的化学防御策略会发生怎样的变化仍需深入探讨. ...
... 牧草在生长过程中, 尤其在生长初期, 需要与外界生物环境竞争有限的资源, 次级代谢过程比初级代谢对碳氮资源的消耗更大, 因此, 牧草面对食草动物的啃食胁迫时需要权衡利弊, 在生长与防御间做出选择(Srinivasa, 2018; Liu et al., 2019a).在6月生长初期, 大针茅和羊草的含碳次级代谢产物含量均在轻度放牧下最低, 这可能是由于生长竞争压力大于啃食胁迫, 所以化学防御机制并没有得到充分的诱导及表达, 更多的资源可能用于补偿生长, 提高了耐牧性(Appel et al., 2012; Zhou et al., 2015; Guo et al., 2018).在8月生长末期, 羊草在重度放牧胁迫下具有较低的C:N, 把更多的碳资源用于次级代谢产物的合成.说明胁迫时间的延长、胁迫强度的增加, 到生长竞争压力不及放牧胁迫时, 化学防御机制才被逐步诱导激活.这一结果与Herms提出的生长-分化平衡假说(growth-differentiation balance hypothesis)相一致(Herms & Mattson, 1992). ...
 ,
, ,1,*, 刘敏1, 侯向阳2, 丁勇2, 杨波1, 张子荷1, 王彪1, 朱趁趁1
,1,*, 刘敏1, 侯向阳2, 丁勇2, 杨波1, 张子荷1, 王彪1, 朱趁趁1
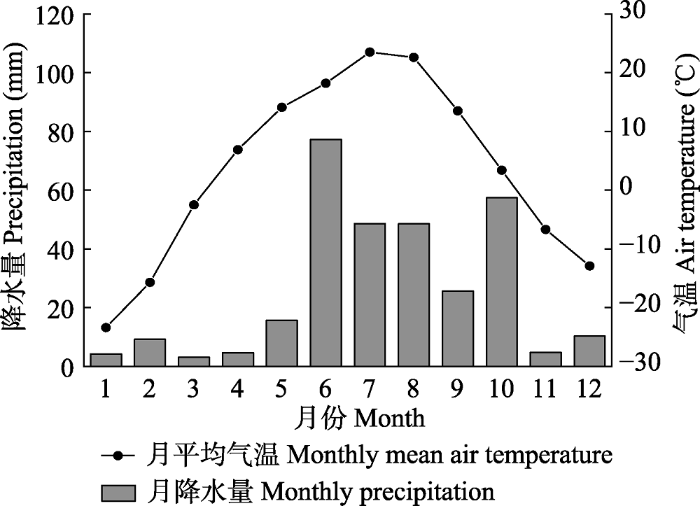 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT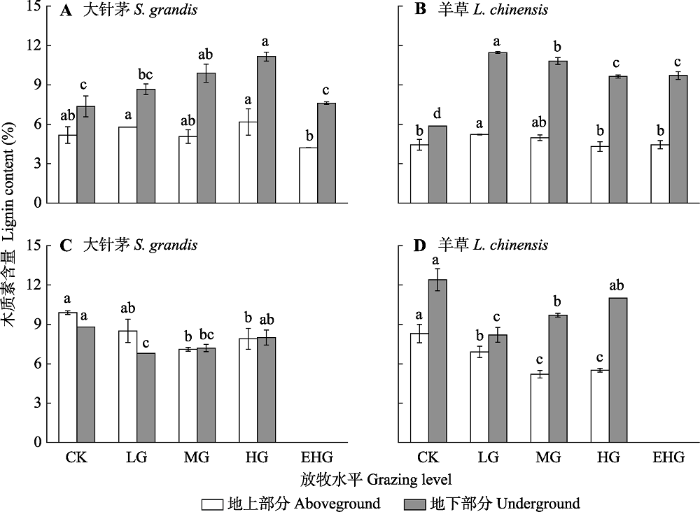 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT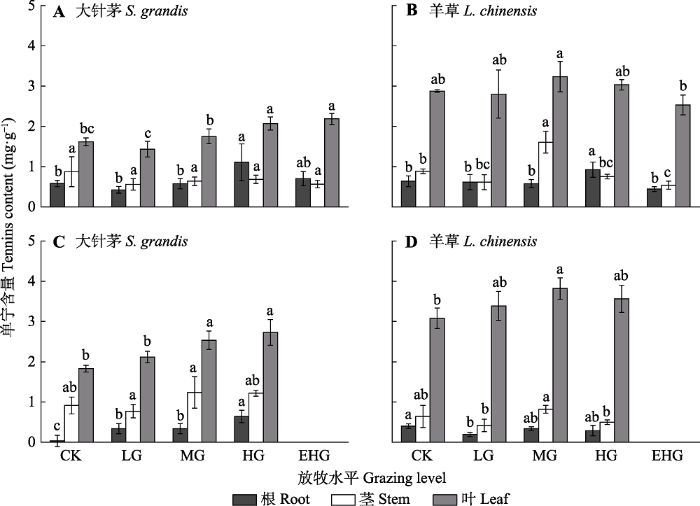 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT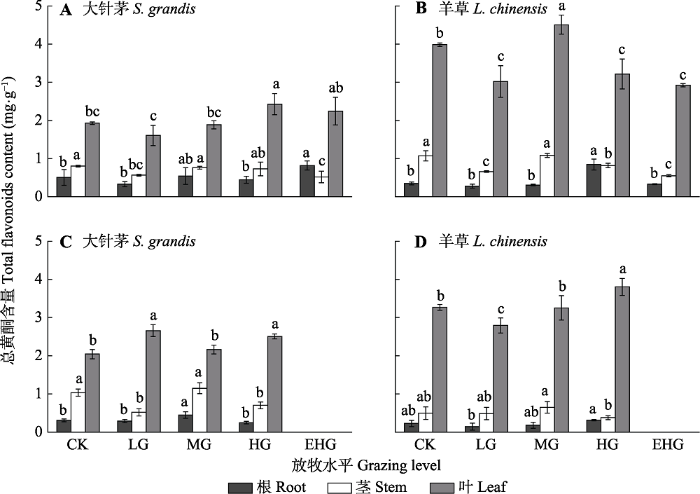 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT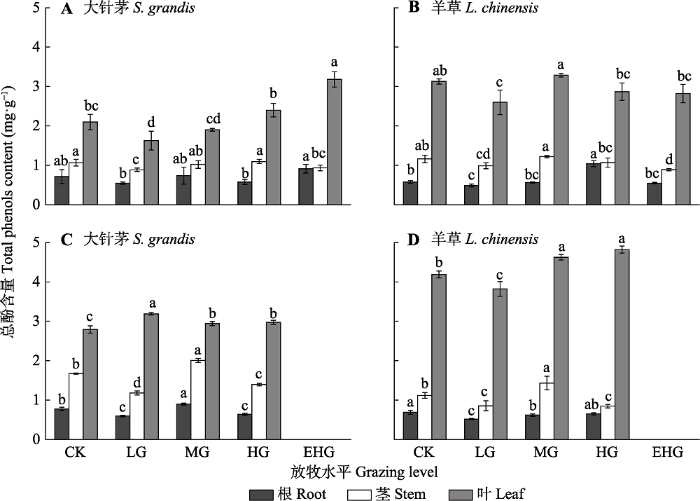 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT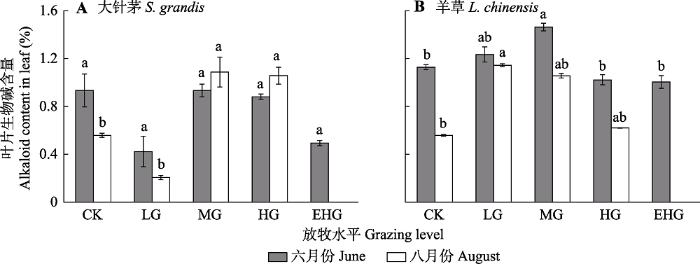 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT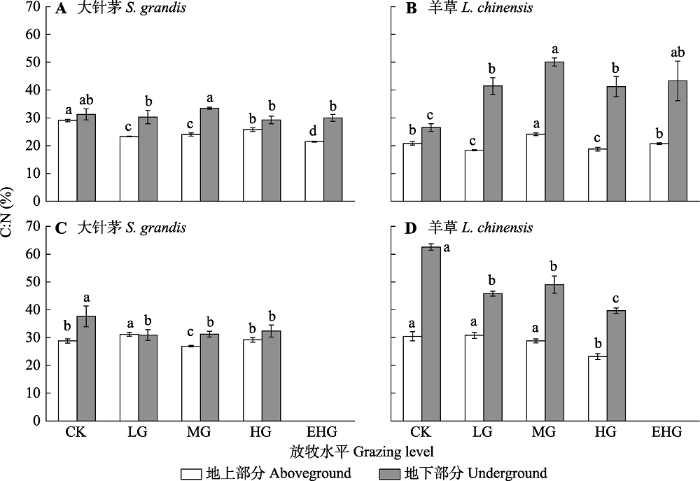 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}