 ,1,*, ����1,2, ����3, ������1, ������1
,1,*, ����1,2, ����3, ������1, ������1Extreme drought effects on nonstructural carbohydrates of dominant plant species in a meadow grassland
Lin SONG1,2, Wen-Tao LUO,1,*, Wang MA1,2, Peng HE3, Xiao-Sa LIANG1, Zheng-Wen WANG1ͨѶ����: *wentaoluo@iae.ac.cn
��ί: ������
���α༭: ����
| ��������: |
Corresponding authors: *wentaoluo@iae.ac.cn
Online:2020-06-20
| Fund supported: |

ժҪ
ֲ�������ò����ķǽṹ��̼ˮ������(NSCs)ˮƽ���Է�ӳֲ�����̬ϵͳ�Ի����仯����Ӧ�̶ȡ�������, ��ԭ���˸ɺ��¼��ķ���Ƶ�ʺͳ���ʱ��������������, ����̬ϵͳ�ṹ���ܲ�����ԶӰ�졣���о������ɹź��ױ����ݵ��ԭΪ�о�����, ͨ������4�����66%��������ˮ���Ŀ���ʵ����ģ�⼫�˸ɺ��¼�, ������ԭ6���������ֺ�ֲ�﹦��ȺNSCs����ֶԼ��˸ɺ�����Ӧ��������ơ������ʾ, ����ֲ������ѧ����������Լ�������̬�����ԵIJ���, ��ͬ���ֶԸɺ�в�ȵ���Ӧ�������Բ��졣������ݵ�ֲ��NSCs��ּ������ò��ԶԸɺ�в�ȵ���Ӧ��������������, �Ӷ��������������IJ�ͬ��Ӧ����6��ֲ���Ϊ�̲ݺͷǺ̲�����, ���ָɺ����������˺̲ݵĵ��ۺ���, ������������Ǻ���������Ӱ��; �෴, �ɺ����������˷Ǻ̲ݹ���Ⱥ�Ŀ������Ǻ���, ������ۺ���������Ӱ��, ������ͬ����Ⱥ��ȡ�˲�ͬ�ĸɺ�Ӧ�Բ��ԡ��̲�ѡ������ù̶����������д�����Ӧ�Ըɺ�в��, ���������Ըɺ���Ӧ������; ���Ǻ̲�ѡ�������Կ������ǵ���ʽֱ�ӹ�ֲ�����������Լ������ɺ�в��, ���������Ըɺ���Ӧ��Ϊ���С���һ���ֿ�ΪԤ����ȫ������仯�����²ݵ��ԭ��̬ϵͳ�ṹ�빦�ܶԼ��˸ɺ�����Ӧ�ṩ��ѧ�ο���
�ؼ��ʣ�
Abstract
Aims Plant nonstructural carbohydrates (NSCs) produced by photosynthesis can reflect the responses of plants and/or ecosystem to environmental changes. Climate models recently predicted an increase in the frequency and duration of extreme drought (ED) events that could profoundly impact ecosystem structure and functions. Yet, less is understood about the response patterns of different plant species and functional groups to extreme drought.
Methods Here we studied the effects of extreme drought on the NSCs of dominant species belonging to different functional groups in grasslands. To achieve ED, we experimentally reduced precipitation amounts by 66% during four consecutive growing seasons in a meadow steppe in Hulunbeier, North China. The NSCs of six plants grouped into two functional groups (i.e., grass and non-grass) were examined.
Important findings We found different species responded differently to drought, due to their differences in plant biological characteristics, photosynthetic characteristics and physiological ecology. This result implied that different species used different NSC-use strategies to cope with drought stress, resulting in different responses of their biomass to extreme drought. Extreme drought significantly increased the starch concentrations, and had no effect on the soluble sugar concentrations of the grass functional group. Contrarily, ED significantly increased the soluble sugar concentrations, and had no significant effects on the starch concentrations of the non-grass functional group. These results indicate that grasses moderately use and store photosynthate to cope with drought stress, hence their biomass was less sensitive. The biomass of the non-grasses was more sensitive perhaps because they maximally utilize soluble sugar for plant growth, defense and reproduction. Our results showed that different species or functional groups exhibit different NSC-use strategies to cope with drought stress. This study could provide scientific data for predicting future ecosystem responses to extreme drought.
Keywords��
PDF (975KB)Ԫ������ά�����������������EndNote|Ris|Bibtex�ղر���
����
����, ������, ����, ����, ������, ������. ���˸ɺ��Բݵ��ԭ����ֲ��ǽṹ��̼ˮ�������Ӱ��. ֲ����̬ѧ��, 2020, 44(6): 669-676. DOI: 10.17521/cjpe.2019.0331
SONG Lin, LUO Wen-Tao, MA Wang, HE Peng, LIANG Xiao-Sa, WANG Zheng-Wen.
ֲ��ͨ��������ý�CO2��ˮ�ϳ�̼ˮ������, ̼ˮ�����ﲻ�������ϸ���ṹ����Ҫ�ɷ�, �������³´�л��������Ҫ����Դ����, ����ά��ֲ������������������м�����Ҫ������(Hartmann & Trumbore, 2016)������̼ˮ��������ֲ�����ڵĴ�����ʽ, ���仮��Ϊ�ṹ��̼ˮ������(SCs)�ͷǽṹ��̼ˮ������(NSCs)��SCs��Ҫ������ά�غ�ľ���صȶ��ָ߷��ӻ�����, ��NSCs��Ҫ�ɿ�������(�����ǡ����ǡ�����)�͵�����ɡ��������Ǻ͵���֮����Ի���ת��, ��ֲ���³´�л���ֿ��澳в�ȹ����з�����Ҫ����(Loewe et al., 2000)��ֲ��NSCs����ֵ��о����ݵ�20��������, Leloir��Cardini (1953)�ڶ�С��(Triticum aestivum)��ѿ���о���, �״η��������Ǻϳ�ø, ��һ��Ҫ����ʹ****��ʼ��ע�ǽṹ��̼��л��ֲ��������������л�е���Ҫ���á�1958��, Hewitt�����ֲ��NSCs�IJⶨ����, ���ֹ��ȼƷ����÷����״�ʹֲ��NSCs���о����Զ�����������30����, ֲ��ѧ�Ҷ���NSCs���о���Ҫ��ũ����̬ϵͳΪ�о�����, �ص��עֲ����ε���NSCs��ʵ��ũ����ĸ߲�������(Raguse & Smith, 1966)��20����90�������, ������̬ѧ�Ŀ��ٷ�չ, �����о�ֲ��NSCs�Ŀռ�ֲ���������Ż�ֲ����������Ӱ��ֲ����ȫ��仯����Ӧ����Ӧ��Ϊ��Ҫ�ķ�չ����1999��, ��̬ѧ��K?rner��̽����ɽ���ߵ��γɹ����������, ����������ġ�̼��������˵���ü�˵��Ϊ, �ɺ������ֲ������̼ѭ��ƽ��, Ӱ��ֲ��������̬����, ���ոı�ֲ��Ⱥ��Ľṹ�빦�ܡ��ڼ��˸ɺ�в����, ֲ������ȱ�����֯�������������Լ�����������, ����һ������������CO2������Ϊ����, ����ֲ��Ϊ��ƥ�����(��������ֳ����)�Լ��μ���л(����������ĺϳ�)��̼���������ϵķǽṹ��̼����, תΪ������֯�ڴ洢��NSCs, ������̼��֧ʧ�������������2008��, McDowell�ȷ�����������������, ���ɺ�����ֲ�������Ļ��ƽ�������, ָ����չ�ɺ���ֲ��NSCs������̬�仯��Ӱ���о����������˽⼫������仯��ֲ��������Ӱ�켰�����û��ơ���������, ĿǰNSCs��̬�����о�����Ҫ������ľ��ֲ��, �����ڲݱ�ֲ��NSCs������о��д�����̽����(Dietze et al., 2014; �������, 2019)�����, ���ڲݱ�ֲ��NSCs���о���ΪԤ����ȫ������仯�����²ݵ��ԭ��̬ϵͳ�ṹ�빦�ܶԼ��˸ɺ�����Ӧ�ṩ��ѧ�ο���
��ԭ��̬ϵͳ���ҹ�����½����̬ϵͳ, ռ�ҹ�½�������41.7%, ���ҹ���Ҫ������ҵ�������غ���ɫ��̬����, �ڱ��Ϲ�����̬��ȫ����ʳ��ȫ����ȫ����̬ƽ���з�����Ҫ����(�ű��, 2014)����ԭ�ǶԽ�ˮ������Ϊ���е���̬ϵͳ����, ���˸ɺ��¼��ᵼ�²�ԭ��̬ϵͳˮ��ʧ��, �Ӿ���̬ϵͳ��ˮ��в�ȳ̶�, ����Ӱ���ԭֲ�������ͷ���(Smith, 2011; Cherwin & Knapp, 2012; ��������, 2015)��������, ��ȫ�������ů�ı�����, ���˸ɺ��¼�������Ƶ�ʺ�ǿ�Ⱦ���������(Jentsch et al., 2009), ����̬ϵͳ�ṹ���ܲ�������Ӱ�졣���ܼ��˸ɺ��¼�ͨ������, �����ֲ��Ⱥ�����̬ϵͳ��Ӱ����ܻᳬԽ���������ɺ�(Jentsch et al., 2004; �ű��, 2014)�����, 21��������, ֲ����ε�����֯��NSCs��ɽṹӦ�Լ��˸ɺ��¼�����Ӧ�ѳ�Ϊ��̬ѧ�Һ�ֲ��ѧ�ҹ�ע���ȵ㡣
���о������ɹź��ױ����ݵ��ԭ��������Ϊ�о�����, ͨ��������������ˮ�ĸɺ�ģ��ʵ��, ̽����ͬ������ֲ�﹦��ȺNSCs�Ըɺ�в�ȵ���Ӧ����, ּ��ΪԤ������仯�²�ԭ��̬ϵͳ�ṹ�����ṩ��ѧ���ݡ�
1 ���Ϻͷ���
1.1 �о����ſ�
�о��ص�λ�����ɹ����������ױ����ж�������о��ڵ��й���ѧԺ����Ӧ����̬�о����������ɭ�ֲ�ԭ���ɴ���̬ϵͳ�о�վ(50.18�� N, 119.38�� E), ����Ϊ�´���½�Լ�������, ��Ҫֲ������Ϊ�ݵ��ԭ, ��������Ϊ�����(Stipa baicalensis)�����(Leymus chinensis)��, ����������ҪΪ�ڸ���, ������>2 m, ����pHΪ7.0-8.0, �����ṹ��Ϊ�ſ�״����״������ϸɰ���õ�������650 m, ��ƽ������Ϊ-2 ��, �꽵ˮ��Ϊ336 mm, ��ˮ�ֲ�����, ��Ҫ������6-9�·�, ����ͬ��, �������档1.2 ʵ�����
���о������й���ѧԺ����Ӧ����̬�о����������վ�ļ��˸ɺ�ʵ��ƽ̨���С�2015��, ѡȡ��Ծ���(������ֲ����)�����е��ʹ����Եĵض�, ������ʵ��ƽ̨��������������, ��������ʵ�鴦��: �ɺ��Ͷ���, ÿ�������ظ�6��, ��12��С��, �ɺ���������������(5-8��) 66%�Ľ�ˮ����С�����Ϊ6 m �� 6 m, ����С�����1 m, ÿ��С������������Ϊ1 m�IJ���ָ����Է�ֹС����ˮ�ֲ���������С���ڲ���С������4 m �� 4 mΪ��������, �ܱ�����1 m������, �Լ�С��ԵЧӦ�������ﶥ�����������θּܵĹ��νṹ, �������ؽ�ˮ���ų��ͼ��ٷ���ƻ�����������8��4 m �� 1.5 m�ĸ�����������ƴ�Ӷ���, ��ֱͶӰ�����С�������ͬ�����������ж�����������ϩ�����ϰ�, �����ϰ���и����ԡ��ͷ��ƶ�ָ��, ��UV��, �ܹ���������������ˮ��, �ּ������ή�����Ч����(��ҪΪɢ���)(Graefe & Sandmann, 2015)��������������üܸ����, ������ߵ�����2.5 m, ���������β�˾����60 cm, �����ڱ�������������ͨ, ��������ЧӦ(Luo et al., 2018, 2019)����������ÿ��5��1�հ�װ, 8��31�ղ��, ͨ���趨���������ϰ���֮�����ʵ�ּ���66%�Ľ�ˮ��, ����Ϊ���������������ȫ����Ȼ���ꡣ1.3 ��Ʒ�ɼ�
2018��8��, ��ÿ��С���ڵĵ����������ɼ�����, �������һ��1 m �� 1 m������, ������������Ϊ4��0.5 m �� 0.5 m��С����, ��б�Խǵ�����С�����н���Ⱥ��������ɺ��������ĵ���ȡ��, ��ʣ������0.5 m �� 0.5 m��С�����н��й�����״�Ĺ۲��ҶƬ�ɼ���ʵ���л�ȡ6������ֲ��, ������ҶҰ����(Thermopsis lanceolata)������(Artemisia frigida)�����(Carex duriuscula)��������о��(Cymbaria dahurica)����ݺ������, �������Ϊ��������Ⱥ: �̲�(��ݡ������)�ͷǺ̲�(����ҶҰ�����������ݡ�������о��)��ÿ��������ѡȡ10�������Բ��溦�Ľ���ֲ��, ȡÿ��ֲ�������������2-3ƬҶƬ, ����ʵ������105 ��ɱ��30 min, 65 ���������������á����պɺ�����ÿ��С�����ɼ�6������ֲ��, ����72��ֲ����Ʒ��1.4 �ǽṹ��̼ˮ������ɷֲⶨ
��ֲ����Ʒ���к�ɷ����, �ⶨֲ���п������Ǻ͵��۵ĺ���������ⶨ����Ϊ: ��ȡ0.1 gֲ����Ʒ��10 mL���Ĺ���, ��5 mL 80%�Ҵ���Һ����, ��80 ���ˮ��ˮԡ30 min, ��ȴ�����º���3 500 r��min-1������10 min, �ظ���ȡ3��, ת������Һ���ݵ�25 mL��ɫ�����ڿ������Ǻ����IJⶨ������������м���2 mL����ˮ�ڷ�ˮ�к���10 min, ��ȴ�����2 mL 9.2 mol��L-1��HClO4��Һ, ������Ⱥ��4 mL����ˮ����, ��4 000 r��min-1������10 min, ������Һת����50 mL����ƿ, ����������м���4.6 mol��L-1 HClO4��Һ, ������Ⱥ��5 mL����ˮ���Ⱥ�, ��4 000 r��min-1������10 min, ת������Һ, ����ˮϴ����2�κ�������Һ������50 mL, ���ڵ��ۺ����IJⶨ(Li et al., 2008)��ֲ���п������Ǻ͵��ۺ����IJⶨ���ô�ͳ����ͪ��ɫ��, ��ͨ������ֹ��ȼ�(620 nm����; 723S, �Ϻ��ӿ������DZ�����˾, �Ϻ�)�ⶨ, ���е��ۺ�����Ҫ����0.9��ת��ϵ����NSCs����Ϊ�������Ǻ͵��ۺ������ܺ͡�1.5 ���ݴ���
����R 3.6.1���������ݽ���ͳ�Ʒ��������û��ЧӦģ�ͷ������, ��ʵ�鴦��������(����Ⱥ)�����߽������ö�ֲ��NSCs���������ۺ������������Ǻ����Ϳ�������/���۵�Ӱ����м���; ����R��������ɺ�в���²�ͬ���ֺ���Ⱥ��NSCs����ֺ��������ڸɺ������µ���Ӧ��[ln(RR)]��95%����������(95% CI)��2 ����ͷ���
2.1 �ɺ���������������������ˮ���ͽ�ˮ����Ӱ��
1972-2018�꽵ˮ����̬�ֲ�ͼ����ʵ�����С���Ľ�ˮ������300 mm����, ������Ƶ�ʾ���68%����, �ɺ�������С���Ľ�ˮ������200 mm����, �ҷ���Ƶ�ʾ�С��15% (ͼ1A)���ɺ�����������������������������ˮ��(ͼ1B)��������ʵ��ƽ̨��ģ��ɺ�������ʼ��2015��, ��������������4�ꡣ�ۺϿ��Ǹɺ��ij���ʱ���ǿ��, �ɺ�����С���Ѵﵽ���˸ɺ�ˮƽ(��ʷ��������С��10%)��ͼ1
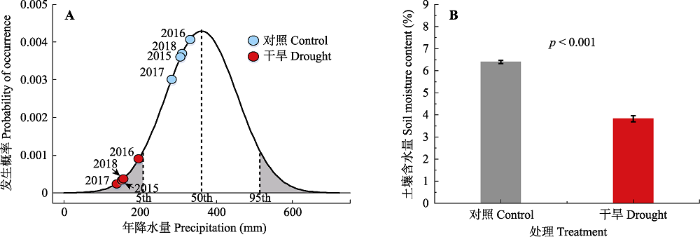 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ1�ɺ������Բݵ��ԭ��ˮ���������ʼ�������ˮ����Ӱ��(ƽ��ֵ�������)��
Fig. 1Effects of drought treatment on precipitation probability and soil moisture content in a meadow grassland (mean �� SE).
2.2 �ɺ�����������ֲ����������NSCs��Ӱ��
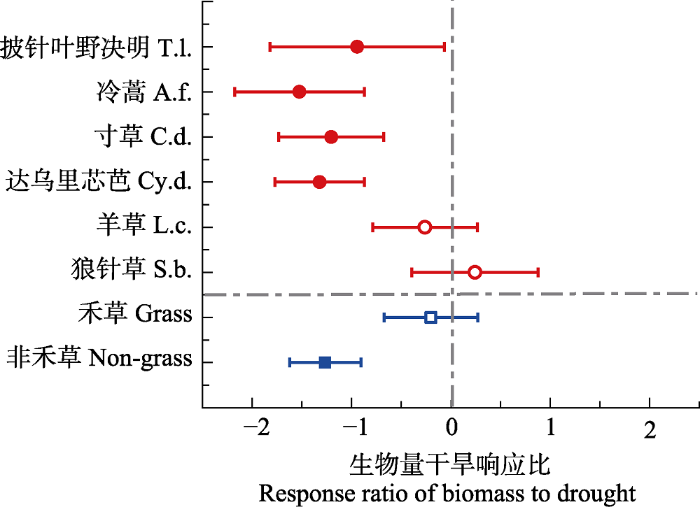
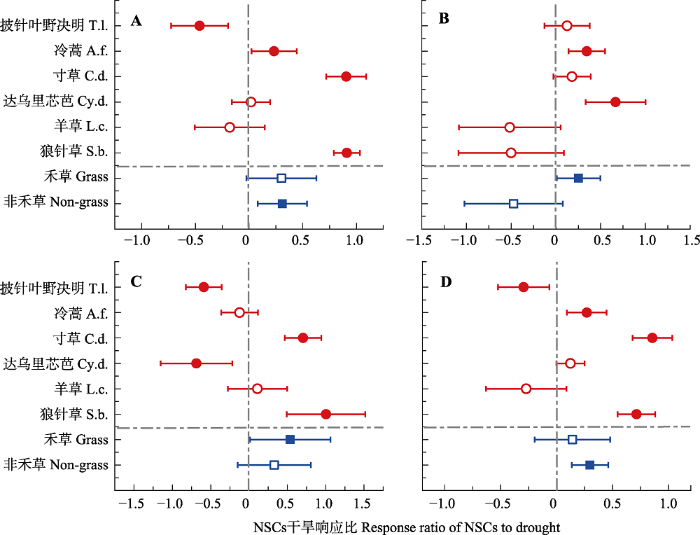
�ɺ����������ֶ��������������Ľ�������, ���������������Ըɺ���������Ӧһ��(��1)�����ֶԸɺ�����Ӧ���ƾ��������ԡ����˸ɺ����������˸����ֵĵ���������, ����������Ӧ������, ����ҶҰ��������ݺʹ�����о�Ŵ�֮, ��ݺ��������Ӧ����(��1; ͼ2)���ɺ����������ֶ�NSCs���������ۺ������������Ǻ����Լ���������/���۾����������Ľ�������(p < 0.05; ��1)����ͼ3��ʾ, ��ֲͬ��ҶƬNSCs����������ֶԸɺ���������Ӧ��ͬ���ɺ����������˴�ݺ�����ݵĿ�������/�����Լ��������ǵĺ�����NSCs�ĺ���, ���Ե��ۺ���������Ӱ�졣�ɺ�������������ҶҰ�����ʹ�����о�ŵĿ�������/����, ������������ҶҰ�����Ŀ������Ǻ�����NSCs����, ���Ե��ۺ���������Ӱ��; �ɺ������´�����о�ŵĿ������Ǻ�����NSCs����δ�����仯, ���ۺ���ȴ��������; �ɺ�δ�����������ݿ�������/���ۺ���ݵĿ������Ǻ����͵��ۺ����ı仯, ȴ�������������Ŀ������Ǻ��������ۺ�����NSCs������Table 1
��1
��1�ɺ�����������/����Ⱥ���佻�����öԲݵ��ԭ����ֲ����������ֲ��ǽṹ��̼ˮ������(NSCs)����ֺ��������ֵӰ��Ļ��ЧӦģ�ͽ��
Table 1
| ������ Biomass | �������� SS | ���� ST | ��������/���� SS/ST | NSCs | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | F | p | F | p | |
| ���� Species | ||||||||||
| �ɺ� Drought (D) | 2.195 | 0.146 | 33.645 | <0.001 | 0.216 | 0.645 | 12.937 | 0.001 | 27.713 | <0.001 |
| ���� Species (S) | 28.212 | <0.001 | 36.017 | <0.001 | 8.115 | <0.001 | 47.012 | <0.001 | 20.478 | <0.001 |
| �ɺ������� D �� S | 0.558 | 0.732 | 25.508 | <0.001 | 3.321 | 0.012 | 13.872 | <0.001 | 19.880 | <0.001 |
| ����Ⱥ Functional group | ||||||||||
| �ɺ� Drought (D) | 0.960 | 0.332 | 5.528 | 0.023 | 0.130 | 0.720 | 2.144 | 0.149 | 6.627 | 0.013 |
| ����Ⱥ Functional (F) | 26.055 | <0.001 | 1.510 | 0.225 | 0.459 | 0.501 | 0.412 | 0.524 | 2.185 | 0.145 |
| �ɺ�������Ⱥ D �� F | 0.154 | 0.696 | 0.014 | 0.905 | 6.236 | 0.016 | 0.326 | 0.571 | 0.280 | 0.599 |
�´��ڴ�|����CSV
ͼ2
 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ2�ݵ��ԭ���ֺ���Ⱥֲ���������Ըɺ���������Ӧ��
Grass, �̲ݹ���Ⱥ; Non-grass, �Ǻ̲ݹ���Ⱥ��������Ӧ��Ϊ�ɺ������С���зǽṹ��̼ˮ������(NSCs)�ı�ֵ, ˮƽ�������ʾ95%����������, ����R�����еġ�metaphor��������ó���Բ��������, �����������Ⱥ; ʵ�ı�ʾ��Ӧ����(p < 0.05), ���ı�ʾ��Ӧ��������
Fig. 2Response ratio of the biomass of six herbaceous species and different plant functional groups to drought in a meadow grassland. T.l., Thermopsis lanceolata; A.f., Artemisia frigida; C.d., Carex duriuscula; Cy.d., Cymbaria dahurica; L.c., Leymus chinensis; S.b., Stipa baicalensis; Grass, grass functional group; Non-grass, non-grass functional group.
The response ratio is biomass (drought)/biomass (control), the horizontal error bars represent the 95% confidence interval, which are calculated by ��metaphor�� in R. Solid circles indicate the significant responses of herbaceous plant species level nonstructural carbohydrates (NSCs) to drought (p < 0.05), while the hollow circles represent no significant response. Solid squares represent significant response of NSCs in different functional groups to drought, while hollow squares represent no significant response.
ͼ3
 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ3�ݵ��ԭ��ͬ���ֺ���Ⱥֲ��ҶƬ�������Ǻ���(A)�����ۺ���(B)����������/����(C)���ǽṹ��̼ˮ������(NSCs) ����(D)�Ըɺ�����Ӧ��
ˮƽ�������ʾ95%���������䡣Բ��������, �����������Ⱥ; ʵ�ı�ʾ��Ӧ����(p < 0.05), ���ı�ʾ��Ӧ��������
Fig. 3Response ratio of soluble sugar content (A), starch concentrations content (B), soluble sugar/starch ratios (C) and nonstructural carbohydrates (NSCs) content (D) in leaves of six herbaceous species and two plant functional groups to drought treatments. T.l., Thermopsis lanceolata; A.f., Artemisia frigida; C.d., Carex duriuscula; Cy.d., Cymbaria dahurica; L.c., Leymus chinensis; S.b., Stipa baicalensis; Grass, grass functional group; Non-grass, non-grass functional group.
The horizontal error bars represent the 95% confidence interval. Solid circles indicate the significant responses of herbaceous plant species level NSCs to drought (p < 0.05), while the hollow circles represent no significant response. Solid squares represent significant response of NSCs in different functional groups to drought, while hollow squares represent no significant response.
2.3 �ɺ������Բ�ֲͬ�﹦��Ⱥ��������NSCs������Ӱ��
�ɺ�������ֲ�﹦��Ⱥ����������Ӱ����������������, �����ɺ������Ը�����Ⱥ��������Ӱ��һ��(��1)���ɺ�������������������Ⱥ�ĵ���������, ���зǺ̲ݹ���Ⱥ����Ӧ�̶ȸ��ں̲�(ͼ2)�����˸ɺ���������Ⱥ�Ľ������öԵ��ۺ�����Ӱ������, ����NSCs��������������/���ۼ��������Ǻ�����Ӱ�첻����(��1)�����պɺ�������Ŀ������Ǻ�����NSCs�������������Բ���, �����ۺ����Ϳ�������/��������������(p > 0.05)����ͬ����ȺNSCs���������ۺ������������Ǻ�������������/ �����������Բ���(ͼ3)���ɺ����������˺̲ݹ���Ⱥ�ĵ��ۺ����Ϳ�������/����, ���Կ������Ǻ�����NSCs����������Ӱ��; �෴, �ɺ����������˷Ǻ̲ݹ���Ⱥ�Ŀ������Ǻ�����NSCs�ĺ���, ���Ե��ۺ����Ϳ�������/����������Ӱ��(ͼ3)��3 ����
3.1 �ɺ��Բ�ԭ��������ҶƬNSCs��Ӱ��
ֲ��Ըɺ�����Ӧ����ֲָ��������������������̬����(����̼���ò���)����Ӧˮ�ֻ����仯������, ��ֲ���ˮ��в�ȵ���Ӧ��ָ�ɺ�в�Ȼᵼ��ֲ��Ĵ��ռ��ռ�ֲ���ַ����仯��ֲ������NSCs���������ֲ�������ͷ�����һϵ�����������Ҫ��Դ����, ����ֲ�����ں����ı仯�����ܷ�ӳֲ������̼���պ�̼���ĵ������״��, ͬʱҲ�ܷ�ӳֲ��Ըɺ�в�ȵ���Ӧ(�������, 2002; ֣���յ�, 2014)����Լ��˸ɺ��¼�, ֲ��ͨ�������ڴ洢һ������NSCs���ṩ���������������ˮ��в��, ά�����������(֣���յ�, 2014)�����о�����, ���˸ɺ�в�ȶ�NSCs��Ӱ��������Ե�����������, ��ͬ���ֲ�ȡ��ͬ��̼���ò�������Ӧ������ˮ��в�ȡ����ڹ���ijһȺ������־��в�ͬ�������������̼�����ѧ����, ֲ���С����Դ���÷�ʽ����ˮ��в�ȵ���Ӧ�ȷ����������������(�Թ�µ�, 2003; McInerny & Etienne, 2012)������, ֲ��Ⱥ������ݺ�������״�����Ըɺ�����Ӧ��ͬ, �����״����ȶ�, ��������״����ˮ�ֵı仯���仯(Zhou et al., 2019)���ڳ��ڵ��ݻ�������, ֲ�����ڡ��ּ侺���ȹ�ϵ��������̬λ�ֻ�, ������̬λ�ֻ�ʹ���в�ͬ��̬�ͻ��Ŵ������Ե������ڸɺ�в���¶�NSCs�����ò��Բ�ͬ(�����ٺ��Ŵ���, 2006; ���Ŀ���������, 2016), ������ҶҰ���������������ݵ�NSCs�����Ըɺ�����Ӧ�̶Ȳ�ͬ(ͼ4)�����, ��ͬ��̬λ�ϵ��������ʽ��������Դ����ʽ�ȷ�����в���(��������, 2004), Ϊ�˱�֤��������������, ��ͬ���ֱ���ͨ����������NSCs�ĺ�����Ӧ�Ըɺ�в��, ���ڲ�ԭ�������ָɺ�в����NSCs��Ӧ��ͬ����Ӧ������Ϊ������̬λģ��Ԥ���ԭ���ַֻ���������̬λ��ģ�ṩ����֧�֡�3.2 �ɺ��Բ�ԭ��ֲͬ�﹦��ȺҶƬNSCsӰ��
�о�����, ��ԭֲ���NSCs��л���̶Լ��˸ɺ�����Ӧ������������������, �ڲ�ͬ����ȺҲ�������Բ���, ���о����ֺ̲ݹ���ȺNSCs�����Ϳ������Ǻ����Ըɺ�в����Ӧ������, ���ɺ�в�����������˵��ۺ������෴, �Ǻ̲ݵ�NSCs�����Ϳ������Ǻ����ڸɺ�в������������, �����ۺ����������仯����˵����ͬ����Ⱥֲ����Ӧ�Ըɺ�в��ʱ��NSCs����ֵĵ��ػ��ƴ��ڲ��졣�澳в����, �̲ݹ���Ⱥ�Էǽṹ��̼ˮ������Ĵ洢������ڷǺ̲ݹ���Ⱥ���෴, �Ǻ̲ݹ���Ⱥ��֯�еĹ�ϲ���(NSCs)��Ҫ�Կ������ǵ���ʽ����, ��ֱ��Ϊ��������ֳ�������Ȼ�ṩ����������, �ڸɺ�в����, ����ľ�ʲ��ṹ����, �Ǻ̲�ֲ���ˮ������ĵ���������ں̲�ֲ��(��������, 2004)�����, �Ǻ̲���֯�е�NSCs��Ҫ�Կ������Ƕ��ǵ��۵���ʽ���ڡ�Ford��Wilson (1981)�о����ֿ���������ֲ�����������������ֱ�ӹ�ϵ, ��ֲ�ﳤ�����ܸɺ���������Ҫ����������, �ɸ�������ˮ�ֵ��������ֲ��ϸ������, ʹֲ����ȱˮ������ܹ�����������������������Ӧ�ɺ�в��(Iannucci et al., 2002)��Li��(2013)�о�����, �ڸɺ�������, ֲ��Ϊ���������ڵ�����������������NSCs������ֵĺ�������ֲͬ�﹦��Ⱥ�������ںͿ��������IJ�������ֲ��ķǽṹ��̼ˮ�������л���̡������ټ��˸ɺ�ʱ, ��ͬ����Ⱥ���ֽ���ȡ��ͬ��̼���ò�������Ӧв��(Chen, 2008; Chen et al., 2013)���̲����Ⱥϳɵ���, �Դ�����������������ɺ����������, ���������Ըɺ�����Ӧ�����С�ͬʱ, һ��������Ϊ�Ͼ��ĸɺ�в��, ��֯�еĵ��ۿ�����ת��Ϊ����Ϊ������ٽ�ľ�ʲ���ˮ�ֵĴ�����NSCs��Ϊ�����ڵ���Ĺ����ѱ�ǰ����֤ʵ(��������, 2004)���෴, �Ǻ̲ݽ�ͬ���Ĺ�ϲ����Կ������ǵ���ʽ�洢, ֱ�����ڵֿ�����в��, ά������������л�������һ�������Ŀ�������ˮƽ������ά��ϸ����ѹ, ����ϸ��ˮ�ֳ���, �������ˮ�ִ���Ч��(Dietze et al., 2014)��
4 ����
ֲ��ͨ��������úϳɵ�NSCs����Ϊ���л��ṩ����, ����һ���̶��Ϸ�ӳ�˸�ֲ�����绷������Ӧ���ԡ�Ȼ��, Ŀǰ�Ըɺ�Ӱ��ǽṹ��̼ˮ�����ﶯ̬���о���Ҫ������ľ��ֲ��, ���Բݱ�ֲ����о����Ա��������о�ͨ���ⶨ��ͬ�ɺ������²ݵ��ԭֲ��ҶƬNSCs������ֵĺ���, ̽���˲ݱ�ֲ��Ըɺ�в�ȵ�������Ӧ����, ���ֲ�ͬ���ֺ���Ⱥͨ����ȡ��ͬ��NSCs���ò�����Ӧ�Ըɺ����̲ݹ���Ⱥ�ڸɺ�в������Ҫ����ϲ����Ե��۵���ʽ���д洢�Ա�֤����ɺ������֯�������Ǻ̲ݹ���Ⱥͨ����߿������ǵĺ�����ά��ֲ��������ˮ�ִ��䡣���о���ʾ�˲ݵ��ԭֲ���ڸɺ������µ�̼��л��������Ӧ����, ���������ֺ���Ⱥ���NSCs���õ���̬λ�ֻ���������ѧ����, ��ڹ�����ֹ�����ƺ�Ⱥ�乹�����������Ҫ���塣
�ο����� ԭ��˳��
������ȵ���
������������
�����ڿ�Ӱ������
[��������: 1]
[��������: 1]
DOI:10.1007/s00442-011-2235-2URLPMID:22223080 [��������: 1]

Global climate models forecast an increase in the frequency and intensity of extreme weather events, including severe droughts. Based on multi-year relationships between precipitation amount and aboveground annual net primary production (ANPP), semi-arid grasslands are projected to be among the most sensitive ecosystems to changes in precipitation. To assess sensitivity to drought, as well as variability within the shortgrass steppe biome, we imposed moderate and severe rainfall reductions for two growing seasons in three undisturbed grasslands that varied in soil type and climate. We predicted strong drought-induced reductions in ANPP at all sites and greater sensitivity to drought in sites with lower average precipitation, consistent with continental-scale patterns. Identical experimental infrastructure at each site reduced growing season rainfall events by 50 or 80%, and significantly reduced average soil moisture in both years (by 21 and 46% of control levels, respectively). Despite reductions in soil moisture, ANPP responses varied unexpectedly-from no reduction in ANPP to a 51% decrease. Although sensitivity to drought was highest in the semi-arid grassland with lowest mean annual precipitation, patterns in responses to drought across these grasslands were also strongly related to rainfall event size. When growing season rainfall patterns were dominated by many smaller events, ANPP was significantly reduced by drought but not when rainfall patterns were characterized by large rain events. This interaction between drought sensitivity and rainfall event size suggests that ANPP responses to future droughts may be reduced if growing season rainfall regimes also become more extreme.
DOI:10.1146/annurev-arplant-050213-040054URLPMID:24274032 [��������: 2]
Nonstructural carbon (NSC) provides the carbon and energy for plant growth and survival. In woody plants, fundamental questions about NSC remain unresolved: Is NSC storage an active or passive process? Do older NSC reserves remain accessible to the plant? How is NSC depletion related to mortality risk? Herein we review conceptual and mathematical models of NSC dynamics, recent observations and experiments at the organismal scale, and advances in plant physiology that have provided a better understanding of the dynamics of woody plant NSC. Plants preferentially use new carbon but can access decade-old carbon when the plant is stressed or physically damaged. In addition to serving as a carbon and energy source, NSC plays important roles in phloem transport, osmoregulation, and cold tolerance, but how plants regulate these competing roles and NSC depletion remains elusive. Moving forward requires greater synthesis of models and data and integration across scales from -omics to ecology.
[��������: 1]
[��������: 1]
URLPMID:27061438 [��������: 1]
URLPMID:13577798
[��������: 1]
[��������: 2]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
DOI:10.1093/treephys/28.8.1287URLPMID:18519260 [��������: 1]
To test whether the altitudinal distribution of trees is determined by a carbon shortage or an insufficient sugar fraction (sugar:starch ratio) in treeline trees, we studied the status of nonstructural carbohydrates (NSC) and their components (total soluble sugars and starch) in Abies fabri (Mast.) Craib and Picea balfouriana var. hirtella Rehd. et Wils. trees along three elevational gradients, ranging from lower elevations to the alpine treeline, on the eastern edge of the Tibetan Plateau. For comparison, we investigated a low-altitude species (Tsuga yunnanensis (Franch.) Pritz.) which served as a warm-climate reference because it is distributed in closed montane forests below 3100 m a.s.l. in the study area. The carbon status of T. yunnanensis responded to altitude differently from that of the treeline species. At the species level, total NSC was not consistently more abundant in treeline trees than in trees of the same species growing at lower elevations. Thus there was no consistent evidence for carbon limitation of growth in treeline trees. For the three treeline species studied (P. balfouriana and A. fabri in the Kang-Ding Valley and A. fabri in the Mo-Xi Valley), winter NSC concentrations in treeline trees were significantly lower than in lower-elevation trees of the same species, suggesting that, in winter, carbon is limited in treeline trees. However, in no case was there total overwinter depletion of NSC or its components in treeline trees. Treeline and low-altitude species had similar sugar:starch ratios of about three at their upper-elevational limits in April. We conclude that survival and growth of trees at the elevational or latitudinal climate limit depend not only on NSC concentration in perennial tissues, but also on the maintenance of an overwintering sugar:starch ratio greater than three.
[��������: 1]
[��������: 1]
DOI:10.1002/ecy.2444URLPMID:30157292 [��������: 1]
The allocation and stoichiometry of plant nutrients in leaves reflect fundamental ecosystem processes, biotic interactions, and environmental drivers such as water availability. Climate change will lead to increases in drought severity and frequency, but how canopy nutrients will respond to drought, and how these responses may vary with community composition along aridity gradients is poorly understood. We experimentally addressed this issue by reducing precipitation amounts by 66% during two consecutive growing seasons at three sites located along a natural aridity gradient. This allowed us to assess drought effects on canopy nitrogen (N) and phosphorus (P) concentrations in arid and semiarid grasslands of northern China. Along the aridity gradient, canopy nutrient concentrations were positively related to aridity, with this pattern was driven primarily by species turnover (i.e., an increase in the relative biomass of N- and P-rich species with increasing aridity). In contrast, drought imposed experimentally increased N but decreased P concentrations in plant canopies. These changes were driven by the combined effects of species turnover and intraspecific variation in leaf nutrient concentrations. In addition, the sensitivity of canopy N and P concentrations to drought varied across the three sites. Canopy nutrient concentrations were less affected by drought at drier than wetter sites, because of the opposing effects of species turnover and intraspecific variation, as well as greater drought tolerance for nutrient-rich species. These contrasting effects of long-term aridity vs. short-term drought on canopy nutrient concentrations, as well as differing sensitivities among sites in the same grassland biome, highlight the challenge of predicting ecosystem responses to future climate change.
DOI:10.1111/j.1469-8137.2008.02436.xURLPMID:18422905
Severe droughts have been associated with regional-scale forest mortality worldwide. Climate change is expected to exacerbate regional mortality events; however, prediction remains difficult because the physiological mechanisms underlying drought survival and mortality are poorly understood. We developed a hydraulically based theory considering carbon balance and insect resistance that allowed development and examination of hypotheses regarding survival and mortality. Multiple mechanisms may cause mortality during drought. A common mechanism for plants with isohydric regulation of water status results from avoidance of drought-induced hydraulic failure via stomatal closure, resulting in carbon starvation and a cascade of downstream effects such as reduced resistance to biotic agents. Mortality by hydraulic failure per se may occur for isohydric seedlings or trees near their maximum height. Although anisohydric plants are relatively drought-tolerant, they are predisposed to hydraulic failure because they operate with narrower hydraulic safety margins during drought. Elevated temperatures should exacerbate carbon starvation and hydraulic failure. Biotic agents may amplify and be amplified by drought-induced plant stress. Wet multidecadal climate oscillations may increase plant susceptibility to drought-induced mortality by stimulating shifts in hydraulic architecture, effectively predisposing plants to water stress. Climate warming and increased frequency of extreme events will probably cause increased regional mortality episodes. Isohydric and anisohydric water potential regulation may partition species between survival and mortality, and, as such, incorporating this hydraulic framework may be effective for modeling plant survival and mortality under future climate conditions.
[��������: 1]
[��������: 1]
[��������: 1]
URLPMID:27228625 [��������: 1]
PMID:27228625 [��������: 1]
DOI:10.1111/gcb.14747URLPMID:31276260
Drought, widely studied as an important driver of ecosystem dynamics, is predicted to increase in frequency and severity globally. To study drought, ecologists must define or at least operationalize what constitutes a drought. How this is accomplished in practice is unclear, particularly given that climatologists have long struggled to agree on definitions of drought, beyond general variants of
DOI:10.1111/j.1365-2745.2011.01798.xURL [��������: 1]
1. Growing recognition of the importance of climate extremes as drivers of contemporary and future ecological dynamics has led to increasing interest in studying these locally and globally important phenomena.2. Many ecological studies examining the impacts of what are deemed climate extremes, such as heat waves and severe drought, do not provide a definition of extremity, either from a statistical context based on the long-term climatic record or from the perspective of the response of the system - are the effects extreme (unusual or profound) in comparison to normal variability?;3. A synthetic definition of an extreme climatic event (ECE) is proposed that includes 'extremeness' in both the driver and the response: an ECE is as an episode or occurrence in which a statistically rare or unusual climatic period alters ecosystem structure and/or function well outside the bounds of what is considered typical or normal variability. This definition is accompanied by a mechanistic framework based on the concept that extreme response thresholds associated with significant community change and altered ecosystem function must be crossed in order for an ECE to occur.4. Synthesis. A definition and mechanistic framework for ECEs is used to identify priorities for future research that will enable ecologists to more fully assess the ecological consequences of climate extremes for ecosystem structure and function today and in a future world where their frequency and intensity are expected to increase.]]>
DOI:10.1093/treephys/19.7.453URLPMID:12651551
Variations in resistance to drought-induced xylem cavitation, xylem air-entry points, stomatal behavior, and hydraulic conductivity were measured in four populations of Populus trichocarpa Torr. & A. Gray collected along an east-west humidity and temperature gradient in Washington State, USA. Xylem air-entry points were less negative in trees from moist environments (-0.71 and -1.32 MPa in the Hoh and Nisqually populations, respectively) than in trees from dry environments (-1.55 and -1.67 MPa in the Palouse and Yakima populations, respectively). Xylem cavitation in response to experimental drought was consistent with air-injection measures of xylem air-entry points for a given population. Populations vulnerable to cavitation also exhibited higher stem specific hydraulic conductivities and limited stomatal control compared with resistant populations. Populations exhibiting vulnerability to cavitation and limited stomatal control desiccated more rapidly during drought compared with resistant populations. This study provides evidence of interpopulation variation in resistance to drought-induced xylem cavitation, stomatal behavior, and hydraulic conductivity within Populus trichocarpa.
DOI:10.1016/j.jclepro.2014.02.059URL
Strategic niche management has been outlined as a policy approach to assist development and diffusion of cleaner new technologies. Based on a case study describing the efforts of a leading actor in the heavy vehicle industry to develop and commercially introduce hybrid-electric vehicles, this paper discusses strategic niche management from the perspective of the manufacturing firm. In particular, the paper analyses experiences from extensive field tests executed in London, tests which involved a number of different manufacturers, hybrid systems and vehicles. The scale of this technological niche added confidence and credibility to the technology and helped opening up an important niche market, which was supported by an extensive subsidy scheme. The analysis shows how the field tests and the subsidies jointly functioned as bridging policies that facilitated an important step in the market formation process. The analysis further points at a critical tension between niches and the volume-oriented business of a major vehicle manufacturer, showing how investments in new technology need to be warranted in market trends and projections of future sales and production volumes to become accepted within a vehicle manufacturer's organization. (C) 2014 Elsevier Ltd.
URL
Stipa grandis�� and ��Leymus chinensis��, were examined based on a seven��year nutrient addition experiment in a semi��arid steppe in Inner Mongolia of China. The experiment included five treatments: control, low P, low N, low N + high P, and high N + low P. Results showed that N, P, SC, starch, and non��structural carbohydrates (NSC) concentrations in leaves of the two plant species were significantly influenced by nutrient addition (P<0.05). A significant interaction effect (P<0.05) was observed between nutrient addition and plant species. Leaf N concentration was increased (P<0.05) for both plant species under the treatments with N addition; starch content was decreased (P<0.05) for both plants in the low N treatment; leaf SC in ��L. chinensis ��was decreased under the low P treatment; while NSC content in leaves of both species significantly decreased (P<0.05) with high N + low P addition. No significant response was observed for all variables in ��S. grandis ��leaf to P addition. The comparatively higher leaf C/N�� C/P, and SC/starch in ��S. grandis�� implied more available carbohydrates and a higher use efficiency of ��S. grandis�� in N and P nutrients, while ��L. chinensis�� exhibited a competitively stronger feature of absorbing nutrients applied.]]>
DOI:10.13287/j.1001-9332.201905.001URLPMID:31107017 [��������: 1]
Non-structural carbohydrates (NSC, including soluble sugars and starch) are key meta-bolites in tree, the storage characteristics of which in tree organs have received extensive attention. It is still unclear how NSC are allocated in the tissues (phloem and xylem) that have different function. In this study, we analyzed the concentration and allocation of NSC in the roots, and in phloem and xylem of the trunk in three dominant species of broadleaved Korean pine forest in the Changbai Mountain, Pinus koraiensis, Fraxinus mandschurica, and Tilia amurensis. The results showed that there was a significant difference in the concentration of NSC between the phloem and xylem. The soluble sugar dominated in the phloem, while starch dominated in the xylem. The concentration of NSC in trunk outside (divided by annual rings, 0-20 years), intermediate (20-40 years) and inner (>40 years) of different tree species was significantly different, but with no difference in the roots. The total soluble sugar concentration in the phloem of P. koraiensis and F. mandschurica was significantly higher than that of T. amurensis, while the difference in xylem was not significant. The results indicated that NSC allocation in the phloem and xylem of the tree had clear tissue differentiation, which might be related to the succession stage of the tree species or the functional evolution of the tissue. These findings would improve our understanding of the carbon storage characteristics and allocation mechanism in temperate trees.
PMID:31107017 [��������: 1]
[��������: 3]
[��������: 3]
[��������: 2]
[��������: 2]
[��������: 1]
[��������: 1]
URLPMID:25011317 [��������: 2]
Non-structural carbohydrates (NSCs) are an important energy source for the metabolism of plants. The size of the NSC pool is likely to mirror the overall carbon supply status and its dynamics strongly influences physiological processes in plants. In order to predict the response and adaptation of trees to climate change, this review summarized the current understanding of NSC pool in trees, and mainly focused on its seasonal and spatial variation for analyzing the relationships between environmental factors and NSC allocation. Moreover, the response and adaptation strategies of NSC pool in trees to climate change were also discussed. Finally, some suggestions were proposed for the potential study orientation of NSC pool in trees in future climate conditions.
PMID:25011317 [��������: 2]
[��������: 1]
[��������: 1]
[��������: 1]
Responses of two dominant plant species to drought stress and defoliation in the Inner Mongolia Steppe of China
1
2013
... �����ټ��˸ɺ�ʱ, ��ͬ����Ⱥ���ֽ���ȡ��ͬ��̼���ò�������Ӧв��(
Comparing physiological responses of two dominant grass species to nitrogen addition in Xilin River Basin of China
1
2005
... �����ټ��˸ɺ�ʱ, ��ͬ����Ⱥ���ֽ���ȡ��ͬ��̼���ò�������Ӧв��(
Unexpected patterns of sensitivity to drought in three semi-arid grasslands
1
2012
... ��ԭ��̬ϵͳ���ҹ�����½����̬ϵͳ, ռ�ҹ�½�������41.7%, ���ҹ���Ҫ������ҵ�������غ���ɫ��̬����, �ڱ��Ϲ�����̬��ȫ����ʳ��ȫ����ȫ����̬ƽ���з�����Ҫ����(
Nonstructural carbon in woody plants
2
2014
... ֲ��ͨ��������ý�CO2��ˮ�ϳ�̼ˮ������, ̼ˮ�����ﲻ�������ϸ���ṹ����Ҫ�ɷ�, �������³´�л��������Ҫ����Դ����, ����ά��ֲ������������������м�����Ҫ������(
... �����ټ��˸ɺ�ʱ, ��ͬ����Ⱥ���ֽ���ȡ��ͬ��̼���ò�������Ӧв��(
Hydrologic regulation of plant rooting depth
2017
Changes in levels of solutes during osmotic adjustment to water-stress in leaves of four tropical pasture species
1
1981
... �о�����, ��ԭֲ���NSCs��л���̶Լ��˸ɺ�����Ӧ������������������, �ڲ�ͬ����ȺҲ�������Բ���, ���о����ֺ̲ݹ���ȺNSCs�����Ϳ������Ǻ����Ըɺ�в����Ӧ������, ���ɺ�в�����������˵��ۺ���.�෴, �Ǻ̲ݵ�NSCs�����Ϳ������Ǻ����ڸɺ�в������������, �����ۺ����������仯.��˵����ͬ����Ⱥֲ����Ӧ�Ըɺ�в��ʱ��NSCs����ֵĵ��ػ��ƴ��ڲ���.�澳в����, �̲ݹ���Ⱥ�Էǽṹ��̼ˮ������Ĵ洢������ڷǺ̲ݹ���Ⱥ.�෴, �Ǻ̲ݹ���Ⱥ��֯�еĹ�ϲ���(NSCs)��Ҫ�Կ������ǵ���ʽ����, ��ֱ��Ϊ��������ֳ�������Ȼ�ṩ����.����, �ڸɺ�в����, ����ľ�ʲ��ṹ����, �Ǻ̲�ֲ���ˮ������ĵ���������ں̲�ֲ��(
Shortwave radiation transfer through a plant canopy covered by single and double layers of plastic
1
2015
... ���о������й���ѧԺ����Ӧ����̬�о����������վ�ļ��˸ɺ�ʵ��ƽ̨����.2015��, ѡȡ��Ծ���(������ֲ����)�����е��ʹ����Եĵض�, �������.ʵ��ƽ̨��������������, ��������ʵ�鴦��: �ɺ��Ͷ���, ÿ�������ظ�6��, ��12��С��, �ɺ���������������(5-8��) 66%�Ľ�ˮ��.С�����Ϊ6 m �� 6 m, ����С�����1 m, ÿ��С������������Ϊ1 m�IJ���ָ����Է�ֹС����ˮ�ֲ���������С���ڲ�.С������4 m �� 4 mΪ��������, �ܱ�����1 m������, �Լ�С��ԵЧӦ.�����ﶥ�����������θּܵĹ��νṹ, �������ؽ�ˮ���ų��ͼ��ٷ���ƻ���.������8��4 m �� 1.5 m�ĸ�����������ƴ�Ӷ���, ��ֱͶӰ�����С�������ͬ.���������ж�����������ϩ�����ϰ�, �����ϰ���и����ԡ��ͷ��ƶ�ָ��, ��UV��, �ܹ���������������ˮ��, �ּ������ή�����Ч����(��ҪΪɢ���)(
Understanding the roles of nonstructural carbohydrates in forest trees-from what we can measure to what we want to know
1
2016
... ֲ��ͨ��������ý�CO2��ˮ�ϳ�̼ˮ������, ̼ˮ�����ﲻ�������ϸ���ṹ����Ҫ�ɷ�, �������³´�л��������Ҫ����Դ����, ����ά��ֲ������������������м�����Ҫ������(
Spectrophotometric determination of total carbohydrate
1958
Water deficit effects on osmotic adjustment and solute accumulation in leaves of annual clovers
1
2002
... �о�����, ��ԭֲ���NSCs��л���̶Լ��˸ɺ�����Ӧ������������������, �ڲ�ͬ����ȺҲ�������Բ���, ���о����ֺ̲ݹ���ȺNSCs�����Ϳ������Ǻ����Ըɺ�в����Ӧ������, ���ɺ�в�����������˵��ۺ���.�෴, �Ǻ̲ݵ�NSCs�����Ϳ������Ǻ����ڸɺ�в������������, �����ۺ����������仯.��˵����ͬ����Ⱥֲ����Ӧ�Ըɺ�в��ʱ��NSCs����ֵĵ��ػ��ƴ��ڲ���.�澳в����, �̲ݹ���Ⱥ�Էǽṹ��̼ˮ������Ĵ洢������ڷǺ̲ݹ���Ⱥ.�෴, �Ǻ̲ݹ���Ⱥ��֯�еĹ�ϲ���(NSCs)��Ҫ�Կ������ǵ���ʽ����, ��ֱ��Ϊ��������ֳ�������Ȼ�ṩ����.����, �ڸɺ�в����, ����ľ�ʲ��ṹ����, �Ǻ̲�ֲ���ˮ������ĵ���������ں̲�ֲ��(
Beyond gradual warming: extreme weather events alter flower phenology of European grassland and heath species
2
2009
... ��ԭ��̬ϵͳ���ҹ�����½����̬ϵͳ, ռ�ҹ�½�������41.7%, ���ҹ���Ҫ������ҵ�������غ���ɫ��̬����, �ڱ��Ϲ�����̬��ȫ����ʳ��ȫ����ȫ����̬ƽ���з�����Ҫ����(
... ), ����̬ϵͳ�ṹ���ܲ�������Ӱ��.���ܼ��˸ɺ��¼�ͨ������, �����ֲ��Ⱥ�����̬ϵͳ��Ӱ����ܻᳬԽ���������ɺ�(
���˸ɺ������ɹ���ݲ�ԭˮ��ƽ���Ӱ��
1
2015
... ��ԭ��̬ϵͳ���ҹ�����½����̬ϵͳ, ռ�ҹ�½�������41.7%, ���ҹ���Ҫ������ҵ�������غ���ɫ��̬����, �ڱ��Ϲ�����̬��ȫ����ʳ��ȫ����ȫ����̬ƽ���з�����Ҫ����(
���˸ɺ������ɹ���ݲ�ԭˮ��ƽ���Ӱ��
1
2015
... ��ԭ��̬ϵͳ���ҹ�����½����̬ϵͳ, ռ�ҹ�½�������41.7%, ���ҹ���Ҫ������ҵ�������غ���ɫ��̬����, �ڱ��Ϲ�����̬��ȫ����ʳ��ȫ����ȫ����̬ƽ���з�����Ҫ����(
Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems
1999
The biosynthesis of sucrose-correction
1
1955
... ֲ��ͨ��������ý�CO2��ˮ�ϳ�̼ˮ������, ̼ˮ�����ﲻ�������ϸ���ṹ����Ҫ�ɷ�, �������³´�л��������Ҫ����Դ����, ����ά��ֲ������������������м�����Ҫ������(
Responses of leaf nitrogen and mobile carbohydrates in different Quercus species/provenances to moderate climate changes
1
2013
... �о�����, ��ԭֲ���NSCs��л���̶Լ��˸ɺ�����Ӧ������������������, �ڲ�ͬ����ȺҲ�������Բ���, ���о����ֺ̲ݹ���ȺNSCs�����Ϳ������Ǻ����Ըɺ�в����Ӧ������, ���ɺ�в�����������˵��ۺ���.�෴, �Ǻ̲ݵ�NSCs�����Ϳ������Ǻ����ڸɺ�в������������, �����ۺ����������仯.��˵����ͬ����Ⱥֲ����Ӧ�Ըɺ�в��ʱ��NSCs����ֵĵ��ػ��ƴ��ڲ���.�澳в����, �̲ݹ���Ⱥ�Էǽṹ��̼ˮ������Ĵ洢������ڷǺ̲ݹ���Ⱥ.�෴, �Ǻ̲ݹ���Ⱥ��֯�еĹ�ϲ���(NSCs)��Ҫ�Կ������ǵ���ʽ����, ��ֱ��Ϊ��������ֳ�������Ȼ�ṩ����.����, �ڸɺ�в����, ����ľ�ʲ��ṹ����, �Ǻ̲�ֲ���ˮ������ĵ���������ں̲�ֲ��(
Mobile carbohydrates in Himalayan treeline trees I. Evidence for carbon gain limitation but not for growth limitation
1
2008
... ��ֲ����Ʒ���к�ɷ����, �ⶨֲ���п������Ǻ͵��۵ĺ���.����ⶨ����Ϊ: ��ȡ0.1 gֲ����Ʒ��10 mL���Ĺ���, ��5 mL 80%�Ҵ���Һ����, ��80 ���ˮ��ˮԡ30 min, ��ȴ�����º���3 500 r��min-1������10 min, �ظ���ȡ3��, ת������Һ���ݵ�25 mL��ɫ�����ڿ������Ǻ����IJⶨ.����������м���2 mL����ˮ�ڷ�ˮ�к���10 min, ��ȴ�����2 mL 9.2 mol��L-1��HClO4��Һ, ������Ⱥ��4 mL����ˮ����, ��4 000 r��min-1������10 min, ������Һת����50 mL����ƿ, ����������м���4.6 mol��L-1 HClO4��Һ, ������Ⱥ��5 mL����ˮ���Ⱥ�, ��4 000 r��min-1������10 min, ת������Һ, ����ˮϴ����2�κ�������Һ������50 mL, ���ڵ��ۺ����IJⶨ(
Mycorrhiza formation and elevated CO2 both increase the capacity for sucrose synthesis in source leaves of spruce and aspen
1
2000
... ֲ��ͨ��������ý�CO2��ˮ�ϳ�̼ˮ������, ̼ˮ�����ﲻ�������ϸ���ṹ����Ҫ�ɷ�, �������³´�л��������Ҫ����Դ����, ����ά��ֲ������������������м�����Ҫ������(
Long term experimental drought alters community plant trait variation, not trait means, across three semiarid grasslands
1
2019
... ���о������й���ѧԺ����Ӧ����̬�о����������վ�ļ��˸ɺ�ʵ��ƽ̨����.2015��, ѡȡ��Ծ���(������ֲ����)�����е��ʹ����Եĵض�, �������.ʵ��ƽ̨��������������, ��������ʵ�鴦��: �ɺ��Ͷ���, ÿ�������ظ�6��, ��12��С��, �ɺ���������������(5-8��) 66%�Ľ�ˮ��.С�����Ϊ6 m �� 6 m, ����С�����1 m, ÿ��С������������Ϊ1 m�IJ���ָ����Է�ֹС����ˮ�ֲ���������С���ڲ�.С������4 m �� 4 mΪ��������, �ܱ�����1 m������, �Լ�С��ԵЧӦ.�����ﶥ�����������θּܵĹ��νṹ, �������ؽ�ˮ���ų��ͼ��ٷ���ƻ���.������8��4 m �� 1.5 m�ĸ�����������ƴ�Ӷ���, ��ֱͶӰ�����С�������ͬ.���������ж�����������ϩ�����ϰ�, �����ϰ���и����ԡ��ͷ��ƶ�ָ��, ��UV��, �ܹ���������������ˮ��, �ּ������ή�����Ч����(��ҪΪɢ���)(
Differential responses of canopy nutrients to experimental drought along a natural aridity gradient
1
2018
... ���о������й���ѧԺ����Ӧ����̬�о����������վ�ļ��˸ɺ�ʵ��ƽ̨����.2015��, ѡȡ��Ծ���(������ֲ����)�����е��ʹ����Եĵض�, �������.ʵ��ƽ̨��������������, ��������ʵ�鴦��: �ɺ��Ͷ���, ÿ�������ظ�6��, ��12��С��, �ɺ���������������(5-8��) 66%�Ľ�ˮ��.С�����Ϊ6 m �� 6 m, ����С�����1 m, ÿ��С������������Ϊ1 m�IJ���ָ����Է�ֹС����ˮ�ֲ���������С���ڲ�.С������4 m �� 4 mΪ��������, �ܱ�����1 m������, �Լ�С��ԵЧӦ.�����ﶥ�����������θּܵĹ��νṹ, �������ؽ�ˮ���ų��ͼ��ٷ���ƻ���.������8��4 m �� 1.5 m�ĸ�����������ƴ�Ӷ���, ��ֱͶӰ�����С�������ͬ.���������ж�����������ϩ�����ϰ�, �����ϰ���и����ԡ��ͷ��ƶ�ָ��, ��UV��, �ܹ���������������ˮ��, �ּ������ή�����Ч����(��ҪΪɢ���)(
Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought?
2008
Pitch the niche-taking responsibility for the concepts we use in ecology and species distribution modelling
1
2012
... ֲ��Ըɺ�����Ӧ����ֲָ��������������������̬����(����̼���ò���)����Ӧˮ�ֻ����仯������, ��ֲ���ˮ��в�ȵ���Ӧ��ָ�ɺ�в�Ȼᵼ��ֲ��Ĵ��ռ��ռ�ֲ���ַ����仯.ֲ������NSCs���������ֲ�������ͷ�����һϵ�����������Ҫ��Դ����, ����ֲ�����ں����ı仯�����ܷ�ӳֲ������̼���պ�̼���ĵ������״��, ͬʱҲ�ܷ�ӳֲ��Ըɺ�в�ȵ���Ӧ(
ֲ��ǽṹ������̼ˮ�������������̬ѧ�о���չ
1
2002
... ֲ��Ըɺ�����Ӧ����ֲָ��������������������̬����(����̼���ò���)����Ӧˮ�ֻ����仯������, ��ֲ���ˮ��в�ȵ���Ӧ��ָ�ɺ�в�Ȼᵼ��ֲ��Ĵ��ռ��ռ�ֲ���ַ����仯.ֲ������NSCs���������ֲ�������ͷ�����һϵ�����������Ҫ��Դ����, ����ֲ�����ں����ı仯�����ܷ�ӳֲ������̼���պ�̼���ĵ������״��, ͬʱҲ�ܷ�ӳֲ��Ըɺ�в�ȵ���Ӧ(
ֲ��ǽṹ������̼ˮ�������������̬ѧ�о���չ
1
2002
... ֲ��Ըɺ�����Ӧ����ֲָ��������������������̬����(����̼���ò���)����Ӧˮ�ֻ����仯������, ��ֲ���ˮ��в�ȵ���Ӧ��ָ�ɺ�в�Ȼᵼ��ֲ��Ĵ��ռ��ռ�ֲ���ַ����仯.ֲ������NSCs���������ֲ�������ͷ�����һϵ�����������Ҫ��Դ����, ����ֲ�����ں����ı仯�����ܷ�ӳֲ������̼���պ�̼���ĵ������״��, ͬʱҲ�ܷ�ӳֲ��Ըɺ�в�ȵ���Ӧ(
��̬λ������ں��ķ�չ��������̬ѧ�еĶ�λ
1
2016
... ֲ��Ըɺ�����Ӧ����ֲָ��������������������̬����(����̼���ò���)����Ӧˮ�ֻ����仯������, ��ֲ���ˮ��в�ȵ���Ӧ��ָ�ɺ�в�Ȼᵼ��ֲ��Ĵ��ռ��ռ�ֲ���ַ����仯.ֲ������NSCs���������ֲ�������ͷ�����һϵ�����������Ҫ��Դ����, ����ֲ�����ں����ı仯�����ܷ�ӳֲ������̼���պ�̼���ĵ������״��, ͬʱҲ�ܷ�ӳֲ��Ըɺ�в�ȵ���Ӧ(
��̬λ������ں��ķ�չ��������̬ѧ�еĶ�λ
1
2016
... ֲ��Ըɺ�����Ӧ����ֲָ��������������������̬����(����̼���ò���)����Ӧˮ�ֻ����仯������, ��ֲ���ˮ��в�ȵ���Ӧ��ָ�ɺ�в�Ȼᵼ��ֲ��Ĵ��ռ��ռ�ֲ���ַ����仯.ֲ������NSCs���������ֲ�������ͷ�����һϵ�����������Ҫ��Դ����, ����ֲ�����ں����ı仯�����ܷ�ӳֲ������̼���պ�̼���ĵ������״��, ͬʱҲ�ܷ�ӳֲ��Ըɺ�в�ȵ���Ӧ(
How ecologists define drought, and why we should do better
2019
An ecological perspective on extreme climatic events: a synthetic definition and framework to guide future research
1
2011
... ��ԭ��̬ϵͳ���ҹ�����½����̬ϵͳ, ռ�ҹ�½�������41.7%, ���ҹ���Ҫ������ҵ�������غ���ɫ��̬����, �ڱ��Ϲ�����̬��ȫ����ʳ��ȫ����ȫ����̬ƽ���з�����Ҫ����(
Regulation of water loss in populations of Populus trichocarpa: the role of stomatal control in preventing xylem cavitation
1999
Strategic niche management from a business perspective: taking cleaner vehicle technologies from prototype to series production
2014
�ߺ��ݵ鲻ͬ�ݵ�������Ⱥ�����Լ���ɶ�ֲ��Ⱥ����������Ӱ��
2004
�ߺ��ݵ鲻ͬ�ݵ�������Ⱥ�����Լ���ɶ�ֲ��Ⱥ����������Ӱ��
2004
��ɺ����Ͳ�ԭ�������Ӷ���������ҶƬ�����ǽṹ��̼ˮ�����ﺬ����Ӱ��
2014
��ɺ����Ͳ�ԭ�������Ӷ���������ҶƬ�����ǽṹ��̼ˮ�����ﺬ����Ӱ��
2014
����ɽ��Ҷ������3����Ҫ���ֵķǽṹ��̼��������
1
2019
... ֲ��ͨ��������ý�CO2��ˮ�ϳ�̼ˮ������, ̼ˮ�����ﲻ�������ϸ���ṹ����Ҫ�ɷ�, �������³´�л��������Ҫ����Դ����, ����ά��ֲ������������������м�����Ҫ������(
����ɽ��Ҷ������3����Ҫ���ֵķǽṹ��̼��������
1
2019
... ֲ��ͨ��������ý�CO2��ˮ�ϳ�̼ˮ������, ̼ˮ�����ﲻ�������ϸ���ṹ����Ҫ�ɷ�, �������³´�л��������Ҫ����Դ����, ����ά��ֲ������������������м�����Ҫ������(
ˮ��в�ȶ���ݹ�ϲ�����估�����彻��������Ӱ��
3
2004
... ֲ��Ըɺ�����Ӧ����ֲָ��������������������̬����(����̼���ò���)����Ӧˮ�ֻ����仯������, ��ֲ���ˮ��в�ȵ���Ӧ��ָ�ɺ�в�Ȼᵼ��ֲ��Ĵ��ռ��ռ�ֲ���ַ����仯.ֲ������NSCs���������ֲ�������ͷ�����һϵ�����������Ҫ��Դ����, ����ֲ�����ں����ı仯�����ܷ�ӳֲ������̼���պ�̼���ĵ������״��, ͬʱҲ�ܷ�ӳֲ��Ըɺ�в�ȵ���Ӧ(
... �о�����, ��ԭֲ���NSCs��л���̶Լ��˸ɺ�����Ӧ������������������, �ڲ�ͬ����ȺҲ�������Բ���, ���о����ֺ̲ݹ���ȺNSCs�����Ϳ������Ǻ����Ըɺ�в����Ӧ������, ���ɺ�в�����������˵��ۺ���.�෴, �Ǻ̲ݵ�NSCs�����Ϳ������Ǻ����ڸɺ�в������������, �����ۺ����������仯.��˵����ͬ����Ⱥֲ����Ӧ�Ըɺ�в��ʱ��NSCs����ֵĵ��ػ��ƴ��ڲ���.�澳в����, �̲ݹ���Ⱥ�Էǽṹ��̼ˮ������Ĵ洢������ڷǺ̲ݹ���Ⱥ.�෴, �Ǻ̲ݹ���Ⱥ��֯�еĹ�ϲ���(NSCs)��Ҫ�Կ������ǵ���ʽ����, ��ֱ��Ϊ��������ֳ�������Ȼ�ṩ����.����, �ڸɺ�в����, ����ľ�ʲ��ṹ����, �Ǻ̲�ֲ���ˮ������ĵ���������ں̲�ֲ��(
... �����ټ��˸ɺ�ʱ, ��ͬ����Ⱥ���ֽ���ȡ��ͬ��̼���ò�������Ӧв��(
ˮ��в�ȶ���ݹ�ϲ�����估�����彻��������Ӱ��
3
2004
... ֲ��Ըɺ�����Ӧ����ֲָ��������������������̬����(����̼���ò���)����Ӧˮ�ֻ����仯������, ��ֲ���ˮ��в�ȵ���Ӧ��ָ�ɺ�в�Ȼᵼ��ֲ��Ĵ��ռ��ռ�ֲ���ַ����仯.ֲ������NSCs���������ֲ�������ͷ�����һϵ�����������Ҫ��Դ����, ����ֲ�����ں����ı仯�����ܷ�ӳֲ������̼���պ�̼���ĵ������״��, ͬʱҲ�ܷ�ӳֲ��Ըɺ�в�ȵ���Ӧ(
... �о�����, ��ԭֲ���NSCs��л���̶Լ��˸ɺ�����Ӧ������������������, �ڲ�ͬ����ȺҲ�������Բ���, ���о����ֺ̲ݹ���ȺNSCs�����Ϳ������Ǻ����Ըɺ�в����Ӧ������, ���ɺ�в�����������˵��ۺ���.�෴, �Ǻ̲ݵ�NSCs�����Ϳ������Ǻ����ڸɺ�в������������, �����ۺ����������仯.��˵����ͬ����Ⱥֲ����Ӧ�Ըɺ�в��ʱ��NSCs����ֵĵ��ػ��ƴ��ڲ���.�澳в����, �̲ݹ���Ⱥ�Էǽṹ��̼ˮ������Ĵ洢������ڷǺ̲ݹ���Ⱥ.�෴, �Ǻ̲ݹ���Ⱥ��֯�еĹ�ϲ���(NSCs)��Ҫ�Կ������ǵ���ʽ����, ��ֱ��Ϊ��������ֳ�������Ȼ�ṩ����.����, �ڸɺ�в����, ����ľ�ʲ��ṹ����, �Ǻ̲�ֲ���ˮ������ĵ���������ں̲�ֲ��(
... �����ټ��˸ɺ�ʱ, ��ͬ����Ⱥ���ֽ���ȡ��ͬ��̼���ò�������Ӧв��(
���˽�ˮ�ͼ��˸ɺ��¼��Բ�ԭ��̬ϵͳ��Ӱ��
2
2014
... ��ԭ��̬ϵͳ���ҹ�����½����̬ϵͳ, ռ�ҹ�½�������41.7%, ���ҹ���Ҫ������ҵ�������غ���ɫ��̬����, �ڱ��Ϲ�����̬��ȫ����ʳ��ȫ����ȫ����̬ƽ���з�����Ҫ����(
... ;
���˽�ˮ�ͼ��˸ɺ��¼��Բ�ԭ��̬ϵͳ��Ӱ��
2
2014
... ��ԭ��̬ϵͳ���ҹ�����½����̬ϵͳ, ռ�ҹ�½�������41.7%, ���ҹ���Ҫ������ҵ�������غ���ɫ��̬����, �ڱ��Ϲ�����̬��ȫ����ʳ��ȫ����ȫ����̬ƽ���з�����Ҫ����(
... ;
ֲ����Ⱥ���ӽ����ж���������Ӧ
1
2003
... ֲ��Ըɺ�����Ӧ����ֲָ��������������������̬����(����̼���ò���)����Ӧˮ�ֻ����仯������, ��ֲ���ˮ��в�ȵ���Ӧ��ָ�ɺ�в�Ȼᵼ��ֲ��Ĵ��ռ��ռ�ֲ���ַ����仯.ֲ������NSCs���������ֲ�������ͷ�����һϵ�����������Ҫ��Դ����, ����ֲ�����ں����ı仯�����ܷ�ӳֲ������̼���պ�̼���ĵ������״��, ͬʱҲ�ܷ�ӳֲ��Ըɺ�в�ȵ���Ӧ(
ֲ����Ⱥ���ӽ����ж���������Ӧ
1
2003
... ֲ��Ըɺ�����Ӧ����ֲָ��������������������̬����(����̼���ò���)����Ӧˮ�ֻ����仯������, ��ֲ���ˮ��в�ȵ���Ӧ��ָ�ɺ�в�Ȼᵼ��ֲ��Ĵ��ռ��ռ�ֲ���ַ����仯.ֲ������NSCs���������ֲ�������ͷ�����һϵ�����������Ҫ��Դ����, ����ֲ�����ں����ı仯�����ܷ�ӳֲ������̼���պ�̼���ĵ������״��, ͬʱҲ�ܷ�ӳֲ��Ըɺ�в�ȵ���Ӧ(
ľ��ֲ��ǽṹ��̼ˮ������仯����Ӱ�������о���չ
2
2014
... ֲ��Ըɺ�����Ӧ����ֲָ��������������������̬����(����̼���ò���)����Ӧˮ�ֻ����仯������, ��ֲ���ˮ��в�ȵ���Ӧ��ָ�ɺ�в�Ȼᵼ��ֲ��Ĵ��ռ��ռ�ֲ���ַ����仯.ֲ������NSCs���������ֲ�������ͷ�����һϵ�����������Ҫ��Դ����, ����ֲ�����ں����ı仯�����ܷ�ӳֲ������̼���պ�̼���ĵ������״��, ͬʱҲ�ܷ�ӳֲ��Ըɺ�в�ȵ���Ӧ(
... ).��Լ��˸ɺ��¼�, ֲ��ͨ�������ڴ洢һ������NSCs���ṩ���������������ˮ��в��, ά�����������(
ľ��ֲ��ǽṹ��̼ˮ������仯����Ӱ�������о���չ
2
2014
... ֲ��Ըɺ�����Ӧ����ֲָ��������������������̬����(����̼���ò���)����Ӧˮ�ֻ����仯������, ��ֲ���ˮ��в�ȵ���Ӧ��ָ�ɺ�в�Ȼᵼ��ֲ��Ĵ��ռ��ռ�ֲ���ַ����仯.ֲ������NSCs���������ֲ�������ͷ�����һϵ�����������Ҫ��Դ����, ����ֲ�����ں����ı仯�����ܷ�ӳֲ������̼���պ�̼���ĵ������״��, ͬʱҲ�ܷ�ӳֲ��Ըɺ�в�ȵ���Ӧ(
... ).��Լ��˸ɺ��¼�, ֲ��ͨ�������ڴ洢һ������NSCs���ṩ���������������ˮ��в��, ά�����������(
The response of root traits to precipitation change of herbaceous species in temperate steppes
1
2019
... ֲ��Ըɺ�����Ӧ����ֲָ��������������������̬����(����̼���ò���)����Ӧˮ�ֻ����仯������, ��ֲ���ˮ��в�ȵ���Ӧ��ָ�ɺ�в�Ȼᵼ��ֲ��Ĵ��ռ��ռ�ֲ���ַ����仯.ֲ������NSCs���������ֲ�������ͷ�����һϵ�����������Ҫ��Դ����, ����ֲ�����ں����ı仯�����ܷ�ӳֲ������̼���պ�̼���ĵ������״��, ͬʱҲ�ܷ�ӳֲ��Ըɺ�в�ȵ���Ӧ(
Ⱥ����̬ѧ����������
1
2006
... ֲ��Ըɺ�����Ӧ����ֲָ��������������������̬����(����̼���ò���)����Ӧˮ�ֻ����仯������, ��ֲ���ˮ��в�ȵ���Ӧ��ָ�ɺ�в�Ȼᵼ��ֲ��Ĵ��ռ��ռ�ֲ���ַ����仯.ֲ������NSCs���������ֲ�������ͷ�����һϵ�����������Ҫ��Դ����, ����ֲ�����ں����ı仯�����ܷ�ӳֲ������̼���պ�̼���ĵ������״��, ͬʱҲ�ܷ�ӳֲ��Ըɺ�в�ȵ���Ӧ(
Ⱥ����̬ѧ����������
1
2006
... ֲ��Ըɺ�����Ӧ����ֲָ��������������������̬����(����̼���ò���)����Ӧˮ�ֻ����仯������, ��ֲ���ˮ��в�ȵ���Ӧ��ָ�ɺ�в�Ȼᵼ��ֲ��Ĵ��ռ��ռ�ֲ���ַ����仯.ֲ������NSCs���������ֲ�������ͷ�����һϵ�����������Ҫ��Դ����, ����ֲ�����ں����ı仯�����ܷ�ӳֲ������̼���պ�̼���ĵ������״��, ͬʱҲ�ܷ�ӳֲ��Ըɺ�в�ȵ���Ӧ(
�ߺ��ݵ鲻ͬ���Ͳݵ�Ⱥ��������������ֲ����̬λ����
2016
�ߺ��ݵ鲻ͬ���Ͳݵ�Ⱥ��������������ֲ����̬λ����
2016

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}