Progress in the effects of elevated ground-level ozone on terrestrial ecosystems
Zhao-Zhong FENG,,1,*, Xiang-Yang YUAN2, Pin LI2, Bo SHANG2, Qin PING2, Ting-Jian HU2, Shuo LIU21Jiangsu Key Laboratory of Agricultural Meteorology, School of Applied Meteorology, Nanjing University of Information Science & Technology, Nanjing 210044, China 2State Key Laboratory of Urban and Regional Ecology, Research Center for Eco-Environmental Science, Chinese Academy of Sciences, Beijing 100085, China
Abstract Rising ground-level ozone (O3) is currently an essential environmental issue in the world, especially in China. While research on the effects of O3 on leaf photosynthetic gas exchange, plant growth and biomass has received a lot of attention, ecosystem-scale studies are however scarce and subject to great uncertainties. This article combs trends and hotpots of ground-level O3 concentration and its effects on plants and ecosystems over the past 40 years. Research techniques and assessment methods for studying the ecological effects of ozone pollution are covered. The most important advances on the impacts of elevated ozone on terrestrial ecosystem are reviewed: plant response mechanisms, effects on grain yield, crop quality, carbon sequestration capacity, community structure and below-ground processes of different terrestrial ecosystems. Finally, regional risk assessment of the O3 pollution is discussed. Considering the main knowledge gaps, future research should focus on belowground ecosystem response to elevated O3 and should also incorporate O3 and multi-factor experiments using Free-Air Ozone Concentration Elevation (FACE) system. More attention should also be paid on food security, establishment of Asian ozone network, standardization of risk assessment approach, and exploration of ecological measures to reduce the negative effects of O3 pollution. This review can help to promote more studies on the ecological effects of ground-level O3 pollution. Keywords:elevated ground-level ozone concentration;terrestrial ecosystem;effect;response;carbon sequestration;risk assessment
PDF (9811KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 冯兆忠, 袁相洋, 李品, 尚博, 平琴, 胡廷剑, 刘硕. 地表臭氧浓度升高对陆地生态系统影响的研究进展. 植物生态学报, 2020, 44(5): 526-542. DOI: 10.17521/cjpe.2019.0144 FENG Zhao-Zhong, YUAN Xiang-Yang, LI Pin, SHANG Bo, PING Qin, HU Ting-Jian, LIU Shuo. Progress in the effects of elevated ground-level ozone on terrestrial ecosystems. Chinese Journal of Plant Ecology, 2020, 44(5): 526-542. DOI: 10.17521/cjpe.2019.0144
基于地表O3与植被、土壤和生态系统研究领域的基本状况, 通过Web of Science核心数据库, 选取检索式TS = ((“vegetation” OR “plant*” OR “soil” OR “ecosystem”) AND (“O3” OR “ozone”))对1990-2018年世界范围内的SCI文章发表状况进行检索, 得到文献10 705篇。然后根据主题“臭氧对植物、土壤及生态系统等的影响”进行人工筛选和逐一排查, 最终获得有效SCI文章2 549篇。自1995年, 中国和世界上发表地表O3污染对植物及生态系统影响研究的SCI文章统计见图1。文章发表最多的时段是1998-2007年, 平均每年150篇左右, 最高达到182篇。2008年文章发表量相对较少, 总计只有17篇, 但之后2009至2018年, 文章数量比较平稳, 平均为每年100余篇。中国的研究成果在SCI源期刊发表数量自1998年开始逐年增加, 2018年达到25篇(截止到2018年11月)。
Fig. 1Articles have been published in the SCI journals about the effects of ground-level O3 on plants or ecosystem from China and the world since 1995.
Table 1 表1 表1O3浓度升高对植物及生态系统影响的SCI文章出现频次最多的50个关键词 Table 1Top 50 most frequent keywords of SCI papers about the effects of elevated ground-level O3 on plants or ecosystem
序号 Number
关键词 Keyword
频次 Frequency
序号 Number
关键词 Key
频次 Frequency
1
臭氧 Ozone
748
26
叶绿素 Chlorophyll
27
2
空气污染 Air pollution
118
27
臭氧通量 Ozone flux
27
3
光合作用 Photosynthesis
110
28
臭氧损耗 Ozone depletion
26
4
气孔导度 Stomatal conductance
103
29
臭氧吸收 Ozone uptake
26
5
对流层臭氧 Tropospheric ozone
95
30
风险评估 Risk assessment
26
6
氧化压力 Oxidative stress
75
31
大豆 Soybean
26
7
产量 Yield
70
32
紫外线B辐射 UV-B radiation
26
8
气候变化 Climate change
62
33
二氧化碳 CO2
25
9
生长 Growth
62
34
欧洲山毛榉 Fagus sylvatica
25
10
小时臭氧浓度超过40 μg·kg-1的积累值 AOT40
56
35
干旱 Drought
24
11
小麦 Wheat
55
36
森林 Forests
24
12
二氧化碳 Carbon dioxide
54
37
开顶气室 Open top chambers
24
13
生物量 Biomass
50
38
抗坏血酸 Ascorbic acid
23
14
臭氧氧化 Ozonation
49
39
消毒/杀菌 Disinfection
23
15
气体交换 Gas exchange
45
40
乙烯二脲的简称 EDU
23
16
臭氧浓度升高 Elevated ozone
45
41
乙烯二脲 Ethylenediurea
22
17
紫外线B辐射 UV-B
41
42
水稻 Rice
22
18
抗氧化物 Antioxidants
39
43
开放式自由空气增强系统 FACE
22
19
氮 Nitrogen
38
44
烟草 Tobacco
21
20
活性氧 Reactive oxygen species
36
45
叶绿素a荧光 Chlorophyll a fluorescence
21
21
叶绿素荧光 Chlorophyll fluorescence
33
46
乙烯 Ethylene
20
22
全球变化 Global change
33
47
过氧化氢 Hydrogen peroxide
20
23
抗坏血酸盐 Ascorbate
30
48
生物监测 Biomonitoring
20
24
二氧化碳浓度升高 Elevated CO2
30
49
膜脂过氧化 Lipid peroxidation
19
25
临界水平 Critical levels
29
50
地表臭氧 Surface ozone
19
AOT40, accumulated hourly O3 concentration over a threshold of 40 μg·kg-1.
Table 2 表2 表2地表O3浓度升高对植物及生态系统影响不同研究方法的优缺点 Table 2Advantages and disadvantages of different study methods about the effects of elevated ground-level O3 on plants or ecosystem
研究方法 Study method
优点 Advantage
缺点 Disadvantage
室内生长箱 Growth chamber
技术简单, 操作容易, 费用低, 可控温湿度、光照 Simple technique, easy to operate, low cost. Temperature, humidity and light intensity can be controlled
空间小, 短期实验为主, 与真实大气环境不符 Small space, suitable for short-term experiments, inconsistent with the real atmospheric environment
开顶气室 Open top chamber
技术简单, 操作容易, 低费用, 高精度, 多因子, 可过滤 O3并进行田间试验 Simple technique, easy to operate, low cost, high precision, suitable for multi-factor and field experimental study, reducing O3 less than ambient air
空间小, 短期幼苗实验, 盆栽为主, 微气候效应 Small space, suitable for short-term and pot experiments such as seedling, significant microclimate effects
自由空气中气体浓度增加系统 Free-Air Concentration Elevation
自然环境, 多因子, 长期实验, 大田研究, 研究尺度囊括叶片、个体、群落或生态系统水平 Natural environment, suitable for multi-factor, field and long-term experimental study, scale covers leaf-, individual-, community- and ecosystem-level
技术要求高, 费用昂贵, 普适性差 Higher technical requirement, high cost, and poor universality
不管研究方法如何更新, 由于空间和时间上的限制, 模拟地表O3浓度升高的控制实验主要探究高浓度O3对植物损伤的机理, 而区域尺度上探究O3对陆地生态系统的影响却很难通过模拟实验获得。为评价区域O3风险和评估O3对生态系统影响带来的损失, ****主要通过不同评估指标评估区域、国家或全球尺度上地表O3污染的生态效应。总体来说, 评估指标的研究主要经历了三个阶段, 分别是浓度响应关系、剂量响应关系和通量响应关系评估指标(Musselman et al., 2006)。浓度响应关系评估指标主要包括M7/M12 (白天7 h (9:00-16:00) O3浓度平均值或白天12 h (8:00-20:00) O3浓度平均值), 早期曾广泛应用于表征O3浓度暴露与农作物产量损失相关性的研究中(Legge et al., 1995)。剂量响应关系评估指标主要有SUM06 (O3小时浓度高于或等于60 μg·kg-1的累积值)(Lefohn & Foley, 1992), W126 (O3小时浓度在规定时段内的加权求和值)和AOTX (整个生长季太阳辐射>50 W·m-2时段内O3小时浓度超过X μg·kg-1的累计值, X普遍设为40 μg·kg-1)(Fuhrer et al., 1997; LRTAR Convention, 2015)。剂量响应指标综合考虑O3浓度和暴露时间对植物生长的影响, 在世界范围内得到广泛应用。
然而, O3对植物的影响不仅依赖于大气O3浓度同时也依赖于叶片气孔导度(O3的吸收)和植物自身抗氧化能力(Matyssek et al., 2007)。近年来, 基于气孔O3吸收量的评估指标PODY (植物毒性O3剂量, 整个生长季单位面积上气孔O3吸收通量超过临界值Y nmol m-2·s-1的积累量)被开发并逐步替代浓度暴露剂量指标, 用于区域或全球O3对农作物和森林树种生长影响的评估(Büker et al., 2015; LRTAR Convention, 2015)。目前, 哪个指标更适合区域O3风险评估的争论一直存在, 但欧洲和亚洲地区多项O3对农作物或植被影响的研究均表明: 考虑水分供应的情况下, O3暴露造成的生物量或作物产量的降低与PODY的拟合度比AOTX更好(Mills et al., 2011; Gao et al., 2017), 能更好表征O3对生态系统的影响。整体来看, 各个评估指标的计算方式和所需参数从简单到复杂, 从单一考虑O3浓度到综合考虑环境因子对植物O3吸收通量的影响, 逐渐可以精确评估地表O3污染对生态系统农林生产力的影响(表3)。
Table 3 表3 表3地表O3对生态系统影响评估指标的优缺点对比 Table 3Advantages and disadvantages of different O3 metrics about the effects of ground-level O3 on ecosystem
评估模型 Evaluation model
评估指标 Evaluation indicator
优点 Advantage
缺点 Disadvantage
浓度响应关系 Concentration-response relationship
白天7 h (9:00-16:00) O3浓度平均值(M7)或白天12 h (8:00-20:00) O3浓度平均值(M12) 7 h (9:00-16:00) seasonal mean O3 concentrations (M7), 12 h (8:00-18:00) seasonal mean O3 concentrations (M12)
计算简单, 直观易懂, 参数需求较少 Simple calculation, easy understanding, less parameters
仅考虑O3浓度唯一因素, 机理性较差 This approach only considers O3 concentration, but no physical mechanism
O3小时浓度高于或等于60 μg·kg-1的累积值(SUM06), O3小时浓度在规定时段内的加权求和值(W126), 整个生长季太阳辐射>50 W·m-2时段内O3小时浓度超过X μg·kg-1的累计值(AOTX) The sum of all hourly average concentrations > 60 μg·kg-1 (SUM06), a cumulative ozone exposure index based on sigmoidally weighted daytime O3 concentrations (W126), accumulated hourly O3 concentration over a threshold of X μg·kg-1 during daylight hours (AOTX)
计算简单, 同时关注O3浓度和暴露时 间, 应用广泛 Simple calculation and wide application, the method concerns O3 concentration and exposure time simultaneously
只探讨O3对植物的影响, 没有考虑其他环境因子的影响, 缺乏生物学意义 This approach only considering the factor O3 but not consider the effect of other environmental factors on plants, and thus no biological meaning is covered
通量响应关系 Flux-based dose-responses relationship
整个生长季单位面积上气孔O3吸收通量超过临界值Y nmol m-2·s-1的积累量(PODY) Phytotoxic O3 dose over a threshold of Y (PODY)
同时考虑环境因子(物候、温度、光照、蒸气压和土壤水势等参数)和植物自身对O3响应的影响 Environmental factors (e.g. phenology, temperature, photosynthetically photon flux density, water vapor pressure deficit, soil water potential) and plants itself were considered
所需参数较多, 计算过程复杂, 树种特异性限制较大 This approach required many parameters with a complex calculation procedure, but varied by species
Fig. 3Effects of elevated O3 on plant processes at the leaf, rhizosphere, whole-plant and ecosystem scales. -, decrease or reduction; +, increase or acceleration; ?, uncertain effect or a question not addressed in current studies.
AgathokleousE, SaitanisCJ, WangXN, WatanabeM, KoikeT (2016). A review study on past 40 years of research on effects of tropospheric O3 on belowground structure, functioning, and processes of trees: a linkage with potential ecological implications Water, Air, & Soil Pollution, 227, 33. DOI: 10.1007/s11270-015-2715-9. [本文引用: 1]
AinsworthEA (2017). Understanding and improving global crop response to ozone pollution The Plant Journal, 90, 886-897. URLPMID:27739639 [本文引用: 2]
AinsworthEA, LemonnierP, WedowJM (2020). The influence of rising tropospheric carbon dioxide and ozone on plant productivity Plant Biology, 22(S1), 5-11. [本文引用: 1]
AinsworthEA, YendrekCR, SitchS (2012). The effects of tropospheric ozone on net primary productivity and implications for climate change Annual Review of Plant Biology, 63, 637-661. URLPMID:22404461 [本文引用: 3]
AndersenCP (2003). Source-sink balance and carbon allocation below ground in plants exposed to ozone: tansley review New Phytologist, 157, 213-228. [本文引用: 2]
AshmoreMR (2005). Assessing the future global impacts of ozone on vegetation Plant, Cell & Environment, 28, 949-964. [本文引用: 2]
AvneryS, MauzerallDL, LiuJF, HorowitzLW (2011). Global crop yield reductions due to surface ozone exposure: 1. Year 2000 crop production losses and economic damage Atmospheric Environment, 45, 2284-2296. [本文引用: 2]
BaiYM, WangCY, LiuL, GuoJP, WenM (2002). A diagnostic experiment and study of the influence of O3 on pakchoi Journal of Applied Meteorological Science, 13, 364-370. [本文引用: 1]
BarboDN, ChappelkaAH, SomersGL, Miller-GoodmanMS, StolteK (1998). Diversity of an early successional plant community as in?uenced by ozone New Phytologist, 138, 653-662. [本文引用: 1]
BergmannE, BenderJ, WeigelHJ (2017). Impact of tropospheric ozone on terrestrial biodiversity: a literature analysis to identify ozone sensitive taxa Journal of Applied Botany and Food Quality, 90, 83-105. [本文引用: 1]
Booker FR. MuntiferingR, McGrathM, BurkeyK, DecoteauD, FiscusE, ManningW, KrupaS, ChappelkaA, GrantzD (2009). The ozone component of global change: potential effects on agricultural and horticultural plant yield, product quality and interactions with invasive species Journal of Integrative Plant Biology, 51, 337-351. [本文引用: 3]
BükerP, FengZZ, UddlingJ, BriolatA, AlonsoR, BraunS, ElviraS, GerosaG, KarlssonPE, ThiecDL, MarzuoliR, MillsG, OksanenE, WieserG, WilkinsonM, EmbersonLD (2015). New flux based dose-response relationships for ozone for European forest tree species Environmental Pollution, 206, 163-174. [本文引用: 1]
CannonWN (1990). Olfactory response of eastern spruce budworm larvae to red spruce needles exposed to acid-rain and elevated levels of ozone Journal of Chemical Ecology, 16, 3255-3261. DOI:10.1007/BF00982096URLPMID:24263427 [本文引用: 1] Second-instar eastern spruce budworm larvae,Choristoneura fumiferana (Clem.), were tested in a two-choice, Y-type, wind-tunnel olfactometer for preferences for red spruce,Picea rubens Sarg., needles exposed for eight weeks to charcoal-filtered air (CFA) or 0.15 ppm ozone (O3) in combination with acidified rainfall at pH 4.2 or 3.0. Volatiles from needles treated with CFA plus pH 4.2 rain (control) were preferred over those from needles exposed to pH 3.0 rain + O3 or O3 alone; O3-treated needles were chosen over those exposed to pH 3.0 rain + O3. No preference was shown between the pH 4.2 and 3.0 rain treatments. Larvae chose purified air flowing through the olfactometer in preference to needle volatiles from the pH 3.0 rain + O3 treatment.
CaoJC, ZhengYF, ZhaoH, XuJX (2017). Impact of elevated ozone concentration on growth and yield of winter wheat and soybean Asian Journal of Ecotoxicology, 12(2), 129-136. [本文引用: 1]
ChameidesWL, KasibhatlaPS, YiengerJ, LevyIIH (1994). Growth of continental-scale metro-agro-plexes, regional ozone pollution and world food production Science, 264, 74-77. [本文引用: 1]
ChappelkaAH, SamuelsonLJ (1998). Ambient ozone effects on forest trees of the eastern United States: a review New Phytologist, 139, 91-108. [本文引用: 1]
CollinsWJ, SitchS, BoucherO (2010). How vegetation impacts affect climate metrics for ozone precursors Journal of Geophysical Research, 115, D23308. DOI: 10.1029/2010JD014187. [本文引用: 1]
CooperOR, ParrishDD, ZiemkeJ, BalashovNV, CupeiroM, GalballyIE, GilgeS, HorowitzL, JensenNR, LamarqueJF, NaikV, OltmansSJ, SchwabJ, ShindellDT, ThompsonAM, ThouretV, WangY, ZbindenRM (2014). Global distribution and trends of tropospheric ozone: an observation- based review Elementa Science of the Anthropocene, 2, 000029. DOI: 10.12952/journal.elementa.000029. [本文引用: 1]
DahlstenDL, RowneuDL, KickertRN (1997). Effects of oxidant air pollutants on western pine beetle (Coleoptera: Scolytidae) populations in southern California Environmental Pollution, 96, 415-423. URLPMID:15093407 [本文引用: 1]
DaiLL, FengZZ, PanXD, XuYS, LiP, LefohnAS, HarmensH, KobayashiK (2019). Increase of apoplastic ascorbate induced by ozone is insufficient to remove the negative effects in tobacco, soybean and poplar Environmental Pollution, 245, 380-388. [本文引用: 1]
EltayebAE, KawanoN, BadawiGH, KaminakaH, SanekataT, MorishimaI, ShibaharaT, InanagaS, TanakaK (2006). Enhanced tolerance to ozone and drought stresses in transgenic tobacco overexpressing dehydroascorbate reductase in cytosol Physiologia Plantarum, 127, 57-65. [本文引用: 1]
EvansPA, AshmoreMR (1992). The effects of ambient air on a seminatural grassland community Agriculture Ecosystems and Environment, 38, 91-97. [本文引用: 1]
FelzerB, KicklighterD, MelilloJ, WangC, ZhuangQL, PrinnRG (2004). Effects of ozone on net primary production and carbon sequestration in the conterminous United States using a biogeochemistry model Tellus, 56B, 230-248. [本文引用: 1]
FelzerB, ReillyJ, MelilloJ, KicklighterDW, WangC, PrinnRG, SarofimMC, ZhuangQ (2005). Future effects of ozone on carbon sequestration and climate change policy using a global biogeochemical model Climate Change, 73, 345-373. [本文引用: 1]
FengZZ, BükerP, PleijelH, EmbersonL, KarlssonPE, UddlingJ (2018a). A unifying explanation for variation in ozone sensitivity among woody plants Global Change Biology, 24, 78-84. [本文引用: 1]
FengZZ, HuEZ, WangXK, JiangLJ, LiuXJ (2015a). Ground-level O3 pollution and its impacts on food crops in China: a review Environmental Pollution, 199, 42-48. [本文引用: 1]
FengZZ, KobayashiK (2009). Assessing the impacts of current and future concentrations of surface ozone on crop yield with meta-analysis Atmospheric Environment, 43, 1510-1519. [本文引用: 2]
FengZZ, LiP, YuanXY, GaoF, JiangLJ, DaiLL (2018). Progress in ecological and environmental effects of ground-level O3 in China Acta Ecologica Sinica, 38, 1530-1541. [本文引用: 6]
FengZZ, LiuXJ, ZhangFS (2015b). Air pollution affects food security in China: taking ozone as an example Frontiers of Agricultural Science and Engineering, 2, 152-158. [本文引用: 1]
FengZZ, TangHY, UddlingJ, PleijelH, KobayashiK, ZhuJG, OueH, GuoW (2012). A stomatal ozone flux-response relationship to assess ozone-induced yield loss of winter wheat in subtropical China Environmental Pollution, 164, 16-23. URLPMID:22310057 [本文引用: 1]
FengZZ, UddlingJ, TangHY, ZhuJG, KobayashiK (2018b). Comparison of crop yield sensitivity to ozone between open-top chamber and free-air experiments Global Change Biology, 24, 2231-2238. DOI:10.1111/gcb.14077URLPMID:29393991 [本文引用: 1] Assessments of the impacts of ozone (O3 ) on regional and global food production are currently based on results from experiments using open-top chambers (OTCs). However, there are concerns that these impact estimates might be biased due to the environmental artifacts imposed by this enclosure system. In this study, we collated O3 exposure and yield data for three major crop species-wheat, rice, and soybean-for which O3 experiments have been conducted with OTCs as well as the ecologically more realistic free-air O3 elevation (O3 -FACE) exposure system; both within the same cultivation region and country. For all three crops, we found that the sensitivity of crop yield to the O3 metric AOT40 (accumulated hourly O3 exposure above a cut-off threshold concentration of 40 ppb) significantly differed between OTC and O3 -FACE experiments. In wheat and rice, O3 sensitivity was higher in O3 -FACE than OTC experiments, while the opposite was the case for soybean. In all three crops, these differences could be linked to factors influencing stomatal conductance (manipulation of water inputs, passive chamber warming, and cultivar differences in gas exchange). Our study thus highlights the importance of accounting for factors that control stomatal O3 flux when applying experimental data to assess O3 impacts on crops at large spatial scales.
FuhrerJ (2009). Ozone risk for crops and pastures in present and future climates Naturwissenschaften, 96, 173-194. [本文引用: 1]
FuhrerJ, Shariat-MadariH, PerlerR, TschannenW, GrubA (1994). Effects of ozone on managed pasture: II. Yield, species composition, canopy structure, and forage quality Environmental Pollution, 86, 307-314. DOI:10.1016/0269-7491(94)90171-6URLPMID:15091622 [本文引用: 1] Managed pasture composed of grasses, clover and weeds was exposed in open-top chambers to different levels of ozone (O(3)) during two consecutive seasons to study changes in yield, species composition, canopy structure, and forage quality. The pasture was established in 1990 and exposed in 1991 and 1992. Ozone treatments included charcoal-filtered air (CF), non-filtered air (NF), and two treatments with O(3) added to NF air during periods with global radiation >/= 400 W m(-2) (NF(+), NF(++)). The ratio between the 2-year cumulative, radiation-weighted O(3) concentration in ambient air (= 365 microl litre(-1) h) and in the different treatments was 0.50 (CF), 0.85 (NF), 1.11 (NF(+)), and 1.64 (NF(++)). Plots were harvested four times in 1991, and five times in 1992. The total forage yield for both seasons was modified little by O(3). The yield reduction in NF(++) was only 10% as compared to the CF treatment. Also, only marginal changes were observed in forage quality (Ca, crude protein, crude fibre), and in leaf area index and fractional light penetration. Ozone strongly reduced the yield of clover (Trifolium repens L. and Trifolium pratense L.). The O(3)-effect on clover growth was small after the first harvest and increased with each growth period. In NF, the 2-year cumulative clover yield was reduced by 24% relative to CF. In NF(++), clover growth almost ceased near the end of the second season. The reduction in clover yield with increasing O(3) was associated with a slight increase in the yield of grasses (mainly Dacytlis glomerata L). The increase in the proportion of invading species (weeds or herbs) (Taraxacum officinale L.) during the experiment was not significantly affected by O(3). A second order polynomial function was fitted to the data to establish an exposure-response model for the cumulative clover yield and the cumulative, radiation-weighted O(3) dose, and linear models were developed for total forage mass, grass yield and yield of weeds. Reducing O(3) from elevated levels (NF(+) and NF(++)) during the first season to near-ambient levels (NF) during the second season resulted in a significant recovery of clover yield after two re-growth periods. It is concluded that continuous exposure to ambient levels of O(3) negatively affects the yield of clover in frequently cut, managed pasture, but because of the relatively small proportion of clover, the shift in species composition only marginally affects total forage yield and forage quality. It is emphasised, however, that limitations of the experimental system must be taken into account before extrapolations to real field situations can be made.
FuhrerJ, SkarbyL, AshmoreM (1997). Critical levels for ozone effects on vegetation in Europe Environmental Pollution, 97, 91-106. [本文引用: 1]
FuhrerJ, Val MartinM, MillsG, HealdCL, HarmensH, HayesF, SharpsK, BenderJ, AshmoreMR (2016). Current and future ozone risks to global terrestrial biodiversity and ecosystem processes Ecology and Evoluton, 6, 8785-8799. [本文引用: 5]
GaoF (2018). Effects of Ozone Pollution and Drought Stress on Growth Regulatory Mechanism of Poplar Saplings PhD dissertation, Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing. 7-8. [本文引用: 4]
GaoF, CalatayudV, García-BreijoF, Reig-ArminanaJ, FengZZ (2016). Effects of elevated ozone on physiological, anatomical and ultrastructural characteristics of four common urban tree species in China Ecological Indicators, 67, 367-379. [本文引用: 1]
GaoF, CatalayudV, PaolettiE, HoshikY, FengZZ (2017). Water stress mitigates the negative effects of ozone on photosynthesis and biomass in poplar plants Environmental Pollution, 230, 268-279. URLPMID:28666133 [本文引用: 3]
GessnerMO, SwanCM, DangCK, McKieBG, BardgetRD, WallDH, H?tenschwilerS (2010). Diversity meets decompositon Trends in Ecology and Evoluton, 25, 372-380. [本文引用: 1]
GhimireRP, KasurinenA, H?iki?E, HolopainenJK, Julkunen-TiittoR, HolopainenT, Kivim?enp??M (2018). Combined effects of elevated ozone, temperature, and nitrogen on stem phenolic concentrations of Scots pine (Pinus sylvestris) seedlings Canadian Journal of Forest Research, 49, 246-255. [本文引用: 1]
GrantzDA, GunnS, VuHB (2006). O3 impacts on plant development: a meta-analysis of root/shoot allocation and growth Plant, Cell & Environment, 29, 1193-1209. [本文引用: 1]
GrulkeNE, HeathRL (2020). Ozone effects on plants in natural ecosystems Plant Biology, 22, 12-37. URLPMID:30730096 [本文引用: 6]
GrulkeNE, MinnichRA, PaineTD, SeyboldSJ, ChavezDJ, FennME, RigganPJ, DunnA (2009). Chapter 17 Air pollution increases forest susceptibility to wildfires: a case study in the San Bernardino Mountains in southern California Developments in Environmental Science, 8, 365-403. [本文引用: 1]
GuoJP, WangCY, WenM, BaiYM (2003). Study on the impacts of ozone concentration on vegetables Chinese Journal of Eco-Agriculture, 11(2), 18-20. [本文引用: 1]
HandleyT, GrulkeNE (2008). Interactive effects of O3 exposure on California black oak (Quercus kelloggii Newb.) seedlings with and without N amendment Environmental Pollution, 156, 53-60. [本文引用: 1]
HayesF, MillsG, AshmoreM (2009). Effects of ozone on inter- and intra-species competition and photosynthesis in mesocosms of Lolium perenne and Trifolium repens Environmental Pollution, 157, 208-214. [本文引用: 1]
HayesF, MillsG, JonesL, AshmoreM (2010). Does a simulated upland grassland community respond to increasing background, peak or accumulated exposure of ozone? Atmospheric Environment, 44, 4155-4164. [本文引用: 1]
HeagleAS, BodyDE, HeckWW (1973). An open-top field chamber to assess the impact of air pollution on plants Journal of Environmental Quality, 2, 365-368. [本文引用: 1]
HeagleAS, PhilbeckRB, RogersHH, LetchworthMB (1979). Dispensing and monitoring ozone in open-top field chambers for plant-effects studies Phytopathology, 69, 15-20. [本文引用: 1]
HendreyGR, KimballB (1994). The FACE program Agricultural and Forest Meteorology, 70, 3-14. [本文引用: 1]
HewittCN, MacKenzieAR, Di CarloP, Di MarcoCF, DorseyJR, EvansM, FowlerD, GallagherMW, HopkinsJR, JonesCE, LangfordB, LeeJD, LewisAC, LimSF, McQuaidJ, MisztalP, MollerSJ, MonksPS, NemitzE, OramDE, OwenSM, PhillipsGJ, PughTAM, PyleJA, ReevesCE, RyderJ, SiongJ, SkibaU, StewartDJ (2009). Nitrogen management is essential to prevent tropical oil palm plantations from causing ground-level ozone pollution Proceedings of the National Academy of Sciences of the United States of America, 106, 18447-18451. DOI:10.1073/pnas.0907541106URLPMID:19841269 [本文引用: 1] More than half the world's rainforest has been lost to agriculture since the Industrial Revolution. Among the most widespread tropical crops is oil palm (Elaeis guineensis): global production now exceeds 35 million tonnes per year. In Malaysia, for example, 13% of land area is now oil palm plantation, compared with 1% in 1974. There are enormous pressures to increase palm oil production for food, domestic products, and, especially, biofuels. Greater use of palm oil for biofuel production is predicated on the assumption that palm oil is an
HofstraG, AliA, WukaschRT, FletcherRA (1981). The rapid inhibition of root respiration after exposure of bean (Phaseolus vulgaris L.) plants to ozone Atmospheric Environment, 15, 483-487. [本文引用: 1]
HoshikaY, WatanabeM, InadaN, KoikeT (2012). Ozone-induced stomatal sluggishness develops progressively in Siebold’s beech (Fagus crenata) Environmental Pollution, 166, 152-156. DOI:10.1016/j.envpol.2012.03.013URLPMID:22504428 [本文引用: 1] We investigated the effects of ozone and leaf senescence on steady-state stomatal conductance and stomatal response to light variation. Measurements were carried out in a free-air ozone exposure experiment on a representative deciduous broadleaved tree species in Japan (Fagus crenata). Both steady-state and dynamic stomatal response to light variation varied intrinsically with season due to leaf senescence. Ozone induced the decrease in steady-state leaf gas exchange and the sluggish stomatal closure progressively. These findings suggest that ozone reduces the ability of plants to adapt to a fluctuating light environment under natural conditions, and therefore impairs plant growth and ability to control water loss.
HuEZ, GaoF, XinY, JiaHX, LiKH, HuJJ, FengZZ (2015). Concentration- and flux-based ozone dose-response relationships for five poplar clones grown in North China Environmental Pollution, 207, 21-30. [本文引用: 1]
ICPVegetation (2011). Ozone pollution: a hidden threat to food security. Programme Coordination Centre for the ICP Vegetation//Mills G, Harmens H. Programme Coordination Centre for the ICP Vegetation NERC/Centre for Ecology and Hydrology, Bangor, UK.116. [本文引用: 1]
ICPVegetation (2013). Ozone pollution: impacts on ecosystem services and biodiversity. Programme Coordination Centre for the ICP Vegetation//Mills G, Wagg S, Harmens H. Programme Coordination Centre for the ICP Vegetation NERC/Centre for Ecology and Hydrology, Bangor, UK.104. [本文引用: 2]
JiaYL (2016). Effects of Ozone Stress on Grain Yield, Quality and Plant Lodging Resistance of Different Wheat Varieties Masters degree dissertation, Yangzhou University, Yangzhou, Jiangsu. 21-38. [本文引用: 2]
Kangasj?rviJ, JaspersP, KollistH (2005). Signalling and cell death in ozone-exposed plants Plant, Cell & Environment, 28, 1021-1036. [本文引用: 1]
KarnoskyDF, PregitzerKS, ZakDR, KubiskeME, HendreyGR, WeinsteinD, NosalM, PercyKE (2005). Scaling ozone responses of forest trees to the ecosystem level in a changing climate Plant, Cell & Environment, 28, 965-981. [本文引用: 2]
KarnoskyDF, ZakDR, PregitzerKS, AwmackCS, BockheimJG, DicksonRE, HendreyGR, HostGE, KingJS, KopperBJ, KrugerEL, KubiskeME, LindrothRL, MattsonWFJ, McdonaldEP, NoormetsA, OksanenE, ParsonsWFJ, PercyKE, PodileGK, RiemenschneiderDE, SharmaP, ThakurR, S?berA, S?berJ, JonesWS, AnttonenS, VapaavuoriE, MankovskaB, HeilmanW, IsebrandsJG (2003). Tropospheric O3 moderates responses of temperate hardwood forests to elevated CO2: a synthesis of molecular to ecosystem results from the Aspen FACE project Functional Ecology, 17, 289-394. [本文引用: 2]
KasurinenA, PeltonenPA, HolopainenJK, VapaavuoriE, HolopainenT (2007). Leaf litter under changing climate: Will increasing levels of CO2 and O3 affect decomposition and nutrient cycling processes? Dynamic Soil, Dynamic Plant, 1, 58-67. [本文引用: 1]
KasurinenA, RiikonenJ, OksanenE, VapaavuoriV, HolopainenT (2006). Chemical composition and decomposition of silver birch leaf litter produced under elevated CO2 and O3 Plant and Soil, 282, 261-280. [本文引用: 1]
KingJS, KubiskeME, PregitzerKS, HendreyGR, McDonaldEP, GiardinaCP, QuinnV, KarnoskyD (2005). Tropospheric O3 compromises net primary production in young stands of trembling aspen, paper birch and sugar maple in response to elevated atmospheric CO2 New Phytologist, 168, 623-636. URLPMID:16313645 [本文引用: 1]
KopperBJ, LindrothRL (2003). Effects of elevated carbon dioxide and ozone on the phytochemistry of aspen and performance of an herbivore Oecologia, 134, 95-103. DOI:10.1007/s00442-002-1090-6URLPMID:12647186 [本文引用: 1] The purpose of this study was to assess the independent and interactive effects of CO(2), O(3), and plant genotype on the foliar quality of a deciduous tree and the performance of a herbivorous insect. Two trembling aspen (Populus tremuloides Michaux) genotypes differing in response to CO(2) and O(3) were grown at the Aspen FACE (Free Air CO(2) Enrichment) site located in northern Wisconsin, USA. Trees were exposed to one of four atmospheric treatments: ambient air (control), elevated carbon dioxide (+CO(2); 560 microl/l), elevated ozone (+O(3); ambient x1.5), and elevated CO(2)+O(3). We measured the effects of CO(2) and O(3) on aspen phytochemistry and on performance of forest tent caterpillar (Malacosoma disstria Hubner) larvae. CO(2) and O(3) treatments influenced foliar quality for both genotypes, with the most notable effects being that elevated CO(2) reduced nitrogen and increased tremulacin levels, whereas elevated O(3) increased early season nitrogen and reduced tremulacin levels, relative to controls. With respect to insects, the +CO(2) treatment had little or no effect on larval performance. Larval performance improved in the +O(3) treatment, but this response was negated by the addition of elevated CO(2) (i.e., +CO(2)+O(3) treatment). We conclude that tent caterpillars will have the greatest impact on aspen under current CO(2) and high O(3) levels, due to increases in insect performance and decreases in tree growth, whereas tent caterpillars will have the least impact on aspen under high CO(2) and low O(3) levels, due to moderate changes in insect performance and increases in tree growth.
KubiskeME, QuinnVS, MarquardtPE, KarnoskyDF (2007). Effects of elevated atmospheric CO2 and/or O3 on intra- and interspecific competitive ability of aspen Plant Biology, 9, 342-355. [本文引用: 2]
LarsonJL, ZakDR, SinsabaughRL (2002). Extracellular enzyme activity beneath temperate trees growing under elevated carbon dioxide and ozone Soil Science Society of America Journal, 66, 1848-1856. [本文引用: 1]
LeedsAR (1880). Lines of discovery in the history of ozone Annals of the New York Academy of Sciences, 1, 363-391. [本文引用: 1]
LefohnAS, FoleyJK (1992). NCLAN results and their application to the standard setting process: protecting vegetation from surface ozone exposures Journal of the Air and Waste Management Association, 42, 1046-1052. [本文引用: 1]
LefohnAS, MalleyCS, SmithL, WellsB, HazuchaM, SimonH, NaikV, MillsG, SchultzMG, PaolettiE, De MarcoA, XuXB, ZhangL, WangT, NeufeldHS, MusselmanRC, TarasickD, BrauerM, FengZZ, TangHY, KobayashiK, SicardP, SolbergS, GerosaG (2018). Tropospheric ozone assessment report: global ozone metrics for climate change, human health, and crop/ecosystem research Elementa Science of the Anthropocene, 6, 28. DOI: 10.1525/elementa.279. [本文引用: 1]
LeggeAH, GrünhageL, NoalM, J?gerHJ, KrupaSV (1995). Ambient ozone and adverse crop response: an evaluation of north American and European data as they relate to exposure indices and critical levels Journal of Applied Botany, 69, 192-205. [本文引用: 1]
LiP, CalatayudV, GaoF, UddlingJ, FengZZ (2016). Differences in ozone sensitivity among woody species are related to leaf morphology and antioxidant levels Tree Physiology, 36, 1105-1116. [本文引用: 1]
LiP, FengZZ, CalatayudV, YuanXY, XuYS, PaolettiE (2017). A meta-analysis on growth, physiological, and biochemical responses of woody species to ground-level ozone highlights the role of plant functional types Plant, Cell & Environment, 40, 2369-2380. URLPMID:28744869 [本文引用: 1]
LiP, FengZZ, ShangB, YuanXY, DaiLL, XuYS (2018). Stomatal characteristics and ozone dose-response relationships for six greening tree species Acta Ecologica Sinica, 38, 2710-2721.
LinYY, JiangF, ZhaoJ, ZhuG, HeXJ, MaXL, LiS, SabelCE, WangHK (2018). Impacts of O3 on premature mortality and crop yield loss across China Atmospheric Environment, 194, 41-47. [本文引用: 1]
LindrothRL (2010). Impacts of elevated atmospheric CO2 and O3 on forests: phytochemistry, trophic interactions, and ecosystem dynamics Journal of Chemical Ecology, 36, 2-21. URLPMID:20054619 [本文引用: 1]
LongSP, AinsworthEA, LeakeyADB, MorganPB (2005). Global food insecurity. Treatment of major food crops with elevated carbon dioxide or ozone under large-scale fully open-air conditions suggests recent models may have overestimated future yields Philosophical Transactions of the Royal Society B: Biological Sciences, 360, 2011-2020. [本文引用: 1]
LRTARConvention (2015). Draft Chapter III: Mapping Critical Levels for Vegetation, of the Manual on Methodologies and Criteria for Modelling and Mapping Critical Loads and Levels and Air Pollution Effects, Risks and Trends. http://icpmapping.org/Mapping_Manual. Cited: 2019-06-13. URL [本文引用: 2]
LuX, HongJY, ZhangL, CooperOR, SchultzMG, XuXB, WangT, GaoM, ZhaoYH, ZhangYH (2018). Severe surface ozone pollution in China: a global perspective Environmental Science and Technology Letters, 5, 487-494. [本文引用: 1]
LuweM (1996). Antioxidants in the apoplast and symplast of beech (Fagus sylvatica L.) leaves: seasonal variations and responses to changing ozone concentrations in air Plant, Cell & Environment, 19, 321-328. [本文引用: 1]
MandleRH (1973). A cylindrical open top chamber for the exposure of plants to air pollutants in the field Journal of Environmental Quality, 2, 371-376. [本文引用: 1]
MatyssekR, BytnerowiczA, KarlssonPE, PaolettiE, SanzeM, SchaubM, WieserG (2007). Promoting the O3 flux concept for European forest trees Environmental Pollution, 146, 587-607. [本文引用: 1]
MatyssekR, WieserG, CeulemansR, RennenbergH, PretzschH, HabererK, LowM, NunnAJ, WernerH, WipflerP, OsswaldW, NikolovaPS, HankeDE, KraigerH, TauszM, BahnwegG, KitaoM, DielerJ, SandermannH, HerbingerK, GrebencT, BlumenrotherM, DeckmynG, GramsTEE, HeerdtC, LeuchnerM, FabianP, HaberleKH (2010). Enhanced ozone strongly reduces carbon sink strength of adult beech (Fagus sylvatica)—Resume from the free-air fumigation study at Kranzberg forest Environmental Pollution, 158, 2527-2532. DOI:10.1016/j.envpol.2010.05.009URLPMID:20570421 [本文引用: 2] Ground-level ozone (O(3)) has gained awareness as an agent of climate change. In this respect, key results are comprehended from a unique 8-year free-air O(3)-fumigation experiment, conducted on adult beech (Fagus sylvatica) at Kranzberg Forest (Germany). A novel canopy O(3) exposure methodology was employed that allowed whole-tree assessment in situ under twice-ambient O(3) levels. Elevated O(3) significantly weakened the C sink strength of the tree-soil system as evidenced by lowered photosynthesis and 44% reduction in whole-stem growth, but increased soil respiration. Associated effects in leaves and roots at the gene, cell and organ level varied from year to year, with drought being a crucial determinant of O(3) responsiveness. Regarding adult individuals of a late-successional tree species, empirical proof is provided first time in relation to recent modelling predictions that enhanced ground-level O(3) can substantially mitigate the C sequestration of forests in view of climate change.
McDonaldEP, KrugerEL, RiemenschneiderDE, IsebrandsJG (2002). Competitive status influences tree-growth responses to elevated CO2 and O3 in aggrading aspen stands Functional Ecology, 16, 792-801. [本文引用: 1]
McLaughlinSB, WullschlegerSD, SunG, NosalM (2007). Interactive effects of ozone and climate on water use, soil moisture content and streamflow in a southern Appalachian forest in the USA New Phytologist, 174, 125-136. [本文引用: 1]
MillsG, BuseA, GimenoB, BermejoV, HollandM, EmbersonL, PleijelH (2007). A synthesis of AOT40-based response functions and critical levels of ozone for agricultural and horticultural crops Atmospheric Environment, 41, 2630-2643. [本文引用: 1]
MillsG, HayesF, SimpsonD, EmbersonL, NorrisD, HarmensH, BukerP (2011). Evidence of widespread effects of ozone on crops and (semi-) natural vegetation in Europe (1990-2006) in relation to AOT40- and flux-based risk maps Global Change Biology, 17, 592-613. [本文引用: 1]
MillsG, SharpsK, SimpsonD, PleijelH, BrobergM, UddlingJ, JaramilloF, DaviesWJ, DentenerF, van den BergM, AgrawalM, AgrawalSB, AinsworthEA, BukerP, EmbersonL, FengZZ, HarmensH, HayesF, KobayashiK, PaolettiE, DingenenRV (2018). Ozone pollution will compromise efforts to increase global wheat production Global Change Biology, 24, 3560-3574. DOI:10.1111/gcb.14157URLPMID:29604158 [本文引用: 3] Introduction of high-performing crop cultivars and crop/soil water management practices that increase the stomatal uptake of carbon dioxide and photosynthesis will be instrumental in realizing the United Nations Sustainable Development Goal (SDG) of achieving food security. To date, however, global assessments of how to increase crop yield have failed to consider the negative effects of tropospheric ozone, a gaseous pollutant that enters the leaf stomatal pores of plants along with carbon dioxide, and is increasing in concentration globally, particularly in rapidly developing countries. Earlier studies have simply estimated that the largest effects are in the areas with the highest ozone concentrations. Using a modelling method that accounts for the effects of soil moisture deficit and meteorological factors on the stomatal uptake of ozone, we show for the first time that ozone impacts on wheat yield are particularly large in humid rain-fed and irrigated areas of major wheat-producing countries (e.g. United States, France, India, China and Russia). Averaged over 2010-2012, we estimate that ozone reduces wheat yields by a mean 9.9% in the northern hemisphere and 6.2% in the southern hemisphere, corresponding to some 85 Tg (million tonnes) of lost grain. Total production losses in developing countries receiving Official Development Assistance are 50% higher than those in developed countries, potentially reducing the possibility of achieving UN SDG2. Crucially, our analysis shows that ozone could reduce the potential yield benefits of increasing irrigation usage in response to climate change because added irrigation increases the uptake and subsequent negative effects of the pollutant. We show that mitigation of air pollution in a changing climate could play a vital role in achieving the above-mentioned UN SDG, while also contributing to other SDGs related to human health and well-being, ecosystems and climate change.
MorganPB, BernacchiCJ, OrtDR, LongSP (2004). An in vivo analysis of the effect of season-long open-air elevation of ozone to anticipated 2050 levels on photosynthesis in soybean Plant Physiology, 135, 2348-2357. DOI:10.1104/pp.104.043968URLPMID:15299126 [本文引用: 1] Rising atmospheric carbon dioxide concentration ([CO(2)]) is widely recognized, but less appreciated is a concomitant rise in tropospheric ozone concentration ([O(3)]). In industrialized countries, [O(3)] has risen by 0.5% to 2.5% per year. Tropospheric [O(3)] is predicted to reach a global mean of >60 nL L(-1) by 2050 with greater averages locally. Previous studies in enclosures suggest that this level of [O(3)] will decrease leaf photosynthesis, thereby limiting growth and yield of Glycine max L. Merr. SoyFACE (Soybean Free Air gas Concentration Enrichment) is the first facility to elevate atmospheric [O(3)] (approximately 1.2x current) in replicated plots under completely open-air conditions within an agricultural field. Measurements of gas exchange (assimilation versus light and assimilation versus intercellular [CO(2)]) were made on excised leaves from control and treatment plots (n = 4). In contrast to expectations from previous chamber studies, elevated [O(3)] did not alter light-saturated photosynthesis (A(sat), P = 0.09), carboxylation capacity (V(c,max), P = 0.82), or maximum electron transport (J(max), P = 0.66) for the topmost most recently fully expanded leaf at any stage of crop development. Leaves formed during the vegetative growth stage did not show a significant ozone-induced loss of photosynthetic capacity as they aged. Leaves formed during flowering did show a more rapid loss of photosynthetic capacity as they aged in elevated [O(3)]. A(sat), V(c,max), and J(max) (P = 0.04, 0.004, and 0.002, respectively) were decreased 20% to 30% by treatment with ozone. This is noteworthy since these leaves provide photosynthate to the developing grain. In conclusion, a small (approximately 20%) increase in tropospheric [O(3)] did not significantly alter photosynthetic capacity of newly expanded leaves, but as these leaves aged, losses in photosynthetic carbon assimilation occurred.
MusselmanRC, LefohnAS, MassmanWJ, HeathRL (2006). A critical review and analysis of the use of exposure- and flux-based ozone indices for predicting vegetation effects Atmospheric Environment, 40, 1869-1888. [本文引用: 1]
OksanenE (2003). Responses of selected birch (Betula pendula) clones to ozone change over time Plant, Cell & Environment, 26, 875-886. DOI:10.1046/j.1365-3040.2003.01020.xURLPMID:12803615 [本文引用: 1] A long-term free air ozone fumigation experiment was conducted to study changes in physiological ozone responses during tree ontogeny and exposure time in ozone sensitive and tolerant clones of European white birch (Betula pendula Roth), originated from south and central Finland. The trees were grown in soil in natural microclimatic conditions under ambient ozone (control) and 1.4-1.7 x ambient (elevated) ozone from May 1996 to October 2001, and were measured for stem and foliage growth, net photosynthesis, stomatal conductance, stomatal density, visible injuries, foliar starch content and bud formation. After 6 years of exposure, the magnitude of ozone-induced growth reductions in the sensitive clone was 12-48% (significant difference), levels similar or greater than those reported earlier for 2- and 3-year-old saplings undergoing shorter exposures. In the tolerant clone, growth of these larger trees was reduced by 1-38% (significant difference in stem volume), although the saplings had previously been unaffected. In both clones, ozone stress led to significantly reduced leaf-level net photosynthesis but significantly increased stomatal conductance rates during the late summer, resulting in a lower carbon gain for bud formation and the onset of visible foliar injuries. Increasing ozone sensitivity with duration of exposure was explained by a change in growth form (relatively reduced foliage mass), a lower photosynthesis to stomatal conductance ratio during the late summer, and deleterious carry-over effects arising from the reduced number of over-wintering buds.
OllingerSV, AberJD, ReichPB, FreuderRJ (2002). Interactive effects of nitrogen deposition, tropospheric ozone, elevated CO2 and land use history on the carbon dynamics of northern hardwood forests Global Change Biology, 8, 545-562. [本文引用: 2]
PaolettiE (2009). Ozone and urban forests in Italy Environmental Pollution, 157, 1506-1512. DOI:10.1016/j.envpol.2008.09.019URLPMID:18977568 [本文引用: 1] Ozone levels along urban-to-rural gradients in three Italian cities (Milan, Florence, Bari) showed that average AOT40 values at rural and suburban sites were 2.6 times higher than those determined at urban sites. However, O(3) also exceeded the European criteria to protect forest health at urban sites, even when the standards for human health protection were met. For protecting street trees in Mediterranean cities, the objectives of measurement at urban sites should extend from the protection of human health to the protection of vegetation as well. A review of forest effects on O(3) pollution and of O(3) pollution on forest conditions in Italian cities showed that it was not possible to distinguish the effect of O(3) in the complex mixture of urban pollutants and stressors. A preliminary list of tree species for urban planning in the Mediterranean area shows the average tree capacity of O(3) removal and VOC emission.
PaolettiE, MaterassiA, FasanoG, HoshikaY, CarrieroG, SilaghiD, BadeaO (2017). A new-generation 3D ozone FACE (Free Air Controlled Exposure) Science of the Total Environment, 575, 1407-1414. [本文引用: 2]
PellEJ, PearsonNS (1984). Ozone-induced reduction in quantity and quality of two potato cultivars Environmental Pollution, 35, 345-352. [本文引用: 1]
PellegriniE, CampanellaA, CotrozziL, TonelliM, NaliC, LorenziniG (2018). What about the detoxification mechanisms underlying ozone sensitivity in Liriodendron tulipifera Environmental Science and Pollution Research, 25, 8148-8160. [本文引用: 1]
PollastriniM, DesotgiuR, CascioC, BussottiF, CherubiniP, SaurerM, GerosaG, MarzuoliR (2010). Growth and physiological responses to ozone and mild drought stress of tree species with different ecological requirements Trees, 24, 695-704. [本文引用: 1]
PretzschH, DielerJ, MatyssekR, WipflerP (2010). Tree and stand growth of mature Norway spruce and European beech under long-term ozone fumigation Environmental Pollution, 158, 1061-1070. DOI:10.1016/j.envpol.2009.07.035URLPMID:19713019 [本文引用: 1] In a 50- to 70-year-old mixed stand of Norway spruce (Picea abies (L.) Karst.) and European beech (Fagus sylvatica L.) in Germany, tree cohorts have been exposed to double ambient ozone (2xO(3)) from 2000 through 2007 and can be compared with trees in the same stand under the ambient ozone regime (1xO(3)). Annual diameter growth, allocation pattern, stem form, and stem volume were quantified at the individual tree and stand level. Ozone fumigation induced a shift in the resource allocation into height growth at the expense of diameter growth. This change in allometry leads to rather cone-shaped stem forms and reduced stem stability in the case of spruce, and even neiloidal stem shapes in the case of beech. Neglect of such ozone-induced changes in stem shape may lead to a flawed estimation of volume growth. On the stand level, 2xO(3) caused, on average, a decrease of 10.2 m(3) ha(-1) yr(-1) in European beech.
ReichPR (1987). Quantifying plant response to ozone: a unifying theory Tree Physiology, 3, 63-91. DOI:10.1093/treephys/3.1.63URLPMID:14975835 [本文引用: 1] Published information about the effects of ozone on plants and ecosystems is synthesized into a conceptual model to explain the response of evergreen conifers, deciduous hardwoods and agricultural crops to ambient levels of ozone pollution. The effects of ozone on carbon balance and growth of individual plants can be quantified on the basis of concentration, external dose (concentration x duration of exposure), or uptake. For an equivalent dose within a single growing season, agricultural crops are the most sensitive to ozone, with hardwoods intermediate and conifers least sensitive. In contrast, all species display a similar decline in photosynthesis and growth in response to equivalent total uptake or uptake per leaf life span, with trees somewhat less sensitive than agricultural crops on a calendar time scale, but slightly more sensitive on a relative (leaf life span) time scale. Among species, differences in ozone uptake and response can be predicted from differences in the inherent leaf diffusive conductance.
RenW, TianHQ, TaoB, ChappelkaAH, SunG, LuCQ, LiuML, ChenGS, XuXF (2011). Impacts of tropospheric ozone and climate change on net primary productivity and net carbon exchange of China’s forest ecosystems Global Ecology and Biogeography, 20, 391-406. [本文引用: 1]
RichardsBL, MiddletonJT, HewittWB (1958). Air pollution with relation to agronomic crops: V. Oxidant stipple of grape Journal of the American Society of Agronomy, 50, 559-560. [本文引用: 1]
RogersLH, RenzettiNA, NeiburgerM (1956). Smog effects and chemical analysis of the Los Angeles atmosphere Journal of the Air Pollution Control Association, 6, 165-170. [本文引用: 1]
SawadaH, TsukaharaK, KohnoY, SuzukiK, NagasawaN, MasanoriT (2016). Elevated ozone deteriorates grain quality of Japonica Rice cv. Koshihikari, even if it does not cause yield reduction Rice, 9, 1-10. [本文引用: 1]
SchloterM, WinklerJB, AnejaM, KochN, FleischmannF, PritschK, HellerW, StichS, GramsTE, G?ttleinA, MatyssekR, MunchJC (2005). Short term effects of ozone on the plant-rhizosphere-bulk soil system of young beech trees Plant Biology, 7, 728-736. [本文引用: 1]
Schmadel-Hageb?llingHE, EngelC, SchmittV, WildA (1998). The combined effects of CO2, ozone and drought on rubisco and nitrogen metabolism of young oak trees (Quercus petraea)—A phytotron study Chemosphere, 36, 789-794. [本文引用: 1]
SicardP, AgathokleousE, AraminieneV, CarrariE, HoshikaY, de MarcoA, PaolettiE (2018). Should we see urban trees as effective solutions to reduce increasing ozone levels in cities? Environmental Pollution, 243, 163-176. [本文引用: 1]
SinghAA, AgrawalSB (2017). Tropospheric ozone pollution in India: effects on crop yield and product quality Environmental Science and Pollution Research, 24, 4367-4382. DOI:10.1007/s11356-016-8178-8URLPMID:27943144 [本文引用: 1] Ozone (O3) in troposphere is the most critical secondary air pollutant, and being phytotoxic causes substantial losses to agricultural productivity. Its increasing concentration in India particularly in Indo-Gangetic plains is an issue of major concern as it is posing a threat to agriculture. In view of the issue of rising surface level of O3 in India, the aim of this compilation is to present the past and the prevailing concentrations of O3 and its important precursor (oxides of nitrogen) over the Indian region. The resulting magnitude of reductions in crop productivity as well as alteration in the quality of the product attributable to tropospheric O3 has also been taken up. Studies in relation to yield measurements have been conducted predominantly in open top chambers (OTCs) and also assessed by using antiozonant ethylene diurea (EDU). There is a substantial spatial difference in O3 distribution at different places displaying variable O3 concentrations due to seasonal and geographical variations. This review further recognizes the major information lacuna and also highlights future perspectives to get the grips with rising trend of ground level O3 pollution and also to formulate the policies to check the emissions of O3 precursors in India.
SitchS, CoxPM, CollinsWJ, HuntingfordC (2007). Indirect radiative forcing of climate change through ozone effects on the land-carbon sink Nature, 448, 791-794. DOI:10.1038/nature06059URLPMID:17653194 [本文引用: 2] The evolution of the Earth's climate over the twenty-first century depends on the rate at which anthropogenic carbon dioxide emissions are removed from the atmosphere by the ocean and land carbon cycles. Coupled climate-carbon cycle models suggest that global warming will act to limit the land-carbon sink, but these first generation models neglected the impacts of changing atmospheric chemistry. Emissions associated with fossil fuel and biomass burning have acted to approximately double the global mean tropospheric ozone concentration, and further increases are expected over the twenty-first century. Tropospheric ozone is known to damage plants, reducing plant primary productivity and crop yields, yet increasing atmospheric carbon dioxide concentrations are thought to stimulate plant primary productivity. Increased carbon dioxide and ozone levels can both lead to stomatal closure, which reduces the uptake of either gas, and in turn limits the damaging effect of ozone and the carbon dioxide fertilization of photosynthesis. Here we estimate the impact of projected changes in ozone levels on the land-carbon sink, using a global land carbon cycle model modified to include the effect of ozone deposition on photosynthesis and to account for interactions between ozone and carbon dioxide through stomatal closure. For a range of sensitivity parameters based on manipulative field experiments, we find a significant suppression of the global land-carbon sink as increases in ozone concentrations affect plant productivity. In consequence, more carbon dioxide accumulates in the atmosphere. We suggest that the resulting indirect radiative forcing by ozone effects on plants could contribute more to global warming than the direct radiative forcing due to tropospheric ozone increases.
SunG, MclaughlinSB, PorterJH, UddlingJ, MulhollandPJ, AdamsMB, PedersonN (2012). Interactive influences of ozone and climate on streamflow of forested watersheds Global Change Biology, 18, 3395-3409. [本文引用: 1]
TaiAPK, Val MartinM, HealdCL (2014). Threat to future global food security from climate change and ozone air pollution Nature Climate Change, 4, 817-821. [本文引用: 1]
TangHY, LiuG, HanY, ZhuJG, KobayashiK (2011). A system for free-air ozone concentration elevation with rice and wheat: control performance and ozone exposure regime Atmospheric Environment, 45, 6276-6282. [本文引用: 1]
TauszM, GrulkeNE, WieserG (2007). Defense and avoidance of ozone under global change Environmental Pollution, 147, 525-531. DOI:10.1016/j.envpol.2006.08.042URLPMID:17055629 [本文引用: 1] The level II approach of the critical loads concept adopted by the UNECE aims at a flux based evaluation and takes into account environmental factors governing stomatal conductance. These factors will probably be affected by global change. The flux concept predicts that a decrease in stomatal conductance would protect trees from air pollution effects by decreasing uptake. However, experimental evidence is inconclusive. Numerous results suggest that pollutants and factors subject to global change (drought, CO(2)) may interact and even exacerbate effects, probably because antioxidative defense systems are involved in both, defense against pollutant effects and protection from natural stress. An effective pollutant dose, which is weighted by physiological defense capacity, would better predict such effects. In this review paper we argue that the flux-based approach is imperfect, because global change effects may also modify the physiological susceptibility to ozone. Instead, a flux concept weighted by defense capacity should be tested.
TomerR, BhatiaA, KumarV, KumarA, SinghR, SinghB, SinghSD (2015). Impact of elevated ozone on growth, yield and nutritional quality of two wheat species in Northern India Aerosol and Air Quality Research, 15, 329-340. [本文引用: 2]
van DingenenR, DentenerFJ, RaesF, KrolMC, EmbersonL, CofalaJ (2009). The global impact of ozone on agricultural crop yields under current and future air quality legislation Atmospheric Environment, 43, 604-618.
VolkM, GeissmannM, BlatterA, ContatF, FuhrerJ (2003). Design and performance of a free-air exposure system to study long-term effects of ozone on grasslands Atmospheric Environment, 37, 1341-1350. [本文引用: 1]
WangCY (1995). Effects of ozone on crops Journal of Applied Meteorological Science, 6, 343-349. [本文引用: 1]
WangN, LyuXP, DengXJ, HuangX, JiangF, DingAJ (2019). Aggravating O3 pollution due to NOx emission control in eastern China Science of the Total Environment, 677, 732-744. DOI:10.1016/j.scitotenv.2019.04.388URLPMID:31075619 [本文引用: 1] During the past five years, China has witnessed a rapid drop of nitrogen oxides (NOx) owing to the wildly-applied rigorous emission control strategies across the country. However, ozone (O3) pollution was found to steadily deteriorate in most part of eastern China, especially in developed regions such as Jing-Jin-Ji (JJJ), Yangtze River Delta region (YRD) and Pearl River Delta region (PRD). To shed more light on current O3 pollution and its responses to precursor emissions, we integrate satellite retrievals, ground-based measurements together with regional numerical simulation in this study. It is indicated by multiple sets of observational data that NOx in eastern China has declined more than 25% from 2012 to 2016. Based on chemical transport modeling, we find that O3 formation in eastern China has changed from volatile organic compounds (VOCs) sensitive regime to the mixed sensitive regime due to NOx reductions, substantially contributing to the recent increasing trend in urban O3. In addition, such transitions tend to bring about an ~1-1.5h earlier peak of net O3 formation rate. We further studied the O3 precursors relationships by conducting tens of sensitivity simulations to explore potential ways for effective O3 mitigation. It is suggested that the past control measures that only focused on NOx may not work or even aggravate O3 pollution in the city clusters. In practice, O3 pollution in the three regions is expected to be effectively mitigated only when the reduction ratio of VOCs/NOx is greater than 2:1, indicating VOCs-targeted control is a more practical and feasible way.
WangXP, MauzerallDL (2004). Characterizing distributions of surface ozone and its impact on grain production in China, Japan and South Korea: 1990 and 2020 Atmospheric Environment, 38, 4383-4402. [本文引用: 1]
WangYX, YangLX, HanY, ZhuJG, KobayashiK, TangHY, WangYL (2012). The impact of elevated tropospheric ozone on grain quality of hybrid rice: a free-air gas concentration enrichment (FACE) experiment Field Crops Research, 129, 81-89. [本文引用: 1]
WatanabeM, HoshikaY, InadaN, WangXN, MaoQZ, KoikeT (2013). Photosynthetic traits of Siebold’s beech and oak saplings grown under free air ozone exposure Environmental Pollution, 174, 50-56. DOI:10.1016/j.envpol.2012.11.006URLPMID:23246746 [本文引用: 1] We set up a free-air ozone (O(3)) exposure system for determining the photosynthetic responses of Siebold's beech (Fagus crenata) and oak (Quercus mongolica var. crispula) to O(3) under field conditions. Ten-year-old saplings of beech and oak were exposed to an elevated O(3) concentration (60 nmol mol(-1)) during daytime from 6 August to 11 November 2011. Ozone significantly reduced the net photosynthetic rate in leaves of both species in October, by 46% for beech and 15% for oak. In beech there were significant decreases in maximum rate of carboxylation, maximum rate of electron transport in photosynthesis, nitrogen content and photosynthetic nitrogen use efficiency, but not in oak. Stomatal limitation of photosynthesis was unaffected by O(3). We therefore concluded photosynthesis in beech is more sensitive to O(3) than that in oak, and the O(3)-induced reduction of photosynthetic activity in beech was due not to stomatal closure, but to biochemical limitation.
WieserG, TegischerK, TauszM, H?berleKH, GramsTE, MatyssekR (2002). Age effects on Norway spruce (Picea abies) susceptibility to ozone uptake: a novel approach relating stress avoidance to defense Tree Physiology, 22, 583-590. DOI:10.1093/treephys/22.8.583URLPMID:12045030 [本文引用: 1] Cumulative ozone (O3) uptake and O3 flux were related to physiological, morphological and biochemical characteristics of Norway spruce (Picea abies (L.) Karst.) trees of different ages. Under ambient CO2 conditions, photosynthetic capacity (Amax) declined in mature trees when cumulative O3 uptake into needles, which provides a measure of effective O3 dose, exceeded 21 mmol m-2 of total needle surface area. A comparable decline in Amax of seedlings occurred when cumulative O(3) uptake was only 4.5 mmol m-2. The threshold O3 flux causing a significant decline in Amax ranged between 2.14 and 2.45 nmol m-2 s-1 in mature trees and seedlings subjected to exposure periods of > or = 70 and > or = 23 days, respectively. The greater O3 sensitivity of young trees compared with mature trees was associated with needle morphology. Biomass of a 100-needle sample increased significantly with tree age, whereas a negative correlation was found for specific leaf area, these changes parallel those observed during differentiation from shade-type to sun-type needles with tree ontogeny. Age-dependent changes in leaf morphology were related to changes in detoxification capacity, with area-based concentrations of ascorbate increasing during tree ontogeny. These findings indicate that the extent of O3-induced injury is related to the ratio of potentially available antioxidants to O3 influx. Because this ratio, when calculated for ascorbate, increased with tree age, we conclude that the ratio may serve as an empirical basis for characterizing age-related differences in tree responses to O3.
WittigVE, AinsworthEA, NaiduSL, KarnoskyDF, LongSP (2009). Quantifying the impact of current and future tropospheric ozone on tree biomass, growth, physiology and biochemistry: a quantitative meta-analysis Global Change Biology, 15, 396-424. [本文引用: 2]
XuYS, FengZZ, TarvainenL, ShangB, DaiLL, UddlingJ (2019). Mesophyll conductance limitation of photosynthesis in poplar under elevated ozone Science of the Total Environment, 657, 136-145. URLPMID:30537576 [本文引用: 1]
YamaguchiM, WatanabeM, MatsumuraH, KohnoY, IzutaT (2010). Effects of ozone on nitrogen metabolism in the leaves of Fagus crenata seedlings under different soil nitrogen loads Trees, 24, 175-184. [本文引用: 1]
YueX, UngerN, HarperK, XiaXG, LiaoH, ZhuT, XiaoJF, FengZZ, LiJ (2017). Ozone and haze pollution weakens net primary productivity in China Atmospheric Chemistry and Physics, 17, 6073-6089. [本文引用: 1]
ZhangRB, HuHJ, ZhaoZ, YangDD, ZhuXK, GuoWS, ZhuJG, KobayashiK (2013). Effects of elevated ozone concentration on starch and starch synthesis enzymes of yangmai 16 under fully open-air field conditions Journal of Integrative Agriculture, 12, 2157-2163. [本文引用: 1]
ZhaoH, ZhengYF, WuXY (2018). Assessment of yield and economic losses for wheat and rice due to ground level O3 exposure in the Yangtze River Delta, China Atmospheric Environment, 191, 241-248. [本文引用: 1]
A review study on past 40 years of research on effects of tropospheric O3 on belowground structure, functioning, and processes of trees: a linkage with potential ecological implications 1 2016
... 地表O3浓度升高对植物光合作用、生长和生物量积累、物质分配等过程的连锁负效应导致陆地生态系统生产力和固碳能力降低(Reich, 1987; Wittig et al., 2009; Ainsworth et al., 2012), 削弱了自然生态系统的碳汇能力(Felzer et al., 2005; Sitch et al., 2007; Collins et al., 2010).基于大量OTCs控制实验下北半球中纬度北方温带树种的meta分析发现, O3浓度升高导致树木的净光合作用和总生物量分别降低11%和7% (Wittig et al., 2009).最近, 有关中国热带、亚热带和温带树种的meta分析则发现, O3浓度升高导致中国木本植物的净光合作用和总生物量分别降低28%和14% (Li et al., 2017). ...
... (1)开展O3-FACE研究: 现有研究多为OTC系统下的实验, OTC气室内“微气候” (如温度、水分、光照和风力等生态因素)与外界环境状态存在明显差异, 这些生态因子的变化可能影响植物应对O3的响应, 缓解或加重O3污染对农作物产量或森林植被生产力的损伤(Long et al., 2005; Feng et al., 2018a).全球模型评估结果显示, OTC实验结果低估了O3对作物产量损失的影响(van Dingenen et al., 2009).最近, Feng等(2018b)也发现, 同样浓度的O3对OTC实验下小麦和水稻产量的影响要低于FACE实验; 而对大豆产量的影响则相反.因此, OTC实验结果并不能完全应用于实际O3区域风险的评估.O3-FACE平台是未来研究实际大田环境下O3浓度升高对农林生态系统影响的主要依托平台.为减少区域O3风险评估的不确定性, 未来应加大O3-FACE平台的实验研究.考虑到目前已开展的FACE实验多坐落在温带地区, 主要探究温带植被或作物对高浓度O3的响应, 较少涉及热带和寒带植物或其他区域典型生态系统(Ainsworth et al., 2020).最理想的情况是, 不同区域、不同生态系统类型共同开展O3-FACE实验研究(Ainsworth et al., 2012). ...
Source-sink balance and carbon allocation below ground in plants exposed to ozone: tansley review 2 2003
... 此外, 虽然地下生态系统并不是直接暴露在O3环境下, 但大量证据表明O3对地上生态系统的影响将会转移至地下(Andersen, 2003; Lindroth, 2010).如高浓度O3环境下, 植物分配到根的生物量将会大大减少, 基于100多组实验数据的meta分析研究表明O3暴露导致生物量分配到地下的量平均减少5.6% (Grantz et al., 2006).另外, 凋落物分解是生态系统营养循环的关键过程.通过影响凋落物的数量、质量和分解速率, 高浓度地表O3会间接改变土壤微生物群落活性、组成和结构, 土壤化学属性及土壤营养循环(Larson et al., 2002; Fuhrer et al., 2016).然而, 比较遗憾的是, 目前有关O3对地下生态过程影响的研究仅集中在农田生态系统, 仅有少部分研究涉及森林土壤(主要来自Aspen FACE)或草地生态系统(ICP Vegetation, 2013).而且, 由于凋落物种类、化学成分、分解者种群、土壤和大气环境条件的不同, 高浓度O3对凋落物分解动态的影响还存在很大争议(Gessner et al., 2010; Fuhrer et al., 2016; Grulke & Heath, 2020).例如, 研究发现O3对欧洲白桦(Betula pendula)叶片凋落物质量和损失率有明显影响(Kasurinen et al., 2006, 2007), 但不影响欧洲山毛榉凋落物的分解速率(Schloter et al., 2005).有关地表O3浓度升高对地下生态系统结构和功能的影响还有待进一步探究. ...
... 除单因子O3胁迫外, 由于全球气候变化和极端气候常态化, 植物经常受到O3和其他环境因子交互作用的影响(如O3与CO2、升温、干旱或氮沉降等)(Schmadel-Hageb?lling et al., 1998; Ollinger et al., 2002; Eltayeb et al., 2006; Ghimire et al., 2018).然而, 植物应对O3与其他因子交互作用的响应并不是两者或单独作用的简单叠加, 往往取决于植物种类、生长时期、对特定环境因子的敏感性、胁迫浓度及时间, 以及具体的微气候条件(列淦文和薛立, 2014).一方面, 通过诱导气孔关闭、限制叶片O3吸收(CO2浓度升高/干旱胁迫), 或提高碳同化速率、促进植物光合作用(CO2浓度升高/氮增加), CO2浓度升高、干旱胁迫和氮沉降经常被报道可增强植物抵御O3的能力, 减缓O3伤害(白月明等, 2005; Handley & Grulke, 2008; Gao et al., 2017).另一方面, 与之相反, 也有研究发现长期高浓度CO2或干旱降低气孔导度的同时, 会引起植物蒸腾速率、光合产物合成和积累的降低(McDonald et al., 2002; Pollastrini et al., 2010); 而超过植物承载阈值的高氮负荷也会对植物的光合生理活性产生负效应(Yamaguchi et al., 2010), 进而加重植物在复合环境下的碳损失、水分蒸散损失和自由基积累, 对植物产生协同伤害(Karnosky et al., 2005; 列淦文和薛立, 2014).高峰(2018)认为O3与其他环境胁迫因子出现的先后次序也会影响植物对复合作用的响应. ...
大豆对臭氧、二氧化碳及其复合效应的响应 1 2005
... 除单因子O3胁迫外, 由于全球气候变化和极端气候常态化, 植物经常受到O3和其他环境因子交互作用的影响(如O3与CO2、升温、干旱或氮沉降等)(Schmadel-Hageb?lling et al., 1998; Ollinger et al., 2002; Eltayeb et al., 2006; Ghimire et al., 2018).然而, 植物应对O3与其他因子交互作用的响应并不是两者或单独作用的简单叠加, 往往取决于植物种类、生长时期、对特定环境因子的敏感性、胁迫浓度及时间, 以及具体的微气候条件(列淦文和薛立, 2014).一方面, 通过诱导气孔关闭、限制叶片O3吸收(CO2浓度升高/干旱胁迫), 或提高碳同化速率、促进植物光合作用(CO2浓度升高/氮增加), CO2浓度升高、干旱胁迫和氮沉降经常被报道可增强植物抵御O3的能力, 减缓O3伤害(白月明等, 2005; Handley & Grulke, 2008; Gao et al., 2017).另一方面, 与之相反, 也有研究发现长期高浓度CO2或干旱降低气孔导度的同时, 会引起植物蒸腾速率、光合产物合成和积累的降低(McDonald et al., 2002; Pollastrini et al., 2010); 而超过植物承载阈值的高氮负荷也会对植物的光合生理活性产生负效应(Yamaguchi et al., 2010), 进而加重植物在复合环境下的碳损失、水分蒸散损失和自由基积累, 对植物产生协同伤害(Karnosky et al., 2005; 列淦文和薛立, 2014).高峰(2018)认为O3与其他环境胁迫因子出现的先后次序也会影响植物对复合作用的响应. ...
Diversity of an early successional plant community as in?uenced by ozone 1 1998
... 植物长期过量摄取O3还可以引起物种组成、冠层结构改变, 影响生态系统种群均匀度和丰富度, 威胁生态系统多样性(Fuhrer et al., 2016; Bergmann et al., 2017), 最终影响生态系统碳、氮和水循环(McLaughlin et al., 2007; Sun et al., 2012; Grulke & Heath, 2020).基于温室或OTCs控制实验下有关草本植物的多项研究都发现, 随着O3暴露时间的延长, 对O3相对敏感的物种/种群将会被对O3相对耐受的物种/种群替代(Fuhrer et al., 1994; Hayes et al., 2009), 非禾本科杂草与禾本科杂草的比例也在发生改变(Hayes et al., 2010).有关Aspen FACE森林生态系统的研究也表明, 尽管整个森林林分结构没有受到明显影响, 但敏感型欧洲山杨树种在O3熏蒸7年后几乎从森林冠层中消失(Kubiske et al., 2007).然而, 由于种内及种间复杂的竞争关系, O3对生态系统群落结构的影响有时候并不是统一的, 高浓度O3暴露环境下O3敏感型种群首先达到较高覆盖度的情况也时有发生(Evans & Ashmore, 1992; Barbo et al., 1998).此外, 由于生态系统各营养级之间的交互作用, 高浓度O3影响植物群落多样性的同时也潜在影响植物组织、化学组成、二次代谢产物(如酚苷类化合物、植物源挥发性有机化合物等)释放, 进而改变植物种群周边与之相关的动植物、真菌、细菌和昆虫行为或活动(ICP Vegetation, 2013; Fuhrer et al., 2016).例如, O3暴露使得西黄松(Pinus ponderosa)更易受到小蠹虫的袭击(Dahlsten et al., 1997), 但O3诱导红云杉(Picea rubens)萜烯物质释放增加的同时减少了其上蚜虫幼虫的数量(Cannon, 1990); 而有关欧洲山杨的研究则发现O3暴露降低柳皮苷和特里杨苷含量, 增加了缩合单宁含量, 引起林分中蚜虫和森林天幕毛虫幼虫数量剧增, 但蚜虫捕食者或天敌(如瓢虫、草蛉和蜘蛛)的数量显著减少(Percy et al., 2002; Kopper & Lindroth, 2003). ...
Impact of tropospheric ozone on terrestrial biodiversity: a literature analysis to identify ozone sensitive taxa 1 2017
... 植物长期过量摄取O3还可以引起物种组成、冠层结构改变, 影响生态系统种群均匀度和丰富度, 威胁生态系统多样性(Fuhrer et al., 2016; Bergmann et al., 2017), 最终影响生态系统碳、氮和水循环(McLaughlin et al., 2007; Sun et al., 2012; Grulke & Heath, 2020).基于温室或OTCs控制实验下有关草本植物的多项研究都发现, 随着O3暴露时间的延长, 对O3相对敏感的物种/种群将会被对O3相对耐受的物种/种群替代(Fuhrer et al., 1994; Hayes et al., 2009), 非禾本科杂草与禾本科杂草的比例也在发生改变(Hayes et al., 2010).有关Aspen FACE森林生态系统的研究也表明, 尽管整个森林林分结构没有受到明显影响, 但敏感型欧洲山杨树种在O3熏蒸7年后几乎从森林冠层中消失(Kubiske et al., 2007).然而, 由于种内及种间复杂的竞争关系, O3对生态系统群落结构的影响有时候并不是统一的, 高浓度O3暴露环境下O3敏感型种群首先达到较高覆盖度的情况也时有发生(Evans & Ashmore, 1992; Barbo et al., 1998).此外, 由于生态系统各营养级之间的交互作用, 高浓度O3影响植物群落多样性的同时也潜在影响植物组织、化学组成、二次代谢产物(如酚苷类化合物、植物源挥发性有机化合物等)释放, 进而改变植物种群周边与之相关的动植物、真菌、细菌和昆虫行为或活动(ICP Vegetation, 2013; Fuhrer et al., 2016).例如, O3暴露使得西黄松(Pinus ponderosa)更易受到小蠹虫的袭击(Dahlsten et al., 1997), 但O3诱导红云杉(Picea rubens)萜烯物质释放增加的同时减少了其上蚜虫幼虫的数量(Cannon, 1990); 而有关欧洲山杨的研究则发现O3暴露降低柳皮苷和特里杨苷含量, 增加了缩合单宁含量, 引起林分中蚜虫和森林天幕毛虫幼虫数量剧增, 但蚜虫捕食者或天敌(如瓢虫、草蛉和蜘蛛)的数量显著减少(Percy et al., 2002; Kopper & Lindroth, 2003). ...
The ozone component of global change: potential effects on agricultural and horticultural plant yield, product quality and interactions with invasive species 3 2009
How vegetation impacts affect climate metrics for ozone precursors 1 2010
... 地表O3浓度升高对植物光合作用、生长和生物量积累、物质分配等过程的连锁负效应导致陆地生态系统生产力和固碳能力降低(Reich, 1987; Wittig et al., 2009; Ainsworth et al., 2012), 削弱了自然生态系统的碳汇能力(Felzer et al., 2005; Sitch et al., 2007; Collins et al., 2010).基于大量OTCs控制实验下北半球中纬度北方温带树种的meta分析发现, O3浓度升高导致树木的净光合作用和总生物量分别降低11%和7% (Wittig et al., 2009).最近, 有关中国热带、亚热带和温带树种的meta分析则发现, O3浓度升高导致中国木本植物的净光合作用和总生物量分别降低28%和14% (Li et al., 2017). ...
Global distribution and trends of tropospheric ozone: an observation- based review 1 2014
Future effects of ozone on carbon sequestration and climate change policy using a global biogeochemical model 1 2005
... 地表O3浓度升高对植物光合作用、生长和生物量积累、物质分配等过程的连锁负效应导致陆地生态系统生产力和固碳能力降低(Reich, 1987; Wittig et al., 2009; Ainsworth et al., 2012), 削弱了自然生态系统的碳汇能力(Felzer et al., 2005; Sitch et al., 2007; Collins et al., 2010).基于大量OTCs控制实验下北半球中纬度北方温带树种的meta分析发现, O3浓度升高导致树木的净光合作用和总生物量分别降低11%和7% (Wittig et al., 2009).最近, 有关中国热带、亚热带和温带树种的meta分析则发现, O3浓度升高导致中国木本植物的净光合作用和总生物量分别降低28%和14% (Li et al., 2017). ...
A unifying explanation for variation in ozone sensitivity among woody plants 1 2018
... (1)开展O3-FACE研究: 现有研究多为OTC系统下的实验, OTC气室内“微气候” (如温度、水分、光照和风力等生态因素)与外界环境状态存在明显差异, 这些生态因子的变化可能影响植物应对O3的响应, 缓解或加重O3污染对农作物产量或森林植被生产力的损伤(Long et al., 2005; Feng et al., 2018a).全球模型评估结果显示, OTC实验结果低估了O3对作物产量损失的影响(van Dingenen et al., 2009).最近, Feng等(2018b)也发现, 同样浓度的O3对OTC实验下小麦和水稻产量的影响要低于FACE实验; 而对大豆产量的影响则相反.因此, OTC实验结果并不能完全应用于实际O3区域风险的评估.O3-FACE平台是未来研究实际大田环境下O3浓度升高对农林生态系统影响的主要依托平台.为减少区域O3风险评估的不确定性, 未来应加大O3-FACE平台的实验研究.考虑到目前已开展的FACE实验多坐落在温带地区, 主要探究温带植被或作物对高浓度O3的响应, 较少涉及热带和寒带植物或其他区域典型生态系统(Ainsworth et al., 2020).最理想的情况是, 不同区域、不同生态系统类型共同开展O3-FACE实验研究(Ainsworth et al., 2012). ...
Ground-level O3 pollution and its impacts on food crops in China: a review 1 2015
Concentration- and flux-based ozone dose-response relationships for five poplar clones grown in North China 1 2015
... 基于OTCs和O3-FACE平台下单个站点的控制实验可探究O3对植物个体、群落或生态系统的影响, 但将站点或样地尺度的研究外推到区域或全球, 评估O3污染区域风险的研究主要通过响应关系和模型.目前, 其研究领域主要集中在地表O3污染对农作物产量和森林生产力等方面.早在1994年, Chameides等(1994)就利用经济模型初步预测O3浓度升高可能导致2025年全球30%-75%的作物面临产量降低的风险.基于O3前体物排放、化学反应传输模型和M12、AOT40指标的评估显示, 2000年全球O3污染引起全球小麦、玉米和大豆产量平均降低2.2%-15%, 每年因上述作物产量降低所导致的经济损失达110-180亿美元(Avnery et al., 2011).最近, 基于PODY评估指标的研究则表明, O3对小麦造成的产量损失北半球(9.9%)明显高于南半球(6.2%), 平均产量损失最高的地区主要位于北半球暖温带湿润地区(12.5%)、热带湿热(12.5%)和热带潮湿气候区(17.2%)(Mills et al., 2018).在区域尺度上, 基于1990年O3浓度数据, Wang和Mauzerall (2004)利用M7和M12指标评估了O3对中国、日本和韩国主要农作物产量的影响, 结果显示O3污染造成小麦、玉米、棉花减产1%-9%, 大豆减产23%-27%, 造成2.4-35亿美元的经济损失, 在维持现有的污染物排放、管理措施和农业生产技术不变的情况下, 预计到2020年小麦、玉米、棉花减产幅度为2%-15%, 大豆为28%-35%.最近, 利用AOT40指标的研究显示, 2014年当前O3浓度导致中国农作物产量损失7 840万t, 小麦、水稻和玉米相对减产量分别为8.5%-14%、3.9%-15%和2.2%-5.5% (Lin et al., 2018).而只聚焦于长江三角洲地区的研究则发现, 2015年区域O3污染分别造成小麦和水稻减产9.9%-36.1%和7.3%- 23.9% (Zhao et al., 2018).虽然国内也有少量研究建立了PODY与个别树种或作物相对生物量或相对产量之间的通量响应关系(Feng et al., 2012; Hu et al., 2015; Gao et al., 2017), 但并没有研究利用PODY指标评估区域O3污染对农作物产量损失的影响. ...
Ozone pollution: a hidden threat to food security. Programme Coordination Centre for the ICP Vegetation//Mills G, Harmens H. Programme Coordination Centre for the ICP Vegetation 1 2011
Ozone pollution: impacts on ecosystem services and biodiversity. Programme Coordination Centre for the ICP Vegetation//Mills G, Wagg S, Harmens H. Programme Coordination Centre for the ICP Vegetation 2 2013
... 植物长期过量摄取O3还可以引起物种组成、冠层结构改变, 影响生态系统种群均匀度和丰富度, 威胁生态系统多样性(Fuhrer et al., 2016; Bergmann et al., 2017), 最终影响生态系统碳、氮和水循环(McLaughlin et al., 2007; Sun et al., 2012; Grulke & Heath, 2020).基于温室或OTCs控制实验下有关草本植物的多项研究都发现, 随着O3暴露时间的延长, 对O3相对敏感的物种/种群将会被对O3相对耐受的物种/种群替代(Fuhrer et al., 1994; Hayes et al., 2009), 非禾本科杂草与禾本科杂草的比例也在发生改变(Hayes et al., 2010).有关Aspen FACE森林生态系统的研究也表明, 尽管整个森林林分结构没有受到明显影响, 但敏感型欧洲山杨树种在O3熏蒸7年后几乎从森林冠层中消失(Kubiske et al., 2007).然而, 由于种内及种间复杂的竞争关系, O3对生态系统群落结构的影响有时候并不是统一的, 高浓度O3暴露环境下O3敏感型种群首先达到较高覆盖度的情况也时有发生(Evans & Ashmore, 1992; Barbo et al., 1998).此外, 由于生态系统各营养级之间的交互作用, 高浓度O3影响植物群落多样性的同时也潜在影响植物组织、化学组成、二次代谢产物(如酚苷类化合物、植物源挥发性有机化合物等)释放, 进而改变植物种群周边与之相关的动植物、真菌、细菌和昆虫行为或活动(ICP Vegetation, 2013; Fuhrer et al., 2016).例如, O3暴露使得西黄松(Pinus ponderosa)更易受到小蠹虫的袭击(Dahlsten et al., 1997), 但O3诱导红云杉(Picea rubens)萜烯物质释放增加的同时减少了其上蚜虫幼虫的数量(Cannon, 1990); 而有关欧洲山杨的研究则发现O3暴露降低柳皮苷和特里杨苷含量, 增加了缩合单宁含量, 引起林分中蚜虫和森林天幕毛虫幼虫数量剧增, 但蚜虫捕食者或天敌(如瓢虫、草蛉和蜘蛛)的数量显著减少(Percy et al., 2002; Kopper & Lindroth, 2003). ...
... 此外, 虽然地下生态系统并不是直接暴露在O3环境下, 但大量证据表明O3对地上生态系统的影响将会转移至地下(Andersen, 2003; Lindroth, 2010).如高浓度O3环境下, 植物分配到根的生物量将会大大减少, 基于100多组实验数据的meta分析研究表明O3暴露导致生物量分配到地下的量平均减少5.6% (Grantz et al., 2006).另外, 凋落物分解是生态系统营养循环的关键过程.通过影响凋落物的数量、质量和分解速率, 高浓度地表O3会间接改变土壤微生物群落活性、组成和结构, 土壤化学属性及土壤营养循环(Larson et al., 2002; Fuhrer et al., 2016).然而, 比较遗憾的是, 目前有关O3对地下生态过程影响的研究仅集中在农田生态系统, 仅有少部分研究涉及森林土壤(主要来自Aspen FACE)或草地生态系统(ICP Vegetation, 2013).而且, 由于凋落物种类、化学成分、分解者种群、土壤和大气环境条件的不同, 高浓度O3对凋落物分解动态的影响还存在很大争议(Gessner et al., 2010; Fuhrer et al., 2016; Grulke & Heath, 2020).例如, 研究发现O3对欧洲白桦(Betula pendula)叶片凋落物质量和损失率有明显影响(Kasurinen et al., 2006, 2007), 但不影响欧洲山毛榉凋落物的分解速率(Schloter et al., 2005).有关地表O3浓度升高对地下生态系统结构和功能的影响还有待进一步探究. ...
... 相对于农作物产量, O3对粮食品质影响的研究集中在几种主要农作物(如小麦、水稻和大豆)(冯兆忠等, 2018).基于北美、欧洲和亚洲已有的O3对小麦影响的meta分析发现, O3显著降低小麦淀粉浓度, 但显著增加蛋白质和一些营养元素浓度(如钾、镁、钙、磷、锌、锰和铜)(Broberg et al., 2015).印度北部热带地区有关小麦的研究则显示, O3显著降低小麦籽粒糖含量, 但没有改变籽粒蛋白质含量(Tomer et al., 2015).有关水稻的研究还发现, O3增加粳稻直链淀粉含量, 但对籼稻直链淀粉含量无显著影响(Sawada et al., 2016).国内研究则表明, 尽管高浓度O3显著降低冬小麦面粉中支链淀粉和淀粉含量, 但是增加直链淀粉含量(Zhang et al., 2013).O3显著降低水稻稻米直链淀粉浓度、分解率, 增加黏度、起糊温度、蛋白质和微量矿物质浓度, 但蛋白质总量没有改变(Wang et al., 2012).此外, 研究还发现O3熏蒸显著降低了小麦籽粒出粉率和硬度, 但增加冬小麦面粉峰值黏度、低谷黏度、最终黏度和反弹值(贾一磊, 2016). ...
Signalling and cell death in ozone-exposed plants 1 2005
... 气孔防御机制失效后, 进入细胞的O3将诱导叶片内部抗氧化系统启动解毒和修复的非气孔防御响应(冯兆忠等, 2018).非气孔防御响应主要是指“质外体-共质体系统”内一些抗氧化组分对O3解毒和修复的防御过程.具体来说, O3进入植物叶片的细胞间隙后会溶解在细胞壁的结合水里, 经过一系列的化学反应形成活性氧分子(ROS), ROS能够破坏细胞壁并与细胞膜上的不饱和脂肪酸发生反应, 从而对叶片造成损伤(Tausz et al., 2007; Pellegrini et al., 2018).植物质外体的抗氧化物质如还原性抗坏血酸(ASC)、抗氧化酶(如超氧化物歧化酶)可与ROS反应进行初级解毒, 成为抵御O3的第二道防线(Kangasj?rvi et al., 2005; Dai et al., 2019), 而剩余的ROS会经过质膜到达共质体, 此时共质体内抗氧化成分如过氧化物酶(POD)、过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)及抗氧化剂(如酚类化合物-谷胱甘肽(GSH)、脯氨酸和多胺等)等抗氧化酶类物质被诱导作为次级解毒响应机制参与诸如“抗坏血酸-谷胱甘肽(ASA-GSH)循环”等氧化还原过程, 进而对ROS进行进一步解毒与消除(Luwe, 1996).气孔闭合除了会减少O3的吸收, 也会抑制CO2吸收和水汽交换, 而非气孔防御响应中的抗氧化系统解毒过程也会消耗大量的能量并对光合反应系统造成不同程度的损伤, 降低羧化速率和电子传递速率, 进而抑制光合和蒸腾作用, 损害植物生长(Gao et al., 2016).然而, 最新研究表明持续整个生长季的长期O3暴露会影响植物叶片的气孔密度、开度和大小等表观属性(李品等, 2018), 而O3从气孔进入到细胞间隙后, 也可以通过影响植物的叶肉导度降低植物的光合能力(Xu et al., 2019).此外, 有关29种落叶和常绿树种应对O3胁迫的研究还发现不同木本植物对O3响应的差异主要来自植物功能叶片比叶质量的大小(即叶片比叶质量越大的树种对O3越不敏感), 与气孔导度无明显相关性(Li et al., 2016). ...
Scaling ozone responses of forest trees to the ecosystem level in a changing climate 2 2005
... 除单因子O3胁迫外, 由于全球气候变化和极端气候常态化, 植物经常受到O3和其他环境因子交互作用的影响(如O3与CO2、升温、干旱或氮沉降等)(Schmadel-Hageb?lling et al., 1998; Ollinger et al., 2002; Eltayeb et al., 2006; Ghimire et al., 2018).然而, 植物应对O3与其他因子交互作用的响应并不是两者或单独作用的简单叠加, 往往取决于植物种类、生长时期、对特定环境因子的敏感性、胁迫浓度及时间, 以及具体的微气候条件(列淦文和薛立, 2014).一方面, 通过诱导气孔关闭、限制叶片O3吸收(CO2浓度升高/干旱胁迫), 或提高碳同化速率、促进植物光合作用(CO2浓度升高/氮增加), CO2浓度升高、干旱胁迫和氮沉降经常被报道可增强植物抵御O3的能力, 减缓O3伤害(白月明等, 2005; Handley & Grulke, 2008; Gao et al., 2017).另一方面, 与之相反, 也有研究发现长期高浓度CO2或干旱降低气孔导度的同时, 会引起植物蒸腾速率、光合产物合成和积累的降低(McDonald et al., 2002; Pollastrini et al., 2010); 而超过植物承载阈值的高氮负荷也会对植物的光合生理活性产生负效应(Yamaguchi et al., 2010), 进而加重植物在复合环境下的碳损失、水分蒸散损失和自由基积累, 对植物产生协同伤害(Karnosky et al., 2005; 列淦文和薛立, 2014).高峰(2018)认为O3与其他环境胁迫因子出现的先后次序也会影响植物对复合作用的响应. ...
... 有关自然环境下O3浓度升高对陆地生态系统影响的研究较为稀缺.但是现有开放式O3浓度增强系统(O3-FACE)观测平台下的长期研究也有着相似结果, Aspen FACE实验发现, O3浓度升高3-4年后, 欧洲山杨(Populus tremuloides)纯林、欧洲山杨-纸皮桦(Betula papyrifera)和欧洲山杨-糖枫(Acer saccharum)混交林的总生物量分别降低23%、13%和14% (Karnosky et al., 2003, 2005); 虽然在最初的7年内生物量的分配格局并未受O3影响(King et al., 2005), 但在实验11年后, 敏感型欧洲山杨几乎从森林冠层中消失(Kubiske et al., 2007).德国克兰兹堡郁闭冠层的成熟挪威云杉(Picea abies)和欧洲山毛榉(Fagus sylvatica)混交林(60年)对O3浓度升高长期响应的研究也表明, 8年O3暴露导致欧洲山毛榉的树干生产力降低了44% (Pretzsch et al., 2010).目前, 国内还没有研究涉及O3-FACE平台下O3浓度升高对森林生态系统碳汇能力的影响. ...
Tropospheric O3 moderates responses of temperate hardwood forests to elevated CO2: a synthesis of molecular to ecosystem results from the Aspen FACE project 2 2003
... 在此期间, 尽管社会经济不断发展、实验设备和研究方法不断更新, 但地表O3污染对植物及生态系统影响的研究主要还是以模拟O3浓度升高的控制实验为主.纵观发展历程, 试验设备与手段的改进和更新大致可分为三个阶段(王春乙, 1995): (1) 1970年之前, 研究主要利用封闭式静态或动态气室或室内生长箱进行模拟实验(Heagle et al., 1973). (2) 1973年之后, 美国****Heagle等(1979)和Mandle (1973)研制开顶式气室(OTC)模拟高浓度O3对植物的损伤和影响机制, 并最终在世界范围内推广, 成为研究地表O3浓度升高对植物及生态系统影响的主流手段.(3)由于OTCs气室内外较大的环境差异, OTCs模拟实验下的熏蒸结果是否适用于大田研究一直备受争议.为解决OTCs内微气候环境带来的影响, 1986年Hendrey和Kimball (1994)提出建立自由空气中气体浓度增加(FACE)系统的想法, 并逐步用于大田实验.由于与外界大气几乎一致的气体流速、温度、光照和风速等条件(Matyssek et al., 2010), FACE系统被普遍认为是当前研究地表O3浓度升高对植物及生态系统影响最为理想的实验平台, 但由于造价昂贵, 技术要求高, 只有少数国家和实验室开展相关研究(有关上述研究方法的优缺点见表2).O3熏蒸的FACE平台仅有10个左右且多数已经停止运行(Paoletti et al., 2017), 主要包括美国威斯康星州Aspen FACE (停止运行; Karnosky et al., 2003), 美国伊利诺伊州Soybean FACE (停止运行; Morgan et al., 2004), 瑞典苏尔草地FACE (停止运行; Volk et al., 2003), 芬兰库奥皮奥FACE (停止运行; Oksanen, 2003), 德国克兰兹堡森林FACE (停止运行; Matyssek et al., 2010), 中国江都水稻(Oryza sativa)/小麦(Triticum aestivum)轮作FACE (停止运行; Tang et al., 2011), 日本札幌森林FACE (仍在运行; Watanabe et al., 2013), 意大利佛罗伦萨3D FACE (仍在运行; Paoletti et al., 2017), 以及刚刚建成的中国延庆唐家堡杨树人工林FACE.由于实验条件和FACE平台规模的限制, 有关地表O3浓度升高对生态系统水平的研究成果主要来自美国Aspen FACE和德国克兰兹堡森林FACE (Grulke & Heath, 2020). ...
... 有关自然环境下O3浓度升高对陆地生态系统影响的研究较为稀缺.但是现有开放式O3浓度增强系统(O3-FACE)观测平台下的长期研究也有着相似结果, Aspen FACE实验发现, O3浓度升高3-4年后, 欧洲山杨(Populus tremuloides)纯林、欧洲山杨-纸皮桦(Betula papyrifera)和欧洲山杨-糖枫(Acer saccharum)混交林的总生物量分别降低23%、13%和14% (Karnosky et al., 2003, 2005); 虽然在最初的7年内生物量的分配格局并未受O3影响(King et al., 2005), 但在实验11年后, 敏感型欧洲山杨几乎从森林冠层中消失(Kubiske et al., 2007).德国克兰兹堡郁闭冠层的成熟挪威云杉(Picea abies)和欧洲山毛榉(Fagus sylvatica)混交林(60年)对O3浓度升高长期响应的研究也表明, 8年O3暴露导致欧洲山毛榉的树干生产力降低了44% (Pretzsch et al., 2010).目前, 国内还没有研究涉及O3-FACE平台下O3浓度升高对森林生态系统碳汇能力的影响. ...
Leaf litter under changing climate: Will increasing levels of CO2 and O3 affect decomposition and nutrient cycling processes? 1 2007
... 此外, 虽然地下生态系统并不是直接暴露在O3环境下, 但大量证据表明O3对地上生态系统的影响将会转移至地下(Andersen, 2003; Lindroth, 2010).如高浓度O3环境下, 植物分配到根的生物量将会大大减少, 基于100多组实验数据的meta分析研究表明O3暴露导致生物量分配到地下的量平均减少5.6% (Grantz et al., 2006).另外, 凋落物分解是生态系统营养循环的关键过程.通过影响凋落物的数量、质量和分解速率, 高浓度地表O3会间接改变土壤微生物群落活性、组成和结构, 土壤化学属性及土壤营养循环(Larson et al., 2002; Fuhrer et al., 2016).然而, 比较遗憾的是, 目前有关O3对地下生态过程影响的研究仅集中在农田生态系统, 仅有少部分研究涉及森林土壤(主要来自Aspen FACE)或草地生态系统(ICP Vegetation, 2013).而且, 由于凋落物种类、化学成分、分解者种群、土壤和大气环境条件的不同, 高浓度O3对凋落物分解动态的影响还存在很大争议(Gessner et al., 2010; Fuhrer et al., 2016; Grulke & Heath, 2020).例如, 研究发现O3对欧洲白桦(Betula pendula)叶片凋落物质量和损失率有明显影响(Kasurinen et al., 2006, 2007), 但不影响欧洲山毛榉凋落物的分解速率(Schloter et al., 2005).有关地表O3浓度升高对地下生态系统结构和功能的影响还有待进一步探究. ...
Chemical composition and decomposition of silver birch leaf litter produced under elevated CO2 and O3 1 2006
... 此外, 虽然地下生态系统并不是直接暴露在O3环境下, 但大量证据表明O3对地上生态系统的影响将会转移至地下(Andersen, 2003; Lindroth, 2010).如高浓度O3环境下, 植物分配到根的生物量将会大大减少, 基于100多组实验数据的meta分析研究表明O3暴露导致生物量分配到地下的量平均减少5.6% (Grantz et al., 2006).另外, 凋落物分解是生态系统营养循环的关键过程.通过影响凋落物的数量、质量和分解速率, 高浓度地表O3会间接改变土壤微生物群落活性、组成和结构, 土壤化学属性及土壤营养循环(Larson et al., 2002; Fuhrer et al., 2016).然而, 比较遗憾的是, 目前有关O3对地下生态过程影响的研究仅集中在农田生态系统, 仅有少部分研究涉及森林土壤(主要来自Aspen FACE)或草地生态系统(ICP Vegetation, 2013).而且, 由于凋落物种类、化学成分、分解者种群、土壤和大气环境条件的不同, 高浓度O3对凋落物分解动态的影响还存在很大争议(Gessner et al., 2010; Fuhrer et al., 2016; Grulke & Heath, 2020).例如, 研究发现O3对欧洲白桦(Betula pendula)叶片凋落物质量和损失率有明显影响(Kasurinen et al., 2006, 2007), 但不影响欧洲山毛榉凋落物的分解速率(Schloter et al., 2005).有关地表O3浓度升高对地下生态系统结构和功能的影响还有待进一步探究. ...
Tropospheric O3 compromises net primary production in young stands of trembling aspen, paper birch and sugar maple in response to elevated atmospheric CO2 1 2005
... 有关自然环境下O3浓度升高对陆地生态系统影响的研究较为稀缺.但是现有开放式O3浓度增强系统(O3-FACE)观测平台下的长期研究也有着相似结果, Aspen FACE实验发现, O3浓度升高3-4年后, 欧洲山杨(Populus tremuloides)纯林、欧洲山杨-纸皮桦(Betula papyrifera)和欧洲山杨-糖枫(Acer saccharum)混交林的总生物量分别降低23%、13%和14% (Karnosky et al., 2003, 2005); 虽然在最初的7年内生物量的分配格局并未受O3影响(King et al., 2005), 但在实验11年后, 敏感型欧洲山杨几乎从森林冠层中消失(Kubiske et al., 2007).德国克兰兹堡郁闭冠层的成熟挪威云杉(Picea abies)和欧洲山毛榉(Fagus sylvatica)混交林(60年)对O3浓度升高长期响应的研究也表明, 8年O3暴露导致欧洲山毛榉的树干生产力降低了44% (Pretzsch et al., 2010).目前, 国内还没有研究涉及O3-FACE平台下O3浓度升高对森林生态系统碳汇能力的影响. ...
Effects of elevated carbon dioxide and ozone on the phytochemistry of aspen and performance of an herbivore 1 2003
... 植物长期过量摄取O3还可以引起物种组成、冠层结构改变, 影响生态系统种群均匀度和丰富度, 威胁生态系统多样性(Fuhrer et al., 2016; Bergmann et al., 2017), 最终影响生态系统碳、氮和水循环(McLaughlin et al., 2007; Sun et al., 2012; Grulke & Heath, 2020).基于温室或OTCs控制实验下有关草本植物的多项研究都发现, 随着O3暴露时间的延长, 对O3相对敏感的物种/种群将会被对O3相对耐受的物种/种群替代(Fuhrer et al., 1994; Hayes et al., 2009), 非禾本科杂草与禾本科杂草的比例也在发生改变(Hayes et al., 2010).有关Aspen FACE森林生态系统的研究也表明, 尽管整个森林林分结构没有受到明显影响, 但敏感型欧洲山杨树种在O3熏蒸7年后几乎从森林冠层中消失(Kubiske et al., 2007).然而, 由于种内及种间复杂的竞争关系, O3对生态系统群落结构的影响有时候并不是统一的, 高浓度O3暴露环境下O3敏感型种群首先达到较高覆盖度的情况也时有发生(Evans & Ashmore, 1992; Barbo et al., 1998).此外, 由于生态系统各营养级之间的交互作用, 高浓度O3影响植物群落多样性的同时也潜在影响植物组织、化学组成、二次代谢产物(如酚苷类化合物、植物源挥发性有机化合物等)释放, 进而改变植物种群周边与之相关的动植物、真菌、细菌和昆虫行为或活动(ICP Vegetation, 2013; Fuhrer et al., 2016).例如, O3暴露使得西黄松(Pinus ponderosa)更易受到小蠹虫的袭击(Dahlsten et al., 1997), 但O3诱导红云杉(Picea rubens)萜烯物质释放增加的同时减少了其上蚜虫幼虫的数量(Cannon, 1990); 而有关欧洲山杨的研究则发现O3暴露降低柳皮苷和特里杨苷含量, 增加了缩合单宁含量, 引起林分中蚜虫和森林天幕毛虫幼虫数量剧增, 但蚜虫捕食者或天敌(如瓢虫、草蛉和蜘蛛)的数量显著减少(Percy et al., 2002; Kopper & Lindroth, 2003). ...
Effects of elevated atmospheric CO2 and/or O3 on intra- and interspecific competitive ability of aspen 2 2007
... 有关自然环境下O3浓度升高对陆地生态系统影响的研究较为稀缺.但是现有开放式O3浓度增强系统(O3-FACE)观测平台下的长期研究也有着相似结果, Aspen FACE实验发现, O3浓度升高3-4年后, 欧洲山杨(Populus tremuloides)纯林、欧洲山杨-纸皮桦(Betula papyrifera)和欧洲山杨-糖枫(Acer saccharum)混交林的总生物量分别降低23%、13%和14% (Karnosky et al., 2003, 2005); 虽然在最初的7年内生物量的分配格局并未受O3影响(King et al., 2005), 但在实验11年后, 敏感型欧洲山杨几乎从森林冠层中消失(Kubiske et al., 2007).德国克兰兹堡郁闭冠层的成熟挪威云杉(Picea abies)和欧洲山毛榉(Fagus sylvatica)混交林(60年)对O3浓度升高长期响应的研究也表明, 8年O3暴露导致欧洲山毛榉的树干生产力降低了44% (Pretzsch et al., 2010).目前, 国内还没有研究涉及O3-FACE平台下O3浓度升高对森林生态系统碳汇能力的影响. ...
... 植物长期过量摄取O3还可以引起物种组成、冠层结构改变, 影响生态系统种群均匀度和丰富度, 威胁生态系统多样性(Fuhrer et al., 2016; Bergmann et al., 2017), 最终影响生态系统碳、氮和水循环(McLaughlin et al., 2007; Sun et al., 2012; Grulke & Heath, 2020).基于温室或OTCs控制实验下有关草本植物的多项研究都发现, 随着O3暴露时间的延长, 对O3相对敏感的物种/种群将会被对O3相对耐受的物种/种群替代(Fuhrer et al., 1994; Hayes et al., 2009), 非禾本科杂草与禾本科杂草的比例也在发生改变(Hayes et al., 2010).有关Aspen FACE森林生态系统的研究也表明, 尽管整个森林林分结构没有受到明显影响, 但敏感型欧洲山杨树种在O3熏蒸7年后几乎从森林冠层中消失(Kubiske et al., 2007).然而, 由于种内及种间复杂的竞争关系, O3对生态系统群落结构的影响有时候并不是统一的, 高浓度O3暴露环境下O3敏感型种群首先达到较高覆盖度的情况也时有发生(Evans & Ashmore, 1992; Barbo et al., 1998).此外, 由于生态系统各营养级之间的交互作用, 高浓度O3影响植物群落多样性的同时也潜在影响植物组织、化学组成、二次代谢产物(如酚苷类化合物、植物源挥发性有机化合物等)释放, 进而改变植物种群周边与之相关的动植物、真菌、细菌和昆虫行为或活动(ICP Vegetation, 2013; Fuhrer et al., 2016).例如, O3暴露使得西黄松(Pinus ponderosa)更易受到小蠹虫的袭击(Dahlsten et al., 1997), 但O3诱导红云杉(Picea rubens)萜烯物质释放增加的同时减少了其上蚜虫幼虫的数量(Cannon, 1990); 而有关欧洲山杨的研究则发现O3暴露降低柳皮苷和特里杨苷含量, 增加了缩合单宁含量, 引起林分中蚜虫和森林天幕毛虫幼虫数量剧增, 但蚜虫捕食者或天敌(如瓢虫、草蛉和蜘蛛)的数量显著减少(Percy et al., 2002; Kopper & Lindroth, 2003). ...
Extracellular enzyme activity beneath temperate trees growing under elevated carbon dioxide and ozone 1 2002
... 此外, 虽然地下生态系统并不是直接暴露在O3环境下, 但大量证据表明O3对地上生态系统的影响将会转移至地下(Andersen, 2003; Lindroth, 2010).如高浓度O3环境下, 植物分配到根的生物量将会大大减少, 基于100多组实验数据的meta分析研究表明O3暴露导致生物量分配到地下的量平均减少5.6% (Grantz et al., 2006).另外, 凋落物分解是生态系统营养循环的关键过程.通过影响凋落物的数量、质量和分解速率, 高浓度地表O3会间接改变土壤微生物群落活性、组成和结构, 土壤化学属性及土壤营养循环(Larson et al., 2002; Fuhrer et al., 2016).然而, 比较遗憾的是, 目前有关O3对地下生态过程影响的研究仅集中在农田生态系统, 仅有少部分研究涉及森林土壤(主要来自Aspen FACE)或草地生态系统(ICP Vegetation, 2013).而且, 由于凋落物种类、化学成分、分解者种群、土壤和大气环境条件的不同, 高浓度O3对凋落物分解动态的影响还存在很大争议(Gessner et al., 2010; Fuhrer et al., 2016; Grulke & Heath, 2020).例如, 研究发现O3对欧洲白桦(Betula pendula)叶片凋落物质量和损失率有明显影响(Kasurinen et al., 2006, 2007), 但不影响欧洲山毛榉凋落物的分解速率(Schloter et al., 2005).有关地表O3浓度升高对地下生态系统结构和功能的影响还有待进一步探究. ...
Ambient ozone and adverse crop response: an evaluation of north American and European data as they relate to exposure indices and critical levels 1 1995
... 不管研究方法如何更新, 由于空间和时间上的限制, 模拟地表O3浓度升高的控制实验主要探究高浓度O3对植物损伤的机理, 而区域尺度上探究O3对陆地生态系统的影响却很难通过模拟实验获得.为评价区域O3风险和评估O3对生态系统影响带来的损失, ****主要通过不同评估指标评估区域、国家或全球尺度上地表O3污染的生态效应.总体来说, 评估指标的研究主要经历了三个阶段, 分别是浓度响应关系、剂量响应关系和通量响应关系评估指标(Musselman et al., 2006).浓度响应关系评估指标主要包括M7/M12 (白天7 h (9:00-16:00) O3浓度平均值或白天12 h (8:00-20:00) O3浓度平均值), 早期曾广泛应用于表征O3浓度暴露与农作物产量损失相关性的研究中(Legge et al., 1995).剂量响应关系评估指标主要有SUM06 (O3小时浓度高于或等于60 μg·kg-1的累积值)(Lefohn & Foley, 1992), W126 (O3小时浓度在规定时段内的加权求和值)和AOTX (整个生长季太阳辐射>50 W·m-2时段内O3小时浓度超过X μg·kg-1的累计值, X普遍设为40 μg·kg-1)(Fuhrer et al., 1997; LRTAR Convention, 2015).剂量响应指标综合考虑O3浓度和暴露时间对植物生长的影响, 在世界范围内得到广泛应用. ...
Differences in ozone sensitivity among woody species are related to leaf morphology and antioxidant levels 1 2016
... 气孔防御机制失效后, 进入细胞的O3将诱导叶片内部抗氧化系统启动解毒和修复的非气孔防御响应(冯兆忠等, 2018).非气孔防御响应主要是指“质外体-共质体系统”内一些抗氧化组分对O3解毒和修复的防御过程.具体来说, O3进入植物叶片的细胞间隙后会溶解在细胞壁的结合水里, 经过一系列的化学反应形成活性氧分子(ROS), ROS能够破坏细胞壁并与细胞膜上的不饱和脂肪酸发生反应, 从而对叶片造成损伤(Tausz et al., 2007; Pellegrini et al., 2018).植物质外体的抗氧化物质如还原性抗坏血酸(ASC)、抗氧化酶(如超氧化物歧化酶)可与ROS反应进行初级解毒, 成为抵御O3的第二道防线(Kangasj?rvi et al., 2005; Dai et al., 2019), 而剩余的ROS会经过质膜到达共质体, 此时共质体内抗氧化成分如过氧化物酶(POD)、过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)及抗氧化剂(如酚类化合物-谷胱甘肽(GSH)、脯氨酸和多胺等)等抗氧化酶类物质被诱导作为次级解毒响应机制参与诸如“抗坏血酸-谷胱甘肽(ASA-GSH)循环”等氧化还原过程, 进而对ROS进行进一步解毒与消除(Luwe, 1996).气孔闭合除了会减少O3的吸收, 也会抑制CO2吸收和水汽交换, 而非气孔防御响应中的抗氧化系统解毒过程也会消耗大量的能量并对光合反应系统造成不同程度的损伤, 降低羧化速率和电子传递速率, 进而抑制光合和蒸腾作用, 损害植物生长(Gao et al., 2016).然而, 最新研究表明持续整个生长季的长期O3暴露会影响植物叶片的气孔密度、开度和大小等表观属性(李品等, 2018), 而O3从气孔进入到细胞间隙后, 也可以通过影响植物的叶肉导度降低植物的光合能力(Xu et al., 2019).此外, 有关29种落叶和常绿树种应对O3胁迫的研究还发现不同木本植物对O3响应的差异主要来自植物功能叶片比叶质量的大小(即叶片比叶质量越大的树种对O3越不敏感), 与气孔导度无明显相关性(Li et al., 2016). ...
A meta-analysis on growth, physiological, and biochemical responses of woody species to ground-level ozone highlights the role of plant functional types 1 2017
... 地表O3浓度升高对植物光合作用、生长和生物量积累、物质分配等过程的连锁负效应导致陆地生态系统生产力和固碳能力降低(Reich, 1987; Wittig et al., 2009; Ainsworth et al., 2012), 削弱了自然生态系统的碳汇能力(Felzer et al., 2005; Sitch et al., 2007; Collins et al., 2010).基于大量OTCs控制实验下北半球中纬度北方温带树种的meta分析发现, O3浓度升高导致树木的净光合作用和总生物量分别降低11%和7% (Wittig et al., 2009).最近, 有关中国热带、亚热带和温带树种的meta分析则发现, O3浓度升高导致中国木本植物的净光合作用和总生物量分别降低28%和14% (Li et al., 2017). ...
6种绿化树种的气孔特性与臭氧剂量的响应关系 2018
6种绿化树种的气孔特性与臭氧剂量的响应关系 2018
臭氧与其他环境因子对植物的交互作用 6 2014
... 气孔防御机制失效后, 进入细胞的O3将诱导叶片内部抗氧化系统启动解毒和修复的非气孔防御响应(冯兆忠等, 2018).非气孔防御响应主要是指“质外体-共质体系统”内一些抗氧化组分对O3解毒和修复的防御过程.具体来说, O3进入植物叶片的细胞间隙后会溶解在细胞壁的结合水里, 经过一系列的化学反应形成活性氧分子(ROS), ROS能够破坏细胞壁并与细胞膜上的不饱和脂肪酸发生反应, 从而对叶片造成损伤(Tausz et al., 2007; Pellegrini et al., 2018).植物质外体的抗氧化物质如还原性抗坏血酸(ASC)、抗氧化酶(如超氧化物歧化酶)可与ROS反应进行初级解毒, 成为抵御O3的第二道防线(Kangasj?rvi et al., 2005; Dai et al., 2019), 而剩余的ROS会经过质膜到达共质体, 此时共质体内抗氧化成分如过氧化物酶(POD)、过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)及抗氧化剂(如酚类化合物-谷胱甘肽(GSH)、脯氨酸和多胺等)等抗氧化酶类物质被诱导作为次级解毒响应机制参与诸如“抗坏血酸-谷胱甘肽(ASA-GSH)循环”等氧化还原过程, 进而对ROS进行进一步解毒与消除(Luwe, 1996).气孔闭合除了会减少O3的吸收, 也会抑制CO2吸收和水汽交换, 而非气孔防御响应中的抗氧化系统解毒过程也会消耗大量的能量并对光合反应系统造成不同程度的损伤, 降低羧化速率和电子传递速率, 进而抑制光合和蒸腾作用, 损害植物生长(Gao et al., 2016).然而, 最新研究表明持续整个生长季的长期O3暴露会影响植物叶片的气孔密度、开度和大小等表观属性(李品等, 2018), 而O3从气孔进入到细胞间隙后, 也可以通过影响植物的叶肉导度降低植物的光合能力(Xu et al., 2019).此外, 有关29种落叶和常绿树种应对O3胁迫的研究还发现不同木本植物对O3响应的差异主要来自植物功能叶片比叶质量的大小(即叶片比叶质量越大的树种对O3越不敏感), 与气孔导度无明显相关性(Li et al., 2016). ...
... 除单因子O3胁迫外, 由于全球气候变化和极端气候常态化, 植物经常受到O3和其他环境因子交互作用的影响(如O3与CO2、升温、干旱或氮沉降等)(Schmadel-Hageb?lling et al., 1998; Ollinger et al., 2002; Eltayeb et al., 2006; Ghimire et al., 2018).然而, 植物应对O3与其他因子交互作用的响应并不是两者或单独作用的简单叠加, 往往取决于植物种类、生长时期、对特定环境因子的敏感性、胁迫浓度及时间, 以及具体的微气候条件(列淦文和薛立, 2014).一方面, 通过诱导气孔关闭、限制叶片O3吸收(CO2浓度升高/干旱胁迫), 或提高碳同化速率、促进植物光合作用(CO2浓度升高/氮增加), CO2浓度升高、干旱胁迫和氮沉降经常被报道可增强植物抵御O3的能力, 减缓O3伤害(白月明等, 2005; Handley & Grulke, 2008; Gao et al., 2017).另一方面, 与之相反, 也有研究发现长期高浓度CO2或干旱降低气孔导度的同时, 会引起植物蒸腾速率、光合产物合成和积累的降低(McDonald et al., 2002; Pollastrini et al., 2010); 而超过植物承载阈值的高氮负荷也会对植物的光合生理活性产生负效应(Yamaguchi et al., 2010), 进而加重植物在复合环境下的碳损失、水分蒸散损失和自由基积累, 对植物产生协同伤害(Karnosky et al., 2005; 列淦文和薛立, 2014).高峰(2018)认为O3与其他环境胁迫因子出现的先后次序也会影响植物对复合作用的响应. ...
Impacts of O3 on premature mortality and crop yield loss across China 1 2018
... 基于OTCs和O3-FACE平台下单个站点的控制实验可探究O3对植物个体、群落或生态系统的影响, 但将站点或样地尺度的研究外推到区域或全球, 评估O3污染区域风险的研究主要通过响应关系和模型.目前, 其研究领域主要集中在地表O3污染对农作物产量和森林生产力等方面.早在1994年, Chameides等(1994)就利用经济模型初步预测O3浓度升高可能导致2025年全球30%-75%的作物面临产量降低的风险.基于O3前体物排放、化学反应传输模型和M12、AOT40指标的评估显示, 2000年全球O3污染引起全球小麦、玉米和大豆产量平均降低2.2%-15%, 每年因上述作物产量降低所导致的经济损失达110-180亿美元(Avnery et al., 2011).最近, 基于PODY评估指标的研究则表明, O3对小麦造成的产量损失北半球(9.9%)明显高于南半球(6.2%), 平均产量损失最高的地区主要位于北半球暖温带湿润地区(12.5%)、热带湿热(12.5%)和热带潮湿气候区(17.2%)(Mills et al., 2018).在区域尺度上, 基于1990年O3浓度数据, Wang和Mauzerall (2004)利用M7和M12指标评估了O3对中国、日本和韩国主要农作物产量的影响, 结果显示O3污染造成小麦、玉米、棉花减产1%-9%, 大豆减产23%-27%, 造成2.4-35亿美元的经济损失, 在维持现有的污染物排放、管理措施和农业生产技术不变的情况下, 预计到2020年小麦、玉米、棉花减产幅度为2%-15%, 大豆为28%-35%.最近, 利用AOT40指标的研究显示, 2014年当前O3浓度导致中国农作物产量损失7 840万t, 小麦、水稻和玉米相对减产量分别为8.5%-14%、3.9%-15%和2.2%-5.5% (Lin et al., 2018).而只聚焦于长江三角洲地区的研究则发现, 2015年区域O3污染分别造成小麦和水稻减产9.9%-36.1%和7.3%- 23.9% (Zhao et al., 2018).虽然国内也有少量研究建立了PODY与个别树种或作物相对生物量或相对产量之间的通量响应关系(Feng et al., 2012; Hu et al., 2015; Gao et al., 2017), 但并没有研究利用PODY指标评估区域O3污染对农作物产量损失的影响. ...
Impacts of elevated atmospheric CO2 and O3 on forests: phytochemistry, trophic interactions, and ecosystem dynamics 1 2010
... 此外, 虽然地下生态系统并不是直接暴露在O3环境下, 但大量证据表明O3对地上生态系统的影响将会转移至地下(Andersen, 2003; Lindroth, 2010).如高浓度O3环境下, 植物分配到根的生物量将会大大减少, 基于100多组实验数据的meta分析研究表明O3暴露导致生物量分配到地下的量平均减少5.6% (Grantz et al., 2006).另外, 凋落物分解是生态系统营养循环的关键过程.通过影响凋落物的数量、质量和分解速率, 高浓度地表O3会间接改变土壤微生物群落活性、组成和结构, 土壤化学属性及土壤营养循环(Larson et al., 2002; Fuhrer et al., 2016).然而, 比较遗憾的是, 目前有关O3对地下生态过程影响的研究仅集中在农田生态系统, 仅有少部分研究涉及森林土壤(主要来自Aspen FACE)或草地生态系统(ICP Vegetation, 2013).而且, 由于凋落物种类、化学成分、分解者种群、土壤和大气环境条件的不同, 高浓度O3对凋落物分解动态的影响还存在很大争议(Gessner et al., 2010; Fuhrer et al., 2016; Grulke & Heath, 2020).例如, 研究发现O3对欧洲白桦(Betula pendula)叶片凋落物质量和损失率有明显影响(Kasurinen et al., 2006, 2007), 但不影响欧洲山毛榉凋落物的分解速率(Schloter et al., 2005).有关地表O3浓度升高对地下生态系统结构和功能的影响还有待进一步探究. ...
Global food insecurity. Treatment of major food crops with elevated carbon dioxide or ozone under large-scale fully open-air conditions suggests recent models may have overestimated future yields 1 2005
... (1)开展O3-FACE研究: 现有研究多为OTC系统下的实验, OTC气室内“微气候” (如温度、水分、光照和风力等生态因素)与外界环境状态存在明显差异, 这些生态因子的变化可能影响植物应对O3的响应, 缓解或加重O3污染对农作物产量或森林植被生产力的损伤(Long et al., 2005; Feng et al., 2018a).全球模型评估结果显示, OTC实验结果低估了O3对作物产量损失的影响(van Dingenen et al., 2009).最近, Feng等(2018b)也发现, 同样浓度的O3对OTC实验下小麦和水稻产量的影响要低于FACE实验; 而对大豆产量的影响则相反.因此, OTC实验结果并不能完全应用于实际O3区域风险的评估.O3-FACE平台是未来研究实际大田环境下O3浓度升高对农林生态系统影响的主要依托平台.为减少区域O3风险评估的不确定性, 未来应加大O3-FACE平台的实验研究.考虑到目前已开展的FACE实验多坐落在温带地区, 主要探究温带植被或作物对高浓度O3的响应, 较少涉及热带和寒带植物或其他区域典型生态系统(Ainsworth et al., 2020).最理想的情况是, 不同区域、不同生态系统类型共同开展O3-FACE实验研究(Ainsworth et al., 2012). ...
2 2015
... 不管研究方法如何更新, 由于空间和时间上的限制, 模拟地表O3浓度升高的控制实验主要探究高浓度O3对植物损伤的机理, 而区域尺度上探究O3对陆地生态系统的影响却很难通过模拟实验获得.为评价区域O3风险和评估O3对生态系统影响带来的损失, ****主要通过不同评估指标评估区域、国家或全球尺度上地表O3污染的生态效应.总体来说, 评估指标的研究主要经历了三个阶段, 分别是浓度响应关系、剂量响应关系和通量响应关系评估指标(Musselman et al., 2006).浓度响应关系评估指标主要包括M7/M12 (白天7 h (9:00-16:00) O3浓度平均值或白天12 h (8:00-20:00) O3浓度平均值), 早期曾广泛应用于表征O3浓度暴露与农作物产量损失相关性的研究中(Legge et al., 1995).剂量响应关系评估指标主要有SUM06 (O3小时浓度高于或等于60 μg·kg-1的累积值)(Lefohn & Foley, 1992), W126 (O3小时浓度在规定时段内的加权求和值)和AOTX (整个生长季太阳辐射>50 W·m-2时段内O3小时浓度超过X μg·kg-1的累计值, X普遍设为40 μg·kg-1)(Fuhrer et al., 1997; LRTAR Convention, 2015).剂量响应指标综合考虑O3浓度和暴露时间对植物生长的影响, 在世界范围内得到广泛应用. ...
... 然而, O3对植物的影响不仅依赖于大气O3浓度同时也依赖于叶片气孔导度(O3的吸收)和植物自身抗氧化能力(Matyssek et al., 2007).近年来, 基于气孔O3吸收量的评估指标PODY (植物毒性O3剂量, 整个生长季单位面积上气孔O3吸收通量超过临界值Y nmol m-2·s-1的积累量)被开发并逐步替代浓度暴露剂量指标, 用于区域或全球O3对农作物和森林树种生长影响的评估(Büker et al., 2015; LRTAR Convention, 2015).目前, 哪个指标更适合区域O3风险评估的争论一直存在, 但欧洲和亚洲地区多项O3对农作物或植被影响的研究均表明: 考虑水分供应的情况下, O3暴露造成的生物量或作物产量的降低与PODY的拟合度比AOTX更好(Mills et al., 2011; Gao et al., 2017), 能更好表征O3对生态系统的影响.整体来看, 各个评估指标的计算方式和所需参数从简单到复杂, 从单一考虑O3浓度到综合考虑环境因子对植物O3吸收通量的影响, 逐渐可以精确评估地表O3污染对生态系统农林生产力的影响(表3). ...
Severe surface ozone pollution in China: a global perspective 1 2018
Antioxidants in the apoplast and symplast of beech (Fagus sylvatica L.) leaves: seasonal variations and responses to changing ozone concentrations in air 1 1996
... 气孔防御机制失效后, 进入细胞的O3将诱导叶片内部抗氧化系统启动解毒和修复的非气孔防御响应(冯兆忠等, 2018).非气孔防御响应主要是指“质外体-共质体系统”内一些抗氧化组分对O3解毒和修复的防御过程.具体来说, O3进入植物叶片的细胞间隙后会溶解在细胞壁的结合水里, 经过一系列的化学反应形成活性氧分子(ROS), ROS能够破坏细胞壁并与细胞膜上的不饱和脂肪酸发生反应, 从而对叶片造成损伤(Tausz et al., 2007; Pellegrini et al., 2018).植物质外体的抗氧化物质如还原性抗坏血酸(ASC)、抗氧化酶(如超氧化物歧化酶)可与ROS反应进行初级解毒, 成为抵御O3的第二道防线(Kangasj?rvi et al., 2005; Dai et al., 2019), 而剩余的ROS会经过质膜到达共质体, 此时共质体内抗氧化成分如过氧化物酶(POD)、过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)及抗氧化剂(如酚类化合物-谷胱甘肽(GSH)、脯氨酸和多胺等)等抗氧化酶类物质被诱导作为次级解毒响应机制参与诸如“抗坏血酸-谷胱甘肽(ASA-GSH)循环”等氧化还原过程, 进而对ROS进行进一步解毒与消除(Luwe, 1996).气孔闭合除了会减少O3的吸收, 也会抑制CO2吸收和水汽交换, 而非气孔防御响应中的抗氧化系统解毒过程也会消耗大量的能量并对光合反应系统造成不同程度的损伤, 降低羧化速率和电子传递速率, 进而抑制光合和蒸腾作用, 损害植物生长(Gao et al., 2016).然而, 最新研究表明持续整个生长季的长期O3暴露会影响植物叶片的气孔密度、开度和大小等表观属性(李品等, 2018), 而O3从气孔进入到细胞间隙后, 也可以通过影响植物的叶肉导度降低植物的光合能力(Xu et al., 2019).此外, 有关29种落叶和常绿树种应对O3胁迫的研究还发现不同木本植物对O3响应的差异主要来自植物功能叶片比叶质量的大小(即叶片比叶质量越大的树种对O3越不敏感), 与气孔导度无明显相关性(Li et al., 2016). ...
A cylindrical open top chamber for the exposure of plants to air pollutants in the field 1 1973
... 在此期间, 尽管社会经济不断发展、实验设备和研究方法不断更新, 但地表O3污染对植物及生态系统影响的研究主要还是以模拟O3浓度升高的控制实验为主.纵观发展历程, 试验设备与手段的改进和更新大致可分为三个阶段(王春乙, 1995): (1) 1970年之前, 研究主要利用封闭式静态或动态气室或室内生长箱进行模拟实验(Heagle et al., 1973). (2) 1973年之后, 美国****Heagle等(1979)和Mandle (1973)研制开顶式气室(OTC)模拟高浓度O3对植物的损伤和影响机制, 并最终在世界范围内推广, 成为研究地表O3浓度升高对植物及生态系统影响的主流手段.(3)由于OTCs气室内外较大的环境差异, OTCs模拟实验下的熏蒸结果是否适用于大田研究一直备受争议.为解决OTCs内微气候环境带来的影响, 1986年Hendrey和Kimball (1994)提出建立自由空气中气体浓度增加(FACE)系统的想法, 并逐步用于大田实验.由于与外界大气几乎一致的气体流速、温度、光照和风速等条件(Matyssek et al., 2010), FACE系统被普遍认为是当前研究地表O3浓度升高对植物及生态系统影响最为理想的实验平台, 但由于造价昂贵, 技术要求高, 只有少数国家和实验室开展相关研究(有关上述研究方法的优缺点见表2).O3熏蒸的FACE平台仅有10个左右且多数已经停止运行(Paoletti et al., 2017), 主要包括美国威斯康星州Aspen FACE (停止运行; Karnosky et al., 2003), 美国伊利诺伊州Soybean FACE (停止运行; Morgan et al., 2004), 瑞典苏尔草地FACE (停止运行; Volk et al., 2003), 芬兰库奥皮奥FACE (停止运行; Oksanen, 2003), 德国克兰兹堡森林FACE (停止运行; Matyssek et al., 2010), 中国江都水稻(Oryza sativa)/小麦(Triticum aestivum)轮作FACE (停止运行; Tang et al., 2011), 日本札幌森林FACE (仍在运行; Watanabe et al., 2013), 意大利佛罗伦萨3D FACE (仍在运行; Paoletti et al., 2017), 以及刚刚建成的中国延庆唐家堡杨树人工林FACE.由于实验条件和FACE平台规模的限制, 有关地表O3浓度升高对生态系统水平的研究成果主要来自美国Aspen FACE和德国克兰兹堡森林FACE (Grulke & Heath, 2020). ...
Promoting the O3 flux concept for European forest trees 1 2007
... 然而, O3对植物的影响不仅依赖于大气O3浓度同时也依赖于叶片气孔导度(O3的吸收)和植物自身抗氧化能力(Matyssek et al., 2007).近年来, 基于气孔O3吸收量的评估指标PODY (植物毒性O3剂量, 整个生长季单位面积上气孔O3吸收通量超过临界值Y nmol m-2·s-1的积累量)被开发并逐步替代浓度暴露剂量指标, 用于区域或全球O3对农作物和森林树种生长影响的评估(Büker et al., 2015; LRTAR Convention, 2015).目前, 哪个指标更适合区域O3风险评估的争论一直存在, 但欧洲和亚洲地区多项O3对农作物或植被影响的研究均表明: 考虑水分供应的情况下, O3暴露造成的生物量或作物产量的降低与PODY的拟合度比AOTX更好(Mills et al., 2011; Gao et al., 2017), 能更好表征O3对生态系统的影响.整体来看, 各个评估指标的计算方式和所需参数从简单到复杂, 从单一考虑O3浓度到综合考虑环境因子对植物O3吸收通量的影响, 逐渐可以精确评估地表O3污染对生态系统农林生产力的影响(表3). ...
Enhanced ozone strongly reduces carbon sink strength of adult beech (Fagus sylvatica)—Resume from the free-air fumigation study at Kranzberg forest 2 2010
... 在此期间, 尽管社会经济不断发展、实验设备和研究方法不断更新, 但地表O3污染对植物及生态系统影响的研究主要还是以模拟O3浓度升高的控制实验为主.纵观发展历程, 试验设备与手段的改进和更新大致可分为三个阶段(王春乙, 1995): (1) 1970年之前, 研究主要利用封闭式静态或动态气室或室内生长箱进行模拟实验(Heagle et al., 1973). (2) 1973年之后, 美国****Heagle等(1979)和Mandle (1973)研制开顶式气室(OTC)模拟高浓度O3对植物的损伤和影响机制, 并最终在世界范围内推广, 成为研究地表O3浓度升高对植物及生态系统影响的主流手段.(3)由于OTCs气室内外较大的环境差异, OTCs模拟实验下的熏蒸结果是否适用于大田研究一直备受争议.为解决OTCs内微气候环境带来的影响, 1986年Hendrey和Kimball (1994)提出建立自由空气中气体浓度增加(FACE)系统的想法, 并逐步用于大田实验.由于与外界大气几乎一致的气体流速、温度、光照和风速等条件(Matyssek et al., 2010), FACE系统被普遍认为是当前研究地表O3浓度升高对植物及生态系统影响最为理想的实验平台, 但由于造价昂贵, 技术要求高, 只有少数国家和实验室开展相关研究(有关上述研究方法的优缺点见表2).O3熏蒸的FACE平台仅有10个左右且多数已经停止运行(Paoletti et al., 2017), 主要包括美国威斯康星州Aspen FACE (停止运行; Karnosky et al., 2003), 美国伊利诺伊州Soybean FACE (停止运行; Morgan et al., 2004), 瑞典苏尔草地FACE (停止运行; Volk et al., 2003), 芬兰库奥皮奥FACE (停止运行; Oksanen, 2003), 德国克兰兹堡森林FACE (停止运行; Matyssek et al., 2010), 中国江都水稻(Oryza sativa)/小麦(Triticum aestivum)轮作FACE (停止运行; Tang et al., 2011), 日本札幌森林FACE (仍在运行; Watanabe et al., 2013), 意大利佛罗伦萨3D FACE (仍在运行; Paoletti et al., 2017), 以及刚刚建成的中国延庆唐家堡杨树人工林FACE.由于实验条件和FACE平台规模的限制, 有关地表O3浓度升高对生态系统水平的研究成果主要来自美国Aspen FACE和德国克兰兹堡森林FACE (Grulke & Heath, 2020). ...
... ), 德国克兰兹堡森林FACE (停止运行; Matyssek et al., 2010), 中国江都水稻(Oryza sativa)/小麦(Triticum aestivum)轮作FACE (停止运行; Tang et al., 2011), 日本札幌森林FACE (仍在运行; Watanabe et al., 2013), 意大利佛罗伦萨3D FACE (仍在运行; Paoletti et al., 2017), 以及刚刚建成的中国延庆唐家堡杨树人工林FACE.由于实验条件和FACE平台规模的限制, 有关地表O3浓度升高对生态系统水平的研究成果主要来自美国Aspen FACE和德国克兰兹堡森林FACE (Grulke & Heath, 2020). ...
Competitive status influences tree-growth responses to elevated CO2 and O3 in aggrading aspen stands 1 2002
... 除单因子O3胁迫外, 由于全球气候变化和极端气候常态化, 植物经常受到O3和其他环境因子交互作用的影响(如O3与CO2、升温、干旱或氮沉降等)(Schmadel-Hageb?lling et al., 1998; Ollinger et al., 2002; Eltayeb et al., 2006; Ghimire et al., 2018).然而, 植物应对O3与其他因子交互作用的响应并不是两者或单独作用的简单叠加, 往往取决于植物种类、生长时期、对特定环境因子的敏感性、胁迫浓度及时间, 以及具体的微气候条件(列淦文和薛立, 2014).一方面, 通过诱导气孔关闭、限制叶片O3吸收(CO2浓度升高/干旱胁迫), 或提高碳同化速率、促进植物光合作用(CO2浓度升高/氮增加), CO2浓度升高、干旱胁迫和氮沉降经常被报道可增强植物抵御O3的能力, 减缓O3伤害(白月明等, 2005; Handley & Grulke, 2008; Gao et al., 2017).另一方面, 与之相反, 也有研究发现长期高浓度CO2或干旱降低气孔导度的同时, 会引起植物蒸腾速率、光合产物合成和积累的降低(McDonald et al., 2002; Pollastrini et al., 2010); 而超过植物承载阈值的高氮负荷也会对植物的光合生理活性产生负效应(Yamaguchi et al., 2010), 进而加重植物在复合环境下的碳损失、水分蒸散损失和自由基积累, 对植物产生协同伤害(Karnosky et al., 2005; 列淦文和薛立, 2014).高峰(2018)认为O3与其他环境胁迫因子出现的先后次序也会影响植物对复合作用的响应. ...
Interactive effects of ozone and climate on water use, soil moisture content and streamflow in a southern Appalachian forest in the USA 1 2007
... 植物长期过量摄取O3还可以引起物种组成、冠层结构改变, 影响生态系统种群均匀度和丰富度, 威胁生态系统多样性(Fuhrer et al., 2016; Bergmann et al., 2017), 最终影响生态系统碳、氮和水循环(McLaughlin et al., 2007; Sun et al., 2012; Grulke & Heath, 2020).基于温室或OTCs控制实验下有关草本植物的多项研究都发现, 随着O3暴露时间的延长, 对O3相对敏感的物种/种群将会被对O3相对耐受的物种/种群替代(Fuhrer et al., 1994; Hayes et al., 2009), 非禾本科杂草与禾本科杂草的比例也在发生改变(Hayes et al., 2010).有关Aspen FACE森林生态系统的研究也表明, 尽管整个森林林分结构没有受到明显影响, 但敏感型欧洲山杨树种在O3熏蒸7年后几乎从森林冠层中消失(Kubiske et al., 2007).然而, 由于种内及种间复杂的竞争关系, O3对生态系统群落结构的影响有时候并不是统一的, 高浓度O3暴露环境下O3敏感型种群首先达到较高覆盖度的情况也时有发生(Evans & Ashmore, 1992; Barbo et al., 1998).此外, 由于生态系统各营养级之间的交互作用, 高浓度O3影响植物群落多样性的同时也潜在影响植物组织、化学组成、二次代谢产物(如酚苷类化合物、植物源挥发性有机化合物等)释放, 进而改变植物种群周边与之相关的动植物、真菌、细菌和昆虫行为或活动(ICP Vegetation, 2013; Fuhrer et al., 2016).例如, O3暴露使得西黄松(Pinus ponderosa)更易受到小蠹虫的袭击(Dahlsten et al., 1997), 但O3诱导红云杉(Picea rubens)萜烯物质释放增加的同时减少了其上蚜虫幼虫的数量(Cannon, 1990); 而有关欧洲山杨的研究则发现O3暴露降低柳皮苷和特里杨苷含量, 增加了缩合单宁含量, 引起林分中蚜虫和森林天幕毛虫幼虫数量剧增, 但蚜虫捕食者或天敌(如瓢虫、草蛉和蜘蛛)的数量显著减少(Percy et al., 2002; Kopper & Lindroth, 2003). ...
A synthesis of AOT40-based response functions and critical levels of ozone for agricultural and horticultural crops 1 2007
Evidence of widespread effects of ozone on crops and (semi-) natural vegetation in Europe (1990-2006) in relation to AOT40- and flux-based risk maps 1 2011
... 然而, O3对植物的影响不仅依赖于大气O3浓度同时也依赖于叶片气孔导度(O3的吸收)和植物自身抗氧化能力(Matyssek et al., 2007).近年来, 基于气孔O3吸收量的评估指标PODY (植物毒性O3剂量, 整个生长季单位面积上气孔O3吸收通量超过临界值Y nmol m-2·s-1的积累量)被开发并逐步替代浓度暴露剂量指标, 用于区域或全球O3对农作物和森林树种生长影响的评估(Büker et al., 2015; LRTAR Convention, 2015).目前, 哪个指标更适合区域O3风险评估的争论一直存在, 但欧洲和亚洲地区多项O3对农作物或植被影响的研究均表明: 考虑水分供应的情况下, O3暴露造成的生物量或作物产量的降低与PODY的拟合度比AOTX更好(Mills et al., 2011; Gao et al., 2017), 能更好表征O3对生态系统的影响.整体来看, 各个评估指标的计算方式和所需参数从简单到复杂, 从单一考虑O3浓度到综合考虑环境因子对植物O3吸收通量的影响, 逐渐可以精确评估地表O3污染对生态系统农林生产力的影响(表3). ...
Ozone pollution will compromise efforts to increase global wheat production 3 2018
An in vivo analysis of the effect of season-long open-air elevation of ozone to anticipated 2050 levels on photosynthesis in soybean 1 2004
... 在此期间, 尽管社会经济不断发展、实验设备和研究方法不断更新, 但地表O3污染对植物及生态系统影响的研究主要还是以模拟O3浓度升高的控制实验为主.纵观发展历程, 试验设备与手段的改进和更新大致可分为三个阶段(王春乙, 1995): (1) 1970年之前, 研究主要利用封闭式静态或动态气室或室内生长箱进行模拟实验(Heagle et al., 1973). (2) 1973年之后, 美国****Heagle等(1979)和Mandle (1973)研制开顶式气室(OTC)模拟高浓度O3对植物的损伤和影响机制, 并最终在世界范围内推广, 成为研究地表O3浓度升高对植物及生态系统影响的主流手段.(3)由于OTCs气室内外较大的环境差异, OTCs模拟实验下的熏蒸结果是否适用于大田研究一直备受争议.为解决OTCs内微气候环境带来的影响, 1986年Hendrey和Kimball (1994)提出建立自由空气中气体浓度增加(FACE)系统的想法, 并逐步用于大田实验.由于与外界大气几乎一致的气体流速、温度、光照和风速等条件(Matyssek et al., 2010), FACE系统被普遍认为是当前研究地表O3浓度升高对植物及生态系统影响最为理想的实验平台, 但由于造价昂贵, 技术要求高, 只有少数国家和实验室开展相关研究(有关上述研究方法的优缺点见表2).O3熏蒸的FACE平台仅有10个左右且多数已经停止运行(Paoletti et al., 2017), 主要包括美国威斯康星州Aspen FACE (停止运行; Karnosky et al., 2003), 美国伊利诺伊州Soybean FACE (停止运行; Morgan et al., 2004), 瑞典苏尔草地FACE (停止运行; Volk et al., 2003), 芬兰库奥皮奥FACE (停止运行; Oksanen, 2003), 德国克兰兹堡森林FACE (停止运行; Matyssek et al., 2010), 中国江都水稻(Oryza sativa)/小麦(Triticum aestivum)轮作FACE (停止运行; Tang et al., 2011), 日本札幌森林FACE (仍在运行; Watanabe et al., 2013), 意大利佛罗伦萨3D FACE (仍在运行; Paoletti et al., 2017), 以及刚刚建成的中国延庆唐家堡杨树人工林FACE.由于实验条件和FACE平台规模的限制, 有关地表O3浓度升高对生态系统水平的研究成果主要来自美国Aspen FACE和德国克兰兹堡森林FACE (Grulke & Heath, 2020). ...
A critical review and analysis of the use of exposure- and flux-based ozone indices for predicting vegetation effects 1 2006
... 不管研究方法如何更新, 由于空间和时间上的限制, 模拟地表O3浓度升高的控制实验主要探究高浓度O3对植物损伤的机理, 而区域尺度上探究O3对陆地生态系统的影响却很难通过模拟实验获得.为评价区域O3风险和评估O3对生态系统影响带来的损失, ****主要通过不同评估指标评估区域、国家或全球尺度上地表O3污染的生态效应.总体来说, 评估指标的研究主要经历了三个阶段, 分别是浓度响应关系、剂量响应关系和通量响应关系评估指标(Musselman et al., 2006).浓度响应关系评估指标主要包括M7/M12 (白天7 h (9:00-16:00) O3浓度平均值或白天12 h (8:00-20:00) O3浓度平均值), 早期曾广泛应用于表征O3浓度暴露与农作物产量损失相关性的研究中(Legge et al., 1995).剂量响应关系评估指标主要有SUM06 (O3小时浓度高于或等于60 μg·kg-1的累积值)(Lefohn & Foley, 1992), W126 (O3小时浓度在规定时段内的加权求和值)和AOTX (整个生长季太阳辐射>50 W·m-2时段内O3小时浓度超过X μg·kg-1的累计值, X普遍设为40 μg·kg-1)(Fuhrer et al., 1997; LRTAR Convention, 2015).剂量响应指标综合考虑O3浓度和暴露时间对植物生长的影响, 在世界范围内得到广泛应用. ...
Responses of selected birch (Betula pendula) clones to ozone change over time 1 2003
... 在此期间, 尽管社会经济不断发展、实验设备和研究方法不断更新, 但地表O3污染对植物及生态系统影响的研究主要还是以模拟O3浓度升高的控制实验为主.纵观发展历程, 试验设备与手段的改进和更新大致可分为三个阶段(王春乙, 1995): (1) 1970年之前, 研究主要利用封闭式静态或动态气室或室内生长箱进行模拟实验(Heagle et al., 1973). (2) 1973年之后, 美国****Heagle等(1979)和Mandle (1973)研制开顶式气室(OTC)模拟高浓度O3对植物的损伤和影响机制, 并最终在世界范围内推广, 成为研究地表O3浓度升高对植物及生态系统影响的主流手段.(3)由于OTCs气室内外较大的环境差异, OTCs模拟实验下的熏蒸结果是否适用于大田研究一直备受争议.为解决OTCs内微气候环境带来的影响, 1986年Hendrey和Kimball (1994)提出建立自由空气中气体浓度增加(FACE)系统的想法, 并逐步用于大田实验.由于与外界大气几乎一致的气体流速、温度、光照和风速等条件(Matyssek et al., 2010), FACE系统被普遍认为是当前研究地表O3浓度升高对植物及生态系统影响最为理想的实验平台, 但由于造价昂贵, 技术要求高, 只有少数国家和实验室开展相关研究(有关上述研究方法的优缺点见表2).O3熏蒸的FACE平台仅有10个左右且多数已经停止运行(Paoletti et al., 2017), 主要包括美国威斯康星州Aspen FACE (停止运行; Karnosky et al., 2003), 美国伊利诺伊州Soybean FACE (停止运行; Morgan et al., 2004), 瑞典苏尔草地FACE (停止运行; Volk et al., 2003), 芬兰库奥皮奥FACE (停止运行; Oksanen, 2003), 德国克兰兹堡森林FACE (停止运行; Matyssek et al., 2010), 中国江都水稻(Oryza sativa)/小麦(Triticum aestivum)轮作FACE (停止运行; Tang et al., 2011), 日本札幌森林FACE (仍在运行; Watanabe et al., 2013), 意大利佛罗伦萨3D FACE (仍在运行; Paoletti et al., 2017), 以及刚刚建成的中国延庆唐家堡杨树人工林FACE.由于实验条件和FACE平台规模的限制, 有关地表O3浓度升高对生态系统水平的研究成果主要来自美国Aspen FACE和德国克兰兹堡森林FACE (Grulke & Heath, 2020). ...
Interactive effects of nitrogen deposition, tropospheric ozone, elevated CO2 and land use history on the carbon dynamics of northern hardwood forests 2 2002
... 除单因子O3胁迫外, 由于全球气候变化和极端气候常态化, 植物经常受到O3和其他环境因子交互作用的影响(如O3与CO2、升温、干旱或氮沉降等)(Schmadel-Hageb?lling et al., 1998; Ollinger et al., 2002; Eltayeb et al., 2006; Ghimire et al., 2018).然而, 植物应对O3与其他因子交互作用的响应并不是两者或单独作用的简单叠加, 往往取决于植物种类、生长时期、对特定环境因子的敏感性、胁迫浓度及时间, 以及具体的微气候条件(列淦文和薛立, 2014).一方面, 通过诱导气孔关闭、限制叶片O3吸收(CO2浓度升高/干旱胁迫), 或提高碳同化速率、促进植物光合作用(CO2浓度升高/氮增加), CO2浓度升高、干旱胁迫和氮沉降经常被报道可增强植物抵御O3的能力, 减缓O3伤害(白月明等, 2005; Handley & Grulke, 2008; Gao et al., 2017).另一方面, 与之相反, 也有研究发现长期高浓度CO2或干旱降低气孔导度的同时, 会引起植物蒸腾速率、光合产物合成和积累的降低(McDonald et al., 2002; Pollastrini et al., 2010); 而超过植物承载阈值的高氮负荷也会对植物的光合生理活性产生负效应(Yamaguchi et al., 2010), 进而加重植物在复合环境下的碳损失、水分蒸散损失和自由基积累, 对植物产生协同伤害(Karnosky et al., 2005; 列淦文和薛立, 2014).高峰(2018)认为O3与其他环境胁迫因子出现的先后次序也会影响植物对复合作用的响应. ...
What about the detoxification mechanisms underlying ozone sensitivity in Liriodendron tulipifera 1 2018
... 气孔防御机制失效后, 进入细胞的O3将诱导叶片内部抗氧化系统启动解毒和修复的非气孔防御响应(冯兆忠等, 2018).非气孔防御响应主要是指“质外体-共质体系统”内一些抗氧化组分对O3解毒和修复的防御过程.具体来说, O3进入植物叶片的细胞间隙后会溶解在细胞壁的结合水里, 经过一系列的化学反应形成活性氧分子(ROS), ROS能够破坏细胞壁并与细胞膜上的不饱和脂肪酸发生反应, 从而对叶片造成损伤(Tausz et al., 2007; Pellegrini et al., 2018).植物质外体的抗氧化物质如还原性抗坏血酸(ASC)、抗氧化酶(如超氧化物歧化酶)可与ROS反应进行初级解毒, 成为抵御O3的第二道防线(Kangasj?rvi et al., 2005; Dai et al., 2019), 而剩余的ROS会经过质膜到达共质体, 此时共质体内抗氧化成分如过氧化物酶(POD)、过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)及抗氧化剂(如酚类化合物-谷胱甘肽(GSH)、脯氨酸和多胺等)等抗氧化酶类物质被诱导作为次级解毒响应机制参与诸如“抗坏血酸-谷胱甘肽(ASA-GSH)循环”等氧化还原过程, 进而对ROS进行进一步解毒与消除(Luwe, 1996).气孔闭合除了会减少O3的吸收, 也会抑制CO2吸收和水汽交换, 而非气孔防御响应中的抗氧化系统解毒过程也会消耗大量的能量并对光合反应系统造成不同程度的损伤, 降低羧化速率和电子传递速率, 进而抑制光合和蒸腾作用, 损害植物生长(Gao et al., 2016).然而, 最新研究表明持续整个生长季的长期O3暴露会影响植物叶片的气孔密度、开度和大小等表观属性(李品等, 2018), 而O3从气孔进入到细胞间隙后, 也可以通过影响植物的叶肉导度降低植物的光合能力(Xu et al., 2019).此外, 有关29种落叶和常绿树种应对O3胁迫的研究还发现不同木本植物对O3响应的差异主要来自植物功能叶片比叶质量的大小(即叶片比叶质量越大的树种对O3越不敏感), 与气孔导度无明显相关性(Li et al., 2016). ...
Altered performance of forest pests under CO2- and O3-enriched atmospheres 1 2002
... 植物长期过量摄取O3还可以引起物种组成、冠层结构改变, 影响生态系统种群均匀度和丰富度, 威胁生态系统多样性(Fuhrer et al., 2016; Bergmann et al., 2017), 最终影响生态系统碳、氮和水循环(McLaughlin et al., 2007; Sun et al., 2012; Grulke & Heath, 2020).基于温室或OTCs控制实验下有关草本植物的多项研究都发现, 随着O3暴露时间的延长, 对O3相对敏感的物种/种群将会被对O3相对耐受的物种/种群替代(Fuhrer et al., 1994; Hayes et al., 2009), 非禾本科杂草与禾本科杂草的比例也在发生改变(Hayes et al., 2010).有关Aspen FACE森林生态系统的研究也表明, 尽管整个森林林分结构没有受到明显影响, 但敏感型欧洲山杨树种在O3熏蒸7年后几乎从森林冠层中消失(Kubiske et al., 2007).然而, 由于种内及种间复杂的竞争关系, O3对生态系统群落结构的影响有时候并不是统一的, 高浓度O3暴露环境下O3敏感型种群首先达到较高覆盖度的情况也时有发生(Evans & Ashmore, 1992; Barbo et al., 1998).此外, 由于生态系统各营养级之间的交互作用, 高浓度O3影响植物群落多样性的同时也潜在影响植物组织、化学组成、二次代谢产物(如酚苷类化合物、植物源挥发性有机化合物等)释放, 进而改变植物种群周边与之相关的动植物、真菌、细菌和昆虫行为或活动(ICP Vegetation, 2013; Fuhrer et al., 2016).例如, O3暴露使得西黄松(Pinus ponderosa)更易受到小蠹虫的袭击(Dahlsten et al., 1997), 但O3诱导红云杉(Picea rubens)萜烯物质释放增加的同时减少了其上蚜虫幼虫的数量(Cannon, 1990); 而有关欧洲山杨的研究则发现O3暴露降低柳皮苷和特里杨苷含量, 增加了缩合单宁含量, 引起林分中蚜虫和森林天幕毛虫幼虫数量剧增, 但蚜虫捕食者或天敌(如瓢虫、草蛉和蜘蛛)的数量显著减少(Percy et al., 2002; Kopper & Lindroth, 2003). ...
Growth and physiological responses to ozone and mild drought stress of tree species with different ecological requirements 1 2010
... 除单因子O3胁迫外, 由于全球气候变化和极端气候常态化, 植物经常受到O3和其他环境因子交互作用的影响(如O3与CO2、升温、干旱或氮沉降等)(Schmadel-Hageb?lling et al., 1998; Ollinger et al., 2002; Eltayeb et al., 2006; Ghimire et al., 2018).然而, 植物应对O3与其他因子交互作用的响应并不是两者或单独作用的简单叠加, 往往取决于植物种类、生长时期、对特定环境因子的敏感性、胁迫浓度及时间, 以及具体的微气候条件(列淦文和薛立, 2014).一方面, 通过诱导气孔关闭、限制叶片O3吸收(CO2浓度升高/干旱胁迫), 或提高碳同化速率、促进植物光合作用(CO2浓度升高/氮增加), CO2浓度升高、干旱胁迫和氮沉降经常被报道可增强植物抵御O3的能力, 减缓O3伤害(白月明等, 2005; Handley & Grulke, 2008; Gao et al., 2017).另一方面, 与之相反, 也有研究发现长期高浓度CO2或干旱降低气孔导度的同时, 会引起植物蒸腾速率、光合产物合成和积累的降低(McDonald et al., 2002; Pollastrini et al., 2010); 而超过植物承载阈值的高氮负荷也会对植物的光合生理活性产生负效应(Yamaguchi et al., 2010), 进而加重植物在复合环境下的碳损失、水分蒸散损失和自由基积累, 对植物产生协同伤害(Karnosky et al., 2005; 列淦文和薛立, 2014).高峰(2018)认为O3与其他环境胁迫因子出现的先后次序也会影响植物对复合作用的响应. ...
Tree and stand growth of mature Norway spruce and European beech under long-term ozone fumigation 1 2010
... 有关自然环境下O3浓度升高对陆地生态系统影响的研究较为稀缺.但是现有开放式O3浓度增强系统(O3-FACE)观测平台下的长期研究也有着相似结果, Aspen FACE实验发现, O3浓度升高3-4年后, 欧洲山杨(Populus tremuloides)纯林、欧洲山杨-纸皮桦(Betula papyrifera)和欧洲山杨-糖枫(Acer saccharum)混交林的总生物量分别降低23%、13%和14% (Karnosky et al., 2003, 2005); 虽然在最初的7年内生物量的分配格局并未受O3影响(King et al., 2005), 但在实验11年后, 敏感型欧洲山杨几乎从森林冠层中消失(Kubiske et al., 2007).德国克兰兹堡郁闭冠层的成熟挪威云杉(Picea abies)和欧洲山毛榉(Fagus sylvatica)混交林(60年)对O3浓度升高长期响应的研究也表明, 8年O3暴露导致欧洲山毛榉的树干生产力降低了44% (Pretzsch et al., 2010).目前, 国内还没有研究涉及O3-FACE平台下O3浓度升高对森林生态系统碳汇能力的影响. ...
Quantifying plant response to ozone: a unifying theory 1 1987
... 地表O3浓度升高对植物光合作用、生长和生物量积累、物质分配等过程的连锁负效应导致陆地生态系统生产力和固碳能力降低(Reich, 1987; Wittig et al., 2009; Ainsworth et al., 2012), 削弱了自然生态系统的碳汇能力(Felzer et al., 2005; Sitch et al., 2007; Collins et al., 2010).基于大量OTCs控制实验下北半球中纬度北方温带树种的meta分析发现, O3浓度升高导致树木的净光合作用和总生物量分别降低11%和7% (Wittig et al., 2009).最近, 有关中国热带、亚热带和温带树种的meta分析则发现, O3浓度升高导致中国木本植物的净光合作用和总生物量分别降低28%和14% (Li et al., 2017). ...
Impacts of tropospheric ozone and climate change on net primary productivity and net carbon exchange of China’s forest ecosystems 1 2011
Indirect radiative forcing of climate change through ozone effects on the land-carbon sink 2 2007
... 地表O3浓度升高对植物光合作用、生长和生物量积累、物质分配等过程的连锁负效应导致陆地生态系统生产力和固碳能力降低(Reich, 1987; Wittig et al., 2009; Ainsworth et al., 2012), 削弱了自然生态系统的碳汇能力(Felzer et al., 2005; Sitch et al., 2007; Collins et al., 2010).基于大量OTCs控制实验下北半球中纬度北方温带树种的meta分析发现, O3浓度升高导致树木的净光合作用和总生物量分别降低11%和7% (Wittig et al., 2009).最近, 有关中国热带、亚热带和温带树种的meta分析则发现, O3浓度升高导致中国木本植物的净光合作用和总生物量分别降低28%和14% (Li et al., 2017). ...
Quantifying the impact of current and future tropospheric ozone on tree biomass, growth, physiology and biochemistry: a quantitative meta-analysis 2 2009
... 地表O3浓度升高对植物光合作用、生长和生物量积累、物质分配等过程的连锁负效应导致陆地生态系统生产力和固碳能力降低(Reich, 1987; Wittig et al., 2009; Ainsworth et al., 2012), 削弱了自然生态系统的碳汇能力(Felzer et al., 2005; Sitch et al., 2007; Collins et al., 2010).基于大量OTCs控制实验下北半球中纬度北方温带树种的meta分析发现, O3浓度升高导致树木的净光合作用和总生物量分别降低11%和7% (Wittig et al., 2009).最近, 有关中国热带、亚热带和温带树种的meta分析则发现, O3浓度升高导致中国木本植物的净光合作用和总生物量分别降低28%和14% (Li et al., 2017). ...
... 浓度升高导致树木的净光合作用和总生物量分别降低11%和7% (Wittig et al., 2009).最近, 有关中国热带、亚热带和温带树种的meta分析则发现, O3浓度升高导致中国木本植物的净光合作用和总生物量分别降低28%和14% (Li et al., 2017). ...
Mesophyll conductance limitation of photosynthesis in poplar under elevated ozone 1 2019
... 气孔防御机制失效后, 进入细胞的O3将诱导叶片内部抗氧化系统启动解毒和修复的非气孔防御响应(冯兆忠等, 2018).非气孔防御响应主要是指“质外体-共质体系统”内一些抗氧化组分对O3解毒和修复的防御过程.具体来说, O3进入植物叶片的细胞间隙后会溶解在细胞壁的结合水里, 经过一系列的化学反应形成活性氧分子(ROS), ROS能够破坏细胞壁并与细胞膜上的不饱和脂肪酸发生反应, 从而对叶片造成损伤(Tausz et al., 2007; Pellegrini et al., 2018).植物质外体的抗氧化物质如还原性抗坏血酸(ASC)、抗氧化酶(如超氧化物歧化酶)可与ROS反应进行初级解毒, 成为抵御O3的第二道防线(Kangasj?rvi et al., 2005; Dai et al., 2019), 而剩余的ROS会经过质膜到达共质体, 此时共质体内抗氧化成分如过氧化物酶(POD)、过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)及抗氧化剂(如酚类化合物-谷胱甘肽(GSH)、脯氨酸和多胺等)等抗氧化酶类物质被诱导作为次级解毒响应机制参与诸如“抗坏血酸-谷胱甘肽(ASA-GSH)循环”等氧化还原过程, 进而对ROS进行进一步解毒与消除(Luwe, 1996).气孔闭合除了会减少O3的吸收, 也会抑制CO2吸收和水汽交换, 而非气孔防御响应中的抗氧化系统解毒过程也会消耗大量的能量并对光合反应系统造成不同程度的损伤, 降低羧化速率和电子传递速率, 进而抑制光合和蒸腾作用, 损害植物生长(Gao et al., 2016).然而, 最新研究表明持续整个生长季的长期O3暴露会影响植物叶片的气孔密度、开度和大小等表观属性(李品等, 2018), 而O3从气孔进入到细胞间隙后, 也可以通过影响植物的叶肉导度降低植物的光合能力(Xu et al., 2019).此外, 有关29种落叶和常绿树种应对O3胁迫的研究还发现不同木本植物对O3响应的差异主要来自植物功能叶片比叶质量的大小(即叶片比叶质量越大的树种对O3越不敏感), 与气孔导度无明显相关性(Li et al., 2016). ...
Effects of ozone on nitrogen metabolism in the leaves of Fagus crenata seedlings under different soil nitrogen loads 1 2010
... 除单因子O3胁迫外, 由于全球气候变化和极端气候常态化, 植物经常受到O3和其他环境因子交互作用的影响(如O3与CO2、升温、干旱或氮沉降等)(Schmadel-Hageb?lling et al., 1998; Ollinger et al., 2002; Eltayeb et al., 2006; Ghimire et al., 2018).然而, 植物应对O3与其他因子交互作用的响应并不是两者或单独作用的简单叠加, 往往取决于植物种类、生长时期、对特定环境因子的敏感性、胁迫浓度及时间, 以及具体的微气候条件(列淦文和薛立, 2014).一方面, 通过诱导气孔关闭、限制叶片O3吸收(CO2浓度升高/干旱胁迫), 或提高碳同化速率、促进植物光合作用(CO2浓度升高/氮增加), CO2浓度升高、干旱胁迫和氮沉降经常被报道可增强植物抵御O3的能力, 减缓O3伤害(白月明等, 2005; Handley & Grulke, 2008; Gao et al., 2017).另一方面, 与之相反, 也有研究发现长期高浓度CO2或干旱降低气孔导度的同时, 会引起植物蒸腾速率、光合产物合成和积累的降低(McDonald et al., 2002; Pollastrini et al., 2010); 而超过植物承载阈值的高氮负荷也会对植物的光合生理活性产生负效应(Yamaguchi et al., 2010), 进而加重植物在复合环境下的碳损失、水分蒸散损失和自由基积累, 对植物产生协同伤害(Karnosky et al., 2005; 列淦文和薛立, 2014).高峰(2018)认为O3与其他环境胁迫因子出现的先后次序也会影响植物对复合作用的响应. ...
Ozone and haze pollution weakens net primary productivity in China 1 2017
 ,
, ,1,*, 袁相洋2, 李品2, 尚博2, 平琴2, 胡廷剑2, 刘硕2
,1,*, 袁相洋2, 李品2, 尚博2, 平琴2, 胡廷剑2, 刘硕2
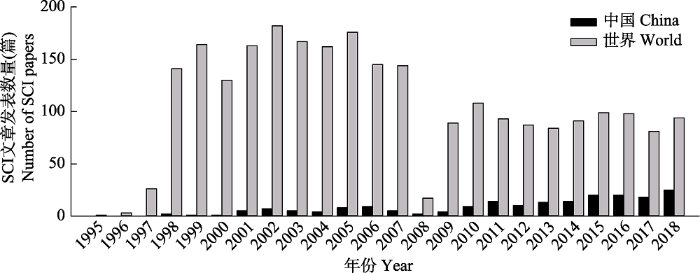 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT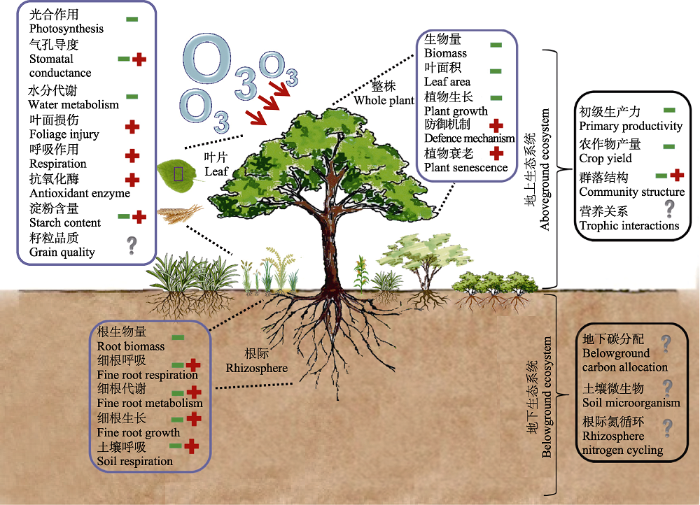 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}