纪若璇, 于笑, 常远, 沈超, 白雪卡, 夏新莉, 尹伟伦, 刘超
, * 北京林业大学生物科学与技术学院林木育种国家工程实验室, 北京 100083
Geographical provenance variation of leaf anatomical structure of Caryopteris mongholica and its significance in response to environmental changes Ruo-Xuan JI, Xiao YU, Yuan CHANG, Chao SHEN, Xue-Qia BAI, Xin-Li XIA, Wei-Lun YIN, Chao LIU
, * National Engineering Laboratory of Tree Breeding, College of Biological Sciences and Technology, Beijing Forestry University, Beijing 100083, China
通讯作者: *
liuchao1306@163.com 编委: 罗天祥
责任编辑: 李敏
收稿日期: 2019-05-18
接受日期: 2020-01-29
网络出版日期: 2020-03-20
基金资助: 国家水体污染控制与治理科技重大专项 (2017ZX07101002 )国家自然科学基金 (31600484 )国家自然科学基金 (31770649 )
Corresponding authors: *
liuchao1306@163.com Received: 2019-05-18
Accepted: 2020-01-29
Online: 2020-03-20
Fund supported: National Science and Technology Major Project for Water Pollution Control and Treatment (2017ZX07101002 )National Natural Science Foundation of China (31600484 )National Natural Science Foundation of China (31770649 )
摘要 长期受到生长环境影响而形成的遗传变异对植物生长发育有着显著的影响。叶片是植物对环境变化最敏感的器官, 了解叶片解剖结构在不同环境中产生的适应性变异是探索植物对环境适应的基础。同质园试验是研究遗传与环境因素对植物生长代谢等影响的一种有效方法, 该研究利用同质园试验排除了环境梯度的影响, 通过常规石蜡切片、多重比较、相关性分析、一般线性模型分析等方法, 对7个不同种源地的蒙古莸(
Caryopteris mongholica )叶片解剖结构及其影响因素进行了定量比较。结果表明, 7个种源地的蒙古莸叶片均为等面叶, 无海绵组织分化, 其上表皮细胞较下表皮细胞厚, 上栅栏组织较下栅栏组织厚; 叶片各解剖结构参数间存在显著的自相关性, 不同种源叶片解剖结构存在显著差异: 随种源地年平均气温升高, 叶厚度、栅栏组织厚度呈增大趋势, 其中, 最西南部的阿左旗种源蒙古莸叶片的上下栅栏组织、叶厚度及叶片结构紧密度值均最大, 表现出明显的抗旱特征。种源地经纬度、气温、降水等对解剖结构指标有显著的影响, 其解释程度为34.09%-81.43%。同质园试验说明, 种源地气候差异驱动的遗传变异是引起不同种源叶片解剖结构差异的重要因素。
关键词: 蒙古莸 ;
同质园 ;
叶片 ;
解剖结构 ;
种源 ;
遗传变异 Abstract Aims The genetic variation caused by long-term adaptation to the environment has significant influence on plant’s growth and development. Leaves serve as the most important and sensitive organs to environmental changes. Understanding the adaptive variation of leaf anatomical structure in different environments is the basis for exploring the adaptation of plants to the environment.
Methods Common garden experiment is an effective method to study the effects of genetic and environmental factors on plant growth and metabolism. In this study, the influence of environmental gradients was eliminated by using the common garden experiment. Then we analyzed anatomical structures of leaves and its driving factors of
Caryopteris mongholica from seven different provenances by using conventional paraffin section, multiple comparisons, correlation analysis, and general linear model analysis method.
Important findings Caryopteris mongholica from seven provenances had typical isolateral leaves, with no differentiation of spongy tissue, and the upper epidermal cell was thicker than the lower epidermal cell. Meanwhile, the upper palisade tissue was thicker than the lower palisade tissue. There was a significant autocorrelation among anatomical parameters of leaves, and there were important differences among the anatomical structures of leaves from different provenances. With increasing mean annual temperature of provenances, the thickness of the leaves and palisade also showed an increasing trend, and the largest thickness of the upper palisade tissues, lower palisade tissues and leaf thickness were in the leaves of Alxa Left Banner, Nei Mongol, indicating a stronger resistance to drought. The latitude, longitude, temperature and precipitation of different provenances had significant effects on the index of anatomical structure, with the explanatory power from 34.09% to 81.43%. The common garden experiment showed that the genetic variation driven by climate was one of the major factors that caused the difference in anatomical structure of leaves from different provenances.
Keywords: Caryopteris mongholica ;
;
leaf ;
anatomical structure ;
provenance ;
genetic variation PDF (1327KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 纪若璇, 于笑, 常远, 沈超, 白雪卡, 夏新莉, 尹伟伦, 刘超. 蒙古莸叶片解剖结构的地理种源变异及其对环境变化响应的意义. 植物生态学报, 2020, 44(3): 277-286. DOI:
10.17521/cjpe.2019.0117 JI Ruo-Xuan, YU Xiao, CHANG Yuan, SHEN Chao, BAI Xue-Qia, XIA Xin-Li, YIN Wei-Lun, LIU Chao.
Geographical provenance variation of leaf anatomical structure of Caryopteris mongholica and its significance in response to environmental changes .
Chinese Journal of Plant Ecology , 2020, 44(3): 277-286. DOI:
10.17521/cjpe.2019.0117 植物在进化过程中, 受到外界温度、水分、光照等环境因素的长期作用而影响其建成, 逐步形成适应环境的表型和遗传特征。即使是同一物种, 长期生长在不同的地理环境中, 也会产生不同程度的基因分化, 以适应各自不同的生态位, 从而形成特定的种源特性(
Smith et al ., 1997 ;
Barboni et al ., 2004 ;
Pigliucci, 2006 ;
胡梦瑶等, 2012 )。植物自然群体分布的环境具有明显差异。从表型来看, 同一结构由于环境分化选择作用, 不同环境条件下的群体最适表型也存在差异, 而表型的差异归根结底是由控制表型的基因型不同所导致的。
叶片作为植物暴露在生活环境中面积最大的器官, 其外部形态特征、内部解剖结构等最能体现植物对环境因子的适应性进化(
Wright et al ., 2001 ;
Chartzoulakis et al ., 2002 ;
钟悦鸣等, 2017 ;
于海燕等, 2019 ), 开展叶片解剖结构分析, 对研究植物在极端生态环境下的适应策略具有重要意义。一般而言, 植物分布范围广泛, 各自生活在温度、水分等条件不相同的自然地理环境中, 在长期对环境适应过程中, 同种植物叶片的解剖结构表现出明显的差异(
季子敬等, 2013 ;
郭婧宇, 2018 )。叶片的这种功能性状差异可能既有自身的可塑性响应也有遗传变异, 以往研究中沿自然环境梯度的采样测定或单一站点控制实验很难区分两者的效应。全缘叶绿绒蒿(
Meconopsis integrifolia )随着海拔升高, 株型变矮, 叶片组织结构紧密度增大(
刘梦颖等, 2018 ); 生长于干旱地区的长梗扁桃(
Amygdalus pedunculata )叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的长梗扁桃, 其叶片栅栏组织排列疏松, 叶厚度较薄(
郭改改等, 2013 )。这种叶片解剖结构特征的差异同时受到环境变异与自身遗传结构两个因素的影响。同质园试验即研究同一基因型的物种在不同环境下的初生与次生代谢的响应, 或不同基因型的物种在相同环境下的初生与次生代谢的响应。将多个种源的植物在同地栽培, 在一定程度上控制环境与基因型间的相互作用, 最大程度地降低立地条件所造成的影响(
王薪琪等, 2015 ;
陈嘉静等, 2019 ), 进而说明不同种源植物叶片解剖结构特征表现出的差异是由于原种源地环境差异造成的植物遗传性基因变异。这与先前关于环境和遗传因素共同影响(包括气候梯度和种源的影响)叶片解剖结构特征的研究有所不同, 这种同质园方法在局部适应性研究中具有重要意义(
de Villemereuil, 2016 ), 有助于我们更好地理解不同地理种源植物对全球变化的响应和适应机制。目前采用此法对不同种源内植物叶片解剖结构差异及其影响因素的相关分析较少。
蒙古莸(
Caryopteris mongholica )属唇形科筋骨草亚科莸属(
Caryopteris ), 是我国干旱、半干旱地区一种耐旱、耐寒兼具观赏性的绿化灌木, 是防风固沙和水土保持的优良造林树种(
李玉俊等, 1991 ;
韩磊等, 2008 )。蒙古莸分布在东起呼伦贝尔典型草原区, 西至甘肃河西走廊西部和青海湖南部的广阔的蒙古高原上。然而, 不同的生长环境导致蒙古莸的生理特征和遗传结构均出现不同程度的变异, 这种对环境的适应性变异表现, 在叶片的解剖结构上尤为明显。
白雪卡等(2018) 对同质园栽培下8个不同种源蒙古莸光响应曲线的研究发现, 蒙古莸光合响应能力在不同种源间表现出明显不同, 光响应曲线也存在显著的差异。蒙古莸这种对环境的适应性变异在叶片的解剖结构上尤为明显, 在前人不同的研究中, 蒙古莸或为等面叶、无海绵组织分化(
符亚儒等, 2005 ;
杨小玉, 2008 ;
李爱平等, 2010 ), 或为异面叶、叶肉中含有海绵组织(
张源润等, 1999 ;
王晓江等, 2006 ;
田英等, 2010 ); 叶片厚度21.6-2 180 μm, 变异极大。
为了研究不同种源植物叶片解剖结构的影响机制, 阐释干旱地区植物对气候变化的响应与适应, 我们将来自华北和西北7个地区的蒙古莸种植在同一环境条件下, 排除外界环境因素差异的作用, 采用石蜡切片法对蒙古莸叶片解剖结构特征进行研究, 探讨以下问题: (1)不同种源蒙古莸叶片解剖结构的差异性; (2)种源地环境条件对叶片解剖结构的影响。以期进一步探讨植物的局域环境适应性进化, 为今后蒙古莸的种质资源保护、遗传改良以及环境变异驱动的植物基因型差异对植物的作用机制提供理论依据。
1 材料和方法 1.1 研究地概况 试验地点位于北京林业大学苗圃(40.01° N, 116.34° E), 属暖温带半湿润大陆性季风气候。年平均气温11.8 ℃, 年最高气温37.5-42.6 ℃, 最低气温-19.5- -14.8 ℃, 年降水量500-650 mm, 多集中于夏季, 占全年降水量的70%, 而春旱情况严重。试验地设置塑料遮雨棚以防止其他水分影响, 雨天关闭, 晴天打开。
1.2 试验材料 试验材料为蒙古莸一年生扦插苗。于2016年3月下旬, 在内蒙古阿巴嘎旗、阿拉善左旗(阿左旗)、东乌旗、鄂托克旗蒙西镇、凉城、河北康保县、陕西神木市(种源地地理环境见
表1 )采取生长良好的蒙古莸植株, 剪取长度约为20 cm左右的枝条茎段, 放置于湿布中保存, 立即带回北京。将不同种源地的扦插枝条经0.1%生根粉ABT1 (中国林业科学研究院林业研究所)处理后扦插于450 mm × 450 mm的花盆中。种植土为沙壤土(草炭土:壤土:沙子体积比为4:4:3), 与蒙古莸的野外土壤成分相似, 所有种源材料种植在同一土壤条件下。
Table 1 表1 表1 蒙古莸种源地基本情况
Table 1
Basic conditions of Caryopteris mongholica provenances 种源地 纬度 经度 海拔 年降水量MAP (mm) 年平均气温MAT (℃) 生长季降水量GSP (mm) 生长季平均气温GST (℃) 最冷月平均气温TCM (℃) 潜在蒸散量PE (mm) 内蒙古阿巴嘎旗 43.90 115.35 1 177 224.03 2.9 434.71 17.74 -19.85 588.04 内蒙古阿拉善左旗 38.88 105.72 1 670 162.54 9.90 315.52 21.05 -7.51 712.81 内蒙古东乌旗 45.73 116.79 1 017 200.72 2.46 415.46 17.76 -20.52 531.06 内蒙古蒙西 40.08 106.92 1 193 118.00 8.92 243.64 20.95 -9.58 702.17 内蒙古凉城 40.66 112.30 1 429 313.75 5.18 611.46 17.13 -13.17 580.80 陕西神木 39.29 110.33 1 209 369.79 9.78 729.85 20.85 -7.64 706.91 河北康保 41.99 114.85 1 590 279.72 3.82 556.53 16.78 -15.22 544.06
GSP, growth season precipitation; GST, growth season temperature; MAP, mean annual precipitation; MAT, mean annual temperature; PE, potential evaporation; TCM, temperature of coldest month.新窗口打开 |
下载CSV 2017年9月中旬, 每个种源挑选长势相似的扦插苗3-5株, 每株取枝条中部成熟叶片3片, 于叶片主脉两侧剪取约1 cm × 1 cm的正方形叶块, 放入FAA固定液(甲醛:乙酸:70%乙醇= 1:1:18)中, 迅速带回实验室用于叶片解剖结构测定。
1.3 试验方法 采用常规石蜡切片法(
冯永庆等, 2008 )制作石蜡切片。将固定24 h以上的叶块, 经70%、85%、95%、100%四个浓度酒精脱水, 二甲苯透明, 石蜡浸透包埋(包埋所用石蜡熔点为56-57 ℃), 待凝固后切片, 切片厚度8-10 μm, 番红-固绿双重染色, 中性树胶封片, 将其制作为永久切片(
de Lima Silva et al ., 2014 )。在Leica DM2500显微镜(Leica, Wezlar, Germany)下观察, 并用Leica LAS AF软件拍照记录。
用Image J软件测量叶片的上角质层厚度, 上、下表皮细胞厚度, 上、下栅栏组织厚度, 叶厚度等叶解剖结构指标, 并计算叶片结构紧密度、栅栏组织总厚度(
李芳兰和包维楷, 2005 ), 每个指标重复测量5组数据。
叶片解剖结构指标:
叶片结构紧密度(
Tight ) =栅栏组织厚度/叶片厚度× 100%
栅栏组织总厚度=上栅栏组织厚度+下栅栏组织厚度
叶片解剖结构可塑性指标:
变异系数=标准差/算数平均值
可塑性指数= (最大值-最小值)/最大值
根据蒙古莸不同种源地的经度与纬度, 从ArcGIS 10.2的WorldCLIM全球高分辨率气候数据库(
Hijmans et al ., 2005 )中选取各样地的月平均气候数据(2007-2016年), 并计算各气候指标大小。气候指标包括: 年降水量(
MAP )、生长季降水量(
GSP )、降水年较差(
ARP )、年平均气温(
MAT )、生长季平均气温(
GST )、气温年较差(
ART )、潜在蒸散量(
PE )。
1.4 数据分析 用方差分析多重比较法, 比较不同种源蒙古莸叶片解剖结构参数差异。用Pearson相关性分析方法分析叶片解剖结构参数与种源地环境因子的相关关系。
结合一般线性模型(GLM) ANOVA分析种源地气候对蒙古莸叶解剖结构参数值的影响作用大小(
He et al ., 2009 )。基于种源地环境因素与叶片解剖结构参数值的相关关系, 选取
MAP 、
MAT 、
PE 、海拔(
Alt )、地理位置(Site)等5个种源地气候环境因子进入模型。
数据分析在Excel、R 3.2.2中完成。
2 结果 2.1 蒙古莸叶片解剖结构特征 观察蒙古莸叶片解剖结构, 7个种源的蒙古莸叶片均为等面叶, 叶片厚度在192.34-270.30 μm之间(
表2 ); 其内部结构可分为表皮、叶肉和叶脉3个基本部分(
图1 )。上表皮(UEC)、下表皮(LEC)均由单层细胞组成, 细胞通常呈长方形或矩圆形, 上表皮细胞厚度9.87-26.46 μm, 其变异系数与可塑性指数最大; 下表皮细胞厚度8.74-17.64 μm, 所有种源地上表皮细胞厚度均大于下表皮细胞厚度; 叶表面密被表皮毛; 上、下表皮细胞外均有角质层, 但上表皮细胞外壁角质层较厚, 上角质层(Cuticle)厚度5.07-11.92 μm; 叶肉由发达的栅栏组织(PT)组成, 无海绵组织。栅栏组织一般为3-4层长柱状薄壁细胞, 排列紧密, 上栅栏组织(UPT)厚度89.24-146.64 μm, 下栅栏组织(LPT)厚度59.57-109.42 μm, 上栅栏组织厚度大于下栅栏组织厚度; 叶片结构紧密度(
Tight )范围0.74-0.90, 其变异系数与可塑性指数最小(
表2 )。主脉含有一个或几个维管束, 其中木质部(MVX)位于近轴面, 韧皮部(MVP)位于远轴面。从
表3 可以看出, 不同种源蒙古莸叶片解剖结构的多数参数值具有明显差异, 且波动范围较大。
Table 2 表2 表2 蒙古莸叶片解剖结构指标
Table 2
Leaf anatomical indices of Caryopteris mongholica 解剖结构指标 观测数目 平均值 标准偏差 均值的95%置信区间 最大值 最小值 变异系数 可塑性指数 上角质层厚度 Cuticle thickness (μm) 70 6.87 1.66 6.05-7.70 11.92 5.07 0.24 0.57 上表皮细胞厚度 UEC thickness (μm) 70 18.53 5.37 15.86-21.20 26.46 9.87 0.29 0.63 下表皮细胞厚度 LEC thickness (μm) 70 11.14 2.32 9.99-12.30 17.64 8.74 0.21 0.50 上栅栏组织厚度 UPT thickness (μm) 70 106.35 17.94 97.43-115.27 146.64 89.24 0.17 0.39 下栅栏组织厚度 LPT thickness (μm) 70 76.17 13.44 69.48-82.85 109.42 59.57 0.18 0.46 栅栏组织总厚度 PT thickness (μm) 70 182.52 29.16 168.02-197.02 243.12 156.05 0.16 0.36 叶厚度 LT (μm) 70 220.02 25.71 207.24-232.80 270.30 192.34 0.12 0.29 叶片结构紧密度 Tight (%) 70 0.83 0.04 0.80-0.85 0.90 0.74 0.05 0.18
LEC, lower epidermal cell; LPT, lower palisade tissue; LT, leaf thickness; PT, palisade tissue; UEC, upper epidermal cell; UPT, upper palisade tissue. 新窗口打开 |
下载CSV 图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1蒙古莸叶片横切面。A, 叶肉结构。B, 主脉结构。C, 主脉及叶肉结构。Co, 厚角组织; Cuticle, 上角质层; LEC, 下表皮细胞; LPT, 下栅栏组织; MVP, 主脉韧皮部; MVX, 主脉木质部; St, 孔下室; UEC, 上表皮细胞; UPT, 上栅栏组织。 Fig. 1Leaf cross section of Caryopteris mongholica. A, Mesophyll structure. B, Main vein structure. C, Mesophyll and main vein structure. Co, collenchyma; LEC, lower epidermal cell; LPT, lower palisade tissue; MVP, main vein phloem; MVX, main vein xylem; St, stomatal chamber; UEC, upper epidermal cell; UPT, upper palisade tissue. Table 3 表3 表3 7个种源蒙古莸叶片解剖结构特征参数(平均值±标准误差)
Table 3
Leaf anatomical parameters of Caryopteris mongholica from seven provenances (mean ± SE ) 种源地 上角质层厚度 上表皮 下表皮 上栅栏 下栅栏 栅栏组织总厚度 叶厚度 LT (μm) 叶片结构紧密度 Tight (%) 内蒙古阿巴嘎旗 8.44 ± 1.74ab 19.63 ± 1.33ab 13.98 ± 2.08a 95.13 ± 1.52cd 63.87 ± 2.15d 159.01 ± 2.07d 200.23 ± 0.56c 0.797 ± 0.012c 内蒙古阿拉善左旗 5.92 ± 0.76bc 13.35 ± 1.21c 9.81 ± 0.54a 137.94 ± 4.35a 102.50 ± 4.58a 240.44 ± 1.56a 269.52 ± 0.60a 0.893 ± 0.006a 内蒙古东乌旗 5.83 ± 0.97bc 22.76 ± 1.23a 12.81 ± 2.04a 100.62 ± 6.13cd 70.81 ± 2.13bc 171.43 ± 7.51cd 212.83 ± 7.49bc 0.803 ± 0.009bc 内蒙古蒙西 5.09 ± 0.20c 16.19 ± 2.17bc 10.61 ± 0.69a 115.80 ± 7.09b 72.99 ± 0.51bc 188.79 ± 7.59c 220.67 ± 8.06b 0.857 ± 0.007ab 内蒙古凉城 8.82 ± 0.35a 23.69 ± 1.27a 11.29 ± 0.08a 104.91 ± 3.71bc 67.20 ± 7.11bc 172.11 ± 10.46cd 224.58 ± 2.62b 0.767 ± 0.038c 陕西神木 6.45 ± 0.69abc 12.42 ± 2.39c 10.09 ± 1.50a 90.30 ± 1.11d 78.22 ± 4.04b 168.52 ± 4.57d 197.48 ± 5.33c 0.853 ± 0.001ab 河北康保 6.73 ± 0.80abc 23.08 ± 1.74a 10.34 ± 0.85a 96.90 ± 3.03cd 68.08 ± 0.17bc 164.98 ± 2.90d 211.67 ± 5.12bc 0.780 ± 0.023c
LEC, lower epidermal cell; LPT, lower palisade tissue; LT, leaf thickness; PT, palisade tissue; UEC, upper epidermal cell; UPT, upper palisade tissue. Letters in the table indicate the Duncan multiple comparison test grouping performed by one-dimensional analysis of variance for each anatomical parameter, with different letters of the same column indicating significant differences (p < 0.05). 表中字母为各解剖结构参数的一维方差分析进行的Duncan多重比较检验分组, 同列不同字母表示差异显著(p < 0.05)。 新窗口打开 |
下载CSV 2.2 叶片解剖结构参数间的自相关性 由
表4 可得, 多数叶片解剖结构参数间存在明显的自相关, 其中, UPT厚度、LPT厚度、PT厚度、
LT 以及
Tight 这5组参数两两之间都极显著正相关(
p < 0.01)。此外,
Cuticle 与LEC厚度间也表现出极显著的相关性(
p < 0.01), 但这两个参数与其他参数均无相关关系。
Table 4 表4 表4 蒙古莸叶片解剖性状相关性分析
Table 4
Correlation analyses of Caryopteris mongholica leaf anatomical characters 相关系数 上角质层 上表皮 下表皮 上栅栏 下栅栏 栅栏组织 叶厚度 LT (μm) 叶片结构Tight (%) 上角质层厚度 Cuticle thickness (μm) 上表皮细胞厚度 UEC thickness (μm) 0.201 下表皮细胞厚度 LEC thickness (μm) 0.590** 0.107 上栅栏组织厚度 UPT thickness (μm) -0.246 -0.355 -0.203 下栅栏组织厚度 LPT thickness (μm) -0.434 -0.545* -0.436 0.721** 栅栏组织总厚度 PT thickness (μm) -0.351 -0.470* -0.326 0.948** 0.904** 叶厚度 LT (μm) -0.260 -0.268 -0.259 0.945** 0.825** 0.962** 叶片结构紧密度 Tight (%) -0.432 -0.770* -0.391 0.683** 0.806** 0.792** 0.595**
LEC, lower epidermal cell; LPT, lower palisade tissue; LT, leaf thickness; PT, palisade tissue; UEC, upper epidermal cell; UPT, upper palisade tissue. **, p < 0.01; *, p < 0.05. 新窗口打开 |
下载CSV 2.3 不同种源蒙古莸叶片解剖结构参数间的差异 由
表3 可得, 除下表皮厚度这一指标外, 其他7个叶片解剖指标间表现出显著的差异(
p < 0.05)。凉城的蒙古莸叶片的
Cuticle 、UEC厚度值最大, 蒙西的
Cuticle 值最小, 神木的UEC厚度值最小; 阿巴嘎的LEC厚度值最大; 阿左旗的UPT厚度、LPT厚度、PT厚度以及
LT 值均最大, 其
Tight 值亦是最大。
2.4 种源地环境对蒙古莸解剖结构参数的影响 由
表5 可得, 蒙古莸叶片解剖结构指标与一个或多个种源地地理位置(经纬度)、温度、降水之间具有显著的相关性。
Table 5 表5 表5 种源地环境因素与蒙古莸叶片解剖结构参数的相关系数
Table 5
Correlation coefficient of environmental conditions in provenances and leaf anatomical characters of Caryopteris mongholica 相关系数 上角质层厚度 上表皮 下表皮 上栅栏 下栅栏 栅栏组织 叶厚度 LT (μm) 叶片结构Tight (%) 纬度 Latitude (°N) 0.328 0.638** 0.451 -0.526* -0.694** -0.643** -0.529* -0.724** 经度 Longitude (°E) 0.437 0.704** 0.366 -0.787** -0.801** -0.854** -0.730** -0.886** 海拔 Altitude (m) -0.124 -0.034 -0.358 0.547* 0.579* 0.604** 0.690** 0.237 年降水量 MAP (mm) 0.261 0.074 -0.103 -0.671** -0.250 -0.528* -0.555* -0.307 生长季降水量 GSP (mm) 0.244 0.085 -0.110 -0.689** -0.269 -0.548* -0.575* -0.321 降水年较差 ARP (mm) 0.348 0.195 -0.064 -0.640** -0.236 -0.503* -0.495* -0.365 年平均气温 MAT (℃) -0.446 -0.803** -0.421 0.534* 0.735** 0.668** 0.495* 0.860** 生长季平均气温 GST (℃) -0.476* -0.863* -0.290 0.512* 0.672** 0.625** 0.424 0.880** 气温年较差 ART (℃) 0.338 0.503* 0.529* -0.426 -0.626** -0.551* -0.465 -0.597**
LEC, lower epidermal cell; LPT, lower palisade tissue; LT, leaf thickness; PT, palisade tissue; UEC, upper epidermal cell; UPT, upper palisade tissue. ARP, annual relatively precipitation; ART, annual relatively temperature; GSP, growth season precipitation; GST, growth season temperature; MAP, mean annual precipitation; MAT, mean annual temperature. ***, p < 0.001; **, p < 0.01; *, p < 0.05. 新窗口打开 |
下载CSV 经度、纬度除与UEC厚度呈正相关关系外, 与UPT厚度、LPT厚度、PT厚度、
LT 、
Tight 5个参数呈负相关关系(
p < 0.05)。地理梯度上呈现自西向东、自南向北, UEC厚度逐渐增大, UPT厚度、LPT厚度、PT厚度、
LT 、
Tight 逐渐减小的趋势。
UPT厚度、LPT厚度、PT厚度、
LT 、
Tight 均与
MAT 显著正相关(
p < 0.05); 而UEC厚度与
MAT 、
GST 显著负相关(
p < 0.05), 与
ART 显著正相关(
p < 0.05)。即随环境温度升高, UPT厚度、LPT厚度、PT厚度、
LT 、
Tight 增大, UEC厚度减小。
降水指标(
MAP 、
GSP 、
ARP )仅与UPT厚度(
p < 0.01)、
LT (
p < 0.05)、PT厚度(
p < 0.05)显著负相关。
2.5 种源地气候对各解剖结构参数的GLM分析 通过
表6 中ANOVA GLM分析可知, 原种源地气候(温度、降水、蒸散)驱动的遗传变异对蒙古莸各解剖结构参数有显著作用, 其解释度在34.09%- 81.43%间, 对LEC厚度的解释度最小(34.09%), 对
Tight 的解释度最大(81.43%)。种源地气候因子中,
MAT 对所选8个叶片解剖结构指标均有显著影响, 其中, 对UEC厚度、UPT厚度、LPT厚度、PT厚度、
LT 、
Tight 这6个指标有极显著影响(
p < 0.001), 解释程度在21.69%-71.89%间。
PE 对UEC厚度、UPT厚度、LPT厚度、PT厚度、
LT 有显著影响(
p < 0.05), 解释程度在5.48%-14.34%间。
MAP 对UPT厚度、LPT厚度、PT厚度、
LT 、
Tight 显著影响的解释程度在4.78%-45.93%间, 其中, 对UPT厚度、PT厚度、
LT 这3个指标有极显著影响(
p < 0.001)。海拔对UPT厚度、LPT厚度、PT厚度
、LT 有显著影响(
p < 0.01), 解释程度在8.33%-16.76%间。温度对蒙古莸叶片解剖结构参数的解释度大于水分与海拔。
Table 6 表6 表6 种源地气候(年降水量、年平均气温、潜在蒸散量、海拔、位置)对蒙古莸叶片各个解剖结构参数的GLM分析
Table 6
Summary of general linear models for the effect of climate variations in different provenances (MAP, MAT, PE, Alt, Site) on individual leaf anatomical characters of Caryopteris mongholica 气候因子 上角质层厚度 Cuticle thickness 上表皮细胞厚度 UEC thickness 下表皮细胞厚度 LEC thickness 叶厚度 LT df %SS Sig. df %SS Sig. df %SS Sig. df %SS Sig. 年降水量 MAP 1 10.13 0.147 1 0.12 0.804 1 0.98 0.646 1 31.40 6.8 × 10-6*** 年平均气温 MAT 1 20.77 0.049* 1 65.82 2.3 × 10-4*** 1 16.78 0.082* 1 21.69 3.0 × 10-5*** 潜在蒸散量 PE 1 4.08 0.339 1 9.20 0.055* 1 16.33 0.085 1 14.34 1.5 × 10-4*** 海拔 Alt 1 2.74 0.430 1 0.00 0.999 1 0.08 0.894 1 16.76 8.3 × 10-5*** 位置 Site 4 26.20 0.248 4 7.86 0.438 4 26.52 0.276 4 12.50 0.004** 残差 Residuals 9 36.09 9 17 9 39.30 9 3.31 气候因子 上栅栏组织厚度 UPT thickness 下栅栏组织厚度 LPT thickness 栅栏组织总厚度 PT thickness 叶片结构紧密度 Tight df %SS Sig. df %SS Sig. df %SS Sig. df %SS Sig. 年降水量 MAP 1 45.93 4.7 × 10-6*** 1 4.78 0.023* 1 27.38 1.6 × 10-7*** 1 8.93 0.028* 年平均气温 MAT 1 24.59 5.8 × 10-5*** 1 57.74 5.5 × 10-6*** 1 44.37 1.9 × 10-8*** 1 71.89 4.1 × 10-5*** 潜在蒸散量 PE 1 6.28 0.006** 1 5.48 0.017* 1 6.78 5.2 × 10-5*** 1 0.61 0.514 海拔 Alt 1 8.65 0.002** 1 8.33 0.006** 1 9.66 1.3 × 10-5*** 1 0.12 0.772 位置 Site 4 10.13 0.019* 4 17.91 0.008** 4 10.62 1.6 × 10-4*** 4 6.65 0.097* 残差 Residuals 9 4.41 9 5.76 9 1.18 9 11.81
LEC, lower epidermal cell; LPT, lower palisade tissue; LT, leaf thickness; PT, palisade tissue; UEC, upper epidermal cell; UPT, upper palisade tissue. Alt, altitude; MAP, mean annual precipitation; MAT, mean annual temperature; PE, potential evaporation. ***, p < 0.001; **, p < 0.01; *, p < 0.05。 df, 自由度; %SS, 解释度; Sig., 显著性。新窗口打开 |
下载CSV 3 讨论 3.1 不同种源间的叶解剖结构特征 叶片是反映植物在不同生态环境条件下特点的首要器官, 其形态结构是自身生理代谢的需要和对外界环境的逐渐适应, 也是植物对环境变化最为敏感的一个器官(
覃凤飞等, 2012 )。同一种植物生长在不同的环境中, 叶片的解剖结构会表现出不同程度的分化差异。
本研究通过对同质园栽培下7个种源蒙古莸叶片解剖结构的观察, 发现7个种源地的蒙古莸叶片均为典型的等面叶, 叶片由紧密排列的上表皮、发达的栅栏组织和下表皮构成, 无海绵组织分化, 是旱生植物最常见的叶片结构(
Delzon, 2015 ), 与
贺一鸣(2018) 、
杨小玉(2008) 、
李爱平等(2010) 、
符亚儒等(2005) 对蒙古莸叶片解剖结构观察结果一致。蒙古莸叶片解剖结构参数的数量化分析表明, 上表皮细胞厚度均大于下表皮细胞厚度, 这种结构有利于气体交换(
陈旭等, 2019 ); 7个种源蒙古莸叶片的7个解剖结构参数存在显著差异(
表3 ), 其中, 叶厚度变化范围为192.34-270.30 μm, 变异系数为0.12, 与
符亚儒等(2005) 、
李爱平等(2010) 的测量结果近似。
3.2 叶解剖结构与种源地环境因子的关系 原种源地的地理位置(经纬度)、温度(
MAT 、
ART )、
MAP 、
PE 、
Alt 与叶片解剖结构指标密切相关(
表6 ), 能够很好地解释其遗传变异程度, 其中种源地气候(
MAT 、
MAP 、
PE )对蒙古莸叶片解剖结构解释程度在34.09%-81.43%之间。
季子敬等(2013) 对兴安落叶松(
Larix gmelinii )针叶解剖结构的分析研究中发现, 针叶长期适应当地环境条件后, 在解剖结构特征上也产生了具有遗传能力的变异。
赵小仙等(2015) 对蒙古沙拐枣(
Calligonum monglicum )叶片解剖结构分析研究中发现, 解剖结构的差异与经纬度、
MAT 、
MAP 的变化有极显著相关性。
白雪卡等(2018) 对不同种源蒙古莸光响应特性的研究中也同样证明, 种源地气候驱动的遗传变异对蒙古莸光合作用参数有显著的解释力, 解释程度在16.65%- 73.52%之间, 与本研究的解释力近似。本研究中, 上角质层厚度、上下表皮细胞厚度、下栅栏组织厚度、栅栏组织总厚度、叶片结构紧密度主要受
MAT 的影响, 解释程度在20.77%-71.89%之间(
表6 ); 上栅栏组织厚度、叶厚度主要受
MAP 影响, 解释程度在31.40%-45.93%之间(
表6 )。相对而言, 温度对蒙古莸叶片解剖结构参数的解释度大于水分, 这与
田丽丽等(2019) 对大叶种茶树叶片解剖结构的分析不谋而合, 其研究发现叶片生长受到温度、降水和光照因子等综合因素的影响, 叶片结构的差异则主要受到温度影响。
3.3 叶解剖结构的适应性 本研究中的
MAT 与叶片结构紧密度相关性最强(
表5 ), 叶片结构紧密度随温度升高而增大, 这可能是由于高温时植物呼吸与代谢作用加强, 加剧了水分散失, 导致叶肉组织向着提高光合效率和水分储量方向发展。而发达的栅栏组织可以有效地储存水分, 栅栏组织的细胞排列越紧密, 细胞间隙越少, 叶片结构紧密度就越大, 能更好地适应高温环境。最西南部的阿拉善左旗属高温少雨地区, 潜在蒸散量最大, 导致该种源的蒙古莸叶片的上下栅栏组织厚度、叶厚度以及叶片结构紧密度均最大(
表1 ,
表3 )。除了与栅栏组织有关的参数外,
GST 与上角质层厚度、上表皮细胞厚度呈现出显著的负相关性, 即上角质层厚度与上表皮细胞厚度随生长季温度的降低而增大。叶表皮外角质层是由表皮细胞合成的脂类和碳水化合物的聚合物组成(
Kolattukudy, 1996 ), 角质层越厚, 蜡被越厚, 相对的抗寒能力就越好; 而较大的表皮细胞厚度有利于光的透射, 且具有较强的贮水作用, 当温度降低时, 植物叶片可能通过增加表皮细胞厚度来保证光合作用的正常进行, 并将水分存储其中来应对栅栏组织厚度的减小。自相关性分析(
表4 )表明, 上角质层厚度与下表皮细胞厚度间存在显著正相关关系(
p < 0.01), 即上角质层厚度发生变化时, 会引起下表皮细胞厚度发生正向变化。
MAP 、
GSP 均与上栅栏组织厚度相关性最强(
表5 ), 与栅栏组织总厚度、叶片厚度显著负相关(
p < 0.05), 即这3个解剖结构指标随降水量的增大而减小, 这可能是因为环境水分充足时, 植物不需要储存过多的水分。由此可见, 受水分含量变化影响最大的是上栅栏组织厚度, 其次是叶厚度与栅栏组织总厚度, 而下栅栏组织厚度与降水量没有表现出明显的相关性。同时, 自相关性分析(
表4 )表明, 上栅栏组织厚度、栅栏组织总厚度、叶厚度这3个指标显著正相关(
p < 0.01), 即任一指标变化, 会引起其他两个指标发生协同变化, 其中上栅栏组织厚度变 化是蒙古莸叶片应对生境水分含量变化最主要的 方式。
4 小结 蒙古莸在我国的分布区域广泛, 生长环境条件差异较大, 为满足自身正常的生长, 对环境和气候条件的适应就至关重要。植物形态结构是植物适应环境而进化出的生存策略, 由于环境分化选择作用, 植物逐步形成适应环境的表型和遗传特征, 表现出独特的种源特性; 即使将不同种源植物移植到相同环境下生长, 其叶片的解剖结构等表型特征仍存在着差异(
马小芬等, 2013 )。这种不同种源间特性的差异正是植物趋异适应的体现, 归根结底是由于控制表型的基因型不同所导致的。本研究采用同质园法, 削弱不同种源地环境对植物个体性状的影响, 将原种源气候差异驱动的遗传变异对蒙古莸叶片解剖结构的影响作为研究重点, 分析结果表明对蒙古莸解剖结构参数有显著的解释力。植物对原生境地气候条件长期适应, 已经产生了遗传结构上的差异, 不同种源蒙古莸叶片解剖结构的差异是其适应特有环境的体现; 同时, 不同解剖结构间存在着协同变化, 反映出植物对干旱环境的良好适应性, 这种适应性策略与近年来提出的叶经济谱概念吻合(
Wright et al ., 2004 ;
Pigliucci, 2006 )。研究结果可为蒙古莸优良抗逆种源选择、遗传资源保护等提供实际应用指导, 并为进一步探究全球变化背景下蒙古莸叶解剖结构对环境变化的响应机制等提供重要的理论依据。此外, 造成不同种源蒙古莸叶片解剖结构差异的因素还有很多, 包括种源地土壤、CO
2 浓度等其他环境因素, 还需在今后的研究中进一步地深入和细化。
[1] Bai XQ Liu C Ji RX Shen C Wang XP 2018 ). Effects of origin climate on light response characteristics of Caryopteris mongholica Acta Ecologica Sinica 8425 -8433 . [本文引用: 2] [ 白雪卡 , 刘超 , 纪若璇 , 沈超 , 王襄平 (2018 ). 种源地气候对蒙古莸光响应特性的影响生态学报 , 38, 8425 -8433 .] [本文引用: 2] [2] Barboni D Harrison SP Bartlein PJ Jalut G New M Prentice IC Sanchez-Go?i MF Spessa A Davis B Stevenson AC 2004 ). Relationships between plant traits and climate in the mediterranean region: a pollen data analysisJournal of Vegetation Science, 635 -646 . DOI:
10.1111/jvs.2004.15.issue-5 URL [本文引用: 1] [3] Chartzoulakis K Patakas A Kofidis G Bosabalidis A Nastou A 2002 ). Water stress affects leaf anatomy, gas exchange, water relations and growth of two avocado cultivarsScientia Horticulturae, 39 -50 . DOI:
10.1016/S0304-4238(02)00016-X URL [本文引用: 1] [4] Chen JJ Yu H Xu CB Zhong QL 2019 ). Effects of provenance and common garden environment on leaf functional traits of Machilus pauhoi seedlingsChinese Journal of Applied and Environmental Biology 648 -654 . [本文引用: 1] [ 陈嘉静 , 余华 , 徐朝斌 , 钟全林 (2019 ). 种源与同质园环境对刨花楠幼苗叶功能性状的影响应用与环境生物学报 , 25, 648 -654 .] [本文引用: 1] [5] Chen X Liu HK Wang Q Zou HY Zhu QL Wang YP 2019 ). Leaf anatomical characteristics of 11 tree species in the homogeneous habitats of eastern Shandong Province hillsChinese Journal of Applied and Environmental Biology, 655 -664 . [本文引用: 1] [ 陈旭 , 刘洪凯 , 王强 , 邹红阳 , 朱启良 , 王延平 (2019 ). 鲁东丘陵同质生境中11个树种叶解剖学特性比较应用与环境生物学报 , 25, 655 -664 .] [本文引用: 1] [6] de Lima Silva A da Silva Alves MV Coan AI 2014 ). Importance of anatomical leaf features for characterization of three species of Mapania(Mapanioideae, Cyperaceae) from the Amazon Forest, BrazilActa Amazonica, 447 -456 . DOI:
10.1590/1809-4392201305953 URL [本文引用: 1] [7] de Villemereuil P Gaggiotti OE Mouterde M Till-Bottraud I 2016 ). Common garden experiments in the genomic era: new perspectives and opportunitiesHeredity, 249 -254 . DOI:
10.1038/hdy.2015.93 URL [本文引用: 1] [8] Delzon S 2015 ). New insight into leaf drought toleranceFunctional Ecology, 1247 -1249 . DOI:
10.1111/fec.2015.29.issue-10 URL [本文引用: 1] [9] Feng YQ Qin L Ma HP 2008 ). The discussion on efficiency and quality of paraffin section in plant microscopy technique courseExperimental Technology and Management, 160 -162 . [本文引用: 1] [ 冯永庆 , 秦岭 , 马焕普 (2008 ). 植物显微技术课程中石蜡制片实验教学效率和质量的探讨实验技术与管理 , 25, 160 -162 .] [本文引用: 1] [10] Fu YR Ma BL Wang ZL Yang W 2005 ). Characteristics of 4 native tree species adapted to arid environment in northern Shaanxi sand-blown area and utilization prospectJournal of Desert Research, 386 -390 . [本文引用: 3] [ 符亚儒 , 麻保林 , 王子玲 , 杨伟 (2005 ). 陕北风沙区4种乡土树种适应干旱环境的特性及利用前景中国沙漠 , 25, 386 -390 .] [本文引用: 3] [11] Guo GG Feng B Ma BL Zhang YL Guo CH Jing ZB 2013 ). Leaf anatomical structures of different regional Amygdalus pedunculata Pall. and their drought resistance analysisActa Botanica Boreali-Occidentalia Sinica 720 -728 . [本文引用: 1] [ 郭改改 , 封斌 , 麻保林 , 张应龙 , 郭春会 , 井赵斌 (2013 ). 不同区域长柄扁桃叶片解剖结构及其抗旱性分析西北植物学报 , 33, 720 -728 .] [本文引用: 1] [12] Guo JY 2018 ). Difference on Leaf Morphology Anatomical Structure and Environmental Heterogeneous of Three Psammophytes Master degree dissertation, Inner Mongolia Agricultural University, Hohhot . [本文引用: 1] [ 郭婧宇 (2018 ). 三种沙生植物叶形态、结构差异性及环境分异硕士学位论文, 内蒙古农业大学, 呼和浩特 .] [本文引用: 1] [13] Han L He KN Lu XJ Wang ZL 2008 ). Changes and environmental effects of leaf water potential of Caryopteris mongolica in Qinghai alpine semi-arid areaBulletin of Soil and Water Conservation 1 -5 . [本文引用: 1] [ 韩磊 , 贺康宁 , 芦新建 , 王占林 (2008 ). 青海高寒半干旱区蒙古莸叶水势变化及其与环境因素的关系水土保持通报 , 28, 1 -5 .] [本文引用: 1] [14] He JS Wang XP Flynn DFB Wang L Schmid B Fang JY 2009 ). Taxonomic, phylogenetic, and environmental trade-offs between leaf productivity and persistenceEcology, 2779 -2791 . DOI:
10.1890/08-1126.1 URL [本文引用: 1] [15] He YM 2018 ). Studies on Ecological Adaptation and Reproductive Strategy of Caryopteris mongolica PhD dissertation, Inner Mongolia Agricultural University, Hohhot . [本文引用: 1] [ 贺一鸣 (2018 ). 蒙古莸种群生态适应及繁殖更新策略博士学位论文, 内蒙古农业大学, 呼和浩特 .] [本文引用: 1] [16] Hijmans RJ Cameron SE Parra JL Jones PG Jarvis A 2005 ). Very high resolution interpolated climate surfaces for global land areasInternational Journal of Climatology, 1965 -1978 . DOI:
10.1002/(ISSN)1097-0088 URL [本文引用: 1] [17] Hu MY Zhang L Luo TX Shen W 2012 ). Variations in leaf functional traits of Stipa purpurea along a rainfall gradient in Xizang, ChinaChinese Journal of Plant Ecology 136 -143 . DOI:
10.3724/SP.J.1258.2012.00136 URL [本文引用: 1] Aims Stipa purpurea is the dominant species in alpine arid and semi-arid grasslands on the Tibetan Plateau. Our objectives are to determine if this species exhibits a strategy shift in its specific leaf area (
SLA ) to nitrogen (N) concentration relationship along a rainfall gradient and to detect possible effects of environmental factors on related leaf traits.
Methods We investigated variations in leaf traits of
S. purpurea associated with climatic and soil factors along an east-west transect with a rainfall gradient (69–479 mm) but similar altitudes (4 300–4 700 m). Five locations from east to west are Damxung, Namco, Gêrzê, Mount Qomolangma and Rutog. We measured
SLA , mass- and area-based leaf N concentration (
N mass ,
N area ), leaf density and thickness and soil total N along the transect.
Important findings In pooled data,
SLA and
N mass varied little with the growing season mean temperature and precipitation and the soil total N concentration. The
SLA-N mass relationship in
S. purpurea did not shift between the semi-humid areas (ratio of rainfall to evaporation > 0.11) and the arid and semi-arid areas (ratio < 0.11), although there was a positive correlation between
SLA and
N mass across the five locations. Variation in
SLA was mainly determined by leaf density in the semi-humid areas and by leaf thickness in the arid and semi-arid areas; both were negatively correlated with
SLA . With increasing temperature or declining precipitation, leaf density decreased and leaf thickness increased, leading to non-significant relationships between
SLA and climatic factors. The increase of leaf density in the semi-humid areas was correlated with the increase of
N area , but the increase of leaf thickness in the arid and semi-arid areas did not lead to change of
N area , resulting in unchanged
N area along the rainfall gradient. A positive correlation was detected between aboveground biomass and Narea in
S. purpurea , indicating that increased
N area may increase plant productivity. Our findings suggest that alpine plants in arid and semi-arid areas may maintain a constant
N area by increased leaf thickness in order to achieve a similar photosynthetic productivity and water use efficiency compared to the relatively humid areas. The relative impacts of leaf density and leaf thickness on
SLA shifted between the semi-humid areas and the arid and semi-arid areas, which may provide insight in detecting the threshold of water limitation in alpine grasslands.
[ 胡梦瑶 , 张林 , 罗天祥 , 沈维 (2012 ). 西藏紫花针茅叶功能性状沿降水梯度的变化植物生态学报 , 36, 136 -143 .] DOI:
10.3724/SP.J.1258.2012.00136 URL [本文引用: 1] Aims Stipa purpurea is the dominant species in alpine arid and semi-arid grasslands on the Tibetan Plateau. Our objectives are to determine if this species exhibits a strategy shift in its specific leaf area (
SLA ) to nitrogen (N) concentration relationship along a rainfall gradient and to detect possible effects of environmental factors on related leaf traits.
Methods We investigated variations in leaf traits of
S. purpurea associated with climatic and soil factors along an east-west transect with a rainfall gradient (69–479 mm) but similar altitudes (4 300–4 700 m). Five locations from east to west are Damxung, Namco, Gêrzê, Mount Qomolangma and Rutog. We measured
SLA , mass- and area-based leaf N concentration (
N mass ,
N area ), leaf density and thickness and soil total N along the transect.
Important findings In pooled data,
SLA and
N mass varied little with the growing season mean temperature and precipitation and the soil total N concentration. The
SLA-N mass relationship in
S. purpurea did not shift between the semi-humid areas (ratio of rainfall to evaporation > 0.11) and the arid and semi-arid areas (ratio < 0.11), although there was a positive correlation between
SLA and
N mass across the five locations. Variation in
SLA was mainly determined by leaf density in the semi-humid areas and by leaf thickness in the arid and semi-arid areas; both were negatively correlated with
SLA . With increasing temperature or declining precipitation, leaf density decreased and leaf thickness increased, leading to non-significant relationships between
SLA and climatic factors. The increase of leaf density in the semi-humid areas was correlated with the increase of
N area , but the increase of leaf thickness in the arid and semi-arid areas did not lead to change of
N area , resulting in unchanged
N area along the rainfall gradient. A positive correlation was detected between aboveground biomass and Narea in
S. purpurea , indicating that increased
N area may increase plant productivity. Our findings suggest that alpine plants in arid and semi-arid areas may maintain a constant
N area by increased leaf thickness in order to achieve a similar photosynthetic productivity and water use efficiency compared to the relatively humid areas. The relative impacts of leaf density and leaf thickness on
SLA shifted between the semi-humid areas and the arid and semi-arid areas, which may provide insight in detecting the threshold of water limitation in alpine grasslands.
[18] Ji ZJ Quan XK Wang CK 2013 ). Variations in leaf anatomy of Larix gmelinii reflect adaptation of its photosynthetic capacity to climate changesActa Ecologica Sinica 6967 -6974 . DOI:
10.5846/stxb URL [本文引用: 2] [ 季子敬 , 全先奎 , 王传宽 (2013 ). 兴安落叶松针叶解剖结构变化及其光合能力对气候变化的适应性生态学报 , 33, 6967 -6974 .] DOI:
10.5846/stxb URL [本文引用: 2] [19] Kolattukudy PE 1996 ). Biosynthetic pathways of cutin and waxes, and their sensitivity to environmental stressesPlant Cuticles, 83 -108 . [本文引用: 1] [20] Li AP Wang XJ Yang XY Zhang L 2010 ). Evaluation of drought resistance capacity of desert shrubs in Hobq desert based on characteristics of leaf anatomical structureJournal of Desert Research, 1405 -1410 . [本文引用: 3] [ 李爱平 , 王晓江 , 杨小玉 , 张雷 (2010 ). 库布齐沙漠几种沙生灌木叶解剖结构耐旱特征研究中国沙漠 , 30, 1405 -1410 .] [本文引用: 3] [21] Li FL Bao WK 2005 ). Responses of the morphological and anatomical structure of the plant leaf to environmental changeChinese Bulletin of Botany, 118 -127 . [本文引用: 1] [ 李芳兰 , 包维楷 (2005 ). 植物叶片形态解剖结构对环境变化的响应与适应植物学通报 , 40, 118 -127 .] [本文引用: 1] [22] Li YJ Li XR Yang XL 1991 ). A study on the character of ecology and physiology of aromatic plant— Caryopteris mongolica BgeJournal of Desert Research 53 -59 . [本文引用: 1] [ 李玉俊 , 李新荣 , 杨喜林 (1991 ). 芳香植物——蒙古莸的生态生理特性及其栽培技术中国沙漠 , 11, 53 -59 .] [本文引用: 1] [23] Liu MY Liu GL Kang YX Zhang S Wu Y Wang Y 2018 ). Responses of leaf morphological and anatomical structure to elevation in an alpine plant Meconopsis integrifolia Chinese Journal of Ecology 35 -42 . [本文引用: 1] [ 刘梦颖 , 刘光立 , 康永祥 , 张硕 , 吴云 , 王玉 (2018 ). 高山植物全缘叶绿绒蒿叶片形态及解剖结构对海拔的响应生态学杂志 , 37, 35 -42 .] [本文引用: 1] [24] Ma XF Wang XF Li Q He X 2013 ). The analysis of drought resistance and the comparison of anatomical structures of the leave of Xanthoceras sorbifolia Bunge introduced from different regionsJournal of Arid Land Resources and Environment 92 -96 . [本文引用: 1] [ 马小芬 , 王兴芳 , 李强 , 贺晓 (2013 ). 不同种源地文冠果叶片解剖结构比较及抗旱性分析干旱区资源与环境 , 27, 92 -96 .] [本文引用: 1] [25] Pigliucci M 2006 ). Phenotypic plasticity and evolution by genetic assimilationJournal of Experimental Biology, 2362 -2367 . DOI:
10.1242/jeb.02070 URL [本文引用: 2] [26] Qin FF Li Q Cui ZM Li HP Yang ZR 2012 ). Leaf anatomical structures and ecological adaptabilities to light of three alfalfa cultivars with different fall dormancies under shading during overwinteringChinese Journal of Plant Ecology, 333 -345 . DOI:
10.3724/SP.J.1258.2012.00333 URL [本文引用: 1] [ 覃凤飞 , 李强 , 崔棹茗 , 李洪萍 , 杨智然 (2012 ). 越冬期遮阴条件下3个不同秋眠型紫花苜蓿品种叶片解剖结构与其光生态适应性植物生态学报 , 36, 333 -345 .] DOI:
10.3724/SP.J.1258.2012.00333 URL [本文引用: 1] [27] Smith WK Vogelmann TC DeLucia EH Bell DT Shepherd KA 1997 ). Leaf form and photosynthesisBioScience, 785 -793 . DOI:
10.2307/1313100 URL [本文引用: 1] [28] Tian LL Li J Lin HY Huang JA Liu ZH 2019 ). Response of leaf anatomical structure of dayezhong tea germplasm to latitudeMolecular Plant Breeding, 7262 -7268 . [本文引用: 1] [ 田丽丽 , 李娟 , 林海燕 , 黄建安 , 刘仲华 (2019 ). 大叶种茶树叶片解剖结构对纬度的响应分子植物育种 , 17, 7262 -7268 .] [本文引用: 1] [29] Tian Y Ni XL Yu HN Shen XD Peng L 2010 ). Studies on drought resistance on morphology anatomical structure of leave of six species shrubsChinese Agricultural Science Bulletin, 113 -117 . [本文引用: 1] [ 田英 , 倪细炉 , 于海宁 , 沈效东 , 彭励 (2010 ). 6种抗旱灌木叶片形态解剖学特征中国农学通报 , 26, 113 -117 .] [本文引用: 1] [30] Wang XJ Li AP Ning MS Zhang JG 2006 ). Study on the biological and ecological characteristics and economic value of the ecological shrub ofCaryopteris mongolica. Journal of Arid Land Resources and Environment, 191 -194 . [本文引用: 1] [ 王晓江 , 李爱平 , 宁明世 , 张纪刚 (2006 ). 生态灌木蒙古莸的生物生态学特性及其经济价值评价干旱区资源与环境 , 20, 191 -194 .] [本文引用: 1] [31] Wang XQ Wang CK Han Y 2015 ). Effects of tree species on soil organic carbon density: a common garden experiment of five temperate tree speciesChinese Journal of Plant Ecology, 1033 -1043 . DOI:
10.17521/cjpe.2015.0100 URL [本文引用: 1] [ 王薪琪 , 王传宽 , 韩轶 (2015 ). 树种对土壤有机碳密度的影响: 5种温带树种同质园试验植物生态学报 , 39, 1033 -1043 .] DOI:
10.17521/cjpe.2015.0100 URL [本文引用: 1] [32] Wright IJ Reich PB Westoby M 2001 ). Strategy shifts in leaf physiology, structure and nutrient content between species of high- and low-rainfall and high- and low-nutrient habitatsFunctional Ecology, 423 -434 . DOI:
10.1046/j.0269-8463.2001.00542.x URL [本文引用: 1] [33] Wright IJ Reich PB Westoby M 2004 ). The worldwide leaf economics spectrumNature, 821 -827 . DOI:
10.1038/nature02403 URL [本文引用: 1] [34] Yang XY 2008 ). Study on Drought Resistance Characteristics of Leaf Anatomical Structure of Five Psammophilic Shrub Species Master degree dissertation, Inner Mongolia Agricultural University, Hohhot . [本文引用: 2] [ 杨小玉 (2008 ). 5种沙生灌木叶片解剖结构与抗旱性研究硕士学位论文, 内蒙古农业大学, 呼和浩特 .] [本文引用: 2] [35] Yu HY Hu XY He CX Cui YF Fan SQ Bi QX Wang LB 2019 ). Differential response of water stress on leaf morphological anatomical structures of varied provenancesXanthocera sorbifolium. Journal of Beijing Forestry University, 1 ), 57 -63 . [本文引用: 1] [ 于海燕 , 胡潇予 , 何春霞 , 崔艺凡 , 范思琪 , 毕泉鑫 , 王利兵 (2019 ). 文冠果不同种源叶片结构对水分胁迫的差异性响应北京林业大学学报 , 41(1 ), 57 -63 .] [本文引用: 1] [36] Zhang YR Dong RC Mai S Mei SG 1999 ). Protection and utilization of Caryopteris mongolica Bunge., one of drought enduring shrub resourcesJournal of Arid Land Resources and Environment 91 -94 . [本文引用: 1] [ 张源润 , 董仁才 , 麦硕 , 梅曙光 (1999 ). 耐旱灌木蒙古莸花的资源及保护利用干旱区资源与环境 , 13, 91 -94 . ] [本文引用: 1] [37] Zhao XX Li Y Su SP Bai X 2015 ). Anatomical structure of assimilating shoots of Calligonum mongolicum from six populations and its relationship with geo-ecological factorsJournal of Arid Land Resources and Environment 55 -60 . [本文引用: 1] [ 赵小仙 , 李毅 , 苏世平 , 白潇 (2015 ). 6个种群蒙古沙拐枣同化枝解剖结构及与地理生态因子的关系干旱区资源与环境 , 29, 55 -60 .] [本文引用: 1] [38] Zhong YM Dong FY Wang WJ Wang JM Li JW Wu B Jia XH 2017 ). Anatomical characteristics and adaptability plasticity of Populus euphratica in different habitatsJournal of Beijing Forestry University 10 ), 53 -61 . [本文引用: 1] [ 钟悦鸣 , 董芳宇 , 王文娟 , 王健铭 , 李景文 , 吴波 , 贾晓红 (2017 ). 不同生境胡杨叶片解剖特征及其适应可塑性北京林业大学学报 , 39(10 ), 53 -61 .] [本文引用: 1] 种源地气候对蒙古莸光响应特性的影响
2
2018
... 蒙古莸(
Caryopteris mongholica )属唇形科筋骨草亚科莸属(
Caryopteris ), 是我国干旱、半干旱地区一种耐旱、耐寒兼具观赏性的绿化灌木, 是防风固沙和水土保持的优良造林树种(
李玉俊等, 1991 ;
韩磊等, 2008 ).蒙古莸分布在东起呼伦贝尔典型草原区, 西至甘肃河西走廊西部和青海湖南部的广阔的蒙古高原上.然而, 不同的生长环境导致蒙古莸的生理特征和遗传结构均出现不同程度的变异, 这种对环境的适应性变异表现, 在叶片的解剖结构上尤为明显.
白雪卡等(2018) 对同质园栽培下8个不同种源蒙古莸光响应曲线的研究发现, 蒙古莸光合响应能力在不同种源间表现出明显不同, 光响应曲线也存在显著的差异.蒙古莸这种对环境的适应性变异在叶片的解剖结构上尤为明显, 在前人不同的研究中, 蒙古莸或为等面叶、无海绵组织分化(
符亚儒等, 2005 ;
杨小玉, 2008 ;
李爱平等, 2010 ), 或为异面叶、叶肉中含有海绵组织(
张源润等, 1999 ;
王晓江等, 2006 ;
田英等, 2010 ); 叶片厚度21.6-2 180 μm, 变异极大. ...
... 原种源地的地理位置(经纬度)、温度(
MAT 、
ART )、
MAP 、
PE 、
Alt 与叶片解剖结构指标密切相关(
表6 ), 能够很好地解释其遗传变异程度, 其中种源地气候(
MAT 、
MAP 、
PE )对蒙古莸叶片解剖结构解释程度在34.09%-81.43%之间.
季子敬等(2013) 对兴安落叶松(
Larix gmelinii )针叶解剖结构的分析研究中发现, 针叶长期适应当地环境条件后, 在解剖结构特征上也产生了具有遗传能力的变异.
赵小仙等(2015) 对蒙古沙拐枣(
Calligonum monglicum )叶片解剖结构分析研究中发现, 解剖结构的差异与经纬度、
MAT 、
MAP 的变化有极显著相关性.
白雪卡等(2018) 对不同种源蒙古莸光响应特性的研究中也同样证明, 种源地气候驱动的遗传变异对蒙古莸光合作用参数有显著的解释力, 解释程度在16.65%- 73.52%之间, 与本研究的解释力近似.本研究中, 上角质层厚度、上下表皮细胞厚度、下栅栏组织厚度、栅栏组织总厚度、叶片结构紧密度主要受
MAT 的影响, 解释程度在20.77%-71.89%之间(
表6 ); 上栅栏组织厚度、叶厚度主要受
MAP 影响, 解释程度在31.40%-45.93%之间(
表6 ).相对而言, 温度对蒙古莸叶片解剖结构参数的解释度大于水分, 这与
田丽丽等(2019) 对大叶种茶树叶片解剖结构的分析不谋而合, 其研究发现叶片生长受到温度、降水和光照因子等综合因素的影响, 叶片结构的差异则主要受到温度影响. ...
种源地气候对蒙古莸光响应特性的影响
2
2018
... 蒙古莸(
Caryopteris mongholica )属唇形科筋骨草亚科莸属(
Caryopteris ), 是我国干旱、半干旱地区一种耐旱、耐寒兼具观赏性的绿化灌木, 是防风固沙和水土保持的优良造林树种(
李玉俊等, 1991 ;
韩磊等, 2008 ).蒙古莸分布在东起呼伦贝尔典型草原区, 西至甘肃河西走廊西部和青海湖南部的广阔的蒙古高原上.然而, 不同的生长环境导致蒙古莸的生理特征和遗传结构均出现不同程度的变异, 这种对环境的适应性变异表现, 在叶片的解剖结构上尤为明显.
白雪卡等(2018) 对同质园栽培下8个不同种源蒙古莸光响应曲线的研究发现, 蒙古莸光合响应能力在不同种源间表现出明显不同, 光响应曲线也存在显著的差异.蒙古莸这种对环境的适应性变异在叶片的解剖结构上尤为明显, 在前人不同的研究中, 蒙古莸或为等面叶、无海绵组织分化(
符亚儒等, 2005 ;
杨小玉, 2008 ;
李爱平等, 2010 ), 或为异面叶、叶肉中含有海绵组织(
张源润等, 1999 ;
王晓江等, 2006 ;
田英等, 2010 ); 叶片厚度21.6-2 180 μm, 变异极大. ...
... 原种源地的地理位置(经纬度)、温度(
MAT 、
ART )、
MAP 、
PE 、
Alt 与叶片解剖结构指标密切相关(
表6 ), 能够很好地解释其遗传变异程度, 其中种源地气候(
MAT 、
MAP 、
PE )对蒙古莸叶片解剖结构解释程度在34.09%-81.43%之间.
季子敬等(2013) 对兴安落叶松(
Larix gmelinii )针叶解剖结构的分析研究中发现, 针叶长期适应当地环境条件后, 在解剖结构特征上也产生了具有遗传能力的变异.
赵小仙等(2015) 对蒙古沙拐枣(
Calligonum monglicum )叶片解剖结构分析研究中发现, 解剖结构的差异与经纬度、
MAT 、
MAP 的变化有极显著相关性.
白雪卡等(2018) 对不同种源蒙古莸光响应特性的研究中也同样证明, 种源地气候驱动的遗传变异对蒙古莸光合作用参数有显著的解释力, 解释程度在16.65%- 73.52%之间, 与本研究的解释力近似.本研究中, 上角质层厚度、上下表皮细胞厚度、下栅栏组织厚度、栅栏组织总厚度、叶片结构紧密度主要受
MAT 的影响, 解释程度在20.77%-71.89%之间(
表6 ); 上栅栏组织厚度、叶厚度主要受
MAP 影响, 解释程度在31.40%-45.93%之间(
表6 ).相对而言, 温度对蒙古莸叶片解剖结构参数的解释度大于水分, 这与
田丽丽等(2019) 对大叶种茶树叶片解剖结构的分析不谋而合, 其研究发现叶片生长受到温度、降水和光照因子等综合因素的影响, 叶片结构的差异则主要受到温度影响. ...
Relationships between plant traits and climate in the mediterranean region: a pollen data analysis
1
2004
... 植物在进化过程中, 受到外界温度、水分、光照等环境因素的长期作用而影响其建成, 逐步形成适应环境的表型和遗传特征.即使是同一物种, 长期生长在不同的地理环境中, 也会产生不同程度的基因分化, 以适应各自不同的生态位, 从而形成特定的种源特性(
Smith et al ., 1997 ;
Barboni et al ., 2004 ;
Pigliucci, 2006 ;
胡梦瑶等, 2012 ).植物自然群体分布的环境具有明显差异.从表型来看, 同一结构由于环境分化选择作用, 不同环境条件下的群体最适表型也存在差异, 而表型的差异归根结底是由控制表型的基因型不同所导致的. ...
Water stress affects leaf anatomy, gas exchange, water relations and growth of two avocado cultivars
1
2002
... 叶片作为植物暴露在生活环境中面积最大的器官, 其外部形态特征、内部解剖结构等最能体现植物对环境因子的适应性进化(
Wright et al ., 2001 ;
Chartzoulakis et al ., 2002 ;
钟悦鸣等, 2017 ;
于海燕等, 2019 ), 开展叶片解剖结构分析, 对研究植物在极端生态环境下的适应策略具有重要意义.一般而言, 植物分布范围广泛, 各自生活在温度、水分等条件不相同的自然地理环境中, 在长期对环境适应过程中, 同种植物叶片的解剖结构表现出明显的差异(
季子敬等, 2013 ;
郭婧宇, 2018 ).叶片的这种功能性状差异可能既有自身的可塑性响应也有遗传变异, 以往研究中沿自然环境梯度的采样测定或单一站点控制实验很难区分两者的效应.全缘叶绿绒蒿(
Meconopsis integrifolia )随着海拔升高, 株型变矮, 叶片组织结构紧密度增大(
刘梦颖等, 2018 ); 生长于干旱地区的长梗扁桃(
Amygdalus pedunculata )叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的长梗扁桃, 其叶片栅栏组织排列疏松, 叶厚度较薄(
郭改改等, 2013 ).这种叶片解剖结构特征的差异同时受到环境变异与自身遗传结构两个因素的影响.同质园试验即研究同一基因型的物种在不同环境下的初生与次生代谢的响应, 或不同基因型的物种在相同环境下的初生与次生代谢的响应.将多个种源的植物在同地栽培, 在一定程度上控制环境与基因型间的相互作用, 最大程度地降低立地条件所造成的影响(
王薪琪等, 2015 ;
陈嘉静等, 2019 ), 进而说明不同种源植物叶片解剖结构特征表现出的差异是由于原种源地环境差异造成的植物遗传性基因变异.这与先前关于环境和遗传因素共同影响(包括气候梯度和种源的影响)叶片解剖结构特征的研究有所不同, 这种同质园方法在局部适应性研究中具有重要意义(
de Villemereuil, 2016 ), 有助于我们更好地理解不同地理种源植物对全球变化的响应和适应机制.目前采用此法对不同种源内植物叶片解剖结构差异及其影响因素的相关分析较少. ...
种源与同质园环境对刨花楠幼苗叶功能性状的影响
1
2019
... 叶片作为植物暴露在生活环境中面积最大的器官, 其外部形态特征、内部解剖结构等最能体现植物对环境因子的适应性进化(
Wright et al ., 2001 ;
Chartzoulakis et al ., 2002 ;
钟悦鸣等, 2017 ;
于海燕等, 2019 ), 开展叶片解剖结构分析, 对研究植物在极端生态环境下的适应策略具有重要意义.一般而言, 植物分布范围广泛, 各自生活在温度、水分等条件不相同的自然地理环境中, 在长期对环境适应过程中, 同种植物叶片的解剖结构表现出明显的差异(
季子敬等, 2013 ;
郭婧宇, 2018 ).叶片的这种功能性状差异可能既有自身的可塑性响应也有遗传变异, 以往研究中沿自然环境梯度的采样测定或单一站点控制实验很难区分两者的效应.全缘叶绿绒蒿(
Meconopsis integrifolia )随着海拔升高, 株型变矮, 叶片组织结构紧密度增大(
刘梦颖等, 2018 ); 生长于干旱地区的长梗扁桃(
Amygdalus pedunculata )叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的长梗扁桃, 其叶片栅栏组织排列疏松, 叶厚度较薄(
郭改改等, 2013 ).这种叶片解剖结构特征的差异同时受到环境变异与自身遗传结构两个因素的影响.同质园试验即研究同一基因型的物种在不同环境下的初生与次生代谢的响应, 或不同基因型的物种在相同环境下的初生与次生代谢的响应.将多个种源的植物在同地栽培, 在一定程度上控制环境与基因型间的相互作用, 最大程度地降低立地条件所造成的影响(
王薪琪等, 2015 ;
陈嘉静等, 2019 ), 进而说明不同种源植物叶片解剖结构特征表现出的差异是由于原种源地环境差异造成的植物遗传性基因变异.这与先前关于环境和遗传因素共同影响(包括气候梯度和种源的影响)叶片解剖结构特征的研究有所不同, 这种同质园方法在局部适应性研究中具有重要意义(
de Villemereuil, 2016 ), 有助于我们更好地理解不同地理种源植物对全球变化的响应和适应机制.目前采用此法对不同种源内植物叶片解剖结构差异及其影响因素的相关分析较少. ...
种源与同质园环境对刨花楠幼苗叶功能性状的影响
1
2019
... 叶片作为植物暴露在生活环境中面积最大的器官, 其外部形态特征、内部解剖结构等最能体现植物对环境因子的适应性进化(
Wright et al ., 2001 ;
Chartzoulakis et al ., 2002 ;
钟悦鸣等, 2017 ;
于海燕等, 2019 ), 开展叶片解剖结构分析, 对研究植物在极端生态环境下的适应策略具有重要意义.一般而言, 植物分布范围广泛, 各自生活在温度、水分等条件不相同的自然地理环境中, 在长期对环境适应过程中, 同种植物叶片的解剖结构表现出明显的差异(
季子敬等, 2013 ;
郭婧宇, 2018 ).叶片的这种功能性状差异可能既有自身的可塑性响应也有遗传变异, 以往研究中沿自然环境梯度的采样测定或单一站点控制实验很难区分两者的效应.全缘叶绿绒蒿(
Meconopsis integrifolia )随着海拔升高, 株型变矮, 叶片组织结构紧密度增大(
刘梦颖等, 2018 ); 生长于干旱地区的长梗扁桃(
Amygdalus pedunculata )叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的长梗扁桃, 其叶片栅栏组织排列疏松, 叶厚度较薄(
郭改改等, 2013 ).这种叶片解剖结构特征的差异同时受到环境变异与自身遗传结构两个因素的影响.同质园试验即研究同一基因型的物种在不同环境下的初生与次生代谢的响应, 或不同基因型的物种在相同环境下的初生与次生代谢的响应.将多个种源的植物在同地栽培, 在一定程度上控制环境与基因型间的相互作用, 最大程度地降低立地条件所造成的影响(
王薪琪等, 2015 ;
陈嘉静等, 2019 ), 进而说明不同种源植物叶片解剖结构特征表现出的差异是由于原种源地环境差异造成的植物遗传性基因变异.这与先前关于环境和遗传因素共同影响(包括气候梯度和种源的影响)叶片解剖结构特征的研究有所不同, 这种同质园方法在局部适应性研究中具有重要意义(
de Villemereuil, 2016 ), 有助于我们更好地理解不同地理种源植物对全球变化的响应和适应机制.目前采用此法对不同种源内植物叶片解剖结构差异及其影响因素的相关分析较少. ...
鲁东丘陵同质生境中11个树种叶解剖学特性比较
1
2019
... 本研究通过对同质园栽培下7个种源蒙古莸叶片解剖结构的观察, 发现7个种源地的蒙古莸叶片均为典型的等面叶, 叶片由紧密排列的上表皮、发达的栅栏组织和下表皮构成, 无海绵组织分化, 是旱生植物最常见的叶片结构(
Delzon, 2015 ), 与
贺一鸣(2018) 、
杨小玉(2008) 、
李爱平等(2010) 、
符亚儒等(2005) 对蒙古莸叶片解剖结构观察结果一致.蒙古莸叶片解剖结构参数的数量化分析表明, 上表皮细胞厚度均大于下表皮细胞厚度, 这种结构有利于气体交换(
陈旭等, 2019 ); 7个种源蒙古莸叶片的7个解剖结构参数存在显著差异(
表3 ), 其中, 叶厚度变化范围为192.34-270.30 μm, 变异系数为0.12, 与
符亚儒等(2005) 、
李爱平等(2010) 的测量结果近似. ...
鲁东丘陵同质生境中11个树种叶解剖学特性比较
1
2019
... 本研究通过对同质园栽培下7个种源蒙古莸叶片解剖结构的观察, 发现7个种源地的蒙古莸叶片均为典型的等面叶, 叶片由紧密排列的上表皮、发达的栅栏组织和下表皮构成, 无海绵组织分化, 是旱生植物最常见的叶片结构(
Delzon, 2015 ), 与
贺一鸣(2018) 、
杨小玉(2008) 、
李爱平等(2010) 、
符亚儒等(2005) 对蒙古莸叶片解剖结构观察结果一致.蒙古莸叶片解剖结构参数的数量化分析表明, 上表皮细胞厚度均大于下表皮细胞厚度, 这种结构有利于气体交换(
陈旭等, 2019 ); 7个种源蒙古莸叶片的7个解剖结构参数存在显著差异(
表3 ), 其中, 叶厚度变化范围为192.34-270.30 μm, 变异系数为0.12, 与
符亚儒等(2005) 、
李爱平等(2010) 的测量结果近似. ...
Importance of anatomical leaf features for characterization of three species of Mapania(Mapanioideae, Cyperaceae) from the Amazon Forest, Brazil
1
2014
... 采用常规石蜡切片法(
冯永庆等, 2008 )制作石蜡切片.将固定24 h以上的叶块, 经70%、85%、95%、100%四个浓度酒精脱水, 二甲苯透明, 石蜡浸透包埋(包埋所用石蜡熔点为56-57 ℃), 待凝固后切片, 切片厚度8-10 μm, 番红-固绿双重染色, 中性树胶封片, 将其制作为永久切片(
de Lima Silva et al ., 2014 ).在Leica DM2500显微镜(Leica, Wezlar, Germany)下观察, 并用Leica LAS AF软件拍照记录. ...
Common garden experiments in the genomic era: new perspectives and opportunities
1
2016
... 叶片作为植物暴露在生活环境中面积最大的器官, 其外部形态特征、内部解剖结构等最能体现植物对环境因子的适应性进化(
Wright et al ., 2001 ;
Chartzoulakis et al ., 2002 ;
钟悦鸣等, 2017 ;
于海燕等, 2019 ), 开展叶片解剖结构分析, 对研究植物在极端生态环境下的适应策略具有重要意义.一般而言, 植物分布范围广泛, 各自生活在温度、水分等条件不相同的自然地理环境中, 在长期对环境适应过程中, 同种植物叶片的解剖结构表现出明显的差异(
季子敬等, 2013 ;
郭婧宇, 2018 ).叶片的这种功能性状差异可能既有自身的可塑性响应也有遗传变异, 以往研究中沿自然环境梯度的采样测定或单一站点控制实验很难区分两者的效应.全缘叶绿绒蒿(
Meconopsis integrifolia )随着海拔升高, 株型变矮, 叶片组织结构紧密度增大(
刘梦颖等, 2018 ); 生长于干旱地区的长梗扁桃(
Amygdalus pedunculata )叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的长梗扁桃, 其叶片栅栏组织排列疏松, 叶厚度较薄(
郭改改等, 2013 ).这种叶片解剖结构特征的差异同时受到环境变异与自身遗传结构两个因素的影响.同质园试验即研究同一基因型的物种在不同环境下的初生与次生代谢的响应, 或不同基因型的物种在相同环境下的初生与次生代谢的响应.将多个种源的植物在同地栽培, 在一定程度上控制环境与基因型间的相互作用, 最大程度地降低立地条件所造成的影响(
王薪琪等, 2015 ;
陈嘉静等, 2019 ), 进而说明不同种源植物叶片解剖结构特征表现出的差异是由于原种源地环境差异造成的植物遗传性基因变异.这与先前关于环境和遗传因素共同影响(包括气候梯度和种源的影响)叶片解剖结构特征的研究有所不同, 这种同质园方法在局部适应性研究中具有重要意义(
de Villemereuil, 2016 ), 有助于我们更好地理解不同地理种源植物对全球变化的响应和适应机制.目前采用此法对不同种源内植物叶片解剖结构差异及其影响因素的相关分析较少. ...
New insight into leaf drought tolerance
1
2015
... 本研究通过对同质园栽培下7个种源蒙古莸叶片解剖结构的观察, 发现7个种源地的蒙古莸叶片均为典型的等面叶, 叶片由紧密排列的上表皮、发达的栅栏组织和下表皮构成, 无海绵组织分化, 是旱生植物最常见的叶片结构(
Delzon, 2015 ), 与
贺一鸣(2018) 、
杨小玉(2008) 、
李爱平等(2010) 、
符亚儒等(2005) 对蒙古莸叶片解剖结构观察结果一致.蒙古莸叶片解剖结构参数的数量化分析表明, 上表皮细胞厚度均大于下表皮细胞厚度, 这种结构有利于气体交换(
陈旭等, 2019 ); 7个种源蒙古莸叶片的7个解剖结构参数存在显著差异(
表3 ), 其中, 叶厚度变化范围为192.34-270.30 μm, 变异系数为0.12, 与
符亚儒等(2005) 、
李爱平等(2010) 的测量结果近似. ...
植物显微技术课程中石蜡制片实验教学效率和质量的探讨
1
2008
... 采用常规石蜡切片法(
冯永庆等, 2008 )制作石蜡切片.将固定24 h以上的叶块, 经70%、85%、95%、100%四个浓度酒精脱水, 二甲苯透明, 石蜡浸透包埋(包埋所用石蜡熔点为56-57 ℃), 待凝固后切片, 切片厚度8-10 μm, 番红-固绿双重染色, 中性树胶封片, 将其制作为永久切片(
de Lima Silva et al ., 2014 ).在Leica DM2500显微镜(Leica, Wezlar, Germany)下观察, 并用Leica LAS AF软件拍照记录. ...
植物显微技术课程中石蜡制片实验教学效率和质量的探讨
1
2008
... 采用常规石蜡切片法(
冯永庆等, 2008 )制作石蜡切片.将固定24 h以上的叶块, 经70%、85%、95%、100%四个浓度酒精脱水, 二甲苯透明, 石蜡浸透包埋(包埋所用石蜡熔点为56-57 ℃), 待凝固后切片, 切片厚度8-10 μm, 番红-固绿双重染色, 中性树胶封片, 将其制作为永久切片(
de Lima Silva et al ., 2014 ).在Leica DM2500显微镜(Leica, Wezlar, Germany)下观察, 并用Leica LAS AF软件拍照记录. ...
陕北风沙区4种乡土树种适应干旱环境的特性及利用前景
3
2005
... 蒙古莸(
Caryopteris mongholica )属唇形科筋骨草亚科莸属(
Caryopteris ), 是我国干旱、半干旱地区一种耐旱、耐寒兼具观赏性的绿化灌木, 是防风固沙和水土保持的优良造林树种(
李玉俊等, 1991 ;
韩磊等, 2008 ).蒙古莸分布在东起呼伦贝尔典型草原区, 西至甘肃河西走廊西部和青海湖南部的广阔的蒙古高原上.然而, 不同的生长环境导致蒙古莸的生理特征和遗传结构均出现不同程度的变异, 这种对环境的适应性变异表现, 在叶片的解剖结构上尤为明显.
白雪卡等(2018) 对同质园栽培下8个不同种源蒙古莸光响应曲线的研究发现, 蒙古莸光合响应能力在不同种源间表现出明显不同, 光响应曲线也存在显著的差异.蒙古莸这种对环境的适应性变异在叶片的解剖结构上尤为明显, 在前人不同的研究中, 蒙古莸或为等面叶、无海绵组织分化(
符亚儒等, 2005 ;
杨小玉, 2008 ;
李爱平等, 2010 ), 或为异面叶、叶肉中含有海绵组织(
张源润等, 1999 ;
王晓江等, 2006 ;
田英等, 2010 ); 叶片厚度21.6-2 180 μm, 变异极大. ...
... 本研究通过对同质园栽培下7个种源蒙古莸叶片解剖结构的观察, 发现7个种源地的蒙古莸叶片均为典型的等面叶, 叶片由紧密排列的上表皮、发达的栅栏组织和下表皮构成, 无海绵组织分化, 是旱生植物最常见的叶片结构(
Delzon, 2015 ), 与
贺一鸣(2018) 、
杨小玉(2008) 、
李爱平等(2010) 、
符亚儒等(2005) 对蒙古莸叶片解剖结构观察结果一致.蒙古莸叶片解剖结构参数的数量化分析表明, 上表皮细胞厚度均大于下表皮细胞厚度, 这种结构有利于气体交换(
陈旭等, 2019 ); 7个种源蒙古莸叶片的7个解剖结构参数存在显著差异(
表3 ), 其中, 叶厚度变化范围为192.34-270.30 μm, 变异系数为0.12, 与
符亚儒等(2005) 、
李爱平等(2010) 的测量结果近似. ...
... ), 其中, 叶厚度变化范围为192.34-270.30 μm, 变异系数为0.12, 与
符亚儒等(2005) 、
李爱平等(2010) 的测量结果近似. ...
陕北风沙区4种乡土树种适应干旱环境的特性及利用前景
3
2005
... 蒙古莸(
Caryopteris mongholica )属唇形科筋骨草亚科莸属(
Caryopteris ), 是我国干旱、半干旱地区一种耐旱、耐寒兼具观赏性的绿化灌木, 是防风固沙和水土保持的优良造林树种(
李玉俊等, 1991 ;
韩磊等, 2008 ).蒙古莸分布在东起呼伦贝尔典型草原区, 西至甘肃河西走廊西部和青海湖南部的广阔的蒙古高原上.然而, 不同的生长环境导致蒙古莸的生理特征和遗传结构均出现不同程度的变异, 这种对环境的适应性变异表现, 在叶片的解剖结构上尤为明显.
白雪卡等(2018) 对同质园栽培下8个不同种源蒙古莸光响应曲线的研究发现, 蒙古莸光合响应能力在不同种源间表现出明显不同, 光响应曲线也存在显著的差异.蒙古莸这种对环境的适应性变异在叶片的解剖结构上尤为明显, 在前人不同的研究中, 蒙古莸或为等面叶、无海绵组织分化(
符亚儒等, 2005 ;
杨小玉, 2008 ;
李爱平等, 2010 ), 或为异面叶、叶肉中含有海绵组织(
张源润等, 1999 ;
王晓江等, 2006 ;
田英等, 2010 ); 叶片厚度21.6-2 180 μm, 变异极大. ...
... 本研究通过对同质园栽培下7个种源蒙古莸叶片解剖结构的观察, 发现7个种源地的蒙古莸叶片均为典型的等面叶, 叶片由紧密排列的上表皮、发达的栅栏组织和下表皮构成, 无海绵组织分化, 是旱生植物最常见的叶片结构(
Delzon, 2015 ), 与
贺一鸣(2018) 、
杨小玉(2008) 、
李爱平等(2010) 、
符亚儒等(2005) 对蒙古莸叶片解剖结构观察结果一致.蒙古莸叶片解剖结构参数的数量化分析表明, 上表皮细胞厚度均大于下表皮细胞厚度, 这种结构有利于气体交换(
陈旭等, 2019 ); 7个种源蒙古莸叶片的7个解剖结构参数存在显著差异(
表3 ), 其中, 叶厚度变化范围为192.34-270.30 μm, 变异系数为0.12, 与
符亚儒等(2005) 、
李爱平等(2010) 的测量结果近似. ...
... ), 其中, 叶厚度变化范围为192.34-270.30 μm, 变异系数为0.12, 与
符亚儒等(2005) 、
李爱平等(2010) 的测量结果近似. ...
不同区域长柄扁桃叶片解剖结构及其抗旱性分析
1
2013
... 叶片作为植物暴露在生活环境中面积最大的器官, 其外部形态特征、内部解剖结构等最能体现植物对环境因子的适应性进化(
Wright et al ., 2001 ;
Chartzoulakis et al ., 2002 ;
钟悦鸣等, 2017 ;
于海燕等, 2019 ), 开展叶片解剖结构分析, 对研究植物在极端生态环境下的适应策略具有重要意义.一般而言, 植物分布范围广泛, 各自生活在温度、水分等条件不相同的自然地理环境中, 在长期对环境适应过程中, 同种植物叶片的解剖结构表现出明显的差异(
季子敬等, 2013 ;
郭婧宇, 2018 ).叶片的这种功能性状差异可能既有自身的可塑性响应也有遗传变异, 以往研究中沿自然环境梯度的采样测定或单一站点控制实验很难区分两者的效应.全缘叶绿绒蒿(
Meconopsis integrifolia )随着海拔升高, 株型变矮, 叶片组织结构紧密度增大(
刘梦颖等, 2018 ); 生长于干旱地区的长梗扁桃(
Amygdalus pedunculata )叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的长梗扁桃, 其叶片栅栏组织排列疏松, 叶厚度较薄(
郭改改等, 2013 ).这种叶片解剖结构特征的差异同时受到环境变异与自身遗传结构两个因素的影响.同质园试验即研究同一基因型的物种在不同环境下的初生与次生代谢的响应, 或不同基因型的物种在相同环境下的初生与次生代谢的响应.将多个种源的植物在同地栽培, 在一定程度上控制环境与基因型间的相互作用, 最大程度地降低立地条件所造成的影响(
王薪琪等, 2015 ;
陈嘉静等, 2019 ), 进而说明不同种源植物叶片解剖结构特征表现出的差异是由于原种源地环境差异造成的植物遗传性基因变异.这与先前关于环境和遗传因素共同影响(包括气候梯度和种源的影响)叶片解剖结构特征的研究有所不同, 这种同质园方法在局部适应性研究中具有重要意义(
de Villemereuil, 2016 ), 有助于我们更好地理解不同地理种源植物对全球变化的响应和适应机制.目前采用此法对不同种源内植物叶片解剖结构差异及其影响因素的相关分析较少. ...
不同区域长柄扁桃叶片解剖结构及其抗旱性分析
1
2013
... 叶片作为植物暴露在生活环境中面积最大的器官, 其外部形态特征、内部解剖结构等最能体现植物对环境因子的适应性进化(
Wright et al ., 2001 ;
Chartzoulakis et al ., 2002 ;
钟悦鸣等, 2017 ;
于海燕等, 2019 ), 开展叶片解剖结构分析, 对研究植物在极端生态环境下的适应策略具有重要意义.一般而言, 植物分布范围广泛, 各自生活在温度、水分等条件不相同的自然地理环境中, 在长期对环境适应过程中, 同种植物叶片的解剖结构表现出明显的差异(
季子敬等, 2013 ;
郭婧宇, 2018 ).叶片的这种功能性状差异可能既有自身的可塑性响应也有遗传变异, 以往研究中沿自然环境梯度的采样测定或单一站点控制实验很难区分两者的效应.全缘叶绿绒蒿(
Meconopsis integrifolia )随着海拔升高, 株型变矮, 叶片组织结构紧密度增大(
刘梦颖等, 2018 ); 生长于干旱地区的长梗扁桃(
Amygdalus pedunculata )叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的长梗扁桃, 其叶片栅栏组织排列疏松, 叶厚度较薄(
郭改改等, 2013 ).这种叶片解剖结构特征的差异同时受到环境变异与自身遗传结构两个因素的影响.同质园试验即研究同一基因型的物种在不同环境下的初生与次生代谢的响应, 或不同基因型的物种在相同环境下的初生与次生代谢的响应.将多个种源的植物在同地栽培, 在一定程度上控制环境与基因型间的相互作用, 最大程度地降低立地条件所造成的影响(
王薪琪等, 2015 ;
陈嘉静等, 2019 ), 进而说明不同种源植物叶片解剖结构特征表现出的差异是由于原种源地环境差异造成的植物遗传性基因变异.这与先前关于环境和遗传因素共同影响(包括气候梯度和种源的影响)叶片解剖结构特征的研究有所不同, 这种同质园方法在局部适应性研究中具有重要意义(
de Villemereuil, 2016 ), 有助于我们更好地理解不同地理种源植物对全球变化的响应和适应机制.目前采用此法对不同种源内植物叶片解剖结构差异及其影响因素的相关分析较少. ...
三种沙生植物叶形态、结构差异性及环境分异
1
2018
... 叶片作为植物暴露在生活环境中面积最大的器官, 其外部形态特征、内部解剖结构等最能体现植物对环境因子的适应性进化(
Wright et al ., 2001 ;
Chartzoulakis et al ., 2002 ;
钟悦鸣等, 2017 ;
于海燕等, 2019 ), 开展叶片解剖结构分析, 对研究植物在极端生态环境下的适应策略具有重要意义.一般而言, 植物分布范围广泛, 各自生活在温度、水分等条件不相同的自然地理环境中, 在长期对环境适应过程中, 同种植物叶片的解剖结构表现出明显的差异(
季子敬等, 2013 ;
郭婧宇, 2018 ).叶片的这种功能性状差异可能既有自身的可塑性响应也有遗传变异, 以往研究中沿自然环境梯度的采样测定或单一站点控制实验很难区分两者的效应.全缘叶绿绒蒿(
Meconopsis integrifolia )随着海拔升高, 株型变矮, 叶片组织结构紧密度增大(
刘梦颖等, 2018 ); 生长于干旱地区的长梗扁桃(
Amygdalus pedunculata )叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的长梗扁桃, 其叶片栅栏组织排列疏松, 叶厚度较薄(
郭改改等, 2013 ).这种叶片解剖结构特征的差异同时受到环境变异与自身遗传结构两个因素的影响.同质园试验即研究同一基因型的物种在不同环境下的初生与次生代谢的响应, 或不同基因型的物种在相同环境下的初生与次生代谢的响应.将多个种源的植物在同地栽培, 在一定程度上控制环境与基因型间的相互作用, 最大程度地降低立地条件所造成的影响(
王薪琪等, 2015 ;
陈嘉静等, 2019 ), 进而说明不同种源植物叶片解剖结构特征表现出的差异是由于原种源地环境差异造成的植物遗传性基因变异.这与先前关于环境和遗传因素共同影响(包括气候梯度和种源的影响)叶片解剖结构特征的研究有所不同, 这种同质园方法在局部适应性研究中具有重要意义(
de Villemereuil, 2016 ), 有助于我们更好地理解不同地理种源植物对全球变化的响应和适应机制.目前采用此法对不同种源内植物叶片解剖结构差异及其影响因素的相关分析较少. ...
三种沙生植物叶形态、结构差异性及环境分异
1
2018
... 叶片作为植物暴露在生活环境中面积最大的器官, 其外部形态特征、内部解剖结构等最能体现植物对环境因子的适应性进化(
Wright et al ., 2001 ;
Chartzoulakis et al ., 2002 ;
钟悦鸣等, 2017 ;
于海燕等, 2019 ), 开展叶片解剖结构分析, 对研究植物在极端生态环境下的适应策略具有重要意义.一般而言, 植物分布范围广泛, 各自生活在温度、水分等条件不相同的自然地理环境中, 在长期对环境适应过程中, 同种植物叶片的解剖结构表现出明显的差异(
季子敬等, 2013 ;
郭婧宇, 2018 ).叶片的这种功能性状差异可能既有自身的可塑性响应也有遗传变异, 以往研究中沿自然环境梯度的采样测定或单一站点控制实验很难区分两者的效应.全缘叶绿绒蒿(
Meconopsis integrifolia )随着海拔升高, 株型变矮, 叶片组织结构紧密度增大(
刘梦颖等, 2018 ); 生长于干旱地区的长梗扁桃(
Amygdalus pedunculata )叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的长梗扁桃, 其叶片栅栏组织排列疏松, 叶厚度较薄(
郭改改等, 2013 ).这种叶片解剖结构特征的差异同时受到环境变异与自身遗传结构两个因素的影响.同质园试验即研究同一基因型的物种在不同环境下的初生与次生代谢的响应, 或不同基因型的物种在相同环境下的初生与次生代谢的响应.将多个种源的植物在同地栽培, 在一定程度上控制环境与基因型间的相互作用, 最大程度地降低立地条件所造成的影响(
王薪琪等, 2015 ;
陈嘉静等, 2019 ), 进而说明不同种源植物叶片解剖结构特征表现出的差异是由于原种源地环境差异造成的植物遗传性基因变异.这与先前关于环境和遗传因素共同影响(包括气候梯度和种源的影响)叶片解剖结构特征的研究有所不同, 这种同质园方法在局部适应性研究中具有重要意义(
de Villemereuil, 2016 ), 有助于我们更好地理解不同地理种源植物对全球变化的响应和适应机制.目前采用此法对不同种源内植物叶片解剖结构差异及其影响因素的相关分析较少. ...
青海高寒半干旱区蒙古莸叶水势变化及其与环境因素的关系
1
2008
... 蒙古莸(
Caryopteris mongholica )属唇形科筋骨草亚科莸属(
Caryopteris ), 是我国干旱、半干旱地区一种耐旱、耐寒兼具观赏性的绿化灌木, 是防风固沙和水土保持的优良造林树种(
李玉俊等, 1991 ;
韩磊等, 2008 ).蒙古莸分布在东起呼伦贝尔典型草原区, 西至甘肃河西走廊西部和青海湖南部的广阔的蒙古高原上.然而, 不同的生长环境导致蒙古莸的生理特征和遗传结构均出现不同程度的变异, 这种对环境的适应性变异表现, 在叶片的解剖结构上尤为明显.
白雪卡等(2018) 对同质园栽培下8个不同种源蒙古莸光响应曲线的研究发现, 蒙古莸光合响应能力在不同种源间表现出明显不同, 光响应曲线也存在显著的差异.蒙古莸这种对环境的适应性变异在叶片的解剖结构上尤为明显, 在前人不同的研究中, 蒙古莸或为等面叶、无海绵组织分化(
符亚儒等, 2005 ;
杨小玉, 2008 ;
李爱平等, 2010 ), 或为异面叶、叶肉中含有海绵组织(
张源润等, 1999 ;
王晓江等, 2006 ;
田英等, 2010 ); 叶片厚度21.6-2 180 μm, 变异极大. ...
青海高寒半干旱区蒙古莸叶水势变化及其与环境因素的关系
1
2008
... 蒙古莸(
Caryopteris mongholica )属唇形科筋骨草亚科莸属(
Caryopteris ), 是我国干旱、半干旱地区一种耐旱、耐寒兼具观赏性的绿化灌木, 是防风固沙和水土保持的优良造林树种(
李玉俊等, 1991 ;
韩磊等, 2008 ).蒙古莸分布在东起呼伦贝尔典型草原区, 西至甘肃河西走廊西部和青海湖南部的广阔的蒙古高原上.然而, 不同的生长环境导致蒙古莸的生理特征和遗传结构均出现不同程度的变异, 这种对环境的适应性变异表现, 在叶片的解剖结构上尤为明显.
白雪卡等(2018) 对同质园栽培下8个不同种源蒙古莸光响应曲线的研究发现, 蒙古莸光合响应能力在不同种源间表现出明显不同, 光响应曲线也存在显著的差异.蒙古莸这种对环境的适应性变异在叶片的解剖结构上尤为明显, 在前人不同的研究中, 蒙古莸或为等面叶、无海绵组织分化(
符亚儒等, 2005 ;
杨小玉, 2008 ;
李爱平等, 2010 ), 或为异面叶、叶肉中含有海绵组织(
张源润等, 1999 ;
王晓江等, 2006 ;
田英等, 2010 ); 叶片厚度21.6-2 180 μm, 变异极大. ...
Taxonomic, phylogenetic, and environmental trade-offs between leaf productivity and persistence
1
2009
... 结合一般线性模型(GLM) ANOVA分析种源地气候对蒙古莸叶解剖结构参数值的影响作用大小(
He et al ., 2009 ).基于种源地环境因素与叶片解剖结构参数值的相关关系, 选取
MAP 、
MAT 、
PE 、海拔(
Alt )、地理位置(Site)等5个种源地气候环境因子进入模型. ...
蒙古莸种群生态适应及繁殖更新策略
1
2018
... 本研究通过对同质园栽培下7个种源蒙古莸叶片解剖结构的观察, 发现7个种源地的蒙古莸叶片均为典型的等面叶, 叶片由紧密排列的上表皮、发达的栅栏组织和下表皮构成, 无海绵组织分化, 是旱生植物最常见的叶片结构(
Delzon, 2015 ), 与
贺一鸣(2018) 、
杨小玉(2008) 、
李爱平等(2010) 、
符亚儒等(2005) 对蒙古莸叶片解剖结构观察结果一致.蒙古莸叶片解剖结构参数的数量化分析表明, 上表皮细胞厚度均大于下表皮细胞厚度, 这种结构有利于气体交换(
陈旭等, 2019 ); 7个种源蒙古莸叶片的7个解剖结构参数存在显著差异(
表3 ), 其中, 叶厚度变化范围为192.34-270.30 μm, 变异系数为0.12, 与
符亚儒等(2005) 、
李爱平等(2010) 的测量结果近似. ...
蒙古莸种群生态适应及繁殖更新策略
1
2018
... 本研究通过对同质园栽培下7个种源蒙古莸叶片解剖结构的观察, 发现7个种源地的蒙古莸叶片均为典型的等面叶, 叶片由紧密排列的上表皮、发达的栅栏组织和下表皮构成, 无海绵组织分化, 是旱生植物最常见的叶片结构(
Delzon, 2015 ), 与
贺一鸣(2018) 、
杨小玉(2008) 、
李爱平等(2010) 、
符亚儒等(2005) 对蒙古莸叶片解剖结构观察结果一致.蒙古莸叶片解剖结构参数的数量化分析表明, 上表皮细胞厚度均大于下表皮细胞厚度, 这种结构有利于气体交换(
陈旭等, 2019 ); 7个种源蒙古莸叶片的7个解剖结构参数存在显著差异(
表3 ), 其中, 叶厚度变化范围为192.34-270.30 μm, 变异系数为0.12, 与
符亚儒等(2005) 、
李爱平等(2010) 的测量结果近似. ...
Very high resolution interpolated climate surfaces for global land areas
1
2005
... 根据蒙古莸不同种源地的经度与纬度, 从ArcGIS 10.2的WorldCLIM全球高分辨率气候数据库(
Hijmans et al ., 2005 )中选取各样地的月平均气候数据(2007-2016年), 并计算各气候指标大小.气候指标包括: 年降水量(
MAP )、生长季降水量(
GSP )、降水年较差(
ARP )、年平均气温(
MAT )、生长季平均气温(
GST )、气温年较差(
ART )、潜在蒸散量(
PE ). ...
西藏紫花针茅叶功能性状沿降水梯度的变化
1
2012
... 植物在进化过程中, 受到外界温度、水分、光照等环境因素的长期作用而影响其建成, 逐步形成适应环境的表型和遗传特征.即使是同一物种, 长期生长在不同的地理环境中, 也会产生不同程度的基因分化, 以适应各自不同的生态位, 从而形成特定的种源特性(
Smith et al ., 1997 ;
Barboni et al ., 2004 ;
Pigliucci, 2006 ;
胡梦瑶等, 2012 ).植物自然群体分布的环境具有明显差异.从表型来看, 同一结构由于环境分化选择作用, 不同环境条件下的群体最适表型也存在差异, 而表型的差异归根结底是由控制表型的基因型不同所导致的. ...
西藏紫花针茅叶功能性状沿降水梯度的变化
1
2012
... 植物在进化过程中, 受到外界温度、水分、光照等环境因素的长期作用而影响其建成, 逐步形成适应环境的表型和遗传特征.即使是同一物种, 长期生长在不同的地理环境中, 也会产生不同程度的基因分化, 以适应各自不同的生态位, 从而形成特定的种源特性(
Smith et al ., 1997 ;
Barboni et al ., 2004 ;
Pigliucci, 2006 ;
胡梦瑶等, 2012 ).植物自然群体分布的环境具有明显差异.从表型来看, 同一结构由于环境分化选择作用, 不同环境条件下的群体最适表型也存在差异, 而表型的差异归根结底是由控制表型的基因型不同所导致的. ...
兴安落叶松针叶解剖结构变化及其光合能力对气候变化的适应性
2
2013
... 叶片作为植物暴露在生活环境中面积最大的器官, 其外部形态特征、内部解剖结构等最能体现植物对环境因子的适应性进化(
Wright et al ., 2001 ;
Chartzoulakis et al ., 2002 ;
钟悦鸣等, 2017 ;
于海燕等, 2019 ), 开展叶片解剖结构分析, 对研究植物在极端生态环境下的适应策略具有重要意义.一般而言, 植物分布范围广泛, 各自生活在温度、水分等条件不相同的自然地理环境中, 在长期对环境适应过程中, 同种植物叶片的解剖结构表现出明显的差异(
季子敬等, 2013 ;
郭婧宇, 2018 ).叶片的这种功能性状差异可能既有自身的可塑性响应也有遗传变异, 以往研究中沿自然环境梯度的采样测定或单一站点控制实验很难区分两者的效应.全缘叶绿绒蒿(
Meconopsis integrifolia )随着海拔升高, 株型变矮, 叶片组织结构紧密度增大(
刘梦颖等, 2018 ); 生长于干旱地区的长梗扁桃(
Amygdalus pedunculata )叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的长梗扁桃, 其叶片栅栏组织排列疏松, 叶厚度较薄(
郭改改等, 2013 ).这种叶片解剖结构特征的差异同时受到环境变异与自身遗传结构两个因素的影响.同质园试验即研究同一基因型的物种在不同环境下的初生与次生代谢的响应, 或不同基因型的物种在相同环境下的初生与次生代谢的响应.将多个种源的植物在同地栽培, 在一定程度上控制环境与基因型间的相互作用, 最大程度地降低立地条件所造成的影响(
王薪琪等, 2015 ;
陈嘉静等, 2019 ), 进而说明不同种源植物叶片解剖结构特征表现出的差异是由于原种源地环境差异造成的植物遗传性基因变异.这与先前关于环境和遗传因素共同影响(包括气候梯度和种源的影响)叶片解剖结构特征的研究有所不同, 这种同质园方法在局部适应性研究中具有重要意义(
de Villemereuil, 2016 ), 有助于我们更好地理解不同地理种源植物对全球变化的响应和适应机制.目前采用此法对不同种源内植物叶片解剖结构差异及其影响因素的相关分析较少. ...
... 原种源地的地理位置(经纬度)、温度(
MAT 、
ART )、
MAP 、
PE 、
Alt 与叶片解剖结构指标密切相关(
表6 ), 能够很好地解释其遗传变异程度, 其中种源地气候(
MAT 、
MAP 、
PE )对蒙古莸叶片解剖结构解释程度在34.09%-81.43%之间.
季子敬等(2013) 对兴安落叶松(
Larix gmelinii )针叶解剖结构的分析研究中发现, 针叶长期适应当地环境条件后, 在解剖结构特征上也产生了具有遗传能力的变异.
赵小仙等(2015) 对蒙古沙拐枣(
Calligonum monglicum )叶片解剖结构分析研究中发现, 解剖结构的差异与经纬度、
MAT 、
MAP 的变化有极显著相关性.
白雪卡等(2018) 对不同种源蒙古莸光响应特性的研究中也同样证明, 种源地气候驱动的遗传变异对蒙古莸光合作用参数有显著的解释力, 解释程度在16.65%- 73.52%之间, 与本研究的解释力近似.本研究中, 上角质层厚度、上下表皮细胞厚度、下栅栏组织厚度、栅栏组织总厚度、叶片结构紧密度主要受
MAT 的影响, 解释程度在20.77%-71.89%之间(
表6 ); 上栅栏组织厚度、叶厚度主要受
MAP 影响, 解释程度在31.40%-45.93%之间(
表6 ).相对而言, 温度对蒙古莸叶片解剖结构参数的解释度大于水分, 这与
田丽丽等(2019) 对大叶种茶树叶片解剖结构的分析不谋而合, 其研究发现叶片生长受到温度、降水和光照因子等综合因素的影响, 叶片结构的差异则主要受到温度影响. ...
兴安落叶松针叶解剖结构变化及其光合能力对气候变化的适应性
2
2013
... 叶片作为植物暴露在生活环境中面积最大的器官, 其外部形态特征、内部解剖结构等最能体现植物对环境因子的适应性进化(
Wright et al ., 2001 ;
Chartzoulakis et al ., 2002 ;
钟悦鸣等, 2017 ;
于海燕等, 2019 ), 开展叶片解剖结构分析, 对研究植物在极端生态环境下的适应策略具有重要意义.一般而言, 植物分布范围广泛, 各自生活在温度、水分等条件不相同的自然地理环境中, 在长期对环境适应过程中, 同种植物叶片的解剖结构表现出明显的差异(
季子敬等, 2013 ;
郭婧宇, 2018 ).叶片的这种功能性状差异可能既有自身的可塑性响应也有遗传变异, 以往研究中沿自然环境梯度的采样测定或单一站点控制实验很难区分两者的效应.全缘叶绿绒蒿(
Meconopsis integrifolia )随着海拔升高, 株型变矮, 叶片组织结构紧密度增大(
刘梦颖等, 2018 ); 生长于干旱地区的长梗扁桃(
Amygdalus pedunculata )叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的长梗扁桃, 其叶片栅栏组织排列疏松, 叶厚度较薄(
郭改改等, 2013 ).这种叶片解剖结构特征的差异同时受到环境变异与自身遗传结构两个因素的影响.同质园试验即研究同一基因型的物种在不同环境下的初生与次生代谢的响应, 或不同基因型的物种在相同环境下的初生与次生代谢的响应.将多个种源的植物在同地栽培, 在一定程度上控制环境与基因型间的相互作用, 最大程度地降低立地条件所造成的影响(
王薪琪等, 2015 ;
陈嘉静等, 2019 ), 进而说明不同种源植物叶片解剖结构特征表现出的差异是由于原种源地环境差异造成的植物遗传性基因变异.这与先前关于环境和遗传因素共同影响(包括气候梯度和种源的影响)叶片解剖结构特征的研究有所不同, 这种同质园方法在局部适应性研究中具有重要意义(
de Villemereuil, 2016 ), 有助于我们更好地理解不同地理种源植物对全球变化的响应和适应机制.目前采用此法对不同种源内植物叶片解剖结构差异及其影响因素的相关分析较少. ...
... 原种源地的地理位置(经纬度)、温度(
MAT 、
ART )、
MAP 、
PE 、
Alt 与叶片解剖结构指标密切相关(
表6 ), 能够很好地解释其遗传变异程度, 其中种源地气候(
MAT 、
MAP 、
PE )对蒙古莸叶片解剖结构解释程度在34.09%-81.43%之间.
季子敬等(2013) 对兴安落叶松(
Larix gmelinii )针叶解剖结构的分析研究中发现, 针叶长期适应当地环境条件后, 在解剖结构特征上也产生了具有遗传能力的变异.
赵小仙等(2015) 对蒙古沙拐枣(
Calligonum monglicum )叶片解剖结构分析研究中发现, 解剖结构的差异与经纬度、
MAT 、
MAP 的变化有极显著相关性.
白雪卡等(2018) 对不同种源蒙古莸光响应特性的研究中也同样证明, 种源地气候驱动的遗传变异对蒙古莸光合作用参数有显著的解释力, 解释程度在16.65%- 73.52%之间, 与本研究的解释力近似.本研究中, 上角质层厚度、上下表皮细胞厚度、下栅栏组织厚度、栅栏组织总厚度、叶片结构紧密度主要受
MAT 的影响, 解释程度在20.77%-71.89%之间(
表6 ); 上栅栏组织厚度、叶厚度主要受
MAP 影响, 解释程度在31.40%-45.93%之间(
表6 ).相对而言, 温度对蒙古莸叶片解剖结构参数的解释度大于水分, 这与
田丽丽等(2019) 对大叶种茶树叶片解剖结构的分析不谋而合, 其研究发现叶片生长受到温度、降水和光照因子等综合因素的影响, 叶片结构的差异则主要受到温度影响. ...
Biosynthetic pathways of cutin and waxes, and their sensitivity to environmental stresses
1
1996
... 本研究中的
MAT 与叶片结构紧密度相关性最强(
表5 ), 叶片结构紧密度随温度升高而增大, 这可能是由于高温时植物呼吸与代谢作用加强, 加剧了水分散失, 导致叶肉组织向着提高光合效率和水分储量方向发展.而发达的栅栏组织可以有效地储存水分, 栅栏组织的细胞排列越紧密, 细胞间隙越少, 叶片结构紧密度就越大, 能更好地适应高温环境.最西南部的阿拉善左旗属高温少雨地区, 潜在蒸散量最大, 导致该种源的蒙古莸叶片的上下栅栏组织厚度、叶厚度以及叶片结构紧密度均最大(
表1 ,
表3 ).除了与栅栏组织有关的参数外,
GST 与上角质层厚度、上表皮细胞厚度呈现出显著的负相关性, 即上角质层厚度与上表皮细胞厚度随生长季温度的降低而增大.叶表皮外角质层是由表皮细胞合成的脂类和碳水化合物的聚合物组成(
Kolattukudy, 1996 ), 角质层越厚, 蜡被越厚, 相对的抗寒能力就越好; 而较大的表皮细胞厚度有利于光的透射, 且具有较强的贮水作用, 当温度降低时, 植物叶片可能通过增加表皮细胞厚度来保证光合作用的正常进行, 并将水分存储其中来应对栅栏组织厚度的减小.自相关性分析(
表4 )表明, 上角质层厚度与下表皮细胞厚度间存在显著正相关关系(
p < 0.01), 即上角质层厚度发生变化时, 会引起下表皮细胞厚度发生正向变化.
MAP 、
GSP 均与上栅栏组织厚度相关性最强(
表5 ), 与栅栏组织总厚度、叶片厚度显著负相关(
p < 0.05), 即这3个解剖结构指标随降水量的增大而减小, 这可能是因为环境水分充足时, 植物不需要储存过多的水分.由此可见, 受水分含量变化影响最大的是上栅栏组织厚度, 其次是叶厚度与栅栏组织总厚度, 而下栅栏组织厚度与降水量没有表现出明显的相关性.同时, 自相关性分析(
表4 )表明, 上栅栏组织厚度、栅栏组织总厚度、叶厚度这3个指标显著正相关(
p < 0.01), 即任一指标变化, 会引起其他两个指标发生协同变化, 其中上栅栏组织厚度变 化是蒙古莸叶片应对生境水分含量变化最主要的 方式. ...
库布齐沙漠几种沙生灌木叶解剖结构耐旱特征研究
3
2010
... 蒙古莸(
Caryopteris mongholica )属唇形科筋骨草亚科莸属(
Caryopteris ), 是我国干旱、半干旱地区一种耐旱、耐寒兼具观赏性的绿化灌木, 是防风固沙和水土保持的优良造林树种(
李玉俊等, 1991 ;
韩磊等, 2008 ).蒙古莸分布在东起呼伦贝尔典型草原区, 西至甘肃河西走廊西部和青海湖南部的广阔的蒙古高原上.然而, 不同的生长环境导致蒙古莸的生理特征和遗传结构均出现不同程度的变异, 这种对环境的适应性变异表现, 在叶片的解剖结构上尤为明显.
白雪卡等(2018) 对同质园栽培下8个不同种源蒙古莸光响应曲线的研究发现, 蒙古莸光合响应能力在不同种源间表现出明显不同, 光响应曲线也存在显著的差异.蒙古莸这种对环境的适应性变异在叶片的解剖结构上尤为明显, 在前人不同的研究中, 蒙古莸或为等面叶、无海绵组织分化(
符亚儒等, 2005 ;
杨小玉, 2008 ;
李爱平等, 2010 ), 或为异面叶、叶肉中含有海绵组织(
张源润等, 1999 ;
王晓江等, 2006 ;
田英等, 2010 ); 叶片厚度21.6-2 180 μm, 变异极大. ...
... 本研究通过对同质园栽培下7个种源蒙古莸叶片解剖结构的观察, 发现7个种源地的蒙古莸叶片均为典型的等面叶, 叶片由紧密排列的上表皮、发达的栅栏组织和下表皮构成, 无海绵组织分化, 是旱生植物最常见的叶片结构(
Delzon, 2015 ), 与
贺一鸣(2018) 、
杨小玉(2008) 、
李爱平等(2010) 、
符亚儒等(2005) 对蒙古莸叶片解剖结构观察结果一致.蒙古莸叶片解剖结构参数的数量化分析表明, 上表皮细胞厚度均大于下表皮细胞厚度, 这种结构有利于气体交换(
陈旭等, 2019 ); 7个种源蒙古莸叶片的7个解剖结构参数存在显著差异(
表3 ), 其中, 叶厚度变化范围为192.34-270.30 μm, 变异系数为0.12, 与
符亚儒等(2005) 、
李爱平等(2010) 的测量结果近似. ...
... 、
李爱平等(2010) 的测量结果近似. ...
库布齐沙漠几种沙生灌木叶解剖结构耐旱特征研究
3
2010
... 蒙古莸(
Caryopteris mongholica )属唇形科筋骨草亚科莸属(
Caryopteris ), 是我国干旱、半干旱地区一种耐旱、耐寒兼具观赏性的绿化灌木, 是防风固沙和水土保持的优良造林树种(
李玉俊等, 1991 ;
韩磊等, 2008 ).蒙古莸分布在东起呼伦贝尔典型草原区, 西至甘肃河西走廊西部和青海湖南部的广阔的蒙古高原上.然而, 不同的生长环境导致蒙古莸的生理特征和遗传结构均出现不同程度的变异, 这种对环境的适应性变异表现, 在叶片的解剖结构上尤为明显.
白雪卡等(2018) 对同质园栽培下8个不同种源蒙古莸光响应曲线的研究发现, 蒙古莸光合响应能力在不同种源间表现出明显不同, 光响应曲线也存在显著的差异.蒙古莸这种对环境的适应性变异在叶片的解剖结构上尤为明显, 在前人不同的研究中, 蒙古莸或为等面叶、无海绵组织分化(
符亚儒等, 2005 ;
杨小玉, 2008 ;
李爱平等, 2010 ), 或为异面叶、叶肉中含有海绵组织(
张源润等, 1999 ;
王晓江等, 2006 ;
田英等, 2010 ); 叶片厚度21.6-2 180 μm, 变异极大. ...
... 本研究通过对同质园栽培下7个种源蒙古莸叶片解剖结构的观察, 发现7个种源地的蒙古莸叶片均为典型的等面叶, 叶片由紧密排列的上表皮、发达的栅栏组织和下表皮构成, 无海绵组织分化, 是旱生植物最常见的叶片结构(
Delzon, 2015 ), 与
贺一鸣(2018) 、
杨小玉(2008) 、
李爱平等(2010) 、
符亚儒等(2005) 对蒙古莸叶片解剖结构观察结果一致.蒙古莸叶片解剖结构参数的数量化分析表明, 上表皮细胞厚度均大于下表皮细胞厚度, 这种结构有利于气体交换(
陈旭等, 2019 ); 7个种源蒙古莸叶片的7个解剖结构参数存在显著差异(
表3 ), 其中, 叶厚度变化范围为192.34-270.30 μm, 变异系数为0.12, 与
符亚儒等(2005) 、
李爱平等(2010) 的测量结果近似. ...
... 、
李爱平等(2010) 的测量结果近似. ...
植物叶片形态解剖结构对环境变化的响应与适应
1
2005
... 用Image J软件测量叶片的上角质层厚度, 上、下表皮细胞厚度, 上、下栅栏组织厚度, 叶厚度等叶解剖结构指标, 并计算叶片结构紧密度、栅栏组织总厚度(
李芳兰和包维楷, 2005 ), 每个指标重复测量5组数据. ...
植物叶片形态解剖结构对环境变化的响应与适应
1
2005
... 用Image J软件测量叶片的上角质层厚度, 上、下表皮细胞厚度, 上、下栅栏组织厚度, 叶厚度等叶解剖结构指标, 并计算叶片结构紧密度、栅栏组织总厚度(
李芳兰和包维楷, 2005 ), 每个指标重复测量5组数据. ...
芳香植物——蒙古莸的生态生理特性及其栽培技术
1
1991
... 蒙古莸(
Caryopteris mongholica )属唇形科筋骨草亚科莸属(
Caryopteris ), 是我国干旱、半干旱地区一种耐旱、耐寒兼具观赏性的绿化灌木, 是防风固沙和水土保持的优良造林树种(
李玉俊等, 1991 ;
韩磊等, 2008 ).蒙古莸分布在东起呼伦贝尔典型草原区, 西至甘肃河西走廊西部和青海湖南部的广阔的蒙古高原上.然而, 不同的生长环境导致蒙古莸的生理特征和遗传结构均出现不同程度的变异, 这种对环境的适应性变异表现, 在叶片的解剖结构上尤为明显.
白雪卡等(2018) 对同质园栽培下8个不同种源蒙古莸光响应曲线的研究发现, 蒙古莸光合响应能力在不同种源间表现出明显不同, 光响应曲线也存在显著的差异.蒙古莸这种对环境的适应性变异在叶片的解剖结构上尤为明显, 在前人不同的研究中, 蒙古莸或为等面叶、无海绵组织分化(
符亚儒等, 2005 ;
杨小玉, 2008 ;
李爱平等, 2010 ), 或为异面叶、叶肉中含有海绵组织(
张源润等, 1999 ;
王晓江等, 2006 ;
田英等, 2010 ); 叶片厚度21.6-2 180 μm, 变异极大. ...
芳香植物——蒙古莸的生态生理特性及其栽培技术
1
1991
... 蒙古莸(
Caryopteris mongholica )属唇形科筋骨草亚科莸属(
Caryopteris ), 是我国干旱、半干旱地区一种耐旱、耐寒兼具观赏性的绿化灌木, 是防风固沙和水土保持的优良造林树种(
李玉俊等, 1991 ;
韩磊等, 2008 ).蒙古莸分布在东起呼伦贝尔典型草原区, 西至甘肃河西走廊西部和青海湖南部的广阔的蒙古高原上.然而, 不同的生长环境导致蒙古莸的生理特征和遗传结构均出现不同程度的变异, 这种对环境的适应性变异表现, 在叶片的解剖结构上尤为明显.
白雪卡等(2018) 对同质园栽培下8个不同种源蒙古莸光响应曲线的研究发现, 蒙古莸光合响应能力在不同种源间表现出明显不同, 光响应曲线也存在显著的差异.蒙古莸这种对环境的适应性变异在叶片的解剖结构上尤为明显, 在前人不同的研究中, 蒙古莸或为等面叶、无海绵组织分化(
符亚儒等, 2005 ;
杨小玉, 2008 ;
李爱平等, 2010 ), 或为异面叶、叶肉中含有海绵组织(
张源润等, 1999 ;
王晓江等, 2006 ;
田英等, 2010 ); 叶片厚度21.6-2 180 μm, 变异极大. ...
高山植物全缘叶绿绒蒿叶片形态及解剖结构对海拔的响应
1
2018
... 叶片作为植物暴露在生活环境中面积最大的器官, 其外部形态特征、内部解剖结构等最能体现植物对环境因子的适应性进化(
Wright et al ., 2001 ;
Chartzoulakis et al ., 2002 ;
钟悦鸣等, 2017 ;
于海燕等, 2019 ), 开展叶片解剖结构分析, 对研究植物在极端生态环境下的适应策略具有重要意义.一般而言, 植物分布范围广泛, 各自生活在温度、水分等条件不相同的自然地理环境中, 在长期对环境适应过程中, 同种植物叶片的解剖结构表现出明显的差异(
季子敬等, 2013 ;
郭婧宇, 2018 ).叶片的这种功能性状差异可能既有自身的可塑性响应也有遗传变异, 以往研究中沿自然环境梯度的采样测定或单一站点控制实验很难区分两者的效应.全缘叶绿绒蒿(
Meconopsis integrifolia )随着海拔升高, 株型变矮, 叶片组织结构紧密度增大(
刘梦颖等, 2018 ); 生长于干旱地区的长梗扁桃(
Amygdalus pedunculata )叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的长梗扁桃, 其叶片栅栏组织排列疏松, 叶厚度较薄(
郭改改等, 2013 ).这种叶片解剖结构特征的差异同时受到环境变异与自身遗传结构两个因素的影响.同质园试验即研究同一基因型的物种在不同环境下的初生与次生代谢的响应, 或不同基因型的物种在相同环境下的初生与次生代谢的响应.将多个种源的植物在同地栽培, 在一定程度上控制环境与基因型间的相互作用, 最大程度地降低立地条件所造成的影响(
王薪琪等, 2015 ;
陈嘉静等, 2019 ), 进而说明不同种源植物叶片解剖结构特征表现出的差异是由于原种源地环境差异造成的植物遗传性基因变异.这与先前关于环境和遗传因素共同影响(包括气候梯度和种源的影响)叶片解剖结构特征的研究有所不同, 这种同质园方法在局部适应性研究中具有重要意义(
de Villemereuil, 2016 ), 有助于我们更好地理解不同地理种源植物对全球变化的响应和适应机制.目前采用此法对不同种源内植物叶片解剖结构差异及其影响因素的相关分析较少. ...
高山植物全缘叶绿绒蒿叶片形态及解剖结构对海拔的响应
1
2018
... 叶片作为植物暴露在生活环境中面积最大的器官, 其外部形态特征、内部解剖结构等最能体现植物对环境因子的适应性进化(
Wright et al ., 2001 ;
Chartzoulakis et al ., 2002 ;
钟悦鸣等, 2017 ;
于海燕等, 2019 ), 开展叶片解剖结构分析, 对研究植物在极端生态环境下的适应策略具有重要意义.一般而言, 植物分布范围广泛, 各自生活在温度、水分等条件不相同的自然地理环境中, 在长期对环境适应过程中, 同种植物叶片的解剖结构表现出明显的差异(
季子敬等, 2013 ;
郭婧宇, 2018 ).叶片的这种功能性状差异可能既有自身的可塑性响应也有遗传变异, 以往研究中沿自然环境梯度的采样测定或单一站点控制实验很难区分两者的效应.全缘叶绿绒蒿(
Meconopsis integrifolia )随着海拔升高, 株型变矮, 叶片组织结构紧密度增大(
刘梦颖等, 2018 ); 生长于干旱地区的长梗扁桃(
Amygdalus pedunculata )叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的长梗扁桃, 其叶片栅栏组织排列疏松, 叶厚度较薄(
郭改改等, 2013 ).这种叶片解剖结构特征的差异同时受到环境变异与自身遗传结构两个因素的影响.同质园试验即研究同一基因型的物种在不同环境下的初生与次生代谢的响应, 或不同基因型的物种在相同环境下的初生与次生代谢的响应.将多个种源的植物在同地栽培, 在一定程度上控制环境与基因型间的相互作用, 最大程度地降低立地条件所造成的影响(
王薪琪等, 2015 ;
陈嘉静等, 2019 ), 进而说明不同种源植物叶片解剖结构特征表现出的差异是由于原种源地环境差异造成的植物遗传性基因变异.这与先前关于环境和遗传因素共同影响(包括气候梯度和种源的影响)叶片解剖结构特征的研究有所不同, 这种同质园方法在局部适应性研究中具有重要意义(
de Villemereuil, 2016 ), 有助于我们更好地理解不同地理种源植物对全球变化的响应和适应机制.目前采用此法对不同种源内植物叶片解剖结构差异及其影响因素的相关分析较少. ...
不同种源地文冠果叶片解剖结构比较及抗旱性分析
1
2013
... 蒙古莸在我国的分布区域广泛, 生长环境条件差异较大, 为满足自身正常的生长, 对环境和气候条件的适应就至关重要.植物形态结构是植物适应环境而进化出的生存策略, 由于环境分化选择作用, 植物逐步形成适应环境的表型和遗传特征, 表现出独特的种源特性; 即使将不同种源植物移植到相同环境下生长, 其叶片的解剖结构等表型特征仍存在着差异(
马小芬等, 2013 ).这种不同种源间特性的差异正是植物趋异适应的体现, 归根结底是由于控制表型的基因型不同所导致的.本研究采用同质园法, 削弱不同种源地环境对植物个体性状的影响, 将原种源气候差异驱动的遗传变异对蒙古莸叶片解剖结构的影响作为研究重点, 分析结果表明对蒙古莸解剖结构参数有显著的解释力.植物对原生境地气候条件长期适应, 已经产生了遗传结构上的差异, 不同种源蒙古莸叶片解剖结构的差异是其适应特有环境的体现; 同时, 不同解剖结构间存在着协同变化, 反映出植物对干旱环境的良好适应性, 这种适应性策略与近年来提出的叶经济谱概念吻合(
Wright et al ., 2004 ;
Pigliucci, 2006 ).研究结果可为蒙古莸优良抗逆种源选择、遗传资源保护等提供实际应用指导, 并为进一步探究全球变化背景下蒙古莸叶解剖结构对环境变化的响应机制等提供重要的理论依据.此外, 造成不同种源蒙古莸叶片解剖结构差异的因素还有很多, 包括种源地土壤、CO
2 浓度等其他环境因素, 还需在今后的研究中进一步地深入和细化. ...
不同种源地文冠果叶片解剖结构比较及抗旱性分析
1
2013
... 蒙古莸在我国的分布区域广泛, 生长环境条件差异较大, 为满足自身正常的生长, 对环境和气候条件的适应就至关重要.植物形态结构是植物适应环境而进化出的生存策略, 由于环境分化选择作用, 植物逐步形成适应环境的表型和遗传特征, 表现出独特的种源特性; 即使将不同种源植物移植到相同环境下生长, 其叶片的解剖结构等表型特征仍存在着差异(
马小芬等, 2013 ).这种不同种源间特性的差异正是植物趋异适应的体现, 归根结底是由于控制表型的基因型不同所导致的.本研究采用同质园法, 削弱不同种源地环境对植物个体性状的影响, 将原种源气候差异驱动的遗传变异对蒙古莸叶片解剖结构的影响作为研究重点, 分析结果表明对蒙古莸解剖结构参数有显著的解释力.植物对原生境地气候条件长期适应, 已经产生了遗传结构上的差异, 不同种源蒙古莸叶片解剖结构的差异是其适应特有环境的体现; 同时, 不同解剖结构间存在着协同变化, 反映出植物对干旱环境的良好适应性, 这种适应性策略与近年来提出的叶经济谱概念吻合(
Wright et al ., 2004 ;
Pigliucci, 2006 ).研究结果可为蒙古莸优良抗逆种源选择、遗传资源保护等提供实际应用指导, 并为进一步探究全球变化背景下蒙古莸叶解剖结构对环境变化的响应机制等提供重要的理论依据.此外, 造成不同种源蒙古莸叶片解剖结构差异的因素还有很多, 包括种源地土壤、CO
2 浓度等其他环境因素, 还需在今后的研究中进一步地深入和细化. ...
Phenotypic plasticity and evolution by genetic assimilation
2
2006
... 植物在进化过程中, 受到外界温度、水分、光照等环境因素的长期作用而影响其建成, 逐步形成适应环境的表型和遗传特征.即使是同一物种, 长期生长在不同的地理环境中, 也会产生不同程度的基因分化, 以适应各自不同的生态位, 从而形成特定的种源特性(
Smith et al ., 1997 ;
Barboni et al ., 2004 ;
Pigliucci, 2006 ;
胡梦瑶等, 2012 ).植物自然群体分布的环境具有明显差异.从表型来看, 同一结构由于环境分化选择作用, 不同环境条件下的群体最适表型也存在差异, 而表型的差异归根结底是由控制表型的基因型不同所导致的. ...
... 蒙古莸在我国的分布区域广泛, 生长环境条件差异较大, 为满足自身正常的生长, 对环境和气候条件的适应就至关重要.植物形态结构是植物适应环境而进化出的生存策略, 由于环境分化选择作用, 植物逐步形成适应环境的表型和遗传特征, 表现出独特的种源特性; 即使将不同种源植物移植到相同环境下生长, 其叶片的解剖结构等表型特征仍存在着差异(
马小芬等, 2013 ).这种不同种源间特性的差异正是植物趋异适应的体现, 归根结底是由于控制表型的基因型不同所导致的.本研究采用同质园法, 削弱不同种源地环境对植物个体性状的影响, 将原种源气候差异驱动的遗传变异对蒙古莸叶片解剖结构的影响作为研究重点, 分析结果表明对蒙古莸解剖结构参数有显著的解释力.植物对原生境地气候条件长期适应, 已经产生了遗传结构上的差异, 不同种源蒙古莸叶片解剖结构的差异是其适应特有环境的体现; 同时, 不同解剖结构间存在着协同变化, 反映出植物对干旱环境的良好适应性, 这种适应性策略与近年来提出的叶经济谱概念吻合(
Wright et al ., 2004 ;
Pigliucci, 2006 ).研究结果可为蒙古莸优良抗逆种源选择、遗传资源保护等提供实际应用指导, 并为进一步探究全球变化背景下蒙古莸叶解剖结构对环境变化的响应机制等提供重要的理论依据.此外, 造成不同种源蒙古莸叶片解剖结构差异的因素还有很多, 包括种源地土壤、CO
2 浓度等其他环境因素, 还需在今后的研究中进一步地深入和细化. ...
越冬期遮阴条件下3个不同秋眠型紫花苜蓿品种叶片解剖结构与其光生态适应性
1
2012
... 叶片是反映植物在不同生态环境条件下特点的首要器官, 其形态结构是自身生理代谢的需要和对外界环境的逐渐适应, 也是植物对环境变化最为敏感的一个器官(
覃凤飞等, 2012 ).同一种植物生长在不同的环境中, 叶片的解剖结构会表现出不同程度的分化差异. ...
越冬期遮阴条件下3个不同秋眠型紫花苜蓿品种叶片解剖结构与其光生态适应性
1
2012
... 叶片是反映植物在不同生态环境条件下特点的首要器官, 其形态结构是自身生理代谢的需要和对外界环境的逐渐适应, 也是植物对环境变化最为敏感的一个器官(
覃凤飞等, 2012 ).同一种植物生长在不同的环境中, 叶片的解剖结构会表现出不同程度的分化差异. ...
Leaf form and photosynthesis
1
1997
... 植物在进化过程中, 受到外界温度、水分、光照等环境因素的长期作用而影响其建成, 逐步形成适应环境的表型和遗传特征.即使是同一物种, 长期生长在不同的地理环境中, 也会产生不同程度的基因分化, 以适应各自不同的生态位, 从而形成特定的种源特性(
Smith et al ., 1997 ;
Barboni et al ., 2004 ;
Pigliucci, 2006 ;
胡梦瑶等, 2012 ).植物自然群体分布的环境具有明显差异.从表型来看, 同一结构由于环境分化选择作用, 不同环境条件下的群体最适表型也存在差异, 而表型的差异归根结底是由控制表型的基因型不同所导致的. ...
大叶种茶树叶片解剖结构对纬度的响应
1
2019
... 原种源地的地理位置(经纬度)、温度(
MAT 、
ART )、
MAP 、
PE 、
Alt 与叶片解剖结构指标密切相关(
表6 ), 能够很好地解释其遗传变异程度, 其中种源地气候(
MAT 、
MAP 、
PE )对蒙古莸叶片解剖结构解释程度在34.09%-81.43%之间.
季子敬等(2013) 对兴安落叶松(
Larix gmelinii )针叶解剖结构的分析研究中发现, 针叶长期适应当地环境条件后, 在解剖结构特征上也产生了具有遗传能力的变异.
赵小仙等(2015) 对蒙古沙拐枣(
Calligonum monglicum )叶片解剖结构分析研究中发现, 解剖结构的差异与经纬度、
MAT 、
MAP 的变化有极显著相关性.
白雪卡等(2018) 对不同种源蒙古莸光响应特性的研究中也同样证明, 种源地气候驱动的遗传变异对蒙古莸光合作用参数有显著的解释力, 解释程度在16.65%- 73.52%之间, 与本研究的解释力近似.本研究中, 上角质层厚度、上下表皮细胞厚度、下栅栏组织厚度、栅栏组织总厚度、叶片结构紧密度主要受
MAT 的影响, 解释程度在20.77%-71.89%之间(
表6 ); 上栅栏组织厚度、叶厚度主要受
MAP 影响, 解释程度在31.40%-45.93%之间(
表6 ).相对而言, 温度对蒙古莸叶片解剖结构参数的解释度大于水分, 这与
田丽丽等(2019) 对大叶种茶树叶片解剖结构的分析不谋而合, 其研究发现叶片生长受到温度、降水和光照因子等综合因素的影响, 叶片结构的差异则主要受到温度影响. ...
大叶种茶树叶片解剖结构对纬度的响应
1
2019
... 原种源地的地理位置(经纬度)、温度(
MAT 、
ART )、
MAP 、
PE 、
Alt 与叶片解剖结构指标密切相关(
表6 ), 能够很好地解释其遗传变异程度, 其中种源地气候(
MAT 、
MAP 、
PE )对蒙古莸叶片解剖结构解释程度在34.09%-81.43%之间.
季子敬等(2013) 对兴安落叶松(
Larix gmelinii )针叶解剖结构的分析研究中发现, 针叶长期适应当地环境条件后, 在解剖结构特征上也产生了具有遗传能力的变异.
赵小仙等(2015) 对蒙古沙拐枣(
Calligonum monglicum )叶片解剖结构分析研究中发现, 解剖结构的差异与经纬度、
MAT 、
MAP 的变化有极显著相关性.
白雪卡等(2018) 对不同种源蒙古莸光响应特性的研究中也同样证明, 种源地气候驱动的遗传变异对蒙古莸光合作用参数有显著的解释力, 解释程度在16.65%- 73.52%之间, 与本研究的解释力近似.本研究中, 上角质层厚度、上下表皮细胞厚度、下栅栏组织厚度、栅栏组织总厚度、叶片结构紧密度主要受
MAT 的影响, 解释程度在20.77%-71.89%之间(
表6 ); 上栅栏组织厚度、叶厚度主要受
MAP 影响, 解释程度在31.40%-45.93%之间(
表6 ).相对而言, 温度对蒙古莸叶片解剖结构参数的解释度大于水分, 这与
田丽丽等(2019) 对大叶种茶树叶片解剖结构的分析不谋而合, 其研究发现叶片生长受到温度、降水和光照因子等综合因素的影响, 叶片结构的差异则主要受到温度影响. ...
6种抗旱灌木叶片形态解剖学特征
1
2010
... 蒙古莸(
Caryopteris mongholica )属唇形科筋骨草亚科莸属(
Caryopteris ), 是我国干旱、半干旱地区一种耐旱、耐寒兼具观赏性的绿化灌木, 是防风固沙和水土保持的优良造林树种(
李玉俊等, 1991 ;
韩磊等, 2008 ).蒙古莸分布在东起呼伦贝尔典型草原区, 西至甘肃河西走廊西部和青海湖南部的广阔的蒙古高原上.然而, 不同的生长环境导致蒙古莸的生理特征和遗传结构均出现不同程度的变异, 这种对环境的适应性变异表现, 在叶片的解剖结构上尤为明显.
白雪卡等(2018) 对同质园栽培下8个不同种源蒙古莸光响应曲线的研究发现, 蒙古莸光合响应能力在不同种源间表现出明显不同, 光响应曲线也存在显著的差异.蒙古莸这种对环境的适应性变异在叶片的解剖结构上尤为明显, 在前人不同的研究中, 蒙古莸或为等面叶、无海绵组织分化(
符亚儒等, 2005 ;
杨小玉, 2008 ;
李爱平等, 2010 ), 或为异面叶、叶肉中含有海绵组织(
张源润等, 1999 ;
王晓江等, 2006 ;
田英等, 2010 ); 叶片厚度21.6-2 180 μm, 变异极大. ...
6种抗旱灌木叶片形态解剖学特征
1
2010
... 蒙古莸(
Caryopteris mongholica )属唇形科筋骨草亚科莸属(
Caryopteris ), 是我国干旱、半干旱地区一种耐旱、耐寒兼具观赏性的绿化灌木, 是防风固沙和水土保持的优良造林树种(
李玉俊等, 1991 ;
韩磊等, 2008 ).蒙古莸分布在东起呼伦贝尔典型草原区, 西至甘肃河西走廊西部和青海湖南部的广阔的蒙古高原上.然而, 不同的生长环境导致蒙古莸的生理特征和遗传结构均出现不同程度的变异, 这种对环境的适应性变异表现, 在叶片的解剖结构上尤为明显.
白雪卡等(2018) 对同质园栽培下8个不同种源蒙古莸光响应曲线的研究发现, 蒙古莸光合响应能力在不同种源间表现出明显不同, 光响应曲线也存在显著的差异.蒙古莸这种对环境的适应性变异在叶片的解剖结构上尤为明显, 在前人不同的研究中, 蒙古莸或为等面叶、无海绵组织分化(
符亚儒等, 2005 ;
杨小玉, 2008 ;
李爱平等, 2010 ), 或为异面叶、叶肉中含有海绵组织(
张源润等, 1999 ;
王晓江等, 2006 ;
田英等, 2010 ); 叶片厚度21.6-2 180 μm, 变异极大. ...
生态灌木蒙古莸的生物生态学特性及其经济价值评价
1
2006
... 蒙古莸(
Caryopteris mongholica )属唇形科筋骨草亚科莸属(
Caryopteris ), 是我国干旱、半干旱地区一种耐旱、耐寒兼具观赏性的绿化灌木, 是防风固沙和水土保持的优良造林树种(
李玉俊等, 1991 ;
韩磊等, 2008 ).蒙古莸分布在东起呼伦贝尔典型草原区, 西至甘肃河西走廊西部和青海湖南部的广阔的蒙古高原上.然而, 不同的生长环境导致蒙古莸的生理特征和遗传结构均出现不同程度的变异, 这种对环境的适应性变异表现, 在叶片的解剖结构上尤为明显.
白雪卡等(2018) 对同质园栽培下8个不同种源蒙古莸光响应曲线的研究发现, 蒙古莸光合响应能力在不同种源间表现出明显不同, 光响应曲线也存在显著的差异.蒙古莸这种对环境的适应性变异在叶片的解剖结构上尤为明显, 在前人不同的研究中, 蒙古莸或为等面叶、无海绵组织分化(
符亚儒等, 2005 ;
杨小玉, 2008 ;
李爱平等, 2010 ), 或为异面叶、叶肉中含有海绵组织(
张源润等, 1999 ;
王晓江等, 2006 ;
田英等, 2010 ); 叶片厚度21.6-2 180 μm, 变异极大. ...
生态灌木蒙古莸的生物生态学特性及其经济价值评价
1
2006
... 蒙古莸(
Caryopteris mongholica )属唇形科筋骨草亚科莸属(
Caryopteris ), 是我国干旱、半干旱地区一种耐旱、耐寒兼具观赏性的绿化灌木, 是防风固沙和水土保持的优良造林树种(
李玉俊等, 1991 ;
韩磊等, 2008 ).蒙古莸分布在东起呼伦贝尔典型草原区, 西至甘肃河西走廊西部和青海湖南部的广阔的蒙古高原上.然而, 不同的生长环境导致蒙古莸的生理特征和遗传结构均出现不同程度的变异, 这种对环境的适应性变异表现, 在叶片的解剖结构上尤为明显.
白雪卡等(2018) 对同质园栽培下8个不同种源蒙古莸光响应曲线的研究发现, 蒙古莸光合响应能力在不同种源间表现出明显不同, 光响应曲线也存在显著的差异.蒙古莸这种对环境的适应性变异在叶片的解剖结构上尤为明显, 在前人不同的研究中, 蒙古莸或为等面叶、无海绵组织分化(
符亚儒等, 2005 ;
杨小玉, 2008 ;
李爱平等, 2010 ), 或为异面叶、叶肉中含有海绵组织(
张源润等, 1999 ;
王晓江等, 2006 ;
田英等, 2010 ); 叶片厚度21.6-2 180 μm, 变异极大. ...
树种对土壤有机碳密度的影响: 5种温带树种同质园试验
1
2015
... 叶片作为植物暴露在生活环境中面积最大的器官, 其外部形态特征、内部解剖结构等最能体现植物对环境因子的适应性进化(
Wright et al ., 2001 ;
Chartzoulakis et al ., 2002 ;
钟悦鸣等, 2017 ;
于海燕等, 2019 ), 开展叶片解剖结构分析, 对研究植物在极端生态环境下的适应策略具有重要意义.一般而言, 植物分布范围广泛, 各自生活在温度、水分等条件不相同的自然地理环境中, 在长期对环境适应过程中, 同种植物叶片的解剖结构表现出明显的差异(
季子敬等, 2013 ;
郭婧宇, 2018 ).叶片的这种功能性状差异可能既有自身的可塑性响应也有遗传变异, 以往研究中沿自然环境梯度的采样测定或单一站点控制实验很难区分两者的效应.全缘叶绿绒蒿(
Meconopsis integrifolia )随着海拔升高, 株型变矮, 叶片组织结构紧密度增大(
刘梦颖等, 2018 ); 生长于干旱地区的长梗扁桃(
Amygdalus pedunculata )叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的长梗扁桃, 其叶片栅栏组织排列疏松, 叶厚度较薄(
郭改改等, 2013 ).这种叶片解剖结构特征的差异同时受到环境变异与自身遗传结构两个因素的影响.同质园试验即研究同一基因型的物种在不同环境下的初生与次生代谢的响应, 或不同基因型的物种在相同环境下的初生与次生代谢的响应.将多个种源的植物在同地栽培, 在一定程度上控制环境与基因型间的相互作用, 最大程度地降低立地条件所造成的影响(
王薪琪等, 2015 ;
陈嘉静等, 2019 ), 进而说明不同种源植物叶片解剖结构特征表现出的差异是由于原种源地环境差异造成的植物遗传性基因变异.这与先前关于环境和遗传因素共同影响(包括气候梯度和种源的影响)叶片解剖结构特征的研究有所不同, 这种同质园方法在局部适应性研究中具有重要意义(
de Villemereuil, 2016 ), 有助于我们更好地理解不同地理种源植物对全球变化的响应和适应机制.目前采用此法对不同种源内植物叶片解剖结构差异及其影响因素的相关分析较少. ...
树种对土壤有机碳密度的影响: 5种温带树种同质园试验
1
2015
... 叶片作为植物暴露在生活环境中面积最大的器官, 其外部形态特征、内部解剖结构等最能体现植物对环境因子的适应性进化(
Wright et al ., 2001 ;
Chartzoulakis et al ., 2002 ;
钟悦鸣等, 2017 ;
于海燕等, 2019 ), 开展叶片解剖结构分析, 对研究植物在极端生态环境下的适应策略具有重要意义.一般而言, 植物分布范围广泛, 各自生活在温度、水分等条件不相同的自然地理环境中, 在长期对环境适应过程中, 同种植物叶片的解剖结构表现出明显的差异(
季子敬等, 2013 ;
郭婧宇, 2018 ).叶片的这种功能性状差异可能既有自身的可塑性响应也有遗传变异, 以往研究中沿自然环境梯度的采样测定或单一站点控制实验很难区分两者的效应.全缘叶绿绒蒿(
Meconopsis integrifolia )随着海拔升高, 株型变矮, 叶片组织结构紧密度增大(
刘梦颖等, 2018 ); 生长于干旱地区的长梗扁桃(
Amygdalus pedunculata )叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的长梗扁桃, 其叶片栅栏组织排列疏松, 叶厚度较薄(
郭改改等, 2013 ).这种叶片解剖结构特征的差异同时受到环境变异与自身遗传结构两个因素的影响.同质园试验即研究同一基因型的物种在不同环境下的初生与次生代谢的响应, 或不同基因型的物种在相同环境下的初生与次生代谢的响应.将多个种源的植物在同地栽培, 在一定程度上控制环境与基因型间的相互作用, 最大程度地降低立地条件所造成的影响(
王薪琪等, 2015 ;
陈嘉静等, 2019 ), 进而说明不同种源植物叶片解剖结构特征表现出的差异是由于原种源地环境差异造成的植物遗传性基因变异.这与先前关于环境和遗传因素共同影响(包括气候梯度和种源的影响)叶片解剖结构特征的研究有所不同, 这种同质园方法在局部适应性研究中具有重要意义(
de Villemereuil, 2016 ), 有助于我们更好地理解不同地理种源植物对全球变化的响应和适应机制.目前采用此法对不同种源内植物叶片解剖结构差异及其影响因素的相关分析较少. ...
Strategy shifts in leaf physiology, structure and nutrient content between species of high- and low-rainfall and high- and low-nutrient habitats
1
2001
... 叶片作为植物暴露在生活环境中面积最大的器官, 其外部形态特征、内部解剖结构等最能体现植物对环境因子的适应性进化(
Wright et al ., 2001 ;
Chartzoulakis et al ., 2002 ;
钟悦鸣等, 2017 ;
于海燕等, 2019 ), 开展叶片解剖结构分析, 对研究植物在极端生态环境下的适应策略具有重要意义.一般而言, 植物分布范围广泛, 各自生活在温度、水分等条件不相同的自然地理环境中, 在长期对环境适应过程中, 同种植物叶片的解剖结构表现出明显的差异(
季子敬等, 2013 ;
郭婧宇, 2018 ).叶片的这种功能性状差异可能既有自身的可塑性响应也有遗传变异, 以往研究中沿自然环境梯度的采样测定或单一站点控制实验很难区分两者的效应.全缘叶绿绒蒿(
Meconopsis integrifolia )随着海拔升高, 株型变矮, 叶片组织结构紧密度增大(
刘梦颖等, 2018 ); 生长于干旱地区的长梗扁桃(
Amygdalus pedunculata )叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的长梗扁桃, 其叶片栅栏组织排列疏松, 叶厚度较薄(
郭改改等, 2013 ).这种叶片解剖结构特征的差异同时受到环境变异与自身遗传结构两个因素的影响.同质园试验即研究同一基因型的物种在不同环境下的初生与次生代谢的响应, 或不同基因型的物种在相同环境下的初生与次生代谢的响应.将多个种源的植物在同地栽培, 在一定程度上控制环境与基因型间的相互作用, 最大程度地降低立地条件所造成的影响(
王薪琪等, 2015 ;
陈嘉静等, 2019 ), 进而说明不同种源植物叶片解剖结构特征表现出的差异是由于原种源地环境差异造成的植物遗传性基因变异.这与先前关于环境和遗传因素共同影响(包括气候梯度和种源的影响)叶片解剖结构特征的研究有所不同, 这种同质园方法在局部适应性研究中具有重要意义(
de Villemereuil, 2016 ), 有助于我们更好地理解不同地理种源植物对全球变化的响应和适应机制.目前采用此法对不同种源内植物叶片解剖结构差异及其影响因素的相关分析较少. ...
The worldwide leaf economics spectrum
1
2004
... 蒙古莸在我国的分布区域广泛, 生长环境条件差异较大, 为满足自身正常的生长, 对环境和气候条件的适应就至关重要.植物形态结构是植物适应环境而进化出的生存策略, 由于环境分化选择作用, 植物逐步形成适应环境的表型和遗传特征, 表现出独特的种源特性; 即使将不同种源植物移植到相同环境下生长, 其叶片的解剖结构等表型特征仍存在着差异(
马小芬等, 2013 ).这种不同种源间特性的差异正是植物趋异适应的体现, 归根结底是由于控制表型的基因型不同所导致的.本研究采用同质园法, 削弱不同种源地环境对植物个体性状的影响, 将原种源气候差异驱动的遗传变异对蒙古莸叶片解剖结构的影响作为研究重点, 分析结果表明对蒙古莸解剖结构参数有显著的解释力.植物对原生境地气候条件长期适应, 已经产生了遗传结构上的差异, 不同种源蒙古莸叶片解剖结构的差异是其适应特有环境的体现; 同时, 不同解剖结构间存在着协同变化, 反映出植物对干旱环境的良好适应性, 这种适应性策略与近年来提出的叶经济谱概念吻合(
Wright et al ., 2004 ;
Pigliucci, 2006 ).研究结果可为蒙古莸优良抗逆种源选择、遗传资源保护等提供实际应用指导, 并为进一步探究全球变化背景下蒙古莸叶解剖结构对环境变化的响应机制等提供重要的理论依据.此外, 造成不同种源蒙古莸叶片解剖结构差异的因素还有很多, 包括种源地土壤、CO
2 浓度等其他环境因素, 还需在今后的研究中进一步地深入和细化. ...
5种沙生灌木叶片解剖结构与抗旱性研究
2
2008
... 蒙古莸(
Caryopteris mongholica )属唇形科筋骨草亚科莸属(
Caryopteris ), 是我国干旱、半干旱地区一种耐旱、耐寒兼具观赏性的绿化灌木, 是防风固沙和水土保持的优良造林树种(
李玉俊等, 1991 ;
韩磊等, 2008 ).蒙古莸分布在东起呼伦贝尔典型草原区, 西至甘肃河西走廊西部和青海湖南部的广阔的蒙古高原上.然而, 不同的生长环境导致蒙古莸的生理特征和遗传结构均出现不同程度的变异, 这种对环境的适应性变异表现, 在叶片的解剖结构上尤为明显.
白雪卡等(2018) 对同质园栽培下8个不同种源蒙古莸光响应曲线的研究发现, 蒙古莸光合响应能力在不同种源间表现出明显不同, 光响应曲线也存在显著的差异.蒙古莸这种对环境的适应性变异在叶片的解剖结构上尤为明显, 在前人不同的研究中, 蒙古莸或为等面叶、无海绵组织分化(
符亚儒等, 2005 ;
杨小玉, 2008 ;
李爱平等, 2010 ), 或为异面叶、叶肉中含有海绵组织(
张源润等, 1999 ;
王晓江等, 2006 ;
田英等, 2010 ); 叶片厚度21.6-2 180 μm, 变异极大. ...
... 本研究通过对同质园栽培下7个种源蒙古莸叶片解剖结构的观察, 发现7个种源地的蒙古莸叶片均为典型的等面叶, 叶片由紧密排列的上表皮、发达的栅栏组织和下表皮构成, 无海绵组织分化, 是旱生植物最常见的叶片结构(
Delzon, 2015 ), 与
贺一鸣(2018) 、
杨小玉(2008) 、
李爱平等(2010) 、
符亚儒等(2005) 对蒙古莸叶片解剖结构观察结果一致.蒙古莸叶片解剖结构参数的数量化分析表明, 上表皮细胞厚度均大于下表皮细胞厚度, 这种结构有利于气体交换(
陈旭等, 2019 ); 7个种源蒙古莸叶片的7个解剖结构参数存在显著差异(
表3 ), 其中, 叶厚度变化范围为192.34-270.30 μm, 变异系数为0.12, 与
符亚儒等(2005) 、
李爱平等(2010) 的测量结果近似. ...
5种沙生灌木叶片解剖结构与抗旱性研究
2
2008
... 蒙古莸(
Caryopteris mongholica )属唇形科筋骨草亚科莸属(
Caryopteris ), 是我国干旱、半干旱地区一种耐旱、耐寒兼具观赏性的绿化灌木, 是防风固沙和水土保持的优良造林树种(
李玉俊等, 1991 ;
韩磊等, 2008 ).蒙古莸分布在东起呼伦贝尔典型草原区, 西至甘肃河西走廊西部和青海湖南部的广阔的蒙古高原上.然而, 不同的生长环境导致蒙古莸的生理特征和遗传结构均出现不同程度的变异, 这种对环境的适应性变异表现, 在叶片的解剖结构上尤为明显.
白雪卡等(2018) 对同质园栽培下8个不同种源蒙古莸光响应曲线的研究发现, 蒙古莸光合响应能力在不同种源间表现出明显不同, 光响应曲线也存在显著的差异.蒙古莸这种对环境的适应性变异在叶片的解剖结构上尤为明显, 在前人不同的研究中, 蒙古莸或为等面叶、无海绵组织分化(
符亚儒等, 2005 ;
杨小玉, 2008 ;
李爱平等, 2010 ), 或为异面叶、叶肉中含有海绵组织(
张源润等, 1999 ;
王晓江等, 2006 ;
田英等, 2010 ); 叶片厚度21.6-2 180 μm, 变异极大. ...
... 本研究通过对同质园栽培下7个种源蒙古莸叶片解剖结构的观察, 发现7个种源地的蒙古莸叶片均为典型的等面叶, 叶片由紧密排列的上表皮、发达的栅栏组织和下表皮构成, 无海绵组织分化, 是旱生植物最常见的叶片结构(
Delzon, 2015 ), 与
贺一鸣(2018) 、
杨小玉(2008) 、
李爱平等(2010) 、
符亚儒等(2005) 对蒙古莸叶片解剖结构观察结果一致.蒙古莸叶片解剖结构参数的数量化分析表明, 上表皮细胞厚度均大于下表皮细胞厚度, 这种结构有利于气体交换(
陈旭等, 2019 ); 7个种源蒙古莸叶片的7个解剖结构参数存在显著差异(
表3 ), 其中, 叶厚度变化范围为192.34-270.30 μm, 变异系数为0.12, 与
符亚儒等(2005) 、
李爱平等(2010) 的测量结果近似. ...
文冠果不同种源叶片结构对水分胁迫的差异性响应
1
2019
... 叶片作为植物暴露在生活环境中面积最大的器官, 其外部形态特征、内部解剖结构等最能体现植物对环境因子的适应性进化(
Wright et al ., 2001 ;
Chartzoulakis et al ., 2002 ;
钟悦鸣等, 2017 ;
于海燕等, 2019 ), 开展叶片解剖结构分析, 对研究植物在极端生态环境下的适应策略具有重要意义.一般而言, 植物分布范围广泛, 各自生活在温度、水分等条件不相同的自然地理环境中, 在长期对环境适应过程中, 同种植物叶片的解剖结构表现出明显的差异(
季子敬等, 2013 ;
郭婧宇, 2018 ).叶片的这种功能性状差异可能既有自身的可塑性响应也有遗传变异, 以往研究中沿自然环境梯度的采样测定或单一站点控制实验很难区分两者的效应.全缘叶绿绒蒿(
Meconopsis integrifolia )随着海拔升高, 株型变矮, 叶片组织结构紧密度增大(
刘梦颖等, 2018 ); 生长于干旱地区的长梗扁桃(
Amygdalus pedunculata )叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的长梗扁桃, 其叶片栅栏组织排列疏松, 叶厚度较薄(
郭改改等, 2013 ).这种叶片解剖结构特征的差异同时受到环境变异与自身遗传结构两个因素的影响.同质园试验即研究同一基因型的物种在不同环境下的初生与次生代谢的响应, 或不同基因型的物种在相同环境下的初生与次生代谢的响应.将多个种源的植物在同地栽培, 在一定程度上控制环境与基因型间的相互作用, 最大程度地降低立地条件所造成的影响(
王薪琪等, 2015 ;
陈嘉静等, 2019 ), 进而说明不同种源植物叶片解剖结构特征表现出的差异是由于原种源地环境差异造成的植物遗传性基因变异.这与先前关于环境和遗传因素共同影响(包括气候梯度和种源的影响)叶片解剖结构特征的研究有所不同, 这种同质园方法在局部适应性研究中具有重要意义(
de Villemereuil, 2016 ), 有助于我们更好地理解不同地理种源植物对全球变化的响应和适应机制.目前采用此法对不同种源内植物叶片解剖结构差异及其影响因素的相关分析较少. ...
文冠果不同种源叶片结构对水分胁迫的差异性响应
1
2019
... 叶片作为植物暴露在生活环境中面积最大的器官, 其外部形态特征、内部解剖结构等最能体现植物对环境因子的适应性进化(
Wright et al ., 2001 ;
Chartzoulakis et al ., 2002 ;
钟悦鸣等, 2017 ;
于海燕等, 2019 ), 开展叶片解剖结构分析, 对研究植物在极端生态环境下的适应策略具有重要意义.一般而言, 植物分布范围广泛, 各自生活在温度、水分等条件不相同的自然地理环境中, 在长期对环境适应过程中, 同种植物叶片的解剖结构表现出明显的差异(
季子敬等, 2013 ;
郭婧宇, 2018 ).叶片的这种功能性状差异可能既有自身的可塑性响应也有遗传变异, 以往研究中沿自然环境梯度的采样测定或单一站点控制实验很难区分两者的效应.全缘叶绿绒蒿(
Meconopsis integrifolia )随着海拔升高, 株型变矮, 叶片组织结构紧密度增大(
刘梦颖等, 2018 ); 生长于干旱地区的长梗扁桃(
Amygdalus pedunculata )叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的长梗扁桃, 其叶片栅栏组织排列疏松, 叶厚度较薄(
郭改改等, 2013 ).这种叶片解剖结构特征的差异同时受到环境变异与自身遗传结构两个因素的影响.同质园试验即研究同一基因型的物种在不同环境下的初生与次生代谢的响应, 或不同基因型的物种在相同环境下的初生与次生代谢的响应.将多个种源的植物在同地栽培, 在一定程度上控制环境与基因型间的相互作用, 最大程度地降低立地条件所造成的影响(
王薪琪等, 2015 ;
陈嘉静等, 2019 ), 进而说明不同种源植物叶片解剖结构特征表现出的差异是由于原种源地环境差异造成的植物遗传性基因变异.这与先前关于环境和遗传因素共同影响(包括气候梯度和种源的影响)叶片解剖结构特征的研究有所不同, 这种同质园方法在局部适应性研究中具有重要意义(
de Villemereuil, 2016 ), 有助于我们更好地理解不同地理种源植物对全球变化的响应和适应机制.目前采用此法对不同种源内植物叶片解剖结构差异及其影响因素的相关分析较少. ...
耐旱灌木蒙古莸花的资源及保护利用
1
1999
... 蒙古莸(
Caryopteris mongholica )属唇形科筋骨草亚科莸属(
Caryopteris ), 是我国干旱、半干旱地区一种耐旱、耐寒兼具观赏性的绿化灌木, 是防风固沙和水土保持的优良造林树种(
李玉俊等, 1991 ;
韩磊等, 2008 ).蒙古莸分布在东起呼伦贝尔典型草原区, 西至甘肃河西走廊西部和青海湖南部的广阔的蒙古高原上.然而, 不同的生长环境导致蒙古莸的生理特征和遗传结构均出现不同程度的变异, 这种对环境的适应性变异表现, 在叶片的解剖结构上尤为明显.
白雪卡等(2018) 对同质园栽培下8个不同种源蒙古莸光响应曲线的研究发现, 蒙古莸光合响应能力在不同种源间表现出明显不同, 光响应曲线也存在显著的差异.蒙古莸这种对环境的适应性变异在叶片的解剖结构上尤为明显, 在前人不同的研究中, 蒙古莸或为等面叶、无海绵组织分化(
符亚儒等, 2005 ;
杨小玉, 2008 ;
李爱平等, 2010 ), 或为异面叶、叶肉中含有海绵组织(
张源润等, 1999 ;
王晓江等, 2006 ;
田英等, 2010 ); 叶片厚度21.6-2 180 μm, 变异极大. ...
耐旱灌木蒙古莸花的资源及保护利用
1
1999
... 蒙古莸(
Caryopteris mongholica )属唇形科筋骨草亚科莸属(
Caryopteris ), 是我国干旱、半干旱地区一种耐旱、耐寒兼具观赏性的绿化灌木, 是防风固沙和水土保持的优良造林树种(
李玉俊等, 1991 ;
韩磊等, 2008 ).蒙古莸分布在东起呼伦贝尔典型草原区, 西至甘肃河西走廊西部和青海湖南部的广阔的蒙古高原上.然而, 不同的生长环境导致蒙古莸的生理特征和遗传结构均出现不同程度的变异, 这种对环境的适应性变异表现, 在叶片的解剖结构上尤为明显.
白雪卡等(2018) 对同质园栽培下8个不同种源蒙古莸光响应曲线的研究发现, 蒙古莸光合响应能力在不同种源间表现出明显不同, 光响应曲线也存在显著的差异.蒙古莸这种对环境的适应性变异在叶片的解剖结构上尤为明显, 在前人不同的研究中, 蒙古莸或为等面叶、无海绵组织分化(
符亚儒等, 2005 ;
杨小玉, 2008 ;
李爱平等, 2010 ), 或为异面叶、叶肉中含有海绵组织(
张源润等, 1999 ;
王晓江等, 2006 ;
田英等, 2010 ); 叶片厚度21.6-2 180 μm, 变异极大. ...
6个种群蒙古沙拐枣同化枝解剖结构及与地理生态因子的关系
1
2015
... 原种源地的地理位置(经纬度)、温度(
MAT 、
ART )、
MAP 、
PE 、
Alt 与叶片解剖结构指标密切相关(
表6 ), 能够很好地解释其遗传变异程度, 其中种源地气候(
MAT 、
MAP 、
PE )对蒙古莸叶片解剖结构解释程度在34.09%-81.43%之间.
季子敬等(2013) 对兴安落叶松(
Larix gmelinii )针叶解剖结构的分析研究中发现, 针叶长期适应当地环境条件后, 在解剖结构特征上也产生了具有遗传能力的变异.
赵小仙等(2015) 对蒙古沙拐枣(
Calligonum monglicum )叶片解剖结构分析研究中发现, 解剖结构的差异与经纬度、
MAT 、
MAP 的变化有极显著相关性.
白雪卡等(2018) 对不同种源蒙古莸光响应特性的研究中也同样证明, 种源地气候驱动的遗传变异对蒙古莸光合作用参数有显著的解释力, 解释程度在16.65%- 73.52%之间, 与本研究的解释力近似.本研究中, 上角质层厚度、上下表皮细胞厚度、下栅栏组织厚度、栅栏组织总厚度、叶片结构紧密度主要受
MAT 的影响, 解释程度在20.77%-71.89%之间(
表6 ); 上栅栏组织厚度、叶厚度主要受
MAP 影响, 解释程度在31.40%-45.93%之间(
表6 ).相对而言, 温度对蒙古莸叶片解剖结构参数的解释度大于水分, 这与
田丽丽等(2019) 对大叶种茶树叶片解剖结构的分析不谋而合, 其研究发现叶片生长受到温度、降水和光照因子等综合因素的影响, 叶片结构的差异则主要受到温度影响. ...
6个种群蒙古沙拐枣同化枝解剖结构及与地理生态因子的关系
1
2015
... 原种源地的地理位置(经纬度)、温度(
MAT 、
ART )、
MAP 、
PE 、
Alt 与叶片解剖结构指标密切相关(
表6 ), 能够很好地解释其遗传变异程度, 其中种源地气候(
MAT 、
MAP 、
PE )对蒙古莸叶片解剖结构解释程度在34.09%-81.43%之间.
季子敬等(2013) 对兴安落叶松(
Larix gmelinii )针叶解剖结构的分析研究中发现, 针叶长期适应当地环境条件后, 在解剖结构特征上也产生了具有遗传能力的变异.
赵小仙等(2015) 对蒙古沙拐枣(
Calligonum monglicum )叶片解剖结构分析研究中发现, 解剖结构的差异与经纬度、
MAT 、
MAP 的变化有极显著相关性.
白雪卡等(2018) 对不同种源蒙古莸光响应特性的研究中也同样证明, 种源地气候驱动的遗传变异对蒙古莸光合作用参数有显著的解释力, 解释程度在16.65%- 73.52%之间, 与本研究的解释力近似.本研究中, 上角质层厚度、上下表皮细胞厚度、下栅栏组织厚度、栅栏组织总厚度、叶片结构紧密度主要受
MAT 的影响, 解释程度在20.77%-71.89%之间(
表6 ); 上栅栏组织厚度、叶厚度主要受
MAP 影响, 解释程度在31.40%-45.93%之间(
表6 ).相对而言, 温度对蒙古莸叶片解剖结构参数的解释度大于水分, 这与
田丽丽等(2019) 对大叶种茶树叶片解剖结构的分析不谋而合, 其研究发现叶片生长受到温度、降水和光照因子等综合因素的影响, 叶片结构的差异则主要受到温度影响. ...
不同生境胡杨叶片解剖特征及其适应可塑性
1
2017
... 叶片作为植物暴露在生活环境中面积最大的器官, 其外部形态特征、内部解剖结构等最能体现植物对环境因子的适应性进化(
Wright et al ., 2001 ;
Chartzoulakis et al ., 2002 ;
钟悦鸣等, 2017 ;
于海燕等, 2019 ), 开展叶片解剖结构分析, 对研究植物在极端生态环境下的适应策略具有重要意义.一般而言, 植物分布范围广泛, 各自生活在温度、水分等条件不相同的自然地理环境中, 在长期对环境适应过程中, 同种植物叶片的解剖结构表现出明显的差异(
季子敬等, 2013 ;
郭婧宇, 2018 ).叶片的这种功能性状差异可能既有自身的可塑性响应也有遗传变异, 以往研究中沿自然环境梯度的采样测定或单一站点控制实验很难区分两者的效应.全缘叶绿绒蒿(
Meconopsis integrifolia )随着海拔升高, 株型变矮, 叶片组织结构紧密度增大(
刘梦颖等, 2018 ); 生长于干旱地区的长梗扁桃(
Amygdalus pedunculata )叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的长梗扁桃, 其叶片栅栏组织排列疏松, 叶厚度较薄(
郭改改等, 2013 ).这种叶片解剖结构特征的差异同时受到环境变异与自身遗传结构两个因素的影响.同质园试验即研究同一基因型的物种在不同环境下的初生与次生代谢的响应, 或不同基因型的物种在相同环境下的初生与次生代谢的响应.将多个种源的植物在同地栽培, 在一定程度上控制环境与基因型间的相互作用, 最大程度地降低立地条件所造成的影响(
王薪琪等, 2015 ;
陈嘉静等, 2019 ), 进而说明不同种源植物叶片解剖结构特征表现出的差异是由于原种源地环境差异造成的植物遗传性基因变异.这与先前关于环境和遗传因素共同影响(包括气候梯度和种源的影响)叶片解剖结构特征的研究有所不同, 这种同质园方法在局部适应性研究中具有重要意义(
de Villemereuil, 2016 ), 有助于我们更好地理解不同地理种源植物对全球变化的响应和适应机制.目前采用此法对不同种源内植物叶片解剖结构差异及其影响因素的相关分析较少. ...
不同生境胡杨叶片解剖特征及其适应可塑性
1
2017
... 叶片作为植物暴露在生活环境中面积最大的器官, 其外部形态特征、内部解剖结构等最能体现植物对环境因子的适应性进化(
Wright et al ., 2001 ;
Chartzoulakis et al ., 2002 ;
钟悦鸣等, 2017 ;
于海燕等, 2019 ), 开展叶片解剖结构分析, 对研究植物在极端生态环境下的适应策略具有重要意义.一般而言, 植物分布范围广泛, 各自生活在温度、水分等条件不相同的自然地理环境中, 在长期对环境适应过程中, 同种植物叶片的解剖结构表现出明显的差异(
季子敬等, 2013 ;
郭婧宇, 2018 ).叶片的这种功能性状差异可能既有自身的可塑性响应也有遗传变异, 以往研究中沿自然环境梯度的采样测定或单一站点控制实验很难区分两者的效应.全缘叶绿绒蒿(
Meconopsis integrifolia )随着海拔升高, 株型变矮, 叶片组织结构紧密度增大(
刘梦颖等, 2018 ); 生长于干旱地区的长梗扁桃(
Amygdalus pedunculata )叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的长梗扁桃, 其叶片栅栏组织排列疏松, 叶厚度较薄(
郭改改等, 2013 ).这种叶片解剖结构特征的差异同时受到环境变异与自身遗传结构两个因素的影响.同质园试验即研究同一基因型的物种在不同环境下的初生与次生代谢的响应, 或不同基因型的物种在相同环境下的初生与次生代谢的响应.将多个种源的植物在同地栽培, 在一定程度上控制环境与基因型间的相互作用, 最大程度地降低立地条件所造成的影响(
王薪琪等, 2015 ;
陈嘉静等, 2019 ), 进而说明不同种源植物叶片解剖结构特征表现出的差异是由于原种源地环境差异造成的植物遗传性基因变异.这与先前关于环境和遗传因素共同影响(包括气候梯度和种源的影响)叶片解剖结构特征的研究有所不同, 这种同质园方法在局部适应性研究中具有重要意义(
de Villemereuil, 2016 ), 有助于我们更好地理解不同地理种源植物对全球变化的响应和适应机制.目前采用此法对不同种源内植物叶片解剖结构差异及其影响因素的相关分析较少. ...
 ,*北京林业大学生物科学与技术学院林木育种国家工程实验室, 北京 100083
,*北京林业大学生物科学与技术学院林木育种国家工程实验室, 北京 100083
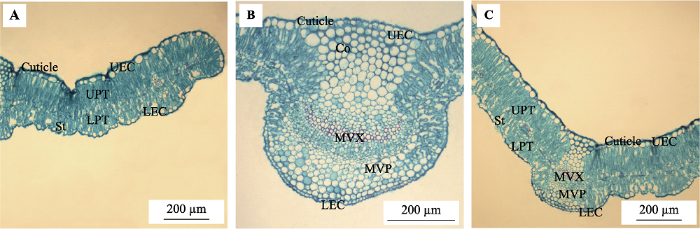 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}