Effects of extreme drought on terrestrial ecosystems: review and prospects
Gui-Yao ZHOU, Ling-Yan ZHOU, Jun-Jiong SHAO, Xu-Hui ZHOU,*and Research Center for Global Change and Ecological Forecasting, East China Normal University, Shanghai 200241, China
National Natural Science Foundation of China(31930072) National Natural Science Foundation of China(31600387) National Natural Science Foundation of China(31600352) Postdoctoral Innovation Talents Program(BX20200133)
Abstract As an important compartment of the Earth?s surface, terrestrial ecosystems act as a vital harbor for human survival and development. Climate change significantly increased the frequency, intensity and duration of drought since the beginning of the 21st century, which have marked impact on ecosystems, leading to serious restriction or even threat on the sustainable development of human beings. Therefore, developing integrative research on effects of drought on terrestrial ecosystems and assessing the associated ecological risk are impressive in global change field. This study reviewed the effects of drought on plant physiological and ecological processes, biogeochemical cycles, biodiversity, and ecosystem structure and functions in terrestrial ecosystems, and discussed current hotspot issues in this field as well as deeply analyzing the existing problems and the potential development direction. This study aims to provide some suggestions for the future observation, manipulative experiments, and modeling prediction on effects of drought on terrestrial ecosystems, and offer new insights to enhance risk assessment and management under drought. Keywords:carbon starvation;extreme drought;biodiversity;ecosystem productivity;carbon cycle
PDF (3152KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 周贵尧, 周灵燕, 邵钧炯, 周旭辉. 极端干旱对陆地生态系统的影响: 进展与展望. 植物生态学报, 2020, 44(5): 515-525. DOI: 10.17521/cjpe.2019.0317 ZHOU Gui-Yao, ZHOU Ling-Yan, SHAO Jun-Jiong, ZHOU Xu-Hui. Effects of extreme drought on terrestrial ecosystems: review and prospects. Chinese Journal of Plant Ecology, 2020, 44(5): 515-525. DOI: 10.17521/cjpe.2019.0317
中国区域与全球陆地生态系统的净初级生产力和净碳汇的时间动态都受到降水量的严重影响(Piao et al., 2009)。大范围的极端干旱事件往往是导致某一区域生态系统生物地球化学循环发生巨大改变的驱动力。例如1983年在亚马孙地区出现的极端干旱事件释放了相当于该地区生态系统需要3-4年才能吸收获得的CO2 (Potter et al., 2001)。2003年夏季在欧洲地区发生的持续高温热浪天气极大地降低了该地区生态系统的生产力, 甚至提升了全球大气CO2浓度的上升速率(Bevan et al., 2014)。目前该领域的研究大多讨论碳循环和水循环过程对极端干旱的响应。由于氮循环在调控生态系统功能中的重要作用, 近年来在区域和全球尺度上干旱对生态系统碳-氮-水耦合过程的影响格局及其调控机制开始逐渐受到关注。
区域和全球的生物地球化学循环对降水格局的响应方式除了在较大的空间尺度上反映了植物生理生态及生态系统结构与功能对降水的响应过程外, 另有其独特的复杂性。首先, 大规模大范围的持续干旱事件的发生往往由厄尔尼诺-南方涛动、太平洋北美涛动等大尺度的气候事件所驱动, 因而这些干旱事件对生物地球化学循环的影响不仅反映了局部的气候效应, 更与全球气候系统关系紧密(Piao et al., 2009)。其次, 越来越频发的干旱事件还能够增加诸如森林火灾、生态系统病虫害等负面事件的发生强度和频度(Schlesinger et al., 2016)。这些灾害事件本身难以有效控制的特性使得它们的波及范围较广, 从而对区域甚至全球的生物地球化学循环造成极其深远的影响(Li et al., 2014)。第三, 与植物个体和生态系统尺度的研究不同, 区域和全球尺度的研究无法开展统一降水受控实验, 因此很大程度上依赖于通过整合多尺度多来源的观测数据模拟和预测生物地球化学循环对降水格局的响应。
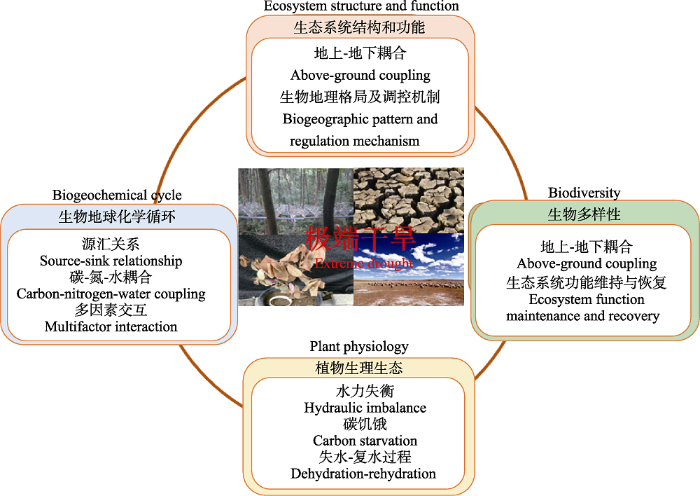
3 极端干旱对陆地生态系统结构和功能的影响
极端干旱对生态系统功能的影响主要表现在生产力、养分含量、凋落物分解等方面(Ciais et al., 2005; Zhou et al., 2009)。这种影响作用可以反映在个体、群落及生态系统等尺度上。近年来, 国内外在不同尺度上围绕极端干旱对生态系统结构和功能的影响开展了一系列研究(Piao et al., 2009; Beier et al., 2012)。例如, 华东师范大学于2013年7月依托浙江天童森林生态系统国家野外科学观测研究站, 建立了大型森林生态系统干旱实验平台, 着重探讨极端干旱对森林生态系统结构和功能, 特别是地下碳过程影响的研究, 目前已取得系列研究成果(聂园园等, 2017; Bu et al., 2018)。研究发现极端干旱显著降低亚热带森林生态系统微生物和凋落物的生物量, 同时导致微生物优势群落从细菌向真菌转移, 进而降低土壤有机碳的分解(聂园园等, 2017)。与此同时, 我们在北美大草原上的研究发现, 根系生物量和土壤碳含量与平均年降水量之间没有显著的相关性, 但土壤呼吸和茎生物量和年降水量之间存在显著的正相关关系, 而地上立枯物和表层凋落物生产量随着降水量的增加呈现先增加后降低的非线性变化趋势(Zhou et al., 2009)。在内蒙古典型草原生态系统中也发现, 年降水量或季节性降水量与植被生产力呈现线性关系(Bai et al., 2004), 但是在更大尺度上也可能会出现非线性关系(Luo et al., 2017)。陆地生态系统功能对干旱响应的敏感性差异主要受到植物生长潜力、物种组成以及资源可利用性等因素的共同影响(Beier et al., 2012)。在水限制区域, 降水的增加会通过解除水分对微生物和植物生长的压力, 进而刺激土壤碳的排放过程; 反之, 干旱可以通过降低土壤水分, 增加含水量较高区域土壤的通透性, 进而刺激土壤呼吸的排放(Knapp et al., 2015)。此外, 由于不同物种的光合作用策略以及对水分响应敏感性的差异, 干旱可能还会引起群落结构的改变, 如C3和C4优势物种相对丰度的变化(龚春梅等, 2009; Maestre et al., 2016)。
在生态系统尺度上, 极端干旱对生态系统结构与功能的影响受到了广泛关注。极端干旱可能通过改变不同大小树木的密度, 或是改变物种间的相互作用来影响森林生态系统的群落结构与生物多样性(Zhao & Running, 2010; Elmendorf et al., 2015; Mcintyre et al., 2015)。植物群落结构及多样性的改变, 以及干旱造成的水分胁迫都能够极大地影响森林生产力(Maestre et al., 2016), 进而影响其生态系统服务功能。然而, 当前关于极端干旱对森林生态系统结构与功能的影响主要集中于地上生产力的研究, 而对地下过程干旱敏感性的研究较少。一般而言, 在极端干旱背景下, 生态系统土壤的异养呼吸及碳周转速率会下降, 可能与微生物活性的下降有关(Zhou et al., 2019a)。但从生态系统整体角度而言, 关于地上与地下生产力对极端干旱的响应是否具有同步性和对称性, 地上-地下耦合过程对极端干旱的反馈调节机制等, 仍是亟待系统深入研究的热点难点问题。
Fig. 4Biodiversity under different precipitation temperateness conditions. Photos from A to D were taken by YANG Jing, ZHOU Gui-Yao, CHEN Zhi-Rong and LIU Hui-Ying, respectively.
AndereggWRL, SchwalmC, BiondiF, CamareroJJ, KochG, LitvakM, OgleK, ShawJD, ShevliakovaE, WilliamsAP, WolfA, ZiacoE, PacalaS (2015). Pervasive drought legacies in forest ecosystems and their implications for carbon cycle models Science, 349, 528-532. [本文引用: 1]
BaiYF, HanXG, WuJG, ChenZZ, LiLH (2004). Ecosystem stability and compensatory effects in the Inner Mongolia grassland Nature, 431, 181-184. [本文引用: 1]
BardgettR, van der PuttenW (2014). Belowground biodiversity and ecosystem functioning Nature, 515, 505-511. [本文引用: 4]
BeierC, BeierkuhnleinC, WohlgemuthT, PenuelasJ, EmmettB, K?rnerC, BoeckH, ChristensenJ, LeuzingerS, JanssensA, HansenK (2012). Precipitation manipulation experiments—Challenges and recommendations for the future Ecology Letters, 15, 899-911. [本文引用: 2]
BevanS, LosS, NorthP (2014). Response of vegetation to the 2003 European drought was mitigated by height Biogeosciences, 11, 2897-2908. DOI:10.5194/bg-11-2897-2014URL [本文引用: 1]
ElmendorfSC, HenryGH, HollisterRD, HollisterRD, FosaaAM, GouldWA, HermanutzL, HofgaardA, JónsdóttirIS, JorgensonJC, LévesqueE, MagnussonB, MolauU, Myers-SmithIH, OberbauerSF, RixenC, TweedieCE, WalkerMD (2015). Experiment, monitoring, and gradient methods used to infer climate change effects on plant communities yield consistent patterns Proceedings of the National Academy of Sciences of United States of America, 112, 448-452. [本文引用: 2]
FangJY, ZhuJL, ShiY (2018). The responses of ecosystems to global warming Chinese Science Bulletin, 63, 136-140. [本文引用: 1]
FuchsluegerL, BahnM, FritzK, HasibederR, RichterA (2014). Experimental drought reduces the transfer of recently fixed plant carbon to soil microbes and alters the bacterial community composition in a mountain meadow New Phytologist, 201, 916-927. [本文引用: 1]
FuentealbaMP, ZhangJ, KenworthyK, EricksonJ, KruseJ, TrenholmL (2015). Transpiration responses of warm- season turfgrass in relation to progressive soil drying Scientia Horticulturae, 198, 249-253. [本文引用: 1]
GalbraithD, LevyPE, SitchS, HuntingfordC, CoxP, WilliamsM, MeirP (2010). Multiple mechanisms of Amazonian forest biomass losses in three dynamic global vegetation models under climate change New Phytologist, 187, 647-665. [本文引用: 1]
Garcia-FornerN, BielC, SavéR, Martínez-VilaltaJ (2017). Isohydric species are not necessarily more carbon limited than anisohydric species during drought Tree Physiology, 37, 441-455. URLPMID:27885172 [本文引用: 1]
GongCM, NingPB, WangGX, LiangZS (2009). A review of adaptable variations and evolution of photo synthetic carbon assimilation pathway in C3 and C4 plants Chinese Journal of Plant Ecology, 33, 206-221. [本文引用: 1]
HuXP, WangSG, XuPP, ShangKZ (2014). Analysis on causes of continuous drought in Southwest China during 2009-2013 Meteorological Monthly, 40, 1216-1229. [本文引用: 1]
HuZM, ShiH, ChengKL, WangYP, PiaoSL, LiY, ZhangL, XiaJY, ZhouL, YuanWP, RunningS, LiLH, HaoYB, HeNP, YuQ, YuGR (2018). Joint structural and physiological control on the interannual variation in productivity in a temperate grassland: a data-model comparison Global Change Biology, 24, 2965-2979. [本文引用: 1]
IPCC (Intergovernmental Panel on Climate Change) (2013). Climate Change 2013: the Physical Science Basis. Cambridge University Press, Cambridge, UK. [本文引用: 1]
JacksonRB, CanadellJ, EhleringerJR, MooneyHA, SalaOE, SchulzeED (1996). A global analysis of root distributions for terrestrial biomes Oecologia, 108, 389-411. [本文引用: 1]
KleinT (2014). The variability of stomatal sensitivity to leaf water potential across tree species indicates a continuum between isohydric and anisohydric behaviours Functional Ecology, 28, 1313-1320. [本文引用: 1]
KnappAK, HooverDL, WilcoxKR, AvolioML, KoernerSE, La PierreKJ, LoikME, LuoYQ, SalaOE, SmithMD (2015). Characterizing differences in precipitation regimes of extreme wet and dry years: implications for climate change experiments Global Change Biology, 21, 2624-2633. [本文引用: 1]
LiF, Bond-LambertyB, LevisS (2014). Quantifying the role of fire in the Earth system—Part 2: Impact on the net carbon balance of global terrestrial ecosystems for the 20th century Biogeosciences, 11, 1345-1360. [本文引用: 1]
LuoYQ, JiangLF, NiuSL, ZhouXH (2017). Nonlinear responses of land ecosystems to variation in precipitation New Phytologist, 214, 5-7. [本文引用: 1]
MaZQ, GuoDL, XuXL, LuMZ, BardgettRD, EissenstatDM, McCormackML, HedinLO (2018). Evolutionary history resolves global organization of root functional traits Nature, 555, 94-97. [本文引用: 1]
MaestreFT, EldridgeDJ, SoliveresS, KéfiS, Delgado- BaquerizoMF, BowkerMA, García-PalaciosPG, GaitánJ, GallardoA, LázaroR, BerdugoM (2016). Structure and functioning of dryland ecosystems in a changing world Annual Review of Ecology Evolution and Systematics, 47, 215-237. [本文引用: 4]
MahfoufJF, CiretC, DucharneA, IrannejadP, NoilhanaJ, ShaoY, ThorntonP, XueY, YangZL (1996). Analysis of transpiration results from the RICE and PILPS workshop Global and Planetary Change, 13, 73-88. [本文引用: 1]
Martínez-VilaltaJM, PoyatosR, AguadéD, RetanaJ, MencucciniM (2014). A new look at water transport regulation in plants New Phytologist, 204, 105-115. [本文引用: 1]
McDowellNG, SevantoS (2010). The mechanisms of carbon starvation: How, when, or does it even occur at all? New Phytologist, 186, 264-266. URLPMID:20409181 [本文引用: 1]
McDowellN, PockmanWT, AllenCD, BreshearsDD, CobbN, KolbT, PlautJ, SperryJ, WestA, WilliamsDG, YepezEA (2008). Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytologist, 178, 719-739. [本文引用: 1]
McintyrePJ, ThorneJH, DolancCR, FlintAL, KellyM, AckerlyDD (2015). Twentieth-century shifts in forest structure in California: denser forests, smaller trees, and increased dominance of oaks Proceedings of the National Academy of Sciences of the United States of America, 112, 1458-1463. [本文引用: 2]
MelilloJM, ButlerS, JohnsonJ, MohanJ, SteudlerP, LuxH, BurrowsE, BowlesF, SmithR, ScottL, VarioC, HillT, BurtonA, ZhouYM, TangJ (2011). Soil warming, carbon- nitrogen interactions, and forest carbon budgets Proceedings of the National Academy of Sciences of the United States of America, 108, 9508-9512. [本文引用: 1]
MencucciniM, MinunnoF, SalmonY, Martínez-VilaltaJ, H?ltt?T (2015). Coordination of physiological traits involved in drought-induced mortality of woody plants New Phytologist, 208, 396-409. [本文引用: 2]
MiyashitaK, TanakamaruS, MaitaniT, KimuraK (2005). Recovery responses of photosynthesis, transpiration, and stomatal conductance in kidney bean following drought stress Environmental and Experimental Botany, 53, 205-214. [本文引用: 1]
NieYY, ZhouGY, ShaoJJ, ZhouLY, LiuRQ, ZhaiDP, ZhouXH (2017). Effects of simulating drought on soil microbial biomass and community structure in subtropical forest Journal of Fudan University(Natural Science), 56, 97-105. [本文引用: 3]
Pérez-RamosIM, VolaireF, FattetM, BlanchardA, RoumetC (2013). Tradeoffs between functional strategies for resource use and drought-survival in Mediterranean rangeland species Environmental and Experimental Botany, 87, 126-136. [本文引用: 1]
PiaoSL, FangJY, CiaisP, PeylinP, HuangY, SitchS (2009). The carbon balance of terrestrial ecosystems in China Nature, 458, 1009-1013. URLPMID:19396142 [本文引用: 6]
PoschS, BennettLT (2009). Photosynthesis, photochemistry and antioxidative defence in response to two drought severities and with re-watering in Allocasuarina luehmannii Plant Biology, 11, 83-93. URLPMID:19121117 [本文引用: 2]
PotterC, KloosterS, CarvalhoCR, GenoveseVB, TorregrosaA, DunganJ, BoboM, CoughlanJ (2001). Modeling seasonal and interannual variability in ecosystem carbon cycling for the Brazilian Amazon region Journal of Geophysical Research, 106, 10423-10446. [本文引用: 1]
RilligMC, RyoM, LehmannA, Aguilar-TriguerosCA, BuchertS, WulfA, IwasakiA, RoyJ, YangGW (2019). The role of multiple global change factors in driving soil functions and microbial biodiversity Science, 366, 886-890 [本文引用: 1]
RowlandL, Da CostaACL, GalbraithDR, OliveiraRS, BinksOJ, OliveiraAAR, PullenAM, DoughtyPCE, MetcalfeDB, VasconcelosSS, FerreiraLV, MalhiY, GraceJ, MencucciniM, MeirP (2015). Death from drought in tropical forests is triggered by hydraulics not carbon starvation Nature, 528, 119-122. [本文引用: 1]
SchlesingerWH, DietzeMC, JacksonRB, PhillipsRP, RhoadesCC, RustadLE, VoseJM (2016). Forest biogeochemistry in response to drought Global Change Biology, 22, 2318-2328. [本文引用: 1]
SperryJS, LoveDM (2015). What plant hydraulics can tell us about responses to climate-change droughts New Phytologist, 207, 14-27. DOI:10.1111/nph.13354URLPMID:25773898 [本文引用: 3] Climate change exposes vegetation to unusual drought, causing declines in productivity and increased mortality. Drought responses are hard to anticipate because canopy transpiration and diffusive conductance (G) respond to drying soil and vapor pressure deficit (D) in complex ways. A growing database of hydraulic traits, combined with a parsimonious theory of tree water transport and its regulation, may improve predictions of at-risk vegetation. The theory uses the physics of flow through soil and xylem to quantify how canopy water supply declines with drought and ceases by hydraulic failure. This transpiration 'supply function' is used to predict a water 'loss function' by assuming that stomatal regulation exploits transport capacity while avoiding failure. Supply-loss theory incorporates root distribution, hydraulic redistribution, cavitation vulnerability, and cavitation reversal. The theory efficiently defines stomatal responses to D, drying soil, and hydraulic vulnerability. Driving the theory with climate predicts drought-induced loss of plant hydraulic conductance (k), canopy G, carbon assimilation, and productivity. Data lead to the 'chronic stress hypothesis' wherein > 60% loss of k increases mortality by multiple mechanisms. Supply-loss theory predicts the climatic conditions that push vegetation over this risk threshold. The theory's simplicity and predictive power encourage testing and application in large-scale modeling.
TaylorPG, ClevelandCC, WiederWR, SullivanBW, DoughtyCE, DobrowskiSZ, TownsendAR (2017). Temperature and rainfall interact to control carbon cycling in tropical forests Ecology Letters, 20, 779-788. URLPMID:28414883 [本文引用: 2]
TronS, PeronaP, GorlaL, SchwarzM, LaioF, RidolfiL (2015). The signature of randomness in riparian plant root distributions Geophysical Research Letters, 42, 7098-7106. [本文引用: 1]
WarrenJM, HansonPJ, IversenCM, KumarJ, WalkerAP, WullschlegerSD (2015). Root structural and functional dynamics in terrestrial biosphere models—Evaluation and recommendations New Phytologist, 205, 59-78. URLPMID:25263989 [本文引用: 2]
XuW, MaZY, JinX, HeJS (2016). Biodiversity and ecosystem multifunctionality: advances and perspectives Biodiversity Science, 24, 55-71. [本文引用: 2]
XuZZ, ZhouGS, ShimizuH (2010). Plant responses to drought and rewatering Plant Signaling and Behavior, 5, 649-654. URLPMID:20404516 [本文引用: 1]
YanY, XiaoF, DuY, LingF, LiXD, LiYZ (2012). Monitoring droughts in the five provinces along the middle-lower reaches of the Yangtze River during spring/summer 2011 using AVCI Resources and Environment in the Yangtze Basin, 21, 1154-1159. [本文引用: 1]
YinJJ, BauerleTL (2017). A global analysis of plant recovery performance from water stress Oikos, 126, 1377-1388. [本文引用: 1]
YuanWP, CaiWW, ChenY, LiuSG, DongWJ, ZhangHC, YuGR, ChenZQ, HeHL, GuoWD, LiuD, LiuSM, XiangWH, XieZH, ZhaoZH, ZhouGM (2016). Severe summer heatwave and drought strongly reduced carbon uptake in Southern China Scientific Reports, 6, 18813. DOI: 10.1038/srep18813. DOI:10.1038/srep18813URLPMID:26739761 [本文引用: 1] Increasing heatwave and drought events can potentially alter the carbon cycle. Few studies have investigated the impacts of hundred-year return heatwaves and droughts, as those events are rare. In the summer of 2013, southern China experienced its strongest drought and heatwave on record for the past 113 years. We show that the record-breaking heatwave and drought lasted two months (from July to August), significantly reduced the satellite-based vegetation index and gross primary production, substantially altered the regional carbon cycle, and produced the largest negative crop yield anomaly since 1960. The event resulted in a net reduction of 101.54 Tg C in carbon sequestration in the region during these two months, which was 39-53% of the annual net carbon sink of China's terrestrial ecosystems (190-260 Tg C yr(-1)). Moreover, model experiments showed that heatwaves and droughts consistently decreased ecosystem vegetation primary production but had opposite impacts on ecosystem respiration (TER), with increased TER by 6.78 +/- 2.15% and decreased TER by 15.34 +/- 3.57% assuming only changed temperature and precipitation, respectively. In light of increasing frequency and severity of future heatwaves and droughts, our study highlights the importance of accounting for the impacts of heatwaves and droughts in assessing the carbon sequestration in terrestrial ecosystems.
ZhangRH, ZhengYJ, MaGS, ZhangXH, LuHD, ShiJT, XueJQ (2011). Effects of drought stress on photosynthetic traits and protective enzyme activity in maize seeding Acta Ecologica Sinica, 31, 1303-1311. [本文引用: 1]
ZhaoMS, RunningSW (2010). Drought-induced reduction in global terrestrial net primary production from 2000 through 2009 Science, 329, 940-943. URLPMID:20724633 [本文引用: 2]
ZhouGY, LuoQ, ChenYJ, HuJQ, HeM, GaoJ, ZhouLY, LiuHY, ZhouXH (2019a). Interactive effects of grazing and global change factors on soil and ecosystems respiration in grassland ecosystems Journal of Applied Ecology, 56, 2007-2019. [本文引用: 3]
ZhouGY, LuoQ, ChenYJ, HeM, ZhouLY, FrankD, HeYH, FuYL, ZhangBC, ZhouXH (2019b). Effects of livestock grazing on grassland carbon storage and release override impacts associated with global climate change Global Change Biology, 25, 1119-1132. [本文引用: 2]
ZhouGY, ZhouXH, HeYH, ShaoJJ, HuZH, LiuRQ, ZhouHM, BaiSH (2017). Grazing intensity significantly affects belowground carbon and nitrogen cycling in grassland ecosystems: a meta-analysis Global Change Biology, 23, 1167-1179. DOI:10.1111/gcb.13431URLPMID:27416555 [本文引用: 1] Livestock grazing activities potentially alter ecosystem carbon (C) and nitrogen (N) cycles in grassland ecosystems. Despite the fact that numerous individual studies and a few meta-analyses had been conducted, how grazing, especially its intensity, affects belowground C and N cycling in grasslands remains unclear. In this study, we performed a comprehensive meta-analysis of 115 published studies to examine the responses of 19 variables associated with belowground C and N cycling to livestock grazing in global grasslands. Our results showed that, on average, grazing significantly decreased belowground C and N pools in grassland ecosystems, with the largest decreases in microbial biomass C and N (21.62% and 24.40%, respectively). In contrast, belowground fluxes, including soil respiration, soil net N mineralization and soil N nitrification increased by 4.25%, 34.67% and 25.87%, respectively, in grazed grasslands compared to ungrazed ones. More importantly, grazing intensity significantly affected the magnitude (even direction) of changes in the majority of the assessed belowground C and N pools and fluxes, and C : N ratio as well as soil moisture. Specifically,light grazing contributed to soil C and N sequestration whereas moderate and heavy grazing significantly increased C and N losses. In addition, soil depth, livestock type and climatic conditions influenced the responses of selected variables to livestock grazing to some degree. Our findings highlight the importance of the effects of grazing intensity on belowground C and N cycling, which may need to be incorporated into regional and global models for predicting effects of human disturbance on global grasslands and assessing the climate-biosphere feedbacks.
ZhouGY, ZhouXH, NieYY, BaiSH, ZhouLY, ShaoJJ, ChengWS, WangJ, HuFQ, FuYL (2018). Drought-induced changes in root biomass largely result from altered root morphological traits: evidence from a synthesis of global field trials Plant, Cell & Environment, 41, 2589-2599. [本文引用: 3]
ZhouSG, DuursmaRA, MedlynBE, KellyJWE, PrenticeIC (2013). How should we model plant responses to drought? An analysis of stomatal and non-stomatal responses to water stress Agricultural and Forest Meteorology, 182, 204-214. [本文引用: 1]
ZhouXH, TalleyM, LuoYQ (2009). Biomass, litter, and soil respiration along a precipitation gradient in southern Great Plains Ecosystems, 12, 1369-1380. [本文引用: 2]
Pervasive drought legacies in forest ecosystems and their implications for carbon cycle models 1 2015
... 极端干旱直接影响到植物的生长和发育, 甚至导致其死亡.当前关于干旱对植物生理生态的影响, 主要在于探讨碳水过程对干旱的响应, 其中树木致死的内在机制是当前关注的核心和热点问题.“水力失衡”假说认为, 干旱导致树木死亡可能来源于木质部导管栓塞引起的“土壤-植物系统”间的水力传输过程失衡; “碳饥饿”假说则认为, 干旱诱发气孔关闭后导致光合作用减弱, 使得的光合产物不足以支撑植物的正常生理代谢过程, 导致植物死亡(图2) (McDowell et al., 2008).近期的研究证据多数倾向于水力失衡是干旱致死的主因(Anderegg et al., 2015; Rowland et al., 2015), 但是由于非结构性碳水化合物在植物生长和防御等生理生态过程中所扮演的复杂角色及其多变的时空动态, 目前关于植物非结构性碳水化合物富足的情况下是否实际上发生了碳饥饿事件, 尚未有明确的定论(McDowell & Sevanto, 2010).与此同时, 不同植物气孔调节叶片水势的策略上的差异也使碳饥饿现象在不同物种中发生的可能性有很大差异(Martínez-Vilalta et al., 2014; Garcia-Forner et al., 2017). ...
Ecosystem stability and compensatory effects in the Inner Mongolia grassland 1 2004
... 极端干旱对生态系统功能的影响主要表现在生产力、养分含量、凋落物分解等方面(Ciais et al., 2005; Zhou et al., 2009).这种影响作用可以反映在个体、群落及生态系统等尺度上.近年来, 国内外在不同尺度上围绕极端干旱对生态系统结构和功能的影响开展了一系列研究(Piao et al., 2009; Beier et al., 2012).例如, 华东师范大学于2013年7月依托浙江天童森林生态系统国家野外科学观测研究站, 建立了大型森林生态系统干旱实验平台, 着重探讨极端干旱对森林生态系统结构和功能, 特别是地下碳过程影响的研究, 目前已取得系列研究成果(聂园园等, 2017; Bu et al., 2018).研究发现极端干旱显著降低亚热带森林生态系统微生物和凋落物的生物量, 同时导致微生物优势群落从细菌向真菌转移, 进而降低土壤有机碳的分解(聂园园等, 2017).与此同时, 我们在北美大草原上的研究发现, 根系生物量和土壤碳含量与平均年降水量之间没有显著的相关性, 但土壤呼吸和茎生物量和年降水量之间存在显著的正相关关系, 而地上立枯物和表层凋落物生产量随着降水量的增加呈现先增加后降低的非线性变化趋势(Zhou et al., 2009).在内蒙古典型草原生态系统中也发现, 年降水量或季节性降水量与植被生产力呈现线性关系(Bai et al., 2004), 但是在更大尺度上也可能会出现非线性关系(Luo et al., 2017).陆地生态系统功能对干旱响应的敏感性差异主要受到植物生长潜力、物种组成以及资源可利用性等因素的共同影响(Beier et al., 2012).在水限制区域, 降水的增加会通过解除水分对微生物和植物生长的压力, 进而刺激土壤碳的排放过程; 反之, 干旱可以通过降低土壤水分, 增加含水量较高区域土壤的通透性, 进而刺激土壤呼吸的排放(Knapp et al., 2015).此外, 由于不同物种的光合作用策略以及对水分响应敏感性的差异, 干旱可能还会引起群落结构的改变, 如C3和C4优势物种相对丰度的变化(龚春梅等, 2009; Maestre et al., 2016). ...
Belowground biodiversity and ecosystem functioning 4 2014
... 极端干旱主要通过调控生态系统中物质循环和能量流通, 以及改变物种间的相互作用来引起生物多样性的改变(Elmendorf et al., 2015).在不同的水热条件下, 植物之间相互作用关系, 养分的生物地球化学循环过程等存在差异, 从而驱动生物多样性分布的空间异质性(图4)(Bardgett & van der Putten, 2014; Taylor et al., 2017).干旱通过对植物的生长、繁殖和物候的调节, 进而改变群落内物种间相互作用来影响和调控地上生物多样性(Zhao & Running, 2010).与此同时, 干旱还可以通过改变生态系统物质循环和养分流通影响地下过程, 如对微生物(细菌、真菌、放线菌、古菌等)多样性、土壤动物(线虫、蚯蚓等)多样性产生重要影响, 并进一步调控地上-地下生物多样性之间的关系(Bardgett & van der Putten, 2014; Mcintyre et al., 2015).但是, 相较于地上多样性而言, 由于测定手段有限以及地下过程的复杂性, 当前人们在区域和全球尺度上就干旱对地下生态系统生物多样性的影响, 以及地上-地下生物多样性之间的协同进化关系的认知尚存争议(徐炜等, 2016).这种争议很大程度来源于地上、地下生物多样性对干旱响应的机制不同, 以及与植物-土壤等的复杂互馈过程(Bardgett & van der Putten, 2014).因此, 优化当前测定分析技术以及开展更大尺度、有针对性的研究, 是解决这一问题的有效途径. ...
... ).与此同时, 干旱还可以通过改变生态系统物质循环和养分流通影响地下过程, 如对微生物(细菌、真菌、放线菌、古菌等)多样性、土壤动物(线虫、蚯蚓等)多样性产生重要影响, 并进一步调控地上-地下生物多样性之间的关系(Bardgett & van der Putten, 2014; Mcintyre et al., 2015).但是, 相较于地上多样性而言, 由于测定手段有限以及地下过程的复杂性, 当前人们在区域和全球尺度上就干旱对地下生态系统生物多样性的影响, 以及地上-地下生物多样性之间的协同进化关系的认知尚存争议(徐炜等, 2016).这种争议很大程度来源于地上、地下生物多样性对干旱响应的机制不同, 以及与植物-土壤等的复杂互馈过程(Bardgett & van der Putten, 2014).因此, 优化当前测定分析技术以及开展更大尺度、有针对性的研究, 是解决这一问题的有效途径. ...
... ).这种争议很大程度来源于地上、地下生物多样性对干旱响应的机制不同, 以及与植物-土壤等的复杂互馈过程(Bardgett & van der Putten, 2014).因此, 优化当前测定分析技术以及开展更大尺度、有针对性的研究, 是解决这一问题的有效途径. ...
... 生物多样性如何调控生态系统功能仍然是当前生态学家研究的重点和热点之一(徐炜等, 2016; Eisenhauer et al., 2018).过去关于干旱背景下生物多样性对生态系统功能影响的研究多集中在站点尺度, 但由于土壤养分、植被等生物和非生物因素对干旱响应的差异(Maestre et al., 2016), 对区域甚至更大尺度上生物多样性的影响及相关驱动机制仍知之甚少.与此同时, 干旱可能会导致地上地下生产力下降、养分循环失衡, 进而改变生态系统的多功能性(Eisenhauer et al., 2018).但是这种变化是否会带来地上地下生物多样性和生态系统关系的同步改变以及相关驱动机制的变化, 当前对其的认识仍不清楚(Maestre et al., 2016).这些知识盲区将在一定程度上导致未来模型在精确模拟和尺度推绎过程中存在很大的不确定性.因此, 未来开展更大尺度的干旱对生物多样性和生态系统功能关系影响的样带实验, 进一步修正和完善相关理论基础, 减少模型结构自身带来的不确定性, 最大程度地改进模型关于干旱对生物多样性影响预测的准确性, 是解决这一瓶颈问题的重要和关键手段(Bardgett & van der Putten, 2014; Eisenhauer et al., 2018). ...
Precipitation manipulation experiments—Challenges and recommendations for the future 2 2012
... 极端干旱对生态系统功能的影响主要表现在生产力、养分含量、凋落物分解等方面(Ciais et al., 2005; Zhou et al., 2009).这种影响作用可以反映在个体、群落及生态系统等尺度上.近年来, 国内外在不同尺度上围绕极端干旱对生态系统结构和功能的影响开展了一系列研究(Piao et al., 2009; Beier et al., 2012).例如, 华东师范大学于2013年7月依托浙江天童森林生态系统国家野外科学观测研究站, 建立了大型森林生态系统干旱实验平台, 着重探讨极端干旱对森林生态系统结构和功能, 特别是地下碳过程影响的研究, 目前已取得系列研究成果(聂园园等, 2017; Bu et al., 2018).研究发现极端干旱显著降低亚热带森林生态系统微生物和凋落物的生物量, 同时导致微生物优势群落从细菌向真菌转移, 进而降低土壤有机碳的分解(聂园园等, 2017).与此同时, 我们在北美大草原上的研究发现, 根系生物量和土壤碳含量与平均年降水量之间没有显著的相关性, 但土壤呼吸和茎生物量和年降水量之间存在显著的正相关关系, 而地上立枯物和表层凋落物生产量随着降水量的增加呈现先增加后降低的非线性变化趋势(Zhou et al., 2009).在内蒙古典型草原生态系统中也发现, 年降水量或季节性降水量与植被生产力呈现线性关系(Bai et al., 2004), 但是在更大尺度上也可能会出现非线性关系(Luo et al., 2017).陆地生态系统功能对干旱响应的敏感性差异主要受到植物生长潜力、物种组成以及资源可利用性等因素的共同影响(Beier et al., 2012).在水限制区域, 降水的增加会通过解除水分对微生物和植物生长的压力, 进而刺激土壤碳的排放过程; 反之, 干旱可以通过降低土壤水分, 增加含水量较高区域土壤的通透性, 进而刺激土壤呼吸的排放(Knapp et al., 2015).此外, 由于不同物种的光合作用策略以及对水分响应敏感性的差异, 干旱可能还会引起群落结构的改变, 如C3和C4优势物种相对丰度的变化(龚春梅等, 2009; Maestre et al., 2016). ...
... ).陆地生态系统功能对干旱响应的敏感性差异主要受到植物生长潜力、物种组成以及资源可利用性等因素的共同影响(Beier et al., 2012).在水限制区域, 降水的增加会通过解除水分对微生物和植物生长的压力, 进而刺激土壤碳的排放过程; 反之, 干旱可以通过降低土壤水分, 增加含水量较高区域土壤的通透性, 进而刺激土壤呼吸的排放(Knapp et al., 2015).此外, 由于不同物种的光合作用策略以及对水分响应敏感性的差异, 干旱可能还会引起群落结构的改变, 如C3和C4优势物种相对丰度的变化(龚春梅等, 2009; Maestre et al., 2016). ...
Response of vegetation to the 2003 European drought was mitigated by height 1 2014
... 中国区域与全球陆地生态系统的净初级生产力和净碳汇的时间动态都受到降水量的严重影响(Piao et al., 2009).大范围的极端干旱事件往往是导致某一区域生态系统生物地球化学循环发生巨大改变的驱动力.例如1983年在亚马孙地区出现的极端干旱事件释放了相当于该地区生态系统需要3-4年才能吸收获得的CO2 (Potter et al., 2001).2003年夏季在欧洲地区发生的持续高温热浪天气极大地降低了该地区生态系统的生产力, 甚至提升了全球大气CO2浓度的上升速率(Bevan et al., 2014).目前该领域的研究大多讨论碳循环和水循环过程对极端干旱的响应.由于氮循环在调控生态系统功能中的重要作用, 近年来在区域和全球尺度上干旱对生态系统碳-氮-水耦合过程的影响格局及其调控机制开始逐渐受到关注. ...
Globally rising soil heterotrophic respiration over recent decades 1 2018
... 过去大量模型和野外控制实验均表明干旱事件会通过改变“植物-土壤-微生物”系统之间的生物地球化学循环过程, 直接或间接地调控生态系统养分(如碳、氮、磷等)循环过程(Piao et al., 2009; 聂园园等, 2017).但是由于实验观测和模型模拟两种方法的目的不完全一致, 使得大量关于生态系统功能的实验数据很难被模型直接采用, 而模型的模拟结果也很少作为产生科学假说的工具来有效指导实验研究(Piao et al., 2009; Taylor et al., 2017).与此同时, 多模型间的相互比较研究和数据-模型间的比对研究结果均显示, 当前人们对生态系统功能与干旱事件相互反馈的认识存在极大的不确定性(Ciais et al., 2005; Bond-Lamberty et al., 2018).为了减小这种不确定性, 一方面有必要利用实验数据评估和改进生态系统模型在模拟和预测生态系统过程上的性能, 从而减小模型间的不确定性(Zhou et al., 2019b).另一方面, 对于导致模型间不确定性的关键生态系统过程, 需要建立起不同区域之间相互协作的多尺度和多层次实验网络, 同时在各个生态系统类型中获取具有统一标准、由统一方法获得的实验数据, 从而进一步约束模型进而达到有效生态预测(Taylor et al., 2017). ...
Extreme drought slightly decreased soil labile organic C and N contents and altered microbial communities in a subtropical evergreen forest 1 2018
... 极端干旱对生态系统功能的影响主要表现在生产力、养分含量、凋落物分解等方面(Ciais et al., 2005; Zhou et al., 2009).这种影响作用可以反映在个体、群落及生态系统等尺度上.近年来, 国内外在不同尺度上围绕极端干旱对生态系统结构和功能的影响开展了一系列研究(Piao et al., 2009; Beier et al., 2012).例如, 华东师范大学于2013年7月依托浙江天童森林生态系统国家野外科学观测研究站, 建立了大型森林生态系统干旱实验平台, 着重探讨极端干旱对森林生态系统结构和功能, 特别是地下碳过程影响的研究, 目前已取得系列研究成果(聂园园等, 2017; Bu et al., 2018).研究发现极端干旱显著降低亚热带森林生态系统微生物和凋落物的生物量, 同时导致微生物优势群落从细菌向真菌转移, 进而降低土壤有机碳的分解(聂园园等, 2017).与此同时, 我们在北美大草原上的研究发现, 根系生物量和土壤碳含量与平均年降水量之间没有显著的相关性, 但土壤呼吸和茎生物量和年降水量之间存在显著的正相关关系, 而地上立枯物和表层凋落物生产量随着降水量的增加呈现先增加后降低的非线性变化趋势(Zhou et al., 2009).在内蒙古典型草原生态系统中也发现, 年降水量或季节性降水量与植被生产力呈现线性关系(Bai et al., 2004), 但是在更大尺度上也可能会出现非线性关系(Luo et al., 2017).陆地生态系统功能对干旱响应的敏感性差异主要受到植物生长潜力、物种组成以及资源可利用性等因素的共同影响(Beier et al., 2012).在水限制区域, 降水的增加会通过解除水分对微生物和植物生长的压力, 进而刺激土壤碳的排放过程; 反之, 干旱可以通过降低土壤水分, 增加含水量较高区域土壤的通透性, 进而刺激土壤呼吸的排放(Knapp et al., 2015).此外, 由于不同物种的光合作用策略以及对水分响应敏感性的差异, 干旱可能还会引起群落结构的改变, 如C3和C4优势物种相对丰度的变化(龚春梅等, 2009; Maestre et al., 2016). ...
Roots, water, and nutrient acquisition: Let’s get physical 1 2012
... 在植物个体水平上, 干旱通过降低土壤水分和养分的可利用性等途径来影响叶片和根系性状(Posch & Bennett, 2009; Zhou et al., 2018).干旱降低了叶片光合速率、气孔导度等可能会导致光合碳固定的减少, 导致植物分配到地下根系的生物量降低, 增加根系死亡率(张仁和等, 2011; Zhou et al., 2018).不同物种之间的叶片性状应对干旱的策略存在一定的差异.例如, 木麻黄属(Casuarina)的一种植物Allocasuarina luehmannii的气孔导度在干旱复水后能恢复到正常水平的60% (Posch & Bennett, 2009).但菜豆(Phaseolus vulgaris)的气孔导度、光合速率和蒸腾速率都显著降低, 并且其经干旱复水后的恢复能力会随着干旱天数的增加而逐渐降低(Miyashita et al., 2005).与此同时, 干旱胁迫下, 植物为了获取更多的水分和养分, 根系直径会变得越来越细, 同时其分布范围也会向更深处扩张(Chapman et al., 2012; Fuentealba et al., 2015).植物较细的根系结构有助于降低植物共生真菌的依赖度, 同时也利于优化植物单位碳投资的获取养分的效率, 进而增强植物对干旱环境的适应与存活能力(Ma et al., 2018).与此同时, 为了应对干旱压力, 多元资源获取策略快速生长型植物往往具有更长的比根长(SRL)和更强的根氮吸收能力, 但其根碳含量和根寿命较低(Pérez-Ramos et al., 2013).干旱导致根系碳、氮化学计量关系的不对称变化, 将进一步改变根系分泌物中有机质的输入, 进而改变与菌根之间的共生关系(Fuchslueger et al., 2014).全球meta分析结果发现在全球尺度上干旱显著降低根系生物量, 这种负面效应主要来源于植物根系性状(如根长、根长密度、比根长、根直径)的改变(Zhou et al., 2018). ...
Europe-wide reduction in primary productivity caused by the heat and drought in 2003 2 2005
... 极端干旱对生态系统功能的影响主要表现在生产力、养分含量、凋落物分解等方面(Ciais et al., 2005; Zhou et al., 2009).这种影响作用可以反映在个体、群落及生态系统等尺度上.近年来, 国内外在不同尺度上围绕极端干旱对生态系统结构和功能的影响开展了一系列研究(Piao et al., 2009; Beier et al., 2012).例如, 华东师范大学于2013年7月依托浙江天童森林生态系统国家野外科学观测研究站, 建立了大型森林生态系统干旱实验平台, 着重探讨极端干旱对森林生态系统结构和功能, 特别是地下碳过程影响的研究, 目前已取得系列研究成果(聂园园等, 2017; Bu et al., 2018).研究发现极端干旱显著降低亚热带森林生态系统微生物和凋落物的生物量, 同时导致微生物优势群落从细菌向真菌转移, 进而降低土壤有机碳的分解(聂园园等, 2017).与此同时, 我们在北美大草原上的研究发现, 根系生物量和土壤碳含量与平均年降水量之间没有显著的相关性, 但土壤呼吸和茎生物量和年降水量之间存在显著的正相关关系, 而地上立枯物和表层凋落物生产量随着降水量的增加呈现先增加后降低的非线性变化趋势(Zhou et al., 2009).在内蒙古典型草原生态系统中也发现, 年降水量或季节性降水量与植被生产力呈现线性关系(Bai et al., 2004), 但是在更大尺度上也可能会出现非线性关系(Luo et al., 2017).陆地生态系统功能对干旱响应的敏感性差异主要受到植物生长潜力、物种组成以及资源可利用性等因素的共同影响(Beier et al., 2012).在水限制区域, 降水的增加会通过解除水分对微生物和植物生长的压力, 进而刺激土壤碳的排放过程; 反之, 干旱可以通过降低土壤水分, 增加含水量较高区域土壤的通透性, 进而刺激土壤呼吸的排放(Knapp et al., 2015).此外, 由于不同物种的光合作用策略以及对水分响应敏感性的差异, 干旱可能还会引起群落结构的改变, 如C3和C4优势物种相对丰度的变化(龚春梅等, 2009; Maestre et al., 2016). ...
... 过去大量模型和野外控制实验均表明干旱事件会通过改变“植物-土壤-微生物”系统之间的生物地球化学循环过程, 直接或间接地调控生态系统养分(如碳、氮、磷等)循环过程(Piao et al., 2009; 聂园园等, 2017).但是由于实验观测和模型模拟两种方法的目的不完全一致, 使得大量关于生态系统功能的实验数据很难被模型直接采用, 而模型的模拟结果也很少作为产生科学假说的工具来有效指导实验研究(Piao et al., 2009; Taylor et al., 2017).与此同时, 多模型间的相互比较研究和数据-模型间的比对研究结果均显示, 当前人们对生态系统功能与干旱事件相互反馈的认识存在极大的不确定性(Ciais et al., 2005; Bond-Lamberty et al., 2018).为了减小这种不确定性, 一方面有必要利用实验数据评估和改进生态系统模型在模拟和预测生态系统过程上的性能, 从而减小模型间的不确定性(Zhou et al., 2019b).另一方面, 对于导致模型间不确定性的关键生态系统过程, 需要建立起不同区域之间相互协作的多尺度和多层次实验网络, 同时在各个生态系统类型中获取具有统一标准、由统一方法获得的实验数据, 从而进一步约束模型进而达到有效生态预测(Taylor et al., 2017). ...
... 生物多样性如何调控生态系统功能仍然是当前生态学家研究的重点和热点之一(徐炜等, 2016; Eisenhauer et al., 2018).过去关于干旱背景下生物多样性对生态系统功能影响的研究多集中在站点尺度, 但由于土壤养分、植被等生物和非生物因素对干旱响应的差异(Maestre et al., 2016), 对区域甚至更大尺度上生物多样性的影响及相关驱动机制仍知之甚少.与此同时, 干旱可能会导致地上地下生产力下降、养分循环失衡, 进而改变生态系统的多功能性(Eisenhauer et al., 2018).但是这种变化是否会带来地上地下生物多样性和生态系统关系的同步改变以及相关驱动机制的变化, 当前对其的认识仍不清楚(Maestre et al., 2016).这些知识盲区将在一定程度上导致未来模型在精确模拟和尺度推绎过程中存在很大的不确定性.因此, 未来开展更大尺度的干旱对生物多样性和生态系统功能关系影响的样带实验, 进一步修正和完善相关理论基础, 减少模型结构自身带来的不确定性, 最大程度地改进模型关于干旱对生物多样性影响预测的准确性, 是解决这一瓶颈问题的重要和关键手段(Bardgett & van der Putten, 2014; Eisenhauer et al., 2018). ...
... ), 对区域甚至更大尺度上生物多样性的影响及相关驱动机制仍知之甚少.与此同时, 干旱可能会导致地上地下生产力下降、养分循环失衡, 进而改变生态系统的多功能性(Eisenhauer et al., 2018).但是这种变化是否会带来地上地下生物多样性和生态系统关系的同步改变以及相关驱动机制的变化, 当前对其的认识仍不清楚(Maestre et al., 2016).这些知识盲区将在一定程度上导致未来模型在精确模拟和尺度推绎过程中存在很大的不确定性.因此, 未来开展更大尺度的干旱对生物多样性和生态系统功能关系影响的样带实验, 进一步修正和完善相关理论基础, 减少模型结构自身带来的不确定性, 最大程度地改进模型关于干旱对生物多样性影响预测的准确性, 是解决这一瓶颈问题的重要和关键手段(Bardgett & van der Putten, 2014; Eisenhauer et al., 2018). ...
... ; Eisenhauer et al., 2018). ...
Experiment, monitoring, and gradient methods used to infer climate change effects on plant communities yield consistent patterns 2 2015
... 在生态系统尺度上, 极端干旱对生态系统结构与功能的影响受到了广泛关注.极端干旱可能通过改变不同大小树木的密度, 或是改变物种间的相互作用来影响森林生态系统的群落结构与生物多样性(Zhao & Running, 2010; Elmendorf et al., 2015; Mcintyre et al., 2015).植物群落结构及多样性的改变, 以及干旱造成的水分胁迫都能够极大地影响森林生产力(Maestre et al., 2016), 进而影响其生态系统服务功能.然而, 当前关于极端干旱对森林生态系统结构与功能的影响主要集中于地上生产力的研究, 而对地下过程干旱敏感性的研究较少.一般而言, 在极端干旱背景下, 生态系统土壤的异养呼吸及碳周转速率会下降, 可能与微生物活性的下降有关(Zhou et al., 2019a).但从生态系统整体角度而言, 关于地上与地下生产力对极端干旱的响应是否具有同步性和对称性, 地上-地下耦合过程对极端干旱的反馈调节机制等, 仍是亟待系统深入研究的热点难点问题. ...
... 极端干旱主要通过调控生态系统中物质循环和能量流通, 以及改变物种间的相互作用来引起生物多样性的改变(Elmendorf et al., 2015).在不同的水热条件下, 植物之间相互作用关系, 养分的生物地球化学循环过程等存在差异, 从而驱动生物多样性分布的空间异质性(图4)(Bardgett & van der Putten, 2014; Taylor et al., 2017).干旱通过对植物的生长、繁殖和物候的调节, 进而改变群落内物种间相互作用来影响和调控地上生物多样性(Zhao & Running, 2010).与此同时, 干旱还可以通过改变生态系统物质循环和养分流通影响地下过程, 如对微生物(细菌、真菌、放线菌、古菌等)多样性、土壤动物(线虫、蚯蚓等)多样性产生重要影响, 并进一步调控地上-地下生物多样性之间的关系(Bardgett & van der Putten, 2014; Mcintyre et al., 2015).但是, 相较于地上多样性而言, 由于测定手段有限以及地下过程的复杂性, 当前人们在区域和全球尺度上就干旱对地下生态系统生物多样性的影响, 以及地上-地下生物多样性之间的协同进化关系的认知尚存争议(徐炜等, 2016).这种争议很大程度来源于地上、地下生物多样性对干旱响应的机制不同, 以及与植物-土壤等的复杂互馈过程(Bardgett & van der Putten, 2014).因此, 优化当前测定分析技术以及开展更大尺度、有针对性的研究, 是解决这一问题的有效途径. ...
Experimental drought reduces the transfer of recently fixed plant carbon to soil microbes and alters the bacterial community composition in a mountain meadow 1 2014
... 在植物个体水平上, 干旱通过降低土壤水分和养分的可利用性等途径来影响叶片和根系性状(Posch & Bennett, 2009; Zhou et al., 2018).干旱降低了叶片光合速率、气孔导度等可能会导致光合碳固定的减少, 导致植物分配到地下根系的生物量降低, 增加根系死亡率(张仁和等, 2011; Zhou et al., 2018).不同物种之间的叶片性状应对干旱的策略存在一定的差异.例如, 木麻黄属(Casuarina)的一种植物Allocasuarina luehmannii的气孔导度在干旱复水后能恢复到正常水平的60% (Posch & Bennett, 2009).但菜豆(Phaseolus vulgaris)的气孔导度、光合速率和蒸腾速率都显著降低, 并且其经干旱复水后的恢复能力会随着干旱天数的增加而逐渐降低(Miyashita et al., 2005).与此同时, 干旱胁迫下, 植物为了获取更多的水分和养分, 根系直径会变得越来越细, 同时其分布范围也会向更深处扩张(Chapman et al., 2012; Fuentealba et al., 2015).植物较细的根系结构有助于降低植物共生真菌的依赖度, 同时也利于优化植物单位碳投资的获取养分的效率, 进而增强植物对干旱环境的适应与存活能力(Ma et al., 2018).与此同时, 为了应对干旱压力, 多元资源获取策略快速生长型植物往往具有更长的比根长(SRL)和更强的根氮吸收能力, 但其根碳含量和根寿命较低(Pérez-Ramos et al., 2013).干旱导致根系碳、氮化学计量关系的不对称变化, 将进一步改变根系分泌物中有机质的输入, 进而改变与菌根之间的共生关系(Fuchslueger et al., 2014).全球meta分析结果发现在全球尺度上干旱显著降低根系生物量, 这种负面效应主要来源于植物根系性状(如根长、根长密度、比根长、根直径)的改变(Zhou et al., 2018). ...
Transpiration responses of warm- season turfgrass in relation to progressive soil drying 1 2015
... 在植物个体水平上, 干旱通过降低土壤水分和养分的可利用性等途径来影响叶片和根系性状(Posch & Bennett, 2009; Zhou et al., 2018).干旱降低了叶片光合速率、气孔导度等可能会导致光合碳固定的减少, 导致植物分配到地下根系的生物量降低, 增加根系死亡率(张仁和等, 2011; Zhou et al., 2018).不同物种之间的叶片性状应对干旱的策略存在一定的差异.例如, 木麻黄属(Casuarina)的一种植物Allocasuarina luehmannii的气孔导度在干旱复水后能恢复到正常水平的60% (Posch & Bennett, 2009).但菜豆(Phaseolus vulgaris)的气孔导度、光合速率和蒸腾速率都显著降低, 并且其经干旱复水后的恢复能力会随着干旱天数的增加而逐渐降低(Miyashita et al., 2005).与此同时, 干旱胁迫下, 植物为了获取更多的水分和养分, 根系直径会变得越来越细, 同时其分布范围也会向更深处扩张(Chapman et al., 2012; Fuentealba et al., 2015).植物较细的根系结构有助于降低植物共生真菌的依赖度, 同时也利于优化植物单位碳投资的获取养分的效率, 进而增强植物对干旱环境的适应与存活能力(Ma et al., 2018).与此同时, 为了应对干旱压力, 多元资源获取策略快速生长型植物往往具有更长的比根长(SRL)和更强的根氮吸收能力, 但其根碳含量和根寿命较低(Pérez-Ramos et al., 2013).干旱导致根系碳、氮化学计量关系的不对称变化, 将进一步改变根系分泌物中有机质的输入, 进而改变与菌根之间的共生关系(Fuchslueger et al., 2014).全球meta分析结果发现在全球尺度上干旱显著降低根系生物量, 这种负面效应主要来源于植物根系性状(如根长、根长密度、比根长、根直径)的改变(Zhou et al., 2018). ...
Multiple mechanisms of Amazonian forest biomass losses in three dynamic global vegetation models under climate change 1 2010
... 目前, 国内外围绕干旱对生态系统过程影响已开展了大量的模型模拟研究.其中模拟生态系统对极端干旱的响应以及干旱后的恢复过程一直是该领域的重点和难点, 存在很大的不确定性(Mencucciniet al., 2015).首先, 由于模型对植被、地表和土壤的诸多水文过程的模拟能力较低(Tron et al., 2015), 限制了对土壤干旱的发生时间和强度的预测准确性.同时, 植被对于干旱胁迫具有强烈的生物调节机制(如根系交互、地上和地下过程的耦合), 并且这种调节机制在不同生长季和区域表现出极大的差异, 这些都增加了模型对生态系统过程的干旱响应方式和强度的模拟难度(Warren et al., 2015).例如, 早在1996年Mahfouf等比较了14个陆地生态系统模型对于夏季作物蒸腾速率的模拟精度, 发现只有一半的模型模拟值在观测值的不确定性范围之内.时隔22年, 研究者在比较6个主流的生态系统模型对于土壤湿度的模拟能力时, 仍然发现所有模型均不能模拟生长季内平均土壤湿度的年际变化(Hu et al., 2018).在对植被的干旱响应的模拟方面, 目前的模拟结果也不尽如人意.Galbraith等(2010)的研究表明, 在亚马孙森林的截雨实验导致生物量减少了20%-30%.相似地, Powell等(2013)利用两个大尺度实验结果检验了5个生态系统模型, 发现所有模型均不能准确反映因干旱导致的地上生物量降低的趋势. ...
Isohydric species are not necessarily more carbon limited than anisohydric species during drought 1 2017
... 极端干旱直接影响到植物的生长和发育, 甚至导致其死亡.当前关于干旱对植物生理生态的影响, 主要在于探讨碳水过程对干旱的响应, 其中树木致死的内在机制是当前关注的核心和热点问题.“水力失衡”假说认为, 干旱导致树木死亡可能来源于木质部导管栓塞引起的“土壤-植物系统”间的水力传输过程失衡; “碳饥饿”假说则认为, 干旱诱发气孔关闭后导致光合作用减弱, 使得的光合产物不足以支撑植物的正常生理代谢过程, 导致植物死亡(图2) (McDowell et al., 2008).近期的研究证据多数倾向于水力失衡是干旱致死的主因(Anderegg et al., 2015; Rowland et al., 2015), 但是由于非结构性碳水化合物在植物生长和防御等生理生态过程中所扮演的复杂角色及其多变的时空动态, 目前关于植物非结构性碳水化合物富足的情况下是否实际上发生了碳饥饿事件, 尚未有明确的定论(McDowell & Sevanto, 2010).与此同时, 不同植物气孔调节叶片水势的策略上的差异也使碳饥饿现象在不同物种中发生的可能性有很大差异(Martínez-Vilalta et al., 2014; Garcia-Forner et al., 2017). ...
C3和C4植物光合途径的适应性变化和进化 1 2009
... 极端干旱对生态系统功能的影响主要表现在生产力、养分含量、凋落物分解等方面(Ciais et al., 2005; Zhou et al., 2009).这种影响作用可以反映在个体、群落及生态系统等尺度上.近年来, 国内外在不同尺度上围绕极端干旱对生态系统结构和功能的影响开展了一系列研究(Piao et al., 2009; Beier et al., 2012).例如, 华东师范大学于2013年7月依托浙江天童森林生态系统国家野外科学观测研究站, 建立了大型森林生态系统干旱实验平台, 着重探讨极端干旱对森林生态系统结构和功能, 特别是地下碳过程影响的研究, 目前已取得系列研究成果(聂园园等, 2017; Bu et al., 2018).研究发现极端干旱显著降低亚热带森林生态系统微生物和凋落物的生物量, 同时导致微生物优势群落从细菌向真菌转移, 进而降低土壤有机碳的分解(聂园园等, 2017).与此同时, 我们在北美大草原上的研究发现, 根系生物量和土壤碳含量与平均年降水量之间没有显著的相关性, 但土壤呼吸和茎生物量和年降水量之间存在显著的正相关关系, 而地上立枯物和表层凋落物生产量随着降水量的增加呈现先增加后降低的非线性变化趋势(Zhou et al., 2009).在内蒙古典型草原生态系统中也发现, 年降水量或季节性降水量与植被生产力呈现线性关系(Bai et al., 2004), 但是在更大尺度上也可能会出现非线性关系(Luo et al., 2017).陆地生态系统功能对干旱响应的敏感性差异主要受到植物生长潜力、物种组成以及资源可利用性等因素的共同影响(Beier et al., 2012).在水限制区域, 降水的增加会通过解除水分对微生物和植物生长的压力, 进而刺激土壤碳的排放过程; 反之, 干旱可以通过降低土壤水分, 增加含水量较高区域土壤的通透性, 进而刺激土壤呼吸的排放(Knapp et al., 2015).此外, 由于不同物种的光合作用策略以及对水分响应敏感性的差异, 干旱可能还会引起群落结构的改变, 如C3和C4优势物种相对丰度的变化(龚春梅等, 2009; Maestre et al., 2016). ...
C3和C4植物光合途径的适应性变化和进化 1 2009
... 极端干旱对生态系统功能的影响主要表现在生产力、养分含量、凋落物分解等方面(Ciais et al., 2005; Zhou et al., 2009).这种影响作用可以反映在个体、群落及生态系统等尺度上.近年来, 国内外在不同尺度上围绕极端干旱对生态系统结构和功能的影响开展了一系列研究(Piao et al., 2009; Beier et al., 2012).例如, 华东师范大学于2013年7月依托浙江天童森林生态系统国家野外科学观测研究站, 建立了大型森林生态系统干旱实验平台, 着重探讨极端干旱对森林生态系统结构和功能, 特别是地下碳过程影响的研究, 目前已取得系列研究成果(聂园园等, 2017; Bu et al., 2018).研究发现极端干旱显著降低亚热带森林生态系统微生物和凋落物的生物量, 同时导致微生物优势群落从细菌向真菌转移, 进而降低土壤有机碳的分解(聂园园等, 2017).与此同时, 我们在北美大草原上的研究发现, 根系生物量和土壤碳含量与平均年降水量之间没有显著的相关性, 但土壤呼吸和茎生物量和年降水量之间存在显著的正相关关系, 而地上立枯物和表层凋落物生产量随着降水量的增加呈现先增加后降低的非线性变化趋势(Zhou et al., 2009).在内蒙古典型草原生态系统中也发现, 年降水量或季节性降水量与植被生产力呈现线性关系(Bai et al., 2004), 但是在更大尺度上也可能会出现非线性关系(Luo et al., 2017).陆地生态系统功能对干旱响应的敏感性差异主要受到植物生长潜力、物种组成以及资源可利用性等因素的共同影响(Beier et al., 2012).在水限制区域, 降水的增加会通过解除水分对微生物和植物生长的压力, 进而刺激土壤碳的排放过程; 反之, 干旱可以通过降低土壤水分, 增加含水量较高区域土壤的通透性, 进而刺激土壤呼吸的排放(Knapp et al., 2015).此外, 由于不同物种的光合作用策略以及对水分响应敏感性的差异, 干旱可能还会引起群落结构的改变, 如C3和C4优势物种相对丰度的变化(龚春梅等, 2009; Maestre et al., 2016). ...
A global analysis of root distributions for terrestrial biomes 1 1996
... 在模拟植物响应土壤干旱过程中, 确定不同植物气孔导度在干旱时的变化特征(等水植物还是非等水植物)处于首要位置(Klein, 2014).植物的气孔调节方式存在着明显的地区差异, 如在湿润地区, 气孔对于干旱的响应敏感, 植物多为等水调节方式(Zhou et al., 2013).然而在目前的模型中, 参数主要按照植被类型区分, 假设在不同水分区域的同一植被类型对干旱的响应方式相同, 忽略了生物对干旱的调节响应差异势必会造成区域模拟的系统偏差(Mencuccini et al., 2015).除了地上部分的模拟偏差外, 在植物地下部分的模拟中, 目前的生态系统模型往往采用静态的根廓线方法(即不同土层的根系生物量随着深度呈现递减趋势, 并且在同一植被类型中根系的分布格局相同)来模拟地下根系(Jackson et al., 1996), 忽略了根系随着水分的变化特征, 无法准确模拟根系动态, 在一定程度上高估了干旱对植被的影响.最近的一些研究采用根系随水分变化的经验方程, 对根系生长的动态模拟进行了改进, 但对于根系的生物调节机制方面的模拟仍然非常欠缺.因此, 构建随水分条件变化的动态根模型也是提高干旱响应模拟准确性的关键(Warren et al., 2015). ...
The variability of stomatal sensitivity to leaf water potential across tree species indicates a continuum between isohydric and anisohydric behaviours 1 2014
... 在模拟植物响应土壤干旱过程中, 确定不同植物气孔导度在干旱时的变化特征(等水植物还是非等水植物)处于首要位置(Klein, 2014).植物的气孔调节方式存在着明显的地区差异, 如在湿润地区, 气孔对于干旱的响应敏感, 植物多为等水调节方式(Zhou et al., 2013).然而在目前的模型中, 参数主要按照植被类型区分, 假设在不同水分区域的同一植被类型对干旱的响应方式相同, 忽略了生物对干旱的调节响应差异势必会造成区域模拟的系统偏差(Mencuccini et al., 2015).除了地上部分的模拟偏差外, 在植物地下部分的模拟中, 目前的生态系统模型往往采用静态的根廓线方法(即不同土层的根系生物量随着深度呈现递减趋势, 并且在同一植被类型中根系的分布格局相同)来模拟地下根系(Jackson et al., 1996), 忽略了根系随着水分的变化特征, 无法准确模拟根系动态, 在一定程度上高估了干旱对植被的影响.最近的一些研究采用根系随水分变化的经验方程, 对根系生长的动态模拟进行了改进, 但对于根系的生物调节机制方面的模拟仍然非常欠缺.因此, 构建随水分条件变化的动态根模型也是提高干旱响应模拟准确性的关键(Warren et al., 2015). ...
Characterizing differences in precipitation regimes of extreme wet and dry years: implications for climate change experiments 1 2015
... 极端干旱对生态系统功能的影响主要表现在生产力、养分含量、凋落物分解等方面(Ciais et al., 2005; Zhou et al., 2009).这种影响作用可以反映在个体、群落及生态系统等尺度上.近年来, 国内外在不同尺度上围绕极端干旱对生态系统结构和功能的影响开展了一系列研究(Piao et al., 2009; Beier et al., 2012).例如, 华东师范大学于2013年7月依托浙江天童森林生态系统国家野外科学观测研究站, 建立了大型森林生态系统干旱实验平台, 着重探讨极端干旱对森林生态系统结构和功能, 特别是地下碳过程影响的研究, 目前已取得系列研究成果(聂园园等, 2017; Bu et al., 2018).研究发现极端干旱显著降低亚热带森林生态系统微生物和凋落物的生物量, 同时导致微生物优势群落从细菌向真菌转移, 进而降低土壤有机碳的分解(聂园园等, 2017).与此同时, 我们在北美大草原上的研究发现, 根系生物量和土壤碳含量与平均年降水量之间没有显著的相关性, 但土壤呼吸和茎生物量和年降水量之间存在显著的正相关关系, 而地上立枯物和表层凋落物生产量随着降水量的增加呈现先增加后降低的非线性变化趋势(Zhou et al., 2009).在内蒙古典型草原生态系统中也发现, 年降水量或季节性降水量与植被生产力呈现线性关系(Bai et al., 2004), 但是在更大尺度上也可能会出现非线性关系(Luo et al., 2017).陆地生态系统功能对干旱响应的敏感性差异主要受到植物生长潜力、物种组成以及资源可利用性等因素的共同影响(Beier et al., 2012).在水限制区域, 降水的增加会通过解除水分对微生物和植物生长的压力, 进而刺激土壤碳的排放过程; 反之, 干旱可以通过降低土壤水分, 增加含水量较高区域土壤的通透性, 进而刺激土壤呼吸的排放(Knapp et al., 2015).此外, 由于不同物种的光合作用策略以及对水分响应敏感性的差异, 干旱可能还会引起群落结构的改变, 如C3和C4优势物种相对丰度的变化(龚春梅等, 2009; Maestre et al., 2016). ...
Quantifying the role of fire in the Earth system—Part 2: Impact on the net carbon balance of global terrestrial ecosystems for the 20th century 1 2014
... 区域和全球的生物地球化学循环对降水格局的响应方式除了在较大的空间尺度上反映了植物生理生态及生态系统结构与功能对降水的响应过程外, 另有其独特的复杂性.首先, 大规模大范围的持续干旱事件的发生往往由厄尔尼诺-南方涛动、太平洋北美涛动等大尺度的气候事件所驱动, 因而这些干旱事件对生物地球化学循环的影响不仅反映了局部的气候效应, 更与全球气候系统关系紧密(Piao et al., 2009).其次, 越来越频发的干旱事件还能够增加诸如森林火灾、生态系统病虫害等负面事件的发生强度和频度(Schlesinger et al., 2016).这些灾害事件本身难以有效控制的特性使得它们的波及范围较广, 从而对区域甚至全球的生物地球化学循环造成极其深远的影响(Li et al., 2014).第三, 与植物个体和生态系统尺度的研究不同, 区域和全球尺度的研究无法开展统一降水受控实验, 因此很大程度上依赖于通过整合多尺度多来源的观测数据模拟和预测生物地球化学循环对降水格局的响应. ...
Nonlinear responses of land ecosystems to variation in precipitation 1 2017
... 极端干旱对生态系统功能的影响主要表现在生产力、养分含量、凋落物分解等方面(Ciais et al., 2005; Zhou et al., 2009).这种影响作用可以反映在个体、群落及生态系统等尺度上.近年来, 国内外在不同尺度上围绕极端干旱对生态系统结构和功能的影响开展了一系列研究(Piao et al., 2009; Beier et al., 2012).例如, 华东师范大学于2013年7月依托浙江天童森林生态系统国家野外科学观测研究站, 建立了大型森林生态系统干旱实验平台, 着重探讨极端干旱对森林生态系统结构和功能, 特别是地下碳过程影响的研究, 目前已取得系列研究成果(聂园园等, 2017; Bu et al., 2018).研究发现极端干旱显著降低亚热带森林生态系统微生物和凋落物的生物量, 同时导致微生物优势群落从细菌向真菌转移, 进而降低土壤有机碳的分解(聂园园等, 2017).与此同时, 我们在北美大草原上的研究发现, 根系生物量和土壤碳含量与平均年降水量之间没有显著的相关性, 但土壤呼吸和茎生物量和年降水量之间存在显著的正相关关系, 而地上立枯物和表层凋落物生产量随着降水量的增加呈现先增加后降低的非线性变化趋势(Zhou et al., 2009).在内蒙古典型草原生态系统中也发现, 年降水量或季节性降水量与植被生产力呈现线性关系(Bai et al., 2004), 但是在更大尺度上也可能会出现非线性关系(Luo et al., 2017).陆地生态系统功能对干旱响应的敏感性差异主要受到植物生长潜力、物种组成以及资源可利用性等因素的共同影响(Beier et al., 2012).在水限制区域, 降水的增加会通过解除水分对微生物和植物生长的压力, 进而刺激土壤碳的排放过程; 反之, 干旱可以通过降低土壤水分, 增加含水量较高区域土壤的通透性, 进而刺激土壤呼吸的排放(Knapp et al., 2015).此外, 由于不同物种的光合作用策略以及对水分响应敏感性的差异, 干旱可能还会引起群落结构的改变, 如C3和C4优势物种相对丰度的变化(龚春梅等, 2009; Maestre et al., 2016). ...
Evolutionary history resolves global organization of root functional traits 1 2018
... 在植物个体水平上, 干旱通过降低土壤水分和养分的可利用性等途径来影响叶片和根系性状(Posch & Bennett, 2009; Zhou et al., 2018).干旱降低了叶片光合速率、气孔导度等可能会导致光合碳固定的减少, 导致植物分配到地下根系的生物量降低, 增加根系死亡率(张仁和等, 2011; Zhou et al., 2018).不同物种之间的叶片性状应对干旱的策略存在一定的差异.例如, 木麻黄属(Casuarina)的一种植物Allocasuarina luehmannii的气孔导度在干旱复水后能恢复到正常水平的60% (Posch & Bennett, 2009).但菜豆(Phaseolus vulgaris)的气孔导度、光合速率和蒸腾速率都显著降低, 并且其经干旱复水后的恢复能力会随着干旱天数的增加而逐渐降低(Miyashita et al., 2005).与此同时, 干旱胁迫下, 植物为了获取更多的水分和养分, 根系直径会变得越来越细, 同时其分布范围也会向更深处扩张(Chapman et al., 2012; Fuentealba et al., 2015).植物较细的根系结构有助于降低植物共生真菌的依赖度, 同时也利于优化植物单位碳投资的获取养分的效率, 进而增强植物对干旱环境的适应与存活能力(Ma et al., 2018).与此同时, 为了应对干旱压力, 多元资源获取策略快速生长型植物往往具有更长的比根长(SRL)和更强的根氮吸收能力, 但其根碳含量和根寿命较低(Pérez-Ramos et al., 2013).干旱导致根系碳、氮化学计量关系的不对称变化, 将进一步改变根系分泌物中有机质的输入, 进而改变与菌根之间的共生关系(Fuchslueger et al., 2014).全球meta分析结果发现在全球尺度上干旱显著降低根系生物量, 这种负面效应主要来源于植物根系性状(如根长、根长密度、比根长、根直径)的改变(Zhou et al., 2018). ...
Structure and functioning of dryland ecosystems in a changing world 4 2016
... 极端干旱对生态系统功能的影响主要表现在生产力、养分含量、凋落物分解等方面(Ciais et al., 2005; Zhou et al., 2009).这种影响作用可以反映在个体、群落及生态系统等尺度上.近年来, 国内外在不同尺度上围绕极端干旱对生态系统结构和功能的影响开展了一系列研究(Piao et al., 2009; Beier et al., 2012).例如, 华东师范大学于2013年7月依托浙江天童森林生态系统国家野外科学观测研究站, 建立了大型森林生态系统干旱实验平台, 着重探讨极端干旱对森林生态系统结构和功能, 特别是地下碳过程影响的研究, 目前已取得系列研究成果(聂园园等, 2017; Bu et al., 2018).研究发现极端干旱显著降低亚热带森林生态系统微生物和凋落物的生物量, 同时导致微生物优势群落从细菌向真菌转移, 进而降低土壤有机碳的分解(聂园园等, 2017).与此同时, 我们在北美大草原上的研究发现, 根系生物量和土壤碳含量与平均年降水量之间没有显著的相关性, 但土壤呼吸和茎生物量和年降水量之间存在显著的正相关关系, 而地上立枯物和表层凋落物生产量随着降水量的增加呈现先增加后降低的非线性变化趋势(Zhou et al., 2009).在内蒙古典型草原生态系统中也发现, 年降水量或季节性降水量与植被生产力呈现线性关系(Bai et al., 2004), 但是在更大尺度上也可能会出现非线性关系(Luo et al., 2017).陆地生态系统功能对干旱响应的敏感性差异主要受到植物生长潜力、物种组成以及资源可利用性等因素的共同影响(Beier et al., 2012).在水限制区域, 降水的增加会通过解除水分对微生物和植物生长的压力, 进而刺激土壤碳的排放过程; 反之, 干旱可以通过降低土壤水分, 增加含水量较高区域土壤的通透性, 进而刺激土壤呼吸的排放(Knapp et al., 2015).此外, 由于不同物种的光合作用策略以及对水分响应敏感性的差异, 干旱可能还会引起群落结构的改变, 如C3和C4优势物种相对丰度的变化(龚春梅等, 2009; Maestre et al., 2016). ...
... 在生态系统尺度上, 极端干旱对生态系统结构与功能的影响受到了广泛关注.极端干旱可能通过改变不同大小树木的密度, 或是改变物种间的相互作用来影响森林生态系统的群落结构与生物多样性(Zhao & Running, 2010; Elmendorf et al., 2015; Mcintyre et al., 2015).植物群落结构及多样性的改变, 以及干旱造成的水分胁迫都能够极大地影响森林生产力(Maestre et al., 2016), 进而影响其生态系统服务功能.然而, 当前关于极端干旱对森林生态系统结构与功能的影响主要集中于地上生产力的研究, 而对地下过程干旱敏感性的研究较少.一般而言, 在极端干旱背景下, 生态系统土壤的异养呼吸及碳周转速率会下降, 可能与微生物活性的下降有关(Zhou et al., 2019a).但从生态系统整体角度而言, 关于地上与地下生产力对极端干旱的响应是否具有同步性和对称性, 地上-地下耦合过程对极端干旱的反馈调节机制等, 仍是亟待系统深入研究的热点难点问题. ...
... 生物多样性如何调控生态系统功能仍然是当前生态学家研究的重点和热点之一(徐炜等, 2016; Eisenhauer et al., 2018).过去关于干旱背景下生物多样性对生态系统功能影响的研究多集中在站点尺度, 但由于土壤养分、植被等生物和非生物因素对干旱响应的差异(Maestre et al., 2016), 对区域甚至更大尺度上生物多样性的影响及相关驱动机制仍知之甚少.与此同时, 干旱可能会导致地上地下生产力下降、养分循环失衡, 进而改变生态系统的多功能性(Eisenhauer et al., 2018).但是这种变化是否会带来地上地下生物多样性和生态系统关系的同步改变以及相关驱动机制的变化, 当前对其的认识仍不清楚(Maestre et al., 2016).这些知识盲区将在一定程度上导致未来模型在精确模拟和尺度推绎过程中存在很大的不确定性.因此, 未来开展更大尺度的干旱对生物多样性和生态系统功能关系影响的样带实验, 进一步修正和完善相关理论基础, 减少模型结构自身带来的不确定性, 最大程度地改进模型关于干旱对生物多样性影响预测的准确性, 是解决这一瓶颈问题的重要和关键手段(Bardgett & van der Putten, 2014; Eisenhauer et al., 2018). ...
... ).但是这种变化是否会带来地上地下生物多样性和生态系统关系的同步改变以及相关驱动机制的变化, 当前对其的认识仍不清楚(Maestre et al., 2016).这些知识盲区将在一定程度上导致未来模型在精确模拟和尺度推绎过程中存在很大的不确定性.因此, 未来开展更大尺度的干旱对生物多样性和生态系统功能关系影响的样带实验, 进一步修正和完善相关理论基础, 减少模型结构自身带来的不确定性, 最大程度地改进模型关于干旱对生物多样性影响预测的准确性, 是解决这一瓶颈问题的重要和关键手段(Bardgett & van der Putten, 2014; Eisenhauer et al., 2018). ...
Analysis of transpiration results from the RICE and PILPS workshop 1 1996
... 目前, 国内外围绕干旱对生态系统过程影响已开展了大量的模型模拟研究.其中模拟生态系统对极端干旱的响应以及干旱后的恢复过程一直是该领域的重点和难点, 存在很大的不确定性(Mencucciniet al., 2015).首先, 由于模型对植被、地表和土壤的诸多水文过程的模拟能力较低(Tron et al., 2015), 限制了对土壤干旱的发生时间和强度的预测准确性.同时, 植被对于干旱胁迫具有强烈的生物调节机制(如根系交互、地上和地下过程的耦合), 并且这种调节机制在不同生长季和区域表现出极大的差异, 这些都增加了模型对生态系统过程的干旱响应方式和强度的模拟难度(Warren et al., 2015).例如, 早在1996年Mahfouf等比较了14个陆地生态系统模型对于夏季作物蒸腾速率的模拟精度, 发现只有一半的模型模拟值在观测值的不确定性范围之内.时隔22年, 研究者在比较6个主流的生态系统模型对于土壤湿度的模拟能力时, 仍然发现所有模型均不能模拟生长季内平均土壤湿度的年际变化(Hu et al., 2018).在对植被的干旱响应的模拟方面, 目前的模拟结果也不尽如人意.Galbraith等(2010)的研究表明, 在亚马孙森林的截雨实验导致生物量减少了20%-30%.相似地, Powell等(2013)利用两个大尺度实验结果检验了5个生态系统模型, 发现所有模型均不能准确反映因干旱导致的地上生物量降低的趋势. ...
A new look at water transport regulation in plants 1 2014
... 极端干旱直接影响到植物的生长和发育, 甚至导致其死亡.当前关于干旱对植物生理生态的影响, 主要在于探讨碳水过程对干旱的响应, 其中树木致死的内在机制是当前关注的核心和热点问题.“水力失衡”假说认为, 干旱导致树木死亡可能来源于木质部导管栓塞引起的“土壤-植物系统”间的水力传输过程失衡; “碳饥饿”假说则认为, 干旱诱发气孔关闭后导致光合作用减弱, 使得的光合产物不足以支撑植物的正常生理代谢过程, 导致植物死亡(图2) (McDowell et al., 2008).近期的研究证据多数倾向于水力失衡是干旱致死的主因(Anderegg et al., 2015; Rowland et al., 2015), 但是由于非结构性碳水化合物在植物生长和防御等生理生态过程中所扮演的复杂角色及其多变的时空动态, 目前关于植物非结构性碳水化合物富足的情况下是否实际上发生了碳饥饿事件, 尚未有明确的定论(McDowell & Sevanto, 2010).与此同时, 不同植物气孔调节叶片水势的策略上的差异也使碳饥饿现象在不同物种中发生的可能性有很大差异(Martínez-Vilalta et al., 2014; Garcia-Forner et al., 2017). ...
The mechanisms of carbon starvation: How, when, or does it even occur at all? 1 2010
... 极端干旱直接影响到植物的生长和发育, 甚至导致其死亡.当前关于干旱对植物生理生态的影响, 主要在于探讨碳水过程对干旱的响应, 其中树木致死的内在机制是当前关注的核心和热点问题.“水力失衡”假说认为, 干旱导致树木死亡可能来源于木质部导管栓塞引起的“土壤-植物系统”间的水力传输过程失衡; “碳饥饿”假说则认为, 干旱诱发气孔关闭后导致光合作用减弱, 使得的光合产物不足以支撑植物的正常生理代谢过程, 导致植物死亡(图2) (McDowell et al., 2008).近期的研究证据多数倾向于水力失衡是干旱致死的主因(Anderegg et al., 2015; Rowland et al., 2015), 但是由于非结构性碳水化合物在植物生长和防御等生理生态过程中所扮演的复杂角色及其多变的时空动态, 目前关于植物非结构性碳水化合物富足的情况下是否实际上发生了碳饥饿事件, 尚未有明确的定论(McDowell & Sevanto, 2010).与此同时, 不同植物气孔调节叶片水势的策略上的差异也使碳饥饿现象在不同物种中发生的可能性有很大差异(Martínez-Vilalta et al., 2014; Garcia-Forner et al., 2017). ...
Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? 1 2008
... 极端干旱直接影响到植物的生长和发育, 甚至导致其死亡.当前关于干旱对植物生理生态的影响, 主要在于探讨碳水过程对干旱的响应, 其中树木致死的内在机制是当前关注的核心和热点问题.“水力失衡”假说认为, 干旱导致树木死亡可能来源于木质部导管栓塞引起的“土壤-植物系统”间的水力传输过程失衡; “碳饥饿”假说则认为, 干旱诱发气孔关闭后导致光合作用减弱, 使得的光合产物不足以支撑植物的正常生理代谢过程, 导致植物死亡(图2) (McDowell et al., 2008).近期的研究证据多数倾向于水力失衡是干旱致死的主因(Anderegg et al., 2015; Rowland et al., 2015), 但是由于非结构性碳水化合物在植物生长和防御等生理生态过程中所扮演的复杂角色及其多变的时空动态, 目前关于植物非结构性碳水化合物富足的情况下是否实际上发生了碳饥饿事件, 尚未有明确的定论(McDowell & Sevanto, 2010).与此同时, 不同植物气孔调节叶片水势的策略上的差异也使碳饥饿现象在不同物种中发生的可能性有很大差异(Martínez-Vilalta et al., 2014; Garcia-Forner et al., 2017). ...
Twentieth-century shifts in forest structure in California: denser forests, smaller trees, and increased dominance of oaks 2 2015
... 在生态系统尺度上, 极端干旱对生态系统结构与功能的影响受到了广泛关注.极端干旱可能通过改变不同大小树木的密度, 或是改变物种间的相互作用来影响森林生态系统的群落结构与生物多样性(Zhao & Running, 2010; Elmendorf et al., 2015; Mcintyre et al., 2015).植物群落结构及多样性的改变, 以及干旱造成的水分胁迫都能够极大地影响森林生产力(Maestre et al., 2016), 进而影响其生态系统服务功能.然而, 当前关于极端干旱对森林生态系统结构与功能的影响主要集中于地上生产力的研究, 而对地下过程干旱敏感性的研究较少.一般而言, 在极端干旱背景下, 生态系统土壤的异养呼吸及碳周转速率会下降, 可能与微生物活性的下降有关(Zhou et al., 2019a).但从生态系统整体角度而言, 关于地上与地下生产力对极端干旱的响应是否具有同步性和对称性, 地上-地下耦合过程对极端干旱的反馈调节机制等, 仍是亟待系统深入研究的热点难点问题. ...
... 极端干旱主要通过调控生态系统中物质循环和能量流通, 以及改变物种间的相互作用来引起生物多样性的改变(Elmendorf et al., 2015).在不同的水热条件下, 植物之间相互作用关系, 养分的生物地球化学循环过程等存在差异, 从而驱动生物多样性分布的空间异质性(图4)(Bardgett & van der Putten, 2014; Taylor et al., 2017).干旱通过对植物的生长、繁殖和物候的调节, 进而改变群落内物种间相互作用来影响和调控地上生物多样性(Zhao & Running, 2010).与此同时, 干旱还可以通过改变生态系统物质循环和养分流通影响地下过程, 如对微生物(细菌、真菌、放线菌、古菌等)多样性、土壤动物(线虫、蚯蚓等)多样性产生重要影响, 并进一步调控地上-地下生物多样性之间的关系(Bardgett & van der Putten, 2014; Mcintyre et al., 2015).但是, 相较于地上多样性而言, 由于测定手段有限以及地下过程的复杂性, 当前人们在区域和全球尺度上就干旱对地下生态系统生物多样性的影响, 以及地上-地下生物多样性之间的协同进化关系的认知尚存争议(徐炜等, 2016).这种争议很大程度来源于地上、地下生物多样性对干旱响应的机制不同, 以及与植物-土壤等的复杂互馈过程(Bardgett & van der Putten, 2014).因此, 优化当前测定分析技术以及开展更大尺度、有针对性的研究, 是解决这一问题的有效途径. ...
... 在自然生态系统中, 干旱事件并不是独立存在的, 往往伴随着其他气候因子(如增温、氮沉降)和人类活动(如放牧、森林砍伐)的同时发生(图5).过去的研究表明, 干旱与其他生物和非生物因素的综合效应, 对陆地生态系统的生物地球化学循环、植物生理生态、生物多样性和生态系统功能等方面都会产生重要影响.例如, Zhou等(2019a)的研究表明, 干旱能够显著降低土壤呼吸和生态系统呼吸, 但是这种负效应在一定程度上能够被增温所缓解.这可能是因为增温能够促进土壤氮的矿化作用, 增加土壤氮的可利用性, 进而缓解干旱导致的土壤氮含量降低(Melillo et al., 2011).与此同时, 干旱与氮沉降、增温、CO2浓度升高在调控草原生态系统呼吸过程中存在明显的交互作用, 这种作用主要以加和效应为主, 而非协同和拮抗作用(Zhou et al., 2019a).在草原生态系统中, 当干旱和放牧活动相伴发生时, 干旱对生态系统地上和地下碳过程的影响, 在一定程度上被放牧活动所主导(Zhou et al., 2019b).这种作用主要来源于植被生物量和地下养分资源对干旱的响应滞后于放牧活动.因此, 未来在评估干旱对生态系统的影响时, 应当充分考虑干旱和其他因素的综合效应. ...
Coordination of physiological traits involved in drought-induced mortality of woody plants 2 2015
... 目前, 国内外围绕干旱对生态系统过程影响已开展了大量的模型模拟研究.其中模拟生态系统对极端干旱的响应以及干旱后的恢复过程一直是该领域的重点和难点, 存在很大的不确定性(Mencucciniet al., 2015).首先, 由于模型对植被、地表和土壤的诸多水文过程的模拟能力较低(Tron et al., 2015), 限制了对土壤干旱的发生时间和强度的预测准确性.同时, 植被对于干旱胁迫具有强烈的生物调节机制(如根系交互、地上和地下过程的耦合), 并且这种调节机制在不同生长季和区域表现出极大的差异, 这些都增加了模型对生态系统过程的干旱响应方式和强度的模拟难度(Warren et al., 2015).例如, 早在1996年Mahfouf等比较了14个陆地生态系统模型对于夏季作物蒸腾速率的模拟精度, 发现只有一半的模型模拟值在观测值的不确定性范围之内.时隔22年, 研究者在比较6个主流的生态系统模型对于土壤湿度的模拟能力时, 仍然发现所有模型均不能模拟生长季内平均土壤湿度的年际变化(Hu et al., 2018).在对植被的干旱响应的模拟方面, 目前的模拟结果也不尽如人意.Galbraith等(2010)的研究表明, 在亚马孙森林的截雨实验导致生物量减少了20%-30%.相似地, Powell等(2013)利用两个大尺度实验结果检验了5个生态系统模型, 发现所有模型均不能准确反映因干旱导致的地上生物量降低的趋势. ...
... 在模拟植物响应土壤干旱过程中, 确定不同植物气孔导度在干旱时的变化特征(等水植物还是非等水植物)处于首要位置(Klein, 2014).植物的气孔调节方式存在着明显的地区差异, 如在湿润地区, 气孔对于干旱的响应敏感, 植物多为等水调节方式(Zhou et al., 2013).然而在目前的模型中, 参数主要按照植被类型区分, 假设在不同水分区域的同一植被类型对干旱的响应方式相同, 忽略了生物对干旱的调节响应差异势必会造成区域模拟的系统偏差(Mencuccini et al., 2015).除了地上部分的模拟偏差外, 在植物地下部分的模拟中, 目前的生态系统模型往往采用静态的根廓线方法(即不同土层的根系生物量随着深度呈现递减趋势, 并且在同一植被类型中根系的分布格局相同)来模拟地下根系(Jackson et al., 1996), 忽略了根系随着水分的变化特征, 无法准确模拟根系动态, 在一定程度上高估了干旱对植被的影响.最近的一些研究采用根系随水分变化的经验方程, 对根系生长的动态模拟进行了改进, 但对于根系的生物调节机制方面的模拟仍然非常欠缺.因此, 构建随水分条件变化的动态根模型也是提高干旱响应模拟准确性的关键(Warren et al., 2015). ...
Recovery responses of photosynthesis, transpiration, and stomatal conductance in kidney bean following drought stress 1 2005
... 在植物个体水平上, 干旱通过降低土壤水分和养分的可利用性等途径来影响叶片和根系性状(Posch & Bennett, 2009; Zhou et al., 2018).干旱降低了叶片光合速率、气孔导度等可能会导致光合碳固定的减少, 导致植物分配到地下根系的生物量降低, 增加根系死亡率(张仁和等, 2011; Zhou et al., 2018).不同物种之间的叶片性状应对干旱的策略存在一定的差异.例如, 木麻黄属(Casuarina)的一种植物Allocasuarina luehmannii的气孔导度在干旱复水后能恢复到正常水平的60% (Posch & Bennett, 2009).但菜豆(Phaseolus vulgaris)的气孔导度、光合速率和蒸腾速率都显著降低, 并且其经干旱复水后的恢复能力会随着干旱天数的增加而逐渐降低(Miyashita et al., 2005).与此同时, 干旱胁迫下, 植物为了获取更多的水分和养分, 根系直径会变得越来越细, 同时其分布范围也会向更深处扩张(Chapman et al., 2012; Fuentealba et al., 2015).植物较细的根系结构有助于降低植物共生真菌的依赖度, 同时也利于优化植物单位碳投资的获取养分的效率, 进而增强植物对干旱环境的适应与存活能力(Ma et al., 2018).与此同时, 为了应对干旱压力, 多元资源获取策略快速生长型植物往往具有更长的比根长(SRL)和更强的根氮吸收能力, 但其根碳含量和根寿命较低(Pérez-Ramos et al., 2013).干旱导致根系碳、氮化学计量关系的不对称变化, 将进一步改变根系分泌物中有机质的输入, 进而改变与菌根之间的共生关系(Fuchslueger et al., 2014).全球meta分析结果发现在全球尺度上干旱显著降低根系生物量, 这种负面效应主要来源于植物根系性状(如根长、根长密度、比根长、根直径)的改变(Zhou et al., 2018). ...
模拟干旱对亚热带森林土壤微生物生物量及群落结构的影响 3 2017
... 极端干旱对生态系统功能的影响主要表现在生产力、养分含量、凋落物分解等方面(Ciais et al., 2005; Zhou et al., 2009).这种影响作用可以反映在个体、群落及生态系统等尺度上.近年来, 国内外在不同尺度上围绕极端干旱对生态系统结构和功能的影响开展了一系列研究(Piao et al., 2009; Beier et al., 2012).例如, 华东师范大学于2013年7月依托浙江天童森林生态系统国家野外科学观测研究站, 建立了大型森林生态系统干旱实验平台, 着重探讨极端干旱对森林生态系统结构和功能, 特别是地下碳过程影响的研究, 目前已取得系列研究成果(聂园园等, 2017; Bu et al., 2018).研究发现极端干旱显著降低亚热带森林生态系统微生物和凋落物的生物量, 同时导致微生物优势群落从细菌向真菌转移, 进而降低土壤有机碳的分解(聂园园等, 2017).与此同时, 我们在北美大草原上的研究发现, 根系生物量和土壤碳含量与平均年降水量之间没有显著的相关性, 但土壤呼吸和茎生物量和年降水量之间存在显著的正相关关系, 而地上立枯物和表层凋落物生产量随着降水量的增加呈现先增加后降低的非线性变化趋势(Zhou et al., 2009).在内蒙古典型草原生态系统中也发现, 年降水量或季节性降水量与植被生产力呈现线性关系(Bai et al., 2004), 但是在更大尺度上也可能会出现非线性关系(Luo et al., 2017).陆地生态系统功能对干旱响应的敏感性差异主要受到植物生长潜力、物种组成以及资源可利用性等因素的共同影响(Beier et al., 2012).在水限制区域, 降水的增加会通过解除水分对微生物和植物生长的压力, 进而刺激土壤碳的排放过程; 反之, 干旱可以通过降低土壤水分, 增加含水量较高区域土壤的通透性, 进而刺激土壤呼吸的排放(Knapp et al., 2015).此外, 由于不同物种的光合作用策略以及对水分响应敏感性的差异, 干旱可能还会引起群落结构的改变, 如C3和C4优势物种相对丰度的变化(龚春梅等, 2009; Maestre et al., 2016). ...
... ).研究发现极端干旱显著降低亚热带森林生态系统微生物和凋落物的生物量, 同时导致微生物优势群落从细菌向真菌转移, 进而降低土壤有机碳的分解(聂园园等, 2017).与此同时, 我们在北美大草原上的研究发现, 根系生物量和土壤碳含量与平均年降水量之间没有显著的相关性, 但土壤呼吸和茎生物量和年降水量之间存在显著的正相关关系, 而地上立枯物和表层凋落物生产量随着降水量的增加呈现先增加后降低的非线性变化趋势(Zhou et al., 2009).在内蒙古典型草原生态系统中也发现, 年降水量或季节性降水量与植被生产力呈现线性关系(Bai et al., 2004), 但是在更大尺度上也可能会出现非线性关系(Luo et al., 2017).陆地生态系统功能对干旱响应的敏感性差异主要受到植物生长潜力、物种组成以及资源可利用性等因素的共同影响(Beier et al., 2012).在水限制区域, 降水的增加会通过解除水分对微生物和植物生长的压力, 进而刺激土壤碳的排放过程; 反之, 干旱可以通过降低土壤水分, 增加含水量较高区域土壤的通透性, 进而刺激土壤呼吸的排放(Knapp et al., 2015).此外, 由于不同物种的光合作用策略以及对水分响应敏感性的差异, 干旱可能还会引起群落结构的改变, 如C3和C4优势物种相对丰度的变化(龚春梅等, 2009; Maestre et al., 2016). ...
... 过去大量模型和野外控制实验均表明干旱事件会通过改变“植物-土壤-微生物”系统之间的生物地球化学循环过程, 直接或间接地调控生态系统养分(如碳、氮、磷等)循环过程(Piao et al., 2009; 聂园园等, 2017).但是由于实验观测和模型模拟两种方法的目的不完全一致, 使得大量关于生态系统功能的实验数据很难被模型直接采用, 而模型的模拟结果也很少作为产生科学假说的工具来有效指导实验研究(Piao et al., 2009; Taylor et al., 2017).与此同时, 多模型间的相互比较研究和数据-模型间的比对研究结果均显示, 当前人们对生态系统功能与干旱事件相互反馈的认识存在极大的不确定性(Ciais et al., 2005; Bond-Lamberty et al., 2018).为了减小这种不确定性, 一方面有必要利用实验数据评估和改进生态系统模型在模拟和预测生态系统过程上的性能, 从而减小模型间的不确定性(Zhou et al., 2019b).另一方面, 对于导致模型间不确定性的关键生态系统过程, 需要建立起不同区域之间相互协作的多尺度和多层次实验网络, 同时在各个生态系统类型中获取具有统一标准、由统一方法获得的实验数据, 从而进一步约束模型进而达到有效生态预测(Taylor et al., 2017). ...
模拟干旱对亚热带森林土壤微生物生物量及群落结构的影响 3 2017
... 极端干旱对生态系统功能的影响主要表现在生产力、养分含量、凋落物分解等方面(Ciais et al., 2005; Zhou et al., 2009).这种影响作用可以反映在个体、群落及生态系统等尺度上.近年来, 国内外在不同尺度上围绕极端干旱对生态系统结构和功能的影响开展了一系列研究(Piao et al., 2009; Beier et al., 2012).例如, 华东师范大学于2013年7月依托浙江天童森林生态系统国家野外科学观测研究站, 建立了大型森林生态系统干旱实验平台, 着重探讨极端干旱对森林生态系统结构和功能, 特别是地下碳过程影响的研究, 目前已取得系列研究成果(聂园园等, 2017; Bu et al., 2018).研究发现极端干旱显著降低亚热带森林生态系统微生物和凋落物的生物量, 同时导致微生物优势群落从细菌向真菌转移, 进而降低土壤有机碳的分解(聂园园等, 2017).与此同时, 我们在北美大草原上的研究发现, 根系生物量和土壤碳含量与平均年降水量之间没有显著的相关性, 但土壤呼吸和茎生物量和年降水量之间存在显著的正相关关系, 而地上立枯物和表层凋落物生产量随着降水量的增加呈现先增加后降低的非线性变化趋势(Zhou et al., 2009).在内蒙古典型草原生态系统中也发现, 年降水量或季节性降水量与植被生产力呈现线性关系(Bai et al., 2004), 但是在更大尺度上也可能会出现非线性关系(Luo et al., 2017).陆地生态系统功能对干旱响应的敏感性差异主要受到植物生长潜力、物种组成以及资源可利用性等因素的共同影响(Beier et al., 2012).在水限制区域, 降水的增加会通过解除水分对微生物和植物生长的压力, 进而刺激土壤碳的排放过程; 反之, 干旱可以通过降低土壤水分, 增加含水量较高区域土壤的通透性, 进而刺激土壤呼吸的排放(Knapp et al., 2015).此外, 由于不同物种的光合作用策略以及对水分响应敏感性的差异, 干旱可能还会引起群落结构的改变, 如C3和C4优势物种相对丰度的变化(龚春梅等, 2009; Maestre et al., 2016). ...
... ).研究发现极端干旱显著降低亚热带森林生态系统微生物和凋落物的生物量, 同时导致微生物优势群落从细菌向真菌转移, 进而降低土壤有机碳的分解(聂园园等, 2017).与此同时, 我们在北美大草原上的研究发现, 根系生物量和土壤碳含量与平均年降水量之间没有显著的相关性, 但土壤呼吸和茎生物量和年降水量之间存在显著的正相关关系, 而地上立枯物和表层凋落物生产量随着降水量的增加呈现先增加后降低的非线性变化趋势(Zhou et al., 2009).在内蒙古典型草原生态系统中也发现, 年降水量或季节性降水量与植被生产力呈现线性关系(Bai et al., 2004), 但是在更大尺度上也可能会出现非线性关系(Luo et al., 2017).陆地生态系统功能对干旱响应的敏感性差异主要受到植物生长潜力、物种组成以及资源可利用性等因素的共同影响(Beier et al., 2012).在水限制区域, 降水的增加会通过解除水分对微生物和植物生长的压力, 进而刺激土壤碳的排放过程; 反之, 干旱可以通过降低土壤水分, 增加含水量较高区域土壤的通透性, 进而刺激土壤呼吸的排放(Knapp et al., 2015).此外, 由于不同物种的光合作用策略以及对水分响应敏感性的差异, 干旱可能还会引起群落结构的改变, 如C3和C4优势物种相对丰度的变化(龚春梅等, 2009; Maestre et al., 2016). ...
... 过去大量模型和野外控制实验均表明干旱事件会通过改变“植物-土壤-微生物”系统之间的生物地球化学循环过程, 直接或间接地调控生态系统养分(如碳、氮、磷等)循环过程(Piao et al., 2009; 聂园园等, 2017).但是由于实验观测和模型模拟两种方法的目的不完全一致, 使得大量关于生态系统功能的实验数据很难被模型直接采用, 而模型的模拟结果也很少作为产生科学假说的工具来有效指导实验研究(Piao et al., 2009; Taylor et al., 2017).与此同时, 多模型间的相互比较研究和数据-模型间的比对研究结果均显示, 当前人们对生态系统功能与干旱事件相互反馈的认识存在极大的不确定性(Ciais et al., 2005; Bond-Lamberty et al., 2018).为了减小这种不确定性, 一方面有必要利用实验数据评估和改进生态系统模型在模拟和预测生态系统过程上的性能, 从而减小模型间的不确定性(Zhou et al., 2019b).另一方面, 对于导致模型间不确定性的关键生态系统过程, 需要建立起不同区域之间相互协作的多尺度和多层次实验网络, 同时在各个生态系统类型中获取具有统一标准、由统一方法获得的实验数据, 从而进一步约束模型进而达到有效生态预测(Taylor et al., 2017). ...
Tradeoffs between functional strategies for resource use and drought-survival in Mediterranean rangeland species 1 2013
... 在植物个体水平上, 干旱通过降低土壤水分和养分的可利用性等途径来影响叶片和根系性状(Posch & Bennett, 2009; Zhou et al., 2018).干旱降低了叶片光合速率、气孔导度等可能会导致光合碳固定的减少, 导致植物分配到地下根系的生物量降低, 增加根系死亡率(张仁和等, 2011; Zhou et al., 2018).不同物种之间的叶片性状应对干旱的策略存在一定的差异.例如, 木麻黄属(Casuarina)的一种植物Allocasuarina luehmannii的气孔导度在干旱复水后能恢复到正常水平的60% (Posch & Bennett, 2009).但菜豆(Phaseolus vulgaris)的气孔导度、光合速率和蒸腾速率都显著降低, 并且其经干旱复水后的恢复能力会随着干旱天数的增加而逐渐降低(Miyashita et al., 2005).与此同时, 干旱胁迫下, 植物为了获取更多的水分和养分, 根系直径会变得越来越细, 同时其分布范围也会向更深处扩张(Chapman et al., 2012; Fuentealba et al., 2015).植物较细的根系结构有助于降低植物共生真菌的依赖度, 同时也利于优化植物单位碳投资的获取养分的效率, 进而增强植物对干旱环境的适应与存活能力(Ma et al., 2018).与此同时, 为了应对干旱压力, 多元资源获取策略快速生长型植物往往具有更长的比根长(SRL)和更强的根氮吸收能力, 但其根碳含量和根寿命较低(Pérez-Ramos et al., 2013).干旱导致根系碳、氮化学计量关系的不对称变化, 将进一步改变根系分泌物中有机质的输入, 进而改变与菌根之间的共生关系(Fuchslueger et al., 2014).全球meta分析结果发现在全球尺度上干旱显著降低根系生物量, 这种负面效应主要来源于植物根系性状(如根长、根长密度、比根长、根直径)的改变(Zhou et al., 2018). ...
The carbon balance of terrestrial ecosystems in China 6 2009
... 中国区域与全球陆地生态系统的净初级生产力和净碳汇的时间动态都受到降水量的严重影响(Piao et al., 2009).大范围的极端干旱事件往往是导致某一区域生态系统生物地球化学循环发生巨大改变的驱动力.例如1983年在亚马孙地区出现的极端干旱事件释放了相当于该地区生态系统需要3-4年才能吸收获得的CO2 (Potter et al., 2001).2003年夏季在欧洲地区发生的持续高温热浪天气极大地降低了该地区生态系统的生产力, 甚至提升了全球大气CO2浓度的上升速率(Bevan et al., 2014).目前该领域的研究大多讨论碳循环和水循环过程对极端干旱的响应.由于氮循环在调控生态系统功能中的重要作用, 近年来在区域和全球尺度上干旱对生态系统碳-氮-水耦合过程的影响格局及其调控机制开始逐渐受到关注. ...
... 区域和全球的生物地球化学循环对降水格局的响应方式除了在较大的空间尺度上反映了植物生理生态及生态系统结构与功能对降水的响应过程外, 另有其独特的复杂性.首先, 大规模大范围的持续干旱事件的发生往往由厄尔尼诺-南方涛动、太平洋北美涛动等大尺度的气候事件所驱动, 因而这些干旱事件对生物地球化学循环的影响不仅反映了局部的气候效应, 更与全球气候系统关系紧密(Piao et al., 2009).其次, 越来越频发的干旱事件还能够增加诸如森林火灾、生态系统病虫害等负面事件的发生强度和频度(Schlesinger et al., 2016).这些灾害事件本身难以有效控制的特性使得它们的波及范围较广, 从而对区域甚至全球的生物地球化学循环造成极其深远的影响(Li et al., 2014).第三, 与植物个体和生态系统尺度的研究不同, 区域和全球尺度的研究无法开展统一降水受控实验, 因此很大程度上依赖于通过整合多尺度多来源的观测数据模拟和预测生物地球化学循环对降水格局的响应. ...
... 极端干旱对生态系统功能的影响主要表现在生产力、养分含量、凋落物分解等方面(Ciais et al., 2005; Zhou et al., 2009).这种影响作用可以反映在个体、群落及生态系统等尺度上.近年来, 国内外在不同尺度上围绕极端干旱对生态系统结构和功能的影响开展了一系列研究(Piao et al., 2009; Beier et al., 2012).例如, 华东师范大学于2013年7月依托浙江天童森林生态系统国家野外科学观测研究站, 建立了大型森林生态系统干旱实验平台, 着重探讨极端干旱对森林生态系统结构和功能, 特别是地下碳过程影响的研究, 目前已取得系列研究成果(聂园园等, 2017; Bu et al., 2018).研究发现极端干旱显著降低亚热带森林生态系统微生物和凋落物的生物量, 同时导致微生物优势群落从细菌向真菌转移, 进而降低土壤有机碳的分解(聂园园等, 2017).与此同时, 我们在北美大草原上的研究发现, 根系生物量和土壤碳含量与平均年降水量之间没有显著的相关性, 但土壤呼吸和茎生物量和年降水量之间存在显著的正相关关系, 而地上立枯物和表层凋落物生产量随着降水量的增加呈现先增加后降低的非线性变化趋势(Zhou et al., 2009).在内蒙古典型草原生态系统中也发现, 年降水量或季节性降水量与植被生产力呈现线性关系(Bai et al., 2004), 但是在更大尺度上也可能会出现非线性关系(Luo et al., 2017).陆地生态系统功能对干旱响应的敏感性差异主要受到植物生长潜力、物种组成以及资源可利用性等因素的共同影响(Beier et al., 2012).在水限制区域, 降水的增加会通过解除水分对微生物和植物生长的压力, 进而刺激土壤碳的排放过程; 反之, 干旱可以通过降低土壤水分, 增加含水量较高区域土壤的通透性, 进而刺激土壤呼吸的排放(Knapp et al., 2015).此外, 由于不同物种的光合作用策略以及对水分响应敏感性的差异, 干旱可能还会引起群落结构的改变, 如C3和C4优势物种相对丰度的变化(龚春梅等, 2009; Maestre et al., 2016). ...
... 过去大量模型和野外控制实验均表明干旱事件会通过改变“植物-土壤-微生物”系统之间的生物地球化学循环过程, 直接或间接地调控生态系统养分(如碳、氮、磷等)循环过程(Piao et al., 2009; 聂园园等, 2017).但是由于实验观测和模型模拟两种方法的目的不完全一致, 使得大量关于生态系统功能的实验数据很难被模型直接采用, 而模型的模拟结果也很少作为产生科学假说的工具来有效指导实验研究(Piao et al., 2009; Taylor et al., 2017).与此同时, 多模型间的相互比较研究和数据-模型间的比对研究结果均显示, 当前人们对生态系统功能与干旱事件相互反馈的认识存在极大的不确定性(Ciais et al., 2005; Bond-Lamberty et al., 2018).为了减小这种不确定性, 一方面有必要利用实验数据评估和改进生态系统模型在模拟和预测生态系统过程上的性能, 从而减小模型间的不确定性(Zhou et al., 2019b).另一方面, 对于导致模型间不确定性的关键生态系统过程, 需要建立起不同区域之间相互协作的多尺度和多层次实验网络, 同时在各个生态系统类型中获取具有统一标准、由统一方法获得的实验数据, 从而进一步约束模型进而达到有效生态预测(Taylor et al., 2017). ...
... ).但是由于实验观测和模型模拟两种方法的目的不完全一致, 使得大量关于生态系统功能的实验数据很难被模型直接采用, 而模型的模拟结果也很少作为产生科学假说的工具来有效指导实验研究(Piao et al., 2009; Taylor et al., 2017).与此同时, 多模型间的相互比较研究和数据-模型间的比对研究结果均显示, 当前人们对生态系统功能与干旱事件相互反馈的认识存在极大的不确定性(Ciais et al., 2005; Bond-Lamberty et al., 2018).为了减小这种不确定性, 一方面有必要利用实验数据评估和改进生态系统模型在模拟和预测生态系统过程上的性能, 从而减小模型间的不确定性(Zhou et al., 2019b).另一方面, 对于导致模型间不确定性的关键生态系统过程, 需要建立起不同区域之间相互协作的多尺度和多层次实验网络, 同时在各个生态系统类型中获取具有统一标准、由统一方法获得的实验数据, 从而进一步约束模型进而达到有效生态预测(Taylor et al., 2017). ...
Photosynthesis, photochemistry and antioxidative defence in response to two drought severities and with re-watering in Allocasuarina luehmannii 2 2009
... 在植物个体水平上, 干旱通过降低土壤水分和养分的可利用性等途径来影响叶片和根系性状(Posch & Bennett, 2009; Zhou et al., 2018).干旱降低了叶片光合速率、气孔导度等可能会导致光合碳固定的减少, 导致植物分配到地下根系的生物量降低, 增加根系死亡率(张仁和等, 2011; Zhou et al., 2018).不同物种之间的叶片性状应对干旱的策略存在一定的差异.例如, 木麻黄属(Casuarina)的一种植物Allocasuarina luehmannii的气孔导度在干旱复水后能恢复到正常水平的60% (Posch & Bennett, 2009).但菜豆(Phaseolus vulgaris)的气孔导度、光合速率和蒸腾速率都显著降低, 并且其经干旱复水后的恢复能力会随着干旱天数的增加而逐渐降低(Miyashita et al., 2005).与此同时, 干旱胁迫下, 植物为了获取更多的水分和养分, 根系直径会变得越来越细, 同时其分布范围也会向更深处扩张(Chapman et al., 2012; Fuentealba et al., 2015).植物较细的根系结构有助于降低植物共生真菌的依赖度, 同时也利于优化植物单位碳投资的获取养分的效率, 进而增强植物对干旱环境的适应与存活能力(Ma et al., 2018).与此同时, 为了应对干旱压力, 多元资源获取策略快速生长型植物往往具有更长的比根长(SRL)和更强的根氮吸收能力, 但其根碳含量和根寿命较低(Pérez-Ramos et al., 2013).干旱导致根系碳、氮化学计量关系的不对称变化, 将进一步改变根系分泌物中有机质的输入, 进而改变与菌根之间的共生关系(Fuchslueger et al., 2014).全球meta分析结果发现在全球尺度上干旱显著降低根系生物量, 这种负面效应主要来源于植物根系性状(如根长、根长密度、比根长、根直径)的改变(Zhou et al., 2018). ...
... 的气孔导度在干旱复水后能恢复到正常水平的60% (Posch & Bennett, 2009).但菜豆(Phaseolus vulgaris)的气孔导度、光合速率和蒸腾速率都显著降低, 并且其经干旱复水后的恢复能力会随着干旱天数的增加而逐渐降低(Miyashita et al., 2005).与此同时, 干旱胁迫下, 植物为了获取更多的水分和养分, 根系直径会变得越来越细, 同时其分布范围也会向更深处扩张(Chapman et al., 2012; Fuentealba et al., 2015).植物较细的根系结构有助于降低植物共生真菌的依赖度, 同时也利于优化植物单位碳投资的获取养分的效率, 进而增强植物对干旱环境的适应与存活能力(Ma et al., 2018).与此同时, 为了应对干旱压力, 多元资源获取策略快速生长型植物往往具有更长的比根长(SRL)和更强的根氮吸收能力, 但其根碳含量和根寿命较低(Pérez-Ramos et al., 2013).干旱导致根系碳、氮化学计量关系的不对称变化, 将进一步改变根系分泌物中有机质的输入, 进而改变与菌根之间的共生关系(Fuchslueger et al., 2014).全球meta分析结果发现在全球尺度上干旱显著降低根系生物量, 这种负面效应主要来源于植物根系性状(如根长、根长密度、比根长、根直径)的改变(Zhou et al., 2018). ...
Modeling seasonal and interannual variability in ecosystem carbon cycling for the Brazilian Amazon region 1 2001
... 中国区域与全球陆地生态系统的净初级生产力和净碳汇的时间动态都受到降水量的严重影响(Piao et al., 2009).大范围的极端干旱事件往往是导致某一区域生态系统生物地球化学循环发生巨大改变的驱动力.例如1983年在亚马孙地区出现的极端干旱事件释放了相当于该地区生态系统需要3-4年才能吸收获得的CO2 (Potter et al., 2001).2003年夏季在欧洲地区发生的持续高温热浪天气极大地降低了该地区生态系统的生产力, 甚至提升了全球大气CO2浓度的上升速率(Bevan et al., 2014).目前该领域的研究大多讨论碳循环和水循环过程对极端干旱的响应.由于氮循环在调控生态系统功能中的重要作用, 近年来在区域和全球尺度上干旱对生态系统碳-氮-水耦合过程的影响格局及其调控机制开始逐渐受到关注. ...
... 过去大量模型和野外控制实验均表明干旱事件会通过改变“植物-土壤-微生物”系统之间的生物地球化学循环过程, 直接或间接地调控生态系统养分(如碳、氮、磷等)循环过程(Piao et al., 2009; 聂园园等, 2017).但是由于实验观测和模型模拟两种方法的目的不完全一致, 使得大量关于生态系统功能的实验数据很难被模型直接采用, 而模型的模拟结果也很少作为产生科学假说的工具来有效指导实验研究(Piao et al., 2009; Taylor et al., 2017).与此同时, 多模型间的相互比较研究和数据-模型间的比对研究结果均显示, 当前人们对生态系统功能与干旱事件相互反馈的认识存在极大的不确定性(Ciais et al., 2005; Bond-Lamberty et al., 2018).为了减小这种不确定性, 一方面有必要利用实验数据评估和改进生态系统模型在模拟和预测生态系统过程上的性能, 从而减小模型间的不确定性(Zhou et al., 2019b).另一方面, 对于导致模型间不确定性的关键生态系统过程, 需要建立起不同区域之间相互协作的多尺度和多层次实验网络, 同时在各个生态系统类型中获取具有统一标准、由统一方法获得的实验数据, 从而进一步约束模型进而达到有效生态预测(Taylor et al., 2017). ...
Temperature and rainfall interact to control carbon cycling in tropical forests 2 2017
... 过去大量模型和野外控制实验均表明干旱事件会通过改变“植物-土壤-微生物”系统之间的生物地球化学循环过程, 直接或间接地调控生态系统养分(如碳、氮、磷等)循环过程(Piao et al., 2009; 聂园园等, 2017).但是由于实验观测和模型模拟两种方法的目的不完全一致, 使得大量关于生态系统功能的实验数据很难被模型直接采用, 而模型的模拟结果也很少作为产生科学假说的工具来有效指导实验研究(Piao et al., 2009; Taylor et al., 2017).与此同时, 多模型间的相互比较研究和数据-模型间的比对研究结果均显示, 当前人们对生态系统功能与干旱事件相互反馈的认识存在极大的不确定性(Ciais et al., 2005; Bond-Lamberty et al., 2018).为了减小这种不确定性, 一方面有必要利用实验数据评估和改进生态系统模型在模拟和预测生态系统过程上的性能, 从而减小模型间的不确定性(Zhou et al., 2019b).另一方面, 对于导致模型间不确定性的关键生态系统过程, 需要建立起不同区域之间相互协作的多尺度和多层次实验网络, 同时在各个生态系统类型中获取具有统一标准、由统一方法获得的实验数据, 从而进一步约束模型进而达到有效生态预测(Taylor et al., 2017). ...
... 极端干旱主要通过调控生态系统中物质循环和能量流通, 以及改变物种间的相互作用来引起生物多样性的改变(Elmendorf et al., 2015).在不同的水热条件下, 植物之间相互作用关系, 养分的生物地球化学循环过程等存在差异, 从而驱动生物多样性分布的空间异质性(图4)(Bardgett & van der Putten, 2014; Taylor et al., 2017).干旱通过对植物的生长、繁殖和物候的调节, 进而改变群落内物种间相互作用来影响和调控地上生物多样性(Zhao & Running, 2010).与此同时, 干旱还可以通过改变生态系统物质循环和养分流通影响地下过程, 如对微生物(细菌、真菌、放线菌、古菌等)多样性、土壤动物(线虫、蚯蚓等)多样性产生重要影响, 并进一步调控地上-地下生物多样性之间的关系(Bardgett & van der Putten, 2014; Mcintyre et al., 2015).但是, 相较于地上多样性而言, 由于测定手段有限以及地下过程的复杂性, 当前人们在区域和全球尺度上就干旱对地下生态系统生物多样性的影响, 以及地上-地下生物多样性之间的协同进化关系的认知尚存争议(徐炜等, 2016).这种争议很大程度来源于地上、地下生物多样性对干旱响应的机制不同, 以及与植物-土壤等的复杂互馈过程(Bardgett & van der Putten, 2014).因此, 优化当前测定分析技术以及开展更大尺度、有针对性的研究, 是解决这一问题的有效途径. ...
The signature of randomness in riparian plant root distributions 1 2015
... 目前, 国内外围绕干旱对生态系统过程影响已开展了大量的模型模拟研究.其中模拟生态系统对极端干旱的响应以及干旱后的恢复过程一直是该领域的重点和难点, 存在很大的不确定性(Mencucciniet al., 2015).首先, 由于模型对植被、地表和土壤的诸多水文过程的模拟能力较低(Tron et al., 2015), 限制了对土壤干旱的发生时间和强度的预测准确性.同时, 植被对于干旱胁迫具有强烈的生物调节机制(如根系交互、地上和地下过程的耦合), 并且这种调节机制在不同生长季和区域表现出极大的差异, 这些都增加了模型对生态系统过程的干旱响应方式和强度的模拟难度(Warren et al., 2015).例如, 早在1996年Mahfouf等比较了14个陆地生态系统模型对于夏季作物蒸腾速率的模拟精度, 发现只有一半的模型模拟值在观测值的不确定性范围之内.时隔22年, 研究者在比较6个主流的生态系统模型对于土壤湿度的模拟能力时, 仍然发现所有模型均不能模拟生长季内平均土壤湿度的年际变化(Hu et al., 2018).在对植被的干旱响应的模拟方面, 目前的模拟结果也不尽如人意.Galbraith等(2010)的研究表明, 在亚马孙森林的截雨实验导致生物量减少了20%-30%.相似地, Powell等(2013)利用两个大尺度实验结果检验了5个生态系统模型, 发现所有模型均不能准确反映因干旱导致的地上生物量降低的趋势. ...
Root structural and functional dynamics in terrestrial biosphere models—Evaluation and recommendations 2 2015
... 目前, 国内外围绕干旱对生态系统过程影响已开展了大量的模型模拟研究.其中模拟生态系统对极端干旱的响应以及干旱后的恢复过程一直是该领域的重点和难点, 存在很大的不确定性(Mencucciniet al., 2015).首先, 由于模型对植被、地表和土壤的诸多水文过程的模拟能力较低(Tron et al., 2015), 限制了对土壤干旱的发生时间和强度的预测准确性.同时, 植被对于干旱胁迫具有强烈的生物调节机制(如根系交互、地上和地下过程的耦合), 并且这种调节机制在不同生长季和区域表现出极大的差异, 这些都增加了模型对生态系统过程的干旱响应方式和强度的模拟难度(Warren et al., 2015).例如, 早在1996年Mahfouf等比较了14个陆地生态系统模型对于夏季作物蒸腾速率的模拟精度, 发现只有一半的模型模拟值在观测值的不确定性范围之内.时隔22年, 研究者在比较6个主流的生态系统模型对于土壤湿度的模拟能力时, 仍然发现所有模型均不能模拟生长季内平均土壤湿度的年际变化(Hu et al., 2018).在对植被的干旱响应的模拟方面, 目前的模拟结果也不尽如人意.Galbraith等(2010)的研究表明, 在亚马孙森林的截雨实验导致生物量减少了20%-30%.相似地, Powell等(2013)利用两个大尺度实验结果检验了5个生态系统模型, 发现所有模型均不能准确反映因干旱导致的地上生物量降低的趋势. ...
... 在模拟植物响应土壤干旱过程中, 确定不同植物气孔导度在干旱时的变化特征(等水植物还是非等水植物)处于首要位置(Klein, 2014).植物的气孔调节方式存在着明显的地区差异, 如在湿润地区, 气孔对于干旱的响应敏感, 植物多为等水调节方式(Zhou et al., 2013).然而在目前的模型中, 参数主要按照植被类型区分, 假设在不同水分区域的同一植被类型对干旱的响应方式相同, 忽略了生物对干旱的调节响应差异势必会造成区域模拟的系统偏差(Mencuccini et al., 2015).除了地上部分的模拟偏差外, 在植物地下部分的模拟中, 目前的生态系统模型往往采用静态的根廓线方法(即不同土层的根系生物量随着深度呈现递减趋势, 并且在同一植被类型中根系的分布格局相同)来模拟地下根系(Jackson et al., 1996), 忽略了根系随着水分的变化特征, 无法准确模拟根系动态, 在一定程度上高估了干旱对植被的影响.最近的一些研究采用根系随水分变化的经验方程, 对根系生长的动态模拟进行了改进, 但对于根系的生物调节机制方面的模拟仍然非常欠缺.因此, 构建随水分条件变化的动态根模型也是提高干旱响应模拟准确性的关键(Warren et al., 2015). ...
生物多样性与生态系统多功能性: 进展与展望 2 2016
... 极端干旱主要通过调控生态系统中物质循环和能量流通, 以及改变物种间的相互作用来引起生物多样性的改变(Elmendorf et al., 2015).在不同的水热条件下, 植物之间相互作用关系, 养分的生物地球化学循环过程等存在差异, 从而驱动生物多样性分布的空间异质性(图4)(Bardgett & van der Putten, 2014; Taylor et al., 2017).干旱通过对植物的生长、繁殖和物候的调节, 进而改变群落内物种间相互作用来影响和调控地上生物多样性(Zhao & Running, 2010).与此同时, 干旱还可以通过改变生态系统物质循环和养分流通影响地下过程, 如对微生物(细菌、真菌、放线菌、古菌等)多样性、土壤动物(线虫、蚯蚓等)多样性产生重要影响, 并进一步调控地上-地下生物多样性之间的关系(Bardgett & van der Putten, 2014; Mcintyre et al., 2015).但是, 相较于地上多样性而言, 由于测定手段有限以及地下过程的复杂性, 当前人们在区域和全球尺度上就干旱对地下生态系统生物多样性的影响, 以及地上-地下生物多样性之间的协同进化关系的认知尚存争议(徐炜等, 2016).这种争议很大程度来源于地上、地下生物多样性对干旱响应的机制不同, 以及与植物-土壤等的复杂互馈过程(Bardgett & van der Putten, 2014).因此, 优化当前测定分析技术以及开展更大尺度、有针对性的研究, 是解决这一问题的有效途径. ...
... 生物多样性如何调控生态系统功能仍然是当前生态学家研究的重点和热点之一(徐炜等, 2016; Eisenhauer et al., 2018).过去关于干旱背景下生物多样性对生态系统功能影响的研究多集中在站点尺度, 但由于土壤养分、植被等生物和非生物因素对干旱响应的差异(Maestre et al., 2016), 对区域甚至更大尺度上生物多样性的影响及相关驱动机制仍知之甚少.与此同时, 干旱可能会导致地上地下生产力下降、养分循环失衡, 进而改变生态系统的多功能性(Eisenhauer et al., 2018).但是这种变化是否会带来地上地下生物多样性和生态系统关系的同步改变以及相关驱动机制的变化, 当前对其的认识仍不清楚(Maestre et al., 2016).这些知识盲区将在一定程度上导致未来模型在精确模拟和尺度推绎过程中存在很大的不确定性.因此, 未来开展更大尺度的干旱对生物多样性和生态系统功能关系影响的样带实验, 进一步修正和完善相关理论基础, 减少模型结构自身带来的不确定性, 最大程度地改进模型关于干旱对生物多样性影响预测的准确性, 是解决这一瓶颈问题的重要和关键手段(Bardgett & van der Putten, 2014; Eisenhauer et al., 2018). ...
生物多样性与生态系统多功能性: 进展与展望 2 2016
... 极端干旱主要通过调控生态系统中物质循环和能量流通, 以及改变物种间的相互作用来引起生物多样性的改变(Elmendorf et al., 2015).在不同的水热条件下, 植物之间相互作用关系, 养分的生物地球化学循环过程等存在差异, 从而驱动生物多样性分布的空间异质性(图4)(Bardgett & van der Putten, 2014; Taylor et al., 2017).干旱通过对植物的生长、繁殖和物候的调节, 进而改变群落内物种间相互作用来影响和调控地上生物多样性(Zhao & Running, 2010).与此同时, 干旱还可以通过改变生态系统物质循环和养分流通影响地下过程, 如对微生物(细菌、真菌、放线菌、古菌等)多样性、土壤动物(线虫、蚯蚓等)多样性产生重要影响, 并进一步调控地上-地下生物多样性之间的关系(Bardgett & van der Putten, 2014; Mcintyre et al., 2015).但是, 相较于地上多样性而言, 由于测定手段有限以及地下过程的复杂性, 当前人们在区域和全球尺度上就干旱对地下生态系统生物多样性的影响, 以及地上-地下生物多样性之间的协同进化关系的认知尚存争议(徐炜等, 2016).这种争议很大程度来源于地上、地下生物多样性对干旱响应的机制不同, 以及与植物-土壤等的复杂互馈过程(Bardgett & van der Putten, 2014).因此, 优化当前测定分析技术以及开展更大尺度、有针对性的研究, 是解决这一问题的有效途径. ...
... 生物多样性如何调控生态系统功能仍然是当前生态学家研究的重点和热点之一(徐炜等, 2016; Eisenhauer et al., 2018).过去关于干旱背景下生物多样性对生态系统功能影响的研究多集中在站点尺度, 但由于土壤养分、植被等生物和非生物因素对干旱响应的差异(Maestre et al., 2016), 对区域甚至更大尺度上生物多样性的影响及相关驱动机制仍知之甚少.与此同时, 干旱可能会导致地上地下生产力下降、养分循环失衡, 进而改变生态系统的多功能性(Eisenhauer et al., 2018).但是这种变化是否会带来地上地下生物多样性和生态系统关系的同步改变以及相关驱动机制的变化, 当前对其的认识仍不清楚(Maestre et al., 2016).这些知识盲区将在一定程度上导致未来模型在精确模拟和尺度推绎过程中存在很大的不确定性.因此, 未来开展更大尺度的干旱对生物多样性和生态系统功能关系影响的样带实验, 进一步修正和完善相关理论基础, 减少模型结构自身带来的不确定性, 最大程度地改进模型关于干旱对生物多样性影响预测的准确性, 是解决这一瓶颈问题的重要和关键手段(Bardgett & van der Putten, 2014; Eisenhauer et al., 2018). ...
... 在植物个体水平上, 干旱通过降低土壤水分和养分的可利用性等途径来影响叶片和根系性状(Posch & Bennett, 2009; Zhou et al., 2018).干旱降低了叶片光合速率、气孔导度等可能会导致光合碳固定的减少, 导致植物分配到地下根系的生物量降低, 增加根系死亡率(张仁和等, 2011; Zhou et al., 2018).不同物种之间的叶片性状应对干旱的策略存在一定的差异.例如, 木麻黄属(Casuarina)的一种植物Allocasuarina luehmannii的气孔导度在干旱复水后能恢复到正常水平的60% (Posch & Bennett, 2009).但菜豆(Phaseolus vulgaris)的气孔导度、光合速率和蒸腾速率都显著降低, 并且其经干旱复水后的恢复能力会随着干旱天数的增加而逐渐降低(Miyashita et al., 2005).与此同时, 干旱胁迫下, 植物为了获取更多的水分和养分, 根系直径会变得越来越细, 同时其分布范围也会向更深处扩张(Chapman et al., 2012; Fuentealba et al., 2015).植物较细的根系结构有助于降低植物共生真菌的依赖度, 同时也利于优化植物单位碳投资的获取养分的效率, 进而增强植物对干旱环境的适应与存活能力(Ma et al., 2018).与此同时, 为了应对干旱压力, 多元资源获取策略快速生长型植物往往具有更长的比根长(SRL)和更强的根氮吸收能力, 但其根碳含量和根寿命较低(Pérez-Ramos et al., 2013).干旱导致根系碳、氮化学计量关系的不对称变化, 将进一步改变根系分泌物中有机质的输入, 进而改变与菌根之间的共生关系(Fuchslueger et al., 2014).全球meta分析结果发现在全球尺度上干旱显著降低根系生物量, 这种负面效应主要来源于植物根系性状(如根长、根长密度、比根长、根直径)的改变(Zhou et al., 2018). ...
干旱胁迫对玉米苗期叶片光合作用和保护酶的影响 1 2011
... 在植物个体水平上, 干旱通过降低土壤水分和养分的可利用性等途径来影响叶片和根系性状(Posch & Bennett, 2009; Zhou et al., 2018).干旱降低了叶片光合速率、气孔导度等可能会导致光合碳固定的减少, 导致植物分配到地下根系的生物量降低, 增加根系死亡率(张仁和等, 2011; Zhou et al., 2018).不同物种之间的叶片性状应对干旱的策略存在一定的差异.例如, 木麻黄属(Casuarina)的一种植物Allocasuarina luehmannii的气孔导度在干旱复水后能恢复到正常水平的60% (Posch & Bennett, 2009).但菜豆(Phaseolus vulgaris)的气孔导度、光合速率和蒸腾速率都显著降低, 并且其经干旱复水后的恢复能力会随着干旱天数的增加而逐渐降低(Miyashita et al., 2005).与此同时, 干旱胁迫下, 植物为了获取更多的水分和养分, 根系直径会变得越来越细, 同时其分布范围也会向更深处扩张(Chapman et al., 2012; Fuentealba et al., 2015).植物较细的根系结构有助于降低植物共生真菌的依赖度, 同时也利于优化植物单位碳投资的获取养分的效率, 进而增强植物对干旱环境的适应与存活能力(Ma et al., 2018).与此同时, 为了应对干旱压力, 多元资源获取策略快速生长型植物往往具有更长的比根长(SRL)和更强的根氮吸收能力, 但其根碳含量和根寿命较低(Pérez-Ramos et al., 2013).干旱导致根系碳、氮化学计量关系的不对称变化, 将进一步改变根系分泌物中有机质的输入, 进而改变与菌根之间的共生关系(Fuchslueger et al., 2014).全球meta分析结果发现在全球尺度上干旱显著降低根系生物量, 这种负面效应主要来源于植物根系性状(如根长、根长密度、比根长、根直径)的改变(Zhou et al., 2018). ...
Drought-induced reduction in global terrestrial net primary production from 2000 through 2009 2 2010
... 在生态系统尺度上, 极端干旱对生态系统结构与功能的影响受到了广泛关注.极端干旱可能通过改变不同大小树木的密度, 或是改变物种间的相互作用来影响森林生态系统的群落结构与生物多样性(Zhao & Running, 2010; Elmendorf et al., 2015; Mcintyre et al., 2015).植物群落结构及多样性的改变, 以及干旱造成的水分胁迫都能够极大地影响森林生产力(Maestre et al., 2016), 进而影响其生态系统服务功能.然而, 当前关于极端干旱对森林生态系统结构与功能的影响主要集中于地上生产力的研究, 而对地下过程干旱敏感性的研究较少.一般而言, 在极端干旱背景下, 生态系统土壤的异养呼吸及碳周转速率会下降, 可能与微生物活性的下降有关(Zhou et al., 2019a).但从生态系统整体角度而言, 关于地上与地下生产力对极端干旱的响应是否具有同步性和对称性, 地上-地下耦合过程对极端干旱的反馈调节机制等, 仍是亟待系统深入研究的热点难点问题. ...
... 极端干旱主要通过调控生态系统中物质循环和能量流通, 以及改变物种间的相互作用来引起生物多样性的改变(Elmendorf et al., 2015).在不同的水热条件下, 植物之间相互作用关系, 养分的生物地球化学循环过程等存在差异, 从而驱动生物多样性分布的空间异质性(图4)(Bardgett & van der Putten, 2014; Taylor et al., 2017).干旱通过对植物的生长、繁殖和物候的调节, 进而改变群落内物种间相互作用来影响和调控地上生物多样性(Zhao & Running, 2010).与此同时, 干旱还可以通过改变生态系统物质循环和养分流通影响地下过程, 如对微生物(细菌、真菌、放线菌、古菌等)多样性、土壤动物(线虫、蚯蚓等)多样性产生重要影响, 并进一步调控地上-地下生物多样性之间的关系(Bardgett & van der Putten, 2014; Mcintyre et al., 2015).但是, 相较于地上多样性而言, 由于测定手段有限以及地下过程的复杂性, 当前人们在区域和全球尺度上就干旱对地下生态系统生物多样性的影响, 以及地上-地下生物多样性之间的协同进化关系的认知尚存争议(徐炜等, 2016).这种争议很大程度来源于地上、地下生物多样性对干旱响应的机制不同, 以及与植物-土壤等的复杂互馈过程(Bardgett & van der Putten, 2014).因此, 优化当前测定分析技术以及开展更大尺度、有针对性的研究, 是解决这一问题的有效途径. ...
Interactive effects of grazing and global change factors on soil and ecosystems respiration in grassland ecosystems 3 2019
... 在生态系统尺度上, 极端干旱对生态系统结构与功能的影响受到了广泛关注.极端干旱可能通过改变不同大小树木的密度, 或是改变物种间的相互作用来影响森林生态系统的群落结构与生物多样性(Zhao & Running, 2010; Elmendorf et al., 2015; Mcintyre et al., 2015).植物群落结构及多样性的改变, 以及干旱造成的水分胁迫都能够极大地影响森林生产力(Maestre et al., 2016), 进而影响其生态系统服务功能.然而, 当前关于极端干旱对森林生态系统结构与功能的影响主要集中于地上生产力的研究, 而对地下过程干旱敏感性的研究较少.一般而言, 在极端干旱背景下, 生态系统土壤的异养呼吸及碳周转速率会下降, 可能与微生物活性的下降有关(Zhou et al., 2019a).但从生态系统整体角度而言, 关于地上与地下生产力对极端干旱的响应是否具有同步性和对称性, 地上-地下耦合过程对极端干旱的反馈调节机制等, 仍是亟待系统深入研究的热点难点问题. ...
... 在自然生态系统中, 干旱事件并不是独立存在的, 往往伴随着其他气候因子(如增温、氮沉降)和人类活动(如放牧、森林砍伐)的同时发生(图5).过去的研究表明, 干旱与其他生物和非生物因素的综合效应, 对陆地生态系统的生物地球化学循环、植物生理生态、生物多样性和生态系统功能等方面都会产生重要影响.例如, Zhou等(2019a)的研究表明, 干旱能够显著降低土壤呼吸和生态系统呼吸, 但是这种负效应在一定程度上能够被增温所缓解.这可能是因为增温能够促进土壤氮的矿化作用, 增加土壤氮的可利用性, 进而缓解干旱导致的土壤氮含量降低(Melillo et al., 2011).与此同时, 干旱与氮沉降、增温、CO2浓度升高在调控草原生态系统呼吸过程中存在明显的交互作用, 这种作用主要以加和效应为主, 而非协同和拮抗作用(Zhou et al., 2019a).在草原生态系统中, 当干旱和放牧活动相伴发生时, 干旱对生态系统地上和地下碳过程的影响, 在一定程度上被放牧活动所主导(Zhou et al., 2019b).这种作用主要来源于植被生物量和地下养分资源对干旱的响应滞后于放牧活动.因此, 未来在评估干旱对生态系统的影响时, 应当充分考虑干旱和其他因素的综合效应. ...
... 浓度升高在调控草原生态系统呼吸过程中存在明显的交互作用, 这种作用主要以加和效应为主, 而非协同和拮抗作用(Zhou et al., 2019a).在草原生态系统中, 当干旱和放牧活动相伴发生时, 干旱对生态系统地上和地下碳过程的影响, 在一定程度上被放牧活动所主导(Zhou et al., 2019b).这种作用主要来源于植被生物量和地下养分资源对干旱的响应滞后于放牧活动.因此, 未来在评估干旱对生态系统的影响时, 应当充分考虑干旱和其他因素的综合效应. ...
Effects of livestock grazing on grassland carbon storage and release override impacts associated with global climate change 2 2019
... 过去大量模型和野外控制实验均表明干旱事件会通过改变“植物-土壤-微生物”系统之间的生物地球化学循环过程, 直接或间接地调控生态系统养分(如碳、氮、磷等)循环过程(Piao et al., 2009; 聂园园等, 2017).但是由于实验观测和模型模拟两种方法的目的不完全一致, 使得大量关于生态系统功能的实验数据很难被模型直接采用, 而模型的模拟结果也很少作为产生科学假说的工具来有效指导实验研究(Piao et al., 2009; Taylor et al., 2017).与此同时, 多模型间的相互比较研究和数据-模型间的比对研究结果均显示, 当前人们对生态系统功能与干旱事件相互反馈的认识存在极大的不确定性(Ciais et al., 2005; Bond-Lamberty et al., 2018).为了减小这种不确定性, 一方面有必要利用实验数据评估和改进生态系统模型在模拟和预测生态系统过程上的性能, 从而减小模型间的不确定性(Zhou et al., 2019b).另一方面, 对于导致模型间不确定性的关键生态系统过程, 需要建立起不同区域之间相互协作的多尺度和多层次实验网络, 同时在各个生态系统类型中获取具有统一标准、由统一方法获得的实验数据, 从而进一步约束模型进而达到有效生态预测(Taylor et al., 2017). ...
... 在自然生态系统中, 干旱事件并不是独立存在的, 往往伴随着其他气候因子(如增温、氮沉降)和人类活动(如放牧、森林砍伐)的同时发生(图5).过去的研究表明, 干旱与其他生物和非生物因素的综合效应, 对陆地生态系统的生物地球化学循环、植物生理生态、生物多样性和生态系统功能等方面都会产生重要影响.例如, Zhou等(2019a)的研究表明, 干旱能够显著降低土壤呼吸和生态系统呼吸, 但是这种负效应在一定程度上能够被增温所缓解.这可能是因为增温能够促进土壤氮的矿化作用, 增加土壤氮的可利用性, 进而缓解干旱导致的土壤氮含量降低(Melillo et al., 2011).与此同时, 干旱与氮沉降、增温、CO2浓度升高在调控草原生态系统呼吸过程中存在明显的交互作用, 这种作用主要以加和效应为主, 而非协同和拮抗作用(Zhou et al., 2019a).在草原生态系统中, 当干旱和放牧活动相伴发生时, 干旱对生态系统地上和地下碳过程的影响, 在一定程度上被放牧活动所主导(Zhou et al., 2019b).这种作用主要来源于植被生物量和地下养分资源对干旱的响应滞后于放牧活动.因此, 未来在评估干旱对生态系统的影响时, 应当充分考虑干旱和其他因素的综合效应. ...
Grazing intensity significantly affects belowground carbon and nitrogen cycling in grassland ecosystems: a meta-analysis 1 2017
 ,*华东师范大学全球变化与生态预测研究中心, 上海 200241
,*华东师范大学全球变化与生态预测研究中心, 上海 200241
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT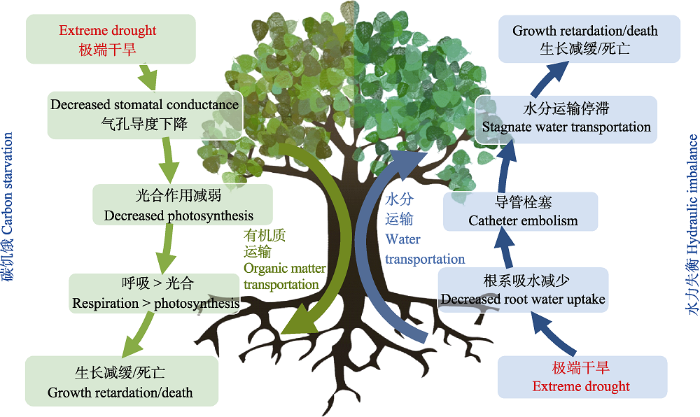 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT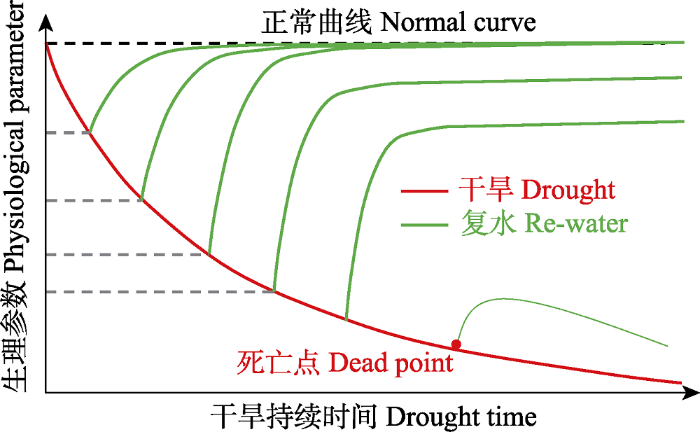 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT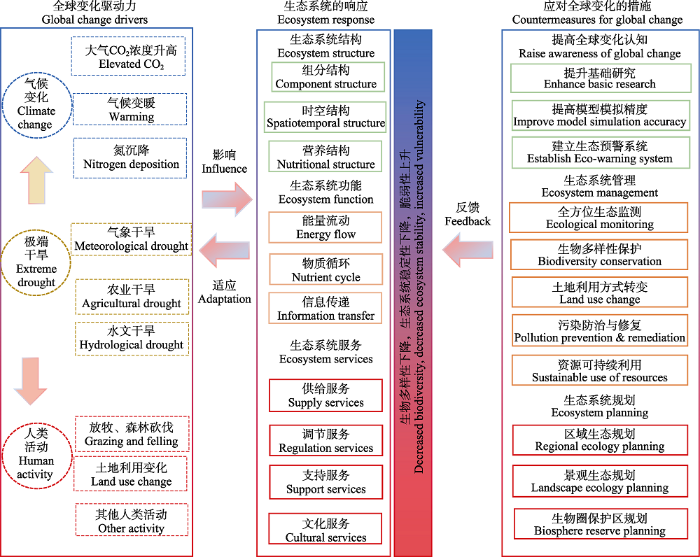 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}