Water isotope analysis for tracing ecosystem processes: measurement techniques, ecological applications, and future challenges
Xian-Hui TANG1, Yong-Le CHEN1,2, Fang LI1,2, Xin SONG,1,3,*1College of Life Sciences and Oceanography, Shenzhen University, Shenzhen, Guangdong 518060, China 2Key Laboratory of Optoelectronic Devices and Systems of Ministry of Education and Guangdong Province, College of Optoelectronic Engineering, Shenzhen University, Shenzhen, Guangdong 518060, China 3Shenzhen Key Laboratory of Marine Biotic Resources and Ecological Environment, Shenzhen University, Shenzhen, Guangdong 518060, China
Abstract Stable oxygen and hydrogen isotope analysis provides an important tool to trace, integrate or indicate water fluxes from leaf, whole-plant to ecosystem levels. Through measuring and analyzing the natural varitions in the hydrogen and oxygen isotope compositions of water from different components of ecosystem, we can partition evapotranspiration of ecosystem, determine source of plant water uptake, and study mechanism of leaf water isotope enrichment. As such, water isotope analysis has emerged as an indispensable technique to study the mechanism and ecological effects of different water cycle processes in ecosystem. In this paper, we briefly reviewed the history in development and application of water isotope analysis for terrestrial ecosystem studies, which then followed by more detailed introduction of the application principles and technical essentials. Furthermore, we reviewed progresses in diverse water-isotope based research field ranging from evapotranspiration partitioning, plant water uptake apportionment, sourcing of dew flux and precipitation vapor, to exploration leaf water isotope enrichment mechanisms and water-carbon coupling. Finally, we summarized technological and methodological challenges to be solved in the future ecological research, so as to fully realize the potential of water isotope analysis in various field of ecological research. Keywords:oxygen isotope;deuterium isotope;tracing;water cycle;ecosystem processes
PDF (1262KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 汤显辉, 陈永乐, 李芳, 宋欣. 水同位素分析与生态系统过程示踪: 技术、应用以及未来挑战. 植物生态学报, 2020, 44(4): 350-359. DOI: 10.17521/cjpe.2019.0204 TANG Xian-Hui, CHEN Yong-Le, LI Fang, SONG Xin. Water isotope analysis for tracing ecosystem processes: measurement techniques, ecological applications, and future challenges. Chinese Journal of Plant Ecology, 2020, 44(4): 350-359. DOI: 10.17521/cjpe.2019.0204
水分是生态系统的核心因子, 影响和驱动着个体的生长、发育和繁殖, 以及生态系统的组成、演化和物质循环, 对于维持生态系统的功能和稳定具有重要意义(Chapin et al., 2011)。然而由于人类活动增加, 全球水循环和水资源格局发生剧烈变化, 水循环过程机理研究对于缓解水资源短缺、调节水循环以及提高生态系统服务功能具有重要意义。降水、植物蒸腾、土壤和水体蒸发、凝露等影响大气和生态系统水汽交换的生态水文过程中, 伴有不同程度的同位素分馏效应, 记录了不同的生态过程信息, 使得水同位素分析成为研究生态系统水循环的重要手段, 其实质是通过测量和分析生态系统关键组分如土壤、植物茎叶或大气中所含水分的氧氢同位素比值(δ)来探究生态系统水循环的过程、机制及生态学效应。例如, 量化土壤蒸发和叶片蒸腾水汽的δ值可为估算生态系统蒸散发来源(即蒸散发拆分)提供实现路径(Wen et al., 2016); 比较分析植物茎干木质部水分与其潜在水源的同位素关系可用来量化植物对不同来源水分的利用比例(Dawson & Ehleringer, 1991); 对叶片水同位素富集机理的探究则为解析树木年轮或叶蜡烷烃中所记录的气候或生理生态过程信息提供了理论基础(Cernusak et al., 2016)。
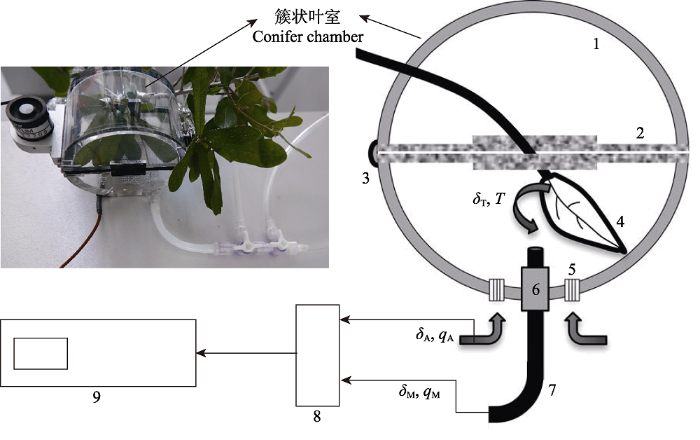
新窗口打开|下载原图ZIP|生成PPT 图1野外环境下簇状叶室与激光同位素仪相连测量蒸腾水汽同位素信号(δT)(引自Wang et al., 2012, 有改动)。实际测量中, 空气(水汽浓度为qA, 同位素信号为δA)以一定流速从叶室的进气口(5)进入叶室, 其与从叶室内叶片蒸腾出的水汽(T, δT)混合后(qM, δM)以同样的流速从叶室出气口(6)排出。进入叶室的空气以及排出叶室的混合气在经由电磁阀控制的多路控制系统(8)后按设定的测量周期(如4 min一个测量周期)交替通往水汽激光同位素仪(9)进行测量。在qA、δA、qM、δM以及气流流速都被测定的情况下, 通过进出叶室的水汽质量守恒可以计算出蒸腾通量, 再通过同位素质量守恒方程即可计算出δT。详细推导过程及注意事项参考Wang et al. (2012)。
Fig. 1Measurement of transpiration vapor isotopic signal (δT) by connecting conifer chamber with isotope ratio laser spectrometer in the field (cited from Wang et al., 2012 with change). In the measurement, the airs (vapor concentration qA and isotopic signal δA) enter the clustered chamber at a velocity from the inlet (5), mixing with the transpiration vapor from leaves (T, δT) in chamber, then (qM, δM) eject from the outlet (6) at the same flow rate. Passing by a solenoid valve controlled multi-channel system (8), the mixed airs are alternately connected to the isotope ratio laser spectrometer (9) in a set measurement period (e.g., a measurement period of 4 min). As qA, δA, qM, δA and flow rate are measured, the transpired vapor flux can be calculated by mass balance between vapor entering and outgoing the chamber, then δT of transpiration vapor can be calculated by isotopic mass balance equation. More details and derivational processes reference to Wang et al. (2012).
AemiseggerF, PfahlS, SodemannH, LehnerI, SeneviratneSI, WernliH (2014). Deuterium excess as a proxy for continental moisture recycling and plant transpiration Atmospheric Chemistry and Physics, 14, 4029-4054. [本文引用: 1]
AngertA, CappaCD, DePaoloDJ (2004). Kinetic 17O effects in the hydrologic cycle: indirect evidence and implications Geochimica et Cosmochimica Acta, 68, 3487-3495. [本文引用: 1]
BaoHM, CaoXB, HaylesJA (2016). Triple oxygen isotopes: fundamental relationships and applications Annual Review of Earth and Planetary Sciences, 44, 463-492. [本文引用: 1]
BarbourMM, EvansJR, SimoninKA, CaemmererS (2016). Online CO2 and H2O oxygen isotope fractionation allows estimation of mesophyll conductance in C4 plants, and reveals that mesophyll conductance decreases as leaves age in both C4 and C3 plants New Phytologist, 210, 875-889. DOI:10.1111/nph.13830URLPMID:26778088 [本文引用: 1] Mesophyll conductance significantly, and variably, limits photosynthesis but we currently have no reliable method of measurement for C4 plants. An online oxygen isotope technique was developed to allow quantification of mesophyll conductance in C4 plants and to provide an alternative estimate in C3 plants. The technique is compared to an established carbon isotope method in three C3 species. Mesophyll conductance of C4 species was similar to that in the C3 species measured, and declined in both C4 and C3 species as leaves aged from fully expanded to senescing. In cotton leaves, simultaneous measurement of carbon and oxygen isotope discrimination allowed the partitioning of total conductance to the chloroplasts into cell wall and plasma membrane versus chloroplast membrane components, if CO2 was assumed to be isotopically equilibrated with cytosolic water, and the partitioning remained stable with leaf age. The oxygen isotope technique allowed estimation of mesophyll conductance in C4 plants and, when combined with well-established carbon isotope techniques, may provide additional information on mesophyll conductance in C3 plants.
BarbourMM, FarquharGD (2000). Relative humidity-and ABA-induced variation in carbon and oxygen isotope ratios of cotton leaves Plant, Cell & Environment, 23, 473-485. [本文引用: 1]
BarkanE, LuzB (2005). High precision measurements of 17O/16O and 18O/16O ratios in H2O Rapid Communications in Mass Spectrometry, 19, 3737-3742. DOI:10.1002/rcm.2250URLPMID:16308852 [本文引用: 2] We have optimized the method of water fluorination using the solid reagent CoF3 to produce O2. This allows isotope ratio measurements by dual-inlet mass spectrometry with very high precision of 0.01 to 0.03/1000 for both delta17O and delta18O. Using this method, delta17O and delta18O of atmospheric O2 were determined as 12.08 and 23.88/1000 vs. VSMOW, respectively. Likewise, delta17O and delta18O of GISP were -13.12 and -24.73/1000, and for SLAP they were -29.48 and -55.11/1000 vs. VSMOW, respectively. Analysis of these data in a ln(delta17O + 1) vs. ln(delta18O + 1) plot yields a line with a regression coefficient (lambda) of 0.5279 +/- 0.0001 (R2 = 0.999999). We also determined the fractionation factors 17alpha and 18alpha in liquid-vapor equilibrium, and found that the ratio ln 17alpha/ln 18alpha is constant (0.529 +/- 0.001) over the temperature range 11.4 to 41.5 degrees C.
BeyerM, DubbertM (2019). X Water Worlds and how to investigate them: a review and future perspective on in situ measurements of water stable isotopes in soils and plants Hydrology and Earth System Sciences Discussions. DOI: 10.5194/hess-2019-600. [本文引用: 1]
BlunierT, BarnettB, BenderML, HendricksMB (2002). Biological oxygen productivity during the last 60,000 years from triple oxygen isotope measurements Global Biogeochemical Cycles, 16, 3. DOI: 10.1029/2001GB001460. [本文引用: 1]
CernusakLA, BarbourMM, ArndtSK, CheesmanAW, EnglishNB, FeildTS, HellikerBR, Holloway-PhillipsMM, HoltumJAM, KahmenA, McInerneyFA, MunksgaardNC, SimoninKA, SongX, Stuart-WilliamsH, WestJB, FarquharGD (2016). Stable isotopes in leaf water of terrestrial plants Plant, Cell & Environment, 39, 1087-1102. DOI:10.1111/pce.12703URLPMID:26715126 [本文引用: 1] Leaf water contains naturally occurring stable isotopes of oxygen and hydrogen in abundances that vary spatially and temporally. When sufficiently understood, these can be harnessed for a wide range of applications. Here, we review the current state of knowledge of stable isotope enrichment of leaf water, and its relevance for isotopic signals incorporated into plant organic matter and atmospheric gases. Models describing evaporative enrichment of leaf water have become increasingly complex over time, reflecting enhanced spatial and temporal resolution. We recommend that practitioners choose a model with a level of complexity suited to their application, and provide guidance. At the same time, there exists some lingering uncertainty about the biophysical processes relevant to patterns of isotopic enrichment in leaf water. An important goal for future research is to link observed variations in isotopic composition to specific anatomical and physiological features of leaves that reflect differences in hydraulic design. New measurement techniques are developing rapidly, enabling determinations of both transpired and leaf water delta(18) O and delta(2) H to be made more easily and at higher temporal resolution than previously possible. We expect these technological advances to spur new developments in our understanding of patterns of stable isotope fractionation in leaf water.
Chapin IIIFS, MatsonPA, VitousekPM (2011). Principles of Terrestrial Ecosystem Ecology Springer Science & Business Media, New York. [本文引用: 1]
CraigH, GordonLL (1965). Deuterium and oxygen-18 variations in the ocean and marine atmosphere//Tongiorgi E. Proceedings of a Conference on Stable Isotopes in Oceanographic Studies and Palaeotemperatures Spoleto, Italy. 9-130. [本文引用: 3]
DavieT (2008). Fundamentals of Hydrology. 2nd ed Routledge, London. [本文引用: 1]
DawsonTE, EhleringerJR (1991). Streamside trees that do not use stream water Nature, 350, 335-337. [本文引用: 2]
DongmannG, NürnbergHW, F?rstelH, WagenerK (1974). On the enrichment of H218O in the leaves of transpiring plants Radiation and Environmental Biophysics, 11, 41-52. DOI:10.1007/BF01323099URLPMID:4832051 [本文引用: 3]
DubbertM, WernerC (2019). Water fluxes mediated by vegetation: emerging isotopic insights at the soil and atmosphere interfaces New Phytologist, 221, 1754-1763. URLPMID:30341780 [本文引用: 1]
EhleringerJR, DawsonTE (1992). Water uptake by plants: perspectives from stable isotope composition Plant, Cell & Environment, 15, 1073-1082. [本文引用: 1]
FarquharGD, CernusakLA (2005). On the isotopic composition of leaf water in the non-steady state Functional Plant Biology, 32, 293-303. DOI:10.1071/FP04232URLPMID:32689132 [本文引用: 2] An expression is derived for the isotopic composition of water in leaves under conditions where the composition of water entering the leaf is not necessarily the same as that of water being transpired. The treatment is simplified and considers the average composition of the lamina and of the sites of evaporation. The concept of 'isostorage' is introduced as the product of leaf water content and the isotopic enrichment of leaf water above source water. It is shown that the rate of increase of isostorage is minus the 'isoflux' through the stomata, with the latter expressed as the product of the transpiration flux and the enrichment of the transpired water beyond source water. The approach of the isostorage to the steady state depends on the deviation of the isotopic enrichment of water at the evaporating sites from the steady value, and on the gross (one way) diffusive flux out of the leaf. To achieve model closure, it is assumed that the relationship between leaf water enrichment and that at the sites of evaporation depends on the radial Peclet number in the same manner as in the steady state. The equations have an analytical solution, and we also show how to calculate the results simply using a commonly available computer tool. The form of the equations emphasises that the one-way fluxes of water into and out of the stomata must sometimes be considered separately, rather than as a net outward flux. In this narrow sense we come to the interesting conclusion that more water usually enters the leaf from the air than from the roots.
FarquharGD, CernusakLA, BarnesB (2007). Heavy water fractionation during transpiration Plant Physiology, 143, 11-18. URLPMID:17210909 [本文引用: 1]
FarquharGD, LloydJ (1993). Carbon and Oxygen Isotope Effects in the Exchange of Carbon Dioxide Between Terrestrial Plants and the Atmosphere. Academic Press, San Diego, USA. 47-70. [本文引用: 1]
FarquharGD, LloydJ, TaylorJA, FlanaganLB, SyvertsenJP, HubickKT, WongSC, EhleringerJR (1993). Vegetation effects on the isotope composition of oxygen in atmospheric CO2 Nature, 363, 439-443. [本文引用: 3]
FarquharGD, RichardsRA (1984). Isotopic composition of plant carbon correlates with water-use efficiency of wheat genotypes Functional Plant Biology, 11, 539-552. [本文引用: 1]
FarrisF, StrainBR (1978). The effects of water-stress on leaf H218O enrichment Radiation and Environmental Biophysics, 15, 167-202. DOI:10.1007/BF01323264URLPMID:724981 [本文引用: 1] Water-stress experiments with Phaseolus vulgaris L. were undertaken to determine the transpiration rate dependency of the naturally occuring leaf H2(18O) fractionation process. Water-stress leaf H2(18O) levels were observed to be unexpectedly higher than controls. Speculations on the cause of this phenomenon are discussed. Since transpiration rate variations should theoretically affect only the rate and not the extent of leaf H2(18O) fractionation, the respective time courses for water-stressed and control leaf H2(18O) accumulations were compared. Water-stressed leaves displayed a slower rate of isotopic enrichment relative to controls, as was predicted from their reduced transpiration rate. In an absolute sense, however, both control and water-stress leaf H2(18O) fractionation rates were markedly greater than projected values from the existing model. Consequently, transpiration rates cannot be derived accurately at present from the observed rates of leaf H2(18O) discrimination. Several modifications of the theory are also considered.
FontesJC, CoqueR, DeverL, FillyA, MamouA (1983). Paléohydrologie isotopique de l'Oued el Akarit (sud tunisie) au Pléistocène supérieur et à l'Holocène Palaeogeography, Palaeoclimatology, Palaeoecology, 43, 41-62. [本文引用: 2]
FranceyRJ, TansPP (1987). Latitudinal variation in oxygen-18 of atmospheric CO2 Nature, 327, 495-497. [本文引用: 1]
GaoJ, Masson-DelmotteV, YaoT, TianL, RisiC, HoffmannG (2011). Precipitation water stable isotopes in the south Tibetan Plateau: observations and modeling Journal of Climate, 24, 3161-3178. [本文引用: 1]
GarrattJR, SegalM (1988). On the contribution of atmospheric moisture to dew formation Boundary-Layer Meteorology, 45, 209-236. [本文引用: 1]
GatJR (1981). Stable Isotope Hydrology: Deuterium and Oxygen-18 in the Water Cycle International Atomic Energy Agency, Vienna. [本文引用: 2]
GatJR (2000). Atmospheric water balance The isotopic perspective Hydrological Processes, 14, 1357-1369. [本文引用: 1]
GatJR, CarmiI (1987). Effect of climate changes on the precipitation patterns and isotopic composition of water in a climate transition zone: case of the Eastern Mediterranean Sea area //Sircoulon J. The Influence of Climate Change and Climatic Variability on the Hydrologic Regime and Water Resources. IAHS-AISH publication, Oxfordshire, UK. 168, 513-523. [本文引用: 1]
GenereuxD (1998). Quantifying uncertainty in tracer-based hydrograph separations Water Resources Research, 34, 915-919. [本文引用: 1]
Gerlein-SafdiC, GauthierPPG, SinklerCJ, CaylorKK (2017). Leaf water 18O and 2H maps show directional enrichment discrepancy in Colocasia esculenta Plant, Cell & Environment, 40, 2095-2108. URLPMID:28658718 [本文引用: 1]
GibsonJJ, ReidR (2014). Water balance along a chain of tundra lakes: a 20-year isotopic perspective Journal of Hydrology, 519, 2148-2164. [本文引用: 1]
GillonJS, YakirD (2001). Influence of carbonic anhydrase activity in terrestrial vegetation on the 18O content of atmospheric CO2 Science, 291, 2584-2587. URLPMID:11283366 [本文引用: 1]
GoodSP, SoderbergK, WangL, CaylorKK (2012). Uncertainties in the assessment of the isotopic composition of surface fluxes: a direct comparison of techniques using laser- based water vapor isotope analyzers Journal of Geophysical Research, 117, D15301. DOI: 10.1029/2011jd017168. [本文引用: 1]
GriffisTJ, BakerJM, SargentSD, TannerBD, ZhangJ (2004). Measuring field-scale isotopic CO2 fluxes with tunable diode laser absorption spectroscopy and micrometeorological techniques Agricultural and Forest Meteorology, 124, 15-29. [本文引用: 1]
GriffisTJ, LeeX, BakerJM, BillmarkK, SchultzN, EricksonM, ZhangX, FassbinderJ, XiaoW, HuN (2011). Oxygen isotope composition of evapotranspiration and its relation to C4 photosynthetic discrimination Journal of Geophysical Research, 116, G01035. DOI: 10.1029/2010JG001514. [本文引用: 1]
GriffisTJ, SargentSD, LeeX, BakerJM, GreeneJ, EricksonM, ZhangX, BillmarkK, SchultzN, XiaoW, HuN (2010). Determining the oxygen isotope composition of evapotranspiration using eddy covariance Boundary-Layer Meteorology, 137, 307-326. [本文引用: 1]
Holloway-PhillipsM, CernusakLA, BarbourM, SongX, CheesmanA, MunksgaardN, Stuart-WilliamsH, FarquharGD (2016). Leaf vein fraction influences the Péclet effect and 18O enrichment in leaf water Plant, Cell & Environment, 39, 2414-2427. DOI:10.1111/pce.12792URLPMID:27391079 [本文引用: 1] The process of evaporation results in the fractionation of water isotopes such that the lighter (16) O isotope preferentially escapes the gas phase leaving the heavier (18) O isotope to accumulate at the sites of evaporation. This applies to transpiration from a leaf with the degree of fractionation dependent on a number of environmental and physiological factors that are well understood. Nevertheless, the (18) O enrichment of bulk leaf water is often less than that predicted for the sites of evaporation. The advection of less enriched water in the transpiration stream has been suggested to limit the back diffusion of enriched evaporative site water (Peclet effect); however, evidence for this effect has been varied. In sampling across a range of species with different vein densities and saturated water contents, we demonstrate the importance of accounting for the relative 'pool' sizes of the vascular and mesophyll water for the interpretation of a Peclet effect. Further, we provide strong evidence for a Peclet signal within the xylem that if unaccounted for can lead to confounding of the estimated enrichment within the mesophyll water. This has important implications for understanding variation in the effective path length of the mesophyll and hence potentially the delta(18) O of organic matter.
HuZM, WenXF, SunXM, LiLH, YuGR, LeeXH, LiSG (2014). Partitioning of evapotranspiration through oxygen isotopic measurements of water pools and fluxes in a temperate grassland Journal of Geophysical Research, 119, 358-372. [本文引用: 1]
KeelingCD (1958). The concentration and isotopic abundances of atmospheric carbon dioxide in rural areas Geochimica et Cosmochimica Acta, 13, 322-334. [本文引用: 1]
KimK, LeeX (2011). Transition of stable isotope ratios of leaf water under simulated dew formation Plant, Cell & Environment, 34, 1790-1801. URLPMID:21675999 [本文引用: 1]
LandaisA, BarkanE, LuzB (2008). Record of δ18O and 17O-excess in ice from Vostok Antarctica during the last 150,000 years Geophysical Research Letters, 35, L02709. DOI: 10.1029/2007GL032096. [本文引用: 1]
LandaisA, BarkanE, YakirD, LuzB (2006). The triple isotopic composition of oxygen in leaf water Geochimica et Cosmochimica Acta, 70, 4105-4115. [本文引用: 1]
LandaisA, RisiC, BonyS, VimeuxF, DescroixL, FalourdS, BouyguesA (2010). Combined measurements of 17O-excess and d-excess in African monsoon precipitation: implications for evaluating convective parameterizations Earth and Planetary Science Letters, 298, 104-112. [本文引用: 1]
LeeX, KimK, SmithR (2007). Temporal variations of the 18O/ 16O signal of the whole-canopy transpiration in a temperate forest Global Biogeochemical Cycles, 21, GB3013. DOI: 10.1029/2006GB002871. [本文引用: 1]
LeeX, SargentS, SmithR, TannerB (2005). In situ measurement of the water vapor 18O/ 16O isotope ratio for atmospheric and ecological applications Journal of Atmospheric and Oceanic Technology, 22, 555-565. [本文引用: 1]
LiSN, LevinNE, SoderbergK, DennisKJ, CaylorKK (2017). Triple oxygen isotope composition of leaf waters in Mpala, central Kenya Earth and Planetary Science Letters, 468, 38-50. [本文引用: 1]
LinGH, SternbergLDSL (1993). Hydrogen isotopic fractionation by plant roots during water uptake in coastal wetland plants//Ebteringer JR, Hall AE, Farquhar GD. Stable Isotopes and Plant Carbon-Water Relations. Academic Press, San Diego, USA. 497-510. [本文引用: 1]
López DíasV, HoangHQ, Martínez-CarrerasN, BarnichF, WirtzT, McDonnellJJ, PfisterL (2019). The use of Nafion membranes to measure 2H/1H and 18O/16O isotopic ratios in water Journal of Membrane Science, 572, 128-139. [本文引用: 1]
LuzB, BarkanE (2000). Assessment of oceanic productivity with the triple-isotope composition of dissolved oxygen Science, 288, 2028-2031. DOI:10.1126/science.288.5473.2028URLPMID:10856212 [本文引用: 1] Plant production in the sea is a primary mechanism of global oxygen formation and carbon fixation. For this reason, and also because the ocean is a major sink for fossil fuel carbon dioxide, much attention has been given to estimating marine primary production. Here, we describe an approach for estimating production of photosynthetic oxygen, based on the isotopic composition of dissolved oxygen of seawater. This method allows the estimation of integrated oceanic productivity on a time scale of weeks.
LuzB, BarkanE, BenderML, ThiemensMH, BoeringKA (1999). Triple-isotope composition of atmospheric oxygen as a tracer of biosphere productivity Nature, 400, 547-550. [本文引用: 1]
MeijerHAJ, LiWJ (1998). The use of electrolysis for accurate δ17O and δ18O isotope measurements in water Isotopes in Environmental and Health Studies, 34, 349-369.
MerlivatL, JouzelJ (1979). Global climatic interpretation of the deuterium-oxygen 18 relationship for precipitation Journal of Geophysical Research, 84, 5029-5033. [本文引用: 2]
MooreJW, SemmensBX (2008). Incorporating uncertainty and prior information into stable isotope mixing models Ecology Letters, 11, 470-480. URLPMID:18294213 [本文引用: 1]
MoreiraMZ, SternbergLDSL, MartinelliLA, VictoriaRL, BarbosaEM, BonatesLCM, NepstadDC (1997). Contribution of transpiration to forest ambient vapour based on isotopic measurements Global Change Biology, 3, 439-450. [本文引用: 3]
NehemyMF, MillarC, JanzenK, GajM, PrattDL, LaroqueCP, McDonnellJJ (2019). 17O-excess as a detector for co-extracted organics in vapor analyses of plant isotope signatures Rapid Communications in Mass Spectrometry, 33, 1301-1310. URLPMID:31017711 [本文引用: 1]
PangHX, HeYQ, ZhangZL, LuAG, GuJ (2004). The origin of summer monsoon rainfall at New Delhi by deuterium excess Hydrology and Earth System Sciences, 8, 115-118. [本文引用: 1]
PengTR, LiuKK, WangCH, ChuangKH (2011). A water isotope approach to assessing moisture recycling in the island-based precipitation of Taiwan: a case study in the western Pacific Water Resources Research, 47, W08507. DOI: 10.1029/2010WR009890. [本文引用: 1]
RileyWJ, StillCJ, HellikerBR, Ribas-CarboM, BerryJA (2003). 18O composition of CO2 and H2O ecosystem pools and fluxes in a tallgrass prairie: simulations and comparisons to measurements Global Change Biology, 9, 1567-1581. [本文引用: 1]
RileyWJ, StillCJ, TornMS, BerryJA (2002). A mechanistic model of H218O and C 18OO fluxes between ecosystems and the atmosphere: model description and sensitivity analyses Global Biogeochemical Cycles, 16, 42. DOI: 10.1029/2002GB001878. [本文引用: 1]
RodenJS, LinG, EhleringerJR (2000). A mechanistic model for interpretation of hydrogen and oxygen isotope ratios in tree-ring cellulose Geochimica et Cosmochimica Acta, 64, 21-35. [本文引用: 1]
RothfussY, JavauxM (2017). Reviews and syntheses: isotopic approaches to quantify root water uptake: a review and comparison of methods Biogeosciences, 14, 2199-2224. [本文引用: 1]
RothfussY, VereeckenH, BrüggemannN (2013). Monitoring water stable isotopic composition in soils using gas-permeable tubing and infrared laser absorption spectroscopy Water Resources Research, 49, 3747-3755. [本文引用: 1]
ScheideggerY, SaurerM, BahnM, SiegwolfR (2000). Linking stable oxygen and carbon isotopes with stomatal conductance and photosynthetic capacity: a conceptual model Oecologia, 125, 350-357. DOI:10.1007/s004420000466URLPMID:28547329 [本文引用: 1] Based on measurements of delta(18)O and delta(13)C in organic matter of C3-plants, we have developed a conceptual model that gives insight into the relationship between stomatal conductance (g l) and photosynthetic capacity (A max) resulting from differing environmental constraints and plant-internal factors. This is a semi-quantitative approach to describing the long-term effects of environmental factors on CO2 and H2O gas exchange, whereby we estimate the intercellular CO2 concentration (c i) from delta(13)C and the air humidity from delta(18)O. Assuming that air humidity is an important factor influencing g l, the model allows us to distinguish whether differences in c i are caused by a response of g l or of A max. As an application of the model we evaluated the isotope data from three species in plots differing in intensity of land use (hay meadows and abandoned areas) at three sites along a south north transect in the Eastern Alps. We found three different delta(18)O-delta(13)C response patterns in native and planted grassland species (cultivated in the greenhouse). After preliminary confirmation by gas-exchange measurements we conclude that the proposed model is a promising tool for deriving carbon water relations in different functional groups from delta(18)O and delta(13)C isotope data.
ScrimgeourCM (1995). Measurement of plant and soil water isotope composition by direct equilibration methods Journal of Hydrology, 172, 261-274. [本文引用: 1]
SongX, LoucosKE, SimoninKA, FarquharGD, BarbourMM (2015a). Measurements of transpiration isotopologues and leaf water to assess enrichment models in cotton New Phytologist, 206, 637-646. URLPMID:25643590 [本文引用: 1]
SongX, SimoninKA, LoucosKE, BarbourMM (2015b). Modelling non-steady-state isotope enrichment of leaf water in a gas-exchange cuvette environment Plant, Cell & Environment, 38, 2618-2628. URLPMID:25993893 [本文引用: 1]
SwartPK, SternbergLDSL, SteinenR, HarrisonSA (1989). Controls on the oxygen and hydrogen isotopic composition of the waters of Florida Bay, USA Chemical Geology: Isotope Geoscience Section, 79, 113-123. [本文引用: 1]
UclésO, VillagarcíaL, MoroMJ, CantonY, DomingoF (2014). Role of dewfall in the water balance of a semiarid coastal steppe ecosystem Hydrological Processes, 28, 2271-2280. [本文引用: 1]
VolkmannTHM, KühnhammerK, HerbstrittB, GesslerA, WeilerM (2016). A method for in situ monitoring of the isotope composition of tree xylem water using laser spectroscopy Plant, Cell & Environment, 39, 2055-2063. DOI:10.1111/pce.12725URLPMID:27260852 [本文引用: 1] Field studies analyzing the stable isotope composition of xylem water are providing important information on ecosystem water relations. However, the capacity of stable isotopes to characterize the functioning of plants in their environment has not been fully explored because of methodological constraints on the extent and resolution at which samples could be collected and analysed. Here, we introduce an in situ method offering the potential to continuously monitor the stable isotope composition of tree xylem water via its vapour phase using a commercial laser-based isotope analyser and compact microporous probes installed into the xylem. Our technique enables efficient high-frequency measurement with intervals of only a few minutes per sample while eliminating the need for costly and cumbersome destructive collection of plant material and laboratory-based processing. We present field observations of xylem water hydrogen and oxygen isotope compositions obtained over several days including a labelled irrigation event and compare them against results from concurrent destructive sampling with cryogenic distillation and mass spectrometric analysis. The data demonstrate that temporal changes as well as spatial patterns of integration in xylem water isotope composition can be resolved through direct measurement. The new technique can therefore present a valuable tool to study the hydraulic architecture and water utilization of trees.
WangLX, GoodSP, CaylorKK, CernusakLA (2012). Direct quantification of leaf transpiration isotopic composition Agricultural and Forest Meteorology, 154, 127-135. [本文引用: 5]
WangP, LiXY, WangL, WuX, HuX, FanY, TongY (2018). Divergent evapotranspiration partition dynamics between shrubs and grasses in a shrub-encroached steppe ecosystem New Phytologist, 219, 1325-1337. DOI:10.1111/nph.15237URLPMID:29862515 [本文引用: 1] Previous evapotranspiration (ET) partitioning studies have usually neglected competitions and interactions between antagonistic plant functional types. This study investigated whether shrubs and grasses have divergent ET partition dynamics impacted by different water-use patterns, canopy structures, and physiological properties in a shrub-encroached steppe ecosystem in Inner Mongolia, China. The soil water-use patterns of shrubs and grasses have been quantified by an isotopic tracing approach and coupled into an improved multisource energy balance model to partition ET fluxes into soil evaporation, grass transpiration, and shrub transpiration. The mean fractional contributions to total ET were 24 +/- 13%, 20 +/- 4%, and 56 +/- 16% for shrub transpiration, grass transpiration, and soil evaporation respectively during the growing season. Difference in ecohydrological connectivity and leaf development both contributed to divergent transpiration partitioning between shrubs and grasses. Shrub-encroachment processes result in larger changes in the ET components than in total ET flux, which could be well explained by changes in canopy resistance, an ecosystem function dominated by the interaction of soil water-use patterns and ecosystem structure. The analyses presented here highlight the crucial effects of vegetation structural changes on the processes of land-atmosphere interaction and climate feedback.
WangSJ, ZhangMJ, CheYJ, ChenFL, QiangF (2016). Contribution of recycled moisture to precipitation in oases of arid central Asia: a stable isotope approach Water Resources Research, 52, 3246-3257. [本文引用: 1]
WangSJ, ZhangMJ, ChenFL, CheYJ, DuMX, LiuYM (2015). Comparison of GCM-simulated isotopic compositions of precipitation in arid central Asia Journal of Geographical Sciences, 25, 771-783. [本文引用: 1]
WassenaarLI, HendryMJ, ChostnerVL, LisGP (2008). High resolution pore water δ2H and δ18O measurements by H2O(liquid)-H2O(vapor) equilibration laser spectroscopy Environmental Science & Technology, 42, 9262-9267. DOI:10.1021/es802065sURLPMID:19174902 [本文引用: 1] A new H2O(liquid)-H2O(vapor) pore water equilibration and laser spectroscopy method provides a fast way to obtain accurate high resolution deltaD and delta18O profiles from single core samples from saturated and unsaturated geologic media. The precision and accuracy of the H2O(liquid)-H2O(vapor) equilibration method was comparable to or better than conventional IRMS-based methods, and it can be conducted on geologic cores that contain volumetric water contents as low as 5%. Significant advantages of the H2O(liquid)-H2O(vapor) pore water equilibration method and laser isotopic analysis method include dual hydrogen- and oxygen-isotope assays on single small core samples, low consumable and instrumentation costs, and the potential for field-based hydrogeologic profiling. A single core is sufficient to obtain detailed vertical isotopic depth profiles in geologic, soil, and lacustrine pore water, dramatically reducing the cost of obtaining pore water by conventional wells or physical water extraction methods. In addition, other inherent problems like contamination of wells by leakage and drilling fluids can be eliminated.
WelpLR, KeelingRF, MeijerHAJ, BollenbacherAF, PiperSC, YoshimuraK, FranceyRJ, AllisonCE, WahlenM (2011). Interannual variability in the oxygen isotopes of atmospheric CO2 driven by El Ni?o Nature, 477, 579-582. URLPMID:21956330 [本文引用: 1]
WenXF, LeeX, SunXM, WangJL, HuZM, LiSG, YuGR (2012). Dew water isotopic ratios and their relationships to ecosystem water pools and fluxes in a cropland and a grassland in China Oecologia, 168, 549-561. URLPMID:21822725 [本文引用: 3]
WenXF, YangB, SunXM, LeeX (2016). Evapotranspiration partitioning through in-situ oxygen isotope measurements in an oasis cropland Agricultural and Forest Meteorology, 230, 89-96. [本文引用: 1]
WershawRL, FriedmanI, HellerSJ, FrankPA (1970). Hydrogen isotopic fractionation of water passing through trees//Hobson GD, Speers GC. Advances in Organic Geochemistry. Pergamon Press, Oxford. 55-67. [本文引用: 1]
WhiteJWC, CookER, LawrenceJR, BroeckerWS (1985). The D/H ratios of sap in trees: implications for water sources and tree ring D/H ratios Geochimica et Cosmochimica Acta, 49, 237-246. [本文引用: 1]
XiaoW, LeeX, GriffisTJ, KimK, WelpLR, YuQ (2010). A modeling investigation of canopy-air oxygen isotopic exchange of water vapor and carbon dioxide in a soybean field Journal of Geophysical Research, 115, G01004. DOI: 10.1029/2009JG001163. [本文引用: 1]
XiaoW, LeeXH, WenXF, SunXM, ZhangSC (2012). Modeling biophysical controls on canopy foliage water 18O enrichment in wheat and corn Global Change Biology, 18, 1769-1780. [本文引用: 1]
XiaoW, WeiZW, WenXF (2018). Evapotranspiration partitioning at the ecosystem scale using the stable isotope method—A review Agricultural and Forest Meteorology, 263, 346-361. [本文引用: 1]
YakirD, SternbergLDSL (2000). The use of stable isotopes to study ecosystem gas exchange Oecologia, 123, 297-311. URLPMID:28308584 [本文引用: 1]
YamanakaT, ShimadaJ, MiyaokaK (2002). Footprint analysis using event-based isotope data for identifying source area of precipitated water Journal of Geophysical Research, 107, D22. DOI: 10.1029/2001JD001187. [本文引用: 1]
ZimmermannU, EhhaltD, MünnichKO (1967). Soil-water movement and evapotranspiration: changes in the isotopic composition of the water//Geophysics in Vienna. Proceedings of the Symposium of Isotopes in Hydrology International Atomic Energy Agency, Vienna. 567-585. [本文引用: 3]
Deuterium excess as a proxy for continental moisture recycling and plant transpiration 1 2014
Kinetic 17O effects in the hydrologic cycle: indirect evidence and implications 1 2004
... 氧元素在自然界有3种同位素, 由于17O自然丰度变异小且测量精度不够(林光辉, 2013), 以往17O在生态系统的研究应用很少.Meijer和Li (1998)通过电解水生成O2首次测量了液态水中的δ17O, 并认为由于δ17O和δ18O之间稳定的比例关系(0.528)而不能为生态系统提供更多有效信息.随着研究发现大气中O2的17O的非质量相关分馏可以用于光合作用和生物生产力的估算(Luz et al., 1999; Luz & Barkan, 2000; Blunier et al., 2002), d-excess可反演水源区温度和相对湿度信息(Merlivat & Jouzel, 1979; Gat, 1981), 17O-excess可用于恢复水源区相对湿度信息(Angert et al., 2004), 三氧同位素因为能提供更多的生态系统过程信息而在生态学研究中应用越来越广泛(Bao et al., 2016).目前水的δ17O的高精度测量主要是通过CoF3将水转化为O2, 再通过同位素比率质谱仪测量得到(Barkan & Luz, 2005), 也可以通过激光同位素仪实现测量和校准(Steig et al., 2014). ...
Triple oxygen isotopes: fundamental relationships and applications 1 2016
... 氧元素在自然界有3种同位素, 由于17O自然丰度变异小且测量精度不够(林光辉, 2013), 以往17O在生态系统的研究应用很少.Meijer和Li (1998)通过电解水生成O2首次测量了液态水中的δ17O, 并认为由于δ17O和δ18O之间稳定的比例关系(0.528)而不能为生态系统提供更多有效信息.随着研究发现大气中O2的17O的非质量相关分馏可以用于光合作用和生物生产力的估算(Luz et al., 1999; Luz & Barkan, 2000; Blunier et al., 2002), d-excess可反演水源区温度和相对湿度信息(Merlivat & Jouzel, 1979; Gat, 1981), 17O-excess可用于恢复水源区相对湿度信息(Angert et al., 2004), 三氧同位素因为能提供更多的生态系统过程信息而在生态学研究中应用越来越广泛(Bao et al., 2016).目前水的δ17O的高精度测量主要是通过CoF3将水转化为O2, 再通过同位素比率质谱仪测量得到(Barkan & Luz, 2005), 也可以通过激光同位素仪实现测量和校准(Steig et al., 2014). ...
Online CO2 and H2O oxygen isotope fractionation allows estimation of mesophyll conductance in C4 plants, and reveals that mesophyll conductance decreases as leaves age in both C4 and C3 plants 1 2016
X Water Worlds and how to investigate them: a review and future perspective on in situ measurements of water stable isotopes in soils and plants 1 2019
Deuterium and oxygen-18 variations in the ocean and marine atmosphere//Tongiorgi E. Proceedings of a Conference on Stable Isotopes in Oceanographic Studies and Palaeotemperatures 3 1965
... 自20世纪60年代以来, 水同位素理论和分析技术不断发展进步, 其在生态学中的应用大致可划分为以下三个不同阶段.第一阶段为水同位素研究的发轫和积累期(1966-1984年).在这一时期, 研究人员研发了低温真空抽提(Wershaw et al., 1970)水分提取技术, 为生态系统中不同“液态水”组分(如土壤和植物茎叶水)的测量清除了技术障碍.在此基础上, 围绕生态系统各组分水δ值的“普查式”测量揭示了以下重要的规律: 1)土壤垂直剖面存在明显的同位素梯度(Zimmermann et al., 1967; Fontes et al., 1983); 2)水分从根系进入植物茎干并在其木质部传输的过程中不存在同位素分馏(Zimmermann et al., 1967); 3)蒸腾导致叶片水的δ值显著高于茎干木质部水, 即叶片水的同位素相对富集(Craig & Gordon, 1965; Dongmann et al., 1974; Farris & Strain, 1978).第二阶段是对水同位素分馏机理认识不断深化的时期(1985-2004年)(Farquhar & Llyod, 1993; Roden et al., 2000), 也是基于同位素分析的生态学应用相继涌现的时期, 如植物水分来源区分(White et al., 1985; Dawson & Ehleringer, 1991), 蒸散发拆分(Moreira et al., 1997)等.第三阶段为水同位素测量技术的创新发展期(2005年至今).稳定同位素红外激光光谱技术的快速发展和推广使得同位素测量(特别是气态水的测量)变得前所未有地便捷(Lee et al., 2005), 这标志着“三高” (高频率、高通量、高精度)时代正式到来, 推动水同位素的研究从机理(Holloway-Phillips et al., 2016; Gerlein-Safdi et al., 2017)到应用(Wang et al., 2018; Dubbert & Werner, 2019)全方位走向繁荣.尽管水同位素分析在生态系统过程中已经有大量研究和进展, 但是水同位素测量技术进入创新发展期后, 对水同位素在生态系统过程中的应用、发展和未来挑战还缺乏系统论述. ...
... 氧元素在自然界有3种同位素, 由于17O自然丰度变异小且测量精度不够(林光辉, 2013), 以往17O在生态系统的研究应用很少.Meijer和Li (1998)通过电解水生成O2首次测量了液态水中的δ17O, 并认为由于δ17O和δ18O之间稳定的比例关系(0.528)而不能为生态系统提供更多有效信息.随着研究发现大气中O2的17O的非质量相关分馏可以用于光合作用和生物生产力的估算(Luz et al., 1999; Luz & Barkan, 2000; Blunier et al., 2002), d-excess可反演水源区温度和相对湿度信息(Merlivat & Jouzel, 1979; Gat, 1981), 17O-excess可用于恢复水源区相对湿度信息(Angert et al., 2004), 三氧同位素因为能提供更多的生态系统过程信息而在生态学研究中应用越来越广泛(Bao et al., 2016).目前水的δ17O的高精度测量主要是通过CoF3将水转化为O2, 再通过同位素比率质谱仪测量得到(Barkan & Luz, 2005), 也可以通过激光同位素仪实现测量和校准(Steig et al., 2014). ...
Atmospheric water balance The isotopic perspective 1 2000
... 同位素瑞利分馏模型可以示踪水汽来源(Dansgaard, 1964), 但是剩余水汽占气团的比例难以准确计算, 也没有考虑地表水体蒸发和云下二次蒸发对大气水汽δ值的影响, 只能模拟气团运行过程中分馏系数和气团内水汽含量变化等因素对降水δ值的影响(Gat, 2000).d-excess因为可以指示水汽源区相对湿度和温度等信息, 也被用于示踪水汽来源(Yamanaka et al., 2002; Pang et al., 2004).此外, δD和δ18O双同位素结合同位素质量平衡提供了更为简化的估算方法, Peng等(2011)研究认为, 假设降水水汽主要是土壤蒸发水汽、植物蒸腾水汽以及上风向平流输送的水汽混合后的结果, 如果3种来源的水汽混合均匀, 则可以建立一个三元混合模型, 通过上述蒸散发拆分中的方法可以确定土壤蒸发水汽的同位素值(δEv)和植物蒸腾水汽的同位素值(δTr), 降水的水汽同位素值(δPV)用降水同位素值经当地水汽蒸发线校准后计算得到(Gibson & Reid, 2014), 上风向平流输送的水汽同位素值(δadv)则可通过风向和两点之间的水汽同位素值得到(Wang et al., 2016).也有一些研究把降水和水汽同位素引入大气环流模型(Gao et al., 2011; Wang et al., 2015), 从更大尺度上确定水汽来源. ...
Effect of climate changes on the precipitation patterns and isotopic composition of water in a climate transition zone: case of the Eastern Mediterranean Sea area 1 1987
Uncertainties in the assessment of the isotopic composition of surface fluxes: a direct comparison of techniques using laser- based water vapor isotope analyzers 1 2012
... )(引自Wang et al., 2012, 有改动).实际测量中, 空气(水汽浓度为qA, 同位素信号为δA)以一定流速从叶室的进气口(5)进入叶室, 其与从叶室内叶片蒸腾出的水汽(T, δT)混合后(qM, δM)以同样的流速从叶室出气口(6)排出.进入叶室的空气以及排出叶室的混合气在经由电磁阀控制的多路控制系统(8)后按设定的测量周期(如4 min一个测量周期)交替通往水汽激光同位素仪(9)进行测量.在qA、δA、qM、δM以及气流流速都被测定的情况下, 通过进出叶室的水汽质量守恒可以计算出蒸腾通量, 再通过同位素质量守恒方程即可计算出δT.详细推导过程及注意事项参考Wang et al. (2012). Measurement of transpiration vapor isotopic signal (<i>δ</i><sub>T</sub>) by connecting conifer chamber with isotope ratio laser spectrometer in the field (cited from <xref ref-type="bibr" rid="b76">Wang <i>et al</i>., 2012</xref> with change). In the measurement, the airs (vapor concentration <i>q</i><sub>A</sub> and isotopic signal <i>δ</i><sub>A</sub>) enter the clustered chamber at a velocity from the inlet (5), mixing with the transpiration vapor from leaves (<i>T, δ</i><sub>T</sub>) in chamber, then (<i>q</i><sub>M</sub>, <i>δ</i><sub>M</sub>) eject from the outlet (6) at the same flow rate. Passing by a solenoid valve controlled multi-channel system (8), the mixed airs are alternately connected to the isotope ratio laser spectrometer (9) in a set measurement period (e.g., a measurement period of 4 min). As <i>q</i><sub>A</sub>, <i>δ</i><sub>A</sub>, <i>q</i><sub>M</sub>, <i>δ</i><sub>A</sub> and flow rate are measured, the transpired vapor flux can be calculated by mass balance between vapor entering and outgoing the chamber, then <i>δ</i><sub>T</sub> of transpiration vapor can be calculated by isotopic mass balance equation. More details and derivational processes reference to <xref ref-type="bibr" rid="b76">Wang <i>et al</i>. (2012)</xref>.Fig. 12.3 露水凝结来源拆分 露水主要是由低空近地层大气水汽、土壤蒸发水汽和植物蒸腾水汽在露点温度以下形成的凝露, 其同位素组成主要受3种来源δ值的影响, 但露水冷凝和蒸发的过程也会影响植物叶片水和大气水汽的同位素组成(Kim & Lee, 2011; Wen et al., 2012), 因此要实现三者拆分并不容易, Wen等(2012)研究认为, 低空近地层大气水汽通量向下, 而蒸散(土壤蒸发和植物蒸腾)的水汽通量向上, 可以通过通量廓线法得到生态系统净水汽通量和同位素值(δn), 通过露水与大气水汽的平衡分馏模型和冠层动力学分馏模型确定露水的同位素值(δd), 再用Craig-Gordon模型确定土壤蒸发和植物蒸腾的同位素值, 最后通过同位素质量守恒确定露水凝结不同来源的比例. ...
... .详细推导过程及注意事项参考Wang et al. (2012). Measurement of transpiration vapor isotopic signal (<i>δ</i><sub>T</sub>) by connecting conifer chamber with isotope ratio laser spectrometer in the field (cited from <xref ref-type="bibr" rid="b76">Wang <i>et al</i>., 2012</xref> with change). In the measurement, the airs (vapor concentration <i>q</i><sub>A</sub> and isotopic signal <i>δ</i><sub>A</sub>) enter the clustered chamber at a velocity from the inlet (5), mixing with the transpiration vapor from leaves (<i>T, δ</i><sub>T</sub>) in chamber, then (<i>q</i><sub>M</sub>, <i>δ</i><sub>M</sub>) eject from the outlet (6) at the same flow rate. Passing by a solenoid valve controlled multi-channel system (8), the mixed airs are alternately connected to the isotope ratio laser spectrometer (9) in a set measurement period (e.g., a measurement period of 4 min). As <i>q</i><sub>A</sub>, <i>δ</i><sub>A</sub>, <i>q</i><sub>M</sub>, <i>δ</i><sub>A</sub> and flow rate are measured, the transpired vapor flux can be calculated by mass balance between vapor entering and outgoing the chamber, then <i>δ</i><sub>T</sub> of transpiration vapor can be calculated by isotopic mass balance equation. More details and derivational processes reference to <xref ref-type="bibr" rid="b76">Wang <i>et al</i>. (2012)</xref>.Fig. 12.3 露水凝结来源拆分 露水主要是由低空近地层大气水汽、土壤蒸发水汽和植物蒸腾水汽在露点温度以下形成的凝露, 其同位素组成主要受3种来源δ值的影响, 但露水冷凝和蒸发的过程也会影响植物叶片水和大气水汽的同位素组成(Kim & Lee, 2011; Wen et al., 2012), 因此要实现三者拆分并不容易, Wen等(2012)研究认为, 低空近地层大气水汽通量向下, 而蒸散(土壤蒸发和植物蒸腾)的水汽通量向上, 可以通过通量廓线法得到生态系统净水汽通量和同位素值(δn), 通过露水与大气水汽的平衡分馏模型和冠层动力学分馏模型确定露水的同位素值(δd), 再用Craig-Gordon模型确定土壤蒸发和植物蒸腾的同位素值, 最后通过同位素质量守恒确定露水凝结不同来源的比例. ...
... ) by connecting conifer chamber with isotope ratio laser spectrometer in the field (cited from Wang et al., 2012 with change). In the measurement, the airs (vapor concentration qA and isotopic signal δA) enter the clustered chamber at a velocity from the inlet (5), mixing with the transpiration vapor from leaves (T, δT) in chamber, then (qM, δM) eject from the outlet (6) at the same flow rate. Passing by a solenoid valve controlled multi-channel system (8), the mixed airs are alternately connected to the isotope ratio laser spectrometer (9) in a set measurement period (e.g., a measurement period of 4 min). As qA, δA, qM, δA and flow rate are measured, the transpired vapor flux can be calculated by mass balance between vapor entering and outgoing the chamber, then δT of transpiration vapor can be calculated by isotopic mass balance equation. More details and derivational processes reference to Wang et al. (2012). Fig. 12.3 露水凝结来源拆分 露水主要是由低空近地层大气水汽、土壤蒸发水汽和植物蒸腾水汽在露点温度以下形成的凝露, 其同位素组成主要受3种来源δ值的影响, 但露水冷凝和蒸发的过程也会影响植物叶片水和大气水汽的同位素组成(Kim & Lee, 2011; Wen et al., 2012), 因此要实现三者拆分并不容易, Wen等(2012)研究认为, 低空近地层大气水汽通量向下, 而蒸散(土壤蒸发和植物蒸腾)的水汽通量向上, 可以通过通量廓线法得到生态系统净水汽通量和同位素值(δn), 通过露水与大气水汽的平衡分馏模型和冠层动力学分馏模型确定露水的同位素值(δd), 再用Craig-Gordon模型确定土壤蒸发和植物蒸腾的同位素值, 最后通过同位素质量守恒确定露水凝结不同来源的比例. ...
... of transpiration vapor can be calculated by isotopic mass balance equation. More details and derivational processes reference to Wang et al. (2012). Fig. 12.3 露水凝结来源拆分 露水主要是由低空近地层大气水汽、土壤蒸发水汽和植物蒸腾水汽在露点温度以下形成的凝露, 其同位素组成主要受3种来源δ值的影响, 但露水冷凝和蒸发的过程也会影响植物叶片水和大气水汽的同位素组成(Kim & Lee, 2011; Wen et al., 2012), 因此要实现三者拆分并不容易, Wen等(2012)研究认为, 低空近地层大气水汽通量向下, 而蒸散(土壤蒸发和植物蒸腾)的水汽通量向上, 可以通过通量廓线法得到生态系统净水汽通量和同位素值(δn), 通过露水与大气水汽的平衡分馏模型和冠层动力学分馏模型确定露水的同位素值(δd), 再用Craig-Gordon模型确定土壤蒸发和植物蒸腾的同位素值, 最后通过同位素质量守恒确定露水凝结不同来源的比例. ...
Divergent evapotranspiration partition dynamics between shrubs and grasses in a shrub-encroached steppe ecosystem 1 2018
... 自20世纪60年代以来, 水同位素理论和分析技术不断发展进步, 其在生态学中的应用大致可划分为以下三个不同阶段.第一阶段为水同位素研究的发轫和积累期(1966-1984年).在这一时期, 研究人员研发了低温真空抽提(Wershaw et al., 1970)水分提取技术, 为生态系统中不同“液态水”组分(如土壤和植物茎叶水)的测量清除了技术障碍.在此基础上, 围绕生态系统各组分水δ值的“普查式”测量揭示了以下重要的规律: 1)土壤垂直剖面存在明显的同位素梯度(Zimmermann et al., 1967; Fontes et al., 1983); 2)水分从根系进入植物茎干并在其木质部传输的过程中不存在同位素分馏(Zimmermann et al., 1967); 3)蒸腾导致叶片水的δ值显著高于茎干木质部水, 即叶片水的同位素相对富集(Craig & Gordon, 1965; Dongmann et al., 1974; Farris & Strain, 1978).第二阶段是对水同位素分馏机理认识不断深化的时期(1985-2004年)(Farquhar & Llyod, 1993; Roden et al., 2000), 也是基于同位素分析的生态学应用相继涌现的时期, 如植物水分来源区分(White et al., 1985; Dawson & Ehleringer, 1991), 蒸散发拆分(Moreira et al., 1997)等.第三阶段为水同位素测量技术的创新发展期(2005年至今).稳定同位素红外激光光谱技术的快速发展和推广使得同位素测量(特别是气态水的测量)变得前所未有地便捷(Lee et al., 2005), 这标志着“三高” (高频率、高通量、高精度)时代正式到来, 推动水同位素的研究从机理(Holloway-Phillips et al., 2016; Gerlein-Safdi et al., 2017)到应用(Wang et al., 2018; Dubbert & Werner, 2019)全方位走向繁荣.尽管水同位素分析在生态系统过程中已经有大量研究和进展, 但是水同位素测量技术进入创新发展期后, 对水同位素在生态系统过程中的应用、发展和未来挑战还缺乏系统论述. ...
Contribution of recycled moisture to precipitation in oases of arid central Asia: a stable isotope approach 1 2016
... 同位素瑞利分馏模型可以示踪水汽来源(Dansgaard, 1964), 但是剩余水汽占气团的比例难以准确计算, 也没有考虑地表水体蒸发和云下二次蒸发对大气水汽δ值的影响, 只能模拟气团运行过程中分馏系数和气团内水汽含量变化等因素对降水δ值的影响(Gat, 2000).d-excess因为可以指示水汽源区相对湿度和温度等信息, 也被用于示踪水汽来源(Yamanaka et al., 2002; Pang et al., 2004).此外, δD和δ18O双同位素结合同位素质量平衡提供了更为简化的估算方法, Peng等(2011)研究认为, 假设降水水汽主要是土壤蒸发水汽、植物蒸腾水汽以及上风向平流输送的水汽混合后的结果, 如果3种来源的水汽混合均匀, 则可以建立一个三元混合模型, 通过上述蒸散发拆分中的方法可以确定土壤蒸发水汽的同位素值(δEv)和植物蒸腾水汽的同位素值(δTr), 降水的水汽同位素值(δPV)用降水同位素值经当地水汽蒸发线校准后计算得到(Gibson & Reid, 2014), 上风向平流输送的水汽同位素值(δadv)则可通过风向和两点之间的水汽同位素值得到(Wang et al., 2016).也有一些研究把降水和水汽同位素引入大气环流模型(Gao et al., 2011; Wang et al., 2015), 从更大尺度上确定水汽来源. ...
Comparison of GCM-simulated isotopic compositions of precipitation in arid central Asia 1 2015
... 同位素瑞利分馏模型可以示踪水汽来源(Dansgaard, 1964), 但是剩余水汽占气团的比例难以准确计算, 也没有考虑地表水体蒸发和云下二次蒸发对大气水汽δ值的影响, 只能模拟气团运行过程中分馏系数和气团内水汽含量变化等因素对降水δ值的影响(Gat, 2000).d-excess因为可以指示水汽源区相对湿度和温度等信息, 也被用于示踪水汽来源(Yamanaka et al., 2002; Pang et al., 2004).此外, δD和δ18O双同位素结合同位素质量平衡提供了更为简化的估算方法, Peng等(2011)研究认为, 假设降水水汽主要是土壤蒸发水汽、植物蒸腾水汽以及上风向平流输送的水汽混合后的结果, 如果3种来源的水汽混合均匀, 则可以建立一个三元混合模型, 通过上述蒸散发拆分中的方法可以确定土壤蒸发水汽的同位素值(δEv)和植物蒸腾水汽的同位素值(δTr), 降水的水汽同位素值(δPV)用降水同位素值经当地水汽蒸发线校准后计算得到(Gibson & Reid, 2014), 上风向平流输送的水汽同位素值(δadv)则可通过风向和两点之间的水汽同位素值得到(Wang et al., 2016).也有一些研究把降水和水汽同位素引入大气环流模型(Gao et al., 2011; Wang et al., 2015), 从更大尺度上确定水汽来源. ...
High resolution pore water δ2H and δ18O measurements by H2O(liquid)-H2O(vapor) equilibration laser spectroscopy 1 2008
Footprint analysis using event-based isotope data for identifying source area of precipitated water 1 2002
... 同位素瑞利分馏模型可以示踪水汽来源(Dansgaard, 1964), 但是剩余水汽占气团的比例难以准确计算, 也没有考虑地表水体蒸发和云下二次蒸发对大气水汽δ值的影响, 只能模拟气团运行过程中分馏系数和气团内水汽含量变化等因素对降水δ值的影响(Gat, 2000).d-excess因为可以指示水汽源区相对湿度和温度等信息, 也被用于示踪水汽来源(Yamanaka et al., 2002; Pang et al., 2004).此外, δD和δ18O双同位素结合同位素质量平衡提供了更为简化的估算方法, Peng等(2011)研究认为, 假设降水水汽主要是土壤蒸发水汽、植物蒸腾水汽以及上风向平流输送的水汽混合后的结果, 如果3种来源的水汽混合均匀, 则可以建立一个三元混合模型, 通过上述蒸散发拆分中的方法可以确定土壤蒸发水汽的同位素值(δEv)和植物蒸腾水汽的同位素值(δTr), 降水的水汽同位素值(δPV)用降水同位素值经当地水汽蒸发线校准后计算得到(Gibson & Reid, 2014), 上风向平流输送的水汽同位素值(δadv)则可通过风向和两点之间的水汽同位素值得到(Wang et al., 2016).也有一些研究把降水和水汽同位素引入大气环流模型(Gao et al., 2011; Wang et al., 2015), 从更大尺度上确定水汽来源. ...
Soil-water movement and evapotranspiration: changes in the isotopic composition of the water//Geophysics in Vienna. Proceedings of the Symposium of Isotopes in Hydrology 3 1967
... 自20世纪60年代以来, 水同位素理论和分析技术不断发展进步, 其在生态学中的应用大致可划分为以下三个不同阶段.第一阶段为水同位素研究的发轫和积累期(1966-1984年).在这一时期, 研究人员研发了低温真空抽提(Wershaw et al., 1970)水分提取技术, 为生态系统中不同“液态水”组分(如土壤和植物茎叶水)的测量清除了技术障碍.在此基础上, 围绕生态系统各组分水δ值的“普查式”测量揭示了以下重要的规律: 1)土壤垂直剖面存在明显的同位素梯度(Zimmermann et al., 1967; Fontes et al., 1983); 2)水分从根系进入植物茎干并在其木质部传输的过程中不存在同位素分馏(Zimmermann et al., 1967); 3)蒸腾导致叶片水的δ值显著高于茎干木质部水, 即叶片水的同位素相对富集(Craig & Gordon, 1965; Dongmann et al., 1974; Farris & Strain, 1978).第二阶段是对水同位素分馏机理认识不断深化的时期(1985-2004年)(Farquhar & Llyod, 1993; Roden et al., 2000), 也是基于同位素分析的生态学应用相继涌现的时期, 如植物水分来源区分(White et al., 1985; Dawson & Ehleringer, 1991), 蒸散发拆分(Moreira et al., 1997)等.第三阶段为水同位素测量技术的创新发展期(2005年至今).稳定同位素红外激光光谱技术的快速发展和推广使得同位素测量(特别是气态水的测量)变得前所未有地便捷(Lee et al., 2005), 这标志着“三高” (高频率、高通量、高精度)时代正式到来, 推动水同位素的研究从机理(Holloway-Phillips et al., 2016; Gerlein-Safdi et al., 2017)到应用(Wang et al., 2018; Dubbert & Werner, 2019)全方位走向繁荣.尽管水同位素分析在生态系统过程中已经有大量研究和进展, 但是水同位素测量技术进入创新发展期后, 对水同位素在生态系统过程中的应用、发展和未来挑战还缺乏系统论述. ...
... ); 2)水分从根系进入植物茎干并在其木质部传输的过程中不存在同位素分馏(Zimmermann et al., 1967); 3)蒸腾导致叶片水的δ值显著高于茎干木质部水, 即叶片水的同位素相对富集(Craig & Gordon, 1965; Dongmann et al., 1974; Farris & Strain, 1978).第二阶段是对水同位素分馏机理认识不断深化的时期(1985-2004年)(Farquhar & Llyod, 1993; Roden et al., 2000), 也是基于同位素分析的生态学应用相继涌现的时期, 如植物水分来源区分(White et al., 1985; Dawson & Ehleringer, 1991), 蒸散发拆分(Moreira et al., 1997)等.第三阶段为水同位素测量技术的创新发展期(2005年至今).稳定同位素红外激光光谱技术的快速发展和推广使得同位素测量(特别是气态水的测量)变得前所未有地便捷(Lee et al., 2005), 这标志着“三高” (高频率、高通量、高精度)时代正式到来, 推动水同位素的研究从机理(Holloway-Phillips et al., 2016; Gerlein-Safdi et al., 2017)到应用(Wang et al., 2018; Dubbert & Werner, 2019)全方位走向繁荣.尽管水同位素分析在生态系统过程中已经有大量研究和进展, 但是水同位素测量技术进入创新发展期后, 对水同位素在生态系统过程中的应用、发展和未来挑战还缺乏系统论述. ...
 ,1,3,*
,1,3,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}