 ,*甘肃省白龙江林业管理局林业科学研究所, 兰州 730046; 甘肃白龙江森林生态系统国家定位观测研究站, 甘肃舟曲 746300
,*甘肃省白龙江林业管理局林业科学研究所, 兰州 730046; 甘肃白龙江森林生态系统国家定位观测研究站, 甘肃舟曲 746300Population structure characteristics of Picea purpurea in the upstream of Taohe River
Yang ZHAO, Jin-Qian LIU, Xue-Long CHEN, Meng-Meng Yang, Jia-Hao CAO, Rui QI, Xiu-Wen CAO,*Institute of Forestry Science of Bailongjiang in Gansu Province, Lanzhou 730046, China; and Gansu Bailongjiang National Forest Ecosystem Research Station, Zhouqu, Gansu 746300, China通讯作者: *gscaoxw@126.com
编委: 邓建明
责任编辑: 李敏
收稿日期:2019-04-8接受日期:2019-12-13网络出版日期:2020-03-20
| 基金资助: |
Corresponding authors: *gscaoxw@126.com
Received:2019-04-8Accepted:2019-12-13Online:2020-03-20
| Fund supported: |

摘要
该研究以洮河上游尕海-则岔自然保护区、卡车林区和冶力关林区的紫果云杉(Picea purpurea)天然种群为研究对象, 通过样地调查和数据统计, 绘制种群结构图, 编制静态生命表, 拟合并分析存活曲线, 运用数量化方法研究种群动态, 揭示种群生存现状, 预测种群发展趋势, 以期为该物种的保护、管理及结构恢复提供理论依据。结果显示: 3个林区紫果云杉种群自然更新能力强, 幼苗、幼树储量丰富, 幼小龄期死亡率普遍偏高; 尕海-则岔种群存活曲线符合Deevey-III型, 种群稳定结构完整, 卡车林区和冶力关林区种群存活曲线均符合Deevey-II型, 且均出现了局部衰退; 3个林区种群动态指数均大于0, 说明种群均属于增长型, 增长潜力为尕海-则岔>卡车>冶力关; 受随机干扰时卡车林区紫果云杉最敏感, 冶力关次之, 尕海-则岔种群最稳定。该研究表明: 竞争和自疏作用是造成紫果云杉幼小龄级个体存活率偏低的普遍因素, 3个林区不同的生存状况反映了紫果云杉种群在不同生境及生活史下生存能力的差异, 保护幼苗生存环境并提高幼苗质量和存活率是种群更新和发展的关键。尕海-则岔紫果云杉生存良好, 种群生存状况主要受自身生物学特性和环境因子的影响; 卡车林区主要受人为影响, 种群结构遭到破坏; 冶力关林区受分布限制, 造成种群结构不稳定, 须采取一定的人工措施来促进种群更新与增长。
关键词:
Abstract
Aims Natural population of Picea purpurea in Gahai-Zecha Nature Reserve and Kache and Yeliguan forest regions in the Taohe River upstream was selected, to reveal the demographic status of P. purpurea populations. The results from the current study will provide theoretical basis for the protection, management and structural restoration of P. purpurea.
Methods Based on field surveys and data analysis, population structure diagram was drawn, static life table was designed, and survival curve of P. purpurea was fitted and analyzed. Quantitative methods were used to study the population dynamics. The population development trend was predicted by time-sequence model.
Important findings In the three forest regions, P. purpurea population showed signs of strong natural regeneration ability with abundant seedling and sapling reserves, yet the mortality rate of saplings was high. The population in Gahai-Zecha forest manifested a Deevey-III type survival curve representing a stable population with a complete population structure, and the population in Kache forest and Yeliguan forest manifested Deevey-II type survival curves with partial declines present in both forests. With population dynamics indexes (Vpi) > 0, the populations of all three forests were in growth. The growth potentials ranking from highest to lowest were Gahai-Zecha > Kache > Yeliguan. The P. purpurea population in Kache forest showed the highest sensitivity to random interference followed by that in Yeliguan forest while Gahai-Zecha population was the most stable. This study showed that competition and self-thinning were the common causes of low survival rate of P. purpurea saplings and that different living situation in the three forests reflected the difference of survivability of P. purpurea populations in different habitats and life histories. The key to regeneration and development of P. purpurea populations was to protect the habitat of seedlings and to improve their quality and survival rate. The P. purpurea population in Gahai-Zecha forest grew well and the survivorship of that population was mainly affected by its own biological characteristics and environmental factors. The population structure of Kache forest was damaged under human interference. The population structure of Yeliguan forest was unstable due to its limited distribution. Appropriate anthropogenic interference measures should be adopted to promote population renewal and growth.
Keywords:
PDF (1139KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
赵阳, 刘锦乾, 陈学龙, 杨萌萌, 曹家豪, 齐瑞, 曹秀文. 洮河上游紫果云杉种群结构特征. 植物生态学报, 2020, 44(3): 266-276. DOI: 10.17521/cjpe.2019.0077
ZHAO Yang, LIU Jin-Qian, CHEN Xue-Long, Yang Meng-Meng, CAO Jia-Hao, QI Rui, CAO Xiu-Wen.
种群结构与动态是种群生态学研究的核心问题, 种群结构反映着种群不同年龄、大小个体的数量配置情况, 也反映着种群数量动态与变化趋势(Kang et al., 2014; 江波等, 2018)。种群动态是指种群大小或数量在时间和空间上的变化规律(滕毅等, 2017), 反映着种群与环境的关系(张亚芳等, 2015)。生命表及其构建的存活曲线可以反映种群当前的存活状态, 体现植物与环境的适合度(周资行等, 2011; Harcombe, 1987), 结合种群动态数量化分析和时间序列预测还可进一步阐明种群经历的生态学过程, 预测种群发展趋势(姜在民等, 2018)。
洮河是黄河的一级支流, 洮河林区是国家天然林资源保护工程重点实施区。该林区森林生态系统对维持洮河乃至黄河上游区域的生态平衡、遏制生态退化发挥着重要的屏障作用, 其森林生态服务功能总价值达到217.28亿元·a-1, 占甘肃全省的10%以上(邱书志等, 2018)。该林区自“天保工程”实施以来, 森林资源恢复良好, 且近年来随着国家对生态文明重视程度的提高, 森林生态恢复加快。然而该林区的森林资源究竟恢复到何种程度, 生存现状如何?近年来鲜有相关的报道。
紫果云杉(Picea purpurea)源于丽江云杉(P. likiangensis)和青扦(P. wilsonii)的自然同倍体杂交, 占据了青藏高原东北部高纬度高海拔极端生境(王婧如等, 2018), 喜阴湿, 耐寒, 幼林耐阴性强, 寿命可达400-500年, 是我国特有树种, 也是甘肃省重点保护植物, 在洮河林区主要分布在海拔2 800-3 600 m的阴坡、半阴坡及河谷地带。作为该地区重要的优势树种和森林建群种, 紫果云杉与云杉(P. asperata)、冷杉(Abies fabri)等混交形成顶极群落, 在白龙江、洮河乃至长江与黄河上游的水源涵养、水土保持及维持区域生态平衡方面发挥着重要作用。为此, 本研究以洮河上游3个林区的紫果云杉天然林种群为研究对象, 分析种群结构特征, 探究种群更新机制, 揭示种群生存现状, 以期为该物种及其群落的保护、管理及可持续经营提供理论依据。
1 材料和方法
1.1 研究区自然概况
本研究选在洮河上游尕海-则岔国家级自然保护区则岔沟、洮河自然保护区卡车林区和冶力关森林公园进行。研究区地处青藏高原东部边缘, 黄土高原与甘肃东南部山区交汇带(102.18°-103.98° E, 34.08°-35.16° N), 属青藏高原湿润气候和高原大陆性季风气候区, 年降水量634-680 mm, 年日照时间2 276-2 364 h, 年平均气温2.3-5.8 ℃。山岭陡峻, 海拔1 000-4 300 m, 水资源极为丰富。土壤多为棕、褐色森林土, 森林植被主要为暗针叶林, 并具有明显的垂直地带性和阴阳坡差异(许幼霞等, 2016)。森林群落主要建群种为云杉、冷杉、油松(Pinus tabuliformis)、落叶松(Larix gmelinii)等, 森林质量总体较高, 野生动植物资源丰富(邱书志等, 2018)。尕海-则岔林区(GZ)是我国少见的集森林资源、珍稀野生动植物资源、高原湿地和高原草甸等多种自然资源于一体的国家级自然保护区(赵阳等, 2018), 紫果云杉纯天然林结构完整, 林下更新良好; 卡车林区(KC)是以紫果云杉为建群种的云杉、冷杉混交林, 人为干扰较大; 冶力关林区(YLG)为紫果云杉分布稀疏, 并多与云杉、冷杉、青扦、桦木(Betula spp.)等形成的混交林。1.2 样地设置与调查
2018年8月, 在对研究区进行了多次踏查的基础上, 根据紫果云杉林的分布特点采用典型抽样法选择具有代表性的林分设置20 m × 20 m的标准样地各10块(GZ: 海拔3 154-3 510 m, 坡度21°-30°, 西南坡; KC: 海拔3 020-3 150 m, 坡度15°-24°, 西南坡; YLG: 海拔3 050-3 150 m, 坡度5°-15°, 西南坡)。采用相邻格子法以5 m × 5 m为基本单元, 记录样地内胸径(DBH) ≥ 5 cm的全部乔木树种的胸径、树高和冠幅等指标, 对DBH < 5 cm的乔木幼苗、幼树逐一测量并记录其地径、高度、冠幅。各样地用GPS定位, 记录其经纬度、海拔高度和郁闭度等因子。1.3 种群龄级的划分
根据同一树种龄级和径级对相同环境反应规律一致的特点(Forst & Rydin, 2000; Wu et al., 2010), 采用空间代替时间和大小结构分析法来研究紫果云杉种群结构。龄(径)级划分: 按胸径大小以5 cm为一级(表1), 即I龄级DBH < 5 cm, II龄级5 cm ≤ DBH < 10 cm, III龄级10 cm ≤ DBH < 15 cm, …, 以此类推。根据调查数据特点, GZ和YLG的紫果云杉各划为16个龄级, KC的划为14个龄级。Table 1
表1
表1洮河上游紫果云杉种群结构级划分
Table 1
| 径级指标 Diameter class indexes (cm) | 龄级 Age class | 生长期 Growing period |
|---|---|---|
| 0 < DBH < 5 | I | 幼苗、幼树 Seedling and sapling |
| 5 ≤ DBH < 10 | II | 小龄期 Small-age |
| 10 ≤ DBH < 15 | III | |
| 15 ≤ DBH < 20 | IV | |
| 20 ≤ DBH < 25 | V | 青年期 Youth-age |
| 25 ≤ DBH < 30 | VI | |
| 30 ≤ DBH < 35 | VII | |
| 35 ≤ DBH < 40 | VIII | |
| 40 ≤ DBH < 45 | IX | 中年期 Middle-age |
| 45 ≤ DBH < 50 | X | |
| 50 ≤ DBH < 55 | XI | |
| 55 ≤ DBH < 60 | XII | |
| 60 ≤ DBH < 65 | XIII | 老年期 Old-age |
| 65 ≤ DBH < 70 | XIV | |
| 70 ≤ DBH < 75 | XV | |
| 75 ≤ DBH | XVI |
新窗口打开|下载CSV
1.4 静态生命表的编制
静态生命表反映的是多个世代重叠的年龄动态历程中的一个特定时间, 因此, 自然种群生命表的编制中常出现死亡率为负的情况, 对此, 许多研究对所得数据进行匀滑, 但当种群各龄级个体数差别较大时, 匀滑后的结果往往掩盖种群的真实信息。以实际存活数编制生命表虽会出现死亡率为负值的情况, 但却能真实反映种群的生态学过程。因此, 本研究以径级对应龄级, 根据各龄级实际存活数编制静态生命表, 编制及计算方法参见张婕等(2014)。并以龄级为纵坐标, 对应个体数为横坐标, 绘制紫果云杉种群结构图。1.5 存活曲线拟合
以龄级为横坐标, 实际存活个体数Ax为纵坐标, 绘制紫果云杉种群存活曲线。采用指数函数和幂函数进行存活数和龄级关系的拟合, 依据决定系数、F检验值来判定模型拟合效果(Silvertown, 1982)。若指数方程Nx = N0e-bx拟合效果好, 存活曲线为Deevey-II型; 若幂函数Nx = N0x-b拟合效果好, 存活曲线为Deevey-III型。1.6 种群动态数量化分析
种群数量化分析是根据种群内两个相邻龄级间的个体数量变化动态指数(Vn)和整个种群年龄结构的数量变化动态指数(Vpi)及考虑未来外部干扰时种群年龄结构动态指数(V′pi)客观精确地定量比较评价种群结构动态(陈晓德, 1998)。计算公式如下:式中, Vn表示种群从n到n+1龄级的个体数量变化动态, Vpi表示整个种群结构的数量变化动态指数, Sn与Sn+1分别为第n与第n+1龄级的个体数, k表示种群大小级数量, max/min(…)取括号中数列极大/极小值, Vn∈[-1, 1], Vn取正、负、零值时分别反映种群两相邻龄级个体数(或整个种群年龄结构)增长、衰退、稳定的结构动态关系。Vpi仅适用于不考虑未来外部环境干扰的种群结构动态比较, 当考虑未来外部干扰时, 种群年龄结构动态(V′pi)还与龄级数量(k)及各龄级个体数(Sn)有关, 因此将式(2)修正为:
式中, V′pi也可作为衡量种群结构动态对随机干扰的敏感性指标。Vpi、V′pi取正、负、零值时反映的动态关系与Vn一致, P为种群对外界随机干扰所承担的风险概率, 只有当P值为最大时才对种群动态Vpi构成最大影响。
1.7 时间序列模型预测
采用时间序列分析中的一次平均推移法预测紫果云杉种群未来2、4、6、8个龄级的动态, 模型为:式中, n为需要预测的未来时间年限; t为龄级; $M_{t}^{1}$为未来n年时t龄级的种群存活数, 是近期n个观测值在t时刻的平均值, 称为第n周期的移动平均; Xk为当前k龄级种群存活数。
2 结果和分析
2.1 种群结构
GZ的紫果云杉共计1 871株, 最大胸径99.8 cm, I级个体数1 135, 占种群个体总数的60.63%, II级318株, 前2个龄级个体数之和占个体总数的77.62%, 前4个龄级个体数随龄级增大迅速减少, IV级之后, 数量变化趋于平缓; KC的紫果云杉共计794株, 最大胸径69.5 cm, I级个体数405, 占种群个体总数的51.01%, 前2个龄级个体数之和占个体总数的63.60%, 随后个体数量剧烈下降, VI级下降至27株, 仅占总数的3.4%, VI、VII级数量回升到总数的10%左右, IX级又下降至5株, 之后数量变化逐渐平缓, XIII级时仅为2株; YLG的紫果云杉共计811株, 最大胸径85.6 cm, I级个体数102, 占种群个体总数的34.28%, II级数量迅速减少至9.86%, 前2个龄级个体数之和占总数的44.14%, III级之后个体数量出现波动, VIII级时个体数达到第二个峰值, 占到总个体数的12.08%, 随后个体数又迅速减少, 到XII级后其比例均不到总数的1%。总体来看, 3个林区紫果云杉种群幼苗、幼树比例高(图1), GZ的个体数量随龄级的增大而减小, 种群结构呈典型的倒“J”形, KC和YLG林区中龄级个体数量有一定波动, 小中龄级个体数量不足, 但由于幼小龄级个体比例高, 因此总体上也属于增长型种群。图1
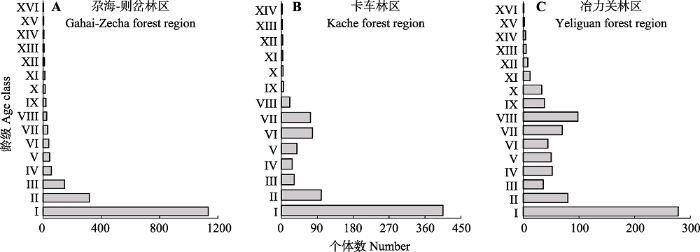 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1洮河上游紫果云杉种群龄级结构。龄级同
Fig. 1Age class structure of Picea purpurea populations in the upstream of Taohe River. Age class see
2.2 生命表与生存分析
2.2.1 生命表由表2可知, 紫果云杉种群不同龄级个体数差别大, 总体均表现为个体数随龄级增大而下降的变化趋势。I-II级时, 竞争、环境筛选等作用使幼苗、幼树大量死亡, 出现第一次死亡高峰(图2), 存活数量迅速下降, 生命期望值较小。从第III级开始, 随着龄级增大不同林区出现不同的变化。GZ的紫果云杉个体数量随龄级增大持续下降但下降趋势减小并在后期逐渐趋于稳定, 生命期望值在较长的龄级区间内(VI-X级)保持在较高水平, VII级后死亡率均低于30.0%, 并保持在相对稳定的水平。KC和YLG林区的小、中龄级个体的缺乏导致中龄林阶段死亡率均出现了负值, IV、V级后个体数量有所回升, 生命期望值增大。青中龄级时存活个体数量达到第二个高峰, 之后随着个体数量快速下降, 出现了第二次死亡高峰, 死亡率最高达到60%以上, 生命期望值减小, 后期死亡率仍有一定波动, 生命期望值较小。总体来看, 幼小龄级个体对种群死亡率贡献最大, 度过幼小龄级后, 随着林木个体对环境的适应, 不同生境林木个体的生存能力出现了显著差异。
Table 2
表2
表2洮河上游紫果云杉种群静态生命表
Table 2
| 不同林区 Different forest | 龄级 Age class | 龄级指标 Age class index (cm) | Ax | lx | lnlx | dx | qx | Lx | Tx | ex | Kx | Sx |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 尕海-则岔林区 Gahai-Zecha forest region | I | 0 < DBH < 5 | 1 135 | 1 000 | 6.908 | 720 | 0.720 | 640 | 1 148 | 1.148 | 1.272 | 0.280 |
| II | 5 ≤ DBH < 10 | 318 | 280 | 5.635 | 151 | 0.538 | 205 | 508 | 1.814 | 0.772 | 0.462 | |
| III | 10 ≤ DBH < 15 | 147 | 130 | 4.864 | 79 | 0.612 | 90 | 304 | 2.344 | 0.947 | 0.388 | |
| IV | 15 ≤ DBH < 20 | 57 | 50 | 3.916 | 11 | 0.211 | 45 | 214 | 4.254 | 0.236 | 0.789 | |
| V | 20 ≤ DBH < 25 | 45 | 40 | 3.680 | 5 | 0.133 | 37 | 169 | 4.256 | 0.143 | 0.867 | |
| VI | 25 ≤ DBH < 30 | 39 | 34 | 3.537 | 6 | 0.179 | 31 | 132 | 3.833 | 0.198 | 0.821 | |
| VII | 30 ≤ DBH < 35 | 32 | 28 | 3.339 | 7 | 0.250 | 25 | 100 | 3.563 | 0.288 | 0.750 | |
| VIII | 35 ≤ DBH < 40 | 24 | 21 | 3.051 | 4 | 0.208 | 19 | 76 | 3.583 | 0.234 | 0.792 | |
| IX | 40 ≤ DBH < 45 | 19 | 17 | 2.818 | 4 | 0.211 | 15 | 57 | 3.395 | 0.236 | 0.789 | |
| X | 45 ≤ DBH < 50 | 15 | 13 | 2.581 | 3 | 0.200 | 12 | 42 | 3.167 | 0.223 | 0.800 | |
| XI | 50 ≤ DBH < 55 | 12 | 11 | 2.358 | 3 | 0.250 | 9 | 30 | 2.833 | 0.288 | 0.750 | |
| XII | 55 ≤ DBH < 60 | 9 | 8 | 2.071 | 2 | 0.222 | 7 | 21 | 2.611 | 0.251 | 0.778 | |
| XIII | 60 ≤ DBH < 65 | 7 | 6 | 1.819 | 2 | 0.286 | 5 | 14 | 2.214 | 0.336 | 0.714 | |
| XIV | 65 ≤ DBH < 70 | 5 | 4 | 1.483 | 1 | 0.200 | 4 | 8 | 1.900 | 0.223 | 0.800 | |
| XV | 70 ≤ DBH < 75 | 4 | 4 | 1.260 | 1 | 0.250 | 3 | 4 | 1.250 | 0.288 | 0.750 | |
| XVI | 75 ≤ DBH | 3 | 3 | 0.972 | 3 | 1 | 1 | 0.500 | ||||
| 卡车林区 Kache forest region | I | 0 < DBH < 5 | 405 | 1 000 | 6.908 | 753 | 0.753 | 503 | 1 006 | 1.006 | 1.399 | 0.247 |
| II | 5 ≤ DBH < 10 | 100 | 247 | 5.509 | 168 | 0.680 | 126 | 503 | 2.036 | 1.139 | 0.320 | |
| III | 10 ≤ DBH < 15 | 32 | 79 | 4.370 | 12 | 0.156 | 42 | 377 | 4.766 | 0.170 | 0.844 | |
| IV | 15 ≤ DBH < 20 | 27 | 67 | 4.200 | -30 | -0.444 | 35 | 335 | 5.023 | -0.368 | 1.444 | |
| V | 20 ≤ DBH < 25 | 39 | 96 | 4.567 | -96 | -1.000 | 50 | 299 | 3.110 | -0.693 | 2.000 | |
| VI | 25 ≤ DBH < 30 | 78 | 193 | 5.261 | 12 | 0.064 | 99 | 249 | 1.293 | 0.066 | 0.936 | |
| VII | 30 ≤ DBH < 35 | 73 | 180 | 5.194 | 128 | 0.712 | 93 | 150 | 0.833 | 1.246 | 0.288 | |
| VIII | 35 ≤ DBH < 40 | 21 | 52 | 3.948 | 40 | 0.762 | 28 | 57 | 1.106 | 1.435 | 0.238 | |
| IX | 40 ≤ DBH < 45 | 5 | 12 | 2.513 | 2 | 0.200 | 7 | 29 | 2.386 | 0.223 | 0.800 | |
| X | 45 ≤ DBH < 50 | 4 | 10 | 2.290 | 2 | 0.250 | 6 | 22 | 2.230 | 0.288 | 0.750 | |
| XI | 50 ≤ DBH < 55 | 3 | 7 | 2.002 | 0 | 0.000 | 5 | 16 | 2.153 | 0.000 | 1.000 | |
| XII | 55 ≤ DBH < 60 | 3 | 7 | 2.002 | 2 | 0.333 | 5 | 11 | 1.517 | 0.405 | 0.667 | |
| XIII | 60 ≤ DBH < 65 | 2 | 5 | 1.597 | 0 | 0.000 | 3 | 7 | 1.323 | 0.000 | 1.000 | |
| XIV | 65 ≤ DBH | 2 | 5 | 1.597 | 5 | 3 | 3 | 0.662 | ||||
| 冶力关林区 Yeliguan forest region | I | 0 < DBH < 5 | 278 | 1 000 | 6.908 | 712 | 0.712 | 503 | 1 494 | 1.494 | 1.246 | 0.288 |
| II | 5 ≤ DBH < 10 | 80 | 288 | 5.662 | 158 | 0.550 | 147 | 990 | 3.441 | 0.799 | 0.450 | |
| III | 10 ≤ DBH < 15 | 36 | 129 | 4.864 | -58 | -0.444 | 67 | 843 | 6.513 | -0.368 | 1.444 | |
| IV | 15 ≤ DBH < 20 | 52 | 187 | 5.231 | 7 | 0.038 | 96 | 776 | 4.150 | 0.039 | 0.962 | |
| V | 20 ≤ DBH < 25 | 50 | 180 | 5.192 | 22 | 0.120 | 93 | 680 | 3.781 | 0.128 | 0.880 | |
| VI | 25 ≤ DBH < 30 | 44 | 158 | 5.064 | -94 | -0.591 | 82 | 588 | 3.713 | -0.464 | 1.591 | |
| VII | 30 ≤ DBH < 35 | 70 | 252 | 5.529 | -101 | -0.400 | 129 | 506 | 2.009 | -0.336 | 1.400 | |
| VIII | 35 ≤ DBH < 40 | 98 | 353 | 5.865 | 216 | 0.612 | 179 | 377 | 1.070 | 0.947 | 0.388 | |
| IX | 40 ≤ DBH < 45 | 38 | 137 | 4.918 | 18 | 0.132 | 71 | 198 | 1.449 | 0.141 | 0.868 | |
| X | 45 ≤ DBH < 50 | 33 | 119 | 4.777 | 76 | 0.636 | 62 | 127 | 1.072 | 1.012 | 0.364 | |
| XI | 50 ≤ DBH < 55 | 12 | 43 | 3.765 | 14 | 0.333 | 23 | 66 | 1.518 | 0.405 | 0.667 | |
| XII | 55 ≤ DBH < 60 | 8 | 29 | 3.360 | 11 | 0.375 | 16 | 42 | 1.461 | 0.470 | 0.625 | |
| XIII | 60 ≤ DBH < 65 | 5 | 18 | 2.890 | 4 | 0.200 | 10 | 26 | 1.445 | 0.223 | 0.800 | |
| XIV | 65 ≤ DBH < 70 | 4 | 14 | 2.666 | 7 | 0.500 | 9 | 16 | 1.081 | 0.693 | 0.500 | |
| XV | 70 ≤ DBH < 75 | 2 | 7 | 1.973 | 4 | 0.500 | 5 | 7 | 0.976 | 0.693 | 0.500 | |
| XVI | 75 ≤ DBH | 1 | 4 | 1.280 | 4 | 2 | 2 | 0.678 |
新窗口打开|下载CSV
图2
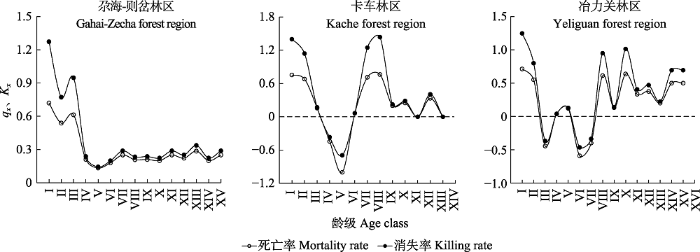 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2洮河上游紫果云杉种群死亡率(qx)和消失率(Kx)曲线。龄级同
Fig. 2Mortality rate (qx) and killing rate (Kx) curves of Picea purpurea populations in the upstream of Taohe River. Age class see
2.2.2 存活曲线由表3可知, GZ的幂函数模型R2和F值大于指数函数, 因此GZ的紫果云杉种群存活曲线符合Deevey-III型, 表明种群幼年死亡率高, 这与种群结构和生命表的分析结果一致。KC和YLG的指数函数模型R2和F值均大于幂函数模型, 因此, 其存活曲线更符合Deevey-II型(图3)。
Table 3
表3
表3洮河上游紫果云杉种群存活曲线方程
Table 3
| 不同林区 Different forest | 曲线方程 Survival curve | 存活曲线类型 Survival type |
|---|---|---|
| 尕海-则岔林区 Gahai-Zecha forest region | Y = 1052.654x -1.868, R2 = 0.989, F = 1 085.471, p < 0.000 1 | Deevey-III |
| Y = 407.192e -1.321x, R2 = 0.856, F = 71.083, p < 0.000 1 | ||
| 卡车林区 Kache forest region | Y = 538.798x -1.935, R2 = 0.773, F = 40.811, p < 0.000 1 | Deevey-II |
| Y = 271.407e -0.373x, R2 = 0.838, F = 61.952, p < 0.000 1 | ||
| 冶力关林区 Yeliguan forest region | Y = 290.048x -1.215, R2 = 0.618, F = 19.429, p < 0.001 | Deevey-II |
| Y = 205.059e -0.245x, R2 = 0.736, F = 33.405, p < 0.000 1 |
新窗口打开|下载CSV
图3
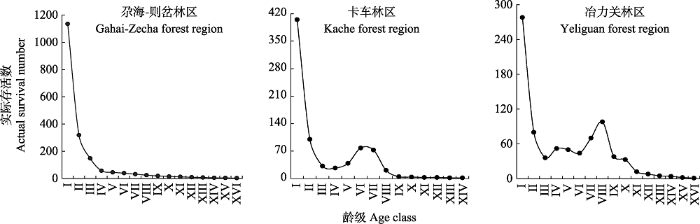 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3洮河上游紫果云杉种群存活曲线。龄级同
Fig. 3Survival curve of Picea purpurea populations in the upstream of Taohe River. Age class see
2.3 种群动态与时间序列预测
2.3.1 种群动态数量化分析不同林区紫果云杉种群相邻龄级数量变化动态指数随龄级的增大变化各异(表4)。GZ的Vn均大于0, 说明种群始终处于增长的结构动态。KC的V4-V5小于0, 说明在IV、V级及其向下一龄级过渡时种群呈衰退的结构动态, 其他龄级种群呈增长状态, 在XIII-XIV级时种群达到稳定。YLG在III-IV和VI-VIII级过渡时种群呈衰退的结构动态, 其他龄级间种群均表现为增长的结构动态。三个林区种群动态指数Vpi均大于0, 表明种群均为增长型, 增长潜力为Vpi (GZ) > Vpi (KC) > Vpi (YLG)。V′pi均大于0, 说明受外界随机干扰时, 种群也均表现为增长型, 随机干扰敏感性指数Pmax (KC) > Pmax (YLG) > Pmax (GZ), 说明对随机干扰敏感度最高的是KC林区, 其次为YLG, GZ敏感度最低, 即表明GZ的紫果云杉种群稳定性最高, KC受随机干扰的概率最大。
Table 4
表4
表4洮河上游紫果云杉种群动态变化指数
Table 4
| 龄级 Age class | 动态指数级 Dynamic index class | 动态指数 Dynamic index value (%) | ||
|---|---|---|---|---|
| 尕海-则岔林区 Gahai-Zecha forest region | 卡车林区 Kache forest region | 冶力关林区 Yeliguan forest region | ||
| I-II | V1 | 71.98 | 75.31 | 71.22 |
| II-III | V2 | 53.77 | 68.00 | 55.00 |
| III-IV | V3 | 61.22 | 15.63 | -30.77 |
| IV-V | V4 | 21.05 | -30.77 | 3.85 |
| V-VI | V5 | 13.33 | -50.00 | 12.00 |
| VI-VII | V6 | 17.95 | 6.41 | -37.14 |
| VII-VIII | V7 | 25.00 | 71.23 | -28.57 |
| VIII-IX | V8 | 20.83 | 76.19 | 61.22 |
| IX-X | V9 | 21.05 | 20.00 | 13.16 |
| X-XI | V10 | 20.00 | 25.00 | 63.64 |
| XI-XII | V11 | 25.00 | 0.00 | 33.33 |
| XII-XIII | V12 | 22.22 | 33.33 | 37.50 |
| XIII-XIV | V13 | 28.57 | 0.00 | 20.00 |
| XIV-XV | V14 | 20.00 | - | 50.00 |
| XV-XVI | V15 | 25.00 | - | 00.00 |
| Vpi | 60.50 | 53.68 | 36.94 | |
| V′pi | 2.91 | 1.92 | 1.15 | |
| Pmax | 2.08 | 3.57 | 3.13 | |
新窗口打开|下载CSV
2.3.2 时间序列预测
以各龄级实际存活数为基数, 按一次平均推移法预测各龄级在未来2、4、6、8个龄级后存活的个体数(表5)。未来2、4、6、8个龄级后, GZ的紫果云杉在VIII级之后各龄级个体数量均呈增加趋势, KC和YLG的种群在VIII-X级出现了局部衰退, 但总体上3个林区紫果云杉种群均呈增长趋势, 随着龄级增大幼苗数量优势逐渐减弱。到8个龄级后, 各种群个体数量增加的平均比例分别为: (GZ) 2.01倍、(KC) 1.53倍、(YLG) 1.25倍, 这与种群动态数量化分析结果一致。由此可见, 各龄级个体数的增加最终是以丰富的幼苗数量为基础, 幼苗比例越高, 增长潜力越大。由于紫果云杉均存在相对丰富的幼苗、幼树, 随着幼龄树木的生长, 中、成年个体数量逐渐得到补充而比例相对提高, 种群结构趋于合理。
Table 5
表5
表5洮河上游紫果云杉种群数量动态时间序列预测
Table 5
| 龄级 Age class | 尕海-则岔林区 Gahai-Zecha forest region | 卡车林区 Kache forest region | 冶力关林区 Yeliguan forest region | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M0 | M2 | M4 | M6 | M8 | M0 | M2 | M4 | M6 | M8 | M0 | M2 | M4 | M6 | M8 | |
| I | 1 135 | 405 | 278 | ||||||||||||
| II | 318 | 727 | 100 | 253 | 80 | 179 | |||||||||
| III | 147 | 233 | 32 | 66 | 36 | 58 | |||||||||
| IV | 57 | 102 | 334 | 27 | 30 | 141 | 52 | 44 | 112 | ||||||
| V | 45 | 51 | 142 | 39 | 33 | 50 | 50 | 51 | 55 | ||||||
| VI | 39 | 42 | 72 | 228 | 78 | 59 | 44 | 114 | 44 | 47 | 46 | 90 | |||
| VII | 32 | 36 | 43 | 106 | 73 | 76 | 54 | 58 | 70 | 57 | 54 | 55 | |||
| VIII | 24 | 28 | 35 | 57 | 178 | 21 | 47 | 53 | 45 | 97 | 98 | 84 | 66 | 58 | 89 |
| IX | 19 | 22 | 29 | 36 | 85 | 5 | 13 | 44 | 41 | 47 | 38 | 68 | 63 | 59 | 59 |
| X | 15 | 17 | 23 | 29 | 47 | 4 | 5 | 26 | 37 | 35 | 33 | 36 | 60 | 56 | 53 |
| XI | 12 | 14 | 18 | 24 | 30 | 3 | 4 | 8 | 31 | 31 | 12 | 23 | 45 | 49 | 50 |
| XII | 9 | 11 | 14 | 19 | 24 | 3 | 3 | 4 | 18 | 28 | 8 | 10 | 23 | 43 | 44 |
| XIII | 7 | 8 | 11 | 14 | 20 | 2 | 3 | 3 | 6 | 24 | 5 | 7 | 15 | 32 | 39 |
| XIV | 5 | 6 | 8 | 11 | 15 | 2 | 2 | 3 | 3 | 14 | 4 | 5 | 7 | 17 | 34 |
| XV | 4 | 5 | 6 | 9 | 12 | - | - | - | - | - | 2 | 3 | 5 | 11 | 25 |
| XVI | 3 | 4 | 5 | 7 | 9 | - | - | - | - | - | 2 | 2 | 3 | 5 | 13 |
新窗口打开|下载CSV
3 结论和讨论
3.1 种群结构特征
3个林区紫果云杉幼苗、幼树均占据极大部分, GZ幼苗比例最高, 占种群个体总数的一半以上, 达到60.73%, KC为51.01%, 比例最低的YLG也达到34.29%。大量幼苗、幼树的存在表明紫果云杉具有很强的自然更新能力, 为种群增长提供了丰富的物质基础(王立龙等, 2015)。幼苗阶段是森林更新和发展过程中最重要和敏感阶段(李晓笑等, 2011; 张婕等, 2014), 幼苗的留存情况往往决定着种群更新的成败(赵阳等, 2018)。3个林区幼苗留存率均不到其数量的30%, 这与长白山云杉、冷杉(李艳丽等, 2014)、雪岭杉(Picea schrenkiana)(李荣等, 2013)和高山松(Pinus densata)(卢杰等, 2013)等种群存在相同的生存特点, 说明幼苗时期为种群更新的瓶颈期。这主要是由幼龄个体的生态学特性以及生境条件共同造成的, 幼苗留存率长期过低造成了中成年林木比例低。此外, 虽然都属于增长型种群, 但相对于GZ个体数随龄级增大的逐级递减, KC和YLG的紫果云杉中小龄级数量不足, 种群结构不合理, 因此须在后期管理经营中对其结构进行调整。3.2 种群动态与发展趋势
结合静态生命表的分析结果可知, 3个林区紫果云杉幼龄期(DBH < 5 cm)死亡率均在70%以上, 幼龄期过后, 经环境筛选留存下的个体生存能力增强, 因此后续龄级死亡率逐渐降低。生命期望值总体均表现为随龄级增大先增大后减小的变化趋势, 幼小龄期迅速增大, 青中龄期时, GZ稳中有降, YLG和KC林区则出现较大波动, 中老龄后期均下降到较低水平, 反映了紫果云杉生命力随龄级变化的生存特征。生境不同造成生理旺盛期不同, 生命期望值大表明林木个体生理旺盛生存力强。GZ的生理旺盛期持续时间长, 表明近自然状态的GZ林区适合紫果云杉生存, KC和YLG的种群死亡率出现负值说明较小龄级筛选强度过大, 同时也证实了干扰存在的可能, 因此生理旺盛期较短。调查中发现, 虽然生境有所差异, 但自然更新的紫果云杉幼苗大都于大树树冠下集中分布, 因此幼龄阶段必会出现剧烈的竞争, 导致高死亡率出现。进入小龄级后, 林木个体需要一定空间和光照进行生长释放, 但是由于生长在大树之下, 受较低枝下高限制, 空间和光照不足进一步加剧了死亡, 造成幼小龄级普遍的高死亡率(赵阳等, 2018)。此外, 甘南地区冬季寒冷漫长, 高海拔地带极端低温可达-30 ℃以下, 造成幼苗越冬困难, 也会加剧苗木死亡。由此可见, 紫果云杉幼龄个体要进入青中年阶段, 必须经过竞争(空间、资源)、干扰、气候等因子多重筛选, 致使幼龄个体不能向下一级林木大量转化(于世川等, 2017), 形成了本研究区种群更新和发展的“瓶颈”, 限制了种群增长。KC和YLG的种群在中后期出现第二次死亡高峰则可能是干扰和生理衰退共同造成。存活曲线能够直观地表达种群的存活过程。李荣等(2013)认为幼苗、幼树具有绝对优势的种群其存活曲线接近Deevey-Ⅲ型, 宋于洋等(2009)认为幼苗期高死亡率的种群存活曲线接近Deevey-II型。本研究中3个林区种群存活曲线不尽相同, 反映了同类种群的生存过程随环境变化的特征。由此可见, 种群的存活曲线并非一成不变, 环境及外界干扰也影响种群生存状况(陈国鹏等, 2016), 生存状况不同进一步反映了紫果云杉在不同生境及生活史下生存能力的差异。
种群数量动态是种群个体生存能力与环境相互作用的结果(Bierzychudek, 2014), 时间序列分析能较准确的预测植物种群动态(卢杰等, 2013; 林勇等, 2017)。数量动态分析表明, GZ的种群始终处于增长状态, 进一步证明GZ适合紫果云杉生长, KC和YLG的种群出现局部衰退, 但未来2、4、6、8个龄级后, 种群个体数量均呈增加趋势, 可见, 由于幼苗基础的存在, 衰退现象会随着龄级的增大逐渐消失转而进入增长阶段。种群数量化动态值均为正进一步说明紫果云杉为增长型种群, 增长潜力为GZ > KC > YLG。此外, 种群数量动态受外界环境影响显著(周资行等, 2011), KC林区在IV-VI级, YLG在III-IV和VI-VIII级时种群结构逐渐稳定, 自疏作用减弱, 因此种群局部衰退必是由外界干扰引起。成年后的林木数量少但却占有较大资源空间, 因此少数个体死亡都将影响种群的动态走向, 但从种群整体发展角度来看, 老龄树消亡有利于小龄级生存力强的个体进入上层空间, 形成性能良好的循环系统(矢佳昱等, 2017), 有利于种群更新和发展。
时间序列分析虽能预测种群的发展趋势, 但其实现条件是以当前种群结构, 尤其是当前幼龄个体数量为基础。如果种群幼龄个体遭到破坏, 失去结构和功能完整性, 其增长趋势将因失去物质基础(幼苗、幼树)而在后续龄级出现衰退, 这对其种群发展趋势预测就失去了实际意义。但现实条件下种群完全不受干扰的状态是不存在的, 因此, 要保证实现种群增长, 必须加强保护, 尤其是幼小龄个体及其生存环境的保护。
3.3 种群生存现状与保护
紫果云杉作为亚高山地区的顶级群落和建群种, 对甘肃南部的生态平衡具有重要作用。本研究表明, 无论纯林还是混交林, 近自然状态还是存在干扰, 均存在一个共同问题就是幼龄个体留存率低, 而幼龄个体留存率长期过低势必影响种群发展。调查研究发现, GZ林区环境优势明显, 紫果云杉种群结构完整, 增长潜力大, 幼小林木存活主要受自身生物学特性和环境因子影响。KC林区除自身生物学特性外主要受人为影响, 由于邻近村庄, 人为活动频繁的林内常见伐桩与活倒木、树枝, 破坏了种群结构和生境, 造成了种群局部衰退, 林下环境的破坏从源头上阻碍了种群更新, 已萌生的幼苗失去有利的生存环境而死亡率过高, 限制了种群增长。YLG的高死亡率主要归咎于种群的分布, 稀疏、散生的林木失去集群效应而在竞争中始终处于劣势地位, 造成中小级林木比例低, 虽然幼苗有一定数量优势, 但如果其质量和存活率长期得不到提高, 种群局部衰退趋势还可能进一步扩大, 甚至有被其他优势树种淘汰的可能。鉴于此种现状, 必须针对各林区存在的主要问题采取目的性经营, 加强幼小林木的抚育和保护。对GZ林区须人工疏苗减小幼苗密度, 提高幼苗质量和存活率; 对KC林区则重点加强保护, 杜绝人为破坏, 清理“脏乱差”的林下空间, 为种群创造相对稳定的环境; YLG林区可通过人工补植使幼苗形成规模, 促进种群增长。同时对各林区的大树人工修枝增加枝下高, 伐除“老弱病残”的个体释放中上层空间, 促进林下“坐待”状态的小径级林木进入林冠层(周建云等, 2012)。同时, 巩固和保护好现有的种群结构及生境, 避免过度的人为干扰造成不可逆的破坏, 保持种群在最小干扰下生存发展。

参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
URLPMID:11662225 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1080/11956860.2000.11682569URL [本文引用: 1]
DOI:10.2307/1310666URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.chnaes.2017.02.003URL [本文引用: 1]
DOI:10.1016/j.chnaes.2017.02.003URL [本文引用: 1]
DOI:10.1038/srep06873URLPMID:25362993 [本文引用: 1]

This study investigated a typical secondary Betula platyphylla forest in the Ziwuling Mountains, Loess Plateau, China. In the sample plot, the DBH (diameter at breast height) class structure of B. platyphylla was bimodal. Individuals with small and large DBH values were abundant. The DBH structures of Quercus wutaishanica and Pinus tabulaeformis were close to that of the logistic model, thus suggesting the increasing population of these species. B. platyphylla and Populus davidiana showed random spatial distributions at almost all scales. However, Q. wutaishanica and P. tabulaeformis were significantly clumped at small scales. B. platyphylla had a negative spatial relation with Q. wutaishanica at small spatial scales. P. tabulaeformis and Q. wutaishanica showed negative spatial correlations at small scales, but they had positive correlations at large scales. These results suggest that P. tabulaeformis and Q. wutaishanica shared habitat preferences at these scales. In the future, the secondary B. platyphylla forest in the Ziwuling Mountains in the Loess Plateau will probably change into a multi-species mixed forest (Quercus-Pinus mixed forest). Assisted restoration strategies must be employed to improve the regeneration dynamics of the forest in the long term.
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.11707/j.1001-7488.20130822URL [本文引用: 2]
After a comprehensive investigation on Pinus densata populations that distribute in southeast Tibet by transects and quadrats, this paper analyzed the diameter class, height class and crown width class, compiled the static life table, drew survival curve, mortality curve, killing power curve, and four survival function curve (Survival rate, accumulated mortality rate, morality density rate and hazard rate), investigated population quantitative characteristics, and predicted population quantitative dynamics by a time sequence prediction model. The results showed that the individual number of P. densata was slight fluctuant with structure level (the diameter class, height class and crown width class), but the number in the three structures was generally presented as the anti-"J" type, and the population belonged to growing type. The survival number of every diameter class had obvious difference, the mortality rate of age Ⅳ was relatively low (13%), that of age Ⅷ was highest (70%), and the average mortality rate accounted for 42%. The survival curve of P. densata population approached to the B1 subtype of Deevey-Ⅱ. With the increase of the population age class, the mortality rate had a similar change trend to the killing power, and they all presented double-peak curves. Four survival function curve for P. densata population indicated that the population would have an early reduction, a mid-term stability, and a late recession because of physiological exhaustion. The time sequence model predicted that P. densata population had strong recovery capability, and the amount of population would have a different rising tendency in the upcoming 3, 6 and 9 age classes.
DOI:10.11707/j.1001-7488.20130822URL [本文引用: 2]
After a comprehensive investigation on Pinus densata populations that distribute in southeast Tibet by transects and quadrats, this paper analyzed the diameter class, height class and crown width class, compiled the static life table, drew survival curve, mortality curve, killing power curve, and four survival function curve (Survival rate, accumulated mortality rate, morality density rate and hazard rate), investigated population quantitative characteristics, and predicted population quantitative dynamics by a time sequence prediction model. The results showed that the individual number of P. densata was slight fluctuant with structure level (the diameter class, height class and crown width class), but the number in the three structures was generally presented as the anti-"J" type, and the population belonged to growing type. The survival number of every diameter class had obvious difference, the mortality rate of age Ⅳ was relatively low (13%), that of age Ⅷ was highest (70%), and the average mortality rate accounted for 42%. The survival curve of P. densata population approached to the B1 subtype of Deevey-Ⅱ. With the increase of the population age class, the mortality rate had a similar change trend to the killing power, and they all presented double-peak curves. Four survival function curve for P. densata population indicated that the population would have an early reduction, a mid-term stability, and a late recession because of physiological exhaustion. The time sequence model predicted that P. densata population had strong recovery capability, and the amount of population would have a different rising tendency in the upcoming 3, 6 and 9 age classes.
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
.
DOI:10.17521/cjpe.2015.0095URL [本文引用: 1]
DOI:10.17521/cjpe.2015.0095URL [本文引用: 1]
DOI:10.3724/SP.J.1227.2010.00250URL [本文引用: 1]
Populus euphratica Oliv. is widely distributed along the Tarim River. Maintaining stability of P. euphratica population is important to local development. This study explored the static life table, survivorship curves and four function curves (survival rate, cumulative mortality rate, mortality density, and hazard rate), and development index of P. euphratica population in the middle reaches of Tarim River. The results indicated that the age structure of P. euphratica population belonged to positive pyramidal type, which meant young age-class individuals occupied most populations. The number ofⅠ-Ⅱage classes accounted for 66.2% of whole population, and this indicated that there were abundant subsequent seedlings resources to support the growth of P. euphratica population in the middle reaches of Tarim River. The survivorship curve of P. euphratica belonged to the Deevey Ⅲ (concave-type) and the development index was 47.72%. Four function curves revealed that the individuals of P. euphratica sharply decreased at the initial stage and then leveled off at the late stage of survival curve. Time sequence prediction models predicted that the number of midlife individuals would increase in future 10, 20, 30 years, and P. euphratica population grew steadily as a result of rich saplings.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 2]
Using the plant survivorship theory, the age structure, and the relationship between tree height and diameter (DBH) of Quercus wutaishanica population in Lingkong Mountain were analyzed, and the static life table was compiled and the survival curve plotted. The shuttle shape in age structure of Q. wutaishanica population suggested its temporal stability. The linear regression significantly fitted the positive correlation between tree height and DBH. The maximal life expectancy was observed among the trees beyond the age of the highest mortality and coincided with the lowest point of mortality density, suggesting the strong vitality of the seedlings and young trees that survived in the natural selection and intraspecific competition. The population stability of the Q. wutaishanica population was characterized by the Deevey-Ⅱ of the survival curve. The dynamic pattern was characterized by the recession in the early phase, growth in the intermediate phase, and stability in the latter phase.
URL [本文引用: 2]
Using the plant survivorship theory, the age structure, and the relationship between tree height and diameter (DBH) of Quercus wutaishanica population in Lingkong Mountain were analyzed, and the static life table was compiled and the survival curve plotted. The shuttle shape in age structure of Q. wutaishanica population suggested its temporal stability. The linear regression significantly fitted the positive correlation between tree height and DBH. The maximal life expectancy was observed among the trees beyond the age of the highest mortality and coincided with the lowest point of mortality density, suggesting the strong vitality of the seedlings and young trees that survived in the natural selection and intraspecific competition. The population stability of the Q. wutaishanica population was characterized by the Deevey-Ⅱ of the survival curve. The dynamic pattern was characterized by the recession in the early phase, growth in the intermediate phase, and stability in the latter phase.
DOI:10.11707/j.1001-7488.20150201URL [本文引用: 1]
【Objective】Population structure and dynamics of plant can reflect the survival of the population and display the interaction between plant and environment. It is an important content of population ecology, and it has significant implications to protection and utilization of plant resources. Juniperus rigida is a wild tree species with priority for protection in Loess Plateau, a comparative study on structure and dynamics of Juniperus rigida populations in different regions of Loess Plateau was carried out to analyze population survival and dynamics of Juniperus rigida in Loess Plateau, in order to reveal the trend of population development. This study would have important theoretical and practical significance for developing strategies of rehabilitating Juniperus rigida population.【Method】A field survey was carried out on 5 populations of Juniperus rigida selected in 5 different regions in Loess Plateau (Wuchuan, Inner Mongolia; Zhuolu, Hebei; Hunyuan, Shanxi; Fugu, Shaanxi; Helanshan, Ningxia) by using sample plots. According to the characteristics of life history of the species and considering results from previous studies, diameter, height and crown width were divided into classes and population structure was constructed using the classes. Static life table, survival curve, mortality curve, disappearance rate curve were produced using space to replace time, i.e. diameter structure instead of age structure. 4 functions (survival, accumulative mortality, mortality density, and hazard rate) of survival analysis were used to analyze population dynamics of Juniperus rigida.【Result】 Different extents of absence of some diameter classes was found in diameter structure of 5 populations; Height structures of Zhuolu and Hunyuan populations were relatively complete; Certain extent of absence of crown-width classes were found in the 5 populations, the crown coverage of Wuchuan, Zhuolu and Hunyuan populations were mostly within 2 m2, while Helanshan population was mostly larger than 3 m2. The static life table showed that there were large differences of number of survival trees among different age classes, the number of survival trees decreases with age; the highest rate of mortality and disappearance rate were both in age class IV; and the highest life expectancy was in age class I. Two types of the survival curves of 5 populations were found, Wuchuan fitted Deevey-III while the others tended fit Deevey-II. The changes of mortality and disappearance rate were in similar trends for the 5 populations. Four survival curves of 5 populations showed that, the populations Wuchuan and Fugu reduced at young ages, grew stably at middle ages and declined at old ages, and Fugu showed a sharper reduction, while the populations Zhuolu and Hunyuan had a steep reduction in early growth and a late recession, and the population Helanshan reduced slightly at young ages and recessed late.【Conclusion】There were some differences among 5 populations of different regions in the structures of diameter class, height class and crown width class. Populations of Zhuolu and Hunyuan showed regeneration potential as the population density of them were relatively large and they also had a certain number of young plants; Fugu population had a considerable number of aging plant; the diameter class structures of Fugu, Wuchuan and Helanshan were relatively complete, while the population density were smaller and young plants were comparatively rare. There were environmental screenings and competitive pressures in the process of young trees growing up into big trees, and the survival ability of the big trees were stronger. The populations Zhuolu and Hunyuan could be restored and updated if the ecological environment could be improved as there were some young trees in Zhuolu and Hunyuan populations; the regeneration and development of Fugu population lacking young trees would be limited without the supplementation of young trees. In order to protect the populations of Juniperus rigida, attentions should be paid to protecting the young trees; and measures should be taken to promote germination as it was low under natural conditions; For populations with serious aging such as Fugu population, supplementary plantings should be used to ensure regeneration and development of the populations.
DOI:10.11707/j.1001-7488.20150201URL [本文引用: 1]
【Objective】Population structure and dynamics of plant can reflect the survival of the population and display the interaction between plant and environment. It is an important content of population ecology, and it has significant implications to protection and utilization of plant resources. Juniperus rigida is a wild tree species with priority for protection in Loess Plateau, a comparative study on structure and dynamics of Juniperus rigida populations in different regions of Loess Plateau was carried out to analyze population survival and dynamics of Juniperus rigida in Loess Plateau, in order to reveal the trend of population development. This study would have important theoretical and practical significance for developing strategies of rehabilitating Juniperus rigida population.【Method】A field survey was carried out on 5 populations of Juniperus rigida selected in 5 different regions in Loess Plateau (Wuchuan, Inner Mongolia; Zhuolu, Hebei; Hunyuan, Shanxi; Fugu, Shaanxi; Helanshan, Ningxia) by using sample plots. According to the characteristics of life history of the species and considering results from previous studies, diameter, height and crown width were divided into classes and population structure was constructed using the classes. Static life table, survival curve, mortality curve, disappearance rate curve were produced using space to replace time, i.e. diameter structure instead of age structure. 4 functions (survival, accumulative mortality, mortality density, and hazard rate) of survival analysis were used to analyze population dynamics of Juniperus rigida.【Result】 Different extents of absence of some diameter classes was found in diameter structure of 5 populations; Height structures of Zhuolu and Hunyuan populations were relatively complete; Certain extent of absence of crown-width classes were found in the 5 populations, the crown coverage of Wuchuan, Zhuolu and Hunyuan populations were mostly within 2 m2, while Helanshan population was mostly larger than 3 m2. The static life table showed that there were large differences of number of survival trees among different age classes, the number of survival trees decreases with age; the highest rate of mortality and disappearance rate were both in age class IV; and the highest life expectancy was in age class I. Two types of the survival curves of 5 populations were found, Wuchuan fitted Deevey-III while the others tended fit Deevey-II. The changes of mortality and disappearance rate were in similar trends for the 5 populations. Four survival curves of 5 populations showed that, the populations Wuchuan and Fugu reduced at young ages, grew stably at middle ages and declined at old ages, and Fugu showed a sharper reduction, while the populations Zhuolu and Hunyuan had a steep reduction in early growth and a late recession, and the population Helanshan reduced slightly at young ages and recessed late.【Conclusion】There were some differences among 5 populations of different regions in the structures of diameter class, height class and crown width class. Populations of Zhuolu and Hunyuan showed regeneration potential as the population density of them were relatively large and they also had a certain number of young plants; Fugu population had a considerable number of aging plant; the diameter class structures of Fugu, Wuchuan and Helanshan were relatively complete, while the population density were smaller and young plants were comparatively rare. There were environmental screenings and competitive pressures in the process of young trees growing up into big trees, and the survival ability of the big trees were stronger. The populations Zhuolu and Hunyuan could be restored and updated if the ecological environment could be improved as there were some young trees in Zhuolu and Hunyuan populations; the regeneration and development of Fugu population lacking young trees would be limited without the supplementation of young trees. In order to protect the populations of Juniperus rigida, attentions should be paid to protecting the young trees; and measures should be taken to promote germination as it was low under natural conditions; For populations with serious aging such as Fugu population, supplementary plantings should be used to ensure regeneration and development of the populations.
[本文引用: 3]
[本文引用: 3]
DOI:10.11707/j.1001-7488.20120425URL [本文引用: 1]
An investigation was conducted in plots of the Quercus wutaishanica forests in the Huanglong Mountain of China. The forests were undertaken the close-to-nature management thinning of 13.4% (light thinning) and 30.0% (heavy thinning). An unthinned forest served as the control. The life table, age structure, survivorship curve and spatial distribution were studied in 6 years after the thinning. The results were as follows. 1) The age structure of the population exhibited that seedlings and saplings were predominated, while the young trees were the scarcest. 2) Population mortality rate was the highest inⅠ~Ⅲ age class. With age increasing, the mortality displayed a "high-low-high" mode, while the expectancy life of populations displayed a "low-high-low" mode. 3) The survivorship curve of populations generally matched the Deevey Type Ⅲ. 4) With sampling dimension increasing, the spatial distribution transformed from clumped or random to clump distribution. 5) As the population grew older, their distribution transformed from the clumped into a random distribution while their clumped intensities reduced. 6) Compared with the control, the light and heavy thinning promoted the number of the juveniles and young trees, and population structure was optimized. Therefore, the secondary Q. wutaishanica forest reaches thinning period. Under natural circumstances, the highest clumped intensity at the 25~50 m2 was the optimal area for the population regeneration. The close-to-nature management thinning 30.0% was more suitable to the management.
DOI:10.11707/j.1001-7488.20120425URL [本文引用: 1]
An investigation was conducted in plots of the Quercus wutaishanica forests in the Huanglong Mountain of China. The forests were undertaken the close-to-nature management thinning of 13.4% (light thinning) and 30.0% (heavy thinning). An unthinned forest served as the control. The life table, age structure, survivorship curve and spatial distribution were studied in 6 years after the thinning. The results were as follows. 1) The age structure of the population exhibited that seedlings and saplings were predominated, while the young trees were the scarcest. 2) Population mortality rate was the highest inⅠ~Ⅲ age class. With age increasing, the mortality displayed a "high-low-high" mode, while the expectancy life of populations displayed a "low-high-low" mode. 3) The survivorship curve of populations generally matched the Deevey Type Ⅲ. 4) With sampling dimension increasing, the spatial distribution transformed from clumped or random to clump distribution. 5) As the population grew older, their distribution transformed from the clumped into a random distribution while their clumped intensities reduced. 6) Compared with the control, the light and heavy thinning promoted the number of the juveniles and young trees, and population structure was optimized. Therefore, the secondary Q. wutaishanica forest reaches thinning period. Under natural circumstances, the highest clumped intensity at the 25~50 m2 was the optimal area for the population regeneration. The close-to-nature management thinning 30.0% was more suitable to the management.
DOI:10.11849/zrzyxb.2011.10.009URL [本文引用: 2]
The quantity dynamics of the natural Reaumuria soongorica populations from different habitats in the south-north hills in Lanzhou were studied. Using the age class structure regression by diameter class, the static life table and survival function curves were determined, the dynamic indexes were calculated and spectrum analysis was done. The results showed: in 4 habitats the Reaumuria soongorica populations all contained more young members and less middle-aged or old members; all populations had highest death rates at age class Ⅲ. As the age increased, the death rates reduced; because of the physiological senescence, the death rates of age class Ⅶ and Ⅷ increased. The survival curve of the populations approached to Deevey type Ⅲ. The populations all belonged to progressive type. The order of growth potential is: at the foot of shady slope (5.90%)> at the foot of sunny slope (5.62%)> at the top of shady slope (4.77%)> at the top of sunny slope (3.79%), and this order indicated obvious periodicity. The dynamic quantity of all Reaumuria soongorica populations were overall controlled by their biological characteristics of lifecycle. Due to the environmental heterogeneity caused by different slope situations and altitudes, Reaumuria soongorica populations at the foot of shady slope grew well, at the top of shady slope and at the foot of sunny slope took the second place. But Reaumuria soongorica populations at the top of sunny slope which were influenced significantly by both environment and intraspecific competition pressure, had the biggest danger coefficient. The Reaumuria soongorica populations should be protected by proper tending practices to enhance sustainable development.
DOI:10.11849/zrzyxb.2011.10.009URL [本文引用: 2]
The quantity dynamics of the natural Reaumuria soongorica populations from different habitats in the south-north hills in Lanzhou were studied. Using the age class structure regression by diameter class, the static life table and survival function curves were determined, the dynamic indexes were calculated and spectrum analysis was done. The results showed: in 4 habitats the Reaumuria soongorica populations all contained more young members and less middle-aged or old members; all populations had highest death rates at age class Ⅲ. As the age increased, the death rates reduced; because of the physiological senescence, the death rates of age class Ⅶ and Ⅷ increased. The survival curve of the populations approached to Deevey type Ⅲ. The populations all belonged to progressive type. The order of growth potential is: at the foot of shady slope (5.90%)> at the foot of sunny slope (5.62%)> at the top of shady slope (4.77%)> at the top of sunny slope (3.79%), and this order indicated obvious periodicity. The dynamic quantity of all Reaumuria soongorica populations were overall controlled by their biological characteristics of lifecycle. Due to the environmental heterogeneity caused by different slope situations and altitudes, Reaumuria soongorica populations at the foot of shady slope grew well, at the top of shady slope and at the foot of sunny slope took the second place. But Reaumuria soongorica populations at the top of sunny slope which were influenced significantly by both environment and intraspecific competition pressure, had the biggest danger coefficient. The Reaumuria soongorica populations should be protected by proper tending practices to enhance sustainable development.
Plant biodiversity and population dynamics//Monson RK. Ecology and the Environment. Springer
1
2014
... 种群数量动态是种群个体生存能力与环境相互作用的结果(
林窗对岷江冷杉幼苗生存过程的影响
1
2016
... 存活曲线能够直观地表达种群的存活过程.
林窗对岷江冷杉幼苗生存过程的影响
1
2016
... 存活曲线能够直观地表达种群的存活过程.
植物种群与群落结构动态量化分析方法研究
1
1998
... 种群数量化分析是根据种群内两个相邻龄级间的个体数量变化动态指数(Vn)和整个种群年龄结构的数量变化动态指数(Vpi)及考虑未来外部干扰时种群年龄结构动态指数(V′pi)客观精确地定量比较评价种群结构动态(
植物种群与群落结构动态量化分析方法研究
1
1998
... 种群数量化分析是根据种群内两个相邻龄级间的个体数量变化动态指数(Vn)和整个种群年龄结构的数量变化动态指数(Vpi)及考虑未来外部干扰时种群年龄结构动态指数(V′pi)客观精确地定量比较评价种群结构动态(
Spatial pattern and size distribution of the animal-dispersed tree Quercus robur in two spruce-dominated forests
1
2000
... 根据同一树种龄级和径级对相同环境反应规律一致的特点(
Tree life tables
1
1987
... 种群结构与动态是种群生态学研究的核心问题, 种群结构反映着种群不同年龄、大小个体的数量配置情况, 也反映着种群数量动态与变化趋势(
中国特有植物巴山榧树的种群结构与动态
1
2018
... 种群结构与动态是种群生态学研究的核心问题, 种群结构反映着种群不同年龄、大小个体的数量配置情况, 也反映着种群数量动态与变化趋势(
中国特有植物巴山榧树的种群结构与动态
1
2018
... 种群结构与动态是种群生态学研究的核心问题, 种群结构反映着种群不同年龄、大小个体的数量配置情况, 也反映着种群数量动态与变化趋势(
濒危植物羽叶丁香种群结构与动态特征
1
2018
... 种群结构与动态是种群生态学研究的核心问题, 种群结构反映着种群不同年龄、大小个体的数量配置情况, 也反映着种群数量动态与变化趋势(
濒危植物羽叶丁香种群结构与动态特征
1
2018
... 种群结构与动态是种群生态学研究的核心问题, 种群结构反映着种群不同年龄、大小个体的数量配置情况, 也反映着种群数量动态与变化趋势(
Population structure and spatial pattern of main tree species in secondary Betula platyphylla forest in Ziwuling Mountains, China
1
2014
... 种群结构与动态是种群生态学研究的核心问题, 种群结构反映着种群不同年龄、大小个体的数量配置情况, 也反映着种群数量动态与变化趋势(
天山北坡天山云杉的种群结构特征和空间分布格局
2
2013
... 3个林区紫果云杉幼苗、幼树均占据极大部分, GZ幼苗比例最高, 占种群个体总数的一半以上, 达到60.73%, KC为51.01%, 比例最低的YLG也达到34.29%.大量幼苗、幼树的存在表明紫果云杉具有很强的自然更新能力, 为种群增长提供了丰富的物质基础(
... 存活曲线能够直观地表达种群的存活过程.
天山北坡天山云杉的种群结构特征和空间分布格局
2
2013
... 3个林区紫果云杉幼苗、幼树均占据极大部分, GZ幼苗比例最高, 占种群个体总数的一半以上, 达到60.73%, KC为51.01%, 比例最低的YLG也达到34.29%.大量幼苗、幼树的存在表明紫果云杉具有很强的自然更新能力, 为种群增长提供了丰富的物质基础(
... 存活曲线能够直观地表达种群的存活过程.
濒危植物梵净山冷杉野生种群结构及动态特征
1
2011
... 3个林区紫果云杉幼苗、幼树均占据极大部分, GZ幼苗比例最高, 占种群个体总数的一半以上, 达到60.73%, KC为51.01%, 比例最低的YLG也达到34.29%.大量幼苗、幼树的存在表明紫果云杉具有很强的自然更新能力, 为种群增长提供了丰富的物质基础(
濒危植物梵净山冷杉野生种群结构及动态特征
1
2011
... 3个林区紫果云杉幼苗、幼树均占据极大部分, GZ幼苗比例最高, 占种群个体总数的一半以上, 达到60.73%, KC为51.01%, 比例最低的YLG也达到34.29%.大量幼苗、幼树的存在表明紫果云杉具有很强的自然更新能力, 为种群增长提供了丰富的物质基础(
长白山云冷杉种群结构和动态分析
1
2014
... 3个林区紫果云杉幼苗、幼树均占据极大部分, GZ幼苗比例最高, 占种群个体总数的一半以上, 达到60.73%, KC为51.01%, 比例最低的YLG也达到34.29%.大量幼苗、幼树的存在表明紫果云杉具有很强的自然更新能力, 为种群增长提供了丰富的物质基础(
长白山云冷杉种群结构和动态分析
1
2014
... 3个林区紫果云杉幼苗、幼树均占据极大部分, GZ幼苗比例最高, 占种群个体总数的一半以上, 达到60.73%, KC为51.01%, 比例最低的YLG也达到34.29%.大量幼苗、幼树的存在表明紫果云杉具有很强的自然更新能力, 为种群增长提供了丰富的物质基础(
水杉原生母树种群结构与动态
1
2017
... 种群数量动态是种群个体生存能力与环境相互作用的结果(
水杉原生母树种群结构与动态
1
2017
... 种群数量动态是种群个体生存能力与环境相互作用的结果(
藏东南高山松种群结构及动态特征
2
2013
... 3个林区紫果云杉幼苗、幼树均占据极大部分, GZ幼苗比例最高, 占种群个体总数的一半以上, 达到60.73%, KC为51.01%, 比例最低的YLG也达到34.29%.大量幼苗、幼树的存在表明紫果云杉具有很强的自然更新能力, 为种群增长提供了丰富的物质基础(
... 种群数量动态是种群个体生存能力与环境相互作用的结果(
藏东南高山松种群结构及动态特征
2
2013
... 3个林区紫果云杉幼苗、幼树均占据极大部分, GZ幼苗比例最高, 占种群个体总数的一半以上, 达到60.73%, KC为51.01%, 比例最低的YLG也达到34.29%.大量幼苗、幼树的存在表明紫果云杉具有很强的自然更新能力, 为种群增长提供了丰富的物质基础(
... 种群数量动态是种群个体生存能力与环境相互作用的结果(
洮河林区森林生态系统服务功能及价值评估
2
2018
... 洮河是黄河的一级支流, 洮河林区是国家天然林资源保护工程重点实施区.该林区森林生态系统对维持洮河乃至黄河上游区域的生态平衡、遏制生态退化发挥着重要的屏障作用, 其森林生态服务功能总价值达到217.28亿元·a-1, 占甘肃全省的10%以上(
... 本研究选在洮河上游尕海-则岔国家级自然保护区则岔沟、洮河自然保护区卡车林区和冶力关森林公园进行.研究区地处青藏高原东部边缘, 黄土高原与甘肃东南部山区交汇带(102.18°-103.98° E, 34.08°-35.16° N), 属青藏高原湿润气候和高原大陆性季风气候区, 年降水量634-680 mm, 年日照时间2 276-2 364 h, 年平均气温2.3-5.8 ℃.山岭陡峻, 海拔1 000-4 300 m, 水资源极为丰富.土壤多为棕、褐色森林土, 森林植被主要为暗针叶林, 并具有明显的垂直地带性和阴阳坡差异(
洮河林区森林生态系统服务功能及价值评估
2
2018
... 洮河是黄河的一级支流, 洮河林区是国家天然林资源保护工程重点实施区.该林区森林生态系统对维持洮河乃至黄河上游区域的生态平衡、遏制生态退化发挥着重要的屏障作用, 其森林生态服务功能总价值达到217.28亿元·a-1, 占甘肃全省的10%以上(
... 本研究选在洮河上游尕海-则岔国家级自然保护区则岔沟、洮河自然保护区卡车林区和冶力关森林公园进行.研究区地处青藏高原东部边缘, 黄土高原与甘肃东南部山区交汇带(102.18°-103.98° E, 34.08°-35.16° N), 属青藏高原湿润气候和高原大陆性季风气候区, 年降水量634-680 mm, 年日照时间2 276-2 364 h, 年平均气温2.3-5.8 ℃.山岭陡峻, 海拔1 000-4 300 m, 水资源极为丰富.土壤多为棕、褐色森林土, 森林植被主要为暗针叶林, 并具有明显的垂直地带性和阴阳坡差异(
河北辽河源自然保护区油松种群年龄结构和种群动态
1
2017
... 种群数量动态是种群个体生存能力与环境相互作用的结果(
河北辽河源自然保护区油松种群年龄结构和种群动态
1
2017
... 种群数量动态是种群个体生存能力与环境相互作用的结果(
Introduction to Plant Population Ecology. Longman Group
1
1982
... 以龄级为横坐标, 实际存活个体数Ax为纵坐标, 绘制紫果云杉种群存活曲线.采用指数函数和幂函数进行存活数和龄级关系的拟合, 依据决定系数、F检验值来判定模型拟合效果(
天山云杉种群数量动态研究
1
2009
... 存活曲线能够直观地表达种群的存活过程.
天山云杉种群数量动态研究
1
2009
... 存活曲线能够直观地表达种群的存活过程.
长白山臭冷杉种群结构与动态
1
2017
... 种群结构与动态是种群生态学研究的核心问题, 种群结构反映着种群不同年龄、大小个体的数量配置情况, 也反映着种群数量动态与变化趋势(
长白山臭冷杉种群结构与动态
1
2017
... 种群结构与动态是种群生态学研究的核心问题, 种群结构反映着种群不同年龄、大小个体的数量配置情况, 也反映着种群数量动态与变化趋势(
同倍体杂交物种紫果云杉的生态位分化及其未来潜在分布区预测
1
2018
... 紫果云杉(Picea purpurea)源于丽江云杉(P. likiangensis)和青扦(P. wilsonii)的自然同倍体杂交, 占据了青藏高原东北部高纬度高海拔极端生境(
同倍体杂交物种紫果云杉的生态位分化及其未来潜在分布区预测
1
2018
... 紫果云杉(Picea purpurea)源于丽江云杉(P. likiangensis)和青扦(P. wilsonii)的自然同倍体杂交, 占据了青藏高原东北部高纬度高海拔极端生境(
不同生境下濒危植物裸果木种群结构及动态特征
1
2015
... 3个林区紫果云杉幼苗、幼树均占据极大部分, GZ幼苗比例最高, 占种群个体总数的一半以上, 达到60.73%, KC为51.01%, 比例最低的YLG也达到34.29%.大量幼苗、幼树的存在表明紫果云杉具有很强的自然更新能力, 为种群增长提供了丰富的物质基础(
不同生境下濒危植物裸果木种群结构及动态特征
1
2015
... 3个林区紫果云杉幼苗、幼树均占据极大部分, GZ幼苗比例最高, 占种群个体总数的一半以上, 达到60.73%, KC为51.01%, 比例最低的YLG也达到34.29%.大量幼苗、幼树的存在表明紫果云杉具有很强的自然更新能力, 为种群增长提供了丰富的物质基础(
Characteristics and dynamics analysis of Populus euphratica populations in the middle reaches of Tarim River
1
2010
... 根据同一树种龄级和径级对相同环境反应规律一致的特点(
半湿润半干旱过渡区洮河流域植被盖度变化特征
1
2016
... 本研究选在洮河上游尕海-则岔国家级自然保护区则岔沟、洮河自然保护区卡车林区和冶力关森林公园进行.研究区地处青藏高原东部边缘, 黄土高原与甘肃东南部山区交汇带(102.18°-103.98° E, 34.08°-35.16° N), 属青藏高原湿润气候和高原大陆性季风气候区, 年降水量634-680 mm, 年日照时间2 276-2 364 h, 年平均气温2.3-5.8 ℃.山岭陡峻, 海拔1 000-4 300 m, 水资源极为丰富.土壤多为棕、褐色森林土, 森林植被主要为暗针叶林, 并具有明显的垂直地带性和阴阳坡差异(
半湿润半干旱过渡区洮河流域植被盖度变化特征
1
2016
... 本研究选在洮河上游尕海-则岔国家级自然保护区则岔沟、洮河自然保护区卡车林区和冶力关森林公园进行.研究区地处青藏高原东部边缘, 黄土高原与甘肃东南部山区交汇带(102.18°-103.98° E, 34.08°-35.16° N), 属青藏高原湿润气候和高原大陆性季风气候区, 年降水量634-680 mm, 年日照时间2 276-2 364 h, 年平均气温2.3-5.8 ℃.山岭陡峻, 海拔1 000-4 300 m, 水资源极为丰富.土壤多为棕、褐色森林土, 森林植被主要为暗针叶林, 并具有明显的垂直地带性和阴阳坡差异(
黄龙山林区不同郁闭度对辽东栎种群结构的影响
1
2017
... 结合静态生命表的分析结果可知, 3个林区紫果云杉幼龄期(DBH < 5 cm)死亡率均在70%以上, 幼龄期过后, 经环境筛选留存下的个体生存能力增强, 因此后续龄级死亡率逐渐降低.生命期望值总体均表现为随龄级增大先增大后减小的变化趋势, 幼小龄期迅速增大, 青中龄期时, GZ稳中有降, YLG和KC林区则出现较大波动, 中老龄后期均下降到较低水平, 反映了紫果云杉生命力随龄级变化的生存特征.生境不同造成生理旺盛期不同, 生命期望值大表明林木个体生理旺盛生存力强.GZ的生理旺盛期持续时间长, 表明近自然状态的GZ林区适合紫果云杉生存, KC和YLG的种群死亡率出现负值说明较小龄级筛选强度过大, 同时也证实了干扰存在的可能, 因此生理旺盛期较短.调查中发现, 虽然生境有所差异, 但自然更新的紫果云杉幼苗大都于大树树冠下集中分布, 因此幼龄阶段必会出现剧烈的竞争, 导致高死亡率出现.进入小龄级后, 林木个体需要一定空间和光照进行生长释放, 但是由于生长在大树之下, 受较低枝下高限制, 空间和光照不足进一步加剧了死亡, 造成幼小龄级普遍的高死亡率(
黄龙山林区不同郁闭度对辽东栎种群结构的影响
1
2017
... 结合静态生命表的分析结果可知, 3个林区紫果云杉幼龄期(DBH < 5 cm)死亡率均在70%以上, 幼龄期过后, 经环境筛选留存下的个体生存能力增强, 因此后续龄级死亡率逐渐降低.生命期望值总体均表现为随龄级增大先增大后减小的变化趋势, 幼小龄期迅速增大, 青中龄期时, GZ稳中有降, YLG和KC林区则出现较大波动, 中老龄后期均下降到较低水平, 反映了紫果云杉生命力随龄级变化的生存特征.生境不同造成生理旺盛期不同, 生命期望值大表明林木个体生理旺盛生存力强.GZ的生理旺盛期持续时间长, 表明近自然状态的GZ林区适合紫果云杉生存, KC和YLG的种群死亡率出现负值说明较小龄级筛选强度过大, 同时也证实了干扰存在的可能, 因此生理旺盛期较短.调查中发现, 虽然生境有所差异, 但自然更新的紫果云杉幼苗大都于大树树冠下集中分布, 因此幼龄阶段必会出现剧烈的竞争, 导致高死亡率出现.进入小龄级后, 林木个体需要一定空间和光照进行生长释放, 但是由于生长在大树之下, 受较低枝下高限制, 空间和光照不足进一步加剧了死亡, 造成幼小龄级普遍的高死亡率(
灵空山辽东栎种群年龄结构与动态
2
2014
... 静态生命表反映的是多个世代重叠的年龄动态历程中的一个特定时间, 因此, 自然种群生命表的编制中常出现死亡率为负的情况, 对此, 许多研究对所得数据进行匀滑, 但当种群各龄级个体数差别较大时, 匀滑后的结果往往掩盖种群的真实信息.以实际存活数编制生命表虽会出现死亡率为负值的情况, 但却能真实反映种群的生态学过程.因此, 本研究以径级对应龄级, 根据各龄级实际存活数编制静态生命表, 编制及计算方法参见
... 3个林区紫果云杉幼苗、幼树均占据极大部分, GZ幼苗比例最高, 占种群个体总数的一半以上, 达到60.73%, KC为51.01%, 比例最低的YLG也达到34.29%.大量幼苗、幼树的存在表明紫果云杉具有很强的自然更新能力, 为种群增长提供了丰富的物质基础(
灵空山辽东栎种群年龄结构与动态
2
2014
... 静态生命表反映的是多个世代重叠的年龄动态历程中的一个特定时间, 因此, 自然种群生命表的编制中常出现死亡率为负的情况, 对此, 许多研究对所得数据进行匀滑, 但当种群各龄级个体数差别较大时, 匀滑后的结果往往掩盖种群的真实信息.以实际存活数编制生命表虽会出现死亡率为负值的情况, 但却能真实反映种群的生态学过程.因此, 本研究以径级对应龄级, 根据各龄级实际存活数编制静态生命表, 编制及计算方法参见
... 3个林区紫果云杉幼苗、幼树均占据极大部分, GZ幼苗比例最高, 占种群个体总数的一半以上, 达到60.73%, KC为51.01%, 比例最低的YLG也达到34.29%.大量幼苗、幼树的存在表明紫果云杉具有很强的自然更新能力, 为种群增长提供了丰富的物质基础(
黄土高原不同地区杜松种群结构与动态
1
2015
... 种群结构与动态是种群生态学研究的核心问题, 种群结构反映着种群不同年龄、大小个体的数量配置情况, 也反映着种群数量动态与变化趋势(
黄土高原不同地区杜松种群结构与动态
1
2015
... 种群结构与动态是种群生态学研究的核心问题, 种群结构反映着种群不同年龄、大小个体的数量配置情况, 也反映着种群数量动态与变化趋势(
尕海-则岔地区紫果云杉种群结构与动态特征
3
2018
... 本研究选在洮河上游尕海-则岔国家级自然保护区则岔沟、洮河自然保护区卡车林区和冶力关森林公园进行.研究区地处青藏高原东部边缘, 黄土高原与甘肃东南部山区交汇带(102.18°-103.98° E, 34.08°-35.16° N), 属青藏高原湿润气候和高原大陆性季风气候区, 年降水量634-680 mm, 年日照时间2 276-2 364 h, 年平均气温2.3-5.8 ℃.山岭陡峻, 海拔1 000-4 300 m, 水资源极为丰富.土壤多为棕、褐色森林土, 森林植被主要为暗针叶林, 并具有明显的垂直地带性和阴阳坡差异(
... 3个林区紫果云杉幼苗、幼树均占据极大部分, GZ幼苗比例最高, 占种群个体总数的一半以上, 达到60.73%, KC为51.01%, 比例最低的YLG也达到34.29%.大量幼苗、幼树的存在表明紫果云杉具有很强的自然更新能力, 为种群增长提供了丰富的物质基础(
... 结合静态生命表的分析结果可知, 3个林区紫果云杉幼龄期(DBH < 5 cm)死亡率均在70%以上, 幼龄期过后, 经环境筛选留存下的个体生存能力增强, 因此后续龄级死亡率逐渐降低.生命期望值总体均表现为随龄级增大先增大后减小的变化趋势, 幼小龄期迅速增大, 青中龄期时, GZ稳中有降, YLG和KC林区则出现较大波动, 中老龄后期均下降到较低水平, 反映了紫果云杉生命力随龄级变化的生存特征.生境不同造成生理旺盛期不同, 生命期望值大表明林木个体生理旺盛生存力强.GZ的生理旺盛期持续时间长, 表明近自然状态的GZ林区适合紫果云杉生存, KC和YLG的种群死亡率出现负值说明较小龄级筛选强度过大, 同时也证实了干扰存在的可能, 因此生理旺盛期较短.调查中发现, 虽然生境有所差异, 但自然更新的紫果云杉幼苗大都于大树树冠下集中分布, 因此幼龄阶段必会出现剧烈的竞争, 导致高死亡率出现.进入小龄级后, 林木个体需要一定空间和光照进行生长释放, 但是由于生长在大树之下, 受较低枝下高限制, 空间和光照不足进一步加剧了死亡, 造成幼小龄级普遍的高死亡率(
尕海-则岔地区紫果云杉种群结构与动态特征
3
2018
... 本研究选在洮河上游尕海-则岔国家级自然保护区则岔沟、洮河自然保护区卡车林区和冶力关森林公园进行.研究区地处青藏高原东部边缘, 黄土高原与甘肃东南部山区交汇带(102.18°-103.98° E, 34.08°-35.16° N), 属青藏高原湿润气候和高原大陆性季风气候区, 年降水量634-680 mm, 年日照时间2 276-2 364 h, 年平均气温2.3-5.8 ℃.山岭陡峻, 海拔1 000-4 300 m, 水资源极为丰富.土壤多为棕、褐色森林土, 森林植被主要为暗针叶林, 并具有明显的垂直地带性和阴阳坡差异(
... 3个林区紫果云杉幼苗、幼树均占据极大部分, GZ幼苗比例最高, 占种群个体总数的一半以上, 达到60.73%, KC为51.01%, 比例最低的YLG也达到34.29%.大量幼苗、幼树的存在表明紫果云杉具有很强的自然更新能力, 为种群增长提供了丰富的物质基础(
... 结合静态生命表的分析结果可知, 3个林区紫果云杉幼龄期(DBH < 5 cm)死亡率均在70%以上, 幼龄期过后, 经环境筛选留存下的个体生存能力增强, 因此后续龄级死亡率逐渐降低.生命期望值总体均表现为随龄级增大先增大后减小的变化趋势, 幼小龄期迅速增大, 青中龄期时, GZ稳中有降, YLG和KC林区则出现较大波动, 中老龄后期均下降到较低水平, 反映了紫果云杉生命力随龄级变化的生存特征.生境不同造成生理旺盛期不同, 生命期望值大表明林木个体生理旺盛生存力强.GZ的生理旺盛期持续时间长, 表明近自然状态的GZ林区适合紫果云杉生存, KC和YLG的种群死亡率出现负值说明较小龄级筛选强度过大, 同时也证实了干扰存在的可能, 因此生理旺盛期较短.调查中发现, 虽然生境有所差异, 但自然更新的紫果云杉幼苗大都于大树树冠下集中分布, 因此幼龄阶段必会出现剧烈的竞争, 导致高死亡率出现.进入小龄级后, 林木个体需要一定空间和光照进行生长释放, 但是由于生长在大树之下, 受较低枝下高限制, 空间和光照不足进一步加剧了死亡, 造成幼小龄级普遍的高死亡率(
不同间伐强度下辽东栎种群结构特征与空间分布格局
1
2012
... 鉴于此种现状, 必须针对各林区存在的主要问题采取目的性经营, 加强幼小林木的抚育和保护.对GZ林区须人工疏苗减小幼苗密度, 提高幼苗质量和存活率; 对KC林区则重点加强保护, 杜绝人为破坏, 清理“脏乱差”的林下空间, 为种群创造相对稳定的环境; YLG林区可通过人工补植使幼苗形成规模, 促进种群增长.同时对各林区的大树人工修枝增加枝下高, 伐除“老弱病残”的个体释放中上层空间, 促进林下“坐待”状态的小径级林木进入林冠层(
不同间伐强度下辽东栎种群结构特征与空间分布格局
1
2012
... 鉴于此种现状, 必须针对各林区存在的主要问题采取目的性经营, 加强幼小林木的抚育和保护.对GZ林区须人工疏苗减小幼苗密度, 提高幼苗质量和存活率; 对KC林区则重点加强保护, 杜绝人为破坏, 清理“脏乱差”的林下空间, 为种群创造相对稳定的环境; YLG林区可通过人工补植使幼苗形成规模, 促进种群增长.同时对各林区的大树人工修枝增加枝下高, 伐除“老弱病残”的个体释放中上层空间, 促进林下“坐待”状态的小径级林木进入林冠层(
兰州市南北两山不同生境红砂种群数量动态研究
2
2011
... 种群结构与动态是种群生态学研究的核心问题, 种群结构反映着种群不同年龄、大小个体的数量配置情况, 也反映着种群数量动态与变化趋势(
... 种群数量动态是种群个体生存能力与环境相互作用的结果(
兰州市南北两山不同生境红砂种群数量动态研究
2
2011
... 种群结构与动态是种群生态学研究的核心问题, 种群结构反映着种群不同年龄、大小个体的数量配置情况, 也反映着种群数量动态与变化趋势(
... 种群数量动态是种群个体生存能力与环境相互作用的结果(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}