Abstract Since the industrial revolution, marine ecosystems have faced unprecedented stress caused by increasing temperature and atmospheric CO2 concentration as a result of anthropogenic activities. In this review, we analyzed the domestic and international research status about impacts of global change on marine ecosystems by bibliometrics, briefly introduced the history of the research on marine ecosystems under global change, and reviewed the main progress in studies about the effects of global change on key processes of marine primary production, focusing on the impacts of ocean warming, ocean acidification, and eutrophication and hypoxia. We also summarized the major issues in current studies and proposed future research directions in the field. Keywords:marine ecosystem;primary production process;ocean warming;ocean acidification;eutrophication and hypoxia
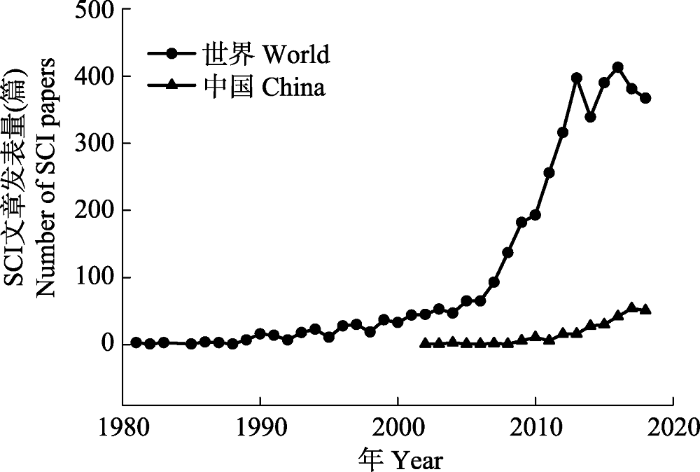
新窗口打开|下载原图ZIP|生成PPT 图11981-2018年发表的与“全球变化对海洋生态系统的影响”相关的SCI论文数。查询数据库: Web of Science; 查询关键词: TI = ((climate AND change) AND (marine OR ocean OR coast OR sea OR estuary))或TI = ((global AND warming) AND (marine OR ocean OR coast OR sea OR estuary))或TI = ((ocean AND acidification) AND (marine OR ocean OR coast OR sea OR estuary))或TI = ((eutrophication AND hypoxia) AND (marine OR ocean OR coast OR sea OR estuary))。
Fig. 1Number of SCI papers published between 1981 and 2018 related to “the impacts of global change on marine ecosystems”. Database: Web of Science; Key words: TI = ((climate AND change) AND (marine OR ocean OR coast OR sea OR estuary)) or TI = ((global AND warming) AND (marine OR ocean OR coast OR sea OR estuary)) or TI = ((ocean AND acidification) AND (marine OR ocean OR coast OR sea OR estuary)) or TI = ((eutrophication AND hypoxia) AND (marine OR ocean OR coast OR sea OR estuary)).
Fig. 2Top 10 countries for SCI papers published between 1981 and 2018 related to “the impacts of global change on marine ecosystems”.
图3
新窗口打开|下载原图ZIP|生成PPT 图31981-2018年发表的与“全球变暖、海洋酸化和富营养化对海洋生态系统的影响”相关的SCI论文数。A, 国际发展态势。B, 中国发展态势。查询数据库: Web of Science; 查询关键词: TI = (climate AND warming) AND (marine OR ocean OR coast OR sea OR estuary), 或TI = (global AND warming) AND (marine OR ocean OR coast OR sea OR estuary); TI = (ocean AND acidification) AND (marine OR ocean OR coast OR sea OR estuary); TI = Eutrophication* AND (marine OR ocean OR coast OR sea OR estuary), 或TI = Hypoxia* AND (marine OR ocean OR coast OR sea OR estuary)) , 或TI = Deoxygenation* AND (marine OR ocean OR coast OR sea OR estuary)。
Fig. 3Number of SCI papers published between 1981 and 2018 related to “the impacts of ocean warming, ocean acidification and eutrophication and hypoxia on marine ecosystems”. A, Development trend of internation. B, Development trend of China. Database: Web of Science; Key wards: TI = (climate AND warming) AND (marine OR ocean OR coast OR sea OR estuary), or TI = (global AND warming) AND (marine OR ocean OR coast OR sea OR estuary); TI = (ocean AND acidification) AND (marine OR ocean OR coast OR sea OR estuary); TI = Eutrophication* AND (marine OR ocean OR coast OR sea OR estuary), or TI = Hypoxia* AND (marine OR ocean OR coast OR sea OR estuary)) , or TI = Deoxygenation* AND (marine OR ocean OR coast OR sea OR estuary).
Fig. 4Top 10 countries for SCI papers published between 1981 and 2018 related to “the impacts of ocean warming, ocean acidification and eutrophication and hypoxia on marine ecosystems”.
BadgerMR, AndrewsTJ, WhitneySM, LudwigM, YellowleesDC, LeggatW, PriceGD (1998). The diversity and coevolution of Rubisco, plastids, pyrenoids, and chloroplast- based CO2-concentrating mechanisms in algae Canadian Journal of Botany, 76, 1052-1071. [本文引用: 1]
BatesNR (2001). Interannual variability of oceanic CO2 and biogeochemical properties in the Western North Atlantic subtropical gyre Deep Sea Research, 48, 1507-1528. [本文引用: 1]
BianchiTS, DiMarcoSF, Cowan JrJH, HetlandRD, ChapmanP, DayJW, AllisonMA (2010). The science of hypoxia in the Northern Gulf of Mexico: a review Science of the Total Environment, 408, 1471-1484. [本文引用: 1]
BreitburgDL, HondorpDW, DaviasLA, DiazRJ (2009). Hypoxia, nitrogen, and fisheries: integrating effects across local and global landscapes Annual Review of Marine Science, 1, 329-349. [本文引用: 1]
BrewerPG (1978). Direct observation of the oceanic CO2 increase Geophysical Research Letters, 5, 997-1000. [本文引用: 1]
BroeckerWS, TakahashiT (1966). Calcium carbonate precipitation on the Bahama Banks Journal of Geophysical Research, 71, 1575-1602. [本文引用: 1]
BrunoJF, StachowiczJJ, BertnessMD (2003). Inclusion of facilitation into ecological theory Trends in Ecology & Evolution, 18, 119-125. [本文引用: 1]
CaoL, CaldeiraK (2008). Atmospheric CO2 stabilization and ocean acidification Geophysical Research Letters, 35, L19609. DOI: 10.1029/2008GL035072. [本文引用: 1]
CloernJE (2001). Our evolving conceptual model of the coastal eutrophication problem Marine Ecology Progress Series, 210, 223-253. [本文引用: 1]
CodispotiLA, BrandesJA, ChristensenJP, DevolAH, NaqviSWA, PaerlHW, YoshinariT (2001). The oceanic fixed nitrogen and nitrous oxide budgets: moving targets as we enter the anthropocene? Scientia Marina, 65, 85-105. [本文引用: 2]
DiazRJ, RosenbergR (2008). Spreading dead zones and consequences for marine ecosystems Science, 321, 926-929. URLPMID:18703733 [本文引用: 1]
DoneySC, FabryVJ, FeelyRA, KleypasJA (2009). Ocean acidification: the other CO2 problem Annual Review of Marine Science, 1, 169-192. [本文引用: 3]
DonnerSD (2009). Coping with commitment: projected thermal stress on coral reefs under different future scenarios PLOS ONE, 4, e5712. DOI: 10.1371/journal.pone.0005712. [本文引用: 1]
EppleyR (1972). Temperature and phytoplankton growth in the sea Fishery Bulletin, 70, 1063-1085. [本文引用: 1]
FlynnKJ, ClarkDR, MitraA, FabianH, HansenPJ, GlibertPM, WheelerGL, StoeckerDK, BlackfordJC, BrownleeC (2015). Ocean acidification with (de)eutrophication will alter future phytoplankton growth and succession Proceedings of the Royal Society B, 282, 20142604. DOI: 10.1098/rspb.2014.2604. URLPMID:25716793 [本文引用: 1]
GruberN (2004). The Ocean Carbon Cycle and Climate Kluwer Academic Publishers, Dordrecht, Netherlands. [本文引用: 1]
HallegraeffGM (2010). Ocean climate change, phytoplankton community responses, and harmful algal blooms: a formidable predictive challenge Journal of Phycology, 46, 220-235. [本文引用: 1]
Hoegh-GuldbergO, BrunoJF (2010). The impact of climate change on the world’s marine ecosystems Science, 328, 1523-1528. URLPMID:20558709 [本文引用: 1]
HofmannGE, BarryJP, EdmundsPJ, GatesRD, HutchinsDA, KlingerT, SewellMA (2010). The effect of ocean acidification on calcifying organisms in marine ecosystems: an organism-to-ecosystem perspective Annual Review of Ecology, Evolution, and Systematics, 41, 127-147. [本文引用: 1]
HongH, ShenR, ZhangF, WenZ, ChangS, LinW, KranzSA, LuoYW, KaoSJ, MorelFMM, ShiD (2017). The complex effects of ocean acidification on the prominent N2-fixing cyanobacterium Trichodesmium Science, 356, 527-531. DOI:10.1126/science.aal2981URLPMID:28450383 [本文引用: 1] Acidification of seawater caused by anthropogenic carbon dioxide (CO2) is anticipated to influence the growth of dinitrogen (N2)-fixing phytoplankton, which contribute a large fraction of primary production in the tropical and subtropical ocean. We found that growth and N2-fixation of the ubiquitous cyanobacterium Trichodesmium decreased under acidified conditions, notwithstanding a beneficial effect of high CO2 Acidification resulted in low cytosolic pH and reduced N2-fixation rates despite elevated nitrogenase concentrations. Low cytosolic pH required increased proton pumping across the thylakoid membrane and elevated adenosine triphosphate production. These requirements were not satisfied under field or experimental iron-limiting conditions, which greatly amplified the negative effect of acidification.
HutchinsDA, BoydPW (2016). Marine phytoplankton and the changing ocean iron cycle Nature Climate Change, 6, 1072-1079. [本文引用: 1]
HyrenbachKD, VeitRR (2003). Ocean warming and seabird communities of the southern California Current System (1987-98): response at multiple temporal scales Deep Sea Research Part II: Topical Studies in Oceanography, 50, 2537-2565. [本文引用: 1]
IshimatsuA, KikkawaT, HayashiM, LeeKS, KitaJ (2004). Effects of CO2 on marine fish: larvae and adults Journal of Oceanography, 60, 731-741. [本文引用: 1]
JinX, GruberN (2003). Offsetting the radiative benefit of ocean iron fertilization by enhancing N2O emissions Geophysical Research Letters, 30, 2249. DOI: 10.1029/2003gl018458. [本文引用: 1]
KeelingRE, K?rtzingerA, GruberN (2010). Ocean deoxygenation in a warming world Annual Review of Marine Science, 2, 199-229. [本文引用: 2]
KleynhansEJ, OttoSP, ReichPB, VellendM (2016). Adaptation to elevated CO2 in different biodiversity contexts Nature Communications, 7, 12358. DOI: 10.1038/ncomms12358. DOI:10.1038/ncomms12358URLPMID:27510545 [本文引用: 1] In the absence of migration, species persistence depends on adaption to a changing environment, but whether and how adaptation to global change is altered by community diversity is not understood. Community diversity may prevent, enhance or alter how species adapt to changing conditions by influencing population sizes, genetic diversity and/or the fitness landscape experienced by focal species. We tested the impact of community diversity on adaptation by performing a reciprocal transplant experiment on grasses that evolved for 14 years under ambient and elevated CO2, in communities of low or high species richness. Using biomass as a fitness proxy, we find evidence for local adaptation to elevated CO2, but only for plants assayed in a community of similar diversity to the one experienced during the period of selection. Our results indicate that the biological community shapes the very nature of the fitness landscape within which species evolve in response to elevated CO2.
KroekerKJ, KordasRL, CrimR, HendriksIE, RamajoL, SinghGS, DuarteCM, GattusoJP (2013). Impacts of ocean acidification on marine organisms: quantifying sensitivities and interaction with warming Global Change Biology, 19, 1884-1896. DOI:10.1111/gcb.12179URLPMID:23505245 [本文引用: 2] Ocean acidification represents a threat to marine species worldwide, and forecasting the ecological impacts of acidification is a high priority for science, management, and policy. As research on the topic expands at an exponential rate, a comprehensive understanding of the variability in organisms' responses and corresponding levels of certainty is necessary to forecast the ecological effects. Here, we perform the most comprehensive meta-analysis to date by synthesizing the results of 228 studies examining biological responses to ocean acidification. The results reveal decreased survival, calcification, growth, development and abundance in response to acidification when the broad range of marine organisms is pooled together. However, the magnitude of these responses varies among taxonomic groups, suggesting there is some predictable trait-based variation in sensitivity, despite the investigation of approximately 100 new species in recent research. The results also reveal an enhanced sensitivity of mollusk larvae, but suggest that an enhanced sensitivity of early life history stages is not universal across all taxonomic groups. In addition, the variability in species' responses is enhanced when they are exposed to acidification in multi-species assemblages, suggesting that it is important to consider indirect effects and exercise caution when forecasting abundance patterns from single-species laboratory experiments. Furthermore, the results suggest that other factors, such as nutritional status or source population, could cause substantial variation in organisms' responses. Last, the results highlight a trend towards enhanced sensitivity to acidification when taxa are concurrently exposed to elevated seawater temperature.
MackeyKRM, MorrisJJ, MorelFMM, KranzSA (2015). Response of photosynthesis to ocean acidification Oceanography, 28, 74-91. [本文引用: 1]
MayorDJ, EverettNR, CookKB (2012). End of century ocean warming and acidification effects on reproductive success in a temperate marine copepod Journal of Plankton Research, 34, 258-262. [本文引用: 1]
MooreB, RoafHE, WhitleyE (1906). On the effects of alkalies and acids, and of alkaline and acid salts, upon growth and cell division in the fertilized eggs of Echinus esculentus —A study in relationship to the causation of malignant disease Proceedings of the Royal Society B, 77, 102-136. [本文引用: 1]
NaylorRL, GoldburgRJ, PrimaveraJH, KautskyN, BeveridgeMCM, ClayJ, FolkeC, LubchencoJ, MooneyH, TroellM (2000). Effect of aquaculture on world fish supplies Nature, 405, 1017-1024. URLPMID:10890435 [本文引用: 1]
NeumannT, EilolaK, GustafssonB, Müller-KarulisB, KuznetsovI, MeierHEM, SavchukOP (2012). Extremes of temperature, oxygen and blooms in the Baltic Sea in a changing climate Ambio, 41, 574-585. URLPMID:22926880 [本文引用: 1]
NevisonC, ButlerJH, ElkinsJW (2003). Global distribution of N2O and the ΔN2O-AOU yield in the subsurface ocean Global Biogeochemical Cycles, 17, 1119. DOI: 10.1029/2003GB002068. [本文引用: 1]
P?rtnerHO (2008). Ecosystem effects of ocean acidification in times of ocean warming: a physiologist’s view Marine Ecology Progress Series, 373, 203-217. [本文引用: 1]
P?rtnerHO, KnustR (2007). Climate change affects marine fishes through the oxygen limitation of thermal tolerance Science, 315, 95-97. DOI:10.1126/science.1135471URLPMID:17204649 [本文引用: 1] A cause-and-effect understanding of climate influences on ecosystems requires evaluation of thermal limits of member species and of their ability to cope with changing temperatures. Laboratory data available for marine fish and invertebrates from various climatic regions led to the hypothesis that, as a unifying principle, a mismatch between the demand for oxygen and the capacity of oxygen supply to tissues is the first mechanism to restrict whole-animal tolerance to thermal extremes. We show in the eelpout, Zoarces viviparus, a bioindicator fish species for environmental monitoring from North and Baltic Seas (Helcom), that thermally limited oxygen delivery closely matches environmental temperatures beyond which growth performance and abundance decrease. Decrements in aerobic performance in warming seas will thus be the first process to cause extinction or relocation to cooler waters.
P?rtnerHO, LangenbuchM (2005). Synergistic effects of temperature extremes, hypoxia, and increases in CO2 on marine animals: from earth history to global change Journal of Geophysical Research, 110, C09S10. DOI: 10.1029/ 2004JC002561. [本文引用: 1]
RabalaisNN, DíazRJ, LevinLA, TurnerRE, GilbertD, ZhangJ (2010). Dynamics and distribution of natural and human- caused hypoxia Biogeosciences, 7, 585-619. [本文引用: 2]
ReinfelderJR (2011). Carbon concentrating mechanisms in eukaryotic marine phytoplankton Annual Review of Marine Science, 3, 291-315. [本文引用: 1]
RevelleR, SuessHE (1957). Carbon dioxide exchange between atmosphere and ocean and the question of an increase of atmospheric CO2 during the past decades Tellus, 9, 18-27. [本文引用: 1]
RiebesellU, SchulzKG, BellerbyRGJ, BotrosM, FritscheP, Meyerh?ferM, NeillC, NondalG, OschliesA, WohlersJ, Z?llnerE (2007). Enhanced biological carbon consumption in a high CO2 ocean Nature, 450, 545-548. [本文引用: 1]
RiebesellU, ZondervanI, RostB, TortellPD, ZeebeRE, MorelFMM (2000). Reduced calcification of marine plankton in response to increased atmospheric CO2 Nature, 407, 364-367. [本文引用: 1]
RosenthalH (1985). Constraints and perspectives in aquaculture development Marine Pollution Bulletin, 16, 227-231. [本文引用: 1]
RossPM, ParkerL, O’ConnorWA, BaileyEA (2011). The impact of ocean acidification on reproduction, early development and settlement of marine organisms Water, 3, 1005-1030. [本文引用: 1]
RubeyWW (1951). Geologic history of sea water: an attempt to state the problem Bulletin of the Geological Society of America, 62, 1111-1148. [本文引用: 1]
Sen GuptaBK, TurnerRE, RabalaisNN (1996). Seasonal oxygen depletion in the continental-shelf waters of Louisiana: historical record of benthic foraminifers Geology, 24, 227-230. [本文引用: 1]
ShiD, LiW, HopkinsonBM, HongH, LiD, KaoSJ, LinW (2015). Interactive effects of light, nitrogen source, and carbon dioxide on energy metabolism in the diatom Thalassiosira pseudonana Limnology and Oceanography, 60, 1805-1822. [本文引用: 2]
ShiD, XuY, HopkinsonBM, MorelFMM (2010). Effect of ocean acidification on iron availability to marine phytoplankton Science, 327, 676-679. [本文引用: 1]
SpivackAJ, YouCF, SmithHJ (1993). Foraminiferal boron isotope ratios as a proxy for surface ocean pH over the past 21 Myr Nature, 363, 149-151. [本文引用: 1]
SteinacherM, JoosF, Fr?licherTL, BoppL, CaduleP, CoccoV, DoneySC, GehlenM, LindsayK, MooreJK, SchneiderB, SegschneiderJ (2010). Projected 21st century decrease in marine productivity: a multi-model analysis Biogeosciences, 7, 979-1005. [本文引用: 1]
SuggettDJ, DongLF, LawsonT, LawrenzE, TorresL, SmithDJ (2013). Light availability determines susceptibility of reef building corals to ocean acidification Coral Reefs, 32, 327-337. [本文引用: 1]
TurnerRE, RabalaisNN (2017). 2017 Forecast: summer hypoxic zone size Northern Gulf of Mexico. http://www.healthygulf.org/sites/healthygulf.org/files/final_lsu_lumcon_ 2017_hypoxia_forecast.pdf. cited: 2019-11-14 URL [本文引用: 1]
WangX, ChengH, CheH, SunJ, LuH, QiangM, HuaT, ZhuB, LiH, MaW, LangL, JiaoL, LiD (2017). Modern dust aerosol availability in northwestern China Scientific Reports, 7, 8741. DOI: 10.1038/s41598-017-09458-w. DOI:10.1038/s41598-017-09458-wURLPMID:28821858 [本文引用: 2] The sources of modern dust aerosols and their emission magnitudes are fundamental for linking dust with climate and environment. Using field sample data, wind tunnel experiments and statistical analysis, we determined the contributions of wadis, gobi (stony desert), lakebeds, riverbeds, and interdunes to modern dust aerosol availability in the three important potential dust sources including the Tarim Basin, Qaidam Basin, and Ala Shan Plateau of China. The results show that riverbeds are the dominant landscape for modern dust aerosol availabilities in the Qaidam Basin, while wadis, gobi, and interdunes are the main landscapes over the Ala Shan Plateau and Tarim Basin. The Ala Shan Plateau and Tarim Basin are potential dust sources in northwestern China, while the Qaidam Basin is not a major source of the modern dust aerosols nowadays, and it is not acting in a significant way to the Loess Plateau presently. Moreover, most of modern dust aerosol emissions from China originated from aeolian processes with low intensities rather than from major dust events.
WeartSR (2003). The Discovery of Global Warming. Harvard University Press, Cambridge, USA. [本文引用: 1]
XiaoW, WangL, LawsE, XieY, ChenJ, LiuX, ChenB, HuangB (2018). Realized niches explain spatial gradients in seasonal abundance of phytoplankton groups in the South China Sea Progress in Oceanography, 162, 223-239. [本文引用: 1]
The diversity and coevolution of Rubisco, plastids, pyrenoids, and chloroplast- based CO2-concentrating mechanisms in algae 1 1998
... 当前, 表层海水中CO2的浓度为10-20 μmol·L-1 (Badger et al., 1998), 低于绝大多数浮游植物中催化固碳反应的关键酶——核酮糖-1,5-二磷酸羧化酶/加氧酶(RubisCO)的半饱和常数.因此, 大多数浮游植物依赖于耗能的CO2浓缩机制(CO2-concentrating mechanism, CCM)以提高胞内RubisCO周围CO2的浓度(Reinfelder, 2011).已有研究表明, 酸化条件下, CO2浓度升高可使CCM下调, 节省能量, 从而对部分浮游植物的生长具有不同程度的促进作用(Riebesell et al., 1993; Mackey et al., 2015).除了CO2浓度升高, 随之降低的海水pH则同样可能对浮游植物产生直接或间接的影响.例如, 近期我国****对海洋优势固氮蓝藻束毛藻(Trichodesmium)开展的研究表明, 海水pH下降导致其固氮酶效率降低, 干扰胞内pH稳态, 影响细胞产能, 且该负效应大于CO2浓度升高所产生的正效应, 故酸化的净效应为抑制束毛藻的固氮作用(Hong et al., 2017).此外, 海水pH下降还可能改变浮游植物对铁、磷等关键营养元素的利用(Shi et al., 2010; McQuaid et al., 2018), 从而间接影响浮游植物生长.除了光合作用和固氮作用外, 海水酸化还会对颗石藻、有孔虫类、珊瑚等钙化生物的钙化作用产生负面影响(Riebesell et al., 2000; Cao & Caldeira, 2008; Hofmann et al., 2010; Kroeker et al., 2013).对于海洋软体动物、棘皮动物、甲壳动物及鱼类等生物而言, 海洋酸化不仅会降低其幼体发育速度和个体大小(Ross et al., 2011), 还会降低部分生物类群的呼吸速率, 扰乱生物体内酸碱平衡, 影响其生长与繁殖(Ishimatsu et al., 2004; P?rtner & Langenbuch, 2005).海洋酸化对生物个体的影响会进一步体现在对海洋生态系统和海洋生物地球化学循环的影响.Riebesell等(2007)通过开展中尺度生态系统(mesocosm) CO2加富实验发现, 与350 mg·kg-1相比, 1 050 mg·kg-1 CO2浓度下的浮游植物群落在12天内多消耗了39%的无机碳, 且浮游植物的C:N从6升高到8, 这一方面将有利于提高生物碳泵的效率, 但另一方面C:N的增加会降低有机物的营养价值, 影响细菌降解与浮游植物繁殖, 改变海洋生态系统的动力学. 此外, 酸化与光辐射(Suggett et al., 2013; Shi et al., 2015; Kleynhans et al., 2016)以及酸化与暖化(P?rtner, 2008; Mayor et al., 2012; Kroeker et al., 2013)之间的耦合作用, 也会从个体、种群、群落等不同水平上对海洋生态系统产生影响. ...
Interannual variability of oceanic CO2 and biogeochemical properties in the Western North Atlantic subtropical gyre 1 2001
... 暖化的海水对生物的影响因种而异.对于大多数浮游植物而言, 在适温范围内, 海水暖化会促进其生长(Eppley, 1972), 但不同浮游植物对温度变化的响应程度不同, 因此全球暖化下浮游植物的生态位将发生改变(Xiao et al., 2018).然而, 对于大多数海洋动物, 暖化将在增加其体内代谢活性的同时加快其能量的消耗, 从而可能导致海洋生物自身生长与繁殖能力的下降(P?rtner & Knust, 2007).这些个体生理生化过程的变化会进一步改变种群的大小、时空分布格局与营养级水平, 最终引起群落结构与功能的改变(Bruno et al., 2003; Hyrenbach & Veit, 2003).从海洋生态系统的层面上来看, 全球变暖会改变海洋的初级生产力, 影响生态系统的稳定性.近期我国科学家采用模型估算发现, 1948-2007年, 全球气候变暖导致海洋水体层化加剧, 使得北大西洋、北太平洋和印度洋初级生产力分别降低了40%、24%和25% (Wang et al., 2017).倘若暖化现象一直持续, 到2100年, 海洋初级生产力将可能进一步减少20% (Steinacher et al., 2010).此外, 已有研究表明, 海水温度升高1 ℃, 珊瑚即会出现白化现象(Hoegh- Guldberg et al., 2007), 这会导致珊瑚礁生态系统多样性降低, 严重时会引发珊瑚礁系统的崩溃(Donner, 2009).海水暖化除了通过温度增加而对海洋生态系统产生直接影响外, 还会造成海洋层化加剧、缺氧区扩增等, 从而间接地对海洋生态系统产生一系列影响(Keeling et al., 2010; Wang et al., 2017). ...
Adaptation to elevated CO2 in different biodiversity contexts 1 2016
... 当前, 表层海水中CO2的浓度为10-20 μmol·L-1 (Badger et al., 1998), 低于绝大多数浮游植物中催化固碳反应的关键酶——核酮糖-1,5-二磷酸羧化酶/加氧酶(RubisCO)的半饱和常数.因此, 大多数浮游植物依赖于耗能的CO2浓缩机制(CO2-concentrating mechanism, CCM)以提高胞内RubisCO周围CO2的浓度(Reinfelder, 2011).已有研究表明, 酸化条件下, CO2浓度升高可使CCM下调, 节省能量, 从而对部分浮游植物的生长具有不同程度的促进作用(Riebesell et al., 1993; Mackey et al., 2015).除了CO2浓度升高, 随之降低的海水pH则同样可能对浮游植物产生直接或间接的影响.例如, 近期我国****对海洋优势固氮蓝藻束毛藻(Trichodesmium)开展的研究表明, 海水pH下降导致其固氮酶效率降低, 干扰胞内pH稳态, 影响细胞产能, 且该负效应大于CO2浓度升高所产生的正效应, 故酸化的净效应为抑制束毛藻的固氮作用(Hong et al., 2017).此外, 海水pH下降还可能改变浮游植物对铁、磷等关键营养元素的利用(Shi et al., 2010; McQuaid et al., 2018), 从而间接影响浮游植物生长.除了光合作用和固氮作用外, 海水酸化还会对颗石藻、有孔虫类、珊瑚等钙化生物的钙化作用产生负面影响(Riebesell et al., 2000; Cao & Caldeira, 2008; Hofmann et al., 2010; Kroeker et al., 2013).对于海洋软体动物、棘皮动物、甲壳动物及鱼类等生物而言, 海洋酸化不仅会降低其幼体发育速度和个体大小(Ross et al., 2011), 还会降低部分生物类群的呼吸速率, 扰乱生物体内酸碱平衡, 影响其生长与繁殖(Ishimatsu et al., 2004; P?rtner & Langenbuch, 2005).海洋酸化对生物个体的影响会进一步体现在对海洋生态系统和海洋生物地球化学循环的影响.Riebesell等(2007)通过开展中尺度生态系统(mesocosm) CO2加富实验发现, 与350 mg·kg-1相比, 1 050 mg·kg-1 CO2浓度下的浮游植物群落在12天内多消耗了39%的无机碳, 且浮游植物的C:N从6升高到8, 这一方面将有利于提高生物碳泵的效率, 但另一方面C:N的增加会降低有机物的营养价值, 影响细菌降解与浮游植物繁殖, 改变海洋生态系统的动力学. 此外, 酸化与光辐射(Suggett et al., 2013; Shi et al., 2015; Kleynhans et al., 2016)以及酸化与暖化(P?rtner, 2008; Mayor et al., 2012; Kroeker et al., 2013)之间的耦合作用, 也会从个体、种群、群落等不同水平上对海洋生态系统产生影响. ...
Impacts of ocean acidification on marine organisms: quantifying sensitivities and interaction with warming 2 2013
... 当前, 表层海水中CO2的浓度为10-20 μmol·L-1 (Badger et al., 1998), 低于绝大多数浮游植物中催化固碳反应的关键酶——核酮糖-1,5-二磷酸羧化酶/加氧酶(RubisCO)的半饱和常数.因此, 大多数浮游植物依赖于耗能的CO2浓缩机制(CO2-concentrating mechanism, CCM)以提高胞内RubisCO周围CO2的浓度(Reinfelder, 2011).已有研究表明, 酸化条件下, CO2浓度升高可使CCM下调, 节省能量, 从而对部分浮游植物的生长具有不同程度的促进作用(Riebesell et al., 1993; Mackey et al., 2015).除了CO2浓度升高, 随之降低的海水pH则同样可能对浮游植物产生直接或间接的影响.例如, 近期我国****对海洋优势固氮蓝藻束毛藻(Trichodesmium)开展的研究表明, 海水pH下降导致其固氮酶效率降低, 干扰胞内pH稳态, 影响细胞产能, 且该负效应大于CO2浓度升高所产生的正效应, 故酸化的净效应为抑制束毛藻的固氮作用(Hong et al., 2017).此外, 海水pH下降还可能改变浮游植物对铁、磷等关键营养元素的利用(Shi et al., 2010; McQuaid et al., 2018), 从而间接影响浮游植物生长.除了光合作用和固氮作用外, 海水酸化还会对颗石藻、有孔虫类、珊瑚等钙化生物的钙化作用产生负面影响(Riebesell et al., 2000; Cao & Caldeira, 2008; Hofmann et al., 2010; Kroeker et al., 2013).对于海洋软体动物、棘皮动物、甲壳动物及鱼类等生物而言, 海洋酸化不仅会降低其幼体发育速度和个体大小(Ross et al., 2011), 还会降低部分生物类群的呼吸速率, 扰乱生物体内酸碱平衡, 影响其生长与繁殖(Ishimatsu et al., 2004; P?rtner & Langenbuch, 2005).海洋酸化对生物个体的影响会进一步体现在对海洋生态系统和海洋生物地球化学循环的影响.Riebesell等(2007)通过开展中尺度生态系统(mesocosm) CO2加富实验发现, 与350 mg·kg-1相比, 1 050 mg·kg-1 CO2浓度下的浮游植物群落在12天内多消耗了39%的无机碳, 且浮游植物的C:N从6升高到8, 这一方面将有利于提高生物碳泵的效率, 但另一方面C:N的增加会降低有机物的营养价值, 影响细菌降解与浮游植物繁殖, 改变海洋生态系统的动力学. 此外, 酸化与光辐射(Suggett et al., 2013; Shi et al., 2015; Kleynhans et al., 2016)以及酸化与暖化(P?rtner, 2008; Mayor et al., 2012; Kroeker et al., 2013)之间的耦合作用, 也会从个体、种群、群落等不同水平上对海洋生态系统产生影响. ...
... ; Kroeker et al., 2013)之间的耦合作用, 也会从个体、种群、群落等不同水平上对海洋生态系统产生影响. ...
Response of photosynthesis to ocean acidification 1 2015
... 当前, 表层海水中CO2的浓度为10-20 μmol·L-1 (Badger et al., 1998), 低于绝大多数浮游植物中催化固碳反应的关键酶——核酮糖-1,5-二磷酸羧化酶/加氧酶(RubisCO)的半饱和常数.因此, 大多数浮游植物依赖于耗能的CO2浓缩机制(CO2-concentrating mechanism, CCM)以提高胞内RubisCO周围CO2的浓度(Reinfelder, 2011).已有研究表明, 酸化条件下, CO2浓度升高可使CCM下调, 节省能量, 从而对部分浮游植物的生长具有不同程度的促进作用(Riebesell et al., 1993; Mackey et al., 2015).除了CO2浓度升高, 随之降低的海水pH则同样可能对浮游植物产生直接或间接的影响.例如, 近期我国****对海洋优势固氮蓝藻束毛藻(Trichodesmium)开展的研究表明, 海水pH下降导致其固氮酶效率降低, 干扰胞内pH稳态, 影响细胞产能, 且该负效应大于CO2浓度升高所产生的正效应, 故酸化的净效应为抑制束毛藻的固氮作用(Hong et al., 2017).此外, 海水pH下降还可能改变浮游植物对铁、磷等关键营养元素的利用(Shi et al., 2010; McQuaid et al., 2018), 从而间接影响浮游植物生长.除了光合作用和固氮作用外, 海水酸化还会对颗石藻、有孔虫类、珊瑚等钙化生物的钙化作用产生负面影响(Riebesell et al., 2000; Cao & Caldeira, 2008; Hofmann et al., 2010; Kroeker et al., 2013).对于海洋软体动物、棘皮动物、甲壳动物及鱼类等生物而言, 海洋酸化不仅会降低其幼体发育速度和个体大小(Ross et al., 2011), 还会降低部分生物类群的呼吸速率, 扰乱生物体内酸碱平衡, 影响其生长与繁殖(Ishimatsu et al., 2004; P?rtner & Langenbuch, 2005).海洋酸化对生物个体的影响会进一步体现在对海洋生态系统和海洋生物地球化学循环的影响.Riebesell等(2007)通过开展中尺度生态系统(mesocosm) CO2加富实验发现, 与350 mg·kg-1相比, 1 050 mg·kg-1 CO2浓度下的浮游植物群落在12天内多消耗了39%的无机碳, 且浮游植物的C:N从6升高到8, 这一方面将有利于提高生物碳泵的效率, 但另一方面C:N的增加会降低有机物的营养价值, 影响细菌降解与浮游植物繁殖, 改变海洋生态系统的动力学. 此外, 酸化与光辐射(Suggett et al., 2013; Shi et al., 2015; Kleynhans et al., 2016)以及酸化与暖化(P?rtner, 2008; Mayor et al., 2012; Kroeker et al., 2013)之间的耦合作用, 也会从个体、种群、群落等不同水平上对海洋生态系统产生影响. ...
End of century ocean warming and acidification effects on reproductive success in a temperate marine copepod 1 2012
... 当前, 表层海水中CO2的浓度为10-20 μmol·L-1 (Badger et al., 1998), 低于绝大多数浮游植物中催化固碳反应的关键酶——核酮糖-1,5-二磷酸羧化酶/加氧酶(RubisCO)的半饱和常数.因此, 大多数浮游植物依赖于耗能的CO2浓缩机制(CO2-concentrating mechanism, CCM)以提高胞内RubisCO周围CO2的浓度(Reinfelder, 2011).已有研究表明, 酸化条件下, CO2浓度升高可使CCM下调, 节省能量, 从而对部分浮游植物的生长具有不同程度的促进作用(Riebesell et al., 1993; Mackey et al., 2015).除了CO2浓度升高, 随之降低的海水pH则同样可能对浮游植物产生直接或间接的影响.例如, 近期我国****对海洋优势固氮蓝藻束毛藻(Trichodesmium)开展的研究表明, 海水pH下降导致其固氮酶效率降低, 干扰胞内pH稳态, 影响细胞产能, 且该负效应大于CO2浓度升高所产生的正效应, 故酸化的净效应为抑制束毛藻的固氮作用(Hong et al., 2017).此外, 海水pH下降还可能改变浮游植物对铁、磷等关键营养元素的利用(Shi et al., 2010; McQuaid et al., 2018), 从而间接影响浮游植物生长.除了光合作用和固氮作用外, 海水酸化还会对颗石藻、有孔虫类、珊瑚等钙化生物的钙化作用产生负面影响(Riebesell et al., 2000; Cao & Caldeira, 2008; Hofmann et al., 2010; Kroeker et al., 2013).对于海洋软体动物、棘皮动物、甲壳动物及鱼类等生物而言, 海洋酸化不仅会降低其幼体发育速度和个体大小(Ross et al., 2011), 还会降低部分生物类群的呼吸速率, 扰乱生物体内酸碱平衡, 影响其生长与繁殖(Ishimatsu et al., 2004; P?rtner & Langenbuch, 2005).海洋酸化对生物个体的影响会进一步体现在对海洋生态系统和海洋生物地球化学循环的影响.Riebesell等(2007)通过开展中尺度生态系统(mesocosm) CO2加富实验发现, 与350 mg·kg-1相比, 1 050 mg·kg-1 CO2浓度下的浮游植物群落在12天内多消耗了39%的无机碳, 且浮游植物的C:N从6升高到8, 这一方面将有利于提高生物碳泵的效率, 但另一方面C:N的增加会降低有机物的营养价值, 影响细菌降解与浮游植物繁殖, 改变海洋生态系统的动力学. 此外, 酸化与光辐射(Suggett et al., 2013; Shi et al., 2015; Kleynhans et al., 2016)以及酸化与暖化(P?rtner, 2008; Mayor et al., 2012; Kroeker et al., 2013)之间的耦合作用, 也会从个体、种群、群落等不同水平上对海洋生态系统产生影响. ...
Carbonate-sensitive phytotransferrin controls high-affinity iron uptake in diatoms 1 2018
... 当前, 表层海水中CO2的浓度为10-20 μmol·L-1 (Badger et al., 1998), 低于绝大多数浮游植物中催化固碳反应的关键酶——核酮糖-1,5-二磷酸羧化酶/加氧酶(RubisCO)的半饱和常数.因此, 大多数浮游植物依赖于耗能的CO2浓缩机制(CO2-concentrating mechanism, CCM)以提高胞内RubisCO周围CO2的浓度(Reinfelder, 2011).已有研究表明, 酸化条件下, CO2浓度升高可使CCM下调, 节省能量, 从而对部分浮游植物的生长具有不同程度的促进作用(Riebesell et al., 1993; Mackey et al., 2015).除了CO2浓度升高, 随之降低的海水pH则同样可能对浮游植物产生直接或间接的影响.例如, 近期我国****对海洋优势固氮蓝藻束毛藻(Trichodesmium)开展的研究表明, 海水pH下降导致其固氮酶效率降低, 干扰胞内pH稳态, 影响细胞产能, 且该负效应大于CO2浓度升高所产生的正效应, 故酸化的净效应为抑制束毛藻的固氮作用(Hong et al., 2017).此外, 海水pH下降还可能改变浮游植物对铁、磷等关键营养元素的利用(Shi et al., 2010; McQuaid et al., 2018), 从而间接影响浮游植物生长.除了光合作用和固氮作用外, 海水酸化还会对颗石藻、有孔虫类、珊瑚等钙化生物的钙化作用产生负面影响(Riebesell et al., 2000; Cao & Caldeira, 2008; Hofmann et al., 2010; Kroeker et al., 2013).对于海洋软体动物、棘皮动物、甲壳动物及鱼类等生物而言, 海洋酸化不仅会降低其幼体发育速度和个体大小(Ross et al., 2011), 还会降低部分生物类群的呼吸速率, 扰乱生物体内酸碱平衡, 影响其生长与繁殖(Ishimatsu et al., 2004; P?rtner & Langenbuch, 2005).海洋酸化对生物个体的影响会进一步体现在对海洋生态系统和海洋生物地球化学循环的影响.Riebesell等(2007)通过开展中尺度生态系统(mesocosm) CO2加富实验发现, 与350 mg·kg-1相比, 1 050 mg·kg-1 CO2浓度下的浮游植物群落在12天内多消耗了39%的无机碳, 且浮游植物的C:N从6升高到8, 这一方面将有利于提高生物碳泵的效率, 但另一方面C:N的增加会降低有机物的营养价值, 影响细菌降解与浮游植物繁殖, 改变海洋生态系统的动力学. 此外, 酸化与光辐射(Suggett et al., 2013; Shi et al., 2015; Kleynhans et al., 2016)以及酸化与暖化(P?rtner, 2008; Mayor et al., 2012; Kroeker et al., 2013)之间的耦合作用, 也会从个体、种群、群落等不同水平上对海洋生态系统产生影响. ...
On the effects of alkalies and acids, and of alkaline and acid salts, upon growth and cell division in the fertilized eggs of Echinus esculentus —A study in relationship to the causation of malignant disease 1 1906
 ,*厦门大学近海海洋环境科学国家重点实验室, 福建厦门 361102
,*厦门大学近海海洋环境科学国家重点实验室, 福建厦门 361102
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT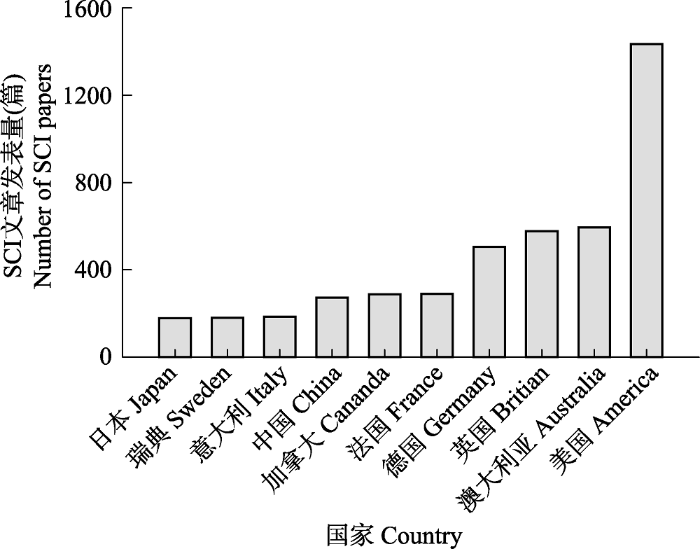 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT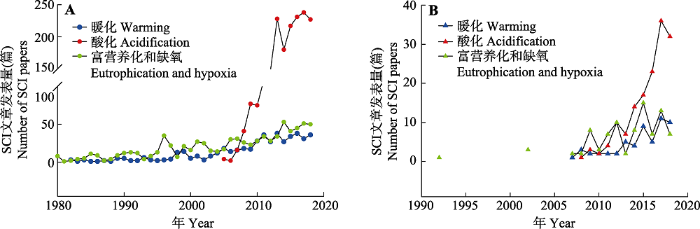 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT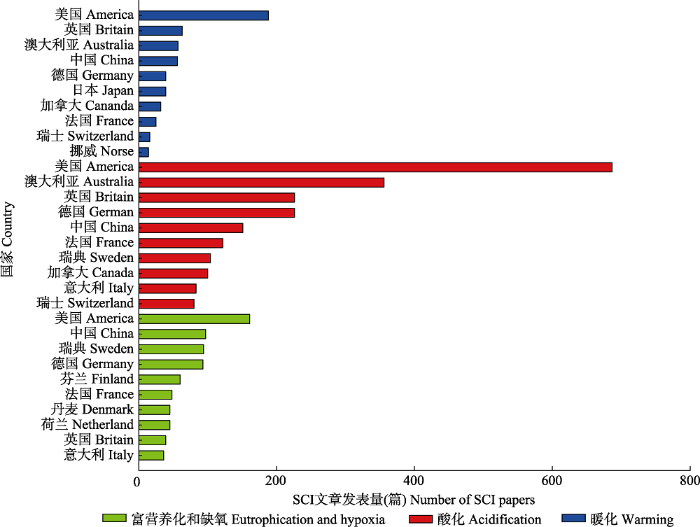 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}