Biomarkers and their applications in ecosystem research
Xiao-Juan FENG,,1,2,*, Yi-Yun WANG1,2, Ting LIU1, Juan JIA1, Guo-Hua DAI1, Tian MA1,2, Zong-Guang LIU11State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China 2College of Resources and Environment, University of Chinese Academy of Sciences, Beijing 100049, China
National Key R&D Program of China(2017YFC0503902) State Key Development Program for Basic Research of China(2015CB954201) Chinese Academy of Sciences Interdisciplinary Project(JCTD-2018-06)
Abstract Biomarkers are biogenic organic compounds that carry the chemical structures specific to their biological sources and survive long-term preservation in environmental and geological systems. The abundance of biomarkers may indicate the relative contribution of specific biological sources to the natural organic matter while their chemical and isotopic compositions may also inform on the transformation stage of organic matter and the environmental settings. Compared with conventional bulk analysis, biomarkers offer highly specific and sensitive tools to track the sources, transformation and dynamic changes of natural organic matter components and have therefore been widely used in ecological and biogeochemical studies in the past decades. In particular, combined with ecosystem observations and control experiments, biomarkers have shown great potentials in revealing changes in microbial activity and carbon sources, soil organic matter dynamics, stabilization mechanisms and response to global changes. The recently-developed biomarker-specific isotope analysis also exhibits a great promise in revealing ecosystem carbon and nitrogen turnover and food web structures. This review summarizes several major categories of commonly used biomarkers, their analytical methods, applications in ecosystem studies and existing pitfalls, and discusses future directions of research to provide guidance for biomarker users in ecology and environmental sciences. Keywords:biomarker;ecosystem research;soil organic matter;compound-specific isotope analysis;carbon cycling;food web
PDF (1165KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 冯晓娟, 王依云, 刘婷, 贾娟, 戴国华, 马田, 刘宗广. 生物标志物及其在生态系统研究中的应用. 植物生态学报, 2020, 44(4): 384-394. DOI: 10.17521/cjpe.2019.0139 FENG Xiao-Juan, WANG Yi-Yun, LIU Ting, JIA Juan, DAI Guo-Hua, MA Tian, LIU Zong-Guang. Biomarkers and their applications in ecosystem research. Chinese Journal of Plant Ecology, 2020, 44(4): 384-394. DOI: 10.17521/cjpe.2019.0139
Fig. 2Examples of typical monomer structures for the main types of biomarkers introduced in this paper. Plant-derived biomarkers: plant wax lipids including long-chain n-alkanes (I) and fatty acids (II), etc. (Otto et al., 2005); cutin monomers such as short-chain ω-hydroxyalkanoic acid (III); lignin phenols such as vanillin (IV) and syringic acid (V), etc. (Thevenot et al., 2010); suberin monomers such as long-chain α,ω-alkanedioic acid (VI; Otto et al., 2005). Micorbial-derived biomarkers: phospholipid fatty acids (PLFAs) such as linoleic acid (VII; Frosteg?rd & B??th, 1996); glycerol dialkyl glycerol tetraethers (GDGTs) such as GDGT-2 (VIII; Schouten et al., 2013); amino sugars, including α-D-glucosamine (IX) and α-D-galactosamine (X), etc. (Joergensen, 2018). In addition, neutral sugars such as α-D-glucose (XI) and amino acids with side chain (R group; XII) can be synthesized by both plants and microbes.
Table 1 表1 表1生态系统研究中常用的生物标志物的分子构成、提取方法、研究意义和关键参数 Table 1Molecular composition, extraction methods, research implications and key parameters of widely-used biomarkers in ecosystem research
生物标志物 Biomarker
分子组成 Molecular composition
提取方法 Extraction method
研究意义 Research implication
关键参数 Key parameter
植物蜡质脂类 Plant lipids
长链(>C20)正构烷烃、脂肪酸、脂肪醇、 固醇等 Long-chain (>C20) n-alkanes, n-alkanoic acids, n-alkanols, steroids, etc.
溶剂萃取 Solvent extraction
代表植物来源的脂类 Indicating lipids from terrestrial vegetation
ACL, CPI
角质单体 Cutin monomers
短链(C14-C18)羟基-环氧酸 Short-chain (C14-C18) hydroxy- and epoxy acids
碱式水解 Base hydrolysis
代表叶片角质来源的脂类 Indicating lipids from leaf cuticles
ω-C16/∑C16, ω-C18/∑C18
软木脂单体 Suberin monomers
长链(C20-C32)脂肪酸、双酸、ω-羟基酸等 Long-chain (C20-C32) aliphatic acids, diacids and ω-hydroxy acids, etc.
碱式水解 Base hydrolysis
代表根系和树皮软木脂来源的脂类 Indicating lipids from suberin in roots and barks
磷脂脂肪酸 Phospholipid fatty acids (PLFAs)
短链脂肪酸(<C20) Short-chain fatty acids (<C20)
Bligh-Dyer
指征活体微生物生物量和群落结构 Indicating microbial biomass and community structure
微生物细胞壁组分, 常指征环境中的微生物残体碳 Components of microbial cell wall; indicating microbial necromass in soils
GluN/MurN
ACL, 平均碳链长度; CPI, 碳优势指数; ACL = Σ(n × Xn)/Σ(Xn), CPI = 0.5 × [Σ(Xeven)/Σ(Xodd) + Σ(Xeven)/Σ(Xodd+2)], 其中X为含有一定碳原子数量的正构烷烃、脂肪酸、脂肪醇的浓度, n是碳原子数量, even代表偶数碳, odd代表奇数碳; ACL和CPI均可指示植物来源脂类的降解程度。ω-C16/ΣC16和ω-C18/ΣC18, 含16个或18个碳原子的ω-羟基酸与含16或18个碳原子的ω-羟基酸、双酸、羟基-环氧酸之和的比值, 可以指示角质的降解程度。F/B, 真菌细菌比, 表征土壤活体微生物群落中真菌和细菌的比例。MBT和CBT, 分别是支链GDGT的甲基化和环化指数, 可用于重建环境温度和pH。GM/AX, (半乳糖+甘露糖)/(阿拉伯糖+木糖); RF/AX, (鼠李糖+岩藻糖)/(阿拉伯糖+木糖); GM/AX和RF/AX均可指示微生物和植物来源糖类对土壤有机质的相对贡献。S/V, 丁香基酚类与香草基酚类的比值, 可区分裸子和被子植物; C/V, 肉桂基酚类与香草基酚类的比值, 可区分木本和非木本组织; (Ad/Al)V和(Ad/Al)S分别是香草基酚和丁香基酚类单体的酸醛比值, 指示木质素的氧化程度。D/L, 右旋(D型)氨基酸与左旋(L型)氨基酸的比值, 用于评价土壤中蛋白类物质的相对年龄。GluN/MurN, 氨基葡萄糖与胞壁酸的比值, 用于评价真菌和细菌残体对土壤有机质的相对贡献。 ACL, average chain length; CPI, carbon preference index; ACL = Σ(n × Xn)/Σ(Xn), CPI = 0.5 × [Σ(Xeven)/Σ(Xodd) + Σ(Xeven)/Σ(Xodd+2)], where X is the concentration of n-alkane, n-alkanol or n-alkanoic acid with a certain number of carbon atoms, whereby n is the number of carbon atoms; ACL and CPI may indicate the decomposition degree of plant-derived lipids. ω-C16/ΣC16 and ω-C18/ΣC18, the ratio of C16 or C18 ω-hydroxy acid to the summation of C16 or C18 ω-hydroxy acids, dicarboxylic acids, and hydroxy- and epoxy acids, both may indicate cutin degradation. F/B, ratio of fungi to bacteria in living soil microbes. MBT and CBT, methylation and cyclization index of branched GDGTs, respectively, proxies for temperature and pH reconstruction. GM/AX, (galactose+mannose)/(arabinose+xylose); RF/AX, (rhamnose+fucose)/(arabinose+xylose)-both ratio can indicate the relative contribution of microbe- and plant-derived sugars to soil organic matter. S/V, ratio of syringyl to vanillyl phenols, different for gymnosperms and angiosperms; C/V, ratio of cinnamyl to vanillyl phenols, different for woody and non-woody tissues; (Ad/Al)V and (Ad/Al)S are acid to aldehyde ratios of vanillyl and syringyl phenols, respectively, which can indicate the oxidation stage of lignin. D/L, the ratio of dextrorotary to levorotatory amino acid, for assessing the relative age of proteinaceous substances in soil. GluN/MurN, the ratio of glucosamine to muramic acid, for evaluating the relative contribution from fungal versus bacterial necromass to soil organic matter.
AmelungW (2003). Nitrogen biomarkers and their fate in soil Journal of Plant Nutrition and Soil Science, 166, 677-686. [本文引用: 1]
AmelungW, ZhangXD (2001). Determination of amino acid enantiomers in soils Soil Biology & Biochemistry, 33, 553-562. [本文引用: 4]
BianchiTS, CuiXQ, BlairNE, BurdigeDJ, EglintonTI, GalyV (2018). Centers of organic carbon burial and oxidation at the land-ocean interface Organic Geochemistry, 115, 138-155. [本文引用: 1]
BlighE, DyerW (1959). A rapid method of total lipid extraction and purification Canadian Journal of Biochemistry and Physiology, 37, 911-917. URLPMID:13671378 [本文引用: 1]
BoecklenWJ, YarnesCT, CookBA, JamesAC (2011). On the use of stable isotopes in trophic ecology Annual Review of Ecology, Evolution, and Systematics, 42, 411-440. [本文引用: 1]
BullID, van BergenPF, NottCJ, PoultonPR, EvershedRP (2000). Organic geochemical studies of soils from the Rothamsted classical experiments—V. The fate of lipids in different long-term experiments Organic Geochemistry, 31, 389-408. [本文引用: 1]
ChikaraishiY, SteffanSA, OgawaNO, IshikawaNF, SasakiY, TsuchiyaM, OhkouchiN (2014). High-resolution food webs based on nitrogen isotopic composition of amino acids Ecology and Evolution, 4, 2423-2449. URLPMID:25360278 [本文引用: 2]
CoppolaAI, WiedemeierDB, GalyV, HaghipourN, HankeUM, NascimentoGS, UsmanM, BlattmannTM, ReisserM, FreymondCV, ZhaoM, VossB, WackerL, Schefu?E, Peucker-EhrenbrinkB, AbivenS, SchmidtMWI, EglintonTI (2018). Global-scale evidence for the refractory nature of riverine black carbon Nature Geoscience, 11, 584-588. [本文引用: 1]
CotrufoMF, SoongJL, HortonAJ, CampbellEE, HaddixML, WallDH, PartonWJ (2015). Formation of soil organic matter via biochemical and physical pathways of litter mass loss Nature Geoscience, 8, 776-779. [本文引用: 1]
CrowSE, LajthaK, FilleyTR, SwanstonCW, BowdenRD, CaldwellBA (2009). Sources of plant-derived carbon and stability of organic matter in soil: implications for global change Global Change Biology, 15, 2003-2019. [本文引用: 2]
DavidsonEA, JanssensIA (2006). Temperature sensitivity of soil carbon decomposition and feedbacks to climate change Nature, 440, 165-173. URLPMID:16525463 [本文引用: 1]
DerenneS, LargeauC (2001). A review of some important families of refractory macromolecules: composition, origin, and fate in soils and sediments Soil Science, 166, 833-847. [本文引用: 1]
DidykB, SimoneitB, BrassellSC, EglintonG (1978). Organic geochemical indicators of palaeoenvironmental conditions of sedimentation Nature, 272, 216-222. [本文引用: 1]
DittmarT, FitznarHP, KattnerG (2001). Origin and biogeochemical cycling of organic nitrogen in the eastern Arctic Ocean as evident from D- and L-amino acids Geochimica et Cosmochimica Acta, 65, 4103-4114. [本文引用: 2]
DolgovaS, PoppBN, CourtoreilleK, EspieRHM, MacleanB, McMasterM, StrakaJR, TetreaultGR, WilkieS, HebertCE (2018). Spatial trends in a biomagnifying contaminant: application of amino acid compound-specific stable nitrogen isotope analysis to the interpretation of bird mercury levels Environmental Toxicology and Chemistry, 37, 1466-1475. URLPMID:29446488 [本文引用: 1]
DouglasPMJ, PaganiM, EglintonTI, BrennerM, CurtisJH, BreckenridgeA, JohnstonK (2018). A long-term decrease in the persistence of soil carbon caused by ancient Maya land use Nature Geoscience, 11, 645-651. [本文引用: 1]
EderE, SpielvogelS, K?lblA, AlbertG, K?gel-KnabnerI (2010). Analysis of hydrolysable neutral sugars in mineral soils: improvement of alditol acetylation for gas chromatographic separation and measurement Organic Geochemistry, 41, 580-585. [本文引用: 3]
EglintonTI, AluwihareLI, BauerJE, DruffelER, McNicholAP (1996). Gas chromatographic isolation of individual compounds from complex matrices for radiocarbon dating Analytical Chemistry, 68, 904-912. DOI:10.1021/ac9508513URLPMID:21619188 [本文引用: 1] This paper describes the application of a novel, practical approach for isolation of individual compounds from complex organic matrices for natural abundance radiocarbon measurement. This is achieved through the use of automated preparative capillary gas chromatography (PCGC) to separate and recover sufficient quantities of individual target compounds for (14)C analysis by accelerator mass spectrometry (AMS). We developed and tested this approach using a suite of samples (plant lipids, petroleums) whose ages spanned the (14)C time scale and which contained a variety of compound types (fatty acids, sterols, hydrocarbons). Comparison of individual compound and bulk radiocarbon signatures for the isotopically homogeneous samples studied revealed that Delta(14)C values generally agreed well (+/-10%). Background contamination was assessed at each stage of the isolation procedure, and incomplete solvent removal prior to combustion was the only significant source of additional carbon. Isotope fractionation was addressed through compound-specific stable carbon isotopic analyses. Fractionation of isotopes during isolation of individual compounds was minimal (<5 per thousand for delta(13)C), provided the entire peak was collected during PCGC. Trapping of partially coeluting peaks did cause errors, and these results highlight the importance of conducting stable carbon isotopic measurements of each trapped compound in concert with AMS for reliable radiocarbon measurements. The addition of carbon accompanying derivatization of functionalized compounds (e.g., fatty acids and sterols) prior to chromatographic separation represents a further source of potential error. This contribution can be removed using a simple isotopic mass balance approach. Based on these preliminary results, the PCGC-based approach holds promise for accurately determining (14)C ages on compounds specific to a given source within complex, heterogeneous samples.
EglintonTI, EglintonG (2008). Molecular proxies for paleoclimatology Earth and Planetary Science Letters, 275, 1-16. [本文引用: 1]
FengXJ, GustafssonO, HolmesRM, VonkJE, van DongenBE, SemiletovIP, DudarevOV, YunkerMB, MacdonaldRW, WackerL, MontluconDB, EglintonTI (2015). Multimolecular tracers of terrestrial carbon transfer across the pan-Arctic: C-14 characteristics of sedimentary carbon components and their environmental controls Global Biogeochemical Cycles, 29, 1855-1873. [本文引用: 1]
FengXJ, SimpsonAJ, WilsonKP, Dudley WilliamsD, SimpsonMJ (2008). Increased cuticular carbon sequestration and lignin oxidation in response to soil warming Nature Geoscience, 1, 836-839. [本文引用: 2]
FengXJ, VonkJE, van DongenBE, GustafssonO, SemiletovIP, DudarevOV, WangZH, MontluconDB, WackerL, EglintonTI (2013). Differential mobilization of terrestrial carbon pools in Eurasian Arctic river basins Proceedings of the National Academy of Sciences of the United States of America, 110, 14168-14173. URLPMID:23940354 [本文引用: 2]
Ferrier-PagesC, LealMC (2019). Stable isotopes as tracers of trophic interactions in marine mutualistic symbioses Ecology and Evolution, 9, 723-740. DOI:10.1002/ece3.4712URLPMID:30680151 [本文引用: 1] Mutualistic nutritional symbioses are widespread in marine ecosystems. They involve the association of a host organism (algae, protists, or marine invertebrates) with symbiotic microorganisms, such as bacteria, cyanobacteria, or dinoflagellates. Nutritional interactions between the partners are difficult to identify in symbioses because they only occur in intact associations. Stable isotope analysis (SIA) has proven to be a useful tool to highlight original nutrient sources and to trace nutrients acquired by and exchanged between the different partners of the association. However, although SIA has been extensively applied to study different marine symbiotic associations, there is no review taking into account of the different types of symbiotic associations, how they have been studied via SIA, methodological issues common among symbiotic associations, and solutions that can be transferred from one type of association with another. The present review aims to fill such gaps in the scientific literature by summarizing the current knowledge of how isotopes have been applied to key marine symbioses to unravel nutrient exchanges between partners, and by describing the difficulties in interpreting the isotopic signal. This review also focuses on the use of compound-specific stable isotope analysis and on statistical advances to analyze stable isotope data. It also highlights the knowledge gaps that would benefit from future research.
FogelML, GriffinPL, NewsomeSD (2016). Hydrogen isotopes in individual amino acids reflect differentiated pools of hydrogen from food and water in Escherichia coli Proceedings of the National Academy of Sciences of the United States of America, 113, 201525703. DOI: 10.1073/pnas.1525703113. [本文引用: 1]
Frosteg?rdA, B??thE (1996). The use of phospholipid fatty acid analysis to estimate bacterial and fungal biomass in soil Biology and Fertility of Soils, 22, 59-65. [本文引用: 3]
GalyV, EglintonT, France-LanordC, SylyaS (2011). The provenance of vegetation and environmental signatures encoded in vascular plant biomarkers carried by the Ganges-Brahmaputra rivers Earth and Planetary Science Letters, 304, 1-12. [本文引用: 2]
GlaserB, TurriónMB, AlefK (2004). Amino sugars and muramic acid—Biomarkers for soil microbial community structure analysis Soil Biology & Biochemistry, 36, 399-407. [本文引用: 1]
GleixnerG, CzimczikCJ, KramerC, LühkerB, SchmidtMW (2001). Plant compounds and their turnover and stabilization as soil organic matter //Schulze ED, Heimann M, Harrison S, Holland E, Lloyd J, Prentice IC, Schimel D. Global Biogeochemical Cycles in the Climate System. Elsevier, Amsterdam, Netherlands. 201-215. [本文引用: 1]
GriepentrogM, BodéS, BoeckxP, HagedornF, HeimA, SchmidtMW (2014). Nitrogen deposition promotes the production of new fungal residues but retards the decomposition of old residues in forest soil fractions Global Change Biology, 20, 327-340. URLPMID:23996910 [本文引用: 1]
GuninaA, KuzyakovY (2015). Sugars in soil and sweets for microorganisms: review of origin, content, composition and fate Soil Biology & Biochemistry, 90, 87-100. [本文引用: 1]
HayesJM, FreemanKH, PoppBN, HohamCH (1990). Compound-specific isotopic analyses: a novel tool for reconstruction of ancient biogeochemical processes Organic Geochemistry, 16, 1115-1128. URLPMID:11540919 [本文引用: 1]
HedgesJI, EglintonG, HatcherPG, KirchmanDL, ArnostiC, DerenneS, EvershedRP, K?gel-KnabnerI, de LeeuwJW, LittkeR, MichaelisW, Rullk?tterJ (2000). The molecularly- uncharacterized component of nonliving organic matter in natural environments Organic Geochemistry, 31, 945-958. [本文引用: 1]
HedgesJI, MannDC (1979). The characterization of plant tissues by their lignin oxidation products Geochimica et Cosmochimica Acta, 43, 1803-1807. [本文引用: 1]
HouJZ, HuangYS, BrodskyC, AlexandreMR, McNicholAP, KingJW, HuFS, ShenJ (2010). Radiocarbon dating of individual lignin phenols: a new approach for establishing chronology of Late Quaternary lake sediments Analytical Chemistry, 82, 7119-7126. DOI:10.1021/ac100494mURLPMID:20684521 [本文引用: 1] The reliability of chronology is a prerequisite for meaningful paleoclimate reconstructions from sedimentary archives. The conventional approach of radiocarbon dating bulk organic carbon in lake sediments is often hampered by the old carbon effect, i.e., the assimilation of ancient dissolved inorganic carbon (DIC) derived from carbonate bedrocks or other sources. Therefore, radiocarbon dating is ideally performed on organic compounds derived from land plants that use atmospheric CO(2) and rapidly delivered to sediments. We demonstrate that lignin phenols isolated from lake sediments using reversed phase high performance liquid chromatography (HPLC) can serve as effective (14)C dating materials for establishing chronology during the late Quaternary. We developed a procedure to purify lignin phenols, building upon a published method. By isolating lignin from standard wood reference substances, we show that our method yields pure lignin phenols and consistent ages as the consensus ages and that our procedure does not introduce radiocarbon contamination. We further demonstrate that lignin phenol ages are compatible with varve counted and macrofossil dated sediment horizons in Steel Lake and Fayetteville Green Lake. Applying the new method to lake sediment cores from Lake Qinghai demonstrates that lignin phenol ages in Lake Qinghai are consistently younger than bulk total organic carbon (TOC) ages which are contaminated by old carbon effect. We also show that the age offset between lignin and bulk organic carbon differs at different Lake Qinghai sedimentary horizons, suggesting a variable hard water effect at different times and that a uniform age correction throughout the core is inappropriate.
HuangXY, MeyersPA (2019). Assessing paleohydrologic controls on the hydrogen isotope compositions of leaf wax n-alkanes in Chinese peat deposits Palaeogeography, Palaeoclimatology, Palaeoecology, 516, 354-363. [本文引用: 1]
IshikawaNF, ChikaraishiY, TakanoY, SasakiY, TakizawaY, TsuchiyaM, TayasuI, NagataT, OhkouchiN (2018). A new analytical method for determination of the nitrogen isotopic composition of methionine: its application to aquatic ecosystems with mixed resources Limnology and Oceanography: Methods, 16, 607-620. [本文引用: 1]
IshikawaNF, TogashiH, KatoY, YoshimuraM, KohmatsuY, YoshimizuC, OgawaNO, OhteN, TokuchiN, OhkouchiN, TayasuI (2016). Terrestrial-aquatic linkage in stream food webs along a forest chronosequence: multi-isotopic evidence Ecology, 97, 1146-1158. URLPMID:27349092 [本文引用: 1]
JansenB, WiesenbergGLB (2017). Opportunities and limitations related to the application of plant-derived lipid molecular proxies in soil science Soil, 3, 211-234. [本文引用: 1]
JexCN, PateGH, BlythAJ, SpencerRGM, HernesPJ, KhanSJ, BakerA (2014). Lignin biogeochemistry: from modern processes to Quaternary archives Quaternary Science Reviews, 87, 46-59. [本文引用: 3]
JoergensenRG (2018). Amino sugars as specific indices for fungal and bacterial residues in soil Biology and Fertility of Soils, 54, 559-568. [本文引用: 4]
KirkTK, FarrellRL (1987). Enzymatic “combustion”: the microbial degradation of lignin Annual Reviews in Microbiology, 41, 465-501. [本文引用: 1]
KlotzbücherT, KalbitzK, CerliC, HernesPJ, KaiserK (2016). Gone or just out of sight? The apparent disappearance of aromatic litter components in soils Soil, 2, 325-335. DOI:10.5194/soil-2-325-2016URL [本文引用: 1]
K?gel-KnabnerI (2002). The macromolecular organic composition of plant and microbial residues as inputs to soil organic matter Soil Biology & Biochemistry, 34, 139-162. [本文引用: 1]
K?gel-KnabnerI (2017). The macromolecular organic composition of plant and microbial residues as inputs to soil organic matter: fourteen years on Soil Biology & Biochemistry, 105, A3-A8. DOI:10.1016/j.soilbio.2016.08.011URL [本文引用: 1]
K?gel-KnabnerI, GuggenbergerG, KleberM, KandelerE, KalbitzK, ScheuS, EusterhuesK, LeinweberP (2008). Organo-mineral associations in temperate soils: integrating biology, mineralogy, and organic matter chemistry Journal of Plant Nutrition and Soil Science, 171, 61-82. [本文引用: 1]
LehmannJ, KleberM (2015). The contentious nature of soil organic matter Nature, 528, 60-68. DOI:10.1038/nature16069URLPMID:26595271 [本文引用: 1] The exchange of nutrients, energy and carbon between soil organic matter, the soil environment, aquatic systems and the atmosphere is important for agricultural productivity, water quality and climate. Long-standing theory suggests that soil organic matter is composed of inherently stable and chemically unique compounds. Here we argue that the available evidence does not support the formation of large-molecular-size and persistent 'humic substances' in soils. Instead, soil organic matter is a continuum of progressively decomposing organic compounds. We discuss implications of this view of the nature of soil organic matter for aquatic health, soil carbon-climate interactions and land management.
LiangC, BalserTC (2012). Warming and nitrogen deposition lessen microbial residue contribution to soil carbon pool Nature Communications, 3, 1222. DOI: 10.1038/ncomms2224. DOI:10.1038/ncomms2224URLPMID:23187622 [本文引用: 1] Microorganisms have a role as gatekeepers for terrestrial carbon fluxes, either causing its release to the atmosphere through their decomposition activities or preventing its release by stabilizing the carbon in a form that cannot be easily decomposed. Although research has focused on microbial sources of greenhouse gas production, somewhat limited attention has been paid to the microbial role in carbon sequestration. However, increasing numbers of reports indicate the importance of incorporating microbial-derived carbon into soil stable carbon pools. Here we investigate microbial residues in a California annual grassland after a continuous 9-year manipulation of three environmental factors (elevated CO(2), warming and nitrogen deposition), singly and in combination. Our results indicate that warming and nitrogen deposition can both alter the fraction of carbon derived from microbes in soils, though for two very different reasons. A reduction in microbial carbon contribution to stable carbon pools may have implications for our predictions of global change impacts on soil stored carbon.
LiangC, ChengG, WixonDL, BalserTC (2011). An Absorbing Markov Chain approach to understanding the microbial role in soil carbon stabilization Biogeochemistry, 106, 303-309. [本文引用: 1]
LiangC, SchimelJP, JastrowJD (2017). The importance of anabolism in microbial control over soil carbon storage Nature Microbiology, 2, 17105. DOI: 10.1038/nmicrobiol.2017.105. URLPMID:28741607 [本文引用: 1]
LiuH, LiuWG (2015). Relationship of plant leaf wax n-alkanes molecular distribution characteristics and vegetation types Journal of Earth Environment, 6, 168-179. [本文引用: 1]
MaT, DaiGH, ZhuSS, ChenDM, ChenLT, LüXT, WangXB, ZhuJT, ZhangYJ, MaWH, HeJS, BaiYF, HanXG, FengXJ (2019). Distribution and preservation of root- and shoot-derived carbon components in soils across the Chinese-Mongolian grasslands Journal of Geophysical Research, 124, 420-431. [本文引用: 2]
MaT, ZhuSS, WangZH, ChenDM, DaiGH, FengBW, SuXY, HuHF, LiKH, HanWX, LiangC, BaiYF, FengXJ (2018). Divergent accumulation of microbial necromass and plant lignin components in grassland soils Nature Communications, 9, 3480. DOI: 10.1038/s41467-018-05891-1. DOI:10.1038/s41467-018-05891-1URLPMID:30154479 [本文引用: 2] The means through which microbes and plants contribute to soil organic carbon (SOC) accumulation remain elusive due to challenges in disentangling the complex components of SOC. Here we use amino sugars and lignin phenols as tracers for microbial necromass and plant lignin components, respectively, and investigate their distribution in the surface soils across Mongolian grasslands in comparison with published data for other grassland soils of the world. While lignin phenols decrease, amino sugars increase with SOC contents in all examined grassland soils, providing continental-scale evidence for the key role of microbial necromass in SOC accumulation. Moreover, in contrast to clay's control on amino sugar accumulation in fine-textured soils, aridity plays a central role in amino sugar accrual and lignin decomposition in the coarse-textured Mongolian soils. Hence, aridity shifts may have differential impacts on microbial-mediated SOC accumulation in grassland soils of varied textures.
McCarthyMD, BennerR, LeeC, FogelML (2007). Amino acid nitrogen isotopic fractionation patterns as indicators of heterotrophy in plankton, particulate, and dissolved organic matter Geochimica et Cosmochimica Acta, 71, 4727-4744. [本文引用: 2]
Medina ContrerasD, Cantera KintzJ, Sánchez GonzálezA, ManceraE (2018). Food web structure and trophic relations in a riverine mangrove system of the tropical eastern Pacific, central coast of Colombia Estuaries and Coasts, 41, 1511-1521. [本文引用: 1]
OhkouchiN, ChikaraishiY, CloseHG, FryB, LarsenT, MadiganDJ, McCarthyMD, McMahonKW, NagataT, NaitoYI, OgawaNO, PoppBN, SteffanS, TakanoY, TayasuI, WyattASJ, YamaguchiYT, YokoyamaY (2017). Advances in the application of amino acid nitrogen isotopic analysis in ecological and biogeochemical studies Organic Geochemistry, 113, 150-174. [本文引用: 3]
OttoA, ShunthirasinghamC, SimpsonMJ (2005). A comparison of plant and microbial biomarkers in grassland soils from the Prairie Ecozone of Canada Organic Geochemistry, 36, 425-448. [本文引用: 6]
OttoA, SimpsonMJ (2005). Degradation and preservation of vascular plant-derived biomarkers in grassland and forest soils from Western Canada Biogeochemistry, 74, 377-409. [本文引用: 1]
OttoA, SimpsonMJ (2006). Sources and composition of hydrolysable aliphatic lipids and phenols in soils from western Canada Organic Geochemistry, 37, 385-407. DOI:10.1016/j.orggeochem.2005.12.011URL [本文引用: 3]
SchmidtMW, TornMS, AbivenS, DittmarT, GuggenbergerG, JanssensIA, KleberM, Kogel-KnabnerI, LehmannJ, ManningDA, NannipieriP, RasseDP, WeinerS, TrumboreSE (2011). Persistence of soil organic matter as an ecosystem property Nature, 478, 49-56. URLPMID:21979045 [本文引用: 1]
SchoutenS, HopmansEC, DamstéJSS (2013). The organic geochemistry of glycerol dialkyl glycerol tetraether lipids: a review Organic Geochemistry, 54, 19-61. [本文引用: 4]
SollinsP, KramerMG, SwanstonC, LajthaK, FilleyT, AufdenkampeAK, WagaiR, BowdenRD (2009). Sequential density fractionation across soils of contrasting mineralogy: evidence for both microbial- and mineral-controlled soil organic matter stabilization Biogeochemistry, 96, 209-231. [本文引用: 2]
SowersTD, HoldenKL, CowardEK, SparksDL (2019). Dissolved organic matter sorption and molecular fractionation by naturally occurring bacteriogenic iron (Oxyhydr) oxides Environmental Science & Technology, 53, 4295-4304. URLPMID:30843682 [本文引用: 2]
SteffanSA, ChikaraishiY, CurrieCR, HornH, Gaines-DayHR, PauliJN, ZalapaJE, OhkouchiN (2015). Microbes are trophic analogs of animals Proceedings of the National Academy of Sciences of the United States of America, 112, 15119-15124. URLPMID:26598691 [本文引用: 1]
StevensonFJ (1982). Organic forms of nitrogen //Stevenson FJ. Nitrogen in Agricultural Soils. American Society of Agronomy, Madison. 67-122. [本文引用: 1]
ThevenotM, DignacMF, RumpelC (2010). Fate of lignins in soils: a review Soil Biology & Biochemistry, 42, 1200-1211. [本文引用: 5]
TierneyJ, SchoutenS, PitcherA, HopmansEC, Sinninghe DamstéJS (2012). Core and intact polar glycerol dialkyl glycerol tetraethers (GDGTs) in Sand Pond, Warwick, Rhode Island (USA): insights into the origin of lacustrine GDGTs Geochimica et Cosmochimica Acta, 77, 561-581. [本文引用: 1]
TippleBJ, BerkeMA, DomanCE, KhachaturyanS, EhleringerJR (2013). Leaf-wax n-alkanes record the plant-water environment at leaf flush Proceedings of the National Academy of Sciences of the United States of America, 110, 2659-2664. URLPMID:23359675 [本文引用: 1]
TreibsA (1936). Chlorophyll and hemin derivatives in organic mineral substances Angewandte Chemie, 49, 682-686. [本文引用: 1]
van der VoortTS, ZellCI, HagedornF, FengXJ, McIntyreCP, HaghipourN, Graf PannatierE, EglintoneTI (2017). Diverse soil carbon dynamics expressed at the molecular level Geophysical Research Letters, 44, 11840-11850. [本文引用: 2]
WangJZ, HoSSH, CaoJJ, HuangRJ, ZhouJM, ZhaoYZ, XuHM, LiuSX, WangGH, ShenZX, HanYM (2015). Characteristics and major sources of carbonaceous aerosols in PM2. 5 from Sanya, China Science of the Total Environment, 530-531, 110-119. DOI:10.1016/j.scitotenv.2015.05.005URLPMID:26026415 [本文引用: 1] PM2.5 samples were collected in Sanya, China in summer and winter in 2012/2013. Organic carbon (OC), elemental carbon (EC), and non-polar organic compounds including n-alkanes (n-C14-n-C40) and polycyclic aromatic hydrocarbons (PAHs) were quantified. The concentrations of these carbonaceous matters were generally higher in winter than summer. The estimated secondary organic carbon (OCsec) accounted for 38% and 54% of the total organic carbon (TOC) in winter and summer, respectively. The higher value of OCsec in addition to the presences of photochemically-produced PAHs in summer supports that photochemical conversions of organics are much active at the higher air temperatures and with stronger intense solar radiation. Carbon preference index (CPI) and percent contribution of wax n-alkanes suggest that anthropogenic sources were more dominant than derivation from terrestrial plants in Sanya. Diagnostic ratios of atmospheric PAHs further indicate that there was a wide mix of pollution sources in winter while fossil fuel combustion was the most dominant in summer. Positive Matrix Factorization (PMF) analysis with 18 PAHs in the winter samples found that motor vehicle emissions and biomass burning were the two main pollution sources, contributing 37.5% and 24.6% of the total quantified PAHs, respectively.
WangYY, WangH, HeJS, FengXJ (2017). Iron-mediated soil carbon response to water-table decline in an alpine wetland Nature Communications, 8, 15972. DOI: 10.1038/ncomms15972. URLPMID:28649988 [本文引用: 1]
WiesenbergG, DorodnikovM, KuzyakovY (2010). Source determination of lipids in bulk soil and soil density fractions after four years of wheat cropping Geoderma, 156, 267-277. [本文引用: 1]
WiesenbergG, SchneckenbergerK, SchwarkL, KuzyakovY (2012). Use of molecular ratios to identify changes in fatty acid composition of Miscanthus × giganteus (Greef et Deu.) plant tissue, rhizosphere and root-free soil during a laboratory experiment Organic Geochemistry, 46, 1-11. [本文引用: 1]
XieSC, LiangB, GuoJQ, YiY, EvershedRP, MaddyD, ChambersFM (2003). Biomarkers and the related global change Quaternary Sciences, 23, 521-528. [本文引用: 2]
XieSC, WangZY, WangHM, ChenFH, AnCB (2002). The occurrence of a grassy vegetation over the Chinese Loess Plateau since the last interglacier: the molecular fossil record Science in China Series D: Earth Sciences, 45, 53-62. [本文引用: 1]
ZhangXD, AmelungW (1996). Gas chromatographic determination of muramic acid, glucosamine, mannosamine, and galactosamine in soils Soil Biology & Biochemistry, 28, 1201-1206. [本文引用: 1]
ZhaoL, WuWC, XuXT, XuYP (2014). Soil organic matter dynamics under different land use in grasslands in Inner Mongolia (northern China) Biogeosciences, 11, 5103-5113. [本文引用: 1]
ZhaoMX, DupontL, EglintonG, TeeceM (2003). n-Alkane and pollen reconstruction of terrestrial climate and vegetation for N. W. Africa over the last 160 kyr Organic Geochemistry, 34, 131-143. DOI:10.1016/S0146-6380(02)00142-0URL [本文引用: 1]
Global-scale evidence for the refractory nature of riverine black carbon 1 2018
... 除了稳定同位素, 放射性同位素(14C)分析与生物标志物的结合是一个新兴的应用方向.Eglinton等(1996)首次发表了利用制备液相色谱开发的分离和纯化脂类生物标志物并进行自然丰度14C测定的分析方法.这一方法在探讨天然有机质的来源和环境驻留时间方面显示了巨大的优势(van der Voort et al., 2017; Douglas et al., 2018), 并逐渐拓展到木质素酚类(Hou et al., 2010; Feng et al., 2013)、黑炭(Coppola et al., 2018)等非脂类的生物标志物.例如, Feng等(2015)对比了环北极河流的河口沉积物中陆源生物标志物的14C含量, 发现木质素酚类和植物叶片角质单体比植物蜡质脂类年轻上万年, 前者主要来源于地表碳库, 而后者则主要追踪了深层的古老冻土碳, 其传输受到了冻土退化程度的影响.此类技术在土壤中的应用将有助于进一步解析土壤有机碳库的天然周转时间和在全球变化下的稳定性变化. ...
Formation of soil organic matter via biochemical and physical pathways of litter mass loss 1 2015
... 土壤有机质的来源和组成非常复杂, 不同生物来源(如微生物和植物、根系和地上凋落物等)对其形成的贡献和积累的影响机制尚不明确(Schmidt et al., 2011; Lehmann & Kleber, 2015).传统的凋落物降解实验和热力学理论都认为, 由酚类构成的木质素是植物凋落物中较难降解的成分, 木质素不仅可以降低凋落物的降解速率(Gleixner et al., 2001), 而且是稳定土壤碳库的主要成分(Derenne & Largeau, 2001).然而, 对土壤生物标志物的研究发现, 木质素酚类只在植物碎屑中大量存在, 而在矿质土壤中并没有发生积累(Klotzbücher et al., 2016); 此外, 在周转时间相对较长的土壤细质颗粒物中, 木质素酚类的相对丰度较小(K?gel-Knabner et al., 2008; Sollins et al., 2009).与之相反, 主要源于微生物残体的氨基糖则易于在矿质土壤和周转较慢的土壤组分中积累(Liang et al., 2011; Cotrufo et al., 2015), 说明微生物和植物残体的相对稳定性及其对土壤有机质的相对贡献仍存在争议.针对这个问题, Ma等(2018)利用生物标志物对比研究了中国-蒙古典型温带草地样带表层土壤中微生物残体(氨基糖)和植物木质素酚类的空间分布特征, 发现在干旱的温带荒漠土壤中, 微生物降解活动受到水分的限制, 土壤有机碳中木质素酚类的相对比例较高、氨基糖较低; 随着湿润度的增加, 木质素酚类的降解和氨基糖的积累同时增强.通过进一步整合全球草地土壤的生物标志物数据发现, 土壤有机碳含量和木质素酚类的丰度呈负相关关系, 与氨基糖则呈正相关关系.以上结果在区域尺度上证明了微生物残体碳在草地土壤有机碳积累中的关键作用, 改变了我们对土壤有机质形成和稳定机制的认识. ...
Sources of plant-derived carbon and stability of organic matter in soil: implications for global change 2 2009
... 生物标志物的检测与定量需根据待测化合物的性质(如是否可气化、是否带有荧光基团等)选择相应的分析仪器, 常用的分析仪器为气相色谱(GC)或高效液相色谱(HPLC)结合火焰离子化检测器(FID)/质谱检测器(MS)或二极管阵列检测器(DAD).目前常用气相色谱-质谱联用仪(GC-MS)测定蜡质脂类、角质单体、软木脂单体、PLFA、木质素酚类以及氨基糖(Otto et al., 2005; Jex et al., 2014); 在GC-MS上机分析前, 可利用三甲基硅烷基三氟乙酰胺(BSTFA)和少量吡啶对含有羟基和羧基的化合物进行衍生化(在70 ℃下加热3 h).质谱检测器通常采用电子电离法, 参照标准化合物的质谱图对目标化合物进行鉴定.根据目标化合物与定量内标的总离子色谱峰(TIC)面积比进行定量分析; 对于含有杂质的化合物色谱峰, 可以使用选择性离子检测模式(SIM)对特征离子进行定量(Crow et al., 2009; Sollins et al., 2009).需要注意的是, 由于结构不同的目标化合物具有不同的离子化特征和质谱响应比, 因此使用GC-MS定量分析时, 需要考虑目标化合物与标准品的响应差异.此外, 应通过过程空白(即不含有样品的提取物)的分析、平行样品的重复上机以及回收率的计算进行样品质量控制: 在过程空白中没有检测到目标化合物, 证明样品无污染; 回收内标的回收率应保证在80%-120%范围内; 平行样品浓度的相对标准偏差小于10%, 表明分析方法满足实验要求(Galy et al., 2011; Ma et al., 2019). ...
... 此外, GDGT常用超高效液相色谱-串联质谱联用仪(UPLC-MS/MS)的SIM模式进行定量分析(Schouten et al., 2013).氨基酸可在不同的衍生化后利用HPLC-DAD或GC-FID进行定量分析(Amelung & Zhang, 2001; Dittmar et al., 2001). ...
Spatial trends in a biomagnifying contaminant: application of amino acid compound-specific stable nitrogen isotope analysis to the interpretation of bird mercury levels 1 2018
A long-term decrease in the persistence of soil carbon caused by ancient Maya land use 1 2018
... 除了稳定同位素, 放射性同位素(14C)分析与生物标志物的结合是一个新兴的应用方向.Eglinton等(1996)首次发表了利用制备液相色谱开发的分离和纯化脂类生物标志物并进行自然丰度14C测定的分析方法.这一方法在探讨天然有机质的来源和环境驻留时间方面显示了巨大的优势(van der Voort et al., 2017; Douglas et al., 2018), 并逐渐拓展到木质素酚类(Hou et al., 2010; Feng et al., 2013)、黑炭(Coppola et al., 2018)等非脂类的生物标志物.例如, Feng等(2015)对比了环北极河流的河口沉积物中陆源生物标志物的14C含量, 发现木质素酚类和植物叶片角质单体比植物蜡质脂类年轻上万年, 前者主要来源于地表碳库, 而后者则主要追踪了深层的古老冻土碳, 其传输受到了冻土退化程度的影响.此类技术在土壤中的应用将有助于进一步解析土壤有机碳库的天然周转时间和在全球变化下的稳定性变化. ...
Analysis of hydrolysable neutral sugars in mineral soils: improvement of alditol acetylation for gas chromatographic separation and measurement 3 2010
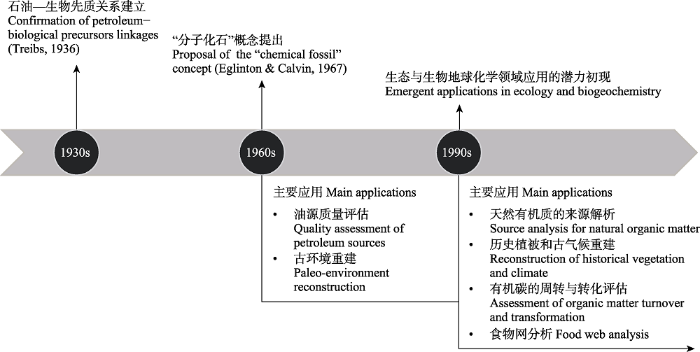
... 生物标志物是环境和地质体中记载了原始生物母质(如高等植物、微生物等)分子结构信息的有机化合物(Eglinton & Calvin, 1967; 谢树成等, 2003).20世纪30年代, Treibs (1936)首次从石油和油页岩样品中分离和鉴定出含金属的卟啉色素化合物, 建立了产物─生物先质的关系, 为生物标志物概念的提出奠定了理论基础(图1).20世纪60年代后期, Eglinton和Calvin (1967)将生物合成的、具有环境指示作用的有机化合物定义为“分子化石”、“生物标志物”或“化学化石”.此后, 生物标志物被广泛应用于评估油源质量(Dahl et al., 1999), 并在古环境重建中大放异彩(Didyk et al., 1978; Xie et al., 2002; Zhao et al., 2003). ...
... ).20世纪60年代后期, Eglinton和Calvin (1967)将生物合成的、具有环境指示作用的有机化合物定义为“分子化石”、“生物标志物”或“化学化石”.此后, 生物标志物被广泛应用于评估油源质量(Dahl et al., 1999), 并在古环境重建中大放异彩(Didyk et al., 1978; Xie et al., 2002; Zhao et al., 2003). ...
Gas chromatographic isolation of individual compounds from complex matrices for radiocarbon dating 1 1996
... 除了稳定同位素, 放射性同位素(14C)分析与生物标志物的结合是一个新兴的应用方向.Eglinton等(1996)首次发表了利用制备液相色谱开发的分离和纯化脂类生物标志物并进行自然丰度14C测定的分析方法.这一方法在探讨天然有机质的来源和环境驻留时间方面显示了巨大的优势(van der Voort et al., 2017; Douglas et al., 2018), 并逐渐拓展到木质素酚类(Hou et al., 2010; Feng et al., 2013)、黑炭(Coppola et al., 2018)等非脂类的生物标志物.例如, Feng等(2015)对比了环北极河流的河口沉积物中陆源生物标志物的14C含量, 发现木质素酚类和植物叶片角质单体比植物蜡质脂类年轻上万年, 前者主要来源于地表碳库, 而后者则主要追踪了深层的古老冻土碳, 其传输受到了冻土退化程度的影响.此类技术在土壤中的应用将有助于进一步解析土壤有机碳库的天然周转时间和在全球变化下的稳定性变化. ...
Molecular proxies for paleoclimatology 1 2008
... 随着环境地球科学和生态系统研究的兴起, 地球化学家逐渐认识到生物标志物在环境和生物地球化学领域中的应用潜力: 生物标志物的结构组成、丰度和同位素含量不仅可以用来指征生物来源, 还可以记录有机质产生时的环境信息以及降解过程中的转化特征(Eglinton & Eglinton, 2008).因此, 自20世纪90年代以来, 生物标志物被广泛地应用到现代生物地球化学和生态学的研究中, 在天然有机质(包括土壤、沉积物、气溶胶等)的来源解析(Wang et al., 2015; K?gel-Knabner, 2017), 历史植被和古气候重建(刘虎和刘卫国, 2015; Huang & Meyers, 2019), 有机碳的周转与转化评估(van der Voort et al., 2017; Bianchi et al., 2018), 食物网分析(Boecklen et al., 2011; Ohkouchi et al., 2017)等方向都有重要的应用, 已成为生态系统研究中新兴的示踪和指示方法.与此同时, 生物标志物的分析仍存在许多亟待突破的技术难点, 其在生态系统研究中的应用也存在一些认识上的误区.基于此, 本文将概述生态系统研究中常用的生物标志物的类型、指示意义、分析方法和典型的应用案例, 并提出生物标志物使用中存在的问题和未来发展方向, 旨在为使用生物标志物的生态学和环境科学研究者提供参考. ...
Multimolecular tracers of terrestrial carbon transfer across the pan-Arctic: C-14 characteristics of sedimentary carbon components and their environmental controls 1 2015
... 除了稳定同位素, 放射性同位素(14C)分析与生物标志物的结合是一个新兴的应用方向.Eglinton等(1996)首次发表了利用制备液相色谱开发的分离和纯化脂类生物标志物并进行自然丰度14C测定的分析方法.这一方法在探讨天然有机质的来源和环境驻留时间方面显示了巨大的优势(van der Voort et al., 2017; Douglas et al., 2018), 并逐渐拓展到木质素酚类(Hou et al., 2010; Feng et al., 2013)、黑炭(Coppola et al., 2018)等非脂类的生物标志物.例如, Feng等(2015)对比了环北极河流的河口沉积物中陆源生物标志物的14C含量, 发现木质素酚类和植物叶片角质单体比植物蜡质脂类年轻上万年, 前者主要来源于地表碳库, 而后者则主要追踪了深层的古老冻土碳, 其传输受到了冻土退化程度的影响.此类技术在土壤中的应用将有助于进一步解析土壤有机碳库的天然周转时间和在全球变化下的稳定性变化. ...
Increased cuticular carbon sequestration and lignin oxidation in response to soil warming 2 2008
... 木质素是维管植物细胞壁的主要成分, 其含量仅次于纤维素和半纤维素, 约占植物生物量干质量的15%-30% (Kirk & Farrell, 1987), 因此是植物凋落物和土壤有机质的重要组分.木质素是一种高分子量聚合物, 目前常用碱性氧化铜氧化法, 将木质素大分子分解为酚类单体(图2(IV)、2(V))进行定量分析(Thevenot et al., 2010).木质素酚类单体包括香草基酚类(Vanillyl, 即V类, 包括香草醛、香草酮和香草酸)、紫丁香基酚类(Syringyl, 即S类, 包括丁香醛、丁香酮和丁香酸)和肉桂基酚类(Cinnamyl, 即C类, 包括对香豆酸和阿魏酸)(Hedges & Mann, 1979; Thevenot et al., 2010).三类单体的总量常用来定量木质素酚类的含量.V类单体存在于所有维管束植物中, S类单体主要存在于被子植物中, C类单体主要存在于木本植物的叶片或草本植物中.因此, S/V可用于区分裸子植物(S/V ≈ 0)和被子植物(S/V = 0.6-4); C/V比值可以区分木本组织(C/V < 0.05)和草本组织(C/V > 0.2)(Thevenot et al., 2010; Jex et al., 2014).土壤中木质素的降解程度可以用V和S类单体的酸醛比(Ad/Al)来表征, 该比值随木质素的降解而增大(Jex et al., 2014).木质素酚类是经典的指示陆源有机碳输入的生物标志物, 在海洋沉积物的来源解析、古植被的重建方面有较为深入的应用.近些年, 木质素酚类也被广泛应用于解析土壤有机质的组成和降解变化(Feng et al., 2008; Zhao et al., 2014; Ma et al., 2018). ...
Differential mobilization of terrestrial carbon pools in Eurasian Arctic river basins 2 2013
... 生物标志物分析可用于土壤、沉积物、溶解性有机质以及生物样品(如植物、动物组织等).样品采集过程中应尽量使用不含有塑化剂等有机污染物的采样器和容器, 并注意防止交叉污染.由于单体化合物容易在湿热和紫外辐射等条件下发生降解(谢树成等, 2003; Eder et al., 2010), 因此以上样品在采集以后应尽快在冷冻(<-20 ℃)和避光条件下保存.在生物标志物提取之前, 应使用冷冻干燥法对土壤、沉积物、生物组织等环境样品进行前处理, 以去除样品中的水分.对于较为稳定的生物标志物(包括角质、软木脂、木质素酚类、氨基糖等), 冷冻干燥后的样品可以在长达数年的时间内保存生物标志物的结构和含量信息.对于易降解的生物标志物(例如PLFA、GDGT等), 则须在冷冻干燥之后尽快提取.糖类化合物(如氨基糖和中性糖)容易在冻融过程中分解, 因此样品不宜反复冻融.对于溶解性有机质, 可以根据拟分析的化合物类型, 选取特定的固相萃取柱对水样进行富集, 也可用冷冻干燥法浓缩溶解性有机质后进行提取(Feng et al., 2013; Sowers et al., 2019). ...
... 除了稳定同位素, 放射性同位素(14C)分析与生物标志物的结合是一个新兴的应用方向.Eglinton等(1996)首次发表了利用制备液相色谱开发的分离和纯化脂类生物标志物并进行自然丰度14C测定的分析方法.这一方法在探讨天然有机质的来源和环境驻留时间方面显示了巨大的优势(van der Voort et al., 2017; Douglas et al., 2018), 并逐渐拓展到木质素酚类(Hou et al., 2010; Feng et al., 2013)、黑炭(Coppola et al., 2018)等非脂类的生物标志物.例如, Feng等(2015)对比了环北极河流的河口沉积物中陆源生物标志物的14C含量, 发现木质素酚类和植物叶片角质单体比植物蜡质脂类年轻上万年, 前者主要来源于地表碳库, 而后者则主要追踪了深层的古老冻土碳, 其传输受到了冻土退化程度的影响.此类技术在土壤中的应用将有助于进一步解析土壤有机碳库的天然周转时间和在全球变化下的稳定性变化. ...
Stable isotopes as tracers of trophic interactions in marine mutualistic symbioses 1 2019
Examples of typical monomer structures for the main types of biomarkers introduced in this paper. Plant-derived biomarkers: plant wax lipids including long-chain <i>n</i>-alkanes (I) and fatty acids (II), etc. (<xref ref-type="bibr" rid="b59">Otto <i>et al</i>., 2005</xref>); cutin monomers such as short-chain ω-hydroxyalkanoic acid (III); lignin phenols such as vanillin (IV) and syringic acid (V), etc. (<xref ref-type="bibr" rid="b68">Thevenot <i>et al</i>., 2010</xref>); suberin monomers such as long-chain α,ω-alkanedioic acid (VI; <xref ref-type="bibr" rid="b59">Otto <i>et al</i>., 2005</xref>). Micorbial-derived biomarkers: phospholipid fatty acids (PLFAs) such as linoleic acid (<xref ref-type="bibr" rid="b27">VII; Frosteg?rd & B??th, 1996</xref>); glycerol dialkyl glycerol tetraethers (GDGTs) such as GDGT-2 (<xref ref-type="bibr" rid="b63">VIII; Schouten <i>et al</i>., 2013</xref>); amino sugars, including α-D-glucosamine (IX) and α-D-galactosamine (X), etc. (<xref ref-type="bibr" rid="b43">Joergensen, 2018</xref>). In addition, neutral sugars such as α-D-glucose (XI) and amino acids with side chain (R group; XII) can be synthesized by both plants and microbes.Fig. 2
生态系统研究中常用的生物标志物的分子构成、提取方法、研究意义和关键参数 ...
... ). Micorbial-derived biomarkers: phospholipid fatty acids (PLFAs) such as linoleic acid (VII; Frosteg?rd & B??th, 1996); glycerol dialkyl glycerol tetraethers (GDGTs) such as GDGT-2 (VIII; Schouten et al., 2013); amino sugars, including α-D-glucosamine (IX) and α-D-galactosamine (X), etc. (Joergensen, 2018). In addition, neutral sugars such as α-D-glucose (XI) and amino acids with side chain (R group; XII) can be synthesized by both plants and microbes. Fig. 2
Compound-specific isotopic analyses: a novel tool for reconstruction of ancient biogeochemical processes 1 1990
... 在生态系统的研究中, 生物标志物在土壤碳氮循环以及食物网中的应用尤为突出, 常用的生物标志物种类包括脂类、糖类、木质素酚类、氨基酸和氨基糖等(图2).这些生物标志物可以指征植物或微生物不同组织来源的有机质在土壤、沉积物或水溶性有机质中的相对丰度和降解状况, 因此可为解析天然有机质的来源、保存机制和动态变化提供重要的信息(表1).此外, 伴随着气相/液相色谱与稳定同位素质谱联用仪的开发(Hayes et al., 1990), 生物标志物的应用已从简单的含量分析发展到单体化合物的稳定同位素分析.与2H、13C、15N、18O等稳定同位素分析相结合, 生物标志物的单体同位素在植物水源示踪(Tipple et al., 2013), 微生物碳源分析(Fogel et al., 2016), 植物与生态系统碳周转(Ishikawa et al., 2016)等方面也有广泛的应用. ...
The molecularly- uncharacterized component of nonliving organic matter in natural environments 1 2000
... 生物标志物在有机质的来源解析和动态监测方面具有巨大的应用前景, 但是生物标志物在土壤中的应用需要充分考虑以下两方面的局限性.首先, 迄今为止, 土壤中大部分(>50%)的有机质在分子水平上仍然不能被解析或提取分析(Hedges et al., 2000); 任何一类生物标志物的总量仅占土壤总碳的很小一部分(通常为0.1%-10%).因此, 尽管生物标志物可以指示和追踪特定有机组分的环境行为, 但不能用以指征土壤碳库的总体特性.其次, 尽管许多生物标志物已经有了非常成熟和常规的分析提取方法, 但是由于土壤、沉积物等环境样品的复杂性, 不同样品之间的提取效率和背景杂质干扰仍然需要经过严格的评估.例如, 黏土矿物和铁氧化物对于木质素具有不同程度的吸附和物理保护作用(Sowers et al., 2019), 进而影响氧化铜催化氧化法的提取效率, 降低木质素酚的产率(Hernes et al., 2013; Wang et al., 2017).因此, 对于生物标志物的应用需要充分考虑样品的特性、提取效率以及影响目标化合物含量的潜在因素. ...
The characterization of plant tissues by their lignin oxidation products 1 1979
... 木质素是维管植物细胞壁的主要成分, 其含量仅次于纤维素和半纤维素, 约占植物生物量干质量的15%-30% (Kirk & Farrell, 1987), 因此是植物凋落物和土壤有机质的重要组分.木质素是一种高分子量聚合物, 目前常用碱性氧化铜氧化法, 将木质素大分子分解为酚类单体(图2(IV)、2(V))进行定量分析(Thevenot et al., 2010).木质素酚类单体包括香草基酚类(Vanillyl, 即V类, 包括香草醛、香草酮和香草酸)、紫丁香基酚类(Syringyl, 即S类, 包括丁香醛、丁香酮和丁香酸)和肉桂基酚类(Cinnamyl, 即C类, 包括对香豆酸和阿魏酸)(Hedges & Mann, 1979; Thevenot et al., 2010).三类单体的总量常用来定量木质素酚类的含量.V类单体存在于所有维管束植物中, S类单体主要存在于被子植物中, C类单体主要存在于木本植物的叶片或草本植物中.因此, S/V可用于区分裸子植物(S/V ≈ 0)和被子植物(S/V = 0.6-4); C/V比值可以区分木本组织(C/V < 0.05)和草本组织(C/V > 0.2)(Thevenot et al., 2010; Jex et al., 2014).土壤中木质素的降解程度可以用V和S类单体的酸醛比(Ad/Al)来表征, 该比值随木质素的降解而增大(Jex et al., 2014).木质素酚类是经典的指示陆源有机碳输入的生物标志物, 在海洋沉积物的来源解析、古植被的重建方面有较为深入的应用.近些年, 木质素酚类也被广泛应用于解析土壤有机质的组成和降解变化(Feng et al., 2008; Zhao et al., 2014; Ma et al., 2018). ...
Molecular trickery in soil organic matter: hidden lignin 1 2013
... 生物标志物在有机质的来源解析和动态监测方面具有巨大的应用前景, 但是生物标志物在土壤中的应用需要充分考虑以下两方面的局限性.首先, 迄今为止, 土壤中大部分(>50%)的有机质在分子水平上仍然不能被解析或提取分析(Hedges et al., 2000); 任何一类生物标志物的总量仅占土壤总碳的很小一部分(通常为0.1%-10%).因此, 尽管生物标志物可以指示和追踪特定有机组分的环境行为, 但不能用以指征土壤碳库的总体特性.其次, 尽管许多生物标志物已经有了非常成熟和常规的分析提取方法, 但是由于土壤、沉积物等环境样品的复杂性, 不同样品之间的提取效率和背景杂质干扰仍然需要经过严格的评估.例如, 黏土矿物和铁氧化物对于木质素具有不同程度的吸附和物理保护作用(Sowers et al., 2019), 进而影响氧化铜催化氧化法的提取效率, 降低木质素酚的产率(Hernes et al., 2013; Wang et al., 2017).因此, 对于生物标志物的应用需要充分考虑样品的特性、提取效率以及影响目标化合物含量的潜在因素. ...
Radiocarbon dating of individual lignin phenols: a new approach for establishing chronology of Late Quaternary lake sediments 1 2010
... 除了稳定同位素, 放射性同位素(14C)分析与生物标志物的结合是一个新兴的应用方向.Eglinton等(1996)首次发表了利用制备液相色谱开发的分离和纯化脂类生物标志物并进行自然丰度14C测定的分析方法.这一方法在探讨天然有机质的来源和环境驻留时间方面显示了巨大的优势(van der Voort et al., 2017; Douglas et al., 2018), 并逐渐拓展到木质素酚类(Hou et al., 2010; Feng et al., 2013)、黑炭(Coppola et al., 2018)等非脂类的生物标志物.例如, Feng等(2015)对比了环北极河流的河口沉积物中陆源生物标志物的14C含量, 发现木质素酚类和植物叶片角质单体比植物蜡质脂类年轻上万年, 前者主要来源于地表碳库, 而后者则主要追踪了深层的古老冻土碳, 其传输受到了冻土退化程度的影响.此类技术在土壤中的应用将有助于进一步解析土壤有机碳库的天然周转时间和在全球变化下的稳定性变化. ...
Assessing paleohydrologic controls on the hydrogen isotope compositions of leaf wax n-alkanes in Chinese peat deposits 1 2019
... 随着环境地球科学和生态系统研究的兴起, 地球化学家逐渐认识到生物标志物在环境和生物地球化学领域中的应用潜力: 生物标志物的结构组成、丰度和同位素含量不仅可以用来指征生物来源, 还可以记录有机质产生时的环境信息以及降解过程中的转化特征(Eglinton & Eglinton, 2008).因此, 自20世纪90年代以来, 生物标志物被广泛地应用到现代生物地球化学和生态学的研究中, 在天然有机质(包括土壤、沉积物、气溶胶等)的来源解析(Wang et al., 2015; K?gel-Knabner, 2017), 历史植被和古气候重建(刘虎和刘卫国, 2015; Huang & Meyers, 2019), 有机碳的周转与转化评估(van der Voort et al., 2017; Bianchi et al., 2018), 食物网分析(Boecklen et al., 2011; Ohkouchi et al., 2017)等方向都有重要的应用, 已成为生态系统研究中新兴的示踪和指示方法.与此同时, 生物标志物的分析仍存在许多亟待突破的技术难点, 其在生态系统研究中的应用也存在一些认识上的误区.基于此, 本文将概述生态系统研究中常用的生物标志物的类型、指示意义、分析方法和典型的应用案例, 并提出生物标志物使用中存在的问题和未来发展方向, 旨在为使用生物标志物的生态学和环境科学研究者提供参考. ...
A new analytical method for determination of the nitrogen isotopic composition of methionine: its application to aquatic ecosystems with mixed resources 1 2018
Lignin biogeochemistry: from modern processes to Quaternary archives 3 2014
... 木质素是维管植物细胞壁的主要成分, 其含量仅次于纤维素和半纤维素, 约占植物生物量干质量的15%-30% (Kirk & Farrell, 1987), 因此是植物凋落物和土壤有机质的重要组分.木质素是一种高分子量聚合物, 目前常用碱性氧化铜氧化法, 将木质素大分子分解为酚类单体(图2(IV)、2(V))进行定量分析(Thevenot et al., 2010).木质素酚类单体包括香草基酚类(Vanillyl, 即V类, 包括香草醛、香草酮和香草酸)、紫丁香基酚类(Syringyl, 即S类, 包括丁香醛、丁香酮和丁香酸)和肉桂基酚类(Cinnamyl, 即C类, 包括对香豆酸和阿魏酸)(Hedges & Mann, 1979; Thevenot et al., 2010).三类单体的总量常用来定量木质素酚类的含量.V类单体存在于所有维管束植物中, S类单体主要存在于被子植物中, C类单体主要存在于木本植物的叶片或草本植物中.因此, S/V可用于区分裸子植物(S/V ≈ 0)和被子植物(S/V = 0.6-4); C/V比值可以区分木本组织(C/V < 0.05)和草本组织(C/V > 0.2)(Thevenot et al., 2010; Jex et al., 2014).土壤中木质素的降解程度可以用V和S类单体的酸醛比(Ad/Al)来表征, 该比值随木质素的降解而增大(Jex et al., 2014).木质素酚类是经典的指示陆源有机碳输入的生物标志物, 在海洋沉积物的来源解析、古植被的重建方面有较为深入的应用.近些年, 木质素酚类也被广泛应用于解析土壤有机质的组成和降解变化(Feng et al., 2008; Zhao et al., 2014; Ma et al., 2018). ...
... ).土壤中木质素的降解程度可以用V和S类单体的酸醛比(Ad/Al)来表征, 该比值随木质素的降解而增大(Jex et al., 2014).木质素酚类是经典的指示陆源有机碳输入的生物标志物, 在海洋沉积物的来源解析、古植被的重建方面有较为深入的应用.近些年, 木质素酚类也被广泛应用于解析土壤有机质的组成和降解变化(Feng et al., 2008; Zhao et al., 2014; Ma et al., 2018). ...
... 生物标志物的检测与定量需根据待测化合物的性质(如是否可气化、是否带有荧光基团等)选择相应的分析仪器, 常用的分析仪器为气相色谱(GC)或高效液相色谱(HPLC)结合火焰离子化检测器(FID)/质谱检测器(MS)或二极管阵列检测器(DAD).目前常用气相色谱-质谱联用仪(GC-MS)测定蜡质脂类、角质单体、软木脂单体、PLFA、木质素酚类以及氨基糖(Otto et al., 2005; Jex et al., 2014); 在GC-MS上机分析前, 可利用三甲基硅烷基三氟乙酰胺(BSTFA)和少量吡啶对含有羟基和羧基的化合物进行衍生化(在70 ℃下加热3 h).质谱检测器通常采用电子电离法, 参照标准化合物的质谱图对目标化合物进行鉴定.根据目标化合物与定量内标的总离子色谱峰(TIC)面积比进行定量分析; 对于含有杂质的化合物色谱峰, 可以使用选择性离子检测模式(SIM)对特征离子进行定量(Crow et al., 2009; Sollins et al., 2009).需要注意的是, 由于结构不同的目标化合物具有不同的离子化特征和质谱响应比, 因此使用GC-MS定量分析时, 需要考虑目标化合物与标准品的响应差异.此外, 应通过过程空白(即不含有样品的提取物)的分析、平行样品的重复上机以及回收率的计算进行样品质量控制: 在过程空白中没有检测到目标化合物, 证明样品无污染; 回收内标的回收率应保证在80%-120%范围内; 平行样品浓度的相对标准偏差小于10%, 表明分析方法满足实验要求(Galy et al., 2011; Ma et al., 2019). ...
Amino sugars as specific indices for fungal and bacterial residues in soil 4 2018
... 在生态系统的研究中, 生物标志物在土壤碳氮循环以及食物网中的应用尤为突出, 常用的生物标志物种类包括脂类、糖类、木质素酚类、氨基酸和氨基糖等(图2).这些生物标志物可以指征植物或微生物不同组织来源的有机质在土壤、沉积物或水溶性有机质中的相对丰度和降解状况, 因此可为解析天然有机质的来源、保存机制和动态变化提供重要的信息(表1).此外, 伴随着气相/液相色谱与稳定同位素质谱联用仪的开发(Hayes et al., 1990), 生物标志物的应用已从简单的含量分析发展到单体化合物的稳定同位素分析.与2H、13C、15N、18O等稳定同位素分析相结合, 生物标志物的单体同位素在植物水源示踪(Tipple et al., 2013), 微生物碳源分析(Fogel et al., 2016), 植物与生态系统碳周转(Ishikawa et al., 2016)等方面也有广泛的应用.
Examples of typical monomer structures for the main types of biomarkers introduced in this paper. Plant-derived biomarkers: plant wax lipids including long-chain <i>n</i>-alkanes (I) and fatty acids (II), etc. (<xref ref-type="bibr" rid="b59">Otto <i>et al</i>., 2005</xref>); cutin monomers such as short-chain ω-hydroxyalkanoic acid (III); lignin phenols such as vanillin (IV) and syringic acid (V), etc. (<xref ref-type="bibr" rid="b68">Thevenot <i>et al</i>., 2010</xref>); suberin monomers such as long-chain α,ω-alkanedioic acid (VI; <xref ref-type="bibr" rid="b59">Otto <i>et al</i>., 2005</xref>). Micorbial-derived biomarkers: phospholipid fatty acids (PLFAs) such as linoleic acid (<xref ref-type="bibr" rid="b27">VII; Frosteg?rd & B??th, 1996</xref>); glycerol dialkyl glycerol tetraethers (GDGTs) such as GDGT-2 (<xref ref-type="bibr" rid="b63">VIII; Schouten <i>et al</i>., 2013</xref>); amino sugars, including α-D-glucosamine (IX) and α-D-galactosamine (X), etc. (<xref ref-type="bibr" rid="b43">Joergensen, 2018</xref>). In addition, neutral sugars such as α-D-glucose (XI) and amino acids with side chain (R group; XII) can be synthesized by both plants and microbes.Fig. 2
生态系统研究中常用的生物标志物的分子构成、提取方法、研究意义和关键参数 ...
... ); amino sugars, including α-D-glucosamine (IX) and α-D-galactosamine (X), etc. (Joergensen, 2018). In addition, neutral sugars such as α-D-glucose (XI) and amino acids with side chain (R group; XII) can be synthesized by both plants and microbes. Fig. 2
Examples of typical monomer structures for the main types of biomarkers introduced in this paper. Plant-derived biomarkers: plant wax lipids including long-chain <i>n</i>-alkanes (I) and fatty acids (II), etc. (<xref ref-type="bibr" rid="b59">Otto <i>et al</i>., 2005</xref>); cutin monomers such as short-chain ω-hydroxyalkanoic acid (III); lignin phenols such as vanillin (IV) and syringic acid (V), etc. (<xref ref-type="bibr" rid="b68">Thevenot <i>et al</i>., 2010</xref>); suberin monomers such as long-chain α,ω-alkanedioic acid (VI; <xref ref-type="bibr" rid="b59">Otto <i>et al</i>., 2005</xref>). Micorbial-derived biomarkers: phospholipid fatty acids (PLFAs) such as linoleic acid (<xref ref-type="bibr" rid="b27">VII; Frosteg?rd & B??th, 1996</xref>); glycerol dialkyl glycerol tetraethers (GDGTs) such as GDGT-2 (<xref ref-type="bibr" rid="b63">VIII; Schouten <i>et al</i>., 2013</xref>); amino sugars, including α-D-glucosamine (IX) and α-D-galactosamine (X), etc. (<xref ref-type="bibr" rid="b43">Joergensen, 2018</xref>). In addition, neutral sugars such as α-D-glucose (XI) and amino acids with side chain (R group; XII) can be synthesized by both plants and microbes.Fig. 2
Examples of typical monomer structures for the main types of biomarkers introduced in this paper. Plant-derived biomarkers: plant wax lipids including long-chain <i>n</i>-alkanes (I) and fatty acids (II), etc. (<xref ref-type="bibr" rid="b59">Otto <i>et al</i>., 2005</xref>); cutin monomers such as short-chain ω-hydroxyalkanoic acid (III); lignin phenols such as vanillin (IV) and syringic acid (V), etc. (<xref ref-type="bibr" rid="b68">Thevenot <i>et al</i>., 2010</xref>); suberin monomers such as long-chain α,ω-alkanedioic acid (VI; <xref ref-type="bibr" rid="b59">Otto <i>et al</i>., 2005</xref>). Micorbial-derived biomarkers: phospholipid fatty acids (PLFAs) such as linoleic acid (<xref ref-type="bibr" rid="b27">VII; Frosteg?rd & B??th, 1996</xref>); glycerol dialkyl glycerol tetraethers (GDGTs) such as GDGT-2 (<xref ref-type="bibr" rid="b63">VIII; Schouten <i>et al</i>., 2013</xref>); amino sugars, including α-D-glucosamine (IX) and α-D-galactosamine (X), etc. (<xref ref-type="bibr" rid="b43">Joergensen, 2018</xref>). In addition, neutral sugars such as α-D-glucose (XI) and amino acids with side chain (R group; XII) can be synthesized by both plants and microbes.Fig. 2
生态系统研究中常用的生物标志物的分子构成、提取方法、研究意义和关键参数 ...
... -alkanes (I) and fatty acids (II), etc. (Otto et al., 2005); cutin monomers such as short-chain ω-hydroxyalkanoic acid (III); lignin phenols such as vanillin (IV) and syringic acid (V), etc. (Thevenot et al., 2010); suberin monomers such as long-chain α,ω-alkanedioic acid (VI; Otto et al., 2005). Micorbial-derived biomarkers: phospholipid fatty acids (PLFAs) such as linoleic acid (VII; Frosteg?rd & B??th, 1996); glycerol dialkyl glycerol tetraethers (GDGTs) such as GDGT-2 (VIII; Schouten et al., 2013); amino sugars, including α-D-glucosamine (IX) and α-D-galactosamine (X), etc. (Joergensen, 2018). In addition, neutral sugars such as α-D-glucose (XI) and amino acids with side chain (R group; XII) can be synthesized by both plants and microbes. Fig. 2
生态系统研究中常用的生物标志物的分子构成、提取方法、研究意义和关键参数 ...
... ); suberin monomers such as long-chain α,ω-alkanedioic acid (VI; Otto et al., 2005). Micorbial-derived biomarkers: phospholipid fatty acids (PLFAs) such as linoleic acid (VII; Frosteg?rd & B??th, 1996); glycerol dialkyl glycerol tetraethers (GDGTs) such as GDGT-2 (VIII; Schouten et al., 2013); amino sugars, including α-D-glucosamine (IX) and α-D-galactosamine (X), etc. (Joergensen, 2018). In addition, neutral sugars such as α-D-glucose (XI) and amino acids with side chain (R group; XII) can be synthesized by both plants and microbes. Fig. 2
Examples of typical monomer structures for the main types of biomarkers introduced in this paper. Plant-derived biomarkers: plant wax lipids including long-chain <i>n</i>-alkanes (I) and fatty acids (II), etc. (<xref ref-type="bibr" rid="b59">Otto <i>et al</i>., 2005</xref>); cutin monomers such as short-chain ω-hydroxyalkanoic acid (III); lignin phenols such as vanillin (IV) and syringic acid (V), etc. (<xref ref-type="bibr" rid="b68">Thevenot <i>et al</i>., 2010</xref>); suberin monomers such as long-chain α,ω-alkanedioic acid (VI; <xref ref-type="bibr" rid="b59">Otto <i>et al</i>., 2005</xref>). Micorbial-derived biomarkers: phospholipid fatty acids (PLFAs) such as linoleic acid (<xref ref-type="bibr" rid="b27">VII; Frosteg?rd & B??th, 1996</xref>); glycerol dialkyl glycerol tetraethers (GDGTs) such as GDGT-2 (<xref ref-type="bibr" rid="b63">VIII; Schouten <i>et al</i>., 2013</xref>); amino sugars, including α-D-glucosamine (IX) and α-D-galactosamine (X), etc. (<xref ref-type="bibr" rid="b43">Joergensen, 2018</xref>). In addition, neutral sugars such as α-D-glucose (XI) and amino acids with side chain (R group; XII) can be synthesized by both plants and microbes.Fig. 2
生态系统研究中常用的生物标志物的分子构成、提取方法、研究意义和关键参数 ...
... ); glycerol dialkyl glycerol tetraethers (GDGTs) such as GDGT-2 (VIII; Schouten et al., 2013); amino sugars, including α-D-glucosamine (IX) and α-D-galactosamine (X), etc. (Joergensen, 2018). In addition, neutral sugars such as α-D-glucose (XI) and amino acids with side chain (R group; XII) can be synthesized by both plants and microbes. Fig. 2
... 此外, GDGT常用超高效液相色谱-串联质谱联用仪(UPLC-MS/MS)的SIM模式进行定量分析(Schouten et al., 2013).氨基酸可在不同的衍生化后利用HPLC-DAD或GC-FID进行定量分析(Amelung & Zhang, 2001; Dittmar et al., 2001). ...
Sequential density fractionation across soils of contrasting mineralogy: evidence for both microbial- and mineral-controlled soil organic matter stabilization 2 2009
... 生物标志物的检测与定量需根据待测化合物的性质(如是否可气化、是否带有荧光基团等)选择相应的分析仪器, 常用的分析仪器为气相色谱(GC)或高效液相色谱(HPLC)结合火焰离子化检测器(FID)/质谱检测器(MS)或二极管阵列检测器(DAD).目前常用气相色谱-质谱联用仪(GC-MS)测定蜡质脂类、角质单体、软木脂单体、PLFA、木质素酚类以及氨基糖(Otto et al., 2005; Jex et al., 2014); 在GC-MS上机分析前, 可利用三甲基硅烷基三氟乙酰胺(BSTFA)和少量吡啶对含有羟基和羧基的化合物进行衍生化(在70 ℃下加热3 h).质谱检测器通常采用电子电离法, 参照标准化合物的质谱图对目标化合物进行鉴定.根据目标化合物与定量内标的总离子色谱峰(TIC)面积比进行定量分析; 对于含有杂质的化合物色谱峰, 可以使用选择性离子检测模式(SIM)对特征离子进行定量(Crow et al., 2009; Sollins et al., 2009).需要注意的是, 由于结构不同的目标化合物具有不同的离子化特征和质谱响应比, 因此使用GC-MS定量分析时, 需要考虑目标化合物与标准品的响应差异.此外, 应通过过程空白(即不含有样品的提取物)的分析、平行样品的重复上机以及回收率的计算进行样品质量控制: 在过程空白中没有检测到目标化合物, 证明样品无污染; 回收内标的回收率应保证在80%-120%范围内; 平行样品浓度的相对标准偏差小于10%, 表明分析方法满足实验要求(Galy et al., 2011; Ma et al., 2019). ...
... 土壤有机质的来源和组成非常复杂, 不同生物来源(如微生物和植物、根系和地上凋落物等)对其形成的贡献和积累的影响机制尚不明确(Schmidt et al., 2011; Lehmann & Kleber, 2015).传统的凋落物降解实验和热力学理论都认为, 由酚类构成的木质素是植物凋落物中较难降解的成分, 木质素不仅可以降低凋落物的降解速率(Gleixner et al., 2001), 而且是稳定土壤碳库的主要成分(Derenne & Largeau, 2001).然而, 对土壤生物标志物的研究发现, 木质素酚类只在植物碎屑中大量存在, 而在矿质土壤中并没有发生积累(Klotzbücher et al., 2016); 此外, 在周转时间相对较长的土壤细质颗粒物中, 木质素酚类的相对丰度较小(K?gel-Knabner et al., 2008; Sollins et al., 2009).与之相反, 主要源于微生物残体的氨基糖则易于在矿质土壤和周转较慢的土壤组分中积累(Liang et al., 2011; Cotrufo et al., 2015), 说明微生物和植物残体的相对稳定性及其对土壤有机质的相对贡献仍存在争议.针对这个问题, Ma等(2018)利用生物标志物对比研究了中国-蒙古典型温带草地样带表层土壤中微生物残体(氨基糖)和植物木质素酚类的空间分布特征, 发现在干旱的温带荒漠土壤中, 微生物降解活动受到水分的限制, 土壤有机碳中木质素酚类的相对比例较高、氨基糖较低; 随着湿润度的增加, 木质素酚类的降解和氨基糖的积累同时增强.通过进一步整合全球草地土壤的生物标志物数据发现, 土壤有机碳含量和木质素酚类的丰度呈负相关关系, 与氨基糖则呈正相关关系.以上结果在区域尺度上证明了微生物残体碳在草地土壤有机碳积累中的关键作用, 改变了我们对土壤有机质形成和稳定机制的认识. ...
Dissolved organic matter sorption and molecular fractionation by naturally occurring bacteriogenic iron (Oxyhydr) oxides 2 2019
... 生物标志物分析可用于土壤、沉积物、溶解性有机质以及生物样品(如植物、动物组织等).样品采集过程中应尽量使用不含有塑化剂等有机污染物的采样器和容器, 并注意防止交叉污染.由于单体化合物容易在湿热和紫外辐射等条件下发生降解(谢树成等, 2003; Eder et al., 2010), 因此以上样品在采集以后应尽快在冷冻(<-20 ℃)和避光条件下保存.在生物标志物提取之前, 应使用冷冻干燥法对土壤、沉积物、生物组织等环境样品进行前处理, 以去除样品中的水分.对于较为稳定的生物标志物(包括角质、软木脂、木质素酚类、氨基糖等), 冷冻干燥后的样品可以在长达数年的时间内保存生物标志物的结构和含量信息.对于易降解的生物标志物(例如PLFA、GDGT等), 则须在冷冻干燥之后尽快提取.糖类化合物(如氨基糖和中性糖)容易在冻融过程中分解, 因此样品不宜反复冻融.对于溶解性有机质, 可以根据拟分析的化合物类型, 选取特定的固相萃取柱对水样进行富集, 也可用冷冻干燥法浓缩溶解性有机质后进行提取(Feng et al., 2013; Sowers et al., 2019). ...
... 生物标志物在有机质的来源解析和动态监测方面具有巨大的应用前景, 但是生物标志物在土壤中的应用需要充分考虑以下两方面的局限性.首先, 迄今为止, 土壤中大部分(>50%)的有机质在分子水平上仍然不能被解析或提取分析(Hedges et al., 2000); 任何一类生物标志物的总量仅占土壤总碳的很小一部分(通常为0.1%-10%).因此, 尽管生物标志物可以指示和追踪特定有机组分的环境行为, 但不能用以指征土壤碳库的总体特性.其次, 尽管许多生物标志物已经有了非常成熟和常规的分析提取方法, 但是由于土壤、沉积物等环境样品的复杂性, 不同样品之间的提取效率和背景杂质干扰仍然需要经过严格的评估.例如, 黏土矿物和铁氧化物对于木质素具有不同程度的吸附和物理保护作用(Sowers et al., 2019), 进而影响氧化铜催化氧化法的提取效率, 降低木质素酚的产率(Hernes et al., 2013; Wang et al., 2017).因此, 对于生物标志物的应用需要充分考虑样品的特性、提取效率以及影响目标化合物含量的潜在因素. ...
Examples of typical monomer structures for the main types of biomarkers introduced in this paper. Plant-derived biomarkers: plant wax lipids including long-chain <i>n</i>-alkanes (I) and fatty acids (II), etc. (<xref ref-type="bibr" rid="b59">Otto <i>et al</i>., 2005</xref>); cutin monomers such as short-chain ω-hydroxyalkanoic acid (III); lignin phenols such as vanillin (IV) and syringic acid (V), etc. (<xref ref-type="bibr" rid="b68">Thevenot <i>et al</i>., 2010</xref>); suberin monomers such as long-chain α,ω-alkanedioic acid (VI; <xref ref-type="bibr" rid="b59">Otto <i>et al</i>., 2005</xref>). Micorbial-derived biomarkers: phospholipid fatty acids (PLFAs) such as linoleic acid (<xref ref-type="bibr" rid="b27">VII; Frosteg?rd & B??th, 1996</xref>); glycerol dialkyl glycerol tetraethers (GDGTs) such as GDGT-2 (<xref ref-type="bibr" rid="b63">VIII; Schouten <i>et al</i>., 2013</xref>); amino sugars, including α-D-glucosamine (IX) and α-D-galactosamine (X), etc. (<xref ref-type="bibr" rid="b43">Joergensen, 2018</xref>). In addition, neutral sugars such as α-D-glucose (XI) and amino acids with side chain (R group; XII) can be synthesized by both plants and microbes.Fig. 2
生态系统研究中常用的生物标志物的分子构成、提取方法、研究意义和关键参数 ...
... ); cutin monomers such as short-chain ω-hydroxyalkanoic acid (III); lignin phenols such as vanillin (IV) and syringic acid (V), etc. (Thevenot et al., 2010); suberin monomers such as long-chain α,ω-alkanedioic acid (VI; Otto et al., 2005). Micorbial-derived biomarkers: phospholipid fatty acids (PLFAs) such as linoleic acid (VII; Frosteg?rd & B??th, 1996); glycerol dialkyl glycerol tetraethers (GDGTs) such as GDGT-2 (VIII; Schouten et al., 2013); amino sugars, including α-D-glucosamine (IX) and α-D-galactosamine (X), etc. (Joergensen, 2018). In addition, neutral sugars such as α-D-glucose (XI) and amino acids with side chain (R group; XII) can be synthesized by both plants and microbes. Fig. 2
生态系统研究中常用的生物标志物的分子构成、提取方法、研究意义和关键参数 ...
... 木质素是维管植物细胞壁的主要成分, 其含量仅次于纤维素和半纤维素, 约占植物生物量干质量的15%-30% (Kirk & Farrell, 1987), 因此是植物凋落物和土壤有机质的重要组分.木质素是一种高分子量聚合物, 目前常用碱性氧化铜氧化法, 将木质素大分子分解为酚类单体(图2(IV)、2(V))进行定量分析(Thevenot et al., 2010).木质素酚类单体包括香草基酚类(Vanillyl, 即V类, 包括香草醛、香草酮和香草酸)、紫丁香基酚类(Syringyl, 即S类, 包括丁香醛、丁香酮和丁香酸)和肉桂基酚类(Cinnamyl, 即C类, 包括对香豆酸和阿魏酸)(Hedges & Mann, 1979; Thevenot et al., 2010).三类单体的总量常用来定量木质素酚类的含量.V类单体存在于所有维管束植物中, S类单体主要存在于被子植物中, C类单体主要存在于木本植物的叶片或草本植物中.因此, S/V可用于区分裸子植物(S/V ≈ 0)和被子植物(S/V = 0.6-4); C/V比值可以区分木本组织(C/V < 0.05)和草本组织(C/V > 0.2)(Thevenot et al., 2010; Jex et al., 2014).土壤中木质素的降解程度可以用V和S类单体的酸醛比(Ad/Al)来表征, 该比值随木质素的降解而增大(Jex et al., 2014).木质素酚类是经典的指示陆源有机碳输入的生物标志物, 在海洋沉积物的来源解析、古植被的重建方面有较为深入的应用.近些年, 木质素酚类也被广泛应用于解析土壤有机质的组成和降解变化(Feng et al., 2008; Zhao et al., 2014; Ma et al., 2018). ...
... ; Thevenot et al., 2010).三类单体的总量常用来定量木质素酚类的含量.V类单体存在于所有维管束植物中, S类单体主要存在于被子植物中, C类单体主要存在于木本植物的叶片或草本植物中.因此, S/V可用于区分裸子植物(S/V ≈ 0)和被子植物(S/V = 0.6-4); C/V比值可以区分木本组织(C/V < 0.05)和草本组织(C/V > 0.2)(Thevenot et al., 2010; Jex et al., 2014).土壤中木质素的降解程度可以用V和S类单体的酸醛比(Ad/Al)来表征, 该比值随木质素的降解而增大(Jex et al., 2014).木质素酚类是经典的指示陆源有机碳输入的生物标志物, 在海洋沉积物的来源解析、古植被的重建方面有较为深入的应用.近些年, 木质素酚类也被广泛应用于解析土壤有机质的组成和降解变化(Feng et al., 2008; Zhao et al., 2014; Ma et al., 2018). ...
... ).三类单体的总量常用来定量木质素酚类的含量.V类单体存在于所有维管束植物中, S类单体主要存在于被子植物中, C类单体主要存在于木本植物的叶片或草本植物中.因此, S/V可用于区分裸子植物(S/V ≈ 0)和被子植物(S/V = 0.6-4); C/V比值可以区分木本组织(C/V < 0.05)和草本组织(C/V > 0.2)(Thevenot et al., 2010; Jex et al., 2014).土壤中木质素的降解程度可以用V和S类单体的酸醛比(Ad/Al)来表征, 该比值随木质素的降解而增大(Jex et al., 2014).木质素酚类是经典的指示陆源有机碳输入的生物标志物, 在海洋沉积物的来源解析、古植被的重建方面有较为深入的应用.近些年, 木质素酚类也被广泛应用于解析土壤有机质的组成和降解变化(Feng et al., 2008; Zhao et al., 2014; Ma et al., 2018). ...
Core and intact polar glycerol dialkyl glycerol tetraethers (GDGTs) in Sand Pond, Warwick, Rhode Island (USA): insights into the origin of lacustrine GDGTs 1 2012
Leaf-wax n-alkanes record the plant-water environment at leaf flush 1 2013
... 在生态系统的研究中, 生物标志物在土壤碳氮循环以及食物网中的应用尤为突出, 常用的生物标志物种类包括脂类、糖类、木质素酚类、氨基酸和氨基糖等(图2).这些生物标志物可以指征植物或微生物不同组织来源的有机质在土壤、沉积物或水溶性有机质中的相对丰度和降解状况, 因此可为解析天然有机质的来源、保存机制和动态变化提供重要的信息(表1).此外, 伴随着气相/液相色谱与稳定同位素质谱联用仪的开发(Hayes et al., 1990), 生物标志物的应用已从简单的含量分析发展到单体化合物的稳定同位素分析.与2H、13C、15N、18O等稳定同位素分析相结合, 生物标志物的单体同位素在植物水源示踪(Tipple et al., 2013), 微生物碳源分析(Fogel et al., 2016), 植物与生态系统碳周转(Ishikawa et al., 2016)等方面也有广泛的应用. ...
Chlorophyll and hemin derivatives in organic mineral substances 1 1936
... 生物标志物是环境和地质体中记载了原始生物母质(如高等植物、微生物等)分子结构信息的有机化合物(Eglinton & Calvin, 1967; 谢树成等, 2003).20世纪30年代, Treibs (1936)首次从石油和油页岩样品中分离和鉴定出含金属的卟啉色素化合物, 建立了产物─生物先质的关系, 为生物标志物概念的提出奠定了理论基础(图1).20世纪60年代后期, Eglinton和Calvin (1967)将生物合成的、具有环境指示作用的有机化合物定义为“分子化石”、“生物标志物”或“化学化石”.此后, 生物标志物被广泛应用于评估油源质量(Dahl et al., 1999), 并在古环境重建中大放异彩(Didyk et al., 1978; Xie et al., 2002; Zhao et al., 2003). ...
Diverse soil carbon dynamics expressed at the molecular level 2 2017
... 随着环境地球科学和生态系统研究的兴起, 地球化学家逐渐认识到生物标志物在环境和生物地球化学领域中的应用潜力: 生物标志物的结构组成、丰度和同位素含量不仅可以用来指征生物来源, 还可以记录有机质产生时的环境信息以及降解过程中的转化特征(Eglinton & Eglinton, 2008).因此, 自20世纪90年代以来, 生物标志物被广泛地应用到现代生物地球化学和生态学的研究中, 在天然有机质(包括土壤、沉积物、气溶胶等)的来源解析(Wang et al., 2015; K?gel-Knabner, 2017), 历史植被和古气候重建(刘虎和刘卫国, 2015; Huang & Meyers, 2019), 有机碳的周转与转化评估(van der Voort et al., 2017; Bianchi et al., 2018), 食物网分析(Boecklen et al., 2011; Ohkouchi et al., 2017)等方向都有重要的应用, 已成为生态系统研究中新兴的示踪和指示方法.与此同时, 生物标志物的分析仍存在许多亟待突破的技术难点, 其在生态系统研究中的应用也存在一些认识上的误区.基于此, 本文将概述生态系统研究中常用的生物标志物的类型、指示意义、分析方法和典型的应用案例, 并提出生物标志物使用中存在的问题和未来发展方向, 旨在为使用生物标志物的生态学和环境科学研究者提供参考. ...
... 除了稳定同位素, 放射性同位素(14C)分析与生物标志物的结合是一个新兴的应用方向.Eglinton等(1996)首次发表了利用制备液相色谱开发的分离和纯化脂类生物标志物并进行自然丰度14C测定的分析方法.这一方法在探讨天然有机质的来源和环境驻留时间方面显示了巨大的优势(van der Voort et al., 2017; Douglas et al., 2018), 并逐渐拓展到木质素酚类(Hou et al., 2010; Feng et al., 2013)、黑炭(Coppola et al., 2018)等非脂类的生物标志物.例如, Feng等(2015)对比了环北极河流的河口沉积物中陆源生物标志物的14C含量, 发现木质素酚类和植物叶片角质单体比植物蜡质脂类年轻上万年, 前者主要来源于地表碳库, 而后者则主要追踪了深层的古老冻土碳, 其传输受到了冻土退化程度的影响.此类技术在土壤中的应用将有助于进一步解析土壤有机碳库的天然周转时间和在全球变化下的稳定性变化. ...
Characteristics and major sources of carbonaceous aerosols in PM2. 5 from Sanya, China 1 2015
... 随着环境地球科学和生态系统研究的兴起, 地球化学家逐渐认识到生物标志物在环境和生物地球化学领域中的应用潜力: 生物标志物的结构组成、丰度和同位素含量不仅可以用来指征生物来源, 还可以记录有机质产生时的环境信息以及降解过程中的转化特征(Eglinton & Eglinton, 2008).因此, 自20世纪90年代以来, 生物标志物被广泛地应用到现代生物地球化学和生态学的研究中, 在天然有机质(包括土壤、沉积物、气溶胶等)的来源解析(Wang et al., 2015; K?gel-Knabner, 2017), 历史植被和古气候重建(刘虎和刘卫国, 2015; Huang & Meyers, 2019), 有机碳的周转与转化评估(van der Voort et al., 2017; Bianchi et al., 2018), 食物网分析(Boecklen et al., 2011; Ohkouchi et al., 2017)等方向都有重要的应用, 已成为生态系统研究中新兴的示踪和指示方法.与此同时, 生物标志物的分析仍存在许多亟待突破的技术难点, 其在生态系统研究中的应用也存在一些认识上的误区.基于此, 本文将概述生态系统研究中常用的生物标志物的类型、指示意义、分析方法和典型的应用案例, 并提出生物标志物使用中存在的问题和未来发展方向, 旨在为使用生物标志物的生态学和环境科学研究者提供参考. ...
Iron-mediated soil carbon response to water-table decline in an alpine wetland 1 2017
... 生物标志物在有机质的来源解析和动态监测方面具有巨大的应用前景, 但是生物标志物在土壤中的应用需要充分考虑以下两方面的局限性.首先, 迄今为止, 土壤中大部分(>50%)的有机质在分子水平上仍然不能被解析或提取分析(Hedges et al., 2000); 任何一类生物标志物的总量仅占土壤总碳的很小一部分(通常为0.1%-10%).因此, 尽管生物标志物可以指示和追踪特定有机组分的环境行为, 但不能用以指征土壤碳库的总体特性.其次, 尽管许多生物标志物已经有了非常成熟和常规的分析提取方法, 但是由于土壤、沉积物等环境样品的复杂性, 不同样品之间的提取效率和背景杂质干扰仍然需要经过严格的评估.例如, 黏土矿物和铁氧化物对于木质素具有不同程度的吸附和物理保护作用(Sowers et al., 2019), 进而影响氧化铜催化氧化法的提取效率, 降低木质素酚的产率(Hernes et al., 2013; Wang et al., 2017).因此, 对于生物标志物的应用需要充分考虑样品的特性、提取效率以及影响目标化合物含量的潜在因素. ...
Source determination of lipids in bulk soil and soil density fractions after four years of wheat cropping 1 2010
Use of molecular ratios to identify changes in fatty acid composition of Miscanthus × giganteus (Greef et Deu.) plant tissue, rhizosphere and root-free soil during a laboratory experiment 1 2012
 ,
, ,1,2,*, 王依云1,2, 刘婷1, 贾娟1, 戴国华1, 马田1,2, 刘宗广1
,1,2,*, 王依云1,2, 刘婷1, 贾娟1, 戴国华1, 马田1,2, 刘宗广1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT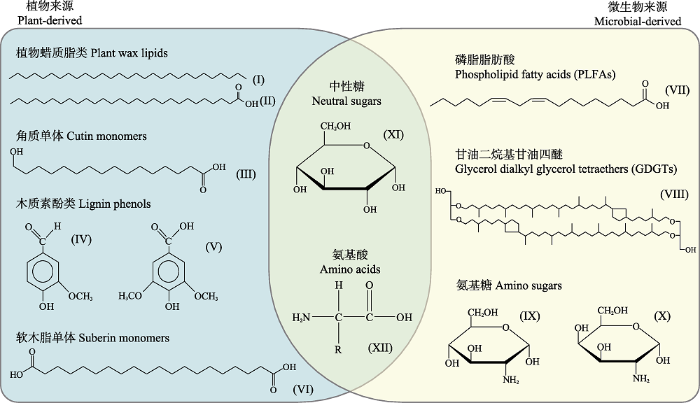 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}