冯银平
1 , 2 , 沈海花
1 , 2 , 罗永开
1 , 2 , 徐龙超
1 , 2 , 刘上石
1 , 2 , 朱言坤
1 , 2 , 赵梦颖
1 , 2 , 邢爱军
1 , 2 , 方精云
, 1 , 2 , 3 , * 1 中国科学院植物研究所植被与环境变化国家重点实验室, 北京 1000932 中国科学院大学, 北京 1000493 北京大学城市与环境学院地表过程分析与模拟教育部重点实验室, 北京 100871Effects of planting density on growth and biomass of Medicago sativa Yin-Ping FENG
1 , 2 , Hai-Hua SHEN
1 , 2 , Yong-Kai LUO
1 , 2 , Long-Chao XU
1 , 2 , Shang-Shi LIU
1 , 2 , Yan-Kun ZHU
1 , 2 , Meng-Ying ZHAO
1 , 2 , Ai-Jun XING
1 , 2 , Jing-Yun FANG
, 1 , 2 , 3 , * 1 State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China2 University of Chinese Academy of Sciences, Beijing 100049, China3 Key Laboratory for Earth Surface Processes of the Ministry of Education, College of Urban and Environmental Sciences, Peking University, Beijing 100871, China通讯作者: *
jyfang@urban.pku.edu.cn 编委: 邓建明
责任编辑: 李敏
收稿日期: 2019-06-25
接受日期: 2020-02-20
网络出版日期: 2020-03-20
基金资助: 中国科学院科技服务网络计划重点项目 (KFJ-STS-ZDTP-004 )中国科学院科技服务网络计划重点项目 (KFJ-STS-ZDTP-056 )
Corresponding authors: *
jyfang@urban.pku.edu.cn Received: 2019-06-25
Accepted: 2020-02-20
Online: 2020-03-20
Fund supported: Key Program of Science and Technology Service Network Initiative of Chinese Academy of Sciences (KFJ-STS-ZDTP-004 )Key Program of Science and Technology Service Network Initiative of Chinese Academy of Sciences (KFJ-STS-ZDTP-056 )
摘要 种植密度作为影响作物产量和品质的重要因素, 会造成植物对于光照、水分和养分的竞争。为研究种植密度对苜蓿生长与产量的影响, 在日光温室环境下, 以紫花苜蓿(
Medicago sativa )为材料, 设置25、100、400、800、1 500、2 000株·m
-2 , 共6个种植密度, 对紫花苜蓿的种群密度和生长状况进行了观测。结果表明, 各处理播种后15天的平均种植密度分别为25、100、373、745、1 255、1 938株·m
-2 ; 随着紫花苜蓿的生长, 除了低密度(25、100株·m
-2 )处理没有发生植株数量的变化外, 其余4个密度处理植株数量均有所减少, 即发生不同程度的自疏, 至第二茬收获时(播种后第187天)种群数量分别减少为297、571、759、839株·m
-2 。植株个体的株高、基径和分枝数量随着现存密度的增加呈指数下降; 个体生物量与现存密度的关系满足竞争密度效应的幂函数关系, 即随着密度的增加而减小。紫花苜蓿单位面积地上生物量符合最终产量恒定法则, 然而, 随着密度的增加, 地下生物量有先增加后减小的趋势。
关键词: 种植密度 ;
紫花苜蓿 ;
生长 ;
生物量 Abstract Aims- Planting density, as one of the most important factors affecting crop yield and quality, will result in plant competition for light, water and nutrients. The objective of this study is to explore the effect of planting density on growth and yield of
Medicago sativa population.
Methods The population density experiments of
M. sativa was conducted in a greenhouse with six planting densities, i.e. 25, 100, 400, 800, 1 500 and 2 000 plants·m
-2 . At each plot, we measured plant height, basal diameter, branch number, biomass, and number of survivors.
Important findings The results showed that the average planting density was 25, 100, 373, 745, 1 255 and 1 938 plants·m
-2 in the 15 days after sowing. With the growth of
M. sativa, except for the low density treatments (25 and 100 plants·m
-2 ), the number of plant individuals under other density treatments decreased, and self-thinning occurred at some degrees. At the second harvest (187 days after sowing), the number of surviving plants decreased to 297, 571, 759 and 839 plants·m
-2 , respectively. The plant height, basal diameter and branch number of individual plants decreased exponentially with the increase of existing density. The relationships between individual biomass and existing density followed the competitive density effect law, that is, individual biomass decreased with the increase of density. The results also showed that the aboveground biomass of
M. sativa per unit area has no significant differences among different densities, but the underground biomass tended to increase first and then decrease with the increase of planting density.
Keywords: planting density ;
Medicago sativa ;
growth ;
biomass PDF (1161KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 冯银平, 沈海花, 罗永开, 徐龙超, 刘上石, 朱言坤, 赵梦颖, 邢爱军, 方精云. 种植密度对苜蓿生长及生物量的影响. 植物生态学报, 2020, 44(3): 248-256. DOI:
10.17521/cjpe.2019.0157 FENG Yin-Ping, SHEN Hai-Hua, LUO Yong-Kai, XU Long-Chao, LIU Shang-Shi, ZHU Yan-Kun, ZHAO Meng-Ying, XING Ai-Jun, FANG Jing-Yun.
Effects of planting density on growth and biomass of Medicago sativa .
Chinese Journal of Plant Ecology , 2020, 44(3): 248-256. DOI:
10.17521/cjpe.2019.0157 豆科苜蓿属(
Medicago )全世界共有56个种(
耿华珠, 1995 ), 分一年生和多年生两种类型。在中国, 尤其是在西北地区, 多年生苜蓿已被种植了2 000多年, 由于其适应性广, 产草量高, 且富含蛋白质、维生素和矿物质等营养物质, 是我国经济价值最高的豆科牧草。与此同时, 苜蓿具有优良的固氮功能, 可有效提高土壤肥力; 株丛密集, 可减少风沙对地面裸土的侵蚀; 根系发达、穿透力强, 有效地保持水土, 改善土壤结构(
洪绂曾等, 2009 )。全球苜蓿种植面积稳定在约2 380万hm
2 , 我国的苜蓿种植面积约占全球总面积的21% (
李新一等, 2015 )。虽然我国苜蓿的种植面积很大, 但高品质的苜蓿种植面积仅有20万hm
2 (
侯龙鱼等, 2018 )。
除了优质苜蓿的种植面积偏少, 我国在苜蓿的种植方式、栽培管理等方面也比较落后, 使得苜蓿产量和质量一直处于较低的水平。有些多年生紫花苜蓿(
Medicago sativa )人工草地不仅生产力低, 而且造成了土壤水分的严重亏缺(
程积民等, 2005 ), 不合理的利用引起了一系列的环境问题。苜蓿作物产量和营养水平受苜蓿品种、施肥量、播种时间、种植株距和行距、刈割高度、刈割时间和次数等多种因素的影响(
Chocarro & Lloveras, 2015 ;
都帅等, 2016 ;
孙万斌等, 2017 ;
魏永鹏等, 2017 ;
Jáuregui et al ., 2019 )。其中, 种植密度作为影响作物产量和品质的重要因素, 会造成植物个体对于光照、水分、养分的竞争, 这种竞争会增加个体大小之间的差异(
Weiner & Damgaard, 2006 ;
Chu et al ., 2009 )。这种差异在个体和群体的性状上都有所体现。
植物种群的密度效应是种群生态学研究的一个核心问题。植物种群在资源有限的情况下, 保证每个个体的存活是困难的, 种群数量会减少, 即产生种群调节作用。在植物种群密度与生物量的关系方面, 涉及很多经典的科学问题, 前人也提出了众多经典的理论, 如最终产量恒定法则、自疏法则以及最适种植密度法则等(
Shinozaki & Kira, 1956 ;
Yoda et al ., 1963 ;
Harper, 1977 ;
Deng et al ., 2012 )。早期研究显示, 适当提高密度可以增加苜蓿的建植率和覆盖率, 是保证其高产的有效措施(
欧阳延生等, 2007 )。虽然在过去的几十年里, 有关种植密度对苜蓿影响的研究已有不少, 但多偏重于探究播种量对苜蓿产量以及品质的影响(
李丽等, 2012 ;
马克成和王秉龙, 2014 ); 从个体和群体两个方面研究种植密度影响苜蓿生长的报道较少。
本文通过观测紫花苜蓿在6个不同种植密度下的密度变化和生长情况, 研究了种群密度和生长之间的关系, 可为确定苜蓿生长的最佳种植密度提供参考。
1 材料和方法 1.1 实验材料 盆栽实验在中国科学院植物研究所温室内进行, 温室气温在16-28 ℃之间, 空气相对湿度为30%-60%。为了模拟野外的生长条件, 本研究使用内蒙古自治区呼伦贝尔农垦特泥河实验站的表层土壤(0-20 cm)。土壤风干后, 过2 mm孔径筛后充分混匀装盆, 填土高度15 cm, 每盆26 kg土。花盆长46 cm, 宽35 cm, 高20 cm。实验开始前测定土壤理化性质, 其中pH使用pH计测定, 全碳和全氮通过元素分析仪(Vario EL III, Elementar, Hanau, Germany)测定; 全磷和全钾利用电感耦合等离子体发射光谱仪(ICAP 6300 ICP-OES Spectrpmeter, Thermo Scientific, Cambridge, UK)测定; 有机质含量使用重铬酸钾氧化-外加热法; 水解氮含量用碱解-扩散法; 速效磷含量利用紫外可见分光光度计(UV-2550, UV-Visible Spectrophotometer, Shimadzu, Japan)测定; 速效钾含量用电感耦合等离子体发射光谱仪(ICAP 6300 ICP-OES Spectrpmeter, Thermo Scientific, Cambridge, UK)测定。
经测定, 土壤pH为7.58, 全碳含量为32.11 g·kg
-1 , 全氮含量为2.97 g·kg
-1 , 全磷含量为0.74 g·kg
-1 , 全钾含量为21.22 g·kg
-1 , 有机质含量为52.96 g·kg
-1 , 水解氮含量为228.85 mg·kg
-1 , 速效磷含量为41.41 mg·kg
-1 , 速效钾含量为280.89 mg·kg
-1 。
实验用紫花苜蓿种子为“科育1号”, 由中国科学院植物研究所提供。种子千粒质量为2.012 5 g, 发芽率为82.75%。根据种子发芽率、千粒质量计算播种量。播种前对种子进行处理, 在浓硫酸中浸泡5 min破坏种皮, 清水冲洗干净后在4 ℃低温下保存48 h。均匀地撒播在花盆中, 覆土5 mm。
1.2 实验设计 实验设A、B、C、D、E、F 6个密度水平, 分别对应种植密度为25、100、400、800、1 500、2 000株·m
-2 。根据发芽率和千粒质量称取相应的种子, 于2017年12月13日播种, 将播种后第15天的实际平均植株数量作为初始密度, 即25、100、373、745、1 255、1 938株·m
-2 。密度A有6次重复, 密度B有5次重复, 其他密度水平4次重复, 共27个花盆, 完全随机排列, 定期定量浇水。播种后0-87天为第一茬(2017年12月13日-2018年3月11日), 88-187天为第二茬(2018年3月12日-6月20日)。本文重点分析了第二茬植株个体性状及生物量与现存密度的关系。
1.3 测定指标和方法 (1)现存植株数量: 记录每盆中存活的植株数量。播种后第15-87天, 每隔20天统计一次, 第87天、第187天各统计一次。
(2)株高: 6月20日(播种后第187天)收获前测量挂牌植株的绝对高度。密度A中的植株全部挂牌, 其他密度在每盆中间部位选挂10株。
(3)基径: 获取地上生物量后, 使用电子游标卡尺测量植物贴地面处主茎的直径, 每盆测量3-5株挂牌植株。
(4)分枝数: 收获当天统计每盆挂牌植株的一级分枝数量。
(5)地上生物量: 随机选择每个花盆里的挂牌植株3株, 将地上部分齐地剪下, 称取鲜质量, 将每株的茎叶分开; 然后将每个花盆里余下植株的地上部分齐地刈割, 称量鲜质量, 然后将茎、叶分开, 置于65 ℃烘箱中烘干至恒质量后, 称量其干质量。
(6)地下生物量: 取出花盆中直径2 mm以上的根系, 在0.045 mm的筛子上用流水完全冲洗干净; 花盆中剩余的部分充分混合均匀, 称质量, 并取300 g土根混合样品, 在0.045 mm的筛子上用流水完全冲洗干净, 得到直径小于2 mm的细根。再把样品放入65 ℃烘箱中烘干至恒质量后, 称量其干质量。
1.4 统计分析 采用Excel 2013软件对数据进行整理和制图; 利用R 3.5.1软件进行单因素方差分析来分析密度对种群生物量及生物量分配的影响, 显著性水平设为0.05。方差分析前对所有数据进行方差齐性检验和正态分布检验, 对于有显著差异的数据用Tukey进行多重比较。利用线性回归分析密度与苜蓿个体平均生物量、株高、基径、分枝数的关系。
2 结果和分析 2.1 植物种群数量的变化 除了A和B处理种群密度没有发生数量变化外, C、D、E、F处理均发生不同程度的株数减少(
图1 )。在第一茬, 随着时间的进行, C、D、E、F 4个密度水平下的植株数量都逐渐下降, 死亡率分别为11.97%、11.25%、26.70%和39.32% (
图2 ), F密度水平下苜蓿经过87天的生长, 下降到1 171株·m
-2 , 减少了767株·m
-2 。在第二茬, 植株数量下降变缓, F密度处理植株数量下降为839株·m
-2 , 减少了332株·m
-2 , 相对于初始密度的死亡率为28.1%; 处理D、E的植株数量略有下降, 下降为571和848株·m
-2 , 死亡率为13.18%和20.5% (
图2 )。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1不同种植密度下紫花苜蓿种群的数量变化(平均值±标准误差)。A、B、C、D、E、F分别表示种植密度为25、100、400、800、1 500、2 000株·m-2 。 Fig. 1Quantity change of Medicago sativa population under different planting densities (mean ± SE ). A, B, C, D, E, F represents 25, 100, 400, 800, 1 500, 2 000 plants·m-2 planting densities. 图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2不同种植密度下的紫花苜蓿种群第一茬和第二茬的平均死亡率(平均值±标准误差)。A、B、C、D、E、F分别表示种植密度为25、100、400、800、1 500、2 000株·m-2 。 Fig. 2Average death rate of Medicago sativa population under different planting densities in the first and second harvests (mean ± SE ). A, B, C, D, E, F represents 25, 100, 400, 800, 1 500, 2 000 plants·m-2 planting densities. 2.2 个体性状与现存密度的关系 苜蓿个体的平均株高、基径和分枝数量与现存密度存在明显的指数关系(
图3 )。随着密度的增加, 植株平均株高呈现迅速下降的趋势, 当密度为200-400株·m
-2 时, 株高的下降趋势逐渐减缓, 而后趋于平稳(
图3 A)。最低密度处理(25株·m
-2 )下, 平均株高为101.6 cm, 而较高密度(739株·m
-2 )下, 平均株高为34.2 cm。因此, 较高的种群密度会抑制植株的高度生长。随着密度的增加, 平均基径呈现迅速下降的趋势, 之后基径的下降趋势逐渐减缓至趋于平稳。当密度为25株·m
-2 时平均基径最大, 为7.95 mm; 当密度为919株·m
-2 时, 平均基径最小, 为1.79 mm (
图3 B)。这表明密度对苜蓿基径的生长产生了明显的负调节作用。密度对于苜蓿单株平均分枝数也存在显著的影响: 密度水平为25株·m
-2 时, 苜蓿的单株平均分枝数量最大, 最高的每株平均达到21个分枝; 而随着密度的增加, 单株平均分枝数量先是急剧下降, 然后逐渐减缓, 最后稳定在每株平均有3-5个分枝(
图3 C)。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3现存密度与紫花苜蓿平均株高(A)、平均基径(B)和平均分枝数(C)的关系。 Fig. 3Relationship between density of survivors and mean plant height (A), basal diameter (B) and branch number (C) for Medicago sativa . 植物个体生物量与密度呈现幂函数的关系, 图中直线的斜率为-0.972 (
图4 )。随着密度的增加, 植物个体生物量显著减小, 密度最低种群的个体为密度最高种群个体的53.4倍。不同密度下, 植物个体生物量与植物的株高、基径、分枝数量有着极为密切的关系。随着株高、基径和分枝数量的增加, 植物个体生物量呈现出指数增长的趋势(
图5 )。
图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4紫花苜蓿植物个体生物量与现存密度的关系(横纵坐标轴均为对数坐标轴)。 Fig. 4Relationships between individual biomass and density of survivors for Medicago sativa. The horizontal and vertical axes are both logarithmic scale. 图5 新窗口打开 |
下载原图ZIP |
生成PPT 图5紫花苜蓿植物个体生物量与平均株高(A)、平均基径(B)和平均分枝数(C)的关系。 Fig. 5Relationships between individual plant biomass and mean plant height (A), basal diameter (B) and branch number (C) for Medicago sativa . 2.3 不同种植密度下种群生物量的变化 苜蓿第二茬生物量测定结果表明, 不同种植密度水平下的单位面积干草生物量差异不显著。但单位面积鲜草生物量在不同密度下有显著差异, F密度的鲜草产量为1 139.5 g·m
-2 , 显著高于密度A、B、C下的鲜草生物量, 最大密度(F密度)种群的生物量比中等密度(C密度)种群的生物量增加了约29% (
表1 )。种植密度对于苜蓿种群的地下生物量有显著影响, 随着密度的增加, 苜蓿种群地下生物量呈现先增加后降低的趋势。A密度的地下生物量显著低于C、D密度(
表1 ); C密度的地下生物量最大, 为146.7 g·m
-2 , 约为最低密度种群的1.5倍。
Table 1 表1 表1 不同种植密度对紫花苜蓿单位面积生物量的影响(平均值±标准误差)
Table 1
Effect of planting density on biomass for Medicago sativa (mean ± SE ) 处理 单位面积干草生物量-2 ) 单位面积鲜草生物量-2 ) 单位面积地下生物量-2 ) A 236.9 ± 41.9a 924.7 ± 62.2a 100.0 ± 23.6a B 256.5 ± 26.4a 902.9 ± 97.6a 125.8 ± 12.9ab C 243.7 ± 8.0a 886.1 ± 81.9a 146.7 ± 16.4b D 246.8 ± 10.1a 968.0 ± 70.2ab 144.5 ± 14.2b E 255.0 ± 19.0a 1 074.3 ± 139.9ab 129.6 ± 7.5ab F 248.4 ± 16.4a 1 139.5 ± 65.8b 133.2 ± 21.6ab
Results based on data from the second harvest. Different lowercase letters (a, b) in the same column indicate significant difference among different planting density at p < 0.05 level. A, B, C, D, E, F represents 25, 100, 400, 800, 1 500, 2 000 plants·m-2 planting density. 第二茬测定结果。同列数字右上角不同小写字母(a, b)表示不同种植密度处理间差异显著(p < 0.05)。A、B、C、D、E、F分别表示种植密度为25、100、400、800、1 500、2 000株·m-2 。 新窗口打开 |
下载CSV 种植密度对苜蓿种群的生物量分配没有显著的影响, 不同密度下苜蓿种群的根冠比、叶茎比之间均无显著差异(
表2 )。
Table 2 表2 表2 不同种植密度对紫花苜蓿生物量分配的影响(平均值±标准误差)
Table 2
Effect of planting density on biomass allocation for Medicago sativa (mean ± SE ) 处理 Treatment 根冠比 Root-shoot ratio 叶茎比 Leaf-stem ratio A 0.45 ± 0.20 0.82 ± 0.05 B 0.49 ± 0.10 0.95 ± 0.12 C 0.60 ± 0.08 0.97 ± 0.13 D 0.58 ± 0.04 0.77 ± 0.05 E 0.53 ± 0.10 0.74 ± 0.07 F 0.53 ± 0.06 0.88 ± 0.11
Results based on data from the second harvest. A, B, C, D, E, F represents 25, 100, 400, 800, 1 500, 2 000 plants·m-2 planting density. 第二茬测定结果, A、B、C、D、E、F分别表示种植密度为25、100、400、800、1 500、2 000株·m-2 。 新窗口打开 |
下载CSV 3 讨论 3.1 种植密度对植物个体的影响 种植密度能引起生长资源的强制分配, 密度的增加导致种内竞争的产生, 影响种群中植物的单株生长量和生物量(
Harper, 1977 ;
李博等, 2000 )。在很大程度上, 密度的制约作用使得种群内部个体的特征发生相应的变化。本研究中, 紫花苜蓿个体的性状随密度的变化发生了很显著的变化, 表现出极显著的密度制约特性: 随着种植密度的增加, 植物个体的株高、分枝数、基径、生物量均减小(
图3 ,
图4 )。在生长空间、光照、土壤水分和养分基本相同的情况下, 不同种植密度的同一植物种群基本具有相同的环境最大容纳量。当空间和资源有限时, 植物个体的生长可能会被抑制, 个体间产生竞争。当密度增加时, 改变自身的形态和结构是植物最易采取的、最经济的适应对策(
Silvertown & Lovett-Doust, 1993 )。尽管有研究表明, 植株高度不受密度的影响(
杜汉强等, 2004 ), 但株高随着种植密度的增加而减小也是一种“耐受”策略, 通过减缓自身的生长速度增加自身对外界胁迫的耐受力, 保证现有的尽可能多的植株的存活, 这与以往的研究结果(
陈静等, 2017 )一致。密度较小的种群在生长过程中能获得的光照、水分、养分以及空间方面的资源比较充足, 个体间的竞争较弱, 植物倾向于增加枝条或分蘖数量来提高个体生物量(
王俊峰和冯玉龙, 2004 )。而在种植密度增加, 资源紧缺的环境下, 植物倾向于减少枝条的数量和质量(
Soleymani et al ., 2011 ), 从而降低植物个体的生物量(
Wang et al ., 2019 )。另外, 关于基径与种群密度的关系,
Reineke (1933) 针对冠层密闭且同龄的植物种群提出了“立地密度法则”, 即
N =
kr -1.605 (
N 为种群密度,
r 为植物的平均直径,
k 为常数)。也就是说, 随着种群密度的增加, 植物的平均单株直径会减小, 两者之间存在权衡关系(
Adams et al ., 2015 )。
植物的形态发育影响个体生物量(
Hecht et al ., 2019 )。构成苜蓿生物量的主要因素有单茎质量、茎长、基径、侧枝数和叶片数(
孟凯等, 2019 )。本实验中, 不同密度下, 植物个体生物量与植物个体的平均株高、基径、分枝数量联系密切。随着密度的降低, 植物个体株高、基径和分枝数量增加, 植物个体生物量逐渐增长。这是因为苜蓿可在根的基部产生根冠芽并成长为分枝, 在外界环境竞争压力较小的情况下, 单株植物可以增加分枝的数量形成一个小的“种群” (
Cain et al ., 1991 ), 进而提高单株个体的生产力(
Kays & Harper, 1974 )。植物生长过程中会争夺有限的资源, 当种内竞争日趋激烈时, 植物个体之间彼此干扰, 许多个体陆续死亡, 种群密度降低(
方精云, 1992 )。在自疏过程中植物的平均个体质量与种群密度间的关系通常符合-3/2 (
Yoda et al ., 1963 )或-4/3 (
Enquist et al ., 1998 )自疏法则。本研究中, 个体平均生物量与密度关系为-1.0, 符合最终产量恒定法则(
Kira et al ., 1953 )。一方面, 自疏法则中存活个体数量与生物量的关系是在种群密度由大变小的过程中得到的; 另一方面, 苜蓿根基部可产生根冠芽, 发育成枝条后会有水平方向的伸展, 出现匍匐生长的情况。该情况下, 植株几何形状发生变化, 会出现平均个体生物量-密度关系的斜率接近-1的情况(
王刚, 1993 ), 即形成最终产量恒定的状态。
3.2 种植密度对现存密度和生物量的影响 密度是自然界的重要选择压力(
Japhet et al ., 2009 )。在密度的作用下, 植物种群内的植株互相竞争可以获得的资源, 主要包括光照、水分、矿物质等(
黎磊等, 2011 )。
Hakl等(2011) 对高密度下的苜 蓿种群数量动态进行了观测, 7年间植株数量由 5 000株·m
-2 降至57株·m
-2 。有研究表明, 苜蓿植株数量在第一生长年急剧下降, 第二生长年缓慢下降, 第三生长年趋于平稳(
王彦华等, 2017 )。本实验的研究结果显示, 初始密度对紫花苜蓿种群数量的动态变化有显著的影响, 前期随着生长时间的增长, 较高密度(1 255、1 938株·m
-2 )水平下的植株数量迅速下降, 随后缓慢下降; 中等密度(373、475株·m
-2 )水平下的植株数量一直较缓慢下降, 而较低密度(25、100株·m
-2 )水平下的植株数量则基本不变。随着生长时间的延长, 初始密度对于植物种群数量的影响有减弱的趋势。产生这种动态变化的原因可能是: 较高密度下当植物种群生长到一定阶段, 种内竞争日趋激烈(
方精云, 1992 ), 植物个体之间彼此相互干扰, 许多个体陆续死亡(
张泽浦等, 2000 ); 而密度处于较低水平时, 植株个体的生存压力较小, 在外界条件没有较大波动的情况下, 种群数量维持着较为平稳的状态, 死亡率为0 (
方精云, 1991 ;
朴顺姬等, 1997 )。
单位面积生物量是评价植物种群生产力的重要指标, 国内外在种植密度对苜蓿产量的影响方面有很多的研究, 但结果有所不同。有研究证明, 增加植株密度, 可以提高作物的产量(
Deng et al ., 2012 ), 有效地抑制杂草生长(
Bybee-Finley et al ., 2017 ;
Gibson et al ., 2017 )。
王彦华等(2017) 的研究则表明, 不同密度水平下苜蓿的产量没有差别。最近的研究结果发现, 当苜蓿的播种量低于10 kg·hm
-2 时, 播种量增加可以提高产量, 而当播种量高于10 kg·hm
-2 时, 产量并不随着播种量的增加而增加(
Berti & Samarappuli, 2018 )。在本实验中, 随着种植密度的增加, 单位面积干草产量间差异不显著, 种群地上与地下的最终产量都保持恒定, 符合最终产量恒定法则。这表明了在外部条件一致的情况下, 苜蓿种群经过充分的生长后, 群体达到了最大的资源利用率, 单位面积生物量保持恒定, 而与初始种植密度无关。此前有大量的以作物为研究对象的实验证明, 在一定密度范围内, 最终产量恒定的法则是适用的(
Kira et al ., 1953 ;
方精云等, 1991 ;
樊星等, 2016 )。
苜蓿是以收获茎叶为主的牧草, 并且叶片中蛋白质的含量远高于茎, 因此, 叶茎比是衡量苜蓿品质的重要指标(
刘东霞等, 2015 )。苜蓿作为多年生植物, 根的基部产生根冠芽发育为分枝, 具有表型可塑性(
Benedek & Englert, 2019 ), 可通过改变植株的形态特征(如: 叶茎比、根冠比等)适应环境, 进一步改变植物生产力和其生物量分配。以往关于种群密度是否影响植物生物量分配的研究结果尚无定论。
范高华等(2017) 认为, 植物各器官生物量均受种群密度的显著影响, 并且茎和地上生物量分配均随种群密度的增大而减小。但在本研究中, 不同密度下苜蓿种群的根冠比、叶茎比均无显著差异, 各器官生物量分配比例不受种群密度影响。同一种植物不同密度水平下, 种群的根冠比主要受环境因素影响(
平晓燕等, 2007 ), 与密度无关(
方华和孔凡斌, 2003 ;
Deru et al ., 2016 )。
4 结论 本研究通过对紫花苜蓿在6个不同初始密度水平下的温室生长情况的观测, 探讨了种植密度对苜蓿生长及单位面积生物量的影响。结果发现, 种植密度对于苜蓿种群的植株数量有显著的影响: 初始密度为25和100株·m
-2 处理的植株数量保持不变; 初始密度越高, 苜蓿生长前期植株数量的变化越大, 随着生长时间的延长, 初始密度为1 938和1 255株·m
-2 的处理植株数量趋于一致。生长后期, 植物种群对资源的利用达到最大化, 密度对苜蓿单位面积的生物量没有显著影响, 符合最终产量恒定法则。
从个体水平上看, 植物个体的株高、基径、分枝数量随着密度的降低呈现增加的趋势, 植物个体平均质量也随着株高、基径、分枝数量的增加而增加。另外, 植物个体平均质量与密度的关系满足竞争密度效应的幂函数关系。本研究为温室中苜蓿生长第一年第二茬的数据, 温室条件下很难反映自然界真实而复杂的状况, 要确认不同的种植密度对苜蓿多年生长的影响及在野外种植中的效果还有待进一步探究。此外, 本实验没有进行生长季内植物生长动态跟踪和生物量取样, 因此, 需要进一步完善。
致谢 感谢中国科学院植物研究所张文浩研究员、潘庆民研究员, 呼伦贝尔生态产业技术研究院郝建玺副院长、武海明站长在实验前期工作中给予的帮助。
[1] Adams CB Erickson JE Campbell DN Singh MP Rebolledo JP 2015 ). Effects of row spacing and population density on yield of sweet sorghum: applications for harvesting as billetsAgronomy Journal, 1831 -1836 . DOI:
10.2134/agronj14.0295 URL [本文引用: 1] [2] Benedek V Englert P 2019 ). The effect of ramet mortality on clonal plant growthTheory in Biosciences, 215 -221 . DOI:
10.1007/s12064-019-00274-6 URL [本文引用: 1] [3] Berti MT Samarappuli D 2018 ). How does sowing rate affect plant and stem density, forage yield, and nutritive value in glyphosate-tolerant alfalfa?Agronomy, 169 . DOI: 10.3390/agronomy8090169 . DOI:
10.3390/agronomy8090169 URL [本文引用: 1] [4] Bybee-Finley KA Mirsky SB Ryan MR 2017 ). Crop biomass not species richness drives weed suppression in warm- season annual grass—Legume intercrops in the northeastWeed Science, 669 -680 . DOI:
10.1017/wsc.2017.25 URL [本文引用: 1] [5] Cain ML Pacala SW Silander JA 1991 ). Stochastic simulation of clonal growth in the tall goldenrod,Solidago altissima. Oecologia, 477 -485 . [本文引用: 1] [6] Chen J Zhao CZ Wang JW Zhao LC 2017 ). Canopy structure and radiation interception of Salix matsudana: stand density dependent relationshipsChinese Journal of Plant Ecology, 661 -669 . DOI:
10.17521/cjpe.2016.0257 URL [本文引用: 1] [ 陈静 , 赵成章 , 王继伟 , 赵连春 (2017 ). 不同密度旱柳的树冠构型与光截获植物生态学报 , 41, 661 -669 .] DOI:
10.17521/cjpe.2016.0257 URL [本文引用: 1] [7] Cheng JM Wan HE Wang J 2005 ). Alfalfa growth and its relation with soil water status in loess hilly and gully regionChinese Journal of Applied Ecology, 435 -438 . [本文引用: 1] [ 程积民 , 万惠娥 , 王静 (2005 ). 黄土丘陵区紫花苜蓿生长与土壤水分变化应用生态学报 , 16, 435 -438 .] [本文引用: 1] [8] Chocarro C Lloveras J 2015 ). The effect of row spacing on alfalfa seed and forage production under irrigated Mediterranean agricultural conditionsGrass and Forage Science, 651 -660 . DOI:
10.1111/gfs.2015.70.issue-4 URL [本文引用: 1] [9] Chu CJ Weiner J Maestre FT Xiao S Wang YS Li Q Yuan JL Zhao LQ Ren ZW Wang G 2009 ). Positive interactions can increase size inequality in plant populationsJournal of Ecology, 1401 -1407 . DOI:
10.1111/jec.2009.97.issue-6 URL [本文引用: 1] [10] Deng JM Ran JZ Wang ZQ Fan ZX Wang GX Ji MF Liu J Wang Y Liu JQ Brown JH 2012 ). Models and tests of optimal density and maximal yield for crop plantsProceedings of the National Academy of Sciences of the United States of America, 15823 -15828 . [本文引用: 2] [11] Deru J Schilder H an der Schoot J R Van Eekeren N 2016 ). No Trade-off Between Root Biomass and Aboveground Production in Lolium perenne. Breeding in a World of Scarcity 289 -292 . [本文引用: 1] [12] Du HQ Niu YC Zhao XL Chen SE 2004 ). Sowing density effects on the major characteristics of alfalfaPratacultural Science, 42 -45 . [本文引用: 1] [ 杜汉强 , 牛一川 , 赵晓玲 , 陈双恩 (2004 ). 不同播种密度对紫花苜蓿主要性状的影响草业科学 , 21, 42 -45 .] [本文引用: 1] [13] Du S You SH Liu Y Sun L Gegentu , Jia YS 2016 ). Effect of different clipping periods and heights on alfalfa qualityActa Agrestia Sinica, 874 -878 . [本文引用: 1] [ 都帅 , 尤思涵 , 刘燕 , 孙林 , 格根图 , 贾玉山 (2016 ). 不同刈割时期与刈割高度对苜蓿品质的影响草地学报 , 24, 874 -878 .] [本文引用: 1] [14] Enquist BJ Brown JH West GB 1998 ). Allometric scaling of plant energetics and population densityNature, 163 -165 . DOI:
10.1038/25977 URL [本文引用: 1] [15] Fan GH Huang YX Zhao XY Shen XJ 2017 ). Effect of population density on the allometric growth of Agriophyllum squarrosum Acta Prataculturae Sinica 3 ), 53 -64 . [本文引用: 1] [ 范高华 , 黄迎新 , 赵学勇 , 神祥金 (2017 ). 种群密度对沙米异速生长的影响草业学报 , 26(3 ), 53 -64 .] [本文引用: 1] [16] Fan X Cai J Liu JP You MH Fan X Wang SS 2016 ). Module traits and biomass allocation of Humulus scandens seedlings during population self-thinningChinese Journal of Ecology 2926 -2934 . URL [本文引用: 1] Humulus scandens is a dioecious climbing herb. In April, a uniformly distributed wild
H. scandens population was selected and managed only for controlling weeds. From the beginning of May, some parameters of
H. scandens seedling, including density and height of population, individual’s module traits and biomass allocation, were determined for six consecutive times at an interval of 10 d. The purpose of this study was to analyze the relationships among population density, individual’s module traits and biomass allocation during population selfthinning, and to explore the selfthinning law of
H. scandens population. The results showed that
H. scandens seedling was characterized by a significant selfthinning phenomenon. Population density decreased by 71% within 60 days. During seedling from upright growth to lateral growth, density and height of population were significantly reduced. Due to selfthinning, stem traits of remained plants were significantly changed in the following order: internode length > main stem length > stem diameter for stem traits; leaf traits varied in the following order: leaf area > petiole length > leaf thickness > leaf width > leaf length > total leaf number > reserved leaf number; and root traits: total root length > root volume > root number > root length > maximum root length. Component biomass and single plant biomass of remained plants were significantly elevated, yet biomass per unit area declined gradually. Allocation ratio of aboveground biomass of remained plants was relatively stable (
P >0.05), while the ratios of root to stem and leaf to stem changed significantly (
P <0.01). There was highly significant allometric relationship among leaf, stem, petiole biomass, root biomass and aboveground biomass (
P <0.01). Leaf and stem presented a constant growth pattern with the increase of aboveground biomass. Meanwhile, leaf, petiole and root presented an allometric growth pattern with the increasing aboveground biomass. There was a highly significant correlation between aboveground biomass and leaf, stem, petiole and root biomass (
P <0.01). The allometric relationship between stem biomass and density abided by the rule of final output constant. Leaf, petiole and root biomass did not follow -3/2 selfthinning rule, nor -4/3 selfthinning rule and -1 selfthinning rule. Both aboveground biomass and total single plant biomass were significantly correlated with the density (
P <0.01). For remained plants, the allometric relationships among aboveground biomass, total biomass and density followed the rule of final output constant.
[ 樊星 , 蔡捡 , 刘金平 , 游明鸿 , 范宣 , 王思思 (2016 ). 葎草种群自疏过程中幼苗构件性状及生物量分配变化生态学杂志 , 35, 2926 -2934 .] URL [本文引用: 1] Humulus scandens is a dioecious climbing herb. In April, a uniformly distributed wild
H. scandens population was selected and managed only for controlling weeds. From the beginning of May, some parameters of
H. scandens seedling, including density and height of population, individual’s module traits and biomass allocation, were determined for six consecutive times at an interval of 10 d. The purpose of this study was to analyze the relationships among population density, individual’s module traits and biomass allocation during population selfthinning, and to explore the selfthinning law of
H. scandens population. The results showed that
H. scandens seedling was characterized by a significant selfthinning phenomenon. Population density decreased by 71% within 60 days. During seedling from upright growth to lateral growth, density and height of population were significantly reduced. Due to selfthinning, stem traits of remained plants were significantly changed in the following order: internode length > main stem length > stem diameter for stem traits; leaf traits varied in the following order: leaf area > petiole length > leaf thickness > leaf width > leaf length > total leaf number > reserved leaf number; and root traits: total root length > root volume > root number > root length > maximum root length. Component biomass and single plant biomass of remained plants were significantly elevated, yet biomass per unit area declined gradually. Allocation ratio of aboveground biomass of remained plants was relatively stable (
P >0.05), while the ratios of root to stem and leaf to stem changed significantly (
P <0.01). There was highly significant allometric relationship among leaf, stem, petiole biomass, root biomass and aboveground biomass (
P <0.01). Leaf and stem presented a constant growth pattern with the increase of aboveground biomass. Meanwhile, leaf, petiole and root presented an allometric growth pattern with the increasing aboveground biomass. There was a highly significant correlation between aboveground biomass and leaf, stem, petiole and root biomass (
P <0.01). The allometric relationship between stem biomass and density abided by the rule of final output constant. Leaf, petiole and root biomass did not follow -3/2 selfthinning rule, nor -4/3 selfthinning rule and -1 selfthinning rule. Both aboveground biomass and total single plant biomass were significantly correlated with the density (
P <0.01). For remained plants, the allometric relationships among aboveground biomass, total biomass and density followed the rule of final output constant.
[17] Fang H Kong FB 2003 ). Study on biomass and its allocation of different density loblolly pineJournal of Fujian College of Forestry, 182 -185 . [本文引用: 1] [ 方华 , 孔凡斌 (2003 ). 不同密度火炬松林生物量及其分配福建林学院学报 , 23, 182 -185 .] [本文引用: 1] [18] Fang JY 1991 ). A Theoretical Approach to the Change of Density in Plant Populations. Transactions of the Ecological Society of Chinese Youth (1) 183 -190 . [本文引用: 1] [ 方精云 (1991 ). 植物种群密度变化的理论. 青年生态****论丛(一) . 中国科学技术出版社, 北京. 183 -190 .] [本文引用: 1] [19] Fang JY 1992 ). Self-thinning rule in plant populationRural Eco-Environment, 2 ), 7 -12 . [本文引用: 2] [ 方精云 (1992 ). 植物种群的自然稀疏法则农村生态环境 , 8(2 ), 7 -12 .] [本文引用: 2] [20] Fang JY Kan M Yamakura T 1991 ). Relationship between population growth and population density in monocultures of Larix leptolepis Acta Botanica Sinica 949 -957 . [本文引用: 1] [ 方精云 , 菅诚 , 山仓拓夫 (1991 ). 日本落叶松模拟种群的生长与密度的关系植物学报 , 33, 949 -957 .] [本文引用: 1] [21] Geng HZ 1995 ). China Alfalfa 334 -338 . [本文引用: 1] [ 耿华珠 (1995 ). 中国苜蓿 . 中国农业出版社, 北京. 334 -338 .] [本文引用: 1] [22] Gibson DJ Young BG Wood AJ Bardgett R 2017 ). Can weeds enhance profitability? Integrating ecological concepts to address crop-weed competition and yield qualityJournal of Ecology, 900 -904 . DOI:
10.1111/1365-2745.12785 URL [本文引用: 1] [23] Hakl J Fuksa P ?antr??ek J Má?ková K 2011 ). The development of lucerne root morphology traits under high initial stand density within a seven year periodPlant Soil and Environment, 2 ), 81 -87 . DOI:
10.17221/PSE URL [本文引用: 1] [24] Harper JL 1977 ). Population biology of plants 151 -236 . [本文引用: 2] [25] Hecht VL Temperton VM Nagel KA Rascher U Pude R Postma JA 2019 ). Plant density modifies root system architecture in spring barley (Hordeum vulgare L.) through a change in nodal root numberPlant and Soil, 179 -200 . DOI:
10.1007/s11104-018-3764-9 URL [本文引用: 1] [26] Hong FZ Lu XS Gao HW 2009 ). Alfalfa Science 13 -23 . [本文引用: 1] [ 洪绂曾 , 卢欣石 , 高洪文 (2009 ). 苜蓿科学 . 中国农业出版社, 北京. 13 -23 .] [本文引用: 1] [27] Hou LY Yang J Zhang QQ Mao XT Song SH Bai WM Pan QM Zhou QP Zhang WH 2018 ). Establishment and management of alfalfa pasture in cold regions of ChinaChinese Science Bulletin, 1 -13 . [本文引用: 1] [ 侯龙鱼 , 杨杰 , 张强强 , 毛小涛 , 宋世环 , 白文明 , 潘庆民 , 周青平 , 张文浩 (2018 ). 高寒地区苜蓿人工草地建植技术科学通报 , 63, 1 -13 .] [本文引用: 1] [28] Japhet W Zhou DW Zhang HX Zhang HX Yu T 2009 ). Evidence of phenotypic plasticity in the response of Fagopyrum esculentum to population density and sowing dateJournal of Plant Biology 303 -311 . DOI:
10.1007/s12374-009-9037-7 URL [本文引用: 1] Phenotypic plasticity is an important attribute that enables plants to survive across a range of environments. We conducted two experiments to investigate the plasticity of architectural traits and biomass ratios for
Fagopyrum esculentum Moench in response to population density and sowing date. These included (1) inter-planting distances of 5, 10, 15, or 20cm; and (2) sowing on 25 July, 1 August, 5 August, or 10 August. Many traits exhibited phenotypic plasticity that was coupled with changes in plant size. However, variations in leaf/mass ratio from either experiment, as well as leaf/root ratios in response to sowing date, were independent of size. When coefficients of variation were computed, some consistency was found in the magnitude of trait plasticity for both density and sowing date. For each experiment, leaf/root ratios, leaf/mass ratios, and stem/mass ratios were most plastic. Although this suggests that biomass ratios could be more responsive to environmental changes, a wide array of traits should be considered if we are to fully understand the mechanism for these phenomena.
[29] Jáuregui JM Mills A Black DBS Wigley K Ridgway HJ Moot DJ 2019 ). Yield components of lucerne were affected by sowing dates and inoculation treatmentsEuropean Journal of Agronomy, 1 -12 . DOI:
10.1016/j.eja.2018.10.005 URL [本文引用: 1] [30] Kays S Harper JL 1974 ). The regulation of plant and tiller density in a grass swardThe Journal of Ecology, 97 -105 . DOI:
10.2307/2258882 URL [本文引用: 1] [31] Kira T Ogawa H Sakazaki N 1953 ). Intraspecific competition among higher plants. I Competition-density-yield interrelationship in regularly dispersed populationJournal of the Institute of Polytechnics, Series D, 1 -16 . [本文引用: 2] [32] Li B Yang C Lin P 2000 ). Ecology [本文引用: 1] [ 李博 , 杨持 , 林鹏 (2000 ). 生态学 . 高等教育出版社, 北京.] [本文引用: 1] [33] Li L Li N Sheng JD Wang H 2012 ). Effects of nitrogen fertilizer and planting density on alfalfa growth and seed yieldActa Agrectia Sinica, 54 -57, 62 . [本文引用: 1] [ 李丽 , 李宁 , 盛建东 , 王皓 (2012 ). 施氮量和种植密度对紫花苜蓿生长及种子产量的影响草地学报 , 20, 54 -57, 62 .] [本文引用: 1] [34] Li L Zhou DW Sheng LX 2011 ). Density dependence- determined plant biomass allocation patternChinese Journal of Ecology, 1579 -1589 . [本文引用: 1] [ 黎磊 , 周道玮 , 盛连喜 (2011 ). 密度制约决定的植物生物量分配格局生态学杂志 , 30, 1579 -1589 .] [本文引用: 1] [35] Li XY Luo J Tian SX Zhang SY 2015 ). Analysis of the overall situation of alfalfa production in ChinaChina Dairy Cattle, 16 ), 58 -64 . [本文引用: 1] [ 李新一 , 罗峻 , 田双喜 , 张书义 (2015 ). 我国苜蓿生产总体形势分析中国奶牛 , (16 ), 58 -64 .] [本文引用: 1] [36] Liu DX Liu GH Yang ZM 2015 ). The effects of planting and harvesting factors on hay yield and stem-leaf ratio of Medicago sativaActa Prataculturae Sinica, 3 ), 48 -57 . [本文引用: 1] [ 刘东霞 , 刘贵河 , 杨志敏 (2015 ). 种植及收获因子对紫花苜蓿干草产量和茎叶比的影响草业学报 , 24(3 ), 48 -57 .] [本文引用: 1] [37] Ma KC Wang BL 2014 ). The effects of row spacing and sowing rates on seed yield and quality ofMedicago sativa. Shaanxi Journal of Agricultural Sciences, 8 ), 6 -8 . [本文引用: 1] [ 马克成 , 王秉龙 (2014 ). 不同行距及播量对紫花苜蓿种子产量和质量的影响陕西农业科学 , 60(8 ), 6 -8 .] [本文引用: 1] [38] Meng K Li XY Jia ZY Jin HQ Mi FG 2019 ). Effects of planting density on the growth, yield and nutritional quality of alfalfa in central Inner MongoliaActa Prataculturae Sinica, 7 ), 73 -81 . [本文引用: 1] [ 孟凯 , 李星月 , 贾振宇 , 靳慧卿 , 米福贵 (2019 ). 种植密度对内蒙古中部地区苜蓿生长,饲草产量及营养品质影响草业学报 , 28(7 ), 73 -81 .] [本文引用: 1] [39] Ouyang YS Yu XG Xu GH Li XH Liu SH Dai ZH 2007 ). The effect of sowing rates and cutting frequency on the fresh yield of the alfalfaActa Agrectia Sinica, 196 -198 . [本文引用: 1] [ 欧阳延生 , 于徐根 , 徐桂花 , 李翔宏 , 刘水华 , 戴征煌 (2007 ). 播种量与刈割对紫花苜蓿产草量的影响草地学报 , 15, 196 -198 .] [本文引用: 1] [40] Piao SJ Yang C Huang SF Song MH 1997 ). Density and growth dynamics of Leymus chinensis populationActa Phytoecologica Sinica 60 -66 . [本文引用: 1] [ 朴顺姬 , 杨持 , 黄绍峰 , 宋明华 (1997 ). 羊草种群密度与生长动态研究植物生态学报 , 21, 60 -66 .] [本文引用: 1] [41] Ping XY Jia BR Yuan WP Wang FY Wang YH Zhou L Xu ZZ Zhou GS 2007 ). Biomass allocation of Leymus chinensis population: a dynamic simulation studyChinese Journal of Applied Ecology 2699 -2704 . URL [本文引用: 1] [ 平晓燕 , 贾丙瑞 , 袁文平 , 王风玉 , 王玉辉 , 周莉 , 许振柱 , 周广胜 (2007 ). 羊草种群生物量分配动态模拟应用生态学报 , 12, 2699 -2704 .] URL [本文引用: 1] [42] Reineke LH 1933 ). Perfecting a stand-density index for even-aged forestsJournal of Agricultural Research, 627 -638 . [本文引用: 1] [43] Shinozaki K Kira T 1956 ). Intraspecific competition among higher plants. VII. Logistic theory of the C-D effectJournal of the Institute of Polytechnics, 69 -82 . [本文引用: 1] [44] Silvertown J Lovett-Doust J 1993 ). Introduction to Plant Population Biology. 3rd ed. Blackwell Science, LondonUK. [本文引用: 1] [45] Soleymani A Shahrajabian MH Naranjani L 2011 ). Determination of the suitable planting date and plant density for different cultivars of barley (Hordeum vulgare L.) in FarsAfrican Journal of Plant Science, 284 -286 . [本文引用: 1] [46] Sun WB Feng GG Ma HL Liu Q Hou XY Mu HB 2017 ). Nutrition characteristics of different alfalfa varieties in different growth stagesGrassland and Turf, 2 ), 63 -68 . [本文引用: 1] [ 孙万斌 , 冯刚刚 , 马晖玲 , 刘强 , 侯向阳 , 穆怀彬 (2017 ). 不同紫花苜蓿品种在不同生育期营养品质特性的比较草原与草坪 , 37(2 ), 63 -68 .] [本文引用: 1] [47] Wang G 1993 ). The general model in the self-thinning process of plantsJournal of Lanzhou University (Natural Sciences )4 ), 215 -218 . [本文引用: 1] [ 王刚 (1993 ). 关于植物自疏过程的一般模型兰州大学学报(自然科学版) , 29(4 ), 215 -218 .] [本文引用: 1] [48] Wang JF Feng YL 2004 ). The effect of light intensity on biomass allocation leaf morphology and relative growth rate of two invasive plantsActa Phytoecologica Sinica, 781 -786 . [本文引用: 1] [ 王俊峰 , 冯玉龙 (2004 ). 光强对两种入侵植物生物量分配、叶片形态和相对生长速率的影响植物生态学报 , 28, 781 -786 .] [本文引用: 1] [49] Wang JF Shi YJ Ao YN Yu DF Wang J Gao S Knops JMH Mu CS Li ZJ 2019 ). Summer drought decreases Leymus chinensis productivity through constraining the bud, tiller and shoot productionJournal of Agronomy and Crop Scien ce, 554 -561 . [本文引用: 1] [50] Wang YH Wang CZ Li DF Zheng AR Qi SL Li GZ 2017 ). Effects of seeding rate on plant number, production performance, and quality of alfalfaActa Prataculturae Sinica, 2 ), 123 -135 . [本文引用: 2] [ 王彦华 , 王成章 , 李德锋 , 郑爱荣 , 齐胜利 , 李冠真 (2017 ). 播种量和品种对紫花苜蓿植株动态变化、产量及品质的影响草业学报 , 26(2 ), 123 -135 .] [本文引用: 2] [51] Wei YP Nan LL Yu C Fu SJ 2017 ). Effect of row spacing and planting density on the yield and quality ofMedicago sativa . Pratacultural Science, 1898 -1905 . [本文引用: 1] [ 魏永鹏 , 南丽丽 , 于闯 , 付双军 (2017 ). 种植密度和行距配置对紫花苜蓿群体产量及品质的影响草业科学 , 34, 1898 -1905 .] [本文引用: 1] [52] Weiner J Damgaard C 2006 ). Size-asymmetric competition and size-asymmetric growth in a spatially explicit zone-of-influence model of plant competitionEcological Research, 707 -712 . DOI:
10.1007/s11284-006-0178-6 URL [本文引用: 1] [53] Yoda K Kira T Ogawa H Hozumi K 1963 ). Self-thinning in over-crowded pure stands under cultivated and natural conditionsJournal of Biology of Osaka City University, 107 -129 . [本文引用: 2] [54] Zhang ZP Fang JY Kan M 2000 ). Effects of competition on growth rate and probability of death of plant individuals: a study based on nursery experiments of Larix leptolepis populationsActa Phytoecologica Sinica 340 -345 . [本文引用: 1] [ 张泽浦 , 方精云 , 菅诚 (2000 ). 邻体竞争对植物个体生长速率和死亡概率的影响: 基于日本落叶松种群试验的研究植物生态学报 , 24, 340 -345 .] [本文引用: 1] Effects of row spacing and population density on yield of sweet sorghum: applications for harvesting as billets
1
2015
... 种植密度能引起生长资源的强制分配, 密度的增加导致种内竞争的产生, 影响种群中植物的单株生长量和生物量(
Harper, 1977 ;
李博等, 2000 ).在很大程度上, 密度的制约作用使得种群内部个体的特征发生相应的变化.本研究中, 紫花苜蓿个体的性状随密度的变化发生了很显著的变化, 表现出极显著的密度制约特性: 随着种植密度的增加, 植物个体的株高、分枝数、基径、生物量均减小(
图3 ,
图4 ).在生长空间、光照、土壤水分和养分基本相同的情况下, 不同种植密度的同一植物种群基本具有相同的环境最大容纳量.当空间和资源有限时, 植物个体的生长可能会被抑制, 个体间产生竞争.当密度增加时, 改变自身的形态和结构是植物最易采取的、最经济的适应对策(
Silvertown & Lovett-Doust, 1993 ).尽管有研究表明, 植株高度不受密度的影响(
杜汉强等, 2004 ), 但株高随着种植密度的增加而减小也是一种“耐受”策略, 通过减缓自身的生长速度增加自身对外界胁迫的耐受力, 保证现有的尽可能多的植株的存活, 这与以往的研究结果(
陈静等, 2017 )一致.密度较小的种群在生长过程中能获得的光照、水分、养分以及空间方面的资源比较充足, 个体间的竞争较弱, 植物倾向于增加枝条或分蘖数量来提高个体生物量(
王俊峰和冯玉龙, 2004 ).而在种植密度增加, 资源紧缺的环境下, 植物倾向于减少枝条的数量和质量(
Soleymani et al ., 2011 ), 从而降低植物个体的生物量(
Wang et al ., 2019 ).另外, 关于基径与种群密度的关系,
Reineke (1933) 针对冠层密闭且同龄的植物种群提出了“立地密度法则”, 即
N =
kr -1.605 (
N 为种群密度,
r 为植物的平均直径,
k 为常数).也就是说, 随着种群密度的增加, 植物的平均单株直径会减小, 两者之间存在权衡关系(
Adams et al ., 2015 ). ...
The effect of ramet mortality on clonal plant growth
1
2019
... 苜蓿是以收获茎叶为主的牧草, 并且叶片中蛋白质的含量远高于茎, 因此, 叶茎比是衡量苜蓿品质的重要指标(
刘东霞等, 2015 ).苜蓿作为多年生植物, 根的基部产生根冠芽发育为分枝, 具有表型可塑性(
Benedek & Englert, 2019 ), 可通过改变植株的形态特征(如: 叶茎比、根冠比等)适应环境, 进一步改变植物生产力和其生物量分配.以往关于种群密度是否影响植物生物量分配的研究结果尚无定论.
范高华等(2017) 认为, 植物各器官生物量均受种群密度的显著影响, 并且茎和地上生物量分配均随种群密度的增大而减小.但在本研究中, 不同密度下苜蓿种群的根冠比、叶茎比均无显著差异, 各器官生物量分配比例不受种群密度影响.同一种植物不同密度水平下, 种群的根冠比主要受环境因素影响(
平晓燕等, 2007 ), 与密度无关(
方华和孔凡斌, 2003 ;
Deru et al ., 2016 ). ...
How does sowing rate affect plant and stem density, forage yield, and nutritive value in glyphosate-tolerant alfalfa?
1
2018
... 单位面积生物量是评价植物种群生产力的重要指标, 国内外在种植密度对苜蓿产量的影响方面有很多的研究, 但结果有所不同.有研究证明, 增加植株密度, 可以提高作物的产量(
Deng et al ., 2012 ), 有效地抑制杂草生长(
Bybee-Finley et al ., 2017 ;
Gibson et al ., 2017 ).
王彦华等(2017) 的研究则表明, 不同密度水平下苜蓿的产量没有差别.最近的研究结果发现, 当苜蓿的播种量低于10 kg·hm
-2 时, 播种量增加可以提高产量, 而当播种量高于10 kg·hm
-2 时, 产量并不随着播种量的增加而增加(
Berti & Samarappuli, 2018 ).在本实验中, 随着种植密度的增加, 单位面积干草产量间差异不显著, 种群地上与地下的最终产量都保持恒定, 符合最终产量恒定法则.这表明了在外部条件一致的情况下, 苜蓿种群经过充分的生长后, 群体达到了最大的资源利用率, 单位面积生物量保持恒定, 而与初始种植密度无关.此前有大量的以作物为研究对象的实验证明, 在一定密度范围内, 最终产量恒定的法则是适用的(
Kira et al ., 1953 ;
方精云等, 1991 ;
樊星等, 2016 ). ...
Crop biomass not species richness drives weed suppression in warm- season annual grass—Legume intercrops in the northeast
1
2017
... 单位面积生物量是评价植物种群生产力的重要指标, 国内外在种植密度对苜蓿产量的影响方面有很多的研究, 但结果有所不同.有研究证明, 增加植株密度, 可以提高作物的产量(
Deng et al ., 2012 ), 有效地抑制杂草生长(
Bybee-Finley et al ., 2017 ;
Gibson et al ., 2017 ).
王彦华等(2017) 的研究则表明, 不同密度水平下苜蓿的产量没有差别.最近的研究结果发现, 当苜蓿的播种量低于10 kg·hm
-2 时, 播种量增加可以提高产量, 而当播种量高于10 kg·hm
-2 时, 产量并不随着播种量的增加而增加(
Berti & Samarappuli, 2018 ).在本实验中, 随着种植密度的增加, 单位面积干草产量间差异不显著, 种群地上与地下的最终产量都保持恒定, 符合最终产量恒定法则.这表明了在外部条件一致的情况下, 苜蓿种群经过充分的生长后, 群体达到了最大的资源利用率, 单位面积生物量保持恒定, 而与初始种植密度无关.此前有大量的以作物为研究对象的实验证明, 在一定密度范围内, 最终产量恒定的法则是适用的(
Kira et al ., 1953 ;
方精云等, 1991 ;
樊星等, 2016 ). ...
Stochastic simulation of clonal growth in the tall goldenrod,
1
1991
... 植物的形态发育影响个体生物量(
Hecht et al ., 2019 ).构成苜蓿生物量的主要因素有单茎质量、茎长、基径、侧枝数和叶片数(
孟凯等, 2019 ).本实验中, 不同密度下, 植物个体生物量与植物个体的平均株高、基径、分枝数量联系密切.随着密度的降低, 植物个体株高、基径和分枝数量增加, 植物个体生物量逐渐增长.这是因为苜蓿可在根的基部产生根冠芽并成长为分枝, 在外界环境竞争压力较小的情况下, 单株植物可以增加分枝的数量形成一个小的“种群” (
Cain et al ., 1991 ), 进而提高单株个体的生产力(
Kays & Harper, 1974 ).植物生长过程中会争夺有限的资源, 当种内竞争日趋激烈时, 植物个体之间彼此干扰, 许多个体陆续死亡, 种群密度降低(
方精云, 1992 ).在自疏过程中植物的平均个体质量与种群密度间的关系通常符合-3/2 (
Yoda et al ., 1963 )或-4/3 (
Enquist et al ., 1998 )自疏法则.本研究中, 个体平均生物量与密度关系为-1.0, 符合最终产量恒定法则(
Kira et al ., 1953 ).一方面, 自疏法则中存活个体数量与生物量的关系是在种群密度由大变小的过程中得到的; 另一方面, 苜蓿根基部可产生根冠芽, 发育成枝条后会有水平方向的伸展, 出现匍匐生长的情况.该情况下, 植株几何形状发生变化, 会出现平均个体生物量-密度关系的斜率接近-1的情况(
王刚, 1993 ), 即形成最终产量恒定的状态. ...
不同密度旱柳的树冠构型与光截获
1
2017
... 种植密度能引起生长资源的强制分配, 密度的增加导致种内竞争的产生, 影响种群中植物的单株生长量和生物量(
Harper, 1977 ;
李博等, 2000 ).在很大程度上, 密度的制约作用使得种群内部个体的特征发生相应的变化.本研究中, 紫花苜蓿个体的性状随密度的变化发生了很显著的变化, 表现出极显著的密度制约特性: 随着种植密度的增加, 植物个体的株高、分枝数、基径、生物量均减小(
图3 ,
图4 ).在生长空间、光照、土壤水分和养分基本相同的情况下, 不同种植密度的同一植物种群基本具有相同的环境最大容纳量.当空间和资源有限时, 植物个体的生长可能会被抑制, 个体间产生竞争.当密度增加时, 改变自身的形态和结构是植物最易采取的、最经济的适应对策(
Silvertown & Lovett-Doust, 1993 ).尽管有研究表明, 植株高度不受密度的影响(
杜汉强等, 2004 ), 但株高随着种植密度的增加而减小也是一种“耐受”策略, 通过减缓自身的生长速度增加自身对外界胁迫的耐受力, 保证现有的尽可能多的植株的存活, 这与以往的研究结果(
陈静等, 2017 )一致.密度较小的种群在生长过程中能获得的光照、水分、养分以及空间方面的资源比较充足, 个体间的竞争较弱, 植物倾向于增加枝条或分蘖数量来提高个体生物量(
王俊峰和冯玉龙, 2004 ).而在种植密度增加, 资源紧缺的环境下, 植物倾向于减少枝条的数量和质量(
Soleymani et al ., 2011 ), 从而降低植物个体的生物量(
Wang et al ., 2019 ).另外, 关于基径与种群密度的关系,
Reineke (1933) 针对冠层密闭且同龄的植物种群提出了“立地密度法则”, 即
N =
kr -1.605 (
N 为种群密度,
r 为植物的平均直径,
k 为常数).也就是说, 随着种群密度的增加, 植物的平均单株直径会减小, 两者之间存在权衡关系(
Adams et al ., 2015 ). ...
不同密度旱柳的树冠构型与光截获
1
2017
... 种植密度能引起生长资源的强制分配, 密度的增加导致种内竞争的产生, 影响种群中植物的单株生长量和生物量(
Harper, 1977 ;
李博等, 2000 ).在很大程度上, 密度的制约作用使得种群内部个体的特征发生相应的变化.本研究中, 紫花苜蓿个体的性状随密度的变化发生了很显著的变化, 表现出极显著的密度制约特性: 随着种植密度的增加, 植物个体的株高、分枝数、基径、生物量均减小(
图3 ,
图4 ).在生长空间、光照、土壤水分和养分基本相同的情况下, 不同种植密度的同一植物种群基本具有相同的环境最大容纳量.当空间和资源有限时, 植物个体的生长可能会被抑制, 个体间产生竞争.当密度增加时, 改变自身的形态和结构是植物最易采取的、最经济的适应对策(
Silvertown & Lovett-Doust, 1993 ).尽管有研究表明, 植株高度不受密度的影响(
杜汉强等, 2004 ), 但株高随着种植密度的增加而减小也是一种“耐受”策略, 通过减缓自身的生长速度增加自身对外界胁迫的耐受力, 保证现有的尽可能多的植株的存活, 这与以往的研究结果(
陈静等, 2017 )一致.密度较小的种群在生长过程中能获得的光照、水分、养分以及空间方面的资源比较充足, 个体间的竞争较弱, 植物倾向于增加枝条或分蘖数量来提高个体生物量(
王俊峰和冯玉龙, 2004 ).而在种植密度增加, 资源紧缺的环境下, 植物倾向于减少枝条的数量和质量(
Soleymani et al ., 2011 ), 从而降低植物个体的生物量(
Wang et al ., 2019 ).另外, 关于基径与种群密度的关系,
Reineke (1933) 针对冠层密闭且同龄的植物种群提出了“立地密度法则”, 即
N =
kr -1.605 (
N 为种群密度,
r 为植物的平均直径,
k 为常数).也就是说, 随着种群密度的增加, 植物的平均单株直径会减小, 两者之间存在权衡关系(
Adams et al ., 2015 ). ...
黄土丘陵区紫花苜蓿生长与土壤水分变化
1
2005
... 除了优质苜蓿的种植面积偏少, 我国在苜蓿的种植方式、栽培管理等方面也比较落后, 使得苜蓿产量和质量一直处于较低的水平.有些多年生紫花苜蓿(
Medicago sativa )人工草地不仅生产力低, 而且造成了土壤水分的严重亏缺(
程积民等, 2005 ), 不合理的利用引起了一系列的环境问题.苜蓿作物产量和营养水平受苜蓿品种、施肥量、播种时间、种植株距和行距、刈割高度、刈割时间和次数等多种因素的影响(
Chocarro & Lloveras, 2015 ;
都帅等, 2016 ;
孙万斌等, 2017 ;
魏永鹏等, 2017 ;
Jáuregui et al ., 2019 ).其中, 种植密度作为影响作物产量和品质的重要因素, 会造成植物个体对于光照、水分、养分的竞争, 这种竞争会增加个体大小之间的差异(
Weiner & Damgaard, 2006 ;
Chu et al ., 2009 ).这种差异在个体和群体的性状上都有所体现. ...
黄土丘陵区紫花苜蓿生长与土壤水分变化
1
2005
... 除了优质苜蓿的种植面积偏少, 我国在苜蓿的种植方式、栽培管理等方面也比较落后, 使得苜蓿产量和质量一直处于较低的水平.有些多年生紫花苜蓿(
Medicago sativa )人工草地不仅生产力低, 而且造成了土壤水分的严重亏缺(
程积民等, 2005 ), 不合理的利用引起了一系列的环境问题.苜蓿作物产量和营养水平受苜蓿品种、施肥量、播种时间、种植株距和行距、刈割高度、刈割时间和次数等多种因素的影响(
Chocarro & Lloveras, 2015 ;
都帅等, 2016 ;
孙万斌等, 2017 ;
魏永鹏等, 2017 ;
Jáuregui et al ., 2019 ).其中, 种植密度作为影响作物产量和品质的重要因素, 会造成植物个体对于光照、水分、养分的竞争, 这种竞争会增加个体大小之间的差异(
Weiner & Damgaard, 2006 ;
Chu et al ., 2009 ).这种差异在个体和群体的性状上都有所体现. ...
The effect of row spacing on alfalfa seed and forage production under irrigated Mediterranean agricultural conditions
1
2015
... 除了优质苜蓿的种植面积偏少, 我国在苜蓿的种植方式、栽培管理等方面也比较落后, 使得苜蓿产量和质量一直处于较低的水平.有些多年生紫花苜蓿(
Medicago sativa )人工草地不仅生产力低, 而且造成了土壤水分的严重亏缺(
程积民等, 2005 ), 不合理的利用引起了一系列的环境问题.苜蓿作物产量和营养水平受苜蓿品种、施肥量、播种时间、种植株距和行距、刈割高度、刈割时间和次数等多种因素的影响(
Chocarro & Lloveras, 2015 ;
都帅等, 2016 ;
孙万斌等, 2017 ;
魏永鹏等, 2017 ;
Jáuregui et al ., 2019 ).其中, 种植密度作为影响作物产量和品质的重要因素, 会造成植物个体对于光照、水分、养分的竞争, 这种竞争会增加个体大小之间的差异(
Weiner & Damgaard, 2006 ;
Chu et al ., 2009 ).这种差异在个体和群体的性状上都有所体现. ...
Positive interactions can increase size inequality in plant populations
1
2009
... 除了优质苜蓿的种植面积偏少, 我国在苜蓿的种植方式、栽培管理等方面也比较落后, 使得苜蓿产量和质量一直处于较低的水平.有些多年生紫花苜蓿(
Medicago sativa )人工草地不仅生产力低, 而且造成了土壤水分的严重亏缺(
程积民等, 2005 ), 不合理的利用引起了一系列的环境问题.苜蓿作物产量和营养水平受苜蓿品种、施肥量、播种时间、种植株距和行距、刈割高度、刈割时间和次数等多种因素的影响(
Chocarro & Lloveras, 2015 ;
都帅等, 2016 ;
孙万斌等, 2017 ;
魏永鹏等, 2017 ;
Jáuregui et al ., 2019 ).其中, 种植密度作为影响作物产量和品质的重要因素, 会造成植物个体对于光照、水分、养分的竞争, 这种竞争会增加个体大小之间的差异(
Weiner & Damgaard, 2006 ;
Chu et al ., 2009 ).这种差异在个体和群体的性状上都有所体现. ...
Models and tests of optimal density and maximal yield for crop plants
2
2012
... 植物种群的密度效应是种群生态学研究的一个核心问题.植物种群在资源有限的情况下, 保证每个个体的存活是困难的, 种群数量会减少, 即产生种群调节作用.在植物种群密度与生物量的关系方面, 涉及很多经典的科学问题, 前人也提出了众多经典的理论, 如最终产量恒定法则、自疏法则以及最适种植密度法则等(
Shinozaki & Kira, 1956 ;
Yoda et al ., 1963 ;
Harper, 1977 ;
Deng et al ., 2012 ).早期研究显示, 适当提高密度可以增加苜蓿的建植率和覆盖率, 是保证其高产的有效措施(
欧阳延生等, 2007 ).虽然在过去的几十年里, 有关种植密度对苜蓿影响的研究已有不少, 但多偏重于探究播种量对苜蓿产量以及品质的影响(
李丽等, 2012 ;
马克成和王秉龙, 2014 ); 从个体和群体两个方面研究种植密度影响苜蓿生长的报道较少. ...
... 单位面积生物量是评价植物种群生产力的重要指标, 国内外在种植密度对苜蓿产量的影响方面有很多的研究, 但结果有所不同.有研究证明, 增加植株密度, 可以提高作物的产量(
Deng et al ., 2012 ), 有效地抑制杂草生长(
Bybee-Finley et al ., 2017 ;
Gibson et al ., 2017 ).
王彦华等(2017) 的研究则表明, 不同密度水平下苜蓿的产量没有差别.最近的研究结果发现, 当苜蓿的播种量低于10 kg·hm
-2 时, 播种量增加可以提高产量, 而当播种量高于10 kg·hm
-2 时, 产量并不随着播种量的增加而增加(
Berti & Samarappuli, 2018 ).在本实验中, 随着种植密度的增加, 单位面积干草产量间差异不显著, 种群地上与地下的最终产量都保持恒定, 符合最终产量恒定法则.这表明了在外部条件一致的情况下, 苜蓿种群经过充分的生长后, 群体达到了最大的资源利用率, 单位面积生物量保持恒定, 而与初始种植密度无关.此前有大量的以作物为研究对象的实验证明, 在一定密度范围内, 最终产量恒定的法则是适用的(
Kira et al ., 1953 ;
方精云等, 1991 ;
樊星等, 2016 ). ...
1
2016
... 苜蓿是以收获茎叶为主的牧草, 并且叶片中蛋白质的含量远高于茎, 因此, 叶茎比是衡量苜蓿品质的重要指标(
刘东霞等, 2015 ).苜蓿作为多年生植物, 根的基部产生根冠芽发育为分枝, 具有表型可塑性(
Benedek & Englert, 2019 ), 可通过改变植株的形态特征(如: 叶茎比、根冠比等)适应环境, 进一步改变植物生产力和其生物量分配.以往关于种群密度是否影响植物生物量分配的研究结果尚无定论.
范高华等(2017) 认为, 植物各器官生物量均受种群密度的显著影响, 并且茎和地上生物量分配均随种群密度的增大而减小.但在本研究中, 不同密度下苜蓿种群的根冠比、叶茎比均无显著差异, 各器官生物量分配比例不受种群密度影响.同一种植物不同密度水平下, 种群的根冠比主要受环境因素影响(
平晓燕等, 2007 ), 与密度无关(
方华和孔凡斌, 2003 ;
Deru et al ., 2016 ). ...
不同播种密度对紫花苜蓿主要性状的影响
1
2004
... 种植密度能引起生长资源的强制分配, 密度的增加导致种内竞争的产生, 影响种群中植物的单株生长量和生物量(
Harper, 1977 ;
李博等, 2000 ).在很大程度上, 密度的制约作用使得种群内部个体的特征发生相应的变化.本研究中, 紫花苜蓿个体的性状随密度的变化发生了很显著的变化, 表现出极显著的密度制约特性: 随着种植密度的增加, 植物个体的株高、分枝数、基径、生物量均减小(
图3 ,
图4 ).在生长空间、光照、土壤水分和养分基本相同的情况下, 不同种植密度的同一植物种群基本具有相同的环境最大容纳量.当空间和资源有限时, 植物个体的生长可能会被抑制, 个体间产生竞争.当密度增加时, 改变自身的形态和结构是植物最易采取的、最经济的适应对策(
Silvertown & Lovett-Doust, 1993 ).尽管有研究表明, 植株高度不受密度的影响(
杜汉强等, 2004 ), 但株高随着种植密度的增加而减小也是一种“耐受”策略, 通过减缓自身的生长速度增加自身对外界胁迫的耐受力, 保证现有的尽可能多的植株的存活, 这与以往的研究结果(
陈静等, 2017 )一致.密度较小的种群在生长过程中能获得的光照、水分、养分以及空间方面的资源比较充足, 个体间的竞争较弱, 植物倾向于增加枝条或分蘖数量来提高个体生物量(
王俊峰和冯玉龙, 2004 ).而在种植密度增加, 资源紧缺的环境下, 植物倾向于减少枝条的数量和质量(
Soleymani et al ., 2011 ), 从而降低植物个体的生物量(
Wang et al ., 2019 ).另外, 关于基径与种群密度的关系,
Reineke (1933) 针对冠层密闭且同龄的植物种群提出了“立地密度法则”, 即
N =
kr -1.605 (
N 为种群密度,
r 为植物的平均直径,
k 为常数).也就是说, 随着种群密度的增加, 植物的平均单株直径会减小, 两者之间存在权衡关系(
Adams et al ., 2015 ). ...
不同播种密度对紫花苜蓿主要性状的影响
1
2004
... 种植密度能引起生长资源的强制分配, 密度的增加导致种内竞争的产生, 影响种群中植物的单株生长量和生物量(
Harper, 1977 ;
李博等, 2000 ).在很大程度上, 密度的制约作用使得种群内部个体的特征发生相应的变化.本研究中, 紫花苜蓿个体的性状随密度的变化发生了很显著的变化, 表现出极显著的密度制约特性: 随着种植密度的增加, 植物个体的株高、分枝数、基径、生物量均减小(
图3 ,
图4 ).在生长空间、光照、土壤水分和养分基本相同的情况下, 不同种植密度的同一植物种群基本具有相同的环境最大容纳量.当空间和资源有限时, 植物个体的生长可能会被抑制, 个体间产生竞争.当密度增加时, 改变自身的形态和结构是植物最易采取的、最经济的适应对策(
Silvertown & Lovett-Doust, 1993 ).尽管有研究表明, 植株高度不受密度的影响(
杜汉强等, 2004 ), 但株高随着种植密度的增加而减小也是一种“耐受”策略, 通过减缓自身的生长速度增加自身对外界胁迫的耐受力, 保证现有的尽可能多的植株的存活, 这与以往的研究结果(
陈静等, 2017 )一致.密度较小的种群在生长过程中能获得的光照、水分、养分以及空间方面的资源比较充足, 个体间的竞争较弱, 植物倾向于增加枝条或分蘖数量来提高个体生物量(
王俊峰和冯玉龙, 2004 ).而在种植密度增加, 资源紧缺的环境下, 植物倾向于减少枝条的数量和质量(
Soleymani et al ., 2011 ), 从而降低植物个体的生物量(
Wang et al ., 2019 ).另外, 关于基径与种群密度的关系,
Reineke (1933) 针对冠层密闭且同龄的植物种群提出了“立地密度法则”, 即
N =
kr -1.605 (
N 为种群密度,
r 为植物的平均直径,
k 为常数).也就是说, 随着种群密度的增加, 植物的平均单株直径会减小, 两者之间存在权衡关系(
Adams et al ., 2015 ). ...
不同刈割时期与刈割高度对苜蓿品质的影响
1
2016
... 除了优质苜蓿的种植面积偏少, 我国在苜蓿的种植方式、栽培管理等方面也比较落后, 使得苜蓿产量和质量一直处于较低的水平.有些多年生紫花苜蓿(
Medicago sativa )人工草地不仅生产力低, 而且造成了土壤水分的严重亏缺(
程积民等, 2005 ), 不合理的利用引起了一系列的环境问题.苜蓿作物产量和营养水平受苜蓿品种、施肥量、播种时间、种植株距和行距、刈割高度、刈割时间和次数等多种因素的影响(
Chocarro & Lloveras, 2015 ;
都帅等, 2016 ;
孙万斌等, 2017 ;
魏永鹏等, 2017 ;
Jáuregui et al ., 2019 ).其中, 种植密度作为影响作物产量和品质的重要因素, 会造成植物个体对于光照、水分、养分的竞争, 这种竞争会增加个体大小之间的差异(
Weiner & Damgaard, 2006 ;
Chu et al ., 2009 ).这种差异在个体和群体的性状上都有所体现. ...
不同刈割时期与刈割高度对苜蓿品质的影响
1
2016
... 除了优质苜蓿的种植面积偏少, 我国在苜蓿的种植方式、栽培管理等方面也比较落后, 使得苜蓿产量和质量一直处于较低的水平.有些多年生紫花苜蓿(
Medicago sativa )人工草地不仅生产力低, 而且造成了土壤水分的严重亏缺(
程积民等, 2005 ), 不合理的利用引起了一系列的环境问题.苜蓿作物产量和营养水平受苜蓿品种、施肥量、播种时间、种植株距和行距、刈割高度、刈割时间和次数等多种因素的影响(
Chocarro & Lloveras, 2015 ;
都帅等, 2016 ;
孙万斌等, 2017 ;
魏永鹏等, 2017 ;
Jáuregui et al ., 2019 ).其中, 种植密度作为影响作物产量和品质的重要因素, 会造成植物个体对于光照、水分、养分的竞争, 这种竞争会增加个体大小之间的差异(
Weiner & Damgaard, 2006 ;
Chu et al ., 2009 ).这种差异在个体和群体的性状上都有所体现. ...
Allometric scaling of plant energetics and population density
1
1998
... 植物的形态发育影响个体生物量(
Hecht et al ., 2019 ).构成苜蓿生物量的主要因素有单茎质量、茎长、基径、侧枝数和叶片数(
孟凯等, 2019 ).本实验中, 不同密度下, 植物个体生物量与植物个体的平均株高、基径、分枝数量联系密切.随着密度的降低, 植物个体株高、基径和分枝数量增加, 植物个体生物量逐渐增长.这是因为苜蓿可在根的基部产生根冠芽并成长为分枝, 在外界环境竞争压力较小的情况下, 单株植物可以增加分枝的数量形成一个小的“种群” (
Cain et al ., 1991 ), 进而提高单株个体的生产力(
Kays & Harper, 1974 ).植物生长过程中会争夺有限的资源, 当种内竞争日趋激烈时, 植物个体之间彼此干扰, 许多个体陆续死亡, 种群密度降低(
方精云, 1992 ).在自疏过程中植物的平均个体质量与种群密度间的关系通常符合-3/2 (
Yoda et al ., 1963 )或-4/3 (
Enquist et al ., 1998 )自疏法则.本研究中, 个体平均生物量与密度关系为-1.0, 符合最终产量恒定法则(
Kira et al ., 1953 ).一方面, 自疏法则中存活个体数量与生物量的关系是在种群密度由大变小的过程中得到的; 另一方面, 苜蓿根基部可产生根冠芽, 发育成枝条后会有水平方向的伸展, 出现匍匐生长的情况.该情况下, 植株几何形状发生变化, 会出现平均个体生物量-密度关系的斜率接近-1的情况(
王刚, 1993 ), 即形成最终产量恒定的状态. ...
种群密度对沙米异速生长的影响
1
2017
... 苜蓿是以收获茎叶为主的牧草, 并且叶片中蛋白质的含量远高于茎, 因此, 叶茎比是衡量苜蓿品质的重要指标(
刘东霞等, 2015 ).苜蓿作为多年生植物, 根的基部产生根冠芽发育为分枝, 具有表型可塑性(
Benedek & Englert, 2019 ), 可通过改变植株的形态特征(如: 叶茎比、根冠比等)适应环境, 进一步改变植物生产力和其生物量分配.以往关于种群密度是否影响植物生物量分配的研究结果尚无定论.
范高华等(2017) 认为, 植物各器官生物量均受种群密度的显著影响, 并且茎和地上生物量分配均随种群密度的增大而减小.但在本研究中, 不同密度下苜蓿种群的根冠比、叶茎比均无显著差异, 各器官生物量分配比例不受种群密度影响.同一种植物不同密度水平下, 种群的根冠比主要受环境因素影响(
平晓燕等, 2007 ), 与密度无关(
方华和孔凡斌, 2003 ;
Deru et al ., 2016 ). ...
种群密度对沙米异速生长的影响
1
2017
... 苜蓿是以收获茎叶为主的牧草, 并且叶片中蛋白质的含量远高于茎, 因此, 叶茎比是衡量苜蓿品质的重要指标(
刘东霞等, 2015 ).苜蓿作为多年生植物, 根的基部产生根冠芽发育为分枝, 具有表型可塑性(
Benedek & Englert, 2019 ), 可通过改变植株的形态特征(如: 叶茎比、根冠比等)适应环境, 进一步改变植物生产力和其生物量分配.以往关于种群密度是否影响植物生物量分配的研究结果尚无定论.
范高华等(2017) 认为, 植物各器官生物量均受种群密度的显著影响, 并且茎和地上生物量分配均随种群密度的增大而减小.但在本研究中, 不同密度下苜蓿种群的根冠比、叶茎比均无显著差异, 各器官生物量分配比例不受种群密度影响.同一种植物不同密度水平下, 种群的根冠比主要受环境因素影响(
平晓燕等, 2007 ), 与密度无关(
方华和孔凡斌, 2003 ;
Deru et al ., 2016 ). ...
葎草种群自疏过程中幼苗构件性状及生物量分配变化
1
2016
... 单位面积生物量是评价植物种群生产力的重要指标, 国内外在种植密度对苜蓿产量的影响方面有很多的研究, 但结果有所不同.有研究证明, 增加植株密度, 可以提高作物的产量(
Deng et al ., 2012 ), 有效地抑制杂草生长(
Bybee-Finley et al ., 2017 ;
Gibson et al ., 2017 ).
王彦华等(2017) 的研究则表明, 不同密度水平下苜蓿的产量没有差别.最近的研究结果发现, 当苜蓿的播种量低于10 kg·hm
-2 时, 播种量增加可以提高产量, 而当播种量高于10 kg·hm
-2 时, 产量并不随着播种量的增加而增加(
Berti & Samarappuli, 2018 ).在本实验中, 随着种植密度的增加, 单位面积干草产量间差异不显著, 种群地上与地下的最终产量都保持恒定, 符合最终产量恒定法则.这表明了在外部条件一致的情况下, 苜蓿种群经过充分的生长后, 群体达到了最大的资源利用率, 单位面积生物量保持恒定, 而与初始种植密度无关.此前有大量的以作物为研究对象的实验证明, 在一定密度范围内, 最终产量恒定的法则是适用的(
Kira et al ., 1953 ;
方精云等, 1991 ;
樊星等, 2016 ). ...
葎草种群自疏过程中幼苗构件性状及生物量分配变化
1
2016
... 单位面积生物量是评价植物种群生产力的重要指标, 国内外在种植密度对苜蓿产量的影响方面有很多的研究, 但结果有所不同.有研究证明, 增加植株密度, 可以提高作物的产量(
Deng et al ., 2012 ), 有效地抑制杂草生长(
Bybee-Finley et al ., 2017 ;
Gibson et al ., 2017 ).
王彦华等(2017) 的研究则表明, 不同密度水平下苜蓿的产量没有差别.最近的研究结果发现, 当苜蓿的播种量低于10 kg·hm
-2 时, 播种量增加可以提高产量, 而当播种量高于10 kg·hm
-2 时, 产量并不随着播种量的增加而增加(
Berti & Samarappuli, 2018 ).在本实验中, 随着种植密度的增加, 单位面积干草产量间差异不显著, 种群地上与地下的最终产量都保持恒定, 符合最终产量恒定法则.这表明了在外部条件一致的情况下, 苜蓿种群经过充分的生长后, 群体达到了最大的资源利用率, 单位面积生物量保持恒定, 而与初始种植密度无关.此前有大量的以作物为研究对象的实验证明, 在一定密度范围内, 最终产量恒定的法则是适用的(
Kira et al ., 1953 ;
方精云等, 1991 ;
樊星等, 2016 ). ...
不同密度火炬松林生物量及其分配
1
2003
... 苜蓿是以收获茎叶为主的牧草, 并且叶片中蛋白质的含量远高于茎, 因此, 叶茎比是衡量苜蓿品质的重要指标(
刘东霞等, 2015 ).苜蓿作为多年生植物, 根的基部产生根冠芽发育为分枝, 具有表型可塑性(
Benedek & Englert, 2019 ), 可通过改变植株的形态特征(如: 叶茎比、根冠比等)适应环境, 进一步改变植物生产力和其生物量分配.以往关于种群密度是否影响植物生物量分配的研究结果尚无定论.
范高华等(2017) 认为, 植物各器官生物量均受种群密度的显著影响, 并且茎和地上生物量分配均随种群密度的增大而减小.但在本研究中, 不同密度下苜蓿种群的根冠比、叶茎比均无显著差异, 各器官生物量分配比例不受种群密度影响.同一种植物不同密度水平下, 种群的根冠比主要受环境因素影响(
平晓燕等, 2007 ), 与密度无关(
方华和孔凡斌, 2003 ;
Deru et al ., 2016 ). ...
不同密度火炬松林生物量及其分配
1
2003
... 苜蓿是以收获茎叶为主的牧草, 并且叶片中蛋白质的含量远高于茎, 因此, 叶茎比是衡量苜蓿品质的重要指标(
刘东霞等, 2015 ).苜蓿作为多年生植物, 根的基部产生根冠芽发育为分枝, 具有表型可塑性(
Benedek & Englert, 2019 ), 可通过改变植株的形态特征(如: 叶茎比、根冠比等)适应环境, 进一步改变植物生产力和其生物量分配.以往关于种群密度是否影响植物生物量分配的研究结果尚无定论.
范高华等(2017) 认为, 植物各器官生物量均受种群密度的显著影响, 并且茎和地上生物量分配均随种群密度的增大而减小.但在本研究中, 不同密度下苜蓿种群的根冠比、叶茎比均无显著差异, 各器官生物量分配比例不受种群密度影响.同一种植物不同密度水平下, 种群的根冠比主要受环境因素影响(
平晓燕等, 2007 ), 与密度无关(
方华和孔凡斌, 2003 ;
Deru et al ., 2016 ). ...
1
1991
... 密度是自然界的重要选择压力(
Japhet et al ., 2009 ).在密度的作用下, 植物种群内的植株互相竞争可以获得的资源, 主要包括光照、水分、矿物质等(
黎磊等, 2011 ).
Hakl等(2011) 对高密度下的苜 蓿种群数量动态进行了观测, 7年间植株数量由 5 000株·m
-2 降至57株·m
-2 .有研究表明, 苜蓿植株数量在第一生长年急剧下降, 第二生长年缓慢下降, 第三生长年趋于平稳(
王彦华等, 2017 ).本实验的研究结果显示, 初始密度对紫花苜蓿种群数量的动态变化有显著的影响, 前期随着生长时间的增长, 较高密度(1 255、1 938株·m
-2 )水平下的植株数量迅速下降, 随后缓慢下降; 中等密度(373、475株·m
-2 )水平下的植株数量一直较缓慢下降, 而较低密度(25、100株·m
-2 )水平下的植株数量则基本不变.随着生长时间的延长, 初始密度对于植物种群数量的影响有减弱的趋势.产生这种动态变化的原因可能是: 较高密度下当植物种群生长到一定阶段, 种内竞争日趋激烈(
方精云, 1992 ), 植物个体之间彼此相互干扰, 许多个体陆续死亡(
张泽浦等, 2000 ); 而密度处于较低水平时, 植株个体的生存压力较小, 在外界条件没有较大波动的情况下, 种群数量维持着较为平稳的状态, 死亡率为0 (
方精云, 1991 ;
朴顺姬等, 1997 ). ...
1
1991
... 密度是自然界的重要选择压力(
Japhet et al ., 2009 ).在密度的作用下, 植物种群内的植株互相竞争可以获得的资源, 主要包括光照、水分、矿物质等(
黎磊等, 2011 ).
Hakl等(2011) 对高密度下的苜 蓿种群数量动态进行了观测, 7年间植株数量由 5 000株·m
-2 降至57株·m
-2 .有研究表明, 苜蓿植株数量在第一生长年急剧下降, 第二生长年缓慢下降, 第三生长年趋于平稳(
王彦华等, 2017 ).本实验的研究结果显示, 初始密度对紫花苜蓿种群数量的动态变化有显著的影响, 前期随着生长时间的增长, 较高密度(1 255、1 938株·m
-2 )水平下的植株数量迅速下降, 随后缓慢下降; 中等密度(373、475株·m
-2 )水平下的植株数量一直较缓慢下降, 而较低密度(25、100株·m
-2 )水平下的植株数量则基本不变.随着生长时间的延长, 初始密度对于植物种群数量的影响有减弱的趋势.产生这种动态变化的原因可能是: 较高密度下当植物种群生长到一定阶段, 种内竞争日趋激烈(
方精云, 1992 ), 植物个体之间彼此相互干扰, 许多个体陆续死亡(
张泽浦等, 2000 ); 而密度处于较低水平时, 植株个体的生存压力较小, 在外界条件没有较大波动的情况下, 种群数量维持着较为平稳的状态, 死亡率为0 (
方精云, 1991 ;
朴顺姬等, 1997 ). ...
植物种群的自然稀疏法则
2
1992
... 植物的形态发育影响个体生物量(
Hecht et al ., 2019 ).构成苜蓿生物量的主要因素有单茎质量、茎长、基径、侧枝数和叶片数(
孟凯等, 2019 ).本实验中, 不同密度下, 植物个体生物量与植物个体的平均株高、基径、分枝数量联系密切.随着密度的降低, 植物个体株高、基径和分枝数量增加, 植物个体生物量逐渐增长.这是因为苜蓿可在根的基部产生根冠芽并成长为分枝, 在外界环境竞争压力较小的情况下, 单株植物可以增加分枝的数量形成一个小的“种群” (
Cain et al ., 1991 ), 进而提高单株个体的生产力(
Kays & Harper, 1974 ).植物生长过程中会争夺有限的资源, 当种内竞争日趋激烈时, 植物个体之间彼此干扰, 许多个体陆续死亡, 种群密度降低(
方精云, 1992 ).在自疏过程中植物的平均个体质量与种群密度间的关系通常符合-3/2 (
Yoda et al ., 1963 )或-4/3 (
Enquist et al ., 1998 )自疏法则.本研究中, 个体平均生物量与密度关系为-1.0, 符合最终产量恒定法则(
Kira et al ., 1953 ).一方面, 自疏法则中存活个体数量与生物量的关系是在种群密度由大变小的过程中得到的; 另一方面, 苜蓿根基部可产生根冠芽, 发育成枝条后会有水平方向的伸展, 出现匍匐生长的情况.该情况下, 植株几何形状发生变化, 会出现平均个体生物量-密度关系的斜率接近-1的情况(
王刚, 1993 ), 即形成最终产量恒定的状态. ...
... 密度是自然界的重要选择压力(
Japhet et al ., 2009 ).在密度的作用下, 植物种群内的植株互相竞争可以获得的资源, 主要包括光照、水分、矿物质等(
黎磊等, 2011 ).
Hakl等(2011) 对高密度下的苜 蓿种群数量动态进行了观测, 7年间植株数量由 5 000株·m
-2 降至57株·m
-2 .有研究表明, 苜蓿植株数量在第一生长年急剧下降, 第二生长年缓慢下降, 第三生长年趋于平稳(
王彦华等, 2017 ).本实验的研究结果显示, 初始密度对紫花苜蓿种群数量的动态变化有显著的影响, 前期随着生长时间的增长, 较高密度(1 255、1 938株·m
-2 )水平下的植株数量迅速下降, 随后缓慢下降; 中等密度(373、475株·m
-2 )水平下的植株数量一直较缓慢下降, 而较低密度(25、100株·m
-2 )水平下的植株数量则基本不变.随着生长时间的延长, 初始密度对于植物种群数量的影响有减弱的趋势.产生这种动态变化的原因可能是: 较高密度下当植物种群生长到一定阶段, 种内竞争日趋激烈(
方精云, 1992 ), 植物个体之间彼此相互干扰, 许多个体陆续死亡(
张泽浦等, 2000 ); 而密度处于较低水平时, 植株个体的生存压力较小, 在外界条件没有较大波动的情况下, 种群数量维持着较为平稳的状态, 死亡率为0 (
方精云, 1991 ;
朴顺姬等, 1997 ). ...
植物种群的自然稀疏法则
2
1992
... 植物的形态发育影响个体生物量(
Hecht et al ., 2019 ).构成苜蓿生物量的主要因素有单茎质量、茎长、基径、侧枝数和叶片数(
孟凯等, 2019 ).本实验中, 不同密度下, 植物个体生物量与植物个体的平均株高、基径、分枝数量联系密切.随着密度的降低, 植物个体株高、基径和分枝数量增加, 植物个体生物量逐渐增长.这是因为苜蓿可在根的基部产生根冠芽并成长为分枝, 在外界环境竞争压力较小的情况下, 单株植物可以增加分枝的数量形成一个小的“种群” (
Cain et al ., 1991 ), 进而提高单株个体的生产力(
Kays & Harper, 1974 ).植物生长过程中会争夺有限的资源, 当种内竞争日趋激烈时, 植物个体之间彼此干扰, 许多个体陆续死亡, 种群密度降低(
方精云, 1992 ).在自疏过程中植物的平均个体质量与种群密度间的关系通常符合-3/2 (
Yoda et al ., 1963 )或-4/3 (
Enquist et al ., 1998 )自疏法则.本研究中, 个体平均生物量与密度关系为-1.0, 符合最终产量恒定法则(
Kira et al ., 1953 ).一方面, 自疏法则中存活个体数量与生物量的关系是在种群密度由大变小的过程中得到的; 另一方面, 苜蓿根基部可产生根冠芽, 发育成枝条后会有水平方向的伸展, 出现匍匐生长的情况.该情况下, 植株几何形状发生变化, 会出现平均个体生物量-密度关系的斜率接近-1的情况(
王刚, 1993 ), 即形成最终产量恒定的状态. ...
... 密度是自然界的重要选择压力(
Japhet et al ., 2009 ).在密度的作用下, 植物种群内的植株互相竞争可以获得的资源, 主要包括光照、水分、矿物质等(
黎磊等, 2011 ).
Hakl等(2011) 对高密度下的苜 蓿种群数量动态进行了观测, 7年间植株数量由 5 000株·m
-2 降至57株·m
-2 .有研究表明, 苜蓿植株数量在第一生长年急剧下降, 第二生长年缓慢下降, 第三生长年趋于平稳(
王彦华等, 2017 ).本实验的研究结果显示, 初始密度对紫花苜蓿种群数量的动态变化有显著的影响, 前期随着生长时间的增长, 较高密度(1 255、1 938株·m
-2 )水平下的植株数量迅速下降, 随后缓慢下降; 中等密度(373、475株·m
-2 )水平下的植株数量一直较缓慢下降, 而较低密度(25、100株·m
-2 )水平下的植株数量则基本不变.随着生长时间的延长, 初始密度对于植物种群数量的影响有减弱的趋势.产生这种动态变化的原因可能是: 较高密度下当植物种群生长到一定阶段, 种内竞争日趋激烈(
方精云, 1992 ), 植物个体之间彼此相互干扰, 许多个体陆续死亡(
张泽浦等, 2000 ); 而密度处于较低水平时, 植株个体的生存压力较小, 在外界条件没有较大波动的情况下, 种群数量维持着较为平稳的状态, 死亡率为0 (
方精云, 1991 ;
朴顺姬等, 1997 ). ...
日本落叶松模拟种群的生长与密度的关系
1
1991
... 单位面积生物量是评价植物种群生产力的重要指标, 国内外在种植密度对苜蓿产量的影响方面有很多的研究, 但结果有所不同.有研究证明, 增加植株密度, 可以提高作物的产量(
Deng et al ., 2012 ), 有效地抑制杂草生长(
Bybee-Finley et al ., 2017 ;
Gibson et al ., 2017 ).
王彦华等(2017) 的研究则表明, 不同密度水平下苜蓿的产量没有差别.最近的研究结果发现, 当苜蓿的播种量低于10 kg·hm
-2 时, 播种量增加可以提高产量, 而当播种量高于10 kg·hm
-2 时, 产量并不随着播种量的增加而增加(
Berti & Samarappuli, 2018 ).在本实验中, 随着种植密度的增加, 单位面积干草产量间差异不显著, 种群地上与地下的最终产量都保持恒定, 符合最终产量恒定法则.这表明了在外部条件一致的情况下, 苜蓿种群经过充分的生长后, 群体达到了最大的资源利用率, 单位面积生物量保持恒定, 而与初始种植密度无关.此前有大量的以作物为研究对象的实验证明, 在一定密度范围内, 最终产量恒定的法则是适用的(
Kira et al ., 1953 ;
方精云等, 1991 ;
樊星等, 2016 ). ...
日本落叶松模拟种群的生长与密度的关系
1
1991
... 单位面积生物量是评价植物种群生产力的重要指标, 国内外在种植密度对苜蓿产量的影响方面有很多的研究, 但结果有所不同.有研究证明, 增加植株密度, 可以提高作物的产量(
Deng et al ., 2012 ), 有效地抑制杂草生长(
Bybee-Finley et al ., 2017 ;
Gibson et al ., 2017 ).
王彦华等(2017) 的研究则表明, 不同密度水平下苜蓿的产量没有差别.最近的研究结果发现, 当苜蓿的播种量低于10 kg·hm
-2 时, 播种量增加可以提高产量, 而当播种量高于10 kg·hm
-2 时, 产量并不随着播种量的增加而增加(
Berti & Samarappuli, 2018 ).在本实验中, 随着种植密度的增加, 单位面积干草产量间差异不显著, 种群地上与地下的最终产量都保持恒定, 符合最终产量恒定法则.这表明了在外部条件一致的情况下, 苜蓿种群经过充分的生长后, 群体达到了最大的资源利用率, 单位面积生物量保持恒定, 而与初始种植密度无关.此前有大量的以作物为研究对象的实验证明, 在一定密度范围内, 最终产量恒定的法则是适用的(
Kira et al ., 1953 ;
方精云等, 1991 ;
樊星等, 2016 ). ...
1
1995
... 豆科苜蓿属(
Medicago )全世界共有56个种(
耿华珠, 1995 ), 分一年生和多年生两种类型.在中国, 尤其是在西北地区, 多年生苜蓿已被种植了2 000多年, 由于其适应性广, 产草量高, 且富含蛋白质、维生素和矿物质等营养物质, 是我国经济价值最高的豆科牧草.与此同时, 苜蓿具有优良的固氮功能, 可有效提高土壤肥力; 株丛密集, 可减少风沙对地面裸土的侵蚀; 根系发达、穿透力强, 有效地保持水土, 改善土壤结构(
洪绂曾等, 2009 ).全球苜蓿种植面积稳定在约2 380万hm
2 , 我国的苜蓿种植面积约占全球总面积的21% (
李新一等, 2015 ).虽然我国苜蓿的种植面积很大, 但高品质的苜蓿种植面积仅有20万hm
2 (
侯龙鱼等, 2018 ). ...
1
1995
... 豆科苜蓿属(
Medicago )全世界共有56个种(
耿华珠, 1995 ), 分一年生和多年生两种类型.在中国, 尤其是在西北地区, 多年生苜蓿已被种植了2 000多年, 由于其适应性广, 产草量高, 且富含蛋白质、维生素和矿物质等营养物质, 是我国经济价值最高的豆科牧草.与此同时, 苜蓿具有优良的固氮功能, 可有效提高土壤肥力; 株丛密集, 可减少风沙对地面裸土的侵蚀; 根系发达、穿透力强, 有效地保持水土, 改善土壤结构(
洪绂曾等, 2009 ).全球苜蓿种植面积稳定在约2 380万hm
2 , 我国的苜蓿种植面积约占全球总面积的21% (
李新一等, 2015 ).虽然我国苜蓿的种植面积很大, 但高品质的苜蓿种植面积仅有20万hm
2 (
侯龙鱼等, 2018 ). ...
Can weeds enhance profitability? Integrating ecological concepts to address crop-weed competition and yield quality
1
2017
... 单位面积生物量是评价植物种群生产力的重要指标, 国内外在种植密度对苜蓿产量的影响方面有很多的研究, 但结果有所不同.有研究证明, 增加植株密度, 可以提高作物的产量(
Deng et al ., 2012 ), 有效地抑制杂草生长(
Bybee-Finley et al ., 2017 ;
Gibson et al ., 2017 ).
王彦华等(2017) 的研究则表明, 不同密度水平下苜蓿的产量没有差别.最近的研究结果发现, 当苜蓿的播种量低于10 kg·hm
-2 时, 播种量增加可以提高产量, 而当播种量高于10 kg·hm
-2 时, 产量并不随着播种量的增加而增加(
Berti & Samarappuli, 2018 ).在本实验中, 随着种植密度的增加, 单位面积干草产量间差异不显著, 种群地上与地下的最终产量都保持恒定, 符合最终产量恒定法则.这表明了在外部条件一致的情况下, 苜蓿种群经过充分的生长后, 群体达到了最大的资源利用率, 单位面积生物量保持恒定, 而与初始种植密度无关.此前有大量的以作物为研究对象的实验证明, 在一定密度范围内, 最终产量恒定的法则是适用的(
Kira et al ., 1953 ;
方精云等, 1991 ;
樊星等, 2016 ). ...
The development of lucerne root morphology traits under high initial stand density within a seven year period
1
2011
... 密度是自然界的重要选择压力(
Japhet et al ., 2009 ).在密度的作用下, 植物种群内的植株互相竞争可以获得的资源, 主要包括光照、水分、矿物质等(
黎磊等, 2011 ).
Hakl等(2011) 对高密度下的苜 蓿种群数量动态进行了观测, 7年间植株数量由 5 000株·m
-2 降至57株·m
-2 .有研究表明, 苜蓿植株数量在第一生长年急剧下降, 第二生长年缓慢下降, 第三生长年趋于平稳(
王彦华等, 2017 ).本实验的研究结果显示, 初始密度对紫花苜蓿种群数量的动态变化有显著的影响, 前期随着生长时间的增长, 较高密度(1 255、1 938株·m
-2 )水平下的植株数量迅速下降, 随后缓慢下降; 中等密度(373、475株·m
-2 )水平下的植株数量一直较缓慢下降, 而较低密度(25、100株·m
-2 )水平下的植株数量则基本不变.随着生长时间的延长, 初始密度对于植物种群数量的影响有减弱的趋势.产生这种动态变化的原因可能是: 较高密度下当植物种群生长到一定阶段, 种内竞争日趋激烈(
方精云, 1992 ), 植物个体之间彼此相互干扰, 许多个体陆续死亡(
张泽浦等, 2000 ); 而密度处于较低水平时, 植株个体的生存压力较小, 在外界条件没有较大波动的情况下, 种群数量维持着较为平稳的状态, 死亡率为0 (
方精云, 1991 ;
朴顺姬等, 1997 ). ...
2
1977
... 植物种群的密度效应是种群生态学研究的一个核心问题.植物种群在资源有限的情况下, 保证每个个体的存活是困难的, 种群数量会减少, 即产生种群调节作用.在植物种群密度与生物量的关系方面, 涉及很多经典的科学问题, 前人也提出了众多经典的理论, 如最终产量恒定法则、自疏法则以及最适种植密度法则等(
Shinozaki & Kira, 1956 ;
Yoda et al ., 1963 ;
Harper, 1977 ;
Deng et al ., 2012 ).早期研究显示, 适当提高密度可以增加苜蓿的建植率和覆盖率, 是保证其高产的有效措施(
欧阳延生等, 2007 ).虽然在过去的几十年里, 有关种植密度对苜蓿影响的研究已有不少, 但多偏重于探究播种量对苜蓿产量以及品质的影响(
李丽等, 2012 ;
马克成和王秉龙, 2014 ); 从个体和群体两个方面研究种植密度影响苜蓿生长的报道较少. ...
... 种植密度能引起生长资源的强制分配, 密度的增加导致种内竞争的产生, 影响种群中植物的单株生长量和生物量(
Harper, 1977 ;
李博等, 2000 ).在很大程度上, 密度的制约作用使得种群内部个体的特征发生相应的变化.本研究中, 紫花苜蓿个体的性状随密度的变化发生了很显著的变化, 表现出极显著的密度制约特性: 随着种植密度的增加, 植物个体的株高、分枝数、基径、生物量均减小(
图3 ,
图4 ).在生长空间、光照、土壤水分和养分基本相同的情况下, 不同种植密度的同一植物种群基本具有相同的环境最大容纳量.当空间和资源有限时, 植物个体的生长可能会被抑制, 个体间产生竞争.当密度增加时, 改变自身的形态和结构是植物最易采取的、最经济的适应对策(
Silvertown & Lovett-Doust, 1993 ).尽管有研究表明, 植株高度不受密度的影响(
杜汉强等, 2004 ), 但株高随着种植密度的增加而减小也是一种“耐受”策略, 通过减缓自身的生长速度增加自身对外界胁迫的耐受力, 保证现有的尽可能多的植株的存活, 这与以往的研究结果(
陈静等, 2017 )一致.密度较小的种群在生长过程中能获得的光照、水分、养分以及空间方面的资源比较充足, 个体间的竞争较弱, 植物倾向于增加枝条或分蘖数量来提高个体生物量(
王俊峰和冯玉龙, 2004 ).而在种植密度增加, 资源紧缺的环境下, 植物倾向于减少枝条的数量和质量(
Soleymani et al ., 2011 ), 从而降低植物个体的生物量(
Wang et al ., 2019 ).另外, 关于基径与种群密度的关系,
Reineke (1933) 针对冠层密闭且同龄的植物种群提出了“立地密度法则”, 即
N =
kr -1.605 (
N 为种群密度,
r 为植物的平均直径,
k 为常数).也就是说, 随着种群密度的增加, 植物的平均单株直径会减小, 两者之间存在权衡关系(
Adams et al ., 2015 ). ...
Plant density modifies root system architecture in spring barley (
Hordeum vulgare L.) through a change in nodal root number
1
2019
... 植物的形态发育影响个体生物量(
Hecht et al ., 2019 ).构成苜蓿生物量的主要因素有单茎质量、茎长、基径、侧枝数和叶片数(
孟凯等, 2019 ).本实验中, 不同密度下, 植物个体生物量与植物个体的平均株高、基径、分枝数量联系密切.随着密度的降低, 植物个体株高、基径和分枝数量增加, 植物个体生物量逐渐增长.这是因为苜蓿可在根的基部产生根冠芽并成长为分枝, 在外界环境竞争压力较小的情况下, 单株植物可以增加分枝的数量形成一个小的“种群” (
Cain et al ., 1991 ), 进而提高单株个体的生产力(
Kays & Harper, 1974 ).植物生长过程中会争夺有限的资源, 当种内竞争日趋激烈时, 植物个体之间彼此干扰, 许多个体陆续死亡, 种群密度降低(
方精云, 1992 ).在自疏过程中植物的平均个体质量与种群密度间的关系通常符合-3/2 (
Yoda et al ., 1963 )或-4/3 (
Enquist et al ., 1998 )自疏法则.本研究中, 个体平均生物量与密度关系为-1.0, 符合最终产量恒定法则(
Kira et al ., 1953 ).一方面, 自疏法则中存活个体数量与生物量的关系是在种群密度由大变小的过程中得到的; 另一方面, 苜蓿根基部可产生根冠芽, 发育成枝条后会有水平方向的伸展, 出现匍匐生长的情况.该情况下, 植株几何形状发生变化, 会出现平均个体生物量-密度关系的斜率接近-1的情况(
王刚, 1993 ), 即形成最终产量恒定的状态. ...
1
2009
... 豆科苜蓿属(
Medicago )全世界共有56个种(
耿华珠, 1995 ), 分一年生和多年生两种类型.在中国, 尤其是在西北地区, 多年生苜蓿已被种植了2 000多年, 由于其适应性广, 产草量高, 且富含蛋白质、维生素和矿物质等营养物质, 是我国经济价值最高的豆科牧草.与此同时, 苜蓿具有优良的固氮功能, 可有效提高土壤肥力; 株丛密集, 可减少风沙对地面裸土的侵蚀; 根系发达、穿透力强, 有效地保持水土, 改善土壤结构(
洪绂曾等, 2009 ).全球苜蓿种植面积稳定在约2 380万hm
2 , 我国的苜蓿种植面积约占全球总面积的21% (
李新一等, 2015 ).虽然我国苜蓿的种植面积很大, 但高品质的苜蓿种植面积仅有20万hm
2 (
侯龙鱼等, 2018 ). ...
1
2009
... 豆科苜蓿属(
Medicago )全世界共有56个种(
耿华珠, 1995 ), 分一年生和多年生两种类型.在中国, 尤其是在西北地区, 多年生苜蓿已被种植了2 000多年, 由于其适应性广, 产草量高, 且富含蛋白质、维生素和矿物质等营养物质, 是我国经济价值最高的豆科牧草.与此同时, 苜蓿具有优良的固氮功能, 可有效提高土壤肥力; 株丛密集, 可减少风沙对地面裸土的侵蚀; 根系发达、穿透力强, 有效地保持水土, 改善土壤结构(
洪绂曾等, 2009 ).全球苜蓿种植面积稳定在约2 380万hm
2 , 我国的苜蓿种植面积约占全球总面积的21% (
李新一等, 2015 ).虽然我国苜蓿的种植面积很大, 但高品质的苜蓿种植面积仅有20万hm
2 (
侯龙鱼等, 2018 ). ...
高寒地区苜蓿人工草地建植技术
1
2018
... 豆科苜蓿属(
Medicago )全世界共有56个种(
耿华珠, 1995 ), 分一年生和多年生两种类型.在中国, 尤其是在西北地区, 多年生苜蓿已被种植了2 000多年, 由于其适应性广, 产草量高, 且富含蛋白质、维生素和矿物质等营养物质, 是我国经济价值最高的豆科牧草.与此同时, 苜蓿具有优良的固氮功能, 可有效提高土壤肥力; 株丛密集, 可减少风沙对地面裸土的侵蚀; 根系发达、穿透力强, 有效地保持水土, 改善土壤结构(
洪绂曾等, 2009 ).全球苜蓿种植面积稳定在约2 380万hm
2 , 我国的苜蓿种植面积约占全球总面积的21% (
李新一等, 2015 ).虽然我国苜蓿的种植面积很大, 但高品质的苜蓿种植面积仅有20万hm
2 (
侯龙鱼等, 2018 ). ...
高寒地区苜蓿人工草地建植技术
1
2018
... 豆科苜蓿属(
Medicago )全世界共有56个种(
耿华珠, 1995 ), 分一年生和多年生两种类型.在中国, 尤其是在西北地区, 多年生苜蓿已被种植了2 000多年, 由于其适应性广, 产草量高, 且富含蛋白质、维生素和矿物质等营养物质, 是我国经济价值最高的豆科牧草.与此同时, 苜蓿具有优良的固氮功能, 可有效提高土壤肥力; 株丛密集, 可减少风沙对地面裸土的侵蚀; 根系发达、穿透力强, 有效地保持水土, 改善土壤结构(
洪绂曾等, 2009 ).全球苜蓿种植面积稳定在约2 380万hm
2 , 我国的苜蓿种植面积约占全球总面积的21% (
李新一等, 2015 ).虽然我国苜蓿的种植面积很大, 但高品质的苜蓿种植面积仅有20万hm
2 (
侯龙鱼等, 2018 ). ...
Evidence of phenotypic plasticity in the response of
Fagopyrum esculentum to population density and sowing date
1
2009
... 密度是自然界的重要选择压力(
Japhet et al ., 2009 ).在密度的作用下, 植物种群内的植株互相竞争可以获得的资源, 主要包括光照、水分、矿物质等(
黎磊等, 2011 ).
Hakl等(2011) 对高密度下的苜 蓿种群数量动态进行了观测, 7年间植株数量由 5 000株·m
-2 降至57株·m
-2 .有研究表明, 苜蓿植株数量在第一生长年急剧下降, 第二生长年缓慢下降, 第三生长年趋于平稳(
王彦华等, 2017 ).本实验的研究结果显示, 初始密度对紫花苜蓿种群数量的动态变化有显著的影响, 前期随着生长时间的增长, 较高密度(1 255、1 938株·m
-2 )水平下的植株数量迅速下降, 随后缓慢下降; 中等密度(373、475株·m
-2 )水平下的植株数量一直较缓慢下降, 而较低密度(25、100株·m
-2 )水平下的植株数量则基本不变.随着生长时间的延长, 初始密度对于植物种群数量的影响有减弱的趋势.产生这种动态变化的原因可能是: 较高密度下当植物种群生长到一定阶段, 种内竞争日趋激烈(
方精云, 1992 ), 植物个体之间彼此相互干扰, 许多个体陆续死亡(
张泽浦等, 2000 ); 而密度处于较低水平时, 植株个体的生存压力较小, 在外界条件没有较大波动的情况下, 种群数量维持着较为平稳的状态, 死亡率为0 (
方精云, 1991 ;
朴顺姬等, 1997 ). ...
Yield components of lucerne were affected by sowing dates and inoculation treatments
1
2019
... 除了优质苜蓿的种植面积偏少, 我国在苜蓿的种植方式、栽培管理等方面也比较落后, 使得苜蓿产量和质量一直处于较低的水平.有些多年生紫花苜蓿(
Medicago sativa )人工草地不仅生产力低, 而且造成了土壤水分的严重亏缺(
程积民等, 2005 ), 不合理的利用引起了一系列的环境问题.苜蓿作物产量和营养水平受苜蓿品种、施肥量、播种时间、种植株距和行距、刈割高度、刈割时间和次数等多种因素的影响(
Chocarro & Lloveras, 2015 ;
都帅等, 2016 ;
孙万斌等, 2017 ;
魏永鹏等, 2017 ;
Jáuregui et al ., 2019 ).其中, 种植密度作为影响作物产量和品质的重要因素, 会造成植物个体对于光照、水分、养分的竞争, 这种竞争会增加个体大小之间的差异(
Weiner & Damgaard, 2006 ;
Chu et al ., 2009 ).这种差异在个体和群体的性状上都有所体现. ...
The regulation of plant and tiller density in a grass sward
1
1974
... 植物的形态发育影响个体生物量(
Hecht et al ., 2019 ).构成苜蓿生物量的主要因素有单茎质量、茎长、基径、侧枝数和叶片数(
孟凯等, 2019 ).本实验中, 不同密度下, 植物个体生物量与植物个体的平均株高、基径、分枝数量联系密切.随着密度的降低, 植物个体株高、基径和分枝数量增加, 植物个体生物量逐渐增长.这是因为苜蓿可在根的基部产生根冠芽并成长为分枝, 在外界环境竞争压力较小的情况下, 单株植物可以增加分枝的数量形成一个小的“种群” (
Cain et al ., 1991 ), 进而提高单株个体的生产力(
Kays & Harper, 1974 ).植物生长过程中会争夺有限的资源, 当种内竞争日趋激烈时, 植物个体之间彼此干扰, 许多个体陆续死亡, 种群密度降低(
方精云, 1992 ).在自疏过程中植物的平均个体质量与种群密度间的关系通常符合-3/2 (
Yoda et al ., 1963 )或-4/3 (
Enquist et al ., 1998 )自疏法则.本研究中, 个体平均生物量与密度关系为-1.0, 符合最终产量恒定法则(
Kira et al ., 1953 ).一方面, 自疏法则中存活个体数量与生物量的关系是在种群密度由大变小的过程中得到的; 另一方面, 苜蓿根基部可产生根冠芽, 发育成枝条后会有水平方向的伸展, 出现匍匐生长的情况.该情况下, 植株几何形状发生变化, 会出现平均个体生物量-密度关系的斜率接近-1的情况(
王刚, 1993 ), 即形成最终产量恒定的状态. ...
Intraspecific competition among higher plants. I Competition-density-yield interrelationship in regularly dispersed population
2
1953
... 植物的形态发育影响个体生物量(
Hecht et al ., 2019 ).构成苜蓿生物量的主要因素有单茎质量、茎长、基径、侧枝数和叶片数(
孟凯等, 2019 ).本实验中, 不同密度下, 植物个体生物量与植物个体的平均株高、基径、分枝数量联系密切.随着密度的降低, 植物个体株高、基径和分枝数量增加, 植物个体生物量逐渐增长.这是因为苜蓿可在根的基部产生根冠芽并成长为分枝, 在外界环境竞争压力较小的情况下, 单株植物可以增加分枝的数量形成一个小的“种群” (
Cain et al ., 1991 ), 进而提高单株个体的生产力(
Kays & Harper, 1974 ).植物生长过程中会争夺有限的资源, 当种内竞争日趋激烈时, 植物个体之间彼此干扰, 许多个体陆续死亡, 种群密度降低(
方精云, 1992 ).在自疏过程中植物的平均个体质量与种群密度间的关系通常符合-3/2 (
Yoda et al ., 1963 )或-4/3 (
Enquist et al ., 1998 )自疏法则.本研究中, 个体平均生物量与密度关系为-1.0, 符合最终产量恒定法则(
Kira et al ., 1953 ).一方面, 自疏法则中存活个体数量与生物量的关系是在种群密度由大变小的过程中得到的; 另一方面, 苜蓿根基部可产生根冠芽, 发育成枝条后会有水平方向的伸展, 出现匍匐生长的情况.该情况下, 植株几何形状发生变化, 会出现平均个体生物量-密度关系的斜率接近-1的情况(
王刚, 1993 ), 即形成最终产量恒定的状态. ...
... 单位面积生物量是评价植物种群生产力的重要指标, 国内外在种植密度对苜蓿产量的影响方面有很多的研究, 但结果有所不同.有研究证明, 增加植株密度, 可以提高作物的产量(
Deng et al ., 2012 ), 有效地抑制杂草生长(
Bybee-Finley et al ., 2017 ;
Gibson et al ., 2017 ).
王彦华等(2017) 的研究则表明, 不同密度水平下苜蓿的产量没有差别.最近的研究结果发现, 当苜蓿的播种量低于10 kg·hm
-2 时, 播种量增加可以提高产量, 而当播种量高于10 kg·hm
-2 时, 产量并不随着播种量的增加而增加(
Berti & Samarappuli, 2018 ).在本实验中, 随着种植密度的增加, 单位面积干草产量间差异不显著, 种群地上与地下的最终产量都保持恒定, 符合最终产量恒定法则.这表明了在外部条件一致的情况下, 苜蓿种群经过充分的生长后, 群体达到了最大的资源利用率, 单位面积生物量保持恒定, 而与初始种植密度无关.此前有大量的以作物为研究对象的实验证明, 在一定密度范围内, 最终产量恒定的法则是适用的(
Kira et al ., 1953 ;
方精云等, 1991 ;
樊星等, 2016 ). ...
1
2000
... 种植密度能引起生长资源的强制分配, 密度的增加导致种内竞争的产生, 影响种群中植物的单株生长量和生物量(
Harper, 1977 ;
李博等, 2000 ).在很大程度上, 密度的制约作用使得种群内部个体的特征发生相应的变化.本研究中, 紫花苜蓿个体的性状随密度的变化发生了很显著的变化, 表现出极显著的密度制约特性: 随着种植密度的增加, 植物个体的株高、分枝数、基径、生物量均减小(
图3 ,
图4 ).在生长空间、光照、土壤水分和养分基本相同的情况下, 不同种植密度的同一植物种群基本具有相同的环境最大容纳量.当空间和资源有限时, 植物个体的生长可能会被抑制, 个体间产生竞争.当密度增加时, 改变自身的形态和结构是植物最易采取的、最经济的适应对策(
Silvertown & Lovett-Doust, 1993 ).尽管有研究表明, 植株高度不受密度的影响(
杜汉强等, 2004 ), 但株高随着种植密度的增加而减小也是一种“耐受”策略, 通过减缓自身的生长速度增加自身对外界胁迫的耐受力, 保证现有的尽可能多的植株的存活, 这与以往的研究结果(
陈静等, 2017 )一致.密度较小的种群在生长过程中能获得的光照、水分、养分以及空间方面的资源比较充足, 个体间的竞争较弱, 植物倾向于增加枝条或分蘖数量来提高个体生物量(
王俊峰和冯玉龙, 2004 ).而在种植密度增加, 资源紧缺的环境下, 植物倾向于减少枝条的数量和质量(
Soleymani et al ., 2011 ), 从而降低植物个体的生物量(
Wang et al ., 2019 ).另外, 关于基径与种群密度的关系,
Reineke (1933) 针对冠层密闭且同龄的植物种群提出了“立地密度法则”, 即
N =
kr -1.605 (
N 为种群密度,
r 为植物的平均直径,
k 为常数).也就是说, 随着种群密度的增加, 植物的平均单株直径会减小, 两者之间存在权衡关系(
Adams et al ., 2015 ). ...
1
2000
... 种植密度能引起生长资源的强制分配, 密度的增加导致种内竞争的产生, 影响种群中植物的单株生长量和生物量(
Harper, 1977 ;
李博等, 2000 ).在很大程度上, 密度的制约作用使得种群内部个体的特征发生相应的变化.本研究中, 紫花苜蓿个体的性状随密度的变化发生了很显著的变化, 表现出极显著的密度制约特性: 随着种植密度的增加, 植物个体的株高、分枝数、基径、生物量均减小(
图3 ,
图4 ).在生长空间、光照、土壤水分和养分基本相同的情况下, 不同种植密度的同一植物种群基本具有相同的环境最大容纳量.当空间和资源有限时, 植物个体的生长可能会被抑制, 个体间产生竞争.当密度增加时, 改变自身的形态和结构是植物最易采取的、最经济的适应对策(
Silvertown & Lovett-Doust, 1993 ).尽管有研究表明, 植株高度不受密度的影响(
杜汉强等, 2004 ), 但株高随着种植密度的增加而减小也是一种“耐受”策略, 通过减缓自身的生长速度增加自身对外界胁迫的耐受力, 保证现有的尽可能多的植株的存活, 这与以往的研究结果(
陈静等, 2017 )一致.密度较小的种群在生长过程中能获得的光照、水分、养分以及空间方面的资源比较充足, 个体间的竞争较弱, 植物倾向于增加枝条或分蘖数量来提高个体生物量(
王俊峰和冯玉龙, 2004 ).而在种植密度增加, 资源紧缺的环境下, 植物倾向于减少枝条的数量和质量(
Soleymani et al ., 2011 ), 从而降低植物个体的生物量(
Wang et al ., 2019 ).另外, 关于基径与种群密度的关系,
Reineke (1933) 针对冠层密闭且同龄的植物种群提出了“立地密度法则”, 即
N =
kr -1.605 (
N 为种群密度,
r 为植物的平均直径,
k 为常数).也就是说, 随着种群密度的增加, 植物的平均单株直径会减小, 两者之间存在权衡关系(
Adams et al ., 2015 ). ...
施氮量和种植密度对紫花苜蓿生长及种子产量的影响
1
2012
... 植物种群的密度效应是种群生态学研究的一个核心问题.植物种群在资源有限的情况下, 保证每个个体的存活是困难的, 种群数量会减少, 即产生种群调节作用.在植物种群密度与生物量的关系方面, 涉及很多经典的科学问题, 前人也提出了众多经典的理论, 如最终产量恒定法则、自疏法则以及最适种植密度法则等(
Shinozaki & Kira, 1956 ;
Yoda et al ., 1963 ;
Harper, 1977 ;
Deng et al ., 2012 ).早期研究显示, 适当提高密度可以增加苜蓿的建植率和覆盖率, 是保证其高产的有效措施(
欧阳延生等, 2007 ).虽然在过去的几十年里, 有关种植密度对苜蓿影响的研究已有不少, 但多偏重于探究播种量对苜蓿产量以及品质的影响(
李丽等, 2012 ;
马克成和王秉龙, 2014 ); 从个体和群体两个方面研究种植密度影响苜蓿生长的报道较少. ...
施氮量和种植密度对紫花苜蓿生长及种子产量的影响
1
2012
... 植物种群的密度效应是种群生态学研究的一个核心问题.植物种群在资源有限的情况下, 保证每个个体的存活是困难的, 种群数量会减少, 即产生种群调节作用.在植物种群密度与生物量的关系方面, 涉及很多经典的科学问题, 前人也提出了众多经典的理论, 如最终产量恒定法则、自疏法则以及最适种植密度法则等(
Shinozaki & Kira, 1956 ;
Yoda et al ., 1963 ;
Harper, 1977 ;
Deng et al ., 2012 ).早期研究显示, 适当提高密度可以增加苜蓿的建植率和覆盖率, 是保证其高产的有效措施(
欧阳延生等, 2007 ).虽然在过去的几十年里, 有关种植密度对苜蓿影响的研究已有不少, 但多偏重于探究播种量对苜蓿产量以及品质的影响(
李丽等, 2012 ;
马克成和王秉龙, 2014 ); 从个体和群体两个方面研究种植密度影响苜蓿生长的报道较少. ...
密度制约决定的植物生物量分配格局
1
2011
... 密度是自然界的重要选择压力(
Japhet et al ., 2009 ).在密度的作用下, 植物种群内的植株互相竞争可以获得的资源, 主要包括光照、水分、矿物质等(
黎磊等, 2011 ).
Hakl等(2011) 对高密度下的苜 蓿种群数量动态进行了观测, 7年间植株数量由 5 000株·m
-2 降至57株·m
-2 .有研究表明, 苜蓿植株数量在第一生长年急剧下降, 第二生长年缓慢下降, 第三生长年趋于平稳(
王彦华等, 2017 ).本实验的研究结果显示, 初始密度对紫花苜蓿种群数量的动态变化有显著的影响, 前期随着生长时间的增长, 较高密度(1 255、1 938株·m
-2 )水平下的植株数量迅速下降, 随后缓慢下降; 中等密度(373、475株·m
-2 )水平下的植株数量一直较缓慢下降, 而较低密度(25、100株·m
-2 )水平下的植株数量则基本不变.随着生长时间的延长, 初始密度对于植物种群数量的影响有减弱的趋势.产生这种动态变化的原因可能是: 较高密度下当植物种群生长到一定阶段, 种内竞争日趋激烈(
方精云, 1992 ), 植物个体之间彼此相互干扰, 许多个体陆续死亡(
张泽浦等, 2000 ); 而密度处于较低水平时, 植株个体的生存压力较小, 在外界条件没有较大波动的情况下, 种群数量维持着较为平稳的状态, 死亡率为0 (
方精云, 1991 ;
朴顺姬等, 1997 ). ...
密度制约决定的植物生物量分配格局
1
2011
... 密度是自然界的重要选择压力(
Japhet et al ., 2009 ).在密度的作用下, 植物种群内的植株互相竞争可以获得的资源, 主要包括光照、水分、矿物质等(
黎磊等, 2011 ).
Hakl等(2011) 对高密度下的苜 蓿种群数量动态进行了观测, 7年间植株数量由 5 000株·m
-2 降至57株·m
-2 .有研究表明, 苜蓿植株数量在第一生长年急剧下降, 第二生长年缓慢下降, 第三生长年趋于平稳(
王彦华等, 2017 ).本实验的研究结果显示, 初始密度对紫花苜蓿种群数量的动态变化有显著的影响, 前期随着生长时间的增长, 较高密度(1 255、1 938株·m
-2 )水平下的植株数量迅速下降, 随后缓慢下降; 中等密度(373、475株·m
-2 )水平下的植株数量一直较缓慢下降, 而较低密度(25、100株·m
-2 )水平下的植株数量则基本不变.随着生长时间的延长, 初始密度对于植物种群数量的影响有减弱的趋势.产生这种动态变化的原因可能是: 较高密度下当植物种群生长到一定阶段, 种内竞争日趋激烈(
方精云, 1992 ), 植物个体之间彼此相互干扰, 许多个体陆续死亡(
张泽浦等, 2000 ); 而密度处于较低水平时, 植株个体的生存压力较小, 在外界条件没有较大波动的情况下, 种群数量维持着较为平稳的状态, 死亡率为0 (
方精云, 1991 ;
朴顺姬等, 1997 ). ...
我国苜蓿生产总体形势分析
1
2015
... 豆科苜蓿属(
Medicago )全世界共有56个种(
耿华珠, 1995 ), 分一年生和多年生两种类型.在中国, 尤其是在西北地区, 多年生苜蓿已被种植了2 000多年, 由于其适应性广, 产草量高, 且富含蛋白质、维生素和矿物质等营养物质, 是我国经济价值最高的豆科牧草.与此同时, 苜蓿具有优良的固氮功能, 可有效提高土壤肥力; 株丛密集, 可减少风沙对地面裸土的侵蚀; 根系发达、穿透力强, 有效地保持水土, 改善土壤结构(
洪绂曾等, 2009 ).全球苜蓿种植面积稳定在约2 380万hm
2 , 我国的苜蓿种植面积约占全球总面积的21% (
李新一等, 2015 ).虽然我国苜蓿的种植面积很大, 但高品质的苜蓿种植面积仅有20万hm
2 (
侯龙鱼等, 2018 ). ...
我国苜蓿生产总体形势分析
1
2015
... 豆科苜蓿属(
Medicago )全世界共有56个种(
耿华珠, 1995 ), 分一年生和多年生两种类型.在中国, 尤其是在西北地区, 多年生苜蓿已被种植了2 000多年, 由于其适应性广, 产草量高, 且富含蛋白质、维生素和矿物质等营养物质, 是我国经济价值最高的豆科牧草.与此同时, 苜蓿具有优良的固氮功能, 可有效提高土壤肥力; 株丛密集, 可减少风沙对地面裸土的侵蚀; 根系发达、穿透力强, 有效地保持水土, 改善土壤结构(
洪绂曾等, 2009 ).全球苜蓿种植面积稳定在约2 380万hm
2 , 我国的苜蓿种植面积约占全球总面积的21% (
李新一等, 2015 ).虽然我国苜蓿的种植面积很大, 但高品质的苜蓿种植面积仅有20万hm
2 (
侯龙鱼等, 2018 ). ...
种植及收获因子对紫花苜蓿干草产量和茎叶比的影响
1
2015
... 苜蓿是以收获茎叶为主的牧草, 并且叶片中蛋白质的含量远高于茎, 因此, 叶茎比是衡量苜蓿品质的重要指标(
刘东霞等, 2015 ).苜蓿作为多年生植物, 根的基部产生根冠芽发育为分枝, 具有表型可塑性(
Benedek & Englert, 2019 ), 可通过改变植株的形态特征(如: 叶茎比、根冠比等)适应环境, 进一步改变植物生产力和其生物量分配.以往关于种群密度是否影响植物生物量分配的研究结果尚无定论.
范高华等(2017) 认为, 植物各器官生物量均受种群密度的显著影响, 并且茎和地上生物量分配均随种群密度的增大而减小.但在本研究中, 不同密度下苜蓿种群的根冠比、叶茎比均无显著差异, 各器官生物量分配比例不受种群密度影响.同一种植物不同密度水平下, 种群的根冠比主要受环境因素影响(
平晓燕等, 2007 ), 与密度无关(
方华和孔凡斌, 2003 ;
Deru et al ., 2016 ). ...
种植及收获因子对紫花苜蓿干草产量和茎叶比的影响
1
2015
... 苜蓿是以收获茎叶为主的牧草, 并且叶片中蛋白质的含量远高于茎, 因此, 叶茎比是衡量苜蓿品质的重要指标(
刘东霞等, 2015 ).苜蓿作为多年生植物, 根的基部产生根冠芽发育为分枝, 具有表型可塑性(
Benedek & Englert, 2019 ), 可通过改变植株的形态特征(如: 叶茎比、根冠比等)适应环境, 进一步改变植物生产力和其生物量分配.以往关于种群密度是否影响植物生物量分配的研究结果尚无定论.
范高华等(2017) 认为, 植物各器官生物量均受种群密度的显著影响, 并且茎和地上生物量分配均随种群密度的增大而减小.但在本研究中, 不同密度下苜蓿种群的根冠比、叶茎比均无显著差异, 各器官生物量分配比例不受种群密度影响.同一种植物不同密度水平下, 种群的根冠比主要受环境因素影响(
平晓燕等, 2007 ), 与密度无关(
方华和孔凡斌, 2003 ;
Deru et al ., 2016 ). ...
不同行距及播量对紫花苜蓿种子产量和质量的影响
1
2014
... 植物种群的密度效应是种群生态学研究的一个核心问题.植物种群在资源有限的情况下, 保证每个个体的存活是困难的, 种群数量会减少, 即产生种群调节作用.在植物种群密度与生物量的关系方面, 涉及很多经典的科学问题, 前人也提出了众多经典的理论, 如最终产量恒定法则、自疏法则以及最适种植密度法则等(
Shinozaki & Kira, 1956 ;
Yoda et al ., 1963 ;
Harper, 1977 ;
Deng et al ., 2012 ).早期研究显示, 适当提高密度可以增加苜蓿的建植率和覆盖率, 是保证其高产的有效措施(
欧阳延生等, 2007 ).虽然在过去的几十年里, 有关种植密度对苜蓿影响的研究已有不少, 但多偏重于探究播种量对苜蓿产量以及品质的影响(
李丽等, 2012 ;
马克成和王秉龙, 2014 ); 从个体和群体两个方面研究种植密度影响苜蓿生长的报道较少. ...
不同行距及播量对紫花苜蓿种子产量和质量的影响
1
2014
... 植物种群的密度效应是种群生态学研究的一个核心问题.植物种群在资源有限的情况下, 保证每个个体的存活是困难的, 种群数量会减少, 即产生种群调节作用.在植物种群密度与生物量的关系方面, 涉及很多经典的科学问题, 前人也提出了众多经典的理论, 如最终产量恒定法则、自疏法则以及最适种植密度法则等(
Shinozaki & Kira, 1956 ;
Yoda et al ., 1963 ;
Harper, 1977 ;
Deng et al ., 2012 ).早期研究显示, 适当提高密度可以增加苜蓿的建植率和覆盖率, 是保证其高产的有效措施(
欧阳延生等, 2007 ).虽然在过去的几十年里, 有关种植密度对苜蓿影响的研究已有不少, 但多偏重于探究播种量对苜蓿产量以及品质的影响(
李丽等, 2012 ;
马克成和王秉龙, 2014 ); 从个体和群体两个方面研究种植密度影响苜蓿生长的报道较少. ...
种植密度对内蒙古中部地区苜蓿生长,饲草产量及营养品质影响
1
2019
... 植物的形态发育影响个体生物量(
Hecht et al ., 2019 ).构成苜蓿生物量的主要因素有单茎质量、茎长、基径、侧枝数和叶片数(
孟凯等, 2019 ).本实验中, 不同密度下, 植物个体生物量与植物个体的平均株高、基径、分枝数量联系密切.随着密度的降低, 植物个体株高、基径和分枝数量增加, 植物个体生物量逐渐增长.这是因为苜蓿可在根的基部产生根冠芽并成长为分枝, 在外界环境竞争压力较小的情况下, 单株植物可以增加分枝的数量形成一个小的“种群” (
Cain et al ., 1991 ), 进而提高单株个体的生产力(
Kays & Harper, 1974 ).植物生长过程中会争夺有限的资源, 当种内竞争日趋激烈时, 植物个体之间彼此干扰, 许多个体陆续死亡, 种群密度降低(
方精云, 1992 ).在自疏过程中植物的平均个体质量与种群密度间的关系通常符合-3/2 (
Yoda et al ., 1963 )或-4/3 (
Enquist et al ., 1998 )自疏法则.本研究中, 个体平均生物量与密度关系为-1.0, 符合最终产量恒定法则(
Kira et al ., 1953 ).一方面, 自疏法则中存活个体数量与生物量的关系是在种群密度由大变小的过程中得到的; 另一方面, 苜蓿根基部可产生根冠芽, 发育成枝条后会有水平方向的伸展, 出现匍匐生长的情况.该情况下, 植株几何形状发生变化, 会出现平均个体生物量-密度关系的斜率接近-1的情况(
王刚, 1993 ), 即形成最终产量恒定的状态. ...
种植密度对内蒙古中部地区苜蓿生长,饲草产量及营养品质影响
1
2019
... 植物的形态发育影响个体生物量(
Hecht et al ., 2019 ).构成苜蓿生物量的主要因素有单茎质量、茎长、基径、侧枝数和叶片数(
孟凯等, 2019 ).本实验中, 不同密度下, 植物个体生物量与植物个体的平均株高、基径、分枝数量联系密切.随着密度的降低, 植物个体株高、基径和分枝数量增加, 植物个体生物量逐渐增长.这是因为苜蓿可在根的基部产生根冠芽并成长为分枝, 在外界环境竞争压力较小的情况下, 单株植物可以增加分枝的数量形成一个小的“种群” (
Cain et al ., 1991 ), 进而提高单株个体的生产力(
Kays & Harper, 1974 ).植物生长过程中会争夺有限的资源, 当种内竞争日趋激烈时, 植物个体之间彼此干扰, 许多个体陆续死亡, 种群密度降低(
方精云, 1992 ).在自疏过程中植物的平均个体质量与种群密度间的关系通常符合-3/2 (
Yoda et al ., 1963 )或-4/3 (
Enquist et al ., 1998 )自疏法则.本研究中, 个体平均生物量与密度关系为-1.0, 符合最终产量恒定法则(
Kira et al ., 1953 ).一方面, 自疏法则中存活个体数量与生物量的关系是在种群密度由大变小的过程中得到的; 另一方面, 苜蓿根基部可产生根冠芽, 发育成枝条后会有水平方向的伸展, 出现匍匐生长的情况.该情况下, 植株几何形状发生变化, 会出现平均个体生物量-密度关系的斜率接近-1的情况(
王刚, 1993 ), 即形成最终产量恒定的状态. ...
播种量与刈割对紫花苜蓿产草量的影响
1
2007
... 植物种群的密度效应是种群生态学研究的一个核心问题.植物种群在资源有限的情况下, 保证每个个体的存活是困难的, 种群数量会减少, 即产生种群调节作用.在植物种群密度与生物量的关系方面, 涉及很多经典的科学问题, 前人也提出了众多经典的理论, 如最终产量恒定法则、自疏法则以及最适种植密度法则等(
Shinozaki & Kira, 1956 ;
Yoda et al ., 1963 ;
Harper, 1977 ;
Deng et al ., 2012 ).早期研究显示, 适当提高密度可以增加苜蓿的建植率和覆盖率, 是保证其高产的有效措施(
欧阳延生等, 2007 ).虽然在过去的几十年里, 有关种植密度对苜蓿影响的研究已有不少, 但多偏重于探究播种量对苜蓿产量以及品质的影响(
李丽等, 2012 ;
马克成和王秉龙, 2014 ); 从个体和群体两个方面研究种植密度影响苜蓿生长的报道较少. ...
播种量与刈割对紫花苜蓿产草量的影响
1
2007
... 植物种群的密度效应是种群生态学研究的一个核心问题.植物种群在资源有限的情况下, 保证每个个体的存活是困难的, 种群数量会减少, 即产生种群调节作用.在植物种群密度与生物量的关系方面, 涉及很多经典的科学问题, 前人也提出了众多经典的理论, 如最终产量恒定法则、自疏法则以及最适种植密度法则等(
Shinozaki & Kira, 1956 ;
Yoda et al ., 1963 ;
Harper, 1977 ;
Deng et al ., 2012 ).早期研究显示, 适当提高密度可以增加苜蓿的建植率和覆盖率, 是保证其高产的有效措施(
欧阳延生等, 2007 ).虽然在过去的几十年里, 有关种植密度对苜蓿影响的研究已有不少, 但多偏重于探究播种量对苜蓿产量以及品质的影响(
李丽等, 2012 ;
马克成和王秉龙, 2014 ); 从个体和群体两个方面研究种植密度影响苜蓿生长的报道较少. ...
羊草种群密度与生长动态研究
1
1997
... 密度是自然界的重要选择压力(
Japhet et al ., 2009 ).在密度的作用下, 植物种群内的植株互相竞争可以获得的资源, 主要包括光照、水分、矿物质等(
黎磊等, 2011 ).
Hakl等(2011) 对高密度下的苜 蓿种群数量动态进行了观测, 7年间植株数量由 5 000株·m
-2 降至57株·m
-2 .有研究表明, 苜蓿植株数量在第一生长年急剧下降, 第二生长年缓慢下降, 第三生长年趋于平稳(
王彦华等, 2017 ).本实验的研究结果显示, 初始密度对紫花苜蓿种群数量的动态变化有显著的影响, 前期随着生长时间的增长, 较高密度(1 255、1 938株·m
-2 )水平下的植株数量迅速下降, 随后缓慢下降; 中等密度(373、475株·m
-2 )水平下的植株数量一直较缓慢下降, 而较低密度(25、100株·m
-2 )水平下的植株数量则基本不变.随着生长时间的延长, 初始密度对于植物种群数量的影响有减弱的趋势.产生这种动态变化的原因可能是: 较高密度下当植物种群生长到一定阶段, 种内竞争日趋激烈(
方精云, 1992 ), 植物个体之间彼此相互干扰, 许多个体陆续死亡(
张泽浦等, 2000 ); 而密度处于较低水平时, 植株个体的生存压力较小, 在外界条件没有较大波动的情况下, 种群数量维持着较为平稳的状态, 死亡率为0 (
方精云, 1991 ;
朴顺姬等, 1997 ). ...
羊草种群密度与生长动态研究
1
1997
... 密度是自然界的重要选择压力(
Japhet et al ., 2009 ).在密度的作用下, 植物种群内的植株互相竞争可以获得的资源, 主要包括光照、水分、矿物质等(
黎磊等, 2011 ).
Hakl等(2011) 对高密度下的苜 蓿种群数量动态进行了观测, 7年间植株数量由 5 000株·m
-2 降至57株·m
-2 .有研究表明, 苜蓿植株数量在第一生长年急剧下降, 第二生长年缓慢下降, 第三生长年趋于平稳(
王彦华等, 2017 ).本实验的研究结果显示, 初始密度对紫花苜蓿种群数量的动态变化有显著的影响, 前期随着生长时间的增长, 较高密度(1 255、1 938株·m
-2 )水平下的植株数量迅速下降, 随后缓慢下降; 中等密度(373、475株·m
-2 )水平下的植株数量一直较缓慢下降, 而较低密度(25、100株·m
-2 )水平下的植株数量则基本不变.随着生长时间的延长, 初始密度对于植物种群数量的影响有减弱的趋势.产生这种动态变化的原因可能是: 较高密度下当植物种群生长到一定阶段, 种内竞争日趋激烈(
方精云, 1992 ), 植物个体之间彼此相互干扰, 许多个体陆续死亡(
张泽浦等, 2000 ); 而密度处于较低水平时, 植株个体的生存压力较小, 在外界条件没有较大波动的情况下, 种群数量维持着较为平稳的状态, 死亡率为0 (
方精云, 1991 ;
朴顺姬等, 1997 ). ...
羊草种群生物量分配动态模拟
1
2007
... 苜蓿是以收获茎叶为主的牧草, 并且叶片中蛋白质的含量远高于茎, 因此, 叶茎比是衡量苜蓿品质的重要指标(
刘东霞等, 2015 ).苜蓿作为多年生植物, 根的基部产生根冠芽发育为分枝, 具有表型可塑性(
Benedek & Englert, 2019 ), 可通过改变植株的形态特征(如: 叶茎比、根冠比等)适应环境, 进一步改变植物生产力和其生物量分配.以往关于种群密度是否影响植物生物量分配的研究结果尚无定论.
范高华等(2017) 认为, 植物各器官生物量均受种群密度的显著影响, 并且茎和地上生物量分配均随种群密度的增大而减小.但在本研究中, 不同密度下苜蓿种群的根冠比、叶茎比均无显著差异, 各器官生物量分配比例不受种群密度影响.同一种植物不同密度水平下, 种群的根冠比主要受环境因素影响(
平晓燕等, 2007 ), 与密度无关(
方华和孔凡斌, 2003 ;
Deru et al ., 2016 ). ...
羊草种群生物量分配动态模拟
1
2007
... 苜蓿是以收获茎叶为主的牧草, 并且叶片中蛋白质的含量远高于茎, 因此, 叶茎比是衡量苜蓿品质的重要指标(
刘东霞等, 2015 ).苜蓿作为多年生植物, 根的基部产生根冠芽发育为分枝, 具有表型可塑性(
Benedek & Englert, 2019 ), 可通过改变植株的形态特征(如: 叶茎比、根冠比等)适应环境, 进一步改变植物生产力和其生物量分配.以往关于种群密度是否影响植物生物量分配的研究结果尚无定论.
范高华等(2017) 认为, 植物各器官生物量均受种群密度的显著影响, 并且茎和地上生物量分配均随种群密度的增大而减小.但在本研究中, 不同密度下苜蓿种群的根冠比、叶茎比均无显著差异, 各器官生物量分配比例不受种群密度影响.同一种植物不同密度水平下, 种群的根冠比主要受环境因素影响(
平晓燕等, 2007 ), 与密度无关(
方华和孔凡斌, 2003 ;
Deru et al ., 2016 ). ...
Perfecting a stand-density index for even-aged forests
1
1933
... 种植密度能引起生长资源的强制分配, 密度的增加导致种内竞争的产生, 影响种群中植物的单株生长量和生物量(
Harper, 1977 ;
李博等, 2000 ).在很大程度上, 密度的制约作用使得种群内部个体的特征发生相应的变化.本研究中, 紫花苜蓿个体的性状随密度的变化发生了很显著的变化, 表现出极显著的密度制约特性: 随着种植密度的增加, 植物个体的株高、分枝数、基径、生物量均减小(
图3 ,
图4 ).在生长空间、光照、土壤水分和养分基本相同的情况下, 不同种植密度的同一植物种群基本具有相同的环境最大容纳量.当空间和资源有限时, 植物个体的生长可能会被抑制, 个体间产生竞争.当密度增加时, 改变自身的形态和结构是植物最易采取的、最经济的适应对策(
Silvertown & Lovett-Doust, 1993 ).尽管有研究表明, 植株高度不受密度的影响(
杜汉强等, 2004 ), 但株高随着种植密度的增加而减小也是一种“耐受”策略, 通过减缓自身的生长速度增加自身对外界胁迫的耐受力, 保证现有的尽可能多的植株的存活, 这与以往的研究结果(
陈静等, 2017 )一致.密度较小的种群在生长过程中能获得的光照、水分、养分以及空间方面的资源比较充足, 个体间的竞争较弱, 植物倾向于增加枝条或分蘖数量来提高个体生物量(
王俊峰和冯玉龙, 2004 ).而在种植密度增加, 资源紧缺的环境下, 植物倾向于减少枝条的数量和质量(
Soleymani et al ., 2011 ), 从而降低植物个体的生物量(
Wang et al ., 2019 ).另外, 关于基径与种群密度的关系,
Reineke (1933) 针对冠层密闭且同龄的植物种群提出了“立地密度法则”, 即
N =
kr -1.605 (
N 为种群密度,
r 为植物的平均直径,
k 为常数).也就是说, 随着种群密度的增加, 植物的平均单株直径会减小, 两者之间存在权衡关系(
Adams et al ., 2015 ). ...
Intraspecific competition among higher plants. VII. Logistic theory of the C-D effect
1
1956
... 植物种群的密度效应是种群生态学研究的一个核心问题.植物种群在资源有限的情况下, 保证每个个体的存活是困难的, 种群数量会减少, 即产生种群调节作用.在植物种群密度与生物量的关系方面, 涉及很多经典的科学问题, 前人也提出了众多经典的理论, 如最终产量恒定法则、自疏法则以及最适种植密度法则等(
Shinozaki & Kira, 1956 ;
Yoda et al ., 1963 ;
Harper, 1977 ;
Deng et al ., 2012 ).早期研究显示, 适当提高密度可以增加苜蓿的建植率和覆盖率, 是保证其高产的有效措施(
欧阳延生等, 2007 ).虽然在过去的几十年里, 有关种植密度对苜蓿影响的研究已有不少, 但多偏重于探究播种量对苜蓿产量以及品质的影响(
李丽等, 2012 ;
马克成和王秉龙, 2014 ); 从个体和群体两个方面研究种植密度影响苜蓿生长的报道较少. ...
Introduction to Plant Population Biology. 3rd ed. Blackwell Science, London
1
1993
... 种植密度能引起生长资源的强制分配, 密度的增加导致种内竞争的产生, 影响种群中植物的单株生长量和生物量(
Harper, 1977 ;
李博等, 2000 ).在很大程度上, 密度的制约作用使得种群内部个体的特征发生相应的变化.本研究中, 紫花苜蓿个体的性状随密度的变化发生了很显著的变化, 表现出极显著的密度制约特性: 随着种植密度的增加, 植物个体的株高、分枝数、基径、生物量均减小(
图3 ,
图4 ).在生长空间、光照、土壤水分和养分基本相同的情况下, 不同种植密度的同一植物种群基本具有相同的环境最大容纳量.当空间和资源有限时, 植物个体的生长可能会被抑制, 个体间产生竞争.当密度增加时, 改变自身的形态和结构是植物最易采取的、最经济的适应对策(
Silvertown & Lovett-Doust, 1993 ).尽管有研究表明, 植株高度不受密度的影响(
杜汉强等, 2004 ), 但株高随着种植密度的增加而减小也是一种“耐受”策略, 通过减缓自身的生长速度增加自身对外界胁迫的耐受力, 保证现有的尽可能多的植株的存活, 这与以往的研究结果(
陈静等, 2017 )一致.密度较小的种群在生长过程中能获得的光照、水分、养分以及空间方面的资源比较充足, 个体间的竞争较弱, 植物倾向于增加枝条或分蘖数量来提高个体生物量(
王俊峰和冯玉龙, 2004 ).而在种植密度增加, 资源紧缺的环境下, 植物倾向于减少枝条的数量和质量(
Soleymani et al ., 2011 ), 从而降低植物个体的生物量(
Wang et al ., 2019 ).另外, 关于基径与种群密度的关系,
Reineke (1933) 针对冠层密闭且同龄的植物种群提出了“立地密度法则”, 即
N =
kr -1.605 (
N 为种群密度,
r 为植物的平均直径,
k 为常数).也就是说, 随着种群密度的增加, 植物的平均单株直径会减小, 两者之间存在权衡关系(
Adams et al ., 2015 ). ...
Determination of the suitable planting date and plant density for different cultivars of barley (
Hordeum vulgare L.) in Fars
1
2011
... 种植密度能引起生长资源的强制分配, 密度的增加导致种内竞争的产生, 影响种群中植物的单株生长量和生物量(
Harper, 1977 ;
李博等, 2000 ).在很大程度上, 密度的制约作用使得种群内部个体的特征发生相应的变化.本研究中, 紫花苜蓿个体的性状随密度的变化发生了很显著的变化, 表现出极显著的密度制约特性: 随着种植密度的增加, 植物个体的株高、分枝数、基径、生物量均减小(
图3 ,
图4 ).在生长空间、光照、土壤水分和养分基本相同的情况下, 不同种植密度的同一植物种群基本具有相同的环境最大容纳量.当空间和资源有限时, 植物个体的生长可能会被抑制, 个体间产生竞争.当密度增加时, 改变自身的形态和结构是植物最易采取的、最经济的适应对策(
Silvertown & Lovett-Doust, 1993 ).尽管有研究表明, 植株高度不受密度的影响(
杜汉强等, 2004 ), 但株高随着种植密度的增加而减小也是一种“耐受”策略, 通过减缓自身的生长速度增加自身对外界胁迫的耐受力, 保证现有的尽可能多的植株的存活, 这与以往的研究结果(
陈静等, 2017 )一致.密度较小的种群在生长过程中能获得的光照、水分、养分以及空间方面的资源比较充足, 个体间的竞争较弱, 植物倾向于增加枝条或分蘖数量来提高个体生物量(
王俊峰和冯玉龙, 2004 ).而在种植密度增加, 资源紧缺的环境下, 植物倾向于减少枝条的数量和质量(
Soleymani et al ., 2011 ), 从而降低植物个体的生物量(
Wang et al ., 2019 ).另外, 关于基径与种群密度的关系,
Reineke (1933) 针对冠层密闭且同龄的植物种群提出了“立地密度法则”, 即
N =
kr -1.605 (
N 为种群密度,
r 为植物的平均直径,
k 为常数).也就是说, 随着种群密度的增加, 植物的平均单株直径会减小, 两者之间存在权衡关系(
Adams et al ., 2015 ). ...
不同紫花苜蓿品种在不同生育期营养品质特性的比较
1
2017
... 除了优质苜蓿的种植面积偏少, 我国在苜蓿的种植方式、栽培管理等方面也比较落后, 使得苜蓿产量和质量一直处于较低的水平.有些多年生紫花苜蓿(
Medicago sativa )人工草地不仅生产力低, 而且造成了土壤水分的严重亏缺(
程积民等, 2005 ), 不合理的利用引起了一系列的环境问题.苜蓿作物产量和营养水平受苜蓿品种、施肥量、播种时间、种植株距和行距、刈割高度、刈割时间和次数等多种因素的影响(
Chocarro & Lloveras, 2015 ;
都帅等, 2016 ;
孙万斌等, 2017 ;
魏永鹏等, 2017 ;
Jáuregui et al ., 2019 ).其中, 种植密度作为影响作物产量和品质的重要因素, 会造成植物个体对于光照、水分、养分的竞争, 这种竞争会增加个体大小之间的差异(
Weiner & Damgaard, 2006 ;
Chu et al ., 2009 ).这种差异在个体和群体的性状上都有所体现. ...
不同紫花苜蓿品种在不同生育期营养品质特性的比较
1
2017
... 除了优质苜蓿的种植面积偏少, 我国在苜蓿的种植方式、栽培管理等方面也比较落后, 使得苜蓿产量和质量一直处于较低的水平.有些多年生紫花苜蓿(
Medicago sativa )人工草地不仅生产力低, 而且造成了土壤水分的严重亏缺(
程积民等, 2005 ), 不合理的利用引起了一系列的环境问题.苜蓿作物产量和营养水平受苜蓿品种、施肥量、播种时间、种植株距和行距、刈割高度、刈割时间和次数等多种因素的影响(
Chocarro & Lloveras, 2015 ;
都帅等, 2016 ;
孙万斌等, 2017 ;
魏永鹏等, 2017 ;
Jáuregui et al ., 2019 ).其中, 种植密度作为影响作物产量和品质的重要因素, 会造成植物个体对于光照、水分、养分的竞争, 这种竞争会增加个体大小之间的差异(
Weiner & Damgaard, 2006 ;
Chu et al ., 2009 ).这种差异在个体和群体的性状上都有所体现. ...
关于植物自疏过程的一般模型
1
1993
... 植物的形态发育影响个体生物量(
Hecht et al ., 2019 ).构成苜蓿生物量的主要因素有单茎质量、茎长、基径、侧枝数和叶片数(
孟凯等, 2019 ).本实验中, 不同密度下, 植物个体生物量与植物个体的平均株高、基径、分枝数量联系密切.随着密度的降低, 植物个体株高、基径和分枝数量增加, 植物个体生物量逐渐增长.这是因为苜蓿可在根的基部产生根冠芽并成长为分枝, 在外界环境竞争压力较小的情况下, 单株植物可以增加分枝的数量形成一个小的“种群” (
Cain et al ., 1991 ), 进而提高单株个体的生产力(
Kays & Harper, 1974 ).植物生长过程中会争夺有限的资源, 当种内竞争日趋激烈时, 植物个体之间彼此干扰, 许多个体陆续死亡, 种群密度降低(
方精云, 1992 ).在自疏过程中植物的平均个体质量与种群密度间的关系通常符合-3/2 (
Yoda et al ., 1963 )或-4/3 (
Enquist et al ., 1998 )自疏法则.本研究中, 个体平均生物量与密度关系为-1.0, 符合最终产量恒定法则(
Kira et al ., 1953 ).一方面, 自疏法则中存活个体数量与生物量的关系是在种群密度由大变小的过程中得到的; 另一方面, 苜蓿根基部可产生根冠芽, 发育成枝条后会有水平方向的伸展, 出现匍匐生长的情况.该情况下, 植株几何形状发生变化, 会出现平均个体生物量-密度关系的斜率接近-1的情况(
王刚, 1993 ), 即形成最终产量恒定的状态. ...
关于植物自疏过程的一般模型
1
1993
... 植物的形态发育影响个体生物量(
Hecht et al ., 2019 ).构成苜蓿生物量的主要因素有单茎质量、茎长、基径、侧枝数和叶片数(
孟凯等, 2019 ).本实验中, 不同密度下, 植物个体生物量与植物个体的平均株高、基径、分枝数量联系密切.随着密度的降低, 植物个体株高、基径和分枝数量增加, 植物个体生物量逐渐增长.这是因为苜蓿可在根的基部产生根冠芽并成长为分枝, 在外界环境竞争压力较小的情况下, 单株植物可以增加分枝的数量形成一个小的“种群” (
Cain et al ., 1991 ), 进而提高单株个体的生产力(
Kays & Harper, 1974 ).植物生长过程中会争夺有限的资源, 当种内竞争日趋激烈时, 植物个体之间彼此干扰, 许多个体陆续死亡, 种群密度降低(
方精云, 1992 ).在自疏过程中植物的平均个体质量与种群密度间的关系通常符合-3/2 (
Yoda et al ., 1963 )或-4/3 (
Enquist et al ., 1998 )自疏法则.本研究中, 个体平均生物量与密度关系为-1.0, 符合最终产量恒定法则(
Kira et al ., 1953 ).一方面, 自疏法则中存活个体数量与生物量的关系是在种群密度由大变小的过程中得到的; 另一方面, 苜蓿根基部可产生根冠芽, 发育成枝条后会有水平方向的伸展, 出现匍匐生长的情况.该情况下, 植株几何形状发生变化, 会出现平均个体生物量-密度关系的斜率接近-1的情况(
王刚, 1993 ), 即形成最终产量恒定的状态. ...
光强对两种入侵植物生物量分配、叶片形态和相对生长速率的影响
1
2004
... 种植密度能引起生长资源的强制分配, 密度的增加导致种内竞争的产生, 影响种群中植物的单株生长量和生物量(
Harper, 1977 ;
李博等, 2000 ).在很大程度上, 密度的制约作用使得种群内部个体的特征发生相应的变化.本研究中, 紫花苜蓿个体的性状随密度的变化发生了很显著的变化, 表现出极显著的密度制约特性: 随着种植密度的增加, 植物个体的株高、分枝数、基径、生物量均减小(
图3 ,
图4 ).在生长空间、光照、土壤水分和养分基本相同的情况下, 不同种植密度的同一植物种群基本具有相同的环境最大容纳量.当空间和资源有限时, 植物个体的生长可能会被抑制, 个体间产生竞争.当密度增加时, 改变自身的形态和结构是植物最易采取的、最经济的适应对策(
Silvertown & Lovett-Doust, 1993 ).尽管有研究表明, 植株高度不受密度的影响(
杜汉强等, 2004 ), 但株高随着种植密度的增加而减小也是一种“耐受”策略, 通过减缓自身的生长速度增加自身对外界胁迫的耐受力, 保证现有的尽可能多的植株的存活, 这与以往的研究结果(
陈静等, 2017 )一致.密度较小的种群在生长过程中能获得的光照、水分、养分以及空间方面的资源比较充足, 个体间的竞争较弱, 植物倾向于增加枝条或分蘖数量来提高个体生物量(
王俊峰和冯玉龙, 2004 ).而在种植密度增加, 资源紧缺的环境下, 植物倾向于减少枝条的数量和质量(
Soleymani et al ., 2011 ), 从而降低植物个体的生物量(
Wang et al ., 2019 ).另外, 关于基径与种群密度的关系,
Reineke (1933) 针对冠层密闭且同龄的植物种群提出了“立地密度法则”, 即
N =
kr -1.605 (
N 为种群密度,
r 为植物的平均直径,
k 为常数).也就是说, 随着种群密度的增加, 植物的平均单株直径会减小, 两者之间存在权衡关系(
Adams et al ., 2015 ). ...
光强对两种入侵植物生物量分配、叶片形态和相对生长速率的影响
1
2004
... 种植密度能引起生长资源的强制分配, 密度的增加导致种内竞争的产生, 影响种群中植物的单株生长量和生物量(
Harper, 1977 ;
李博等, 2000 ).在很大程度上, 密度的制约作用使得种群内部个体的特征发生相应的变化.本研究中, 紫花苜蓿个体的性状随密度的变化发生了很显著的变化, 表现出极显著的密度制约特性: 随着种植密度的增加, 植物个体的株高、分枝数、基径、生物量均减小(
图3 ,
图4 ).在生长空间、光照、土壤水分和养分基本相同的情况下, 不同种植密度的同一植物种群基本具有相同的环境最大容纳量.当空间和资源有限时, 植物个体的生长可能会被抑制, 个体间产生竞争.当密度增加时, 改变自身的形态和结构是植物最易采取的、最经济的适应对策(
Silvertown & Lovett-Doust, 1993 ).尽管有研究表明, 植株高度不受密度的影响(
杜汉强等, 2004 ), 但株高随着种植密度的增加而减小也是一种“耐受”策略, 通过减缓自身的生长速度增加自身对外界胁迫的耐受力, 保证现有的尽可能多的植株的存活, 这与以往的研究结果(
陈静等, 2017 )一致.密度较小的种群在生长过程中能获得的光照、水分、养分以及空间方面的资源比较充足, 个体间的竞争较弱, 植物倾向于增加枝条或分蘖数量来提高个体生物量(
王俊峰和冯玉龙, 2004 ).而在种植密度增加, 资源紧缺的环境下, 植物倾向于减少枝条的数量和质量(
Soleymani et al ., 2011 ), 从而降低植物个体的生物量(
Wang et al ., 2019 ).另外, 关于基径与种群密度的关系,
Reineke (1933) 针对冠层密闭且同龄的植物种群提出了“立地密度法则”, 即
N =
kr -1.605 (
N 为种群密度,
r 为植物的平均直径,
k 为常数).也就是说, 随着种群密度的增加, 植物的平均单株直径会减小, 两者之间存在权衡关系(
Adams et al ., 2015 ). ...
Summer drought decreases
Leymus chinensis productivity through constraining the bud, tiller and shoot production
1
2019
... 种植密度能引起生长资源的强制分配, 密度的增加导致种内竞争的产生, 影响种群中植物的单株生长量和生物量(
Harper, 1977 ;
李博等, 2000 ).在很大程度上, 密度的制约作用使得种群内部个体的特征发生相应的变化.本研究中, 紫花苜蓿个体的性状随密度的变化发生了很显著的变化, 表现出极显著的密度制约特性: 随着种植密度的增加, 植物个体的株高、分枝数、基径、生物量均减小(
图3 ,
图4 ).在生长空间、光照、土壤水分和养分基本相同的情况下, 不同种植密度的同一植物种群基本具有相同的环境最大容纳量.当空间和资源有限时, 植物个体的生长可能会被抑制, 个体间产生竞争.当密度增加时, 改变自身的形态和结构是植物最易采取的、最经济的适应对策(
Silvertown & Lovett-Doust, 1993 ).尽管有研究表明, 植株高度不受密度的影响(
杜汉强等, 2004 ), 但株高随着种植密度的增加而减小也是一种“耐受”策略, 通过减缓自身的生长速度增加自身对外界胁迫的耐受力, 保证现有的尽可能多的植株的存活, 这与以往的研究结果(
陈静等, 2017 )一致.密度较小的种群在生长过程中能获得的光照、水分、养分以及空间方面的资源比较充足, 个体间的竞争较弱, 植物倾向于增加枝条或分蘖数量来提高个体生物量(
王俊峰和冯玉龙, 2004 ).而在种植密度增加, 资源紧缺的环境下, 植物倾向于减少枝条的数量和质量(
Soleymani et al ., 2011 ), 从而降低植物个体的生物量(
Wang et al ., 2019 ).另外, 关于基径与种群密度的关系,
Reineke (1933) 针对冠层密闭且同龄的植物种群提出了“立地密度法则”, 即
N =
kr -1.605 (
N 为种群密度,
r 为植物的平均直径,
k 为常数).也就是说, 随着种群密度的增加, 植物的平均单株直径会减小, 两者之间存在权衡关系(
Adams et al ., 2015 ). ...
播种量和品种对紫花苜蓿植株动态变化、产量及品质的影响
2
2017
... 密度是自然界的重要选择压力(
Japhet et al ., 2009 ).在密度的作用下, 植物种群内的植株互相竞争可以获得的资源, 主要包括光照、水分、矿物质等(
黎磊等, 2011 ).
Hakl等(2011) 对高密度下的苜 蓿种群数量动态进行了观测, 7年间植株数量由 5 000株·m
-2 降至57株·m
-2 .有研究表明, 苜蓿植株数量在第一生长年急剧下降, 第二生长年缓慢下降, 第三生长年趋于平稳(
王彦华等, 2017 ).本实验的研究结果显示, 初始密度对紫花苜蓿种群数量的动态变化有显著的影响, 前期随着生长时间的增长, 较高密度(1 255、1 938株·m
-2 )水平下的植株数量迅速下降, 随后缓慢下降; 中等密度(373、475株·m
-2 )水平下的植株数量一直较缓慢下降, 而较低密度(25、100株·m
-2 )水平下的植株数量则基本不变.随着生长时间的延长, 初始密度对于植物种群数量的影响有减弱的趋势.产生这种动态变化的原因可能是: 较高密度下当植物种群生长到一定阶段, 种内竞争日趋激烈(
方精云, 1992 ), 植物个体之间彼此相互干扰, 许多个体陆续死亡(
张泽浦等, 2000 ); 而密度处于较低水平时, 植株个体的生存压力较小, 在外界条件没有较大波动的情况下, 种群数量维持着较为平稳的状态, 死亡率为0 (
方精云, 1991 ;
朴顺姬等, 1997 ). ...
... 单位面积生物量是评价植物种群生产力的重要指标, 国内外在种植密度对苜蓿产量的影响方面有很多的研究, 但结果有所不同.有研究证明, 增加植株密度, 可以提高作物的产量(
Deng et al ., 2012 ), 有效地抑制杂草生长(
Bybee-Finley et al ., 2017 ;
Gibson et al ., 2017 ).
王彦华等(2017) 的研究则表明, 不同密度水平下苜蓿的产量没有差别.最近的研究结果发现, 当苜蓿的播种量低于10 kg·hm
-2 时, 播种量增加可以提高产量, 而当播种量高于10 kg·hm
-2 时, 产量并不随着播种量的增加而增加(
Berti & Samarappuli, 2018 ).在本实验中, 随着种植密度的增加, 单位面积干草产量间差异不显著, 种群地上与地下的最终产量都保持恒定, 符合最终产量恒定法则.这表明了在外部条件一致的情况下, 苜蓿种群经过充分的生长后, 群体达到了最大的资源利用率, 单位面积生物量保持恒定, 而与初始种植密度无关.此前有大量的以作物为研究对象的实验证明, 在一定密度范围内, 最终产量恒定的法则是适用的(
Kira et al ., 1953 ;
方精云等, 1991 ;
樊星等, 2016 ). ...
播种量和品种对紫花苜蓿植株动态变化、产量及品质的影响
2
2017
... 密度是自然界的重要选择压力(
Japhet et al ., 2009 ).在密度的作用下, 植物种群内的植株互相竞争可以获得的资源, 主要包括光照、水分、矿物质等(
黎磊等, 2011 ).
Hakl等(2011) 对高密度下的苜 蓿种群数量动态进行了观测, 7年间植株数量由 5 000株·m
-2 降至57株·m
-2 .有研究表明, 苜蓿植株数量在第一生长年急剧下降, 第二生长年缓慢下降, 第三生长年趋于平稳(
王彦华等, 2017 ).本实验的研究结果显示, 初始密度对紫花苜蓿种群数量的动态变化有显著的影响, 前期随着生长时间的增长, 较高密度(1 255、1 938株·m
-2 )水平下的植株数量迅速下降, 随后缓慢下降; 中等密度(373、475株·m
-2 )水平下的植株数量一直较缓慢下降, 而较低密度(25、100株·m
-2 )水平下的植株数量则基本不变.随着生长时间的延长, 初始密度对于植物种群数量的影响有减弱的趋势.产生这种动态变化的原因可能是: 较高密度下当植物种群生长到一定阶段, 种内竞争日趋激烈(
方精云, 1992 ), 植物个体之间彼此相互干扰, 许多个体陆续死亡(
张泽浦等, 2000 ); 而密度处于较低水平时, 植株个体的生存压力较小, 在外界条件没有较大波动的情况下, 种群数量维持着较为平稳的状态, 死亡率为0 (
方精云, 1991 ;
朴顺姬等, 1997 ). ...
... 单位面积生物量是评价植物种群生产力的重要指标, 国内外在种植密度对苜蓿产量的影响方面有很多的研究, 但结果有所不同.有研究证明, 增加植株密度, 可以提高作物的产量(
Deng et al ., 2012 ), 有效地抑制杂草生长(
Bybee-Finley et al ., 2017 ;
Gibson et al ., 2017 ).
王彦华等(2017) 的研究则表明, 不同密度水平下苜蓿的产量没有差别.最近的研究结果发现, 当苜蓿的播种量低于10 kg·hm
-2 时, 播种量增加可以提高产量, 而当播种量高于10 kg·hm
-2 时, 产量并不随着播种量的增加而增加(
Berti & Samarappuli, 2018 ).在本实验中, 随着种植密度的增加, 单位面积干草产量间差异不显著, 种群地上与地下的最终产量都保持恒定, 符合最终产量恒定法则.这表明了在外部条件一致的情况下, 苜蓿种群经过充分的生长后, 群体达到了最大的资源利用率, 单位面积生物量保持恒定, 而与初始种植密度无关.此前有大量的以作物为研究对象的实验证明, 在一定密度范围内, 最终产量恒定的法则是适用的(
Kira et al ., 1953 ;
方精云等, 1991 ;
樊星等, 2016 ). ...
种植密度和行距配置对紫花苜蓿群体产量及品质的影响
1
2017
... 除了优质苜蓿的种植面积偏少, 我国在苜蓿的种植方式、栽培管理等方面也比较落后, 使得苜蓿产量和质量一直处于较低的水平.有些多年生紫花苜蓿(
Medicago sativa )人工草地不仅生产力低, 而且造成了土壤水分的严重亏缺(
程积民等, 2005 ), 不合理的利用引起了一系列的环境问题.苜蓿作物产量和营养水平受苜蓿品种、施肥量、播种时间、种植株距和行距、刈割高度、刈割时间和次数等多种因素的影响(
Chocarro & Lloveras, 2015 ;
都帅等, 2016 ;
孙万斌等, 2017 ;
魏永鹏等, 2017 ;
Jáuregui et al ., 2019 ).其中, 种植密度作为影响作物产量和品质的重要因素, 会造成植物个体对于光照、水分、养分的竞争, 这种竞争会增加个体大小之间的差异(
Weiner & Damgaard, 2006 ;
Chu et al ., 2009 ).这种差异在个体和群体的性状上都有所体现. ...
种植密度和行距配置对紫花苜蓿群体产量及品质的影响
1
2017
... 除了优质苜蓿的种植面积偏少, 我国在苜蓿的种植方式、栽培管理等方面也比较落后, 使得苜蓿产量和质量一直处于较低的水平.有些多年生紫花苜蓿(
Medicago sativa )人工草地不仅生产力低, 而且造成了土壤水分的严重亏缺(
程积民等, 2005 ), 不合理的利用引起了一系列的环境问题.苜蓿作物产量和营养水平受苜蓿品种、施肥量、播种时间、种植株距和行距、刈割高度、刈割时间和次数等多种因素的影响(
Chocarro & Lloveras, 2015 ;
都帅等, 2016 ;
孙万斌等, 2017 ;
魏永鹏等, 2017 ;
Jáuregui et al ., 2019 ).其中, 种植密度作为影响作物产量和品质的重要因素, 会造成植物个体对于光照、水分、养分的竞争, 这种竞争会增加个体大小之间的差异(
Weiner & Damgaard, 2006 ;
Chu et al ., 2009 ).这种差异在个体和群体的性状上都有所体现. ...
Size-asymmetric competition and size-asymmetric growth in a spatially explicit zone-of-influence model of plant competition
1
2006
... 除了优质苜蓿的种植面积偏少, 我国在苜蓿的种植方式、栽培管理等方面也比较落后, 使得苜蓿产量和质量一直处于较低的水平.有些多年生紫花苜蓿(
Medicago sativa )人工草地不仅生产力低, 而且造成了土壤水分的严重亏缺(
程积民等, 2005 ), 不合理的利用引起了一系列的环境问题.苜蓿作物产量和营养水平受苜蓿品种、施肥量、播种时间、种植株距和行距、刈割高度、刈割时间和次数等多种因素的影响(
Chocarro & Lloveras, 2015 ;
都帅等, 2016 ;
孙万斌等, 2017 ;
魏永鹏等, 2017 ;
Jáuregui et al ., 2019 ).其中, 种植密度作为影响作物产量和品质的重要因素, 会造成植物个体对于光照、水分、养分的竞争, 这种竞争会增加个体大小之间的差异(
Weiner & Damgaard, 2006 ;
Chu et al ., 2009 ).这种差异在个体和群体的性状上都有所体现. ...
Self-thinning in over-crowded pure stands under cultivated and natural conditions
2
1963
... 植物种群的密度效应是种群生态学研究的一个核心问题.植物种群在资源有限的情况下, 保证每个个体的存活是困难的, 种群数量会减少, 即产生种群调节作用.在植物种群密度与生物量的关系方面, 涉及很多经典的科学问题, 前人也提出了众多经典的理论, 如最终产量恒定法则、自疏法则以及最适种植密度法则等(
Shinozaki & Kira, 1956 ;
Yoda et al ., 1963 ;
Harper, 1977 ;
Deng et al ., 2012 ).早期研究显示, 适当提高密度可以增加苜蓿的建植率和覆盖率, 是保证其高产的有效措施(
欧阳延生等, 2007 ).虽然在过去的几十年里, 有关种植密度对苜蓿影响的研究已有不少, 但多偏重于探究播种量对苜蓿产量以及品质的影响(
李丽等, 2012 ;
马克成和王秉龙, 2014 ); 从个体和群体两个方面研究种植密度影响苜蓿生长的报道较少. ...
... 植物的形态发育影响个体生物量(
Hecht et al ., 2019 ).构成苜蓿生物量的主要因素有单茎质量、茎长、基径、侧枝数和叶片数(
孟凯等, 2019 ).本实验中, 不同密度下, 植物个体生物量与植物个体的平均株高、基径、分枝数量联系密切.随着密度的降低, 植物个体株高、基径和分枝数量增加, 植物个体生物量逐渐增长.这是因为苜蓿可在根的基部产生根冠芽并成长为分枝, 在外界环境竞争压力较小的情况下, 单株植物可以增加分枝的数量形成一个小的“种群” (
Cain et al ., 1991 ), 进而提高单株个体的生产力(
Kays & Harper, 1974 ).植物生长过程中会争夺有限的资源, 当种内竞争日趋激烈时, 植物个体之间彼此干扰, 许多个体陆续死亡, 种群密度降低(
方精云, 1992 ).在自疏过程中植物的平均个体质量与种群密度间的关系通常符合-3/2 (
Yoda et al ., 1963 )或-4/3 (
Enquist et al ., 1998 )自疏法则.本研究中, 个体平均生物量与密度关系为-1.0, 符合最终产量恒定法则(
Kira et al ., 1953 ).一方面, 自疏法则中存活个体数量与生物量的关系是在种群密度由大变小的过程中得到的; 另一方面, 苜蓿根基部可产生根冠芽, 发育成枝条后会有水平方向的伸展, 出现匍匐生长的情况.该情况下, 植株几何形状发生变化, 会出现平均个体生物量-密度关系的斜率接近-1的情况(
王刚, 1993 ), 即形成最终产量恒定的状态. ...
邻体竞争对植物个体生长速率和死亡概率的影响: 基于日本落叶松种群试验的研究
1
2000
... 密度是自然界的重要选择压力(
Japhet et al ., 2009 ).在密度的作用下, 植物种群内的植株互相竞争可以获得的资源, 主要包括光照、水分、矿物质等(
黎磊等, 2011 ).
Hakl等(2011) 对高密度下的苜 蓿种群数量动态进行了观测, 7年间植株数量由 5 000株·m
-2 降至57株·m
-2 .有研究表明, 苜蓿植株数量在第一生长年急剧下降, 第二生长年缓慢下降, 第三生长年趋于平稳(
王彦华等, 2017 ).本实验的研究结果显示, 初始密度对紫花苜蓿种群数量的动态变化有显著的影响, 前期随着生长时间的增长, 较高密度(1 255、1 938株·m
-2 )水平下的植株数量迅速下降, 随后缓慢下降; 中等密度(373、475株·m
-2 )水平下的植株数量一直较缓慢下降, 而较低密度(25、100株·m
-2 )水平下的植株数量则基本不变.随着生长时间的延长, 初始密度对于植物种群数量的影响有减弱的趋势.产生这种动态变化的原因可能是: 较高密度下当植物种群生长到一定阶段, 种内竞争日趋激烈(
方精云, 1992 ), 植物个体之间彼此相互干扰, 许多个体陆续死亡(
张泽浦等, 2000 ); 而密度处于较低水平时, 植株个体的生存压力较小, 在外界条件没有较大波动的情况下, 种群数量维持着较为平稳的状态, 死亡率为0 (
方精云, 1991 ;
朴顺姬等, 1997 ). ...
邻体竞争对植物个体生长速率和死亡概率的影响: 基于日本落叶松种群试验的研究
1
2000
... 密度是自然界的重要选择压力(
Japhet et al ., 2009 ).在密度的作用下, 植物种群内的植株互相竞争可以获得的资源, 主要包括光照、水分、矿物质等(
黎磊等, 2011 ).
Hakl等(2011) 对高密度下的苜 蓿种群数量动态进行了观测, 7年间植株数量由 5 000株·m
-2 降至57株·m
-2 .有研究表明, 苜蓿植株数量在第一生长年急剧下降, 第二生长年缓慢下降, 第三生长年趋于平稳(
王彦华等, 2017 ).本实验的研究结果显示, 初始密度对紫花苜蓿种群数量的动态变化有显著的影响, 前期随着生长时间的增长, 较高密度(1 255、1 938株·m
-2 )水平下的植株数量迅速下降, 随后缓慢下降; 中等密度(373、475株·m
-2 )水平下的植株数量一直较缓慢下降, 而较低密度(25、100株·m
-2 )水平下的植株数量则基本不变.随着生长时间的延长, 初始密度对于植物种群数量的影响有减弱的趋势.产生这种动态变化的原因可能是: 较高密度下当植物种群生长到一定阶段, 种内竞争日趋激烈(
方精云, 1992 ), 植物个体之间彼此相互干扰, 许多个体陆续死亡(
张泽浦等, 2000 ); 而密度处于较低水平时, 植株个体的生存压力较小, 在外界条件没有较大波动的情况下, 种群数量维持着较为平稳的状态, 死亡率为0 (
方精云, 1991 ;
朴顺姬等, 1997 ). ...
 ,1,2,3,*
,1,2,3,*
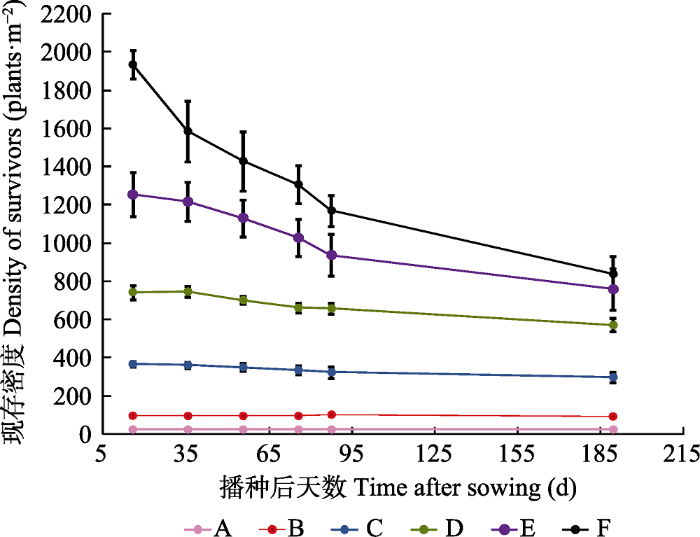 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT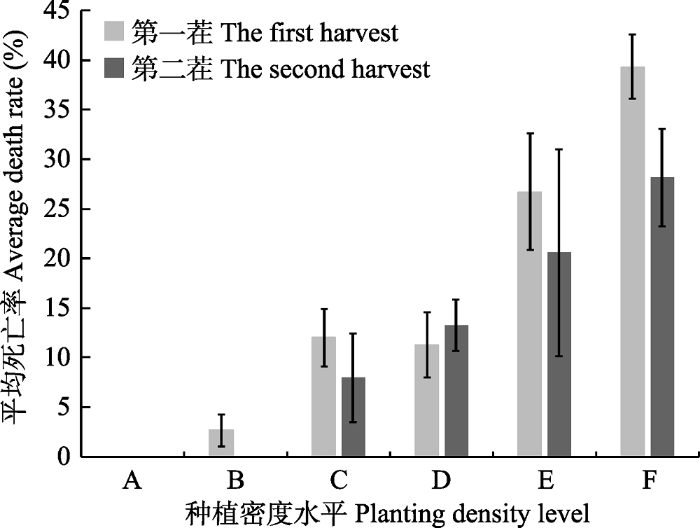 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT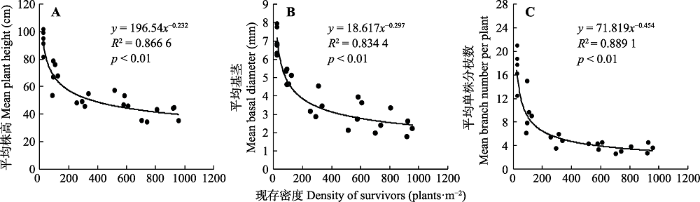 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT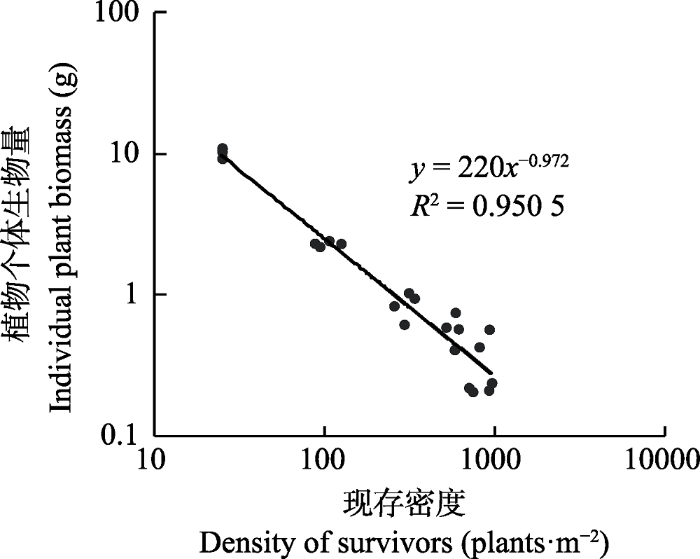 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT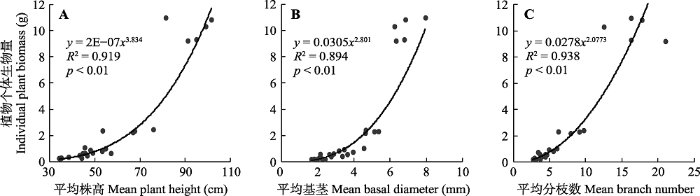 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}