彭书时,,1,*, 岳超2, 常锦峰31北京大学城市与环境学院, 北京大学地表过程分析与模拟教育部重点实验室, 北京 100871 2西北农林科技大学水土保持研究所, 陕西杨凌, 712100 3International Institute for Applied Systems Analysis (IIASA), A-2361 Laxenburg, Austria
Developments and applications of terrestrial biosphere model
Shu-Shi PENG,,1,*, Chao YUE2, Jin-Feng CHANG31College of Urban and Environmental Sciences, Key Laboratory for Earth Surface Processes of Ministry of Education, Peking University, Beijing 100871, China 2Institute of Soil & Water Conservation, Northwest A&F University, Yangling, Shannxi 712100, China 3International Institute for Applied Systems Analysis (IIASA), A-2361 Laxenburg, Austria
Abstract Exchanges of energy and matter between terrestrial biosphere and atmosphere and hydrosphere create critical feedbacks to Earth’s climates. To quantify how terrestrial ecosystems respond and feedback to global changes, terrestrial biosphere model (TBM) has been developed and applied in global change ecology during the past decades. In TBMs, myriad of biogeophysical, biogeochemical, hydrological cycles and dynamics processes on different spatial and temporal scales are represented. The TBMs have been applied on assessing and attributing past changes in terrestrial biosphere, and on predicting future changes and their feedbacks to climates. Here, we provide an overview of processes included in TBMs and TBMs applications on carbon and hydrological cycles, as well as their application on exploring human impacts on terrestrial ecosystems. Finally, we outline perspectives for future development and application of TBMs. Keywords:terrestrial biosphere model;terrestrial biosphere;ecosystem ecology;ecosystem research;global change;carbon cycle
土地利用和土地管理强度的变化影响生物地球化学循环并通过改变地表状态产生生物物理反馈, 历史时期土地利用变化贡献了约25%的人为碳排放(Le Quéré et al., 2018), 但对区域气候的影响仍有待进一步评估(Pitman et al., 2009)。未来随着人口增加, 土地资源将变得更加稀缺, 如何在不同利用类型间分配土地资源以及能否通过土地管理减缓气候变化, 这些都是应对气候变化需要回答的科学问题。国际上在CMIP6框架下发起了土地利用模型比较计划 (Land Use Model Intercomparison Project, LUMIP), 以推动不同陆地生物圈模型使用相同的驱动数据和模拟方法, 回答气候政策中与土地利用和管理密切相关的科学问题(Lawrence et al., 2016)。
多个融合了农作物模块的陆地生物圈模型(CLM-crop, LPJ-GUESS, LPJmL, ORCHIDEE-crop)深入参与了全球格点作物模型比较计划(The Global Gridded Crop Model Inter-comparison (GGCMI); Elliott et al., 2015)。该计划旨在评估现有作物模型所模拟的作物产量、环境影响等对气候变化、大气CO2含量升高、灌溉和施肥的响应, 分析不确定性来源, 并探讨模型发展方向(Muller et al., 2017), 为准确评估未来气候变化和人为管理下作物产量、减产风险和相关粮食安全等问题提供科学参考。
BastrikovV, MacBeanN, BacourC, SantarenD, KuppelS, PeylinP (2018). Land surface model parameter optimisation using in situ flux data: comparison of gradient-based versus random search algorithms (a case study using ORCHIDEE v1.9.5.2) Geoscientific Model Development, 11, 4739-4754. [本文引用: 1]
BlankeJ, Boke-OlénN, OlinS, ChangJF, SahlinU, LindeskogM, LehstenV (2018). Implications of accounting for management intensity on carbon and nitrogen balances of European grasslands PLOS ONE, 13, e0201058. DOI: 10.1371/journal.pone.0201058. URLPMID:30102732 [本文引用: 1]
BodmanRW, RaynerPJ, KarolyDJ (2013). Uncertainty in temperature projections reduced using carbon cycle and climate observations Nature Climate Change, 3, 725-729. [本文引用: 1]
BowringSPK, LauerwaldR, GuenetB, ZhuD, GuimberteauM, TootchiA, DucharneA, CiaisP (2019). ORCHIDEE MICT-LEAK (r5459), a global model for the production, transport, and transformation of Dissolved organic carbon from Arctic permafrost regions—Part 1: Rationale, model description, and simulation protocol Geoscientific Model Development, 12, 3503-3521. [本文引用: 1]
BrissonN, MaryB, RipocheD, JeuffroyMH, RugetF, NicoullaudB, GateP, Devienne-BarretF, AntoniolettiR, DurrC, RichardG, BeaudoinN, RecousS, TayotX, PlenetD, CellierP, MacHetJM, MeynardJM, DelécolleR (1998). STICS: a generic model for the simulation of crops and their water and nitrogen balances. I. Theory and parameterization applied to wheat and corn Agronomie, 18, 311-346. [本文引用: 1]
CarlsonKM, GerberJS, MuellerND, HerreroM, MacDonaldGK, BraumanKA, HavlikP, O’ConnellCS, JohnsonJA, SaatchiS, WestPC (2017). Greenhouse gas emissions intensity of global croplands Nature Climate Change, 7, 63-68. [本文引用: 1]
ChangJF, CiaisP, HerreroM, HavlikP, CampioliM, ZhangXZ, BaiYF, ViovyN, JoinerJ, WangXH, PengSS, YueC, PiaoSL, WangT, HauglustaineDA, SoussanaJF, PeregonA, KosykhN, Mironycheva-TokarevaN (2016). Combining livestock production information in a process-based vegetation model to reconstruct the history of grassland management Biogeosciences, 13, 3757-3776. DOI:10.5194/bg-13-3757-2016URL [本文引用: 1]
ChangJF, CiaisP, ViovyN, SoussanaJF, KlumppK, SultanB (2017). Future productivity and phenology changes in European grasslands for different warming levels: implications for grassland management and carbon balance Carbon Balance and Management, 12. DOI:10.1186/s13021-017-0079-8. [本文引用: 1]
ChangJF, CiaisP, ViovyN, VuichardN, SultanB, SoussanaJF (2015). The greenhouse gas balance of European grasslands Global Change Biology, 21, 3748-3761. URLPMID:26059550 [本文引用: 3]
ChangJF, ViovyN, VuichardN, CiaisP, WangT, CozicA, LardyR, GrauxAI, KlumppK, MartinR, SoussanaJF (2013). Incorporating grassland management in ORCHIDEE: model description and evaluation at 11 eddy-covariance sites in Europe Geoscientific Model Development, 6, 2165-2181. DOI:10.5194/gmd-6-2165-2013URL [本文引用: 1] This study describes how management of grasslands is included in the Organizing Carbon and Hydrology in Dynamic Ecosystems (ORCHIDEE) process-based ecosystem model designed for large-scale applications, and how management affects modeled grassland-atmosphere CO2 fluxes. The new model, ORCHIDEE-GM (grassland management) is enabled with a management module inspired from a grassland model (PaSim, version 5.0), with two grassland management practices being considered, cutting and grazing. The evaluation of the results from ORCHIDEE compared with those of ORCHIDEE-GM at 11 European sites, equipped with eddy covariance and biometric measurements, shows that ORCHIDEE-GM can realistically capture the cut-induced seasonal variation in biometric variables (LAI: leaf area index; AGB: aboveground biomass) and in CO2 fluxes (GPP: gross primary productivity; TER: total ecosystem respiration; and NEE: net ecosystem exchange). However, improvements at grazing sites are only marginal in ORCHIDEE-GM due to the difficulty in accounting for continuous grazing disturbance and its induced complex animal-vegetation interactions. Both NEE and GPP on monthly to annual timescales can be better simulated in ORCHIDEE-GM than in ORCHIDEE without management. For annual CO2 fluxes, the NEE bias and RMSE (root mean square error) in ORCHIDEE-GM are reduced by 53% and 20%, respectively, compared to ORCHIDEE. ORCHIDEE-GM is capable of modeling the net carbon balance (NBP) of managed temperate grasslands (37 +/- 30 gC m(-2) yr(-1) (P < 0.01) over the 11 sites) because the management module contains provisions to simulate the carbon fluxes of forage yield, herbage consumption, animal respiration and methane emissions.
DangalSRS, TianHQ, LuCQ, PanSF, PedersonN, HesslA (2016). Synergistic effects of climate change and grazing on net primary production of Mongolian grasslands Ecosphere, 7, e01274. DOI: 10.1002/ecs2.1274. [本文引用: 1]
de RosnayP, PolcherJ (1998). Modelling root water uptake in a complex land surface scheme coupled to a GCM Hydrology and Earth System Sciences, 2, 239-255. [本文引用: 1]
de RosnayP, PolcherJ, BruenM, LavalK (2002). Impact of a physically based soil water flow and soil-plant interaction representation for modeling large-scale land surface processes Journal of Geophysical Research, 107, 4118. DOI: 10.1029/2001JD000634. [本文引用: 1]
DucoudréNI, LavalK, PerrierA (1993). SECHIBA, a new set of parameterizations of the hydrologic exchanges at the land-atmosphere interface within the LMD atmospheric general circulation model Journal of Climate, 6, 248-273. [本文引用: 1]
ElliottJ, MüllerC, DeryngD, ChryssanthacopoulosJ, BooteKJ, BüchnerM, FosterI, GlotterM, HeinkeJ, IizumiT, IzaurraldeRC, MuellerND, RayDK, RosenzweigC, RuaneAC, SheffieldJ (2015). The Global Gridded Crop Model Intercomparison: data and modeling protocols for Phase 1 (v1.0) Geoscientific Model Development, 8, 261-277. [本文引用: 1]
FisherJB, HuntzingerDN, SchwalmCR, SitchS (2014). Modeling the terrestrial biosphere Annual Review of Environment and Resources, 39, 91-123. [本文引用: 5]
FleischerK, RammigA, de KauweMG, WalkerAP, DominguesTF, FuchsluegerL, GarciaS, GollDS, GrandisA, JiangMK, HaverdV, HofhanslF, HolmJA, KruijtB, LeungF, MedlynBE, MercadoLM, NorbyRJ, PakB, von RandowC, QuesadaCA, SchaapKJ, Valverde-BarrantesOJ, WangYP, YangXJ, ZaehleS, ZhuQ, LapolaDM (2019). Amazon forest response to CO2 fertilization dependent on plant phosphorus acquisition Nature Geoscience, 12, 736-741. [本文引用: 2]
FriedlingsteinP, CoxP, BettsR, BoppL, von BlohW, BrovkinV, CaduleP, DoneyS, EbyM, FungI, BalaG, JohnJ, JonesC, JoosF, KatoT, KawamiyaM, KnorrW, LindsayK, MatthewsHD, RaddatzT, RaynerP, ReickC, RoecknerE, SchnitzlerKG, SchnurR, StrassmannK, WeaverAJ, YoshikawaC, ZengN (2006). Climate-carbon cycle feedback analysis: results from the C4MIP model intercomparison Journal of Climate, 19, 3337-3353. [本文引用: 1]
FriedlingsteinP, MeinshausenM, AroraVK, JonesCD, AnavA, LiddicoatSK, KnuttiR (2014). Uncertainties in CMIP5 climate projections due to carbon cycle feedbacks Journal of Climate, 27, 511-526. [本文引用: 1]
FriendAD, StevensAK, KnoxRG, CannellMGR (1997). A process-based, terrestrial biosphere model of ecosystem dynamics (Hybrid v3.0) Ecological Modelling, 95, 249-287. [本文引用: 1]
GollDS, BrovkinV, ParidaBR, ReickCH, KattgeJ, ReichPB, van BodegomPM, Niinemetsü (2012). Nutrient limitation reduces land carbon uptake in simulations with a model of combined carbon, nitrogen and phosphorus cycling Biogeosciences, 9, 3547-3569. [本文引用: 1]
GollDS, VuichardN, MaignanF, Jornet-PuigA, SardansJ, VioletteA, PengSS, SunY, KvakicM, GuimberteauM, GuenetB, ZaehleS, PenuelasJ, JanssensI, CiaisP (2017). A representation of the phosphorus cycle for ORCHIDEE (revision 4520) Geoscientific Model Development, 10, 3745-3770. [本文引用: 2]
GrauxAI, GaurutM, AgabrielJ, BaumontR, DelagardeR, DelabyL, SoussanaJF (2011). Development of the Pasture Simulation Model for assessing livestock production under climate change Agriculture, Ecosystems & Environment, 144, 69-91. [本文引用: 1]
GuimberteauM, CiaisP, DucharneA, BoisierJP, Dutra AguiarAP, BiemansH, de DeurwaerderH, GalbraithD, KruijtB, LangerwischF, PovedaG, RammigA, RodriguezDA, TejadaG, ThonickeK, von RandowC, von RandowRCS, ZhangK, VerbeeckH (2017). Impacts of future deforestation and climate change on the hydrology of the Amazon Basin: a multi-model analysis with a new set of land-cover change scenarios Hydrology and Earth System Sciences, 21, 1455-1475. DOI:10.5194/hess-21-1455-2017URL [本文引用: 1]
GuimberteauM, ZhuD, MaignanF, HuangY, YueC, Dantec- NédélecS, OttléC, Jornet-PuigA, BastosA, LaurentP, GollD, BowringS, ChangJ, GuenetB, TifafiM, PengS, KrinnerG, DucharneA, WangF, WangT, WangX, WangY, YinZ, LauerwaldR, JoetzjerE, QiuC, KimH, CiaisP (2018). ORCHIDEE-MICT (v8.4.1), a land surface model for the high latitudes: model description and validation Geoscientific Model Development, 11, 121-163. [本文引用: 1]
HuntzingerDN, SchwalmC, MichalakAM, SchaeferK, KingAW, WeiY, JacobsonA, LiuS, CookRB, PostWM, BerthierG, HayesD, HuangM, ItoA, LeiH, LuC, MaoJ, PengCH, PengS, PoulterB, RiccuitoD, ShiX, TianH, WangW, ZengN, ZhaoF, ZhuQ (2013). The north American carbon program multi-scale synthesis and terrestrial model intercomparison project—Part 1: overview and experimental design Geoscientific Model Development, 6, 2121-2133. [本文引用: 3]
IPCC (2013). Climate Change 2013: the Physical Science Basis. Cambridge University Press, Cambridge, UK. [本文引用: 3]
KrinnerG, ViovyN, de Noblet-DucoudréN, OgéeJ, PolcherJ, FriedlingsteinP, CiaisP, SitchS, PrenticeIC (2005). A dynamic global vegetation model for studies of the coupled atmosphere-biosphere system Global Biogeochemical Cycles, 19, GB1015. DOI: 10.1029/2003gb002199. [本文引用: 1]
KuppelS, PeylinP, MaignanF, ChevallierF, KielyG, MontagnaniL, CescattiA (2014). Model-data fusion across ecosystems: from multisite optimizations to global simulations Geoscientific Model Development, 7, 2581-2597. [本文引用: 1]
LawrenceDM, HurttGC, ArnethA, BrovkinV, CalvinKV, JonesAD, JonesCD, LawrencePJ, de Noblet-DucoudréN, PongratzJ, SeneviratneSI, ShevliakovaE (2016). The Land Use Model Intercomparison Project (LUMIP) contribution to CMIP6: rationale and experimental design Geoscientific Model Development, 9, 2973-2998. [本文引用: 1]
LehmannJ, KleberM (2015). The contentious nature of soil organic matter Nature, 528, 60-68. DOI:10.1038/nature16069URLPMID:26595271 [本文引用: 1] The exchange of nutrients, energy and carbon between soil organic matter, the soil environment, aquatic systems and the atmosphere is important for agricultural productivity, water quality and climate. Long-standing theory suggests that soil organic matter is composed of inherently stable and chemically unique compounds. Here we argue that the available evidence does not support the formation of large-molecular-size and persistent 'humic substances' in soils. Instead, soil organic matter is a continuum of progressively decomposing organic compounds. We discuss implications of this view of the nature of soil organic matter for aquatic health, soil carbon-climate interactions and land management.
LiF, LawrenceDM (2017). Role of fire in the global land water budget during the twentieth century due to changing ecosystems Journal of Climate, 30, 1893-1908. DOI:10.1175/JCLI-D-16-0460.1URL [本文引用: 1]
McGuireAD, LawrenceDM, KovenC, CleinJS, BurkeE, ChenGS, JafarovE, MacDougallAH, MarchenkoS, NicolskyD, PengSS, RinkeA, CiaisP, GouttevinI, HayesDJ, JiDY, KrinnerG, MooreJC, RomanovskyV, Sch?delC, SchaeferK, SchuurEAG, ZhuangQL (2018). Dependence of the evolution of carbon dynamics in the northern permafrost region on the trajectory of climate change Proceedings of the National Academy of Sciences of the United States of America, 115, 3882-3887. URLPMID:29581283 [本文引用: 1]
NorbyRJ, WarrenJM, IversenCM, MedlynBE, McMurtrieRE (2010). CO2 enhancement of forest productivity constrained by limited nitrogen availability Proceedings of the National Academy of Sciences of the United States of America, 107, 19368-19373. URLPMID:20974944 [本文引用: 1]
PartonWJ, OjimaDS, ColeCV, SchimelDS (1994). A General Model for Soil Organic Matter Dynamics: Sensitivity to Litter Chemistry, Texture and Management. Quantitative Modeling of Soil Forming Processes. Soil Science Society of America, Madison, Wisconsin, USA. [本文引用: 1]
PiaoS, FriedlingsteinP, CiaisP, de Noblet-DucoudréN, LabatD, ZaehleS (2007). Changes in climate and land use have a larger direct impact than rising CO2 on global river runoff trends Proceedings of the National Academy of Sciences of the United States of America, 104, 15242-15247. DOI:10.1073/pnas.0707213104URLPMID:17878298 [本文引用: 1] The significant worldwide increase in observed river runoff has been tentatively attributed to the stomatal
PitmanAJ, de Noblet-DucoudréN, CruzFT, DavinEL, BonanGB, BrovkinV, ClaussenM, DelireC, GanzeveldL, GaylerV, van den HurkBJJM, LawrencePJ, van der MolenMK, MüllerC, ReickCH, SeneviratneSI, StrengersBJ, VoldoireA (2009). Uncertainties in climate responses to past land cover change: first results from the LUCID intercomparison study Geophysical Research Letters, 36, L14814. DOI: 10.1029/2009GL039076. DOI:10.1029/2009GL039076URL [本文引用: 1]
PrenticeIC, BondeauA, CramerW, HarrisonSP, HicklerT, LuchtW, SitchS, SmithB, SykesMT (2007). Dynamic Global Vegetation Modeling: Quantifying Terrestrial Ecosystem Responses to Large-scale Environmental Change Terrestrial Ecosystems in a Changing World. Springer, Berlin. [本文引用: 1]
PrenticeIC, CowlingSA (2013). Dynamic global vegetation models //Levin SA. Encyclopedia of Biodiversity. 2nd ed. Elsevier, Amsterdam. [本文引用: 3]
RabinSS, MeltonJR, LasslopG, BacheletD, ForrestM, HantsonS, KaplanJO, LiF, MangeonS, WardDS, YueC, AroraVK, HicklerT, KlosterS, KnorrW, NieradzikL, SpessaA, FolberthGA, SheehanT, VoulgarakisA, KelleyDI, PrenticeIC, SitchS, HarrisonS, ArnethA (2017). The Fire Modeling Intercomparison Project (FireMIP), phase 1: experimental and analytical protocols with detailed model descriptions Geoscientific Model Development, 10, 1175-1197. DOI:10.5194/gmd-10-1175-2017URL [本文引用: 1]
RiedoM, GrubA, RossetM, FuhrerJ (1998). A pasture simulation model for dry matter production, and fluxes of carbon, nitrogen, water and energy Ecological Modelling, 105, 141-183. DOI:10.1016/S0304-3800(97)00110-5URL [本文引用: 1]
RolinskiS, MüllerC, HeinkeJ, WeindlI, BiewaldA, BodirskyBL, BondeauA, Boons-PrinsER, BouwmanAF, LeffelaarPA, te RollerJA, SchaphoffS, ThonickeK (2018). Modeling vegetation and carbon dynamics of managed grasslands at the global scale with LPJmL 3.6 Geoscientific Model Development, 11, 429-451. DOI:10.5194/gmd-11-429-2018URL [本文引用: 2]
RothermelRC (1972). A Mathematical Model for Predicting Fire Spread in Wildland Fuels. USDA Forest Service General Technical Report INT-115. Intermountain Forest and Range Experiment Station, Forest Service, US Dept. of Agriculture, Ogden, Utah. [本文引用: 1]
RyderJ, PolcherJ, PeylinP, OttléC, ChenY, van GorselE, HaverdV, McGrathMJ, NaudtsK, OttoJ, ValadeA, LuyssaertS (2016). A multi-layer land surface energy budget model for implicit coupling with global atmospheric simulations Geoscientific Model Development, 9, 223-245. DOI:10.5194/gmd-9-223-2016URL [本文引用: 2]
SantiniM, CaporasoL (2018). Evaluation of freshwater flow from rivers to the sea in CMIP5 simulations: insights from the Congo river basin Journal of Geophysical Research, 123, 10278-10300. [本文引用: 1]
SitchS, SmithB, PrenticeIC, ArnethA, BondeauA, CramerW, KaplanJO, LevisS, LuchtW, SykesMT, ThonickeK, VenevskyS (2003). Evaluation of ecosystem dynamics, plant geography and terrestrial carbon cycling in the LPJ dynamic global vegetation model Global Change Biology, 9, 161-185. DOI:10.1046/j.1365-2486.2003.00569.xURL [本文引用: 2]
SokolNW, SandermanJ, BradfordMA (2019). Pathways of mineral-associated soil organic matter formation: integrating the role of plant carbon source, chemistry, and point of entry Global Change Biology, 25, 12-24. URLPMID:30338884 [本文引用: 1]
TangJY, RileyWJ (2017). SUPECA kinetics for scaling redox reactions in networks of mixed substrates and consumers and an example application to aerobic soil respiration Geoscientific Model Development, 10, 3277-3295. DOI:10.5194/gmd-10-3277-2017URL [本文引用: 1]
TerrerC, ViccaS, StockerBD, HungateBA, PhillipsRP, ReichPB, FinziAC, PrenticeIC (2018). Ecosystem responses to elevated CO2 governed by plant-soil interactions and the cost of nitrogen acquisition New Phytologist, 217, 507-522. URLPMID:29105765 [本文引用: 1]
VitousekPM, PorderS, HoultonBZ, ChadwickOA (2010). Terrestrial phosphorus limitation: mechanisms, implications, and nitrogen-phosphorus interactions Ecological Applications, 20, 5-15. DOI:10.1890/08-0127.1URLPMID:20349827 [本文引用: 1] Nutrient limitation to primary productivity and other biological processes is widespread in terrestrial ecosystems, and nitrogen (N) and phosphorus (P) are the most common limiting elements, both individually and in combination. Mechanisms that drive P limitation, and their interactions with the N cycle, have received less attention than mechanisms causing N limitation. We identify and discuss six mechanisms that could drive P limitation in terrestrial ecosystems. The best known of these is depletion-driven limitation, in which accumulated P losses during long-term soil and ecosystem development contribute to what Walker and Syers termed a
WattAS (1947). Pattern and process in the plant community Journal of Ecology, 35, 1-22. DOI:10.2307/2256497URL [本文引用: 1]
WiederWR, BonanGB, AllisonSD (2013). Global soil carbon projections are improved by modelling microbial processes Nature Climate Change, 3, 909-912. DOI:10.1038/NCLIMATE1951URL [本文引用: 1] Society relies on Earth system models (ESMs) to project future climate and carbon (C) cycle feedbacks. However, the soil C response to climate change is highly uncertain in these models(1,2) and they omit key biogeochemical mechanisms(3-5). Specifically, the traditional approach in ESMs lacks direct microbial control over soil C dynamics(6-8). Thus, we tested a new model that explicitly represents microbial mechanisms of soil C cycling on the global scale. Compared with traditional models, the microbial model simulates soil C pools that more closely match contemporary observations. It also projects a much wider range of soil C responses to climate change over the twenty-first century. Global soils accumulate C if microbial growth efficiency declines with warming in the microbial model. If growth efficiency adapts to warming, the microbial model projects large soil C losses. By comparison, traditional models project modest soil C losses with global warming. Microbes also change the soil response to increased C inputs, as might occur with CO2 or nutrient fertilization. In the microbial model, microbes consume these additional inputs; whereas in traditional models, additional inputs lead to C storage. Our results indicate that ESMs should simulate microbial physiology to more accurately project climate change feedbacks.
WiederWR, HartmanMD, SulmanBN, WangYP, KovenCD, BonanGB (2018). Carbon cycle confidence and uncertainty: exploring variation among soil biogeochemical models Global Change Biology, 24, 1563-1579. URLPMID:29120516 [本文引用: 1]
WuX, VuichardN, CiaisP, ViovyN, de Noblet-DucoudréN, WangX, MagliuloV, WattenbachM, VitaleL, di TommasiP, MoorsEJ, JansW, ElbersJ, CeschiaE, TallecT, BernhoferC, GrünwaldT, MoureauxC, ManiseT, LigneA, CellierP, LoubetB, LarmanouE, RipocheD (2016). ORCHIDEE-CROP (v0), a new process-based agro-land surface model: model description and evaluation over Europe Geoscientific Model Development, 9, 857-873. DOI:10.5194/gmd-9-857-2016URL [本文引用: 1]
YinXY, StruikPC, RomeroP, HarbinsonJ, EversJB, van der PuttenPEL, VosJ (2009). Using combined measurements of gas exchange and chlorophyll fluorescence to estimate parameters of a biochemical C3 photosynthesis model: a critical appraisal and a new integrated approach applied to leaves in a wheat (Triticum aestivum) canopy Plant, Cell & Environment, 32, 448-464. URLPMID:19183300 [本文引用: 1]
ZaehleS, JonesCD, HoultonB, LamarqueJF, RobertsonE (2015). Nitrogen availability reduces CMIP5 projections of twenty-first-century land carbon uptake Journal of Climate, 28, 2494-2511. DOI:10.1175/JCLI-D-13-00776.1URL [本文引用: 2]
ZaehleS, MedlynBE, de KauweMG, WalkerAP, DietzeMC, HicklerT, LuoYQ, WangYP, El-MasriB, ThorntonP, JainA, WangSS, WarlindD, WengES, PartonW, IversenCM, Gallet-BudynekA, McCarthyH, FinziA, HansonPJ, PrenticeIC, OrenR, NorbyRJ (2014). Evaluation of 11 terrestrial carbon-nitrogen cycle models against observations from two temperate Free-Air CO2 Enrichment studies New Phytologist, 202, 803-822. DOI:10.1111/nph.12697URLPMID:24467623 [本文引用: 1] We analysed the responses of 11 ecosystem models to elevated atmospheric [CO2 ] (eCO2 ) at two temperate forest ecosystems (Duke and Oak Ridge National Laboratory (ORNL) Free-Air CO2 Enrichment (FACE) experiments) to test alternative representations of carbon (C)-nitrogen (N) cycle processes. We decomposed the model responses into component processes affecting the response to eCO2 and confronted these with observations from the FACE experiments. Most of the models reproduced the observed initial enhancement of net primary production (NPP) at both sites, but none was able to simulate both the sustained 10-yr enhancement at Duke and the declining response at ORNL: models generally showed signs of progressive N limitation as a result of lower than observed plant N uptake. Nonetheless, many models showed qualitative agreement with observed component processes. The results suggest that improved representation of above-ground-below-ground interactions and better constraints on plant stoichiometry are important for a predictive understanding of eCO2 effects. Improved accuracy of soil organic matter inventories is pivotal to reduce uncertainty in the observed C-N budgets. The two FACE experiments are insufficient to fully constrain terrestrial responses to eCO2 , given the complexity of factors leading to the observed diverging trends, and the consequential inability of the models to explain these trends. Nevertheless, the ecosystem models were able to capture important features of the experiments, lending some support to their projections.
ZhuD, CiaisP, ChangJF, KrinnerG, PengSS, ViovyN, Pe?uelasJ, ZimovS (2018). The large mean body size of mammalian herbivores explains the productivity paradox during the Last Glacial Maximum Nature Ecology & Evolution, 2, 640-649. DOI:10.1038/s41559-018-0481-yURLPMID:29483680 [本文引用: 1] Large herbivores are a major agent in ecosystems, influencing vegetation structure, and carbon and nutrient flows. During the last glacial period, a mammoth steppe ecosystem prevailed in the unglaciated northern lands, supporting a high diversity and density of megafaunal herbivores. The apparent discrepancy between abundant megafauna and the expected low vegetation productivity under a generally harsher climate with a lower CO2 concentration, termed the productivity paradox, requires large-scale quantitative analysis using process-based ecosystem models. However, most of the current global dynamic vegetation models (DGVMs) lack explicit representation of large herbivores. Here we incorporated a grazing module in a DGVM based on physiological and demographic equations for wild large grazers, taking into account feedbacks of large grazers on vegetation. The model was applied globally for present-day and the Last Glacial Maximum (LGM). The present-day results of potential grazer biomass, combined with an empirical land-use map, infer a reduction in wild grazer biomass by 79-93% owing to anthropogenic land replacement of natural grasslands. For the LGM, we find that the larger mean body size of mammalian herbivores than today is the crucial clue to explain the productivity paradox, due to a more efficient exploitation of grass production by grazers with a large body size.
Land surface model parameter optimisation using in situ flux data: comparison of gradient-based versus random search algorithms (a case study using ORCHIDEE v1.9.5.2) 1 2018
... 随着观测数据的积累和大数据时代的来临, 如何利用不同时空尺度的观测数据提高模型的模拟精度也是当前的研究热点和难点(Prentice & Cowling, 2013; Medlyn et al., 2015).如何降低陆地生物圈模型的不确定性是科学界面临的巨大挑战.陆地生物圈模型的不确定性主要来源于模型结构、过程、参数、气候和土壤驱动数据.基于观测数据和数据模型融化算法优化模型参数(Kuppel et al., 2014; Bastrikov et al., 2018)可以显著改进陆地生物圈模型的模拟精度.全球越来越多从叶片到生态系统尺度的观测数据和全球变化控制实验等数据为模型参数优化和模型数据融合提供了基础(Prentice & Cowling, 2013).但目前观测数据和全球变化控制实验主要集中在温带和寒带, 降低陆地生物圈模型在热带模拟的不确定性需要加强热带的观测和模型数据融合研究(Fleischer et al., 2019). ...
Implications of accounting for management intensity on carbon and nitrogen balances of European grasslands 1 2018
... 融入了草地管理模块的陆地生物圈模型(DLEM, LPJ-GUESS, LPJmL, ORCHIDEE)已被用于定量评估全球及区域牧草生产潜力对气候变化、大气CO2含量升高、草地利用强度等因子的响应(Chang et al., 2015; Dangal et al., 2016; Rolinski et al., 2018), 模拟区域草地碳动态及温室气体排放(Chang et al., 2015, 2017; Blanke et al., 2018), 以及对草地管理模式和利用强度进行优化(Chang et al., 2015; Rolinski et al., 2018)和重建(Chang et al., 2016).ORCHIDEE-GM模型还被用于探讨末次冰期猛犸草原大型食草动物生物量动态及其影响下的草地碳平衡(Zhu et al., 2018). ...
Uncertainty in temperature projections reduced using carbon cycle and climate observations 1 2013
... 已有研究表明未来气候变化预测的不确定性有一半来自于陆地生物圈, 例如陆地生物圈碳循环对气候变化的响应与反馈(Friedlingstein et al., 2006, 2014; Bodman et al., 2013).由于地球气候对陆地生态系统的变化非常敏感, 从IPCC第一次评估报告到第五次评估报告, 陆地生物圈模型也逐步成为地球系统模式中的关键部分(IPCC, 2013).发展陆地生物圈模型和降低陆地生物圈模型的不确定性也被认为是准确预测未来地球气候变化的关键(Fisher et al., 2014).陆地生物圈模型不仅是生态系统生态学研究的重要工具, 也是地球系统模式中的重要模块.近20年陆地生物圈模型在国际地圈与生物圈计划(IGBP)、全球环境变化计划(GECP)等多个国际项目的推动下, 融合了生态学、环境科学和地球系统科学等多学科基础和交叉前沿, 迅速发展并形成了耦合多种过程的复杂模型.为了推动陆地生物圈模型的进一步发展和与生态学有关的应用, 本文将简要回顾陆地生物圈模型的发展历史, 对陆地生物圈模型的主要过程进行简介, 总结模型在生态系统生态学中的应用, 并展望模型未来的研究方向. ...
ORCHIDEE MICT-LEAK (r5459), a global model for the production, transport, and transformation of Dissolved organic carbon from Arctic permafrost regions—Part 1: Rationale, model description, and simulation protocol 1 2019
... 另外, ORCHIDEE模型针对冻土(Guimberteau et al., 2018), 泥炭地(Qiu et al., 2018), 有机和无机碳通过河流的横向输送等过程(Bowring et al., 2019)发展了不同的版本, 具体过程可以参考相应文章.不同陆地生物圈模型在所涵盖的过程和模拟机制上有所不同, 具体的模型过程可以参考附录I当前主要陆地生物圈模型的文献列表和Huntzinger等(2013)正文及其附录表1-4总结的模型过程具体算法. ...
Managing uncertainty in soil carbon feedbacks to climate change 1 2016
... 土壤团聚体等物理过程对土壤物理和碳氮磷循环过程有着重要影响(Lehmann & Kleber, 2015; Sokol et al., 2019), 但目前模型仍缺乏对土壤团聚体形成等物理过程的模拟(Bradford et al., 2016).土壤微生物对土壤有机碳的分解和形成至关重要.研究发现在陆地生物圈模型中耦合土壤微生物过程可能会降低陆地对气候变化的正反馈强度(Wieder et al., 2013), 但这一研究结果不确定性很大, 需要进一步研究.目前如何在陆地生物圈模型中模拟土壤物理过程和微生物过程及其对生态系统碳氮磷循环的影响是当前的研究热点和难点(Tang & Riley, 2017; Wieder et al., 2018). ...
STICS: a generic model for the simulation of crops and their water and nitrogen balances. I. Theory and parameterization applied to wheat and corn 1 1998
... 农田覆盖了超过11%的陆地表面.农田作为保障全球粮食安全的重要基础, 受到气候变化和土地利用变化的强烈影响.同时, 农田开发和利用也对气候系统产生了重要影响.农田N2O和甲烷排放以及泥炭地开发造成的CO2排放已占人为温室气体排放的4.5% (Carlson et al., 2017).另外, 农业活动灌溉消耗了全球近70%的总用水量, 极大程度地改变了水循环格局.ORCHIDEE-CROP模型版本(Wu et al., 2016)融入了农作物模型STICS (Brisson et al., 1998)的作物生长及管理模块, 考虑小麦、玉米、大豆、水稻等主要作物的发育阶段和物候等生理过程, 以及播种及收获日期、施肥、灌溉、收获指数等管理模式, 从而模拟不同管理模式下作物产量对气候变化的响应以及对农田碳水循环的影响. ...
Greenhouse gas emissions intensity of global croplands 1 2017
... 农田覆盖了超过11%的陆地表面.农田作为保障全球粮食安全的重要基础, 受到气候变化和土地利用变化的强烈影响.同时, 农田开发和利用也对气候系统产生了重要影响.农田N2O和甲烷排放以及泥炭地开发造成的CO2排放已占人为温室气体排放的4.5% (Carlson et al., 2017).另外, 农业活动灌溉消耗了全球近70%的总用水量, 极大程度地改变了水循环格局.ORCHIDEE-CROP模型版本(Wu et al., 2016)融入了农作物模型STICS (Brisson et al., 1998)的作物生长及管理模块, 考虑小麦、玉米、大豆、水稻等主要作物的发育阶段和物候等生理过程, 以及播种及收获日期、施肥、灌溉、收获指数等管理模式, 从而模拟不同管理模式下作物产量对气候变化的响应以及对农田碳水循环的影响. ...
Combining livestock production information in a process-based vegetation model to reconstruct the history of grassland management 1 2016
... 融入了草地管理模块的陆地生物圈模型(DLEM, LPJ-GUESS, LPJmL, ORCHIDEE)已被用于定量评估全球及区域牧草生产潜力对气候变化、大气CO2含量升高、草地利用强度等因子的响应(Chang et al., 2015; Dangal et al., 2016; Rolinski et al., 2018), 模拟区域草地碳动态及温室气体排放(Chang et al., 2015, 2017; Blanke et al., 2018), 以及对草地管理模式和利用强度进行优化(Chang et al., 2015; Rolinski et al., 2018)和重建(Chang et al., 2016).ORCHIDEE-GM模型还被用于探讨末次冰期猛犸草原大型食草动物生物量动态及其影响下的草地碳平衡(Zhu et al., 2018). ...
Future productivity and phenology changes in European grasslands for different warming levels: implications for grassland management and carbon balance 1 2017
... 融入了草地管理模块的陆地生物圈模型(DLEM, LPJ-GUESS, LPJmL, ORCHIDEE)已被用于定量评估全球及区域牧草生产潜力对气候变化、大气CO2含量升高、草地利用强度等因子的响应(Chang et al., 2015; Dangal et al., 2016; Rolinski et al., 2018), 模拟区域草地碳动态及温室气体排放(Chang et al., 2015, 2017; Blanke et al., 2018), 以及对草地管理模式和利用强度进行优化(Chang et al., 2015; Rolinski et al., 2018)和重建(Chang et al., 2016).ORCHIDEE-GM模型还被用于探讨末次冰期猛犸草原大型食草动物生物量动态及其影响下的草地碳平衡(Zhu et al., 2018). ...
The greenhouse gas balance of European grasslands 3 2015
... 融入了草地管理模块的陆地生物圈模型(DLEM, LPJ-GUESS, LPJmL, ORCHIDEE)已被用于定量评估全球及区域牧草生产潜力对气候变化、大气CO2含量升高、草地利用强度等因子的响应(Chang et al., 2015; Dangal et al., 2016; Rolinski et al., 2018), 模拟区域草地碳动态及温室气体排放(Chang et al., 2015, 2017; Blanke et al., 2018), 以及对草地管理模式和利用强度进行优化(Chang et al., 2015; Rolinski et al., 2018)和重建(Chang et al., 2016).ORCHIDEE-GM模型还被用于探讨末次冰期猛犸草原大型食草动物生物量动态及其影响下的草地碳平衡(Zhu et al., 2018). ...
... ), 模拟区域草地碳动态及温室气体排放(Chang et al., 2015, 2017; Blanke et al., 2018), 以及对草地管理模式和利用强度进行优化(Chang et al., 2015; Rolinski et al., 2018)和重建(Chang et al., 2016).ORCHIDEE-GM模型还被用于探讨末次冰期猛犸草原大型食草动物生物量动态及其影响下的草地碳平衡(Zhu et al., 2018). ...
... ), 以及对草地管理模式和利用强度进行优化(Chang et al., 2015; Rolinski et al., 2018)和重建(Chang et al., 2016).ORCHIDEE-GM模型还被用于探讨末次冰期猛犸草原大型食草动物生物量动态及其影响下的草地碳平衡(Zhu et al., 2018). ...
Incorporating grassland management in ORCHIDEE: model description and evaluation at 11 eddy-covariance sites in Europe 1 2013
... 全球37% (30%-47%)的陆地被草地覆盖, 其中很大一部分被用于进行畜牧业生产.ORCHIDEE- GM模型版本(Chang et al., 2013)融入了PaSim草地模型(Riedo et al., 1998; Graux et al., 2011)的管理模块, 用以模拟草料收割、放牧、施肥等过程, 并考虑这些过程对光合速率(通过施肥), 叶面积指数, 叶龄, 生物量(通过吃草、踩踏等), 生物量储存、分配及再生长等的影响.改进后的模型可以用于模拟气候和草地管理强度变化下区域及全球草地碳平衡. ...
Synergistic effects of climate change and grazing on net primary production of Mongolian grasslands 1 2016
... 融入了草地管理模块的陆地生物圈模型(DLEM, LPJ-GUESS, LPJmL, ORCHIDEE)已被用于定量评估全球及区域牧草生产潜力对气候变化、大气CO2含量升高、草地利用强度等因子的响应(Chang et al., 2015; Dangal et al., 2016; Rolinski et al., 2018), 模拟区域草地碳动态及温室气体排放(Chang et al., 2015, 2017; Blanke et al., 2018), 以及对草地管理模式和利用强度进行优化(Chang et al., 2015; Rolinski et al., 2018)和重建(Chang et al., 2016).ORCHIDEE-GM模型还被用于探讨末次冰期猛犸草原大型食草动物生物量动态及其影响下的草地碳平衡(Zhu et al., 2018). ...
Modelling root water uptake in a complex land surface scheme coupled to a GCM 1 1998
... ORCHIDEE模型的生物地球物理过程主要包括地表能量平衡和土壤热传导.地表能量平衡模块根据输入的向下短波和长波辐射、风速、空气温湿度以及地表状态计算出向上短波和长波辐射、感热和潜热通量, 以及土壤热通量(Ducoudré et al., 1993; de Rosnay & Polcher, 1998).为了计算土壤热传导和土壤温度, 用户可以设置土壤热传导深度和分层厚度; 根据热传导偏微分方程和其傅里叶求解方法, 土壤热传导深度一般设置较深, 默认土壤热传导深度为90 m, 按照指数增加的间距分为46层.其中每层土壤的热传导率和热容量与土壤质地和土壤含水量有关, 每30 min计算出每层土壤的热通量和土壤温度.用户可以设置土壤热传导底部的地热通量和是否考虑深层基岩热通量. ...
Impact of a physically based soil water flow and soil-plant interaction representation for modeling large-scale land surface processes 1 2002
... 地表水分平衡包括了由穿透雨和积雪融水形成的输入以及由入渗和地表径流形成的输出, 当水分输入大于入渗时, 就会形成地表径流.在个别坡度较低区域, 地表径流会再次向土壤入渗(再入渗过程), 构成地表水输入的第三个来源.在每个格点内, 土壤水库按照植被类型分为相互独立的三个部分: 木本、草本和裸地, 即水分平衡在每部分独立核算.默认土壤深度为2 m, 按照指数增加的间距分为11层.地表蒸发仅仅发生在最上层土壤, 该层以下的土层湿度变化取决于垂向水分交换和蒸腾量.土层间垂向水分交换由描述一维非饱和土壤水分运动的Fokker-Planck方程进行模拟(de Rosney et al., 2002), 不同格点间的水分横向运动予以忽略, 后者在大尺度上应用模型时是适当的.根据不同土层的根系分布状况和土壤湿度, 将植被蒸腾消耗的水量分配至各土壤层(Seidl et al., 2017).最下层土壤水分饱和时会形成重力排水, 重力排水和地表径流一起, 构成河川径流. ...
SECHIBA, a new set of parameterizations of the hydrologic exchanges at the land-atmosphere interface within the LMD atmospheric general circulation model 1 1993
... ORCHIDEE模型的生物地球物理过程主要包括地表能量平衡和土壤热传导.地表能量平衡模块根据输入的向下短波和长波辐射、风速、空气温湿度以及地表状态计算出向上短波和长波辐射、感热和潜热通量, 以及土壤热通量(Ducoudré et al., 1993; de Rosnay & Polcher, 1998).为了计算土壤热传导和土壤温度, 用户可以设置土壤热传导深度和分层厚度; 根据热传导偏微分方程和其傅里叶求解方法, 土壤热传导深度一般设置较深, 默认土壤热传导深度为90 m, 按照指数增加的间距分为46层.其中每层土壤的热传导率和热容量与土壤质地和土壤含水量有关, 每30 min计算出每层土壤的热通量和土壤温度.用户可以设置土壤热传导底部的地热通量和是否考虑深层基岩热通量. ...
The Global Gridded Crop Model Intercomparison: data and modeling protocols for Phase 1 (v1.0) 1 2015
... 多个融合了农作物模块的陆地生物圈模型(CLM-crop, LPJ-GUESS, LPJmL, ORCHIDEE-crop)深入参与了全球格点作物模型比较计划(The Global Gridded Crop Model Inter-comparison (GGCMI); Elliott et al., 2015).该计划旨在评估现有作物模型所模拟的作物产量、环境影响等对气候变化、大气CO2含量升高、灌溉和施肥的响应, 分析不确定性来源, 并探讨模型发展方向(Muller et al., 2017), 为准确评估未来气候变化和人为管理下作物产量、减产风险和相关粮食安全等问题提供科学参考. ...
Modeling the terrestrial biosphere 5 2014
... 已有研究表明未来气候变化预测的不确定性有一半来自于陆地生物圈, 例如陆地生物圈碳循环对气候变化的响应与反馈(Friedlingstein et al., 2006, 2014; Bodman et al., 2013).由于地球气候对陆地生态系统的变化非常敏感, 从IPCC第一次评估报告到第五次评估报告, 陆地生物圈模型也逐步成为地球系统模式中的关键部分(IPCC, 2013).发展陆地生物圈模型和降低陆地生物圈模型的不确定性也被认为是准确预测未来地球气候变化的关键(Fisher et al., 2014).陆地生物圈模型不仅是生态系统生态学研究的重要工具, 也是地球系统模式中的重要模块.近20年陆地生物圈模型在国际地圈与生物圈计划(IGBP)、全球环境变化计划(GECP)等多个国际项目的推动下, 融合了生态学、环境科学和地球系统科学等多学科基础和交叉前沿, 迅速发展并形成了耦合多种过程的复杂模型.为了推动陆地生物圈模型的进一步发展和与生态学有关的应用, 本文将简要回顾陆地生物圈模型的发展历史, 对陆地生物圈模型的主要过程进行简介, 总结模型在生态系统生态学中的应用, 并展望模型未来的研究方向. ...
... 自1849年亚历山大·冯·洪堡(Alexander von Humboldt)首次建立气候与植物地理关系后, 经奧古斯特·格里瑟巴赫(August Grisebach)和弗拉迪米尔·彼得·柯本(Wladimir Peter K?ppen)细化完善后, 形成了植物地理预测模型的雏形.植物地理模型能够描述气候平衡态植被类型的静态空间分布, 但不能模拟碳氮水循环, 也不能模拟能量等生物物理过程.与大尺度植物地理模型相反, 描述小尺度植被动态的模型主要关注植物个体生长和死亡, 对光、水和营养元素的竞争和群落演替等过程, 这些小尺度模型逐渐模拟了小尺度上森林植被动态(Watt, 1947), 例如HYBRID和ED模型(Friend et al., 1997; Moorcroft et al., 2001).这些小尺度植被动态模型一般不能保证物质或能量守恒, 随后发展的生物地球化学循环模型建立在物质和能量守恒的基础上, 研究碳氮水在生态系统中的循环过程, 例如CENTURY模型(Parton et al., 1994).同时, 早期为了模拟陆地与大气边界层之间能量、水汽和动量过程, 与气候和大气环流模型(GCMs)相耦合, 陆面模式(LSMs)基于生物物理模型的基础应运而生, 例如SiB模型(Sellers et al., 1997).早期的陆面模式侧重陆地生态系统生物物理过程的模拟, 缺乏植被动态和生物地球化学循环过程.关于这些不同类型的模型定义和区分可进一步参考Fisher等(2014)文中的表1. ...
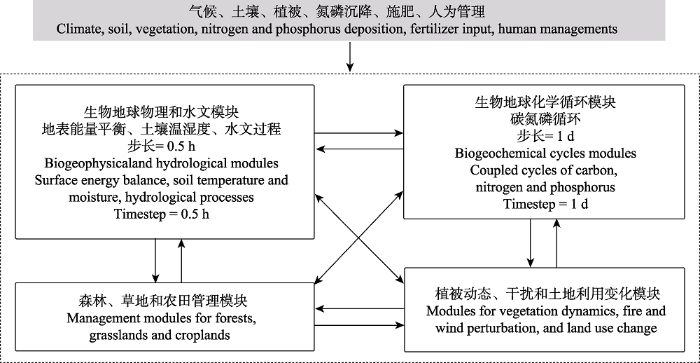
... 虽然不同的陆地生物圈模型对不同过程的参数化方案或算法有所不同, 但基本框架和结构是类似的(Huntzinger et al., 2013; Fisher et al., 2014).本节以法国的陆地生物圈模型ORCHIDEE为例, 简要介绍模型中生物地球物理过程, 水文过程, 生物地球化学循环, 植被动态、干扰和土地利用模块, 森林、草地和农田管理模块(图1).ORCHIDEE默认版本(trunk)及不同开发版本(branch)的代码下载和使用说明文档请见该模型网站http://forge.ipsl.jussieu.fr/orchidee/wiki. ...
... 地球陆地表层42%-68%的面积受人类活动影响, 人类对森林、草地和农田的经营管理, 如人工林管理、木材采集、放牧、割草、施肥、灌溉等措施直接影响了生态系统结构和功能.另外, 迁徙耕作、耕地过度开垦、过度放牧、泥炭地开发(包括火干扰)以及气候变化(全球变暖、降水强度变化、干旱等)可能造成土地退化(水蚀、风蚀、营养流失、质地变化)并对生态系统碳循环产生显著影响.随着人类对生态系统的影响日益加剧, 如何在陆地生物圈模型中模拟人类活动也是当前的研究热点和难点(Fisher et al., 2014; Naudts et al., 2016). ...
Amazon forest response to CO2 fertilization dependent on plant phosphorus acquisition 2 2019
... 研究发现大气CO2浓度上升对植被生长的施肥效应会受到氮磷营养元素的限制(Norby et al., 2010; Vitousek et al., 2010; Terrer et al., 2018), 从而可能限制未来陆地碳汇强度, 同时增加模型对未来陆地生态系统碳平衡评估的不确定性(Goll et al., 2012; Zaehle et al., 2015).多个陆地生物圈模型在已有碳循环的基础上耦合了氮循环过程, 但已有的模型对碳氮耦合模拟的不确定性仍然很大, 改进模型碳氮耦合是模型研究的挑战(Zaehle et al., 2014).磷循环也会影响植被生长等碳循环过程, 在陆地生物圈模型中耦合磷循环也是目前模型的研究热点(Goll et al., 2017; Fleischer et al., 2019). ...
... 随着观测数据的积累和大数据时代的来临, 如何利用不同时空尺度的观测数据提高模型的模拟精度也是当前的研究热点和难点(Prentice & Cowling, 2013; Medlyn et al., 2015).如何降低陆地生物圈模型的不确定性是科学界面临的巨大挑战.陆地生物圈模型的不确定性主要来源于模型结构、过程、参数、气候和土壤驱动数据.基于观测数据和数据模型融化算法优化模型参数(Kuppel et al., 2014; Bastrikov et al., 2018)可以显著改进陆地生物圈模型的模拟精度.全球越来越多从叶片到生态系统尺度的观测数据和全球变化控制实验等数据为模型参数优化和模型数据融合提供了基础(Prentice & Cowling, 2013).但目前观测数据和全球变化控制实验主要集中在温带和寒带, 降低陆地生物圈模型在热带模拟的不确定性需要加强热带的观测和模型数据融合研究(Fleischer et al., 2019). ...
Climate-carbon cycle feedback analysis: results from the C4MIP model intercomparison 1 2006
... 已有研究表明未来气候变化预测的不确定性有一半来自于陆地生物圈, 例如陆地生物圈碳循环对气候变化的响应与反馈(Friedlingstein et al., 2006, 2014; Bodman et al., 2013).由于地球气候对陆地生态系统的变化非常敏感, 从IPCC第一次评估报告到第五次评估报告, 陆地生物圈模型也逐步成为地球系统模式中的关键部分(IPCC, 2013).发展陆地生物圈模型和降低陆地生物圈模型的不确定性也被认为是准确预测未来地球气候变化的关键(Fisher et al., 2014).陆地生物圈模型不仅是生态系统生态学研究的重要工具, 也是地球系统模式中的重要模块.近20年陆地生物圈模型在国际地圈与生物圈计划(IGBP)、全球环境变化计划(GECP)等多个国际项目的推动下, 融合了生态学、环境科学和地球系统科学等多学科基础和交叉前沿, 迅速发展并形成了耦合多种过程的复杂模型.为了推动陆地生物圈模型的进一步发展和与生态学有关的应用, 本文将简要回顾陆地生物圈模型的发展历史, 对陆地生物圈模型的主要过程进行简介, 总结模型在生态系统生态学中的应用, 并展望模型未来的研究方向. ...
Uncertainties in CMIP5 climate projections due to carbon cycle feedbacks 1 2014
... 已有研究表明未来气候变化预测的不确定性有一半来自于陆地生物圈, 例如陆地生物圈碳循环对气候变化的响应与反馈(Friedlingstein et al., 2006, 2014; Bodman et al., 2013).由于地球气候对陆地生态系统的变化非常敏感, 从IPCC第一次评估报告到第五次评估报告, 陆地生物圈模型也逐步成为地球系统模式中的关键部分(IPCC, 2013).发展陆地生物圈模型和降低陆地生物圈模型的不确定性也被认为是准确预测未来地球气候变化的关键(Fisher et al., 2014).陆地生物圈模型不仅是生态系统生态学研究的重要工具, 也是地球系统模式中的重要模块.近20年陆地生物圈模型在国际地圈与生物圈计划(IGBP)、全球环境变化计划(GECP)等多个国际项目的推动下, 融合了生态学、环境科学和地球系统科学等多学科基础和交叉前沿, 迅速发展并形成了耦合多种过程的复杂模型.为了推动陆地生物圈模型的进一步发展和与生态学有关的应用, 本文将简要回顾陆地生物圈模型的发展历史, 对陆地生物圈模型的主要过程进行简介, 总结模型在生态系统生态学中的应用, 并展望模型未来的研究方向. ...
A process-based, terrestrial biosphere model of ecosystem dynamics (Hybrid v3.0) 1 1997
... 自1849年亚历山大·冯·洪堡(Alexander von Humboldt)首次建立气候与植物地理关系后, 经奧古斯特·格里瑟巴赫(August Grisebach)和弗拉迪米尔·彼得·柯本(Wladimir Peter K?ppen)细化完善后, 形成了植物地理预测模型的雏形.植物地理模型能够描述气候平衡态植被类型的静态空间分布, 但不能模拟碳氮水循环, 也不能模拟能量等生物物理过程.与大尺度植物地理模型相反, 描述小尺度植被动态的模型主要关注植物个体生长和死亡, 对光、水和营养元素的竞争和群落演替等过程, 这些小尺度模型逐渐模拟了小尺度上森林植被动态(Watt, 1947), 例如HYBRID和ED模型(Friend et al., 1997; Moorcroft et al., 2001).这些小尺度植被动态模型一般不能保证物质或能量守恒, 随后发展的生物地球化学循环模型建立在物质和能量守恒的基础上, 研究碳氮水在生态系统中的循环过程, 例如CENTURY模型(Parton et al., 1994).同时, 早期为了模拟陆地与大气边界层之间能量、水汽和动量过程, 与气候和大气环流模型(GCMs)相耦合, 陆面模式(LSMs)基于生物物理模型的基础应运而生, 例如SiB模型(Sellers et al., 1997).早期的陆面模式侧重陆地生态系统生物物理过程的模拟, 缺乏植被动态和生物地球化学循环过程.关于这些不同类型的模型定义和区分可进一步参考Fisher等(2014)文中的表1. ...
Nutrient limitation reduces land carbon uptake in simulations with a model of combined carbon, nitrogen and phosphorus cycling 1 2012
... 研究发现大气CO2浓度上升对植被生长的施肥效应会受到氮磷营养元素的限制(Norby et al., 2010; Vitousek et al., 2010; Terrer et al., 2018), 从而可能限制未来陆地碳汇强度, 同时增加模型对未来陆地生态系统碳平衡评估的不确定性(Goll et al., 2012; Zaehle et al., 2015).多个陆地生物圈模型在已有碳循环的基础上耦合了氮循环过程, 但已有的模型对碳氮耦合模拟的不确定性仍然很大, 改进模型碳氮耦合是模型研究的挑战(Zaehle et al., 2014).磷循环也会影响植被生长等碳循环过程, 在陆地生物圈模型中耦合磷循环也是目前模型的研究热点(Goll et al., 2017; Fleischer et al., 2019). ...
A representation of the phosphorus cycle for ORCHIDEE (revision 4520) 2 2017
... 研究发现大气CO2浓度上升对植被生长的施肥效应会受到氮磷营养元素的限制(Norby et al., 2010; Vitousek et al., 2010; Terrer et al., 2018), 从而可能限制未来陆地碳汇强度, 同时增加模型对未来陆地生态系统碳平衡评估的不确定性(Goll et al., 2012; Zaehle et al., 2015).多个陆地生物圈模型在已有碳循环的基础上耦合了氮循环过程, 但已有的模型对碳氮耦合模拟的不确定性仍然很大, 改进模型碳氮耦合是模型研究的挑战(Zaehle et al., 2014).磷循环也会影响植被生长等碳循环过程, 在陆地生物圈模型中耦合磷循环也是目前模型的研究热点(Goll et al., 2017; Fleischer et al., 2019). ...
Development of the Pasture Simulation Model for assessing livestock production under climate change 1 2011
... 全球37% (30%-47%)的陆地被草地覆盖, 其中很大一部分被用于进行畜牧业生产.ORCHIDEE- GM模型版本(Chang et al., 2013)融入了PaSim草地模型(Riedo et al., 1998; Graux et al., 2011)的管理模块, 用以模拟草料收割、放牧、施肥等过程, 并考虑这些过程对光合速率(通过施肥), 叶面积指数, 叶龄, 生物量(通过吃草、踩踏等), 生物量储存、分配及再生长等的影响.改进后的模型可以用于模拟气候和草地管理强度变化下区域及全球草地碳平衡. ...
Impacts of future deforestation and climate change on the hydrology of the Amazon Basin: a multi-model analysis with a new set of land-cover change scenarios 1 2017
Using ecosystem experiments to improve vegetation models 1 2015
... 随着观测数据的积累和大数据时代的来临, 如何利用不同时空尺度的观测数据提高模型的模拟精度也是当前的研究热点和难点(Prentice & Cowling, 2013; Medlyn et al., 2015).如何降低陆地生物圈模型的不确定性是科学界面临的巨大挑战.陆地生物圈模型的不确定性主要来源于模型结构、过程、参数、气候和土壤驱动数据.基于观测数据和数据模型融化算法优化模型参数(Kuppel et al., 2014; Bastrikov et al., 2018)可以显著改进陆地生物圈模型的模拟精度.全球越来越多从叶片到生态系统尺度的观测数据和全球变化控制实验等数据为模型参数优化和模型数据融合提供了基础(Prentice & Cowling, 2013).但目前观测数据和全球变化控制实验主要集中在温带和寒带, 降低陆地生物圈模型在热带模拟的不确定性需要加强热带的观测和模型数据融合研究(Fleischer et al., 2019). ...
A method for scaling vegetation dynamics: the ecosystem demography model (ED) 1 2001
... 自1849年亚历山大·冯·洪堡(Alexander von Humboldt)首次建立气候与植物地理关系后, 经奧古斯特·格里瑟巴赫(August Grisebach)和弗拉迪米尔·彼得·柯本(Wladimir Peter K?ppen)细化完善后, 形成了植物地理预测模型的雏形.植物地理模型能够描述气候平衡态植被类型的静态空间分布, 但不能模拟碳氮水循环, 也不能模拟能量等生物物理过程.与大尺度植物地理模型相反, 描述小尺度植被动态的模型主要关注植物个体生长和死亡, 对光、水和营养元素的竞争和群落演替等过程, 这些小尺度模型逐渐模拟了小尺度上森林植被动态(Watt, 1947), 例如HYBRID和ED模型(Friend et al., 1997; Moorcroft et al., 2001).这些小尺度植被动态模型一般不能保证物质或能量守恒, 随后发展的生物地球化学循环模型建立在物质和能量守恒的基础上, 研究碳氮水在生态系统中的循环过程, 例如CENTURY模型(Parton et al., 1994).同时, 早期为了模拟陆地与大气边界层之间能量、水汽和动量过程, 与气候和大气环流模型(GCMs)相耦合, 陆面模式(LSMs)基于生物物理模型的基础应运而生, 例如SiB模型(Sellers et al., 1997).早期的陆面模式侧重陆地生态系统生物物理过程的模拟, 缺乏植被动态和生物地球化学循环过程.关于这些不同类型的模型定义和区分可进一步参考Fisher等(2014)文中的表1. ...
Strategies for feeding the world more sustainably with organic agriculture 1 2017
... 多个融合了农作物模块的陆地生物圈模型(CLM-crop, LPJ-GUESS, LPJmL, ORCHIDEE-crop)深入参与了全球格点作物模型比较计划(The Global Gridded Crop Model Inter-comparison (GGCMI); Elliott et al., 2015).该计划旨在评估现有作物模型所模拟的作物产量、环境影响等对气候变化、大气CO2含量升高、灌溉和施肥的响应, 分析不确定性来源, 并探讨模型发展方向(Muller et al., 2017), 为准确评估未来气候变化和人为管理下作物产量、减产风险和相关粮食安全等问题提供科学参考. ...
Europes forest management did not mitigate climate warming 1 2016
... 地球陆地表层42%-68%的面积受人类活动影响, 人类对森林、草地和农田的经营管理, 如人工林管理、木材采集、放牧、割草、施肥、灌溉等措施直接影响了生态系统结构和功能.另外, 迁徙耕作、耕地过度开垦、过度放牧、泥炭地开发(包括火干扰)以及气候变化(全球变暖、降水强度变化、干旱等)可能造成土地退化(水蚀、风蚀、营养流失、质地变化)并对生态系统碳循环产生显著影响.随着人类对生态系统的影响日益加剧, 如何在陆地生物圈模型中模拟人类活动也是当前的研究热点和难点(Fisher et al., 2014; Naudts et al., 2016). ...
CO2 enhancement of forest productivity constrained by limited nitrogen availability 1 2010
... 研究发现大气CO2浓度上升对植被生长的施肥效应会受到氮磷营养元素的限制(Norby et al., 2010; Vitousek et al., 2010; Terrer et al., 2018), 从而可能限制未来陆地碳汇强度, 同时增加模型对未来陆地生态系统碳平衡评估的不确定性(Goll et al., 2012; Zaehle et al., 2015).多个陆地生物圈模型在已有碳循环的基础上耦合了氮循环过程, 但已有的模型对碳氮耦合模拟的不确定性仍然很大, 改进模型碳氮耦合是模型研究的挑战(Zaehle et al., 2014).磷循环也会影响植被生长等碳循环过程, 在陆地生物圈模型中耦合磷循环也是目前模型的研究热点(Goll et al., 2017; Fleischer et al., 2019). ...
A General Model for Soil Organic Matter Dynamics: Sensitivity to Litter Chemistry, Texture and Management. Quantitative Modeling of Soil Forming Processes. 1 1994
... 自1849年亚历山大·冯·洪堡(Alexander von Humboldt)首次建立气候与植物地理关系后, 经奧古斯特·格里瑟巴赫(August Grisebach)和弗拉迪米尔·彼得·柯本(Wladimir Peter K?ppen)细化完善后, 形成了植物地理预测模型的雏形.植物地理模型能够描述气候平衡态植被类型的静态空间分布, 但不能模拟碳氮水循环, 也不能模拟能量等生物物理过程.与大尺度植物地理模型相反, 描述小尺度植被动态的模型主要关注植物个体生长和死亡, 对光、水和营养元素的竞争和群落演替等过程, 这些小尺度模型逐渐模拟了小尺度上森林植被动态(Watt, 1947), 例如HYBRID和ED模型(Friend et al., 1997; Moorcroft et al., 2001).这些小尺度植被动态模型一般不能保证物质或能量守恒, 随后发展的生物地球化学循环模型建立在物质和能量守恒的基础上, 研究碳氮水在生态系统中的循环过程, 例如CENTURY模型(Parton et al., 1994).同时, 早期为了模拟陆地与大气边界层之间能量、水汽和动量过程, 与气候和大气环流模型(GCMs)相耦合, 陆面模式(LSMs)基于生物物理模型的基础应运而生, 例如SiB模型(Sellers et al., 1997).早期的陆面模式侧重陆地生态系统生物物理过程的模拟, 缺乏植被动态和生物地球化学循环过程.关于这些不同类型的模型定义和区分可进一步参考Fisher等(2014)文中的表1. ...
Changes in climate and land use have a larger direct impact than rising CO2 on global river runoff trends 1 2007
... 随着观测数据的积累和大数据时代的来临, 如何利用不同时空尺度的观测数据提高模型的模拟精度也是当前的研究热点和难点(Prentice & Cowling, 2013; Medlyn et al., 2015).如何降低陆地生物圈模型的不确定性是科学界面临的巨大挑战.陆地生物圈模型的不确定性主要来源于模型结构、过程、参数、气候和土壤驱动数据.基于观测数据和数据模型融化算法优化模型参数(Kuppel et al., 2014; Bastrikov et al., 2018)可以显著改进陆地生物圈模型的模拟精度.全球越来越多从叶片到生态系统尺度的观测数据和全球变化控制实验等数据为模型参数优化和模型数据融合提供了基础(Prentice & Cowling, 2013).但目前观测数据和全球变化控制实验主要集中在温带和寒带, 降低陆地生物圈模型在热带模拟的不确定性需要加强热带的观测和模型数据融合研究(Fleischer et al., 2019). ...
... )可以显著改进陆地生物圈模型的模拟精度.全球越来越多从叶片到生态系统尺度的观测数据和全球变化控制实验等数据为模型参数优化和模型数据融合提供了基础(Prentice & Cowling, 2013).但目前观测数据和全球变化控制实验主要集中在温带和寒带, 降低陆地生物圈模型在热带模拟的不确定性需要加强热带的观测和模型数据融合研究(Fleischer et al., 2019). ...
ORCHIDEE-PEAT (revision 4596), a model for northern peatland CO2, water, and energy fluxes on daily to annual scales 1 2018
... 另外, ORCHIDEE模型针对冻土(Guimberteau et al., 2018), 泥炭地(Qiu et al., 2018), 有机和无机碳通过河流的横向输送等过程(Bowring et al., 2019)发展了不同的版本, 具体过程可以参考相应文章.不同陆地生物圈模型在所涵盖的过程和模拟机制上有所不同, 具体的模型过程可以参考附录I当前主要陆地生物圈模型的文献列表和Huntzinger等(2013)正文及其附录表1-4总结的模型过程具体算法. ...
The Fire Modeling Intercomparison Project (FireMIP), phase 1: experimental and analytical protocols with detailed model descriptions 1 2017
... 干扰(火灾、风灾、虫灾、雪灾、干旱、病原性灾害等)是森林生态系统的固有组成部分, 对森林生态系统造成破坏性影响, 可以改变森林结构、物种组成并影响其生态系统服务功能.全球变化既可以通过气候变化影响干扰发生的频率、范围和强度, 又可以通过影响植被组成间接影响干扰的发生(Seidl et al., 2017).然而, 干扰在陆地生物圈模型中的表征仍不完善.目前多数模型包括了基于机制的火灾模块(Rabin et al., 2017), 然而对于其他形式干扰的模拟仍普遍欠缺.模拟某些自然灾害(如风灾)、以及模拟多重干扰的相互作用需要解析森林冠层结构, 以表征灾害过后冠层结构变化对次生灾害发生的影响.可以预见, 不断完善对森林干扰的模拟将是下一阶段模型发展的一个重点方向. ...
A pasture simulation model for dry matter production, and fluxes of carbon, nitrogen, water and energy 1 1998
... 全球37% (30%-47%)的陆地被草地覆盖, 其中很大一部分被用于进行畜牧业生产.ORCHIDEE- GM模型版本(Chang et al., 2013)融入了PaSim草地模型(Riedo et al., 1998; Graux et al., 2011)的管理模块, 用以模拟草料收割、放牧、施肥等过程, 并考虑这些过程对光合速率(通过施肥), 叶面积指数, 叶龄, 生物量(通过吃草、踩踏等), 生物量储存、分配及再生长等的影响.改进后的模型可以用于模拟气候和草地管理强度变化下区域及全球草地碳平衡. ...
Modeling vegetation and carbon dynamics of managed grasslands at the global scale with LPJmL 3.6 2 2018
... 融入了草地管理模块的陆地生物圈模型(DLEM, LPJ-GUESS, LPJmL, ORCHIDEE)已被用于定量评估全球及区域牧草生产潜力对气候变化、大气CO2含量升高、草地利用强度等因子的响应(Chang et al., 2015; Dangal et al., 2016; Rolinski et al., 2018), 模拟区域草地碳动态及温室气体排放(Chang et al., 2015, 2017; Blanke et al., 2018), 以及对草地管理模式和利用强度进行优化(Chang et al., 2015; Rolinski et al., 2018)和重建(Chang et al., 2016).ORCHIDEE-GM模型还被用于探讨末次冰期猛犸草原大型食草动物生物量动态及其影响下的草地碳平衡(Zhu et al., 2018). ...
... ; Rolinski et al., 2018)和重建(Chang et al., 2016).ORCHIDEE-GM模型还被用于探讨末次冰期猛犸草原大型食草动物生物量动态及其影响下的草地碳平衡(Zhu et al., 2018). ...
A Mathematical Model for Predicting Fire Spread in Wildland Fuels. USDA Forest Service General Technical Report INT-115. 1 1972
Pathways of mineral-associated soil organic matter formation: integrating the role of plant carbon source, chemistry, and point of entry 1 2019
... 土壤团聚体等物理过程对土壤物理和碳氮磷循环过程有着重要影响(Lehmann & Kleber, 2015; Sokol et al., 2019), 但目前模型仍缺乏对土壤团聚体形成等物理过程的模拟(Bradford et al., 2016).土壤微生物对土壤有机碳的分解和形成至关重要.研究发现在陆地生物圈模型中耦合土壤微生物过程可能会降低陆地对气候变化的正反馈强度(Wieder et al., 2013), 但这一研究结果不确定性很大, 需要进一步研究.目前如何在陆地生物圈模型中模拟土壤物理过程和微生物过程及其对生态系统碳氮磷循环的影响是当前的研究热点和难点(Tang & Riley, 2017; Wieder et al., 2018). ...
SUPECA kinetics for scaling redox reactions in networks of mixed substrates and consumers and an example application to aerobic soil respiration 1 2017
... 土壤团聚体等物理过程对土壤物理和碳氮磷循环过程有着重要影响(Lehmann & Kleber, 2015; Sokol et al., 2019), 但目前模型仍缺乏对土壤团聚体形成等物理过程的模拟(Bradford et al., 2016).土壤微生物对土壤有机碳的分解和形成至关重要.研究发现在陆地生物圈模型中耦合土壤微生物过程可能会降低陆地对气候变化的正反馈强度(Wieder et al., 2013), 但这一研究结果不确定性很大, 需要进一步研究.目前如何在陆地生物圈模型中模拟土壤物理过程和微生物过程及其对生态系统碳氮磷循环的影响是当前的研究热点和难点(Tang & Riley, 2017; Wieder et al., 2018). ...
Ecosystem responses to elevated CO2 governed by plant-soil interactions and the cost of nitrogen acquisition 1 2018
... 研究发现大气CO2浓度上升对植被生长的施肥效应会受到氮磷营养元素的限制(Norby et al., 2010; Vitousek et al., 2010; Terrer et al., 2018), 从而可能限制未来陆地碳汇强度, 同时增加模型对未来陆地生态系统碳平衡评估的不确定性(Goll et al., 2012; Zaehle et al., 2015).多个陆地生物圈模型在已有碳循环的基础上耦合了氮循环过程, 但已有的模型对碳氮耦合模拟的不确定性仍然很大, 改进模型碳氮耦合是模型研究的挑战(Zaehle et al., 2014).磷循环也会影响植被生长等碳循环过程, 在陆地生物圈模型中耦合磷循环也是目前模型的研究热点(Goll et al., 2017; Fleischer et al., 2019). ...
Terrestrial phosphorus limitation: mechanisms, implications, and nitrogen-phosphorus interactions 1 2010
... 研究发现大气CO2浓度上升对植被生长的施肥效应会受到氮磷营养元素的限制(Norby et al., 2010; Vitousek et al., 2010; Terrer et al., 2018), 从而可能限制未来陆地碳汇强度, 同时增加模型对未来陆地生态系统碳平衡评估的不确定性(Goll et al., 2012; Zaehle et al., 2015).多个陆地生物圈模型在已有碳循环的基础上耦合了氮循环过程, 但已有的模型对碳氮耦合模拟的不确定性仍然很大, 改进模型碳氮耦合是模型研究的挑战(Zaehle et al., 2014).磷循环也会影响植被生长等碳循环过程, 在陆地生物圈模型中耦合磷循环也是目前模型的研究热点(Goll et al., 2017; Fleischer et al., 2019). ...
Pattern and process in the plant community 1 1947
... 自1849年亚历山大·冯·洪堡(Alexander von Humboldt)首次建立气候与植物地理关系后, 经奧古斯特·格里瑟巴赫(August Grisebach)和弗拉迪米尔·彼得·柯本(Wladimir Peter K?ppen)细化完善后, 形成了植物地理预测模型的雏形.植物地理模型能够描述气候平衡态植被类型的静态空间分布, 但不能模拟碳氮水循环, 也不能模拟能量等生物物理过程.与大尺度植物地理模型相反, 描述小尺度植被动态的模型主要关注植物个体生长和死亡, 对光、水和营养元素的竞争和群落演替等过程, 这些小尺度模型逐渐模拟了小尺度上森林植被动态(Watt, 1947), 例如HYBRID和ED模型(Friend et al., 1997; Moorcroft et al., 2001).这些小尺度植被动态模型一般不能保证物质或能量守恒, 随后发展的生物地球化学循环模型建立在物质和能量守恒的基础上, 研究碳氮水在生态系统中的循环过程, 例如CENTURY模型(Parton et al., 1994).同时, 早期为了模拟陆地与大气边界层之间能量、水汽和动量过程, 与气候和大气环流模型(GCMs)相耦合, 陆面模式(LSMs)基于生物物理模型的基础应运而生, 例如SiB模型(Sellers et al., 1997).早期的陆面模式侧重陆地生态系统生物物理过程的模拟, 缺乏植被动态和生物地球化学循环过程.关于这些不同类型的模型定义和区分可进一步参考Fisher等(2014)文中的表1. ...
Global soil carbon projections are improved by modelling microbial processes 1 2013
... 土壤团聚体等物理过程对土壤物理和碳氮磷循环过程有着重要影响(Lehmann & Kleber, 2015; Sokol et al., 2019), 但目前模型仍缺乏对土壤团聚体形成等物理过程的模拟(Bradford et al., 2016).土壤微生物对土壤有机碳的分解和形成至关重要.研究发现在陆地生物圈模型中耦合土壤微生物过程可能会降低陆地对气候变化的正反馈强度(Wieder et al., 2013), 但这一研究结果不确定性很大, 需要进一步研究.目前如何在陆地生物圈模型中模拟土壤物理过程和微生物过程及其对生态系统碳氮磷循环的影响是当前的研究热点和难点(Tang & Riley, 2017; Wieder et al., 2018). ...
Carbon cycle confidence and uncertainty: exploring variation among soil biogeochemical models 1 2018
... 土壤团聚体等物理过程对土壤物理和碳氮磷循环过程有着重要影响(Lehmann & Kleber, 2015; Sokol et al., 2019), 但目前模型仍缺乏对土壤团聚体形成等物理过程的模拟(Bradford et al., 2016).土壤微生物对土壤有机碳的分解和形成至关重要.研究发现在陆地生物圈模型中耦合土壤微生物过程可能会降低陆地对气候变化的正反馈强度(Wieder et al., 2013), 但这一研究结果不确定性很大, 需要进一步研究.目前如何在陆地生物圈模型中模拟土壤物理过程和微生物过程及其对生态系统碳氮磷循环的影响是当前的研究热点和难点(Tang & Riley, 2017; Wieder et al., 2018). ...
ORCHIDEE-CROP (v0), a new process-based agro-land surface model: model description and evaluation over Europe 1 2016
... 农田覆盖了超过11%的陆地表面.农田作为保障全球粮食安全的重要基础, 受到气候变化和土地利用变化的强烈影响.同时, 农田开发和利用也对气候系统产生了重要影响.农田N2O和甲烷排放以及泥炭地开发造成的CO2排放已占人为温室气体排放的4.5% (Carlson et al., 2017).另外, 农业活动灌溉消耗了全球近70%的总用水量, 极大程度地改变了水循环格局.ORCHIDEE-CROP模型版本(Wu et al., 2016)融入了农作物模型STICS (Brisson et al., 1998)的作物生长及管理模块, 考虑小麦、玉米、大豆、水稻等主要作物的发育阶段和物候等生理过程, 以及播种及收获日期、施肥、灌溉、收获指数等管理模式, 从而模拟不同管理模式下作物产量对气候变化的响应以及对农田碳水循环的影响. ...
Multicriteria evaluation of discharge simulation in dynamic global vegetation models 1 2015
Using combined measurements of gas exchange and chlorophyll fluorescence to estimate parameters of a biochemical C3 photosynthesis model: a critical appraisal and a new integrated approach applied to leaves in a wheat (Triticum aestivum) canopy 1 2009
... ORCHIDEE对碳循环的模拟主要包括生态系统植被、凋落物和土壤碳库、各碳库之间的碳通量和生态系统与大气之间的碳通量.默认版本中碳库包括8个生物量碳库(叶、根、地上茎、地下茎、地上心材、地下心材、花和果实、储藏库)、6个凋落物碳库和3个土壤碳库.凋落物碳库根据凋落物的来源分成了地上和地下各3个周转周期的碳库; 土壤碳库的3个碳库分别是活性碳库、缓性碳库和惰性碳库(Krinner et al., 2005).光合作用模拟采用Ball Berry的计算方案(Yin et al., 2009), 光合作用的产物通过分配进入植物生物量碳库的各个部分, 同时植物进行自养呼吸(包括生长呼吸和维持呼吸)向大气中释放CO2; 而植物的器官凋落或植物死亡后, 进入凋落物碳库; 凋落物一部分经过分解以CO2的形式释放到大气中, 剩下的部分进入土壤的3个碳库.土壤3个碳库通过分解作用在3个碳库之间重新分配, 并向大气释放CO2.凋落物和土壤碳库的分解为异养呼吸.物候模块控制着植物光合作用开始和结束的时间及叶片的寿命. ...
Nitrogen availability reduces CMIP5 projections of twenty-first-century land carbon uptake 2 2015
... 研究发现大气CO2浓度上升对植被生长的施肥效应会受到氮磷营养元素的限制(Norby et al., 2010; Vitousek et al., 2010; Terrer et al., 2018), 从而可能限制未来陆地碳汇强度, 同时增加模型对未来陆地生态系统碳平衡评估的不确定性(Goll et al., 2012; Zaehle et al., 2015).多个陆地生物圈模型在已有碳循环的基础上耦合了氮循环过程, 但已有的模型对碳氮耦合模拟的不确定性仍然很大, 改进模型碳氮耦合是模型研究的挑战(Zaehle et al., 2014).磷循环也会影响植被生长等碳循环过程, 在陆地生物圈模型中耦合磷循环也是目前模型的研究热点(Goll et al., 2017; Fleischer et al., 2019). ...
Evaluation of 11 terrestrial carbon-nitrogen cycle models against observations from two temperate Free-Air CO2 Enrichment studies 1 2014
... 研究发现大气CO2浓度上升对植被生长的施肥效应会受到氮磷营养元素的限制(Norby et al., 2010; Vitousek et al., 2010; Terrer et al., 2018), 从而可能限制未来陆地碳汇强度, 同时增加模型对未来陆地生态系统碳平衡评估的不确定性(Goll et al., 2012; Zaehle et al., 2015).多个陆地生物圈模型在已有碳循环的基础上耦合了氮循环过程, 但已有的模型对碳氮耦合模拟的不确定性仍然很大, 改进模型碳氮耦合是模型研究的挑战(Zaehle et al., 2014).磷循环也会影响植被生长等碳循环过程, 在陆地生物圈模型中耦合磷循环也是目前模型的研究热点(Goll et al., 2017; Fleischer et al., 2019). ...
The large mean body size of mammalian herbivores explains the productivity paradox during the Last Glacial Maximum 1 2018
... 融入了草地管理模块的陆地生物圈模型(DLEM, LPJ-GUESS, LPJmL, ORCHIDEE)已被用于定量评估全球及区域牧草生产潜力对气候变化、大气CO2含量升高、草地利用强度等因子的响应(Chang et al., 2015; Dangal et al., 2016; Rolinski et al., 2018), 模拟区域草地碳动态及温室气体排放(Chang et al., 2015, 2017; Blanke et al., 2018), 以及对草地管理模式和利用强度进行优化(Chang et al., 2015; Rolinski et al., 2018)和重建(Chang et al., 2016).ORCHIDEE-GM模型还被用于探讨末次冰期猛犸草原大型食草动物生物量动态及其影响下的草地碳平衡(Zhu et al., 2018). ...
 ,
, ,1,*, 岳超2, 常锦峰3
,1,*, 岳超2, 常锦峰3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}