Applications of nitrogen stable isotope techniques in the study of nitrogen cycling in terrestrial ecosystems
Yun-Ting FANG,1,2,3,*, Dong-Wei LIU1,2,3, Fei-Fei ZHU1,2,3, Ying TU1,3,4, Shan-Long LI1,2, Shao-Nan HUANG1,3,4, Zhi QUAN1,2,3, Ang WANG1,2,31Key Laboratory of Forest Ecology and Management, Institute of Applied Ecology, Chinese Academy of Sciences, Shenyang 110016 2Qingyuan Forest CERN, Chinese Academy of Sciences, Shenyang 110016 China 3Key Laboratory of Stable Isotope Techniques and Applications, Liaoning Province, Shenyang, 110016 4University of Chinese Academy of Sciences, Beijing 100049, China
National Key R&D Program of China(2016YFA0600802) National Natural Science Foundation of China(41773094) Key Research Program of Frontier Sciences of Chinese Academy of Sciences(QYZDB-SSW-DQC002) K.C.Wong Education Foundation(GJTD-2018-07)
Abstract In the past several decades, the development of nitrogen (N) stable isotope techniques has improved the understanding of N cycling in terrestrial ecosystems. This review briefly introduced the history of N stable isotope techniques in studying N cycling in terrestrial ecosystems and summarized typical studies focusing on different aspects of ecosystem N cycling in recent years, including using 1) 15N natural abundance to identify plant N sources, indicate N status of ecosystems, and quantify N transformation rates; 2) 15N enriched tracers to study N fates, redistribution and gaseous loss from ecosystems. In the end, this review points out challenges and future applications of N stable isotope techniques on studying N cycling in terrestrial ecosystems. Keywords:nitrogen stable isotope technique;natural abundance technique;15N labelling technique
PDF (1080KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 方运霆, 刘冬伟, 朱飞飞, 图影, 李善龙, 黄韶楠, 全智, 王盎. 氮稳定同位素技术在陆地生态系统氮循环研究中的应用. 植物生态学报, 2020, 44(4): 373-383. DOI: 10.17521/cjpe.2019.0249 FANG Yun-Ting, LIU Dong-Wei, ZHU Fei-Fei, TU Ying, LI Shan-Long, HUANG Shao-Nan, QUAN Zhi, WANG Ang. Applications of nitrogen stable isotope techniques in the study of nitrogen cycling in terrestrial ecosystems. Chinese Journal of Plant Ecology, 2020, 44(4): 373-383. DOI: 10.17521/cjpe.2019.0249
新窗口打开|下载原图ZIP|生成PPT 图1氮循环过程中的同位素分馏效应(15ε, ‰, 修改自Denk et al., 2017)。英文字母代表生态系统氮循环过程: a, 矿化作用; b,硝化作用; c, 反硝化作用; d, 生物固氮; e, NH3和NH4+同位素交换; f, 硝酸盐异化还原成铵作用; g, 微生物固定; h, 植物吸收; i, 厌氧氨氧化作用; j, NH3挥发。
Fig. 1Isotope fractionation effects in nitrogen processes (15ε, ‰, modified by Denk et al., 2017). Processes involved in terrestrial ecosystem N cycling are: a, mineralization; b, nitrification; c, denitrification; d, biological N2 fixation; e, isotope exchange between NH3 and NH4+; f, dissimilatory nitrate reduction to ammonium; g, immobilization by microbes; h, plant uptake; i, anaerobic ammonia oxidation; j, NH3 volatilization. SOM, soil organic matter.
1.2 15N标记技术
15N标记技术是将一定数量的15N富集的含氮化合物添加到系统中(植物、土壤或整个生态系统), 经过一段时间后分析其去向(Templer et al., 2012)。在生态系统氮循环研究方面, 15N标记技术不受稳定同位素分馏效应的影响, 具有15N自然丰度法和传统示踪法所不具备的优点。但15N标记技术的最大不足之处是标记药品及样品分析费用较高, 因而限制其广泛应用。此外, 实验中还应考虑标记样品的均匀性、标记期长短和条件、示踪期长短和各库转化, 同时注意追踪的各个库要全面等问题。近年来, 随着稳定同位素技术的不断革新, 氮同位素标记物和仪器检测成本的下降, 15N标记技术已应用于陆地生态系统尺度上的各个方面, 如分析氮的来源、分配和去向、土壤氮的转化以及氮损失等研究(He et al., 2011; Yang et al., 2014; Li et al., 2019; Zhu et al., 2019)。下文以陆地生态系统中森林生态系统(自然生态系统)和农田生态系统(人工生态系统)为例, 介绍近年来15N标记技术在生态系统尺度上的应用。
近些年来, 通过配对15N标记(15NH4+和15NO3-), 越来越多的研究发现, 不仅植物和土壤对于两种沉降氮的吸收和滞留率差异明显, 而且不同类型乔木植物对两种形态氮的吸收利用效率也明显不同(Liu et al., 2017b; Goodale, 2017; Wang et al., 2018)。在温带森林, 地表凋落物层对15NH4+的吸收固持率更高, 而对15NO3-吸收固持率明显更低, 导致其在表层矿物质土壤滞留率更高(Feng et al., 2008; Liu et al., 2017b)。而在凋落物层较薄的亚热带和热带森林, 无论是地表凋落物层还是矿物质层土壤, 都更偏向吸收固持15NH4+ (Liu et al., 2017c; Wang et al., 2018)。目前, 在同一森林的铵态氮和硝态氮配对的实验较少, 但几乎所有的配对标记结果都显示, 地上植物对15NO3-的总吸收固持率明显比15NH4+更高。其主要原因被认为是沉降氮进入森林生态系统后, 15NH4+更易于被土壤中的微生物固持, 以及发生无机过程的吸附或置换, 更难以到达植物根系被根系吸收。Liu等(2017b)在吉林长白山的研究发现, 与针叶树种相比, 阔叶树种对15NO3-的吸收利用速率更快。Goodale (2017)在美国纽约州Arnot森林的研究发现, 侵染丛枝菌根真菌(AM)的树种比侵染内生菌根真菌(ECM)的树种更偏向利用NO3-。Li等(2019)在辽宁清原次生林的研究也表明, AM树种对标记的15NO3-的吸收利用速率明显快于ECM树种。其中, 蒙古栎(Quercus mongolica)作为阔叶的ECM树种, 也符合这一规律。另外, 通过15N标记后不同时间序列的研究结果表明, 沉降氮进入森林生态系统后, 其损失主要发生在第一个生长季内。在标记后的1-7年的时间内, 地表凋落物层和表层矿物质土壤中固持的沉降氮, 逐渐向更深层土壤和植物层迁移, 而通过气体释放或淋溶过程损失的比例很小, 能够较长时间地滞留在整个生态系统内(Nadelhoffer et al., 2004; Goodale, 2017; Liu et al., 2017b; Wang et al., 2018; Li et al., 2019)。
最近十年, 陆地生态系统氮稳定同位素标记技术得到长足的发展, 如利用15N交叉标记技术研究土壤氮转化速率、气态氮损失和氮在生态系统内部的去向及分配等(Müller et al., 2007; Templer et al., 2012; Sebilo et al., 2013; Yang et al., 2014; Xi et al., 2016)。然而, 目前生态系统尺度上的研究样方面积仍然在几十到几百平方米。随着同位素标记产品的多样化和成本降低, 未来的研究可以标记不同的氮形态, 也可发展到集水区尺度的示踪研究, 如集水区或景观尺度上不同形态氮损失的研究。另外, 区域尺度15N示踪研究也有必要进一步发展, 如方运霆等正在开展的中国森林15N示踪网络研究, 在国家尺度上利用15N标记技术研究不同气候区森林生态系统沉降氮去向和分配。
AckermanD, MilletDB, ChenX (2019). Global estimates of inorganic nitrogen deposition across four decades Global Biogeochemical Cycles, 33, 100-107. DOI:10.1029/2018GB005990URL
BaiE, HoultonBZ (2009). Coupled isotopic and process-based modeling of gaseous nitrogen losses from tropical rain forests Global Biogeochemical Cycles, 23, GB2011. DOI: 10.1029/2008gb003361. [本文引用: 1]
BaiE, HoultonBZ, WangYP (2012). Isotopic identification of nitrogen hotspots across natural terrestrial ecosystems BiogeoSciences, 9, 3287-3304. DOI:10.5194/bg-9-3287-2012URL [本文引用: 2]
BleshJ, DrinkwaterLE (2014). Retention of 15N-labeled fertilizer in an Illinois prairie soil with winter rye Soil Science Society of America Journal, 78, 496-508. DOI:10.2136/sssaj2013.09.0403URL [本文引用: 1] Human production of reactive nitrogen (N), particularly for agriculture, has led to consequences for air and water pollution. Winter annual cover crops show promise for reducing N losses from grain agroecosystems. We applied N-15-labeled ammonium sulfate at corn (Zea mays L.) planting in May 2009 to a long-term corn and soybean (Glycine max [ L.] Merr.) variety trial on an Illinois Mollisol. We tracked the fate of the labeled fertilizer in treatments comparing winter rye (Secale cereal) cover to the typical winter bare fallow (BF). We measured fertilizer recovery in corn at harvest in the fall, in rye biomass the following spring (May 2010), and in a range of heterogeneous soil organic matter (SOM) pools at both sampling dates. In the spring, total recovery of added N-15 in crops and soil was low (37-45%). Due to unfavorable conditions for cover crop (CC) establishment and an unusually rainy October, rye growth missed the optimal window for 15N recovery and little tracer N-15 was recovered in the rye. However, the CC significantly reduced soil inorganic N (SIN) pools in the spring (11.1 kg N ha(-1) in the BF treatment compared with 1.9 kg N ha(-1) in the CC treatment) by an amount similar in magnitude to total N uptake by rye biomass (23.7 kg N ha(-1)), suggesting that a key role of CCs is scavenging inorganic N mineralized from SOM pools. This study reflected current challenges for incorporating CCs into grain rotations with corn varieties that have longer growing seasons.
BrunnerB, ContrerasS, LehmannMF, MatantsevaO, RollogM, KalvelageT, KlockgetherG, LavikG, JettenMSM, KartalB, KuypersMMM (2013). Nitrogen isotope effects induced by anammox bacteria Proceedings of the National Academy of Sciences of the United States of America, 110, 18994-18999. URLPMID:24191043 [本文引用: 1]
BurkeIC, O’DeenLA, MosierAR, PorterLK (1990). Diffusion of soil extracts for nitrogen and nitrogen-15 analyses by automated combustion/mass spectrometry Soil Science Society of America Journal, 54, 1190-1192. DOI:10.2136/sssaj1990.03615995005400040047xURL [本文引用: 1]
ChalkPM, CraswellET, PolidoroJC, ChenDL (2015). Fate and efficiency of 15N-labelled slow- and controlled-release fertilizers Nutrient Cycling in Agroecosystems, 102, 167-178. DOI:10.1007/s10705-015-9697-2URL [本文引用: 1]
ChenZM, WangHY, LiuXW, LuDJ, ZhouJM (2016). The fates of 15N-labeled fertilizer in a wheat-soil system as influenced by fertilization practice in a loamy soil Scientific Reports, 6, 34754. DOI: 10.1038/srep34754. URLPMID:27713476 [本文引用: 1]
CoyleJS, DijkstraP, DoucettRR, SchwartzE, HartSC, HungateBA (2009). Relationships between C and N availability, substrate age, and natural abundance 13C and 15N signatures of soil microbial biomass in a semiarid climate Soil Biology & Biochemistry, 41, 1605-1611. [本文引用: 1]
CraineJM, ElmoreAJ, AidarMPM, BustamanteM, DawsonTE, HobbieEA, KahmenA, MackMC, McLauchlanKK, MichelsenA, NardotoGB, PardoLH, Pe?uelasJ, ReichPB, SchuurEAG, StockWD, TemplerPH, VirginiaRA, WelkerJM, WrightIJ (2009). Global patterns of foliar nitrogen isotopes and their relationships with climate, mycorrhizal fungi, foliar nutrient concentrations, and nitrogen availability New Phytologist, 183, 980-992. URLPMID:19563444 [本文引用: 2]
CraineJM, ElmoreAJ, WangLX, AugustoL, BaisdenWT, BrookshireENJ, CramerMD, HasselquistNJ, HobbieEA, KahmenA, KobaK, KranabetterJM, MackMC, Marin-SpiottaE, MayorJR, McLauchlanKK, MichelsenA, NardotoGB, OliveiraRS, PerakisSS, PeriPL, QuesadaCA, RichterA, SchipperLA, StevensonBA, TurnerBL, VianiRAG, WanekW, ZellerB (2015). Convergence of soil nitrogen isotopes across global climate gradients Scientific Reports, 5, 8280. DOI: 10.1038/srep08280. DOI:10.1038/srep08280URLPMID:25655192 [本文引用: 1] Quantifying global patterns of terrestrial nitrogen (N) cycling is central to predicting future patterns of primary productivity, carbon sequestration, nutrient fluxes to aquatic systems, and climate forcing. With limited direct measures of soil N cycling at the global scale, syntheses of the (15)N:(14)N ratio of soil organic matter across climate gradients provide key insights into understanding global patterns of N cycling. In synthesizing data from over 6000 soil samples, we show strong global relationships among soil N isotopes, mean annual temperature (MAT), mean annual precipitation (MAP), and the concentrations of organic carbon and clay in soil. In both hot ecosystems and dry ecosystems, soil organic matter was more enriched in (15)N than in corresponding cold ecosystems or wet ecosystems. Below a MAT of 9.8 degrees C, soil delta(15)N was invariant with MAT. At the global scale, soil organic C concentrations also declined with increasing MAT and decreasing MAP. After standardizing for variation among mineral soils in soil C and clay concentrations, soil delta(15)N showed no consistent trends across global climate and latitudinal gradients. Our analyses could place new constraints on interpretations of patterns of ecosystem N cycling and global budgets of gaseous N loss.
de OliveiraSM, de AlmeidaREM, CiampittiIA, Pierozan JuniorC, LagoBC, TrivelinPCO, FavarinJL (2018). Understanding N timing in corn yield and fertilizer N recovery: an insight from an isotopic labeled-N determination PLOS ONE, 13, e0192776. DOI: 10.1371/journal.pone.0192776. URLPMID:29462178 [本文引用: 1]
DenkTRA, MohnJ, DecockC, Lewicka-SzczebakD, HarrisE, Butterbach-BahlK, KieseR, WolfB (2017). The nitrogen cycle: a review of isotope effects and isotope modeling approaches Soil Biology & Biochemistry, 105, 121-137. DOI:10.1016/j.soilbio.2016.11.015URL [本文引用: 4]
DijkstraP, IshizuA, DoucettR, HartSC, SchwartzE, MenyailoOV, HungateBA (2006). 13C and 15N natural abundance of the soil microbial biomass Soil Biology & Biochemistry, 38, 3257-3266. DOI:10.1016/j.soilbio.2006.04.005URL [本文引用: 2]
DingLJ, AnXL, LiS, ZhangGL, ZhuYG (2014). Nitrogen loss through anaerobic ammonium oxidation coupled to iron reduction from paddy soils in a chronosequence Environmental Science & Technology, 48, 10641-10647. DOI:10.1021/es503113sURLPMID:25158120 [本文引用: 1] Anaerobic ammonium oxidation coupled to iron(III) reduction (termed Feammox) with dinitrogen, nitrite, or nitrate as the end-product is a recently discovered process of nitrogen cycling. However, Feammox has not been described in paddy soils, which are rich in iron(III) oxides and subjected to intensive nitrogen fertilization. Here, evidence for Feammox in a paddy soil chronosequence with a gradient of microbially reducible iron(III) levels was obtained in Southern China using (15)N-labeled ammonium-based isotopic tracing and acetylene inhibition techniques. Our study demonstrated the occurrence of Feammox in the chronosequence, and direct dinitrogen production was shown to be the dominant Feammox pathway. Within the chronosequence, three paddy soils with higher microbially reducible iron(III) levels had higher Feammox rates (ranged from 0.17 to 0.59 mg N kg(-1) d(-1)) compared to an uncultivated soil (0.04 mg N kg(-1) d(-1)). It is estimated that a loss of 7.8-61 kg N ha(-1) year(-1) is associated with Feammox in the examined paddy soils. Overall, we discover that rice cultivation could enrich microbially reducible iron(III), accelerate Feammox reaction and thus fuel nitrogen loss from soils, and suggest that Feammox could be a potentially important pathway for nitrogen loss in paddy soils.
FangYT, KobaK, MakabeA, TakahashiC, ZhuWX, HayashiT, HokariAA, UrakawaR, BaiE, HoultonBZ, XiD, ZhangSS, MatsushitaK, TuY, LiuDW, ZhuFF, WangZY, ZhouGY, ChenDX, MakitaT, TodaH, LiuXY, ChenQS, ZhangDQ, LiYD, YohM (2015). Microbial denitrification dominates nitrate losses from forest ecosystems Proceedings of the National Academy of Sciences of the United States of America, 112, 1470-1474. URLPMID:25605898 [本文引用: 3]
FengZ, BrummeR, XuYJ, LamersdorfN (2008). Tracing the fate of mineral N compounds under high ambient N deposition in a Norway spruce forest at Solling/Germany Forest Ecology and Management, 255, 2061-2073. DOI:10.1016/j.foreco.2007.12.049URL [本文引用: 1]
FryB (2006). Stable Isotope Ecology. Springer, New York.
GabrielJL, Alonso-AyusoM, García-GonzálezI, HontoriaC, QuemadaM (2016). Nitrogen use efficiency and fertiliser fate in a long-term experiment with winter cover crops European Journal of Agronomy, 79, 14-22. [本文引用: 1]
GallowayJN, DentenerFJ, CaponeDG, BoyerEW, HowarthRW, SeitzingerSP, AsnerGP, ClevelandCC, GreenPA, HollandEA, KarlDM, MichaelsAF, PorterJH, TownsendAR, V?r?smartyCJ (2004). Nitrogen cycles: past, present, and future Biogeochemistry, 70, 153-226. DOI:10.1007/s10533-004-0370-0URL This paper contrasts the natural and anthropogenic controls on the conversion of unreactive N2 to more reactive forms of nitrogen (Nr). A variety of data sets are used to construct global N budgets for 1860 and the early 1990s and to make projections for the global N budget in 2050. Regional N budgets for Asia, North America, and other major regions for the early 1990s, as well as the marine N budget, are presented to Highlight the dominant fluxes of nitrogen in each region. Important findings are that human activities increasingly dominate the N budget at the global and at most regional scales, the terrestrial and open ocean N budgets are essentially disconnected, and the fixed forms of N are accumulating in most environmental reservoirs. The largest uncertainties in our understanding of the N budget at most scales are the rates of natural biological nitrogen fixation, the amount of Nr storage in most environmental reservoirs, and the production rates of N2 by denitrification.
GardnerJB, DrinkwaterLE (2009). The fate of nitrogen in grain cropping systems: a meta-analysis of 15N field experiments Ecological Applications, 19, 2167-2184. DOI:10.1890/08-1122.1URLPMID:20014586 [本文引用: 2] Intensively managed grain farms are saturated with large inputs of nitrogen (N) fertilizer, leading to N losses and environmental degradation. Despite decades of research directed toward reducing N losses from agroecosystems, progress has been minimal, and the currently promoted best management practices are not necessarily the most effective. We investigated the fate of N additions to temperate grain agroecosystems using a meta-analysis of 217 field-scale studies that followed the stable isotope 15N in crops and soil. We compared management practices that alter inorganic fertilizer additions, such as application timing or reduced N fertilizer rates, to practices that re-couple the biogeochemical cycles of carbon (C) and N, such as organic N sources and diversified crop rotations, and analyzed the following response variables: 15N recovery in crops, total recovery of 15N in crops and soil, and crop yield. More of the literature (94%) emphasized crop recovery of 15N than total 15N recovery in crops and soil (58%), though total recovery is a more ecologically appropriate indicator for assessing N losses. Findings show wide differences in the ability of management practices to improve N use efficiency. Practices that aimed to increase crop uptake of commercial fertilizer had a lower impact on total 15N recovery (3-21% increase) than practices that re-coupled C and N cycling (30-42% increase). A majority of studies (66%) were only one growing season long, which poses a particular problem when organic N sources are used because crops recover N from these sources over several years. These short-term studies neglect significant ecological processes that occur over longer time scales. Field-scale mass balance calculations using the 15N data set show that, on average, 43 kg N x ha(-1) x yr(-1) was unaccounted for at the end of one growing season out of 114 kg N x ha(-1) x yr(-1), representing approximately 38% of the total 15N applied. This comprehensive assessment of stable-isotope research on agroecosystem N management can inform the development of policies to mitigate nonpoint source pollution. Nitrogen management practices that most effectively increase N retention are not currently being promoted and are rare on the landscape in the United States.
GoodaleCL (2017). Multiyear fate of a 15N tracer in a mixed deciduous forest: retention, redistribution, and differences by mycorrhizal association Global Change Biology, 23, 867-880. DOI:10.1111/gcb.13483URLPMID:27564989 [本文引用: 4] The impact of atmospheric nitrogen deposition on forest ecosystems depends in large part on its fate. Past tracer studies show that litter and soils dominate the short-term fate of added (15) N, yet few have examined its longer term dynamics or differences among forest types. This study examined the fate of a (15) N-NO3- tracer over 5-6 years in a mixed deciduous stand that was evenly composed of trees with ectomycorrhizal and arbuscular mycorrhizal associations. The tracer was expected to slowly mineralize from its main initial fate in litter and surface soil, with some (15) N moving to trees, some to deeper soil, and some net losses. Recovery of added (15) N in trees and litterfall totaled 11.3% both 1 and 5-6 years after the tracer addition, as (15) N redistributed from fine and especially coarse roots into cumulative litterfall and small accumulations in woody tissues. Estimates of potential carbon sequestration from tree (15) N recovery amounted to 12-14 kg C per kg of N deposition. Tree (15) N acquisition occurred within the first year after the tracer addition, with no subsequent additional net transfer of (15) N from detrital to plant pools. In both years, ectomycorrhizal trees gained 50% more of the tracer than did trees with arbuscular mycorrhizae. Much of the (15) N recovered in wood occurred in tree rings formed prior to the (15) N addition, demonstrating the mobility of N in wood. Tracer recovery rapidly decreased over time in surface litter material and accumulated in both shallow and deep soil, perhaps through mixing by earthworms. Overall, results showed redistribution of tracer (15) N through trees and surface soils without any losses, as whole-ecosystem recovery remained constant between 1 and 5-6 years at 70% of the (15) N addition. These results demonstrate the persistent ecosystem retention of N deposition even as it redistributes, without additional plant uptake over this timescale.
GroffmanPM, AltabetMA, B?hlkeJK, Butterbach-BahlK, DavidMB, FirestoneMK, GiblinAE, KanaTM, NielsenLP, VoytekMA (2006). Methods for measuring denitrification: diverse approaches to a difficult problem Ecological Applications, 16, 2091-2122. DOI:10.1890/1051-0761(2006)016[2091:mfmdda]2.0.co;2URLPMID:17205891 [本文引用: 1] Denitrification, the reduction of the nitrogen (N) oxides, nitrate (NO3-) and nitrite (NO2-), to the gases nitric oxide (NO), nitrous oxide (N2O), and dinitrogen (N2), is important to primary production, water quality, and the chemistry and physics of the atmosphere at ecosystem, landscape, regional, and global scales. Unfortunately, this process is very difficult to measure, and existing methods are problematic for different reasons in different places at different times. In this paper, we review the major approaches that have been taken to measure denitrification in terrestrial and aquatic environments and discuss the strengths, weaknesses, and future prospects for the different methods. Methodological approaches covered include (1) acetylene-based methods, (2) 15N tracers, (3) direct N2 quantification, (4) N2:Ar ratio quantification, (5) mass balance approaches, (6) stoichiometric approaches, (7) methods based on stable isotopes, (8) in situ gradients with atmospheric environmental tracers, and (9) molecular approaches. Our review makes it clear that the prospects for improved quantification of denitrification vary greatly in different environments and at different scales. While current methodology allows for the production of accurate estimates of denitrification at scales relevant to water and air quality and ecosystem fertility questions in some systems (e.g., aquatic sediments, well-defined aquifers), methodology for other systems, especially upland terrestrial areas, still needs development. Comparison of mass balance and stoichiometric approaches that constrain estimates of denitrification at large scales with point measurements (made using multiple methods), in multiple systems, is likely to propel more improvement in denitrification methods over the next few years.
GurmesaGA, LuXK, GundersenP, MaoQG, ZhouKJ, FangYT, MoJM (2016). High retention of 15N-labeled nitrogen deposition in a nitrogen saturated old-growth tropical forest Global Change Biology, 22, 3608-3620. URLPMID:27097744 [本文引用: 1]
HastingsMG (2010). Evaluating source, chemistry and climate change based upon the isotopic composition of nitrate in ice cores IOP Conference Series: Earth and Environmental Science, 9, 012002. DOI: 10.1088/1755-1315/9/1/012002. DOI:10.1088/1755-1315/9/1/012002URL [本文引用: 1]
HeHB, LüH, ZhangW, HouSM, ZhangXD (2011). A liquid chromatographic/mass spectrometric method to evaluate 13C and 15N incorporation into soil amino acids Journal of Soils and Sediments, 11, 731-740. DOI:10.1007/s11368-011-0360-5URL [本文引用: 2] Amino acids are highly associated with biogeochemical cycling and represent an important potential source and sink of carbon (C) and nitrogen (N) in terrestrial ecosystems. Tracing the isotope dynamics of amino acids can improve the understanding of the origin and transformation of amino acids in soil matrix at process-levels; hence, the liquid chromatographic/mass spectrometric (LC/MS) method to evaluate C-13 or N-15 enrichment in amino acids is necessary to be established. Laboratory incubations of a Mollisol (sampled from 0-20 cm) were conducted with either (NH)-N-15 (4) (+) plus glucose or NH (4) (+) plus U-C-13-glucose as substrates. The substrates were added weekly until the soils were sampled after 1 and 4 weeks, respectively. The soil samples were then ground to < 0.25 mm and hydrolyzed to release amino acids. After being purified, the amino acids were derivatized with 6-aminoquinolyl-N-hydroxysuccinimidyl carbamate and then identified by LC/MS. The air-dried original soil was used as control and analyzed in the same assay. The amino acids were quantified based on the total current ion chromatograms. To fractionate and quantify the C and N isotopes in the amino acids, the intensities of the quasi-molecular peaks and the minor fragments were measured under both scan and selective ion monitoring modes. The intensities of the corresponding isotope fragments in amino acids increased significantly after labile substrate addition, indicating the N-15 and C-13 incorporation into soil amino acids, including both free and proteinaceous or peptide forms during the incubations. The synchronous utilization of glucose was dominant to structure amino acid skeleton and the conversion from NH (4) (+) to multi-nitrogen-containing amino acids exhibited the same pattern. The isotope enrichment in the amino acids was calculated according to the relative intensity increase of the isotope fragment and expressed as atom percentage excess (APE). The amount of isotope-labeled amino acids was furthermore differentiated from the native portion based on both APE and the concentration of individual compounds. The isotope-based LC/MS technique was useful to assess N-15 and C-13 enrichment in soil amino acids at relative high levels. The quantification of isotope incorporation makes it possible to evaluate the C or N turnover velocity of individual amino acids induced by the available substrates. Furthermore, the differentiation between the labeled and unlabeled amino acids was quite helpful to investigate the synthesis-decomposition dynamics of amino acid pools and explore the pathways of biological transformation of soil amino acids.
HeHB, XieHT, ZhangXD (2006). A novel GC/MS technique to assess 15N and 13C incorporation into soil amino sugars Soil Biology & Biochemistry, 38, 1083-1091. DOI:10.1016/j.soilbio.2005.09.007URL [本文引用: 1]
HietzP, TurnerBL, WanekW, RichterA, NockCA, WrightSJ (2011). Long-term change in the nitrogen cycle of tropical forests Science, 334, 664-666. DOI:10.1126/science.1211979URLPMID:22053047 [本文引用: 1] Deposition of reactive nitrogen (N) from human activities has large effects on temperate forests where low natural N availability limits productivity but is not known to affect tropical forests where natural N availability is often much greater. Leaf N and the ratio of N isotopes (delta(15)N) increased substantially in a moist forest in Panama between ~1968 and 2007, as did tree-ring delta(15)N in a dry forest in Thailand over the past century. A decade of fertilization of a nearby Panamanian forest with N caused similar increases in leaf N and delta(15)N. Therefore, our results indicate regional increases in N availability due to anthropogenic N deposition. Atmospheric nitrogen dioxide measurements and increased emissions of anthropogenic reactive N over tropical land areas suggest that these changes are widespread in tropical forests.
HoeringT (1955). Variations of nitrogen-15 abundance in naturally occurring substances Science, 122, 1233-1234. URLPMID:17754703 [本文引用: 1]
HoultonBZ, BaiE (2009). Imprint of denitrifying bacteria on the global terrestrial biosphere Proceedings of the National Academy of Sciences of the United States of America, 106, 21713-21716. URLPMID:19995974 [本文引用: 3]
HoultonBZ, MarkleinAR, BaiE (2015). Representation of nitrogen in climate change forecasts Nature Climate Change, 5, 398-401. DOI:10.1038/nclimate2538URL [本文引用: 1]
HoultonBZ, SigmanDM, HedinLO (2006). Isotopic evidence for large gaseous nitrogen losses from tropical rainforests Proceedings of the National Academy of Sciences of the United States of America, 103, 8745-8750. URLPMID:16728510 [本文引用: 4]
HoultonBZ, SigmanDM, SchuurEAG, HedinLO (2007). A climate-driven switch in plant nitrogen acquisition within tropical forest communities Proceedings of the National Academy of Sciences of the United States of America, 104, 8902-8906. URLPMID:17502607 [本文引用: 4]
HuangSN, ElliottEM, FelixJD, PanYP, LiuDW, LiSL, LiZJ, ZhuFF, ZhangN, FuPQ, FangYT (2019). Seasonal pattern of ammonium 15N natural abundance in precipitation at a rural forested site and implications for NH3 source partitioning Environmental Pollutant, 247, 541-549. [本文引用: 1]
JuXT (2014). The concept and meanings of nitrogen fertilizer availability ratio—Discussing misunderstanding of traditional nitrogen use efficiency Acta Pedologica Sinica, 51, 921-933. [本文引用: 1]
JuXT, ChristieP (2011). Calculation of theoretical nitrogen rate for simple nitrogen recommendations in intensive cropping systems: a case study on the North China Plain Field Crops Research, 124, 450-458. DOI:10.1016/j.fcr.2011.08.002URL [本文引用: 1] Nitrogen (N) is a crucial nutrient that requires careful management in intensive cropping systems because of its diverse beneficial and detrimental effects. Here we propose the concept of theoretical N rate (TNR) to answer the important question of how much fertilizer N should be applied to intensive systems based on the N fluxes due to transformation processes in the soil crop environment continuum. We define TNR as the theoretically calculated fertilizer N rate with the quantitative relationships of the core N fluxes among fertilizer N, soil N and crop uptake N in the crop root zone to obtain high target yield, maintain soil N balance and minimize environmental risk. We deduced one basic mathematical expression (N-fert = N-uptake - N-straw - N-fert3) and two simplified expressions [N-fert = (N-uptake - N-straw)/(1 - Coeff); N-fert congruent to N-uptake] for calculating the TNR. These expressions do not need much field experimentation or elaborate soil and plant testing to obtain information on crop N demand and soil N supply, and are simple to implement in farming practice to provide a very cost-effective approach. We consider this scheme to be a useful contribution to rational fertilizer practice, especially in developing countries where other N recommendation systems are usually not available and agricultural extension services are poorly developed or absent. (C) 2011 Elsevier B.V.
KobaK, InagakiK, SasakiY, TakebayashiY, YohM (2010). Nitrogen isotopic analysis of dissolved inorganic and organic nitrogen in soil extracts//Ohkouchi N, Tayasu I, Koba K. Earth, Life and Isotopes. Kyoto University Press, Kyoto, Japan. 17-36. [本文引用: 1]
KoolDM, DolfingJ, WrageN, van GroenigenJW (2011). Nitrifier denitrification as a distinct and significant source of nitrous oxide from soil Soil Biology & Biochemistry, 43, 174-178. DOI:10.1016/j.soilbio.2010.09.030URL [本文引用: 1]
KrauseK, ProvidoliI, CurrieWS, BugmannH, SchleppiP (2012). Long-term tracing of whole catchment 15N additions in a mountain spruce forest: measurements and simulations with the TRACE model Trees, 26, 1683-1702. DOI:10.1007/s00468-012-0737-0URL [本文引用: 1]
LadhaJK, PathakH, KrupnikTJ, SixJ, van KesselC (2005). Efficiency of fertilizer nitrogen in cereal production: retrospects and prospects Advances in Agronomy, 87, 85-156. [本文引用: 1]
LeBauerDS, TresederKK (2008). Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed Ecology, 89, 371-379. URLPMID:18409427
LiSL, GurmesaGA, ZhuWX, GundersenP, ZhangSS, XiD, HuangSN, WangA, ZhuFF, JiangY, ZhuJJ, FangYT (2019). Fate of atmospherically deposited NH4+ and NO3- in two temperate forests in China: temporal pattern and redistribution Ecological Applications, 29, e01920. DOI: 10.1002/eap.1920. DOI:10.1002/eap.1920URLPMID:31058370 [本文引用: 4] The impacts of anthropogenic nitrogen (N) deposition on forest ecosystems depend in large part on its fate. However, our understanding of the fates of different forms of deposited N as well as the redistribution over time within different ecosystems is limited. In this study, we used the (15) N-tracer method to investigate both the short-term (1 week to 3 months) and long-term (1-3 yr) fates of deposited NH4 (+) or NO3 (-) by following the recovery of the (15) N in different ecosystem compartments in a larch plantation forest and a mixed forest located in northeastern China. The results showed similar total ecosystem retention for deposited NH4 (+) and NO3 (-) , but their distribution within the ecosystems (plants vs. soil) differed distinctly particularly in the short-term, with higher (15) NO3 (-) recoveries in plants (while lower recoveries in organic layer) than found for (15) NH4 (+) . The different short-term fate was likely related to the higher mobility of (15) NO3 (-) than (15) NH4 (+) in soils instead of plant uptake preferences for NO3 (-) over NH4 (+) . In the long-term, differences between N forms became less prevalent but higher recoveries in trees (particularly in the larch forest) of (15) NO3 (-) than (15) NH4 (+) tracer persisted, suggesting that incoming NO3 (-) may contribute more to plant biomass increment and forest carbon sequestration than incoming NH4 (+) . Differences between the two forests in recoveries were largely driven by a higher (15) N recovery in the organic layer (both N forms) and in trees (for (15) NO3 (-) ) in the larch forest compared to the mixed forest. This was due to a more abundant organic layer and possibly higher tree N demand in the larch forest than in the mixed forest. Leachate (15) N loss was minor (<1% of the added (15) N) for both N forms and in both forests. Total (15) N recovery averaged 78% in the short-term and decreased to 55% in the long-term but with increasing amount of (15) N label (re)-redistributed into slow turn-over pools (e.g., trees and mineral soil). The different retention dynamics of deposited NH4 (+) and NO3 (-) may have implications in environmental policy related to the anthropogenic emissions of the two N forms.
LiangB, ZhaoW, YangXY, ZhouJB (2013). Fate of nitrogen- 15 as influenced by soil and nutrient management history in a 19-year wheat-maize experiment Field Crops Research, 144, 126-134. DOI:10.1016/j.fcr.2012.12.007URL [本文引用: 1] High soil organic matter content may improve synchronization between N supply and crop demand. To test this hypothesis, we compared the fate of N-15-labeled fertilizer in soil with different management history. The soils had received no fertilizer (No-F soil), inorganic N, P. and K fertilizer (NPK soil), or manure plus N, P. and K fertilizer (MNPK soil) as part of a 19-year long-term fertilization trial. The N use efficiency (NUE) of wheat (Triticum aestiyum L.) was 62% in the MNPK soil, higher than that in the NPK soil (50% NUE), and in the No-F soil (13% NUE). At wheat harvest, 38% of the fertilizer N-15 remained in the 0-100 cm depth of the MNPK soil, significantly less that the amount of fertilizer N-15 that remained in the NPK soil (45%) or in the No-F soil (88%). More than 50% of the fertilizer N-15 in the No-F soil had leached below the 20 cm depth by wheat harvest, significantly more than in the NPK or MNPK soils. The amount of immobilized N-15 at wheat stem elongation was significantly (P<0.05) greater in the MNPK soil than in the NPK soil. The mineralization of immobilized N-15 between stem elongation and flowering was also significantly higher in the MNPK soil than in the NPK soil (P<0.05). The succeeding maize (Zea mays L.) crop took up 9% of the fertilizer N-15 in the No-F soil, 6% of the fertilizer N-15 in the NPK soil, and 2% of the fertilizer N-15 in the MNPK soil. Combined soil profile and crop removal analyses at wheat harvest accounted for nearly 100% of the fertilizer N-15 for all three soils. However, only 45% of the fertilizer N-15 added to the No-F soil could be accounted for at maize harvest, significantly less than the recovery rate in the NPK (83%) and MNPK (85%) soils (P<0.01). These results indicate that the fertilizer N-15 was mainly lost from these soils during the maize growing season. We conclude that the combined application of manure and inorganic fertilizers improves synchrony between N supply and crop demand, thus reducing N losses from agriculture. (C) 2012 Elsevier B.V.
LiuDW, TuY, FangYT (2017). Isotope analysis of ammonium and nitrate: a review on measured methods and their application Chinese Journal of Applied Ecology, 28, 2353-2360. URLPMID:29741070 [本文引用: 1]
LiuDW, ZhuWX, WangXB, PanYP, WangC, XiD, BaiE, WangYS, HanXG, FangYT (2017a). Abiotic versus biotic controls on soil nitrogen cycling in drylands along a 3200 km transect Biogeosciences, 14, 989-1001. DOI:10.5194/bg-14-989-2017URL [本文引用: 2]
LiuJ, PengB, XiaZW, SunJF, GaoDC, DaiWW, JiangP, BaiE (2017b). Different fates of deposited NH4+ and NO3- in a temperate forest in northeast China: a 15N tracer study Global Change Biology, 23, 2441-2449. DOI:10.1111/gcb.13533URLPMID:27753166 [本文引用: 4] Increasing atmospheric reactive nitrogen (N) deposition due to human activities could change N cycling in terrestrial ecosystems. However, the differences between the fates of deposited NH4+ and NO3- are still not fully understood. Here, we investigated the fates of deposited NH4+ and NO3-, respectively, via the application of (15) NH4 NO3 and NH4(15) NO3 in a temperate forest ecosystem. Results showed that at 410 days after tracer application, most 15NH4+ was immobilized in litter layer (50 +/- 2%), while a considerable amount of 15NO3- penetrated into 0-5 cm mineral soil (42 +/- 2%), indicating that litter layer and 0-5 cm mineral soil were the major N sinks of NH4+ and NO3-, respectively. Broad-leaved trees assimilated more (15) N under NH4(15) NO3 treatment compared to under (15) NH4 NO3 treatment, indicating their preference for NO3--N. At 410 days after tracer application, 16 +/- 4% added (15) N was found in aboveground biomass under 15NO3- treatment, which was twice more than that under 15NH4+ treatment (6 +/- 1%). At the same time, approximately 80% added (15) N was recovered in soil and plants under both treatments, which suggested that this forest had high potential for retention of deposited N. These results provided evidence that there were great differences between the fates of deposited NH4+ and NO3-, which could help us better understand the mechanisms and capability of forest ecosystems as a sink of reactive nitrogen.
LiuWJ, YuLF, ZhangT, KangRH, ZhuJ, MulderJ, HuangYM, DuanL (2017c). In situ15N labeling experiment reveals different long-term responses to ammonium and nitrate inputs in N-saturated subtropical forest Journal of Geophysical Research, 122, 2251-2264. [本文引用: 1]
LiuXE, LiXG, GuoRY, KuzyakovY, LiFM (2015). The effect of plastic mulch on the fate of urea-N in rain-fed maize production in a semiarid environment as assessed by 15N-labeling European Journal of Agronomy, 70, 71-77. DOI:10.1016/j.eja.2015.07.006URL [本文引用: 1]
LiuXY, KobaK, KoyamaLA, HobbieSE, WeissMS, InagakiY, ShaverGR, GiblinAE, HobaraS, NadelhofferKJ, SommerkornM, RastetterEB, KlingGW, LaundreJA, YanoY, MakabeA, YanoM, LiuCQ (2018). Nitrate is an important nitrogen source for Arctic tundra plants Proceedings of the National Academy of Sciences of the United States of America, 115, 3398-3403. DOI:10.1073/pnas.1715382115URLPMID:29540568 [本文引用: 1] Plant nitrogen (N) use is a key component of the N cycle in terrestrial ecosystems. The supply of N to plants affects community species composition and ecosystem processes such as photosynthesis and carbon (C) accumulation. However, the availabilities and relative importance of different N forms to plants are not well understood. While nitrate (NO3(-)) is a major N form used by plants worldwide, it is discounted as a N source for Arctic tundra plants because of extremely low NO3(-) concentrations in Arctic tundra soils, undetectable soil nitrification, and plant-tissue NO3(-) that is typically below detection limits. Here we reexamine NO3(-) use by tundra plants using a sensitive denitrifier method to analyze plant-tissue NO3(-) Soil-derived NO3(-) was detected in tundra plant tissues, and tundra plants took up soil NO3(-) at comparable rates to plants from relatively NO3(-)-rich ecosystems in other biomes. Nitrate assimilation determined by (15)N enrichments of leaf NO3(-) relative to soil NO3(-) accounted for 4 to 52% (as estimated by a Bayesian isotope-mixing model) of species-specific total leaf N of Alaskan tundra plants. Our finding that in situ soil NO3(-) availability for tundra plants is high has important implications for Arctic ecosystems, not only in determining species compositions, but also in determining the loss of N from soils via leaching and denitrification. Plant N uptake and soil N losses can strongly influence C uptake and accumulation in tundra soils. Accordingly, this evidence of NO3(-) availability in tundra soils is crucial for predicting C storage in tundra.
MaryB, RecousS, RobinD (1998). A model for calculating nitrogen fluxes in soil using 15N tracing Soil Biology & Biochemistry, 30, 1963-1979. DOI:10.1016/S0038-0717(98)00068-6URL [本文引用: 1]
MayorJR, SchuurEAG, MackMC, HollingsworthTN, B??thE (2012). Nitrogen isotope patterns in Alaskan black spruce reflect organic nitrogen sources and the activity of ectomycorrhizal fungi Ecosystems, 15, 819-831. DOI:10.1007/s10021-012-9548-9URL [本文引用: 1] Global patterns in soil, plant, and fungal stable isotopes of N (delta N-15) show promise as integrated metrics of N cycling, particularly the activity of ectomycorrhizal (ECM) fungi. At small spatial scales, however, it remains difficult to differentiate the underlying causes of plant delta N-15 variability and this limits the application of such measurements to better understand N cycling. We conducted a landscape-scale analysis of delta N-15 values from 31 putatively N-limited monospecific black spruce (Picea mariana) stands in central Alaska to assess the two main hypothesized sources of plant delta N-15 variation: differing sources and ECM fractionation. We found roughly 20% of the variability in black spruce foliar N and delta N-15 values to be correlated with the concentration and delta N-15 values of soil NH4 (+) and dissolved organic N (DON) pools, respectively. However, N-15-based mixing models from 24 of the stands suggested that fractionation by ECM fungi obscures the N-15 signature of soil N pools. Models, regressions, and N abundance data all suggested that increasing dependence on soil DON to meet black spruce growth demands predicates increasing reliance on ECM-derived N and that black spruce, on average, received 53% of its N from ECM fungi. Future research should partition the delta N-15 values within the soil DON pool to determine how choice of soil delta N-15 values influence modeled ECM activity. The C balance of boreal forests is tightly linked to N cycling and delta N-15 values may be useful metrics of changes to these connections.
McIlvinMR, AltabetMA (2005). Chemical conversion of nitrate and nitrite to nitrous oxide for nitrogen and oxygen isotopic analysis in freshwater and seawater Analytical Chemistry, 77, 5589-5595. DOI:10.1021/ac050528sURLPMID:16131070 [本文引用: 1] We present a novel method for nitrogen and oxygen natural isotopic abundance analysis of nitrate and nitrite of seawater and freshwater at environmental concentrations. The method involves the reduction of nitrate to nitrite using spongy cadmium with further reduction to nitrous oxide using sodium azide in an acetic acid buffer. For separate nitrite analysis, the cadmium reduction step is simply bypassed. Nitrous oxide is purged from the water sample and trapped cryogenically using an automated system with subsequent release into a gas chromatography column. The isolated nitrous oxide is then analyzed on a continuous flow isotope ratio mass spectrometer via an open split. This paper describes the basic protocol and reaction conditions required to obtain reproducible natural abundance level nitrogen and oxygen isotopic ratios from nitrate, nitrite, or both, and the results obtained to support these conclusions. A standard deviation less than 0.2 per thousand for nitrogen and 0.5 per thousand for oxygen was found for nitrate samples ranging in concentration from 40 to 0.5 microM (better for nitrite), with a blank of 2 nmol for 50-mL samples. Nitrogen and oxygen isotopic fractionation and oxygen atom exchange were consistent within each batch of analysis. There was no interference from any seawater matrixes. Only one other method published to date can measure the nitrate oxygen isotopic abundance in seawater and none that do so for nitrite alone in the presence of nitrate. This method may prove to be simpler, faster, and obtain isotopic information for lower concentrations of nitrate and nitrite than other methods.
McLauchlanKK, CraineJM, OswaldWW, LeavittPR, LikensGE (2007). Changes in nitrogen cycling during the past century in a northern hardwood forest Proceedings of the National Academy of Sciences of the United States of America, 104, 7466-7470. DOI:10.1073/pnas.0701779104URLPMID:17446271 [本文引用: 1] Nitrogen (N) availability, defined here as the supply of N to terrestrial plants and soil microorganisms relative to their N demands, limits the productivity of many temperate zone forests and in part determines ecosystem carbon (C) content. Despite multidecadal monitoring of N in streams, the long-term record of N availability in forests of the northeastern United States is largely unknown. Therefore, although these forests have been receiving anthropogenic N deposition for the past few decades, it is still uncertain whether terrestrial N availability has changed during this time and, subsequently, whether forest ecosystems have responded to increased N deposition. Here, we used stable N isotopes in tree rings and lake sediments to demonstrate that N availability in a northeastern forest has declined over the past 75 years, likely because of ecosystem recovery from Euro-American land use. Forest N availability has only recently returned to levels forecast from presettlement trajectories, rendering the trajectory of future forest N cycling uncertain. Our results suggest that chronic disturbances caused by humans, especially logging and agriculture, are major drivers of terrestrial N cycling in forest ecosystems today, even a century after cessation.
McLauchlanKK, FergusonCJ, WilsonIE, OcheltreeTW, CraineJM (2010). Thirteen decades of foliar isotopes indicate declining nitrogen availability in central North American grasslands New Phytologist, 187, 1135-1145. URLPMID:20553396 [本文引用: 1]
McLauchlanKK, GerhartLM, BattlesJJ, CraineJM, ElmoreAJ, HigueraPE, MackMC, McNeilBE, NelsonDM, PedersonN, PerakisSS (2017). Centennial-scale reductions in nitrogen availability in temperate forests of the United States Scientific Report, 7, 7856. DOI: 10.1038/s41598-017-08170-z. [本文引用: 1]
MichenerR, LajthaAK (2007). Stable Isotopes in Ecology and Environmental Science. Blackwell Publications, Oxford.
M?biusJ (2013). Isotope fractionation during nitrogen remineralization (ammonification): implications for nitrogen isotope biogeochemistry Geochimica et Cosmochimica Acta, 105, 422-432. DOI:10.1016/j.gca.2012.11.048URL [本文引用: 1] The N-15/N-14 isotopic ratio of marine particulate organic matter usually increases as a consequence of preferential N-14-remineralization (ammonification) during decay in the oxic water-column and at the oxic seafloor. However, a fractionation factor (15)epsilon of sediment ammonification in natural environments has not been determined yet. Here, the (15)epsilon of sedimentary ammonification is calculated in a set of seven sapropel containing sediment cores, two subrecent sediment cores and four sediment trap deployments from the Eastern Mediterranean Sea. Data clearly reveal Rayleigh type isotope fractionation and a fractionation factor (15)epsilon between -1.43 parts per thousand and -2.3 parts per thousand. Hence, sediments - and probably sinking particulate organic matter as well - can be important sources of isotopically depleted reactive nitrogen. The knowledge of (15)epsilon further enables to reconstruct paleo export fluxes from sediment cores if original and actual delta N-15 as well as N accumulation rates are known. (C) 2012 Elsevier Ltd.
MüllerC, LaughlinRJ, SpottO, RüttingT (2014). Quantification of N2O emission pathways via a 15N tracing model Soil Biology & Biochemistry, 72, 44-54. DOI:10.1016/j.soilbio.2014.01.013URL [本文引用: 1]
MüllerC, RüttingT, KattgeJ, LaughlinRJ, StevensRJ (2007). Estimation of parameters in complex 15N tracing models by Monte Carlo sampling Soil Biology & Biochemistry, 39, 715-726. DOI:10.1016/j.soilbio.2006.09.021URL [本文引用: 2]
MurphyRP, Montes-MolinaJA, GovaertsB, SixJ, van KesselC, FonteSJ (2016). Crop residue retention enhances soil properties and nitrogen cycling in smallholder maize systems of Chiapas, Mexico Applied Soil Ecology, 103, 110-116. DOI:10.1016/j.apsoil.2016.03.014URL [本文引用: 1]
NadelhofferKJ, ColmanBP, CurrieWS, MagillA, AberJD (2004). Decadal-scale fates of 15N tracers added to oak and pine stands under ambient and elevated N inputs at the Harvard Forest (USA) Forest Ecology and Management, 196, 89-107. DOI:10.1016/j.foreco.2004.03.014URL [本文引用: 2]
NieSA, LiH, YangXR, ZhangZJ, WengBS, HuangFY, ZhuGB, ZhuYG (2015). Nitrogen loss by anaerobic oxidation of ammonium in rice rhizosphere The ISME Journal, 9, 2059-2067. DOI:10.1038/ismej.2015.25URLPMID:25689022 [本文引用: 1] Anaerobic oxidation of ammonium (anammox) is recognized as an important process for nitrogen (N) cycling, yet its role in agricultural ecosystems, which are intensively fertilized, remains unclear. In this study, we investigated the presence, activity, functional gene abundance and role of anammox bacteria in rhizosphere and non-rhizosphere paddy soils using catalyzed reporter deposition-fluorescence in situ hybridization, isotope-tracing technique, quantitative PCR assay and 16S rRNA gene clone libraries. Results showed that rhizosphere anammox contributed to 31-41% N2 production with activities of 0.33-0.64 nmol N2 g(-1) soil h(-1), whereas the non-rhizosphere anammox bacteria contributed to only 2-3% N2 production with lower activities of 0.08-0.26 nmol N2 g(-1) soil h(-1). Higher anammox bacterial cells were observed (0.75-1.4 x 10(7) copies g(-1) soil) in the rhizosphere, which were twofold higher compared with the non-rhizosphere soil (3.7-5.9 x 10(6) copies g(-1) soil). Phylogenetic analysis of the anammox bacterial 16S rRNA genes indicated that two genera of 'Candidatus Kuenenia' and 'Candidatus Brocadia' and the family of Planctomycetaceae were identified. We suggest the rhizosphere provides a favorable niche for anammox bacteria, which are important to N cycling, but were previously largely overlooked.
NiuSL, ClassenAT, DukesJS, KardolP, LiuLL, LuoYQ, RustadL, SunJ, TangJW, TemplerPH, ThomasRQ, TianDS, ViccaS, WangYP, XiaJY, ZaehleS (2016). Global patterns and substrate-based mechanisms of the terrestrial nitrogen cycle Ecology Letters, 19, 697-709. DOI:10.1111/ele.12591URLPMID:26932540 [本文引用: 1] Nitrogen (N) deposition is impacting the services that ecosystems provide to humanity. However, the mechanisms determining impacts on the N cycle are not fully understood. To explore the mechanistic underpinnings of N impacts on N cycle processes, we reviewed and synthesised recent progress in ecosystem N research through empirical studies, conceptual analysis and model simulations. Experimental and observational studies have revealed that the stimulation of plant N uptake and soil retention generally diminishes as N loading increases, while dissolved and gaseous losses of N occur at low N availability but increase exponentially and become the dominant fate of N at high loading rates. The original N saturation hypothesis emphasises sequential N saturation from plant uptake to soil retention before N losses occur. However, biogeochemical models that simulate simultaneous competition for soil N substrates by multiple processes match the observed patterns of N losses better than models based on sequential competition. To enable better prediction of terrestrial N cycle responses to N loading, we recommend that future research identifies the response functions of different N processes to substrate availability using manipulative experiments, and incorporates the measured N saturation response functions into conceptual, theoretical and quantitative analyses.
PanYP, TianSL, LiuDW, FangYT, ZhuXY, ZhangQ, ZhengB, MichalskiG, WangYS (2016). Fossil fuel combustion-related emissions dominate atmospheric ammonia sources during severe haze episodes: evidence from 15N-stable isotope in size-resolved aerosol ammonium Environmental Science & Technology, 50, 8049-8056. DOI:10.1021/acs.est.6b00634URLPMID:27359161 [本文引用: 1] The reduction of ammonia (NH3) emissions is urgently needed due to its role in aerosol nucleation and growth causing haze formation during its conversion into ammonium (NH4(+)). However, the relative contributions of individual NH3 sources are unclear, and debate remains over whether agricultural emissions dominate atmospheric NH3 in urban areas. Based on the chemical and isotopic measurements of size-resolved aerosols in urban Beijing, China, we find that the natural abundance of (15)N (expressed using delta(15)N values) of NH4(+) in fine particles varies with the development of haze episodes, ranging from -37.1 per thousand to -21.7 per thousand during clean/dusty days (relative humidity: approximately 40%), to -13.1 per thousand to +5.8 per thousand during hazy days (relative humidity: 70-90%). After accounting for the isotope exchange between NH3 gas and aerosol NH4(+), the delta(15)N value of the initial NH3 during hazy days is found to be -14.5 per thousand to -1.6 per thousand, which indicates fossil fuel-based emissions. These emissions contribute 90% of the total NH3 during hazy days in urban Beijing. This work demonstrates the analysis of delta(15)N values of aerosol NH4(+) to be a promising new tool for partitioning atmospheric NH3 sources, providing policy makers with insights into NH3 emissions and secondary aerosols for regulation in urban environments.
QuanZ, LiSL, ZhuFF, ZhangLM, HeJZ, WeiWX, FangYT (2018). Fates of 15N-labeled fertilizer in a black soil-maize system and the response to straw incorporation in Northeast China Journal of Soils and Sediments, 18, 1441-1452. DOI:10.1007/s11368-017-1857-3URL [本文引用: 1]
Rimski-KorsakovH, RubioG, LavadoRS (2012). Fate of the nitrogen from fertilizers in field-grown maize Nutrient Cycling in Agroecosystems, 93, 253-263. [本文引用: 1]
RobinsonD (2001). δ15N as an integrator of the nitrogen cycle Trends in Ecology & Evolution, 16, 153-162. URLPMID:11179580 [本文引用: 3]
RuisiP, SaiaS, BadagliaccaG, AmatoG, FrendaAS, GiambalvoD, di MiceliG (2016). Long-term effects of no tillage treatment on soil N availability, N uptake, and 15N-fertilizer recovery of durum wheat differ in relation to crop sequence Field Crops Research, 189, 51-58. [本文引用: 1]
SebiloM, MayerB, NicolardotB, PinayG, MariottiA (2013). Long-term fate of nitrate fertilizer in agricultural soils Proceedings of the National Academy of Sciences of the United States of America, 110, 18185-18189. URLPMID:24145428 [本文引用: 3]
SigmanDM, CasciottiKL, AndreaniM, BarfordC, GalanterM, B?hlkeJK (2001). A bacterial method for the nitrogen isotopic analysis of nitrate in seawater and freshwater Analytical Chemistry, 73, 4145-4153. URLPMID:11569803 [本文引用: 1]
StevensRJ, LaughlinRJ (1998). Measurement of nitrous oxide and di-nitrogen emissions from agricultural soils Nutrient Cycling in Agroecosystems, 52, 131-139. [本文引用: 1]
TakebayashiY, KobaK, SasakiY, FangYT, YohM (2010). The natural abundance of 15N in plant and soil-available N indicates a shift of main plant N resources to NO3- from NH4+ along the N leaching gradient Rapid Communications in Mass Spectrometry, 24, 1001-1008. URLPMID:20213633 [本文引用: 1]
TemplerPH, MackMC, ChapinFS, ChristensonLM, ComptonJE, CrookHD, CurrieWS, CurtisCJ, DailDB, D’antonioCM, EmmettBA, EpsteinHE, GoodaleCL, GundersenP, HobbieSE, HollandK, HooperDU, HungateBA, LamontagneS, NadelhofferKJ, OsenbergCW, PerakisSS, SchleppiP, SchimelJ, SchmidtIK, SommerkornM, SpoelstraJ, TietemaA, WesselWW, ZakDR (2012). Sinks for nitrogen inputs in terrestrial ecosystems: a meta-analysis of 15N tracer field studies Ecology, 93, 1816-1829. DOI:10.1890/11-1146.1URLPMID:22928411 [本文引用: 3] Effects of anthropogenic nitrogen (N) deposition and the ability of terrestrial ecosystems to store carbon (C) depend in part on the amount of N retained in the system and its partitioning among plant and soil pools. We conducted a meta-analysis of studies at 48 sites across four continents that used enriched 15N isotope tracers in order to synthesize information about total ecosystem N retention (i.e., total ecosystem 15N recovery in plant and soil pools) across natural systems and N partitioning among ecosystem pools. The greatest recoveries of ecosystem 15N tracer occurred in shrublands (mean, 89.5%) and wetlands (84.8%) followed by forests (74.9%) and grasslands (51.8%). In the short term (< 1 week after 15N tracer application), total ecosystem 15N recovery was negatively correlated with fine-root and soil 15N natural abundance, and organic soil C and N concentration but was positively correlated with mean annual temperature and mineral soil C:N. In the longer term (3-18 months after 15N tracer application), total ecosystem 15N retention was negatively correlated with foliar natural-abundance 15N but was positively correlated with mineral soil C and N concentration and C:N, showing that plant and soil natural-abundance 15N and soil C:N are good indicators of total ecosystem N retention. Foliar N concentration was not significantly related to ecosystem 15N tracer recovery, suggesting that plant N status is not a good predictor of total ecosystem N retention. Because the largest ecosystem sinks for 15N tracer were below ground in forests, shrublands, and grasslands, we conclude that growth enhancement and potential for increased C storage in aboveground biomass from atmospheric N deposition is likely to be modest in these ecosystems. Total ecosystem 15N recovery decreased with N fertilization, with an apparent threshold fertilization rate of 46 kg N x ha(-1) x yr(-1) above which most ecosystems showed net losses of applied 15N tracer in response to N fertilizer addition.
TietemaA, EmmettBA, GundersenP, Kj?naasOJ, KoopmansCJ (1998). The fate of 15N-labelled nitrogen deposition in coniferous forest ecosystems Forest Ecology and Management, 101, 19-27. DOI:10.1016/S0378-1127(97)00123-0URL [本文引用: 1]
WangA, ZhuWX, GundersenP, PhillipsOL, ChenDX, FangYT (2018). Fates of atmospheric deposited nitrogen in an Asian tropical primary forest Forest Ecology and Management, 411, 213-222. [本文引用: 3]
WangSJ, LuoSS, YueSC, ShenYF, LiSQ (2016a). Fate of 15N fertilizer under different nitrogen split applications to plastic mulched maize in semiarid farmland Nutrient Cycling in Agroecosystems, 105, 129-140. [本文引用: 1]
WangXB, ZhouW, LiangGQ, PeiXX, LiKJ (2016b). The fate of 15N-labelled urea in an alkaline calcareous soil under different N application rates and N splits Nutrient Cycling in Agroecosystems, 106, 311-324. [本文引用: 1]
WangXT, SuoYY, FengY, ShohagMJI, GaoJ, ZhangQC, XieS, LinXY (2011). Recovery of 15N-labeled urea and soil nitrogen dynamics as affected by irrigation management and nitrogen application rate in a double rice cropping system Plant and Soil, 343, 195-208. [本文引用: 1]
XiD, BaiR, ZhangLM, FangYT (2016). Contribution of anammox to nitrogen removal in two temperate forest soils Applied and Environmental Microbiology, 82, 4602-4612. URLPMID:27208117 [本文引用: 1]
XuHG, ZhongGR, LinJJ, DingYF, LiGH, WangSH, LiuZH, TangS, DingCQ (2015). Effect of nitrogen management during the panicle stage in rice on the nitrogen utilization of rice and succeeding wheat crops European Journal of Agronomy, 70, 41-47. [本文引用: 1]
YangL, GuoS, ChenQW, ChenFJ, YuanLX, MiGH (2016). Use of the stable nitrogen isotope to reveal the source-sink regulation of nitrogen uptake and remobilization during grain filling phase in maize PLOS ONE, 11, e0162201. DOI: 10.1371/journal.pone.0162201. DOI:10.1371/journal.pone.0162201URLPMID:27606628 [本文引用: 1] Although the remobilization of vegetative nitrogen (N) and post-silking N both contribute to grain N in maize (Zea mays L.), their regulation by grain sink strength is poorly understood. Here we use 15N labeling to analyze the dynamic behaviors of both pre- and post-silking N in relation to source and sink manipulation in maize plants. The results showed that the remobilization of pre-silking N started immediately after silking and the remobilized pre-silking N had a greater contribution to grain N during early grain filling, with post-silking N importance increasing during the later filling stage. The amount of post-silking N uptake was largely driven by post-silking dry matter accumulation in both grain as well as vegetative organs. Prevention of pollination during silking had less effect on post-silking N uptake, as a consequence of compensatory growth of stems, husk + cob and roots. Also, leaves continuously export N even though grain sink was removed. The remobilization efficiency of N in the leaf and stem increased with increasing grain yield (hence N requirement). It is suggested that the remobilization of N in the leaf is controlled by sink strength but not the leaf per se. Enhancing post-silking N uptake rather than N remobilization is more likely to increase grain N accumulation.
YangWH, McDowellAC, BrooksPD, SilverWL (2014). New high precision approach for measuring 15N-N2 gas fluxes from terrestrial ecosystems Soil Biology & Biochemistry, 69, 234-241. [本文引用: 3]
YaoFY, ZhuB, DuEZ (2012). Use of 15N natural abundance in nitrogen cycling of terrestrial ecosystems Chinese Journal of Plant Ecology, 36, 346-352. [本文引用: 1]
ZhangL, AltabetMA, WuTX, HadasO (2007a). Sensitive measurement of NH4+15N/14N (δ15NH4+) at natural abundance levels in fresh and saltwaters Analytical Chemistry, 79, 5297-5303. URLPMID:17567102 [本文引用: 1]
ZhangL, WuZ, JiangY, ChenL, SongY, WangL, XieJ, MaX (2010). Fate of applied urea 15N in a soil-maize system as affected by urease inhibitor and nitrification inhibitor Plant, Soil & Environment, 56, 8-15. [本文引用: 1]
ZhangSS, FangYT, XiD (2015). Adaptation of microdiffusion method for the analysis of 15N natural abundance of ammonium in samples with small volume Rapid Communications in Mass Spectrometry, 29, 1297-1306. URLPMID:26405791 [本文引用: 1]
ZhangXD, AmelungW (1996). Gas chromatographic determination of muramic acid, glucosamine, mannosamine, and galactosamine in soils Soil Biology & Biochemistry, 28, 1201-1206. [本文引用: 1]
ZhangXD, HeHB, AmelungW (2007b). A GC/MS method for the assessment of 15N and 13C incorporation into soil amino acid enantiomers Soil Biology & Biochemistry, 39, 2785-2796. [本文引用: 1]
ZhangZL, LiN, XiaoJ, ZhaoCZ, ZouTT, LiDD, LiuQ, YinHJ (2018). Changes in plant nitrogen acquisition strategies during the restoration of spruce plantations on the eastern Tibetan Plateau, China Soil Biology & Biochemistry, 119, 50-58. DOI:10.1016/j.soilbio.2018.01.002URL [本文引用: 1]
ZhuFF, DaiLM, HobbieEA, KobaK, LiuXY, GurmesaGA, HuangSN, LiSL, LiYH, HanSJ, FangYT (2019). Uptake patterns of glycine, ammonium, and nitrate differ among four common tree species of northeast China Frontiers in Plant Science, 10, 799. DOI: 10.3389/fpls.2019.00799. URLPMID:31333684 [本文引用: 1]
ZhuGB, WangSY, WangY, WangCX, Risgaard-PetersenN, JettenMS, YinCQ (2011). Anaerobic ammonia oxidation in a fertilized paddy soil The ISME Journal, 5, 1905-1912. URLPMID:21593796 [本文引用: 1]
ZongZ, WangXP, TianCG, ChenYJ, FangYT, ZhangF, LiC, SunJZ, LiJ, ZhangG (2017). First assessment of NOx sources at a regional background site in North China using isotopic analysis linked with modeling Environmental Science & Technology, 51, 5923-5931. DOI:10.1021/acs.est.6b06316URLPMID:28516763 [本文引用: 1] Nitrogen oxides (NOx, including NO and NO2) play an important role in the formation of atmospheric particles. Thus, NOx emission reduction is critical for improving air quality, especially in severely air-polluted regions (e.g., North China). In this study, the source of NOx was investigated by the isotopic composition (delta(15)N) of particulate nitrate (p-NO3(-)) at Beihuangcheng Island (BH), a regional background site in North China. It was found that the delta(15)N-NO3(-) (n = 120) values varied between -1.7 per thousand and +24.0 per thousand and the delta(18)O-NO3(-) values ranged from 49.4 per thousand to 103.9 per thousand. On the basis of the Bayesian mixing model, 27.78 +/- 8.89%, 36.53 +/- 6.66%, 22.01 +/- 6.92%, and 13.68 +/- 3.16% of annual NOx could be attributed to biomass burning, coal combustion, mobile sources, and biogenic soil emissions, respectively. Seasonally, the four sources were similar in spring and fall. Biogenic soil emissions were augmented in summer in association with the hot and rainy weather. Coal combustion increased significantly in winter with other sources showing an obvious decline. This study confirmed that isotope-modeling by delta(15)N-NO3(-) is a promising tool for partitioning NOx sources and provides guidance to policymakers with regard to options for NOx reduction in North China. Global estimates of inorganic nitrogen deposition across four decades 2019
Coupled isotopic and process-based modeling of gaseous nitrogen losses from tropical rain forests 1 2009
... 然而, 利用氮稳定同位素研究氮循环过程, 必须考虑氮循环各个过程的氮同位素分馏效应(图1).例如, 利用近十年来发展的生态系统氮循环同位素模型和硝酸盐氮氧同位素自然丰度法量化生态系统反硝化速率时, 必须考虑反硝化作用过程中氮同位素分馏效应(Houlton et al., 2006; Houlton & Bai, 2009; Bai et al., 2012; Fang et al., 2015); 利用植物氮同位素研究植物氮吸收策略时, 必须考虑植物氮吸收以及植物内部的氮转移过程中的氮同位素分馏效应(Houlton et al., 2007).目前国内外对于氮同位素分馏效应缺乏系统性的研究, 使得氮循环过程中的氮同位素分馏效应存在很大的不确定性, 如厌氧氨氧化过程(Brunner et al., 2013)和矿化过程(M?bius, 2013; Zhang et al., 2015)的同位素分馏效应研究较少, DNRA过程中的同位素分馏效应尚无报道, 而反硝化过程的同位素分馏效应则随实验条件和计算方法的不同呈现较大的变异性(Denk et al., 2017).因此, 在未来对于氮循环的研究中, 应重点考虑各个过程中氮同位素分馏效应的研究, 而进一步将含氮化合物的氮同位素组成、同位素分馏效应与生物地球化学模型相结合, 也会加深我们对氮循环的理解, 并为这些模型的验证提供综合约束(Bai & Houlton, 2009; Houlton et al., 2015; Denk et al., 2017). ...
Isotopic identification of nitrogen hotspots across natural terrestrial ecosystems 2 2012
... 目前氮循环研究中不确定性最大的过程是生态系统气态氮损失(Niu et al., 2016).反硝化作用是造成气态氮损失的最主要途径.以往量化反硝化速率的方法有乙炔抑制法和15N标记法, 但由于空间异质性无法应用到生态系统尺度(Groffman et al., 2006).Houlton等(2006)建立的概念模型认为土壤总氮的15N受到氮沉降、氮淋失和气态氮损失的影响, 通过同位素混合模型可以量化生态系统尺度反硝化作用速率(Houlton et al., 2006; Houlton & Bai, 2009).但是, 由于土壤总氮库是长期积累的结果, 极其稳定, 因此土壤总氮的15N自然丰度法反映的是生态系统尺度反硝化作用在长时间上的平均水平(Houlton et al., 2006; Houlton & Bai, 2009; Bai et al., 2012).随后, Fang等(2015)提出利用硝酸盐氮氧同位素自然丰度技术(δ15N、δ18O和Δ17O), 结合传统的氮沉降和氮流失的监测, 再根据质量平衡和同位素质量平衡原理来量化生态系统尺度硝化和反硝化作用速率, 该方法的优势在于可以同时量化不同时间段的硝化和反硝化速率. ...
... 然而, 利用氮稳定同位素研究氮循环过程, 必须考虑氮循环各个过程的氮同位素分馏效应(图1).例如, 利用近十年来发展的生态系统氮循环同位素模型和硝酸盐氮氧同位素自然丰度法量化生态系统反硝化速率时, 必须考虑反硝化作用过程中氮同位素分馏效应(Houlton et al., 2006; Houlton & Bai, 2009; Bai et al., 2012; Fang et al., 2015); 利用植物氮同位素研究植物氮吸收策略时, 必须考虑植物氮吸收以及植物内部的氮转移过程中的氮同位素分馏效应(Houlton et al., 2007).目前国内外对于氮同位素分馏效应缺乏系统性的研究, 使得氮循环过程中的氮同位素分馏效应存在很大的不确定性, 如厌氧氨氧化过程(Brunner et al., 2013)和矿化过程(M?bius, 2013; Zhang et al., 2015)的同位素分馏效应研究较少, DNRA过程中的同位素分馏效应尚无报道, 而反硝化过程的同位素分馏效应则随实验条件和计算方法的不同呈现较大的变异性(Denk et al., 2017).因此, 在未来对于氮循环的研究中, 应重点考虑各个过程中氮同位素分馏效应的研究, 而进一步将含氮化合物的氮同位素组成、同位素分馏效应与生物地球化学模型相结合, 也会加深我们对氮循环的理解, 并为这些模型的验证提供综合约束(Bai & Houlton, 2009; Houlton et al., 2015; Denk et al., 2017). ...
Retention of 15N-labeled fertilizer in an Illinois prairie soil with winter rye 1 2014
... 在原位小区试验条件下, 研究者们主要是通过15N脉冲标记来模拟氮肥施用, 从而追踪土壤中肥料氮的流向和命运.由于农田生态系统结构单一、人为活动影响大, 田间原位15N的去向很不稳定, 变异较大(Gardner & Drinkwater, 2009).过去和目前的大部分研究主要探讨农艺措施对肥料氮去向的影响, 相关措施包括施用增效肥料(Zhang et al., 2010; Chalk et al., 2015), 优化施氮方式或时间(Wang et al., 2016a; de Oliveira et al., 2018), 优化施肥量(Rimski-Korsakov et al., 2012; Chen et al., 2016; Wang et al., 2016b), 少免耕(Ruisi et al., 2016), 秸秆还田(Gabriel et al., 2016; Murphy et al., 2016; Quan et al., 2018), 覆膜灌水(Wang et al., 2011; Liu et al., 2015), 轮间作(Xu et al., 2015), 作物品种(Yang et al., 2016), 耕作历史(Liang et al., 2013)等.采用15N示踪法计算出来的作物回收率通常只是肥料氮的当季利用效率, 真实评价氮肥的供氮功能还需要考虑其残留效应.在土壤氮平衡条件下, 残留效应的本质就是当季残留肥料氮对当季土壤氮消耗的补偿, 可以理解为肥料氮的间接供氮效应.该效应被认为是维持土壤长久供氮能力的重要机制(Ju & Christie, 2011; 巨晓棠, 2014).残留肥料氮的后季利用可通过15N示踪定量, Ladha等(2005)总结了前人多季15N示踪研究的结果, 发现当季残留肥料氮在随后五季作物的累积利用率平均仅在6.5%左右(逐年平均分别为3.3%、1.3%、1.0%、0.4%和0.5%).但不能由此就认为残留肥料氮的后季利用就很低, 因为绝大多数的15N示踪结果均显示作物吸收的氮主要还是来自土壤氮库, 肥料氮当季的贡献只占37%-44% (Gardner & Drinkwater, 2009; Blesh & Drinkwater, 2014).为了证明肥料氮的长期效应, Sebilo等(2013)研究了硝态氮肥施入农田土壤后30年的利用和损失情况, 他发现肥料氮的利用和损失是长期的过程, 30年连续作物种植仍有12%-15%的肥料氮残留在土壤中, 第一季肥料氮30年的累积利用率达到61%-65%. ...
Nitrogen isotope effects induced by anammox bacteria 1 2013
... 然而, 利用氮稳定同位素研究氮循环过程, 必须考虑氮循环各个过程的氮同位素分馏效应(图1).例如, 利用近十年来发展的生态系统氮循环同位素模型和硝酸盐氮氧同位素自然丰度法量化生态系统反硝化速率时, 必须考虑反硝化作用过程中氮同位素分馏效应(Houlton et al., 2006; Houlton & Bai, 2009; Bai et al., 2012; Fang et al., 2015); 利用植物氮同位素研究植物氮吸收策略时, 必须考虑植物氮吸收以及植物内部的氮转移过程中的氮同位素分馏效应(Houlton et al., 2007).目前国内外对于氮同位素分馏效应缺乏系统性的研究, 使得氮循环过程中的氮同位素分馏效应存在很大的不确定性, 如厌氧氨氧化过程(Brunner et al., 2013)和矿化过程(M?bius, 2013; Zhang et al., 2015)的同位素分馏效应研究较少, DNRA过程中的同位素分馏效应尚无报道, 而反硝化过程的同位素分馏效应则随实验条件和计算方法的不同呈现较大的变异性(Denk et al., 2017).因此, 在未来对于氮循环的研究中, 应重点考虑各个过程中氮同位素分馏效应的研究, 而进一步将含氮化合物的氮同位素组成、同位素分馏效应与生物地球化学模型相结合, 也会加深我们对氮循环的理解, 并为这些模型的验证提供综合约束(Bai & Houlton, 2009; Houlton et al., 2015; Denk et al., 2017). ...
Diffusion of soil extracts for nitrogen and nitrogen-15 analyses by automated combustion/mass spectrometry 1 1990
... 氮稳定同位素技术在研究生态系统氮循环中有比较长的应用历史, 早在20世纪50年代, Hoering就发表了首篇关于自然物质15N自然丰度的论文(Hoering, 1955).根据分馏方法和分析的气体类型不同, 同位素测定方法的发展大致经历了三个时期.20世纪50年代至80年代, 科学家主要利用碱性次溴酸盐(Rittenberg)氧化法将离子转化成N2, 20世纪80年代至20世纪末期, 科学家利用杜马斯(Dumas)燃烧法把固体样品转化成N2, 实现氮稳定同位素在线测定(Burke et al., 1990), 该方法也沿用至今.自21世纪开始, 科学家进一步发展了测定可溶性有机和无机氮同位素的快速分析技术, 成功通过化学或者生物的方法将溶液中的含氮物质直接转化成N2O, 再利用商业化的气体吹扫捕集系统连接稳定同位素质谱仪进行在线同位素测定(Sigman et al., 2001; McIlvin & Altabet, 2005; Zhang et al., 2007a; Koba et al., 2010; Liu et al., 2017a).新技术对样品的需求是传统方法的1/100甚至1/1 000, 因此适合样品量少和氮含量低的样品, 也使得氮循环的研究从海洋和陆地生态系统扩展到大气和冰芯的研究中(Hastings, 2010; Huang et al., 2019). ...
Fate and efficiency of 15N-labelled slow- and controlled-release fertilizers 1 2015
... 在原位小区试验条件下, 研究者们主要是通过15N脉冲标记来模拟氮肥施用, 从而追踪土壤中肥料氮的流向和命运.由于农田生态系统结构单一、人为活动影响大, 田间原位15N的去向很不稳定, 变异较大(Gardner & Drinkwater, 2009).过去和目前的大部分研究主要探讨农艺措施对肥料氮去向的影响, 相关措施包括施用增效肥料(Zhang et al., 2010; Chalk et al., 2015), 优化施氮方式或时间(Wang et al., 2016a; de Oliveira et al., 2018), 优化施肥量(Rimski-Korsakov et al., 2012; Chen et al., 2016; Wang et al., 2016b), 少免耕(Ruisi et al., 2016), 秸秆还田(Gabriel et al., 2016; Murphy et al., 2016; Quan et al., 2018), 覆膜灌水(Wang et al., 2011; Liu et al., 2015), 轮间作(Xu et al., 2015), 作物品种(Yang et al., 2016), 耕作历史(Liang et al., 2013)等.采用15N示踪法计算出来的作物回收率通常只是肥料氮的当季利用效率, 真实评价氮肥的供氮功能还需要考虑其残留效应.在土壤氮平衡条件下, 残留效应的本质就是当季残留肥料氮对当季土壤氮消耗的补偿, 可以理解为肥料氮的间接供氮效应.该效应被认为是维持土壤长久供氮能力的重要机制(Ju & Christie, 2011; 巨晓棠, 2014).残留肥料氮的后季利用可通过15N示踪定量, Ladha等(2005)总结了前人多季15N示踪研究的结果, 发现当季残留肥料氮在随后五季作物的累积利用率平均仅在6.5%左右(逐年平均分别为3.3%、1.3%、1.0%、0.4%和0.5%).但不能由此就认为残留肥料氮的后季利用就很低, 因为绝大多数的15N示踪结果均显示作物吸收的氮主要还是来自土壤氮库, 肥料氮当季的贡献只占37%-44% (Gardner & Drinkwater, 2009; Blesh & Drinkwater, 2014).为了证明肥料氮的长期效应, Sebilo等(2013)研究了硝态氮肥施入农田土壤后30年的利用和损失情况, 他发现肥料氮的利用和损失是长期的过程, 30年连续作物种植仍有12%-15%的肥料氮残留在土壤中, 第一季肥料氮30年的累积利用率达到61%-65%. ...
The fates of 15N-labeled fertilizer in a wheat-soil system as influenced by fertilization practice in a loamy soil 1 2016
... 在原位小区试验条件下, 研究者们主要是通过15N脉冲标记来模拟氮肥施用, 从而追踪土壤中肥料氮的流向和命运.由于农田生态系统结构单一、人为活动影响大, 田间原位15N的去向很不稳定, 变异较大(Gardner & Drinkwater, 2009).过去和目前的大部分研究主要探讨农艺措施对肥料氮去向的影响, 相关措施包括施用增效肥料(Zhang et al., 2010; Chalk et al., 2015), 优化施氮方式或时间(Wang et al., 2016a; de Oliveira et al., 2018), 优化施肥量(Rimski-Korsakov et al., 2012; Chen et al., 2016; Wang et al., 2016b), 少免耕(Ruisi et al., 2016), 秸秆还田(Gabriel et al., 2016; Murphy et al., 2016; Quan et al., 2018), 覆膜灌水(Wang et al., 2011; Liu et al., 2015), 轮间作(Xu et al., 2015), 作物品种(Yang et al., 2016), 耕作历史(Liang et al., 2013)等.采用15N示踪法计算出来的作物回收率通常只是肥料氮的当季利用效率, 真实评价氮肥的供氮功能还需要考虑其残留效应.在土壤氮平衡条件下, 残留效应的本质就是当季残留肥料氮对当季土壤氮消耗的补偿, 可以理解为肥料氮的间接供氮效应.该效应被认为是维持土壤长久供氮能力的重要机制(Ju & Christie, 2011; 巨晓棠, 2014).残留肥料氮的后季利用可通过15N示踪定量, Ladha等(2005)总结了前人多季15N示踪研究的结果, 发现当季残留肥料氮在随后五季作物的累积利用率平均仅在6.5%左右(逐年平均分别为3.3%、1.3%、1.0%、0.4%和0.5%).但不能由此就认为残留肥料氮的后季利用就很低, 因为绝大多数的15N示踪结果均显示作物吸收的氮主要还是来自土壤氮库, 肥料氮当季的贡献只占37%-44% (Gardner & Drinkwater, 2009; Blesh & Drinkwater, 2014).为了证明肥料氮的长期效应, Sebilo等(2013)研究了硝态氮肥施入农田土壤后30年的利用和损失情况, 他发现肥料氮的利用和损失是长期的过程, 30年连续作物种植仍有12%-15%的肥料氮残留在土壤中, 第一季肥料氮30年的累积利用率达到61%-65%. ...
Relationships between C and N availability, substrate age, and natural abundance 13C and 15N signatures of soil microbial biomass in a semiarid climate 1 2009
Global patterns of foliar nitrogen isotopes and their relationships with climate, mycorrhizal fungi, foliar nutrient concentrations, and nitrogen availability 2 2009
Understanding N timing in corn yield and fertilizer N recovery: an insight from an isotopic labeled-N determination 1 2018
... 在原位小区试验条件下, 研究者们主要是通过15N脉冲标记来模拟氮肥施用, 从而追踪土壤中肥料氮的流向和命运.由于农田生态系统结构单一、人为活动影响大, 田间原位15N的去向很不稳定, 变异较大(Gardner & Drinkwater, 2009).过去和目前的大部分研究主要探讨农艺措施对肥料氮去向的影响, 相关措施包括施用增效肥料(Zhang et al., 2010; Chalk et al., 2015), 优化施氮方式或时间(Wang et al., 2016a; de Oliveira et al., 2018), 优化施肥量(Rimski-Korsakov et al., 2012; Chen et al., 2016; Wang et al., 2016b), 少免耕(Ruisi et al., 2016), 秸秆还田(Gabriel et al., 2016; Murphy et al., 2016; Quan et al., 2018), 覆膜灌水(Wang et al., 2011; Liu et al., 2015), 轮间作(Xu et al., 2015), 作物品种(Yang et al., 2016), 耕作历史(Liang et al., 2013)等.采用15N示踪法计算出来的作物回收率通常只是肥料氮的当季利用效率, 真实评价氮肥的供氮功能还需要考虑其残留效应.在土壤氮平衡条件下, 残留效应的本质就是当季残留肥料氮对当季土壤氮消耗的补偿, 可以理解为肥料氮的间接供氮效应.该效应被认为是维持土壤长久供氮能力的重要机制(Ju & Christie, 2011; 巨晓棠, 2014).残留肥料氮的后季利用可通过15N示踪定量, Ladha等(2005)总结了前人多季15N示踪研究的结果, 发现当季残留肥料氮在随后五季作物的累积利用率平均仅在6.5%左右(逐年平均分别为3.3%、1.3%、1.0%、0.4%和0.5%).但不能由此就认为残留肥料氮的后季利用就很低, 因为绝大多数的15N示踪结果均显示作物吸收的氮主要还是来自土壤氮库, 肥料氮当季的贡献只占37%-44% (Gardner & Drinkwater, 2009; Blesh & Drinkwater, 2014).为了证明肥料氮的长期效应, Sebilo等(2013)研究了硝态氮肥施入农田土壤后30年的利用和损失情况, 他发现肥料氮的利用和损失是长期的过程, 30年连续作物种植仍有12%-15%的肥料氮残留在土壤中, 第一季肥料氮30年的累积利用率达到61%-65%. ...
The nitrogen cycle: a review of isotope effects and isotope modeling approaches 4 2017
氮循环过程中的同位素分馏效应(<sup>15</sup>ε, ‰, 修改自<xref ref-type="bibr" rid="b14">Denk <i>et al</i>., 2017</xref>).英文字母代表生态系统氮循环过程: a, 矿化作用; b,硝化作用; c, 反硝化作用; d, 生物固氮; e, NH<sub>3</sub>和NH<sub>4</sub><sup>+</sup>同位素交换; f, 硝酸盐异化还原成铵作用; g, 微生物固定; h, 植物吸收; i, 厌氧氨氧化作用; j, NH<sub>3</sub>挥发.
Isotope fractionation effects in nitrogen processes (<sup>15</sup>ε, ‰, modified by <xref ref-type="bibr" rid="b14">Denk <i>et al</i>., 2017</xref>). Processes involved in terrestrial ecosystem N cycling are: a, mineralization; b, nitrification; c, denitrification; d, biological N<sub>2</sub> fixation; e, isotope exchange between NH<sub>3</sub> and NH<sub>4</sub><sup>+</sup>; f, dissimilatory nitrate reduction to ammonium; g, immobilization by microbes; h, plant uptake; i, anaerobic ammonia oxidation; j, NH<sub>3</sub> volatilization. SOM, soil organic matter.Fig. 11.2 <sup>15</sup>N标记技术15N标记技术是将一定数量的15N富集的含氮化合物添加到系统中(植物、土壤或整个生态系统), 经过一段时间后分析其去向(Templer et al., 2012).在生态系统氮循环研究方面, 15N标记技术不受稳定同位素分馏效应的影响, 具有15N自然丰度法和传统示踪法所不具备的优点.但15N标记技术的最大不足之处是标记药品及样品分析费用较高, 因而限制其广泛应用.此外, 实验中还应考虑标记样品的均匀性、标记期长短和条件、示踪期长短和各库转化, 同时注意追踪的各个库要全面等问题.近年来, 随着稳定同位素技术的不断革新, 氮同位素标记物和仪器检测成本的下降, 15N标记技术已应用于陆地生态系统尺度上的各个方面, 如分析氮的来源、分配和去向、土壤氮的转化以及氮损失等研究(He et al., 2011; Yang et al., 2014; Li et al., 2019; Zhu et al., 2019).下文以陆地生态系统中森林生态系统(自然生态系统)和农田生态系统(人工生态系统)为例, 介绍近年来15N标记技术在生态系统尺度上的应用. ...
... ε, ‰, modified by Denk et al., 2017). Processes involved in terrestrial ecosystem N cycling are: a, mineralization; b, nitrification; c, denitrification; d, biological N2 fixation; e, isotope exchange between NH3 and NH4+; f, dissimilatory nitrate reduction to ammonium; g, immobilization by microbes; h, plant uptake; i, anaerobic ammonia oxidation; j, NH3 volatilization. SOM, soil organic matter. Fig. 11.2 <sup>15</sup>N标记技术15N标记技术是将一定数量的15N富集的含氮化合物添加到系统中(植物、土壤或整个生态系统), 经过一段时间后分析其去向(Templer et al., 2012).在生态系统氮循环研究方面, 15N标记技术不受稳定同位素分馏效应的影响, 具有15N自然丰度法和传统示踪法所不具备的优点.但15N标记技术的最大不足之处是标记药品及样品分析费用较高, 因而限制其广泛应用.此外, 实验中还应考虑标记样品的均匀性、标记期长短和条件、示踪期长短和各库转化, 同时注意追踪的各个库要全面等问题.近年来, 随着稳定同位素技术的不断革新, 氮同位素标记物和仪器检测成本的下降, 15N标记技术已应用于陆地生态系统尺度上的各个方面, 如分析氮的来源、分配和去向、土壤氮的转化以及氮损失等研究(He et al., 2011; Yang et al., 2014; Li et al., 2019; Zhu et al., 2019).下文以陆地生态系统中森林生态系统(自然生态系统)和农田生态系统(人工生态系统)为例, 介绍近年来15N标记技术在生态系统尺度上的应用. ...
... 然而, 利用氮稳定同位素研究氮循环过程, 必须考虑氮循环各个过程的氮同位素分馏效应(图1).例如, 利用近十年来发展的生态系统氮循环同位素模型和硝酸盐氮氧同位素自然丰度法量化生态系统反硝化速率时, 必须考虑反硝化作用过程中氮同位素分馏效应(Houlton et al., 2006; Houlton & Bai, 2009; Bai et al., 2012; Fang et al., 2015); 利用植物氮同位素研究植物氮吸收策略时, 必须考虑植物氮吸收以及植物内部的氮转移过程中的氮同位素分馏效应(Houlton et al., 2007).目前国内外对于氮同位素分馏效应缺乏系统性的研究, 使得氮循环过程中的氮同位素分馏效应存在很大的不确定性, 如厌氧氨氧化过程(Brunner et al., 2013)和矿化过程(M?bius, 2013; Zhang et al., 2015)的同位素分馏效应研究较少, DNRA过程中的同位素分馏效应尚无报道, 而反硝化过程的同位素分馏效应则随实验条件和计算方法的不同呈现较大的变异性(Denk et al., 2017).因此, 在未来对于氮循环的研究中, 应重点考虑各个过程中氮同位素分馏效应的研究, 而进一步将含氮化合物的氮同位素组成、同位素分馏效应与生物地球化学模型相结合, 也会加深我们对氮循环的理解, 并为这些模型的验证提供综合约束(Bai & Houlton, 2009; Houlton et al., 2015; Denk et al., 2017). ...
... ; Denk et al., 2017). ...
13C and 15N natural abundance of the soil microbial biomass 2 2006
Nitrogen loss through anaerobic ammonium oxidation coupled to iron reduction from paddy soils in a chronosequence 1 2014
... 在实验室培养条件下, 研究者们主要通过15N的生物标志物和同位素模型来研究土壤氮的转化动态.其中, Zhang和Amelung (1996)、Zhang等(2007b)和He等(2006, 2011)将同位素示踪技术和土壤功能组分相结合, 建立了测定土壤氨基糖和氨基酸转化与更新的GC-MS和HPLC-MS方法, 并成功应用该方法区分了不同微生物对土壤有机碳氮转化的贡献.Mary等(1998)、Müller等(2007)通过NH4NO3-15N交叉标记技术, 发展了定量土壤氮转化过程的数值计算模型, 成功模拟出土壤中矿化、固持、再矿化、自养硝化、异养硝化、反硝化、硝酸盐异化还原成铵(DNRA)等多个过程的转化速率.通过15N同位素质量守恒和数值模型, Stevens和Laughlin (1998)、Kool等(2011)、Yang等(2014)、Müller等(2014)则建立了区分土壤释放N2O和N2微生物途径的新方法.此外, 还有研究者将同位素示踪技术与现代分子生物学技术相结合, 来研究更为复杂的微生物氮转化过程, 例如厌氧氨氧化(Zhu et al., 2011; Ding et al., 2014; Nie et al., 2015). ...
Microbial denitrification dominates nitrate losses from forest ecosystems 3 2015
... 目前氮循环研究中不确定性最大的过程是生态系统气态氮损失(Niu et al., 2016).反硝化作用是造成气态氮损失的最主要途径.以往量化反硝化速率的方法有乙炔抑制法和15N标记法, 但由于空间异质性无法应用到生态系统尺度(Groffman et al., 2006).Houlton等(2006)建立的概念模型认为土壤总氮的15N受到氮沉降、氮淋失和气态氮损失的影响, 通过同位素混合模型可以量化生态系统尺度反硝化作用速率(Houlton et al., 2006; Houlton & Bai, 2009).但是, 由于土壤总氮库是长期积累的结果, 极其稳定, 因此土壤总氮的15N自然丰度法反映的是生态系统尺度反硝化作用在长时间上的平均水平(Houlton et al., 2006; Houlton & Bai, 2009; Bai et al., 2012).随后, Fang等(2015)提出利用硝酸盐氮氧同位素自然丰度技术(δ15N、δ18O和Δ17O), 结合传统的氮沉降和氮流失的监测, 再根据质量平衡和同位素质量平衡原理来量化生态系统尺度硝化和反硝化作用速率, 该方法的优势在于可以同时量化不同时间段的硝化和反硝化速率. ...
... 氮稳定同位素自然丰度法被证明是研究氮循环过程的有效手段(Robinson, 2001).由于氮循环的物理、化学和生物过程均伴随氮的同位素分馏, 使生态系统不同含氮化合物的氮同位素组成(15N/14N)呈现动态变化, 因此氮稳定同位素自然丰度被用于指示生态系统中氮的来源和周转过程, 如分析植物生长策略(Houlton et al., 2007)、追踪NOx和NH3的排放源 (Pan et al., 2016; Zong et al., 2017)和量化生态系统尺度上的反硝化速率(Fang et al., 2015). ...
... 然而, 利用氮稳定同位素研究氮循环过程, 必须考虑氮循环各个过程的氮同位素分馏效应(图1).例如, 利用近十年来发展的生态系统氮循环同位素模型和硝酸盐氮氧同位素自然丰度法量化生态系统反硝化速率时, 必须考虑反硝化作用过程中氮同位素分馏效应(Houlton et al., 2006; Houlton & Bai, 2009; Bai et al., 2012; Fang et al., 2015); 利用植物氮同位素研究植物氮吸收策略时, 必须考虑植物氮吸收以及植物内部的氮转移过程中的氮同位素分馏效应(Houlton et al., 2007).目前国内外对于氮同位素分馏效应缺乏系统性的研究, 使得氮循环过程中的氮同位素分馏效应存在很大的不确定性, 如厌氧氨氧化过程(Brunner et al., 2013)和矿化过程(M?bius, 2013; Zhang et al., 2015)的同位素分馏效应研究较少, DNRA过程中的同位素分馏效应尚无报道, 而反硝化过程的同位素分馏效应则随实验条件和计算方法的不同呈现较大的变异性(Denk et al., 2017).因此, 在未来对于氮循环的研究中, 应重点考虑各个过程中氮同位素分馏效应的研究, 而进一步将含氮化合物的氮同位素组成、同位素分馏效应与生物地球化学模型相结合, 也会加深我们对氮循环的理解, 并为这些模型的验证提供综合约束(Bai & Houlton, 2009; Houlton et al., 2015; Denk et al., 2017). ...
Tracing the fate of mineral N compounds under high ambient N deposition in a Norway spruce forest at Solling/Germany 1 2008
... 近些年来, 通过配对15N标记(15NH4+和15NO3-), 越来越多的研究发现, 不仅植物和土壤对于两种沉降氮的吸收和滞留率差异明显, 而且不同类型乔木植物对两种形态氮的吸收利用效率也明显不同(Liu et al., 2017b; Goodale, 2017; Wang et al., 2018).在温带森林, 地表凋落物层对15NH4+的吸收固持率更高, 而对15NO3-吸收固持率明显更低, 导致其在表层矿物质土壤滞留率更高(Feng et al., 2008; Liu et al., 2017b).而在凋落物层较薄的亚热带和热带森林, 无论是地表凋落物层还是矿物质层土壤, 都更偏向吸收固持15NH4+ (Liu et al., 2017c; Wang et al., 2018).目前, 在同一森林的铵态氮和硝态氮配对的实验较少, 但几乎所有的配对标记结果都显示, 地上植物对15NO3-的总吸收固持率明显比15NH4+更高.其主要原因被认为是沉降氮进入森林生态系统后, 15NH4+更易于被土壤中的微生物固持, 以及发生无机过程的吸附或置换, 更难以到达植物根系被根系吸收.Liu等(2017b)在吉林长白山的研究发现, 与针叶树种相比, 阔叶树种对15NO3-的吸收利用速率更快.Goodale (2017)在美国纽约州Arnot森林的研究发现, 侵染丛枝菌根真菌(AM)的树种比侵染内生菌根真菌(ECM)的树种更偏向利用NO3-.Li等(2019)在辽宁清原次生林的研究也表明, AM树种对标记的15NO3-的吸收利用速率明显快于ECM树种.其中, 蒙古栎(Quercus mongolica)作为阔叶的ECM树种, 也符合这一规律.另外, 通过15N标记后不同时间序列的研究结果表明, 沉降氮进入森林生态系统后, 其损失主要发生在第一个生长季内.在标记后的1-7年的时间内, 地表凋落物层和表层矿物质土壤中固持的沉降氮, 逐渐向更深层土壤和植物层迁移, 而通过气体释放或淋溶过程损失的比例很小, 能够较长时间地滞留在整个生态系统内(Nadelhoffer et al., 2004; Goodale, 2017; Liu et al., 2017b; Wang et al., 2018; Li et al., 2019). ...
Stable Isotope Ecology. 2006
Nitrogen use efficiency and fertiliser fate in a long-term experiment with winter cover crops 1 2016
... 在原位小区试验条件下, 研究者们主要是通过15N脉冲标记来模拟氮肥施用, 从而追踪土壤中肥料氮的流向和命运.由于农田生态系统结构单一、人为活动影响大, 田间原位15N的去向很不稳定, 变异较大(Gardner & Drinkwater, 2009).过去和目前的大部分研究主要探讨农艺措施对肥料氮去向的影响, 相关措施包括施用增效肥料(Zhang et al., 2010; Chalk et al., 2015), 优化施氮方式或时间(Wang et al., 2016a; de Oliveira et al., 2018), 优化施肥量(Rimski-Korsakov et al., 2012; Chen et al., 2016; Wang et al., 2016b), 少免耕(Ruisi et al., 2016), 秸秆还田(Gabriel et al., 2016; Murphy et al., 2016; Quan et al., 2018), 覆膜灌水(Wang et al., 2011; Liu et al., 2015), 轮间作(Xu et al., 2015), 作物品种(Yang et al., 2016), 耕作历史(Liang et al., 2013)等.采用15N示踪法计算出来的作物回收率通常只是肥料氮的当季利用效率, 真实评价氮肥的供氮功能还需要考虑其残留效应.在土壤氮平衡条件下, 残留效应的本质就是当季残留肥料氮对当季土壤氮消耗的补偿, 可以理解为肥料氮的间接供氮效应.该效应被认为是维持土壤长久供氮能力的重要机制(Ju & Christie, 2011; 巨晓棠, 2014).残留肥料氮的后季利用可通过15N示踪定量, Ladha等(2005)总结了前人多季15N示踪研究的结果, 发现当季残留肥料氮在随后五季作物的累积利用率平均仅在6.5%左右(逐年平均分别为3.3%、1.3%、1.0%、0.4%和0.5%).但不能由此就认为残留肥料氮的后季利用就很低, 因为绝大多数的15N示踪结果均显示作物吸收的氮主要还是来自土壤氮库, 肥料氮当季的贡献只占37%-44% (Gardner & Drinkwater, 2009; Blesh & Drinkwater, 2014).为了证明肥料氮的长期效应, Sebilo等(2013)研究了硝态氮肥施入农田土壤后30年的利用和损失情况, 他发现肥料氮的利用和损失是长期的过程, 30年连续作物种植仍有12%-15%的肥料氮残留在土壤中, 第一季肥料氮30年的累积利用率达到61%-65%. ...
Nitrogen cycles: past, present, and future 2004
The fate of nitrogen in grain cropping systems: a meta-analysis of 15N field experiments 2 2009
... 在原位小区试验条件下, 研究者们主要是通过15N脉冲标记来模拟氮肥施用, 从而追踪土壤中肥料氮的流向和命运.由于农田生态系统结构单一、人为活动影响大, 田间原位15N的去向很不稳定, 变异较大(Gardner & Drinkwater, 2009).过去和目前的大部分研究主要探讨农艺措施对肥料氮去向的影响, 相关措施包括施用增效肥料(Zhang et al., 2010; Chalk et al., 2015), 优化施氮方式或时间(Wang et al., 2016a; de Oliveira et al., 2018), 优化施肥量(Rimski-Korsakov et al., 2012; Chen et al., 2016; Wang et al., 2016b), 少免耕(Ruisi et al., 2016), 秸秆还田(Gabriel et al., 2016; Murphy et al., 2016; Quan et al., 2018), 覆膜灌水(Wang et al., 2011; Liu et al., 2015), 轮间作(Xu et al., 2015), 作物品种(Yang et al., 2016), 耕作历史(Liang et al., 2013)等.采用15N示踪法计算出来的作物回收率通常只是肥料氮的当季利用效率, 真实评价氮肥的供氮功能还需要考虑其残留效应.在土壤氮平衡条件下, 残留效应的本质就是当季残留肥料氮对当季土壤氮消耗的补偿, 可以理解为肥料氮的间接供氮效应.该效应被认为是维持土壤长久供氮能力的重要机制(Ju & Christie, 2011; 巨晓棠, 2014).残留肥料氮的后季利用可通过15N示踪定量, Ladha等(2005)总结了前人多季15N示踪研究的结果, 发现当季残留肥料氮在随后五季作物的累积利用率平均仅在6.5%左右(逐年平均分别为3.3%、1.3%、1.0%、0.4%和0.5%).但不能由此就认为残留肥料氮的后季利用就很低, 因为绝大多数的15N示踪结果均显示作物吸收的氮主要还是来自土壤氮库, 肥料氮当季的贡献只占37%-44% (Gardner & Drinkwater, 2009; Blesh & Drinkwater, 2014).为了证明肥料氮的长期效应, Sebilo等(2013)研究了硝态氮肥施入农田土壤后30年的利用和损失情况, 他发现肥料氮的利用和损失是长期的过程, 30年连续作物种植仍有12%-15%的肥料氮残留在土壤中, 第一季肥料氮30年的累积利用率达到61%-65%. ...
Multiyear fate of a 15N tracer in a mixed deciduous forest: retention, redistribution, and differences by mycorrhizal association 4 2017
... 近些年来, 通过配对15N标记(15NH4+和15NO3-), 越来越多的研究发现, 不仅植物和土壤对于两种沉降氮的吸收和滞留率差异明显, 而且不同类型乔木植物对两种形态氮的吸收利用效率也明显不同(Liu et al., 2017b; Goodale, 2017; Wang et al., 2018).在温带森林, 地表凋落物层对15NH4+的吸收固持率更高, 而对15NO3-吸收固持率明显更低, 导致其在表层矿物质土壤滞留率更高(Feng et al., 2008; Liu et al., 2017b).而在凋落物层较薄的亚热带和热带森林, 无论是地表凋落物层还是矿物质层土壤, 都更偏向吸收固持15NH4+ (Liu et al., 2017c; Wang et al., 2018).目前, 在同一森林的铵态氮和硝态氮配对的实验较少, 但几乎所有的配对标记结果都显示, 地上植物对15NO3-的总吸收固持率明显比15NH4+更高.其主要原因被认为是沉降氮进入森林生态系统后, 15NH4+更易于被土壤中的微生物固持, 以及发生无机过程的吸附或置换, 更难以到达植物根系被根系吸收.Liu等(2017b)在吉林长白山的研究发现, 与针叶树种相比, 阔叶树种对15NO3-的吸收利用速率更快.Goodale (2017)在美国纽约州Arnot森林的研究发现, 侵染丛枝菌根真菌(AM)的树种比侵染内生菌根真菌(ECM)的树种更偏向利用NO3-.Li等(2019)在辽宁清原次生林的研究也表明, AM树种对标记的15NO3-的吸收利用速率明显快于ECM树种.其中, 蒙古栎(Quercus mongolica)作为阔叶的ECM树种, 也符合这一规律.另外, 通过15N标记后不同时间序列的研究结果表明, 沉降氮进入森林生态系统后, 其损失主要发生在第一个生长季内.在标记后的1-7年的时间内, 地表凋落物层和表层矿物质土壤中固持的沉降氮, 逐渐向更深层土壤和植物层迁移, 而通过气体释放或淋溶过程损失的比例很小, 能够较长时间地滞留在整个生态系统内(Nadelhoffer et al., 2004; Goodale, 2017; Liu et al., 2017b; Wang et al., 2018; Li et al., 2019). ...
... 的吸收利用速率更快.Goodale (2017)在美国纽约州Arnot森林的研究发现, 侵染丛枝菌根真菌(AM)的树种比侵染内生菌根真菌(ECM)的树种更偏向利用NO3-.Li等(2019)在辽宁清原次生林的研究也表明, AM树种对标记的15NO3-的吸收利用速率明显快于ECM树种.其中, 蒙古栎(Quercus mongolica)作为阔叶的ECM树种, 也符合这一规律.另外, 通过15N标记后不同时间序列的研究结果表明, 沉降氮进入森林生态系统后, 其损失主要发生在第一个生长季内.在标记后的1-7年的时间内, 地表凋落物层和表层矿物质土壤中固持的沉降氮, 逐渐向更深层土壤和植物层迁移, 而通过气体释放或淋溶过程损失的比例很小, 能够较长时间地滞留在整个生态系统内(Nadelhoffer et al., 2004; Goodale, 2017; Liu et al., 2017b; Wang et al., 2018; Li et al., 2019). ...
... ; Goodale, 2017; Liu et al., 2017b; Wang et al., 2018; Li et al., 2019). ...
... 另外, 目前已有的15N示踪研究主要集中于短期氮去向研究, 缺乏长时间尺度的研究.然而, 已有研究表明在农田生态系统中, 肥料氮的利用和损失是长期的过程(Sebilo et al., 2013); 在森林生态系统中, 沉降氮能较长时间地滞留在生态系统内部且在不同组分中分配比例发生变化(Nadelhoffer et al., 2004; Krause et al., 2012; Goodale, 2017; Li et al., 2019).因此, 在全球变化背景下, 深入了解长时间尺度下氮在生态系统内部的滞留、损失以及周转对于评估氮的生态环境效应意义深远. ...
Methods for measuring denitrification: diverse approaches to a difficult problem 1 2006
... 目前氮循环研究中不确定性最大的过程是生态系统气态氮损失(Niu et al., 2016).反硝化作用是造成气态氮损失的最主要途径.以往量化反硝化速率的方法有乙炔抑制法和15N标记法, 但由于空间异质性无法应用到生态系统尺度(Groffman et al., 2006).Houlton等(2006)建立的概念模型认为土壤总氮的15N受到氮沉降、氮淋失和气态氮损失的影响, 通过同位素混合模型可以量化生态系统尺度反硝化作用速率(Houlton et al., 2006; Houlton & Bai, 2009).但是, 由于土壤总氮库是长期积累的结果, 极其稳定, 因此土壤总氮的15N自然丰度法反映的是生态系统尺度反硝化作用在长时间上的平均水平(Houlton et al., 2006; Houlton & Bai, 2009; Bai et al., 2012).随后, Fang等(2015)提出利用硝酸盐氮氧同位素自然丰度技术(δ15N、δ18O和Δ17O), 结合传统的氮沉降和氮流失的监测, 再根据质量平衡和同位素质量平衡原理来量化生态系统尺度硝化和反硝化作用速率, 该方法的优势在于可以同时量化不同时间段的硝化和反硝化速率. ...
High retention of 15N-labeled nitrogen deposition in a nitrogen saturated old-growth tropical forest 1 2016
Evaluating source, chemistry and climate change based upon the isotopic composition of nitrate in ice cores 1 2010
... 氮稳定同位素技术在研究生态系统氮循环中有比较长的应用历史, 早在20世纪50年代, Hoering就发表了首篇关于自然物质15N自然丰度的论文(Hoering, 1955).根据分馏方法和分析的气体类型不同, 同位素测定方法的发展大致经历了三个时期.20世纪50年代至80年代, 科学家主要利用碱性次溴酸盐(Rittenberg)氧化法将离子转化成N2, 20世纪80年代至20世纪末期, 科学家利用杜马斯(Dumas)燃烧法把固体样品转化成N2, 实现氮稳定同位素在线测定(Burke et al., 1990), 该方法也沿用至今.自21世纪开始, 科学家进一步发展了测定可溶性有机和无机氮同位素的快速分析技术, 成功通过化学或者生物的方法将溶液中的含氮物质直接转化成N2O, 再利用商业化的气体吹扫捕集系统连接稳定同位素质谱仪进行在线同位素测定(Sigman et al., 2001; McIlvin & Altabet, 2005; Zhang et al., 2007a; Koba et al., 2010; Liu et al., 2017a).新技术对样品的需求是传统方法的1/100甚至1/1 000, 因此适合样品量少和氮含量低的样品, 也使得氮循环的研究从海洋和陆地生态系统扩展到大气和冰芯的研究中(Hastings, 2010; Huang et al., 2019). ...
A liquid chromatographic/mass spectrometric method to evaluate 13C and 15N incorporation into soil amino acids 2 2011
... 15N标记技术是将一定数量的15N富集的含氮化合物添加到系统中(植物、土壤或整个生态系统), 经过一段时间后分析其去向(Templer et al., 2012).在生态系统氮循环研究方面, 15N标记技术不受稳定同位素分馏效应的影响, 具有15N自然丰度法和传统示踪法所不具备的优点.但15N标记技术的最大不足之处是标记药品及样品分析费用较高, 因而限制其广泛应用.此外, 实验中还应考虑标记样品的均匀性、标记期长短和条件、示踪期长短和各库转化, 同时注意追踪的各个库要全面等问题.近年来, 随着稳定同位素技术的不断革新, 氮同位素标记物和仪器检测成本的下降, 15N标记技术已应用于陆地生态系统尺度上的各个方面, 如分析氮的来源、分配和去向、土壤氮的转化以及氮损失等研究(He et al., 2011; Yang et al., 2014; Li et al., 2019; Zhu et al., 2019).下文以陆地生态系统中森林生态系统(自然生态系统)和农田生态系统(人工生态系统)为例, 介绍近年来15N标记技术在生态系统尺度上的应用. ...
... 在实验室培养条件下, 研究者们主要通过15N的生物标志物和同位素模型来研究土壤氮的转化动态.其中, Zhang和Amelung (1996)、Zhang等(2007b)和He等(2006, 2011)将同位素示踪技术和土壤功能组分相结合, 建立了测定土壤氨基糖和氨基酸转化与更新的GC-MS和HPLC-MS方法, 并成功应用该方法区分了不同微生物对土壤有机碳氮转化的贡献.Mary等(1998)、Müller等(2007)通过NH4NO3-15N交叉标记技术, 发展了定量土壤氮转化过程的数值计算模型, 成功模拟出土壤中矿化、固持、再矿化、自养硝化、异养硝化、反硝化、硝酸盐异化还原成铵(DNRA)等多个过程的转化速率.通过15N同位素质量守恒和数值模型, Stevens和Laughlin (1998)、Kool等(2011)、Yang等(2014)、Müller等(2014)则建立了区分土壤释放N2O和N2微生物途径的新方法.此外, 还有研究者将同位素示踪技术与现代分子生物学技术相结合, 来研究更为复杂的微生物氮转化过程, 例如厌氧氨氧化(Zhu et al., 2011; Ding et al., 2014; Nie et al., 2015). ...
A novel GC/MS technique to assess 15N and 13C incorporation into soil amino sugars 1 2006
... 在实验室培养条件下, 研究者们主要通过15N的生物标志物和同位素模型来研究土壤氮的转化动态.其中, Zhang和Amelung (1996)、Zhang等(2007b)和He等(2006, 2011)将同位素示踪技术和土壤功能组分相结合, 建立了测定土壤氨基糖和氨基酸转化与更新的GC-MS和HPLC-MS方法, 并成功应用该方法区分了不同微生物对土壤有机碳氮转化的贡献.Mary等(1998)、Müller等(2007)通过NH4NO3-15N交叉标记技术, 发展了定量土壤氮转化过程的数值计算模型, 成功模拟出土壤中矿化、固持、再矿化、自养硝化、异养硝化、反硝化、硝酸盐异化还原成铵(DNRA)等多个过程的转化速率.通过15N同位素质量守恒和数值模型, Stevens和Laughlin (1998)、Kool等(2011)、Yang等(2014)、Müller等(2014)则建立了区分土壤释放N2O和N2微生物途径的新方法.此外, 还有研究者将同位素示踪技术与现代分子生物学技术相结合, 来研究更为复杂的微生物氮转化过程, 例如厌氧氨氧化(Zhu et al., 2011; Ding et al., 2014; Nie et al., 2015). ...
Long-term change in the nitrogen cycle of tropical forests 1 2011
... 氮稳定同位素自然丰度法被证明是研究氮循环过程的有效手段(Robinson, 2001).由于氮循环的物理、化学和生物过程均伴随氮的同位素分馏, 使生态系统不同含氮化合物的氮同位素组成(15N/14N)呈现动态变化, 因此氮稳定同位素自然丰度被用于指示生态系统中氮的来源和周转过程, 如分析植物生长策略(Houlton et al., 2007)、追踪NOx和NH3的排放源 (Pan et al., 2016; Zong et al., 2017)和量化生态系统尺度上的反硝化速率(Fang et al., 2015). ...
... 然而, 利用氮稳定同位素研究氮循环过程, 必须考虑氮循环各个过程的氮同位素分馏效应(图1).例如, 利用近十年来发展的生态系统氮循环同位素模型和硝酸盐氮氧同位素自然丰度法量化生态系统反硝化速率时, 必须考虑反硝化作用过程中氮同位素分馏效应(Houlton et al., 2006; Houlton & Bai, 2009; Bai et al., 2012; Fang et al., 2015); 利用植物氮同位素研究植物氮吸收策略时, 必须考虑植物氮吸收以及植物内部的氮转移过程中的氮同位素分馏效应(Houlton et al., 2007).目前国内外对于氮同位素分馏效应缺乏系统性的研究, 使得氮循环过程中的氮同位素分馏效应存在很大的不确定性, 如厌氧氨氧化过程(Brunner et al., 2013)和矿化过程(M?bius, 2013; Zhang et al., 2015)的同位素分馏效应研究较少, DNRA过程中的同位素分馏效应尚无报道, 而反硝化过程的同位素分馏效应则随实验条件和计算方法的不同呈现较大的变异性(Denk et al., 2017).因此, 在未来对于氮循环的研究中, 应重点考虑各个过程中氮同位素分馏效应的研究, 而进一步将含氮化合物的氮同位素组成、同位素分馏效应与生物地球化学模型相结合, 也会加深我们对氮循环的理解, 并为这些模型的验证提供综合约束(Bai & Houlton, 2009; Houlton et al., 2015; Denk et al., 2017). ...
Seasonal pattern of ammonium 15N natural abundance in precipitation at a rural forested site and implications for NH3 source partitioning 1 2019
... 氮稳定同位素技术在研究生态系统氮循环中有比较长的应用历史, 早在20世纪50年代, Hoering就发表了首篇关于自然物质15N自然丰度的论文(Hoering, 1955).根据分馏方法和分析的气体类型不同, 同位素测定方法的发展大致经历了三个时期.20世纪50年代至80年代, 科学家主要利用碱性次溴酸盐(Rittenberg)氧化法将离子转化成N2, 20世纪80年代至20世纪末期, 科学家利用杜马斯(Dumas)燃烧法把固体样品转化成N2, 实现氮稳定同位素在线测定(Burke et al., 1990), 该方法也沿用至今.自21世纪开始, 科学家进一步发展了测定可溶性有机和无机氮同位素的快速分析技术, 成功通过化学或者生物的方法将溶液中的含氮物质直接转化成N2O, 再利用商业化的气体吹扫捕集系统连接稳定同位素质谱仪进行在线同位素测定(Sigman et al., 2001; McIlvin & Altabet, 2005; Zhang et al., 2007a; Koba et al., 2010; Liu et al., 2017a).新技术对样品的需求是传统方法的1/100甚至1/1 000, 因此适合样品量少和氮含量低的样品, 也使得氮循环的研究从海洋和陆地生态系统扩展到大气和冰芯的研究中(Hastings, 2010; Huang et al., 2019). ...
氮肥有效性的概念和意义——兼论对传统氮肥利用率的理解误区 1 2014
... 在原位小区试验条件下, 研究者们主要是通过15N脉冲标记来模拟氮肥施用, 从而追踪土壤中肥料氮的流向和命运.由于农田生态系统结构单一、人为活动影响大, 田间原位15N的去向很不稳定, 变异较大(Gardner & Drinkwater, 2009).过去和目前的大部分研究主要探讨农艺措施对肥料氮去向的影响, 相关措施包括施用增效肥料(Zhang et al., 2010; Chalk et al., 2015), 优化施氮方式或时间(Wang et al., 2016a; de Oliveira et al., 2018), 优化施肥量(Rimski-Korsakov et al., 2012; Chen et al., 2016; Wang et al., 2016b), 少免耕(Ruisi et al., 2016), 秸秆还田(Gabriel et al., 2016; Murphy et al., 2016; Quan et al., 2018), 覆膜灌水(Wang et al., 2011; Liu et al., 2015), 轮间作(Xu et al., 2015), 作物品种(Yang et al., 2016), 耕作历史(Liang et al., 2013)等.采用15N示踪法计算出来的作物回收率通常只是肥料氮的当季利用效率, 真实评价氮肥的供氮功能还需要考虑其残留效应.在土壤氮平衡条件下, 残留效应的本质就是当季残留肥料氮对当季土壤氮消耗的补偿, 可以理解为肥料氮的间接供氮效应.该效应被认为是维持土壤长久供氮能力的重要机制(Ju & Christie, 2011; 巨晓棠, 2014).残留肥料氮的后季利用可通过15N示踪定量, Ladha等(2005)总结了前人多季15N示踪研究的结果, 发现当季残留肥料氮在随后五季作物的累积利用率平均仅在6.5%左右(逐年平均分别为3.3%、1.3%、1.0%、0.4%和0.5%).但不能由此就认为残留肥料氮的后季利用就很低, 因为绝大多数的15N示踪结果均显示作物吸收的氮主要还是来自土壤氮库, 肥料氮当季的贡献只占37%-44% (Gardner & Drinkwater, 2009; Blesh & Drinkwater, 2014).为了证明肥料氮的长期效应, Sebilo等(2013)研究了硝态氮肥施入农田土壤后30年的利用和损失情况, 他发现肥料氮的利用和损失是长期的过程, 30年连续作物种植仍有12%-15%的肥料氮残留在土壤中, 第一季肥料氮30年的累积利用率达到61%-65%. ...
氮肥有效性的概念和意义——兼论对传统氮肥利用率的理解误区 1 2014
... 在原位小区试验条件下, 研究者们主要是通过15N脉冲标记来模拟氮肥施用, 从而追踪土壤中肥料氮的流向和命运.由于农田生态系统结构单一、人为活动影响大, 田间原位15N的去向很不稳定, 变异较大(Gardner & Drinkwater, 2009).过去和目前的大部分研究主要探讨农艺措施对肥料氮去向的影响, 相关措施包括施用增效肥料(Zhang et al., 2010; Chalk et al., 2015), 优化施氮方式或时间(Wang et al., 2016a; de Oliveira et al., 2018), 优化施肥量(Rimski-Korsakov et al., 2012; Chen et al., 2016; Wang et al., 2016b), 少免耕(Ruisi et al., 2016), 秸秆还田(Gabriel et al., 2016; Murphy et al., 2016; Quan et al., 2018), 覆膜灌水(Wang et al., 2011; Liu et al., 2015), 轮间作(Xu et al., 2015), 作物品种(Yang et al., 2016), 耕作历史(Liang et al., 2013)等.采用15N示踪法计算出来的作物回收率通常只是肥料氮的当季利用效率, 真实评价氮肥的供氮功能还需要考虑其残留效应.在土壤氮平衡条件下, 残留效应的本质就是当季残留肥料氮对当季土壤氮消耗的补偿, 可以理解为肥料氮的间接供氮效应.该效应被认为是维持土壤长久供氮能力的重要机制(Ju & Christie, 2011; 巨晓棠, 2014).残留肥料氮的后季利用可通过15N示踪定量, Ladha等(2005)总结了前人多季15N示踪研究的结果, 发现当季残留肥料氮在随后五季作物的累积利用率平均仅在6.5%左右(逐年平均分别为3.3%、1.3%、1.0%、0.4%和0.5%).但不能由此就认为残留肥料氮的后季利用就很低, 因为绝大多数的15N示踪结果均显示作物吸收的氮主要还是来自土壤氮库, 肥料氮当季的贡献只占37%-44% (Gardner & Drinkwater, 2009; Blesh & Drinkwater, 2014).为了证明肥料氮的长期效应, Sebilo等(2013)研究了硝态氮肥施入农田土壤后30年的利用和损失情况, 他发现肥料氮的利用和损失是长期的过程, 30年连续作物种植仍有12%-15%的肥料氮残留在土壤中, 第一季肥料氮30年的累积利用率达到61%-65%. ...
Calculation of theoretical nitrogen rate for simple nitrogen recommendations in intensive cropping systems: a case study on the North China Plain 1 2011
... 在原位小区试验条件下, 研究者们主要是通过15N脉冲标记来模拟氮肥施用, 从而追踪土壤中肥料氮的流向和命运.由于农田生态系统结构单一、人为活动影响大, 田间原位15N的去向很不稳定, 变异较大(Gardner & Drinkwater, 2009).过去和目前的大部分研究主要探讨农艺措施对肥料氮去向的影响, 相关措施包括施用增效肥料(Zhang et al., 2010; Chalk et al., 2015), 优化施氮方式或时间(Wang et al., 2016a; de Oliveira et al., 2018), 优化施肥量(Rimski-Korsakov et al., 2012; Chen et al., 2016; Wang et al., 2016b), 少免耕(Ruisi et al., 2016), 秸秆还田(Gabriel et al., 2016; Murphy et al., 2016; Quan et al., 2018), 覆膜灌水(Wang et al., 2011; Liu et al., 2015), 轮间作(Xu et al., 2015), 作物品种(Yang et al., 2016), 耕作历史(Liang et al., 2013)等.采用15N示踪法计算出来的作物回收率通常只是肥料氮的当季利用效率, 真实评价氮肥的供氮功能还需要考虑其残留效应.在土壤氮平衡条件下, 残留效应的本质就是当季残留肥料氮对当季土壤氮消耗的补偿, 可以理解为肥料氮的间接供氮效应.该效应被认为是维持土壤长久供氮能力的重要机制(Ju & Christie, 2011; 巨晓棠, 2014).残留肥料氮的后季利用可通过15N示踪定量, Ladha等(2005)总结了前人多季15N示踪研究的结果, 发现当季残留肥料氮在随后五季作物的累积利用率平均仅在6.5%左右(逐年平均分别为3.3%、1.3%、1.0%、0.4%和0.5%).但不能由此就认为残留肥料氮的后季利用就很低, 因为绝大多数的15N示踪结果均显示作物吸收的氮主要还是来自土壤氮库, 肥料氮当季的贡献只占37%-44% (Gardner & Drinkwater, 2009; Blesh & Drinkwater, 2014).为了证明肥料氮的长期效应, Sebilo等(2013)研究了硝态氮肥施入农田土壤后30年的利用和损失情况, 他发现肥料氮的利用和损失是长期的过程, 30年连续作物种植仍有12%-15%的肥料氮残留在土壤中, 第一季肥料氮30年的累积利用率达到61%-65%. ...
1 2010
... 氮稳定同位素技术在研究生态系统氮循环中有比较长的应用历史, 早在20世纪50年代, Hoering就发表了首篇关于自然物质15N自然丰度的论文(Hoering, 1955).根据分馏方法和分析的气体类型不同, 同位素测定方法的发展大致经历了三个时期.20世纪50年代至80年代, 科学家主要利用碱性次溴酸盐(Rittenberg)氧化法将离子转化成N2, 20世纪80年代至20世纪末期, 科学家利用杜马斯(Dumas)燃烧法把固体样品转化成N2, 实现氮稳定同位素在线测定(Burke et al., 1990), 该方法也沿用至今.自21世纪开始, 科学家进一步发展了测定可溶性有机和无机氮同位素的快速分析技术, 成功通过化学或者生物的方法将溶液中的含氮物质直接转化成N2O, 再利用商业化的气体吹扫捕集系统连接稳定同位素质谱仪进行在线同位素测定(Sigman et al., 2001; McIlvin & Altabet, 2005; Zhang et al., 2007a; Koba et al., 2010; Liu et al., 2017a).新技术对样品的需求是传统方法的1/100甚至1/1 000, 因此适合样品量少和氮含量低的样品, 也使得氮循环的研究从海洋和陆地生态系统扩展到大气和冰芯的研究中(Hastings, 2010; Huang et al., 2019). ...
Nitrifier denitrification as a distinct and significant source of nitrous oxide from soil 1 2011
... 在实验室培养条件下, 研究者们主要通过15N的生物标志物和同位素模型来研究土壤氮的转化动态.其中, Zhang和Amelung (1996)、Zhang等(2007b)和He等(2006, 2011)将同位素示踪技术和土壤功能组分相结合, 建立了测定土壤氨基糖和氨基酸转化与更新的GC-MS和HPLC-MS方法, 并成功应用该方法区分了不同微生物对土壤有机碳氮转化的贡献.Mary等(1998)、Müller等(2007)通过NH4NO3-15N交叉标记技术, 发展了定量土壤氮转化过程的数值计算模型, 成功模拟出土壤中矿化、固持、再矿化、自养硝化、异养硝化、反硝化、硝酸盐异化还原成铵(DNRA)等多个过程的转化速率.通过15N同位素质量守恒和数值模型, Stevens和Laughlin (1998)、Kool等(2011)、Yang等(2014)、Müller等(2014)则建立了区分土壤释放N2O和N2微生物途径的新方法.此外, 还有研究者将同位素示踪技术与现代分子生物学技术相结合, 来研究更为复杂的微生物氮转化过程, 例如厌氧氨氧化(Zhu et al., 2011; Ding et al., 2014; Nie et al., 2015). ...
Long-term tracing of whole catchment 15N additions in a mountain spruce forest: measurements and simulations with the TRACE model 1 2012
... 另外, 目前已有的15N示踪研究主要集中于短期氮去向研究, 缺乏长时间尺度的研究.然而, 已有研究表明在农田生态系统中, 肥料氮的利用和损失是长期的过程(Sebilo et al., 2013); 在森林生态系统中, 沉降氮能较长时间地滞留在生态系统内部且在不同组分中分配比例发生变化(Nadelhoffer et al., 2004; Krause et al., 2012; Goodale, 2017; Li et al., 2019).因此, 在全球变化背景下, 深入了解长时间尺度下氮在生态系统内部的滞留、损失以及周转对于评估氮的生态环境效应意义深远. ...
Efficiency of fertilizer nitrogen in cereal production: retrospects and prospects 1 2005
... 在原位小区试验条件下, 研究者们主要是通过15N脉冲标记来模拟氮肥施用, 从而追踪土壤中肥料氮的流向和命运.由于农田生态系统结构单一、人为活动影响大, 田间原位15N的去向很不稳定, 变异较大(Gardner & Drinkwater, 2009).过去和目前的大部分研究主要探讨农艺措施对肥料氮去向的影响, 相关措施包括施用增效肥料(Zhang et al., 2010; Chalk et al., 2015), 优化施氮方式或时间(Wang et al., 2016a; de Oliveira et al., 2018), 优化施肥量(Rimski-Korsakov et al., 2012; Chen et al., 2016; Wang et al., 2016b), 少免耕(Ruisi et al., 2016), 秸秆还田(Gabriel et al., 2016; Murphy et al., 2016; Quan et al., 2018), 覆膜灌水(Wang et al., 2011; Liu et al., 2015), 轮间作(Xu et al., 2015), 作物品种(Yang et al., 2016), 耕作历史(Liang et al., 2013)等.采用15N示踪法计算出来的作物回收率通常只是肥料氮的当季利用效率, 真实评价氮肥的供氮功能还需要考虑其残留效应.在土壤氮平衡条件下, 残留效应的本质就是当季残留肥料氮对当季土壤氮消耗的补偿, 可以理解为肥料氮的间接供氮效应.该效应被认为是维持土壤长久供氮能力的重要机制(Ju & Christie, 2011; 巨晓棠, 2014).残留肥料氮的后季利用可通过15N示踪定量, Ladha等(2005)总结了前人多季15N示踪研究的结果, 发现当季残留肥料氮在随后五季作物的累积利用率平均仅在6.5%左右(逐年平均分别为3.3%、1.3%、1.0%、0.4%和0.5%).但不能由此就认为残留肥料氮的后季利用就很低, 因为绝大多数的15N示踪结果均显示作物吸收的氮主要还是来自土壤氮库, 肥料氮当季的贡献只占37%-44% (Gardner & Drinkwater, 2009; Blesh & Drinkwater, 2014).为了证明肥料氮的长期效应, Sebilo等(2013)研究了硝态氮肥施入农田土壤后30年的利用和损失情况, 他发现肥料氮的利用和损失是长期的过程, 30年连续作物种植仍有12%-15%的肥料氮残留在土壤中, 第一季肥料氮30年的累积利用率达到61%-65%. ...
Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed 2008
Fate of atmospherically deposited NH4+ and NO3- in two temperate forests in China: temporal pattern and redistribution 4 2019
... 15N标记技术是将一定数量的15N富集的含氮化合物添加到系统中(植物、土壤或整个生态系统), 经过一段时间后分析其去向(Templer et al., 2012).在生态系统氮循环研究方面, 15N标记技术不受稳定同位素分馏效应的影响, 具有15N自然丰度法和传统示踪法所不具备的优点.但15N标记技术的最大不足之处是标记药品及样品分析费用较高, 因而限制其广泛应用.此外, 实验中还应考虑标记样品的均匀性、标记期长短和条件、示踪期长短和各库转化, 同时注意追踪的各个库要全面等问题.近年来, 随着稳定同位素技术的不断革新, 氮同位素标记物和仪器检测成本的下降, 15N标记技术已应用于陆地生态系统尺度上的各个方面, 如分析氮的来源、分配和去向、土壤氮的转化以及氮损失等研究(He et al., 2011; Yang et al., 2014; Li et al., 2019; Zhu et al., 2019).下文以陆地生态系统中森林生态系统(自然生态系统)和农田生态系统(人工生态系统)为例, 介绍近年来15N标记技术在生态系统尺度上的应用. ...
... 近些年来, 通过配对15N标记(15NH4+和15NO3-), 越来越多的研究发现, 不仅植物和土壤对于两种沉降氮的吸收和滞留率差异明显, 而且不同类型乔木植物对两种形态氮的吸收利用效率也明显不同(Liu et al., 2017b; Goodale, 2017; Wang et al., 2018).在温带森林, 地表凋落物层对15NH4+的吸收固持率更高, 而对15NO3-吸收固持率明显更低, 导致其在表层矿物质土壤滞留率更高(Feng et al., 2008; Liu et al., 2017b).而在凋落物层较薄的亚热带和热带森林, 无论是地表凋落物层还是矿物质层土壤, 都更偏向吸收固持15NH4+ (Liu et al., 2017c; Wang et al., 2018).目前, 在同一森林的铵态氮和硝态氮配对的实验较少, 但几乎所有的配对标记结果都显示, 地上植物对15NO3-的总吸收固持率明显比15NH4+更高.其主要原因被认为是沉降氮进入森林生态系统后, 15NH4+更易于被土壤中的微生物固持, 以及发生无机过程的吸附或置换, 更难以到达植物根系被根系吸收.Liu等(2017b)在吉林长白山的研究发现, 与针叶树种相比, 阔叶树种对15NO3-的吸收利用速率更快.Goodale (2017)在美国纽约州Arnot森林的研究发现, 侵染丛枝菌根真菌(AM)的树种比侵染内生菌根真菌(ECM)的树种更偏向利用NO3-.Li等(2019)在辽宁清原次生林的研究也表明, AM树种对标记的15NO3-的吸收利用速率明显快于ECM树种.其中, 蒙古栎(Quercus mongolica)作为阔叶的ECM树种, 也符合这一规律.另外, 通过15N标记后不同时间序列的研究结果表明, 沉降氮进入森林生态系统后, 其损失主要发生在第一个生长季内.在标记后的1-7年的时间内, 地表凋落物层和表层矿物质土壤中固持的沉降氮, 逐渐向更深层土壤和植物层迁移, 而通过气体释放或淋溶过程损失的比例很小, 能够较长时间地滞留在整个生态系统内(Nadelhoffer et al., 2004; Goodale, 2017; Liu et al., 2017b; Wang et al., 2018; Li et al., 2019). ...
... ; Li et al., 2019). ...
... 另外, 目前已有的15N示踪研究主要集中于短期氮去向研究, 缺乏长时间尺度的研究.然而, 已有研究表明在农田生态系统中, 肥料氮的利用和损失是长期的过程(Sebilo et al., 2013); 在森林生态系统中, 沉降氮能较长时间地滞留在生态系统内部且在不同组分中分配比例发生变化(Nadelhoffer et al., 2004; Krause et al., 2012; Goodale, 2017; Li et al., 2019).因此, 在全球变化背景下, 深入了解长时间尺度下氮在生态系统内部的滞留、损失以及周转对于评估氮的生态环境效应意义深远. ...
Fate of nitrogen- 15 as influenced by soil and nutrient management history in a 19-year wheat-maize experiment 1 2013
... 在原位小区试验条件下, 研究者们主要是通过15N脉冲标记来模拟氮肥施用, 从而追踪土壤中肥料氮的流向和命运.由于农田生态系统结构单一、人为活动影响大, 田间原位15N的去向很不稳定, 变异较大(Gardner & Drinkwater, 2009).过去和目前的大部分研究主要探讨农艺措施对肥料氮去向的影响, 相关措施包括施用增效肥料(Zhang et al., 2010; Chalk et al., 2015), 优化施氮方式或时间(Wang et al., 2016a; de Oliveira et al., 2018), 优化施肥量(Rimski-Korsakov et al., 2012; Chen et al., 2016; Wang et al., 2016b), 少免耕(Ruisi et al., 2016), 秸秆还田(Gabriel et al., 2016; Murphy et al., 2016; Quan et al., 2018), 覆膜灌水(Wang et al., 2011; Liu et al., 2015), 轮间作(Xu et al., 2015), 作物品种(Yang et al., 2016), 耕作历史(Liang et al., 2013)等.采用15N示踪法计算出来的作物回收率通常只是肥料氮的当季利用效率, 真实评价氮肥的供氮功能还需要考虑其残留效应.在土壤氮平衡条件下, 残留效应的本质就是当季残留肥料氮对当季土壤氮消耗的补偿, 可以理解为肥料氮的间接供氮效应.该效应被认为是维持土壤长久供氮能力的重要机制(Ju & Christie, 2011; 巨晓棠, 2014).残留肥料氮的后季利用可通过15N示踪定量, Ladha等(2005)总结了前人多季15N示踪研究的结果, 发现当季残留肥料氮在随后五季作物的累积利用率平均仅在6.5%左右(逐年平均分别为3.3%、1.3%、1.0%、0.4%和0.5%).但不能由此就认为残留肥料氮的后季利用就很低, 因为绝大多数的15N示踪结果均显示作物吸收的氮主要还是来自土壤氮库, 肥料氮当季的贡献只占37%-44% (Gardner & Drinkwater, 2009; Blesh & Drinkwater, 2014).为了证明肥料氮的长期效应, Sebilo等(2013)研究了硝态氮肥施入农田土壤后30年的利用和损失情况, 他发现肥料氮的利用和损失是长期的过程, 30年连续作物种植仍有12%-15%的肥料氮残留在土壤中, 第一季肥料氮30年的累积利用率达到61%-65%. ...
Different fates of deposited NH4+ and NO3- in a temperate forest in northeast China: a 15N tracer study 4 2017b
... 近些年来, 通过配对15N标记(15NH4+和15NO3-), 越来越多的研究发现, 不仅植物和土壤对于两种沉降氮的吸收和滞留率差异明显, 而且不同类型乔木植物对两种形态氮的吸收利用效率也明显不同(Liu et al., 2017b; Goodale, 2017; Wang et al., 2018).在温带森林, 地表凋落物层对15NH4+的吸收固持率更高, 而对15NO3-吸收固持率明显更低, 导致其在表层矿物质土壤滞留率更高(Feng et al., 2008; Liu et al., 2017b).而在凋落物层较薄的亚热带和热带森林, 无论是地表凋落物层还是矿物质层土壤, 都更偏向吸收固持15NH4+ (Liu et al., 2017c; Wang et al., 2018).目前, 在同一森林的铵态氮和硝态氮配对的实验较少, 但几乎所有的配对标记结果都显示, 地上植物对15NO3-的总吸收固持率明显比15NH4+更高.其主要原因被认为是沉降氮进入森林生态系统后, 15NH4+更易于被土壤中的微生物固持, 以及发生无机过程的吸附或置换, 更难以到达植物根系被根系吸收.Liu等(2017b)在吉林长白山的研究发现, 与针叶树种相比, 阔叶树种对15NO3-的吸收利用速率更快.Goodale (2017)在美国纽约州Arnot森林的研究发现, 侵染丛枝菌根真菌(AM)的树种比侵染内生菌根真菌(ECM)的树种更偏向利用NO3-.Li等(2019)在辽宁清原次生林的研究也表明, AM树种对标记的15NO3-的吸收利用速率明显快于ECM树种.其中, 蒙古栎(Quercus mongolica)作为阔叶的ECM树种, 也符合这一规律.另外, 通过15N标记后不同时间序列的研究结果表明, 沉降氮进入森林生态系统后, 其损失主要发生在第一个生长季内.在标记后的1-7年的时间内, 地表凋落物层和表层矿物质土壤中固持的沉降氮, 逐渐向更深层土壤和植物层迁移, 而通过气体释放或淋溶过程损失的比例很小, 能够较长时间地滞留在整个生态系统内(Nadelhoffer et al., 2004; Goodale, 2017; Liu et al., 2017b; Wang et al., 2018; Li et al., 2019). ...
... ; Liu et al., 2017b).而在凋落物层较薄的亚热带和热带森林, 无论是地表凋落物层还是矿物质层土壤, 都更偏向吸收固持15NH4+ (Liu et al., 2017c; Wang et al., 2018).目前, 在同一森林的铵态氮和硝态氮配对的实验较少, 但几乎所有的配对标记结果都显示, 地上植物对15NO3-的总吸收固持率明显比15NH4+更高.其主要原因被认为是沉降氮进入森林生态系统后, 15NH4+更易于被土壤中的微生物固持, 以及发生无机过程的吸附或置换, 更难以到达植物根系被根系吸收.Liu等(2017b)在吉林长白山的研究发现, 与针叶树种相比, 阔叶树种对15NO3-的吸收利用速率更快.Goodale (2017)在美国纽约州Arnot森林的研究发现, 侵染丛枝菌根真菌(AM)的树种比侵染内生菌根真菌(ECM)的树种更偏向利用NO3-.Li等(2019)在辽宁清原次生林的研究也表明, AM树种对标记的15NO3-的吸收利用速率明显快于ECM树种.其中, 蒙古栎(Quercus mongolica)作为阔叶的ECM树种, 也符合这一规律.另外, 通过15N标记后不同时间序列的研究结果表明, 沉降氮进入森林生态系统后, 其损失主要发生在第一个生长季内.在标记后的1-7年的时间内, 地表凋落物层和表层矿物质土壤中固持的沉降氮, 逐渐向更深层土壤和植物层迁移, 而通过气体释放或淋溶过程损失的比例很小, 能够较长时间地滞留在整个生态系统内(Nadelhoffer et al., 2004; Goodale, 2017; Liu et al., 2017b; Wang et al., 2018; Li et al., 2019). ...
... 更易于被土壤中的微生物固持, 以及发生无机过程的吸附或置换, 更难以到达植物根系被根系吸收.Liu等(2017b)在吉林长白山的研究发现, 与针叶树种相比, 阔叶树种对15NO3-的吸收利用速率更快.Goodale (2017)在美国纽约州Arnot森林的研究发现, 侵染丛枝菌根真菌(AM)的树种比侵染内生菌根真菌(ECM)的树种更偏向利用NO3-.Li等(2019)在辽宁清原次生林的研究也表明, AM树种对标记的15NO3-的吸收利用速率明显快于ECM树种.其中, 蒙古栎(Quercus mongolica)作为阔叶的ECM树种, 也符合这一规律.另外, 通过15N标记后不同时间序列的研究结果表明, 沉降氮进入森林生态系统后, 其损失主要发生在第一个生长季内.在标记后的1-7年的时间内, 地表凋落物层和表层矿物质土壤中固持的沉降氮, 逐渐向更深层土壤和植物层迁移, 而通过气体释放或淋溶过程损失的比例很小, 能够较长时间地滞留在整个生态系统内(Nadelhoffer et al., 2004; Goodale, 2017; Liu et al., 2017b; Wang et al., 2018; Li et al., 2019). ...
... ; Liu et al., 2017b; Wang et al., 2018; Li et al., 2019). ...
In situ15N labeling experiment reveals different long-term responses to ammonium and nitrate inputs in N-saturated subtropical forest 1 2017c
... 近些年来, 通过配对15N标记(15NH4+和15NO3-), 越来越多的研究发现, 不仅植物和土壤对于两种沉降氮的吸收和滞留率差异明显, 而且不同类型乔木植物对两种形态氮的吸收利用效率也明显不同(Liu et al., 2017b; Goodale, 2017; Wang et al., 2018).在温带森林, 地表凋落物层对15NH4+的吸收固持率更高, 而对15NO3-吸收固持率明显更低, 导致其在表层矿物质土壤滞留率更高(Feng et al., 2008; Liu et al., 2017b).而在凋落物层较薄的亚热带和热带森林, 无论是地表凋落物层还是矿物质层土壤, 都更偏向吸收固持15NH4+ (Liu et al., 2017c; Wang et al., 2018).目前, 在同一森林的铵态氮和硝态氮配对的实验较少, 但几乎所有的配对标记结果都显示, 地上植物对15NO3-的总吸收固持率明显比15NH4+更高.其主要原因被认为是沉降氮进入森林生态系统后, 15NH4+更易于被土壤中的微生物固持, 以及发生无机过程的吸附或置换, 更难以到达植物根系被根系吸收.Liu等(2017b)在吉林长白山的研究发现, 与针叶树种相比, 阔叶树种对15NO3-的吸收利用速率更快.Goodale (2017)在美国纽约州Arnot森林的研究发现, 侵染丛枝菌根真菌(AM)的树种比侵染内生菌根真菌(ECM)的树种更偏向利用NO3-.Li等(2019)在辽宁清原次生林的研究也表明, AM树种对标记的15NO3-的吸收利用速率明显快于ECM树种.其中, 蒙古栎(Quercus mongolica)作为阔叶的ECM树种, 也符合这一规律.另外, 通过15N标记后不同时间序列的研究结果表明, 沉降氮进入森林生态系统后, 其损失主要发生在第一个生长季内.在标记后的1-7年的时间内, 地表凋落物层和表层矿物质土壤中固持的沉降氮, 逐渐向更深层土壤和植物层迁移, 而通过气体释放或淋溶过程损失的比例很小, 能够较长时间地滞留在整个生态系统内(Nadelhoffer et al., 2004; Goodale, 2017; Liu et al., 2017b; Wang et al., 2018; Li et al., 2019). ...
The effect of plastic mulch on the fate of urea-N in rain-fed maize production in a semiarid environment as assessed by 15N-labeling 1 2015
... 在原位小区试验条件下, 研究者们主要是通过15N脉冲标记来模拟氮肥施用, 从而追踪土壤中肥料氮的流向和命运.由于农田生态系统结构单一、人为活动影响大, 田间原位15N的去向很不稳定, 变异较大(Gardner & Drinkwater, 2009).过去和目前的大部分研究主要探讨农艺措施对肥料氮去向的影响, 相关措施包括施用增效肥料(Zhang et al., 2010; Chalk et al., 2015), 优化施氮方式或时间(Wang et al., 2016a; de Oliveira et al., 2018), 优化施肥量(Rimski-Korsakov et al., 2012; Chen et al., 2016; Wang et al., 2016b), 少免耕(Ruisi et al., 2016), 秸秆还田(Gabriel et al., 2016; Murphy et al., 2016; Quan et al., 2018), 覆膜灌水(Wang et al., 2011; Liu et al., 2015), 轮间作(Xu et al., 2015), 作物品种(Yang et al., 2016), 耕作历史(Liang et al., 2013)等.采用15N示踪法计算出来的作物回收率通常只是肥料氮的当季利用效率, 真实评价氮肥的供氮功能还需要考虑其残留效应.在土壤氮平衡条件下, 残留效应的本质就是当季残留肥料氮对当季土壤氮消耗的补偿, 可以理解为肥料氮的间接供氮效应.该效应被认为是维持土壤长久供氮能力的重要机制(Ju & Christie, 2011; 巨晓棠, 2014).残留肥料氮的后季利用可通过15N示踪定量, Ladha等(2005)总结了前人多季15N示踪研究的结果, 发现当季残留肥料氮在随后五季作物的累积利用率平均仅在6.5%左右(逐年平均分别为3.3%、1.3%、1.0%、0.4%和0.5%).但不能由此就认为残留肥料氮的后季利用就很低, 因为绝大多数的15N示踪结果均显示作物吸收的氮主要还是来自土壤氮库, 肥料氮当季的贡献只占37%-44% (Gardner & Drinkwater, 2009; Blesh & Drinkwater, 2014).为了证明肥料氮的长期效应, Sebilo等(2013)研究了硝态氮肥施入农田土壤后30年的利用和损失情况, 他发现肥料氮的利用和损失是长期的过程, 30年连续作物种植仍有12%-15%的肥料氮残留在土壤中, 第一季肥料氮30年的累积利用率达到61%-65%. ...
Nitrate is an important nitrogen source for Arctic tundra plants 1 2018
... 随着土壤有效态氮(NH4+和NO3-) 15N分析方法的稳步和快速发展, 植物和土壤15N自然丰度法也越来越多地被用于反映和推测植物氮利用特点和偏好(Houlton et al., 2007).该方法通过分析植物各部位15N自然丰度, 结合土壤NH4+和NO3-及有机氮的15N自然丰度值, 根据同位素混合模型和同位素分馏作用, 进而确定不同物种氮吸收偏好及不同土壤氮形态对植物氮利用的贡献(Takebayashi et al., 2010; Liu et al., 2018).Houlton等(2007)采用该方法研究了位于夏威夷毛依岛(Maui)的降水梯度上(2 200-5 050 mm; 6个站点)热带山地雨林(优势植物种均为Metrosideros polymorpha)的植物氮吸收特点, 发现降水量3 350 mm是植物利用氮形态变化的转折点, 即在降水量少于3 350 mm的站点, NO3-是植物吸收利用的主要形式, 而在降水量大于3 350 mm的站点, NH4+是植物利用氮的主要形式.Zhang等(2018)通过分析来自中国青藏高原东部不同林龄的云杉(Picea asperata)种植林植物叶片δ15N和土壤NH4+和NO3-及有机氮的15N自然丰度值, 根据同位素二元和三元混合模型计算发现, 在林龄20和30年的种植林内, 植物优先利用NO3-, 在林龄大于30年的40、50、70年的云杉种植林内植物则更偏向利用NH4+ (Zhang et al., 2018), 而土壤有机氮也是植物氮利用的重要组成部分, 占23%-44%.以上来自不同类型生态系统(热带山地林和高山云杉林)的研究结果充分表明植物氮利用策略上具有可塑性, 即利用土壤中最主要的氮形式, 这代表了植物具备一定的适应性以对抗环境选择压力. ...
A model for calculating nitrogen fluxes in soil using 15N tracing 1 1998
... 在实验室培养条件下, 研究者们主要通过15N的生物标志物和同位素模型来研究土壤氮的转化动态.其中, Zhang和Amelung (1996)、Zhang等(2007b)和He等(2006, 2011)将同位素示踪技术和土壤功能组分相结合, 建立了测定土壤氨基糖和氨基酸转化与更新的GC-MS和HPLC-MS方法, 并成功应用该方法区分了不同微生物对土壤有机碳氮转化的贡献.Mary等(1998)、Müller等(2007)通过NH4NO3-15N交叉标记技术, 发展了定量土壤氮转化过程的数值计算模型, 成功模拟出土壤中矿化、固持、再矿化、自养硝化、异养硝化、反硝化、硝酸盐异化还原成铵(DNRA)等多个过程的转化速率.通过15N同位素质量守恒和数值模型, Stevens和Laughlin (1998)、Kool等(2011)、Yang等(2014)、Müller等(2014)则建立了区分土壤释放N2O和N2微生物途径的新方法.此外, 还有研究者将同位素示踪技术与现代分子生物学技术相结合, 来研究更为复杂的微生物氮转化过程, 例如厌氧氨氧化(Zhu et al., 2011; Ding et al., 2014; Nie et al., 2015). ...
Nitrogen isotope patterns in Alaskan black spruce reflect organic nitrogen sources and the activity of ectomycorrhizal fungi 1 2012
Stable Isotopes in Ecology and Environmental Science. 2007
Isotope fractionation during nitrogen remineralization (ammonification): implications for nitrogen isotope biogeochemistry 1 2013
... 然而, 利用氮稳定同位素研究氮循环过程, 必须考虑氮循环各个过程的氮同位素分馏效应(图1).例如, 利用近十年来发展的生态系统氮循环同位素模型和硝酸盐氮氧同位素自然丰度法量化生态系统反硝化速率时, 必须考虑反硝化作用过程中氮同位素分馏效应(Houlton et al., 2006; Houlton & Bai, 2009; Bai et al., 2012; Fang et al., 2015); 利用植物氮同位素研究植物氮吸收策略时, 必须考虑植物氮吸收以及植物内部的氮转移过程中的氮同位素分馏效应(Houlton et al., 2007).目前国内外对于氮同位素分馏效应缺乏系统性的研究, 使得氮循环过程中的氮同位素分馏效应存在很大的不确定性, 如厌氧氨氧化过程(Brunner et al., 2013)和矿化过程(M?bius, 2013; Zhang et al., 2015)的同位素分馏效应研究较少, DNRA过程中的同位素分馏效应尚无报道, 而反硝化过程的同位素分馏效应则随实验条件和计算方法的不同呈现较大的变异性(Denk et al., 2017).因此, 在未来对于氮循环的研究中, 应重点考虑各个过程中氮同位素分馏效应的研究, 而进一步将含氮化合物的氮同位素组成、同位素分馏效应与生物地球化学模型相结合, 也会加深我们对氮循环的理解, 并为这些模型的验证提供综合约束(Bai & Houlton, 2009; Houlton et al., 2015; Denk et al., 2017). ...
Quantification of N2O emission pathways via a 15N tracing model 1 2014
... 在实验室培养条件下, 研究者们主要通过15N的生物标志物和同位素模型来研究土壤氮的转化动态.其中, Zhang和Amelung (1996)、Zhang等(2007b)和He等(2006, 2011)将同位素示踪技术和土壤功能组分相结合, 建立了测定土壤氨基糖和氨基酸转化与更新的GC-MS和HPLC-MS方法, 并成功应用该方法区分了不同微生物对土壤有机碳氮转化的贡献.Mary等(1998)、Müller等(2007)通过NH4NO3-15N交叉标记技术, 发展了定量土壤氮转化过程的数值计算模型, 成功模拟出土壤中矿化、固持、再矿化、自养硝化、异养硝化、反硝化、硝酸盐异化还原成铵(DNRA)等多个过程的转化速率.通过15N同位素质量守恒和数值模型, Stevens和Laughlin (1998)、Kool等(2011)、Yang等(2014)、Müller等(2014)则建立了区分土壤释放N2O和N2微生物途径的新方法.此外, 还有研究者将同位素示踪技术与现代分子生物学技术相结合, 来研究更为复杂的微生物氮转化过程, 例如厌氧氨氧化(Zhu et al., 2011; Ding et al., 2014; Nie et al., 2015). ...
Estimation of parameters in complex 15N tracing models by Monte Carlo sampling 2 2007
... 在实验室培养条件下, 研究者们主要通过15N的生物标志物和同位素模型来研究土壤氮的转化动态.其中, Zhang和Amelung (1996)、Zhang等(2007b)和He等(2006, 2011)将同位素示踪技术和土壤功能组分相结合, 建立了测定土壤氨基糖和氨基酸转化与更新的GC-MS和HPLC-MS方法, 并成功应用该方法区分了不同微生物对土壤有机碳氮转化的贡献.Mary等(1998)、Müller等(2007)通过NH4NO3-15N交叉标记技术, 发展了定量土壤氮转化过程的数值计算模型, 成功模拟出土壤中矿化、固持、再矿化、自养硝化、异养硝化、反硝化、硝酸盐异化还原成铵(DNRA)等多个过程的转化速率.通过15N同位素质量守恒和数值模型, Stevens和Laughlin (1998)、Kool等(2011)、Yang等(2014)、Müller等(2014)则建立了区分土壤释放N2O和N2微生物途径的新方法.此外, 还有研究者将同位素示踪技术与现代分子生物学技术相结合, 来研究更为复杂的微生物氮转化过程, 例如厌氧氨氧化(Zhu et al., 2011; Ding et al., 2014; Nie et al., 2015). ...
... 最近十年, 陆地生态系统氮稳定同位素标记技术得到长足的发展, 如利用15N交叉标记技术研究土壤氮转化速率、气态氮损失和氮在生态系统内部的去向及分配等(Müller et al., 2007; Templer et al., 2012; Sebilo et al., 2013; Yang et al., 2014; Xi et al., 2016).然而, 目前生态系统尺度上的研究样方面积仍然在几十到几百平方米.随着同位素标记产品的多样化和成本降低, 未来的研究可以标记不同的氮形态, 也可发展到集水区尺度的示踪研究, 如集水区或景观尺度上不同形态氮损失的研究.另外, 区域尺度15N示踪研究也有必要进一步发展, 如方运霆等正在开展的中国森林15N示踪网络研究, 在国家尺度上利用15N标记技术研究不同气候区森林生态系统沉降氮去向和分配. ...
Crop residue retention enhances soil properties and nitrogen cycling in smallholder maize systems of Chiapas, Mexico 1 2016
... 在原位小区试验条件下, 研究者们主要是通过15N脉冲标记来模拟氮肥施用, 从而追踪土壤中肥料氮的流向和命运.由于农田生态系统结构单一、人为活动影响大, 田间原位15N的去向很不稳定, 变异较大(Gardner & Drinkwater, 2009).过去和目前的大部分研究主要探讨农艺措施对肥料氮去向的影响, 相关措施包括施用增效肥料(Zhang et al., 2010; Chalk et al., 2015), 优化施氮方式或时间(Wang et al., 2016a; de Oliveira et al., 2018), 优化施肥量(Rimski-Korsakov et al., 2012; Chen et al., 2016; Wang et al., 2016b), 少免耕(Ruisi et al., 2016), 秸秆还田(Gabriel et al., 2016; Murphy et al., 2016; Quan et al., 2018), 覆膜灌水(Wang et al., 2011; Liu et al., 2015), 轮间作(Xu et al., 2015), 作物品种(Yang et al., 2016), 耕作历史(Liang et al., 2013)等.采用15N示踪法计算出来的作物回收率通常只是肥料氮的当季利用效率, 真实评价氮肥的供氮功能还需要考虑其残留效应.在土壤氮平衡条件下, 残留效应的本质就是当季残留肥料氮对当季土壤氮消耗的补偿, 可以理解为肥料氮的间接供氮效应.该效应被认为是维持土壤长久供氮能力的重要机制(Ju & Christie, 2011; 巨晓棠, 2014).残留肥料氮的后季利用可通过15N示踪定量, Ladha等(2005)总结了前人多季15N示踪研究的结果, 发现当季残留肥料氮在随后五季作物的累积利用率平均仅在6.5%左右(逐年平均分别为3.3%、1.3%、1.0%、0.4%和0.5%).但不能由此就认为残留肥料氮的后季利用就很低, 因为绝大多数的15N示踪结果均显示作物吸收的氮主要还是来自土壤氮库, 肥料氮当季的贡献只占37%-44% (Gardner & Drinkwater, 2009; Blesh & Drinkwater, 2014).为了证明肥料氮的长期效应, Sebilo等(2013)研究了硝态氮肥施入农田土壤后30年的利用和损失情况, 他发现肥料氮的利用和损失是长期的过程, 30年连续作物种植仍有12%-15%的肥料氮残留在土壤中, 第一季肥料氮30年的累积利用率达到61%-65%. ...
Decadal-scale fates of 15N tracers added to oak and pine stands under ambient and elevated N inputs at the Harvard Forest (USA) 2 2004
... 近些年来, 通过配对15N标记(15NH4+和15NO3-), 越来越多的研究发现, 不仅植物和土壤对于两种沉降氮的吸收和滞留率差异明显, 而且不同类型乔木植物对两种形态氮的吸收利用效率也明显不同(Liu et al., 2017b; Goodale, 2017; Wang et al., 2018).在温带森林, 地表凋落物层对15NH4+的吸收固持率更高, 而对15NO3-吸收固持率明显更低, 导致其在表层矿物质土壤滞留率更高(Feng et al., 2008; Liu et al., 2017b).而在凋落物层较薄的亚热带和热带森林, 无论是地表凋落物层还是矿物质层土壤, 都更偏向吸收固持15NH4+ (Liu et al., 2017c; Wang et al., 2018).目前, 在同一森林的铵态氮和硝态氮配对的实验较少, 但几乎所有的配对标记结果都显示, 地上植物对15NO3-的总吸收固持率明显比15NH4+更高.其主要原因被认为是沉降氮进入森林生态系统后, 15NH4+更易于被土壤中的微生物固持, 以及发生无机过程的吸附或置换, 更难以到达植物根系被根系吸收.Liu等(2017b)在吉林长白山的研究发现, 与针叶树种相比, 阔叶树种对15NO3-的吸收利用速率更快.Goodale (2017)在美国纽约州Arnot森林的研究发现, 侵染丛枝菌根真菌(AM)的树种比侵染内生菌根真菌(ECM)的树种更偏向利用NO3-.Li等(2019)在辽宁清原次生林的研究也表明, AM树种对标记的15NO3-的吸收利用速率明显快于ECM树种.其中, 蒙古栎(Quercus mongolica)作为阔叶的ECM树种, 也符合这一规律.另外, 通过15N标记后不同时间序列的研究结果表明, 沉降氮进入森林生态系统后, 其损失主要发生在第一个生长季内.在标记后的1-7年的时间内, 地表凋落物层和表层矿物质土壤中固持的沉降氮, 逐渐向更深层土壤和植物层迁移, 而通过气体释放或淋溶过程损失的比例很小, 能够较长时间地滞留在整个生态系统内(Nadelhoffer et al., 2004; Goodale, 2017; Liu et al., 2017b; Wang et al., 2018; Li et al., 2019). ...
... 另外, 目前已有的15N示踪研究主要集中于短期氮去向研究, 缺乏长时间尺度的研究.然而, 已有研究表明在农田生态系统中, 肥料氮的利用和损失是长期的过程(Sebilo et al., 2013); 在森林生态系统中, 沉降氮能较长时间地滞留在生态系统内部且在不同组分中分配比例发生变化(Nadelhoffer et al., 2004; Krause et al., 2012; Goodale, 2017; Li et al., 2019).因此, 在全球变化背景下, 深入了解长时间尺度下氮在生态系统内部的滞留、损失以及周转对于评估氮的生态环境效应意义深远. ...
Sinks for 15N-enriched additions to an oak forest and a red pine plantation 1 1999
Nitrogen loss by anaerobic oxidation of ammonium in rice rhizosphere 1 2015
... 在实验室培养条件下, 研究者们主要通过15N的生物标志物和同位素模型来研究土壤氮的转化动态.其中, Zhang和Amelung (1996)、Zhang等(2007b)和He等(2006, 2011)将同位素示踪技术和土壤功能组分相结合, 建立了测定土壤氨基糖和氨基酸转化与更新的GC-MS和HPLC-MS方法, 并成功应用该方法区分了不同微生物对土壤有机碳氮转化的贡献.Mary等(1998)、Müller等(2007)通过NH4NO3-15N交叉标记技术, 发展了定量土壤氮转化过程的数值计算模型, 成功模拟出土壤中矿化、固持、再矿化、自养硝化、异养硝化、反硝化、硝酸盐异化还原成铵(DNRA)等多个过程的转化速率.通过15N同位素质量守恒和数值模型, Stevens和Laughlin (1998)、Kool等(2011)、Yang等(2014)、Müller等(2014)则建立了区分土壤释放N2O和N2微生物途径的新方法.此外, 还有研究者将同位素示踪技术与现代分子生物学技术相结合, 来研究更为复杂的微生物氮转化过程, 例如厌氧氨氧化(Zhu et al., 2011; Ding et al., 2014; Nie et al., 2015). ...
Global patterns and substrate-based mechanisms of the terrestrial nitrogen cycle 1 2016
... 目前氮循环研究中不确定性最大的过程是生态系统气态氮损失(Niu et al., 2016).反硝化作用是造成气态氮损失的最主要途径.以往量化反硝化速率的方法有乙炔抑制法和15N标记法, 但由于空间异质性无法应用到生态系统尺度(Groffman et al., 2006).Houlton等(2006)建立的概念模型认为土壤总氮的15N受到氮沉降、氮淋失和气态氮损失的影响, 通过同位素混合模型可以量化生态系统尺度反硝化作用速率(Houlton et al., 2006; Houlton & Bai, 2009).但是, 由于土壤总氮库是长期积累的结果, 极其稳定, 因此土壤总氮的15N自然丰度法反映的是生态系统尺度反硝化作用在长时间上的平均水平(Houlton et al., 2006; Houlton & Bai, 2009; Bai et al., 2012).随后, Fang等(2015)提出利用硝酸盐氮氧同位素自然丰度技术(δ15N、δ18O和Δ17O), 结合传统的氮沉降和氮流失的监测, 再根据质量平衡和同位素质量平衡原理来量化生态系统尺度硝化和反硝化作用速率, 该方法的优势在于可以同时量化不同时间段的硝化和反硝化速率. ...
Fossil fuel combustion-related emissions dominate atmospheric ammonia sources during severe haze episodes: evidence from 15N-stable isotope in size-resolved aerosol ammonium 1 2016
... 氮稳定同位素自然丰度法被证明是研究氮循环过程的有效手段(Robinson, 2001).由于氮循环的物理、化学和生物过程均伴随氮的同位素分馏, 使生态系统不同含氮化合物的氮同位素组成(15N/14N)呈现动态变化, 因此氮稳定同位素自然丰度被用于指示生态系统中氮的来源和周转过程, 如分析植物生长策略(Houlton et al., 2007)、追踪NOx和NH3的排放源 (Pan et al., 2016; Zong et al., 2017)和量化生态系统尺度上的反硝化速率(Fang et al., 2015). ...
Fates of 15N-labeled fertilizer in a black soil-maize system and the response to straw incorporation in Northeast China 1 2018
... 在原位小区试验条件下, 研究者们主要是通过15N脉冲标记来模拟氮肥施用, 从而追踪土壤中肥料氮的流向和命运.由于农田生态系统结构单一、人为活动影响大, 田间原位15N的去向很不稳定, 变异较大(Gardner & Drinkwater, 2009).过去和目前的大部分研究主要探讨农艺措施对肥料氮去向的影响, 相关措施包括施用增效肥料(Zhang et al., 2010; Chalk et al., 2015), 优化施氮方式或时间(Wang et al., 2016a; de Oliveira et al., 2018), 优化施肥量(Rimski-Korsakov et al., 2012; Chen et al., 2016; Wang et al., 2016b), 少免耕(Ruisi et al., 2016), 秸秆还田(Gabriel et al., 2016; Murphy et al., 2016; Quan et al., 2018), 覆膜灌水(Wang et al., 2011; Liu et al., 2015), 轮间作(Xu et al., 2015), 作物品种(Yang et al., 2016), 耕作历史(Liang et al., 2013)等.采用15N示踪法计算出来的作物回收率通常只是肥料氮的当季利用效率, 真实评价氮肥的供氮功能还需要考虑其残留效应.在土壤氮平衡条件下, 残留效应的本质就是当季残留肥料氮对当季土壤氮消耗的补偿, 可以理解为肥料氮的间接供氮效应.该效应被认为是维持土壤长久供氮能力的重要机制(Ju & Christie, 2011; 巨晓棠, 2014).残留肥料氮的后季利用可通过15N示踪定量, Ladha等(2005)总结了前人多季15N示踪研究的结果, 发现当季残留肥料氮在随后五季作物的累积利用率平均仅在6.5%左右(逐年平均分别为3.3%、1.3%、1.0%、0.4%和0.5%).但不能由此就认为残留肥料氮的后季利用就很低, 因为绝大多数的15N示踪结果均显示作物吸收的氮主要还是来自土壤氮库, 肥料氮当季的贡献只占37%-44% (Gardner & Drinkwater, 2009; Blesh & Drinkwater, 2014).为了证明肥料氮的长期效应, Sebilo等(2013)研究了硝态氮肥施入农田土壤后30年的利用和损失情况, 他发现肥料氮的利用和损失是长期的过程, 30年连续作物种植仍有12%-15%的肥料氮残留在土壤中, 第一季肥料氮30年的累积利用率达到61%-65%. ...
Exacerbated nitrogen limitation ends transient stimulation of grassland productivity by increased precipitation 1 2017
Fate of the nitrogen from fertilizers in field-grown maize 1 2012
... 在原位小区试验条件下, 研究者们主要是通过15N脉冲标记来模拟氮肥施用, 从而追踪土壤中肥料氮的流向和命运.由于农田生态系统结构单一、人为活动影响大, 田间原位15N的去向很不稳定, 变异较大(Gardner & Drinkwater, 2009).过去和目前的大部分研究主要探讨农艺措施对肥料氮去向的影响, 相关措施包括施用增效肥料(Zhang et al., 2010; Chalk et al., 2015), 优化施氮方式或时间(Wang et al., 2016a; de Oliveira et al., 2018), 优化施肥量(Rimski-Korsakov et al., 2012; Chen et al., 2016; Wang et al., 2016b), 少免耕(Ruisi et al., 2016), 秸秆还田(Gabriel et al., 2016; Murphy et al., 2016; Quan et al., 2018), 覆膜灌水(Wang et al., 2011; Liu et al., 2015), 轮间作(Xu et al., 2015), 作物品种(Yang et al., 2016), 耕作历史(Liang et al., 2013)等.采用15N示踪法计算出来的作物回收率通常只是肥料氮的当季利用效率, 真实评价氮肥的供氮功能还需要考虑其残留效应.在土壤氮平衡条件下, 残留效应的本质就是当季残留肥料氮对当季土壤氮消耗的补偿, 可以理解为肥料氮的间接供氮效应.该效应被认为是维持土壤长久供氮能力的重要机制(Ju & Christie, 2011; 巨晓棠, 2014).残留肥料氮的后季利用可通过15N示踪定量, Ladha等(2005)总结了前人多季15N示踪研究的结果, 发现当季残留肥料氮在随后五季作物的累积利用率平均仅在6.5%左右(逐年平均分别为3.3%、1.3%、1.0%、0.4%和0.5%).但不能由此就认为残留肥料氮的后季利用就很低, 因为绝大多数的15N示踪结果均显示作物吸收的氮主要还是来自土壤氮库, 肥料氮当季的贡献只占37%-44% (Gardner & Drinkwater, 2009; Blesh & Drinkwater, 2014).为了证明肥料氮的长期效应, Sebilo等(2013)研究了硝态氮肥施入农田土壤后30年的利用和损失情况, 他发现肥料氮的利用和损失是长期的过程, 30年连续作物种植仍有12%-15%的肥料氮残留在土壤中, 第一季肥料氮30年的累积利用率达到61%-65%. ...
δ15N as an integrator of the nitrogen cycle 3 2001
... 氮稳定同位素自然丰度法被证明是研究氮循环过程的有效手段(Robinson, 2001).由于氮循环的物理、化学和生物过程均伴随氮的同位素分馏, 使生态系统不同含氮化合物的氮同位素组成(15N/14N)呈现动态变化, 因此氮稳定同位素自然丰度被用于指示生态系统中氮的来源和周转过程, 如分析植物生长策略(Houlton et al., 2007)、追踪NOx和NH3的排放源 (Pan et al., 2016; Zong et al., 2017)和量化生态系统尺度上的反硝化速率(Fang et al., 2015). ...
Long-term effects of no tillage treatment on soil N availability, N uptake, and 15N-fertilizer recovery of durum wheat differ in relation to crop sequence 1 2016
... 在原位小区试验条件下, 研究者们主要是通过15N脉冲标记来模拟氮肥施用, 从而追踪土壤中肥料氮的流向和命运.由于农田生态系统结构单一、人为活动影响大, 田间原位15N的去向很不稳定, 变异较大(Gardner & Drinkwater, 2009).过去和目前的大部分研究主要探讨农艺措施对肥料氮去向的影响, 相关措施包括施用增效肥料(Zhang et al., 2010; Chalk et al., 2015), 优化施氮方式或时间(Wang et al., 2016a; de Oliveira et al., 2018), 优化施肥量(Rimski-Korsakov et al., 2012; Chen et al., 2016; Wang et al., 2016b), 少免耕(Ruisi et al., 2016), 秸秆还田(Gabriel et al., 2016; Murphy et al., 2016; Quan et al., 2018), 覆膜灌水(Wang et al., 2011; Liu et al., 2015), 轮间作(Xu et al., 2015), 作物品种(Yang et al., 2016), 耕作历史(Liang et al., 2013)等.采用15N示踪法计算出来的作物回收率通常只是肥料氮的当季利用效率, 真实评价氮肥的供氮功能还需要考虑其残留效应.在土壤氮平衡条件下, 残留效应的本质就是当季残留肥料氮对当季土壤氮消耗的补偿, 可以理解为肥料氮的间接供氮效应.该效应被认为是维持土壤长久供氮能力的重要机制(Ju & Christie, 2011; 巨晓棠, 2014).残留肥料氮的后季利用可通过15N示踪定量, Ladha等(2005)总结了前人多季15N示踪研究的结果, 发现当季残留肥料氮在随后五季作物的累积利用率平均仅在6.5%左右(逐年平均分别为3.3%、1.3%、1.0%、0.4%和0.5%).但不能由此就认为残留肥料氮的后季利用就很低, 因为绝大多数的15N示踪结果均显示作物吸收的氮主要还是来自土壤氮库, 肥料氮当季的贡献只占37%-44% (Gardner & Drinkwater, 2009; Blesh & Drinkwater, 2014).为了证明肥料氮的长期效应, Sebilo等(2013)研究了硝态氮肥施入农田土壤后30年的利用和损失情况, 他发现肥料氮的利用和损失是长期的过程, 30年连续作物种植仍有12%-15%的肥料氮残留在土壤中, 第一季肥料氮30年的累积利用率达到61%-65%. ...
Long-term fate of nitrate fertilizer in agricultural soils 3 2013
... 在原位小区试验条件下, 研究者们主要是通过15N脉冲标记来模拟氮肥施用, 从而追踪土壤中肥料氮的流向和命运.由于农田生态系统结构单一、人为活动影响大, 田间原位15N的去向很不稳定, 变异较大(Gardner & Drinkwater, 2009).过去和目前的大部分研究主要探讨农艺措施对肥料氮去向的影响, 相关措施包括施用增效肥料(Zhang et al., 2010; Chalk et al., 2015), 优化施氮方式或时间(Wang et al., 2016a; de Oliveira et al., 2018), 优化施肥量(Rimski-Korsakov et al., 2012; Chen et al., 2016; Wang et al., 2016b), 少免耕(Ruisi et al., 2016), 秸秆还田(Gabriel et al., 2016; Murphy et al., 2016; Quan et al., 2018), 覆膜灌水(Wang et al., 2011; Liu et al., 2015), 轮间作(Xu et al., 2015), 作物品种(Yang et al., 2016), 耕作历史(Liang et al., 2013)等.采用15N示踪法计算出来的作物回收率通常只是肥料氮的当季利用效率, 真实评价氮肥的供氮功能还需要考虑其残留效应.在土壤氮平衡条件下, 残留效应的本质就是当季残留肥料氮对当季土壤氮消耗的补偿, 可以理解为肥料氮的间接供氮效应.该效应被认为是维持土壤长久供氮能力的重要机制(Ju & Christie, 2011; 巨晓棠, 2014).残留肥料氮的后季利用可通过15N示踪定量, Ladha等(2005)总结了前人多季15N示踪研究的结果, 发现当季残留肥料氮在随后五季作物的累积利用率平均仅在6.5%左右(逐年平均分别为3.3%、1.3%、1.0%、0.4%和0.5%).但不能由此就认为残留肥料氮的后季利用就很低, 因为绝大多数的15N示踪结果均显示作物吸收的氮主要还是来自土壤氮库, 肥料氮当季的贡献只占37%-44% (Gardner & Drinkwater, 2009; Blesh & Drinkwater, 2014).为了证明肥料氮的长期效应, Sebilo等(2013)研究了硝态氮肥施入农田土壤后30年的利用和损失情况, 他发现肥料氮的利用和损失是长期的过程, 30年连续作物种植仍有12%-15%的肥料氮残留在土壤中, 第一季肥料氮30年的累积利用率达到61%-65%. ...
... 最近十年, 陆地生态系统氮稳定同位素标记技术得到长足的发展, 如利用15N交叉标记技术研究土壤氮转化速率、气态氮损失和氮在生态系统内部的去向及分配等(Müller et al., 2007; Templer et al., 2012; Sebilo et al., 2013; Yang et al., 2014; Xi et al., 2016).然而, 目前生态系统尺度上的研究样方面积仍然在几十到几百平方米.随着同位素标记产品的多样化和成本降低, 未来的研究可以标记不同的氮形态, 也可发展到集水区尺度的示踪研究, 如集水区或景观尺度上不同形态氮损失的研究.另外, 区域尺度15N示踪研究也有必要进一步发展, 如方运霆等正在开展的中国森林15N示踪网络研究, 在国家尺度上利用15N标记技术研究不同气候区森林生态系统沉降氮去向和分配. ...
... 另外, 目前已有的15N示踪研究主要集中于短期氮去向研究, 缺乏长时间尺度的研究.然而, 已有研究表明在农田生态系统中, 肥料氮的利用和损失是长期的过程(Sebilo et al., 2013); 在森林生态系统中, 沉降氮能较长时间地滞留在生态系统内部且在不同组分中分配比例发生变化(Nadelhoffer et al., 2004; Krause et al., 2012; Goodale, 2017; Li et al., 2019).因此, 在全球变化背景下, 深入了解长时间尺度下氮在生态系统内部的滞留、损失以及周转对于评估氮的生态环境效应意义深远. ...
A bacterial method for the nitrogen isotopic analysis of nitrate in seawater and freshwater 1 2001
... 氮稳定同位素技术在研究生态系统氮循环中有比较长的应用历史, 早在20世纪50年代, Hoering就发表了首篇关于自然物质15N自然丰度的论文(Hoering, 1955).根据分馏方法和分析的气体类型不同, 同位素测定方法的发展大致经历了三个时期.20世纪50年代至80年代, 科学家主要利用碱性次溴酸盐(Rittenberg)氧化法将离子转化成N2, 20世纪80年代至20世纪末期, 科学家利用杜马斯(Dumas)燃烧法把固体样品转化成N2, 实现氮稳定同位素在线测定(Burke et al., 1990), 该方法也沿用至今.自21世纪开始, 科学家进一步发展了测定可溶性有机和无机氮同位素的快速分析技术, 成功通过化学或者生物的方法将溶液中的含氮物质直接转化成N2O, 再利用商业化的气体吹扫捕集系统连接稳定同位素质谱仪进行在线同位素测定(Sigman et al., 2001; McIlvin & Altabet, 2005; Zhang et al., 2007a; Koba et al., 2010; Liu et al., 2017a).新技术对样品的需求是传统方法的1/100甚至1/1 000, 因此适合样品量少和氮含量低的样品, 也使得氮循环的研究从海洋和陆地生态系统扩展到大气和冰芯的研究中(Hastings, 2010; Huang et al., 2019). ...
Measurement of nitrous oxide and di-nitrogen emissions from agricultural soils 1 1998
... 在实验室培养条件下, 研究者们主要通过15N的生物标志物和同位素模型来研究土壤氮的转化动态.其中, Zhang和Amelung (1996)、Zhang等(2007b)和He等(2006, 2011)将同位素示踪技术和土壤功能组分相结合, 建立了测定土壤氨基糖和氨基酸转化与更新的GC-MS和HPLC-MS方法, 并成功应用该方法区分了不同微生物对土壤有机碳氮转化的贡献.Mary等(1998)、Müller等(2007)通过NH4NO3-15N交叉标记技术, 发展了定量土壤氮转化过程的数值计算模型, 成功模拟出土壤中矿化、固持、再矿化、自养硝化、异养硝化、反硝化、硝酸盐异化还原成铵(DNRA)等多个过程的转化速率.通过15N同位素质量守恒和数值模型, Stevens和Laughlin (1998)、Kool等(2011)、Yang等(2014)、Müller等(2014)则建立了区分土壤释放N2O和N2微生物途径的新方法.此外, 还有研究者将同位素示踪技术与现代分子生物学技术相结合, 来研究更为复杂的微生物氮转化过程, 例如厌氧氨氧化(Zhu et al., 2011; Ding et al., 2014; Nie et al., 2015). ...
The natural abundance of 15N in plant and soil-available N indicates a shift of main plant N resources to NO3- from NH4+ along the N leaching gradient 1 2010
... 随着土壤有效态氮(NH4+和NO3-) 15N分析方法的稳步和快速发展, 植物和土壤15N自然丰度法也越来越多地被用于反映和推测植物氮利用特点和偏好(Houlton et al., 2007).该方法通过分析植物各部位15N自然丰度, 结合土壤NH4+和NO3-及有机氮的15N自然丰度值, 根据同位素混合模型和同位素分馏作用, 进而确定不同物种氮吸收偏好及不同土壤氮形态对植物氮利用的贡献(Takebayashi et al., 2010; Liu et al., 2018).Houlton等(2007)采用该方法研究了位于夏威夷毛依岛(Maui)的降水梯度上(2 200-5 050 mm; 6个站点)热带山地雨林(优势植物种均为Metrosideros polymorpha)的植物氮吸收特点, 发现降水量3 350 mm是植物利用氮形态变化的转折点, 即在降水量少于3 350 mm的站点, NO3-是植物吸收利用的主要形式, 而在降水量大于3 350 mm的站点, NH4+是植物利用氮的主要形式.Zhang等(2018)通过分析来自中国青藏高原东部不同林龄的云杉(Picea asperata)种植林植物叶片δ15N和土壤NH4+和NO3-及有机氮的15N自然丰度值, 根据同位素二元和三元混合模型计算发现, 在林龄20和30年的种植林内, 植物优先利用NO3-, 在林龄大于30年的40、50、70年的云杉种植林内植物则更偏向利用NH4+ (Zhang et al., 2018), 而土壤有机氮也是植物氮利用的重要组成部分, 占23%-44%.以上来自不同类型生态系统(热带山地林和高山云杉林)的研究结果充分表明植物氮利用策略上具有可塑性, 即利用土壤中最主要的氮形式, 这代表了植物具备一定的适应性以对抗环境选择压力. ...
Sinks for nitrogen inputs in terrestrial ecosystems: a meta-analysis of 15N tracer field studies 3 2012
... 15N标记技术是将一定数量的15N富集的含氮化合物添加到系统中(植物、土壤或整个生态系统), 经过一段时间后分析其去向(Templer et al., 2012).在生态系统氮循环研究方面, 15N标记技术不受稳定同位素分馏效应的影响, 具有15N自然丰度法和传统示踪法所不具备的优点.但15N标记技术的最大不足之处是标记药品及样品分析费用较高, 因而限制其广泛应用.此外, 实验中还应考虑标记样品的均匀性、标记期长短和条件、示踪期长短和各库转化, 同时注意追踪的各个库要全面等问题.近年来, 随着稳定同位素技术的不断革新, 氮同位素标记物和仪器检测成本的下降, 15N标记技术已应用于陆地生态系统尺度上的各个方面, 如分析氮的来源、分配和去向、土壤氮的转化以及氮损失等研究(He et al., 2011; Yang et al., 2014; Li et al., 2019; Zhu et al., 2019).下文以陆地生态系统中森林生态系统(自然生态系统)和农田生态系统(人工生态系统)为例, 介绍近年来15N标记技术在生态系统尺度上的应用. ...
... 最近十年, 陆地生态系统氮稳定同位素标记技术得到长足的发展, 如利用15N交叉标记技术研究土壤氮转化速率、气态氮损失和氮在生态系统内部的去向及分配等(Müller et al., 2007; Templer et al., 2012; Sebilo et al., 2013; Yang et al., 2014; Xi et al., 2016).然而, 目前生态系统尺度上的研究样方面积仍然在几十到几百平方米.随着同位素标记产品的多样化和成本降低, 未来的研究可以标记不同的氮形态, 也可发展到集水区尺度的示踪研究, 如集水区或景观尺度上不同形态氮损失的研究.另外, 区域尺度15N示踪研究也有必要进一步发展, 如方运霆等正在开展的中国森林15N示踪网络研究, 在国家尺度上利用15N标记技术研究不同气候区森林生态系统沉降氮去向和分配. ...
The fate of 15N-labelled nitrogen deposition in coniferous forest ecosystems 1 1998
Fates of atmospheric deposited nitrogen in an Asian tropical primary forest 3 2018
... 近些年来, 通过配对15N标记(15NH4+和15NO3-), 越来越多的研究发现, 不仅植物和土壤对于两种沉降氮的吸收和滞留率差异明显, 而且不同类型乔木植物对两种形态氮的吸收利用效率也明显不同(Liu et al., 2017b; Goodale, 2017; Wang et al., 2018).在温带森林, 地表凋落物层对15NH4+的吸收固持率更高, 而对15NO3-吸收固持率明显更低, 导致其在表层矿物质土壤滞留率更高(Feng et al., 2008; Liu et al., 2017b).而在凋落物层较薄的亚热带和热带森林, 无论是地表凋落物层还是矿物质层土壤, 都更偏向吸收固持15NH4+ (Liu et al., 2017c; Wang et al., 2018).目前, 在同一森林的铵态氮和硝态氮配对的实验较少, 但几乎所有的配对标记结果都显示, 地上植物对15NO3-的总吸收固持率明显比15NH4+更高.其主要原因被认为是沉降氮进入森林生态系统后, 15NH4+更易于被土壤中的微生物固持, 以及发生无机过程的吸附或置换, 更难以到达植物根系被根系吸收.Liu等(2017b)在吉林长白山的研究发现, 与针叶树种相比, 阔叶树种对15NO3-的吸收利用速率更快.Goodale (2017)在美国纽约州Arnot森林的研究发现, 侵染丛枝菌根真菌(AM)的树种比侵染内生菌根真菌(ECM)的树种更偏向利用NO3-.Li等(2019)在辽宁清原次生林的研究也表明, AM树种对标记的15NO3-的吸收利用速率明显快于ECM树种.其中, 蒙古栎(Quercus mongolica)作为阔叶的ECM树种, 也符合这一规律.另外, 通过15N标记后不同时间序列的研究结果表明, 沉降氮进入森林生态系统后, 其损失主要发生在第一个生长季内.在标记后的1-7年的时间内, 地表凋落物层和表层矿物质土壤中固持的沉降氮, 逐渐向更深层土壤和植物层迁移, 而通过气体释放或淋溶过程损失的比例很小, 能够较长时间地滞留在整个生态系统内(Nadelhoffer et al., 2004; Goodale, 2017; Liu et al., 2017b; Wang et al., 2018; Li et al., 2019). ...
... ; Wang et al., 2018).目前, 在同一森林的铵态氮和硝态氮配对的实验较少, 但几乎所有的配对标记结果都显示, 地上植物对15NO3-的总吸收固持率明显比15NH4+更高.其主要原因被认为是沉降氮进入森林生态系统后, 15NH4+更易于被土壤中的微生物固持, 以及发生无机过程的吸附或置换, 更难以到达植物根系被根系吸收.Liu等(2017b)在吉林长白山的研究发现, 与针叶树种相比, 阔叶树种对15NO3-的吸收利用速率更快.Goodale (2017)在美国纽约州Arnot森林的研究发现, 侵染丛枝菌根真菌(AM)的树种比侵染内生菌根真菌(ECM)的树种更偏向利用NO3-.Li等(2019)在辽宁清原次生林的研究也表明, AM树种对标记的15NO3-的吸收利用速率明显快于ECM树种.其中, 蒙古栎(Quercus mongolica)作为阔叶的ECM树种, 也符合这一规律.另外, 通过15N标记后不同时间序列的研究结果表明, 沉降氮进入森林生态系统后, 其损失主要发生在第一个生长季内.在标记后的1-7年的时间内, 地表凋落物层和表层矿物质土壤中固持的沉降氮, 逐渐向更深层土壤和植物层迁移, 而通过气体释放或淋溶过程损失的比例很小, 能够较长时间地滞留在整个生态系统内(Nadelhoffer et al., 2004; Goodale, 2017; Liu et al., 2017b; Wang et al., 2018; Li et al., 2019). ...
... ; Wang et al., 2018; Li et al., 2019). ...
Fate of 15N fertilizer under different nitrogen split applications to plastic mulched maize in semiarid farmland 1 2016a
... 在原位小区试验条件下, 研究者们主要是通过15N脉冲标记来模拟氮肥施用, 从而追踪土壤中肥料氮的流向和命运.由于农田生态系统结构单一、人为活动影响大, 田间原位15N的去向很不稳定, 变异较大(Gardner & Drinkwater, 2009).过去和目前的大部分研究主要探讨农艺措施对肥料氮去向的影响, 相关措施包括施用增效肥料(Zhang et al., 2010; Chalk et al., 2015), 优化施氮方式或时间(Wang et al., 2016a; de Oliveira et al., 2018), 优化施肥量(Rimski-Korsakov et al., 2012; Chen et al., 2016; Wang et al., 2016b), 少免耕(Ruisi et al., 2016), 秸秆还田(Gabriel et al., 2016; Murphy et al., 2016; Quan et al., 2018), 覆膜灌水(Wang et al., 2011; Liu et al., 2015), 轮间作(Xu et al., 2015), 作物品种(Yang et al., 2016), 耕作历史(Liang et al., 2013)等.采用15N示踪法计算出来的作物回收率通常只是肥料氮的当季利用效率, 真实评价氮肥的供氮功能还需要考虑其残留效应.在土壤氮平衡条件下, 残留效应的本质就是当季残留肥料氮对当季土壤氮消耗的补偿, 可以理解为肥料氮的间接供氮效应.该效应被认为是维持土壤长久供氮能力的重要机制(Ju & Christie, 2011; 巨晓棠, 2014).残留肥料氮的后季利用可通过15N示踪定量, Ladha等(2005)总结了前人多季15N示踪研究的结果, 发现当季残留肥料氮在随后五季作物的累积利用率平均仅在6.5%左右(逐年平均分别为3.3%、1.3%、1.0%、0.4%和0.5%).但不能由此就认为残留肥料氮的后季利用就很低, 因为绝大多数的15N示踪结果均显示作物吸收的氮主要还是来自土壤氮库, 肥料氮当季的贡献只占37%-44% (Gardner & Drinkwater, 2009; Blesh & Drinkwater, 2014).为了证明肥料氮的长期效应, Sebilo等(2013)研究了硝态氮肥施入农田土壤后30年的利用和损失情况, 他发现肥料氮的利用和损失是长期的过程, 30年连续作物种植仍有12%-15%的肥料氮残留在土壤中, 第一季肥料氮30年的累积利用率达到61%-65%. ...
The fate of 15N-labelled urea in an alkaline calcareous soil under different N application rates and N splits 1 2016b
... 在原位小区试验条件下, 研究者们主要是通过15N脉冲标记来模拟氮肥施用, 从而追踪土壤中肥料氮的流向和命运.由于农田生态系统结构单一、人为活动影响大, 田间原位15N的去向很不稳定, 变异较大(Gardner & Drinkwater, 2009).过去和目前的大部分研究主要探讨农艺措施对肥料氮去向的影响, 相关措施包括施用增效肥料(Zhang et al., 2010; Chalk et al., 2015), 优化施氮方式或时间(Wang et al., 2016a; de Oliveira et al., 2018), 优化施肥量(Rimski-Korsakov et al., 2012; Chen et al., 2016; Wang et al., 2016b), 少免耕(Ruisi et al., 2016), 秸秆还田(Gabriel et al., 2016; Murphy et al., 2016; Quan et al., 2018), 覆膜灌水(Wang et al., 2011; Liu et al., 2015), 轮间作(Xu et al., 2015), 作物品种(Yang et al., 2016), 耕作历史(Liang et al., 2013)等.采用15N示踪法计算出来的作物回收率通常只是肥料氮的当季利用效率, 真实评价氮肥的供氮功能还需要考虑其残留效应.在土壤氮平衡条件下, 残留效应的本质就是当季残留肥料氮对当季土壤氮消耗的补偿, 可以理解为肥料氮的间接供氮效应.该效应被认为是维持土壤长久供氮能力的重要机制(Ju & Christie, 2011; 巨晓棠, 2014).残留肥料氮的后季利用可通过15N示踪定量, Ladha等(2005)总结了前人多季15N示踪研究的结果, 发现当季残留肥料氮在随后五季作物的累积利用率平均仅在6.5%左右(逐年平均分别为3.3%、1.3%、1.0%、0.4%和0.5%).但不能由此就认为残留肥料氮的后季利用就很低, 因为绝大多数的15N示踪结果均显示作物吸收的氮主要还是来自土壤氮库, 肥料氮当季的贡献只占37%-44% (Gardner & Drinkwater, 2009; Blesh & Drinkwater, 2014).为了证明肥料氮的长期效应, Sebilo等(2013)研究了硝态氮肥施入农田土壤后30年的利用和损失情况, 他发现肥料氮的利用和损失是长期的过程, 30年连续作物种植仍有12%-15%的肥料氮残留在土壤中, 第一季肥料氮30年的累积利用率达到61%-65%. ...
Recovery of 15N-labeled urea and soil nitrogen dynamics as affected by irrigation management and nitrogen application rate in a double rice cropping system 1 2011
... 在原位小区试验条件下, 研究者们主要是通过15N脉冲标记来模拟氮肥施用, 从而追踪土壤中肥料氮的流向和命运.由于农田生态系统结构单一、人为活动影响大, 田间原位15N的去向很不稳定, 变异较大(Gardner & Drinkwater, 2009).过去和目前的大部分研究主要探讨农艺措施对肥料氮去向的影响, 相关措施包括施用增效肥料(Zhang et al., 2010; Chalk et al., 2015), 优化施氮方式或时间(Wang et al., 2016a; de Oliveira et al., 2018), 优化施肥量(Rimski-Korsakov et al., 2012; Chen et al., 2016; Wang et al., 2016b), 少免耕(Ruisi et al., 2016), 秸秆还田(Gabriel et al., 2016; Murphy et al., 2016; Quan et al., 2018), 覆膜灌水(Wang et al., 2011; Liu et al., 2015), 轮间作(Xu et al., 2015), 作物品种(Yang et al., 2016), 耕作历史(Liang et al., 2013)等.采用15N示踪法计算出来的作物回收率通常只是肥料氮的当季利用效率, 真实评价氮肥的供氮功能还需要考虑其残留效应.在土壤氮平衡条件下, 残留效应的本质就是当季残留肥料氮对当季土壤氮消耗的补偿, 可以理解为肥料氮的间接供氮效应.该效应被认为是维持土壤长久供氮能力的重要机制(Ju & Christie, 2011; 巨晓棠, 2014).残留肥料氮的后季利用可通过15N示踪定量, Ladha等(2005)总结了前人多季15N示踪研究的结果, 发现当季残留肥料氮在随后五季作物的累积利用率平均仅在6.5%左右(逐年平均分别为3.3%、1.3%、1.0%、0.4%和0.5%).但不能由此就认为残留肥料氮的后季利用就很低, 因为绝大多数的15N示踪结果均显示作物吸收的氮主要还是来自土壤氮库, 肥料氮当季的贡献只占37%-44% (Gardner & Drinkwater, 2009; Blesh & Drinkwater, 2014).为了证明肥料氮的长期效应, Sebilo等(2013)研究了硝态氮肥施入农田土壤后30年的利用和损失情况, 他发现肥料氮的利用和损失是长期的过程, 30年连续作物种植仍有12%-15%的肥料氮残留在土壤中, 第一季肥料氮30年的累积利用率达到61%-65%. ...
Contribution of anammox to nitrogen removal in two temperate forest soils 1 2016
... 最近十年, 陆地生态系统氮稳定同位素标记技术得到长足的发展, 如利用15N交叉标记技术研究土壤氮转化速率、气态氮损失和氮在生态系统内部的去向及分配等(Müller et al., 2007; Templer et al., 2012; Sebilo et al., 2013; Yang et al., 2014; Xi et al., 2016).然而, 目前生态系统尺度上的研究样方面积仍然在几十到几百平方米.随着同位素标记产品的多样化和成本降低, 未来的研究可以标记不同的氮形态, 也可发展到集水区尺度的示踪研究, 如集水区或景观尺度上不同形态氮损失的研究.另外, 区域尺度15N示踪研究也有必要进一步发展, 如方运霆等正在开展的中国森林15N示踪网络研究, 在国家尺度上利用15N标记技术研究不同气候区森林生态系统沉降氮去向和分配. ...
Effect of nitrogen management during the panicle stage in rice on the nitrogen utilization of rice and succeeding wheat crops 1 2015
... 在原位小区试验条件下, 研究者们主要是通过15N脉冲标记来模拟氮肥施用, 从而追踪土壤中肥料氮的流向和命运.由于农田生态系统结构单一、人为活动影响大, 田间原位15N的去向很不稳定, 变异较大(Gardner & Drinkwater, 2009).过去和目前的大部分研究主要探讨农艺措施对肥料氮去向的影响, 相关措施包括施用增效肥料(Zhang et al., 2010; Chalk et al., 2015), 优化施氮方式或时间(Wang et al., 2016a; de Oliveira et al., 2018), 优化施肥量(Rimski-Korsakov et al., 2012; Chen et al., 2016; Wang et al., 2016b), 少免耕(Ruisi et al., 2016), 秸秆还田(Gabriel et al., 2016; Murphy et al., 2016; Quan et al., 2018), 覆膜灌水(Wang et al., 2011; Liu et al., 2015), 轮间作(Xu et al., 2015), 作物品种(Yang et al., 2016), 耕作历史(Liang et al., 2013)等.采用15N示踪法计算出来的作物回收率通常只是肥料氮的当季利用效率, 真实评价氮肥的供氮功能还需要考虑其残留效应.在土壤氮平衡条件下, 残留效应的本质就是当季残留肥料氮对当季土壤氮消耗的补偿, 可以理解为肥料氮的间接供氮效应.该效应被认为是维持土壤长久供氮能力的重要机制(Ju & Christie, 2011; 巨晓棠, 2014).残留肥料氮的后季利用可通过15N示踪定量, Ladha等(2005)总结了前人多季15N示踪研究的结果, 发现当季残留肥料氮在随后五季作物的累积利用率平均仅在6.5%左右(逐年平均分别为3.3%、1.3%、1.0%、0.4%和0.5%).但不能由此就认为残留肥料氮的后季利用就很低, 因为绝大多数的15N示踪结果均显示作物吸收的氮主要还是来自土壤氮库, 肥料氮当季的贡献只占37%-44% (Gardner & Drinkwater, 2009; Blesh & Drinkwater, 2014).为了证明肥料氮的长期效应, Sebilo等(2013)研究了硝态氮肥施入农田土壤后30年的利用和损失情况, 他发现肥料氮的利用和损失是长期的过程, 30年连续作物种植仍有12%-15%的肥料氮残留在土壤中, 第一季肥料氮30年的累积利用率达到61%-65%. ...
Use of the stable nitrogen isotope to reveal the source-sink regulation of nitrogen uptake and remobilization during grain filling phase in maize 1 2016
... 在原位小区试验条件下, 研究者们主要是通过15N脉冲标记来模拟氮肥施用, 从而追踪土壤中肥料氮的流向和命运.由于农田生态系统结构单一、人为活动影响大, 田间原位15N的去向很不稳定, 变异较大(Gardner & Drinkwater, 2009).过去和目前的大部分研究主要探讨农艺措施对肥料氮去向的影响, 相关措施包括施用增效肥料(Zhang et al., 2010; Chalk et al., 2015), 优化施氮方式或时间(Wang et al., 2016a; de Oliveira et al., 2018), 优化施肥量(Rimski-Korsakov et al., 2012; Chen et al., 2016; Wang et al., 2016b), 少免耕(Ruisi et al., 2016), 秸秆还田(Gabriel et al., 2016; Murphy et al., 2016; Quan et al., 2018), 覆膜灌水(Wang et al., 2011; Liu et al., 2015), 轮间作(Xu et al., 2015), 作物品种(Yang et al., 2016), 耕作历史(Liang et al., 2013)等.采用15N示踪法计算出来的作物回收率通常只是肥料氮的当季利用效率, 真实评价氮肥的供氮功能还需要考虑其残留效应.在土壤氮平衡条件下, 残留效应的本质就是当季残留肥料氮对当季土壤氮消耗的补偿, 可以理解为肥料氮的间接供氮效应.该效应被认为是维持土壤长久供氮能力的重要机制(Ju & Christie, 2011; 巨晓棠, 2014).残留肥料氮的后季利用可通过15N示踪定量, Ladha等(2005)总结了前人多季15N示踪研究的结果, 发现当季残留肥料氮在随后五季作物的累积利用率平均仅在6.5%左右(逐年平均分别为3.3%、1.3%、1.0%、0.4%和0.5%).但不能由此就认为残留肥料氮的后季利用就很低, 因为绝大多数的15N示踪结果均显示作物吸收的氮主要还是来自土壤氮库, 肥料氮当季的贡献只占37%-44% (Gardner & Drinkwater, 2009; Blesh & Drinkwater, 2014).为了证明肥料氮的长期效应, Sebilo等(2013)研究了硝态氮肥施入农田土壤后30年的利用和损失情况, 他发现肥料氮的利用和损失是长期的过程, 30年连续作物种植仍有12%-15%的肥料氮残留在土壤中, 第一季肥料氮30年的累积利用率达到61%-65%. ...
New high precision approach for measuring 15N-N2 gas fluxes from terrestrial ecosystems 3 2014
... 15N标记技术是将一定数量的15N富集的含氮化合物添加到系统中(植物、土壤或整个生态系统), 经过一段时间后分析其去向(Templer et al., 2012).在生态系统氮循环研究方面, 15N标记技术不受稳定同位素分馏效应的影响, 具有15N自然丰度法和传统示踪法所不具备的优点.但15N标记技术的最大不足之处是标记药品及样品分析费用较高, 因而限制其广泛应用.此外, 实验中还应考虑标记样品的均匀性、标记期长短和条件、示踪期长短和各库转化, 同时注意追踪的各个库要全面等问题.近年来, 随着稳定同位素技术的不断革新, 氮同位素标记物和仪器检测成本的下降, 15N标记技术已应用于陆地生态系统尺度上的各个方面, 如分析氮的来源、分配和去向、土壤氮的转化以及氮损失等研究(He et al., 2011; Yang et al., 2014; Li et al., 2019; Zhu et al., 2019).下文以陆地生态系统中森林生态系统(自然生态系统)和农田生态系统(人工生态系统)为例, 介绍近年来15N标记技术在生态系统尺度上的应用. ...
... 在实验室培养条件下, 研究者们主要通过15N的生物标志物和同位素模型来研究土壤氮的转化动态.其中, Zhang和Amelung (1996)、Zhang等(2007b)和He等(2006, 2011)将同位素示踪技术和土壤功能组分相结合, 建立了测定土壤氨基糖和氨基酸转化与更新的GC-MS和HPLC-MS方法, 并成功应用该方法区分了不同微生物对土壤有机碳氮转化的贡献.Mary等(1998)、Müller等(2007)通过NH4NO3-15N交叉标记技术, 发展了定量土壤氮转化过程的数值计算模型, 成功模拟出土壤中矿化、固持、再矿化、自养硝化、异养硝化、反硝化、硝酸盐异化还原成铵(DNRA)等多个过程的转化速率.通过15N同位素质量守恒和数值模型, Stevens和Laughlin (1998)、Kool等(2011)、Yang等(2014)、Müller等(2014)则建立了区分土壤释放N2O和N2微生物途径的新方法.此外, 还有研究者将同位素示踪技术与现代分子生物学技术相结合, 来研究更为复杂的微生物氮转化过程, 例如厌氧氨氧化(Zhu et al., 2011; Ding et al., 2014; Nie et al., 2015). ...
... 最近十年, 陆地生态系统氮稳定同位素标记技术得到长足的发展, 如利用15N交叉标记技术研究土壤氮转化速率、气态氮损失和氮在生态系统内部的去向及分配等(Müller et al., 2007; Templer et al., 2012; Sebilo et al., 2013; Yang et al., 2014; Xi et al., 2016).然而, 目前生态系统尺度上的研究样方面积仍然在几十到几百平方米.随着同位素标记产品的多样化和成本降低, 未来的研究可以标记不同的氮形态, 也可发展到集水区尺度的示踪研究, 如集水区或景观尺度上不同形态氮损失的研究.另外, 区域尺度15N示踪研究也有必要进一步发展, 如方运霆等正在开展的中国森林15N示踪网络研究, 在国家尺度上利用15N标记技术研究不同气候区森林生态系统沉降氮去向和分配. ...
Sensitive measurement of NH4+15N/14N (δ15NH4+) at natural abundance levels in fresh and saltwaters 1 2007a
... 氮稳定同位素技术在研究生态系统氮循环中有比较长的应用历史, 早在20世纪50年代, Hoering就发表了首篇关于自然物质15N自然丰度的论文(Hoering, 1955).根据分馏方法和分析的气体类型不同, 同位素测定方法的发展大致经历了三个时期.20世纪50年代至80年代, 科学家主要利用碱性次溴酸盐(Rittenberg)氧化法将离子转化成N2, 20世纪80年代至20世纪末期, 科学家利用杜马斯(Dumas)燃烧法把固体样品转化成N2, 实现氮稳定同位素在线测定(Burke et al., 1990), 该方法也沿用至今.自21世纪开始, 科学家进一步发展了测定可溶性有机和无机氮同位素的快速分析技术, 成功通过化学或者生物的方法将溶液中的含氮物质直接转化成N2O, 再利用商业化的气体吹扫捕集系统连接稳定同位素质谱仪进行在线同位素测定(Sigman et al., 2001; McIlvin & Altabet, 2005; Zhang et al., 2007a; Koba et al., 2010; Liu et al., 2017a).新技术对样品的需求是传统方法的1/100甚至1/1 000, 因此适合样品量少和氮含量低的样品, 也使得氮循环的研究从海洋和陆地生态系统扩展到大气和冰芯的研究中(Hastings, 2010; Huang et al., 2019). ...
Fate of applied urea 15N in a soil-maize system as affected by urease inhibitor and nitrification inhibitor 1 2010
... 在原位小区试验条件下, 研究者们主要是通过15N脉冲标记来模拟氮肥施用, 从而追踪土壤中肥料氮的流向和命运.由于农田生态系统结构单一、人为活动影响大, 田间原位15N的去向很不稳定, 变异较大(Gardner & Drinkwater, 2009).过去和目前的大部分研究主要探讨农艺措施对肥料氮去向的影响, 相关措施包括施用增效肥料(Zhang et al., 2010; Chalk et al., 2015), 优化施氮方式或时间(Wang et al., 2016a; de Oliveira et al., 2018), 优化施肥量(Rimski-Korsakov et al., 2012; Chen et al., 2016; Wang et al., 2016b), 少免耕(Ruisi et al., 2016), 秸秆还田(Gabriel et al., 2016; Murphy et al., 2016; Quan et al., 2018), 覆膜灌水(Wang et al., 2011; Liu et al., 2015), 轮间作(Xu et al., 2015), 作物品种(Yang et al., 2016), 耕作历史(Liang et al., 2013)等.采用15N示踪法计算出来的作物回收率通常只是肥料氮的当季利用效率, 真实评价氮肥的供氮功能还需要考虑其残留效应.在土壤氮平衡条件下, 残留效应的本质就是当季残留肥料氮对当季土壤氮消耗的补偿, 可以理解为肥料氮的间接供氮效应.该效应被认为是维持土壤长久供氮能力的重要机制(Ju & Christie, 2011; 巨晓棠, 2014).残留肥料氮的后季利用可通过15N示踪定量, Ladha等(2005)总结了前人多季15N示踪研究的结果, 发现当季残留肥料氮在随后五季作物的累积利用率平均仅在6.5%左右(逐年平均分别为3.3%、1.3%、1.0%、0.4%和0.5%).但不能由此就认为残留肥料氮的后季利用就很低, 因为绝大多数的15N示踪结果均显示作物吸收的氮主要还是来自土壤氮库, 肥料氮当季的贡献只占37%-44% (Gardner & Drinkwater, 2009; Blesh & Drinkwater, 2014).为了证明肥料氮的长期效应, Sebilo等(2013)研究了硝态氮肥施入农田土壤后30年的利用和损失情况, 他发现肥料氮的利用和损失是长期的过程, 30年连续作物种植仍有12%-15%的肥料氮残留在土壤中, 第一季肥料氮30年的累积利用率达到61%-65%. ...
Adaptation of microdiffusion method for the analysis of 15N natural abundance of ammonium in samples with small volume 1 2015
... 然而, 利用氮稳定同位素研究氮循环过程, 必须考虑氮循环各个过程的氮同位素分馏效应(图1).例如, 利用近十年来发展的生态系统氮循环同位素模型和硝酸盐氮氧同位素自然丰度法量化生态系统反硝化速率时, 必须考虑反硝化作用过程中氮同位素分馏效应(Houlton et al., 2006; Houlton & Bai, 2009; Bai et al., 2012; Fang et al., 2015); 利用植物氮同位素研究植物氮吸收策略时, 必须考虑植物氮吸收以及植物内部的氮转移过程中的氮同位素分馏效应(Houlton et al., 2007).目前国内外对于氮同位素分馏效应缺乏系统性的研究, 使得氮循环过程中的氮同位素分馏效应存在很大的不确定性, 如厌氧氨氧化过程(Brunner et al., 2013)和矿化过程(M?bius, 2013; Zhang et al., 2015)的同位素分馏效应研究较少, DNRA过程中的同位素分馏效应尚无报道, 而反硝化过程的同位素分馏效应则随实验条件和计算方法的不同呈现较大的变异性(Denk et al., 2017).因此, 在未来对于氮循环的研究中, 应重点考虑各个过程中氮同位素分馏效应的研究, 而进一步将含氮化合物的氮同位素组成、同位素分馏效应与生物地球化学模型相结合, 也会加深我们对氮循环的理解, 并为这些模型的验证提供综合约束(Bai & Houlton, 2009; Houlton et al., 2015; Denk et al., 2017). ...
Gas chromatographic determination of muramic acid, glucosamine, mannosamine, and galactosamine in soils 1 1996
... 在实验室培养条件下, 研究者们主要通过15N的生物标志物和同位素模型来研究土壤氮的转化动态.其中, Zhang和Amelung (1996)、Zhang等(2007b)和He等(2006, 2011)将同位素示踪技术和土壤功能组分相结合, 建立了测定土壤氨基糖和氨基酸转化与更新的GC-MS和HPLC-MS方法, 并成功应用该方法区分了不同微生物对土壤有机碳氮转化的贡献.Mary等(1998)、Müller等(2007)通过NH4NO3-15N交叉标记技术, 发展了定量土壤氮转化过程的数值计算模型, 成功模拟出土壤中矿化、固持、再矿化、自养硝化、异养硝化、反硝化、硝酸盐异化还原成铵(DNRA)等多个过程的转化速率.通过15N同位素质量守恒和数值模型, Stevens和Laughlin (1998)、Kool等(2011)、Yang等(2014)、Müller等(2014)则建立了区分土壤释放N2O和N2微生物途径的新方法.此外, 还有研究者将同位素示踪技术与现代分子生物学技术相结合, 来研究更为复杂的微生物氮转化过程, 例如厌氧氨氧化(Zhu et al., 2011; Ding et al., 2014; Nie et al., 2015). ...
A GC/MS method for the assessment of 15N and 13C incorporation into soil amino acid enantiomers 1 2007b
... 在实验室培养条件下, 研究者们主要通过15N的生物标志物和同位素模型来研究土壤氮的转化动态.其中, Zhang和Amelung (1996)、Zhang等(2007b)和He等(2006, 2011)将同位素示踪技术和土壤功能组分相结合, 建立了测定土壤氨基糖和氨基酸转化与更新的GC-MS和HPLC-MS方法, 并成功应用该方法区分了不同微生物对土壤有机碳氮转化的贡献.Mary等(1998)、Müller等(2007)通过NH4NO3-15N交叉标记技术, 发展了定量土壤氮转化过程的数值计算模型, 成功模拟出土壤中矿化、固持、再矿化、自养硝化、异养硝化、反硝化、硝酸盐异化还原成铵(DNRA)等多个过程的转化速率.通过15N同位素质量守恒和数值模型, Stevens和Laughlin (1998)、Kool等(2011)、Yang等(2014)、Müller等(2014)则建立了区分土壤释放N2O和N2微生物途径的新方法.此外, 还有研究者将同位素示踪技术与现代分子生物学技术相结合, 来研究更为复杂的微生物氮转化过程, 例如厌氧氨氧化(Zhu et al., 2011; Ding et al., 2014; Nie et al., 2015). ...
Changes in plant nitrogen acquisition strategies during the restoration of spruce plantations on the eastern Tibetan Plateau, China 1 2018
... 随着土壤有效态氮(NH4+和NO3-) 15N分析方法的稳步和快速发展, 植物和土壤15N自然丰度法也越来越多地被用于反映和推测植物氮利用特点和偏好(Houlton et al., 2007).该方法通过分析植物各部位15N自然丰度, 结合土壤NH4+和NO3-及有机氮的15N自然丰度值, 根据同位素混合模型和同位素分馏作用, 进而确定不同物种氮吸收偏好及不同土壤氮形态对植物氮利用的贡献(Takebayashi et al., 2010; Liu et al., 2018).Houlton等(2007)采用该方法研究了位于夏威夷毛依岛(Maui)的降水梯度上(2 200-5 050 mm; 6个站点)热带山地雨林(优势植物种均为Metrosideros polymorpha)的植物氮吸收特点, 发现降水量3 350 mm是植物利用氮形态变化的转折点, 即在降水量少于3 350 mm的站点, NO3-是植物吸收利用的主要形式, 而在降水量大于3 350 mm的站点, NH4+是植物利用氮的主要形式.Zhang等(2018)通过分析来自中国青藏高原东部不同林龄的云杉(Picea asperata)种植林植物叶片δ15N和土壤NH4+和NO3-及有机氮的15N自然丰度值, 根据同位素二元和三元混合模型计算发现, 在林龄20和30年的种植林内, 植物优先利用NO3-, 在林龄大于30年的40、50、70年的云杉种植林内植物则更偏向利用NH4+ (Zhang et al., 2018), 而土壤有机氮也是植物氮利用的重要组成部分, 占23%-44%.以上来自不同类型生态系统(热带山地林和高山云杉林)的研究结果充分表明植物氮利用策略上具有可塑性, 即利用土壤中最主要的氮形式, 这代表了植物具备一定的适应性以对抗环境选择压力. ...
Uptake patterns of glycine, ammonium, and nitrate differ among four common tree species of northeast China 1 2019
... 15N标记技术是将一定数量的15N富集的含氮化合物添加到系统中(植物、土壤或整个生态系统), 经过一段时间后分析其去向(Templer et al., 2012).在生态系统氮循环研究方面, 15N标记技术不受稳定同位素分馏效应的影响, 具有15N自然丰度法和传统示踪法所不具备的优点.但15N标记技术的最大不足之处是标记药品及样品分析费用较高, 因而限制其广泛应用.此外, 实验中还应考虑标记样品的均匀性、标记期长短和条件、示踪期长短和各库转化, 同时注意追踪的各个库要全面等问题.近年来, 随着稳定同位素技术的不断革新, 氮同位素标记物和仪器检测成本的下降, 15N标记技术已应用于陆地生态系统尺度上的各个方面, 如分析氮的来源、分配和去向、土壤氮的转化以及氮损失等研究(He et al., 2011; Yang et al., 2014; Li et al., 2019; Zhu et al., 2019).下文以陆地生态系统中森林生态系统(自然生态系统)和农田生态系统(人工生态系统)为例, 介绍近年来15N标记技术在生态系统尺度上的应用. ...
Anaerobic ammonia oxidation in a fertilized paddy soil 1 2011
... 在实验室培养条件下, 研究者们主要通过15N的生物标志物和同位素模型来研究土壤氮的转化动态.其中, Zhang和Amelung (1996)、Zhang等(2007b)和He等(2006, 2011)将同位素示踪技术和土壤功能组分相结合, 建立了测定土壤氨基糖和氨基酸转化与更新的GC-MS和HPLC-MS方法, 并成功应用该方法区分了不同微生物对土壤有机碳氮转化的贡献.Mary等(1998)、Müller等(2007)通过NH4NO3-15N交叉标记技术, 发展了定量土壤氮转化过程的数值计算模型, 成功模拟出土壤中矿化、固持、再矿化、自养硝化、异养硝化、反硝化、硝酸盐异化还原成铵(DNRA)等多个过程的转化速率.通过15N同位素质量守恒和数值模型, Stevens和Laughlin (1998)、Kool等(2011)、Yang等(2014)、Müller等(2014)则建立了区分土壤释放N2O和N2微生物途径的新方法.此外, 还有研究者将同位素示踪技术与现代分子生物学技术相结合, 来研究更为复杂的微生物氮转化过程, 例如厌氧氨氧化(Zhu et al., 2011; Ding et al., 2014; Nie et al., 2015). ...
First assessment of NOx sources at a regional background site in North China using isotopic analysis linked with modeling 1 2017
... 氮稳定同位素自然丰度法被证明是研究氮循环过程的有效手段(Robinson, 2001).由于氮循环的物理、化学和生物过程均伴随氮的同位素分馏, 使生态系统不同含氮化合物的氮同位素组成(15N/14N)呈现动态变化, 因此氮稳定同位素自然丰度被用于指示生态系统中氮的来源和周转过程, 如分析植物生长策略(Houlton et al., 2007)、追踪NOx和NH3的排放源 (Pan et al., 2016; Zong et al., 2017)和量化生态系统尺度上的反硝化速率(Fang et al., 2015). ...
 ,1,2,3,*, 刘冬伟1,2,3, 朱飞飞1,2,3, 图影1,3,4, 李善龙1,2, 黄韶楠1,3,4, 全智1,2,3, 王盎1,2,3
,1,2,3,*, 刘冬伟1,2,3, 朱飞飞1,2,3, 图影1,3,4, 李善龙1,2, 黄韶楠1,3,4, 全智1,2,3, 王盎1,2,3
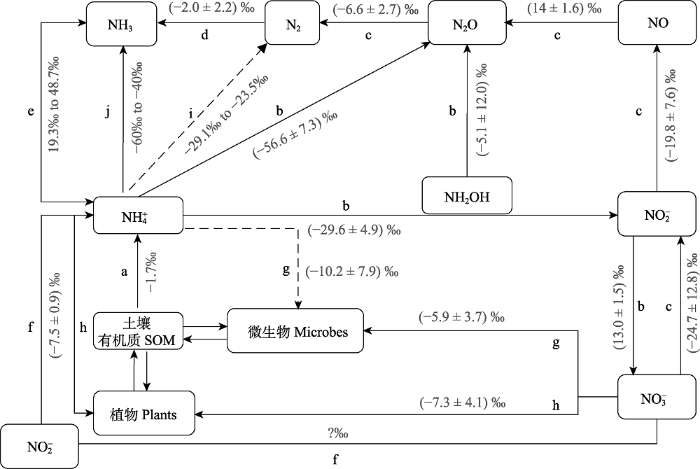 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}