朱林
, , 1 , 2 , * , 王甜甜
1 , 2 , 赵学琳
1 , 2 , 祁亚淑
3 , 许兴
1 , 2 1 宁夏大学西北土地退化与生态恢复国家重点实验室培育基地, 银川 7500212 宁夏大学西北退化生态系统恢复与重建教育部重点实验室, 银川 7500213 宁夏中卫市农业技术推广与培训中心, 宁夏中卫 755099Hydraulic lift of Medicago sativa and Astragalus laxmannii and its effect on their neighborhood plants Lin ZHU
, , 1 , 2 , * , Tian-Tian WANG
1 , 2 , Xue-Lin ZHAO
1 , 2 , Ya-Shu QI
3 , Xing XU
1 , 2 1 State Key Laboratory Breeding Base of Land Degradation and Ecological Restoration of Northwestern China, Ningxia University, Yinchuan 750021, China 2 Key Laboratory for Restoration and Reconstruction of Degraded Ecosystem in North-western China, Ministry of Education, Ningxia University, Yinchuan 750021, China 3 Center for Extension and Training of Agricultural Technique, Zhongwei, Ningxia 755099, China 通讯作者: *朱林: ORCID:0000-0002-1234-5837,E-mail:
zhulinscience@126.com 编委: 李彦
责任编辑: 李敏
收稿日期: 2019-12-3
接受日期: 2020-06-17
网络出版日期: 2020-07-20
基金资助: 国家自然科学基金 (31160478 )国家自然科学基金 (31830135 )宁夏回族自治区牧草育种专项 (2014NYYZ0401 )宁夏自然科学基金 (NZ17040 )
Corresponding authors: >*ORCID:0000-0002-1234-5837,E-mail:
zhulinscience@126.com Received: 2019-12-3
Accepted: 2020-06-17
Online: 2020-07-20
Fund supported: National Natural Science Foundation of China (31160478 )National Natural Science Foundation of China (31830135 )Forage Breeding Program of Ningxia Hui Autonomous Region (2014NYYZ0401 )Natural Science Foundation of Ningxia Hui Autonomous Region (NZ17040 )
摘要 为探讨紫花苜蓿(
Medicago sativa )及斜茎黄耆(沙打旺,
Astragalus laxmannii )与禾本科牧草混播后的水力提升现象, 揭示深、浅根性牧草的种间关系, 为混播草地的建植提供理论依据, 该研究开展了室外“上下盆”分根盆栽试验、采用土壤水分测定及“重水” (D
2 O, 氘(D)含量>99.9%)标记法估算了苜蓿及斜茎黄耆分别与‘冬牧70’黑麦(
Secale cereal ‘Dongmu 70’)不同混播比例(豆科:禾本科分别为3:7、5:5、7:3)条件下水力提升的发生情况及其对伴生牧草生长生理性状的影响。结果表明: ‘冬牧70’黑麦与斜茎黄耆混播后的产量显著高于其与紫花苜蓿混播后的产量, 同一种禾豆牧草混播组合不同混播比例中, 以AC2 (紫花苜蓿:‘冬牧70’黑麦为5:5)和BC3 (斜茎黄耆:‘冬牧70’黑麦为7:3)混播组合的总产量最高。不同单混播组合的单株整个生育期内日均提水量存在显著差异, 两种豆科牧草在混播时日均提水量均高于单播时, 斜茎黄耆单混播时的日均提水量显著高于紫花苜蓿, BC2组合(斜茎黄耆:‘冬牧70’黑麦为5:5)的日均提水量高于其他混播组合。在用标记水处理下盆土壤后, 各组合上下盆土壤水氢稳定同位素比率(
δ D)值显著升高。不同禾豆牧草组合上盆土壤水
δ D及禾本科牧草茎秆水
δ D、整株碳同位素分辨率(Δ
13 C)和产量数据表明, 在斜茎黄耆与‘冬牧70’黑麦混播比例为3:7、紫花苜蓿与‘冬牧70’黑麦混播比例为5:5时, 禾本科牧草水分状况或产量好于其他混播比例。以上结果表明, 两种深浅根豆科牧草与浅根性禾本科牧草混播种植时发生了水力提升现象, 两种豆科牧草提升的水分可以被伴生的禾本科牧草所吸收利用。
关键词: 禾豆牧草混播 ;
水力提升 ;
稳定同位素技术 Abstract Aims A variety of plant species can release soil water which is absorbed from wet soil layers to the dry soil layers during the period when transpiration rate is low (hydraulic lift). The purpose of this study is to explore the phenomenon of hydraulic lift for the mixed sowing combinations of two leguminous forages with gramineous forage, uncover the inter-specific relationship between forages with deep and shallow roots and provide theoretical basis for the establishment of mixed grassland.
Methods A split-root pot experiment was conducted outdoors. Two leguminous forages
Medicago sativa and
Astragalus laxmannii were sowed individually or sowed with a gramineous forage (
Secale cereal ‘Dongmu 70’) with proportions of 3:7, 5:5 and 7:3. While the soil water content was measured continuously, deuteroxide (D
2 O, D% > 99.9%) tracing technique was adopted to test the occurrence of hydraulic lift and to calculate the lift water quantity.
Important findings The yield of
S. Cereal ‘Dongmu 70’ in mixed sowing with
A. laxmannii was higher than that with
M. sativa. The yield of AC2 (
Medicago sativa :gramineae being 5:5) and BC3 (
A. laxmannii : gramineae being 7:3) was higher than other mixed sowing proportions in the same mixed sowing combination. There was a significant difference in the average daily water lift during the whole growth period among different single sowing or mixed sowing combinations. The lift water amount for a given leguminous forage was higher in mixed sowing combinations than in single sowing combinations.
A. laxmannii displayed higher daily water lift quantity regardless of single sowing or mixed sowing. The average daily water lift of BC2 (
A. laxmannii : gramineae being 5:5) was significantly higher than other combinations. Hydrogen stable isotope ratio (
δ D) of soil water in upper and bottom pots significantly increased after treated with deuteroxide tracer. The values of soil water
δ D in the upper pot, xylem water
δ D, whole plant carbon isotope discrimination (Δ
13 C) and yield of
Secale cereal ‘Dongmu 70’ have provided the evidence that the water status or yielding performance of
S. cereal ‘Dongmu 70’ in the combinations of BC1 (
A. laxmannii : gramineae being 3:7) and AC2 was better than other mixed combinations. This study validated the hypothesis that the phenomenon of hydraulic lift occurred when the deep-rooting leguminous and gramineous forages with shallow root were sown together. The water lift by the tested leguminous forages has been absorbed by their neighboring gramineous forage.
Keywords: mixed sowing between leguminous forage and gramineous forage ;
hydraulic lift ;
stable isotopic technique PDF (1803KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 朱林, 王甜甜, 赵学琳, 祁亚淑, 许兴. 紫花苜蓿和斜茎黄耆水力提升作用及其对伴生植物的效应. 植物生态学报, 2020, 44(7): 752-762. DOI:
10.17521/cjpe.2019.0337 ZHU Lin, WANG Tian-Tian, ZHAO Xue-Lin, QI Ya-Shu, XU Xing.
Hydraulic lift of Medicago sativa and Astragalus laxmannii and its effect on their neighborhood plants .
Chinese Journal of Plant Ecology , 2020, 44(7): 752-762. DOI:
10.17521/cjpe.2019.0337 植物处于深层湿润土壤中的根系吸收的水分可以在夜间蒸腾降低后通过输导组织运输并释放到浅层较干燥的土壤中, 即存在水力提升的现象(
Caldwell et al ., 1998 )。迄今为止, 越来越多的实验证实了水力提升现象的存在, 研究者们观测到很多物种, 包括乔木、灌木、藤本、牧草和农作物等约60种植物存在水分提升作用(
李唯等, 2007 ;
刘小莉等, 2007 ;
郝兴明等, 2009 ;
阿拉木萨和周丽芳, 2011 ;
Brooksbank et al ., 2011 ;
Couvreur et al ., 2014 ;
鱼腾飞等, 2014 ;
Meunier et al ., 2017 ;
李宏侠等, 2018 )。
深根性植物水力提升作用所释放的水分可以改善水分在土壤中的分布格局, 促进土壤养分分解和矿化过程, 有利于植物对养分的吸收, 促进养分循环(
Caldwell et al ., 1998 ;
何维明和张新时, 2001 ;
李宏侠等, 2018 )。旱区土壤水分是植物生存发展的关键生态因子, 深根植物提升的水分有利于维持相邻浅根植物的生存, 在调节生态系统水分平衡方面有重要的作用。在旱地农业或草地农业生产中, 将具有水力提升功能的深根植物与浅根植物套种, 通过发展粮-粮、草-粮或者草-草等间混套作模式, 可能是充分利用干旱半干旱地区土壤养分和自然降水资源, 提高作物产量的一种有效途径。
豆科与禾本科牧草混播可以有效地提高单位面积草地饲草产量、改善饲用品质、提高土壤肥力, 在世界上许多地区备受重视(
王旭等, 2007 )。研究表明, 紫花苜蓿(
Medicago sativa )与无芒雀麦(
Bromus inermis )混播后增产效果显著, 但混播组合中紫花苜蓿竞争优势明显, 要降低其比例才能使混播组分稳定(
张永亮等, 2019 )。在宁夏中部半干旱带有灌溉条件下斜茎黄耆(沙打旺,
Astragalus laxmannii )与禾本科牧草混播草地中不同物种生长较为均衡, 混播草地持续性好, 并且有较好的产量效应(
朱林等, 2014 )。
由于豆科与禾本科牧草地下生态位存在明显的不同, 当深根性的豆科牧草与浅根性的禾本科植物混播时, 植物根系分布格局的变化有可能导致资源在时间和空间上的协调分配(
吴序卉, 1999 )。研究表明在豆科/禾本科混播系统中存在一种氮共享的通道, 即豆科植物将固定的氮通过各种途径为伴生的禾本科植物提供氮源(
Eriksen et al ., 2010 )。例如, 在建植不久的白三叶(
Trifolium repens )/黑麦草(
Lolium perenne )混播草地中, 黑麦草通过吸收白三叶根系渗出液从而实现氮由豆科牧草向禾草的转移(
Paynel et al ., 2001 )。而水是植物种间营养物质运输的媒介, 禾豆牧草间氮再分配过程必然伴随着水分的共享。
李唯等(2007) 和
刘小莉等(2007) 分别通过上下桶分根试验证明了紫花苜蓿和斜茎黄耆都具有根系水力提升现象, 这就为豆禾牧草混、间作模式提供了可能的理论基础。
前人关于豆科牧草水力提升效应的研究仅限于通过土壤含水量的昼夜变化估算出来, 缺乏通过标记方法进行验证的过程。本研究通过设置室外“上下盆”分根盆栽试验, 采用土壤水分昼夜差值法及“重水”标记法研究紫花苜蓿、斜茎黄耆与‘冬牧70’黑麦(
Secale cereal ‘Dongmu 70’)不同混播比例条件下水力提升的发生情况及其对伴生牧草生长生理性状的影响。本研究旨在回答以下科学问题: 深根性豆科牧草紫花苜蓿及斜茎黄耆是否发生水力提升作用? 其提升的水分能否被伴生的浅根性植物所利用? 不同豆科牧草的水力提升量是否存在差异? 豆科牧草的水力提升对伴生的植物产生哪些效应? 不同豆科牧草的水力提升效应是否有所不同? 通过对上述科学问题的解答可以丰富生态学关于植物间正相互作用的理论, 揭示生态水文学中关于植物与土壤以及植物种间水分循环和互馈机制。在实践上, 对于混播草地建植也可以提供理论依据和技术支撑。
1 材料和方法 1.1 材料 该实验供试的牧草有3个, 其中, 豆科作物采用紫花苜蓿和斜茎黄耆, 紫花苜蓿及斜茎黄耆均为优良的豆科牧草, 对于促进畜牧业发展、生态环境建设和保护等方面具有重要的意义。这两种豆科牧草的根系都十分发达, 对深层土壤水分的利用能力很强(
山仑等, 2008 ;
杨永东等, 2008 )。本实验采用的紫花苜蓿品种是‘美国牧歌401’, 在实验中的编号为A, 斜茎黄耆为当地栽培种, 在实验中的编号为B。混播的禾本科作物是‘冬牧70’黑麦, 在实验中的编号为C。
1.2 研究区概况 该盆栽实验在宁夏大学农科实训基地进行, 该实训基地位于宁夏银川市, 地处西北内陆, 属于典型的中温带大陆性气候, 年平均气温8-9 ℃, 无霜期较短, 一般在157-170天, 全年日照时间2 800-3 000 h, 年降水量200 mm左右, 主要集中在7-9月。供试土壤全磷含量0.353 g·kg
-1 , 全氮含量0.187 g·kg
-1 , 全钾含量17.0 g·kg
-1 , 有机质含量2.65 g·kg
-1 , 速效磷含量4.63 mg·kg
-1 , 速效钾含量74.7 mg·kg
-1 , 碱解氮含量23.0 mg·kg
-1 。
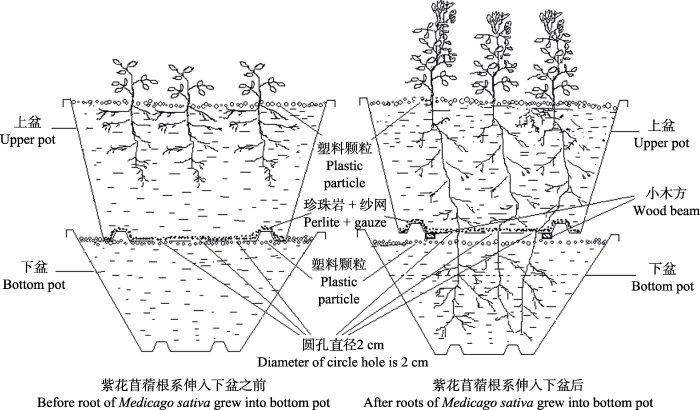
1.3 实验设计 采用两因素随机区组实验设计, 选用树脂花盆做成上下盆分根实验装置, 上盆规格为28 cm × 22 cm × 31 cm (上径×下径×高), 下盆内径32 cm、高17 cm的装置。上盆底钻5个直径为2 cm的孔, 将孔洞用尼龙纱网覆盖并在盆底铺0.5 cm厚的珍珠岩, 以便上盆植物根系穿过珍珠岩和纱网顺利向下生长, 将土壤按计算的量装入盆中(上盆是10.44 kg, 下盆是6.5 kg)。2014年5月5日将供试紫花苜蓿及斜茎黄耆种子分别播在上盆中, 土表用聚乙烯颗粒覆盖以防止土表水分蒸发。播种后保持上盆水分充足, 以便豆科作物充分生长。2015年4月20日, 将‘冬牧70’黑麦种子按照混播比例播在上盆中紫花苜蓿与斜茎黄耆的周围。待禾本科作物定苗后, 将H21土壤水分自动监测探头埋入上下盆土壤中, 以便连续监测土壤含水量。等豆科作物的根系扎入下盆后, 将上盆用小木块垫起来, 使上下盆隔离开, 为了防止土壤毛细管作用对上盆水分的补给, 在上下盆连接处用水把土冲掉, 只留根系与上盆相连(如
图1 )。为了确保豆科作物根系水力提升现象发生的有利条件, 将下盆土壤含水量均保持田间持水量的75%。对所有进行实验的盆栽进行遮雨处理以便隔绝降水对禾本科牧草生长生理指标的影响。为了验证豆科作物根系提水作用, 在7月17日采样前, 利用D
2 O (氘(D)含量>99.9%)稀释配制成氢稳定同位素比率(
δ D)值为277.9‰的标记水, 对下盆进行灌溉, 然后对上盆的植物、上下盆的土样进行氢同位素组成分析。该实验共设置8个单混播组合, 3次重复, AC1、AC2、AC3与BC1、BC2、BC3分别表示苜蓿、斜茎黄耆与黑麦的混播比例为3:7、5:5、7:3, AA及BB分别表示单播苜蓿及单播斜茎黄耆, 定苗后每盆均保留10株植物。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1上下盆提水实验示意图。 Fig. 1Upper and bottom pot test design for hydraulic lift. 1.4 测定项目及方法 (1)产量: 分别于2015年7月17日, 10月18日, 对每个盆里的所有植物都进行刈割采样, 3次重复, 将采集的样品在105 ℃下杀青10 min , 再在65 ℃下连续烘干至恒质量, 最后测定干质量并计算产量。
(2)碳同位素分辨率的测定: 2015年7月17日采样, 将采集的植物整株样品烘干粉碎并过0.25 mm的筛子, 各个植物样品在Flash 2000 HT元素分析仪(Thermo Scientific, Waltham, USA)中高温燃烧后生成CO
2 , 用Delta V Advantage同位素比率质谱仪(Thermo Scientific, Waltham, USA)检测CO
2 中
13 C与
12 C的比率(测定精度: 碳稳定同位素比率(
δ 13 C) ± <0.1‰), 参照国际标准物(Pee Dee Belnite)比对计算
δ 13 C)。各处理的碳同位素分辨率(?
13 C)则依据各自的
δ 13 C值并参照
Farquhar和Richards (1984) 的公式推导得出。
(3)氢稳定同位素比率(
δ D)的测定: 先对下盆灌适量稀释好的D
2 O, 留水样用于测定灌溉水的
δ D, 2015年7月17日采集植物样品和土壤样品。植物样品的采集方法是选取5-6株有代表性的植株, 刮去韧皮部取5 cm长的木栓化茎秆。为了验证标记水对上盆土壤水氢同位素组成的影响, 分别于标记前后采集上、下盆土壤样品, 采样深度为10-15 cm。所有采集的样品都及时装到8 mL的硼硅酸盐棕色玻璃瓶中, 用胶膜密封, 随后把灌溉水样在4 ℃下冷藏, 土壤样品和植物样品在-20 ℃条件下冷冻, 以备进行样品的水分提取。测定时使用同位素比率质谱法, 水在Flash 2000HT元素分析仪(Thermo Scientific, Waltham, USA)中经高温裂解后生成H
2 , 用质谱仪通过检测H
2 中的
2 H(D)与
1 H比率(测定精度±<1‰), 并与国际标准物(SMOW)比对后计算出样品的
δ 2 H (
δ D)值。
(5)土壤体积含水量(
θ, %)的测定以及监测: 为了连续监测上盆土壤体积含水量的变化趋势, 在定苗之后将土壤水分自动监测器的探头(HOBO H21- USB, Onset, Pocasset, USA)埋入上盆土壤中, 每小时记录一次数据。
(6)提水量
两种豆科牧草水分提升作用检测是通过上盆土壤水分的变化来确定的, 在间隔时段的观测中, 此次土壤水分的观测数值减去前一次观测数值, 如差值为正值, 则认为此期间发生了水分提升作用, 其差值为植物水分提升作用产生的土壤水分增加量(提升量)。再根据上盆土壤体积, 可以推算出提升的水量。根据参试豆科牧草的株数, 结合单位时间、单位体积土壤内水分含量的增加量, 获得单株豆科牧草产生的水分提升数量, 以合理地比较不同物种以及不同禾豆牧草组合中豆科牧草的水分提升能力(
阿拉木萨和周丽芳, 2011 )。
单株豆科牧草水力提升量的计算方法如下:
m water = (
ρ × Δ
θ ×
V soil )/
n 式中,
m 水 为每株豆科作物日水力提升量, 单位为g·plant
-1 ·d
-1 ); Δ
θ 为平均昼夜土壤含水量差值, 即Δ
θ =
θ night -
θ daytime , 单位为%;
V water 表示每个昼夜提升水的体积,
V soil 表示盆栽土壤的体积, 单位为cm
3 ;
ρ 表示水的密度, 其值为1 g·cm
-3 ;
n 为每盆豆科作物的株数。
1.5 数据处理及分析方法 采用Excel 2003以及SPSS 11.5数据处理软件对实验数据进行方差分析、计算标准偏差、相关分析等统计分析工作, 分别对混播组合间进行单因素方差分析, 采用Ducan新复极差法对各测定数据进行多重比较。采用SigmaPlot 10.0制图。
δ 13 C经计算转换成?
13 C之后再进行数据的分析处理, 二者之间的转换公式如下(
Farquhar & Richards, 1984 ):
δ 13 C(‰) = (
R plant /
R PDB - 1) × 1000?
13 C(‰) = [(
δ 13 C
air -
δ 13 C
plant ) × 1000]/(1 +
δ 13 C
plant )
其中,
R plant 表示植物样品的
13 C/
12 C值,
R PDB 表示国际标准物的
13 C/
12 C值,
δ 13 C
air 与
δ 13 C
plant 分别代表大气、植物的稳定碳同位素比率, 其中
δ 13 C
air = -7‰。
2 结果和分析 2.1 不同混播比例下各混播组合的产量 由
表1 可知, 禾豆牧草混播后, 豆科牧草的产量总体上高于禾本科牧草‘冬牧70’黑麦。斜茎黄耆单播产量显著高于紫花苜蓿, 两种豆科牧草与‘冬牧70’黑麦混播后产量显著下降(
p < 0.05)。斜茎黄耆与‘冬牧70’黑麦混播后的总产量显著高于紫花苜蓿与‘冬牧70’黑麦混播后的总产量(
p < 0.05), 同一种禾豆牧草混播组合不同混播比例中, 以AC2和BC3混播组合的总产最高。斜茎黄耆-‘冬牧70’黑麦混播组合中的禾本科牧草产量显著高于紫花苜蓿-‘冬牧70’黑麦混播组合中的禾本科牧草产量。在同一种豆科牧草与‘冬牧70’黑麦混播时, 以AC2和BC1组合中的禾本科产量为最高。
Table 1 表1 表1 不同混播组合下各混播组合的产量(平均值±标准偏差)
Table 1
Yield of forages in different sowing combinations (Mean ± SD ) 组合 Combination 豆科 Leguminosae 禾本科 Gramineae 禾+豆总产量 Sum of Leguminosae and Gramineae AA 5 802.90 ± 326.7b 5 802.9 ± 326.70b BB 7 861.99 ± 504.85a 7 861.99 ± 504.85a AC1 4 365.35 ± 232.90c 525.89 ± 52.98c 4 891.25 ± 285.88b AC2 4 832.31 ± 511.98c 703.87 ± 57.05c 5 536.18 ± 454.93b AC3 4 125.48 ± 518.69c 284.56 ± 20.69c 4 410.05 ± 539.38b BC1 4 829.60 ± 538.53c 2 617.29 ± 142.73a 7 266.63 ± 268.33a BC2 5 082.16 ± 184.05bc 1 501.70 ± 665.95b 6 583.86 ± 481.90a BC3 5 982.88 ± 490.54b 1 386.05 ± 545.77b 7 549.20 ± 72.24a
AA、BB表示紫花苜蓿及斜茎黄耆单播, AC1、AC2、AC3与BC1、BC2、BC3分别表示比例为3:7、5:5、7:3的禾豆牧草混播组合。不同小写字母表示不同混播方式间的产量差异显著(p < 0.05)。 AA and BB represent single sowing of Medicago sativa and Astragalus laxmannii, respectively. AC1, AC2, AC3, BC1, BC2 and BC3 represent the mixed sowing combinations of Medicago sativa and Astragalus laxmannii with gramineous forage in proportions of 3:7, 5:5 and 7:3, respectively. Different lowercase letters represent differences in yield at the level of 0.05. 新窗口打开 |
下载CSV 2.2 不同水分条件下各单混播组合土壤含水量的昼夜变化 图2 中两条曲线代表牧草生长期间上盆土壤水分的最大值(午夜)和最小值(正午)。不同单混播组合的上盆土壤含水量在7月下旬至8月上旬、9月上旬和9月下旬出现了3次峰值, 这主要与人工补水有关。总体说来, 8月份之前斜茎黄耆单播时上盆土壤含水量高于紫花苜蓿单播时的上盆土壤含水量。在禾豆混播组合中, BC1的上盆土壤含水量在9月中旬前高于AC1上盆土壤含水量; BC2上盆土壤含水量在整个生育期内均高于AC2上盆土壤含水量; BC3上盆土壤含水量仅在9月下旬低于AC3混播组合的上盆土壤含水量。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图24-10月份各单混播组合土壤含水量的昼夜变化。 AA、BB表示紫花苜蓿及斜茎黄耆单播, AC1、AC2、AC3与BC1、BC2、BC3分别表示比例为3:7、5:5、7:3的禾豆牧草混播组合。
Fig. 2Differences in the soil water content between the mid-day and mid-night for the different mixed sowing or single sowing combinations from April to October. AA and BB represent single sowing of Medicago sativa and Astragalus laxmannii, respectively. AC1, AC2, AC3, BC1, BC2 and BC3 represent the mixed sowing combinations of Medicago sativa and Astragalus laxmannii with gramineous forage in proportions of 3:7, 5:5 and 7:3, respectively.
根据
图3 , 两个单混播组合(AA和BB)的土壤含水量昼夜差值(上盆土壤含水量的最大值-最小值, 是计算水力提升量的依据)分别为1.95%和2.33%; 紫花苜蓿与黑麦混播的3个组合土壤含水量昼夜差值分别为1.99%、2.64%和2.46%; 斜茎黄耆与黑麦混播的3个组合土壤含水量昼夜差值分别为2.42%、3.70%和3.31%。从以上数据可以得出以下规律: 斜茎黄耆单播处理的土壤含水量昼夜差值大于紫花苜蓿单播(
p < 0.05), BC2的土壤昼夜含水量差值显著高于其他组合, BC3的平均土壤含水量最高(
p < 0.05)。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图34-10月份各单混播组合土壤含水量日最大值、最小值及昼夜差值(平均值+标准偏差)。 AA、BB表示紫花苜蓿及斜茎黄耆单播, AC1、AC2、AC3与BC1、BC2、BC3分别表示比例为3:7、5:5、7:3的禾豆牧草混播组合。不同小写字母表示不同混播方式间的土壤含水量差异显著(
p < 0.05)。
Fig. 3The maximum, minimum of daily soil water content and differences in the soil water content between the mid-day and mid-night for the different mixed sowing or single sowing combinations from April to October (mean + SD ). AA and BB represent single sowing of
Medicago sativa and
Astragalus laxmannii , respectively. AC1, AC2, AC3, BC1, BC2 and BC3 represent the mixed sowing combinations of
Medicago sativa and
Astragalus laxmannii with gramineous forage in lowercase proportions of 3:7, 5:5 and 7:3, respectively. Different lowercase letters represent differences in soil water content between different mixed sowing combinations at the level of 0.05.
2.3 不同水分条件下各混播组合中豆科作物根系每天的提水量 表2 是两种豆科牧草单播及与‘冬牧70’黑麦混播后的全生育期日平均提水量, 在整个生育期间, 不同单混播组合的单株日平均提水量存在显著的差异。两种豆科牧草在混播条件下日平均提水量均高于单播条件, 斜茎黄耆单混播条件下的日平均提水量均高于苜蓿, BC2的日平均提水量显著高于其他组合。
Table 2 表2 表2 各混播组合中平均每株豆科作物根系每天的提水量
Table 2
Hydraulic lift water of leguminous crops in mixed combinations of different water treatments 组合 Combination 平均值 Mean (g·plant-1 ·d-1 ) 标准偏差 SD AA 36.63d 3.07 AC1 37.55d 4.51 AC2 46.58c 8.66 AC3 47.36c 6.37 BB 44.41c 7.15 BC1 45.65c 5.08 BC2 70.09a 6.92 BC3 62.70b 10.11 F (df = 3)37.25**
AA、BB表示紫花苜蓿及斜茎黄耆单播, AC1、AC2、AC3与BC1、BC2、BC3分别表示比例为3:7、5:5、7:3的禾豆牧草混播组合。不同小写字母表示不同混播方式间的提水量差异显著(p < 0.05)。 AA and BB represent single sowing of Medicago sativa and Astragalus laxmannii, respectively. AC1, AC2, AC3, BC1, BC2 and BC3 represent the mixed sowing combinations of Medicago sativa and Astragalus laxmannii with gramineous forage in proportions of 3:7, 5:5 and 7:3, respectively. Different lowercase letters represent differences in hydraulic lift water between different mixed sowing combinations at the level of 0.05. 新窗口打开 |
下载CSV 2.4 不同水分处理及混播比例下各混播组合上下盆土壤水氢同位素比率(δ D)的变化 由
图4 可知, 不同组合上下盆土壤
δ D值在用标记处理后显著升高, 下盆土壤
δ D值在23.38‰- 31.76‰, 而上盆土壤
δ D值相比处理前的-79.59‰- -70.82‰也显著地增加到-29.94‰- -15.62‰。总体说来, 斜茎黄耆与禾本科牧草单混播后上盆土壤水
δ D值高于紫花苜蓿与禾本科牧草混播的上盆土壤水
δ D值, 斜茎黄耆与‘冬牧70’黑麦混播后上盆土壤水
δ D值的排序为BC1 > BC3 > BC2, 紫花苜蓿与‘冬牧70’黑麦混播后上盆
δ D值的排序为AC2 > AC1 > AC3。说明在同位素标记期间斜茎黄耆与‘冬牧70’黑麦混播比例为3:7时下盆土壤水对上盆的贡献率最高, 而紫花苜蓿与‘冬牧70’黑麦混播比例为5:5时下盆土壤水对上盆的贡献率最高。
图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4不同单混播组合下盆、上盆土壤水氢稳定同位素比率(δ D)(平均值+标准偏差)。16个土壤样品来自8个单混播组合的上下盆土壤。 AA、BB表示紫花苜蓿及斜茎黄耆单播, AC1、AC2、AC3与BC1、BC2、BC3分别表示比例为3:7、5:5、7:3的禾豆牧草混播组合。
Fig. 4Soil water hydrogen stable isotope ratio (δ D) in the bottom and upper pots of different combinations (mean + SD ). Sixteen soil samples from the upper and bottom pots of eight single or mixed sowing combinations, totally. AA and BB represent single sowing of
Medicago sativa and
Astragalus laxmannii, respectively. AC1, AC2, AC3, BC1, BC2 and BC3 represent the mixed sowing combinations of
Medicago sativa and
Astragalus laxmannii with gramineous forage in proportions of 3:7, 5:5 and 7:3, respectively.
2.5 不同混播组合植物茎秆水δ D的变化 图5 为第一次刈割时不同组合中植物茎秆水
δ D值, 总体来说, 参试的禾豆牧草茎秆水
δ D值相比处理前土壤水
δ D值均显著升高, 说明标记水已被这3种植物所吸收利用。豆科牧草茎秆水
δ D值低于禾本科牧草, 说明豆科牧草与禾本科牧草相比对上盆土壤水利用较多, 而禾本科牧草的生长较多地依赖豆科牧草从下盆吸收并释放到上盆的水分。单播条件下斜茎黄耆的茎秆水
δ D值低于紫花苜蓿, 说明单播斜茎黄耆对上盆土壤水分利用率高于紫花苜蓿。对于紫花苜蓿与‘冬牧70’黑麦的混播组合, 禾豆比例为5:5时紫花苜蓿及‘冬牧70’黑麦茎秆水
δ D值为最高; 对于斜茎黄耆与‘冬牧70’黑麦混播组合而言, 在豆禾比为3:7时, ‘冬牧70’黑麦茎秆水
δ D值最高, 说明在以上两种禾豆混播比例时, 豆科牧草水力提升作用对伴生的禾本科牧草水分贡献率最高。
图5 新窗口打开 |
下载原图ZIP |
生成PPT 图5各单混播组合植物茎秆氢稳定同位素比率(δ D)(平均值+标准偏差)。14个植物样品来自8个单混播组合。 AA、BB表示紫花苜蓿及斜茎黄耆单播, AC1、AC2、AC3与BC1、BC2、BC3分别表示比例为3:7、5:5、7:3的禾豆牧草混播组合。
Fig. 5Plant stem water hydrogen stable isotope ratio (δ D) of single or mixed sowing combinations (mean + SD ). Fourteen plant samples from eight single or mixed sowing combinations, totally. AA and BB represent single sowing of
Medicago sativa and
Astragalus laxmannii, respectively. AC1, AC2, AC3, BC1, BC2 and BC3 represent the mixed sowing combinations of
Medicago sativa and
Astragalus laxmannii with gramineous forage in proportions of 3:7, 5:5 and 7:3, respectively.
2.6 不同水分处理、混播比例下各混播组合中?13 C的变化 由
图6 可知, 两种豆科牧草单播条件下, 紫花苜蓿的整株?
13 C显著高于斜茎黄耆, 说明前者的蒸腾效率低于后者。禾豆混播后, 除AC1组合禾豆牧草整株?
13 C差异不显著外其他各混播组合中的禾本科牧草整株?
13 C均高于豆科牧草。紫花苜蓿与‘冬牧70’黑麦混播时其整株?
13 C值在AC1和AC2组合中有所增加, 而斜茎黄耆与‘冬牧70’黑麦混播后在BC3组合中其整株?
13 C与单播时的整株?
13 C差异不显著, 而在其他两个组合中斜茎黄耆整株?
13 C均显著低于其单播整株?
13 C。BC1及BC3组合中的‘冬牧70’黑麦整株?
13 C分别略高于AC1和AC3中‘冬牧70’黑麦整株?
13 C, 说明在豆禾比为3:7和7:3时, 斜茎黄耆与‘冬牧70’黑麦混播组合中禾本科牧草的水分状况好于紫花苜蓿与‘冬牧70’黑麦混播组合中的禾本科牧草水分状况。对于同一种豆科牧草, 紫花苜蓿的整株?
13 C值随着混播组合中豆科比例的增加呈先升后降的趋势, 而斜茎黄耆的整株?
13 C值随着其在混播组合中比例的增加而增加。AC2组合中‘冬牧70’黑麦整株?
13 C高于AC1和AC2组合中的‘冬牧70’黑麦整株?
13 C。
图6 新窗口打开 |
下载原图ZIP |
生成PPT 图6不同水分处理、混播比例下各混播组合中碳同位素分辨率(?13 C)的变化(平均值+标准偏差)。14个植物样品来自8个单混播组合。 AA、BB表示紫花苜蓿及斜茎黄耆单播, AC1、AC2、AC3与BC1、BC2、BC3分别表示比例为3:7、5:5、7:3的禾豆牧草混播组合。不同小写字母表示不同混播方式间的植物整株?
13 C值差异显著(
p < 0.05)。
Fig. 6Comparison of ?13 C in each combination under different water treatments and mixed seeding rates (mean + SD ). Fourteen plant samples from eight single or mixed sowing combinations, totally. AA and BB represent single sowing of
Medicago sativa and
Astragalus laxmannii, respectively. AC1, AC2, AC3, BC1, BC2 and BC3 represent the mixed sowing combinations of
Medicago sativa and
Astragalus laxmannii with gramineous forage in lowercase proportions of 3:7, 5:5 and 7:3, respectively. Different lowercase letters represent differences in whole plant ?
13 C between different mixed sowing combinations at the level of 0.05.
3 讨论 3.1 各混播组合中豆科牧草水力提升作用的判定 已有的水力提升测定方法有很多种, 采用较多的是以下两种方法: 1)测定植物根系周围土壤水分含量或水势的变化, 即通过将植物地上部遮阴或在夜间植物蒸腾速率为0或很低时测定和计算根区土壤含水量或水势的昼夜差值来反映水力提升作用; 2)氢同位素示踪方法, 即通过对植物深层根系施以重同位素标记水(
Pe?uelas & Filella, 2003 )或酸性品红作为示踪剂处理(
何维明和张新时, 2001 ), 再测定浅层土壤水以及植物茎秆水氢同位素组成的变化, 来判明是否发生了水力提升现象并避免了土壤毛管水的影响(
Dawson, 1996 )。
即时域反射技术(TDR)具有数据采集准确及时、相对误差小、对土壤扰动小、操作简单、结果可靠等优势, 目前在测定土壤含水量方面得到普遍应用。近年来, 许多研究者采用分根装置与TDR水分测定系统相结合研究了不同植物的根系提水作用(
Wan et al ., 2000 ;
Hirota et al ., 2004 ;
薛小红等, 2007 ;
阿拉木萨和周丽芳, 2011 )。此后, TDR与分根装置的结合成为研究水分提升作用的一种新方法, 这种方法操作简便, 结果可靠, 但关键在于分根装置设计时要注意消除土壤毛管上升水对实验结果的影响(
薛小红等, 2007 )。根据已发表的上下桶实验结果, 上层干燥土壤含水量呈现出有规律的昼夜变化趋势, 即在下午土壤含水量急剧下降, 而在午夜至凌晨时段土壤含水量最高(
李唯等, 2007 ;
薛小红等, 2007 ;
阿拉木萨和周丽芳, 2011 )。我们在制作“上下盆”实验装置时, 在豆科牧草根系长入下盆后, 将上盆用小木块垫起, 使上下盆分离, 并用水把上下盆连接处的土壤冲掉, 只留根系与上盆相连, 从而防止了下盆土壤毛细管作用对上盆水分的补给。同时, 在实验过程中对上盆做了遮雨处理, 因而上盆土壤水分的变化只受植物因素的影响。在我们的实验结果中, 斜茎黄耆土壤含水量昼夜差值与
薛小红等(2007) 报道的最大土壤含水量昼夜差值相近, 两种豆科牧草上盆土壤含水量昼夜差值及提水量存在显著差异, 斜茎黄耆无论在单播还是混播时上盆土壤含水量昼夜差值及日平均提水量均显著高于紫花苜蓿(
表2 ), 说明斜茎黄耆的水力提升效果好于紫花苜蓿。
在对下盆土壤进行D
2 O标记后, 不同混播组合的上下盆土壤水
δ D值在用标记处理后均显著升高, 证明紫花苜蓿和斜茎黄耆都发生了水力提升作用。斜茎黄耆单混播时上盆土壤水
δ D值高于紫花苜蓿, 说明更多的标记水被斜茎黄耆从下盆土壤中吸收并释放到上盆土壤中, 这与斜茎黄耆单混播时日平均提水量更高的结果是一致的。结合紫花苜蓿在单混播时的提水量以及上盆土壤含水量均低于斜茎黄耆的情况分析, AA和AC3的上盆土壤水
δ D值低于其他组合, 主要与标记时下盆对上盆较低的土壤水分贡献率有关。
在我们试验结果中, 单播紫花苜蓿茎秆水
δ D值为-11.71‰, 单播斜茎黄耆茎秆水
δ D值为-22.12‰, 紫花苜蓿混播时茎秆水
δ D值在-8.39‰- -4.60‰之间, 斜茎黄耆混播时茎秆水
δ D值在-19.35‰- -6.64‰之间, 两种豆科牧草在单混播时的茎秆水
δ D值均在上下盆土壤
δ D值区间内, 说明两种豆科牧草生长所利用的水源为上盆或下盆中的土壤水。同时, 参试的豆科牧草茎秆水
δ D值相比处理前土壤水
δ D值均显著升高(
图2 ,
图4 ), 说明标记水已被这两种植物所吸收利用。BB、BC2和BC3组合中斜茎黄耆的茎秆水
δ D值均较低(
图4 ), 说明在这些组合中斜茎黄耆对上盆土壤水利用程度比其他组合高, 这可以用同位素短期标记与豆科牧草水力提升作用长期效果的矛盾来解释。在同位素标记之前, 斜茎黄耆通过较强的水力提升作用(
表2 )将下盆水分提升并释放到上盆土壤中, 使上盆土壤水分含量增加(
图2 ), 而此时还没有进行同位素标记, 土壤水
δ D值是较低的。在同位素标记时, 由于上盆土壤含水量较高, 导致豆科植物释放的标记水较少, 或者是上盆原有的土壤水“稀释”了标记水, 使上盆土壤水
δ D值较低或者升高不显著, 而斜茎黄耆对这部分水吸收利用率较高, 使其茎秆水
δ D值较低。同时, 也可能由于重力的作用, 标记前提升的水分主要存在于上盆底部而被斜茎黄耆吸收利用, 使其茎秆水
δ D值较低。
3.2 两种豆科牧草水力提升作用对伴生植物的效应 在我们的实验结果中, ‘冬牧70’黑麦与不同豆科牧草混播后的产量存在显著差异, 斜茎黄耆与‘冬牧70’黑麦混播组合中的禾本科牧草产量显著高于紫花苜蓿与‘冬牧70’黑麦混播组合中的禾本科牧草产量。以上结果一方面与紫花苜蓿地上部分对于禾草的竞争力较高, 其旺盛的枝叶对禾草遮阴效应较强而影响了伴生禾草生长(
朱林等, 2014 ;
张永亮等, 2019 ), 另一方面也可能与斜茎黄耆水力提升作用高于紫花苜蓿有关。在整个生长季节, 斜茎黄耆与‘冬牧70’黑麦混播组合的土壤含水量总体说来高于紫花苜蓿-‘冬牧70’黑麦(
图2 ), 而且斜茎黄耆的提水量总体上比紫花苜蓿的高, 因此, 斜茎黄耆与‘冬牧70’黑麦组合中上盆土壤较高的含水量促进了混播组合中禾本科牧草的生长。在同一种豆科牧草与‘冬牧70’黑麦混播时, 以AC2和BC1组合中的禾本科产量为最高, 说明在紫花苜蓿与‘冬牧70’黑麦比例为5:5、斜茎黄耆与‘冬牧70’黑麦比例为3:7混播时, 豆科牧草水力提升作用对伴生的禾本科牧草生长的贡献最大, 这可能是不同禾豆比例的盆栽中豆科牧草群体产生的提水量与其对禾本科牧草水分竞争相“权衡”的结果。在不同混播组合中, ‘冬牧70’黑麦茎秆水
δ D值与处理前土壤水
δ D值相比均显著上升, 并介于上盆土壤水
δ D值和下盆土壤水
δ D值之间, 说明两种豆科牧草提升的标记水被伴生的禾本科牧草所吸收利用。在紫花苜蓿与‘冬牧70’黑麦的不同混播比例中, 以AC2组合中禾本科牧草茎秆水
δ D值为最高; 在斜茎黄耆与‘冬牧70’黑麦的不同混播比例中, 以BC1组合中禾本科牧草茎秆水
δ D值最高(
图4 ), 说明以上两个混播组合中的禾本科牧草生长所利用的水分主要来自于豆科牧草的水力提升作用。对于同一种豆科牧草与‘冬牧70’黑麦的混播组合, AC2组合中禾本科整株Δ
13 C和产量最高(
图5 ), BC1组合中禾本科产量最高(
表1 ), 这一结果支持了上述结论。
3.3 各混播组合中不同牧草的水分利用特征 C
3 植物的Δ
13 C与植物蒸腾效率及环境水分状况有密切的关系, 较低的气孔导度或较高的光合速率 会降低Δ
13 C (
Farquhar et al ., 1989 ;
Badeck et al ., 2005 ;
孙谷畴等, 2008 ; 朱林等, 2016)。对于C
3 植物, Δ
13 C与
C i /
C a 呈正相关关系, 而
C i /
C a 与气孔导度(
G s )正相关而与蒸腾效率(
TE )负相关。水分胁迫会使
G s 下降, 从而导致更低的Δ
13 C值和更高的
TE (
Farquhar & Richards, 1984 ;
Farquhar et al ., 1989 )。在本实验结果中, 斜茎黄耆整株?
13 C比紫花苜蓿的低(
图5 ), 说明斜茎黄耆气孔导度较低, 蒸腾效率较高。这可能与其遗传特性有关,
徐炳成等(2005) 和
山仑等(2008) 研究发现紫花苜蓿对水分条件变化的敏感性高于斜茎黄耆, 斜茎黄耆在低水条件下蒸腾效率高于紫花苜蓿, 本实验中斜茎黄耆整株?
13 C显著低于紫花苜蓿的结果与之相一致。在“上干下湿”的“上下盆”土壤水分格局下, 紫花苜蓿对水分的高度敏感性促使其根系向下盆湿润土层生长, 并增加了其对下盆水分的吸收利用, 这也是紫花苜蓿有更高的整株Δ
13 C值的原因之一。
在本实验结果中, 禾本科整株?
13 C显著高于豆科牧草, 这种情况除了与物种间蒸腾效率的差异有关, 也可能是因为豆科牧草水力提升作用改善了上盆土壤水分状况使伴生的禾本牧草?
13 C增加。在各混播组合中, 随着‘冬牧70’黑麦比例的增加其整株Δ
13 C呈先升高后下降的趋势, 这可能是豆科牧草与伴生的禾本科牧草间水分竞争及水力提升双重作用的结果。对于同一种豆科牧草与‘冬牧70’黑麦混播时, 在禾豆比为5:5时, 禾本科整株Δ
13 C值均高于其他两个组合, 说明在这一比例下豆科牧草的水力提升作用对伴生的禾本科牧草生长的促进作用最大。在禾豆比为3:7时, 禾本科整株Δ
13 C值的下降可能是因为豆科牧草与其竞争水分, 使其受到水分胁迫, 导致气孔导度降低和较低的Δ
13 C值, 同时, AC1和BC1组合较低的提水量(
表2 )和较低的土壤含水量(
图1 )也说明这两个组合中的禾本科牧草水分状况较差, 导致其气孔导度和整株Δ
13 C值的下降。在禾豆比为7:3时, 禾本科整株Δ
13 C值的下降可能是禾本科牧草密度效应使其群体间水分竞争加剧而使禾本科牧草整株?
13 C值降低。
BC1混播组合中斜茎黄耆整株?
13 C值最低,可能是在斜茎黄耆比例较低时受到伴生的禾本科牧草水分竞争作用, 同时, 在BC1组合中, 单株提水量以及每盆斜茎黄耆的株数均较低, 暗示该混播群体中总的提水量较低, 上盆土壤水处于“入不敷出”状态, 而使植株水分状况变差, 导致斜茎黄耆整株?
13 C值较低; 在BC2和BC3组合中, 随着单株提水量和每盆总株数的增加, 土壤水分状况改善, 使斜茎黄耆整株?
13 C值增加。紫花苜蓿整株?
13 C随着混播组合中紫花苜蓿比例的增加呈先升后降的趋势, 暗示在AC1组合中紫花苜蓿比例低而禾本科比例高时, 因紫花苜蓿比禾本科牧草有更强的水分竞争力, 其水分状况较好; 而在AC2组合中, 随着紫花苜蓿在混播组合中比例的增加, 由于紫花苜蓿的单株提水量和株数的增加而使每盆总提水量增加, 因而改善了紫花苜蓿的水分状况; 对于AC3组合, 由于紫花苜蓿株数较多, 其在生长过程中过多地消耗了土壤水分导致其水分状况变差而使其整株?
13 C值下降。
综上所述, 采用土壤水分的昼夜变化及稳定同位素示踪法都指示紫花苜蓿及斜茎黄耆发生了水力提升现象, 提升的水分被伴生的浅根性牧草所吸收利用。不同物种间水力提升作用也有显著差异, 斜茎黄耆在单混播时的日平均提水量显著高于紫花苜蓿, 在斜茎黄耆与‘冬牧70’黑麦混播组合中禾本科牧草有更高的产量及?
13 C值, 斜茎黄耆水力提升效应高于紫花苜蓿。不同禾豆混播方式下豆科牧草的水力提升作用是不同的, 在紫花苜蓿与‘冬牧70’黑麦混播比例为7:3、斜茎黄耆与‘冬牧70’黑麦混播比例为5:5时豆科牧草提水量最大。在斜茎黄耆与‘冬牧70’黑麦混播比例为3:7、紫花苜蓿与‘冬牧70’黑麦混播比例为5:5时, 禾本科牧草水分状况或产量好于其他混播比例。本研究结果揭示了深、浅根性牧草种间的水分互作关系, 丰富了生态学以及生态水文学的相关理论, 同时也为混播草地的建植提供了理论依据。
[1] Alamusa Zhou LF 2011 ). Empirical test of hydraulic lift in 21 plant species in the Horqin sandy land, Inner MongoliaJournal of Beijing Forestry University 1 ), 70 -77 . [本文引用: 4] [ 阿拉木萨 , 周丽芳 (2011 ). 科尔沁沙地21种植物水分提升作用的实证检验北京林业大学学报 , 33(1 ), 70 -77 .] [本文引用: 4] [2] Badeck FW Tcherkez G Nogués S Piel C Ghashghaie J 2005 ). Post-photosynthetic fractionation of stable carbon isotopes between plant organs—A wide spread phenomenonRapid Communications in Mass Spectrometry 1381 -1391 . URL PMID:
15880634 [本文引用: 1] [3] Brooksbank K White DA Veneklaas EJ Carter JL 2011 ). Hydraulic redistribution in Eucalyptus kochii subsp. borealis with variable access to fresh groundwaterTrees 735 -744 . [本文引用: 1] [4] Caldwell MM Dawson TE Richards JH 1998 ). Hydraulic lift: consequences of water efflux from the roots of plantsOecologia 151 -161 . URL PMID:
28308192 [本文引用: 2] [5] Couvreur V Vanderborght J Draye X Javaux M 2014 ). Dynamic aspects of soil water availability for isohydric plants: focus on root hydraulic resistancesWater Resources Research 8891 -8906 . [本文引用: 1] [6] Dawson TE 1996 ). Determining water use by trees and forests from isotopic, energy balance and transpiration analyses: the roles of tree size and hydraulic liftTree Physiology 263 -272 . URL PMID:
14871771 [本文引用: 1] [7] Eriksen J Askegaard M S?egaard K 2010 ). Residual effect and nitrate leaching in grass-arable rotations: effect of grassland proportion, sward type and fertilizer historySoil Use & Management 373 -382 . [本文引用: 1] [8] Farquhar GD Ehleringer FR Hubick KT 1989 ). Carbon isotope discrimination and photosynthesisAnnual Review of Plant Biology 503 -537 . [本文引用: 2] [9] Farquhar GD Richards RA 1984 ). Isotopic composition of plant carbon correlates with water-use efficiency of wheat genotypesAustralian Journal of Plant Physiology 539 -552 . [本文引用: 3] [10] Hao XM Chen YN Li WH Guo B Zhao RF 2009 ). Evidence and ecological effects of hydraulic lift inPopulus euphratica . Chinese Journal of Plant Ecology 1125 -1131 . [本文引用: 1] [ 郝兴明 , 陈亚宁 , 李卫红 , 郭斌 , 赵锐锋 (2009 ). 胡杨根系水力提升作用的证据及其生态学意义植物生态学报 , 33, 1125 -1131 .] [本文引用: 1] [11] He WM Zhang XS 2001 ). Water sharing in the roots of four shrubs of the mu us sandy desertActa Phytoecologica Sinica 630 -633 . [本文引用: 2] [ 何维明 , 张新时 (2001 ). 水分共享在毛乌素沙地4种灌木根系中的存在状况植物生态学报 , 25, 630 -633 .] [本文引用: 2] [12] Hirota I Sakuratani T Sato T Higuchi H Nawata E 2004 ). A split-root apparatus for examining the effects of hydraulic lift by trees on the water status of neighboring cropsAgroforestry Systems 181 -187 . [本文引用: 1] [13] Li HX Yang XD Lü GH 2018 ). Relationships between hydraulic lift of Haloxylon ammodendron with growth condition, abundance and richness of shallow-rooted plantsBulletin of Soil and Water Conservation 75 -81 . [本文引用: 2] [ 李宏侠 , 杨晓东 , 吕光辉 (2018 ). 梭梭水力提升与浅根系植物优势度、丰富度和多度的关系水土保持通报 , 38, 75 -81 . ] [本文引用: 2] [14] Li W Hu ZZ Ni Y Li SZ Ni SL 2007 ). The relationship of hydraulic lift in alfalfa and maize and their drought resistance study on mechanism of hydraulic lift in root system of plant (II)Acta Agrestia Sinica 515 -518 . [本文引用: 3] [ 李唯 , 胡自治 , 倪郁 , 李尚忠 , 倪胜利 (2007 ). 苜蓿、玉米根系提水作用与耐旱性的关系——植物根系提水作用机理研究II草地学报 , 15, 515 -518 .] [本文引用: 3] [15] Liu XL Zhang HR Fu H 2007 ). Root hydraulic lift and nutrition activity in Astraglus adsurgens with fertilizationActa Botanica Boreali-Occidentalia Sinica 2507 -2513 . [本文引用: 2] [ 刘小莉 , 张洪荣 , 傅华 (2007 ). 施肥对沙打旺根系提水及土壤养分活性的影响西北植物学报 , 27, 2507 -2513 .] [本文引用: 2] [16] Meunier F Rothfuss Y Bariac T Biron P Richard P Durand JL Couvreur V Vanderborght J Javaux M 2017 ). Measuring and modeling hydraulic lift of Lolium multiflorum using stable water isotopesVadose Zone Journal 1 -15 . [本文引用: 1] [17] Paynel F Murray PJ Cliquet JB 2001 ). Root exudates: a pathway for short-term N transfer from clover to ryegrassPlant and Soil 235 -243 . [本文引用: 1] [18] Pe?uelas J Filella I 2003 ). Deuterium labeling of roots provides evidence of deep water access and hydraulic lift by Pinus nigra, in a Mediterranean forest of NE SpainEnvironmental and Experimental Botany 201 -208 . [本文引用: 1] [19] Shan L Zhang SQ Li WR 2008 ). Productivity and drought resistance of alfalfaJournal of Agricultural Science and Technology 12 -17 . [本文引用: 2] [ 山仑 , 张岁岐 , 李文娆 (2008 ). 论苜蓿的生产力与抗旱性中国农业科技导报 , 10, 12 -17 .] [本文引用: 2] [20] Sun GC Zhao P Cai XA Zeng XP Liu XJ Rao XQ Wang H 2008 ). Carbon isotope discrimination in leaf juice of Acacia mangium and its relation to water-use efficiencyChinese Journal of Ecology 497 -503 . [本文引用: 1] [ 孙谷畴 , 赵平 , 蔡锡安 , 曾小平 , 刘晓静 , 饶兴权 , 王华 (2008 ). 马占相思叶片液汁碳同位素甄别率和水分利用效率生态学杂志 , 27, 497 -503 .] [本文引用: 1] [21] Wan CG Xu WW Sosebee RE Machado S Archer T 2000 ). Hydraulic lift in drought-tolerant and susceptible maize hybridsPlant and Soil 117 -126 . [本文引用: 1] [22] Wang X Zeng ZH Hu YG Zhu B 2007 ). Progress and prospect on mixture of Gramineae herbage and Leguminosae herbageChinese Journal of Grassland 92 -98 . [本文引用: 1] [ 王旭 , 曾昭海 , 胡跃高 , 朱波 (2007 ). 豆科与禾本科牧草混播效应研究进展中国草地学报 , 29, 92 -98 .] [本文引用: 1] [23] Wu XH 1999 ). Root development of pea and oat mixture sward with plastic mulchingJournal of Gansu Agricultural University 120 -124 . [本文引用: 1] [ 吴序卉 (1999 ). 地膜覆盖下豆禾混播草地根系的动态研究甘肃农业大学学报 , 34, 120 -124 .] [本文引用: 1] [24] Xu BC Shan L Li FM 2005 ). Responses of Medicago sativa and Astragalus adsurgens seedlings growth and water use to soil moisture regimeChinese Journal of Applied Ecology 2328 -2332 . URL PMID:
16515181 [本文引用: 1] [ 徐炳成 , 山仑 , 李凤民 (2005 ). 苜蓿与沙打旺苗期生长和水分利用对土壤水分变化的反应应用生态学报 , 16, 2328 -2332 .] PMID:
16515181 [本文引用: 1] [25] Xue XH Niu DC Fu H Zhang HR 2007 ). Studies on mechanism of hydraulic lift by Astragalus laxmannii Acta Botanica Boreali-Occidentalia Sinica 2269 -2274 . [本文引用: 4] [ 薛小红 , 牛得草 , 傅华 , 张洪荣 (2007 ). 沙打旺根系提水作用及其机理研究西北植物学报 , 27, 2269 -2274 .] [本文引用: 4] [26] Yang YD Zhang JS Cai GJ Mo BR Chai CS Wang ZT 2008 ). Soil moisture dynamics of alfalfa pasture at different eco- sites in Gullied Loess AreaPratural Science 10 ), 25 -28 . [本文引用: 1] [ 杨永东 , 张建生 , 蔡国军 , 莫保儒 , 柴春山 , 王子婷 (2008 ). 黄土丘陵区不同立地条件下紫花苜蓿地土壤水分动态变化草业科学 , 25(10 ), 25 -28 .] [本文引用: 1] [27] Yu TF Feng Q Si JH Zhang XY 2014 ). Patterns, magnitude and controlling factors of hydraulic redistribution by Populus euphratica rootsJournal of Beijing Forestry University 2 ), 22 -29 . [本文引用: 1] [ 鱼腾飞 , 冯起 , 司建华 , 张小由 (2014 ). 胡杨根系水力再分配的模式、大小及其影响因子北京林业大学学报 , 36(2 ), 22 -29 .] [本文引用: 1] [28] Zhang YL Zhang LJ Yu TF Pan D 2019 ). Effects of grass- legume combinations and intercropping patterns on the forage yield and yield stabilityActa Agrestia Sinica 1410 -1418 . [本文引用: 2] [ 张永亮 , 张丽娟 , 于铁峰 , 潘东 (2019 ). 禾豆组合与间作方式对牧草产量及产量稳定性的影响草地学报 , 27, 1410 -1418 .] [本文引用: 2] [29] Zhu L Zheng SX Xu X Hou ZJ 2014 ). Effects of irrigation on mixed sowing of leguminous and graminaceous foragePratacultural Science 1752 -1760 . [本文引用: 2] [ 朱林 , 郑淑欣 , 许兴 , 侯志军 (2014 ). 宁夏中部灌水量对豆-禾牧草混播的影响草业科学 , 31, 1752 -1760 .] [本文引用: 2] 科尔沁沙地21种植物水分提升作用的实证检验
4
2011
... 植物处于深层湿润土壤中的根系吸收的水分可以在夜间蒸腾降低后通过输导组织运输并释放到浅层较干燥的土壤中, 即存在水力提升的现象(
Caldwell et al ., 1998 ).迄今为止, 越来越多的实验证实了水力提升现象的存在, 研究者们观测到很多物种, 包括乔木、灌木、藤本、牧草和农作物等约60种植物存在水分提升作用(
李唯等, 2007 ;
刘小莉等, 2007 ;
郝兴明等, 2009 ;
阿拉木萨和周丽芳, 2011 ;
Brooksbank et al ., 2011 ;
Couvreur et al ., 2014 ;
鱼腾飞等, 2014 ;
Meunier et al ., 2017 ;
李宏侠等, 2018 ). ...
... 两种豆科牧草水分提升作用检测是通过上盆土壤水分的变化来确定的, 在间隔时段的观测中, 此次土壤水分的观测数值减去前一次观测数值, 如差值为正值, 则认为此期间发生了水分提升作用, 其差值为植物水分提升作用产生的土壤水分增加量(提升量).再根据上盆土壤体积, 可以推算出提升的水量.根据参试豆科牧草的株数, 结合单位时间、单位体积土壤内水分含量的增加量, 获得单株豆科牧草产生的水分提升数量, 以合理地比较不同物种以及不同禾豆牧草组合中豆科牧草的水分提升能力(
阿拉木萨和周丽芳, 2011 ). ...
... 即时域反射技术(TDR)具有数据采集准确及时、相对误差小、对土壤扰动小、操作简单、结果可靠等优势, 目前在测定土壤含水量方面得到普遍应用.近年来, 许多研究者采用分根装置与TDR水分测定系统相结合研究了不同植物的根系提水作用(
Wan et al ., 2000 ;
Hirota et al ., 2004 ;
薛小红等, 2007 ;
阿拉木萨和周丽芳, 2011 ).此后, TDR与分根装置的结合成为研究水分提升作用的一种新方法, 这种方法操作简便, 结果可靠, 但关键在于分根装置设计时要注意消除土壤毛管上升水对实验结果的影响(
薛小红等, 2007 ).根据已发表的上下桶实验结果, 上层干燥土壤含水量呈现出有规律的昼夜变化趋势, 即在下午土壤含水量急剧下降, 而在午夜至凌晨时段土壤含水量最高(
李唯等, 2007 ;
薛小红等, 2007 ;
阿拉木萨和周丽芳, 2011 ).我们在制作“上下盆”实验装置时, 在豆科牧草根系长入下盆后, 将上盆用小木块垫起, 使上下盆分离, 并用水把上下盆连接处的土壤冲掉, 只留根系与上盆相连, 从而防止了下盆土壤毛细管作用对上盆水分的补给.同时, 在实验过程中对上盆做了遮雨处理, 因而上盆土壤水分的变化只受植物因素的影响.在我们的实验结果中, 斜茎黄耆土壤含水量昼夜差值与
薛小红等(2007) 报道的最大土壤含水量昼夜差值相近, 两种豆科牧草上盆土壤含水量昼夜差值及提水量存在显著差异, 斜茎黄耆无论在单播还是混播时上盆土壤含水量昼夜差值及日平均提水量均显著高于紫花苜蓿(
表2 ), 说明斜茎黄耆的水力提升效果好于紫花苜蓿. ...
... ;
阿拉木萨和周丽芳, 2011 ).我们在制作“上下盆”实验装置时, 在豆科牧草根系长入下盆后, 将上盆用小木块垫起, 使上下盆分离, 并用水把上下盆连接处的土壤冲掉, 只留根系与上盆相连, 从而防止了下盆土壤毛细管作用对上盆水分的补给.同时, 在实验过程中对上盆做了遮雨处理, 因而上盆土壤水分的变化只受植物因素的影响.在我们的实验结果中, 斜茎黄耆土壤含水量昼夜差值与
薛小红等(2007) 报道的最大土壤含水量昼夜差值相近, 两种豆科牧草上盆土壤含水量昼夜差值及提水量存在显著差异, 斜茎黄耆无论在单播还是混播时上盆土壤含水量昼夜差值及日平均提水量均显著高于紫花苜蓿(
表2 ), 说明斜茎黄耆的水力提升效果好于紫花苜蓿. ...
科尔沁沙地21种植物水分提升作用的实证检验
4
2011
... 植物处于深层湿润土壤中的根系吸收的水分可以在夜间蒸腾降低后通过输导组织运输并释放到浅层较干燥的土壤中, 即存在水力提升的现象(
Caldwell et al ., 1998 ).迄今为止, 越来越多的实验证实了水力提升现象的存在, 研究者们观测到很多物种, 包括乔木、灌木、藤本、牧草和农作物等约60种植物存在水分提升作用(
李唯等, 2007 ;
刘小莉等, 2007 ;
郝兴明等, 2009 ;
阿拉木萨和周丽芳, 2011 ;
Brooksbank et al ., 2011 ;
Couvreur et al ., 2014 ;
鱼腾飞等, 2014 ;
Meunier et al ., 2017 ;
李宏侠等, 2018 ). ...
... 两种豆科牧草水分提升作用检测是通过上盆土壤水分的变化来确定的, 在间隔时段的观测中, 此次土壤水分的观测数值减去前一次观测数值, 如差值为正值, 则认为此期间发生了水分提升作用, 其差值为植物水分提升作用产生的土壤水分增加量(提升量).再根据上盆土壤体积, 可以推算出提升的水量.根据参试豆科牧草的株数, 结合单位时间、单位体积土壤内水分含量的增加量, 获得单株豆科牧草产生的水分提升数量, 以合理地比较不同物种以及不同禾豆牧草组合中豆科牧草的水分提升能力(
阿拉木萨和周丽芳, 2011 ). ...
... 即时域反射技术(TDR)具有数据采集准确及时、相对误差小、对土壤扰动小、操作简单、结果可靠等优势, 目前在测定土壤含水量方面得到普遍应用.近年来, 许多研究者采用分根装置与TDR水分测定系统相结合研究了不同植物的根系提水作用(
Wan et al ., 2000 ;
Hirota et al ., 2004 ;
薛小红等, 2007 ;
阿拉木萨和周丽芳, 2011 ).此后, TDR与分根装置的结合成为研究水分提升作用的一种新方法, 这种方法操作简便, 结果可靠, 但关键在于分根装置设计时要注意消除土壤毛管上升水对实验结果的影响(
薛小红等, 2007 ).根据已发表的上下桶实验结果, 上层干燥土壤含水量呈现出有规律的昼夜变化趋势, 即在下午土壤含水量急剧下降, 而在午夜至凌晨时段土壤含水量最高(
李唯等, 2007 ;
薛小红等, 2007 ;
阿拉木萨和周丽芳, 2011 ).我们在制作“上下盆”实验装置时, 在豆科牧草根系长入下盆后, 将上盆用小木块垫起, 使上下盆分离, 并用水把上下盆连接处的土壤冲掉, 只留根系与上盆相连, 从而防止了下盆土壤毛细管作用对上盆水分的补给.同时, 在实验过程中对上盆做了遮雨处理, 因而上盆土壤水分的变化只受植物因素的影响.在我们的实验结果中, 斜茎黄耆土壤含水量昼夜差值与
薛小红等(2007) 报道的最大土壤含水量昼夜差值相近, 两种豆科牧草上盆土壤含水量昼夜差值及提水量存在显著差异, 斜茎黄耆无论在单播还是混播时上盆土壤含水量昼夜差值及日平均提水量均显著高于紫花苜蓿(
表2 ), 说明斜茎黄耆的水力提升效果好于紫花苜蓿. ...
... ;
阿拉木萨和周丽芳, 2011 ).我们在制作“上下盆”实验装置时, 在豆科牧草根系长入下盆后, 将上盆用小木块垫起, 使上下盆分离, 并用水把上下盆连接处的土壤冲掉, 只留根系与上盆相连, 从而防止了下盆土壤毛细管作用对上盆水分的补给.同时, 在实验过程中对上盆做了遮雨处理, 因而上盆土壤水分的变化只受植物因素的影响.在我们的实验结果中, 斜茎黄耆土壤含水量昼夜差值与
薛小红等(2007) 报道的最大土壤含水量昼夜差值相近, 两种豆科牧草上盆土壤含水量昼夜差值及提水量存在显著差异, 斜茎黄耆无论在单播还是混播时上盆土壤含水量昼夜差值及日平均提水量均显著高于紫花苜蓿(
表2 ), 说明斜茎黄耆的水力提升效果好于紫花苜蓿. ...
Post-photosynthetic fractionation of stable carbon isotopes between plant organs—A wide spread phenomenon
1
2005
... C
3 植物的Δ
13 C与植物蒸腾效率及环境水分状况有密切的关系, 较低的气孔导度或较高的光合速率 会降低Δ
13 C (
Farquhar et al ., 1989 ;
Badeck et al ., 2005 ;
孙谷畴等, 2008 ; 朱林等, 2016).对于C
3 植物, Δ
13 C与
C i /
C a 呈正相关关系, 而
C i /
C a 与气孔导度(
G s )正相关而与蒸腾效率(
TE )负相关.水分胁迫会使
G s 下降, 从而导致更低的Δ
13 C值和更高的
TE (
Farquhar & Richards, 1984 ;
Farquhar et al ., 1989 ).在本实验结果中, 斜茎黄耆整株?
13 C比紫花苜蓿的低(
图5 ), 说明斜茎黄耆气孔导度较低, 蒸腾效率较高.这可能与其遗传特性有关,
徐炳成等(2005) 和
山仑等(2008) 研究发现紫花苜蓿对水分条件变化的敏感性高于斜茎黄耆, 斜茎黄耆在低水条件下蒸腾效率高于紫花苜蓿, 本实验中斜茎黄耆整株?
13 C显著低于紫花苜蓿的结果与之相一致.在“上干下湿”的“上下盆”土壤水分格局下, 紫花苜蓿对水分的高度敏感性促使其根系向下盆湿润土层生长, 并增加了其对下盆水分的吸收利用, 这也是紫花苜蓿有更高的整株Δ
13 C值的原因之一. ...
Hydraulic redistribution in
Eucalyptus kochii subsp.
borealis with variable access to fresh groundwater
1
2011
... 植物处于深层湿润土壤中的根系吸收的水分可以在夜间蒸腾降低后通过输导组织运输并释放到浅层较干燥的土壤中, 即存在水力提升的现象(
Caldwell et al ., 1998 ).迄今为止, 越来越多的实验证实了水力提升现象的存在, 研究者们观测到很多物种, 包括乔木、灌木、藤本、牧草和农作物等约60种植物存在水分提升作用(
李唯等, 2007 ;
刘小莉等, 2007 ;
郝兴明等, 2009 ;
阿拉木萨和周丽芳, 2011 ;
Brooksbank et al ., 2011 ;
Couvreur et al ., 2014 ;
鱼腾飞等, 2014 ;
Meunier et al ., 2017 ;
李宏侠等, 2018 ). ...
Hydraulic lift: consequences of water efflux from the roots of plants
2
1998
... 植物处于深层湿润土壤中的根系吸收的水分可以在夜间蒸腾降低后通过输导组织运输并释放到浅层较干燥的土壤中, 即存在水力提升的现象(
Caldwell et al ., 1998 ).迄今为止, 越来越多的实验证实了水力提升现象的存在, 研究者们观测到很多物种, 包括乔木、灌木、藤本、牧草和农作物等约60种植物存在水分提升作用(
李唯等, 2007 ;
刘小莉等, 2007 ;
郝兴明等, 2009 ;
阿拉木萨和周丽芳, 2011 ;
Brooksbank et al ., 2011 ;
Couvreur et al ., 2014 ;
鱼腾飞等, 2014 ;
Meunier et al ., 2017 ;
李宏侠等, 2018 ). ...
... 深根性植物水力提升作用所释放的水分可以改善水分在土壤中的分布格局, 促进土壤养分分解和矿化过程, 有利于植物对养分的吸收, 促进养分循环(
Caldwell et al ., 1998 ;
何维明和张新时, 2001 ;
李宏侠等, 2018 ).旱区土壤水分是植物生存发展的关键生态因子, 深根植物提升的水分有利于维持相邻浅根植物的生存, 在调节生态系统水分平衡方面有重要的作用.在旱地农业或草地农业生产中, 将具有水力提升功能的深根植物与浅根植物套种, 通过发展粮-粮、草-粮或者草-草等间混套作模式, 可能是充分利用干旱半干旱地区土壤养分和自然降水资源, 提高作物产量的一种有效途径. ...
Dynamic aspects of soil water availability for isohydric plants: focus on root hydraulic resistances
1
2014
... 植物处于深层湿润土壤中的根系吸收的水分可以在夜间蒸腾降低后通过输导组织运输并释放到浅层较干燥的土壤中, 即存在水力提升的现象(
Caldwell et al ., 1998 ).迄今为止, 越来越多的实验证实了水力提升现象的存在, 研究者们观测到很多物种, 包括乔木、灌木、藤本、牧草和农作物等约60种植物存在水分提升作用(
李唯等, 2007 ;
刘小莉等, 2007 ;
郝兴明等, 2009 ;
阿拉木萨和周丽芳, 2011 ;
Brooksbank et al ., 2011 ;
Couvreur et al ., 2014 ;
鱼腾飞等, 2014 ;
Meunier et al ., 2017 ;
李宏侠等, 2018 ). ...
Determining water use by trees and forests from isotopic, energy balance and transpiration analyses: the roles of tree size and hydraulic lift
1
1996
... 已有的水力提升测定方法有很多种, 采用较多的是以下两种方法: 1)测定植物根系周围土壤水分含量或水势的变化, 即通过将植物地上部遮阴或在夜间植物蒸腾速率为0或很低时测定和计算根区土壤含水量或水势的昼夜差值来反映水力提升作用; 2)氢同位素示踪方法, 即通过对植物深层根系施以重同位素标记水(
Pe?uelas & Filella, 2003 )或酸性品红作为示踪剂处理(
何维明和张新时, 2001 ), 再测定浅层土壤水以及植物茎秆水氢同位素组成的变化, 来判明是否发生了水力提升现象并避免了土壤毛管水的影响(
Dawson, 1996 ). ...
Residual effect and nitrate leaching in grass-arable rotations: effect of grassland proportion, sward type and fertilizer history
1
2010
... 由于豆科与禾本科牧草地下生态位存在明显的不同, 当深根性的豆科牧草与浅根性的禾本科植物混播时, 植物根系分布格局的变化有可能导致资源在时间和空间上的协调分配(
吴序卉, 1999 ).研究表明在豆科/禾本科混播系统中存在一种氮共享的通道, 即豆科植物将固定的氮通过各种途径为伴生的禾本科植物提供氮源(
Eriksen et al ., 2010 ).例如, 在建植不久的白三叶(
Trifolium repens )/黑麦草(
Lolium perenne )混播草地中, 黑麦草通过吸收白三叶根系渗出液从而实现氮由豆科牧草向禾草的转移(
Paynel et al ., 2001 ).而水是植物种间营养物质运输的媒介, 禾豆牧草间氮再分配过程必然伴随着水分的共享.
李唯等(2007) 和
刘小莉等(2007) 分别通过上下桶分根试验证明了紫花苜蓿和斜茎黄耆都具有根系水力提升现象, 这就为豆禾牧草混、间作模式提供了可能的理论基础. ...
Carbon isotope discrimination and photosynthesis
2
1989
... C
3 植物的Δ
13 C与植物蒸腾效率及环境水分状况有密切的关系, 较低的气孔导度或较高的光合速率 会降低Δ
13 C (
Farquhar et al ., 1989 ;
Badeck et al ., 2005 ;
孙谷畴等, 2008 ; 朱林等, 2016).对于C
3 植物, Δ
13 C与
C i /
C a 呈正相关关系, 而
C i /
C a 与气孔导度(
G s )正相关而与蒸腾效率(
TE )负相关.水分胁迫会使
G s 下降, 从而导致更低的Δ
13 C值和更高的
TE (
Farquhar & Richards, 1984 ;
Farquhar et al ., 1989 ).在本实验结果中, 斜茎黄耆整株?
13 C比紫花苜蓿的低(
图5 ), 说明斜茎黄耆气孔导度较低, 蒸腾效率较高.这可能与其遗传特性有关,
徐炳成等(2005) 和
山仑等(2008) 研究发现紫花苜蓿对水分条件变化的敏感性高于斜茎黄耆, 斜茎黄耆在低水条件下蒸腾效率高于紫花苜蓿, 本实验中斜茎黄耆整株?
13 C显著低于紫花苜蓿的结果与之相一致.在“上干下湿”的“上下盆”土壤水分格局下, 紫花苜蓿对水分的高度敏感性促使其根系向下盆湿润土层生长, 并增加了其对下盆水分的吸收利用, 这也是紫花苜蓿有更高的整株Δ
13 C值的原因之一. ...
... ;
Farquhar et al ., 1989 ).在本实验结果中, 斜茎黄耆整株?
13 C比紫花苜蓿的低(
图5 ), 说明斜茎黄耆气孔导度较低, 蒸腾效率较高.这可能与其遗传特性有关,
徐炳成等(2005) 和
山仑等(2008) 研究发现紫花苜蓿对水分条件变化的敏感性高于斜茎黄耆, 斜茎黄耆在低水条件下蒸腾效率高于紫花苜蓿, 本实验中斜茎黄耆整株?
13 C显著低于紫花苜蓿的结果与之相一致.在“上干下湿”的“上下盆”土壤水分格局下, 紫花苜蓿对水分的高度敏感性促使其根系向下盆湿润土层生长, 并增加了其对下盆水分的吸收利用, 这也是紫花苜蓿有更高的整株Δ
13 C值的原因之一. ...
Isotopic composition of plant carbon correlates with water-use efficiency of wheat genotypes
3
1984
... (2)碳同位素分辨率的测定: 2015年7月17日采样, 将采集的植物整株样品烘干粉碎并过0.25 mm的筛子, 各个植物样品在Flash 2000 HT元素分析仪(Thermo Scientific, Waltham, USA)中高温燃烧后生成CO
2 , 用Delta V Advantage同位素比率质谱仪(Thermo Scientific, Waltham, USA)检测CO
2 中
13 C与
12 C的比率(测定精度: 碳稳定同位素比率(
δ 13 C) ± <0.1‰), 参照国际标准物(Pee Dee Belnite)比对计算
δ 13 C).各处理的碳同位素分辨率(?
13 C)则依据各自的
δ 13 C值并参照
Farquhar和Richards (1984) 的公式推导得出. ...
...
δ 13 C经计算转换成?
13 C之后再进行数据的分析处理, 二者之间的转换公式如下(
Farquhar & Richards, 1984 ): ...
... C
3 植物的Δ
13 C与植物蒸腾效率及环境水分状况有密切的关系, 较低的气孔导度或较高的光合速率 会降低Δ
13 C (
Farquhar et al ., 1989 ;
Badeck et al ., 2005 ;
孙谷畴等, 2008 ; 朱林等, 2016).对于C
3 植物, Δ
13 C与
C i /
C a 呈正相关关系, 而
C i /
C a 与气孔导度(
G s )正相关而与蒸腾效率(
TE )负相关.水分胁迫会使
G s 下降, 从而导致更低的Δ
13 C值和更高的
TE (
Farquhar & Richards, 1984 ;
Farquhar et al ., 1989 ).在本实验结果中, 斜茎黄耆整株?
13 C比紫花苜蓿的低(
图5 ), 说明斜茎黄耆气孔导度较低, 蒸腾效率较高.这可能与其遗传特性有关,
徐炳成等(2005) 和
山仑等(2008) 研究发现紫花苜蓿对水分条件变化的敏感性高于斜茎黄耆, 斜茎黄耆在低水条件下蒸腾效率高于紫花苜蓿, 本实验中斜茎黄耆整株?
13 C显著低于紫花苜蓿的结果与之相一致.在“上干下湿”的“上下盆”土壤水分格局下, 紫花苜蓿对水分的高度敏感性促使其根系向下盆湿润土层生长, 并增加了其对下盆水分的吸收利用, 这也是紫花苜蓿有更高的整株Δ
13 C值的原因之一. ...
胡杨根系水力提升作用的证据及其生态学意义
1
2009
... 植物处于深层湿润土壤中的根系吸收的水分可以在夜间蒸腾降低后通过输导组织运输并释放到浅层较干燥的土壤中, 即存在水力提升的现象(
Caldwell et al ., 1998 ).迄今为止, 越来越多的实验证实了水力提升现象的存在, 研究者们观测到很多物种, 包括乔木、灌木、藤本、牧草和农作物等约60种植物存在水分提升作用(
李唯等, 2007 ;
刘小莉等, 2007 ;
郝兴明等, 2009 ;
阿拉木萨和周丽芳, 2011 ;
Brooksbank et al ., 2011 ;
Couvreur et al ., 2014 ;
鱼腾飞等, 2014 ;
Meunier et al ., 2017 ;
李宏侠等, 2018 ). ...
胡杨根系水力提升作用的证据及其生态学意义
1
2009
... 植物处于深层湿润土壤中的根系吸收的水分可以在夜间蒸腾降低后通过输导组织运输并释放到浅层较干燥的土壤中, 即存在水力提升的现象(
Caldwell et al ., 1998 ).迄今为止, 越来越多的实验证实了水力提升现象的存在, 研究者们观测到很多物种, 包括乔木、灌木、藤本、牧草和农作物等约60种植物存在水分提升作用(
李唯等, 2007 ;
刘小莉等, 2007 ;
郝兴明等, 2009 ;
阿拉木萨和周丽芳, 2011 ;
Brooksbank et al ., 2011 ;
Couvreur et al ., 2014 ;
鱼腾飞等, 2014 ;
Meunier et al ., 2017 ;
李宏侠等, 2018 ). ...
水分共享在毛乌素沙地4种灌木根系中的存在状况
2
2001
... 深根性植物水力提升作用所释放的水分可以改善水分在土壤中的分布格局, 促进土壤养分分解和矿化过程, 有利于植物对养分的吸收, 促进养分循环(
Caldwell et al ., 1998 ;
何维明和张新时, 2001 ;
李宏侠等, 2018 ).旱区土壤水分是植物生存发展的关键生态因子, 深根植物提升的水分有利于维持相邻浅根植物的生存, 在调节生态系统水分平衡方面有重要的作用.在旱地农业或草地农业生产中, 将具有水力提升功能的深根植物与浅根植物套种, 通过发展粮-粮、草-粮或者草-草等间混套作模式, 可能是充分利用干旱半干旱地区土壤养分和自然降水资源, 提高作物产量的一种有效途径. ...
... 已有的水力提升测定方法有很多种, 采用较多的是以下两种方法: 1)测定植物根系周围土壤水分含量或水势的变化, 即通过将植物地上部遮阴或在夜间植物蒸腾速率为0或很低时测定和计算根区土壤含水量或水势的昼夜差值来反映水力提升作用; 2)氢同位素示踪方法, 即通过对植物深层根系施以重同位素标记水(
Pe?uelas & Filella, 2003 )或酸性品红作为示踪剂处理(
何维明和张新时, 2001 ), 再测定浅层土壤水以及植物茎秆水氢同位素组成的变化, 来判明是否发生了水力提升现象并避免了土壤毛管水的影响(
Dawson, 1996 ). ...
水分共享在毛乌素沙地4种灌木根系中的存在状况
2
2001
... 深根性植物水力提升作用所释放的水分可以改善水分在土壤中的分布格局, 促进土壤养分分解和矿化过程, 有利于植物对养分的吸收, 促进养分循环(
Caldwell et al ., 1998 ;
何维明和张新时, 2001 ;
李宏侠等, 2018 ).旱区土壤水分是植物生存发展的关键生态因子, 深根植物提升的水分有利于维持相邻浅根植物的生存, 在调节生态系统水分平衡方面有重要的作用.在旱地农业或草地农业生产中, 将具有水力提升功能的深根植物与浅根植物套种, 通过发展粮-粮、草-粮或者草-草等间混套作模式, 可能是充分利用干旱半干旱地区土壤养分和自然降水资源, 提高作物产量的一种有效途径. ...
... 已有的水力提升测定方法有很多种, 采用较多的是以下两种方法: 1)测定植物根系周围土壤水分含量或水势的变化, 即通过将植物地上部遮阴或在夜间植物蒸腾速率为0或很低时测定和计算根区土壤含水量或水势的昼夜差值来反映水力提升作用; 2)氢同位素示踪方法, 即通过对植物深层根系施以重同位素标记水(
Pe?uelas & Filella, 2003 )或酸性品红作为示踪剂处理(
何维明和张新时, 2001 ), 再测定浅层土壤水以及植物茎秆水氢同位素组成的变化, 来判明是否发生了水力提升现象并避免了土壤毛管水的影响(
Dawson, 1996 ). ...
A split-root apparatus for examining the effects of hydraulic lift by trees on the water status of neighboring crops
1
2004
... 即时域反射技术(TDR)具有数据采集准确及时、相对误差小、对土壤扰动小、操作简单、结果可靠等优势, 目前在测定土壤含水量方面得到普遍应用.近年来, 许多研究者采用分根装置与TDR水分测定系统相结合研究了不同植物的根系提水作用(
Wan et al ., 2000 ;
Hirota et al ., 2004 ;
薛小红等, 2007 ;
阿拉木萨和周丽芳, 2011 ).此后, TDR与分根装置的结合成为研究水分提升作用的一种新方法, 这种方法操作简便, 结果可靠, 但关键在于分根装置设计时要注意消除土壤毛管上升水对实验结果的影响(
薛小红等, 2007 ).根据已发表的上下桶实验结果, 上层干燥土壤含水量呈现出有规律的昼夜变化趋势, 即在下午土壤含水量急剧下降, 而在午夜至凌晨时段土壤含水量最高(
李唯等, 2007 ;
薛小红等, 2007 ;
阿拉木萨和周丽芳, 2011 ).我们在制作“上下盆”实验装置时, 在豆科牧草根系长入下盆后, 将上盆用小木块垫起, 使上下盆分离, 并用水把上下盆连接处的土壤冲掉, 只留根系与上盆相连, 从而防止了下盆土壤毛细管作用对上盆水分的补给.同时, 在实验过程中对上盆做了遮雨处理, 因而上盆土壤水分的变化只受植物因素的影响.在我们的实验结果中, 斜茎黄耆土壤含水量昼夜差值与
薛小红等(2007) 报道的最大土壤含水量昼夜差值相近, 两种豆科牧草上盆土壤含水量昼夜差值及提水量存在显著差异, 斜茎黄耆无论在单播还是混播时上盆土壤含水量昼夜差值及日平均提水量均显著高于紫花苜蓿(
表2 ), 说明斜茎黄耆的水力提升效果好于紫花苜蓿. ...
梭梭水力提升与浅根系植物优势度、丰富度和多度的关系
2
2018
... 植物处于深层湿润土壤中的根系吸收的水分可以在夜间蒸腾降低后通过输导组织运输并释放到浅层较干燥的土壤中, 即存在水力提升的现象(
Caldwell et al ., 1998 ).迄今为止, 越来越多的实验证实了水力提升现象的存在, 研究者们观测到很多物种, 包括乔木、灌木、藤本、牧草和农作物等约60种植物存在水分提升作用(
李唯等, 2007 ;
刘小莉等, 2007 ;
郝兴明等, 2009 ;
阿拉木萨和周丽芳, 2011 ;
Brooksbank et al ., 2011 ;
Couvreur et al ., 2014 ;
鱼腾飞等, 2014 ;
Meunier et al ., 2017 ;
李宏侠等, 2018 ). ...
... 深根性植物水力提升作用所释放的水分可以改善水分在土壤中的分布格局, 促进土壤养分分解和矿化过程, 有利于植物对养分的吸收, 促进养分循环(
Caldwell et al ., 1998 ;
何维明和张新时, 2001 ;
李宏侠等, 2018 ).旱区土壤水分是植物生存发展的关键生态因子, 深根植物提升的水分有利于维持相邻浅根植物的生存, 在调节生态系统水分平衡方面有重要的作用.在旱地农业或草地农业生产中, 将具有水力提升功能的深根植物与浅根植物套种, 通过发展粮-粮、草-粮或者草-草等间混套作模式, 可能是充分利用干旱半干旱地区土壤养分和自然降水资源, 提高作物产量的一种有效途径. ...
梭梭水力提升与浅根系植物优势度、丰富度和多度的关系
2
2018
... 植物处于深层湿润土壤中的根系吸收的水分可以在夜间蒸腾降低后通过输导组织运输并释放到浅层较干燥的土壤中, 即存在水力提升的现象(
Caldwell et al ., 1998 ).迄今为止, 越来越多的实验证实了水力提升现象的存在, 研究者们观测到很多物种, 包括乔木、灌木、藤本、牧草和农作物等约60种植物存在水分提升作用(
李唯等, 2007 ;
刘小莉等, 2007 ;
郝兴明等, 2009 ;
阿拉木萨和周丽芳, 2011 ;
Brooksbank et al ., 2011 ;
Couvreur et al ., 2014 ;
鱼腾飞等, 2014 ;
Meunier et al ., 2017 ;
李宏侠等, 2018 ). ...
... 深根性植物水力提升作用所释放的水分可以改善水分在土壤中的分布格局, 促进土壤养分分解和矿化过程, 有利于植物对养分的吸收, 促进养分循环(
Caldwell et al ., 1998 ;
何维明和张新时, 2001 ;
李宏侠等, 2018 ).旱区土壤水分是植物生存发展的关键生态因子, 深根植物提升的水分有利于维持相邻浅根植物的生存, 在调节生态系统水分平衡方面有重要的作用.在旱地农业或草地农业生产中, 将具有水力提升功能的深根植物与浅根植物套种, 通过发展粮-粮、草-粮或者草-草等间混套作模式, 可能是充分利用干旱半干旱地区土壤养分和自然降水资源, 提高作物产量的一种有效途径. ...
苜蓿、玉米根系提水作用与耐旱性的关系——植物根系提水作用机理研究II
3
2007
... 植物处于深层湿润土壤中的根系吸收的水分可以在夜间蒸腾降低后通过输导组织运输并释放到浅层较干燥的土壤中, 即存在水力提升的现象(
Caldwell et al ., 1998 ).迄今为止, 越来越多的实验证实了水力提升现象的存在, 研究者们观测到很多物种, 包括乔木、灌木、藤本、牧草和农作物等约60种植物存在水分提升作用(
李唯等, 2007 ;
刘小莉等, 2007 ;
郝兴明等, 2009 ;
阿拉木萨和周丽芳, 2011 ;
Brooksbank et al ., 2011 ;
Couvreur et al ., 2014 ;
鱼腾飞等, 2014 ;
Meunier et al ., 2017 ;
李宏侠等, 2018 ). ...
... 由于豆科与禾本科牧草地下生态位存在明显的不同, 当深根性的豆科牧草与浅根性的禾本科植物混播时, 植物根系分布格局的变化有可能导致资源在时间和空间上的协调分配(
吴序卉, 1999 ).研究表明在豆科/禾本科混播系统中存在一种氮共享的通道, 即豆科植物将固定的氮通过各种途径为伴生的禾本科植物提供氮源(
Eriksen et al ., 2010 ).例如, 在建植不久的白三叶(
Trifolium repens )/黑麦草(
Lolium perenne )混播草地中, 黑麦草通过吸收白三叶根系渗出液从而实现氮由豆科牧草向禾草的转移(
Paynel et al ., 2001 ).而水是植物种间营养物质运输的媒介, 禾豆牧草间氮再分配过程必然伴随着水分的共享.
李唯等(2007) 和
刘小莉等(2007) 分别通过上下桶分根试验证明了紫花苜蓿和斜茎黄耆都具有根系水力提升现象, 这就为豆禾牧草混、间作模式提供了可能的理论基础. ...
... 即时域反射技术(TDR)具有数据采集准确及时、相对误差小、对土壤扰动小、操作简单、结果可靠等优势, 目前在测定土壤含水量方面得到普遍应用.近年来, 许多研究者采用分根装置与TDR水分测定系统相结合研究了不同植物的根系提水作用(
Wan et al ., 2000 ;
Hirota et al ., 2004 ;
薛小红等, 2007 ;
阿拉木萨和周丽芳, 2011 ).此后, TDR与分根装置的结合成为研究水分提升作用的一种新方法, 这种方法操作简便, 结果可靠, 但关键在于分根装置设计时要注意消除土壤毛管上升水对实验结果的影响(
薛小红等, 2007 ).根据已发表的上下桶实验结果, 上层干燥土壤含水量呈现出有规律的昼夜变化趋势, 即在下午土壤含水量急剧下降, 而在午夜至凌晨时段土壤含水量最高(
李唯等, 2007 ;
薛小红等, 2007 ;
阿拉木萨和周丽芳, 2011 ).我们在制作“上下盆”实验装置时, 在豆科牧草根系长入下盆后, 将上盆用小木块垫起, 使上下盆分离, 并用水把上下盆连接处的土壤冲掉, 只留根系与上盆相连, 从而防止了下盆土壤毛细管作用对上盆水分的补给.同时, 在实验过程中对上盆做了遮雨处理, 因而上盆土壤水分的变化只受植物因素的影响.在我们的实验结果中, 斜茎黄耆土壤含水量昼夜差值与
薛小红等(2007) 报道的最大土壤含水量昼夜差值相近, 两种豆科牧草上盆土壤含水量昼夜差值及提水量存在显著差异, 斜茎黄耆无论在单播还是混播时上盆土壤含水量昼夜差值及日平均提水量均显著高于紫花苜蓿(
表2 ), 说明斜茎黄耆的水力提升效果好于紫花苜蓿. ...
苜蓿、玉米根系提水作用与耐旱性的关系——植物根系提水作用机理研究II
3
2007
... 植物处于深层湿润土壤中的根系吸收的水分可以在夜间蒸腾降低后通过输导组织运输并释放到浅层较干燥的土壤中, 即存在水力提升的现象(
Caldwell et al ., 1998 ).迄今为止, 越来越多的实验证实了水力提升现象的存在, 研究者们观测到很多物种, 包括乔木、灌木、藤本、牧草和农作物等约60种植物存在水分提升作用(
李唯等, 2007 ;
刘小莉等, 2007 ;
郝兴明等, 2009 ;
阿拉木萨和周丽芳, 2011 ;
Brooksbank et al ., 2011 ;
Couvreur et al ., 2014 ;
鱼腾飞等, 2014 ;
Meunier et al ., 2017 ;
李宏侠等, 2018 ). ...
... 由于豆科与禾本科牧草地下生态位存在明显的不同, 当深根性的豆科牧草与浅根性的禾本科植物混播时, 植物根系分布格局的变化有可能导致资源在时间和空间上的协调分配(
吴序卉, 1999 ).研究表明在豆科/禾本科混播系统中存在一种氮共享的通道, 即豆科植物将固定的氮通过各种途径为伴生的禾本科植物提供氮源(
Eriksen et al ., 2010 ).例如, 在建植不久的白三叶(
Trifolium repens )/黑麦草(
Lolium perenne )混播草地中, 黑麦草通过吸收白三叶根系渗出液从而实现氮由豆科牧草向禾草的转移(
Paynel et al ., 2001 ).而水是植物种间营养物质运输的媒介, 禾豆牧草间氮再分配过程必然伴随着水分的共享.
李唯等(2007) 和
刘小莉等(2007) 分别通过上下桶分根试验证明了紫花苜蓿和斜茎黄耆都具有根系水力提升现象, 这就为豆禾牧草混、间作模式提供了可能的理论基础. ...
... 即时域反射技术(TDR)具有数据采集准确及时、相对误差小、对土壤扰动小、操作简单、结果可靠等优势, 目前在测定土壤含水量方面得到普遍应用.近年来, 许多研究者采用分根装置与TDR水分测定系统相结合研究了不同植物的根系提水作用(
Wan et al ., 2000 ;
Hirota et al ., 2004 ;
薛小红等, 2007 ;
阿拉木萨和周丽芳, 2011 ).此后, TDR与分根装置的结合成为研究水分提升作用的一种新方法, 这种方法操作简便, 结果可靠, 但关键在于分根装置设计时要注意消除土壤毛管上升水对实验结果的影响(
薛小红等, 2007 ).根据已发表的上下桶实验结果, 上层干燥土壤含水量呈现出有规律的昼夜变化趋势, 即在下午土壤含水量急剧下降, 而在午夜至凌晨时段土壤含水量最高(
李唯等, 2007 ;
薛小红等, 2007 ;
阿拉木萨和周丽芳, 2011 ).我们在制作“上下盆”实验装置时, 在豆科牧草根系长入下盆后, 将上盆用小木块垫起, 使上下盆分离, 并用水把上下盆连接处的土壤冲掉, 只留根系与上盆相连, 从而防止了下盆土壤毛细管作用对上盆水分的补给.同时, 在实验过程中对上盆做了遮雨处理, 因而上盆土壤水分的变化只受植物因素的影响.在我们的实验结果中, 斜茎黄耆土壤含水量昼夜差值与
薛小红等(2007) 报道的最大土壤含水量昼夜差值相近, 两种豆科牧草上盆土壤含水量昼夜差值及提水量存在显著差异, 斜茎黄耆无论在单播还是混播时上盆土壤含水量昼夜差值及日平均提水量均显著高于紫花苜蓿(
表2 ), 说明斜茎黄耆的水力提升效果好于紫花苜蓿. ...
施肥对沙打旺根系提水及土壤养分活性的影响
2
2007
... 植物处于深层湿润土壤中的根系吸收的水分可以在夜间蒸腾降低后通过输导组织运输并释放到浅层较干燥的土壤中, 即存在水力提升的现象(
Caldwell et al ., 1998 ).迄今为止, 越来越多的实验证实了水力提升现象的存在, 研究者们观测到很多物种, 包括乔木、灌木、藤本、牧草和农作物等约60种植物存在水分提升作用(
李唯等, 2007 ;
刘小莉等, 2007 ;
郝兴明等, 2009 ;
阿拉木萨和周丽芳, 2011 ;
Brooksbank et al ., 2011 ;
Couvreur et al ., 2014 ;
鱼腾飞等, 2014 ;
Meunier et al ., 2017 ;
李宏侠等, 2018 ). ...
... 由于豆科与禾本科牧草地下生态位存在明显的不同, 当深根性的豆科牧草与浅根性的禾本科植物混播时, 植物根系分布格局的变化有可能导致资源在时间和空间上的协调分配(
吴序卉, 1999 ).研究表明在豆科/禾本科混播系统中存在一种氮共享的通道, 即豆科植物将固定的氮通过各种途径为伴生的禾本科植物提供氮源(
Eriksen et al ., 2010 ).例如, 在建植不久的白三叶(
Trifolium repens )/黑麦草(
Lolium perenne )混播草地中, 黑麦草通过吸收白三叶根系渗出液从而实现氮由豆科牧草向禾草的转移(
Paynel et al ., 2001 ).而水是植物种间营养物质运输的媒介, 禾豆牧草间氮再分配过程必然伴随着水分的共享.
李唯等(2007) 和
刘小莉等(2007) 分别通过上下桶分根试验证明了紫花苜蓿和斜茎黄耆都具有根系水力提升现象, 这就为豆禾牧草混、间作模式提供了可能的理论基础. ...
施肥对沙打旺根系提水及土壤养分活性的影响
2
2007
... 植物处于深层湿润土壤中的根系吸收的水分可以在夜间蒸腾降低后通过输导组织运输并释放到浅层较干燥的土壤中, 即存在水力提升的现象(
Caldwell et al ., 1998 ).迄今为止, 越来越多的实验证实了水力提升现象的存在, 研究者们观测到很多物种, 包括乔木、灌木、藤本、牧草和农作物等约60种植物存在水分提升作用(
李唯等, 2007 ;
刘小莉等, 2007 ;
郝兴明等, 2009 ;
阿拉木萨和周丽芳, 2011 ;
Brooksbank et al ., 2011 ;
Couvreur et al ., 2014 ;
鱼腾飞等, 2014 ;
Meunier et al ., 2017 ;
李宏侠等, 2018 ). ...
... 由于豆科与禾本科牧草地下生态位存在明显的不同, 当深根性的豆科牧草与浅根性的禾本科植物混播时, 植物根系分布格局的变化有可能导致资源在时间和空间上的协调分配(
吴序卉, 1999 ).研究表明在豆科/禾本科混播系统中存在一种氮共享的通道, 即豆科植物将固定的氮通过各种途径为伴生的禾本科植物提供氮源(
Eriksen et al ., 2010 ).例如, 在建植不久的白三叶(
Trifolium repens )/黑麦草(
Lolium perenne )混播草地中, 黑麦草通过吸收白三叶根系渗出液从而实现氮由豆科牧草向禾草的转移(
Paynel et al ., 2001 ).而水是植物种间营养物质运输的媒介, 禾豆牧草间氮再分配过程必然伴随着水分的共享.
李唯等(2007) 和
刘小莉等(2007) 分别通过上下桶分根试验证明了紫花苜蓿和斜茎黄耆都具有根系水力提升现象, 这就为豆禾牧草混、间作模式提供了可能的理论基础. ...
Measuring and modeling hydraulic lift of
Lolium multiflorum using stable water isotopes
1
2017
... 植物处于深层湿润土壤中的根系吸收的水分可以在夜间蒸腾降低后通过输导组织运输并释放到浅层较干燥的土壤中, 即存在水力提升的现象(
Caldwell et al ., 1998 ).迄今为止, 越来越多的实验证实了水力提升现象的存在, 研究者们观测到很多物种, 包括乔木、灌木、藤本、牧草和农作物等约60种植物存在水分提升作用(
李唯等, 2007 ;
刘小莉等, 2007 ;
郝兴明等, 2009 ;
阿拉木萨和周丽芳, 2011 ;
Brooksbank et al ., 2011 ;
Couvreur et al ., 2014 ;
鱼腾飞等, 2014 ;
Meunier et al ., 2017 ;
李宏侠等, 2018 ). ...
Root exudates: a pathway for short-term N transfer from clover to ryegrass
1
2001
... 由于豆科与禾本科牧草地下生态位存在明显的不同, 当深根性的豆科牧草与浅根性的禾本科植物混播时, 植物根系分布格局的变化有可能导致资源在时间和空间上的协调分配(
吴序卉, 1999 ).研究表明在豆科/禾本科混播系统中存在一种氮共享的通道, 即豆科植物将固定的氮通过各种途径为伴生的禾本科植物提供氮源(
Eriksen et al ., 2010 ).例如, 在建植不久的白三叶(
Trifolium repens )/黑麦草(
Lolium perenne )混播草地中, 黑麦草通过吸收白三叶根系渗出液从而实现氮由豆科牧草向禾草的转移(
Paynel et al ., 2001 ).而水是植物种间营养物质运输的媒介, 禾豆牧草间氮再分配过程必然伴随着水分的共享.
李唯等(2007) 和
刘小莉等(2007) 分别通过上下桶分根试验证明了紫花苜蓿和斜茎黄耆都具有根系水力提升现象, 这就为豆禾牧草混、间作模式提供了可能的理论基础. ...
Deuterium labeling of roots provides evidence of deep water access and hydraulic lift by
Pinus nigra, in a Mediterranean forest of NE Spain
1
2003
... 已有的水力提升测定方法有很多种, 采用较多的是以下两种方法: 1)测定植物根系周围土壤水分含量或水势的变化, 即通过将植物地上部遮阴或在夜间植物蒸腾速率为0或很低时测定和计算根区土壤含水量或水势的昼夜差值来反映水力提升作用; 2)氢同位素示踪方法, 即通过对植物深层根系施以重同位素标记水(
Pe?uelas & Filella, 2003 )或酸性品红作为示踪剂处理(
何维明和张新时, 2001 ), 再测定浅层土壤水以及植物茎秆水氢同位素组成的变化, 来判明是否发生了水力提升现象并避免了土壤毛管水的影响(
Dawson, 1996 ). ...
论苜蓿的生产力与抗旱性
2
2008
... 该实验供试的牧草有3个, 其中, 豆科作物采用紫花苜蓿和斜茎黄耆, 紫花苜蓿及斜茎黄耆均为优良的豆科牧草, 对于促进畜牧业发展、生态环境建设和保护等方面具有重要的意义.这两种豆科牧草的根系都十分发达, 对深层土壤水分的利用能力很强(
山仑等, 2008 ;
杨永东等, 2008 ).本实验采用的紫花苜蓿品种是‘美国牧歌401’, 在实验中的编号为A, 斜茎黄耆为当地栽培种, 在实验中的编号为B.混播的禾本科作物是‘冬牧70’黑麦, 在实验中的编号为C. ...
... C
3 植物的Δ
13 C与植物蒸腾效率及环境水分状况有密切的关系, 较低的气孔导度或较高的光合速率 会降低Δ
13 C (
Farquhar et al ., 1989 ;
Badeck et al ., 2005 ;
孙谷畴等, 2008 ; 朱林等, 2016).对于C
3 植物, Δ
13 C与
C i /
C a 呈正相关关系, 而
C i /
C a 与气孔导度(
G s )正相关而与蒸腾效率(
TE )负相关.水分胁迫会使
G s 下降, 从而导致更低的Δ
13 C值和更高的
TE (
Farquhar & Richards, 1984 ;
Farquhar et al ., 1989 ).在本实验结果中, 斜茎黄耆整株?
13 C比紫花苜蓿的低(
图5 ), 说明斜茎黄耆气孔导度较低, 蒸腾效率较高.这可能与其遗传特性有关,
徐炳成等(2005) 和
山仑等(2008) 研究发现紫花苜蓿对水分条件变化的敏感性高于斜茎黄耆, 斜茎黄耆在低水条件下蒸腾效率高于紫花苜蓿, 本实验中斜茎黄耆整株?
13 C显著低于紫花苜蓿的结果与之相一致.在“上干下湿”的“上下盆”土壤水分格局下, 紫花苜蓿对水分的高度敏感性促使其根系向下盆湿润土层生长, 并增加了其对下盆水分的吸收利用, 这也是紫花苜蓿有更高的整株Δ
13 C值的原因之一. ...
论苜蓿的生产力与抗旱性
2
2008
... 该实验供试的牧草有3个, 其中, 豆科作物采用紫花苜蓿和斜茎黄耆, 紫花苜蓿及斜茎黄耆均为优良的豆科牧草, 对于促进畜牧业发展、生态环境建设和保护等方面具有重要的意义.这两种豆科牧草的根系都十分发达, 对深层土壤水分的利用能力很强(
山仑等, 2008 ;
杨永东等, 2008 ).本实验采用的紫花苜蓿品种是‘美国牧歌401’, 在实验中的编号为A, 斜茎黄耆为当地栽培种, 在实验中的编号为B.混播的禾本科作物是‘冬牧70’黑麦, 在实验中的编号为C. ...
... C
3 植物的Δ
13 C与植物蒸腾效率及环境水分状况有密切的关系, 较低的气孔导度或较高的光合速率 会降低Δ
13 C (
Farquhar et al ., 1989 ;
Badeck et al ., 2005 ;
孙谷畴等, 2008 ; 朱林等, 2016).对于C
3 植物, Δ
13 C与
C i /
C a 呈正相关关系, 而
C i /
C a 与气孔导度(
G s )正相关而与蒸腾效率(
TE )负相关.水分胁迫会使
G s 下降, 从而导致更低的Δ
13 C值和更高的
TE (
Farquhar & Richards, 1984 ;
Farquhar et al ., 1989 ).在本实验结果中, 斜茎黄耆整株?
13 C比紫花苜蓿的低(
图5 ), 说明斜茎黄耆气孔导度较低, 蒸腾效率较高.这可能与其遗传特性有关,
徐炳成等(2005) 和
山仑等(2008) 研究发现紫花苜蓿对水分条件变化的敏感性高于斜茎黄耆, 斜茎黄耆在低水条件下蒸腾效率高于紫花苜蓿, 本实验中斜茎黄耆整株?
13 C显著低于紫花苜蓿的结果与之相一致.在“上干下湿”的“上下盆”土壤水分格局下, 紫花苜蓿对水分的高度敏感性促使其根系向下盆湿润土层生长, 并增加了其对下盆水分的吸收利用, 这也是紫花苜蓿有更高的整株Δ
13 C值的原因之一. ...
马占相思叶片液汁碳同位素甄别率和水分利用效率
1
2008
... C
3 植物的Δ
13 C与植物蒸腾效率及环境水分状况有密切的关系, 较低的气孔导度或较高的光合速率 会降低Δ
13 C (
Farquhar et al ., 1989 ;
Badeck et al ., 2005 ;
孙谷畴等, 2008 ; 朱林等, 2016).对于C
3 植物, Δ
13 C与
C i /
C a 呈正相关关系, 而
C i /
C a 与气孔导度(
G s )正相关而与蒸腾效率(
TE )负相关.水分胁迫会使
G s 下降, 从而导致更低的Δ
13 C值和更高的
TE (
Farquhar & Richards, 1984 ;
Farquhar et al ., 1989 ).在本实验结果中, 斜茎黄耆整株?
13 C比紫花苜蓿的低(
图5 ), 说明斜茎黄耆气孔导度较低, 蒸腾效率较高.这可能与其遗传特性有关,
徐炳成等(2005) 和
山仑等(2008) 研究发现紫花苜蓿对水分条件变化的敏感性高于斜茎黄耆, 斜茎黄耆在低水条件下蒸腾效率高于紫花苜蓿, 本实验中斜茎黄耆整株?
13 C显著低于紫花苜蓿的结果与之相一致.在“上干下湿”的“上下盆”土壤水分格局下, 紫花苜蓿对水分的高度敏感性促使其根系向下盆湿润土层生长, 并增加了其对下盆水分的吸收利用, 这也是紫花苜蓿有更高的整株Δ
13 C值的原因之一. ...
马占相思叶片液汁碳同位素甄别率和水分利用效率
1
2008
... C
3 植物的Δ
13 C与植物蒸腾效率及环境水分状况有密切的关系, 较低的气孔导度或较高的光合速率 会降低Δ
13 C (
Farquhar et al ., 1989 ;
Badeck et al ., 2005 ;
孙谷畴等, 2008 ; 朱林等, 2016).对于C
3 植物, Δ
13 C与
C i /
C a 呈正相关关系, 而
C i /
C a 与气孔导度(
G s )正相关而与蒸腾效率(
TE )负相关.水分胁迫会使
G s 下降, 从而导致更低的Δ
13 C值和更高的
TE (
Farquhar & Richards, 1984 ;
Farquhar et al ., 1989 ).在本实验结果中, 斜茎黄耆整株?
13 C比紫花苜蓿的低(
图5 ), 说明斜茎黄耆气孔导度较低, 蒸腾效率较高.这可能与其遗传特性有关,
徐炳成等(2005) 和
山仑等(2008) 研究发现紫花苜蓿对水分条件变化的敏感性高于斜茎黄耆, 斜茎黄耆在低水条件下蒸腾效率高于紫花苜蓿, 本实验中斜茎黄耆整株?
13 C显著低于紫花苜蓿的结果与之相一致.在“上干下湿”的“上下盆”土壤水分格局下, 紫花苜蓿对水分的高度敏感性促使其根系向下盆湿润土层生长, 并增加了其对下盆水分的吸收利用, 这也是紫花苜蓿有更高的整株Δ
13 C值的原因之一. ...
Hydraulic lift in drought-tolerant and susceptible maize hybrids
1
2000
... 即时域反射技术(TDR)具有数据采集准确及时、相对误差小、对土壤扰动小、操作简单、结果可靠等优势, 目前在测定土壤含水量方面得到普遍应用.近年来, 许多研究者采用分根装置与TDR水分测定系统相结合研究了不同植物的根系提水作用(
Wan et al ., 2000 ;
Hirota et al ., 2004 ;
薛小红等, 2007 ;
阿拉木萨和周丽芳, 2011 ).此后, TDR与分根装置的结合成为研究水分提升作用的一种新方法, 这种方法操作简便, 结果可靠, 但关键在于分根装置设计时要注意消除土壤毛管上升水对实验结果的影响(
薛小红等, 2007 ).根据已发表的上下桶实验结果, 上层干燥土壤含水量呈现出有规律的昼夜变化趋势, 即在下午土壤含水量急剧下降, 而在午夜至凌晨时段土壤含水量最高(
李唯等, 2007 ;
薛小红等, 2007 ;
阿拉木萨和周丽芳, 2011 ).我们在制作“上下盆”实验装置时, 在豆科牧草根系长入下盆后, 将上盆用小木块垫起, 使上下盆分离, 并用水把上下盆连接处的土壤冲掉, 只留根系与上盆相连, 从而防止了下盆土壤毛细管作用对上盆水分的补给.同时, 在实验过程中对上盆做了遮雨处理, 因而上盆土壤水分的变化只受植物因素的影响.在我们的实验结果中, 斜茎黄耆土壤含水量昼夜差值与
薛小红等(2007) 报道的最大土壤含水量昼夜差值相近, 两种豆科牧草上盆土壤含水量昼夜差值及提水量存在显著差异, 斜茎黄耆无论在单播还是混播时上盆土壤含水量昼夜差值及日平均提水量均显著高于紫花苜蓿(
表2 ), 说明斜茎黄耆的水力提升效果好于紫花苜蓿. ...
豆科与禾本科牧草混播效应研究进展
1
2007
... 豆科与禾本科牧草混播可以有效地提高单位面积草地饲草产量、改善饲用品质、提高土壤肥力, 在世界上许多地区备受重视(
王旭等, 2007 ).研究表明, 紫花苜蓿(
Medicago sativa )与无芒雀麦(
Bromus inermis )混播后增产效果显著, 但混播组合中紫花苜蓿竞争优势明显, 要降低其比例才能使混播组分稳定(
张永亮等, 2019 ).在宁夏中部半干旱带有灌溉条件下斜茎黄耆(沙打旺,
Astragalus laxmannii )与禾本科牧草混播草地中不同物种生长较为均衡, 混播草地持续性好, 并且有较好的产量效应(
朱林等, 2014 ). ...
豆科与禾本科牧草混播效应研究进展
1
2007
... 豆科与禾本科牧草混播可以有效地提高单位面积草地饲草产量、改善饲用品质、提高土壤肥力, 在世界上许多地区备受重视(
王旭等, 2007 ).研究表明, 紫花苜蓿(
Medicago sativa )与无芒雀麦(
Bromus inermis )混播后增产效果显著, 但混播组合中紫花苜蓿竞争优势明显, 要降低其比例才能使混播组分稳定(
张永亮等, 2019 ).在宁夏中部半干旱带有灌溉条件下斜茎黄耆(沙打旺,
Astragalus laxmannii )与禾本科牧草混播草地中不同物种生长较为均衡, 混播草地持续性好, 并且有较好的产量效应(
朱林等, 2014 ). ...
地膜覆盖下豆禾混播草地根系的动态研究
1
1999
... 由于豆科与禾本科牧草地下生态位存在明显的不同, 当深根性的豆科牧草与浅根性的禾本科植物混播时, 植物根系分布格局的变化有可能导致资源在时间和空间上的协调分配(
吴序卉, 1999 ).研究表明在豆科/禾本科混播系统中存在一种氮共享的通道, 即豆科植物将固定的氮通过各种途径为伴生的禾本科植物提供氮源(
Eriksen et al ., 2010 ).例如, 在建植不久的白三叶(
Trifolium repens )/黑麦草(
Lolium perenne )混播草地中, 黑麦草通过吸收白三叶根系渗出液从而实现氮由豆科牧草向禾草的转移(
Paynel et al ., 2001 ).而水是植物种间营养物质运输的媒介, 禾豆牧草间氮再分配过程必然伴随着水分的共享.
李唯等(2007) 和
刘小莉等(2007) 分别通过上下桶分根试验证明了紫花苜蓿和斜茎黄耆都具有根系水力提升现象, 这就为豆禾牧草混、间作模式提供了可能的理论基础. ...
地膜覆盖下豆禾混播草地根系的动态研究
1
1999
... 由于豆科与禾本科牧草地下生态位存在明显的不同, 当深根性的豆科牧草与浅根性的禾本科植物混播时, 植物根系分布格局的变化有可能导致资源在时间和空间上的协调分配(
吴序卉, 1999 ).研究表明在豆科/禾本科混播系统中存在一种氮共享的通道, 即豆科植物将固定的氮通过各种途径为伴生的禾本科植物提供氮源(
Eriksen et al ., 2010 ).例如, 在建植不久的白三叶(
Trifolium repens )/黑麦草(
Lolium perenne )混播草地中, 黑麦草通过吸收白三叶根系渗出液从而实现氮由豆科牧草向禾草的转移(
Paynel et al ., 2001 ).而水是植物种间营养物质运输的媒介, 禾豆牧草间氮再分配过程必然伴随着水分的共享.
李唯等(2007) 和
刘小莉等(2007) 分别通过上下桶分根试验证明了紫花苜蓿和斜茎黄耆都具有根系水力提升现象, 这就为豆禾牧草混、间作模式提供了可能的理论基础. ...
苜蓿与沙打旺苗期生长和水分利用对土壤水分变化的反应
1
2005
... C
3 植物的Δ
13 C与植物蒸腾效率及环境水分状况有密切的关系, 较低的气孔导度或较高的光合速率 会降低Δ
13 C (
Farquhar et al ., 1989 ;
Badeck et al ., 2005 ;
孙谷畴等, 2008 ; 朱林等, 2016).对于C
3 植物, Δ
13 C与
C i /
C a 呈正相关关系, 而
C i /
C a 与气孔导度(
G s )正相关而与蒸腾效率(
TE )负相关.水分胁迫会使
G s 下降, 从而导致更低的Δ
13 C值和更高的
TE (
Farquhar & Richards, 1984 ;
Farquhar et al ., 1989 ).在本实验结果中, 斜茎黄耆整株?
13 C比紫花苜蓿的低(
图5 ), 说明斜茎黄耆气孔导度较低, 蒸腾效率较高.这可能与其遗传特性有关,
徐炳成等(2005) 和
山仑等(2008) 研究发现紫花苜蓿对水分条件变化的敏感性高于斜茎黄耆, 斜茎黄耆在低水条件下蒸腾效率高于紫花苜蓿, 本实验中斜茎黄耆整株?
13 C显著低于紫花苜蓿的结果与之相一致.在“上干下湿”的“上下盆”土壤水分格局下, 紫花苜蓿对水分的高度敏感性促使其根系向下盆湿润土层生长, 并增加了其对下盆水分的吸收利用, 这也是紫花苜蓿有更高的整株Δ
13 C值的原因之一. ...
苜蓿与沙打旺苗期生长和水分利用对土壤水分变化的反应
1
2005
... C
3 植物的Δ
13 C与植物蒸腾效率及环境水分状况有密切的关系, 较低的气孔导度或较高的光合速率 会降低Δ
13 C (
Farquhar et al ., 1989 ;
Badeck et al ., 2005 ;
孙谷畴等, 2008 ; 朱林等, 2016).对于C
3 植物, Δ
13 C与
C i /
C a 呈正相关关系, 而
C i /
C a 与气孔导度(
G s )正相关而与蒸腾效率(
TE )负相关.水分胁迫会使
G s 下降, 从而导致更低的Δ
13 C值和更高的
TE (
Farquhar & Richards, 1984 ;
Farquhar et al ., 1989 ).在本实验结果中, 斜茎黄耆整株?
13 C比紫花苜蓿的低(
图5 ), 说明斜茎黄耆气孔导度较低, 蒸腾效率较高.这可能与其遗传特性有关,
徐炳成等(2005) 和
山仑等(2008) 研究发现紫花苜蓿对水分条件变化的敏感性高于斜茎黄耆, 斜茎黄耆在低水条件下蒸腾效率高于紫花苜蓿, 本实验中斜茎黄耆整株?
13 C显著低于紫花苜蓿的结果与之相一致.在“上干下湿”的“上下盆”土壤水分格局下, 紫花苜蓿对水分的高度敏感性促使其根系向下盆湿润土层生长, 并增加了其对下盆水分的吸收利用, 这也是紫花苜蓿有更高的整株Δ
13 C值的原因之一. ...
沙打旺根系提水作用及其机理研究
4
2007
... 即时域反射技术(TDR)具有数据采集准确及时、相对误差小、对土壤扰动小、操作简单、结果可靠等优势, 目前在测定土壤含水量方面得到普遍应用.近年来, 许多研究者采用分根装置与TDR水分测定系统相结合研究了不同植物的根系提水作用(
Wan et al ., 2000 ;
Hirota et al ., 2004 ;
薛小红等, 2007 ;
阿拉木萨和周丽芳, 2011 ).此后, TDR与分根装置的结合成为研究水分提升作用的一种新方法, 这种方法操作简便, 结果可靠, 但关键在于分根装置设计时要注意消除土壤毛管上升水对实验结果的影响(
薛小红等, 2007 ).根据已发表的上下桶实验结果, 上层干燥土壤含水量呈现出有规律的昼夜变化趋势, 即在下午土壤含水量急剧下降, 而在午夜至凌晨时段土壤含水量最高(
李唯等, 2007 ;
薛小红等, 2007 ;
阿拉木萨和周丽芳, 2011 ).我们在制作“上下盆”实验装置时, 在豆科牧草根系长入下盆后, 将上盆用小木块垫起, 使上下盆分离, 并用水把上下盆连接处的土壤冲掉, 只留根系与上盆相连, 从而防止了下盆土壤毛细管作用对上盆水分的补给.同时, 在实验过程中对上盆做了遮雨处理, 因而上盆土壤水分的变化只受植物因素的影响.在我们的实验结果中, 斜茎黄耆土壤含水量昼夜差值与
薛小红等(2007) 报道的最大土壤含水量昼夜差值相近, 两种豆科牧草上盆土壤含水量昼夜差值及提水量存在显著差异, 斜茎黄耆无论在单播还是混播时上盆土壤含水量昼夜差值及日平均提水量均显著高于紫花苜蓿(
表2 ), 说明斜茎黄耆的水力提升效果好于紫花苜蓿. ...
... ).此后, TDR与分根装置的结合成为研究水分提升作用的一种新方法, 这种方法操作简便, 结果可靠, 但关键在于分根装置设计时要注意消除土壤毛管上升水对实验结果的影响(
薛小红等, 2007 ).根据已发表的上下桶实验结果, 上层干燥土壤含水量呈现出有规律的昼夜变化趋势, 即在下午土壤含水量急剧下降, 而在午夜至凌晨时段土壤含水量最高(
李唯等, 2007 ;
薛小红等, 2007 ;
阿拉木萨和周丽芳, 2011 ).我们在制作“上下盆”实验装置时, 在豆科牧草根系长入下盆后, 将上盆用小木块垫起, 使上下盆分离, 并用水把上下盆连接处的土壤冲掉, 只留根系与上盆相连, 从而防止了下盆土壤毛细管作用对上盆水分的补给.同时, 在实验过程中对上盆做了遮雨处理, 因而上盆土壤水分的变化只受植物因素的影响.在我们的实验结果中, 斜茎黄耆土壤含水量昼夜差值与
薛小红等(2007) 报道的最大土壤含水量昼夜差值相近, 两种豆科牧草上盆土壤含水量昼夜差值及提水量存在显著差异, 斜茎黄耆无论在单播还是混播时上盆土壤含水量昼夜差值及日平均提水量均显著高于紫花苜蓿(
表2 ), 说明斜茎黄耆的水力提升效果好于紫花苜蓿. ...
... ;
薛小红等, 2007 ;
阿拉木萨和周丽芳, 2011 ).我们在制作“上下盆”实验装置时, 在豆科牧草根系长入下盆后, 将上盆用小木块垫起, 使上下盆分离, 并用水把上下盆连接处的土壤冲掉, 只留根系与上盆相连, 从而防止了下盆土壤毛细管作用对上盆水分的补给.同时, 在实验过程中对上盆做了遮雨处理, 因而上盆土壤水分的变化只受植物因素的影响.在我们的实验结果中, 斜茎黄耆土壤含水量昼夜差值与
薛小红等(2007) 报道的最大土壤含水量昼夜差值相近, 两种豆科牧草上盆土壤含水量昼夜差值及提水量存在显著差异, 斜茎黄耆无论在单播还是混播时上盆土壤含水量昼夜差值及日平均提水量均显著高于紫花苜蓿(
表2 ), 说明斜茎黄耆的水力提升效果好于紫花苜蓿. ...
... ).我们在制作“上下盆”实验装置时, 在豆科牧草根系长入下盆后, 将上盆用小木块垫起, 使上下盆分离, 并用水把上下盆连接处的土壤冲掉, 只留根系与上盆相连, 从而防止了下盆土壤毛细管作用对上盆水分的补给.同时, 在实验过程中对上盆做了遮雨处理, 因而上盆土壤水分的变化只受植物因素的影响.在我们的实验结果中, 斜茎黄耆土壤含水量昼夜差值与
薛小红等(2007) 报道的最大土壤含水量昼夜差值相近, 两种豆科牧草上盆土壤含水量昼夜差值及提水量存在显著差异, 斜茎黄耆无论在单播还是混播时上盆土壤含水量昼夜差值及日平均提水量均显著高于紫花苜蓿(
表2 ), 说明斜茎黄耆的水力提升效果好于紫花苜蓿. ...
沙打旺根系提水作用及其机理研究
4
2007
... 即时域反射技术(TDR)具有数据采集准确及时、相对误差小、对土壤扰动小、操作简单、结果可靠等优势, 目前在测定土壤含水量方面得到普遍应用.近年来, 许多研究者采用分根装置与TDR水分测定系统相结合研究了不同植物的根系提水作用(
Wan et al ., 2000 ;
Hirota et al ., 2004 ;
薛小红等, 2007 ;
阿拉木萨和周丽芳, 2011 ).此后, TDR与分根装置的结合成为研究水分提升作用的一种新方法, 这种方法操作简便, 结果可靠, 但关键在于分根装置设计时要注意消除土壤毛管上升水对实验结果的影响(
薛小红等, 2007 ).根据已发表的上下桶实验结果, 上层干燥土壤含水量呈现出有规律的昼夜变化趋势, 即在下午土壤含水量急剧下降, 而在午夜至凌晨时段土壤含水量最高(
李唯等, 2007 ;
薛小红等, 2007 ;
阿拉木萨和周丽芳, 2011 ).我们在制作“上下盆”实验装置时, 在豆科牧草根系长入下盆后, 将上盆用小木块垫起, 使上下盆分离, 并用水把上下盆连接处的土壤冲掉, 只留根系与上盆相连, 从而防止了下盆土壤毛细管作用对上盆水分的补给.同时, 在实验过程中对上盆做了遮雨处理, 因而上盆土壤水分的变化只受植物因素的影响.在我们的实验结果中, 斜茎黄耆土壤含水量昼夜差值与
薛小红等(2007) 报道的最大土壤含水量昼夜差值相近, 两种豆科牧草上盆土壤含水量昼夜差值及提水量存在显著差异, 斜茎黄耆无论在单播还是混播时上盆土壤含水量昼夜差值及日平均提水量均显著高于紫花苜蓿(
表2 ), 说明斜茎黄耆的水力提升效果好于紫花苜蓿. ...
... ).此后, TDR与分根装置的结合成为研究水分提升作用的一种新方法, 这种方法操作简便, 结果可靠, 但关键在于分根装置设计时要注意消除土壤毛管上升水对实验结果的影响(
薛小红等, 2007 ).根据已发表的上下桶实验结果, 上层干燥土壤含水量呈现出有规律的昼夜变化趋势, 即在下午土壤含水量急剧下降, 而在午夜至凌晨时段土壤含水量最高(
李唯等, 2007 ;
薛小红等, 2007 ;
阿拉木萨和周丽芳, 2011 ).我们在制作“上下盆”实验装置时, 在豆科牧草根系长入下盆后, 将上盆用小木块垫起, 使上下盆分离, 并用水把上下盆连接处的土壤冲掉, 只留根系与上盆相连, 从而防止了下盆土壤毛细管作用对上盆水分的补给.同时, 在实验过程中对上盆做了遮雨处理, 因而上盆土壤水分的变化只受植物因素的影响.在我们的实验结果中, 斜茎黄耆土壤含水量昼夜差值与
薛小红等(2007) 报道的最大土壤含水量昼夜差值相近, 两种豆科牧草上盆土壤含水量昼夜差值及提水量存在显著差异, 斜茎黄耆无论在单播还是混播时上盆土壤含水量昼夜差值及日平均提水量均显著高于紫花苜蓿(
表2 ), 说明斜茎黄耆的水力提升效果好于紫花苜蓿. ...
... ;
薛小红等, 2007 ;
阿拉木萨和周丽芳, 2011 ).我们在制作“上下盆”实验装置时, 在豆科牧草根系长入下盆后, 将上盆用小木块垫起, 使上下盆分离, 并用水把上下盆连接处的土壤冲掉, 只留根系与上盆相连, 从而防止了下盆土壤毛细管作用对上盆水分的补给.同时, 在实验过程中对上盆做了遮雨处理, 因而上盆土壤水分的变化只受植物因素的影响.在我们的实验结果中, 斜茎黄耆土壤含水量昼夜差值与
薛小红等(2007) 报道的最大土壤含水量昼夜差值相近, 两种豆科牧草上盆土壤含水量昼夜差值及提水量存在显著差异, 斜茎黄耆无论在单播还是混播时上盆土壤含水量昼夜差值及日平均提水量均显著高于紫花苜蓿(
表2 ), 说明斜茎黄耆的水力提升效果好于紫花苜蓿. ...
... ).我们在制作“上下盆”实验装置时, 在豆科牧草根系长入下盆后, 将上盆用小木块垫起, 使上下盆分离, 并用水把上下盆连接处的土壤冲掉, 只留根系与上盆相连, 从而防止了下盆土壤毛细管作用对上盆水分的补给.同时, 在实验过程中对上盆做了遮雨处理, 因而上盆土壤水分的变化只受植物因素的影响.在我们的实验结果中, 斜茎黄耆土壤含水量昼夜差值与
薛小红等(2007) 报道的最大土壤含水量昼夜差值相近, 两种豆科牧草上盆土壤含水量昼夜差值及提水量存在显著差异, 斜茎黄耆无论在单播还是混播时上盆土壤含水量昼夜差值及日平均提水量均显著高于紫花苜蓿(
表2 ), 说明斜茎黄耆的水力提升效果好于紫花苜蓿. ...
黄土丘陵区不同立地条件下紫花苜蓿地土壤水分动态变化
1
2008
... 该实验供试的牧草有3个, 其中, 豆科作物采用紫花苜蓿和斜茎黄耆, 紫花苜蓿及斜茎黄耆均为优良的豆科牧草, 对于促进畜牧业发展、生态环境建设和保护等方面具有重要的意义.这两种豆科牧草的根系都十分发达, 对深层土壤水分的利用能力很强(
山仑等, 2008 ;
杨永东等, 2008 ).本实验采用的紫花苜蓿品种是‘美国牧歌401’, 在实验中的编号为A, 斜茎黄耆为当地栽培种, 在实验中的编号为B.混播的禾本科作物是‘冬牧70’黑麦, 在实验中的编号为C. ...
黄土丘陵区不同立地条件下紫花苜蓿地土壤水分动态变化
1
2008
... 该实验供试的牧草有3个, 其中, 豆科作物采用紫花苜蓿和斜茎黄耆, 紫花苜蓿及斜茎黄耆均为优良的豆科牧草, 对于促进畜牧业发展、生态环境建设和保护等方面具有重要的意义.这两种豆科牧草的根系都十分发达, 对深层土壤水分的利用能力很强(
山仑等, 2008 ;
杨永东等, 2008 ).本实验采用的紫花苜蓿品种是‘美国牧歌401’, 在实验中的编号为A, 斜茎黄耆为当地栽培种, 在实验中的编号为B.混播的禾本科作物是‘冬牧70’黑麦, 在实验中的编号为C. ...
胡杨根系水力再分配的模式、大小及其影响因子
1
2014
... 植物处于深层湿润土壤中的根系吸收的水分可以在夜间蒸腾降低后通过输导组织运输并释放到浅层较干燥的土壤中, 即存在水力提升的现象(
Caldwell et al ., 1998 ).迄今为止, 越来越多的实验证实了水力提升现象的存在, 研究者们观测到很多物种, 包括乔木、灌木、藤本、牧草和农作物等约60种植物存在水分提升作用(
李唯等, 2007 ;
刘小莉等, 2007 ;
郝兴明等, 2009 ;
阿拉木萨和周丽芳, 2011 ;
Brooksbank et al ., 2011 ;
Couvreur et al ., 2014 ;
鱼腾飞等, 2014 ;
Meunier et al ., 2017 ;
李宏侠等, 2018 ). ...
胡杨根系水力再分配的模式、大小及其影响因子
1
2014
... 植物处于深层湿润土壤中的根系吸收的水分可以在夜间蒸腾降低后通过输导组织运输并释放到浅层较干燥的土壤中, 即存在水力提升的现象(
Caldwell et al ., 1998 ).迄今为止, 越来越多的实验证实了水力提升现象的存在, 研究者们观测到很多物种, 包括乔木、灌木、藤本、牧草和农作物等约60种植物存在水分提升作用(
李唯等, 2007 ;
刘小莉等, 2007 ;
郝兴明等, 2009 ;
阿拉木萨和周丽芳, 2011 ;
Brooksbank et al ., 2011 ;
Couvreur et al ., 2014 ;
鱼腾飞等, 2014 ;
Meunier et al ., 2017 ;
李宏侠等, 2018 ). ...
禾豆组合与间作方式对牧草产量及产量稳定性的影响
2
2019
... 豆科与禾本科牧草混播可以有效地提高单位面积草地饲草产量、改善饲用品质、提高土壤肥力, 在世界上许多地区备受重视(
王旭等, 2007 ).研究表明, 紫花苜蓿(
Medicago sativa )与无芒雀麦(
Bromus inermis )混播后增产效果显著, 但混播组合中紫花苜蓿竞争优势明显, 要降低其比例才能使混播组分稳定(
张永亮等, 2019 ).在宁夏中部半干旱带有灌溉条件下斜茎黄耆(沙打旺,
Astragalus laxmannii )与禾本科牧草混播草地中不同物种生长较为均衡, 混播草地持续性好, 并且有较好的产量效应(
朱林等, 2014 ). ...
... 在我们的实验结果中, ‘冬牧70’黑麦与不同豆科牧草混播后的产量存在显著差异, 斜茎黄耆与‘冬牧70’黑麦混播组合中的禾本科牧草产量显著高于紫花苜蓿与‘冬牧70’黑麦混播组合中的禾本科牧草产量.以上结果一方面与紫花苜蓿地上部分对于禾草的竞争力较高, 其旺盛的枝叶对禾草遮阴效应较强而影响了伴生禾草生长(
朱林等, 2014 ;
张永亮等, 2019 ), 另一方面也可能与斜茎黄耆水力提升作用高于紫花苜蓿有关.在整个生长季节, 斜茎黄耆与‘冬牧70’黑麦混播组合的土壤含水量总体说来高于紫花苜蓿-‘冬牧70’黑麦(
图2 ), 而且斜茎黄耆的提水量总体上比紫花苜蓿的高, 因此, 斜茎黄耆与‘冬牧70’黑麦组合中上盆土壤较高的含水量促进了混播组合中禾本科牧草的生长.在同一种豆科牧草与‘冬牧70’黑麦混播时, 以AC2和BC1组合中的禾本科产量为最高, 说明在紫花苜蓿与‘冬牧70’黑麦比例为5:5、斜茎黄耆与‘冬牧70’黑麦比例为3:7混播时, 豆科牧草水力提升作用对伴生的禾本科牧草生长的贡献最大, 这可能是不同禾豆比例的盆栽中豆科牧草群体产生的提水量与其对禾本科牧草水分竞争相“权衡”的结果.在不同混播组合中, ‘冬牧70’黑麦茎秆水
δ D值与处理前土壤水
δ D值相比均显著上升, 并介于上盆土壤水
δ D值和下盆土壤水
δ D值之间, 说明两种豆科牧草提升的标记水被伴生的禾本科牧草所吸收利用.在紫花苜蓿与‘冬牧70’黑麦的不同混播比例中, 以AC2组合中禾本科牧草茎秆水
δ D值为最高; 在斜茎黄耆与‘冬牧70’黑麦的不同混播比例中, 以BC1组合中禾本科牧草茎秆水
δ D值最高(
图4 ), 说明以上两个混播组合中的禾本科牧草生长所利用的水分主要来自于豆科牧草的水力提升作用.对于同一种豆科牧草与‘冬牧70’黑麦的混播组合, AC2组合中禾本科整株Δ
13 C和产量最高(
图5 ), BC1组合中禾本科产量最高(
表1 ), 这一结果支持了上述结论. ...
禾豆组合与间作方式对牧草产量及产量稳定性的影响
2
2019
... 豆科与禾本科牧草混播可以有效地提高单位面积草地饲草产量、改善饲用品质、提高土壤肥力, 在世界上许多地区备受重视(
王旭等, 2007 ).研究表明, 紫花苜蓿(
Medicago sativa )与无芒雀麦(
Bromus inermis )混播后增产效果显著, 但混播组合中紫花苜蓿竞争优势明显, 要降低其比例才能使混播组分稳定(
张永亮等, 2019 ).在宁夏中部半干旱带有灌溉条件下斜茎黄耆(沙打旺,
Astragalus laxmannii )与禾本科牧草混播草地中不同物种生长较为均衡, 混播草地持续性好, 并且有较好的产量效应(
朱林等, 2014 ). ...
... 在我们的实验结果中, ‘冬牧70’黑麦与不同豆科牧草混播后的产量存在显著差异, 斜茎黄耆与‘冬牧70’黑麦混播组合中的禾本科牧草产量显著高于紫花苜蓿与‘冬牧70’黑麦混播组合中的禾本科牧草产量.以上结果一方面与紫花苜蓿地上部分对于禾草的竞争力较高, 其旺盛的枝叶对禾草遮阴效应较强而影响了伴生禾草生长(
朱林等, 2014 ;
张永亮等, 2019 ), 另一方面也可能与斜茎黄耆水力提升作用高于紫花苜蓿有关.在整个生长季节, 斜茎黄耆与‘冬牧70’黑麦混播组合的土壤含水量总体说来高于紫花苜蓿-‘冬牧70’黑麦(
图2 ), 而且斜茎黄耆的提水量总体上比紫花苜蓿的高, 因此, 斜茎黄耆与‘冬牧70’黑麦组合中上盆土壤较高的含水量促进了混播组合中禾本科牧草的生长.在同一种豆科牧草与‘冬牧70’黑麦混播时, 以AC2和BC1组合中的禾本科产量为最高, 说明在紫花苜蓿与‘冬牧70’黑麦比例为5:5、斜茎黄耆与‘冬牧70’黑麦比例为3:7混播时, 豆科牧草水力提升作用对伴生的禾本科牧草生长的贡献最大, 这可能是不同禾豆比例的盆栽中豆科牧草群体产生的提水量与其对禾本科牧草水分竞争相“权衡”的结果.在不同混播组合中, ‘冬牧70’黑麦茎秆水
δ D值与处理前土壤水
δ D值相比均显著上升, 并介于上盆土壤水
δ D值和下盆土壤水
δ D值之间, 说明两种豆科牧草提升的标记水被伴生的禾本科牧草所吸收利用.在紫花苜蓿与‘冬牧70’黑麦的不同混播比例中, 以AC2组合中禾本科牧草茎秆水
δ D值为最高; 在斜茎黄耆与‘冬牧70’黑麦的不同混播比例中, 以BC1组合中禾本科牧草茎秆水
δ D值最高(
图4 ), 说明以上两个混播组合中的禾本科牧草生长所利用的水分主要来自于豆科牧草的水力提升作用.对于同一种豆科牧草与‘冬牧70’黑麦的混播组合, AC2组合中禾本科整株Δ
13 C和产量最高(
图5 ), BC1组合中禾本科产量最高(
表1 ), 这一结果支持了上述结论. ...
宁夏中部灌水量对豆-禾牧草混播的影响
2
2014
... 豆科与禾本科牧草混播可以有效地提高单位面积草地饲草产量、改善饲用品质、提高土壤肥力, 在世界上许多地区备受重视(
王旭等, 2007 ).研究表明, 紫花苜蓿(
Medicago sativa )与无芒雀麦(
Bromus inermis )混播后增产效果显著, 但混播组合中紫花苜蓿竞争优势明显, 要降低其比例才能使混播组分稳定(
张永亮等, 2019 ).在宁夏中部半干旱带有灌溉条件下斜茎黄耆(沙打旺,
Astragalus laxmannii )与禾本科牧草混播草地中不同物种生长较为均衡, 混播草地持续性好, 并且有较好的产量效应(
朱林等, 2014 ). ...
... 在我们的实验结果中, ‘冬牧70’黑麦与不同豆科牧草混播后的产量存在显著差异, 斜茎黄耆与‘冬牧70’黑麦混播组合中的禾本科牧草产量显著高于紫花苜蓿与‘冬牧70’黑麦混播组合中的禾本科牧草产量.以上结果一方面与紫花苜蓿地上部分对于禾草的竞争力较高, 其旺盛的枝叶对禾草遮阴效应较强而影响了伴生禾草生长(
朱林等, 2014 ;
张永亮等, 2019 ), 另一方面也可能与斜茎黄耆水力提升作用高于紫花苜蓿有关.在整个生长季节, 斜茎黄耆与‘冬牧70’黑麦混播组合的土壤含水量总体说来高于紫花苜蓿-‘冬牧70’黑麦(
图2 ), 而且斜茎黄耆的提水量总体上比紫花苜蓿的高, 因此, 斜茎黄耆与‘冬牧70’黑麦组合中上盆土壤较高的含水量促进了混播组合中禾本科牧草的生长.在同一种豆科牧草与‘冬牧70’黑麦混播时, 以AC2和BC1组合中的禾本科产量为最高, 说明在紫花苜蓿与‘冬牧70’黑麦比例为5:5、斜茎黄耆与‘冬牧70’黑麦比例为3:7混播时, 豆科牧草水力提升作用对伴生的禾本科牧草生长的贡献最大, 这可能是不同禾豆比例的盆栽中豆科牧草群体产生的提水量与其对禾本科牧草水分竞争相“权衡”的结果.在不同混播组合中, ‘冬牧70’黑麦茎秆水
δ D值与处理前土壤水
δ D值相比均显著上升, 并介于上盆土壤水
δ D值和下盆土壤水
δ D值之间, 说明两种豆科牧草提升的标记水被伴生的禾本科牧草所吸收利用.在紫花苜蓿与‘冬牧70’黑麦的不同混播比例中, 以AC2组合中禾本科牧草茎秆水
δ D值为最高; 在斜茎黄耆与‘冬牧70’黑麦的不同混播比例中, 以BC1组合中禾本科牧草茎秆水
δ D值最高(
图4 ), 说明以上两个混播组合中的禾本科牧草生长所利用的水分主要来自于豆科牧草的水力提升作用.对于同一种豆科牧草与‘冬牧70’黑麦的混播组合, AC2组合中禾本科整株Δ
13 C和产量最高(
图5 ), BC1组合中禾本科产量最高(
表1 ), 这一结果支持了上述结论. ...
宁夏中部灌水量对豆-禾牧草混播的影响
2
2014
... 豆科与禾本科牧草混播可以有效地提高单位面积草地饲草产量、改善饲用品质、提高土壤肥力, 在世界上许多地区备受重视(
王旭等, 2007 ).研究表明, 紫花苜蓿(
Medicago sativa )与无芒雀麦(
Bromus inermis )混播后增产效果显著, 但混播组合中紫花苜蓿竞争优势明显, 要降低其比例才能使混播组分稳定(
张永亮等, 2019 ).在宁夏中部半干旱带有灌溉条件下斜茎黄耆(沙打旺,
Astragalus laxmannii )与禾本科牧草混播草地中不同物种生长较为均衡, 混播草地持续性好, 并且有较好的产量效应(
朱林等, 2014 ). ...
... 在我们的实验结果中, ‘冬牧70’黑麦与不同豆科牧草混播后的产量存在显著差异, 斜茎黄耆与‘冬牧70’黑麦混播组合中的禾本科牧草产量显著高于紫花苜蓿与‘冬牧70’黑麦混播组合中的禾本科牧草产量.以上结果一方面与紫花苜蓿地上部分对于禾草的竞争力较高, 其旺盛的枝叶对禾草遮阴效应较强而影响了伴生禾草生长(
朱林等, 2014 ;
张永亮等, 2019 ), 另一方面也可能与斜茎黄耆水力提升作用高于紫花苜蓿有关.在整个生长季节, 斜茎黄耆与‘冬牧70’黑麦混播组合的土壤含水量总体说来高于紫花苜蓿-‘冬牧70’黑麦(
图2 ), 而且斜茎黄耆的提水量总体上比紫花苜蓿的高, 因此, 斜茎黄耆与‘冬牧70’黑麦组合中上盆土壤较高的含水量促进了混播组合中禾本科牧草的生长.在同一种豆科牧草与‘冬牧70’黑麦混播时, 以AC2和BC1组合中的禾本科产量为最高, 说明在紫花苜蓿与‘冬牧70’黑麦比例为5:5、斜茎黄耆与‘冬牧70’黑麦比例为3:7混播时, 豆科牧草水力提升作用对伴生的禾本科牧草生长的贡献最大, 这可能是不同禾豆比例的盆栽中豆科牧草群体产生的提水量与其对禾本科牧草水分竞争相“权衡”的结果.在不同混播组合中, ‘冬牧70’黑麦茎秆水
δ D值与处理前土壤水
δ D值相比均显著上升, 并介于上盆土壤水
δ D值和下盆土壤水
δ D值之间, 说明两种豆科牧草提升的标记水被伴生的禾本科牧草所吸收利用.在紫花苜蓿与‘冬牧70’黑麦的不同混播比例中, 以AC2组合中禾本科牧草茎秆水
δ D值为最高; 在斜茎黄耆与‘冬牧70’黑麦的不同混播比例中, 以BC1组合中禾本科牧草茎秆水
δ D值最高(
图4 ), 说明以上两个混播组合中的禾本科牧草生长所利用的水分主要来自于豆科牧草的水力提升作用.对于同一种豆科牧草与‘冬牧70’黑麦的混播组合, AC2组合中禾本科整株Δ
13 C和产量最高(
图5 ), BC1组合中禾本科产量最高(
表1 ), 这一结果支持了上述结论. ...
 ,
, ,1,2,*, 王甜甜1,2, 赵学琳1,2, 祁亚淑3, 许兴1,2
,1,2,*, 王甜甜1,2, 赵学琳1,2, 祁亚淑3, 许兴1,2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT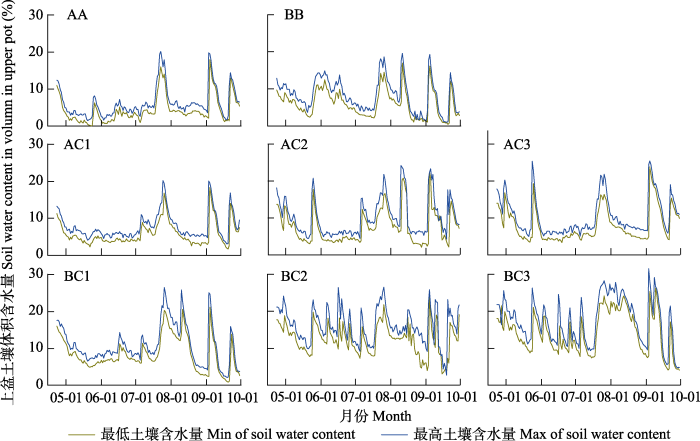 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT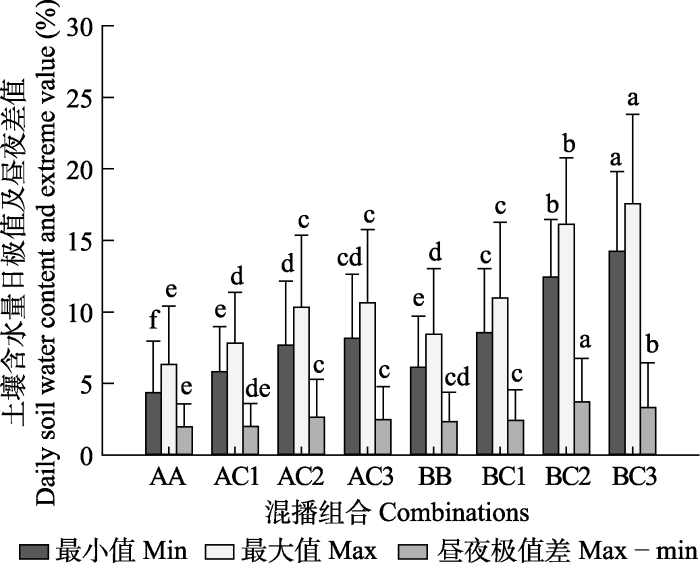 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT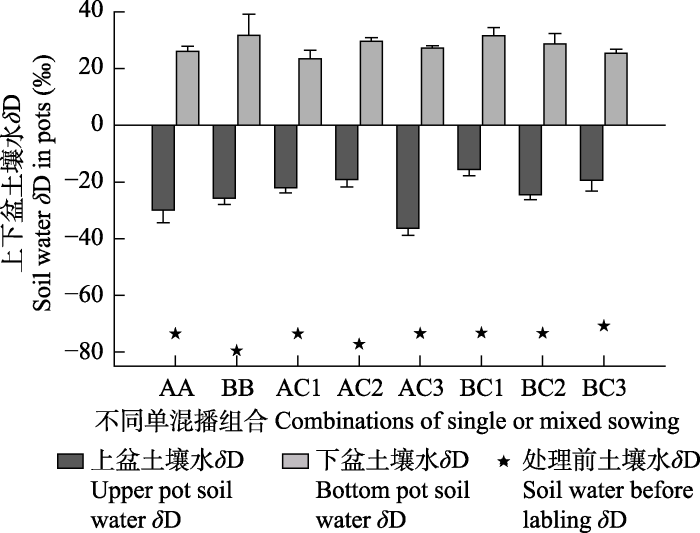 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT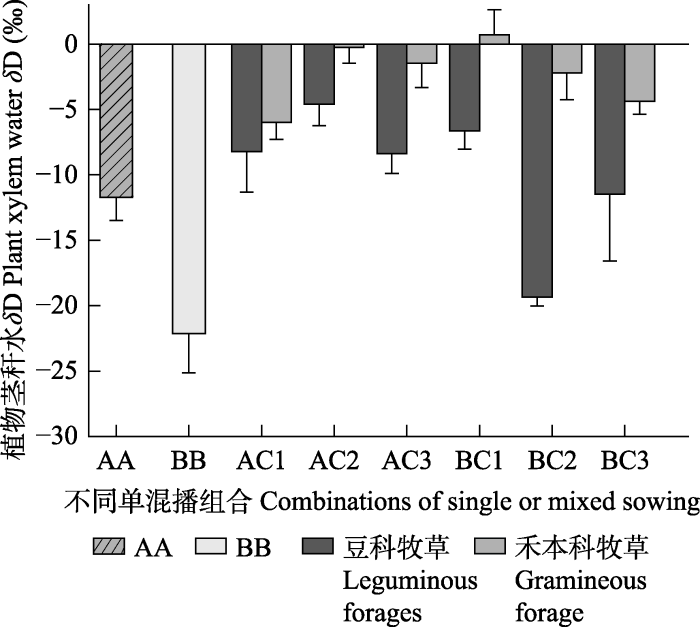 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT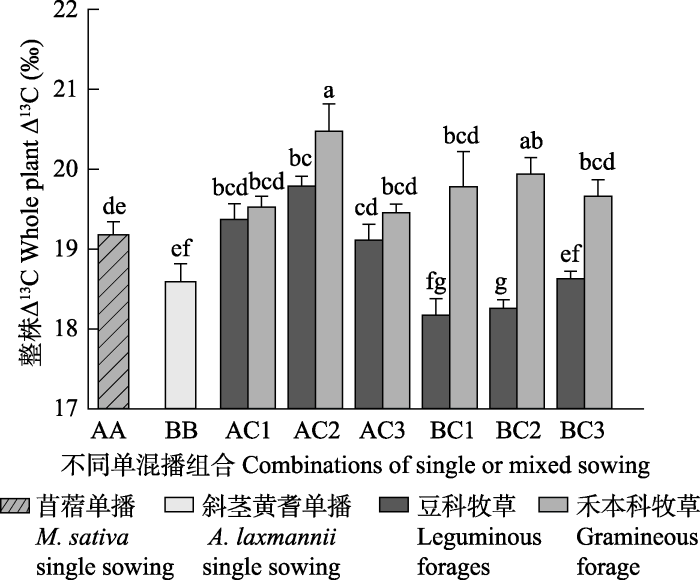 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}