Hydraulic and photosynthetic characteristics differ between co-generic tree and liana species: a case study of Millettia and Gnetum in tropical forest
Hui-Qing SONG, Ming-Yuan NI, Shi-Dan ZHU,,*Guangxi Key Laboratory of Forest Ecology and Conservation, College of Forestry, Guangxi University, Nanning 530004, China; and State Key Laboratory for Conservation and Utilization of Subtropical Agro-bioresources, Guangxi University, Nanning 530004, China
Abstract Aims Liana is an important component of tropical forest, and exert a significant impact on community structure and function. Previous studies have found significant differences in hydraulic traits between lianas and trees, as indicated that lianas tended to have large and long vessels to compensate hydraulically to their thin stems, resulting in high hydraulic conductivity but low resistant to drought-induced cavitation. In order to reduce the influence of different genotypes on the comparative results, we aimed to compare the differences in hydraulic and photosynthetic characteristics between the two life forms from two genera Millettia and Gnetum. Methods We measured branch and leaf hydraulic properties, sapwood density, gas exchange rates in the dry season for nine tree and liana species grown in common garden. We compared the hydraulic and photosynthetic traits between each species using one-way ANOVA. In addition, we analyzed hydraulic efficiency-safety trade-off, and the relationship between dry-season photosynthetic rates and hydraulic traits. Important findings (1) There was a significant variations in hydraulic traits in genus Millettia, which was related to their light requirements and life forms. Compared with trees, the shade-tolerant liana species had lower hydraulic conductivity and higher resistance to cavitation. (2) Despite angiosperm-like characteristics such as vessels and broad pinnate-veined leaves, the Gnetum tree species had the lowest hydraulic conductivity among the nine species. However, the Gnetum liana species had higher hydraulic conductivity, comparable to light-demanding angiosperm species in this study. (3) There was no significant trade-off between hydraulic conductivity efficiency and hydraulic safety in both branch- and leaf-level across all the species, or within each plant group. (4) Compared to co-generic tree species, liana species’ leaves were more resistant to cavitation than branches, as indicated by higher maximum net photosynthetic rates and stomatal conductance during the dry season. These results support the hypothesis of “growth advantages at dry season” for liana species. This study reveals the high diversity and significance of hydraulic functioning in tropical lianas. Extensive measurements of hydraulic properties are needed to promote understanding of tropical species response to environmental change. Keywords:hydraulic function;cavitation resistance;trade-off;vulnerability segmentation;photosynthetic rates
PDF (1542KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 宋慧清, 倪鸣源, 朱师丹. 乔木与木质藤本的水力与光合性状的差异: 以热带森林崖豆藤属和买麻藤属为例. 植物生态学报, 2020, 44(3): 192-204. DOI: 10.17521/cjpe.2019.0304 SONG Hui-Qing, NI Ming-Yuan, ZHU Shi-Dan. Hydraulic and photosynthetic characteristics differ between co-generic tree and liana species: a case study of Millettia and Gnetum in tropical forest. Chinese Journal of Plant Ecology, 2020, 44(3): 192-204. DOI: 10.17521/cjpe.2019.0304
Table 1 表1 表19种植物的生活型、名称缩写、最大导管长度以及生态学特性描述 Table 1Life form, species abbreviations, maximum vessel length and ecological descriptions for the nine woody species
物种 Species
生活型 Life form
缩写 Abbreviation
最大导管长度 Maximum vessel length (m)
原生生境分布 Native habitat of adult in the forest
崖豆藤属(豆科) Millettia (Fabaceae)
思茅崖豆 Millettia leptobotrya
乔木 Tree
M. lep
0.69
山坡疏林或常绿阔叶林中, 海拔300-1 000 m Open forest on slopes or evergreen broad-leaved forest; 300-1 000 m a.s.l.
红河崖豆 Millettia cubittii
乔木 Tree
M. cub
0.49
河边的林地或路边, 海拔300-1 000 m Riparian forest or roadside; 300-1 000 m a.s.l.
变色鸡血藤 Millettia versicolor
乔木 Tree
M. ver
引种于非洲南部热带次生林、稀树草原 Tropical secondary forests and savannas of southern Africa
厚果崖豆藤 Millettia pachycarpa
藤本 Liana
M. pac
1.50
山坡疏林、阔叶林内或路边, 海拔100-2 000 m Open forest on slopes, broad-leaved forest or roadside; 100-2 000 m a.s.l.
香花崖豆藤 Millettia dielsiana
藤本 Liana
M. die
0.82
山坡杂木林与灌丛中, 海拔300-2 500 m Tree-shrub mixed forest on slopes; 300-2 500 m a.s.l.
海南崖豆藤 Millettia pachyloba
藤本 Liana
M. pab
沟谷常绿阔叶林中, 海拔1 500 m以下 Evergreen broad-leaved forests in valleys; below 1 500 m a.s.l.
买麻藤属(买麻藤科) Gnetum (Gnetaceae)
灌状买麻藤 Gnetum gnemon
乔木 Tree
G. gne
湿润的常绿次生森林下, 海拔1 600-2 000 m Moist evergreen secondary forests; 1 600-2 000 m a.s.l.
少苞买麻藤 Gnetum brunonianum
乔木 Tree
G. bru
1.14
海拔350 m的阔叶林下 Broad-leaved forest; below 350 m a.s.l.
小叶买麻藤 Gnetum parvifolium
藤本 Liana
G. par
2.00
海拔较低的干燥平地或湿润谷地的森林,海拔100-1 000m Dry flat or moist valleys forests at lower altitude; 100-1 000 m a.s.l.
Native habitat descriptions of these species were extracted from Flora Reipublicae Popularis Sinicae (Editorial Committee of Flora of China, Chinese Academy of Sciences, 1993) and Li et al. (1996). 原生生境信息来源于《中国植物志》(中国科学院中国植物志编辑委员会, 1993)和《引种植物名录》(李延辉等, 1996)。
本试验在中国科学院西双版纳热带植物园(21.68° N, 101.42° E, 海拔580 m)内进行。研究地点位于热带北缘, 年降水量1 550 mm, 年平均气温21.7 ℃, 相对湿度85%。受西南季风的影响, 一年中有明显的干季(11月-翌年4月)和雨季(5-10月)之分, 雨季降水量占全年的80%以上。
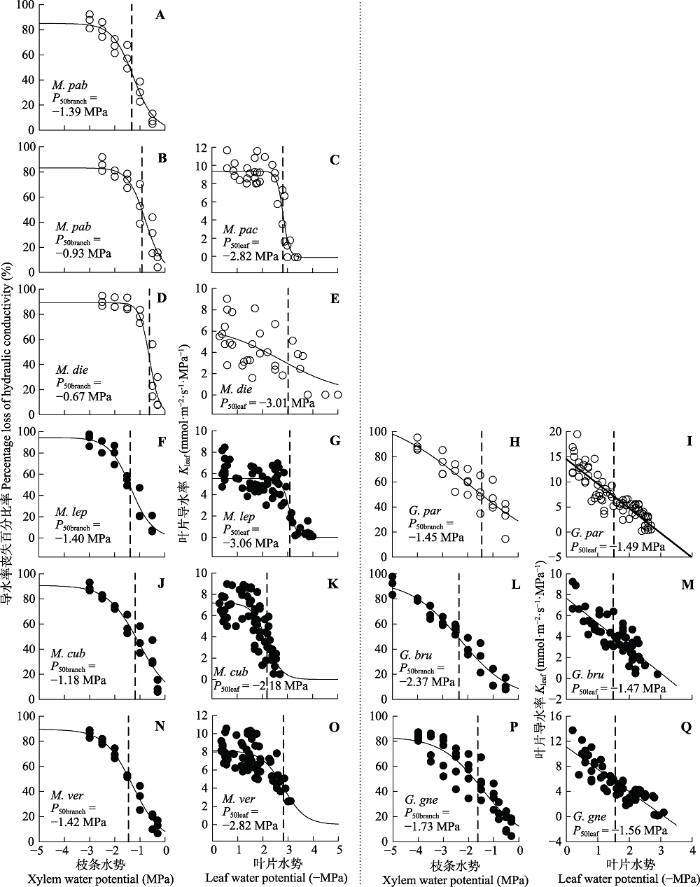
Fig. 1Branch and leaf vulnerability curves of the woody species in Millettia (left) and Gnetum (right). Filled and open circles indicate tree and liana species, respectively. Species abbreviations are shown in Table 1. Water potential at 50% loss of branch hydraulic conductivity (P50branch) and leaf hydraulic conductance (P50leaf) are indicated by vertical dashed lines. Kleaf, leaf hydraulic conductivity.
Fig. 2Comparison in branch hydraulic traits among the nine woody species in Millettia and Gnetum (mean + SE). ks, sapwood specific hydraulic conductivity; kl, leaf specific hydraulic conductivity; Al/As, leaf area/sapwood area ratio; WD, sapwood density. Filled and open bars indicate tree and liana species, respectively. Different letters indicate significant difference at p < 0.05. Species abbreviations are shown in Table 1.
Fig. 3Relationship between sapwood specific hydraulic conductivity (ks) and vulnerability to cavitation in branches (A), and relationship between maximum hydraulic leaf conductance (Kleaf-max) and vulnerability to cavitation in leaves (B). P50branch, xylem water potential at 50% loss of branch hydraulic conductivity; P50leaf, leaf water potential at 50% loss of leaf hydraulic conductance. Error bars are standard errors.
Fig. 4Relationships between maximum net photosynthetic rates during the dry season (Amax) and leaf specific hydraulic conductivity (kl)(A), or maximum leaf hydraulic conductivity (Kleaf-max)(B), and relationships between the difference in P50 between leaves and branches (P50leaf-branch) and Amax (C) or maximum stomatal conductance during the dry season (gs-max)(D). Error bars are standard errors.
Supplement I Mean value, standard error and comparisons significance of six hydraulic traits between tree and liana in different groups of Millettia and Gnetun
SSupplement Ⅱ Trait correlation between function traits across all species, within each genus and within each life in Millettia and Gnetun of this study
AllenCD, MacAladyAK, ChenchouniH, BacheletD, McDowellN, VennetierM, KitzbergerT, RiglingA, BreshearsDD, HoggEHT, GonzalezP, FenshamR, ZhangZ, CastroJ, DemidovaN, LimJH, AllardG, RunningSW, SemerciA, CobbN (2010). A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests Forest Ecology and Management, 259, 660-684. DOI:10.1016/j.foreco.2009.09.001URL [本文引用: 1]
AngyalossyV, PaceMR, LimaAC (2014). Liana anatomy: a broad perspective on structural evolution of the vascular system//Schnitzer SA, Bongers F, Burnham RJ, Putz FE Ecology of Lianas. John Wiley & Sons, Chichester, UK. 253-287. [本文引用: 1]
BittencourtPRL, PereiraL, OliveiraRS (2016). On xylem hydraulic efficiencies, wood space-use and the safety-efficiency tradeoff New Phytologist, 211, 1152-1155. DOI:10.1111/nph.2016.211.issue-4URL [本文引用: 1]
BlackmanCJ, BrodribbTJ, JordanGJ (2010). Leaf hydraulic vulnerability is related to conduit dimensions and drought resistance across a diverse range of woody angiosperms New Phytologist, 188, 1113-1123. DOI:10.1111/j.1469-8137.2010.03439.xURL [本文引用: 1]
BrodribbTJ, BowmanDJMS, NicholsS, DelzonS, BurlettR (2010). Xylem function and growth rate interact to determine recovery rates after exposure to extreme water deficit New Phytologist, 188, 533-542. DOI:10.1111/j.1469-8137.2010.03393.xURL [本文引用: 1]
BrodribbTJ, FeildTS (2000). Stem hydraulic supply is linked to leaf photosynthetic capacity: evidence from New Caledonian and Tasmanian rainforests Plant, Cell & Environment, 23, 1381-1388. [本文引用: 1]
BrodribbTJ, HolbrookNM (2003). Stomatal closure during leaf dehydration, correlation with other leaf physiological traits Plant Physiology, 132, 2166-2173. DOI:10.1104/pp.103.023879URL [本文引用: 2]
BrodribbTJ, HolbrookNM (2004). Stomatal protection against hydraulic failure: a comparison of coexisting ferns and angiosperms New Phytologist, 162, 663-670. DOI:10.1111/nph.2004.162.issue-3URL [本文引用: 1]
BrodribbTJ, HolbrookNM (2006). Declining hydraulic efficiency as transpiring leaves desiccate: two types of response Plant, Cell & Environment, 29, 2205-2215. [本文引用: 1]
BucciSJ, ScholzFG, CampanelloPI, MonttiL, Jimenez- CastilloM, RockwellFA, MannaLL, GuerraP, BernalPL, TroncosoO, EnricciJ, HolbrookMN, GoldsteinG (2012). Hydraulic differences along the water transport system of South American Nothofagus species: Do leaves protect the stem functionality? Tree Physiology, 32, 880-893. DOI:10.1093/treephys/tps054URL [本文引用: 1]
BuckleyTN, JohnGP, ScoffoniC, SackL (2015). How does leaf anatomy influence water transport outside the xylem? Plant Physiology, 168, 1616-1635. DOI:10.1104/pp.15.00731URL [本文引用: 1]
CaiZQ, SchnitzerSA, BongersF (2009). Seasonal differences in leaf-level physiology give lianas a competitive advantage over trees in a tropical seasonal forest Oecologia, 161, 25-33. DOI:10.1007/s00442-009-1355-4URL [本文引用: 1]
CampbellMJ, EdwardsW, MagrachA, AlamgirM, PorolakG, MohandassD, LauranceWF (2018). Edge disturbance drives liana abundance increase and alteration of Liana- host tree interactions in tropical forest fragments Ecology and Evolution, 8, 4237-4251. DOI:10.1002/ece3.2018.8.issue-8URL [本文引用: 1]
CelisG, AvalosG (2013). Acclimation of seedlings of Gnetum leyboldii Tul.(Gnetaceae) to light changes in a tropical rain forest Revista De Biologia Tropical, 61, 1859-1868. [本文引用: 1]
ChenYJ, CaoKF, SchnitzerSA, FanZX, ZhangJL, BongersF (2015). Water-use advantage for lianas over trees in tropical seasonal forests New Phytologist, 205, 128-136. DOI:10.1111/nph.13036URL [本文引用: 1]
ChoatB, DraytonWM, BrodersenC, MattthewsMA, ShackelKA, WadaH, McElroneAJ (2010). Measurement of vulnerability to water stress-induced cavitation in grapevine: a comparison of four techniques applied to a long-vesseled species Plant, Cell & Environment, 33, 1502-1512. [本文引用: 1]
ChoatB, JansenS, BrodribbTJ, CochardH, DelzonS, BhaskarR, BucciSJ, FeildTS, GleasonSM, HackeUG, JacobsenAL, LensF, MaheraliH, Martínez-VilaltaJ, MayrS, MencucciniM, MitchellPJ, NardiniA, PittermannJ, PrattRB, SperryJS, WestobyM, WrightIJ, ZanneAE (2012). Global convergence in the vulnerability of forests to drought Nature, 491, 752-755. DOI:10.1038/nature11688URL [本文引用: 1]
CochardH, CruiziatP, TyreeMT (1992). Use of positive pressures to establish vulnerability curves: further support for the air-seeding hypothesis and implications for pressure- volume analysis Plant Physiology, 100, 205-209. DOI:10.1104/pp.100.1.205URL [本文引用: 1]
CollinsCG, WrightSJ, WurzburgerN (2016). Root and leaf traits reflect distinct resource acquisition strategies in tropical lianas and trees Oecologia, 180, 1037-1047. DOI:10.1007/s00442-015-3410-7URL [本文引用: 1]
CruiziatP, CochardH, AméglioT (2002). Hydraulic architecture of trees: main concepts and results Annals of Forest Science, 59, 723-752. DOI:10.1051/forest:2002060URL [本文引用: 1]
de GuzmanME, SantiagoLS, SchnitzerSA, álvarez-CansinoL (2017). Trade-offs between water transport capacity and drought resistance in neotropical canopy liana and tree species Tree Physiology, 37, 1404-1414. [本文引用: 1]
DengN, HouC, LiuCX, LiMH, BartishI, TianYX, ChenW, DuCJ, JiangZP, ShiSQ (2019). Significance of photosynthetic characters in the evolution of Asian Gnetum(Gnetales) Frontiers in Plant Science, 10, 39. DOI: 10.3389/fpls.2019.00039. DOI:10.3389/fpls.2019.00039URL [本文引用: 1]
DoyleJA, DonoghueMJ (1986). Seed plant phylogeny and the origin of angiosperms: an experimental cladistic approach The Botanical Review, 52, 321-431. DOI:10.1007/BF02861082URL [本文引用: 1]
Editorial Committee of Flora of China, Chinese Academy of Sciences (1993). Flora Reipublicae Popularis Sinicae. Science Press, Beijing. [本文引用: 4]
EnnajehM, Sim?esF, KhemiraH, CochardH (2011). How reliable is the double-ended pressure sleeve technique for assessing xylem vulnerability to cavitation in woody angiosperms? Physiologia Plantarum, 142, 205-210. DOI:10.1111/ppl.2011.142.issue-3URL [本文引用: 2]
EwersFW, RosellJA, OlsonME (2015). Lianas as structural parasites//Hacke U Functional and Ecological Xylem Anatomy. Springer,Cham. 163-188. [本文引用: 1]
FeildTS, BalunL (2008). Xylem hydraulic and photosynthetic function of Gnetum(Gnetales) species from Papua New Guinea New Phytologist, 177, 665-675. DOI:10.1111/nph.2008.177.issue-3URL [本文引用: 2]
FuPL, JiangYJ, WangAY, BrodribbTJ, ZhangJL, ZhuSD, CaoKF (2012). Stem hydraulic traits and leaf water-stress tolerance are co-ordinated with the leaf phenology of angiosperm trees in an Asian tropical dry karst forest Annals of Botany, 110, 189-199. DOI:10.1093/aob/mcs092URL [本文引用: 1]
GleasonSM, WestobyM, JansenS, ChoatB, HackeUG, PrattRB, BhaskarR, BrodribbTJ, BucciSJ, CaoKF, CochardH, DelzonS, DomecJC, FanZX, FeildTS, JacobsenAL, JohnsonDM, LensF, MaheraliH, Martínez-VilaltaJ, MayrS, McCullohKA, MencucciniM, MitchellPJ, MorrisH, NardiniA, PittermannJ, PlavcováL, SchreiberSG, SperryJS, WrightIJ, ZanneAE (2016). Weak tradeoff between xylem safety and xylem-specific hydraulic efficiency across the world’s woody plant species New Phytologist, 209, 123-136. DOI:10.1111/nph.13646URL [本文引用: 1]
IsnardS, FeildTS (2014). The evolution of angiosperm lianescence: a perspective from xylem structure-function //Schnitzer SA, Bongers F, Burnham RJ, Putz FE Ecology of Lianas. John Wiley & Sons,Chichester,UK. 221-238. [本文引用: 1]
IsnardS, SilkWK (2009). Moving with climbing plants from Charles Darwin’s time into the 21st century American Journal of Botany, 96, 1205-1221. DOI:10.3732/ajb.0900045URL [本文引用: 1]
KotowskaMM, HertelD, RajabYA, BarusH, SchuldtB (2015). Patterns in hydraulic architecture from roots to branches in six tropical tree species from cacao agroforestry and their relation to wood density and stem growth Frontiers in Plant Science, 6, 191. DOI: 10.3389/fpls.2015.00191. [本文引用: 1]
LiYH, PeiSJ, XuZF (1996). List of Palnt in Xishuangbanna. Yunnan Nationality Press, Kunming. [本文引用: 4]
MaRY, ZhangJL, CavaleriMA, SterckF, StrijkJS, CaoKF (2015). Convergent evolution towards high net carbon gain efficiency contributes to the shade tolerance of palms (Arecaceae) PLOS ONE, 10, e0140384. DOI: 10.1371/journal.pone.0140384. DOI:10.1371/journal.pone.0140384URL [本文引用: 1]
MaréchauxI, BartlettMK, IribarA, SackL, ChaveJ (2017). Stronger seasonal adjustment in leaf turgor loss point in lianas than trees in an Amazonian forest Biology Letters, 13, 20160819. DOI: 10.1098/rsbl.2016.0819. DOI:10.1098/rsbl.2016.0819URL [本文引用: 1]
MarkesteijnL, PoorterL, BongersF, PazH, SackL (2011). Hydraulics and life history of tropical dry forest tree species: coordination of species’ drought and shade tolerance New Phytologist, 191, 480-495. DOI:10.1111/j.1469-8137.2011.03708.xURL [本文引用: 1]
McCullohKA, JohnsonDM, MeinzerFC, WoodruffDR (2014). The dynamic pipeline: hydraulic capacitance and xylem hydraulic safety in four tall conifer species Plant, Cell & Environment, 37, 1171-1183. [本文引用: 1]
MeinzerFC, McCullohKA, LachenbruchB, WoodruffDR, JohnsonDM (2010). The blind men and the elephant: the impact of context and scale in evaluating conflicts between plant hydraulic safety and efficiency Oecologia, 164, 287-296. DOI:10.1007/s00442-010-1734-xURL [本文引用: 1]
MencucciniM, MinunnoF, SalmonY, Martínez-VilaltaJ, H?ltt?T (2015). Coordination of physiological traits involved in drought-induced mortality of woody plants New Phytologist, 208, 396-409. DOI:10.1111/nph.2015.208.issue-2URL [本文引用: 1]
PammenterNW, van der WilligenC (1998). A mathematical and statistical analysis of the curves illustrating vulnerability of xylem to cavitation Tree Physiology, 18, 589-593. DOI:10.1093/treephys/18.8-9.589URL [本文引用: 1]
PivovaroffAL, PasquiniSC, de GuzmanME, AlstadKP, StemkeJS, SantiagoLS (2016). Multiple strategies for drought survival among woody plant species Functional Ecology, 30, 517-526. DOI:10.1111/fec.2016.30.issue-4URL [本文引用: 1]
PutzFE (1984). The natural history of lianas on barro Colorado Island, Panama Ecology, 65, 1713-1724. DOI:10.2307/1937767URL [本文引用: 1]
Rodríguez-RonderosME, BohrerG, Sanchez-AzofeifaA, PowersJS, SchnitzerSA (2016). Contribution of lianas to plant area index and canopy structure in a Panamanian forest Ecology, 97, 3271-3277. DOI:10.1002/ecy.1597URL [本文引用: 1]
SackL, FroleK (2006). Leaf structural diversity is related to hydraulic capacity in tropical rain forest trees Ecology, 87, 483-491. DOI:10.1890/05-0710URL [本文引用: 1]
SantiagoLS, de GuzmanME, BaralotoC, VogenbergJE, BrodieM, HéraultB, FortunelC, BonalD (2018). Coordination and trade-offs among hydraulic safety, efficiency and drought avoidance traits in Amazonian rainforest canopy tree species New Phytologist, 218, 1015-1024. DOI:10.1111/nph.2018.218.issue-3URL [本文引用: 1]
SantiagoLS, GoldsteinG, MeinzerFC, FisherJB, MacHadoK, WoodruffD, JonesT (2004). Leaf photosynthetic traits scale with hydraulic conductivity and wood density in Panamanian forest canopy trees Oecologia, 140, 543-550. DOI:10.1007/s00442-004-1624-1URL [本文引用: 1]
SantiagoLS, PasquiniSC, de GuzmanME (2014). Physiological implications of the liana growth form//Schnitzer SA, Bongers F, Burnham RJ, Putz FE. Ecology of Lianas. John Wiley & Sons, Chichester, UK. 288-298. [本文引用: 1]
SantiagoLS, WrightSJ (2007). Leaf functional traits of tropical forest plants in relation to growth form Functional Ecology, 21, 19-27. [本文引用: 1]
SchnitzerSA (2005). A mechanistic explanation for global patterns of liana abundance and distribution The American Naturalist, 166, 262-276. DOI:10.1086/431250URL [本文引用: 1]
SchnitzerSA, BongersF (2011). Increasing liana abundance and biomass in tropical forests: emerging patterns and putative mechanisms Ecology Letters, 14, 397-406. DOI:10.1111/ele.2011.14.issue-4URL [本文引用: 1]
SchnitzerSA, DallingJW, CarsonWP (2000). The impact of lianas on tree regeneration in tropical forest canopy gaps: evidence for an alternative pathway of gap-phase regeneration Journal of Ecology, 88, 655-666. DOI:10.1046/j.1365-2745.2000.00489.xURL [本文引用: 1]
SchnitzerSA, van der HeijdenGMF (2019). Lianas have a seasonal growth advantage over co-occurring trees Ecology, 100, e02655. DOI: 10.1002/ecy.2655. DOI:10.1002/ecy.2019.100.issue-5URL [本文引用: 2]
SchultePJ, HinckleyTM (1985). A comparison of pressure- volume curve data analysis techniques Journal of Experimental Botany, 36, 1590-1602. DOI:10.1093/jxb/36.10.1590URL [本文引用: 1]
ScoffoniC, AlbuquerqueC, BrodersenCR, TownesSV, JohnGP, CochardH, BuckleyTN, McElroneAJ, SackL (2017b). Leaf vein xylem conduit diameter influences susceptibility to embolism and hydraulic decline New Phytologist, 213, 1076-1092. DOI:10.1111/nph.14256URL [本文引用: 1]
ScoffoniC, VuongC, DiepS, CochardH, SackL (2014). Leaf shrinkage with dehydration: coordination with hydraulic vulnerability and drought tolerance Plant Physiology, 164, 1772-1788. DOI:10.1104/pp.113.221424URL [本文引用: 1]
Smith-MartinCM, SkeltonRP, JohnsonKM, LucaniC, BrodribbTJ (2020). Lack of vulnerability segmentation among woody species in a diverse dry sclerophyll woodland community Functional Ecology, 34, 777-787. DOI:10.1111/fec.v34.4URL [本文引用: 1]
SperryJS, DonnellyJR, TyreeMT (1988). A method for measuring hydraulic conductivity and embolism in xylem Plant, Cell & Environment, 11, 35-40. [本文引用: 4]
SperryJS, SaliendraNZ (1994). Intra- and inter-plant variation in xylem cavitation in Betula occidentalis Plant, Cell & Environment, 17, 1233-1241. DOI:10.1105/tpc.104.030452URL [本文引用: 1]
TomlinsonPB, FisherJB (2005). Development of nonlignified fibers in leaves of Gnetum gnemon(Gnetales) American Journal of Botany, 92, 383-389. DOI:10.3732/ajb.92.3.383URL [本文引用: 1]
TrifilóP, RaimondoF, SaviT, Lo GulloMA, NardiniA (2016). The contribution of vascular and Extra-vascular water pathways to drought-induced decline of leaf hydraulic conductance Journal of Experimental Botany, 67, 5029-5039. DOI:10.1093/jxb/erw268URL [本文引用: 1]
van der SandeMT, PoorterL, SchnitzerSA, EngelbrechtBMJ, MarkesteijnL (2019). The hydraulic efficiency-safety trade-off differs between lianas and trees Ecology, 100, e02666. DOI: 10.1002/ecy.2666. DOI:10.1002/ecy.2019.100.issue-5URL [本文引用: 1]
VillagraM, CampanelloPI, BucciSJ, GoldsteinG (2013). Functional relationships between leaf hydraulics and leaf economic traits in response to nutrient addition in subtropical tree species Tree Physiology, 33, 1308-1318. DOI:10.1093/treephys/tpt098URL [本文引用: 1]
YinPX, CaiJ (2018). New possible mechanisms of embolism formation when measuring vulnerability curves by air injection in a pressure sleeve Plant, Cell & Environment, 41, 1361-1368. [本文引用: 2]
ZhangL, ChenYJ, MaKP, BongersF, SterckFJ (2019). Fully exposed canopy tree and liana branches in a tropical forest differ in mechanical traits but are similar in hydraulic traits Tree Physiology, 39, 1713-1724. DOI:10.1093/treephys/tpz070URL [本文引用: 1]
ZhuSD, CaoKF (2009). Hydraulic properties and photosynthetic rates in co-occurring lianas and trees in a seasonal tropical rainforest in southwestern China Plant Ecology, 204, 295-304. DOI:10.1007/s11258-009-9592-5URL [本文引用: 1]
ZhuSD, ChenYJ, CaoKF, YeQ (2015). Interspecific variation in branch and leaf traits among three Syzygium tree species from different successional tropical forests Functional Plant Biology, 42, 423-432. DOI:10.1071/FP14201URL [本文引用: 1]
ZhuSD, ChenYJ, FuPL, CaoKF (2017). Different hydraulic traits of woody plants from tropical forests with contrasting soil water availability Tree Physiology, 37, 1469-1477. DOI:10.1093/treephys/tpx094URL [本文引用: 2]
ZhuSD, LiuH, XuQY, CaoKF, YeQ (2016). Are leaves more vulnerable to cavitation than branches? Functional Ecology, 30, 1740-1744. DOI:10.1111/fec.2016.30.issue-11URL [本文引用: 3]
ZimmermanMH, BrownCL (1971). Trees: Structure and Function. Springer-Verlag, New York. [本文引用: 2]
A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests 1 2010
... 木质藤本植物是热带森林的重要组成部分(Schnitzer & Bongers, 2011).木质藤本具有木质化的茎, 但是不能直立生长, 通过攀附或缠绕其他植物(主要是乔木)的茎到达森林冠层, 由于投资到支撑结构的生物量较少, 木质藤本把较多的生物量配置到叶片和根系上(Rodríguez-Ronderos et al., 2016).木质藤本植物在水力结构和碳经济学方面与乔木显著不同(Angyalossy et al., 2014).与乔木相比, 木质藤本具有较高的比叶面积和光合速率(Cai et al., 2009), 具有发达且高效的维管系统(高的水力传导效率)为叶片运输水分和营养(Isnard & Feild, 2014), 但该结构导致木质部较低的抗栓塞能力(Isnard & Silk, 2009).基于木质藤本对季节性干旱的生理生态响应, Schnitzer (2005)提出了木质藤本植物的“旱季生长优势”假说, 即木质藤本在干旱季节的生长速率要显著高于其他直立树木.这可能与木质藤本具备有效的水力适应策略有关, 如发达的根系保证其在旱季获得深层土壤水分, 以及较强的气孔和渗透调节能力维持旱季光合作用并减少过度的水分损失(Maréchaux et al., 2017)等.长期的森林监测数据表明, 随着全球气候日益干热化对森林结构影响加剧(Allen et al., 2010), 木质藤本植物在热带季节性森林中的丰富度在增加, 将显著影响热带森林的群落结构和功能(Campbell et al., 2018), 这一现象印证了“旱季生长优势”假说的同时, 也使木质藤本的生理生态特征受到广泛的关注(Schnitzer & van der Heijden, 2019). ...
Liana anatomy: a broad perspective on structural evolution of the vascular system//Schnitzer SA, Bongers F, Burnham RJ, Putz FE 1 2014
... 木质藤本植物是热带森林的重要组成部分(Schnitzer & Bongers, 2011).木质藤本具有木质化的茎, 但是不能直立生长, 通过攀附或缠绕其他植物(主要是乔木)的茎到达森林冠层, 由于投资到支撑结构的生物量较少, 木质藤本把较多的生物量配置到叶片和根系上(Rodríguez-Ronderos et al., 2016).木质藤本植物在水力结构和碳经济学方面与乔木显著不同(Angyalossy et al., 2014).与乔木相比, 木质藤本具有较高的比叶面积和光合速率(Cai et al., 2009), 具有发达且高效的维管系统(高的水力传导效率)为叶片运输水分和营养(Isnard & Feild, 2014), 但该结构导致木质部较低的抗栓塞能力(Isnard & Silk, 2009).基于木质藤本对季节性干旱的生理生态响应, Schnitzer (2005)提出了木质藤本植物的“旱季生长优势”假说, 即木质藤本在干旱季节的生长速率要显著高于其他直立树木.这可能与木质藤本具备有效的水力适应策略有关, 如发达的根系保证其在旱季获得深层土壤水分, 以及较强的气孔和渗透调节能力维持旱季光合作用并减少过度的水分损失(Maréchaux et al., 2017)等.长期的森林监测数据表明, 随着全球气候日益干热化对森林结构影响加剧(Allen et al., 2010), 木质藤本植物在热带季节性森林中的丰富度在增加, 将显著影响热带森林的群落结构和功能(Campbell et al., 2018), 这一现象印证了“旱季生长优势”假说的同时, 也使木质藤本的生理生态特征受到广泛的关注(Schnitzer & van der Heijden, 2019). ...
On xylem hydraulic efficiencies, wood space-use and the safety-efficiency tradeoff 1 2016
... 植物枝条和叶片的水分传导有效性与安全性之间均没有显著的权衡关系(图3A、3B), 不支持第二个研究假设.对于枝条来说, 部分树种枝条木质部同时具有较低的水分传导效率和抗栓塞能力, 如生长在林下的思茅崖豆.木质部的水分传导效率主要决定于木质部导管特征, 而木质部水力安全是各种水力策略密切协同的结果, 如根系吸水、茎部储水能力和气孔调节(Mencuccini et al., 2015)等, 因此植物整株水平的水力抗旱机制可能会导致枝条水平上缺乏安全性和有效性的权衡关系(Meinzer et al., 2010).另外, 由于木质藤本木质部结构的特殊性(Ewers et al., 2015), 它们在木质部支撑和输导组织的分配上受到的限制较少, 在相似的抗栓塞能力的情况下, 可能存在更多的水分传导效率的变化(Bittencourt et al., 2016), 导致木质藤本不受水力权衡关系的约束(Zhu et al., 2017; Zhang et al., 2019). ...
Leaf hydraulic vulnerability is related to conduit dimensions and drought resistance across a diverse range of woody angiosperms 1 2010
... 叶片水分传导过程分为木质部途径与外木质部途径(Trifiló et al., 2016), 共同影响着叶片水分运输的有效性与安全性(Sack & Frole, 2006).叶片细胞特征(如原生质体大小和细胞壁厚度)与外木质部途径密切相关(Scoffoni et al., 2017a).叶片水容(Pivovaroff et al., 2016)与叶片膨压(Scoffoni et al., 2014)影响叶片的渗透调节, 在一定干旱条件下对水势的变化起缓冲作用, 能够影响叶片抗栓塞能力, 也影响了叶片的水分传导效率(Buckley et al., 2015).在木质部途径中, 叶脉导管壁厚度与导管大小及其比值, 以及叶脉密度(Blackman et al., 2010)等都会影响水分在导管内的传导效率, 同时各指标变化还会伴随着安全性的变化(Scoffoni et al., 2017b).外木质部途径和木质部途径的作用机制因不同叶片结构而变化(Sack & Holbrook, 2006), 因此影响叶片水分传导有效性与安全性的因素错综复杂, 而它们之间的权衡关系并不能简单地从叶片抗栓塞能力(P50leaf)与叶片最大水力导度(Kleaf-max)两个指标之间的线性关系来探讨, 叶片内各组织结构及其生理过程之间的关系需要更深入的研究. ...
Xylem function and growth rate interact to determine recovery rates after exposure to extreme water deficit 1 2010
... 植物的水力特征是探讨植物生理生态适应的核心内容(Choat et al., 2012).植物水力学性状主要包括: (1)水力传导能力(有效性), 如边材和叶片比导率(ks、kl), 它是衡量植物水分传导效率的指标(Sperry et al., 1988).水分传导效率与叶片的光合能力密切相关(Santiago et al., 2004), 并最终影响植物的生长速率(Brodribb et al., 2010).(2)抗栓塞能力(安全性), 用植物输导组织损失50%导水能力时的水势值(P50)表示, 与植物的干旱适应策略有关.植物水分传导有效性与安全性之间存在权衡关系, 是探讨植物的生长策略以及沿水分梯度分布等方面的重要的生理生态机制(Markesteijn et al., 2011).但是基于全球数据分析发现这种权衡关系较弱(Gleason et al., 2016; Santiago et al., 2018), 在水分胁迫不严重的热带森林中甚至没有权衡(van der Sande et al., 2019).植物不同器官的水力特征之间的差异或者协同关系是水力学研究的重要内容(Bucci et al., 2012).经典的脆弱性分段假说认为植物茎的抗栓塞能力要强于末端的叶片, 在水分胁迫状况下, 叶片更容易发生栓塞, 从而减少水分进一步损失, 保证了茎的水力安全(Zimmerman & Brown, 1971; Tyree & Ewers, 1991).因此, 叶片在植物整个水分运输通路中起到了“安全阀”的作用(McCulloh et al., 2014).但是, 也有部分研究发现热带亚热带森林的植物并不符合脆弱性分段假说: 叶片的抗栓塞能力与枝条相近(Villagra et al., 2013), 甚至要强于枝条(Zhu et al., 2015), 原因在于枝-叶的脆弱性分段与生境的干旱指数相关, 生长在湿润地区的植物具有稳定的水分来源, 环境选择压力小; 另外, 缺乏枝-叶脆弱性分段的植物具有丰富的水力补偿策略维持叶片的水分供给, 在潜在的干旱胁迫下可能具有生长优势(Zhu et al., 2016).基于水力性状可以分析植物的水分适应策略, 被广泛地用于解释植物一系列的生态学表现(de Guzman et al., 2017). ...
Stem hydraulic supply is linked to leaf photosynthetic capacity: evidence from New Caledonian and Tasmanian rainforests 1 2000
Hydraulic differences along the water transport system of South American Nothofagus species: Do leaves protect the stem functionality? 1 2012
... 植物的水力特征是探讨植物生理生态适应的核心内容(Choat et al., 2012).植物水力学性状主要包括: (1)水力传导能力(有效性), 如边材和叶片比导率(ks、kl), 它是衡量植物水分传导效率的指标(Sperry et al., 1988).水分传导效率与叶片的光合能力密切相关(Santiago et al., 2004), 并最终影响植物的生长速率(Brodribb et al., 2010).(2)抗栓塞能力(安全性), 用植物输导组织损失50%导水能力时的水势值(P50)表示, 与植物的干旱适应策略有关.植物水分传导有效性与安全性之间存在权衡关系, 是探讨植物的生长策略以及沿水分梯度分布等方面的重要的生理生态机制(Markesteijn et al., 2011).但是基于全球数据分析发现这种权衡关系较弱(Gleason et al., 2016; Santiago et al., 2018), 在水分胁迫不严重的热带森林中甚至没有权衡(van der Sande et al., 2019).植物不同器官的水力特征之间的差异或者协同关系是水力学研究的重要内容(Bucci et al., 2012).经典的脆弱性分段假说认为植物茎的抗栓塞能力要强于末端的叶片, 在水分胁迫状况下, 叶片更容易发生栓塞, 从而减少水分进一步损失, 保证了茎的水力安全(Zimmerman & Brown, 1971; Tyree & Ewers, 1991).因此, 叶片在植物整个水分运输通路中起到了“安全阀”的作用(McCulloh et al., 2014).但是, 也有部分研究发现热带亚热带森林的植物并不符合脆弱性分段假说: 叶片的抗栓塞能力与枝条相近(Villagra et al., 2013), 甚至要强于枝条(Zhu et al., 2015), 原因在于枝-叶的脆弱性分段与生境的干旱指数相关, 生长在湿润地区的植物具有稳定的水分来源, 环境选择压力小; 另外, 缺乏枝-叶脆弱性分段的植物具有丰富的水力补偿策略维持叶片的水分供给, 在潜在的干旱胁迫下可能具有生长优势(Zhu et al., 2016).基于水力性状可以分析植物的水分适应策略, 被广泛地用于解释植物一系列的生态学表现(de Guzman et al., 2017). ...
How does leaf anatomy influence water transport outside the xylem? 1 2015
... 叶片水分传导过程分为木质部途径与外木质部途径(Trifiló et al., 2016), 共同影响着叶片水分运输的有效性与安全性(Sack & Frole, 2006).叶片细胞特征(如原生质体大小和细胞壁厚度)与外木质部途径密切相关(Scoffoni et al., 2017a).叶片水容(Pivovaroff et al., 2016)与叶片膨压(Scoffoni et al., 2014)影响叶片的渗透调节, 在一定干旱条件下对水势的变化起缓冲作用, 能够影响叶片抗栓塞能力, 也影响了叶片的水分传导效率(Buckley et al., 2015).在木质部途径中, 叶脉导管壁厚度与导管大小及其比值, 以及叶脉密度(Blackman et al., 2010)等都会影响水分在导管内的传导效率, 同时各指标变化还会伴随着安全性的变化(Scoffoni et al., 2017b).外木质部途径和木质部途径的作用机制因不同叶片结构而变化(Sack & Holbrook, 2006), 因此影响叶片水分传导有效性与安全性的因素错综复杂, 而它们之间的权衡关系并不能简单地从叶片抗栓塞能力(P50leaf)与叶片最大水力导度(Kleaf-max)两个指标之间的线性关系来探讨, 叶片内各组织结构及其生理过程之间的关系需要更深入的研究. ...
Seasonal differences in leaf-level physiology give lianas a competitive advantage over trees in a tropical seasonal forest 1 2009
... 木质藤本植物是热带森林的重要组成部分(Schnitzer & Bongers, 2011).木质藤本具有木质化的茎, 但是不能直立生长, 通过攀附或缠绕其他植物(主要是乔木)的茎到达森林冠层, 由于投资到支撑结构的生物量较少, 木质藤本把较多的生物量配置到叶片和根系上(Rodríguez-Ronderos et al., 2016).木质藤本植物在水力结构和碳经济学方面与乔木显著不同(Angyalossy et al., 2014).与乔木相比, 木质藤本具有较高的比叶面积和光合速率(Cai et al., 2009), 具有发达且高效的维管系统(高的水力传导效率)为叶片运输水分和营养(Isnard & Feild, 2014), 但该结构导致木质部较低的抗栓塞能力(Isnard & Silk, 2009).基于木质藤本对季节性干旱的生理生态响应, Schnitzer (2005)提出了木质藤本植物的“旱季生长优势”假说, 即木质藤本在干旱季节的生长速率要显著高于其他直立树木.这可能与木质藤本具备有效的水力适应策略有关, 如发达的根系保证其在旱季获得深层土壤水分, 以及较强的气孔和渗透调节能力维持旱季光合作用并减少过度的水分损失(Maréchaux et al., 2017)等.长期的森林监测数据表明, 随着全球气候日益干热化对森林结构影响加剧(Allen et al., 2010), 木质藤本植物在热带季节性森林中的丰富度在增加, 将显著影响热带森林的群落结构和功能(Campbell et al., 2018), 这一现象印证了“旱季生长优势”假说的同时, 也使木质藤本的生理生态特征受到广泛的关注(Schnitzer & van der Heijden, 2019). ...
Edge disturbance drives liana abundance increase and alteration of Liana- host tree interactions in tropical forest fragments 1 2018
... 木质藤本植物是热带森林的重要组成部分(Schnitzer & Bongers, 2011).木质藤本具有木质化的茎, 但是不能直立生长, 通过攀附或缠绕其他植物(主要是乔木)的茎到达森林冠层, 由于投资到支撑结构的生物量较少, 木质藤本把较多的生物量配置到叶片和根系上(Rodríguez-Ronderos et al., 2016).木质藤本植物在水力结构和碳经济学方面与乔木显著不同(Angyalossy et al., 2014).与乔木相比, 木质藤本具有较高的比叶面积和光合速率(Cai et al., 2009), 具有发达且高效的维管系统(高的水力传导效率)为叶片运输水分和营养(Isnard & Feild, 2014), 但该结构导致木质部较低的抗栓塞能力(Isnard & Silk, 2009).基于木质藤本对季节性干旱的生理生态响应, Schnitzer (2005)提出了木质藤本植物的“旱季生长优势”假说, 即木质藤本在干旱季节的生长速率要显著高于其他直立树木.这可能与木质藤本具备有效的水力适应策略有关, 如发达的根系保证其在旱季获得深层土壤水分, 以及较强的气孔和渗透调节能力维持旱季光合作用并减少过度的水分损失(Maréchaux et al., 2017)等.长期的森林监测数据表明, 随着全球气候日益干热化对森林结构影响加剧(Allen et al., 2010), 木质藤本植物在热带季节性森林中的丰富度在增加, 将显著影响热带森林的群落结构和功能(Campbell et al., 2018), 这一现象印证了“旱季生长优势”假说的同时, 也使木质藤本的生理生态特征受到广泛的关注(Schnitzer & van der Heijden, 2019). ...
Acclimation of seedlings of Gnetum leyboldii Tul.(Gnetaceae) to light changes in a tropical rain forest 1 2013
... 买麻藤属植物是特殊的植物类群, 其生理生态适应机制引起广泛的关注(Kotowska et al., 2015).Feild和Balun (2008)对几种买麻藤植物的水力和光合相关性状进行了测定, 结果发现它们的水分传导效率和光合速率都很低.虽然买麻藤植物进化出与被子植物相似的导管系统和阔叶, 但是还没有进化出类似热带雨林需光种的典型特征, 如: 高繁殖速率与高光合速率, 因此常分布于热带森林林下阴生湿润生境中(Feild & Balun, 2008; Celis & Avalos, 2013).本研究中买麻藤属乔木树种灌状买麻藤与少苞买麻藤的测定结果符合这一结论.但是, 买麻藤属木质藤本小叶买麻藤则表现出很高的水分传导效率, 与阳生性的被子植物相当.这可能与其已进化出粗大的导管有关, 其平均水力导管直径为171 mm (未发表数据), 远高于热带森林植物的平均值(100 mm; Zhu et al., 2017).但其光合速率仍无法与阳生性的被子植物相比, 这可能与其进化史有关, 因为多功能基因的缺失导致的低光合速率可能是买麻藤属固有的特性(Deng et al., 2019). ...
Water-use advantage for lianas over trees in tropical seasonal forests 1 2015
... 本研究使用注气法测定木质部脆弱性曲线, 但是该方法的准确性受到质疑.在测量过程中, 当使用的茎段长度小于最大导管长度时产生的导管开口现象, 以及加压过程中形成的气泡效应, 会导致PLC被高估(抗栓塞能力被低估; Choat et al., 2010; Yin & Cai, 2018).另外, 加压的时间以及压力套的长度都会对曲线造成影响(Yin & Cai, 2018).由于本试验物种(部分为引种)可破坏性采集的样本量少, 无法选择较为精确但是枝条样本量需求较多的自然干燥法(Sperry et al., 1988).另外, 本研究尽量使用足够长的枝条以防止开口导管产生的影响(Ennajeh et al., 2011).已有研究利用自然干燥法测定了部分物种的P50branch, 如红河崖豆(Fu et al., 2012)、厚果崖豆藤(Zhu & Cao, 2009)和小叶买麻藤(Chen et al., 2015), 本研究测定的结果与之相似. ...
Measurement of vulnerability to water stress-induced cavitation in grapevine: a comparison of four techniques applied to a long-vesseled species 1 2010
... 本研究使用注气法测定木质部脆弱性曲线, 但是该方法的准确性受到质疑.在测量过程中, 当使用的茎段长度小于最大导管长度时产生的导管开口现象, 以及加压过程中形成的气泡效应, 会导致PLC被高估(抗栓塞能力被低估; Choat et al., 2010; Yin & Cai, 2018).另外, 加压的时间以及压力套的长度都会对曲线造成影响(Yin & Cai, 2018).由于本试验物种(部分为引种)可破坏性采集的样本量少, 无法选择较为精确但是枝条样本量需求较多的自然干燥法(Sperry et al., 1988).另外, 本研究尽量使用足够长的枝条以防止开口导管产生的影响(Ennajeh et al., 2011).已有研究利用自然干燥法测定了部分物种的P50branch, 如红河崖豆(Fu et al., 2012)、厚果崖豆藤(Zhu & Cao, 2009)和小叶买麻藤(Chen et al., 2015), 本研究测定的结果与之相似. ...
Global convergence in the vulnerability of forests to drought 1 2012
... 植物的水力特征是探讨植物生理生态适应的核心内容(Choat et al., 2012).植物水力学性状主要包括: (1)水力传导能力(有效性), 如边材和叶片比导率(ks、kl), 它是衡量植物水分传导效率的指标(Sperry et al., 1988).水分传导效率与叶片的光合能力密切相关(Santiago et al., 2004), 并最终影响植物的生长速率(Brodribb et al., 2010).(2)抗栓塞能力(安全性), 用植物输导组织损失50%导水能力时的水势值(P50)表示, 与植物的干旱适应策略有关.植物水分传导有效性与安全性之间存在权衡关系, 是探讨植物的生长策略以及沿水分梯度分布等方面的重要的生理生态机制(Markesteijn et al., 2011).但是基于全球数据分析发现这种权衡关系较弱(Gleason et al., 2016; Santiago et al., 2018), 在水分胁迫不严重的热带森林中甚至没有权衡(van der Sande et al., 2019).植物不同器官的水力特征之间的差异或者协同关系是水力学研究的重要内容(Bucci et al., 2012).经典的脆弱性分段假说认为植物茎的抗栓塞能力要强于末端的叶片, 在水分胁迫状况下, 叶片更容易发生栓塞, 从而减少水分进一步损失, 保证了茎的水力安全(Zimmerman & Brown, 1971; Tyree & Ewers, 1991).因此, 叶片在植物整个水分运输通路中起到了“安全阀”的作用(McCulloh et al., 2014).但是, 也有部分研究发现热带亚热带森林的植物并不符合脆弱性分段假说: 叶片的抗栓塞能力与枝条相近(Villagra et al., 2013), 甚至要强于枝条(Zhu et al., 2015), 原因在于枝-叶的脆弱性分段与生境的干旱指数相关, 生长在湿润地区的植物具有稳定的水分来源, 环境选择压力小; 另外, 缺乏枝-叶脆弱性分段的植物具有丰富的水力补偿策略维持叶片的水分供给, 在潜在的干旱胁迫下可能具有生长优势(Zhu et al., 2016).基于水力性状可以分析植物的水分适应策略, 被广泛地用于解释植物一系列的生态学表现(de Guzman et al., 2017). ...
Use of positive pressures to establish vulnerability curves: further support for the air-seeding hypothesis and implications for pressure- volume analysis 1 1992
Trade-offs between water transport capacity and drought resistance in neotropical canopy liana and tree species 1 2017
... 植物的水力特征是探讨植物生理生态适应的核心内容(Choat et al., 2012).植物水力学性状主要包括: (1)水力传导能力(有效性), 如边材和叶片比导率(ks、kl), 它是衡量植物水分传导效率的指标(Sperry et al., 1988).水分传导效率与叶片的光合能力密切相关(Santiago et al., 2004), 并最终影响植物的生长速率(Brodribb et al., 2010).(2)抗栓塞能力(安全性), 用植物输导组织损失50%导水能力时的水势值(P50)表示, 与植物的干旱适应策略有关.植物水分传导有效性与安全性之间存在权衡关系, 是探讨植物的生长策略以及沿水分梯度分布等方面的重要的生理生态机制(Markesteijn et al., 2011).但是基于全球数据分析发现这种权衡关系较弱(Gleason et al., 2016; Santiago et al., 2018), 在水分胁迫不严重的热带森林中甚至没有权衡(van der Sande et al., 2019).植物不同器官的水力特征之间的差异或者协同关系是水力学研究的重要内容(Bucci et al., 2012).经典的脆弱性分段假说认为植物茎的抗栓塞能力要强于末端的叶片, 在水分胁迫状况下, 叶片更容易发生栓塞, 从而减少水分进一步损失, 保证了茎的水力安全(Zimmerman & Brown, 1971; Tyree & Ewers, 1991).因此, 叶片在植物整个水分运输通路中起到了“安全阀”的作用(McCulloh et al., 2014).但是, 也有部分研究发现热带亚热带森林的植物并不符合脆弱性分段假说: 叶片的抗栓塞能力与枝条相近(Villagra et al., 2013), 甚至要强于枝条(Zhu et al., 2015), 原因在于枝-叶的脆弱性分段与生境的干旱指数相关, 生长在湿润地区的植物具有稳定的水分来源, 环境选择压力小; 另外, 缺乏枝-叶脆弱性分段的植物具有丰富的水力补偿策略维持叶片的水分供给, 在潜在的干旱胁迫下可能具有生长优势(Zhu et al., 2016).基于水力性状可以分析植物的水分适应策略, 被广泛地用于解释植物一系列的生态学表现(de Guzman et al., 2017). ...
Significance of photosynthetic characters in the evolution of Asian Gnetum(Gnetales) 1 2019
... 买麻藤属植物是特殊的植物类群, 其生理生态适应机制引起广泛的关注(Kotowska et al., 2015).Feild和Balun (2008)对几种买麻藤植物的水力和光合相关性状进行了测定, 结果发现它们的水分传导效率和光合速率都很低.虽然买麻藤植物进化出与被子植物相似的导管系统和阔叶, 但是还没有进化出类似热带雨林需光种的典型特征, 如: 高繁殖速率与高光合速率, 因此常分布于热带森林林下阴生湿润生境中(Feild & Balun, 2008; Celis & Avalos, 2013).本研究中买麻藤属乔木树种灌状买麻藤与少苞买麻藤的测定结果符合这一结论.但是, 买麻藤属木质藤本小叶买麻藤则表现出很高的水分传导效率, 与阳生性的被子植物相当.这可能与其已进化出粗大的导管有关, 其平均水力导管直径为171 mm (未发表数据), 远高于热带森林植物的平均值(100 mm; Zhu et al., 2017).但其光合速率仍无法与阳生性的被子植物相比, 这可能与其进化史有关, 因为多功能基因的缺失导致的低光合速率可能是买麻藤属固有的特性(Deng et al., 2019). ...
Seed plant phylogeny and the origin of angiosperms: an experimental cladistic approach 1 1986
... Native habitat descriptions of these species were extracted from Flora Reipublicae Popularis Sinicae (Editorial Committee of Flora of China, Chinese Academy of Sciences, 1993) and Li et al. (1996). ...
... Native habitat descriptions of these species were extracted from Flora Reipublicae Popularis Sinicae (Editorial Committee of Flora of China, Chinese Academy of Sciences, 1993) and Li et al. (1996). ...
Stem hydraulic traits and leaf water-stress tolerance are co-ordinated with the leaf phenology of angiosperm trees in an Asian tropical dry karst forest 1 2012
... 本研究使用注气法测定木质部脆弱性曲线, 但是该方法的准确性受到质疑.在测量过程中, 当使用的茎段长度小于最大导管长度时产生的导管开口现象, 以及加压过程中形成的气泡效应, 会导致PLC被高估(抗栓塞能力被低估; Choat et al., 2010; Yin & Cai, 2018).另外, 加压的时间以及压力套的长度都会对曲线造成影响(Yin & Cai, 2018).由于本试验物种(部分为引种)可破坏性采集的样本量少, 无法选择较为精确但是枝条样本量需求较多的自然干燥法(Sperry et al., 1988).另外, 本研究尽量使用足够长的枝条以防止开口导管产生的影响(Ennajeh et al., 2011).已有研究利用自然干燥法测定了部分物种的P50branch, 如红河崖豆(Fu et al., 2012)、厚果崖豆藤(Zhu & Cao, 2009)和小叶买麻藤(Chen et al., 2015), 本研究测定的结果与之相似. ...
Weak tradeoff between xylem safety and xylem-specific hydraulic efficiency across the world’s woody plant species 1 2016
... 植物的水力特征是探讨植物生理生态适应的核心内容(Choat et al., 2012).植物水力学性状主要包括: (1)水力传导能力(有效性), 如边材和叶片比导率(ks、kl), 它是衡量植物水分传导效率的指标(Sperry et al., 1988).水分传导效率与叶片的光合能力密切相关(Santiago et al., 2004), 并最终影响植物的生长速率(Brodribb et al., 2010).(2)抗栓塞能力(安全性), 用植物输导组织损失50%导水能力时的水势值(P50)表示, 与植物的干旱适应策略有关.植物水分传导有效性与安全性之间存在权衡关系, 是探讨植物的生长策略以及沿水分梯度分布等方面的重要的生理生态机制(Markesteijn et al., 2011).但是基于全球数据分析发现这种权衡关系较弱(Gleason et al., 2016; Santiago et al., 2018), 在水分胁迫不严重的热带森林中甚至没有权衡(van der Sande et al., 2019).植物不同器官的水力特征之间的差异或者协同关系是水力学研究的重要内容(Bucci et al., 2012).经典的脆弱性分段假说认为植物茎的抗栓塞能力要强于末端的叶片, 在水分胁迫状况下, 叶片更容易发生栓塞, 从而减少水分进一步损失, 保证了茎的水力安全(Zimmerman & Brown, 1971; Tyree & Ewers, 1991).因此, 叶片在植物整个水分运输通路中起到了“安全阀”的作用(McCulloh et al., 2014).但是, 也有部分研究发现热带亚热带森林的植物并不符合脆弱性分段假说: 叶片的抗栓塞能力与枝条相近(Villagra et al., 2013), 甚至要强于枝条(Zhu et al., 2015), 原因在于枝-叶的脆弱性分段与生境的干旱指数相关, 生长在湿润地区的植物具有稳定的水分来源, 环境选择压力小; 另外, 缺乏枝-叶脆弱性分段的植物具有丰富的水力补偿策略维持叶片的水分供给, 在潜在的干旱胁迫下可能具有生长优势(Zhu et al., 2016).基于水力性状可以分析植物的水分适应策略, 被广泛地用于解释植物一系列的生态学表现(de Guzman et al., 2017). ...
The evolution of angiosperm lianescence: a perspective from xylem structure-function //Schnitzer SA, Bongers F, Burnham RJ, Putz FE 1 2014
... 木质藤本植物是热带森林的重要组成部分(Schnitzer & Bongers, 2011).木质藤本具有木质化的茎, 但是不能直立生长, 通过攀附或缠绕其他植物(主要是乔木)的茎到达森林冠层, 由于投资到支撑结构的生物量较少, 木质藤本把较多的生物量配置到叶片和根系上(Rodríguez-Ronderos et al., 2016).木质藤本植物在水力结构和碳经济学方面与乔木显著不同(Angyalossy et al., 2014).与乔木相比, 木质藤本具有较高的比叶面积和光合速率(Cai et al., 2009), 具有发达且高效的维管系统(高的水力传导效率)为叶片运输水分和营养(Isnard & Feild, 2014), 但该结构导致木质部较低的抗栓塞能力(Isnard & Silk, 2009).基于木质藤本对季节性干旱的生理生态响应, Schnitzer (2005)提出了木质藤本植物的“旱季生长优势”假说, 即木质藤本在干旱季节的生长速率要显著高于其他直立树木.这可能与木质藤本具备有效的水力适应策略有关, 如发达的根系保证其在旱季获得深层土壤水分, 以及较强的气孔和渗透调节能力维持旱季光合作用并减少过度的水分损失(Maréchaux et al., 2017)等.长期的森林监测数据表明, 随着全球气候日益干热化对森林结构影响加剧(Allen et al., 2010), 木质藤本植物在热带季节性森林中的丰富度在增加, 将显著影响热带森林的群落结构和功能(Campbell et al., 2018), 这一现象印证了“旱季生长优势”假说的同时, 也使木质藤本的生理生态特征受到广泛的关注(Schnitzer & van der Heijden, 2019). ...
Moving with climbing plants from Charles Darwin’s time into the 21st century 1 2009
... 木质藤本植物是热带森林的重要组成部分(Schnitzer & Bongers, 2011).木质藤本具有木质化的茎, 但是不能直立生长, 通过攀附或缠绕其他植物(主要是乔木)的茎到达森林冠层, 由于投资到支撑结构的生物量较少, 木质藤本把较多的生物量配置到叶片和根系上(Rodríguez-Ronderos et al., 2016).木质藤本植物在水力结构和碳经济学方面与乔木显著不同(Angyalossy et al., 2014).与乔木相比, 木质藤本具有较高的比叶面积和光合速率(Cai et al., 2009), 具有发达且高效的维管系统(高的水力传导效率)为叶片运输水分和营养(Isnard & Feild, 2014), 但该结构导致木质部较低的抗栓塞能力(Isnard & Silk, 2009).基于木质藤本对季节性干旱的生理生态响应, Schnitzer (2005)提出了木质藤本植物的“旱季生长优势”假说, 即木质藤本在干旱季节的生长速率要显著高于其他直立树木.这可能与木质藤本具备有效的水力适应策略有关, 如发达的根系保证其在旱季获得深层土壤水分, 以及较强的气孔和渗透调节能力维持旱季光合作用并减少过度的水分损失(Maréchaux et al., 2017)等.长期的森林监测数据表明, 随着全球气候日益干热化对森林结构影响加剧(Allen et al., 2010), 木质藤本植物在热带季节性森林中的丰富度在增加, 将显著影响热带森林的群落结构和功能(Campbell et al., 2018), 这一现象印证了“旱季生长优势”假说的同时, 也使木质藤本的生理生态特征受到广泛的关注(Schnitzer & van der Heijden, 2019). ...
Patterns in hydraulic architecture from roots to branches in six tropical tree species from cacao agroforestry and their relation to wood density and stem growth 1 2015
... 买麻藤属植物是特殊的植物类群, 其生理生态适应机制引起广泛的关注(Kotowska et al., 2015).Feild和Balun (2008)对几种买麻藤植物的水力和光合相关性状进行了测定, 结果发现它们的水分传导效率和光合速率都很低.虽然买麻藤植物进化出与被子植物相似的导管系统和阔叶, 但是还没有进化出类似热带雨林需光种的典型特征, 如: 高繁殖速率与高光合速率, 因此常分布于热带森林林下阴生湿润生境中(Feild & Balun, 2008; Celis & Avalos, 2013).本研究中买麻藤属乔木树种灌状买麻藤与少苞买麻藤的测定结果符合这一结论.但是, 买麻藤属木质藤本小叶买麻藤则表现出很高的水分传导效率, 与阳生性的被子植物相当.这可能与其已进化出粗大的导管有关, 其平均水力导管直径为171 mm (未发表数据), 远高于热带森林植物的平均值(100 mm; Zhu et al., 2017).但其光合速率仍无法与阳生性的被子植物相比, 这可能与其进化史有关, 因为多功能基因的缺失导致的低光合速率可能是买麻藤属固有的特性(Deng et al., 2019). ...
4 1996
... Native habitat descriptions of these species were extracted from Flora Reipublicae Popularis Sinicae (Editorial Committee of Flora of China, Chinese Academy of Sciences, 1993) and Li et al. (1996). ...
... Native habitat descriptions of these species were extracted from Flora Reipublicae Popularis Sinicae (Editorial Committee of Flora of China, Chinese Academy of Sciences, 1993) and Li et al. (1996). ...
Stronger seasonal adjustment in leaf turgor loss point in lianas than trees in an Amazonian forest 1 2017
... 木质藤本植物是热带森林的重要组成部分(Schnitzer & Bongers, 2011).木质藤本具有木质化的茎, 但是不能直立生长, 通过攀附或缠绕其他植物(主要是乔木)的茎到达森林冠层, 由于投资到支撑结构的生物量较少, 木质藤本把较多的生物量配置到叶片和根系上(Rodríguez-Ronderos et al., 2016).木质藤本植物在水力结构和碳经济学方面与乔木显著不同(Angyalossy et al., 2014).与乔木相比, 木质藤本具有较高的比叶面积和光合速率(Cai et al., 2009), 具有发达且高效的维管系统(高的水力传导效率)为叶片运输水分和营养(Isnard & Feild, 2014), 但该结构导致木质部较低的抗栓塞能力(Isnard & Silk, 2009).基于木质藤本对季节性干旱的生理生态响应, Schnitzer (2005)提出了木质藤本植物的“旱季生长优势”假说, 即木质藤本在干旱季节的生长速率要显著高于其他直立树木.这可能与木质藤本具备有效的水力适应策略有关, 如发达的根系保证其在旱季获得深层土壤水分, 以及较强的气孔和渗透调节能力维持旱季光合作用并减少过度的水分损失(Maréchaux et al., 2017)等.长期的森林监测数据表明, 随着全球气候日益干热化对森林结构影响加剧(Allen et al., 2010), 木质藤本植物在热带季节性森林中的丰富度在增加, 将显著影响热带森林的群落结构和功能(Campbell et al., 2018), 这一现象印证了“旱季生长优势”假说的同时, 也使木质藤本的生理生态特征受到广泛的关注(Schnitzer & van der Heijden, 2019). ...
Hydraulics and life history of tropical dry forest tree species: coordination of species’ drought and shade tolerance 1 2011
... 植物的水力特征是探讨植物生理生态适应的核心内容(Choat et al., 2012).植物水力学性状主要包括: (1)水力传导能力(有效性), 如边材和叶片比导率(ks、kl), 它是衡量植物水分传导效率的指标(Sperry et al., 1988).水分传导效率与叶片的光合能力密切相关(Santiago et al., 2004), 并最终影响植物的生长速率(Brodribb et al., 2010).(2)抗栓塞能力(安全性), 用植物输导组织损失50%导水能力时的水势值(P50)表示, 与植物的干旱适应策略有关.植物水分传导有效性与安全性之间存在权衡关系, 是探讨植物的生长策略以及沿水分梯度分布等方面的重要的生理生态机制(Markesteijn et al., 2011).但是基于全球数据分析发现这种权衡关系较弱(Gleason et al., 2016; Santiago et al., 2018), 在水分胁迫不严重的热带森林中甚至没有权衡(van der Sande et al., 2019).植物不同器官的水力特征之间的差异或者协同关系是水力学研究的重要内容(Bucci et al., 2012).经典的脆弱性分段假说认为植物茎的抗栓塞能力要强于末端的叶片, 在水分胁迫状况下, 叶片更容易发生栓塞, 从而减少水分进一步损失, 保证了茎的水力安全(Zimmerman & Brown, 1971; Tyree & Ewers, 1991).因此, 叶片在植物整个水分运输通路中起到了“安全阀”的作用(McCulloh et al., 2014).但是, 也有部分研究发现热带亚热带森林的植物并不符合脆弱性分段假说: 叶片的抗栓塞能力与枝条相近(Villagra et al., 2013), 甚至要强于枝条(Zhu et al., 2015), 原因在于枝-叶的脆弱性分段与生境的干旱指数相关, 生长在湿润地区的植物具有稳定的水分来源, 环境选择压力小; 另外, 缺乏枝-叶脆弱性分段的植物具有丰富的水力补偿策略维持叶片的水分供给, 在潜在的干旱胁迫下可能具有生长优势(Zhu et al., 2016).基于水力性状可以分析植物的水分适应策略, 被广泛地用于解释植物一系列的生态学表现(de Guzman et al., 2017). ...
The dynamic pipeline: hydraulic capacitance and xylem hydraulic safety in four tall conifer species 1 2014
... 植物的水力特征是探讨植物生理生态适应的核心内容(Choat et al., 2012).植物水力学性状主要包括: (1)水力传导能力(有效性), 如边材和叶片比导率(ks、kl), 它是衡量植物水分传导效率的指标(Sperry et al., 1988).水分传导效率与叶片的光合能力密切相关(Santiago et al., 2004), 并最终影响植物的生长速率(Brodribb et al., 2010).(2)抗栓塞能力(安全性), 用植物输导组织损失50%导水能力时的水势值(P50)表示, 与植物的干旱适应策略有关.植物水分传导有效性与安全性之间存在权衡关系, 是探讨植物的生长策略以及沿水分梯度分布等方面的重要的生理生态机制(Markesteijn et al., 2011).但是基于全球数据分析发现这种权衡关系较弱(Gleason et al., 2016; Santiago et al., 2018), 在水分胁迫不严重的热带森林中甚至没有权衡(van der Sande et al., 2019).植物不同器官的水力特征之间的差异或者协同关系是水力学研究的重要内容(Bucci et al., 2012).经典的脆弱性分段假说认为植物茎的抗栓塞能力要强于末端的叶片, 在水分胁迫状况下, 叶片更容易发生栓塞, 从而减少水分进一步损失, 保证了茎的水力安全(Zimmerman & Brown, 1971; Tyree & Ewers, 1991).因此, 叶片在植物整个水分运输通路中起到了“安全阀”的作用(McCulloh et al., 2014).但是, 也有部分研究发现热带亚热带森林的植物并不符合脆弱性分段假说: 叶片的抗栓塞能力与枝条相近(Villagra et al., 2013), 甚至要强于枝条(Zhu et al., 2015), 原因在于枝-叶的脆弱性分段与生境的干旱指数相关, 生长在湿润地区的植物具有稳定的水分来源, 环境选择压力小; 另外, 缺乏枝-叶脆弱性分段的植物具有丰富的水力补偿策略维持叶片的水分供给, 在潜在的干旱胁迫下可能具有生长优势(Zhu et al., 2016).基于水力性状可以分析植物的水分适应策略, 被广泛地用于解释植物一系列的生态学表现(de Guzman et al., 2017). ...
The blind men and the elephant: the impact of context and scale in evaluating conflicts between plant hydraulic safety and efficiency 1 2010
... 植物枝条和叶片的水分传导有效性与安全性之间均没有显著的权衡关系(图3A、3B), 不支持第二个研究假设.对于枝条来说, 部分树种枝条木质部同时具有较低的水分传导效率和抗栓塞能力, 如生长在林下的思茅崖豆.木质部的水分传导效率主要决定于木质部导管特征, 而木质部水力安全是各种水力策略密切协同的结果, 如根系吸水、茎部储水能力和气孔调节(Mencuccini et al., 2015)等, 因此植物整株水平的水力抗旱机制可能会导致枝条水平上缺乏安全性和有效性的权衡关系(Meinzer et al., 2010).另外, 由于木质藤本木质部结构的特殊性(Ewers et al., 2015), 它们在木质部支撑和输导组织的分配上受到的限制较少, 在相似的抗栓塞能力的情况下, 可能存在更多的水分传导效率的变化(Bittencourt et al., 2016), 导致木质藤本不受水力权衡关系的约束(Zhu et al., 2017; Zhang et al., 2019). ...
Coordination of physiological traits involved in drought-induced mortality of woody plants 1 2015
... 植物枝条和叶片的水分传导有效性与安全性之间均没有显著的权衡关系(图3A、3B), 不支持第二个研究假设.对于枝条来说, 部分树种枝条木质部同时具有较低的水分传导效率和抗栓塞能力, 如生长在林下的思茅崖豆.木质部的水分传导效率主要决定于木质部导管特征, 而木质部水力安全是各种水力策略密切协同的结果, 如根系吸水、茎部储水能力和气孔调节(Mencuccini et al., 2015)等, 因此植物整株水平的水力抗旱机制可能会导致枝条水平上缺乏安全性和有效性的权衡关系(Meinzer et al., 2010).另外, 由于木质藤本木质部结构的特殊性(Ewers et al., 2015), 它们在木质部支撑和输导组织的分配上受到的限制较少, 在相似的抗栓塞能力的情况下, 可能存在更多的水分传导效率的变化(Bittencourt et al., 2016), 导致木质藤本不受水力权衡关系的约束(Zhu et al., 2017; Zhang et al., 2019). ...
A mathematical and statistical analysis of the curves illustrating vulnerability of xylem to cavitation 1 1998
... 在SigmaPlot 12.5中使用Sigmoid模型拟合PLC与所对应的压力值之间的关系(Pammenter & van der Willigen, 1998), 并计算PLC为50%时的木质部水势值(P50branch)用于表征枝条木质部的抗栓塞能力. ...
Multiple strategies for drought survival among woody plant species 1 2016
... 叶片水分传导过程分为木质部途径与外木质部途径(Trifiló et al., 2016), 共同影响着叶片水分运输的有效性与安全性(Sack & Frole, 2006).叶片细胞特征(如原生质体大小和细胞壁厚度)与外木质部途径密切相关(Scoffoni et al., 2017a).叶片水容(Pivovaroff et al., 2016)与叶片膨压(Scoffoni et al., 2014)影响叶片的渗透调节, 在一定干旱条件下对水势的变化起缓冲作用, 能够影响叶片抗栓塞能力, 也影响了叶片的水分传导效率(Buckley et al., 2015).在木质部途径中, 叶脉导管壁厚度与导管大小及其比值, 以及叶脉密度(Blackman et al., 2010)等都会影响水分在导管内的传导效率, 同时各指标变化还会伴随着安全性的变化(Scoffoni et al., 2017b).外木质部途径和木质部途径的作用机制因不同叶片结构而变化(Sack & Holbrook, 2006), 因此影响叶片水分传导有效性与安全性的因素错综复杂, 而它们之间的权衡关系并不能简单地从叶片抗栓塞能力(P50leaf)与叶片最大水力导度(Kleaf-max)两个指标之间的线性关系来探讨, 叶片内各组织结构及其生理过程之间的关系需要更深入的研究. ...
The natural history of lianas on barro Colorado Island, Panama 1 1984
Contribution of lianas to plant area index and canopy structure in a Panamanian forest 1 2016
... 木质藤本植物是热带森林的重要组成部分(Schnitzer & Bongers, 2011).木质藤本具有木质化的茎, 但是不能直立生长, 通过攀附或缠绕其他植物(主要是乔木)的茎到达森林冠层, 由于投资到支撑结构的生物量较少, 木质藤本把较多的生物量配置到叶片和根系上(Rodríguez-Ronderos et al., 2016).木质藤本植物在水力结构和碳经济学方面与乔木显著不同(Angyalossy et al., 2014).与乔木相比, 木质藤本具有较高的比叶面积和光合速率(Cai et al., 2009), 具有发达且高效的维管系统(高的水力传导效率)为叶片运输水分和营养(Isnard & Feild, 2014), 但该结构导致木质部较低的抗栓塞能力(Isnard & Silk, 2009).基于木质藤本对季节性干旱的生理生态响应, Schnitzer (2005)提出了木质藤本植物的“旱季生长优势”假说, 即木质藤本在干旱季节的生长速率要显著高于其他直立树木.这可能与木质藤本具备有效的水力适应策略有关, 如发达的根系保证其在旱季获得深层土壤水分, 以及较强的气孔和渗透调节能力维持旱季光合作用并减少过度的水分损失(Maréchaux et al., 2017)等.长期的森林监测数据表明, 随着全球气候日益干热化对森林结构影响加剧(Allen et al., 2010), 木质藤本植物在热带季节性森林中的丰富度在增加, 将显著影响热带森林的群落结构和功能(Campbell et al., 2018), 这一现象印证了“旱季生长优势”假说的同时, 也使木质藤本的生理生态特征受到广泛的关注(Schnitzer & van der Heijden, 2019). ...
Leaf structural diversity is related to hydraulic capacity in tropical rain forest trees 1 2006
... 叶片水分传导过程分为木质部途径与外木质部途径(Trifiló et al., 2016), 共同影响着叶片水分运输的有效性与安全性(Sack & Frole, 2006).叶片细胞特征(如原生质体大小和细胞壁厚度)与外木质部途径密切相关(Scoffoni et al., 2017a).叶片水容(Pivovaroff et al., 2016)与叶片膨压(Scoffoni et al., 2014)影响叶片的渗透调节, 在一定干旱条件下对水势的变化起缓冲作用, 能够影响叶片抗栓塞能力, 也影响了叶片的水分传导效率(Buckley et al., 2015).在木质部途径中, 叶脉导管壁厚度与导管大小及其比值, 以及叶脉密度(Blackman et al., 2010)等都会影响水分在导管内的传导效率, 同时各指标变化还会伴随着安全性的变化(Scoffoni et al., 2017b).外木质部途径和木质部途径的作用机制因不同叶片结构而变化(Sack & Holbrook, 2006), 因此影响叶片水分传导有效性与安全性的因素错综复杂, 而它们之间的权衡关系并不能简单地从叶片抗栓塞能力(P50leaf)与叶片最大水力导度(Kleaf-max)两个指标之间的线性关系来探讨, 叶片内各组织结构及其生理过程之间的关系需要更深入的研究. ...
Leaf hydraulics 1 2006
... 叶片水分传导过程分为木质部途径与外木质部途径(Trifiló et al., 2016), 共同影响着叶片水分运输的有效性与安全性(Sack & Frole, 2006).叶片细胞特征(如原生质体大小和细胞壁厚度)与外木质部途径密切相关(Scoffoni et al., 2017a).叶片水容(Pivovaroff et al., 2016)与叶片膨压(Scoffoni et al., 2014)影响叶片的渗透调节, 在一定干旱条件下对水势的变化起缓冲作用, 能够影响叶片抗栓塞能力, 也影响了叶片的水分传导效率(Buckley et al., 2015).在木质部途径中, 叶脉导管壁厚度与导管大小及其比值, 以及叶脉密度(Blackman et al., 2010)等都会影响水分在导管内的传导效率, 同时各指标变化还会伴随着安全性的变化(Scoffoni et al., 2017b).外木质部途径和木质部途径的作用机制因不同叶片结构而变化(Sack & Holbrook, 2006), 因此影响叶片水分传导有效性与安全性的因素错综复杂, 而它们之间的权衡关系并不能简单地从叶片抗栓塞能力(P50leaf)与叶片最大水力导度(Kleaf-max)两个指标之间的线性关系来探讨, 叶片内各组织结构及其生理过程之间的关系需要更深入的研究. ...
Coordination and trade-offs among hydraulic safety, efficiency and drought avoidance traits in Amazonian rainforest canopy tree species 1 2018
... 植物的水力特征是探讨植物生理生态适应的核心内容(Choat et al., 2012).植物水力学性状主要包括: (1)水力传导能力(有效性), 如边材和叶片比导率(ks、kl), 它是衡量植物水分传导效率的指标(Sperry et al., 1988).水分传导效率与叶片的光合能力密切相关(Santiago et al., 2004), 并最终影响植物的生长速率(Brodribb et al., 2010).(2)抗栓塞能力(安全性), 用植物输导组织损失50%导水能力时的水势值(P50)表示, 与植物的干旱适应策略有关.植物水分传导有效性与安全性之间存在权衡关系, 是探讨植物的生长策略以及沿水分梯度分布等方面的重要的生理生态机制(Markesteijn et al., 2011).但是基于全球数据分析发现这种权衡关系较弱(Gleason et al., 2016; Santiago et al., 2018), 在水分胁迫不严重的热带森林中甚至没有权衡(van der Sande et al., 2019).植物不同器官的水力特征之间的差异或者协同关系是水力学研究的重要内容(Bucci et al., 2012).经典的脆弱性分段假说认为植物茎的抗栓塞能力要强于末端的叶片, 在水分胁迫状况下, 叶片更容易发生栓塞, 从而减少水分进一步损失, 保证了茎的水力安全(Zimmerman & Brown, 1971; Tyree & Ewers, 1991).因此, 叶片在植物整个水分运输通路中起到了“安全阀”的作用(McCulloh et al., 2014).但是, 也有部分研究发现热带亚热带森林的植物并不符合脆弱性分段假说: 叶片的抗栓塞能力与枝条相近(Villagra et al., 2013), 甚至要强于枝条(Zhu et al., 2015), 原因在于枝-叶的脆弱性分段与生境的干旱指数相关, 生长在湿润地区的植物具有稳定的水分来源, 环境选择压力小; 另外, 缺乏枝-叶脆弱性分段的植物具有丰富的水力补偿策略维持叶片的水分供给, 在潜在的干旱胁迫下可能具有生长优势(Zhu et al., 2016).基于水力性状可以分析植物的水分适应策略, 被广泛地用于解释植物一系列的生态学表现(de Guzman et al., 2017). ...
Leaf photosynthetic traits scale with hydraulic conductivity and wood density in Panamanian forest canopy trees 1 2004
... 植物的水力特征是探讨植物生理生态适应的核心内容(Choat et al., 2012).植物水力学性状主要包括: (1)水力传导能力(有效性), 如边材和叶片比导率(ks、kl), 它是衡量植物水分传导效率的指标(Sperry et al., 1988).水分传导效率与叶片的光合能力密切相关(Santiago et al., 2004), 并最终影响植物的生长速率(Brodribb et al., 2010).(2)抗栓塞能力(安全性), 用植物输导组织损失50%导水能力时的水势值(P50)表示, 与植物的干旱适应策略有关.植物水分传导有效性与安全性之间存在权衡关系, 是探讨植物的生长策略以及沿水分梯度分布等方面的重要的生理生态机制(Markesteijn et al., 2011).但是基于全球数据分析发现这种权衡关系较弱(Gleason et al., 2016; Santiago et al., 2018), 在水分胁迫不严重的热带森林中甚至没有权衡(van der Sande et al., 2019).植物不同器官的水力特征之间的差异或者协同关系是水力学研究的重要内容(Bucci et al., 2012).经典的脆弱性分段假说认为植物茎的抗栓塞能力要强于末端的叶片, 在水分胁迫状况下, 叶片更容易发生栓塞, 从而减少水分进一步损失, 保证了茎的水力安全(Zimmerman & Brown, 1971; Tyree & Ewers, 1991).因此, 叶片在植物整个水分运输通路中起到了“安全阀”的作用(McCulloh et al., 2014).但是, 也有部分研究发现热带亚热带森林的植物并不符合脆弱性分段假说: 叶片的抗栓塞能力与枝条相近(Villagra et al., 2013), 甚至要强于枝条(Zhu et al., 2015), 原因在于枝-叶的脆弱性分段与生境的干旱指数相关, 生长在湿润地区的植物具有稳定的水分来源, 环境选择压力小; 另外, 缺乏枝-叶脆弱性分段的植物具有丰富的水力补偿策略维持叶片的水分供给, 在潜在的干旱胁迫下可能具有生长优势(Zhu et al., 2016).基于水力性状可以分析植物的水分适应策略, 被广泛地用于解释植物一系列的生态学表现(de Guzman et al., 2017). ...
Physiological implications of the liana growth form//Schnitzer SA, Bongers F, Burnham RJ, Putz FE. Ecology of Lianas. John Wiley & Sons, 1 2014
Outside-xylem vulnerability, not xylem embolism, controls leaf hydraulic decline during dehydration 1 2017
... 叶片水分传导过程分为木质部途径与外木质部途径(Trifiló et al., 2016), 共同影响着叶片水分运输的有效性与安全性(Sack & Frole, 2006).叶片细胞特征(如原生质体大小和细胞壁厚度)与外木质部途径密切相关(Scoffoni et al., 2017a).叶片水容(Pivovaroff et al., 2016)与叶片膨压(Scoffoni et al., 2014)影响叶片的渗透调节, 在一定干旱条件下对水势的变化起缓冲作用, 能够影响叶片抗栓塞能力, 也影响了叶片的水分传导效率(Buckley et al., 2015).在木质部途径中, 叶脉导管壁厚度与导管大小及其比值, 以及叶脉密度(Blackman et al., 2010)等都会影响水分在导管内的传导效率, 同时各指标变化还会伴随着安全性的变化(Scoffoni et al., 2017b).外木质部途径和木质部途径的作用机制因不同叶片结构而变化(Sack & Holbrook, 2006), 因此影响叶片水分传导有效性与安全性的因素错综复杂, 而它们之间的权衡关系并不能简单地从叶片抗栓塞能力(P50leaf)与叶片最大水力导度(Kleaf-max)两个指标之间的线性关系来探讨, 叶片内各组织结构及其生理过程之间的关系需要更深入的研究. ...
Leaf vein xylem conduit diameter influences susceptibility to embolism and hydraulic decline 1 2017
... 叶片水分传导过程分为木质部途径与外木质部途径(Trifiló et al., 2016), 共同影响着叶片水分运输的有效性与安全性(Sack & Frole, 2006).叶片细胞特征(如原生质体大小和细胞壁厚度)与外木质部途径密切相关(Scoffoni et al., 2017a).叶片水容(Pivovaroff et al., 2016)与叶片膨压(Scoffoni et al., 2014)影响叶片的渗透调节, 在一定干旱条件下对水势的变化起缓冲作用, 能够影响叶片抗栓塞能力, 也影响了叶片的水分传导效率(Buckley et al., 2015).在木质部途径中, 叶脉导管壁厚度与导管大小及其比值, 以及叶脉密度(Blackman et al., 2010)等都会影响水分在导管内的传导效率, 同时各指标变化还会伴随着安全性的变化(Scoffoni et al., 2017b).外木质部途径和木质部途径的作用机制因不同叶片结构而变化(Sack & Holbrook, 2006), 因此影响叶片水分传导有效性与安全性的因素错综复杂, 而它们之间的权衡关系并不能简单地从叶片抗栓塞能力(P50leaf)与叶片最大水力导度(Kleaf-max)两个指标之间的线性关系来探讨, 叶片内各组织结构及其生理过程之间的关系需要更深入的研究. ...
Leaf shrinkage with dehydration: coordination with hydraulic vulnerability and drought tolerance 1 2014
... 叶片水分传导过程分为木质部途径与外木质部途径(Trifiló et al., 2016), 共同影响着叶片水分运输的有效性与安全性(Sack & Frole, 2006).叶片细胞特征(如原生质体大小和细胞壁厚度)与外木质部途径密切相关(Scoffoni et al., 2017a).叶片水容(Pivovaroff et al., 2016)与叶片膨压(Scoffoni et al., 2014)影响叶片的渗透调节, 在一定干旱条件下对水势的变化起缓冲作用, 能够影响叶片抗栓塞能力, 也影响了叶片的水分传导效率(Buckley et al., 2015).在木质部途径中, 叶脉导管壁厚度与导管大小及其比值, 以及叶脉密度(Blackman et al., 2010)等都会影响水分在导管内的传导效率, 同时各指标变化还会伴随着安全性的变化(Scoffoni et al., 2017b).外木质部途径和木质部途径的作用机制因不同叶片结构而变化(Sack & Holbrook, 2006), 因此影响叶片水分传导有效性与安全性的因素错综复杂, 而它们之间的权衡关系并不能简单地从叶片抗栓塞能力(P50leaf)与叶片最大水力导度(Kleaf-max)两个指标之间的线性关系来探讨, 叶片内各组织结构及其生理过程之间的关系需要更深入的研究. ...
Lack of vulnerability segmentation among woody species in a diverse dry sclerophyll woodland community 1 2020
... 与第三个研究假设相符, 两个属的木质藤本植物均不符合传统的枝-叶脆弱性分段假说, 即叶片比枝条具有更强的抗栓塞能力(图4C).枝-叶脆弱性分段不存在的原因可能与本研究测定脆弱性曲线的方法有关(可能低估了枝条的抗栓塞能力), 但是最近的研究采用相同的方法(光学法)测定枝条和叶片的脆弱性曲线, 同样发现很多树种不符合传统的 枝-叶脆弱性分段(Smith-Martin et al., 2020).另外, 前期研究表明这些植物在增强叶片抗栓塞能力的同时, 拥有有效的水力策略以保证茎的水力安全, 在干旱胁迫的情况下可能会具有生理优势: 能较大限度地推迟关闭气孔, 从而保持光合作用和生长(Zhu et al., 2016).本研究发现枝-叶脆弱性分段与旱季最大净光合速率和最大气孔导度呈显著负相关关系, 直接支持Zhu等(2016)的研究论断.另外, 由于木质藤本具有比同属乔木更强的旱季光合能力, 也支持了木质藤本植物的旱季优势生长假说(Zimmerman & Brown, 1971; Tyree & Ewers, 1991; Schnitzer & van der Heijden, 2019). ...
A method for measuring hydraulic conductivity and embolism in xylem 4 1988
... 植物的水力特征是探讨植物生理生态适应的核心内容(Choat et al., 2012).植物水力学性状主要包括: (1)水力传导能力(有效性), 如边材和叶片比导率(ks、kl), 它是衡量植物水分传导效率的指标(Sperry et al., 1988).水分传导效率与叶片的光合能力密切相关(Santiago et al., 2004), 并最终影响植物的生长速率(Brodribb et al., 2010).(2)抗栓塞能力(安全性), 用植物输导组织损失50%导水能力时的水势值(P50)表示, 与植物的干旱适应策略有关.植物水分传导有效性与安全性之间存在权衡关系, 是探讨植物的生长策略以及沿水分梯度分布等方面的重要的生理生态机制(Markesteijn et al., 2011).但是基于全球数据分析发现这种权衡关系较弱(Gleason et al., 2016; Santiago et al., 2018), 在水分胁迫不严重的热带森林中甚至没有权衡(van der Sande et al., 2019).植物不同器官的水力特征之间的差异或者协同关系是水力学研究的重要内容(Bucci et al., 2012).经典的脆弱性分段假说认为植物茎的抗栓塞能力要强于末端的叶片, 在水分胁迫状况下, 叶片更容易发生栓塞, 从而减少水分进一步损失, 保证了茎的水力安全(Zimmerman & Brown, 1971; Tyree & Ewers, 1991).因此, 叶片在植物整个水分运输通路中起到了“安全阀”的作用(McCulloh et al., 2014).但是, 也有部分研究发现热带亚热带森林的植物并不符合脆弱性分段假说: 叶片的抗栓塞能力与枝条相近(Villagra et al., 2013), 甚至要强于枝条(Zhu et al., 2015), 原因在于枝-叶的脆弱性分段与生境的干旱指数相关, 生长在湿润地区的植物具有稳定的水分来源, 环境选择压力小; 另外, 缺乏枝-叶脆弱性分段的植物具有丰富的水力补偿策略维持叶片的水分供给, 在潜在的干旱胁迫下可能具有生长优势(Zhu et al., 2016).基于水力性状可以分析植物的水分适应策略, 被广泛地用于解释植物一系列的生态学表现(de Guzman et al., 2017). ...
The contribution of vascular and Extra-vascular water pathways to drought-induced decline of leaf hydraulic conductance 1 2016
... 叶片水分传导过程分为木质部途径与外木质部途径(Trifiló et al., 2016), 共同影响着叶片水分运输的有效性与安全性(Sack & Frole, 2006).叶片细胞特征(如原生质体大小和细胞壁厚度)与外木质部途径密切相关(Scoffoni et al., 2017a).叶片水容(Pivovaroff et al., 2016)与叶片膨压(Scoffoni et al., 2014)影响叶片的渗透调节, 在一定干旱条件下对水势的变化起缓冲作用, 能够影响叶片抗栓塞能力, 也影响了叶片的水分传导效率(Buckley et al., 2015).在木质部途径中, 叶脉导管壁厚度与导管大小及其比值, 以及叶脉密度(Blackman et al., 2010)等都会影响水分在导管内的传导效率, 同时各指标变化还会伴随着安全性的变化(Scoffoni et al., 2017b).外木质部途径和木质部途径的作用机制因不同叶片结构而变化(Sack & Holbrook, 2006), 因此影响叶片水分传导有效性与安全性的因素错综复杂, 而它们之间的权衡关系并不能简单地从叶片抗栓塞能力(P50leaf)与叶片最大水力导度(Kleaf-max)两个指标之间的线性关系来探讨, 叶片内各组织结构及其生理过程之间的关系需要更深入的研究. ...
The hydraulic architecture of trees and other woody plants 2 1991
... 植物的水力特征是探讨植物生理生态适应的核心内容(Choat et al., 2012).植物水力学性状主要包括: (1)水力传导能力(有效性), 如边材和叶片比导率(ks、kl), 它是衡量植物水分传导效率的指标(Sperry et al., 1988).水分传导效率与叶片的光合能力密切相关(Santiago et al., 2004), 并最终影响植物的生长速率(Brodribb et al., 2010).(2)抗栓塞能力(安全性), 用植物输导组织损失50%导水能力时的水势值(P50)表示, 与植物的干旱适应策略有关.植物水分传导有效性与安全性之间存在权衡关系, 是探讨植物的生长策略以及沿水分梯度分布等方面的重要的生理生态机制(Markesteijn et al., 2011).但是基于全球数据分析发现这种权衡关系较弱(Gleason et al., 2016; Santiago et al., 2018), 在水分胁迫不严重的热带森林中甚至没有权衡(van der Sande et al., 2019).植物不同器官的水力特征之间的差异或者协同关系是水力学研究的重要内容(Bucci et al., 2012).经典的脆弱性分段假说认为植物茎的抗栓塞能力要强于末端的叶片, 在水分胁迫状况下, 叶片更容易发生栓塞, 从而减少水分进一步损失, 保证了茎的水力安全(Zimmerman & Brown, 1971; Tyree & Ewers, 1991).因此, 叶片在植物整个水分运输通路中起到了“安全阀”的作用(McCulloh et al., 2014).但是, 也有部分研究发现热带亚热带森林的植物并不符合脆弱性分段假说: 叶片的抗栓塞能力与枝条相近(Villagra et al., 2013), 甚至要强于枝条(Zhu et al., 2015), 原因在于枝-叶的脆弱性分段与生境的干旱指数相关, 生长在湿润地区的植物具有稳定的水分来源, 环境选择压力小; 另外, 缺乏枝-叶脆弱性分段的植物具有丰富的水力补偿策略维持叶片的水分供给, 在潜在的干旱胁迫下可能具有生长优势(Zhu et al., 2016).基于水力性状可以分析植物的水分适应策略, 被广泛地用于解释植物一系列的生态学表现(de Guzman et al., 2017). ...
... 与第三个研究假设相符, 两个属的木质藤本植物均不符合传统的枝-叶脆弱性分段假说, 即叶片比枝条具有更强的抗栓塞能力(图4C).枝-叶脆弱性分段不存在的原因可能与本研究测定脆弱性曲线的方法有关(可能低估了枝条的抗栓塞能力), 但是最近的研究采用相同的方法(光学法)测定枝条和叶片的脆弱性曲线, 同样发现很多树种不符合传统的 枝-叶脆弱性分段(Smith-Martin et al., 2020).另外, 前期研究表明这些植物在增强叶片抗栓塞能力的同时, 拥有有效的水力策略以保证茎的水力安全, 在干旱胁迫的情况下可能会具有生理优势: 能较大限度地推迟关闭气孔, 从而保持光合作用和生长(Zhu et al., 2016).本研究发现枝-叶脆弱性分段与旱季最大净光合速率和最大气孔导度呈显著负相关关系, 直接支持Zhu等(2016)的研究论断.另外, 由于木质藤本具有比同属乔木更强的旱季光合能力, 也支持了木质藤本植物的旱季优势生长假说(Zimmerman & Brown, 1971; Tyree & Ewers, 1991; Schnitzer & van der Heijden, 2019). ...
The hydraulic efficiency-safety trade-off differs between lianas and trees 1 2019
... 植物的水力特征是探讨植物生理生态适应的核心内容(Choat et al., 2012).植物水力学性状主要包括: (1)水力传导能力(有效性), 如边材和叶片比导率(ks、kl), 它是衡量植物水分传导效率的指标(Sperry et al., 1988).水分传导效率与叶片的光合能力密切相关(Santiago et al., 2004), 并最终影响植物的生长速率(Brodribb et al., 2010).(2)抗栓塞能力(安全性), 用植物输导组织损失50%导水能力时的水势值(P50)表示, 与植物的干旱适应策略有关.植物水分传导有效性与安全性之间存在权衡关系, 是探讨植物的生长策略以及沿水分梯度分布等方面的重要的生理生态机制(Markesteijn et al., 2011).但是基于全球数据分析发现这种权衡关系较弱(Gleason et al., 2016; Santiago et al., 2018), 在水分胁迫不严重的热带森林中甚至没有权衡(van der Sande et al., 2019).植物不同器官的水力特征之间的差异或者协同关系是水力学研究的重要内容(Bucci et al., 2012).经典的脆弱性分段假说认为植物茎的抗栓塞能力要强于末端的叶片, 在水分胁迫状况下, 叶片更容易发生栓塞, 从而减少水分进一步损失, 保证了茎的水力安全(Zimmerman & Brown, 1971; Tyree & Ewers, 1991).因此, 叶片在植物整个水分运输通路中起到了“安全阀”的作用(McCulloh et al., 2014).但是, 也有部分研究发现热带亚热带森林的植物并不符合脆弱性分段假说: 叶片的抗栓塞能力与枝条相近(Villagra et al., 2013), 甚至要强于枝条(Zhu et al., 2015), 原因在于枝-叶的脆弱性分段与生境的干旱指数相关, 生长在湿润地区的植物具有稳定的水分来源, 环境选择压力小; 另外, 缺乏枝-叶脆弱性分段的植物具有丰富的水力补偿策略维持叶片的水分供给, 在潜在的干旱胁迫下可能具有生长优势(Zhu et al., 2016).基于水力性状可以分析植物的水分适应策略, 被广泛地用于解释植物一系列的生态学表现(de Guzman et al., 2017). ...
Functional relationships between leaf hydraulics and leaf economic traits in response to nutrient addition in subtropical tree species 1 2013
... 植物的水力特征是探讨植物生理生态适应的核心内容(Choat et al., 2012).植物水力学性状主要包括: (1)水力传导能力(有效性), 如边材和叶片比导率(ks、kl), 它是衡量植物水分传导效率的指标(Sperry et al., 1988).水分传导效率与叶片的光合能力密切相关(Santiago et al., 2004), 并最终影响植物的生长速率(Brodribb et al., 2010).(2)抗栓塞能力(安全性), 用植物输导组织损失50%导水能力时的水势值(P50)表示, 与植物的干旱适应策略有关.植物水分传导有效性与安全性之间存在权衡关系, 是探讨植物的生长策略以及沿水分梯度分布等方面的重要的生理生态机制(Markesteijn et al., 2011).但是基于全球数据分析发现这种权衡关系较弱(Gleason et al., 2016; Santiago et al., 2018), 在水分胁迫不严重的热带森林中甚至没有权衡(van der Sande et al., 2019).植物不同器官的水力特征之间的差异或者协同关系是水力学研究的重要内容(Bucci et al., 2012).经典的脆弱性分段假说认为植物茎的抗栓塞能力要强于末端的叶片, 在水分胁迫状况下, 叶片更容易发生栓塞, 从而减少水分进一步损失, 保证了茎的水力安全(Zimmerman & Brown, 1971; Tyree & Ewers, 1991).因此, 叶片在植物整个水分运输通路中起到了“安全阀”的作用(McCulloh et al., 2014).但是, 也有部分研究发现热带亚热带森林的植物并不符合脆弱性分段假说: 叶片的抗栓塞能力与枝条相近(Villagra et al., 2013), 甚至要强于枝条(Zhu et al., 2015), 原因在于枝-叶的脆弱性分段与生境的干旱指数相关, 生长在湿润地区的植物具有稳定的水分来源, 环境选择压力小; 另外, 缺乏枝-叶脆弱性分段的植物具有丰富的水力补偿策略维持叶片的水分供给, 在潜在的干旱胁迫下可能具有生长优势(Zhu et al., 2016).基于水力性状可以分析植物的水分适应策略, 被广泛地用于解释植物一系列的生态学表现(de Guzman et al., 2017). ...
New possible mechanisms of embolism formation when measuring vulnerability curves by air injection in a pressure sleeve 2 2018
... 本研究使用注气法测定木质部脆弱性曲线, 但是该方法的准确性受到质疑.在测量过程中, 当使用的茎段长度小于最大导管长度时产生的导管开口现象, 以及加压过程中形成的气泡效应, 会导致PLC被高估(抗栓塞能力被低估; Choat et al., 2010; Yin & Cai, 2018).另外, 加压的时间以及压力套的长度都会对曲线造成影响(Yin & Cai, 2018).由于本试验物种(部分为引种)可破坏性采集的样本量少, 无法选择较为精确但是枝条样本量需求较多的自然干燥法(Sperry et al., 1988).另外, 本研究尽量使用足够长的枝条以防止开口导管产生的影响(Ennajeh et al., 2011).已有研究利用自然干燥法测定了部分物种的P50branch, 如红河崖豆(Fu et al., 2012)、厚果崖豆藤(Zhu & Cao, 2009)和小叶买麻藤(Chen et al., 2015), 本研究测定的结果与之相似. ...
... ).另外, 加压的时间以及压力套的长度都会对曲线造成影响(Yin & Cai, 2018).由于本试验物种(部分为引种)可破坏性采集的样本量少, 无法选择较为精确但是枝条样本量需求较多的自然干燥法(Sperry et al., 1988).另外, 本研究尽量使用足够长的枝条以防止开口导管产生的影响(Ennajeh et al., 2011).已有研究利用自然干燥法测定了部分物种的P50branch, 如红河崖豆(Fu et al., 2012)、厚果崖豆藤(Zhu & Cao, 2009)和小叶买麻藤(Chen et al., 2015), 本研究测定的结果与之相似. ...
Fully exposed canopy tree and liana branches in a tropical forest differ in mechanical traits but are similar in hydraulic traits 1 2019
... 植物枝条和叶片的水分传导有效性与安全性之间均没有显著的权衡关系(图3A、3B), 不支持第二个研究假设.对于枝条来说, 部分树种枝条木质部同时具有较低的水分传导效率和抗栓塞能力, 如生长在林下的思茅崖豆.木质部的水分传导效率主要决定于木质部导管特征, 而木质部水力安全是各种水力策略密切协同的结果, 如根系吸水、茎部储水能力和气孔调节(Mencuccini et al., 2015)等, 因此植物整株水平的水力抗旱机制可能会导致枝条水平上缺乏安全性和有效性的权衡关系(Meinzer et al., 2010).另外, 由于木质藤本木质部结构的特殊性(Ewers et al., 2015), 它们在木质部支撑和输导组织的分配上受到的限制较少, 在相似的抗栓塞能力的情况下, 可能存在更多的水分传导效率的变化(Bittencourt et al., 2016), 导致木质藤本不受水力权衡关系的约束(Zhu et al., 2017; Zhang et al., 2019). ...
Hydraulic properties and photosynthetic rates in co-occurring lianas and trees in a seasonal tropical rainforest in southwestern China 1 2009
... 本研究使用注气法测定木质部脆弱性曲线, 但是该方法的准确性受到质疑.在测量过程中, 当使用的茎段长度小于最大导管长度时产生的导管开口现象, 以及加压过程中形成的气泡效应, 会导致PLC被高估(抗栓塞能力被低估; Choat et al., 2010; Yin & Cai, 2018).另外, 加压的时间以及压力套的长度都会对曲线造成影响(Yin & Cai, 2018).由于本试验物种(部分为引种)可破坏性采集的样本量少, 无法选择较为精确但是枝条样本量需求较多的自然干燥法(Sperry et al., 1988).另外, 本研究尽量使用足够长的枝条以防止开口导管产生的影响(Ennajeh et al., 2011).已有研究利用自然干燥法测定了部分物种的P50branch, 如红河崖豆(Fu et al., 2012)、厚果崖豆藤(Zhu & Cao, 2009)和小叶买麻藤(Chen et al., 2015), 本研究测定的结果与之相似. ...
Interspecific variation in branch and leaf traits among three Syzygium tree species from different successional tropical forests 1 2015
... 植物的水力特征是探讨植物生理生态适应的核心内容(Choat et al., 2012).植物水力学性状主要包括: (1)水力传导能力(有效性), 如边材和叶片比导率(ks、kl), 它是衡量植物水分传导效率的指标(Sperry et al., 1988).水分传导效率与叶片的光合能力密切相关(Santiago et al., 2004), 并最终影响植物的生长速率(Brodribb et al., 2010).(2)抗栓塞能力(安全性), 用植物输导组织损失50%导水能力时的水势值(P50)表示, 与植物的干旱适应策略有关.植物水分传导有效性与安全性之间存在权衡关系, 是探讨植物的生长策略以及沿水分梯度分布等方面的重要的生理生态机制(Markesteijn et al., 2011).但是基于全球数据分析发现这种权衡关系较弱(Gleason et al., 2016; Santiago et al., 2018), 在水分胁迫不严重的热带森林中甚至没有权衡(van der Sande et al., 2019).植物不同器官的水力特征之间的差异或者协同关系是水力学研究的重要内容(Bucci et al., 2012).经典的脆弱性分段假说认为植物茎的抗栓塞能力要强于末端的叶片, 在水分胁迫状况下, 叶片更容易发生栓塞, 从而减少水分进一步损失, 保证了茎的水力安全(Zimmerman & Brown, 1971; Tyree & Ewers, 1991).因此, 叶片在植物整个水分运输通路中起到了“安全阀”的作用(McCulloh et al., 2014).但是, 也有部分研究发现热带亚热带森林的植物并不符合脆弱性分段假说: 叶片的抗栓塞能力与枝条相近(Villagra et al., 2013), 甚至要强于枝条(Zhu et al., 2015), 原因在于枝-叶的脆弱性分段与生境的干旱指数相关, 生长在湿润地区的植物具有稳定的水分来源, 环境选择压力小; 另外, 缺乏枝-叶脆弱性分段的植物具有丰富的水力补偿策略维持叶片的水分供给, 在潜在的干旱胁迫下可能具有生长优势(Zhu et al., 2016).基于水力性状可以分析植物的水分适应策略, 被广泛地用于解释植物一系列的生态学表现(de Guzman et al., 2017). ...
Different hydraulic traits of woody plants from tropical forests with contrasting soil water availability 2 2017
... 买麻藤属植物是特殊的植物类群, 其生理生态适应机制引起广泛的关注(Kotowska et al., 2015).Feild和Balun (2008)对几种买麻藤植物的水力和光合相关性状进行了测定, 结果发现它们的水分传导效率和光合速率都很低.虽然买麻藤植物进化出与被子植物相似的导管系统和阔叶, 但是还没有进化出类似热带雨林需光种的典型特征, 如: 高繁殖速率与高光合速率, 因此常分布于热带森林林下阴生湿润生境中(Feild & Balun, 2008; Celis & Avalos, 2013).本研究中买麻藤属乔木树种灌状买麻藤与少苞买麻藤的测定结果符合这一结论.但是, 买麻藤属木质藤本小叶买麻藤则表现出很高的水分传导效率, 与阳生性的被子植物相当.这可能与其已进化出粗大的导管有关, 其平均水力导管直径为171 mm (未发表数据), 远高于热带森林植物的平均值(100 mm; Zhu et al., 2017).但其光合速率仍无法与阳生性的被子植物相比, 这可能与其进化史有关, 因为多功能基因的缺失导致的低光合速率可能是买麻藤属固有的特性(Deng et al., 2019). ...
... 植物枝条和叶片的水分传导有效性与安全性之间均没有显著的权衡关系(图3A、3B), 不支持第二个研究假设.对于枝条来说, 部分树种枝条木质部同时具有较低的水分传导效率和抗栓塞能力, 如生长在林下的思茅崖豆.木质部的水分传导效率主要决定于木质部导管特征, 而木质部水力安全是各种水力策略密切协同的结果, 如根系吸水、茎部储水能力和气孔调节(Mencuccini et al., 2015)等, 因此植物整株水平的水力抗旱机制可能会导致枝条水平上缺乏安全性和有效性的权衡关系(Meinzer et al., 2010).另外, 由于木质藤本木质部结构的特殊性(Ewers et al., 2015), 它们在木质部支撑和输导组织的分配上受到的限制较少, 在相似的抗栓塞能力的情况下, 可能存在更多的水分传导效率的变化(Bittencourt et al., 2016), 导致木质藤本不受水力权衡关系的约束(Zhu et al., 2017; Zhang et al., 2019). ...
Are leaves more vulnerable to cavitation than branches? 3 2016
... 植物的水力特征是探讨植物生理生态适应的核心内容(Choat et al., 2012).植物水力学性状主要包括: (1)水力传导能力(有效性), 如边材和叶片比导率(ks、kl), 它是衡量植物水分传导效率的指标(Sperry et al., 1988).水分传导效率与叶片的光合能力密切相关(Santiago et al., 2004), 并最终影响植物的生长速率(Brodribb et al., 2010).(2)抗栓塞能力(安全性), 用植物输导组织损失50%导水能力时的水势值(P50)表示, 与植物的干旱适应策略有关.植物水分传导有效性与安全性之间存在权衡关系, 是探讨植物的生长策略以及沿水分梯度分布等方面的重要的生理生态机制(Markesteijn et al., 2011).但是基于全球数据分析发现这种权衡关系较弱(Gleason et al., 2016; Santiago et al., 2018), 在水分胁迫不严重的热带森林中甚至没有权衡(van der Sande et al., 2019).植物不同器官的水力特征之间的差异或者协同关系是水力学研究的重要内容(Bucci et al., 2012).经典的脆弱性分段假说认为植物茎的抗栓塞能力要强于末端的叶片, 在水分胁迫状况下, 叶片更容易发生栓塞, 从而减少水分进一步损失, 保证了茎的水力安全(Zimmerman & Brown, 1971; Tyree & Ewers, 1991).因此, 叶片在植物整个水分运输通路中起到了“安全阀”的作用(McCulloh et al., 2014).但是, 也有部分研究发现热带亚热带森林的植物并不符合脆弱性分段假说: 叶片的抗栓塞能力与枝条相近(Villagra et al., 2013), 甚至要强于枝条(Zhu et al., 2015), 原因在于枝-叶的脆弱性分段与生境的干旱指数相关, 生长在湿润地区的植物具有稳定的水分来源, 环境选择压力小; 另外, 缺乏枝-叶脆弱性分段的植物具有丰富的水力补偿策略维持叶片的水分供给, 在潜在的干旱胁迫下可能具有生长优势(Zhu et al., 2016).基于水力性状可以分析植物的水分适应策略, 被广泛地用于解释植物一系列的生态学表现(de Guzman et al., 2017). ...
... 与第三个研究假设相符, 两个属的木质藤本植物均不符合传统的枝-叶脆弱性分段假说, 即叶片比枝条具有更强的抗栓塞能力(图4C).枝-叶脆弱性分段不存在的原因可能与本研究测定脆弱性曲线的方法有关(可能低估了枝条的抗栓塞能力), 但是最近的研究采用相同的方法(光学法)测定枝条和叶片的脆弱性曲线, 同样发现很多树种不符合传统的 枝-叶脆弱性分段(Smith-Martin et al., 2020).另外, 前期研究表明这些植物在增强叶片抗栓塞能力的同时, 拥有有效的水力策略以保证茎的水力安全, 在干旱胁迫的情况下可能会具有生理优势: 能较大限度地推迟关闭气孔, 从而保持光合作用和生长(Zhu et al., 2016).本研究发现枝-叶脆弱性分段与旱季最大净光合速率和最大气孔导度呈显著负相关关系, 直接支持Zhu等(2016)的研究论断.另外, 由于木质藤本具有比同属乔木更强的旱季光合能力, 也支持了木质藤本植物的旱季优势生长假说(Zimmerman & Brown, 1971; Tyree & Ewers, 1991; Schnitzer & van der Heijden, 2019). ...
... ).本研究发现枝-叶脆弱性分段与旱季最大净光合速率和最大气孔导度呈显著负相关关系, 直接支持Zhu等(2016)的研究论断.另外, 由于木质藤本具有比同属乔木更强的旱季光合能力, 也支持了木质藤本植物的旱季优势生长假说(Zimmerman & Brown, 1971; Tyree & Ewers, 1991; Schnitzer & van der Heijden, 2019). ...
2 1971
... 植物的水力特征是探讨植物生理生态适应的核心内容(Choat et al., 2012).植物水力学性状主要包括: (1)水力传导能力(有效性), 如边材和叶片比导率(ks、kl), 它是衡量植物水分传导效率的指标(Sperry et al., 1988).水分传导效率与叶片的光合能力密切相关(Santiago et al., 2004), 并最终影响植物的生长速率(Brodribb et al., 2010).(2)抗栓塞能力(安全性), 用植物输导组织损失50%导水能力时的水势值(P50)表示, 与植物的干旱适应策略有关.植物水分传导有效性与安全性之间存在权衡关系, 是探讨植物的生长策略以及沿水分梯度分布等方面的重要的生理生态机制(Markesteijn et al., 2011).但是基于全球数据分析发现这种权衡关系较弱(Gleason et al., 2016; Santiago et al., 2018), 在水分胁迫不严重的热带森林中甚至没有权衡(van der Sande et al., 2019).植物不同器官的水力特征之间的差异或者协同关系是水力学研究的重要内容(Bucci et al., 2012).经典的脆弱性分段假说认为植物茎的抗栓塞能力要强于末端的叶片, 在水分胁迫状况下, 叶片更容易发生栓塞, 从而减少水分进一步损失, 保证了茎的水力安全(Zimmerman & Brown, 1971; Tyree & Ewers, 1991).因此, 叶片在植物整个水分运输通路中起到了“安全阀”的作用(McCulloh et al., 2014).但是, 也有部分研究发现热带亚热带森林的植物并不符合脆弱性分段假说: 叶片的抗栓塞能力与枝条相近(Villagra et al., 2013), 甚至要强于枝条(Zhu et al., 2015), 原因在于枝-叶的脆弱性分段与生境的干旱指数相关, 生长在湿润地区的植物具有稳定的水分来源, 环境选择压力小; 另外, 缺乏枝-叶脆弱性分段的植物具有丰富的水力补偿策略维持叶片的水分供给, 在潜在的干旱胁迫下可能具有生长优势(Zhu et al., 2016).基于水力性状可以分析植物的水分适应策略, 被广泛地用于解释植物一系列的生态学表现(de Guzman et al., 2017). ...
... 与第三个研究假设相符, 两个属的木质藤本植物均不符合传统的枝-叶脆弱性分段假说, 即叶片比枝条具有更强的抗栓塞能力(图4C).枝-叶脆弱性分段不存在的原因可能与本研究测定脆弱性曲线的方法有关(可能低估了枝条的抗栓塞能力), 但是最近的研究采用相同的方法(光学法)测定枝条和叶片的脆弱性曲线, 同样发现很多树种不符合传统的 枝-叶脆弱性分段(Smith-Martin et al., 2020).另外, 前期研究表明这些植物在增强叶片抗栓塞能力的同时, 拥有有效的水力策略以保证茎的水力安全, 在干旱胁迫的情况下可能会具有生理优势: 能较大限度地推迟关闭气孔, 从而保持光合作用和生长(Zhu et al., 2016).本研究发现枝-叶脆弱性分段与旱季最大净光合速率和最大气孔导度呈显著负相关关系, 直接支持Zhu等(2016)的研究论断.另外, 由于木质藤本具有比同属乔木更强的旱季光合能力, 也支持了木质藤本植物的旱季优势生长假说(Zimmerman & Brown, 1971; Tyree & Ewers, 1991; Schnitzer & van der Heijden, 2019). ...
 ,
, ,*广西大学林学院广西森林生态与保育重点实验室, 南宁 530004; 广西大学亚热带农业生物资源保护与利用国家重点实验室, 南宁 530004
,*广西大学林学院广西森林生态与保育重点实验室, 南宁 530004; 广西大学亚热带农业生物资源保护与利用国家重点实验室, 南宁 530004
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT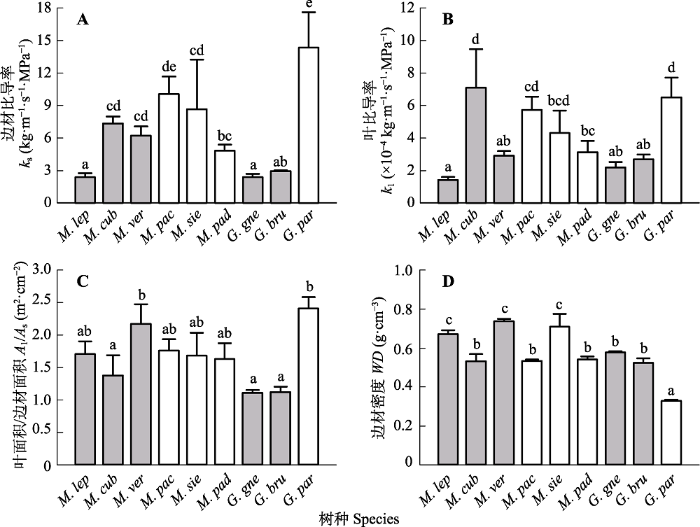 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT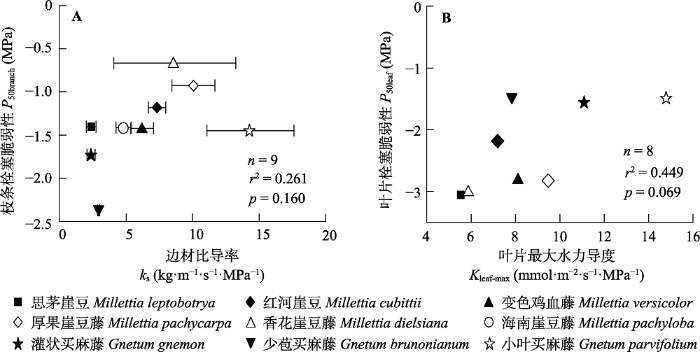 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT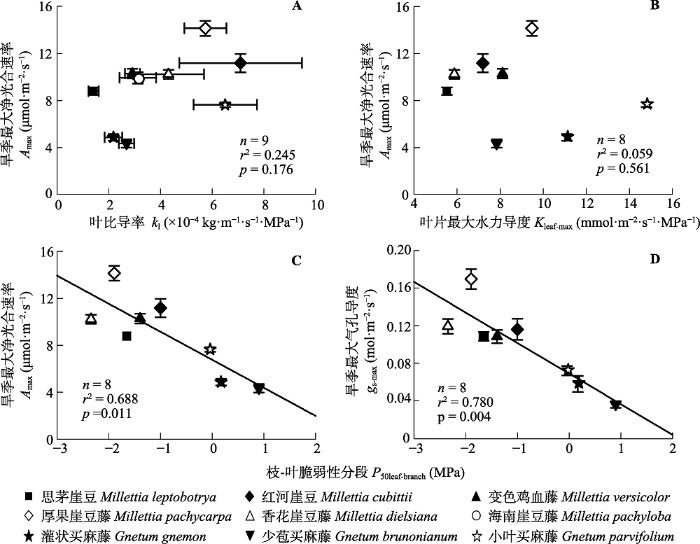 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}