Nitrogen-fixing bacteria alleviates competition between arbuscular mycorrhizal fungi and Solidago canadensis for nutrients under nitrogen limitation
Fang PANG1, Wei-Kang XIA1, Min HE1, Shan-Shan QI2, Zhi-Cong DAI,,1,2,*, Dao-Lin DU11 School of the Environment and Safety Engineering, Jiangsu University, Zhenjiang, Jiangsu 212013, China 2 Key Laboratory of Modern Agricultural Equipment and Technology, Ministry of Education, School of Agricultural Engineering, Jiangsu University, Zhenjiang, Jiangsu 212013, China
National Natural Science Foundation of China(31700342) National Natural Science Foundation of China(31770446) China Postdoctoral Science Foundation(2017T100329) Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD)
Abstract Aims Arbuscular mycorrhizal fungi (AMF) form symbiotic relationships with most terrestrial plants, contributing to the nutrient uptake of host plants. While little is known on how rhizospheric microorganisms affect the relationships between AMF and host plants under nutritional stress. We hypothesize that AMF may compete for nutrients with host plants in extremely nutrient-limited environments, such as nitrogen deficient habitats, and nitrogen-fixing bacteria will alleviate the competition. Methods In order to test our hypotheses, we grew Solidago canadensis plants under nitrogen deficient treatments. We inoculated plants with AMF and a nitrogen-fixing bacterium to test the relationships among the host plant and microorganisms. Important findings Under the lowest nitrogen level (0.025 mmol·L-1 N of ammonium nitrogen), the growth of S. canadensis was more restricted with AMF colonization, suggesting competition between AMF and the host. However, with the inoculation of nitrogen-fixing bacterium, AMF colonization was promoted and plant growth was increased. These results indicate that nitrogen-fixing bacteria could moderate the competition for nutrients between AMF and their host under nitrogen deficiency. This study improves our understanding of the invasion mechanisms of alien plants, where they interact with different microorganisms under extreme nutrient stress. Keywords:arbuscular mycorrhizal fungi;nitrogen limitation;nutrients competition;nitrogen-fixing bacteria;invasive plant
PDF (1434KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 庞芳, 夏维康, 何敏, 祁珊珊, 戴志聪, 杜道林. 固氮菌缓解氮限制环境中丛枝菌根真菌对加拿大一枝黄花的营养竞争. 植物生态学报, 2020, 44(7): 782-790. DOI: 10.17521/cjpe.2020.0114 PANG Fang, XIA Wei-Kang, HE Min, QI Shan-Shan, DAI Zhi-Cong, DU Dao-Lin. Nitrogen-fixing bacteria alleviates competition between arbuscular mycorrhizal fungi and Solidago canadensis for nutrients under nitrogen limitation. Chinese Journal of Plant Ecology, 2020, 44(7): 782-790. DOI: 10.17521/cjpe.2020.0114
Fig. 2Neighbour-joining tree of the isolated nitrogen-fixing bacterium, ScRB-JS106, based on 16S rDNA. Sequences for the other isolates were downloaded from GenBank.
CK, 对照处理(10 mmol·L-1 N)。不同小写字母表示差异显著(p < 0.05)。 Fig. 3Growth of Solidago canadensis under different concentrations of ammonium chloride (mean ± SE, n = 4).
CK, negative control (10 mmol·L-1 N). Different lowercase letters indicate significant differences (Duncan’s test, p < 0.05).
CK, 对照处理(10 mmol·L-1 N)。不同小写字母表示差异显著(p < 0.05)。 Fig. 4Growth of Solidago canadensis under different concentrations of potassium nitrate (mean ± SE, n = 4).
CK, negative control (10 mmol·L-1 N). Different lowercase letters indicate significant differences (Duncan’s test, p < 0.05).
2.3 固氮菌对AMF与加拿大一枝黄花间关系的影响
低氮限制处理下, 3种AMF对加拿大一枝黄花根部均有侵染(图5), 其中以GI的侵染率最高(约33.3%, 图5B)。而接种固氮菌后, 在一定程度上提高了GE (+6.7%, p = 0.439)和GI (+11.7%, p = 0.251)对加拿大一枝黄花根部的侵染(图5A、5B)。
图5
新窗口打开|下载原图ZIP|生成PPT 图5不同菌剂接种方式下加拿大一枝黄花根部的丛枝菌根真菌侵染率(平均值±标准偏差, n = 6)。
A, 幼套球囊霉(GE)。 B, 根内球囊霉(GI)。C, 摩西球囊霉(GM)。JS106, 星孢类芽孢杆菌ScRB-JS106。ns, 无显著性差异(p > 0.05)。 Fig. 5Root colonization rate of Solidago canadensis inoculated with various arbuscular mycorrhizal fungi and nitrogen-fixing bacteria (mean ± SE, n = 6).
新窗口打开|下载原图ZIP|生成PPT 图6氮限制环境中接种幼套球囊霉及同时添加固氮菌处理加拿大一枝黄花的生长情况(平均值±标准偏差, n = 6)。
GE, 幼套球囊霉。JS106, 星孢类芽孢杆菌ScRB-JS106。CK, 未添加GE处理。ns, 无显著性差异(p > 0.05)。 Fig. 6Growth of Solidago canadensis inoculated with Glomus etunicatum and nitrogen-fixing bacteria under nitrogen limitation (mean ± SE, n = 6).
GE, G. etunicatum. JS106, Paenibacillus stellifer ScRB-JS106. CK, un-inoculated control. ns, no significant difference (p > 0.05).
图7
新窗口打开|下载原图ZIP|生成PPT 图7氮限制环境中接种根内球囊霉及同时添加固氮菌处理加拿大一枝黄花的生长情况(平均值±标准偏差, n = 6)。
GI, 根内球囊霉。JS106, 星孢类芽孢杆菌ScRB-JS106。CK, 未添加GI处理。ns, 无显著性差异(p > 0.05)。 Fig. 7Growth of Solidago canadensis inoculated with Glomus intraradices and nitrogen-fixing bacteria (mean ± SE, n = 6).
新窗口打开|下载原图ZIP|生成PPT 图8氮限制环境中接种摩西球囊霉及同时添加固氮菌处理加拿大一枝黄花的生长情况(平均值±标准偏差, n = 6)。
GM, 摩西球囊霉。JS106, 星孢类芽孢杆菌ScRB-JS106。CK, 未添加GM处理。ns, 无显著性差异(p > 0.05)。 Fig. 8Growth of Solidago canadensis inoculated with Glomus mosseae and nitrogen-fixing bacteria (mean ± SE, n = 6).
GM, G. mosseae. JS106, Paenibacillus stellifer ScRB-JS106. CK, un-inoculated control. ns, no significant difference (p > 0.05).
3 讨论
根际最佳分配理论认为, 植物生长过程中会因生存环境变化而对资源分配进行动态调节, 它们可以通过将光合作用制造的有机物分配给各营养器官或生殖器官, 以提高获取稀缺资源的能力(Thornley, 1972; Bloom et al., 1985)。相比生长在富营养环境下的植物, 那些生长在贫瘠的、低氮土壤上的植物往往倾向于将更多的资源投入到根系生长, 表现出根部具有更高的质量分数(Tilman & Wedin, 1991; Cambui et al., 2011; Sun et al., 2014)。在加拿大一枝黄花氮受限条件筛选实验中, 我们发现在低浓度铵态氮处理下加拿大一枝黄花地下部分生物量显著高于对照组, 而地上部分的生物量明显低于对照组(图3)。这表明加拿大一枝黄花在氮缺乏环境下可能是通过促进根系生长来扩大其在基质中的营养有效吸收面积, 从而提高自身的环境适应能力, 这可能是加拿大一枝黄花成功入侵的机制之一。
BattiniF, Gr?nlundM, AgnolucciM, GiovannettiM, JakobsenI (2017). Facilitation of phosphorus uptake in maize plants by mycorrhizosphere bacteria Scientific Reports, 7, 4686. DOI: 10.1038/s41598-017-04959-0. DOI:10.1038/s41598-017-04959-0URLPMID:28680077 [本文引用: 1] A major challenge for agriculture is to provide sufficient plant nutrients such as phosphorus (P) to meet the global food demand. The sufficiency of P is a concern because of it's essential role in plant growth, the finite availability of P-rock for fertilizer production and the poor plant availability of soil P. This study investigated whether biofertilizers and bioenhancers, such as arbuscular mycorrhizal fungi (AMF) and their associated bacteria could enhance growth and P uptake in maize. Plants were grown with or without mycorrhizas in compartmented pots with radioactive P tracers and were inoculated with each of 10 selected bacteria isolated from AMF spores. Root colonization by AMF produced large plant growth responses, while seven bacterial strains further facilitated root growth and P uptake by promoting the development of AMF extraradical mycelium. Among the tested strains, Streptomyces sp. W94 produced the largest increases in uptake and translocation of (33)P, while Streptomyces sp. W77 highly enhanced hyphal length specific uptake of (33)P. The positive relationship between AMF-mediated P absorption and shoot P content was significantly influenced by the bacteria inoculants and such results emphasize the potential importance of managing both AMF and their microbiota for improving P acquisition by crops.
BloomAJ, ChapinIII FS, MooneyHA (1985). Resource limitation in plants—An economic analogy Annual Review of Ecology, Evolution, and Systematics, 16, 363-392. [本文引用: 1]
BonfanteP, GenreA (2010). Mechanisms underlying beneficial plant-fungus interactions in mycorrhizal symbiosis Nature Communications, 1, 48. DOI: 10.1038/ncomms1046. DOI:10.1038/ncomms1046URLPMID:20975705 [本文引用: 1] Mycorrhizal fungi are a heterogeneous group of diverse fungal taxa, associated with the roots of over 90% of all plant species. Recently, state-of-the-art molecular and genetic tools, coupled to high-throughput sequencing and advanced microscopy, have led to the genome and transcriptome analysis of several symbionts. Signalling pathways between plants and fungi have now been described and the identification of several novel nutrient transporters has revealed some of the cellular processes that underlie symbiosis. Thus, the contributions of each partner in a mycorrhizal association are starting to be unravelled. This new knowledge is now available for use in agricultural practices.
BunnRA, LekbergY, GallagherC, RosendahlS, RamseyPW (2014). Grassland invaders and their mycorrhizal symbionts: a study across climate and invasion gradients Ecology and Evolution, 4, 794-805. URLPMID:24683461 [本文引用: 1]
ChandrasekaranM, BoughattasS, HuSJ, OhSH, SaTM (2014). A meta-analysis of arbuscular mycorrhizal effects on plants grown under salt stress Mycorrhiza, 24, 611-625. URLPMID:24770494 [本文引用: 1]
ChenQ, WuWW, QiSS, ChengH, LiQ, RanQ, DaiZC, DuDL, EganS, ThomasT (2020). Arbuscular mycorrhizal fungi improve the growth and disease resistance of the invasive plantWedelia trilobata Journal of Applied Microbiology, 2019,0665. DOI: 10.1111/jam.14415. [本文引用: 4]
ChenYL, ChenBD, LiuL, HuYJ, XuTL, ZhangX (2014). The role of arbuscular mycorrhizal fungi in soil nitrogen cycling Acta Ecologica Sinica, 34, 4807-4815. [本文引用: 1]
ChengJK, YueMF, YangHR, ChenBM, XinGR (2019). Do arbuscular mycorrhizal fungi help the native speciesBidens biternata resist the invasion of Bidens alba? Plant and Soil, 444, 443-455. [本文引用: 1]
CiE, GaoM (2004). Research progress on biological nitrogen fixation Chinese Agricultural Science Bulletin, 20(1), 25-28. [本文引用: 1]
CockingEC (2003). Endophytic colonization of plant roots by nitrogen-fixing bacteria Plant and Soil, 252, 169-175. [本文引用: 1]
DaiZC, FuW, QiSS, ZhaiDL, ChenSC, WanLY, HuangP, DuDL (2016a). Different responses of an invasive clonal plant Wedelia trilobata and its native congener to gibberellin: implications for biological invasion Journal of Chemical Ecology, 42, 85-94. URLPMID:26879680 [本文引用: 1]
DaiZC, FuW, WanLY, CaiHH, WangN, QiSS, DuDL (2016b). Different growth promoting effects of endophytic bacteria on invasive and native clonal plants Frontiers in Plant Science, 7, 706. DOI: 10.3389/fpls.2016. 00706. DOI:10.3389/fpls.2016.00706URLPMID:27252722 [本文引用: 2] The role of the interactions between endophytes and alien plants has been unclear yet in plant invasion. We used a completely germ-free culture system to quantify the plant growth-promoting (PGP) effects of endophytic bacteria Bacillus sp. on aseptic seedlings of Wedelia trilobata and of its native clonal congener W. chinensis. The endophytic bacteria did not affect the growth of W. chinensis, but they significantly promoted the growth of W. trilobata. With the PGP effects of endophytic bacteria, relative change ratios of the clonal traits and the ramets' growth traits of W. trilobata were significantly greater than those of W. chinensis. Our results indicate that the growth-promoting effects of endophytes may differ between invasive and native clonal plants, and the endophytes of invasive plant may be host-specific to facilitate plant invasion.
DingY, WangJ, LiuY, ChenS (2005). Isolation and identification of nitrogen-fixing bacilli from plant rhizospheres in Beijing region Journal of Applied Microbiology, 99, 1271-1281. DOI:10.1111/j.1365-2672.2005.02738.xURLPMID:16238759 [本文引用: 1] AIMS: To isolate and identify nitrogen-fixing bacilli from the plant rhizospheres in Beijing region of China. METHODS AND RESULTS: A total of 29 isolates were selectively obtained from the rhizospheres of wheat, maize, ryegrass and willow based on their growth on nitrogen-free medium and their resistance to 100 degrees C for 10 min. Of the 29 isolates, seven had nifH gene determined by PCR amplification. The seven isolates were found to belong to the genera Bacillus and Paenibacillus based on phenotypic characterization, 16S rDNA sequence, G+C content and DNA-DNA hybridization. Isolates T1 and W5 were identified as Bacillus cereus and Bacillus marisflavi respectively. Isolates G1, C4 and C5 were identified as Bacillus megaterium. Isolate G2 was identified as Paenibacillus polymyxa and isolate T7 as Paenibacillus massiliensis. CONCLUSIONS: This study suggests that nifH gene could be detected in the both genera Bacillus and Paenibacillus. These degenerate primers for nifH gene fragment used in this study were shown to be useful for identifying nitrogen-fixing bacilli. SIGNIFICANCE AND IMPACT OF THE STUDY: It is the first demonstration that nitrogen fixation exists in B. marisflavi and P. massiliensis and the first report of the sequences of the nifH gene from B. megaterium and B. cereus. The nitrogen-fixing bacilli obtained in this study will be used in our future research for investigating the mechanisms of nitrogen fixation in bacilli.
FrancheC, Lindstr?mK, ElmerichC (2009). Nitrogen-fixing bacteria associated with leguminous and non-leguminous plants Plant and Soil, 321, 35-59. [本文引用: 1]
HodgeA, CampbellCD, FitterAH (2001). An arbuscular mycorrhizal fungus accelerates decomposition and acquires nitrogen directly from organic material Nature, 413, 297-299. URLPMID:11565029 [本文引用: 1]
JinL (2005). Ecology of Arbuscular Mycorrhizal Associations in Solidago canadensis, an Invasive Alien Plant PhD dissertation, Fudan University, Shanghai. [本文引用: 2]
JinL, GuYJ, XiaoM, ChenJK, LiB (2004). The history of Solidago canadensis invasion and the development of its mycorrhizal associations in newly-reclaimed land Functional Plant Biology, 31, 979-986. URLPMID:32688966 [本文引用: 1]
JohnsonN, WilsonG, BowkerM, WilsonJ, MillerR (2010). Resource limitation is a driver of local adaptation in mycorrhizal symbioses Proceedings of the National Academy of Sciences of the United States of America, 107, 2093-2098. URLPMID:20133855 [本文引用: 2]
JohnsonNC (2010). Resource stoichiometry elucidates the structure and function of arbuscular mycorrhizas across scales New Phytologist, 185, 631-647. [本文引用: 1]
KhanAG (1974). The occurrence of mycorrhizas in halophytes, hydrophytes and xerophytes, and of Endogone spores in adjacent soils Microbiology, 81, 7-14. [本文引用: 1]
LeeMR, TuC, ChenX, HuSJ (2014). Arbuscular mycorrhizal fungi enhance P uptake and alter plant morphology in the invasive plant Microstegium vimineum Biological Invasions, 16, 1083-1093. [本文引用: 2]
LeighJ, HodgeA, FitterAH (2009). Arbuscular mycorrhizal fungi can transfer substantial amounts of nitrogen to their host plant from organic material The New Phytologist, 181, 199-207. [本文引用: 1]
LiuCL, ZuoWY, ZhaoZY, QiuLH (2012). Bacterial diversity of different successional stage forest soils in Dinghushan Acta Microbiologica Sinica, 52, 1489-1496. URLPMID:23457798 [本文引用: 1]
MurugesanC, BoughattasS, HuSJ, OhSH, SaT (2014). A meta-analysis of arbuscular mycorrhizal effects on plants grown under salt stress Mycorrhiza, 24, 611-625. DOI:10.1007/s00572-014-0582-7URLPMID:24770494 [本文引用: 1] Salt stress limits crop yield and sustainable agriculture in most arid and semiarid regions of the world. Arbuscular mycorrhizal fungi (AMF) are considered bio-ameliorators of soil salinity tolerance in plants. In evaluating AMF as significant predictors of mycorrhizal ecology, precise quantifiable changes in plant biomass and nutrient uptake under salt stress are crucial factors. Therefore, the objective of the present study was to analyze the magnitude of the effects of AMF inoculation on growth and nutrient uptake of plants under salt stress through meta-analyses. For this, data were compared in the context of mycorrhizal host plant species, plant family and functional group, herbaceous vs. woody plants, annual vs. perennial plants, and the level of salinity across 43 studies. Results indicate that, under saline conditions, AMF inoculation significantly increased total, shoot, and root biomass as well as phosphorous (P), nitrogen (N), and potassium (K) uptake. Activities of the antioxidant enzymes superoxide dismutase, catalase, peroxidase, and ascorbate peroxidase also increased significantly in mycorrhizal compared to nonmycorrhizal plants growing under salt stress. In addition, sodium (Na) uptake decreased significantly in mycorrhizal plants, while changes in proline accumulation were not significant. Across most subsets of the data analysis, identities of AMF (Glomus fasciculatum) and host plants (Acacia nilotica, herbs, woody and perennial) were found to be essential in understanding plant responses to salinity stress. For the analyzed dataset, it is concluded that under salt stress, mycorrhizal plants have extensive root traits and mycorrhizal morphological traits which help the uptake of more P and K, together with the enhanced production of antioxidant enzymes resulting in salt stress alleviation and increased plant biomass.
PepeA, SbranaC, FerrolN, GiovannettiM (2017). An in vivo whole-plant experimental system for the analysis of gene expression in extraradical mycorrhizal mycelium Mycorrhiza, 27, 659-668. URLPMID:28573458 [本文引用: 1]
PhillipsJM, HaymanDS (1970). Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection Transactions of the British Mycological Society, 55, 158-161. [本文引用: 1]
SchmidB, PuttickGM, BurgessKH, BazzazFA (1988). Clonal integration and effects of simulated herbivory in old-field perennials Oecologia, 75, 465-471. URLPMID:28312698 [本文引用: 1]
ShahM, ReshiZ, RashidI (2008a). Mycorrhizal source and neighbour identity differently influence Anthemis cotula L. invasion in the Kashmir Himalaya, India Applied Soil Ecology, 40, 330-337. [本文引用: 1]
ShahMA, ReshiZ, RashidI (2008b). Mycorrhizosphere mediated Mayweed chamomile invasion in the Kashmir himalaya, India Plant and Soil, 312, 219-225. [本文引用: 1]
SmithFA, GraceEJ, SmithSE (2009). More than a carbon economy: nutrient trade and ecological sustainability in facultative arbuscular mycorrhizal symbioses New Phytologist, 182, 347-358. DOI:10.1111/j.1469-8137.2008.02753.xURLPMID:19207688 [本文引用: 1] Symbiosis is well recognized as a major force in plant ecology and evolution. However, there is considerable uncertainty about the functional, ecological and evolutionary benefits of the very widespread facultative arbuscular mycorrhizal (AM) associations, in which the plants can grow and reproduce whether or not they are colonized by AM fungi. Here we address the significance of new research findings that are overturning conventional views that facultative AM associations can be likened to parasitic fungus-plant associations. Specifically, we address the occurrence and importance of phosphate uptake via AM fungi that does not result in increases in total phosphorus (P) uptake or in plant growth, and possible signalling between AM fungi and plants that can result in plant growth depressions even when fungal colonization remains very low. We conclude that, depending on the individual AM fungi that are present, the role of facultative AM associations in the field, especially in relation to plant competition, may be much more subtle than has been previously envisaged.
SmithSE, SmithFA (2011). Roles of arbuscular mycorrhizas in plant nutrition and growth: new paradigms from cellular to ecosystem scales Annual Review of Plant Biology, 62, 227-250. URLPMID:21391813 [本文引用: 1]
TanakaY, YanoK (2005). Nitrogen delivery to maize via mycorrhizal hyphae depends on the form of N supplied Plant Cell and Environment, 28, 1247-1254. [本文引用: 2]
TangJJ, ZhangQ, YangRY, ChenX (2009). Effects of exotic plant Solidago canadensis L. on local arbuscular mycorrhizal fungi Bulletin of Science and Technology, 25, 233-237. [本文引用: 1]
ThornleyJHM (1972). A balanced quantitative model for root:shoot ratios in vegetative plants Annals of Botany, 36, 431-441. [本文引用: 1]
TilmanD, WedinDA (1991). Plant traits and resource reduction for five grasses growing on a nitrogen gradient Ecology, 72, 685-700. [本文引用: 1]
WangB (2013). Effects of Allelopathy and Arbuscular Mycorrhizae in the Invasion of Solidago canadensis L Master degree dissertation, Zhejiang University, Hangzhou. [本文引用: 1]
WangCQ, LiuS, WangXL, ChenHW, WuYJ, WangZG (2015). Isolation, identification and characteristics of authigenic azotobacters from the rhizosphere of soybean Soybean Science, 34, 850-854. [本文引用: 1]
WangYH, WangMQ, LiY, WuAP, HuangJY (2018). Effects of arbuscular mycorrhizal fungi on growth and nitrogen uptake of Chrysanthemum morifolium under salt stress PLOS ONE, 13, e0196408. DOI: 10.1371/journal.pone. 0196408. URLPMID:29698448 [本文引用: 1]
WernerPA, GrossRS, BradburyIK (1980). The biology of Canadian.: 45.Solidago canadensis L Canadian Journal of Plant Science, 60, 1393-1409. [本文引用: 1]
WilsonGWT, HickmanKR, WilliamsonMM (2012). Invasive warm-season grasses reduce mycorrhizal root colonization and biomass production of native prairie grasses Mycorrhiza, 22, 327-336. URLPMID:21845465 [本文引用: 1]
YangJY, WangYH, WenGS, YiLT (2013). Effects of AM fungi and simulated nitrogen deposition on the growth and biomass accumulation of Solidago canadensis seedings Chinese Journal of Ecology, 32, 2953-2958. [本文引用: 2]
ZhangFJ, LiQ, ChenFX, XuHY, Inderjit, WanFH (2017). Arbuscular mycorrhizal fungi facilitate growth and competitive ability of an exotic species Flaveria bidentis Soil Biology & Biochemistry, 115, 275-284. [本文引用: 1]
ZhouG, ZanS, GuoFY, SuNN, LiJ (2014). Arbuscular mycorrhizal fungi facilitate the invasion of Solidago canadensis L. in southeastern China Acta Oecologica, 61, 71-77. [本文引用: 1]
ZhuC, TianGL, LuoGW, KongYL, GuoJJ, WangM, GuoSW, LingN, ShenQR (2018). N-fertilizer-driven association between the arbuscular mycorrhizal fungal community and diazotrophic community impacts wheat yield Agriculture Ecosystems & Environment, 254, 191-201. [本文引用: 2]
Facilitation of phosphorus uptake in maize plants by mycorrhizosphere bacteria 1 2017
... 丛枝菌根真菌(AMF)是一种普遍存在的根际微生物, 在所有陆地生态系统中70%-90%的植物能被AMF定殖(Khan, 1974; Smith & Read, 2008; Bonfante & Genre, 2010), 能在植物根部形成复杂的菌丝网络(CMNs), 与植物形成互惠共生体.这种高效的共生关系除了能够明显改善宿主植物的营养水平(氮、磷、硫等)外, 还能帮助宿主植物适应多种逆境胁迫(Smith & Read, 2008; Battini et al., 2017; Pepe et al., 2017).AMF能够利用土壤中游离的NH4+、NO3-、简单形态的氨基酸和有机氮(Hodge et al., 2001; Leigh et al., 2009; 陈永亮等, 2014), 并且会优先利用NH4+ (Tanaka & Yano, 2005), 与宿主植物交换碳水化合物(Johnson et al., 2010), 改善宿主植物对氮的吸收水平(Murugesan et al., 2014; Wang et al., 2018; Chen et al., 2020), 形成互利共生的关系(Johnson, 2010; Smith et al., 2009). ...
Resource limitation in plants—An economic analogy 1 1985
... 根际最佳分配理论认为, 植物生长过程中会因生存环境变化而对资源分配进行动态调节, 它们可以通过将光合作用制造的有机物分配给各营养器官或生殖器官, 以提高获取稀缺资源的能力(Thornley, 1972; Bloom et al., 1985).相比生长在富营养环境下的植物, 那些生长在贫瘠的、低氮土壤上的植物往往倾向于将更多的资源投入到根系生长, 表现出根部具有更高的质量分数(Tilman & Wedin, 1991; Cambui et al., 2011; Sun et al., 2014).在加拿大一枝黄花氮受限条件筛选实验中, 我们发现在低浓度铵态氮处理下加拿大一枝黄花地下部分生物量显著高于对照组, 而地上部分的生物量明显低于对照组(图3).这表明加拿大一枝黄花在氮缺乏环境下可能是通过促进根系生长来扩大其在基质中的营养有效吸收面积, 从而提高自身的环境适应能力, 这可能是加拿大一枝黄花成功入侵的机制之一. ...
Mechanisms underlying beneficial plant-fungus interactions in mycorrhizal symbiosis 1 2010
... 丛枝菌根真菌(AMF)是一种普遍存在的根际微生物, 在所有陆地生态系统中70%-90%的植物能被AMF定殖(Khan, 1974; Smith & Read, 2008; Bonfante & Genre, 2010), 能在植物根部形成复杂的菌丝网络(CMNs), 与植物形成互惠共生体.这种高效的共生关系除了能够明显改善宿主植物的营养水平(氮、磷、硫等)外, 还能帮助宿主植物适应多种逆境胁迫(Smith & Read, 2008; Battini et al., 2017; Pepe et al., 2017).AMF能够利用土壤中游离的NH4+、NO3-、简单形态的氨基酸和有机氮(Hodge et al., 2001; Leigh et al., 2009; 陈永亮等, 2014), 并且会优先利用NH4+ (Tanaka & Yano, 2005), 与宿主植物交换碳水化合物(Johnson et al., 2010), 改善宿主植物对氮的吸收水平(Murugesan et al., 2014; Wang et al., 2018; Chen et al., 2020), 形成互利共生的关系(Johnson, 2010; Smith et al., 2009). ...
Grassland invaders and their mycorrhizal symbionts: a study across climate and invasion gradients 1 2014
... 多数研究表明AMF能促进许多外来植物的生长和适应性进而增强其入侵能力(Shah et al., 2008a, 2008b; Smith & Read, 2008; Bunn et al., 2014; Yang et al., 2014; Zhang et al., 2017; Chen et al., 2020), 如, 通过增加匍匐茎和气生根的数量来改变入侵植物的形态特征(Lee et al., 2014), 甚至能通过影响植物对磷的吸收来改变入侵种与本地种之间的竞争关系(Wilson et al., 2012; Chandrasekaran et al., 2014; Lee et al., 2014; Cheng et al., 2019).研究表明, 加拿大一枝黄花(Solidago canadensis)是一种菌根营养植物(Werner et al., 1980; Schmid et al., 1988; Jin et al., 2004), 它能与其入侵地的菌根真菌形成互惠共生体, 这不仅能够改变入侵地中的优势菌根真菌组成, 还能够降低入侵地中本土植物根部的AMF侵染率, 破坏AMF与本地植物的共生关系(杨贤燕, 2007; 唐建军等, 2009), 从而提高加拿大一枝黄花的入侵能力(王兵, 2013; Zhou et al., 2014). ...
Patterns of plant biomass partitioning depend on nitrogen source 1 2011
... 根际最佳分配理论认为, 植物生长过程中会因生存环境变化而对资源分配进行动态调节, 它们可以通过将光合作用制造的有机物分配给各营养器官或生殖器官, 以提高获取稀缺资源的能力(Thornley, 1972; Bloom et al., 1985).相比生长在富营养环境下的植物, 那些生长在贫瘠的、低氮土壤上的植物往往倾向于将更多的资源投入到根系生长, 表现出根部具有更高的质量分数(Tilman & Wedin, 1991; Cambui et al., 2011; Sun et al., 2014).在加拿大一枝黄花氮受限条件筛选实验中, 我们发现在低浓度铵态氮处理下加拿大一枝黄花地下部分生物量显著高于对照组, 而地上部分的生物量明显低于对照组(图3).这表明加拿大一枝黄花在氮缺乏环境下可能是通过促进根系生长来扩大其在基质中的营养有效吸收面积, 从而提高自身的环境适应能力, 这可能是加拿大一枝黄花成功入侵的机制之一. ...
A meta-analysis of arbuscular mycorrhizal effects on plants grown under salt stress 1 2014
... 多数研究表明AMF能促进许多外来植物的生长和适应性进而增强其入侵能力(Shah et al., 2008a, 2008b; Smith & Read, 2008; Bunn et al., 2014; Yang et al., 2014; Zhang et al., 2017; Chen et al., 2020), 如, 通过增加匍匐茎和气生根的数量来改变入侵植物的形态特征(Lee et al., 2014), 甚至能通过影响植物对磷的吸收来改变入侵种与本地种之间的竞争关系(Wilson et al., 2012; Chandrasekaran et al., 2014; Lee et al., 2014; Cheng et al., 2019).研究表明, 加拿大一枝黄花(Solidago canadensis)是一种菌根营养植物(Werner et al., 1980; Schmid et al., 1988; Jin et al., 2004), 它能与其入侵地的菌根真菌形成互惠共生体, 这不仅能够改变入侵地中的优势菌根真菌组成, 还能够降低入侵地中本土植物根部的AMF侵染率, 破坏AMF与本地植物的共生关系(杨贤燕, 2007; 唐建军等, 2009), 从而提高加拿大一枝黄花的入侵能力(王兵, 2013; Zhou et al., 2014). ...
Arbuscular mycorrhizal fungi improve the growth and disease resistance of the invasive plantWedelia trilobata 4 2019
... 丛枝菌根真菌(AMF)是一种普遍存在的根际微生物, 在所有陆地生态系统中70%-90%的植物能被AMF定殖(Khan, 1974; Smith & Read, 2008; Bonfante & Genre, 2010), 能在植物根部形成复杂的菌丝网络(CMNs), 与植物形成互惠共生体.这种高效的共生关系除了能够明显改善宿主植物的营养水平(氮、磷、硫等)外, 还能帮助宿主植物适应多种逆境胁迫(Smith & Read, 2008; Battini et al., 2017; Pepe et al., 2017).AMF能够利用土壤中游离的NH4+、NO3-、简单形态的氨基酸和有机氮(Hodge et al., 2001; Leigh et al., 2009; 陈永亮等, 2014), 并且会优先利用NH4+ (Tanaka & Yano, 2005), 与宿主植物交换碳水化合物(Johnson et al., 2010), 改善宿主植物对氮的吸收水平(Murugesan et al., 2014; Wang et al., 2018; Chen et al., 2020), 形成互利共生的关系(Johnson, 2010; Smith et al., 2009). ...
... 多数研究表明AMF能促进许多外来植物的生长和适应性进而增强其入侵能力(Shah et al., 2008a, 2008b; Smith & Read, 2008; Bunn et al., 2014; Yang et al., 2014; Zhang et al., 2017; Chen et al., 2020), 如, 通过增加匍匐茎和气生根的数量来改变入侵植物的形态特征(Lee et al., 2014), 甚至能通过影响植物对磷的吸收来改变入侵种与本地种之间的竞争关系(Wilson et al., 2012; Chandrasekaran et al., 2014; Lee et al., 2014; Cheng et al., 2019).研究表明, 加拿大一枝黄花(Solidago canadensis)是一种菌根营养植物(Werner et al., 1980; Schmid et al., 1988; Jin et al., 2004), 它能与其入侵地的菌根真菌形成互惠共生体, 这不仅能够改变入侵地中的优势菌根真菌组成, 还能够降低入侵地中本土植物根部的AMF侵染率, 破坏AMF与本地植物的共生关系(杨贤燕, 2007; 唐建军等, 2009), 从而提高加拿大一枝黄花的入侵能力(王兵, 2013; Zhou et al., 2014). ...
... 丛枝菌根真菌(AMF)是一种普遍存在的根际微生物, 在所有陆地生态系统中70%-90%的植物能被AMF定殖(Khan, 1974; Smith & Read, 2008; Bonfante & Genre, 2010), 能在植物根部形成复杂的菌丝网络(CMNs), 与植物形成互惠共生体.这种高效的共生关系除了能够明显改善宿主植物的营养水平(氮、磷、硫等)外, 还能帮助宿主植物适应多种逆境胁迫(Smith & Read, 2008; Battini et al., 2017; Pepe et al., 2017).AMF能够利用土壤中游离的NH4+、NO3-、简单形态的氨基酸和有机氮(Hodge et al., 2001; Leigh et al., 2009; 陈永亮等, 2014), 并且会优先利用NH4+ (Tanaka & Yano, 2005), 与宿主植物交换碳水化合物(Johnson et al., 2010), 改善宿主植物对氮的吸收水平(Murugesan et al., 2014; Wang et al., 2018; Chen et al., 2020), 形成互利共生的关系(Johnson, 2010; Smith et al., 2009). ...
丛枝菌根真菌在土壤氮素循环中的作用 1 2014
... 丛枝菌根真菌(AMF)是一种普遍存在的根际微生物, 在所有陆地生态系统中70%-90%的植物能被AMF定殖(Khan, 1974; Smith & Read, 2008; Bonfante & Genre, 2010), 能在植物根部形成复杂的菌丝网络(CMNs), 与植物形成互惠共生体.这种高效的共生关系除了能够明显改善宿主植物的营养水平(氮、磷、硫等)外, 还能帮助宿主植物适应多种逆境胁迫(Smith & Read, 2008; Battini et al., 2017; Pepe et al., 2017).AMF能够利用土壤中游离的NH4+、NO3-、简单形态的氨基酸和有机氮(Hodge et al., 2001; Leigh et al., 2009; 陈永亮等, 2014), 并且会优先利用NH4+ (Tanaka & Yano, 2005), 与宿主植物交换碳水化合物(Johnson et al., 2010), 改善宿主植物对氮的吸收水平(Murugesan et al., 2014; Wang et al., 2018; Chen et al., 2020), 形成互利共生的关系(Johnson, 2010; Smith et al., 2009). ...
Do arbuscular mycorrhizal fungi help the native speciesBidens biternata resist the invasion of Bidens alba? 1 2019
... 多数研究表明AMF能促进许多外来植物的生长和适应性进而增强其入侵能力(Shah et al., 2008a, 2008b; Smith & Read, 2008; Bunn et al., 2014; Yang et al., 2014; Zhang et al., 2017; Chen et al., 2020), 如, 通过增加匍匐茎和气生根的数量来改变入侵植物的形态特征(Lee et al., 2014), 甚至能通过影响植物对磷的吸收来改变入侵种与本地种之间的竞争关系(Wilson et al., 2012; Chandrasekaran et al., 2014; Lee et al., 2014; Cheng et al., 2019).研究表明, 加拿大一枝黄花(Solidago canadensis)是一种菌根营养植物(Werner et al., 1980; Schmid et al., 1988; Jin et al., 2004), 它能与其入侵地的菌根真菌形成互惠共生体, 这不仅能够改变入侵地中的优势菌根真菌组成, 还能够降低入侵地中本土植物根部的AMF侵染率, 破坏AMF与本地植物的共生关系(杨贤燕, 2007; 唐建军等, 2009), 从而提高加拿大一枝黄花的入侵能力(王兵, 2013; Zhou et al., 2014). ...
The history of Solidago canadensis invasion and the development of its mycorrhizal associations in newly-reclaimed land 1 2004
... 多数研究表明AMF能促进许多外来植物的生长和适应性进而增强其入侵能力(Shah et al., 2008a, 2008b; Smith & Read, 2008; Bunn et al., 2014; Yang et al., 2014; Zhang et al., 2017; Chen et al., 2020), 如, 通过增加匍匐茎和气生根的数量来改变入侵植物的形态特征(Lee et al., 2014), 甚至能通过影响植物对磷的吸收来改变入侵种与本地种之间的竞争关系(Wilson et al., 2012; Chandrasekaran et al., 2014; Lee et al., 2014; Cheng et al., 2019).研究表明, 加拿大一枝黄花(Solidago canadensis)是一种菌根营养植物(Werner et al., 1980; Schmid et al., 1988; Jin et al., 2004), 它能与其入侵地的菌根真菌形成互惠共生体, 这不仅能够改变入侵地中的优势菌根真菌组成, 还能够降低入侵地中本土植物根部的AMF侵染率, 破坏AMF与本地植物的共生关系(杨贤燕, 2007; 唐建军等, 2009), 从而提高加拿大一枝黄花的入侵能力(王兵, 2013; Zhou et al., 2014). ...
Resource limitation is a driver of local adaptation in mycorrhizal symbioses 2 2010
... 丛枝菌根真菌(AMF)是一种普遍存在的根际微生物, 在所有陆地生态系统中70%-90%的植物能被AMF定殖(Khan, 1974; Smith & Read, 2008; Bonfante & Genre, 2010), 能在植物根部形成复杂的菌丝网络(CMNs), 与植物形成互惠共生体.这种高效的共生关系除了能够明显改善宿主植物的营养水平(氮、磷、硫等)外, 还能帮助宿主植物适应多种逆境胁迫(Smith & Read, 2008; Battini et al., 2017; Pepe et al., 2017).AMF能够利用土壤中游离的NH4+、NO3-、简单形态的氨基酸和有机氮(Hodge et al., 2001; Leigh et al., 2009; 陈永亮等, 2014), 并且会优先利用NH4+ (Tanaka & Yano, 2005), 与宿主植物交换碳水化合物(Johnson et al., 2010), 改善宿主植物对氮的吸收水平(Murugesan et al., 2014; Wang et al., 2018; Chen et al., 2020), 形成互利共生的关系(Johnson, 2010; Smith et al., 2009). ...
Resource stoichiometry elucidates the structure and function of arbuscular mycorrhizas across scales 1 2010
... 丛枝菌根真菌(AMF)是一种普遍存在的根际微生物, 在所有陆地生态系统中70%-90%的植物能被AMF定殖(Khan, 1974; Smith & Read, 2008; Bonfante & Genre, 2010), 能在植物根部形成复杂的菌丝网络(CMNs), 与植物形成互惠共生体.这种高效的共生关系除了能够明显改善宿主植物的营养水平(氮、磷、硫等)外, 还能帮助宿主植物适应多种逆境胁迫(Smith & Read, 2008; Battini et al., 2017; Pepe et al., 2017).AMF能够利用土壤中游离的NH4+、NO3-、简单形态的氨基酸和有机氮(Hodge et al., 2001; Leigh et al., 2009; 陈永亮等, 2014), 并且会优先利用NH4+ (Tanaka & Yano, 2005), 与宿主植物交换碳水化合物(Johnson et al., 2010), 改善宿主植物对氮的吸收水平(Murugesan et al., 2014; Wang et al., 2018; Chen et al., 2020), 形成互利共生的关系(Johnson, 2010; Smith et al., 2009). ...
The occurrence of mycorrhizas in halophytes, hydrophytes and xerophytes, and of Endogone spores in adjacent soils 1 1974
... 丛枝菌根真菌(AMF)是一种普遍存在的根际微生物, 在所有陆地生态系统中70%-90%的植物能被AMF定殖(Khan, 1974; Smith & Read, 2008; Bonfante & Genre, 2010), 能在植物根部形成复杂的菌丝网络(CMNs), 与植物形成互惠共生体.这种高效的共生关系除了能够明显改善宿主植物的营养水平(氮、磷、硫等)外, 还能帮助宿主植物适应多种逆境胁迫(Smith & Read, 2008; Battini et al., 2017; Pepe et al., 2017).AMF能够利用土壤中游离的NH4+、NO3-、简单形态的氨基酸和有机氮(Hodge et al., 2001; Leigh et al., 2009; 陈永亮等, 2014), 并且会优先利用NH4+ (Tanaka & Yano, 2005), 与宿主植物交换碳水化合物(Johnson et al., 2010), 改善宿主植物对氮的吸收水平(Murugesan et al., 2014; Wang et al., 2018; Chen et al., 2020), 形成互利共生的关系(Johnson, 2010; Smith et al., 2009). ...
Arbuscular mycorrhizal fungi enhance P uptake and alter plant morphology in the invasive plant Microstegium vimineum 2 2014
... 多数研究表明AMF能促进许多外来植物的生长和适应性进而增强其入侵能力(Shah et al., 2008a, 2008b; Smith & Read, 2008; Bunn et al., 2014; Yang et al., 2014; Zhang et al., 2017; Chen et al., 2020), 如, 通过增加匍匐茎和气生根的数量来改变入侵植物的形态特征(Lee et al., 2014), 甚至能通过影响植物对磷的吸收来改变入侵种与本地种之间的竞争关系(Wilson et al., 2012; Chandrasekaran et al., 2014; Lee et al., 2014; Cheng et al., 2019).研究表明, 加拿大一枝黄花(Solidago canadensis)是一种菌根营养植物(Werner et al., 1980; Schmid et al., 1988; Jin et al., 2004), 它能与其入侵地的菌根真菌形成互惠共生体, 这不仅能够改变入侵地中的优势菌根真菌组成, 还能够降低入侵地中本土植物根部的AMF侵染率, 破坏AMF与本地植物的共生关系(杨贤燕, 2007; 唐建军等, 2009), 从而提高加拿大一枝黄花的入侵能力(王兵, 2013; Zhou et al., 2014). ...
... ; Lee et al., 2014; Cheng et al., 2019).研究表明, 加拿大一枝黄花(Solidago canadensis)是一种菌根营养植物(Werner et al., 1980; Schmid et al., 1988; Jin et al., 2004), 它能与其入侵地的菌根真菌形成互惠共生体, 这不仅能够改变入侵地中的优势菌根真菌组成, 还能够降低入侵地中本土植物根部的AMF侵染率, 破坏AMF与本地植物的共生关系(杨贤燕, 2007; 唐建军等, 2009), 从而提高加拿大一枝黄花的入侵能力(王兵, 2013; Zhou et al., 2014). ...
Arbuscular mycorrhizal fungi can transfer substantial amounts of nitrogen to their host plant from organic material 1 2009
... 丛枝菌根真菌(AMF)是一种普遍存在的根际微生物, 在所有陆地生态系统中70%-90%的植物能被AMF定殖(Khan, 1974; Smith & Read, 2008; Bonfante & Genre, 2010), 能在植物根部形成复杂的菌丝网络(CMNs), 与植物形成互惠共生体.这种高效的共生关系除了能够明显改善宿主植物的营养水平(氮、磷、硫等)外, 还能帮助宿主植物适应多种逆境胁迫(Smith & Read, 2008; Battini et al., 2017; Pepe et al., 2017).AMF能够利用土壤中游离的NH4+、NO3-、简单形态的氨基酸和有机氮(Hodge et al., 2001; Leigh et al., 2009; 陈永亮等, 2014), 并且会优先利用NH4+ (Tanaka & Yano, 2005), 与宿主植物交换碳水化合物(Johnson et al., 2010), 改善宿主植物对氮的吸收水平(Murugesan et al., 2014; Wang et al., 2018; Chen et al., 2020), 形成互利共生的关系(Johnson, 2010; Smith et al., 2009). ...
Clonal integration and effects of simulated herbivory in old-field perennials 1 1988
... 多数研究表明AMF能促进许多外来植物的生长和适应性进而增强其入侵能力(Shah et al., 2008a, 2008b; Smith & Read, 2008; Bunn et al., 2014; Yang et al., 2014; Zhang et al., 2017; Chen et al., 2020), 如, 通过增加匍匐茎和气生根的数量来改变入侵植物的形态特征(Lee et al., 2014), 甚至能通过影响植物对磷的吸收来改变入侵种与本地种之间的竞争关系(Wilson et al., 2012; Chandrasekaran et al., 2014; Lee et al., 2014; Cheng et al., 2019).研究表明, 加拿大一枝黄花(Solidago canadensis)是一种菌根营养植物(Werner et al., 1980; Schmid et al., 1988; Jin et al., 2004), 它能与其入侵地的菌根真菌形成互惠共生体, 这不仅能够改变入侵地中的优势菌根真菌组成, 还能够降低入侵地中本土植物根部的AMF侵染率, 破坏AMF与本地植物的共生关系(杨贤燕, 2007; 唐建军等, 2009), 从而提高加拿大一枝黄花的入侵能力(王兵, 2013; Zhou et al., 2014). ...
Mycorrhizal source and neighbour identity differently influence Anthemis cotula L. invasion in the Kashmir Himalaya, India 1 2008
... 多数研究表明AMF能促进许多外来植物的生长和适应性进而增强其入侵能力(Shah et al., 2008a, 2008b; Smith & Read, 2008; Bunn et al., 2014; Yang et al., 2014; Zhang et al., 2017; Chen et al., 2020), 如, 通过增加匍匐茎和气生根的数量来改变入侵植物的形态特征(Lee et al., 2014), 甚至能通过影响植物对磷的吸收来改变入侵种与本地种之间的竞争关系(Wilson et al., 2012; Chandrasekaran et al., 2014; Lee et al., 2014; Cheng et al., 2019).研究表明, 加拿大一枝黄花(Solidago canadensis)是一种菌根营养植物(Werner et al., 1980; Schmid et al., 1988; Jin et al., 2004), 它能与其入侵地的菌根真菌形成互惠共生体, 这不仅能够改变入侵地中的优势菌根真菌组成, 还能够降低入侵地中本土植物根部的AMF侵染率, 破坏AMF与本地植物的共生关系(杨贤燕, 2007; 唐建军等, 2009), 从而提高加拿大一枝黄花的入侵能力(王兵, 2013; Zhou et al., 2014). ...
Mycorrhizosphere mediated Mayweed chamomile invasion in the Kashmir himalaya, India 1 2008
... 多数研究表明AMF能促进许多外来植物的生长和适应性进而增强其入侵能力(Shah et al., 2008a, 2008b; Smith & Read, 2008; Bunn et al., 2014; Yang et al., 2014; Zhang et al., 2017; Chen et al., 2020), 如, 通过增加匍匐茎和气生根的数量来改变入侵植物的形态特征(Lee et al., 2014), 甚至能通过影响植物对磷的吸收来改变入侵种与本地种之间的竞争关系(Wilson et al., 2012; Chandrasekaran et al., 2014; Lee et al., 2014; Cheng et al., 2019).研究表明, 加拿大一枝黄花(Solidago canadensis)是一种菌根营养植物(Werner et al., 1980; Schmid et al., 1988; Jin et al., 2004), 它能与其入侵地的菌根真菌形成互惠共生体, 这不仅能够改变入侵地中的优势菌根真菌组成, 还能够降低入侵地中本土植物根部的AMF侵染率, 破坏AMF与本地植物的共生关系(杨贤燕, 2007; 唐建军等, 2009), 从而提高加拿大一枝黄花的入侵能力(王兵, 2013; Zhou et al., 2014). ...
More than a carbon economy: nutrient trade and ecological sustainability in facultative arbuscular mycorrhizal symbioses 1 2009
... 丛枝菌根真菌(AMF)是一种普遍存在的根际微生物, 在所有陆地生态系统中70%-90%的植物能被AMF定殖(Khan, 1974; Smith & Read, 2008; Bonfante & Genre, 2010), 能在植物根部形成复杂的菌丝网络(CMNs), 与植物形成互惠共生体.这种高效的共生关系除了能够明显改善宿主植物的营养水平(氮、磷、硫等)外, 还能帮助宿主植物适应多种逆境胁迫(Smith & Read, 2008; Battini et al., 2017; Pepe et al., 2017).AMF能够利用土壤中游离的NH4+、NO3-、简单形态的氨基酸和有机氮(Hodge et al., 2001; Leigh et al., 2009; 陈永亮等, 2014), 并且会优先利用NH4+ (Tanaka & Yano, 2005), 与宿主植物交换碳水化合物(Johnson et al., 2010), 改善宿主植物对氮的吸收水平(Murugesan et al., 2014; Wang et al., 2018; Chen et al., 2020), 形成互利共生的关系(Johnson, 2010; Smith et al., 2009). ...
3 2008
... 丛枝菌根真菌(AMF)是一种普遍存在的根际微生物, 在所有陆地生态系统中70%-90%的植物能被AMF定殖(Khan, 1974; Smith & Read, 2008; Bonfante & Genre, 2010), 能在植物根部形成复杂的菌丝网络(CMNs), 与植物形成互惠共生体.这种高效的共生关系除了能够明显改善宿主植物的营养水平(氮、磷、硫等)外, 还能帮助宿主植物适应多种逆境胁迫(Smith & Read, 2008; Battini et al., 2017; Pepe et al., 2017).AMF能够利用土壤中游离的NH4+、NO3-、简单形态的氨基酸和有机氮(Hodge et al., 2001; Leigh et al., 2009; 陈永亮等, 2014), 并且会优先利用NH4+ (Tanaka & Yano, 2005), 与宿主植物交换碳水化合物(Johnson et al., 2010), 改善宿主植物对氮的吸收水平(Murugesan et al., 2014; Wang et al., 2018; Chen et al., 2020), 形成互利共生的关系(Johnson, 2010; Smith et al., 2009). ...
... ), 能在植物根部形成复杂的菌丝网络(CMNs), 与植物形成互惠共生体.这种高效的共生关系除了能够明显改善宿主植物的营养水平(氮、磷、硫等)外, 还能帮助宿主植物适应多种逆境胁迫(Smith & Read, 2008; Battini et al., 2017; Pepe et al., 2017).AMF能够利用土壤中游离的NH4+、NO3-、简单形态的氨基酸和有机氮(Hodge et al., 2001; Leigh et al., 2009; 陈永亮等, 2014), 并且会优先利用NH4+ (Tanaka & Yano, 2005), 与宿主植物交换碳水化合物(Johnson et al., 2010), 改善宿主植物对氮的吸收水平(Murugesan et al., 2014; Wang et al., 2018; Chen et al., 2020), 形成互利共生的关系(Johnson, 2010; Smith et al., 2009). ...
... 多数研究表明AMF能促进许多外来植物的生长和适应性进而增强其入侵能力(Shah et al., 2008a, 2008b; Smith & Read, 2008; Bunn et al., 2014; Yang et al., 2014; Zhang et al., 2017; Chen et al., 2020), 如, 通过增加匍匐茎和气生根的数量来改变入侵植物的形态特征(Lee et al., 2014), 甚至能通过影响植物对磷的吸收来改变入侵种与本地种之间的竞争关系(Wilson et al., 2012; Chandrasekaran et al., 2014; Lee et al., 2014; Cheng et al., 2019).研究表明, 加拿大一枝黄花(Solidago canadensis)是一种菌根营养植物(Werner et al., 1980; Schmid et al., 1988; Jin et al., 2004), 它能与其入侵地的菌根真菌形成互惠共生体, 这不仅能够改变入侵地中的优势菌根真菌组成, 还能够降低入侵地中本土植物根部的AMF侵染率, 破坏AMF与本地植物的共生关系(杨贤燕, 2007; 唐建军等, 2009), 从而提高加拿大一枝黄花的入侵能力(王兵, 2013; Zhou et al., 2014). ...
Roles of arbuscular mycorrhizas in plant nutrition and growth: new paradigms from cellular to ecosystem scales 1 2011
... 多数研究表明AMF能促进许多外来植物的生长和适应性进而增强其入侵能力(Shah et al., 2008a, 2008b; Smith & Read, 2008; Bunn et al., 2014; Yang et al., 2014; Zhang et al., 2017; Chen et al., 2020), 如, 通过增加匍匐茎和气生根的数量来改变入侵植物的形态特征(Lee et al., 2014), 甚至能通过影响植物对磷的吸收来改变入侵种与本地种之间的竞争关系(Wilson et al., 2012; Chandrasekaran et al., 2014; Lee et al., 2014; Cheng et al., 2019).研究表明, 加拿大一枝黄花(Solidago canadensis)是一种菌根营养植物(Werner et al., 1980; Schmid et al., 1988; Jin et al., 2004), 它能与其入侵地的菌根真菌形成互惠共生体, 这不仅能够改变入侵地中的优势菌根真菌组成, 还能够降低入侵地中本土植物根部的AMF侵染率, 破坏AMF与本地植物的共生关系(杨贤燕, 2007; 唐建军等, 2009), 从而提高加拿大一枝黄花的入侵能力(王兵, 2013; Zhou et al., 2014). ...
外来植物加拿大一枝黄花对入侵地丛枝菌根真菌(AMF)的影响 1 2009
... 多数研究表明AMF能促进许多外来植物的生长和适应性进而增强其入侵能力(Shah et al., 2008a, 2008b; Smith & Read, 2008; Bunn et al., 2014; Yang et al., 2014; Zhang et al., 2017; Chen et al., 2020), 如, 通过增加匍匐茎和气生根的数量来改变入侵植物的形态特征(Lee et al., 2014), 甚至能通过影响植物对磷的吸收来改变入侵种与本地种之间的竞争关系(Wilson et al., 2012; Chandrasekaran et al., 2014; Lee et al., 2014; Cheng et al., 2019).研究表明, 加拿大一枝黄花(Solidago canadensis)是一种菌根营养植物(Werner et al., 1980; Schmid et al., 1988; Jin et al., 2004), 它能与其入侵地的菌根真菌形成互惠共生体, 这不仅能够改变入侵地中的优势菌根真菌组成, 还能够降低入侵地中本土植物根部的AMF侵染率, 破坏AMF与本地植物的共生关系(杨贤燕, 2007; 唐建军等, 2009), 从而提高加拿大一枝黄花的入侵能力(王兵, 2013; Zhou et al., 2014). ...
A balanced quantitative model for root:shoot ratios in vegetative plants 1 1972
... 根际最佳分配理论认为, 植物生长过程中会因生存环境变化而对资源分配进行动态调节, 它们可以通过将光合作用制造的有机物分配给各营养器官或生殖器官, 以提高获取稀缺资源的能力(Thornley, 1972; Bloom et al., 1985).相比生长在富营养环境下的植物, 那些生长在贫瘠的、低氮土壤上的植物往往倾向于将更多的资源投入到根系生长, 表现出根部具有更高的质量分数(Tilman & Wedin, 1991; Cambui et al., 2011; Sun et al., 2014).在加拿大一枝黄花氮受限条件筛选实验中, 我们发现在低浓度铵态氮处理下加拿大一枝黄花地下部分生物量显著高于对照组, 而地上部分的生物量明显低于对照组(图3).这表明加拿大一枝黄花在氮缺乏环境下可能是通过促进根系生长来扩大其在基质中的营养有效吸收面积, 从而提高自身的环境适应能力, 这可能是加拿大一枝黄花成功入侵的机制之一. ...
Plant traits and resource reduction for five grasses growing on a nitrogen gradient 1 1991
... 根际最佳分配理论认为, 植物生长过程中会因生存环境变化而对资源分配进行动态调节, 它们可以通过将光合作用制造的有机物分配给各营养器官或生殖器官, 以提高获取稀缺资源的能力(Thornley, 1972; Bloom et al., 1985).相比生长在富营养环境下的植物, 那些生长在贫瘠的、低氮土壤上的植物往往倾向于将更多的资源投入到根系生长, 表现出根部具有更高的质量分数(Tilman & Wedin, 1991; Cambui et al., 2011; Sun et al., 2014).在加拿大一枝黄花氮受限条件筛选实验中, 我们发现在低浓度铵态氮处理下加拿大一枝黄花地下部分生物量显著高于对照组, 而地上部分的生物量明显低于对照组(图3).这表明加拿大一枝黄花在氮缺乏环境下可能是通过促进根系生长来扩大其在基质中的营养有效吸收面积, 从而提高自身的环境适应能力, 这可能是加拿大一枝黄花成功入侵的机制之一. ...
化感作用与丛枝菌根在加拿大一枝黄花入侵过程中的作用研究 1 2013
... 多数研究表明AMF能促进许多外来植物的生长和适应性进而增强其入侵能力(Shah et al., 2008a, 2008b; Smith & Read, 2008; Bunn et al., 2014; Yang et al., 2014; Zhang et al., 2017; Chen et al., 2020), 如, 通过增加匍匐茎和气生根的数量来改变入侵植物的形态特征(Lee et al., 2014), 甚至能通过影响植物对磷的吸收来改变入侵种与本地种之间的竞争关系(Wilson et al., 2012; Chandrasekaran et al., 2014; Lee et al., 2014; Cheng et al., 2019).研究表明, 加拿大一枝黄花(Solidago canadensis)是一种菌根营养植物(Werner et al., 1980; Schmid et al., 1988; Jin et al., 2004), 它能与其入侵地的菌根真菌形成互惠共生体, 这不仅能够改变入侵地中的优势菌根真菌组成, 还能够降低入侵地中本土植物根部的AMF侵染率, 破坏AMF与本地植物的共生关系(杨贤燕, 2007; 唐建军等, 2009), 从而提高加拿大一枝黄花的入侵能力(王兵, 2013; Zhou et al., 2014). ...
化感作用与丛枝菌根在加拿大一枝黄花入侵过程中的作用研究 1 2013
... 多数研究表明AMF能促进许多外来植物的生长和适应性进而增强其入侵能力(Shah et al., 2008a, 2008b; Smith & Read, 2008; Bunn et al., 2014; Yang et al., 2014; Zhang et al., 2017; Chen et al., 2020), 如, 通过增加匍匐茎和气生根的数量来改变入侵植物的形态特征(Lee et al., 2014), 甚至能通过影响植物对磷的吸收来改变入侵种与本地种之间的竞争关系(Wilson et al., 2012; Chandrasekaran et al., 2014; Lee et al., 2014; Cheng et al., 2019).研究表明, 加拿大一枝黄花(Solidago canadensis)是一种菌根营养植物(Werner et al., 1980; Schmid et al., 1988; Jin et al., 2004), 它能与其入侵地的菌根真菌形成互惠共生体, 这不仅能够改变入侵地中的优势菌根真菌组成, 还能够降低入侵地中本土植物根部的AMF侵染率, 破坏AMF与本地植物的共生关系(杨贤燕, 2007; 唐建军等, 2009), 从而提高加拿大一枝黄花的入侵能力(王兵, 2013; Zhou et al., 2014). ...
Effects of arbuscular mycorrhizal fungi on growth and nitrogen uptake of Chrysanthemum morifolium under salt stress 1 2018
... 丛枝菌根真菌(AMF)是一种普遍存在的根际微生物, 在所有陆地生态系统中70%-90%的植物能被AMF定殖(Khan, 1974; Smith & Read, 2008; Bonfante & Genre, 2010), 能在植物根部形成复杂的菌丝网络(CMNs), 与植物形成互惠共生体.这种高效的共生关系除了能够明显改善宿主植物的营养水平(氮、磷、硫等)外, 还能帮助宿主植物适应多种逆境胁迫(Smith & Read, 2008; Battini et al., 2017; Pepe et al., 2017).AMF能够利用土壤中游离的NH4+、NO3-、简单形态的氨基酸和有机氮(Hodge et al., 2001; Leigh et al., 2009; 陈永亮等, 2014), 并且会优先利用NH4+ (Tanaka & Yano, 2005), 与宿主植物交换碳水化合物(Johnson et al., 2010), 改善宿主植物对氮的吸收水平(Murugesan et al., 2014; Wang et al., 2018; Chen et al., 2020), 形成互利共生的关系(Johnson, 2010; Smith et al., 2009). ...
The biology of Canadian.: 45.Solidago canadensis L 1 1980
... 多数研究表明AMF能促进许多外来植物的生长和适应性进而增强其入侵能力(Shah et al., 2008a, 2008b; Smith & Read, 2008; Bunn et al., 2014; Yang et al., 2014; Zhang et al., 2017; Chen et al., 2020), 如, 通过增加匍匐茎和气生根的数量来改变入侵植物的形态特征(Lee et al., 2014), 甚至能通过影响植物对磷的吸收来改变入侵种与本地种之间的竞争关系(Wilson et al., 2012; Chandrasekaran et al., 2014; Lee et al., 2014; Cheng et al., 2019).研究表明, 加拿大一枝黄花(Solidago canadensis)是一种菌根营养植物(Werner et al., 1980; Schmid et al., 1988; Jin et al., 2004), 它能与其入侵地的菌根真菌形成互惠共生体, 这不仅能够改变入侵地中的优势菌根真菌组成, 还能够降低入侵地中本土植物根部的AMF侵染率, 破坏AMF与本地植物的共生关系(杨贤燕, 2007; 唐建军等, 2009), 从而提高加拿大一枝黄花的入侵能力(王兵, 2013; Zhou et al., 2014). ...
Invasive warm-season grasses reduce mycorrhizal root colonization and biomass production of native prairie grasses 1 2012
... 多数研究表明AMF能促进许多外来植物的生长和适应性进而增强其入侵能力(Shah et al., 2008a, 2008b; Smith & Read, 2008; Bunn et al., 2014; Yang et al., 2014; Zhang et al., 2017; Chen et al., 2020), 如, 通过增加匍匐茎和气生根的数量来改变入侵植物的形态特征(Lee et al., 2014), 甚至能通过影响植物对磷的吸收来改变入侵种与本地种之间的竞争关系(Wilson et al., 2012; Chandrasekaran et al., 2014; Lee et al., 2014; Cheng et al., 2019).研究表明, 加拿大一枝黄花(Solidago canadensis)是一种菌根营养植物(Werner et al., 1980; Schmid et al., 1988; Jin et al., 2004), 它能与其入侵地的菌根真菌形成互惠共生体, 这不仅能够改变入侵地中的优势菌根真菌组成, 还能够降低入侵地中本土植物根部的AMF侵染率, 破坏AMF与本地植物的共生关系(杨贤燕, 2007; 唐建军等, 2009), 从而提高加拿大一枝黄花的入侵能力(王兵, 2013; Zhou et al., 2014). ...
Arbuscular mycorrhizal fungi facilitate the invasion of Solidago canadensis L. in southeastern China 1 2014
... 多数研究表明AMF能促进许多外来植物的生长和适应性进而增强其入侵能力(Shah et al., 2008a, 2008b; Smith & Read, 2008; Bunn et al., 2014; Yang et al., 2014; Zhang et al., 2017; Chen et al., 2020), 如, 通过增加匍匐茎和气生根的数量来改变入侵植物的形态特征(Lee et al., 2014), 甚至能通过影响植物对磷的吸收来改变入侵种与本地种之间的竞争关系(Wilson et al., 2012; Chandrasekaran et al., 2014; Lee et al., 2014; Cheng et al., 2019).研究表明, 加拿大一枝黄花(Solidago canadensis)是一种菌根营养植物(Werner et al., 1980; Schmid et al., 1988; Jin et al., 2004), 它能与其入侵地的菌根真菌形成互惠共生体, 这不仅能够改变入侵地中的优势菌根真菌组成, 还能够降低入侵地中本土植物根部的AMF侵染率, 破坏AMF与本地植物的共生关系(杨贤燕, 2007; 唐建军等, 2009), 从而提高加拿大一枝黄花的入侵能力(王兵, 2013; Zhou et al., 2014). ...
外来杂草对本地植物丛枝菌根的化感作用 1 2007
... 多数研究表明AMF能促进许多外来植物的生长和适应性进而增强其入侵能力(Shah et al., 2008a, 2008b; Smith & Read, 2008; Bunn et al., 2014; Yang et al., 2014; Zhang et al., 2017; Chen et al., 2020), 如, 通过增加匍匐茎和气生根的数量来改变入侵植物的形态特征(Lee et al., 2014), 甚至能通过影响植物对磷的吸收来改变入侵种与本地种之间的竞争关系(Wilson et al., 2012; Chandrasekaran et al., 2014; Lee et al., 2014; Cheng et al., 2019).研究表明, 加拿大一枝黄花(Solidago canadensis)是一种菌根营养植物(Werner et al., 1980; Schmid et al., 1988; Jin et al., 2004), 它能与其入侵地的菌根真菌形成互惠共生体, 这不仅能够改变入侵地中的优势菌根真菌组成, 还能够降低入侵地中本土植物根部的AMF侵染率, 破坏AMF与本地植物的共生关系(杨贤燕, 2007; 唐建军等, 2009), 从而提高加拿大一枝黄花的入侵能力(王兵, 2013; Zhou et al., 2014). ...
外来杂草对本地植物丛枝菌根的化感作用 1 2007
... 多数研究表明AMF能促进许多外来植物的生长和适应性进而增强其入侵能力(Shah et al., 2008a, 2008b; Smith & Read, 2008; Bunn et al., 2014; Yang et al., 2014; Zhang et al., 2017; Chen et al., 2020), 如, 通过增加匍匐茎和气生根的数量来改变入侵植物的形态特征(Lee et al., 2014), 甚至能通过影响植物对磷的吸收来改变入侵种与本地种之间的竞争关系(Wilson et al., 2012; Chandrasekaran et al., 2014; Lee et al., 2014; Cheng et al., 2019).研究表明, 加拿大一枝黄花(Solidago canadensis)是一种菌根营养植物(Werner et al., 1980; Schmid et al., 1988; Jin et al., 2004), 它能与其入侵地的菌根真菌形成互惠共生体, 这不仅能够改变入侵地中的优势菌根真菌组成, 还能够降低入侵地中本土植物根部的AMF侵染率, 破坏AMF与本地植物的共生关系(杨贤燕, 2007; 唐建军等, 2009), 从而提高加拿大一枝黄花的入侵能力(王兵, 2013; Zhou et al., 2014). ...
Arbuscular mycorrhizal fungi facilitate growth and competitive ability of an exotic species Flaveria bidentis 1 2017
... 多数研究表明AMF能促进许多外来植物的生长和适应性进而增强其入侵能力(Shah et al., 2008a, 2008b; Smith & Read, 2008; Bunn et al., 2014; Yang et al., 2014; Zhang et al., 2017; Chen et al., 2020), 如, 通过增加匍匐茎和气生根的数量来改变入侵植物的形态特征(Lee et al., 2014), 甚至能通过影响植物对磷的吸收来改变入侵种与本地种之间的竞争关系(Wilson et al., 2012; Chandrasekaran et al., 2014; Lee et al., 2014; Cheng et al., 2019).研究表明, 加拿大一枝黄花(Solidago canadensis)是一种菌根营养植物(Werner et al., 1980; Schmid et al., 1988; Jin et al., 2004), 它能与其入侵地的菌根真菌形成互惠共生体, 这不仅能够改变入侵地中的优势菌根真菌组成, 还能够降低入侵地中本土植物根部的AMF侵染率, 破坏AMF与本地植物的共生关系(杨贤燕, 2007; 唐建军等, 2009), 从而提高加拿大一枝黄花的入侵能力(王兵, 2013; Zhou et al., 2014). ...
Arbuscular mycorrhizal fungi facilitate the invasion of Solidago canadensis L. in southeastern China 1 2014
... 多数研究表明AMF能促进许多外来植物的生长和适应性进而增强其入侵能力(Shah et al., 2008a, 2008b; Smith & Read, 2008; Bunn et al., 2014; Yang et al., 2014; Zhang et al., 2017; Chen et al., 2020), 如, 通过增加匍匐茎和气生根的数量来改变入侵植物的形态特征(Lee et al., 2014), 甚至能通过影响植物对磷的吸收来改变入侵种与本地种之间的竞争关系(Wilson et al., 2012; Chandrasekaran et al., 2014; Lee et al., 2014; Cheng et al., 2019).研究表明, 加拿大一枝黄花(Solidago canadensis)是一种菌根营养植物(Werner et al., 1980; Schmid et al., 1988; Jin et al., 2004), 它能与其入侵地的菌根真菌形成互惠共生体, 这不仅能够改变入侵地中的优势菌根真菌组成, 还能够降低入侵地中本土植物根部的AMF侵染率, 破坏AMF与本地植物的共生关系(杨贤燕, 2007; 唐建军等, 2009), 从而提高加拿大一枝黄花的入侵能力(王兵, 2013; Zhou et al., 2014). ...
N-fertilizer-driven association between the arbuscular mycorrhizal fungal community and diazotrophic community impacts wheat yield 2 2018
 ,
, ,1,2,*, 杜道林1
,1,2,*, 杜道林1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT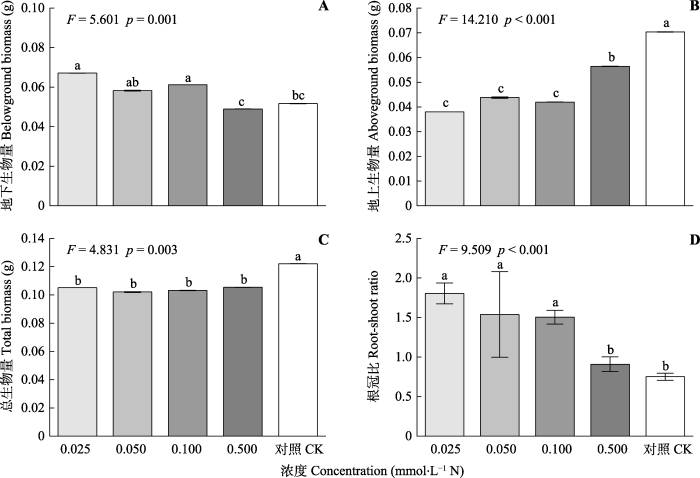 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT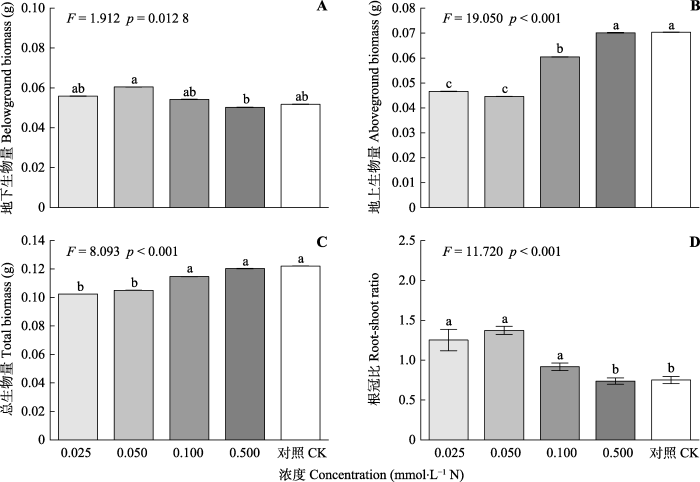 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT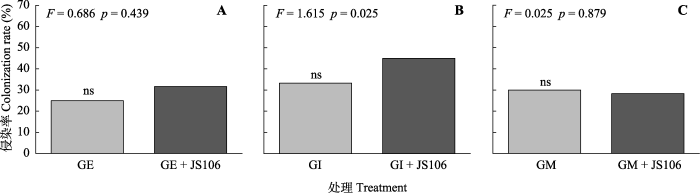 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT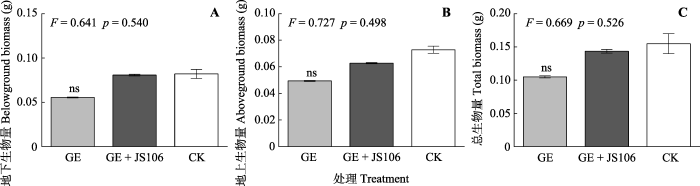 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT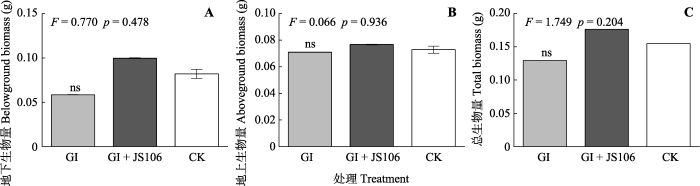 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT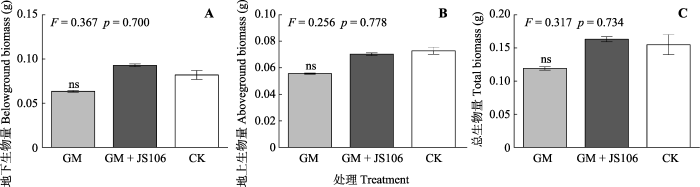 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}