刘凌
1 , 樊英杰
1 , 宋晓彤
1 , 李敏
2 , 邵小明
1 , 王晓蕊
, 3 , * 1 中国农业大学资源与环境学院, 生物多样性与有机农业北京市重点实验室, 北京 1001932 河北师范大学生命科学学院, 石家庄 0500243 石家庄学院资源与环境科学学院, 石家庄 050035Bryophyte societies on the fallen logs of Pinus armandii with different decay classes in Sygera Mountains Ling LIU
1 , Ying-Jie FAN
1 , Xiao-Tong SONG
1 , Min LI
2 , Xiao-Ming SHAO
1 , Xiao-Rui WANG
, 3 , * 1 College of Resources and Environmental Sciences, Beijing Key Laboratory of Biodiversity and Organic Farming, China Agricultural University, Beijing 100193, China2 College of Life Sciences, Hebei Normal University, Shijiazhuang 050024, China3 College of Resources and Environmental Science, Shijiazhuang University, Shijiazhuang 050035, China通讯作者: *
wxr_wxr@126.com 编委: 石培礼
责任编辑: 赵航
收稿日期: 2019-11-25
接受日期: 2020-04-27
网络出版日期: 2020-08-20
基金资助: 国家自然科学基金 (41771054 )国家自然科学基金 (31570474 )河北省自然科学基金 (D2019106006 )
Corresponding authors: *
wxr_wxr@126.com Received: 2019-11-25
Accepted: 2020-04-27
Online: 2020-08-20
Fund supported: National Natural Science Foundation of China (41771054 )National Natural Science Foundation of China (31570474 )Natural Science Foundation of Hebei Province (D2019106006 )
摘要 倒木是森林生态系统维持健康和更新的重要组成部分, 在倒木不同腐解阶段, 倒木上定植的苔藓植物组合的差异尚不清楚。为深入探讨倒木上苔藓植物组合变化规律及其影响因素, 该文对西藏色季拉山同一地点不同腐解程度的4株华山松(
Pinus armandii )倒木上的苔藓植物进行了样方调查和数据分析。结果表明: 4株倒木上40个样方共有苔藓植物22科52种, 其中藓类13科38种, 苔类9科14种; 以恒有优势种为特征进行划分, 共得到14个物种组合。随着倒木腐解程度加大以及周围环境的不断改变, 生长其上的苔藓植物物种多样性逐渐增加, 但组合数量和苔藓总盖度却呈现先增大后减小的趋势。苔藓物种由耐旱性强的丛生型藓类, 如木灵藓科、白发藓科, 逐渐演替为喜阴湿环境、快速生长的垫状或毯状藓类, 如提灯藓科、羽藓科和青藓科。倒木苔藓植物组合空间结构复杂程度逐渐增大。倒木是多种苔藓植物选择的重要生长基质, 其上苔藓植物受倒木的腐解程度、水分和光照等条件的影响而发生相应的物种组合变化。
关键词: 苔藓植物 ;
倒木 ;
生活型 ;
物种多样性 ;
环境因子 Abstract Aims Fallen logs play important roles in maintaining the health and the renewal of forest ecosystems, but the difference of bryophyte societies on fallen logs with different decay classes is unclear.
Methods To understand the regularity and influencing factors of changes in bryophyte societies, we investigated the bryophytes with quadrats along the trunks on four fallen logs of
Pinus armandii with different decay classes at the same site in Sygera Mountains, Xizang. Then, the measured data were collected and analyzed.
Important findings There were 22 families and 52 species of bryophytes inhabiting the four fallen logs, including 13 families and 38 species of Musci, 9 families and 14 species of Hepaticae. According to the characteristics of the shared dominant species, 14 societies were obtained. With the increase of decay classes of fallen logs and changes in surrounding environment, the bryophyte diversity increased gradually, while the number of societies and the total bryophyte coverage fluctuated. Bryophyte species varied from drought-tolerant tall turfs, such as Orthotrichaceae and Leucobryaceae, to fast-growing cushions or mats in humid and shady environments, such as Mniaceae, Thuidiaceae and Brachytheciaceae. The complexity of spatial structure of bryophytic society increased gradually. Fallen logs are important growth substrates for a variety of bryophytes. Bryophyte societies on fallen logs vary with decay classes, moisture and light conditions.
Keywords: bryophyte ;
fallen log ;
life form ;
biodiversity ;
environmental factor PDF (1171KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 刘凌, 樊英杰, 宋晓彤, 李敏, 邵小明, 王晓蕊. 色季拉山不同腐解等级华山松倒木上的苔藓植物组合. 植物生态学报, 2020, 44(8): 842-853. DOI:
10.17521/cjpe.2019.0327 LIU Ling, FAN Ying-Jie, SONG Xiao-Tong, LI Min, SHAO Xiao-Ming, WANG Xiao-Rui.
Bryophyte societies on the fallen logs of Pinus armandii with different decay classes in Sygera Mountains .
Chinese Journal of Plant Ecology , 2020, 44(8): 842-853. DOI:
10.17521/cjpe.2019.0327 倒木属于粗死木质残体(coarse woody debris, CWD)的一种(
闫恩荣等, 2005 )。在传统的森林管理过程中, 倒木常被误认为容易引起病虫害和火灾而被清除。近年来, 许多****对森林生态系统的深入研究结果表明, 倒木是森林生态系统不可缺少的重要组成部分, 在减少水土流失、提高林地土壤肥力和加速养分循环等方面发挥着重要作用(
Sandstr?m et al ., 2007 ;
Vanha-Majamaa et al ., 2007 ;
Davis et al ., 2010 )。倒木作为森林生态系统中的持续营养源, 是种类繁多的生物赖以生存的蔽护所和繁殖场地, 也是林木更新的良好苗床, 对森林生态系统的完整性和生物多样性的维持具有重要意义(
Takahashi et al ., 2000 ;
何东进等, 2009 )。
在早期森林生态系统的研究中, ****们对倒木的关注极少。1948年, McCullough研究了美国天然云杉(
Picea asperata )林3种生境下腐烂倒木上的植物演替, 认为其模式为: 地衣和苔类植物→藓类植物→草本和矮灌木→云杉、冷杉(
Abies fabri )等乔木树种。
Fogel和Cromack (1977) 提出倒木腐烂等级的五级系统, 并研究了倒木的养分变化。20世纪90年代, 倒木的生态功能以及动态特征的研究开始逐渐增加。倒木与生物多样性维持的研究, 主要围绕微生物多样性(木材腐朽菌及大型真菌)和动物多样性展开研究, 也有少量研究涉及植物(如苔藓、地衣)多样性(
游惠明, 2013 ), 在倒木上栖息的植物类型中, 苔藓植物是最大的植物类群(
Dittrich et al ., 2014 )。
倒木主要存在于地表并与土壤层联系紧密, 具有相对稳定的环境, 是森林中苔藓植物生长的重要基质(
Preik?a et al ., 2015 ); 它也是一个不断发展的基质, 木材腐烂程度、树皮覆盖度、离地面高度以及一些小气候条件随时间连续变化(
闫恩荣等, 2005 ;
何东进等, 2009 ), 其上生长的苔藓植物也随着环境改变持续变化(
Kumar et al ., 2018 )。苔藓植物组合作为倒木群落内部次优势物种所构成的群落局部集合体(
李博等, 2000 ), 更是随着倒木的腐解进程发生着演替(
Kushnevskaya et al ., 2007 )。一系列的研究探讨了倒木上苔藓植物物种组成和丰度的变化与倒木自身性质以及林地环境的相互关系(
McAlister, 1997 ;
Dittrich et al ., 2014 ;
Preik?a et al ., 2015 )。但由于变量间存在复杂的共线性关系, 要找到直接作用于苔藓组合的生态因子, 并从中抽离出倒木上苔藓植物组合演替规律是困难的(
Botting & DeLong, 2009 )。
2018年7月, 我们于西藏色季拉山进行野外调查时, 很幸运地发现在同一地点有4株倒伏的华山松(
Pinus armandii ), 其倒伏时间不同, 并表现出不同的腐解等级。每株倒木间相距仅2-4 m, 其上定植有多种苔藓植物, 森林管理状况、物种区系、气候条件等大环境一致的前提下, 这4株倒木成为倒木上苔藓组合演替的可视样本。为了能更深入地了解这一过程, 我们对倒木上的苔藓进行了采样调查与分析。
1 材料和方法 1.1 自然概况 研究地点位于藏东南色季拉山鲁朗兵站往通麦方向约8 km处(29.87° N, 94.77° E), 海拔2 867 m。本区域属亚高山寒温带湿润气候区, 年平均气温-0.73 ℃, 最暖月(7月)平均气温9.8 ℃, 年降水量约1 134.1 mm, 年蒸发量544.0 mm, 年平均相对湿度78% (
马豪霞等, 2016 ); 研究地点植被是以华山松为优势树种的森林, 间或分布急尖长苞冷杉(
Abies georgei var.
smithii ); 灌木层有高山栎(
Quercus semecarpifolia )、西藏箭竹(
Fargesia macclureana )、西南花楸(
Sorbus rehderiana )等; 草本层有鬼灯檠(
Rodgersia podophylla )、小檗属(
Berberis )、悬钩子属(
Rubus )、野草莓(
Fragaria vesca )、高山露珠草(
Circaea alpina )、薹草属(
Carex )、鳞毛蕨属(
Dryopteris )等。坡向朝南, 坡度约为23°。
1.2 取样方法 倒木均为由坡上向坡下倾倒, 对每株倒木朝上一侧的苔藓进行了调查与采集, 调查方法为从基部到梢部, 在倒木正上方每隔1 m做一个样方, 样方大小为10 cm × 10 cm, 每株倒木共记10个样方, 估测且记录每个样方内苔藓的总盖度, 并采集样方内苔藓植物装入编号的牛皮纸袋内; 另外记录倒木腐解程度数据、倒木倒伏方向、倒木与林窗相对位置、遮阴程度等信息。
1.3 数据获得与分析 1.3.1 样方苔藓植物的室内鉴定
将苔藓植物标本带回实验室阴干、装袋, 登记采集信息, 参考《西藏苔藓植物志》(
中国科学院青藏高原综合科学考察队, 1985 )、《中国苔藓志》(
高谦, 1994 ,
1996 ,
2003 ;
吴鹏程, 2002 ;
吴鹏程和贾渝, 2004 ,
2011 ;
胡人亮和王幼芳, 2005 ;
黎兴江, 2006 ;
高谦和吴玉环, 2008 )、《中国生物物种名录: 1. 植物: 苔藓植物》(
贾渝和何思, 2013 )以及近年来相关物种的研究资料(
李敏等, 2016 )对标本进行解剖鉴定, 参考美国密苏里植物园Tropicos网站(http://www.tropicos. org/)对苔藓物种名录进行整理归纳, 并根据当时所拍摄照片, 确定每个样方中每种苔藓的分盖度。本论文所采用的标本存放于中国农业大学标本室(BAU)。
1.3.2 倒木基本信息及其腐解等级的确定
依据所记录倒木的腐解程度数据, 参照
闫恩荣等(2005) 和
马豪霞等(2016) 对森林粗木质残体腐解等级的划分及判定标准, 确定每株倒木的腐解等级, 并根据腐解等级对倒木进行编号。
1.3.3 倒木苔藓组合演替
1.3.3.1 倒木苔藓生长情况 按照倒木上苔藓的生长情况将苔藓分成不同的生活型, 研究样本中共出现7种生活型: 大垫状(large cushion)、矮藓丛状(short turf)、直立高藓丛状(tall turf)、毯状(rough mat)、交织状(weft)、丝状(thread-like form)、紧密簇状(dense form)(
吴鹏程, 1998 )。统计不同倒木上着生的不同类型苔藓物种数目及盖度情况。
相对盖度为单株倒木10个样方内该物种的分盖度值之和与该倒木样方内各物种的分盖度之和的比值。
倒木苔藓盖度为单株倒木10个样方内苔藓盖度的平均值。
1.3.3.2 倒木苔藓组合 以恒有优势种为特征对样方进行划分及归类, 并按照优势种命名苔藓组合。
对不同组合内各苔藓植物重要值进行计算(
Song et al ., 2015 ;
范苗等, 2017 )。
重要值= (相对盖度+相对频度)/2
相对盖度=组合内该物种的分盖度/组合内各个物种的分盖度之和。
相对频度=组合内该物种的出现次数/组合内各个物种出现次数之和。
统计各苔藓组合内不同生活型苔藓的重要值, 并根据苔藓组合内不同苔藓生活型的数量划分苔藓组合空间结构的复杂程度。具体标准为: 6种及以上生活型, 复杂; 5种生活型, 较复杂; 3或4种生活型, 较简单; 1或2种生活型, 简单。
2 结果 2.1 倒木基本信息 依据野外记录数据, 结合参考资料, 整理出4株倒木的基本信息及编号(
表1 )。
Table 1 表1 表1 色季拉山4株倒木基本信息
Table 1
Essential information of the four fallen logs in Sygera Mountains 倒木 Fallen log 倒木1 Log 1 倒木2 Log 2 倒木3 Log 3 倒木4 Log 4 针叶 Needle 存在 Present 无 Absent 无 Absent 无 Absent 树皮 Bark 与活体无显著区别 少部分脱落 仅在粗大树枝上残存 无 Absent 树枝 小树枝均存在 中等树枝存在 粗大树枝存在 仅粗大树枝基部存在 主干形状 Bole shape 圆形 Round 圆形 Round 圆形 Round 圆形至卵形 Round to oval 与地面位置 被某点抬高 被某点抬高 接近地面 整体落在地面 结构完整性 木质体新鲜, 刀片可刺进 开始腐解, 刀片可刺进 刀片可刺进约2 cm 严重腐烂, 刀片可刺进 腐解等级 Decay class Ⅰ Ⅱ Ⅲ Ⅳ 胸径 Diameter at breast height 35 cm 23 cm 32 cm 27 cm 倒伏方向 南偏东15° 南偏东25° 南偏东15° 南偏东10° 倒木位置 林窗中心 林窗中心 林内 林窗边缘* Edge of forest gap* 遮阴程度 Shade degree 10% 50% 90% 60%*
*, 倒木4处于倒木2倒下所形成的林窗边缘。 *, Log 4 was located at the edge of a forest gap formed by Log 2. 新窗口打开 |
下载CSV 4株倒木的位置, 从坡下至坡上依次是倒木1、倒木3、倒木4和倒木2, 两两相距约2-4 m。
倒木1大部分树皮完整光滑且干燥, 中上部树皮内存在大量树脂囊泡; 树干结构完整, 木质体新鲜, 刀片刺入很浅; 针叶仍在树枝上未脱落; 粗大树枝较多, 使得树干梢部离地1.5 m左右, 枝干上小树枝和针叶完整, 依据以上特征, 判断此倒木腐解等级为I。倒木1位于山坡最下方, 倒伏后横跨一个季节性流水沟, 位于林窗中心, 周围高大乔木和灌木均较少, 草本层稀疏, 倒木整体遮阴程度较低, 仅10%。
倒木2仅少部分树皮脱落, 保留的树皮已被水汽浸润, 不见或偶见树脂囊泡; 树干结构完整, 部分木质体开始腐解, 刀片可刺进略深; 针叶已脱落; 部分粗大树枝尚存, 使得树干梢部离地0.5-1 m, 枝干上仅存部分小树枝, 依据以上特征, 判断此倒木腐解等级为II。倒木2位于山坡最上方, 自身倒伏后形成的林窗中心, 林窗内尚未有大的乔木或灌木形成, 只有一些小灌木、幼树和少数草本植物, 周围树木枝杈向林窗伸展, 遮阴程度50%。
倒木3绝大部分树皮已脱落, 仅在粗大树枝基部残存, 且已被水汽浸润, 不见树脂囊泡或被枯枝落叶覆盖; 树干结构尚完整, 大部分木质体已经腐解, 刀片易刺入较深; 针叶完全脱落; 少部分粗大树枝仅基部存在使得树干接近地面, 仅梢部离地面不到0.5 m, 依据以上特征, 判断此倒木腐解等级为Ⅲ。倒木3倒伏时间较长, 位于以华山松为优势的乔木层之下, 且灌木和草本植物生长旺盛, 遮阴程度高, 达到90%, 并且树干部分区域被枯枝落叶覆盖, 偶见菌类和其他植物幼苗着生树干上。
倒木4已完全不见树皮; 树干被水汽浸润, 木质体腐解程度较大, 部分区域边材已脱落, 可见开始腐解的心材, 刀片可轻易刺入达5 cm; 针叶完全脱落; 偶见粗大树枝基部, 但不能撑起树干而使得树干整体落地, 依据以上特征, 判断此倒木腐解等级为IV。尽管倒木4倒伏时间最长, 但位于倒木2和3之间, 在倒木2倒伏形成林窗后, 倒木4暴露在林窗边缘, 因此遮阴程度小于倒木3为60%, 树干部分区域重新暴露于阳光之下。树干上的枯枝落叶层最多最厚, 部分覆盖了苔藓植物, 少量菌类和其他种子植物着生于树干上。
2.2 倒木苔藓生长情况 4株倒木上共鉴定出苔藓22科52种, 其中藓类13科38种, 苔类9科14种, 苔藓植物名录、物种代码、生活型及相对盖度见
表2 。
Table 2 表2 表2 色季拉山4株倒木苔藓生长情况统计表
Table 2
Statistics of growth situation of bryophytes on the four fallen logs in Sygera Mountains 科 物种 代码 生活型 相对盖度 Relative coverage (%) Log 1 Log 2 Log 3 Log 4 曲背藓科 Oncophoraceae 曲背藓 Oncophorus wahlenbergii OW T-turf 9.74 - - - 南亚合睫藓 Symblepharis reinwardtii SR T-turf - - - 1.99 合睫藓 Symblepharis vaginata SV T-turf - 44.18 - 2.99 曲尾藓科 Dicranaceae 波叶曲尾藓 Dicranum polysetum DP T-turf - - 3.97 - 白发藓科 Leucobryaceae 丛叶青毛藓 Dicranodontium caespitosum DC T-turf - 8.52 4.77 6.97 青毛藓 Dicranodontium denudatum DiDe T-turf - 6.39 - 36.82 长叶青毛藓 Dicranodontium didymodon DiDi T-turf - 3.69 0.64 5.47 丛藓科 Pottiaceae 高山红叶藓 Bryoerythrophyllum alpigenum BA S-turf - - 0.32 - 齿叶薄齿藓 Leptodontium handelii LH S-turf 0.49 - - - 狭叶拟合睫藓 Pseudosymblepharis angustata PA S-turf - 0.14 4.45 4.23 真藓科 Bryaceae 拟大叶真藓 Bryum salakense BryS T-turf - - 0.32 - 提灯藓科 Mniaceae 侧枝匐灯藓 Plagiomnium maximoviczii PM L-cushion - - 8.10 - 全缘匐灯藓 Plagiomnium integrum PI L-cushion - - - 1.00 钝叶匐灯藓 Plagiomnium rostratum PR L-cushion - - 7.15 - 具缘提灯藓 Mnium marginatum MM L-cushion - - 2.54 - 木灵藓科 Orthotrichaceae 毛帽木灵藓 Orthotrichum dasymitrium OD D-form 13.92 - - - 白齿藓科 Leucodontaceae 中华白齿藓 Leucodon sinensis LS Weft 11.68 - - - 蔓藓科 Meteoriaceae 新丝藓 Neodicladiella pendula NP R-mat 11.68 - - - 羽藓科 Thuidiaceae 毛羽藓 Bryonoguchia molkenboeri BM R-mat - - - 0.50 大羽藓 Thuidium cymbifolium TC R-mat - 12.07 34.95 9.70 毛尖羽藓 Thuidium plumulosum TP R-mat - - - 0.50 狭叶小羽藓 Haplocladium angustifolium HA R-mat - 0.28 - - 细叶小羽藓 Haplocladium microphyllum HM R-mat - - 8.26 5.22 青藓科 Brachytheciaceae 多褶青藓 Brachythecium buchananii BB L-cushion - 0.21 6.04 0.50 平枝青藓 Brachythecium helminthocladum BH L-cushion - - 2.38 - 燕尾青藓 Brachythecium novae-angliae BraN L-cushion - - 4.29 - 褶叶青藓 Brachythecium salebrosum BraS L-cushion - - - 1.24 拟异叶藓 Pseudokindbergia dumosa PD L-cushion - - 0.64 - 灰藓科 Hypnaceae 圆枝粗枝藓 Gollania tereticaulis GT R-mat - 2.13 - - 灰藓 Hypnum cupressiforme HC R-mat - - 2.22 0.50 密枝灰藓 Hypnum densirameum HD R-mat - 2.13 2.86 7.46 毛梳藓 Ptilium crista-castrensis PC R-mat 14.61 - - - 拟硬叶藓 Stereodontopsis pseudorevoluta SP R-mat 0.19 - - - 丝金灰藓 Pylaisia levieri PL R-mat 34.08 7.39 - - 毛锦藓科 Pylaisiadelphaceae 曲叶小锦藓 Brotherella curvirostris BC R-mat - 2.70 1.59 2.24 弯叶小锦藓 Brotherella falcata BF R-mat 0.97 - - - 垂蒴小锦藓 Brotherella nictans BroN R-mat - - 2.38 7.46 暗绿毛锦藓 Pylaisiadelpha tristoviridis PT R-mat - 4.12 - - 叶苔科 Jungermanniaceae 深绿叶苔 Jungermannia atrovirens JA D-form - 4.69 0.16 1.24 小萼叶苔 Jungermannia parviperiantha JP D-form - - - 0.25 方叶无褶苔 Leiocolea bantriensis LB D-form - - - 0.25 挺叶苔科 Anastrophyllaceae 小挺叶苔 Anastrophyllum minutum AM R-mat - - - 0.37 裂叶苔科 Lophoziaceae 三瓣苔 Tritomaria exsecta TE T-form - 0.85 - 0.62 折叶苔科 Scapaniaceae 刺边合叶苔 Scapania ciliata SC T-form - - 1.27 - 睫毛苔科 Blepharostomataceae 睫毛苔 Blepharostoma trichophyllum BT R-mat - 0.50 0.08 1.37 指叶苔科 Lepidoziaceae 指叶苔 Lepidozia reptans LR R-mat - - - 0.62 光萼苔科 Porellaceae 光萼苔 Porella pinnata PP R-mat 0.19 - - - 耳叶苔科 Frullaniaceae 细茎耳叶苔 Frullania bolanderi FB T-form 0.19 - - - 达乌里耳叶苔 Frullania davurica FD T-form 1.27 - 0.64 - 列胞耳叶苔 Frullania moniliata FM T-form 0.49 - - - 细鳞苔科 Lejeuneaceae 角齿细鳞苔 Lejeunea otiana LO T-form - - - 0.50 南亚瓦鳞苔 Trocholejeunea sandvicensis TS T-form 0.49 - - -
D-form, 紧密簇状; L-cushion, 大垫状; R-mat, 毯状; S-turf, 矮藓丛状; T-form, 丝状; T-turf, 直立高藓丛状; Weft, 交织状。 D-form, dense form; L-cushion, large cushion; R-mat, rough mat; S-turf, short turf; T-form, thread-like form; T-turf, tall turf. 新窗口打开 |
下载CSV 依据
表2 数据, 对4株倒木不同生活型苔藓植物物种数目和盖度情况进行了统计和比较(
图1 )。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1色季拉山4株倒木不同生活型苔藓物种数目(A)及盖度(B)柱状图。D-form, 紧密簇状; L-cushion, 大垫状; R-mat, 毯状; S-turf, 矮藓丛状; T-form, 丝状; T-turf, 直立高藓丛状; Weft, 交织状。 Fig. 1The richness (A) and coverage (B) of bryophyte species with different growth forms on the four fallen logs in Sygera Mountains. D-form, dense form; L-cushion, large cushion; R-mat, rough mat; S-turf, short turf; T-form, thread-like form; T-turf, tall turf. 由
表2 和
图1 可以看出, 倒木1上生长的苔藓植物种类最少, 仅有13种, 其中毯状和丝状生活型占多数。盖度最大的物种为毯状生长的丝金灰藓(
Pylaisia levieri ), 相对盖度34.08%, 拟硬叶藓(
Stereodontopsis pseudorevoluta )、毛帽木灵藓(
Orthotrichum dasymitrium )、新丝藓(
Neodicladiella pendula )、中华白齿藓(
Leucodon sinensis )、曲背藓(
Oncophorus wahlenbergii )也有较高的盖度, 另外还着生有几种茎叶体苔类植物。
倒木2上生长有苔藓16种, 毯状生长的藓类植物最为多样, 其次是直立高藓丛状的。盖度最大的物种为合睫藓(
Symblepharis vaginata ), 相对盖度44.18%, 直立高藓丛状生长, 此外, 毯状生长的大羽藓(
Thuidium cymbifolium )也有较高的盖度。
倒木3上着生有苔藓23种, 以毯状和大垫状的为主, 盖度最大的为大羽藓, 相对盖度34.95%, 毯状生长, 而细叶小羽藓(
Haplocladium microphyllum )、侧枝匐灯藓(
Plagiomnium maximoviczii )、钝叶匐灯藓(
Plagiomnium rostratum )和多褶青藓(
Brachythecium buchananii ) 4种大垫状生长的藓类植物也有较高的盖度。
倒木4上着生的苔藓植物物种最为丰富, 共有25种, 毯状生长的藓类植物最为多样, 直立高藓丛状、大垫状和紧密簇状的苔藓种类也较多。盖度最大的种类为青毛藓(
Dicranodontium denudatum ), 相对盖度36.82%, 大羽藓、密枝灰藓(
Hypnum densirameum )、垂蒴小锦藓(
Brotherella nictans )、丛叶青毛藓(
Dicranodontium caespitosum )也有较高的盖度。
随着倒木自身和周围环境的变化, 倒木上原有物种被淘汰, 新物种定植, 整体来说, 随倒木腐解等级增大和水分光照等因子的改变, 倒木上物种数目增多, 倒木苔藓的盖度变化也较大, 在倒木1上苔藓盖度非常低, 而在倒木2达到最大, 随后再又逐渐减少。
2.3 倒木苔藓组合 以恒有优势种为特征对4株倒木上共40个样方进行了划分, 共得到14个苔藓组合(
表3 )。将14个组合内同生活型物种的重要值求和以表征各生活型在组合内的优势程度, 综合组合内生活型数量及优势程度, 得到各组合空间结构的复杂程度(
表4 )。
Table 3 表3 表3 色季拉山4株倒木苔藓组合
Table 3
Bryophyte societies on the four fallen logs in Sygera Mountains 组合名称 物种组成 重要值 分布样方 组合名称 物种组成 重要值 分布样方 合睫藓组合Symblepharis vaginata society SV 0.425 Log 2-4, Log 2-5, Log 2-6, Log 2-7, Log 2-8, Log 2-9, Log 2-10 丛叶青毛藓组合Dicranodontium vaginata society DC 0.250 Log 3-10, Log 4-7 DC 0.135 PA 0.167 JA 0.100 BC 0.114 BC 0.076 JA 0.092 PL 0.065 PM 0.084 DiDi 0.043 TC 0.076 PT 0.043 BroN 0.069 DiDe 0.034 SV 0.053 HD 0.034 TP 0.053 TE 0.025 BT 0.042 BT 0.020 侧枝匐灯藓组合Plagiomnium maximoviczii society PM 0.346 Log 3-5, Log 3-7 大羽藓组合Thuidium cymbifolium society TC 0.423 Log 2-2, Log 3-2, Log 3-3, Log 3-8, Log 3-9, Log 4-10 BB 0.207 DC 0.069 TC 0.164 BroN 0.069 PA 0.126 HD 0.052 HC 0.086 PM 0.048 BraN 0.071 DP 0.042 丝金灰藓组合Pylaisia levieri society PL 0.427 Log 1-1, Log 1-7, Log 1-9 PA 0.042 PC 0.172 JA 0.037 OW 0.132 BT 0.036 OD 0.058 BH 0.032 LH 0.054 BB 0.027 FD 0.054 SC 0.025 TS 0.052 PT 0.021 FB 0.051 LR 0.018 密枝灰藓组合Hypnum densirameum society HD 0.389 Log 4-4, Log 4-8 SV 0.018 DiDe 0.130 LB 0.018 BC 0.130 DiDi 0.018 TC 0.093 青毛藓组合Dicranodontium denudatum society DiDe 0.391 Log 2-3, Log 4-2, Log 4-5, Log 4-6, Log 4-9 DiDi 0.074 SR 0.064 BT 0.065 PL 0.062 TE 0.060 BT 0.056 LR 0.060 JA 0.051 钝叶匐灯藓组合Plagiomnium rostratum society PR 0.575 Log 3-6 GT 0.044 HM 0.145 BroN 0.044 BB 0.145 SV 0.035 TC 0.135 PA 0.035 燕尾青藓组合Brachythecium novae-angliae society BraN 0.378 Log 3-4 BraS 0.026 HC 0.211 HC 0.020 TC 0.156 BM 0.020 PA 0.133 TE 0.020 BryS 0.122 BB 0.020 毛帽木灵藓组合Orthotrichum dasymitrium society OD 0.509 Log 1-4, Log 1-5, Log 1-10 AM 0.019 FD 0.139 HM 0.018 TS 0.086 LB 0.018 PL 0.068 BC 0.018 BF 0.068 LR 0.018 PP 0.044 JP 0.018 SP 0.044 细叶小羽藓组合Haplocladium microphylllum society HM 0.282 Log 3-1, Log 4-1 FB 0.041 DiDi 0.136 新丝藓组合Neodicladiella pendula society NP 0.730 Log 1-8 PA 0.091 FM 0.270 MM 0.081 SV 0.061 BB 0.049 中华白齿藓组合Leucodon sinensis society LS 1.000 Log 1-2 PI 0.042 PD 0.042 FD 0.042 狭叶小羽藓组合Haplocladium angustifolium society HA 0.389 Log 2-1 BA 0.036 BB 0.333 LO 0.036 PA 0.278 TC 0.036 BT 0.036 LR 0.031
物种代码见表2 。 Species code refer to Table 2 . 新窗口打开 |
下载CSV Table 4 表4 表4 苔藓组合内各生活型物种重要值之和
Table 4
Sum of the important values of different growth forms in different bryophyte societies 物种重要值之和 大垫状 矮藓丛状 直立高 毯状 交织状 丝状 紧密簇状 组合空间结构 合睫藓组合 Symblepharis vaginata society - - 0.637 0.238 - 0.025 0.100 较简单 大羽藓组合 Thuidium cymbifolium society 0.108 0.042 0.148 0.621 - 0.025 0.055 复杂 青毛藓组合 Dicranodontium denudatum society 0.046 0.035 0.491 0.321 - 0.020 0.088 复杂 细叶小羽藓组合 Haplocladium microphyllum society 0.213 0.091 0.233 0.385 - 0.078 - 较复杂 丛叶青毛藓组合 Dicranodontium caespitosum society 0.084 0.167 0.303 0.354 - - 0.092 较复杂 侧枝匐灯藓组合 Plagiomnium maximoviczii society 0.624 0.126 - 0.250 - - - 较简单 丝金灰藓组合 Pylaisia levieri society - 0.054 0.132 0.600 - 0.157 0.058 较复杂 密枝灰藓组合 Hypnum densirameum society - - 0.204 0.736 - 0.060 - 较简单 钝叶匐灯藓组合 Plagiomnium rostratum society 0.720 - - 0.28 - - - 简单 燕尾青藓组合 Brachythecium novae-angliae society - - - 0.225 - 0.266 0.509 较简单 毛帽木灵藓组合 Orthotrichum dasymitrium society 0.378 0.133 0.122 0.367 - - - 较简单 新丝藓组合 Neodicladiella pendula society - - - 0.730 - 0.270 - 简单 中华白齿藓组合 Leucodon sinensis society - - - - 1.00 - - 简单 狭叶小羽藓组合 Haplocladium angustifolium society 0.333 0.278 - 0.389 - - - 较简单
新窗口打开 |
下载CSV 苔藓组合排序按照分布样方盖度之和的大小, 14个组合依次为:
合睫藓组合: 分布在倒木2的中部及梢部共7个样方内, 共有苔藓种类11种, 优势苔藓物种合睫藓重要值为0.425。组合内包含有直立高藓丛状、毯状、丝状、紧密簇状生长的物种, 直立高藓丛状物种最具优势, 重要值之和为0.637, 其次是毯状生长藓类, 重要值之和0.238, 组合空间结构较简单。
大羽藓组合: 分布在倒木2基部、倒木3基部及梢部和倒木4梢部共6个样方内, 共有苔藓种类17种, 优势苔藓物种大羽藓重要值为0.423。组合内包含有大垫状、矮藓丛状、直立高藓丛状、毯状、丝状、紧密簇状生长的物种, 毯状物种最具优势, 重要值之和为0.621, 其次是直立高藓丛状和大垫状, 重要值之和分别为0.148、0.108, 组合空间结构复杂。
青毛藓组合: 分布在倒木2基部、倒木4不同部位共5个样方内, 共有苔藓种类20种, 优势苔藓物种青毛藓重要值为0.391。组合内包含有大垫状、矮藓丛状、直立高藓丛状、毯状、丝状、紧密簇状生长的物种, 直立高藓丛状和毯状物种较具优势, 重要值之和分别为0.491、0.321, 组合空间结构复杂。
细叶小羽藓组合: 分布在倒木3和倒木4基部共2个样方内, 共有苔藓种类14种, 优势苔藓物种细叶小羽藓重要值为0.282。组合内包含有大垫状、矮藓丛状、直立高藓丛状、毯状、丝状生长的物种, 毯状、直立高藓丛状、大垫状物种较具优势, 重要值之和分别为0.385、0.233、0.213, 组合空间结构较复杂。
丛叶青毛藓组合: 分布在倒木3和倒木4梢部共2个样方内, 共有苔藓种类10种, 优势苔藓物种丛叶青毛藓重要值为0.250。组合内包含有大垫状、矮藓丛状、直立高藓丛状、毯状、紧密粗簇状生长的物种, 毯状和直立高藓丛状物种较具优势, 重要值之和分别为0.354、0.303, 组合空间结构较复杂。
侧枝匐灯藓组合: 分布在倒木3中部2个样方内, 共有苔藓种类6种, 优势苔藓物种侧枝匐灯藓重要值为0.346。组合内包含有大垫状、矮藓丛状、毯状生长的物种, 大垫状物种最具优势, 重要值之和为0.624, 组合空间结构较简单。
丝金灰藓组合: 分布在倒木1基部及梢部共3个样方内, 共有苔藓种类8种, 优势苔藓物种丝金灰藓重要值为0.427。组合内包含有矮藓丛状、直立高藓丛状、毯状、丝状、紧密粗簇状生长的物种, 毯状物种最具优势, 重要值之和为0.600, 组合空间结构较复杂。
密枝灰藓组合: 分布在倒木4中部及梢部共2个样方内, 共有苔藓种类8种, 优势苔藓物种密枝灰藓重要值为0.389。组合内包含有直立高藓丛状、毯状、丝状生长的物种, 毯状物种最具优势, 重要值之和为0.736, 组合空间结构较简单。
钝叶匐灯藓组合: 分布在倒木3中部1个样方内, 共有苔藓种类4种, 优势苔藓物种钝叶匐灯藓重要值为0.575。组合内包含有大垫状、毯状生长的物种, 大垫状物种最具优势, 重要值之和为0.720, 组合空间结构简单。
燕尾青藓组合: 分布在倒木3中部1个样方内, 共有苔藓种类5种, 优势苔藓物种燕尾青藓(
Brachythecium novae-angliae )重要值为0.378。组合内包含有毯状、丝状、紧密簇状生长的物种, 紧密簇状物种最具优势, 重要值之和为0.509, 组合空间结构较简单。
毛帽木灵藓组合: 分布在倒木1中部及梢部共3个样方内, 共有苔藓种类8种, 优势苔藓物种毛帽木灵藓重要值为0.509。组合内包含有大垫状、矮藓丛状、直立高藓丛状、毯状生长的物种, 大垫状和毯状物种较具优势, 重要值之和分别为0.378、0.367, 组合空间结构较简单。
新丝藓组合: 分布在倒木1梢部1个样方内, 共有苔藓种类2种, 优势苔藓物种新丝藓重要值为0.730, 列胞耳叶苔重要值为0.270, 两者分别为毯状、丝状生长藓类, 组合空间结构简单。
中华白齿藓组合: 分布在倒木1基部1个样方内, 仅有中华白齿藓一个物种, 生活型为交织状, 组合空间结构简单。
狭叶小羽藓组合: 分布在倒木2基部1个样方内, 共有苔藓种类3种——狭叶小羽藓(
Haplocladium angustifolium )、多褶青藓、狭叶拟合睫藓(
Pseudosymblepharis angustata ), 三者重要值分别为0.389、0.333和0.278, 生活型分别为毯状、大垫状、矮藓丛状, 组合空间结构较简单。
倒木1的样方3和样方6无苔藓植物分布, 是因为倒木腐解程度低, 在这两个部位, 尚无苔藓定植。倒木4的样方3内无苔藓植物分布, 是此倒木过于腐解在样方3处树皮和边材已腐烂脱落, 露出的心材还未有苔藓植物生长。
2.4 倒木苔藓组合演替 根据研究结果分析, 西藏色季拉山华山松倒木的苔藓植物组合的演替过程如下:
倒木1, 腐解等级低, 树表干燥, 位于开阔的林窗中心, 遮阴程度低。其上定植的苔藓种类较少, 总盖度较低, 表面仅有少量喜光耐旱的藓类, 以及紧贴于树干表面稀疏着生的苔类, 毛帽木灵藓、中华白齿藓、新丝藓和光萼苔较为优势; 着生的苔藓组合类型同种活体树附生苔藓组合相似, 说明本研究中倒木1倒伏年限较低, 仅1-2年。随着腐解等级增加和其他生境变化, 倒木1的优势物种和物种组合会逐渐被其他优势物种和物种组合替换。
倒木2, 腐解程度加深, 树皮被水汽浸润, 位于开始闭合的林窗中心, 遮阴程度较倒木1增加。其上着生的苔藓物种数目和总盖度有所增加, 物种组合、物种种类和相应的生活型也发生明显变化。在倒木基部苔藓组合较多, 包括狭叶小羽藓组合、大羽藓组合和青毛藓组合, 倒木中部以上直至稍部均为合睫藓组合。藓类植物以直立高藓丛状生长的曲背藓科、白发藓科和毯状生长的羽藓科和毛锦藓科为主, 苔类以混生或附生于其他藓类植物的叶苔科、裂叶苔科和睫毛苔科为主。合睫藓组合虽然分布面积大, 但组合的空间结构较倒木基部组合简单(
图2 )。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2色季拉山倒木苔藓盖度及组合空间结构复杂程度变化图。 Fig. 2Variation in moss coverage and the complexity of society spatial structure in Sygera Mountains. 倒木3, 腐解等级进一步加大, 树干表层被水汽浸润, 倒伏形成的林窗闭合, 位于林内, 遮阴程度高。倒木上苔藓组合发生较为明显变化, 只有大羽藓组合与前一倒木相同, 分布于树干基部和梢部, 新出现的组合包括细叶小羽藓组合, 分布在树干基部; 侧枝匐灯藓组合、钝叶匐灯藓组合分布在树干中部; 丛叶青毛藓组合分布在树干梢部。这些组合所包含的物种数目及生活型种类都很多, 组合空间结构整体较倒木2更为复杂。苔藓植物物种数目继续增加, 物种种类和生活型也随之变化, 较为喜光的直立高藓丛状生长的曲背藓科逐渐被极度耐阴湿的大垫状着生的提灯藓科和青藓科藓类所替代; 生境条件和分布基质多样的丛藓科也定植下来; 羽藓科种类有所变化, 但盖度进一步加大; 而白发藓科盖度却较上一阶段减小。深绿叶苔(
Jungermannia atrovirens )和睫毛苔(
Blepharostoma trichophyllum )仍然占据极微小的盖度, 另外还出现了刺边合叶苔(
Scapania ciliata )和达乌里耳叶苔(
Frullania davurica )这种喜湿的苔类。
倒木4, 腐解程度最大, 树干整体被水汽浸润, 但由于周边存在新形成的林窗, 整体位于林窗边缘, 遮阴程度60%。藓类仍然以生活型为毯状的羽藓科为主, 毯状生长的灰藓科也成为了重要类群。组合类型与倒木2、倒木3相似, 但优势组合更为明显, 其中青毛藓组合从基部到梢部均有分布, 丛叶青毛藓组合与青毛藓组合占据不同空间位置, 还能见到细叶小羽藓组合和大羽藓组合, 分别分布在倒木基部和梢部的一个样方内。密枝灰藓组合是所观察的演替中最晚出现的组合, 所含物种数目和生活型都较少, 结构简单。因该倒木处于新形成的林窗边缘, 倒木上所出现的大量白发藓科物种可能与光照增强有关。
3 讨论 倒木作为苔藓植物的着生基质和微生境, 是苔藓多样性高度集中的区域, 仅在我们研究的4株倒木上就发现了苔藓植物22科52种, 其中藓类13科38种, 苔类9科14种(
表2 ), 在所研究的倒木上的苔藓植物区系成分以温带成分为主, 占比达到52%, 同时含有一定热带成分以及少量世界广布和中国特有成分。随着倒木腐解程度的加深, 以及其他生境条件的变化, 苔藓物种组成及物种组合都发生了明显的变化, 总体表现为: 物种多样性逐渐增加, 组合数量和苔藓总盖度先增大, 并随着腐木腐烂、斑块状脱落和其他植物的定植而减小(
图1 ,
图2 ), 同时倒木上苔藓物种由耐旱性相对较强的丛生型藓类逐渐为喜阴湿环境、快速生长的垫状或毯状藓类所替代(
图1 ); 苔藓组合空间结构随着倒木腐解等级增大以及其他生境条件的变化, 复杂程度逐渐增大(
图2 )。
这一系列变化与苔藓生长微生境的改变是密不可分的。倒木基质随着时间不断变化, 树皮的粗糙程度及覆盖度、结构的完整性、养分含量等都在改变(
何东进等, 2009 ), 这些基质的变化对苔藓的定植存在影响(
Dittrich et al ., 2014 ;
Kushnevskaya & Shorohova, 2018 )。另外, 倒木结构完整性、倒木离地面的距离以及倒木距离水源的距离都会影响倒木上的水分情况, 在本研究中4株倒木的水分情况随腐解程度加深而增加。而水分对于苔藓的定植很重要, 会显著影响苔藓植物的多样性(
范苗等, 2017 )。林窗作为森林中常见的小尺度干扰机制, 对于森林物种多样性维持有着重要的意义(
李强等, 2019 ), 树木倒下形成的林窗随着时间推移逐渐闭合, 这也导致不同腐解等级倒木上生长的苔藓受到的光照情况在改变。一般来说, 苔藓喜好相对遮阴的环境, 在这种环境下苔藓植物较维管植物有更大的生长优势(
白学良等, 1998 ;
刘俊华等, 2005 )。在微生境不断变化的情况下, 本研究案例中苔藓植物物种多样性、组合数量和苔藓总盖度在腐解中后期达到最大, 这与其他不同气候下、不同区域的研究趋势一致, 是倒木上苔藓植物演替的一般规律(
Cole et al ., 2008 ;
Botting & DeLong, 2009 ;
游惠明, 2013 ;
Kumar et al ., 2018 )。
苔藓植物物种组成在演替中变化明显。该变化取决于苔藓物种自身生物学和生态学属性, 并与环境密切相关。不同的苔藓物种有不同的定植方式、定植速率以及环境偏好。生长缓慢的苔类和顶蒴藓类可以在开放的倒木表面及完整的苔藓层的缝隙中定植, 而快速生长的侧蒴藓类会在短时间内构建形成大型斑块; 苔藓物种根据其对不同腐解等级倒木的定植偏好可以分为: 树表附生种、腐木泛生种、优先腐木生种和严格腐木生种(
ódor & van Hees, 2004 )。在本研究中, 毛帽木灵藓、中华白齿藓和新丝藓属于典型的树表附生种, 它们只在活体树木树干或倒伏不久、腐解等级较低的倒木的树皮上着生; 严格腐木生种如南亚合睫藓、全缘匐灯藓、毛羽藓、毛尖羽藓、褶叶青藓, 它们在腐解等级较低的倒木上不出现, 而在腐解等级较高的倒木上出现频率高, 甚至表现为优势类群的种类。但就4株倒木上苔藓的着生情况讨论物种定植偏好只是初步的, 是否具有普遍性还需要有更多的研究样本来验证。除了倒木腐解等级外, 光照和水分对苔藓植物的分布也存在较大影响(
李小芳等, 2018 ), 不同生活型的苔藓植物对光照的偏好在本研究中非常明显。倒木4由于新形成的林窗, 遮阴程度与倒木2类似, 它们在苔藓着生情况上表现为共有多种直立高藓丛状生长的植物, 直立高藓丛状有利于苔藓吸收水分而减少水分缺失对生长造成的影响, 因此能在光照相对较充足的条件下更好地生长(
M?gdefrau, 1982 ), 但是由于基质、水分含量及其他环境因子的影响, 可以看到两株倒木上占优势的直立高藓丛状苔藓种类还是存在区别的, 倒木2上较具优势的物种为合睫藓, 倒木4上较具优势物种为青毛藓。
植物之间的相互作用也在影响着演替的过程。随着时间的推移, 倒木木质素等难分解组分降解程度增加, 结构相对疏松, 树干湿度增加, 倒木上环境逐渐优化, 蕨类植物、种子植物开始在倒木上及倒木周边定植, 一定程度上挤压了苔藓的生存空间, 加之枯落物的不断累积, 占据了部分可定植的空间, 对苔藓植物的生长造成了负面的影响(
Natalia et al ., 2008 ;
Márialigeti et al ., 2009 ), 使得苔藓植物盖度在到达极大值后逐渐减小。另外苔藓植物之间也存在合作、竞争等关系(
卜兆君等, 2009 )。倒木作为一个相对变化快速的生境, 其上生长的苔藓物种和组合通常是不稳定的, 在这种情况下种间关系常趋于负相关, 即种间竞争或排他作用强烈(
刘艳等, 2019 ), 例如青毛藓、丛叶青毛藓和长叶青毛藓都分布于光照较为充足的倒木2和倒木4, 但是同一个样方内通常仅有一种青毛藓属植物。苔藓植物的竞争等级还会随着环境的变化而改变(
Breeuwer et al ., 2008 ), 从而推动不同演替阶段苔藓物种以及物种组合的变化。早期优势的丛生型藓类能较好地适应相对干旱、强光照的环境, 但随着环境向潮湿荫蔽转变, 个体更大且生长快速的垫状毯状藓类出现, 它们能获取更多的资源, 不对称竞争导致竞争等级的改变。随着更多苔藓物种的定植, 倒木上苔藓间竞争压力逐渐增大, 这种情况下物种间生态位的分化是物种共存的重要机理之一(
Rydin, 1986 ), 本研究中苔藓植物组合内生活型种类和数量即整体空间结构复杂程度随着腐解等级及其他生境条件的变化而逐渐增大, 呈现出类似于植物演替过程中的生态位分化。
4 结论 在微生境适宜的条件下, 倒木是苔藓植物选择的生长基质之一; 定植在倒木上的苔藓植物多样性及其组合空间结构的复杂程度会随着倒木的腐解进程逐渐增加; 苔藓植物组合类型及生活型的变化除了与倒木的腐解程度有关, 还与水分、光照条件以及植物之间的相互作用密切关联。
致谢 感谢华东师范大学王幼芳教授在野外考察和论文写作工作中给予的帮助与建议。
[1] Bai XL Zhao LM Sun W Sun WG 1998 ). A preliminary study on the species diversity, phytomass and ecological effect of bryophytes in Helan Mountain, ChinaActa Scientiarum Naturalium Universitatis Neimongol 118 -124 . [本文引用: 1] [ 白学良 , 赵连梅 , 孙维 , 孙卫国 (1998 ). 贺兰山苔藓植物物种多样性、生物量及生态学作用的研究内蒙古大学学报(自然科学版) 118 -124 .] [本文引用: 1] [2] Botting RS DeLong C 2009 ). Macrolichen and bryophyte responses to coarse woody debris characteristics in sub- boreal spruce forestForest Ecology and Management S85 -S94 . [本文引用: 2] [3] Breeuwer A Heijmans MMPD Robroek BJM Berendse F 2008 ). The effect of temperature on growth and competition between Sphagnum speciesOecologia 155 -167 . URL PMID:
18283501 [本文引用: 1] [4] Bu ZJ Chen X Jiang LH Li HK Zhao HY 2009 ). Research advances on interactions among bryophytesChinese Journal of Applied Ecology 460 -466 . URL PMID:
19459391 [本文引用: 1] This paper summarized the present research status and advances on the intra- and interspecific positive interaction, intra- and inter-specific competition, niche, and coexistence of bryophytes. Bryophytes are generally the dominant plants in harsh environments, and there is a trade-off between their water retention and light and nutrient resource availability. Because of the lesser importance of competition in harsh environments, the positive interaction among bryophytes is common, but the intra- and inter-specific competition among bryophytes and the competition between bryophytes and vascular plants are not rare. Competition hierarchy may exist among some bryophytes, but often changes with environments. In the process of bryophyte community formation, the random process, nature of colonization, and difference in regeneration strategy can result in the niche overlap and coexistence of bryophytes, and the niche differentiation resulted from competition is also one of the mechanisms for bryophytes coexistence. Bryophytes should not be simply classified as stress tolerated-ruderal life history strategists, and competition is still one of important factors for constructing some bryophyte communities and vegetations co-existed by bryophytes and vascular plants.
[ 卜兆君 , 陈旭 , 姜丽红 , 李鸿凯 , 赵红艳 (2009 ). 苔藓植物相互作用的研究进展应用生态学报 460 -466 .] PMID:
19459391 [本文引用: 1] [5] Cole HA Newmaster SG Bell FW Pitt D Stinson A 2008 ). Influence of microhabitat on bryophyte diversity in Ontario mixedwood boreal forestCanadian Journal of Forest Research 1867 -1876 . [本文引用: 1] [6] Davis JC Castleberry SB Kilgo JC 2010 ). Influence of coarse woody debris on herpetofaunal communities in upland pine stands of the southeastern Coastal PlainForest Ecology and Management 1111 -1117 . [本文引用: 1] [7] Dittrich S Jacob M Bade C Leuschner C Hauck M 2014 ). The significance of deadwood for total bryophyte, lichen, and vascular plant diversity in an old-growth spruce forestPlant Ecology 1123 -1137 . [本文引用: 3] [8] Fan M Wu YP Hu RG Jiang YB 2017 ). Diversity and distribution of bryophytes and their relationship with environmental factors in WuhanPlant Science Journal 825 -834 . [本文引用: 2] [ 范苗 , 伍玉鹏 , 胡荣桂 , 姜炎彬 (2017 ). 武汉市城区苔藓植物多样性和分布及与环境因子的关系植物科学学报 825 -834 .] [本文引用: 2] [9] Fogel R Cromack Jr K 1977 ). Effect of habitat and substrate quality on Douglas-fir litter decomposition in western OregonCanadian Journal of Botany 1632 -1640 . [本文引用: 1] [10] Gao Q 1994 ). Flora Bryophytarum Sinicorum: Vol. 1 [本文引用: 1] [ 高谦 (1994 ). 中国苔藓志: 1卷 [本文引用: 1] [11] Gao Q 1996 ). Flora Bryophytarum Sinicorum:. Vol. 2 [本文引用: 1] [ 高谦 (1996 ). 中国苔藓志: 2卷 [本文引用: 1] [12] Gao Q 2003 ). Flora Bryophytarum Sinicorum:. Vol. 9 [本文引用: 1] [ 高谦 (2003 ). 中国苔藓志: 9卷 [本文引用: 1] [13] Gao Q Wu YH 2008 ). Flora Bryophytarum Sinicorum:. Vol. 10 [本文引用: 1] [ 高谦 , 吴玉环 (2008 ). 中国苔藓志: 10卷 [本文引用: 1] [14] He DJ He XJ Hong W Liu YS Bian LL Qin DH You HM 2009 ). Research progress of coarse woody debris in forest ecosystemsForest Research 715 -721 . [本文引用: 3] [ 何东进 , 何小娟 , 洪伟 , 刘勇生 , 卞莉莉 , 覃德华 , 游惠明 (2009 ). 森林生态系统粗死木质残体的研究进展林业科学研究 715 -721 .] [本文引用: 3] [15] Hu RL Wang YF 2005 ). Flora Bryophytarum Sinicorum: Vol. 7 [本文引用: 1] [ 胡人亮 , 王幼芳 (2005 ). 中国苔藓志: 7卷 [本文引用: 1] [16] Jia Y He S 2013 ). Catalogue of Life China: Vol. 1. Plants: Bryophytes [本文引用: 1] [ 贾渝 , 何思 (2013 ). 中国生物物种名录: 1. 植物: 苔藓植物 [本文引用: 1] [17] Kumar P Chen HYH Thomas SC Shahi C 2018 ). Epixylic vegetation abundance, diversity, and composition vary with coarse woody debris decay class and substrate species in boreal forestCanadian Journal of Forest Research 399 -411 . [本文引用: 2] [18] Kushnevskaya H Mirin D Shorohova E 2007 ). Patterns of epixylic vegetation on spruce logs in late-successional boreal forestsForest Ecology and Management 25 -33 . [本文引用: 1] [19] Kushnevskaya H Shorohova E 2018 ). Presence of bark influences the succession of cryptogamic wood-inhabiting communities on conifer fallen logsFolia Geobotanica 175 -190 . [本文引用: 1] [20] Li B Yang C Lin P 2000 ). Ecology [本文引用: 1] [ 李博 , 杨持 , 林鹏 (2000 ). 生态学 [本文引用: 1] [21] Li M Wei QQ Wang XR Wang YF 2016 ). A rivision of Chinese Brachytheciaceae (Bryophyta) II: Brynia KaurinActa Botanica Boreali-Occidentalia Sinica 1257 -1262 . [本文引用: 1] [ 李敏 , 魏倩倩 , 王晓蕊 , 王幼芳 (2016 ). 中国青藓科植物分类学修订(Ⅱ): 燕尾藓属西北植物学报 1257 -1262 .] [本文引用: 1] [22] Li Q Wang B Deng Y Lin LX Dawa ZX Zhang ZM 2019 ). Correlation between spatial distribution of forest canopy gaps and plant diversity indices in Xishuangbanna tropical forestsBiodiversity Science 273 -285 . [本文引用: 1] [ 李强 , 王彬 , 邓云 , 林露湘 , 达佤扎喜 , 张志明 (2019 ). 西双版纳热带雨林林窗空间分布格局及其特征指数与林窗下植物多样性的相关性生物多样性 273 -285 .] [本文引用: 1] [23] Li XF Zhang ZH Li CY Wu J Wang ZH 2018 ). Diversity of bryophytes in underground forest of Monkey-ear TiankengActa Botanica Boreali-Occidentalia Sinica 2324 -2333 . [本文引用: 1] [ 李小芳 , 张朝晖 , 李承义 , 吴金 , 王智慧 (2018 ). 贵州猴耳天坑地下森林苔藓植物多样性特征研究西北植物学报 2324 -2333 .] [本文引用: 1] [24] Li XJ 2006 ). Flora Bryophytarum Sinicorum: Vol. 4 [本文引用: 1] [ 黎兴江 (2006 ). 中国苔藓志: 4卷 [本文引用: 1] [25] Liu JH Bao WK Li FL 2005 ). Major bryophyte patch structures and their relationships with environmental factors under a coniferous forest of eastern Tibetan PlateauEcology and Environment 735 -741 . [本文引用: 1] [ 刘俊华 , 包维楷 , 李芳兰 (2005 ). 青藏高原东部原始林下地表主要苔藓斑块特征及其影响因素生态环境 735 -741 .] [本文引用: 1] [26] Liu Y Zheng YY Ao YY 2019 ). Niche and interspecific association of dominant bryophytes on different substratesActa Ecologica Sinica 286 -293 . [本文引用: 1] [ 刘艳 , 郑越月 , 敖艳艳 (2019 ). 不同生长基质的苔藓植物优势种生态位与种间联结生态学报 286 -293 .] [本文引用: 1] [27] Ma HX Ren YH Hou L Wang BW Zheng WL 2016 ). Studies on the reserves of coarse woody debris and decomposition of fallen trees in Abies georgei var. smithii forest of Sygera Mountains in TibetJournal of Northwest Forestry University 68 -73 . [本文引用: 2] [ 马豪霞 , 任毅华 , 侯磊 , 王博文 , 郑维列 (2016 ). 西藏色季拉山急尖长苞冷杉林粗木质残体储量与倒木分解研究西北林学院学报 68 -73 .] [本文引用: 2] [28] M?gdefrau K 1982 ). Life-forms of bryophytes//Smith AJE. Bryophyte EcologyChapman and Hall 45 -58 . [本文引用: 1] [29] Márialigeti S Németh B Tinya F ódor P 2009 ). The effects of stand structure on ground-floor bryophyte assemblages in temperate mixed forestsBiodiversity and Conservation 2223 -2241 . [本文引用: 1] [30] McAlister S 1997 ). Cryptogam communities on fallen logs in the Duke Forest, North CarolinaJournal of Vegetation Science 115 -124 . [本文引用: 1] [31] McCullough HA 1948 ). Plant succession on fallen logs in a virgin spruce-fir forestEcology 508 -513 . [32] Natalia S Lieffers VJ Landh?usser SM 2008 ). Effects of leaf litter on the growth of boreal feather mosses: implication for forest ?oor developmentJournal of Vegetation Science 253 -260 . [本文引用: 1] [33] ódor P van Hees AFM 2004 ). Preferences of dead wood inhabiting bryophytes for decay stage, log size and habitat types in Hungarian beech forestsJournal of Bryology 79 -95 . [本文引用: 1] [34] Preik?a Z Brazaitis G Marozas V Jaroszewicz B 2015 ). Dead wood quality influences species diversity of rare cryptogams in temperate broadleaved forestsiForest 276 -285 . [本文引用: 2] [35] Rydin H 1986 ). Competition and niche separation in SphagnumCanadian Journal of Botany 1817 -1824 . [本文引用: 1] [36] Sandstr?m F Petersson H Kruys N St?hl G 2007 ). Biomass conversion factors (density and carbon concentration) by decay classes for dead wood of Pinus sylvestris, Picea abies and Betula spp. in boreal forests of SwedenForest Ecology and Management 19 -27 . [本文引用: 1] [37] Song L Ma WZ Yao YL Liu WY Li S Chen K Lu HZ Cao M Sun ZH Tan ZH Nakamura A 2015 ). Bole bryophyte diversity and distribution patterns along three altitudinal gradients in Yunnan, ChinaJournal of Vegetation Science 576 -587 . [本文引用: 1] [38] Takahashi M Sakai Y Ootomo R Shiozaki M 2000 ). Establishment of tree seedlings and water-soluble nutrients in coarse woody debris in an old-growth Picea-Abies forest in Hokkaido, northern JapanCanadian Journal of Forest Research 1148 -1155 . [本文引用: 1] [39] The Comprehensive Scientific Expedition to the Qinghai- Xizang Plateau, Academia, Sinica (1985 ). Bryoflora of Xizang , Science Press. Beijing. [本文引用: 1] [ 中国科学院青藏高原综合科学考察队 (1985 ). 西藏苔藓植物志 [本文引用: 1] [40] Vanha-Majamaa I Lilja S Ry?m? R Kotiaho JS Laaka- Lindberg S Lindberg H Puttonen P Tamminen P Toivanen T Kuuluvainen T 2007 ). Rehabilitating boreal forest structure and species composition in Finland through logging, dead wood creation and fire: the EVO experimentForest Ecology and Management 77 -88 . [本文引用: 1] [41] Wu PC 1998 ). Bryophyte Biology [本文引用: 2] [ 吴鹏程 (1998 ). 苔藓植物生物学 [本文引用: 2] [42] Wu PC 1998 ). Flora Bryophytarum Sinicorum: Vol. 6 [ 吴鹏程 (1998 ). 中国苔藓志: 6卷 [43] Wu PC Jia Y 2004 ). Flora Bryophytarum Sinicorum: Vol. 8 [本文引用: 1] [ 吴鹏程 , 贾渝 (2004 ). 中国苔藓志: 8卷 [本文引用: 1] [44] Wu PC Jia Y 2011 ). Flora Bryophytarum Sinicorum: Vol. 5 [本文引用: 1] [ 吴鹏程 , 贾渝 (2011 ). 中国苔藓志: 5卷 [本文引用: 1] [45] Yan ER Wang XH Huang JJ 2005 ). Concept and classification of coarse woody debris in forest ecosystemsActa Ecologica Sinica 158 -167 . [本文引用: 3] [ 闫恩荣 , 王希华 , 黄建军 (2005 ). 森林粗死木质残体的概念及其分类生态学报 158 -167 .] [本文引用: 3] [46] You HM 2013 ). Basical Quality and Environmental Effects of Fallen Logs in Tsuga longibracteata Forest in Tianbaoyan National Nature Reserve [本文引用: 2] [ 游惠明 (2013 ). 天宝岩长苞铁杉林倒木的基础特征及其环境效应研究 . 博士学位论文, 福建农林大学, 福州.] [本文引用: 2] 贺兰山苔藓植物物种多样性、生物量及生态学作用的研究
1
1998
... 这一系列变化与苔藓生长微生境的改变是密不可分的.倒木基质随着时间不断变化, 树皮的粗糙程度及覆盖度、结构的完整性、养分含量等都在改变(
何东进等, 2009 ), 这些基质的变化对苔藓的定植存在影响(
Dittrich et al ., 2014 ;
Kushnevskaya & Shorohova, 2018 ).另外, 倒木结构完整性、倒木离地面的距离以及倒木距离水源的距离都会影响倒木上的水分情况, 在本研究中4株倒木的水分情况随腐解程度加深而增加.而水分对于苔藓的定植很重要, 会显著影响苔藓植物的多样性(
范苗等, 2017 ).林窗作为森林中常见的小尺度干扰机制, 对于森林物种多样性维持有着重要的意义(
李强等, 2019 ), 树木倒下形成的林窗随着时间推移逐渐闭合, 这也导致不同腐解等级倒木上生长的苔藓受到的光照情况在改变.一般来说, 苔藓喜好相对遮阴的环境, 在这种环境下苔藓植物较维管植物有更大的生长优势(
白学良等, 1998 ;
刘俊华等, 2005 ).在微生境不断变化的情况下, 本研究案例中苔藓植物物种多样性、组合数量和苔藓总盖度在腐解中后期达到最大, 这与其他不同气候下、不同区域的研究趋势一致, 是倒木上苔藓植物演替的一般规律(
Cole et al ., 2008 ;
Botting & DeLong, 2009 ;
游惠明, 2013 ;
Kumar et al ., 2018 ). ...
贺兰山苔藓植物物种多样性、生物量及生态学作用的研究
1
1998
... 这一系列变化与苔藓生长微生境的改变是密不可分的.倒木基质随着时间不断变化, 树皮的粗糙程度及覆盖度、结构的完整性、养分含量等都在改变(
何东进等, 2009 ), 这些基质的变化对苔藓的定植存在影响(
Dittrich et al ., 2014 ;
Kushnevskaya & Shorohova, 2018 ).另外, 倒木结构完整性、倒木离地面的距离以及倒木距离水源的距离都会影响倒木上的水分情况, 在本研究中4株倒木的水分情况随腐解程度加深而增加.而水分对于苔藓的定植很重要, 会显著影响苔藓植物的多样性(
范苗等, 2017 ).林窗作为森林中常见的小尺度干扰机制, 对于森林物种多样性维持有着重要的意义(
李强等, 2019 ), 树木倒下形成的林窗随着时间推移逐渐闭合, 这也导致不同腐解等级倒木上生长的苔藓受到的光照情况在改变.一般来说, 苔藓喜好相对遮阴的环境, 在这种环境下苔藓植物较维管植物有更大的生长优势(
白学良等, 1998 ;
刘俊华等, 2005 ).在微生境不断变化的情况下, 本研究案例中苔藓植物物种多样性、组合数量和苔藓总盖度在腐解中后期达到最大, 这与其他不同气候下、不同区域的研究趋势一致, 是倒木上苔藓植物演替的一般规律(
Cole et al ., 2008 ;
Botting & DeLong, 2009 ;
游惠明, 2013 ;
Kumar et al ., 2018 ). ...
Macrolichen and bryophyte responses to coarse woody debris characteristics in sub- boreal spruce forest
2
2009
... 倒木主要存在于地表并与土壤层联系紧密, 具有相对稳定的环境, 是森林中苔藓植物生长的重要基质(
Preik?a et al ., 2015 ); 它也是一个不断发展的基质, 木材腐烂程度、树皮覆盖度、离地面高度以及一些小气候条件随时间连续变化(
闫恩荣等, 2005 ;
何东进等, 2009 ), 其上生长的苔藓植物也随着环境改变持续变化(
Kumar et al ., 2018 ).苔藓植物组合作为倒木群落内部次优势物种所构成的群落局部集合体(
李博等, 2000 ), 更是随着倒木的腐解进程发生着演替(
Kushnevskaya et al ., 2007 ).一系列的研究探讨了倒木上苔藓植物物种组成和丰度的变化与倒木自身性质以及林地环境的相互关系(
McAlister, 1997 ;
Dittrich et al ., 2014 ;
Preik?a et al ., 2015 ).但由于变量间存在复杂的共线性关系, 要找到直接作用于苔藓组合的生态因子, 并从中抽离出倒木上苔藓植物组合演替规律是困难的(
Botting & DeLong, 2009 ). ...
... 这一系列变化与苔藓生长微生境的改变是密不可分的.倒木基质随着时间不断变化, 树皮的粗糙程度及覆盖度、结构的完整性、养分含量等都在改变(
何东进等, 2009 ), 这些基质的变化对苔藓的定植存在影响(
Dittrich et al ., 2014 ;
Kushnevskaya & Shorohova, 2018 ).另外, 倒木结构完整性、倒木离地面的距离以及倒木距离水源的距离都会影响倒木上的水分情况, 在本研究中4株倒木的水分情况随腐解程度加深而增加.而水分对于苔藓的定植很重要, 会显著影响苔藓植物的多样性(
范苗等, 2017 ).林窗作为森林中常见的小尺度干扰机制, 对于森林物种多样性维持有着重要的意义(
李强等, 2019 ), 树木倒下形成的林窗随着时间推移逐渐闭合, 这也导致不同腐解等级倒木上生长的苔藓受到的光照情况在改变.一般来说, 苔藓喜好相对遮阴的环境, 在这种环境下苔藓植物较维管植物有更大的生长优势(
白学良等, 1998 ;
刘俊华等, 2005 ).在微生境不断变化的情况下, 本研究案例中苔藓植物物种多样性、组合数量和苔藓总盖度在腐解中后期达到最大, 这与其他不同气候下、不同区域的研究趋势一致, 是倒木上苔藓植物演替的一般规律(
Cole et al ., 2008 ;
Botting & DeLong, 2009 ;
游惠明, 2013 ;
Kumar et al ., 2018 ). ...
The effect of temperature on growth and competition between Sphagnum species
1
2008
... 植物之间的相互作用也在影响着演替的过程.随着时间的推移, 倒木木质素等难分解组分降解程度增加, 结构相对疏松, 树干湿度增加, 倒木上环境逐渐优化, 蕨类植物、种子植物开始在倒木上及倒木周边定植, 一定程度上挤压了苔藓的生存空间, 加之枯落物的不断累积, 占据了部分可定植的空间, 对苔藓植物的生长造成了负面的影响(
Natalia et al ., 2008 ;
Márialigeti et al ., 2009 ), 使得苔藓植物盖度在到达极大值后逐渐减小.另外苔藓植物之间也存在合作、竞争等关系(
卜兆君等, 2009 ).倒木作为一个相对变化快速的生境, 其上生长的苔藓物种和组合通常是不稳定的, 在这种情况下种间关系常趋于负相关, 即种间竞争或排他作用强烈(
刘艳等, 2019 ), 例如青毛藓、丛叶青毛藓和长叶青毛藓都分布于光照较为充足的倒木2和倒木4, 但是同一个样方内通常仅有一种青毛藓属植物.苔藓植物的竞争等级还会随着环境的变化而改变(
Breeuwer et al ., 2008 ), 从而推动不同演替阶段苔藓物种以及物种组合的变化.早期优势的丛生型藓类能较好地适应相对干旱、强光照的环境, 但随着环境向潮湿荫蔽转变, 个体更大且生长快速的垫状毯状藓类出现, 它们能获取更多的资源, 不对称竞争导致竞争等级的改变.随着更多苔藓物种的定植, 倒木上苔藓间竞争压力逐渐增大, 这种情况下物种间生态位的分化是物种共存的重要机理之一(
Rydin, 1986 ), 本研究中苔藓植物组合内生活型种类和数量即整体空间结构复杂程度随着腐解等级及其他生境条件的变化而逐渐增大, 呈现出类似于植物演替过程中的生态位分化. ...
苔藓植物相互作用的研究进展
1
2009
... 植物之间的相互作用也在影响着演替的过程.随着时间的推移, 倒木木质素等难分解组分降解程度增加, 结构相对疏松, 树干湿度增加, 倒木上环境逐渐优化, 蕨类植物、种子植物开始在倒木上及倒木周边定植, 一定程度上挤压了苔藓的生存空间, 加之枯落物的不断累积, 占据了部分可定植的空间, 对苔藓植物的生长造成了负面的影响(
Natalia et al ., 2008 ;
Márialigeti et al ., 2009 ), 使得苔藓植物盖度在到达极大值后逐渐减小.另外苔藓植物之间也存在合作、竞争等关系(
卜兆君等, 2009 ).倒木作为一个相对变化快速的生境, 其上生长的苔藓物种和组合通常是不稳定的, 在这种情况下种间关系常趋于负相关, 即种间竞争或排他作用强烈(
刘艳等, 2019 ), 例如青毛藓、丛叶青毛藓和长叶青毛藓都分布于光照较为充足的倒木2和倒木4, 但是同一个样方内通常仅有一种青毛藓属植物.苔藓植物的竞争等级还会随着环境的变化而改变(
Breeuwer et al ., 2008 ), 从而推动不同演替阶段苔藓物种以及物种组合的变化.早期优势的丛生型藓类能较好地适应相对干旱、强光照的环境, 但随着环境向潮湿荫蔽转变, 个体更大且生长快速的垫状毯状藓类出现, 它们能获取更多的资源, 不对称竞争导致竞争等级的改变.随着更多苔藓物种的定植, 倒木上苔藓间竞争压力逐渐增大, 这种情况下物种间生态位的分化是物种共存的重要机理之一(
Rydin, 1986 ), 本研究中苔藓植物组合内生活型种类和数量即整体空间结构复杂程度随着腐解等级及其他生境条件的变化而逐渐增大, 呈现出类似于植物演替过程中的生态位分化. ...
苔藓植物相互作用的研究进展
1
2009
... 植物之间的相互作用也在影响着演替的过程.随着时间的推移, 倒木木质素等难分解组分降解程度增加, 结构相对疏松, 树干湿度增加, 倒木上环境逐渐优化, 蕨类植物、种子植物开始在倒木上及倒木周边定植, 一定程度上挤压了苔藓的生存空间, 加之枯落物的不断累积, 占据了部分可定植的空间, 对苔藓植物的生长造成了负面的影响(
Natalia et al ., 2008 ;
Márialigeti et al ., 2009 ), 使得苔藓植物盖度在到达极大值后逐渐减小.另外苔藓植物之间也存在合作、竞争等关系(
卜兆君等, 2009 ).倒木作为一个相对变化快速的生境, 其上生长的苔藓物种和组合通常是不稳定的, 在这种情况下种间关系常趋于负相关, 即种间竞争或排他作用强烈(
刘艳等, 2019 ), 例如青毛藓、丛叶青毛藓和长叶青毛藓都分布于光照较为充足的倒木2和倒木4, 但是同一个样方内通常仅有一种青毛藓属植物.苔藓植物的竞争等级还会随着环境的变化而改变(
Breeuwer et al ., 2008 ), 从而推动不同演替阶段苔藓物种以及物种组合的变化.早期优势的丛生型藓类能较好地适应相对干旱、强光照的环境, 但随着环境向潮湿荫蔽转变, 个体更大且生长快速的垫状毯状藓类出现, 它们能获取更多的资源, 不对称竞争导致竞争等级的改变.随着更多苔藓物种的定植, 倒木上苔藓间竞争压力逐渐增大, 这种情况下物种间生态位的分化是物种共存的重要机理之一(
Rydin, 1986 ), 本研究中苔藓植物组合内生活型种类和数量即整体空间结构复杂程度随着腐解等级及其他生境条件的变化而逐渐增大, 呈现出类似于植物演替过程中的生态位分化. ...
Influence of microhabitat on bryophyte diversity in Ontario mixedwood boreal forest
1
2008
... 这一系列变化与苔藓生长微生境的改变是密不可分的.倒木基质随着时间不断变化, 树皮的粗糙程度及覆盖度、结构的完整性、养分含量等都在改变(
何东进等, 2009 ), 这些基质的变化对苔藓的定植存在影响(
Dittrich et al ., 2014 ;
Kushnevskaya & Shorohova, 2018 ).另外, 倒木结构完整性、倒木离地面的距离以及倒木距离水源的距离都会影响倒木上的水分情况, 在本研究中4株倒木的水分情况随腐解程度加深而增加.而水分对于苔藓的定植很重要, 会显著影响苔藓植物的多样性(
范苗等, 2017 ).林窗作为森林中常见的小尺度干扰机制, 对于森林物种多样性维持有着重要的意义(
李强等, 2019 ), 树木倒下形成的林窗随着时间推移逐渐闭合, 这也导致不同腐解等级倒木上生长的苔藓受到的光照情况在改变.一般来说, 苔藓喜好相对遮阴的环境, 在这种环境下苔藓植物较维管植物有更大的生长优势(
白学良等, 1998 ;
刘俊华等, 2005 ).在微生境不断变化的情况下, 本研究案例中苔藓植物物种多样性、组合数量和苔藓总盖度在腐解中后期达到最大, 这与其他不同气候下、不同区域的研究趋势一致, 是倒木上苔藓植物演替的一般规律(
Cole et al ., 2008 ;
Botting & DeLong, 2009 ;
游惠明, 2013 ;
Kumar et al ., 2018 ). ...
Influence of coarse woody debris on herpetofaunal communities in upland pine stands of the southeastern Coastal Plain
1
2010
... 倒木属于粗死木质残体(coarse woody debris, CWD)的一种(
闫恩荣等, 2005 ).在传统的森林管理过程中, 倒木常被误认为容易引起病虫害和火灾而被清除.近年来, 许多****对森林生态系统的深入研究结果表明, 倒木是森林生态系统不可缺少的重要组成部分, 在减少水土流失、提高林地土壤肥力和加速养分循环等方面发挥着重要作用(
Sandstr?m et al ., 2007 ;
Vanha-Majamaa et al ., 2007 ;
Davis et al ., 2010 ).倒木作为森林生态系统中的持续营养源, 是种类繁多的生物赖以生存的蔽护所和繁殖场地, 也是林木更新的良好苗床, 对森林生态系统的完整性和生物多样性的维持具有重要意义(
Takahashi et al ., 2000 ;
何东进等, 2009 ). ...
The significance of deadwood for total bryophyte, lichen, and vascular plant diversity in an old-growth spruce forest
3
2014
... 在早期森林生态系统的研究中, ****们对倒木的关注极少.1948年, McCullough研究了美国天然云杉(
Picea asperata )林3种生境下腐烂倒木上的植物演替, 认为其模式为: 地衣和苔类植物→藓类植物→草本和矮灌木→云杉、冷杉(
Abies fabri )等乔木树种.
Fogel和Cromack (1977) 提出倒木腐烂等级的五级系统, 并研究了倒木的养分变化.20世纪90年代, 倒木的生态功能以及动态特征的研究开始逐渐增加.倒木与生物多样性维持的研究, 主要围绕微生物多样性(木材腐朽菌及大型真菌)和动物多样性展开研究, 也有少量研究涉及植物(如苔藓、地衣)多样性(
游惠明, 2013 ), 在倒木上栖息的植物类型中, 苔藓植物是最大的植物类群(
Dittrich et al ., 2014 ). ...
... 倒木主要存在于地表并与土壤层联系紧密, 具有相对稳定的环境, 是森林中苔藓植物生长的重要基质(
Preik?a et al ., 2015 ); 它也是一个不断发展的基质, 木材腐烂程度、树皮覆盖度、离地面高度以及一些小气候条件随时间连续变化(
闫恩荣等, 2005 ;
何东进等, 2009 ), 其上生长的苔藓植物也随着环境改变持续变化(
Kumar et al ., 2018 ).苔藓植物组合作为倒木群落内部次优势物种所构成的群落局部集合体(
李博等, 2000 ), 更是随着倒木的腐解进程发生着演替(
Kushnevskaya et al ., 2007 ).一系列的研究探讨了倒木上苔藓植物物种组成和丰度的变化与倒木自身性质以及林地环境的相互关系(
McAlister, 1997 ;
Dittrich et al ., 2014 ;
Preik?a et al ., 2015 ).但由于变量间存在复杂的共线性关系, 要找到直接作用于苔藓组合的生态因子, 并从中抽离出倒木上苔藓植物组合演替规律是困难的(
Botting & DeLong, 2009 ). ...
... 这一系列变化与苔藓生长微生境的改变是密不可分的.倒木基质随着时间不断变化, 树皮的粗糙程度及覆盖度、结构的完整性、养分含量等都在改变(
何东进等, 2009 ), 这些基质的变化对苔藓的定植存在影响(
Dittrich et al ., 2014 ;
Kushnevskaya & Shorohova, 2018 ).另外, 倒木结构完整性、倒木离地面的距离以及倒木距离水源的距离都会影响倒木上的水分情况, 在本研究中4株倒木的水分情况随腐解程度加深而增加.而水分对于苔藓的定植很重要, 会显著影响苔藓植物的多样性(
范苗等, 2017 ).林窗作为森林中常见的小尺度干扰机制, 对于森林物种多样性维持有着重要的意义(
李强等, 2019 ), 树木倒下形成的林窗随着时间推移逐渐闭合, 这也导致不同腐解等级倒木上生长的苔藓受到的光照情况在改变.一般来说, 苔藓喜好相对遮阴的环境, 在这种环境下苔藓植物较维管植物有更大的生长优势(
白学良等, 1998 ;
刘俊华等, 2005 ).在微生境不断变化的情况下, 本研究案例中苔藓植物物种多样性、组合数量和苔藓总盖度在腐解中后期达到最大, 这与其他不同气候下、不同区域的研究趋势一致, 是倒木上苔藓植物演替的一般规律(
Cole et al ., 2008 ;
Botting & DeLong, 2009 ;
游惠明, 2013 ;
Kumar et al ., 2018 ). ...
武汉市城区苔藓植物多样性和分布及与环境因子的关系
2
2017
... 对不同组合内各苔藓植物重要值进行计算(
Song et al ., 2015 ;
范苗等, 2017 ). ...
... 这一系列变化与苔藓生长微生境的改变是密不可分的.倒木基质随着时间不断变化, 树皮的粗糙程度及覆盖度、结构的完整性、养分含量等都在改变(
何东进等, 2009 ), 这些基质的变化对苔藓的定植存在影响(
Dittrich et al ., 2014 ;
Kushnevskaya & Shorohova, 2018 ).另外, 倒木结构完整性、倒木离地面的距离以及倒木距离水源的距离都会影响倒木上的水分情况, 在本研究中4株倒木的水分情况随腐解程度加深而增加.而水分对于苔藓的定植很重要, 会显著影响苔藓植物的多样性(
范苗等, 2017 ).林窗作为森林中常见的小尺度干扰机制, 对于森林物种多样性维持有着重要的意义(
李强等, 2019 ), 树木倒下形成的林窗随着时间推移逐渐闭合, 这也导致不同腐解等级倒木上生长的苔藓受到的光照情况在改变.一般来说, 苔藓喜好相对遮阴的环境, 在这种环境下苔藓植物较维管植物有更大的生长优势(
白学良等, 1998 ;
刘俊华等, 2005 ).在微生境不断变化的情况下, 本研究案例中苔藓植物物种多样性、组合数量和苔藓总盖度在腐解中后期达到最大, 这与其他不同气候下、不同区域的研究趋势一致, 是倒木上苔藓植物演替的一般规律(
Cole et al ., 2008 ;
Botting & DeLong, 2009 ;
游惠明, 2013 ;
Kumar et al ., 2018 ). ...
武汉市城区苔藓植物多样性和分布及与环境因子的关系
2
2017
... 对不同组合内各苔藓植物重要值进行计算(
Song et al ., 2015 ;
范苗等, 2017 ). ...
... 这一系列变化与苔藓生长微生境的改变是密不可分的.倒木基质随着时间不断变化, 树皮的粗糙程度及覆盖度、结构的完整性、养分含量等都在改变(
何东进等, 2009 ), 这些基质的变化对苔藓的定植存在影响(
Dittrich et al ., 2014 ;
Kushnevskaya & Shorohova, 2018 ).另外, 倒木结构完整性、倒木离地面的距离以及倒木距离水源的距离都会影响倒木上的水分情况, 在本研究中4株倒木的水分情况随腐解程度加深而增加.而水分对于苔藓的定植很重要, 会显著影响苔藓植物的多样性(
范苗等, 2017 ).林窗作为森林中常见的小尺度干扰机制, 对于森林物种多样性维持有着重要的意义(
李强等, 2019 ), 树木倒下形成的林窗随着时间推移逐渐闭合, 这也导致不同腐解等级倒木上生长的苔藓受到的光照情况在改变.一般来说, 苔藓喜好相对遮阴的环境, 在这种环境下苔藓植物较维管植物有更大的生长优势(
白学良等, 1998 ;
刘俊华等, 2005 ).在微生境不断变化的情况下, 本研究案例中苔藓植物物种多样性、组合数量和苔藓总盖度在腐解中后期达到最大, 这与其他不同气候下、不同区域的研究趋势一致, 是倒木上苔藓植物演替的一般规律(
Cole et al ., 2008 ;
Botting & DeLong, 2009 ;
游惠明, 2013 ;
Kumar et al ., 2018 ). ...
Effect of habitat and substrate quality on Douglas-fir litter decomposition in western Oregon
1
1977
... 在早期森林生态系统的研究中, ****们对倒木的关注极少.1948年, McCullough研究了美国天然云杉(
Picea asperata )林3种生境下腐烂倒木上的植物演替, 认为其模式为: 地衣和苔类植物→藓类植物→草本和矮灌木→云杉、冷杉(
Abies fabri )等乔木树种.
Fogel和Cromack (1977) 提出倒木腐烂等级的五级系统, 并研究了倒木的养分变化.20世纪90年代, 倒木的生态功能以及动态特征的研究开始逐渐增加.倒木与生物多样性维持的研究, 主要围绕微生物多样性(木材腐朽菌及大型真菌)和动物多样性展开研究, 也有少量研究涉及植物(如苔藓、地衣)多样性(
游惠明, 2013 ), 在倒木上栖息的植物类型中, 苔藓植物是最大的植物类群(
Dittrich et al ., 2014 ). ...
1
1994
... 将苔藓植物标本带回实验室阴干、装袋, 登记采集信息, 参考《西藏苔藓植物志》(
中国科学院青藏高原综合科学考察队, 1985 )、《中国苔藓志》(
高谦, 1994 ,
1996 ,
2003 ;
吴鹏程, 2002 ;
吴鹏程和贾渝, 2004 ,
2011 ;
胡人亮和王幼芳, 2005 ;
黎兴江, 2006 ;
高谦和吴玉环, 2008 )、《中国生物物种名录: 1. 植物: 苔藓植物》(
贾渝和何思, 2013 )以及近年来相关物种的研究资料(
李敏等, 2016 )对标本进行解剖鉴定, 参考美国密苏里植物园Tropicos网站(http://www.tropicos. org/)对苔藓物种名录进行整理归纳, 并根据当时所拍摄照片, 确定每个样方中每种苔藓的分盖度.本论文所采用的标本存放于中国农业大学标本室(BAU). ...
1
1994
... 将苔藓植物标本带回实验室阴干、装袋, 登记采集信息, 参考《西藏苔藓植物志》(
中国科学院青藏高原综合科学考察队, 1985 )、《中国苔藓志》(
高谦, 1994 ,
1996 ,
2003 ;
吴鹏程, 2002 ;
吴鹏程和贾渝, 2004 ,
2011 ;
胡人亮和王幼芳, 2005 ;
黎兴江, 2006 ;
高谦和吴玉环, 2008 )、《中国生物物种名录: 1. 植物: 苔藓植物》(
贾渝和何思, 2013 )以及近年来相关物种的研究资料(
李敏等, 2016 )对标本进行解剖鉴定, 参考美国密苏里植物园Tropicos网站(http://www.tropicos. org/)对苔藓物种名录进行整理归纳, 并根据当时所拍摄照片, 确定每个样方中每种苔藓的分盖度.本论文所采用的标本存放于中国农业大学标本室(BAU). ...
1
1996
... 将苔藓植物标本带回实验室阴干、装袋, 登记采集信息, 参考《西藏苔藓植物志》(
中国科学院青藏高原综合科学考察队, 1985 )、《中国苔藓志》(
高谦, 1994 ,
1996 ,
2003 ;
吴鹏程, 2002 ;
吴鹏程和贾渝, 2004 ,
2011 ;
胡人亮和王幼芳, 2005 ;
黎兴江, 2006 ;
高谦和吴玉环, 2008 )、《中国生物物种名录: 1. 植物: 苔藓植物》(
贾渝和何思, 2013 )以及近年来相关物种的研究资料(
李敏等, 2016 )对标本进行解剖鉴定, 参考美国密苏里植物园Tropicos网站(http://www.tropicos. org/)对苔藓物种名录进行整理归纳, 并根据当时所拍摄照片, 确定每个样方中每种苔藓的分盖度.本论文所采用的标本存放于中国农业大学标本室(BAU). ...
1
1996
... 将苔藓植物标本带回实验室阴干、装袋, 登记采集信息, 参考《西藏苔藓植物志》(
中国科学院青藏高原综合科学考察队, 1985 )、《中国苔藓志》(
高谦, 1994 ,
1996 ,
2003 ;
吴鹏程, 2002 ;
吴鹏程和贾渝, 2004 ,
2011 ;
胡人亮和王幼芳, 2005 ;
黎兴江, 2006 ;
高谦和吴玉环, 2008 )、《中国生物物种名录: 1. 植物: 苔藓植物》(
贾渝和何思, 2013 )以及近年来相关物种的研究资料(
李敏等, 2016 )对标本进行解剖鉴定, 参考美国密苏里植物园Tropicos网站(http://www.tropicos. org/)对苔藓物种名录进行整理归纳, 并根据当时所拍摄照片, 确定每个样方中每种苔藓的分盖度.本论文所采用的标本存放于中国农业大学标本室(BAU). ...
1
2003
... 将苔藓植物标本带回实验室阴干、装袋, 登记采集信息, 参考《西藏苔藓植物志》(
中国科学院青藏高原综合科学考察队, 1985 )、《中国苔藓志》(
高谦, 1994 ,
1996 ,
2003 ;
吴鹏程, 2002 ;
吴鹏程和贾渝, 2004 ,
2011 ;
胡人亮和王幼芳, 2005 ;
黎兴江, 2006 ;
高谦和吴玉环, 2008 )、《中国生物物种名录: 1. 植物: 苔藓植物》(
贾渝和何思, 2013 )以及近年来相关物种的研究资料(
李敏等, 2016 )对标本进行解剖鉴定, 参考美国密苏里植物园Tropicos网站(http://www.tropicos. org/)对苔藓物种名录进行整理归纳, 并根据当时所拍摄照片, 确定每个样方中每种苔藓的分盖度.本论文所采用的标本存放于中国农业大学标本室(BAU). ...
1
2003
... 将苔藓植物标本带回实验室阴干、装袋, 登记采集信息, 参考《西藏苔藓植物志》(
中国科学院青藏高原综合科学考察队, 1985 )、《中国苔藓志》(
高谦, 1994 ,
1996 ,
2003 ;
吴鹏程, 2002 ;
吴鹏程和贾渝, 2004 ,
2011 ;
胡人亮和王幼芳, 2005 ;
黎兴江, 2006 ;
高谦和吴玉环, 2008 )、《中国生物物种名录: 1. 植物: 苔藓植物》(
贾渝和何思, 2013 )以及近年来相关物种的研究资料(
李敏等, 2016 )对标本进行解剖鉴定, 参考美国密苏里植物园Tropicos网站(http://www.tropicos. org/)对苔藓物种名录进行整理归纳, 并根据当时所拍摄照片, 确定每个样方中每种苔藓的分盖度.本论文所采用的标本存放于中国农业大学标本室(BAU). ...
1
2008
... 将苔藓植物标本带回实验室阴干、装袋, 登记采集信息, 参考《西藏苔藓植物志》(
中国科学院青藏高原综合科学考察队, 1985 )、《中国苔藓志》(
高谦, 1994 ,
1996 ,
2003 ;
吴鹏程, 2002 ;
吴鹏程和贾渝, 2004 ,
2011 ;
胡人亮和王幼芳, 2005 ;
黎兴江, 2006 ;
高谦和吴玉环, 2008 )、《中国生物物种名录: 1. 植物: 苔藓植物》(
贾渝和何思, 2013 )以及近年来相关物种的研究资料(
李敏等, 2016 )对标本进行解剖鉴定, 参考美国密苏里植物园Tropicos网站(http://www.tropicos. org/)对苔藓物种名录进行整理归纳, 并根据当时所拍摄照片, 确定每个样方中每种苔藓的分盖度.本论文所采用的标本存放于中国农业大学标本室(BAU). ...
1
2008
... 将苔藓植物标本带回实验室阴干、装袋, 登记采集信息, 参考《西藏苔藓植物志》(
中国科学院青藏高原综合科学考察队, 1985 )、《中国苔藓志》(
高谦, 1994 ,
1996 ,
2003 ;
吴鹏程, 2002 ;
吴鹏程和贾渝, 2004 ,
2011 ;
胡人亮和王幼芳, 2005 ;
黎兴江, 2006 ;
高谦和吴玉环, 2008 )、《中国生物物种名录: 1. 植物: 苔藓植物》(
贾渝和何思, 2013 )以及近年来相关物种的研究资料(
李敏等, 2016 )对标本进行解剖鉴定, 参考美国密苏里植物园Tropicos网站(http://www.tropicos. org/)对苔藓物种名录进行整理归纳, 并根据当时所拍摄照片, 确定每个样方中每种苔藓的分盖度.本论文所采用的标本存放于中国农业大学标本室(BAU). ...
森林生态系统粗死木质残体的研究进展
3
2009
... 倒木属于粗死木质残体(coarse woody debris, CWD)的一种(
闫恩荣等, 2005 ).在传统的森林管理过程中, 倒木常被误认为容易引起病虫害和火灾而被清除.近年来, 许多****对森林生态系统的深入研究结果表明, 倒木是森林生态系统不可缺少的重要组成部分, 在减少水土流失、提高林地土壤肥力和加速养分循环等方面发挥着重要作用(
Sandstr?m et al ., 2007 ;
Vanha-Majamaa et al ., 2007 ;
Davis et al ., 2010 ).倒木作为森林生态系统中的持续营养源, 是种类繁多的生物赖以生存的蔽护所和繁殖场地, 也是林木更新的良好苗床, 对森林生态系统的完整性和生物多样性的维持具有重要意义(
Takahashi et al ., 2000 ;
何东进等, 2009 ). ...
... 倒木主要存在于地表并与土壤层联系紧密, 具有相对稳定的环境, 是森林中苔藓植物生长的重要基质(
Preik?a et al ., 2015 ); 它也是一个不断发展的基质, 木材腐烂程度、树皮覆盖度、离地面高度以及一些小气候条件随时间连续变化(
闫恩荣等, 2005 ;
何东进等, 2009 ), 其上生长的苔藓植物也随着环境改变持续变化(
Kumar et al ., 2018 ).苔藓植物组合作为倒木群落内部次优势物种所构成的群落局部集合体(
李博等, 2000 ), 更是随着倒木的腐解进程发生着演替(
Kushnevskaya et al ., 2007 ).一系列的研究探讨了倒木上苔藓植物物种组成和丰度的变化与倒木自身性质以及林地环境的相互关系(
McAlister, 1997 ;
Dittrich et al ., 2014 ;
Preik?a et al ., 2015 ).但由于变量间存在复杂的共线性关系, 要找到直接作用于苔藓组合的生态因子, 并从中抽离出倒木上苔藓植物组合演替规律是困难的(
Botting & DeLong, 2009 ). ...
... 这一系列变化与苔藓生长微生境的改变是密不可分的.倒木基质随着时间不断变化, 树皮的粗糙程度及覆盖度、结构的完整性、养分含量等都在改变(
何东进等, 2009 ), 这些基质的变化对苔藓的定植存在影响(
Dittrich et al ., 2014 ;
Kushnevskaya & Shorohova, 2018 ).另外, 倒木结构完整性、倒木离地面的距离以及倒木距离水源的距离都会影响倒木上的水分情况, 在本研究中4株倒木的水分情况随腐解程度加深而增加.而水分对于苔藓的定植很重要, 会显著影响苔藓植物的多样性(
范苗等, 2017 ).林窗作为森林中常见的小尺度干扰机制, 对于森林物种多样性维持有着重要的意义(
李强等, 2019 ), 树木倒下形成的林窗随着时间推移逐渐闭合, 这也导致不同腐解等级倒木上生长的苔藓受到的光照情况在改变.一般来说, 苔藓喜好相对遮阴的环境, 在这种环境下苔藓植物较维管植物有更大的生长优势(
白学良等, 1998 ;
刘俊华等, 2005 ).在微生境不断变化的情况下, 本研究案例中苔藓植物物种多样性、组合数量和苔藓总盖度在腐解中后期达到最大, 这与其他不同气候下、不同区域的研究趋势一致, 是倒木上苔藓植物演替的一般规律(
Cole et al ., 2008 ;
Botting & DeLong, 2009 ;
游惠明, 2013 ;
Kumar et al ., 2018 ). ...
森林生态系统粗死木质残体的研究进展
3
2009
... 倒木属于粗死木质残体(coarse woody debris, CWD)的一种(
闫恩荣等, 2005 ).在传统的森林管理过程中, 倒木常被误认为容易引起病虫害和火灾而被清除.近年来, 许多****对森林生态系统的深入研究结果表明, 倒木是森林生态系统不可缺少的重要组成部分, 在减少水土流失、提高林地土壤肥力和加速养分循环等方面发挥着重要作用(
Sandstr?m et al ., 2007 ;
Vanha-Majamaa et al ., 2007 ;
Davis et al ., 2010 ).倒木作为森林生态系统中的持续营养源, 是种类繁多的生物赖以生存的蔽护所和繁殖场地, 也是林木更新的良好苗床, 对森林生态系统的完整性和生物多样性的维持具有重要意义(
Takahashi et al ., 2000 ;
何东进等, 2009 ). ...
... 倒木主要存在于地表并与土壤层联系紧密, 具有相对稳定的环境, 是森林中苔藓植物生长的重要基质(
Preik?a et al ., 2015 ); 它也是一个不断发展的基质, 木材腐烂程度、树皮覆盖度、离地面高度以及一些小气候条件随时间连续变化(
闫恩荣等, 2005 ;
何东进等, 2009 ), 其上生长的苔藓植物也随着环境改变持续变化(
Kumar et al ., 2018 ).苔藓植物组合作为倒木群落内部次优势物种所构成的群落局部集合体(
李博等, 2000 ), 更是随着倒木的腐解进程发生着演替(
Kushnevskaya et al ., 2007 ).一系列的研究探讨了倒木上苔藓植物物种组成和丰度的变化与倒木自身性质以及林地环境的相互关系(
McAlister, 1997 ;
Dittrich et al ., 2014 ;
Preik?a et al ., 2015 ).但由于变量间存在复杂的共线性关系, 要找到直接作用于苔藓组合的生态因子, 并从中抽离出倒木上苔藓植物组合演替规律是困难的(
Botting & DeLong, 2009 ). ...
... 这一系列变化与苔藓生长微生境的改变是密不可分的.倒木基质随着时间不断变化, 树皮的粗糙程度及覆盖度、结构的完整性、养分含量等都在改变(
何东进等, 2009 ), 这些基质的变化对苔藓的定植存在影响(
Dittrich et al ., 2014 ;
Kushnevskaya & Shorohova, 2018 ).另外, 倒木结构完整性、倒木离地面的距离以及倒木距离水源的距离都会影响倒木上的水分情况, 在本研究中4株倒木的水分情况随腐解程度加深而增加.而水分对于苔藓的定植很重要, 会显著影响苔藓植物的多样性(
范苗等, 2017 ).林窗作为森林中常见的小尺度干扰机制, 对于森林物种多样性维持有着重要的意义(
李强等, 2019 ), 树木倒下形成的林窗随着时间推移逐渐闭合, 这也导致不同腐解等级倒木上生长的苔藓受到的光照情况在改变.一般来说, 苔藓喜好相对遮阴的环境, 在这种环境下苔藓植物较维管植物有更大的生长优势(
白学良等, 1998 ;
刘俊华等, 2005 ).在微生境不断变化的情况下, 本研究案例中苔藓植物物种多样性、组合数量和苔藓总盖度在腐解中后期达到最大, 这与其他不同气候下、不同区域的研究趋势一致, 是倒木上苔藓植物演替的一般规律(
Cole et al ., 2008 ;
Botting & DeLong, 2009 ;
游惠明, 2013 ;
Kumar et al ., 2018 ). ...
1
2005
... 将苔藓植物标本带回实验室阴干、装袋, 登记采集信息, 参考《西藏苔藓植物志》(
中国科学院青藏高原综合科学考察队, 1985 )、《中国苔藓志》(
高谦, 1994 ,
1996 ,
2003 ;
吴鹏程, 2002 ;
吴鹏程和贾渝, 2004 ,
2011 ;
胡人亮和王幼芳, 2005 ;
黎兴江, 2006 ;
高谦和吴玉环, 2008 )、《中国生物物种名录: 1. 植物: 苔藓植物》(
贾渝和何思, 2013 )以及近年来相关物种的研究资料(
李敏等, 2016 )对标本进行解剖鉴定, 参考美国密苏里植物园Tropicos网站(http://www.tropicos. org/)对苔藓物种名录进行整理归纳, 并根据当时所拍摄照片, 确定每个样方中每种苔藓的分盖度.本论文所采用的标本存放于中国农业大学标本室(BAU). ...
1
2005
... 将苔藓植物标本带回实验室阴干、装袋, 登记采集信息, 参考《西藏苔藓植物志》(
中国科学院青藏高原综合科学考察队, 1985 )、《中国苔藓志》(
高谦, 1994 ,
1996 ,
2003 ;
吴鹏程, 2002 ;
吴鹏程和贾渝, 2004 ,
2011 ;
胡人亮和王幼芳, 2005 ;
黎兴江, 2006 ;
高谦和吴玉环, 2008 )、《中国生物物种名录: 1. 植物: 苔藓植物》(
贾渝和何思, 2013 )以及近年来相关物种的研究资料(
李敏等, 2016 )对标本进行解剖鉴定, 参考美国密苏里植物园Tropicos网站(http://www.tropicos. org/)对苔藓物种名录进行整理归纳, 并根据当时所拍摄照片, 确定每个样方中每种苔藓的分盖度.本论文所采用的标本存放于中国农业大学标本室(BAU). ...
1
2013
... 将苔藓植物标本带回实验室阴干、装袋, 登记采集信息, 参考《西藏苔藓植物志》(
中国科学院青藏高原综合科学考察队, 1985 )、《中国苔藓志》(
高谦, 1994 ,
1996 ,
2003 ;
吴鹏程, 2002 ;
吴鹏程和贾渝, 2004 ,
2011 ;
胡人亮和王幼芳, 2005 ;
黎兴江, 2006 ;
高谦和吴玉环, 2008 )、《中国生物物种名录: 1. 植物: 苔藓植物》(
贾渝和何思, 2013 )以及近年来相关物种的研究资料(
李敏等, 2016 )对标本进行解剖鉴定, 参考美国密苏里植物园Tropicos网站(http://www.tropicos. org/)对苔藓物种名录进行整理归纳, 并根据当时所拍摄照片, 确定每个样方中每种苔藓的分盖度.本论文所采用的标本存放于中国农业大学标本室(BAU). ...
1
2013
... 将苔藓植物标本带回实验室阴干、装袋, 登记采集信息, 参考《西藏苔藓植物志》(
中国科学院青藏高原综合科学考察队, 1985 )、《中国苔藓志》(
高谦, 1994 ,
1996 ,
2003 ;
吴鹏程, 2002 ;
吴鹏程和贾渝, 2004 ,
2011 ;
胡人亮和王幼芳, 2005 ;
黎兴江, 2006 ;
高谦和吴玉环, 2008 )、《中国生物物种名录: 1. 植物: 苔藓植物》(
贾渝和何思, 2013 )以及近年来相关物种的研究资料(
李敏等, 2016 )对标本进行解剖鉴定, 参考美国密苏里植物园Tropicos网站(http://www.tropicos. org/)对苔藓物种名录进行整理归纳, 并根据当时所拍摄照片, 确定每个样方中每种苔藓的分盖度.本论文所采用的标本存放于中国农业大学标本室(BAU). ...
Epixylic vegetation abundance, diversity, and composition vary with coarse woody debris decay class and substrate species in boreal forest
2
2018
... 倒木主要存在于地表并与土壤层联系紧密, 具有相对稳定的环境, 是森林中苔藓植物生长的重要基质(
Preik?a et al ., 2015 ); 它也是一个不断发展的基质, 木材腐烂程度、树皮覆盖度、离地面高度以及一些小气候条件随时间连续变化(
闫恩荣等, 2005 ;
何东进等, 2009 ), 其上生长的苔藓植物也随着环境改变持续变化(
Kumar et al ., 2018 ).苔藓植物组合作为倒木群落内部次优势物种所构成的群落局部集合体(
李博等, 2000 ), 更是随着倒木的腐解进程发生着演替(
Kushnevskaya et al ., 2007 ).一系列的研究探讨了倒木上苔藓植物物种组成和丰度的变化与倒木自身性质以及林地环境的相互关系(
McAlister, 1997 ;
Dittrich et al ., 2014 ;
Preik?a et al ., 2015 ).但由于变量间存在复杂的共线性关系, 要找到直接作用于苔藓组合的生态因子, 并从中抽离出倒木上苔藓植物组合演替规律是困难的(
Botting & DeLong, 2009 ). ...
... 这一系列变化与苔藓生长微生境的改变是密不可分的.倒木基质随着时间不断变化, 树皮的粗糙程度及覆盖度、结构的完整性、养分含量等都在改变(
何东进等, 2009 ), 这些基质的变化对苔藓的定植存在影响(
Dittrich et al ., 2014 ;
Kushnevskaya & Shorohova, 2018 ).另外, 倒木结构完整性、倒木离地面的距离以及倒木距离水源的距离都会影响倒木上的水分情况, 在本研究中4株倒木的水分情况随腐解程度加深而增加.而水分对于苔藓的定植很重要, 会显著影响苔藓植物的多样性(
范苗等, 2017 ).林窗作为森林中常见的小尺度干扰机制, 对于森林物种多样性维持有着重要的意义(
李强等, 2019 ), 树木倒下形成的林窗随着时间推移逐渐闭合, 这也导致不同腐解等级倒木上生长的苔藓受到的光照情况在改变.一般来说, 苔藓喜好相对遮阴的环境, 在这种环境下苔藓植物较维管植物有更大的生长优势(
白学良等, 1998 ;
刘俊华等, 2005 ).在微生境不断变化的情况下, 本研究案例中苔藓植物物种多样性、组合数量和苔藓总盖度在腐解中后期达到最大, 这与其他不同气候下、不同区域的研究趋势一致, 是倒木上苔藓植物演替的一般规律(
Cole et al ., 2008 ;
Botting & DeLong, 2009 ;
游惠明, 2013 ;
Kumar et al ., 2018 ). ...
Patterns of epixylic vegetation on spruce logs in late-successional boreal forests
1
2007
... 倒木主要存在于地表并与土壤层联系紧密, 具有相对稳定的环境, 是森林中苔藓植物生长的重要基质(
Preik?a et al ., 2015 ); 它也是一个不断发展的基质, 木材腐烂程度、树皮覆盖度、离地面高度以及一些小气候条件随时间连续变化(
闫恩荣等, 2005 ;
何东进等, 2009 ), 其上生长的苔藓植物也随着环境改变持续变化(
Kumar et al ., 2018 ).苔藓植物组合作为倒木群落内部次优势物种所构成的群落局部集合体(
李博等, 2000 ), 更是随着倒木的腐解进程发生着演替(
Kushnevskaya et al ., 2007 ).一系列的研究探讨了倒木上苔藓植物物种组成和丰度的变化与倒木自身性质以及林地环境的相互关系(
McAlister, 1997 ;
Dittrich et al ., 2014 ;
Preik?a et al ., 2015 ).但由于变量间存在复杂的共线性关系, 要找到直接作用于苔藓组合的生态因子, 并从中抽离出倒木上苔藓植物组合演替规律是困难的(
Botting & DeLong, 2009 ). ...
Presence of bark influences the succession of cryptogamic wood-inhabiting communities on conifer fallen logs
1
2018
... 这一系列变化与苔藓生长微生境的改变是密不可分的.倒木基质随着时间不断变化, 树皮的粗糙程度及覆盖度、结构的完整性、养分含量等都在改变(
何东进等, 2009 ), 这些基质的变化对苔藓的定植存在影响(
Dittrich et al ., 2014 ;
Kushnevskaya & Shorohova, 2018 ).另外, 倒木结构完整性、倒木离地面的距离以及倒木距离水源的距离都会影响倒木上的水分情况, 在本研究中4株倒木的水分情况随腐解程度加深而增加.而水分对于苔藓的定植很重要, 会显著影响苔藓植物的多样性(
范苗等, 2017 ).林窗作为森林中常见的小尺度干扰机制, 对于森林物种多样性维持有着重要的意义(
李强等, 2019 ), 树木倒下形成的林窗随着时间推移逐渐闭合, 这也导致不同腐解等级倒木上生长的苔藓受到的光照情况在改变.一般来说, 苔藓喜好相对遮阴的环境, 在这种环境下苔藓植物较维管植物有更大的生长优势(
白学良等, 1998 ;
刘俊华等, 2005 ).在微生境不断变化的情况下, 本研究案例中苔藓植物物种多样性、组合数量和苔藓总盖度在腐解中后期达到最大, 这与其他不同气候下、不同区域的研究趋势一致, 是倒木上苔藓植物演替的一般规律(
Cole et al ., 2008 ;
Botting & DeLong, 2009 ;
游惠明, 2013 ;
Kumar et al ., 2018 ). ...
1
2000
... 倒木主要存在于地表并与土壤层联系紧密, 具有相对稳定的环境, 是森林中苔藓植物生长的重要基质(
Preik?a et al ., 2015 ); 它也是一个不断发展的基质, 木材腐烂程度、树皮覆盖度、离地面高度以及一些小气候条件随时间连续变化(
闫恩荣等, 2005 ;
何东进等, 2009 ), 其上生长的苔藓植物也随着环境改变持续变化(
Kumar et al ., 2018 ).苔藓植物组合作为倒木群落内部次优势物种所构成的群落局部集合体(
李博等, 2000 ), 更是随着倒木的腐解进程发生着演替(
Kushnevskaya et al ., 2007 ).一系列的研究探讨了倒木上苔藓植物物种组成和丰度的变化与倒木自身性质以及林地环境的相互关系(
McAlister, 1997 ;
Dittrich et al ., 2014 ;
Preik?a et al ., 2015 ).但由于变量间存在复杂的共线性关系, 要找到直接作用于苔藓组合的生态因子, 并从中抽离出倒木上苔藓植物组合演替规律是困难的(
Botting & DeLong, 2009 ). ...
1
2000
... 倒木主要存在于地表并与土壤层联系紧密, 具有相对稳定的环境, 是森林中苔藓植物生长的重要基质(
Preik?a et al ., 2015 ); 它也是一个不断发展的基质, 木材腐烂程度、树皮覆盖度、离地面高度以及一些小气候条件随时间连续变化(
闫恩荣等, 2005 ;
何东进等, 2009 ), 其上生长的苔藓植物也随着环境改变持续变化(
Kumar et al ., 2018 ).苔藓植物组合作为倒木群落内部次优势物种所构成的群落局部集合体(
李博等, 2000 ), 更是随着倒木的腐解进程发生着演替(
Kushnevskaya et al ., 2007 ).一系列的研究探讨了倒木上苔藓植物物种组成和丰度的变化与倒木自身性质以及林地环境的相互关系(
McAlister, 1997 ;
Dittrich et al ., 2014 ;
Preik?a et al ., 2015 ).但由于变量间存在复杂的共线性关系, 要找到直接作用于苔藓组合的生态因子, 并从中抽离出倒木上苔藓植物组合演替规律是困难的(
Botting & DeLong, 2009 ). ...
中国青藓科植物分类学修订(Ⅱ): 燕尾藓属
1
2016
... 将苔藓植物标本带回实验室阴干、装袋, 登记采集信息, 参考《西藏苔藓植物志》(
中国科学院青藏高原综合科学考察队, 1985 )、《中国苔藓志》(
高谦, 1994 ,
1996 ,
2003 ;
吴鹏程, 2002 ;
吴鹏程和贾渝, 2004 ,
2011 ;
胡人亮和王幼芳, 2005 ;
黎兴江, 2006 ;
高谦和吴玉环, 2008 )、《中国生物物种名录: 1. 植物: 苔藓植物》(
贾渝和何思, 2013 )以及近年来相关物种的研究资料(
李敏等, 2016 )对标本进行解剖鉴定, 参考美国密苏里植物园Tropicos网站(http://www.tropicos. org/)对苔藓物种名录进行整理归纳, 并根据当时所拍摄照片, 确定每个样方中每种苔藓的分盖度.本论文所采用的标本存放于中国农业大学标本室(BAU). ...
中国青藓科植物分类学修订(Ⅱ): 燕尾藓属
1
2016
... 将苔藓植物标本带回实验室阴干、装袋, 登记采集信息, 参考《西藏苔藓植物志》(
中国科学院青藏高原综合科学考察队, 1985 )、《中国苔藓志》(
高谦, 1994 ,
1996 ,
2003 ;
吴鹏程, 2002 ;
吴鹏程和贾渝, 2004 ,
2011 ;
胡人亮和王幼芳, 2005 ;
黎兴江, 2006 ;
高谦和吴玉环, 2008 )、《中国生物物种名录: 1. 植物: 苔藓植物》(
贾渝和何思, 2013 )以及近年来相关物种的研究资料(
李敏等, 2016 )对标本进行解剖鉴定, 参考美国密苏里植物园Tropicos网站(http://www.tropicos. org/)对苔藓物种名录进行整理归纳, 并根据当时所拍摄照片, 确定每个样方中每种苔藓的分盖度.本论文所采用的标本存放于中国农业大学标本室(BAU). ...
西双版纳热带雨林林窗空间分布格局及其特征指数与林窗下植物多样性的相关性
1
2019
... 这一系列变化与苔藓生长微生境的改变是密不可分的.倒木基质随着时间不断变化, 树皮的粗糙程度及覆盖度、结构的完整性、养分含量等都在改变(
何东进等, 2009 ), 这些基质的变化对苔藓的定植存在影响(
Dittrich et al ., 2014 ;
Kushnevskaya & Shorohova, 2018 ).另外, 倒木结构完整性、倒木离地面的距离以及倒木距离水源的距离都会影响倒木上的水分情况, 在本研究中4株倒木的水分情况随腐解程度加深而增加.而水分对于苔藓的定植很重要, 会显著影响苔藓植物的多样性(
范苗等, 2017 ).林窗作为森林中常见的小尺度干扰机制, 对于森林物种多样性维持有着重要的意义(
李强等, 2019 ), 树木倒下形成的林窗随着时间推移逐渐闭合, 这也导致不同腐解等级倒木上生长的苔藓受到的光照情况在改变.一般来说, 苔藓喜好相对遮阴的环境, 在这种环境下苔藓植物较维管植物有更大的生长优势(
白学良等, 1998 ;
刘俊华等, 2005 ).在微生境不断变化的情况下, 本研究案例中苔藓植物物种多样性、组合数量和苔藓总盖度在腐解中后期达到最大, 这与其他不同气候下、不同区域的研究趋势一致, 是倒木上苔藓植物演替的一般规律(
Cole et al ., 2008 ;
Botting & DeLong, 2009 ;
游惠明, 2013 ;
Kumar et al ., 2018 ). ...
西双版纳热带雨林林窗空间分布格局及其特征指数与林窗下植物多样性的相关性
1
2019
... 这一系列变化与苔藓生长微生境的改变是密不可分的.倒木基质随着时间不断变化, 树皮的粗糙程度及覆盖度、结构的完整性、养分含量等都在改变(
何东进等, 2009 ), 这些基质的变化对苔藓的定植存在影响(
Dittrich et al ., 2014 ;
Kushnevskaya & Shorohova, 2018 ).另外, 倒木结构完整性、倒木离地面的距离以及倒木距离水源的距离都会影响倒木上的水分情况, 在本研究中4株倒木的水分情况随腐解程度加深而增加.而水分对于苔藓的定植很重要, 会显著影响苔藓植物的多样性(
范苗等, 2017 ).林窗作为森林中常见的小尺度干扰机制, 对于森林物种多样性维持有着重要的意义(
李强等, 2019 ), 树木倒下形成的林窗随着时间推移逐渐闭合, 这也导致不同腐解等级倒木上生长的苔藓受到的光照情况在改变.一般来说, 苔藓喜好相对遮阴的环境, 在这种环境下苔藓植物较维管植物有更大的生长优势(
白学良等, 1998 ;
刘俊华等, 2005 ).在微生境不断变化的情况下, 本研究案例中苔藓植物物种多样性、组合数量和苔藓总盖度在腐解中后期达到最大, 这与其他不同气候下、不同区域的研究趋势一致, 是倒木上苔藓植物演替的一般规律(
Cole et al ., 2008 ;
Botting & DeLong, 2009 ;
游惠明, 2013 ;
Kumar et al ., 2018 ). ...
贵州猴耳天坑地下森林苔藓植物多样性特征研究
1
2018
... 苔藓植物物种组成在演替中变化明显.该变化取决于苔藓物种自身生物学和生态学属性, 并与环境密切相关.不同的苔藓物种有不同的定植方式、定植速率以及环境偏好.生长缓慢的苔类和顶蒴藓类可以在开放的倒木表面及完整的苔藓层的缝隙中定植, 而快速生长的侧蒴藓类会在短时间内构建形成大型斑块; 苔藓物种根据其对不同腐解等级倒木的定植偏好可以分为: 树表附生种、腐木泛生种、优先腐木生种和严格腐木生种(
ódor & van Hees, 2004 ).在本研究中, 毛帽木灵藓、中华白齿藓和新丝藓属于典型的树表附生种, 它们只在活体树木树干或倒伏不久、腐解等级较低的倒木的树皮上着生; 严格腐木生种如南亚合睫藓、全缘匐灯藓、毛羽藓、毛尖羽藓、褶叶青藓, 它们在腐解等级较低的倒木上不出现, 而在腐解等级较高的倒木上出现频率高, 甚至表现为优势类群的种类.但就4株倒木上苔藓的着生情况讨论物种定植偏好只是初步的, 是否具有普遍性还需要有更多的研究样本来验证.除了倒木腐解等级外, 光照和水分对苔藓植物的分布也存在较大影响(
李小芳等, 2018 ), 不同生活型的苔藓植物对光照的偏好在本研究中非常明显.倒木4由于新形成的林窗, 遮阴程度与倒木2类似, 它们在苔藓着生情况上表现为共有多种直立高藓丛状生长的植物, 直立高藓丛状有利于苔藓吸收水分而减少水分缺失对生长造成的影响, 因此能在光照相对较充足的条件下更好地生长(
M?gdefrau, 1982 ), 但是由于基质、水分含量及其他环境因子的影响, 可以看到两株倒木上占优势的直立高藓丛状苔藓种类还是存在区别的, 倒木2上较具优势的物种为合睫藓, 倒木4上较具优势物种为青毛藓. ...
贵州猴耳天坑地下森林苔藓植物多样性特征研究
1
2018
... 苔藓植物物种组成在演替中变化明显.该变化取决于苔藓物种自身生物学和生态学属性, 并与环境密切相关.不同的苔藓物种有不同的定植方式、定植速率以及环境偏好.生长缓慢的苔类和顶蒴藓类可以在开放的倒木表面及完整的苔藓层的缝隙中定植, 而快速生长的侧蒴藓类会在短时间内构建形成大型斑块; 苔藓物种根据其对不同腐解等级倒木的定植偏好可以分为: 树表附生种、腐木泛生种、优先腐木生种和严格腐木生种(
ódor & van Hees, 2004 ).在本研究中, 毛帽木灵藓、中华白齿藓和新丝藓属于典型的树表附生种, 它们只在活体树木树干或倒伏不久、腐解等级较低的倒木的树皮上着生; 严格腐木生种如南亚合睫藓、全缘匐灯藓、毛羽藓、毛尖羽藓、褶叶青藓, 它们在腐解等级较低的倒木上不出现, 而在腐解等级较高的倒木上出现频率高, 甚至表现为优势类群的种类.但就4株倒木上苔藓的着生情况讨论物种定植偏好只是初步的, 是否具有普遍性还需要有更多的研究样本来验证.除了倒木腐解等级外, 光照和水分对苔藓植物的分布也存在较大影响(
李小芳等, 2018 ), 不同生活型的苔藓植物对光照的偏好在本研究中非常明显.倒木4由于新形成的林窗, 遮阴程度与倒木2类似, 它们在苔藓着生情况上表现为共有多种直立高藓丛状生长的植物, 直立高藓丛状有利于苔藓吸收水分而减少水分缺失对生长造成的影响, 因此能在光照相对较充足的条件下更好地生长(
M?gdefrau, 1982 ), 但是由于基质、水分含量及其他环境因子的影响, 可以看到两株倒木上占优势的直立高藓丛状苔藓种类还是存在区别的, 倒木2上较具优势的物种为合睫藓, 倒木4上较具优势物种为青毛藓. ...
1
2006
... 将苔藓植物标本带回实验室阴干、装袋, 登记采集信息, 参考《西藏苔藓植物志》(
中国科学院青藏高原综合科学考察队, 1985 )、《中国苔藓志》(
高谦, 1994 ,
1996 ,
2003 ;
吴鹏程, 2002 ;
吴鹏程和贾渝, 2004 ,
2011 ;
胡人亮和王幼芳, 2005 ;
黎兴江, 2006 ;
高谦和吴玉环, 2008 )、《中国生物物种名录: 1. 植物: 苔藓植物》(
贾渝和何思, 2013 )以及近年来相关物种的研究资料(
李敏等, 2016 )对标本进行解剖鉴定, 参考美国密苏里植物园Tropicos网站(http://www.tropicos. org/)对苔藓物种名录进行整理归纳, 并根据当时所拍摄照片, 确定每个样方中每种苔藓的分盖度.本论文所采用的标本存放于中国农业大学标本室(BAU). ...
1
2006
... 将苔藓植物标本带回实验室阴干、装袋, 登记采集信息, 参考《西藏苔藓植物志》(
中国科学院青藏高原综合科学考察队, 1985 )、《中国苔藓志》(
高谦, 1994 ,
1996 ,
2003 ;
吴鹏程, 2002 ;
吴鹏程和贾渝, 2004 ,
2011 ;
胡人亮和王幼芳, 2005 ;
黎兴江, 2006 ;
高谦和吴玉环, 2008 )、《中国生物物种名录: 1. 植物: 苔藓植物》(
贾渝和何思, 2013 )以及近年来相关物种的研究资料(
李敏等, 2016 )对标本进行解剖鉴定, 参考美国密苏里植物园Tropicos网站(http://www.tropicos. org/)对苔藓物种名录进行整理归纳, 并根据当时所拍摄照片, 确定每个样方中每种苔藓的分盖度.本论文所采用的标本存放于中国农业大学标本室(BAU). ...
青藏高原东部原始林下地表主要苔藓斑块特征及其影响因素
1
2005
... 这一系列变化与苔藓生长微生境的改变是密不可分的.倒木基质随着时间不断变化, 树皮的粗糙程度及覆盖度、结构的完整性、养分含量等都在改变(
何东进等, 2009 ), 这些基质的变化对苔藓的定植存在影响(
Dittrich et al ., 2014 ;
Kushnevskaya & Shorohova, 2018 ).另外, 倒木结构完整性、倒木离地面的距离以及倒木距离水源的距离都会影响倒木上的水分情况, 在本研究中4株倒木的水分情况随腐解程度加深而增加.而水分对于苔藓的定植很重要, 会显著影响苔藓植物的多样性(
范苗等, 2017 ).林窗作为森林中常见的小尺度干扰机制, 对于森林物种多样性维持有着重要的意义(
李强等, 2019 ), 树木倒下形成的林窗随着时间推移逐渐闭合, 这也导致不同腐解等级倒木上生长的苔藓受到的光照情况在改变.一般来说, 苔藓喜好相对遮阴的环境, 在这种环境下苔藓植物较维管植物有更大的生长优势(
白学良等, 1998 ;
刘俊华等, 2005 ).在微生境不断变化的情况下, 本研究案例中苔藓植物物种多样性、组合数量和苔藓总盖度在腐解中后期达到最大, 这与其他不同气候下、不同区域的研究趋势一致, 是倒木上苔藓植物演替的一般规律(
Cole et al ., 2008 ;
Botting & DeLong, 2009 ;
游惠明, 2013 ;
Kumar et al ., 2018 ). ...
青藏高原东部原始林下地表主要苔藓斑块特征及其影响因素
1
2005
... 这一系列变化与苔藓生长微生境的改变是密不可分的.倒木基质随着时间不断变化, 树皮的粗糙程度及覆盖度、结构的完整性、养分含量等都在改变(
何东进等, 2009 ), 这些基质的变化对苔藓的定植存在影响(
Dittrich et al ., 2014 ;
Kushnevskaya & Shorohova, 2018 ).另外, 倒木结构完整性、倒木离地面的距离以及倒木距离水源的距离都会影响倒木上的水分情况, 在本研究中4株倒木的水分情况随腐解程度加深而增加.而水分对于苔藓的定植很重要, 会显著影响苔藓植物的多样性(
范苗等, 2017 ).林窗作为森林中常见的小尺度干扰机制, 对于森林物种多样性维持有着重要的意义(
李强等, 2019 ), 树木倒下形成的林窗随着时间推移逐渐闭合, 这也导致不同腐解等级倒木上生长的苔藓受到的光照情况在改变.一般来说, 苔藓喜好相对遮阴的环境, 在这种环境下苔藓植物较维管植物有更大的生长优势(
白学良等, 1998 ;
刘俊华等, 2005 ).在微生境不断变化的情况下, 本研究案例中苔藓植物物种多样性、组合数量和苔藓总盖度在腐解中后期达到最大, 这与其他不同气候下、不同区域的研究趋势一致, 是倒木上苔藓植物演替的一般规律(
Cole et al ., 2008 ;
Botting & DeLong, 2009 ;
游惠明, 2013 ;
Kumar et al ., 2018 ). ...
不同生长基质的苔藓植物优势种生态位与种间联结
1
2019
... 植物之间的相互作用也在影响着演替的过程.随着时间的推移, 倒木木质素等难分解组分降解程度增加, 结构相对疏松, 树干湿度增加, 倒木上环境逐渐优化, 蕨类植物、种子植物开始在倒木上及倒木周边定植, 一定程度上挤压了苔藓的生存空间, 加之枯落物的不断累积, 占据了部分可定植的空间, 对苔藓植物的生长造成了负面的影响(
Natalia et al ., 2008 ;
Márialigeti et al ., 2009 ), 使得苔藓植物盖度在到达极大值后逐渐减小.另外苔藓植物之间也存在合作、竞争等关系(
卜兆君等, 2009 ).倒木作为一个相对变化快速的生境, 其上生长的苔藓物种和组合通常是不稳定的, 在这种情况下种间关系常趋于负相关, 即种间竞争或排他作用强烈(
刘艳等, 2019 ), 例如青毛藓、丛叶青毛藓和长叶青毛藓都分布于光照较为充足的倒木2和倒木4, 但是同一个样方内通常仅有一种青毛藓属植物.苔藓植物的竞争等级还会随着环境的变化而改变(
Breeuwer et al ., 2008 ), 从而推动不同演替阶段苔藓物种以及物种组合的变化.早期优势的丛生型藓类能较好地适应相对干旱、强光照的环境, 但随着环境向潮湿荫蔽转变, 个体更大且生长快速的垫状毯状藓类出现, 它们能获取更多的资源, 不对称竞争导致竞争等级的改变.随着更多苔藓物种的定植, 倒木上苔藓间竞争压力逐渐增大, 这种情况下物种间生态位的分化是物种共存的重要机理之一(
Rydin, 1986 ), 本研究中苔藓植物组合内生活型种类和数量即整体空间结构复杂程度随着腐解等级及其他生境条件的变化而逐渐增大, 呈现出类似于植物演替过程中的生态位分化. ...
不同生长基质的苔藓植物优势种生态位与种间联结
1
2019
... 植物之间的相互作用也在影响着演替的过程.随着时间的推移, 倒木木质素等难分解组分降解程度增加, 结构相对疏松, 树干湿度增加, 倒木上环境逐渐优化, 蕨类植物、种子植物开始在倒木上及倒木周边定植, 一定程度上挤压了苔藓的生存空间, 加之枯落物的不断累积, 占据了部分可定植的空间, 对苔藓植物的生长造成了负面的影响(
Natalia et al ., 2008 ;
Márialigeti et al ., 2009 ), 使得苔藓植物盖度在到达极大值后逐渐减小.另外苔藓植物之间也存在合作、竞争等关系(
卜兆君等, 2009 ).倒木作为一个相对变化快速的生境, 其上生长的苔藓物种和组合通常是不稳定的, 在这种情况下种间关系常趋于负相关, 即种间竞争或排他作用强烈(
刘艳等, 2019 ), 例如青毛藓、丛叶青毛藓和长叶青毛藓都分布于光照较为充足的倒木2和倒木4, 但是同一个样方内通常仅有一种青毛藓属植物.苔藓植物的竞争等级还会随着环境的变化而改变(
Breeuwer et al ., 2008 ), 从而推动不同演替阶段苔藓物种以及物种组合的变化.早期优势的丛生型藓类能较好地适应相对干旱、强光照的环境, 但随着环境向潮湿荫蔽转变, 个体更大且生长快速的垫状毯状藓类出现, 它们能获取更多的资源, 不对称竞争导致竞争等级的改变.随着更多苔藓物种的定植, 倒木上苔藓间竞争压力逐渐增大, 这种情况下物种间生态位的分化是物种共存的重要机理之一(
Rydin, 1986 ), 本研究中苔藓植物组合内生活型种类和数量即整体空间结构复杂程度随着腐解等级及其他生境条件的变化而逐渐增大, 呈现出类似于植物演替过程中的生态位分化. ...
西藏色季拉山急尖长苞冷杉林粗木质残体储量与倒木分解研究
2
2016
... 研究地点位于藏东南色季拉山鲁朗兵站往通麦方向约8 km处(29.87° N, 94.77° E), 海拔2 867 m.本区域属亚高山寒温带湿润气候区, 年平均气温-0.73 ℃, 最暖月(7月)平均气温9.8 ℃, 年降水量约1 134.1 mm, 年蒸发量544.0 mm, 年平均相对湿度78% (
马豪霞等, 2016 ); 研究地点植被是以华山松为优势树种的森林, 间或分布急尖长苞冷杉(
Abies georgei var.
smithii ); 灌木层有高山栎(
Quercus semecarpifolia )、西藏箭竹(
Fargesia macclureana )、西南花楸(
Sorbus rehderiana )等; 草本层有鬼灯檠(
Rodgersia podophylla )、小檗属(
Berberis )、悬钩子属(
Rubus )、野草莓(
Fragaria vesca )、高山露珠草(
Circaea alpina )、薹草属(
Carex )、鳞毛蕨属(
Dryopteris )等.坡向朝南, 坡度约为23°. ...
... 依据所记录倒木的腐解程度数据, 参照
闫恩荣等(2005) 和
马豪霞等(2016) 对森林粗木质残体腐解等级的划分及判定标准, 确定每株倒木的腐解等级, 并根据腐解等级对倒木进行编号. ...
西藏色季拉山急尖长苞冷杉林粗木质残体储量与倒木分解研究
2
2016
... 研究地点位于藏东南色季拉山鲁朗兵站往通麦方向约8 km处(29.87° N, 94.77° E), 海拔2 867 m.本区域属亚高山寒温带湿润气候区, 年平均气温-0.73 ℃, 最暖月(7月)平均气温9.8 ℃, 年降水量约1 134.1 mm, 年蒸发量544.0 mm, 年平均相对湿度78% (
马豪霞等, 2016 ); 研究地点植被是以华山松为优势树种的森林, 间或分布急尖长苞冷杉(
Abies georgei var.
smithii ); 灌木层有高山栎(
Quercus semecarpifolia )、西藏箭竹(
Fargesia macclureana )、西南花楸(
Sorbus rehderiana )等; 草本层有鬼灯檠(
Rodgersia podophylla )、小檗属(
Berberis )、悬钩子属(
Rubus )、野草莓(
Fragaria vesca )、高山露珠草(
Circaea alpina )、薹草属(
Carex )、鳞毛蕨属(
Dryopteris )等.坡向朝南, 坡度约为23°. ...
... 依据所记录倒木的腐解程度数据, 参照
闫恩荣等(2005) 和
马豪霞等(2016) 对森林粗木质残体腐解等级的划分及判定标准, 确定每株倒木的腐解等级, 并根据腐解等级对倒木进行编号. ...
Life-forms of bryophytes//Smith AJE. Bryophyte Ecology
1
1982
... 苔藓植物物种组成在演替中变化明显.该变化取决于苔藓物种自身生物学和生态学属性, 并与环境密切相关.不同的苔藓物种有不同的定植方式、定植速率以及环境偏好.生长缓慢的苔类和顶蒴藓类可以在开放的倒木表面及完整的苔藓层的缝隙中定植, 而快速生长的侧蒴藓类会在短时间内构建形成大型斑块; 苔藓物种根据其对不同腐解等级倒木的定植偏好可以分为: 树表附生种、腐木泛生种、优先腐木生种和严格腐木生种(
ódor & van Hees, 2004 ).在本研究中, 毛帽木灵藓、中华白齿藓和新丝藓属于典型的树表附生种, 它们只在活体树木树干或倒伏不久、腐解等级较低的倒木的树皮上着生; 严格腐木生种如南亚合睫藓、全缘匐灯藓、毛羽藓、毛尖羽藓、褶叶青藓, 它们在腐解等级较低的倒木上不出现, 而在腐解等级较高的倒木上出现频率高, 甚至表现为优势类群的种类.但就4株倒木上苔藓的着生情况讨论物种定植偏好只是初步的, 是否具有普遍性还需要有更多的研究样本来验证.除了倒木腐解等级外, 光照和水分对苔藓植物的分布也存在较大影响(
李小芳等, 2018 ), 不同生活型的苔藓植物对光照的偏好在本研究中非常明显.倒木4由于新形成的林窗, 遮阴程度与倒木2类似, 它们在苔藓着生情况上表现为共有多种直立高藓丛状生长的植物, 直立高藓丛状有利于苔藓吸收水分而减少水分缺失对生长造成的影响, 因此能在光照相对较充足的条件下更好地生长(
M?gdefrau, 1982 ), 但是由于基质、水分含量及其他环境因子的影响, 可以看到两株倒木上占优势的直立高藓丛状苔藓种类还是存在区别的, 倒木2上较具优势的物种为合睫藓, 倒木4上较具优势物种为青毛藓. ...
The effects of stand structure on ground-floor bryophyte assemblages in temperate mixed forests
1
2009
... 植物之间的相互作用也在影响着演替的过程.随着时间的推移, 倒木木质素等难分解组分降解程度增加, 结构相对疏松, 树干湿度增加, 倒木上环境逐渐优化, 蕨类植物、种子植物开始在倒木上及倒木周边定植, 一定程度上挤压了苔藓的生存空间, 加之枯落物的不断累积, 占据了部分可定植的空间, 对苔藓植物的生长造成了负面的影响(
Natalia et al ., 2008 ;
Márialigeti et al ., 2009 ), 使得苔藓植物盖度在到达极大值后逐渐减小.另外苔藓植物之间也存在合作、竞争等关系(
卜兆君等, 2009 ).倒木作为一个相对变化快速的生境, 其上生长的苔藓物种和组合通常是不稳定的, 在这种情况下种间关系常趋于负相关, 即种间竞争或排他作用强烈(
刘艳等, 2019 ), 例如青毛藓、丛叶青毛藓和长叶青毛藓都分布于光照较为充足的倒木2和倒木4, 但是同一个样方内通常仅有一种青毛藓属植物.苔藓植物的竞争等级还会随着环境的变化而改变(
Breeuwer et al ., 2008 ), 从而推动不同演替阶段苔藓物种以及物种组合的变化.早期优势的丛生型藓类能较好地适应相对干旱、强光照的环境, 但随着环境向潮湿荫蔽转变, 个体更大且生长快速的垫状毯状藓类出现, 它们能获取更多的资源, 不对称竞争导致竞争等级的改变.随着更多苔藓物种的定植, 倒木上苔藓间竞争压力逐渐增大, 这种情况下物种间生态位的分化是物种共存的重要机理之一(
Rydin, 1986 ), 本研究中苔藓植物组合内生活型种类和数量即整体空间结构复杂程度随着腐解等级及其他生境条件的变化而逐渐增大, 呈现出类似于植物演替过程中的生态位分化. ...
Cryptogam communities on fallen logs in the Duke Forest, North Carolina
1
1997
... 倒木主要存在于地表并与土壤层联系紧密, 具有相对稳定的环境, 是森林中苔藓植物生长的重要基质(
Preik?a et al ., 2015 ); 它也是一个不断发展的基质, 木材腐烂程度、树皮覆盖度、离地面高度以及一些小气候条件随时间连续变化(
闫恩荣等, 2005 ;
何东进等, 2009 ), 其上生长的苔藓植物也随着环境改变持续变化(
Kumar et al ., 2018 ).苔藓植物组合作为倒木群落内部次优势物种所构成的群落局部集合体(
李博等, 2000 ), 更是随着倒木的腐解进程发生着演替(
Kushnevskaya et al ., 2007 ).一系列的研究探讨了倒木上苔藓植物物种组成和丰度的变化与倒木自身性质以及林地环境的相互关系(
McAlister, 1997 ;
Dittrich et al ., 2014 ;
Preik?a et al ., 2015 ).但由于变量间存在复杂的共线性关系, 要找到直接作用于苔藓组合的生态因子, 并从中抽离出倒木上苔藓植物组合演替规律是困难的(
Botting & DeLong, 2009 ). ...
Plant succession on fallen logs in a virgin spruce-fir forest
1948
Effects of leaf litter on the growth of boreal feather mosses: implication for forest ?oor development
1
2008
... 植物之间的相互作用也在影响着演替的过程.随着时间的推移, 倒木木质素等难分解组分降解程度增加, 结构相对疏松, 树干湿度增加, 倒木上环境逐渐优化, 蕨类植物、种子植物开始在倒木上及倒木周边定植, 一定程度上挤压了苔藓的生存空间, 加之枯落物的不断累积, 占据了部分可定植的空间, 对苔藓植物的生长造成了负面的影响(
Natalia et al ., 2008 ;
Márialigeti et al ., 2009 ), 使得苔藓植物盖度在到达极大值后逐渐减小.另外苔藓植物之间也存在合作、竞争等关系(
卜兆君等, 2009 ).倒木作为一个相对变化快速的生境, 其上生长的苔藓物种和组合通常是不稳定的, 在这种情况下种间关系常趋于负相关, 即种间竞争或排他作用强烈(
刘艳等, 2019 ), 例如青毛藓、丛叶青毛藓和长叶青毛藓都分布于光照较为充足的倒木2和倒木4, 但是同一个样方内通常仅有一种青毛藓属植物.苔藓植物的竞争等级还会随着环境的变化而改变(
Breeuwer et al ., 2008 ), 从而推动不同演替阶段苔藓物种以及物种组合的变化.早期优势的丛生型藓类能较好地适应相对干旱、强光照的环境, 但随着环境向潮湿荫蔽转变, 个体更大且生长快速的垫状毯状藓类出现, 它们能获取更多的资源, 不对称竞争导致竞争等级的改变.随着更多苔藓物种的定植, 倒木上苔藓间竞争压力逐渐增大, 这种情况下物种间生态位的分化是物种共存的重要机理之一(
Rydin, 1986 ), 本研究中苔藓植物组合内生活型种类和数量即整体空间结构复杂程度随着腐解等级及其他生境条件的变化而逐渐增大, 呈现出类似于植物演替过程中的生态位分化. ...
Preferences of dead wood inhabiting bryophytes for decay stage, log size and habitat types in Hungarian beech forests
1
2004
... 苔藓植物物种组成在演替中变化明显.该变化取决于苔藓物种自身生物学和生态学属性, 并与环境密切相关.不同的苔藓物种有不同的定植方式、定植速率以及环境偏好.生长缓慢的苔类和顶蒴藓类可以在开放的倒木表面及完整的苔藓层的缝隙中定植, 而快速生长的侧蒴藓类会在短时间内构建形成大型斑块; 苔藓物种根据其对不同腐解等级倒木的定植偏好可以分为: 树表附生种、腐木泛生种、优先腐木生种和严格腐木生种(
ódor & van Hees, 2004 ).在本研究中, 毛帽木灵藓、中华白齿藓和新丝藓属于典型的树表附生种, 它们只在活体树木树干或倒伏不久、腐解等级较低的倒木的树皮上着生; 严格腐木生种如南亚合睫藓、全缘匐灯藓、毛羽藓、毛尖羽藓、褶叶青藓, 它们在腐解等级较低的倒木上不出现, 而在腐解等级较高的倒木上出现频率高, 甚至表现为优势类群的种类.但就4株倒木上苔藓的着生情况讨论物种定植偏好只是初步的, 是否具有普遍性还需要有更多的研究样本来验证.除了倒木腐解等级外, 光照和水分对苔藓植物的分布也存在较大影响(
李小芳等, 2018 ), 不同生活型的苔藓植物对光照的偏好在本研究中非常明显.倒木4由于新形成的林窗, 遮阴程度与倒木2类似, 它们在苔藓着生情况上表现为共有多种直立高藓丛状生长的植物, 直立高藓丛状有利于苔藓吸收水分而减少水分缺失对生长造成的影响, 因此能在光照相对较充足的条件下更好地生长(
M?gdefrau, 1982 ), 但是由于基质、水分含量及其他环境因子的影响, 可以看到两株倒木上占优势的直立高藓丛状苔藓种类还是存在区别的, 倒木2上较具优势的物种为合睫藓, 倒木4上较具优势物种为青毛藓. ...
Dead wood quality influences species diversity of rare cryptogams in temperate broadleaved forests
2
2015
... 倒木主要存在于地表并与土壤层联系紧密, 具有相对稳定的环境, 是森林中苔藓植物生长的重要基质(
Preik?a et al ., 2015 ); 它也是一个不断发展的基质, 木材腐烂程度、树皮覆盖度、离地面高度以及一些小气候条件随时间连续变化(
闫恩荣等, 2005 ;
何东进等, 2009 ), 其上生长的苔藓植物也随着环境改变持续变化(
Kumar et al ., 2018 ).苔藓植物组合作为倒木群落内部次优势物种所构成的群落局部集合体(
李博等, 2000 ), 更是随着倒木的腐解进程发生着演替(
Kushnevskaya et al ., 2007 ).一系列的研究探讨了倒木上苔藓植物物种组成和丰度的变化与倒木自身性质以及林地环境的相互关系(
McAlister, 1997 ;
Dittrich et al ., 2014 ;
Preik?a et al ., 2015 ).但由于变量间存在复杂的共线性关系, 要找到直接作用于苔藓组合的生态因子, 并从中抽离出倒木上苔藓植物组合演替规律是困难的(
Botting & DeLong, 2009 ). ...
... ;
Preik?a et al ., 2015 ).但由于变量间存在复杂的共线性关系, 要找到直接作用于苔藓组合的生态因子, 并从中抽离出倒木上苔藓植物组合演替规律是困难的(
Botting & DeLong, 2009 ). ...
Competition and niche separation in Sphagnum
1
1986
... 植物之间的相互作用也在影响着演替的过程.随着时间的推移, 倒木木质素等难分解组分降解程度增加, 结构相对疏松, 树干湿度增加, 倒木上环境逐渐优化, 蕨类植物、种子植物开始在倒木上及倒木周边定植, 一定程度上挤压了苔藓的生存空间, 加之枯落物的不断累积, 占据了部分可定植的空间, 对苔藓植物的生长造成了负面的影响(
Natalia et al ., 2008 ;
Márialigeti et al ., 2009 ), 使得苔藓植物盖度在到达极大值后逐渐减小.另外苔藓植物之间也存在合作、竞争等关系(
卜兆君等, 2009 ).倒木作为一个相对变化快速的生境, 其上生长的苔藓物种和组合通常是不稳定的, 在这种情况下种间关系常趋于负相关, 即种间竞争或排他作用强烈(
刘艳等, 2019 ), 例如青毛藓、丛叶青毛藓和长叶青毛藓都分布于光照较为充足的倒木2和倒木4, 但是同一个样方内通常仅有一种青毛藓属植物.苔藓植物的竞争等级还会随着环境的变化而改变(
Breeuwer et al ., 2008 ), 从而推动不同演替阶段苔藓物种以及物种组合的变化.早期优势的丛生型藓类能较好地适应相对干旱、强光照的环境, 但随着环境向潮湿荫蔽转变, 个体更大且生长快速的垫状毯状藓类出现, 它们能获取更多的资源, 不对称竞争导致竞争等级的改变.随着更多苔藓物种的定植, 倒木上苔藓间竞争压力逐渐增大, 这种情况下物种间生态位的分化是物种共存的重要机理之一(
Rydin, 1986 ), 本研究中苔藓植物组合内生活型种类和数量即整体空间结构复杂程度随着腐解等级及其他生境条件的变化而逐渐增大, 呈现出类似于植物演替过程中的生态位分化. ...
Biomass conversion factors (density and carbon concentration) by decay classes for dead wood of Pinus sylvestris, Picea abies and Betula spp. in boreal forests of Sweden
1
2007
... 倒木属于粗死木质残体(coarse woody debris, CWD)的一种(
闫恩荣等, 2005 ).在传统的森林管理过程中, 倒木常被误认为容易引起病虫害和火灾而被清除.近年来, 许多****对森林生态系统的深入研究结果表明, 倒木是森林生态系统不可缺少的重要组成部分, 在减少水土流失、提高林地土壤肥力和加速养分循环等方面发挥着重要作用(
Sandstr?m et al ., 2007 ;
Vanha-Majamaa et al ., 2007 ;
Davis et al ., 2010 ).倒木作为森林生态系统中的持续营养源, 是种类繁多的生物赖以生存的蔽护所和繁殖场地, 也是林木更新的良好苗床, 对森林生态系统的完整性和生物多样性的维持具有重要意义(
Takahashi et al ., 2000 ;
何东进等, 2009 ). ...
Bole bryophyte diversity and distribution patterns along three altitudinal gradients in Yunnan, China
1
2015
... 对不同组合内各苔藓植物重要值进行计算(
Song et al ., 2015 ;
范苗等, 2017 ). ...
Establishment of tree seedlings and water-soluble nutrients in coarse woody debris in an old-growth Picea-Abies forest in Hokkaido, northern Japan
1
2000
... 倒木属于粗死木质残体(coarse woody debris, CWD)的一种(
闫恩荣等, 2005 ).在传统的森林管理过程中, 倒木常被误认为容易引起病虫害和火灾而被清除.近年来, 许多****对森林生态系统的深入研究结果表明, 倒木是森林生态系统不可缺少的重要组成部分, 在减少水土流失、提高林地土壤肥力和加速养分循环等方面发挥着重要作用(
Sandstr?m et al ., 2007 ;
Vanha-Majamaa et al ., 2007 ;
Davis et al ., 2010 ).倒木作为森林生态系统中的持续营养源, 是种类繁多的生物赖以生存的蔽护所和繁殖场地, 也是林木更新的良好苗床, 对森林生态系统的完整性和生物多样性的维持具有重要意义(
Takahashi et al ., 2000 ;
何东进等, 2009 ). ...
1
1985
... 将苔藓植物标本带回实验室阴干、装袋, 登记采集信息, 参考《西藏苔藓植物志》(
中国科学院青藏高原综合科学考察队, 1985 )、《中国苔藓志》(
高谦, 1994 ,
1996 ,
2003 ;
吴鹏程, 2002 ;
吴鹏程和贾渝, 2004 ,
2011 ;
胡人亮和王幼芳, 2005 ;
黎兴江, 2006 ;
高谦和吴玉环, 2008 )、《中国生物物种名录: 1. 植物: 苔藓植物》(
贾渝和何思, 2013 )以及近年来相关物种的研究资料(
李敏等, 2016 )对标本进行解剖鉴定, 参考美国密苏里植物园Tropicos网站(http://www.tropicos. org/)对苔藓物种名录进行整理归纳, 并根据当时所拍摄照片, 确定每个样方中每种苔藓的分盖度.本论文所采用的标本存放于中国农业大学标本室(BAU). ...
1
1985
... 将苔藓植物标本带回实验室阴干、装袋, 登记采集信息, 参考《西藏苔藓植物志》(
中国科学院青藏高原综合科学考察队, 1985 )、《中国苔藓志》(
高谦, 1994 ,
1996 ,
2003 ;
吴鹏程, 2002 ;
吴鹏程和贾渝, 2004 ,
2011 ;
胡人亮和王幼芳, 2005 ;
黎兴江, 2006 ;
高谦和吴玉环, 2008 )、《中国生物物种名录: 1. 植物: 苔藓植物》(
贾渝和何思, 2013 )以及近年来相关物种的研究资料(
李敏等, 2016 )对标本进行解剖鉴定, 参考美国密苏里植物园Tropicos网站(http://www.tropicos. org/)对苔藓物种名录进行整理归纳, 并根据当时所拍摄照片, 确定每个样方中每种苔藓的分盖度.本论文所采用的标本存放于中国农业大学标本室(BAU). ...
Rehabilitating boreal forest structure and species composition in Finland through logging, dead wood creation and fire: the EVO experiment
1
2007
... 倒木属于粗死木质残体(coarse woody debris, CWD)的一种(
闫恩荣等, 2005 ).在传统的森林管理过程中, 倒木常被误认为容易引起病虫害和火灾而被清除.近年来, 许多****对森林生态系统的深入研究结果表明, 倒木是森林生态系统不可缺少的重要组成部分, 在减少水土流失、提高林地土壤肥力和加速养分循环等方面发挥着重要作用(
Sandstr?m et al ., 2007 ;
Vanha-Majamaa et al ., 2007 ;
Davis et al ., 2010 ).倒木作为森林生态系统中的持续营养源, 是种类繁多的生物赖以生存的蔽护所和繁殖场地, 也是林木更新的良好苗床, 对森林生态系统的完整性和生物多样性的维持具有重要意义(
Takahashi et al ., 2000 ;
何东进等, 2009 ). ...
2
1998
... 将苔藓植物标本带回实验室阴干、装袋, 登记采集信息, 参考《西藏苔藓植物志》(
中国科学院青藏高原综合科学考察队, 1985 )、《中国苔藓志》(
高谦, 1994 ,
1996 ,
2003 ;
吴鹏程, 2002 ;
吴鹏程和贾渝, 2004 ,
2011 ;
胡人亮和王幼芳, 2005 ;
黎兴江, 2006 ;
高谦和吴玉环, 2008 )、《中国生物物种名录: 1. 植物: 苔藓植物》(
贾渝和何思, 2013 )以及近年来相关物种的研究资料(
李敏等, 2016 )对标本进行解剖鉴定, 参考美国密苏里植物园Tropicos网站(http://www.tropicos. org/)对苔藓物种名录进行整理归纳, 并根据当时所拍摄照片, 确定每个样方中每种苔藓的分盖度.本论文所采用的标本存放于中国农业大学标本室(BAU). ...
... 1.3.3.1 倒木苔藓生长情况 按照倒木上苔藓的生长情况将苔藓分成不同的生活型, 研究样本中共出现7种生活型: 大垫状(large cushion)、矮藓丛状(short turf)、直立高藓丛状(tall turf)、毯状(rough mat)、交织状(weft)、丝状(thread-like form)、紧密簇状(dense form)(
吴鹏程, 1998 ).统计不同倒木上着生的不同类型苔藓物种数目及盖度情况. ...
2
1998
... 将苔藓植物标本带回实验室阴干、装袋, 登记采集信息, 参考《西藏苔藓植物志》(
中国科学院青藏高原综合科学考察队, 1985 )、《中国苔藓志》(
高谦, 1994 ,
1996 ,
2003 ;
吴鹏程, 2002 ;
吴鹏程和贾渝, 2004 ,
2011 ;
胡人亮和王幼芳, 2005 ;
黎兴江, 2006 ;
高谦和吴玉环, 2008 )、《中国生物物种名录: 1. 植物: 苔藓植物》(
贾渝和何思, 2013 )以及近年来相关物种的研究资料(
李敏等, 2016 )对标本进行解剖鉴定, 参考美国密苏里植物园Tropicos网站(http://www.tropicos. org/)对苔藓物种名录进行整理归纳, 并根据当时所拍摄照片, 确定每个样方中每种苔藓的分盖度.本论文所采用的标本存放于中国农业大学标本室(BAU). ...
... 1.3.3.1 倒木苔藓生长情况 按照倒木上苔藓的生长情况将苔藓分成不同的生活型, 研究样本中共出现7种生活型: 大垫状(large cushion)、矮藓丛状(short turf)、直立高藓丛状(tall turf)、毯状(rough mat)、交织状(weft)、丝状(thread-like form)、紧密簇状(dense form)(
吴鹏程, 1998 ).统计不同倒木上着生的不同类型苔藓物种数目及盖度情况. ...
1998
1998
1
2004
... 将苔藓植物标本带回实验室阴干、装袋, 登记采集信息, 参考《西藏苔藓植物志》(
中国科学院青藏高原综合科学考察队, 1985 )、《中国苔藓志》(
高谦, 1994 ,
1996 ,
2003 ;
吴鹏程, 2002 ;
吴鹏程和贾渝, 2004 ,
2011 ;
胡人亮和王幼芳, 2005 ;
黎兴江, 2006 ;
高谦和吴玉环, 2008 )、《中国生物物种名录: 1. 植物: 苔藓植物》(
贾渝和何思, 2013 )以及近年来相关物种的研究资料(
李敏等, 2016 )对标本进行解剖鉴定, 参考美国密苏里植物园Tropicos网站(http://www.tropicos. org/)对苔藓物种名录进行整理归纳, 并根据当时所拍摄照片, 确定每个样方中每种苔藓的分盖度.本论文所采用的标本存放于中国农业大学标本室(BAU). ...
1
2004
... 将苔藓植物标本带回实验室阴干、装袋, 登记采集信息, 参考《西藏苔藓植物志》(
中国科学院青藏高原综合科学考察队, 1985 )、《中国苔藓志》(
高谦, 1994 ,
1996 ,
2003 ;
吴鹏程, 2002 ;
吴鹏程和贾渝, 2004 ,
2011 ;
胡人亮和王幼芳, 2005 ;
黎兴江, 2006 ;
高谦和吴玉环, 2008 )、《中国生物物种名录: 1. 植物: 苔藓植物》(
贾渝和何思, 2013 )以及近年来相关物种的研究资料(
李敏等, 2016 )对标本进行解剖鉴定, 参考美国密苏里植物园Tropicos网站(http://www.tropicos. org/)对苔藓物种名录进行整理归纳, 并根据当时所拍摄照片, 确定每个样方中每种苔藓的分盖度.本论文所采用的标本存放于中国农业大学标本室(BAU). ...
1
2011
... 将苔藓植物标本带回实验室阴干、装袋, 登记采集信息, 参考《西藏苔藓植物志》(
中国科学院青藏高原综合科学考察队, 1985 )、《中国苔藓志》(
高谦, 1994 ,
1996 ,
2003 ;
吴鹏程, 2002 ;
吴鹏程和贾渝, 2004 ,
2011 ;
胡人亮和王幼芳, 2005 ;
黎兴江, 2006 ;
高谦和吴玉环, 2008 )、《中国生物物种名录: 1. 植物: 苔藓植物》(
贾渝和何思, 2013 )以及近年来相关物种的研究资料(
李敏等, 2016 )对标本进行解剖鉴定, 参考美国密苏里植物园Tropicos网站(http://www.tropicos. org/)对苔藓物种名录进行整理归纳, 并根据当时所拍摄照片, 确定每个样方中每种苔藓的分盖度.本论文所采用的标本存放于中国农业大学标本室(BAU). ...
1
2011
... 将苔藓植物标本带回实验室阴干、装袋, 登记采集信息, 参考《西藏苔藓植物志》(
中国科学院青藏高原综合科学考察队, 1985 )、《中国苔藓志》(
高谦, 1994 ,
1996 ,
2003 ;
吴鹏程, 2002 ;
吴鹏程和贾渝, 2004 ,
2011 ;
胡人亮和王幼芳, 2005 ;
黎兴江, 2006 ;
高谦和吴玉环, 2008 )、《中国生物物种名录: 1. 植物: 苔藓植物》(
贾渝和何思, 2013 )以及近年来相关物种的研究资料(
李敏等, 2016 )对标本进行解剖鉴定, 参考美国密苏里植物园Tropicos网站(http://www.tropicos. org/)对苔藓物种名录进行整理归纳, 并根据当时所拍摄照片, 确定每个样方中每种苔藓的分盖度.本论文所采用的标本存放于中国农业大学标本室(BAU). ...
森林粗死木质残体的概念及其分类
3
2005
... 倒木属于粗死木质残体(coarse woody debris, CWD)的一种(
闫恩荣等, 2005 ).在传统的森林管理过程中, 倒木常被误认为容易引起病虫害和火灾而被清除.近年来, 许多****对森林生态系统的深入研究结果表明, 倒木是森林生态系统不可缺少的重要组成部分, 在减少水土流失、提高林地土壤肥力和加速养分循环等方面发挥着重要作用(
Sandstr?m et al ., 2007 ;
Vanha-Majamaa et al ., 2007 ;
Davis et al ., 2010 ).倒木作为森林生态系统中的持续营养源, 是种类繁多的生物赖以生存的蔽护所和繁殖场地, 也是林木更新的良好苗床, 对森林生态系统的完整性和生物多样性的维持具有重要意义(
Takahashi et al ., 2000 ;
何东进等, 2009 ). ...
... 倒木主要存在于地表并与土壤层联系紧密, 具有相对稳定的环境, 是森林中苔藓植物生长的重要基质(
Preik?a et al ., 2015 ); 它也是一个不断发展的基质, 木材腐烂程度、树皮覆盖度、离地面高度以及一些小气候条件随时间连续变化(
闫恩荣等, 2005 ;
何东进等, 2009 ), 其上生长的苔藓植物也随着环境改变持续变化(
Kumar et al ., 2018 ).苔藓植物组合作为倒木群落内部次优势物种所构成的群落局部集合体(
李博等, 2000 ), 更是随着倒木的腐解进程发生着演替(
Kushnevskaya et al ., 2007 ).一系列的研究探讨了倒木上苔藓植物物种组成和丰度的变化与倒木自身性质以及林地环境的相互关系(
McAlister, 1997 ;
Dittrich et al ., 2014 ;
Preik?a et al ., 2015 ).但由于变量间存在复杂的共线性关系, 要找到直接作用于苔藓组合的生态因子, 并从中抽离出倒木上苔藓植物组合演替规律是困难的(
Botting & DeLong, 2009 ). ...
... 依据所记录倒木的腐解程度数据, 参照
闫恩荣等(2005) 和
马豪霞等(2016) 对森林粗木质残体腐解等级的划分及判定标准, 确定每株倒木的腐解等级, 并根据腐解等级对倒木进行编号. ...
森林粗死木质残体的概念及其分类
3
2005
... 倒木属于粗死木质残体(coarse woody debris, CWD)的一种(
闫恩荣等, 2005 ).在传统的森林管理过程中, 倒木常被误认为容易引起病虫害和火灾而被清除.近年来, 许多****对森林生态系统的深入研究结果表明, 倒木是森林生态系统不可缺少的重要组成部分, 在减少水土流失、提高林地土壤肥力和加速养分循环等方面发挥着重要作用(
Sandstr?m et al ., 2007 ;
Vanha-Majamaa et al ., 2007 ;
Davis et al ., 2010 ).倒木作为森林生态系统中的持续营养源, 是种类繁多的生物赖以生存的蔽护所和繁殖场地, 也是林木更新的良好苗床, 对森林生态系统的完整性和生物多样性的维持具有重要意义(
Takahashi et al ., 2000 ;
何东进等, 2009 ). ...
... 倒木主要存在于地表并与土壤层联系紧密, 具有相对稳定的环境, 是森林中苔藓植物生长的重要基质(
Preik?a et al ., 2015 ); 它也是一个不断发展的基质, 木材腐烂程度、树皮覆盖度、离地面高度以及一些小气候条件随时间连续变化(
闫恩荣等, 2005 ;
何东进等, 2009 ), 其上生长的苔藓植物也随着环境改变持续变化(
Kumar et al ., 2018 ).苔藓植物组合作为倒木群落内部次优势物种所构成的群落局部集合体(
李博等, 2000 ), 更是随着倒木的腐解进程发生着演替(
Kushnevskaya et al ., 2007 ).一系列的研究探讨了倒木上苔藓植物物种组成和丰度的变化与倒木自身性质以及林地环境的相互关系(
McAlister, 1997 ;
Dittrich et al ., 2014 ;
Preik?a et al ., 2015 ).但由于变量间存在复杂的共线性关系, 要找到直接作用于苔藓组合的生态因子, 并从中抽离出倒木上苔藓植物组合演替规律是困难的(
Botting & DeLong, 2009 ). ...
... 依据所记录倒木的腐解程度数据, 参照
闫恩荣等(2005) 和
马豪霞等(2016) 对森林粗木质残体腐解等级的划分及判定标准, 确定每株倒木的腐解等级, 并根据腐解等级对倒木进行编号. ...
2
2013
... 在早期森林生态系统的研究中, ****们对倒木的关注极少.1948年, McCullough研究了美国天然云杉(
Picea asperata )林3种生境下腐烂倒木上的植物演替, 认为其模式为: 地衣和苔类植物→藓类植物→草本和矮灌木→云杉、冷杉(
Abies fabri )等乔木树种.
Fogel和Cromack (1977) 提出倒木腐烂等级的五级系统, 并研究了倒木的养分变化.20世纪90年代, 倒木的生态功能以及动态特征的研究开始逐渐增加.倒木与生物多样性维持的研究, 主要围绕微生物多样性(木材腐朽菌及大型真菌)和动物多样性展开研究, 也有少量研究涉及植物(如苔藓、地衣)多样性(
游惠明, 2013 ), 在倒木上栖息的植物类型中, 苔藓植物是最大的植物类群(
Dittrich et al ., 2014 ). ...
... 这一系列变化与苔藓生长微生境的改变是密不可分的.倒木基质随着时间不断变化, 树皮的粗糙程度及覆盖度、结构的完整性、养分含量等都在改变(
何东进等, 2009 ), 这些基质的变化对苔藓的定植存在影响(
Dittrich et al ., 2014 ;
Kushnevskaya & Shorohova, 2018 ).另外, 倒木结构完整性、倒木离地面的距离以及倒木距离水源的距离都会影响倒木上的水分情况, 在本研究中4株倒木的水分情况随腐解程度加深而增加.而水分对于苔藓的定植很重要, 会显著影响苔藓植物的多样性(
范苗等, 2017 ).林窗作为森林中常见的小尺度干扰机制, 对于森林物种多样性维持有着重要的意义(
李强等, 2019 ), 树木倒下形成的林窗随着时间推移逐渐闭合, 这也导致不同腐解等级倒木上生长的苔藓受到的光照情况在改变.一般来说, 苔藓喜好相对遮阴的环境, 在这种环境下苔藓植物较维管植物有更大的生长优势(
白学良等, 1998 ;
刘俊华等, 2005 ).在微生境不断变化的情况下, 本研究案例中苔藓植物物种多样性、组合数量和苔藓总盖度在腐解中后期达到最大, 这与其他不同气候下、不同区域的研究趋势一致, 是倒木上苔藓植物演替的一般规律(
Cole et al ., 2008 ;
Botting & DeLong, 2009 ;
游惠明, 2013 ;
Kumar et al ., 2018 ). ...
2
2013
... 在早期森林生态系统的研究中, ****们对倒木的关注极少.1948年, McCullough研究了美国天然云杉(
Picea asperata )林3种生境下腐烂倒木上的植物演替, 认为其模式为: 地衣和苔类植物→藓类植物→草本和矮灌木→云杉、冷杉(
Abies fabri )等乔木树种.
Fogel和Cromack (1977) 提出倒木腐烂等级的五级系统, 并研究了倒木的养分变化.20世纪90年代, 倒木的生态功能以及动态特征的研究开始逐渐增加.倒木与生物多样性维持的研究, 主要围绕微生物多样性(木材腐朽菌及大型真菌)和动物多样性展开研究, 也有少量研究涉及植物(如苔藓、地衣)多样性(
游惠明, 2013 ), 在倒木上栖息的植物类型中, 苔藓植物是最大的植物类群(
Dittrich et al ., 2014 ). ...
... 这一系列变化与苔藓生长微生境的改变是密不可分的.倒木基质随着时间不断变化, 树皮的粗糙程度及覆盖度、结构的完整性、养分含量等都在改变(
何东进等, 2009 ), 这些基质的变化对苔藓的定植存在影响(
Dittrich et al ., 2014 ;
Kushnevskaya & Shorohova, 2018 ).另外, 倒木结构完整性、倒木离地面的距离以及倒木距离水源的距离都会影响倒木上的水分情况, 在本研究中4株倒木的水分情况随腐解程度加深而增加.而水分对于苔藓的定植很重要, 会显著影响苔藓植物的多样性(
范苗等, 2017 ).林窗作为森林中常见的小尺度干扰机制, 对于森林物种多样性维持有着重要的意义(
李强等, 2019 ), 树木倒下形成的林窗随着时间推移逐渐闭合, 这也导致不同腐解等级倒木上生长的苔藓受到的光照情况在改变.一般来说, 苔藓喜好相对遮阴的环境, 在这种环境下苔藓植物较维管植物有更大的生长优势(
白学良等, 1998 ;
刘俊华等, 2005 ).在微生境不断变化的情况下, 本研究案例中苔藓植物物种多样性、组合数量和苔藓总盖度在腐解中后期达到最大, 这与其他不同气候下、不同区域的研究趋势一致, 是倒木上苔藓植物演替的一般规律(
Cole et al ., 2008 ;
Botting & DeLong, 2009 ;
游惠明, 2013 ;
Kumar et al ., 2018 ). ...
 ,3,*
,3,*
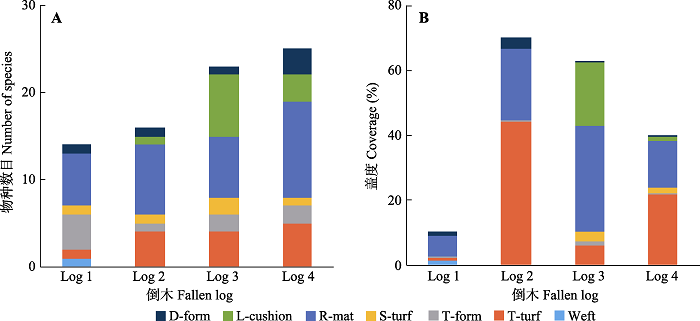 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT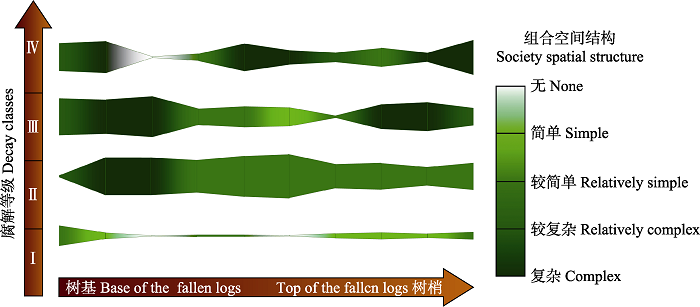 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}