 ,2,*
,2,*Effects of plant interspecific interaction and warming on soil microbial community in root zone soil of two dominant tree species in the subalpine coniferous forest in southwestern China
Lin LUO1,3, Yan HUANG1,3, Jin LIANG1, En-Tao WANG4, Jun HU1, He-Liang HE1,3, Chun-Zhang ZHAO,2,*通讯作者: *zhaochzh04@126.com
编委: 郭良栋
责任编辑: 赵航
收稿日期:2019-12-31接受日期:2020-03-25网络出版日期:2020-08-20
| 基金资助: |
Corresponding authors: *zhaochzh04@126.com
Received:2019-12-31Accepted:2020-03-25Online:2020-08-20
| Fund supported: |

摘要
温度与植物种类是生态系统土壤微生物群落组成与结构的重要影响因子。气候变暖背景下, 不同树种及树种互作对土壤微生物群落产生的影响仍不清楚。该文以西南亚高山针叶林主要建群种粗枝云杉(Picea asperata)和岷江冷杉(Abies faxoniana)为研究对象, 采用红外加热器模拟增温, 通过不同种植方式(云杉、冷杉单种和二者混种, 以及裸地对照), 研究不同物种及增温对土壤微生物磷脂脂肪酸(PLFAs)含量与群落结构的影响。结果表明: (1)无论增温与否, 与裸地相比, 云杉与冷杉单种均显著增加了土壤微生物群落主要类群及总PLFAs含量, 而混种仅在非增温条件下增加了微生物群落PLFAs含量; 另一方面, 增温显著促进了裸地真菌(F)和云杉根区革兰氏阴性菌(GN)的生长, 但对冷杉与冷杉-云杉混种小区微生物群落具有显著的抑制作用。(2)主成分分析(PCA)表明, 非增温条件下, 植物种植对土壤微生物群落组成的影响更为明显。非增温情况下云杉、冷杉单种和混种均对微生物群落结构有显著影响, 显著降低了土壤革兰氏阳性菌/阴性菌(GP/GN), 增加了土壤真菌细菌比(F/B)(64.29%-35.71%), 而增温时, 仅冷杉单种对GP/GN和F/B有显著影响。(3) PLFAs含量与土壤碳含量显著正相关, 微生物群落结构(F/B)则与土壤pH及无机氮含量有显著相关关系。以上结果说明, 在非增温情况下, 无论单种还是混种均有利于土壤微生物生长, 但在增温情况下混种对微生物群落PLFAs含量无显著影响, 两个物种对微生物群落结构的影响在增温条件下也有减弱的趋势。
关键词:
Abstract
Aims Soil microbial community composition and structure were regulated by temperature and plant species. Picea asperata and Abies faxoniana were planted in the monoculture and mixture plantations of the subalpine region in southwestern China. However, the effects of these two species and their interactions on soil microbial community under future climate warming remain unclear.
Methods An experiment was conducted to examine the effects of warming and plant species on soil microbial community composition with two levels of temperature (unwarming and warming with infrared heater) and four planting patterns (single A. faxoniana, single P. asperata, mixture of A. faxoniana and P. asperata, and unplanted bare land). Root zone soil of different planting treatments were sampled to estimating the microbial biomass and microbial community composition by the phospholipid fatty acids (PLFAs) content analysis.
Important findings The results indicated that: (1) Both P. asperata and A. faxoniana mono-planting significantly increased the biomass (PLFAs content) of main soil microbial groups and the whole community, regardless of warming, but the PLFAs content was only increased by mixed planting in unwarming plots. On the other hand, warming enhanced fungi (F) in unplanted plots and gram-negative bacteria (GN) in the P. asperata plots, respectively. However, warming significantly decreased soil microbial biomass in A. faxoniana and the mixed planting plots. (2) Principal component analysis (PCA) showed that effects of planting P. asperata and A. faxoniana on soil microbial community composition were greater under unwarming than under warming conditions. All the planting treatments significantly decreased the ratio of gram-positive/gram-negative bacteria (GP/GN) and increased the ratio of fungi/bacteria (F/B) in unwarming plots. However, significant effects on GP/GN and F/B ratios were only observed in A. faxoniana plots under warming condition. (3) PLFAs content was positively correlated with soil organic carbon, and F/B ratio was significantly correlated with soil pH and inorganic N. These results showed that the effects of warming on soil microbial biomass and composition varied among the tree species, and the effects of P. asperata and A. faxoniana were weakened under warming condition than under unwarming condition. Our results provide a vital theoretical basis for further study on the responses of soil microbial communities to vegetation and global climate change in southwestern China.
Keywords:
PDF (1117KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
罗林, 黄艳, 梁进, 汪恩涛, 胡君, 贺合亮, 赵春章. 西南亚高山针叶林主要树种互作及增温对根区土壤微生物群落的影响. 植物生态学报, 2020, 44(8): 875-884. DOI: 10.17521/cjpe.2019.0369
LUO Lin, HUANG Yan, LIANG Jin, WANG En-Tao, HU Jun, HE He-Liang, ZHAO Chun-Zhang.
土壤微生物被认为是地球元素生物地球化学循环的引擎, 对陆地生态系统碳(C)、氮(N)、磷(P)、硫(S)等养分循环起着关键作用(Falkowski et al., 2008; 沈菊培和贺纪正, 2011; Delgado-Baquerizo et al., 2016)。同时土壤微生物对维持生态系统服务功能也不可或缺, 是“环境污染物净化器”和“陆地生态系统稳定调节器” (宋长青等, 2013; Rui et al., 2015)。研究生物和非生物因素对土壤微生物群落组成的影响机制, 有利于预测环境变化对土壤微生物群落及其相关的土壤物质循环过程的影响(Lladó et al., 2018)。
据IPCC预测, 到21世纪末全球平均气温将上升1.5-4.8 ℃, 全球变暖已进入高速模式(IPCC, 2013)。目前, 全球变暖对土壤微生物群落组成的影响已有许多研究, 如有****发现土壤持续20年升温5 ℃后, 土壤有机层的细菌群落结构发生了明显变化(DeAngelis et al., 2015); Wang等(2017)发现模拟升温会使青藏高原高寒草甸土壤微生物群落从细菌向真菌转移; Zhao等(2014)发现4年夜间增温对青藏高原东部粗枝云杉(Picea asperata)根际微生物群落没有显著影响, 却对岷江冷杉(Abies faxoniana)微生物群落产生了明显的负效应。另一方面, 已有研究报道植物能通过自身生命活动, 如根系周转、根系分泌物及凋落物的类型和数量改变土壤的理化性质, 直接或间接影响土壤微生物群落结构与组成(Lladó et al., 2018), 并且不同植物对微生物群落结构产生不同影响, 如有****在研究北极两种先锋植物山蓼(Oxyria digyna)和虎耳草(Saxifraga oppositifolia)对土壤细菌群落结构影响时发现, 这两种植物根际土壤的细菌群落结构存在显著差异(Kumar et al., 2016); 在凋落物和土壤中, 树种对细菌和真菌的群落组成也有显著影响(Urbanová et al., 2015)。然而, 自然生态系统中往往多种植物共存, 植物之间通过物质交换、资源争夺互相影响, 植物之间的互作必然会对根际土壤微生物产生影响(Treseder et al., 2018), 如Guo等(2019)在研究日本落叶松(Larix kaempferi)和长白落叶松(Larix olgensis)单种和混种对土壤真菌和细菌群落影响时发现两种植物混种比日本落叶松单种有更高的细菌和真菌多样性, 这种差异很有可能是种间互作通过改变根系活动、根际土壤化学性质而导致的; 还有研究发现欧洲赤杨(Alnus glutinosa)-英国橡树(Quercus robur)与白桦(Betula pendula)-橡树混种土壤的微生物群落总生物量均显著低于橡树单种(Gunina et al., 2017)。
增温可以通过直接改变土壤理化特性而影响土壤微生物群落, 也可以通过影响植物根系的生长和生理特征等间接影响根际土壤微生物群落结构(Yin et al., 2012b; Wu et al., 2014)。已有的关于增温对土壤微生物群落的研究结果往往因为关注的生态系统的不同而差异较大, 如Schindlbacher等(2011)对以成熟云杉林为主的温带山地森林生态系统土壤增温4-6年后发现, 增温对云杉林土壤微生物生物量及大多数微生物群落的丰度没有显著影响; 但Allison和Treseder (2008)在对阿拉斯加北部以黑云杉(Picea mariana)为主的森林进行增温后发现, 土壤真菌和细菌的丰度下降超过50%。而且这些研究更多只关注了增温对某种植物或某种植被类型土壤微生物群落的影响, 同时比较分析植物互作及增温对根际土壤微生物群落影响的研究较少见。
西南亚高山针叶林位于青藏高原东部, 是全球变暖的敏感地带, 也是研究全球气候变化对森林生态系统影响的重要地域(刘庆等, 2001)。岷江冷杉和粗枝云杉是西南亚高山针叶林的主要建群种, 二者或单独形成纯林或交互存在形成针叶混交林。云杉和冷杉均是浅根系植物, 对环境温度变化较敏感(D’Amore et al., 2009)。前人对该地区主要树种单独生长时地下生物过程对增温的响应已经有过很多的研究, 如Yin等(2013a)发现增温能显著增加云杉和冷杉的根系分泌物; 在增温条件下, 与冷杉相比, 云杉根际土壤中有更高的硝态氮和更低的铵态氮含量(Yin et al., 2012a); 李月蛟等(2015)发现夜间增温对云杉外生菌根侵染率的影响因季节和根级不同而存在差异。但对该地区主要树种单种和混种时根区土壤微生物群落及其对增温的响应研究较少。我们前期研究发现, 增温对人工云杉纯林及云杉、冷杉为建群种的天然林土壤微生物群落影响具有显著差异(Sun et al., 2016)。然而, 因为云杉纯林与天然林土壤背景如土壤有机质含量、N及微量元素差异显著, 造成土壤微生物群落对增温响应的差异的原因仍不清楚。因此, 本文以冷杉和云杉作为实验对象, 利用红外辐射增温装置模拟气候变暖, 并结合磷脂脂肪酸分析技术, 探究增温条件下西南亚高山主要树种互作对土壤微生物群落的影响, 旨在为全球变暖背景下该地区主要树种根际土壤微生物群落研究提供理论依据。
1 材料和方法
1.1 样地概况与实验设计
研究区域位于青藏高原东南部, 邛崃山脉东部边缘的四川省阿坝州理县东南部杂谷脑镇(31.42° N, 103.20° E), 平均海拔2 700 m, 春夏季降水量多, 冬季无霜期短, 年平均气温6.9 ℃, 年降水量为600- 1 100 mm。样地设置在四川省阿坝州理县米亚罗区杂谷脑镇的一个开阔地上。以西南亚高山针叶林主要建群种粗枝云杉和岷江冷杉为试验对象, 两种树种均来自当地林场苗圃。人工挖掘4对2 m × 2 m × 50 cm (长×宽×深)的开阔小区, 利用客土法将实验地土壤全部换为该林区人工云杉林下过5 mm筛后混匀的表层土, 土壤pH为5.85, 土壤有机碳62.70 mg·g-1, 总氮3.66 mg·g-1, 总磷0.43 mg·g-1, 总钾7.92 mg·g-1。4对2 m × 2 m小区, 每对设置增温与对照处理, 处理之间间隔5 m, 每个2 m × 2 m的小区又分成4个1 m × 1 m亚小区。为避免亚小区内植物根系之间及周围环境的影响, 在亚小区四周埋入深50 cm的双层塑料膜进行隔离。2013年6月挑选长势、大小一致的3年生冷杉及2年生云杉幼苗, 在亚小区内按20 cm × 30 cm间距分别种植冷杉、云杉、云杉和冷杉以及裸地对照, 每个亚小区24株幼苗。2016年预实验发现云杉、冷杉根系已经比较发达, 当年开始对增温小区进行增温处理。增温方式采用悬挂于小区上方距地面1.8 m的165 cm × 15 cm红外辐射加热器(Kalglo Electronics, Bethlehem, USA)进行全天24 h连续增温, 实验期间空气温度和土壤温度分别增加2.0和3.7 ℃ (Yin et al., 2012a)。
试验设计共8个处理, 分别为: 1) U-C: 对照裸地; 2) U-L: 对照冷杉单种; 3) U-Y: 对照云杉单种; 4) U-H: 对照混种; 5) W-C: 增温裸地; 6) W-L: 增温冷杉单种; 7) W-Y: 增温云杉单种; 8) W-H: 增温混种。
1.2 取样与研究方法
2017年3月分别在增温小区与对照小区各亚小区内进行“S”形随机取样, 借助土钻钻取根系周围0-15 cm表层土, 4次重复取样混合为一个土样。所有土壤去除肉眼可见的根系和石块并过2 mm筛后分成两部分储存在4 ℃的保温箱中尽快运回实验室进行进一步分析。土壤总碳(TC)、总氮(TN)含量利用元素分析仪(MACRO cube, Elementar, Germany)测定; 土壤硝态氮(NO3--N)与铵态氮(NH4+-N)含量采用比色法测定(鲁如坤, 2000)。微生物的生物量采用磷脂脂肪酸(PLFA)生物标记法测定(White et al., 1979), 借助以下指示脂肪酸指标对土壤微生物进行分类(Kaiser et al., 2010; Willers et al., 2015): 细菌(Bacteria): 16:1ω11c、16:1ω7c、18:1ω7c、cy17:0、i17:1ω9c、a17:1ω9c、18:1ω5c、i14:0、i15:0、a15:0、i16:0、i17:0、a17:0; 革兰氏阴性菌(GN): 16:1ω11c、16:1ω7c、18:1ω7c、cy17:0、i17:1ω9c、a17:1ω9c、18:1ω5c; 革兰氏阳性菌(GP): i14:0、i15:0、a15:0、i16:0、i17:0、a17:0; 真菌(F): 18:2ω6c和18:1ω9c; 放线菌(Ac): 10Me18:0。其余不确定的碳链算入总PLFAs。
1.3 数据分析
首先, 采用双因素检验分析不同栽种处理、增温及二者交互作用对微生物群落和土壤理化性质的影响; 接着分别对未增温与增温下不同栽种处理对根区土壤微生物群落PLFAs (每种微生物群落及每种碳链)和土壤理化性质的影响进行单因素方差分析。若p < 0.05, 再进行最小显著差异(LSD)多重比较, 并对不同栽种处理未增温与增温时各微生物群落PLFAs和理化指标进行t检验; 然后利用各微生物群落PLFAs数据进行主成分分析(PCA); 最后对土壤微生物群落和环境因子进行Pearson相关性分析。所有数据分析均借助R 3.5.3, 图形绘制利用ggplot2软件包。2 结果和分析
2.1 不同栽种处理和增温对微生物群落PLFAs的影响
栽种处理和增温及二者的交互作用对土壤中总PLFAs及主要类群PLFAs含量具有显著影响(图1)。未增温条件下, 云杉、冷杉单种和二者混种土壤与裸地比较, 总PLFAs、Bacteria、GP、GN、F和AC均显著增加, 增加幅度为42.89% (Bacteria)- 129.98% (F)(图1)。同时冷杉对细菌的促进作用较云杉更强, 在U-L小区, 表征细菌的碳链16:1ω7c、18:1ω7c、cy17:0、i17:1ω9c、a17:1ω9c、18:1ω5c、i14:0、i15:0、a15:0、a17:0均显著增加(图2)。增温条件下与裸地比较(图1), 云杉单种小区所有脂肪酸含量均有显著的提高(图1, 图2), 提高幅度为85.67% (GN)-61.14% (GP); 冷杉单种土壤GP和F没有明显变化, 其他PLFAs含量显著增加(54.41%- 34.30%); 二者混种仅对根区土壤中AC含量有显著的促进作用, 而对其余微生物类群PLFAs含量没有显著影响。图1
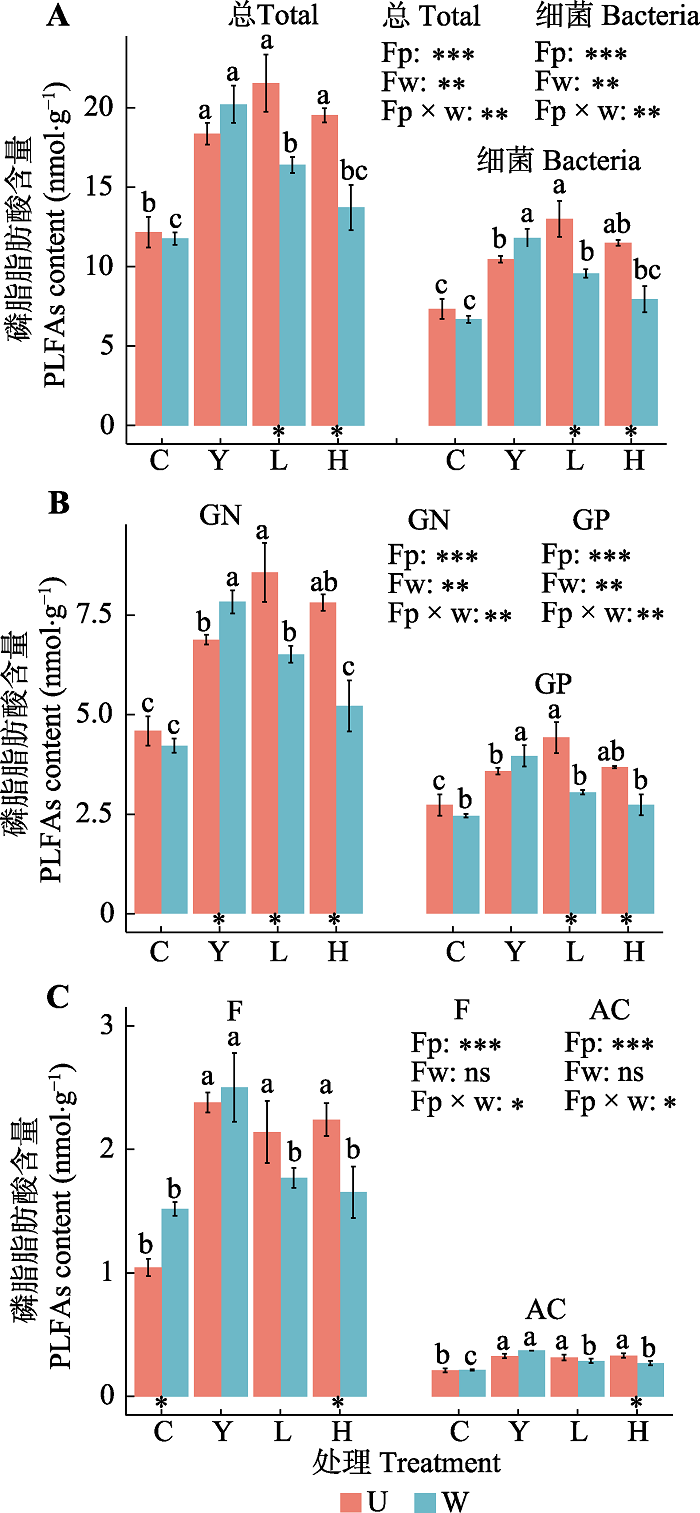 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1未增温和增温时不同栽种处理根区土壤各微生物群落磷脂脂肪酸(PLFAs)含量(平均值±标准误差)。A, 总PLFAs和细菌的PLFAs含量。B, 革兰氏阴性菌(GN)和革兰氏阳性菌(GP)的PLFAs含量。C, 真菌(F)和放线菌(AC)的PLFAs含量。U, 未增温; W, 增温。C, 裸地; Y, 云杉单种; L, 冷杉单种; H, 混种; Fp, 树种效应; Fw, 增温效应; Fp × w, 树种和增温的交互效应; *, p < 0.05; **, p < 0.01; ***, p < 0.001; ns, p > 0.05。不同小写字母表示未增温和增温时不同栽种处理根区土壤微生物群落PLFAs含量有显著差异(p < 0.05)。柱下*表示t检验有显著差异(p < 0.05)。
Fig. 1Contents of phospholipid fatty acids (PLFAs) (mean ± SE) in root zone soil of different planting treatments with/ without warming. A, The content of total and bacteria PLFAs. B, The content of PLFAs of gram-negative bacteria (GN) and gram-positive bacteria (GP). C, The content of PLFAs of fungi (F) and actinomycetes (AC). U, unwarming; W, warming. C, unplanted area; Y, Picea asperata; L, Abies faxoniana; H, mixed; Fp, plant types effect; Fw, warming effect; Fp × w, plant types × warming interaction effect; *, p < 0.05; **, p < 0.01; ***, p < 0.001; ns, p > 0.05. Different lowercase letters indicate significant differences in PLFAs content in root zone of different planting treatments with and without warming (p < 0.05). * under the column indicates a significant difference in the t-test (p < 0.05).
图2
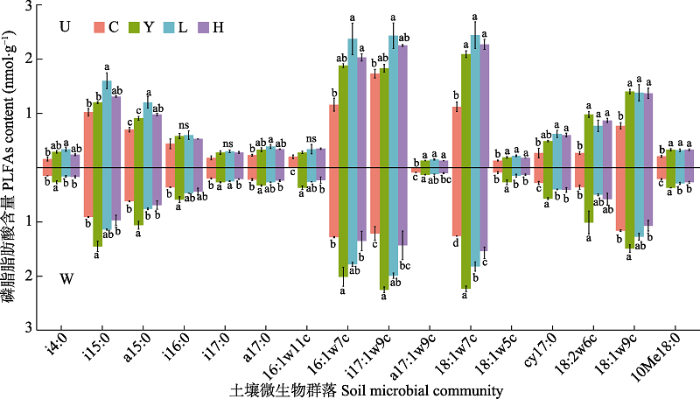 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2不同栽种处理和增温对微生物群落磷脂脂肪酸(PLFAs)含量的影响(平均值±标准误差)。不同小写字母表示未增温和增温时不同栽种处理根区土壤微生物群落PLFAs含量有显著差异(p < 0.05)。U, 未增温; W, 增温。C, 裸地; Y, 云杉单种; L, 冷杉单种; H, 混种。
Fig. 2Effects of different planting treatments and warming on phospholipid fatty acids (PLFAs) content of microbial community (mean ± SE). Different lowercase letters indicate significant differences in PLFAs cotent in root zone of different planting treatments with and without warming (p < 0.05). U, unwarming; W, warming. C, unplanted area; Y, Picea asperata; L, Abies faxoniana; H, mixed.
与未增温相比, 增温条件下裸地土壤中真菌明显增加, 云杉根区土壤中GN的PLFAs含量显著增加了13.79%。但增温显著降低了冷杉单种根区土壤中总PLFAs、Bacteria、GP和GN含量(减少23.90%- 31.08%)以及混种根区土壤中所有微生物群落的PLFAs含量(减少18.34%-33.20%)(图1)。
2.2 不同栽种处理和增温对微生物群落结构的影响
PCA分析中轴1和轴2分别解释了微生物群落结构变异的76.16%和9.19%, 总解释率为85.35% (图3)。图3
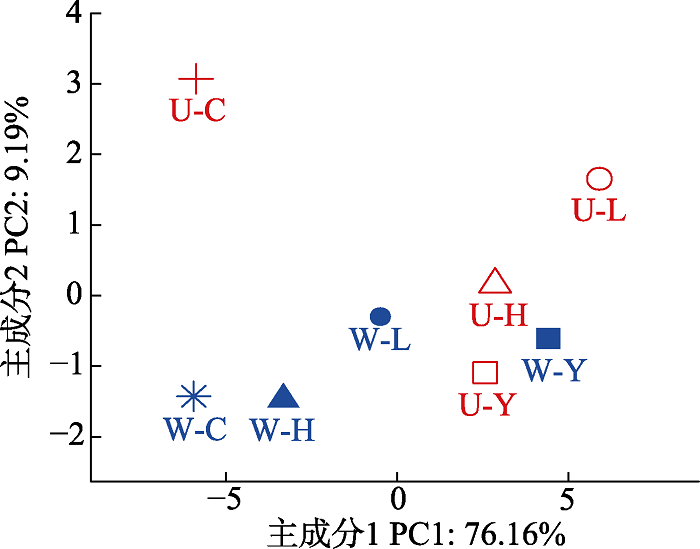 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3不同栽种处理和增温对微生物群落结构的影响。U-C, 不增温裸地; U-H, 不增温混种; U-L, 不增温冷杉; U-Y, 不增温云杉; W-C, 增温裸地; W-H, 增温混种; W-L, 增温冷杉; W-Y, 增温云杉。
Fig. 3Effects of different planting treatments and warming on soil microbial community structure. U-C, unwarming-empty area; U-H, unwarming-mixed area; U-L, unwarming-Abies faxoniana; U-Y, unwarming-Picea asperata; W-C, warming- empty area; W-H, warming-mixed area; W-L, warming-Abies faxoniana; W-Y, warming-Picea asperata.
由图3可见, 在对照条件下云杉、冷杉单种及二者混种均使微生物群落结构与裸地具有明显差异; 增温条件下, 云杉和冷杉单种使土壤微生物群落结构发生了明显变化, 但二者混种对微生物群落结构的影响则不明显。此外, 增温对裸地、冷杉单种及冷杉-云杉混种土壤微生物群落结构均有不同程度的影响, 裸地尤为明显。
对表征微生物群落结构的指标即F/B、GP/GN的分析(图4)表明, 栽种处理和增温及二者的交互作用对F/B影响显著, 仅栽种处理显著了影响GP/GN。对照条件下, 与裸地相比, 云杉单种和云杉-冷杉混种分别使根区土壤中F/B显著增大58.44%和35.55%, 但冷杉单种对F/B没有显著影响; 同时, 云杉、冷杉单种和混种分别导致根区土壤中GP/GN显著减小了12.38%、13.01%和20.41%。增温条件下, 与裸地比较, 仅冷杉单种显著降低了根区土壤中F/B和GP/GN。增温显著增加了裸地土壤中F/B, 并使冷杉单种根区土壤中的GP/GN显著降低。
图4
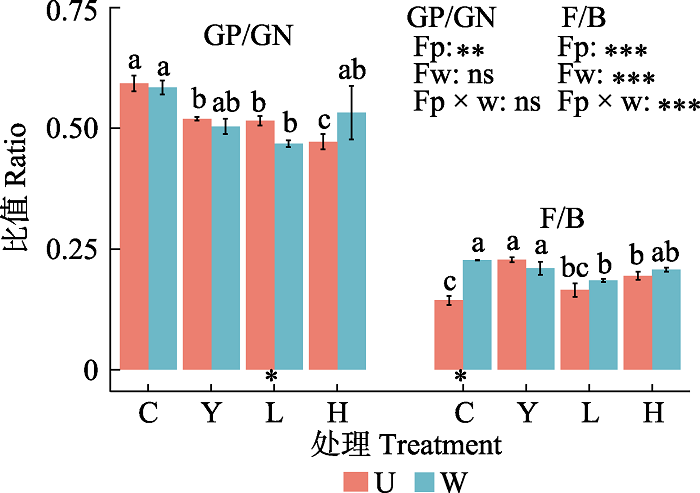 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4未增温和增温时不同栽种处理根区土壤革兰氏阳性菌/革兰氏阴性菌(GP/GN)和真菌/细菌(F/B)(平均值±标准误差)。U, 未增温; W, 增温。C, 裸地; Y, 云杉单种; L, 冷杉单种; H, 混种。Fp, 树种效应; Fw, 增温效应; Fp × w, 树种和增温的交互效应; **, p < 0.01; ***, p < 0.001; ns, p > 0.05。不同小写字母表示未增温和增温时不同栽种处理根区GP/GN和F/B有显著差异(p < 0.05)。柱下*表示t检验有显著差异(p < 0.05)。
Fig. 4The GP/GN and F/B ratios (mean ± SE) in root zone soil of different planting treatments with and without warming. GN, gram-negative bacteria; GP, gram-positive bacteria. F, fungi; B, bacteria. U, unwarming; W, warming. C, unplanted area; Y, Picea asperata; L, Abies faxoniana; H, mixed. Fp, plant types effect; Fw, warming effect; Fp × w, plant types × warming interaction effect; **, p < 0.01; ***, p < 0.001; ns, p > 0.05. Different lowercase letters indicate significant differences in the GP/GN and F/B in root zone of different planting treatments under unwarming and warming (p < 0.05). * under the column indicates a significant difference in the t-test (p < 0.05).
2.3 不同栽种处理和增温对土壤理化性质的影响
不同栽种处理及其与增温的交互作用对根区土壤pH、含水量和硝态氮含量有显著影响; 增温对土壤含水量、总碳、总氮和硝态氮含量影响显著(表1)。Table 1
表1
表1不同栽种处理和增温对土壤理化性质的影响
Table 1
| 土壤因子 Soil factor | 处理 Treatment | C (mean ± SD) | Y (mean ± SD) | L (mean ± SD) | H (mean ± SD) | P | W | P × W |
|---|---|---|---|---|---|---|---|---|
| pH | U | 7.69 ± 0.02b | 7.80 ± 0.04a | 7.70 ± 0.02b | 7.81 ± 0.02a | 10.82*** | 1.08 | 4.65* |
| W | 7.81 ± 0.02a* | 7.81 ± 0.01a | 7.67 ± 0.02b | 7.78 ± 0.03a | ||||
| SM (%) | U | 30.80 ± 0.00bc | 29.60 ± 0.01c | 34.86 ± 0.01a | 32.11 ± 0.00b | 9.51*** | 637.00*** | 41.51*** |
| W | 0.27 ± 0.00a* | 0.24 ± 0.00b* | 0.22 ± 0.00c* | 0.23 ± 0.01bc* | ||||
| TN (mg·kg-1) | U | 2.90 ± 0.04ab | 2.93 ± 0.15ab | 3.15 ± 0.17a | 2.70 ± 0.04b | 0.66 | 7.31* | 2.28 |
| W | 2.64 ± 0.15a | 2.63 ± 0.11a | 2.66 ± 0.08a* | 2.79 ± 0.08a | ||||
| TC (mg·kg-1) | U | 29.86 ± 0.37a | 31.59 ± 0.91a | 32.38 ± 1.35a | 30.06 ± 0.63a | 1.81 | 12.54** | 1.10 |
| W | 28.37 ± 1.04a | 28.55 ± 0.36a* | 29.51 ± 0.22a | 29.48 ± 0.76a | ||||
| NH4+-N (mg·kg-1) | U | 11.29 ± 1.17a | 12.70 ± 1.08a | 9.93 ± 0.38a | 11.48 ± 1.07a | 3.09 | 0.02 | 2.91 |
| W | 11.03 ± 0.38b | 9.93 ± 0.51b | 8.93 ± 0.21b | 15.06 ± 2.36a | ||||
| NO3--N (mg·kg-1) | U | 5.76 ± 0.17b | 8.07 ± 0.38a | 7.82 ± 0.31a | 6.35 ± 0.26b | 108.88*** | 945.64*** | 53.29*** |
| W | 13.39 ± 0.27b* | 15.06 ± 0.07a* | 13.69 ± 0.07b* | 8.10 ± 0.32c* | ||||
| TC/TN | U | 10.31 ± 0.19b | 10.80 ± 0.38ab | 10.29 ± 0.12b | 11.15 ± 0.18a | 0.50 | 0.87 | 2.42 |
| W | 10.77 ± 0.30a | 10.90 ± 0.38a | 11.11 ± 0.34a | 10.57 ± 0.03a |
新窗口打开|下载CSV
在不增温条件下与裸地相比, 云杉单种根区土壤中硝态氮含量显著增加了40.10%, pH也明显增大; 冷杉单种根区土壤含水量和硝态氮含量显著增加了12.90%和35.7%; 云杉和冷杉混种的根区土壤中, pH和TC/TN均显著增大。增温条件下, 云杉、冷杉单种和混种均显著降低了土壤含水量, 此外, 云杉单种显著增加了土壤硝态氮含量(12.47%), 冷杉单种使土壤pH显著降低, 混种的土壤中铵态氮显著增加39.26%而硝态氮显著减少39.51%。增温显著降低了云杉、冷杉单种和混种的土壤含水量(20%-37%), 但是显著增加了3种种植小区土壤硝态氮含量, 分别增加了86.62%、75.06%和27.56% (表1)。增温使裸地pH显著升高、使云杉根区TC含量和冷杉根区TN含量显著下降。
2.4 微生物群落与土壤理化因子的关系
将微生物磷脂脂肪酸(PLFAs)含量与土壤理化因子进行Pearson相关性分析, 结果(表2)表明, F/B与土壤pH、TC/TN和NO3--N含量表现出显著的正相关关系(p < 0.05), 而与TN含量呈显著负相关关系(p < 0.05)。此外, GP含量与土壤含水量及总PLFAs、Bacteria、GP和GN含量与TC含量呈显著正相关关系(p < 0.05)。Table 2
表2
表2微生物群落磷脂脂肪酸(PLFAs)与环境因子的Pearson相关性分析(相关系数)
Table 2
| pH | SM (%) | TN | TC | NH4+-N | NO3--N | TC/TN | |
|---|---|---|---|---|---|---|---|
| Total | 0.05 | 0.33 | 0.25 | 0.47* | -0.22 | 0.01 | 0.16 |
| B | 0.01 | 0.36 | 0.30 | 0.50* | -0.23 | -0.02 | 0.12 |
| GP | 0.04 | 0.44* | 0.36 | 0.53** | -0.27 | -0.05 | 0.04 |
| GN | 0.00 | 0.32 | 0.27 | 0.49* | -0.21 | -0.01 | 0.15 |
| F | 0.35 | 0.08 | -0.04 | 0.21 | -0.07 | 0.18 | 0.37 |
| AC | 0.21 | 0.03 | 0.12 | 0.31 | -0.08 | 0.16 | 0.20 |
| F/B | 0.64*** | -0.38 | -0.46* | -0.31 | 0.21 | 0.41* | 0.45* |
| GP/GN | 0.12 | 0.13 | 0.01 | -0.23 | -0.07 | -0.10 | -0.29 |
新窗口打开|下载CSV
3 讨论
3.1 增温对土壤微生物群落的影响
前人研究发现, 在增温处理下, 根分泌、细根周转比较强烈, 同时与微生物代谢相关的酶的活性会提高, 从而促进微生物群落的生长(Majdi & ?hrvik, 2004; Nazaries et al., 2015)。这有可能是本研究中增温后裸地的F、F/B和云杉单种的GN含量显著增大的原因。但增温后, 裸地和云杉单种小区土壤中总PLFAs含量及主要微生物类群PLFAs含量并没有发生明显变化, 且增温对冷杉及混种根区微生物群落生物量的影响要强于云杉单种(图1): 增温后冷杉单种和混种土壤中总PLFAs含量和主要微生物类群PLFAs含量显著减少。一方面, 微生物群落生长受土壤含水量的影响很大(Brockett et al., 2012), 如有研究发现, 在正常降水和降水减少时期对土壤增温, 土壤微生物生物量发生了截然不同的变化: 正常降水时微生物群落生物量显著增加而缺水年份显著减少(Sheik et al., 2011)。本研究与对照相比, 增温后, 裸地、云杉、冷杉单种和混种土壤含水量均显著减少, 所以有可能是受到水分的限制, 导致增温后裸地(F除外)、云杉单种(GN除外)、冷杉单种和混种土壤中总PLFAs含量和主要微生物群落没有明显变化或显著减少。另一方面, 我们发现增温条件下云杉的根系分泌物显著多于冷杉(Yin et al., 2013b), 更多的根系分泌物能为土壤微生物提供更充足的养分(Marschner et al., 2004; Yin et al., 2013b), 所以有可能即使受到水分的限制, 云杉仍能维持其根区总PLFAs含量和主要微生物群落的相对稳定, 而冷杉单种和混种土壤中微生物群落则表现出对增温显著的负响应, 这与赵春章等(2014)的研究结果一致。3.2 不同栽种处理对土壤微生物群落的影响
首先, 植物通过多种机制直接或间接改变土壤微生物生物量和群落组成, 包括增加地上凋落物和地下根系分泌物等为土壤微生物生长提供所需的营养物质, 从而引起微生物生物量的快速增长(Grayston et al., 1997; van der Wal et al., 2006; Macdonald et al., 2009; Jangid et al., 2011)。本研究中相关性分析表明, 微生物生物量与土壤TC含量呈显著正相关关系(表2)。U-Y、U-L、W-Y、W-L和U-H小区中土壤微生物群落总PLFAs含量和主要微生物类群的含量均显著增加, 但相应处理中土壤总碳的积累增加不显著(表1), 这可能意味着源于植物凋落物和根系分泌物中的碳等营养物质已经被微生物分解释放或转化为其生物量。W-H对总PLFAs含量和主要微生物群落的PLFAs含量没有显著影响。前人在研究植物单种和混种对土壤细菌、真菌群落PLFAs含量的影响时也得出相似的结果, 发现单种的促进作用比混种更明显, 他们认为树种之间的“认同”比树种多样性对微生物群落有更强的作用(Gunina et al., 2017)。本文中云杉和冷杉混种在增温条件下对根区土壤主要微生物群落PLFAs含量的影响弱于单种, 可能是混种时种间互作减弱或掩盖了单种对微生物群落的促进作用, 而这种种间互作在本研究中未增温时并没有发现。其次, 不同树种对微生物群落结构的影响存在差异(Uroz et al., 2016)。实验中未增温时, 冷杉根区细菌的增加显著大于云杉, 进而导致冷杉根区F/B显著低于云杉。这可能反映了不同树种通过其根系活动诸如根系周转、分泌物组成等影响土壤理化性质, 进而直接或间接改变土壤微生物群落结构(Lladó et al., 2018)。有研究发现土壤含水量与F/B存在显著的相关关系, 在良好的水分条件下, 细菌竞争更占优势, 而在缺水的环境中, 真菌更耐旱(Hackl et al., 2005; 张地等, 2012)。在本研究中, 与云杉单种比较, 冷杉单种根区土壤中细菌增加更明显, 可能是种植冷杉后土壤含水量显著增加(表1), 良好的水资源更有利于细菌的生长; 而云杉单种时其土壤含水量与裸地比较没有明显变化且显著低于冷杉单种, 所以真菌表现出更好的竞争优势, 云杉和冷杉单种土壤含水量的差异可能是二者F/B不同的原因。另外微生物群落组成的差异往往与植物种类有关(Wan et al., 2015), 我们前期研究发现云杉和冷杉两种幼苗生长和根系特性、生理过程如根生物量、根系类型、细根周转以及凋落物和根系分泌物数量和质量(土壤微生物群落的主要碳源)等具有显著差异(Nakadai et al., 2002; Zhao & Liu, 2008; Bardgett et al., 2014; Uroz et al., 2016), 这些差异都可能导致云杉和冷杉对微生物群落产生不同程度的影响。
本研究还发现增温弱化了植物对微生物群落结构的影响, 与对照相比, 增温后有植物处理小区的微生物群落结构与裸地更相似。未增温条件下, 单种和混种F/B与GP/GN均发生了显著变化, 但与W-C比较, F/B和GP/GN仅在W-L小区显著减小, 而在W-Y和W-H小区保持稳定(图4)。温度升高通过多种方式影响土壤微生物群落结构, 如改变土壤理化性质等对微生物群落产生影响(Rui et al., 2015)。本文相关性分析表明, F/B与土壤pH、GP与土壤含水量均表现出显著正相关关系。与W-C比较, W-L和W-H的土壤pH保持稳定, 但W-L的土壤pH显著降低; 同时W-L含水量最低。所以有可能是增温条件下, 土壤酸碱度和含水量的差异导致云杉和混种根区微生物群落结构与W-C更接近, 而冷杉的微生物群落结构发生显著变化。
4 结论
本研究利用红外辐射加热器模拟增温, 研究气候变暖对西南亚高山主要建群树种互作对根区土壤微生物群落生物量和结构的影响。结果表明, 不论增温与否, 云杉、冷杉单种均对土壤微生物生物量有显著的促进作用, 使微生物群落结构发生显著变化; 混种仅在未增温时显著促进土壤微生物PLFAs含量的增加, 明显改变微生物群落结构, 而在增温时对微生物群落PLFAs含量和结构没有明显作用。植物种植对土壤微生物群落结构的影响在增温条件下有减弱的趋势。此外, 增温对冷杉和混种根区微生物群落有显著的负效应, 而对裸地F和云杉GN生长表现出显著的正效应。在以后的研究中, 可以考虑结合高通量测序等技术, 在基因水平上更深入地探究西南亚高山针叶林土壤微生物群落对树种的不同种植方式及全球变暖的响应。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1111/j.1365-2486.2008.01716.xURL [本文引用: 1]
DOI:10.1016/j.tree.2014.10.006URLPMID:25459399 [本文引用: 1]

Ecologists are increasingly adopting trait-based approaches to understand how community change influences ecosystem processes. However, most of this research has focussed on aboveground plant traits, whereas it is becoming clear that root traits are important drivers of many ecosystem processes, such as carbon (C) and nutrient cycling, and the formation and structural stability of soil. Here, we synthesise emerging evidence that illustrates how root traits impact ecosystem processes, and propose a pathway to unravel the complex roles of root traits in driving ecosystem processes and their response to global change. Finally, we identify research challenges and novel technologies to address them.
DOI:10.1016/j.soilbio.2011.09.003URL [本文引用: 1]
DOI:10.1016/j.foreco.2009.03.006URL [本文引用: 1]
DOI:10.3389/fmicb.2015.00104URLPMID:25762989 [本文引用: 1]
Soil microbes are major drivers of soil carbon cycling, yet we lack an understanding of how climate warming will affect microbial communities. Three ongoing field studies at the Harvard Forest Long-term Ecological Research (LTER) site (Petersham, MA) have warmed soils 5 degrees C above ambient temperatures for 5, 8, and 20 years. We used this chronosequence to test the hypothesis that soil microbial communities have changed in response to chronic warming. Bacterial community composition was studied using Illumina sequencing of the 16S ribosomal RNA gene, and bacterial and fungal abundance were assessed using quantitative PCR. Only the 20-year warmed site exhibited significant change in bacterial community structure in the organic soil horizon, with no significant changes in the mineral soil. The dominant taxa, abundant at 0.1% or greater, represented 0.3% of the richness but nearly 50% of the observations (sequences). Individual members of the Actinobacteria, Alphaproteobacteria and Acidobacteria showed strong warming responses, with one Actinomycete decreasing from 4.5 to 1% relative abundance with warming. Ribosomal RNA copy number can obfuscate community profiles, but is also correlated with maximum growth rate or trophic strategy among bacteria. Ribosomal RNA copy number correction did not affect community profiles, but rRNA copy number was significantly decreased in warming plots compared to controls. Increased bacterial evenness, shifting beta diversity, decreased fungal abundance and increased abundance of bacteria with low rRNA operon copy number, including Alphaproteobacteria and Acidobacteria, together suggest that more or alternative niche space is being created over the course of long-term warming.
DOI:10.1038/ncomms10541URLPMID:26817514 [本文引用: 1]
Despite the importance of microbial communities for ecosystem services and human welfare, the relationship between microbial diversity and multiple ecosystem functions and services (that is, multifunctionality) at the global scale has yet to be evaluated. Here we use two independent, large-scale databases with contrasting geographic coverage (from 78 global drylands and from 179 locations across Scotland, respectively), and report that soil microbial diversity positively relates to multifunctionality in terrestrial ecosystems. The direct positive effects of microbial diversity were maintained even when accounting simultaneously for multiple multifunctionality drivers (climate, soil abiotic factors and spatial predictors). Our findings provide empirical evidence that any loss in microbial diversity will likely reduce multifunctionality, negatively impacting the provision of services such as climate regulation, soil fertility and food and fibre production by terrestrial ecosystems.
DOI:10.1126/science.1153213URLPMID:18497287 [本文引用: 1]
Virtually all nonequilibrium electron transfers on Earth are driven by a set of nanobiological machines composed largely of multimeric protein complexes associated with a small number of prosthetic groups. These machines evolved exclusively in microbes early in our planet's history yet, despite their antiquity, are highly conserved. Hence, although there is enormous genetic diversity in nature, there remains a relatively stable set of core genes coding for the major redox reactions essential for life and biogeochemical cycles. These genes created and coevolved with biogeochemical cycles and were passed from microbe to microbe primarily by horizontal gene transfer. A major challenge in the coming decades is to understand how these machines evolved, how they work, and the processes that control their activity on both molecular and planetary scales.
DOI:10.1016/S0929-1393(96)00126-6URL [本文引用: 1]
DOI:10.1007/s11104-016-3073-0URL [本文引用: 2]
DOI:10.1016/j.soilbio.2018.10.018URL [本文引用: 1]
DOI:10.1016/j.soilbio.2004.08.023URL [本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.soilbio.2011.06.022URL [本文引用: 1]
DOI:10.1111/j.1469-8137.2010.03321.xURL [本文引用: 1]
DOI:10.1007/s11104-015-2702-3URL [本文引用: 1]
[本文引用: 2]
[本文引用: 2]
DOI:10.1007/s00253-018-8950-4URLPMID:29600493 [本文引用: 3]
Forests are essential biomes for global biogeochemical cycles, and belowground microorganisms have a key role in providing relevant ecosystem services. To predict the effects of environmental changes on these ecosystem services requires a comprehensive understanding of how biotic and abiotic factors drive the composition of microbial communities in soil. However, microorganisms are not homogeneously distributed in complex environments such as soil, with different features affecting microbes at different extent depending on the niche they occupy. Indeed, this spatial heterogeneity hampers the extrapolation of microbial diversity study results from particular habitats to the ecosystem level, even if the resolution of the more recent studies has increased significantly after the standardization of high-throughput sequencing techniques. The present work intends to give a comprehensive view of the knowledge accumulated until date defining the more important drivers determining the structure of forest soil microbial communities from fine to continental scales.
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.soilbio.2009.05.003URL [本文引用: 1]
DOI:10.1111/gcb.2004.10.issue-2URL [本文引用: 1]
DOI:10.1023/B:PLSO.0000035569.80747.c5URL [本文引用: 1]
Rhizosphere microbial communities are important for plant nutrition and plant health. Using the culture-independent method of PCR-DGGE of 16S rDNA for community analyses, we conducted several experiments to investigate the importance of pH, soil type, soil amendment, nutritional status of the plant, plant species and plant age on the structure of the bacterial community in the rhizosphere. At the same time, we assessed the spatial variability of bacterial communities in different root zone locations. Our results showed that the bacterial community structure is influenced by soil pH and type of P fertilization. In a short-term experiment (15–22 days) with cucumber and barley growing in a N deficient or a P deficient soil, the bacterial community structure in the rhizosphere was affected by soil type and fertilization but not by plant species. In a 7.5-week experiment with three plant species (chickpea, canola, Sudan grass) growing in three different soils (a sand, a loam and a clay), the complex interactions between soil and plant effects on the rhizosphere community were apparent. In the sand and the loam, the three plant species had distinct rhizosphere communities while in the clay soil the rhizosphere community structures of canola and Sudan grass were similar and differed from those of chickpea. In all soils, the rhizosphere community structures of the root tip were different from those in the mature root zone. In white lupin, the bacterial community structure of the non-cluster roots differed from those of the cluster roots. As plants matured, different cluster root age classes (young, mature, old) had distinct rhizosphere communities. We conclude that many different factors will contribute to shaping the species composition in the rhizosphere, but that the plant itself exerts a highly selective effect that is at least as great as that of the soil. Root exudate amount and composition are the key drivers for the differences in community structure observed in this study.]]>
DOI:10.1016/S0929-1393(01)00180-9URL [本文引用: 1]
DOI:10.1016/j.soilbio.2015.06.027URL [本文引用: 1]
DOI:10.1128/AEM.00557-15URLPMID:26116682 [本文引用: 2]
The soil microbial community plays an important role in terrestrial carbon and nitrogen cycling. However, microbial responses to climate warming or cooling remain poorly understood, limiting our ability to predict the consequences of future climate changes. To address this issue, it is critical to identify microbes sensitive to climate change and key driving factors shifting microbial communities. In this study, alpine soil transplant experiments were conducted downward or upward along an elevation gradient between 3,200 and 3,800 m in the Qinghai-Tibet plateau to simulate climate warming or cooling. After a 2-year soil transplant experiment, soil bacterial communities were analyzed by pyrosequencing of 16S rRNA gene amplicons. The results showed that the transplanted soil bacterial communities became more similar to those in their destination sites and more different from those in their
DOI:10.1016/j.soilbio.2011.03.005URLPMID:21760644 [本文引用: 1]
Soil microbial communities mediate the decomposition of soil organic matter (SOM). The amount of carbon (C) that is respired leaves the soil as CO(2) (soil respiration) and causes one of the greatest fluxes in the global carbon cycle. How soil microbial communities will respond to global warming, however, is not well understood. To elucidate the effect of warming on the microbial community we analyzed soil from the soil warming experiment Achenkirch, Austria. Soil of a mature spruce forest was warmed by 4 degrees C during snow-free seasons since 2004. Repeated soil sampling from control and warmed plots took place from 2008 until 2010. We monitored microbial biomass C and nitrogen (N). Microbial community composition was assessed by phospholipid fatty acid analysis (PLFA) and by quantitative real time polymerase chain reaction (qPCR) of ribosomal RNA genes. Microbial metabolic activity was estimated by soil respiration to biomass ratios and RNA to DNA ratios. Soil warming did not affect microbial biomass, nor did warming affect the abundances of most microbial groups. Warming significantly enhanced microbial metabolic activity in terms of soil respiration per amount of microbial biomass C. Microbial stress biomarkers were elevated in warmed plots. In summary, the 4 degrees C increase in soil temperature during the snow-free season had no influence on microbial community composition and biomass but strongly increased microbial metabolic activity and hence reduced carbon use efficiency.
DOI:10.1038/ismej.2011.32URLPMID:21451582 [本文引用: 2]
The soil microbiome is responsible for mediating key ecological processes; however, little is known about its sensitivity to climate change. Observed increases in global temperatures and alteration to rainfall patterns, due to anthropogenic release of greenhouse gases, will likely have a strong influence on soil microbial communities and ultimately the ecosystem services they provide. Therefore, it is vital to understand how soil microbial communities will respond to future climate change scenarios. To this end, we surveyed the abundance, diversity and structure of microbial communities over a 2-year period from a long-term in situ warming experiment that experienced a moderate natural drought. We found the warming treatment and soil water budgets strongly influence bacterial population size and diversity. In normal precipitation years, the warming treatment significantly increased microbial population size 40-150% but decreased diversity and significantly changed the composition of the community when compared with the unwarmed controls. However during drought conditions, the warming treatment significantly reduced soil moisture thereby creating unfavorable growth conditions that led to a 50-80% reduction in the microbial population size when compared with the control. Warmed plots also saw an increase in species richness, diversity and evenness; however, community composition was unaffected suggesting that few phylotypes may be active under these stressful conditions. Our results indicate that under warmed conditions, ecosystem water budget regulates the abundance and diversity of microbial populations and that rainfall timing is critical at the onset of drought for sustaining microbial populations.
[本文引用: 1]
31,
[本文引用: 1]
DOI:10.11867/j.issn.1001-8166.2013.10.1087URL
Soils are fundamental to preservation and sustainability of life-support system on Earth. Soils develop as the most dynamic and complex interface linking atmosphere, hydrosphere, lithosphere and biosphere. Soils harbor enormous diversities of microbial communities as the primary driving forces for global exchanges of matter and energy on our planet. Despite of its profound importance, the invisible soil microbes have for long been underappreciated. In the early 2000s, there has been growing awareness that soil microbiology has attracted huge interest from nonsoil scientists due to the introduction of threedomain phylogeny. It is also known as tree of life theory which is widely recognized as the most accurate reflection of the relatedness of all organisms and provides us with a tool to classify and elucidate the largely untapped resource of soil microbial communities. In January 2005, the Department of Earth Sciences of National Natural Science Foundation of China organized a workshop of ‘Soil Biology and Soil Processes’ with focused discussion on soil microbiology research frontiers. The workshop outlined research priorities, crossdisciplinary research opportunities, technological needs and potential breakthroughs within soil microbiology. This workshop has witnessed the rapid advances of soil microbiology in soil nutrient transformation, global environmental changes and environmental remediation over the last decade in China. This article will give a brief review on soil microbial researches in the past decade in China, present the status quo of funding system and highlight the challenge and opportunities for future soil microbiology in China.
DOI:10.1007/s11629-015-3733-1URL [本文引用: 1]
DOI:10.1111/1365-2745.12919URL [本文引用: 1]
DOI:10.1016/j.soilbio.2015.02.011URL [本文引用: 1]
DOI:10.1038/srep27756URLPMID:27302652 [本文引用: 2]
The impacts of plant species on the microbial communities and physico-chemical characteristics of soil are well documented for many herbs, grasses and legumes but much less so for tree species. Here, we investigate by rRNA and ITS amplicon sequencing the diversity of microorganisms from the three domains of life (Archaea, Bacteria and Eukaryota:Fungi) in soil samples taken from the forest experimental site of Breuil-Chenue (France). We discovered significant differences in the abundance, composition and structure of the microbial communities associated with two phylogenetically distant tree species of the same age, deciduous European beech (Fagus sylvatica) and coniferous Norway spruce (Picea abies Karst), planted in the same soil. Our results suggest a significant effect of tree species on soil microbiota though in different ways for each of the three microbial groups. Fungal and archaeal community structures and compositions are mainly determined according to tree species, whereas bacterial communities differ to a great degree between rhizosphere and bulk soils, regardless of the tree species. These results were confirmed by quantitative PCR, which revealed significant enrichment of specific bacterial genera, such as Burkholderia and Collimonas, known for their ability to weather minerals within the tree root vicinity.
DOI:10.1016/j.soilbio.2005.04.017URL [本文引用: 1]
DOI:10.1007/s11104-014-2277-4URL [本文引用: 1]
DOI:10.1016/j.apsoil.2017.03.005URL [本文引用: 1]
DOI:10.1007/BF00388810URLPMID:28309603 [本文引用: 1]
The measurement of lipid phosphate is proposed as an indicator of microbial biomass in marine and estuarine sediments. This relatively simple assay can be performed on fresh, frozen or frozen-lyophilized sediment samples with chloroform methanol extraction and subsequent phosphate determination. The sedimentary lipid phosphate recovery correlates with the extractible ATP and the rate of DNA synthesis. Pulse-chase experiments show active metabolism of the sedimentary phospholipids. The recovery of added (14)C-labeled bacterial lipids from sediments is quantitative. Replicate analyses from a single sediment sample gave a standard deviation of 11%. The lipid extract can be fractionated by relatively simple procedures and the plasmalogen, diacyl phospholipid, phosphonolipid and non-hydrolyzable phospholipid content determined. The relative fatty acid composition can be readily determined by gas-liquid chromatography.The lipid composition can be used to define the microbial community structure. For example, the absence of polyenoic fatty acids indicates minimal contamination with benthic micro-eukaryotes. Therefore the high content of plasmalogen phospholipids in these sediments suggests that the anaerobic prokaryotic Clostridia are found in the aerobic sedimentary horizon. This would require anaerobic microhabitats in the aerated zones.
DOI:10.1111/jam.12902URLPMID:26184497 [本文引用: 1]
Profiling of microbial communities in environmental samples often utilizes phospholipid fatty acid (PLFA) analysis. This method has been used for more than 35 years and is still popular as a means to characterize microbial communities in a diverse range of environmental matrices. This review examines the various recent applications of PLFA analysis in environmental studies with specific reference to the interpretation of the PLFA results. It is evident that interpretations of PLFA results do not always correlate between different investigations. These discrepancies in interpretation and their subsequent applications to environmental studies are discussed. However, in spite of limitations to the manner in which PLFA data are applied, the approach remains one with great potential for improving our understanding of the relationship between microbial populations and the environment. This review highlights the caveats and provides suggestions towards the practicable application of PLFA data interpretation.
DOI:10.1007/s11258-014-0364-5URL [本文引用: 1]
Roots form one of the most important carbon (C) pools in alpine ecosystems. Uncertainty about the effects of warming on root dynamics limits our ability to predict how C will transfer between biological and atmospheric pools in alpine regions under global warming. We used a minirhizotron technique to gain a better understanding of the response of alpine plant roots to warming. We looked for effects on root diameter, root depth in the soil, and root lifespan under a controlled asymmetrical warming (1.2/1.7 A degrees C during daytime/nighttime) experiment during the growing season of 2009 in an alpine meadow on the northern Tibetan plateau. Roots became smaller in diameter, moved toward the upper soil layers, and showed significantly shorter lifespans in heated (H) than in unheated (UH) plots. Furthermore, in H treatment plots root lifespan was more strongly influenced by the time of root emergence rather than by root diameter. These results provide evidence that alpine plants may respond to climate change by altering their roots so that they are thinner, distributed shallower and turning over faster, suggesting that soil C flow and nutrient cycling would be accelerated resulting from the fast turnover of fine roots under elevated temperatures.
DOI:10.1016/j.soilbio.2012.03.004URL [本文引用: 2]
DOI:10.1111/gcb.12161URL
Despite the perceived importance of exudation to forest ecosystem function, few studies have attempted to examine the effects of elevated temperature and nutrition availability on the rates of root exudation and associated microbial processes. In this study, we performed an experiment in which in situ exudates were collected from Picea asperata seedlings that were transplanted in disturbed soils exposed to two levels of temperature (ambient temperature and infrared heater warming) and two nitrogen levels (unfertilized and 25g Nm-2a-1). Here, we show that the trees exposed to an elevated temperature increased their exudation rates I (gCg-1 root biomassh-1), II (gCcm-1root lengthh-1) and III (gCcm-2root areah-1) in the unfertilized plots. The altered morphological and physiological traits of the roots exposed to experimental warming could be responsible for this variation in root exudation. Moreover, these increases in root-derived C were positively correlated with the microbial release of extracellular enzymes involved in the breakdown of organic N (R2=0.790; P=0.038), which was coupled with stimulated microbial activity and accelerated N transformations in the unfertilized soils. In contrast, the trees exposed to both experimental warming and N fertilization did not show increased exudation rates or soil enzyme activity, indicating that the stimulatory effects of experimental warming on root exudation depend on soil fertility. Collectively, our results provide preliminary evidence that an increase in the release of root exudates into the soil may be an important physiological adjustment by which the sustained growth responses of plants to experimental warming may be maintained via enhanced soil microbial activity and soil N transformation. Accordingly, the underlying mechanisms by which plant root-microbe interactions influence soil organic matter decomposition and N cycling should be incorporated into climate-carbon cycle models to determine reliable estimates of long-term C storage in forests.
DOI:10.1016/j.agrformet.2013.06.016URL [本文引用: 2]
DOI:10.1016/j.apsoil.2012.03.013URL [本文引用: 2]
Tree species can exert a strong influence on rhizosphere nutrient cycling through root and rhizosphere processes and create feedback in the patterns of nutrient cycling in forest ecosystems. In this study, we conducted an experiment to compare the rhizosphere effects of two coniferous species on N transformation as well as their responses to experimental warming using infrared heaters in the Eastern Tibetan Plateau. We examined the potential net N mineralization and nitrification rates. N availability, and microbial biomass C (MBC) and N (MBN) in rhizosphere soils of Picea asperata and Abies faxoniana plots and compared them to bulk soils. The infrared heater increased both the mean air and the soil temperatures by 1.5 degrees C and 2.1 degrees C respectively. Potential net N mineralization and net nitrification rates were generally greater in rhizosphere soils for the two conifers than in bulk soil, especially in the warmed plots. This led to higher NH4+ and NO3- concentrations in the rhizosphere soils. MBC and MBN were markedly higher in the rhizosphere soils relative to bulk soil in the study plots. In the control subplots of P. asperata, MBC, MBN, potential net N mineralization and net nitrification rates in the rhizosphere were 9.6%, 21.7%, 33.3% and 20.1% greater than in the bulk soil, respectively. MBC, MBN, potential net N mineralization and net nitrification rates in the control subplots of A. faxoniana, however, were 2.0%, 7.7%, 22.0% and 11.8% higher, respectively, in the rhizosphere than in the bulk soil; all of the variables were significantly lower than those of P. asperata subplots. Warming significantly promoted N transformation and nutrient availability by enhancing the rhizosphere priming effects for the two conifers, but the magnitudes of the rhizosphere effects on soil N transformation stimulated by warming were generally greater in P. asperata than in A. faxoniana subplots. Differences in the altered morphological and functional characteristics of the roots between the two species under experimental warming could be largely responsible for this variation. Taken together, the results indicated that the two species exhibited similar patterns but with considerably different magnitudes of rhizosphere effects on N transformations in response to experimental warming, implying different capacities of the two conifers to acquire nutrients and thereby altered the competitive and adaptive relationships between the tree species under climate change. (c) 2012 Elsevier B.V.
URL
Quercus liaotungensis forest soil in Dongling Mountain of Beijing as the object, and by using chloroform fumigationextraction and phospholipid fatty acid (PLFA) analysis methods, this paper studied the variation characteristics of soil microbial community along an altitudinal gradient in the tree growth season. With increasing altitude, the soil microbial biomass carbon and nitrogen and the quantities of various soil microbial groups in the forest had definite differences but not significant. The ratio of soil bacteria to fungi increased, but the ratio of G+ to G- bacteria decreased. The soil microbial biomass carbon and nitrogen and the quantities of soil bacteria, fungi, and G+ and G- bacteria had significant positive correlations with the contents of soil moisture, organic carbon, and total nitrogen, and the quantity of soil fungi was positively correlated with soil carbon/nitrogen ratio. The variations of the soil microbial community structure (bacteria/fungi and G+/G- bacteria) were mainly affected by soil temperature and moisture content, which meant that the soil microbial community structure was sensitive to the environmental conditions. Along with the global warming, the proportions of soil fungi and G+ bacteria in the Q. liaotungensis forests in warm temperate zone would have an increase.]]>
DOI:10.1007/s11738-008-0217-8URL [本文引用: 1]
DOI:10.1007/s11104-014-2153-2URL [本文引用: 1]
An experiment was conducted to examine soil microbial communities and their related soil processes in rhizospheric soil of two coniferous species that were exposed to two levels of temperature (unwarmed and infrared heater warming) and two levels of nitrogen (unfertilized and 25 g N m(-2) a(-1)) from April 2007.Four-year night warming alone slightly affected the phospholipid fatty acid contents of the microbial community. However, the combination of nitrogen addition and soil warming significantly affected soil microbial composition while reducing the biomass of major microbial groups and the activities of most enzymes, especially in Abies faxoniana plots. The combination of warming and nitrogen addition increased soil labile C and N pools in Picea asperata plots and was beneficial for soil recalcitrant C, as well as for labile and total C and N pools in A. faxoniana plots.Results indicated that future warming will slightly affect soil microbial communities and their related soil processes. However, warming combined with high nitrogen deposition will significantly constrain soil microbial biomass and enzyme activities, consequently increasing soil C and N pools in sub-alpine coniferous forests of this region.]]>
Warming and drying suppress microbial activity and carbon cycling in boreal forest soils
1
2008
... 增温可以通过直接改变土壤理化特性而影响土壤微生物群落, 也可以通过影响植物根系的生长和生理特征等间接影响根际土壤微生物群落结构(
Going underground: root traits as drivers of ecosystem processes
1
2014
... 其次, 不同树种对微生物群落结构的影响存在差异(
Soil moisture is the major factor influencing microbial community structure and enzyme activities across seven biogeoclimatic zones in western Canada
1
2012
... 前人研究发现, 在增温处理下, 根分泌、细根周转比较强烈, 同时与微生物代谢相关的酶的活性会提高, 从而促进微生物群落的生长(
Adaptation to exploit nitrate in surface soils predisposes yellow-cedar to climate-induced decline while enhancing the survival of western redcedar: a new hypothesis
1
2009
... 西南亚高山针叶林位于青藏高原东部, 是全球变暖的敏感地带, 也是研究全球气候变化对森林生态系统影响的重要地域(
Long-term forest soil warming alters microbial communities in temperate forest soils
1
2015
... 据IPCC预测, 到21世纪末全球平均气温将上升1.5-4.8 ℃, 全球变暖已进入高速模式(
Microbial diversity drives multifunctionality in terrestrial ecosystems
1
2016
... 土壤微生物被认为是地球元素生物地球化学循环的引擎, 对陆地生态系统碳(C)、氮(N)、磷(P)、硫(S)等养分循环起着关键作用(
The microbial engines that drive Earth?s biogeochemical cycles
1
2008
... 土壤微生物被认为是地球元素生物地球化学循环的引擎, 对陆地生态系统碳(C)、氮(N)、磷(P)、硫(S)等养分循环起着关键作用(
Rhizosphere carbon flow in trees, in comparison with annual plants: the importance of root exudation and its impact on microbial activity and nutrient availability
1
1997
... 首先, 植物通过多种机制直接或间接改变土壤微生物生物量和群落组成, 包括增加地上凋落物和地下根系分泌物等为土壤微生物生长提供所需的营养物质, 从而引起微生物生物量的快速增长(
Response of soil microbial community to afforestation with pure and mixed species
2
2017
... 据IPCC预测, 到21世纪末全球平均气温将上升1.5-4.8 ℃, 全球变暖已进入高速模式(
... 首先, 植物通过多种机制直接或间接改变土壤微生物生物量和群落组成, 包括增加地上凋落物和地下根系分泌物等为土壤微生物生长提供所需的营养物质, 从而引起微生物生物量的快速增长(
Plant-plant interactions and N fertilization shape soil bacterial and fungal communities
1
2019
... 据IPCC预测, 到21世纪末全球平均气温将上升1.5-4.8 ℃, 全球变暖已进入高速模式(
Composition of the microbial communities in the mineral soil under different types of natural forest
1
2005
... 其次, 不同树种对微生物群落结构的影响存在差异(
1
2013
... 据IPCC预测, 到21世纪末全球平均气温将上升1.5-4.8 ℃, 全球变暖已进入高速模式(
Land-use history has a stronger impact on soil microbial community composition than aboveground vegetation and soil properties
1
2011
... 首先, 植物通过多种机制直接或间接改变土壤微生物生物量和群落组成, 包括增加地上凋落物和地下根系分泌物等为土壤微生物生长提供所需的营养物质, 从而引起微生物生物量的快速增长(
Belowground carbon allocation by trees drives seasonal patterns of extracellular enzyme activities by altering microbial community composition in a beech forest soil
1
2010
... 土壤总碳(TC)、总氮(TN)含量利用元素分析仪(MACRO cube, Elementar, Germany)测定; 土壤硝态氮(NO3--N)与铵态氮(NH4+-N)含量采用比色法测定(
Plants impact structure and function of bacterial communities in Arctic soils
1
2016
... 据IPCC预测, 到21世纪末全球平均气温将上升1.5-4.8 ℃, 全球变暖已进入高速模式(
连续三年夜间增温和施氮对云杉外生菌根及菌根真菌多样性的影响
2
2015
... 西南亚高山针叶林位于青藏高原东部, 是全球变暖的敏感地带, 也是研究全球气候变化对森林生态系统影响的重要地域(
... );
连续三年夜间增温和施氮对云杉外生菌根及菌根真菌多样性的影响
2
2015
... 西南亚高山针叶林位于青藏高原东部, 是全球变暖的敏感地带, 也是研究全球气候变化对森林生态系统影响的重要地域(
... );
中国西南亚高山针叶林的生态学问题
2001
中国西南亚高山针叶林的生态学问题
2001
Drivers of microbial community structure in forest soils
3
2018
... 土壤微生物被认为是地球元素生物地球化学循环的引擎, 对陆地生态系统碳(C)、氮(N)、磷(P)、硫(S)等养分循环起着关键作用(
... 据IPCC预测, 到21世纪末全球平均气温将上升1.5-4.8 ℃, 全球变暖已进入高速模式(
... 其次, 不同树种对微生物群落结构的影响存在差异(
1
2000
... 土壤总碳(TC)、总氮(TN)含量利用元素分析仪(MACRO cube, Elementar, Germany)测定; 土壤硝态氮(NO3--N)与铵态氮(NH4+-N)含量采用比色法测定(
1
2000
... 土壤总碳(TC)、总氮(TN)含量利用元素分析仪(MACRO cube, Elementar, Germany)测定; 土壤硝态氮(NO3--N)与铵态氮(NH4+-N)含量采用比色法测定(
Physiological, biochemical and molecular responses of the soil microbial community after afforestation of pastures with Pinus radiata
1
2009
... 首先, 植物通过多种机制直接或间接改变土壤微生物生物量和群落组成, 包括增加地上凋落物和地下根系分泌物等为土壤微生物生长提供所需的营养物质, 从而引起微生物生物量的快速增长(
Interactive effects of soil warming and fertilization on root production, mortality, and longevity in a Norway spruce stand in Northern Sweden
1
2004
... 前人研究发现, 在增温处理下, 根分泌、细根周转比较强烈, 同时与微生物代谢相关的酶的活性会提高, 从而促进微生物群落的生长(
Development of specific rhizosphere bacterial communities in relation to plant species, nutrition and soil type
1
2004
... 前人研究发现, 在增温处理下, 根分泌、细根周转比较强烈, 同时与微生物代谢相关的酶的活性会提高, 从而促进微生物群落的生长(
Diurnal changes of carbon dioxide flux from bare soil in agricultural field in Japan
1
2002
... 其次, 不同树种对微生物群落结构的影响存在差异(
Shifts in the microbial community structure explain the response of soil respiration to land-use change but not to climate warming
1
2015
... 前人研究发现, 在增温处理下, 根分泌、细根周转比较强烈, 同时与微生物代谢相关的酶的活性会提高, 从而促进微生物群落的生长(
Responses of bacterial communities to simulated climate changes in alpine meadow soil of the Qinghai-Tibet Plateau
2
2015
... 土壤微生物被认为是地球元素生物地球化学循环的引擎, 对陆地生态系统碳(C)、氮(N)、磷(P)、硫(S)等养分循环起着关键作用(
... 本研究还发现增温弱化了植物对微生物群落结构的影响, 与对照相比, 增温后有植物处理小区的微生物群落结构与裸地更相似.未增温条件下, 单种和混种F/B与GP/GN均发生了显著变化, 但与W-C比较, F/B和GP/GN仅在W-L小区显著减小, 而在W-Y和W-H小区保持稳定(
Experimental warming effects on the microbial community of a temperate mountain forest soil
1
2011
... 增温可以通过直接改变土壤理化特性而影响土壤微生物群落, 也可以通过影响植物根系的生长和生理特征等间接影响根际土壤微生物群落结构(
Effect of warming and drought on grassland microbial communities
2
2011
... 土壤微生物被认为是地球元素生物地球化学循环的引擎, 对陆地生态系统碳(C)、氮(N)、磷(P)、硫(S)等养分循环起着关键作用(
... 前人研究发现, 在增温处理下, 根分泌、细根周转比较强烈, 同时与微生物代谢相关的酶的活性会提高, 从而促进微生物群落的生长(
生态学报
1
2011
... 土壤微生物被认为是地球元素生物地球化学循环的引擎, 对陆地生态系统碳(C)、氮(N)、磷(P)、硫(S)等养分循环起着关键作用(
生态学报
1
2011
... 土壤微生物被认为是地球元素生物地球化学循环的引擎, 对陆地生态系统碳(C)、氮(N)、磷(P)、硫(S)等养分循环起着关键作用(
中国土壤微生物学研究10年回顾
2013
中国土壤微生物学研究10年回顾
2013
Effects of experimental warming on soil microbial communities in two contrasting subalpine forest ecosystems, eastern Tibetan Plateau, China
1
2016
... 西南亚高山针叶林位于青藏高原东部, 是全球变暖的敏感地带, 也是研究全球气候变化对森林生态系统影响的重要地域(
Arbuscular mycorrhizal fungi as mediators of ecosystem responses to nitrogen deposition: a trait-based predictive framework
1
2018
... 据IPCC预测, 到21世纪末全球平均气温将上升1.5-4.8 ℃, 全球变暖已进入高速模式(
Composition of fungal and bacterial communities in forest litter and soil is largely determined by dominant trees
1
2015
... 据IPCC预测, 到21世纪末全球平均气温将上升1.5-4.8 ℃, 全球变暖已进入高速模式(
Specific impacts of beech and Norway spruce on the structure and diversity of the rhizosphere and soil microbial communities
2
2016
... 其次, 不同树种对微生物群落结构的影响存在差异(
... ;
Fungal biomass development in a chronosequence of land abandonment
1
2006
... 首先, 植物通过多种机制直接或间接改变土壤微生物生物量和群落组成, 包括增加地上凋落物和地下根系分泌物等为土壤微生物生长提供所需的营养物质, 从而引起微生物生物量的快速增长(
Soil C:N ratio is the major determinant of soil microbial community structure in subtropical coniferous and broadleaf forest plantations
1
2015
... 其次, 不同树种对微生物群落结构的影响存在差异(
The effect of simulated warming on root dynamics and soil microbial community in an alpine meadow of the Qinghai-Tibet Plateau
1
2017
... 据IPCC预测, 到21世纪末全球平均气温将上升1.5-4.8 ℃, 全球变暖已进入高速模式(
Determination of the sedimentary microbial biomass by extractible lipid phosphate
1
1979
... 土壤总碳(TC)、总氮(TN)含量利用元素分析仪(MACRO cube, Elementar, Germany)测定; 土壤硝态氮(NO3--N)与铵态氮(NH4+-N)含量采用比色法测定(
Phospholipid fatty acid profiling of microbial communities—A review of interpretations and recent applications
1
2015
... 土壤总碳(TC)、总氮(TN)含量利用元素分析仪(MACRO cube, Elementar, Germany)测定; 土壤硝态氮(NO3--N)与铵态氮(NH4+-N)含量采用比色法测定(
Effects of warming on root diameter, distribution, and longevity in an alpine meadow
1
2014
... 增温可以通过直接改变土壤理化特性而影响土壤微生物群落, 也可以通过影响植物根系的生长和生理特征等间接影响根际土壤微生物群落结构(
Effects of experimental warming on soil N transformations of two coniferous species, Eastern Tibetan Plateau, China
2
2012
... 西南亚高山针叶林位于青藏高原东部, 是全球变暖的敏感地带, 也是研究全球气候变化对森林生态系统影响的重要地域(
... 人工挖掘4对2 m × 2 m × 50 cm (长×宽×深)的开阔小区, 利用客土法将实验地土壤全部换为该林区人工云杉林下过5 mm筛后混匀的表层土, 土壤pH为5.85, 土壤有机碳62.70 mg·g-1, 总氮3.66 mg·g-1, 总磷0.43 mg·g-1, 总钾7.92 mg·g-1.4对2 m × 2 m小区, 每对设置增温与对照处理, 处理之间间隔5 m, 每个2 m × 2 m的小区又分成4个1 m × 1 m亚小区.为避免亚小区内植物根系之间及周围环境的影响, 在亚小区四周埋入深50 cm的双层塑料膜进行隔离.2013年6月挑选长势、大小一致的3年生冷杉及2年生云杉幼苗, 在亚小区内按20 cm × 30 cm间距分别种植冷杉、云杉、云杉和冷杉以及裸地对照, 每个亚小区24株幼苗.2016年预实验发现云杉、冷杉根系已经比较发达, 当年开始对增温小区进行增温处理.增温方式采用悬挂于小区上方距地面1.8 m的165 cm × 15 cm红外辐射加热器(Kalglo Electronics, Bethlehem, USA)进行全天24 h连续增温, 实验期间空气温度和土壤温度分别增加2.0和3.7 ℃ (
Enhanced root exudation stimulates soil nitrogen transformations in a subalpine coniferous forest under experimental warming
2013
Warming effects on root morphological and physiological traits: the potential consequences on soil C dynamics as altered root exudation
2
2013
... 前人研究发现, 在增温处理下, 根分泌、细根周转比较强烈, 同时与微生物代谢相关的酶的活性会提高, 从而促进微生物群落的生长(
... ;
Nitrogen transformation in the rhizospheres of two subalpine coniferous species under experimental warming
2
2012
... 增温可以通过直接改变土壤理化特性而影响土壤微生物群落, 也可以通过影响植物根系的生长和生理特征等间接影响根际土壤微生物群落结构(
... 其次, 不同树种对微生物群落结构的影响存在差异(
海拔对辽东栎林地土壤土壤微生物群落的影响
2012
海拔对辽东栎林地土壤土壤微生物群落的影响
2012
Growth and physiological responses of Picea asperata seedlings to elevated temperature and to nitrogen fertilization
1
2008
... 其次, 不同树种对微生物群落结构的影响存在差异(
Effects of experimental warming and nitrogen fertilization on soil microbial communities and processes of two subalpine coniferous species in Eastern Tibetan Plateau, China
1
2014
... 据IPCC预测, 到21世纪末全球平均气温将上升1.5-4.8 ℃, 全球变暖已进入高速模式(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}