邢磊
1 , 2 , 段娜
3 , 李清河
, , 1 , 2 , * , 刘成功
1 , 2 , 李慧卿
1 , 2 , 孙高洁
1 , 2 1 中国林业科学研究院林业研究所, 北京 1000912 国家林业和草原局林木培育重点实验室, 北京 1000913 中国林业科学研究院沙漠林业实验中心, 内蒙古磴口 015200Variation in biomass allocation of Nitraria tangutorum during different phenological phases Lei XING
1 , 2 , Na DUAN
3 , Qing-He LI
, , 1 , 2 , * , Cheng-Gong LIU
1 , 2 , Hui-Qing LI
1 , 2 , Gao-Jie SUN
1 , 2 1 Research Institute of Forestry, Chinese Academy of Forestry, Beijing 100091, China 2 Key Laboratory of Tree Breeding and Cultivation, National Forestry and Grassland Administration, Beijing 100091, China 3 Experimental Center of Desert Forestry, Chinese Academy of Forestry, Dengkou, Nei Mongol 015200, China 通讯作者: *李清河: ORCID:0000-0001-8011-1322,
tsinghel@caf.ac.cn 编委: 邓建明
责任编辑: 李敏
收稿日期: 2019-06-26
接受日期: 2020-05-1
网络出版日期: 2020-07-20
基金资助: 国家自然科学基金 (31470622 )国家重点研发计划 (31470622 )中央级公益性科研院所基本科研业务费专项 (31470622 )
Corresponding authors: *ORCID:0000-0001-8011-1322,
tsinghel@caf.ac.cn Received: 2019-06-26
Accepted: 2020-05-1
Online: 2020-07-20
Fund supported: National Natural Science Foundation of China (31470622 )National Key R&D Program of China (31470622 )Fundamental Research Funds for the Central Nonprofit Research Institution of Chinese Academy of Forestry (31470622 )
摘要 植物生物量的分配模式是植物对环境适应的结果, 并伴随着植物生活史的每一个阶段, 与植物的生长和发育息息相关。目前关于植物生物量分配的大小依赖性已有相关报道, 但关于其对物候期的响应尚鲜有报道。该研究以乌兰布和沙漠地区白刺(
Nitraria tangutorum )为研究对象, 通过对其2016与2017年连续2个生长季里盛花期、盛果期与营养生长期3个物候期的根、压条、新枝、老枝、叶、繁殖器官等部分的生物量测定, 采用标准化主轴回归方程的斜率和截距的显著性比较, 分别探讨了白刺在不同物候期的异速生长的大小依赖程度和相对生物量分配比例, 特别是地上与地下部分之间、支持与同化器官之间, 在不同物候期的生物量分配规律。结果表明: 白刺的繁殖生长对生物量分配模式造成的影响主要体现在相对生物量分配比例(36.00%)而不是个体大小的依赖性程度上(16.67%), 且对新枝的影响较大, 使其不同物候期的大小依赖性程度发生了改变, 但是变化趋势不一致, 同时繁殖生长增加对叶片的相对生物量分配比例, 减少对老枝的相对生物量分配比例, 但并没有改变他们的大小依赖性程度。白刺生长过程中的地下部分生物量分配率随个体生物量的累积均增大, 而繁殖分配会在一定程度内减弱这种速率。白刺随着个体生物量的增大其生物量向支持器官分配率也越大, 但随着生长时间推移, 更倾向于将生物量分配给同化吸收器官。
关键词: 异速生长 ;
大小依赖性 ;
物候期 ;
生物量分配 ;
白刺 Abstract Aims The pattern of plant biomass allocation represents the plant response to the environment and is accompanied by every stage of plant life history. So it is closely related to the growth and development of plants. There have been reports on the size dependence of plant biomass allocation, but few studies have reported on its responses to different phenological phase.
Methods In this study,
Nitraria tangutorum in Ulan Buh Desert was used as the research object. The biomass of different organs (root, layering, newborn stem, older stem, leaf and reproductive organ) in the flowering, fruiting and vegetative growth stages in two consecutive years of 2016 and 2017 were measured. The significant differences of slope and intercept of the fitted equation with Standardized-major-axis were respectively used to discuss the varied size dependence and the biomass allocation in different phenological phase or ages, especially between the aboveground and belowground biomass and between the support and assimilation organs.
Important findings The results showed that the effect of reproductive allocation on biomass allocation pattern of
N. tangutorum is mainly reflected in the scale of relative biomass allocation (36.00%) rather than the extent of size dependence (16.67%). The reproductive growth has a greater impact on the biomass allocation pattern of newborn stems, which changes the size dependence extent among different phenological phases, but the trend is inconsistent. The reproductive growth increases the scale of relative biomass allocation to leaves and reduces that to older stems, but did not change their extent of size dependence. The biomass allocation rate in the belowground part of
N. tangutorum increased with the accumulation of its biomass, but the reproductive allocation slightly decreased this rate.
Nitraria tangutorum exhibited a higher rate of biomass allocation to supporting organs as plant biomass increases. Over time it is more likely for biomass to be allocated to assimilating organs.
Keywords: allometric growth ;
size dependence ;
phenological phase ;
biomass allocation ;
Nitraria tangutorum PDF (953KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 邢磊, 段娜, 李清河, 刘成功, 李慧卿, 孙高洁. 白刺不同物候期的生物量分配规律. 植物生态学报, 2020, 44(7): 763-771. DOI:
10.17521/cjpe.2019.0162 XING Lei, DUAN Na, LI Qing-He, LIU Cheng-Gong, LI Hui-Qing, SUN Gao-Jie.
Variation in biomass allocation of Nitraria tangutorum during different phenological phases .
Chinese Journal of Plant Ecology , 2020, 44(7): 763-771. DOI:
10.17521/cjpe.2019.0162 作为植物最基本的生态学特性和功能性状, 生物量累积是生态系统生产力的直观体现(
Klinkhamer et al ., 1992 )。植物生物量的分配模式是植物对环境适应的结果, 并伴随着植物生活史的每一个阶段, 与植物的生长和发育相互影响(
Bonser & Aarssen, 2009 ;
Poorter et al ., 2012 )。同时, 生物量分配模式实质上也反映出植物在各器官生物量分配中的权衡问题。关于植物生物量分配机制的探讨, 一种理论认为植物倾向于将生物量分配给能够获取稀缺资源的器官以最大化适应外界环境的变化(
Fraterrigo et al ., 2006 ;
Poorter et al ., 2012 ;
Reich et al ., 2014 ), 另一种理论认为生物量分配是由植物本身的生长模式可塑性主导的(
McCarthy & Enquist, 2007 )。但事实上, 它们并非对立而是相互补充的(
Niklas, 2006 )。但不可否认的是无论生物量分配模式是为了适应外界环境的变化还是由于生物量分配模式本身的可塑性所致, 每当生物量分配模式显示部分或全部发育的异速生长增长时, 生物量分配特征将显示个体发育漂变(即, 不管环境条件如何, 生物量分配模式将随植物生长和发育而变化), 只有当生物量积累在各个组成部分之间的函数关系表现为等距关系或线性时, 才可以忽略个体发育漂变(
Gedroc et al ., 1996 ;
McConnaughay & Coleman; 1999 ;
Xie et al ., 2012 )。但大多数研究基于集中在温带森林物种的数据, 而忽视了适应干旱条件的沙漠物种的数据。生活环境的差异也必然导致植物的生长规律发生适应性的变化(
Fu et al ., 2015 ), 而植物在不同物候期的生物量分配变化尚不明确。
近年来, 异速生长已成为研究生物量分配及权衡的重要方法。一些研究结果也表明植物的生物量分配规律可能符合异速分配理论(
Shipley & Meziane, 2002 ;
Peng & Yang, 2016 ), 表明不同器官生物量的分配比例是依赖于植物大小的。如以叶片为例, 假设其生物量分配是依赖于个体大小的, 那么随着其个体生物量的累积(
x ), 叶片的生物量累积(
y )形式已经明确, 并可以用函数ln
y =
c +
b ln
x 表示。而任何一个时期自然界的每一种植物都存在不同大小的个体, 因此, 可通过某一时期不同个体大小之间的拟合来将其异速生长方程表达出来(
Cheverud, 1982 ), 当然对于某些植物在不同年份之间的差异可归因于年龄大小造成的分配模式差异。同时, 植物在长期进化的过程中, 不同的物候期形成了一种响应环境季节性变化的生长发育阶段(如抽芽、开花、结果、落叶、休眠等), 这种生长阶段容易受到环境因子的影响(
Menzel et al ., 2006 ;
Wolkovich et al ., 2012 ), 但却是其本身生长模式对环境的一种适应。大量的研究也表明植物的不同生长发育期之间, 特别是营养生长期与繁殖生长期之间的生物量分配有着显著的差异(
Bonser & Aarssen, 2009 ;
Reekie & Bazzaz, 2011 ;
李清河等; 2012 )。
由此可见, 植物的生物量分配在依赖于个体大小的同时不可或缺的会受到其自身物候期的影响, 此时的异速生长方程是否会发生改变或将如何改变则需要进一步的研究数据来论证。但是当前对于大小依赖性的研究很少区分不同物候期之间的差异, 而且大多仅以营养生长期的数据支持, 鲜有涉及繁殖分配对其造成的影响, 同时当前的研究对象较少针对木本植物。因此, 若想更好地了解和预测植物的生长模式就有必要结合异速生长理论针对多年生木本植物生物量分配的个体大小依赖性在不同物候期的差异来开展大量试验研究。
白刺(
Nitraria tangutorum )是我国干旱荒漠地区的重要建群植物种, 在其一年的生长发育过程中营养生长和有性生殖生长均具有明显的时期且指标可观测, 同时它还是一种克隆植物, 其通过压条克隆繁殖形成的沙包是沙区的独特景观, 对荒漠地区的植被生态系统稳定性具有重要作用。同时鉴于木本植物生长特点, 当年的植物生物量与上一年的植物能量储备息息相关。因此, 本文通过连续两个生长季不同物候期的白刺各器官的异速生长函数拟合, 探讨了其生物量分配变化规律, 以期为理解干旱地区木本植物生存策略提供依据。
1 材料和方法 1.1 研究区概况 试验地点位于乌兰布和沙漠东北缘, 行政区划隶属于内蒙古巴彦淖尔市磴口县(40.40° N, 106.72° E, 海拔1 050 m), 是我国西北地区荒漠与半荒漠的过渡地带。该地区气候属于中温带荒漠气候, 夏季炎热, 年平均气温7.4 ℃; 日照充足, 年日照时间3 300 h以上; 多风少雨, 年平均风速4 m·s
-1 , 年降水量149.0 mm, 且降水多集中在7-9月, 年蒸发量2 351.9 mm。地貌以流动沙丘、半固定沙丘和平缓沙地为主。土壤类型为风沙土和灰棕漠土。试验地主要是以白刺为建群种的植被生态系统, 其他植物种还有油蒿(
Artemisia ordosica )、沙鞭(
Psammochloa villosa )等。
1.2 样方布设 2015年4月初, 在白刺沙包聚集地, 选取生长健壮的沙包沿其边缘处设置1 m × 1 m样方: 样方四角用铁丝固定标记, 用修枝剪将样方内植株的压条与外界断开, 并松开样方边缘及其样方内每株分株基部的土壤, 根据压条上的分株与根系分布状态将萌生分株的压条均匀剪断, 使每株分株成独立植株, 然后将土壤回覆, 确保每个样方内均包含3-6株白刺植株。试验在3个白刺沙包分布片区, 共计设置594个样方。不同白刺沙包之间的遗传差异性确保了试验植株的多样性。样方内各独立分株经过1年的生长, 消除克隆植物的生理整合影响后, 于次年进行取样调查。
1.3 样品采集 2016与2017年的5月(盛花期)、7月(盛果期)、9月(营养生长期), 分别在样地随机选取30-60个样方, 采用全挖法获取样方内全部存活植株, 并用土壤筛筛取土壤中遗留根系, 初步去除沙土后, 用信封盛装带回实验室。洗净表土后将整株(T)区分为根(R)、压条(La)、新枝(S)、老枝(B)、叶(Le)、繁殖器官(F)等部分, 并置于75 ℃烘箱烘干至恒质量(24 h)后测定干质量。其中F在盛花期为花(Fl), 在盛果期为果实(Fr), 在营养生长期则没有。各时期样本数据统计结果如
表1 所示。
Table 1 表1 表1 白刺各时期样本量及各器官平均生物量
Table 1
Sample size and average biomass of each organ in six stages of Nitraria tangutorum 时期 样本数N R Le S B La F T M SD M SD M SD M SD M SD M SD M SD 2016 5月 May 45 3.726 3.026 4.138 3.256 0.686 0.574 8.847 6.28 6.274 4.691 0.575 0.948 24.248 15.255 7月 July 213 4.351 4.232 6.182 6.549 2.498 3.18 12.506 9.714 10.284 9.866 1.593 2.018 37.415 26.952 9月 Sept. 47 2.217 1.746 8.877 9.514 8.618 10.456 17.173 16.67 8.670 6.188 - - 50.640 44.827 2017 5月 May 112 7.417 5.936 7.738 8.726 2.309 3.237 22.260 27.997 25.045 26.766 1.048 1.370 65.817 65.777 7月 July 100 10.334 9.637 12.198 10.636 7.098 6.978 23.631 20.556 37.957 24.793 3.078 6.407 94.296 70.004 9月 Sept. 95 10.008 6.787 7.316 9.089 9.963 14.905 35.305 26.269 33.716 22.517 - - 96.308 80.532
B, 老枝; F, 繁殖器官; La, 压条; Le, 叶片; R, 根; S, 新枝; T, 整株。M , 平均生物量(g); SD, 标准偏差。 B, older stem; F, reproductive organ; La, layering; Le, leaf; R, root; S, newborn stem; T, whole plant. M, mean biomass (g); SD, standard deviation. 新窗口打开 |
下载CSV 1.4 数据处理 各器官以及整株生物量之间的异速生长关系采用常用对数(lg)转换后的值, 由R 3.5.2软件中的smatr包中的SMA函数进行标准化主轴回归(SMA)分析。该函数可根据需要对多组数据回归方程的斜率与截距进行似然比(
LR )检验与多重比较。对于不同时期SMA回归方程的斜率以及截距的显著性比较解释参照
Xie等(2012) : 若某一器官的异速生长SMA的斜率在不同时期之间具有显著性差异, 则认为其大小依赖性程度在不同时期产生了漂变, 即该器官在不同生长时期有着不同的生长轨迹。若其斜率之间不具有显著性差异, 则进一步对其截距进行差异显著性分析。若截距之间差异显著, 则认为不同时期其生物量分配的大小依赖性的变化率不变, 但相对生物量分配比例产生了变化(截距变大则相对增加了该器官的生物量分配比例, 反之, 截距变小则相对降低了该器官的生物量分配比例); 若截距之间的差异也不显著, 则认为不同时期生物量分配规律无显著变化。讨论地上部分和地下部分、同化器官和支持器官的异速生长关系时仅对比斜率之间的差异, 在此, 斜率可以表征生物量分配率, 说明在植物生物量累积的过程中, 某一器官生物量占比的变化速率。图表制作在Excel 2012中完成。
2 结果和分析 2.1 白刺各器官与整株生物量SMA回归的异速生长关系 2016和2017年白刺不同物候期各器官生物量与整株生物量之间的SMA回归方程如
表2 所示。不同物候期各器官与整株间SMA回归方程的斜率在3个物候期之间的似然比检验结果(
表3 )显示, 在2016和2017年的3个物候期之间, 白刺的Le和B的生物量分配在2年的不同物候期之间均无显著差异, 而S和F的生物量分配的大小依赖性程度均发生了改变, 表明物候期对白刺生物量分配模式产生了显著影响。去掉繁殖器官之后进一步对斜率间差异显著的器官生物量分配进行多重比较(
表4 ), 结果表明, 有5组的异速生长指数达到显著差异水平(
p < 0.05), 占全部器官(除F外)回归方程组数的16.67%。而斜率差异不显著的25组中有9组的截距产生了显著差异(
p < 0.05), 占36.00% (
表5 )。表明白刺在不同物候期之间的繁殖分配对生物量分配模式造成的影响主要体现在相对生物量分配比例而不是个体大小的依赖性程度上, 且对S影响较大, 其在盛果期的SMA回归斜率要么增大要么降低, 表明繁殖生长对S生物量分配, 特别是盛果期的S生物量分配大小依赖性程度影响明显, 但变化趋势不一致。依据Le和B生物量分配的截距显著性来看, 繁殖生长增加对Le的相对生物量分配比例, 减少对B的相对生物量分配比例, 但并没有改变他们的大小依赖性程度。
Table 2 表2 表2 2016与2017年白刺不同物候期各器官与整株生物量的标准化主轴回归方程
Table 2
Standardized-major-axis results of biomass between organs and whole plant of Nitraria tangutorum of different phenological periods in 2016 and 2017 器官 时期 2016 2017 截距及置信区间CI ) 斜率及置信区间CI ) r 2 p 截距及置信区间CI ) 斜率及置信区间CI ) r 2 p R/T 5月 May -1.226 5 -1.476 6 -0.976 3 1.280 0 1.106 9 1.480 2 0.775 2 0.000 0 -0.601 6 -0.791 4 -0.411 8 0.823 8 0.719 7 0.942 8 0.486 4 0.000 0 7月 July -1.576 5 -1.770 5 -1.382 4 1.379 3 1.256 6 1.513 8 0.528 1 0.000 0 -1.080 8 -1.350 7 -0.810 8 1.052 9 0.919 6 1.205 5 0.541 4 0.000 0 9月 Sept. -1.213 2 -1.461 2 -0.965 3 1.200 2 1.055 7 1.364 5 0.816 5 0.000 0 -0.719 0 -0.953 0 -0.485 1 0.875 3 0.760 3 1.007 7 0.529 0 0.000 0 Le/T 5月 May -1.313 1 -1.613 4 -1.012 7 1.373 4 1.167 5 1.615 6 0.718 7 0.000 0 -1.520 7 -1.699 2 -1.342 3 1.303 6 1.202 6 1.413 1 0.817 5 0.000 0 7月 July -1.359 6 -1.514 9 -1.204 2 1.335 1 1.236 0 1.442 2 0.676 4 0.000 0 -1.624 6 -1.826 1 -1.423 1 1.354 2 1.251 6 1.465 3 0.845 0 0.000 0 9月 Sept. -1.171 0 -1.394 9 -0.947 1 1.222 4 1.090 8 1.369 7 0.855 6 0.000 0 -1.684 3 -1.969 7 -1.398 9 1.253 3 1.111 3 1.413 3 0.657 7 0.000 0 S/T 5月 May -2.529 4 -3.053 9 -2.005 0 1.680 9 1.335 3 2.116 1 0.429 8 0.000 0 -2.762 3 -3.048 4 -2.476 2 1.655 0 1.495 0 1.832 2 0.709 5 0.000 0 7月 July -2.424 0 -2.666 1 -2.181 8 1.720 7 1.567 7 1.888 6 0.527 9 0.000 0 -2.016 4 -2.279 6 -1.753 1 1.421 7 1.289 0 1.568 1 0.760 1 0.000 0 9月 Sept. -1.406 8 -1.737 6 -1.076 1 1.336 0 1.145 9 1.557 5 0.736 7 0.000 0 -2.929 8 -3.352 1 -2.507 6 1.902 3 1.692 0 2.138 8 0.674 7 0.000 0 B/T 5月 May -0.829 4 -1.114 9 -0.543 8 1.272 9 1.077 6 1.503 6 0.704 0 0.000 0 -0.970 3 -1.157 9 -0.782 7 1.254 0 1.148 3 1.369 5 0.782 2 0.000 0 7月 July -0.841 9 -1.017 0 -0.666 7 1.228 4 1.118 3 1.349 4 0.531 1 0.000 0 -0.934 7 -1.176 9 -0.692 4 1.157 7 1.036 4 1.293 2 0.693 9 0.000 0 9月 Sept. -0.820 6 -1.060 8 -0.580 5 1.198 1 1.057 9 1.356 9 0.827 2 0.000 0 -0.797 0 -1.006 5 -0.587 5 1.167 1 1.061 4 1.283 3 0.787 0 0.000 0 La/T 5月 May -0.949 8 -1.355 6 -0.544 0 1.251 7 0.985 7 1.589 5 0.385 0 0.000 0 -0.681 4 -0.817 8 -0.545 0 1.137 8 1.060 2 1.221 1 0.859 9 0.000 0 7月 July -1.071 1 -1.260 4 -0.881 8 1.305 1 1.185 7 1.436 5 0.498 7 0.000 0 -0.522 1 -0.688 6 -0.355 6 1.061 3 0.976 6 1.153 3 0.827 7 0.000 0 9月 Sept. -0.602 8 -0.955 4 -0.250 2 0.915 6 0.722 7 1.160 0 0.367 4 0.000 0 -0.614 8 -0.811 5 -0.418 1 1.081 9 0.982 7 1.191 0 0.781 5 0.000 0 F/T 5月 May -4.740 7 -5.773 3 -3.708 0 2.994 2 2.323 4 3.858 7 0.305 1 0.000 1 -2.925 5 -3.413 8 -2.437 2 1.608 8 1.354 5 1.910 8 0.333 8 0.000 0 7月 July -2.630 6 -2.949 3 -2.311 8 1.749 9 1.558 1 1.965 3 0.381 2 0.000 0 -4.587 9 -5.612 5 -3.563 2 2.455 7 2.008 1 3.003 1 0.470 1 0.000 0 9月 Sept. - - - - - - - - - - - - - - - -
B, 老枝; F, 繁殖器官; La, 压条; Le, 叶片; R, 根; S, 新枝; T, 整株。 B, older stem; F, reproductive organ; La, layering; Le, leaf; R, root; S, newborn stem; T, whole plant. 新窗口打开 |
下载CSV Table 3 表3 表3 白刺不同物候期各器官与整株标准化主轴回归斜率的似然比检验
Table 3
Likelihood ratio test of the slope of Standardized-major-axis of biomass between organs and whole plant of Nitraria tangutorum in three phenological phases 器官 2016 2017 df LR p df LR p R/T 2 3.104 0.211 8 2 6.844 0.032 7 S/T 2 7.665 0.021 7 2 14.02 0.000 9 B/T 2 0.339 6 0.843 8 2 1.716 0.424 1 Le/T 2 2.026 0.363 1 2 1.211 0.545 8 La/T 2 7.311 0.025 8 2 1.73 0.421 0 F/T 1 13.7 0.000 2 1 9.737 0.001 8
LR , 似然比。B, 老枝; F, 繁殖器官; La, 压条; Le, 叶片; R, 根; S, 新枝; T, 整株。LR , likelihood ratio. B, older stem; F, reproductive organ; La, layering; Le, leaf; R, root; S, newborn stem; T, whole plant.新窗口打开 |
下载CSV Table 4 表4 表4 白刺不同物候期之间部分器官与整株标准化主轴回归斜率的多重比较
Table 4
Multiple comparisons of slopes of Standardized-major-axis of biomass between partial organs and whole plant of Nitraria tangutorum in three phenological phases 2016 2017 器官 Organ 时期 Time df LR p 器官 Organ 时期 Time df LR p S/T 5月、7月 May vs. July 1 0.098 1 0.851 9 R/T 5月、7月 May vs. July 1 6.343 7 0.011 8 5月、9月 May vs. Sept. 1 1.366 5 0.100 5 5月、9月 May vs. Sept. 1 0.375 7 0.539 9 7月、9月 July vs. Sept. 1 1.616 4 0.006 0 7月、9月 July vs. Sept. 1 3.462 5 0.062 8 La/T 5月、7月 May vs. July 1 0.103 6 0.747 6 S/T 5月、7月 May vs. July 1 4.475 5 0.034 4 5月、9月 May vs. Sept. 1 3.370 0 0.066 4 5月、9月 May vs. Sept. 1 3.122 1 0.077 2 7月、9月 July vs. Sept. 1 7.309 6 0.006 9 7月、9月 July vs. Sept. 1 13.763 3 0.000 2
La, 压条; R, 根; S, 新枝; T, 整株。LR , 似然比。 La, layering; R, root; S, newborn stem; T, whole plant. LR , likelihood ratio. 新窗口打开 |
下载CSV Table 5 表5 表5 白刺不同物候期之间部分器官与整株标准化主轴回归截距的似然比检验
Table 5
Likelihood ratio test of the intercept of the Standardized-major-axis of biomass between organs and whole plant of Nitraria tangutorum in three phenological phases 器官 时期 2016 2017 df LR p df LR p R/T 5月、7月 May vs. July 1 0.099 3 0.752 7 1 - - 5月、9月 May vs. Sept. 1 3.502 9 0.061 3 1 1.326 6 0.249 4 7月、9月 July vs. Sept. 1 7.672 1 0.005 6 1 0.056 8 0.811 7 S/T 5月、7月 May vs. July 1 0.821 5 0.364 8 1 - - 5月、9月 May vs. Sept. 1 3.533 1 0.060 2 1 3.162 5 0.075 3 7月、9月 July vs. Sept. 1 - - 1 - - B/T 5月、7月 May vs. July 1 0.385 0 0.535 0 1 2.435 1 0.118 6 5月、9月 May vs. Sept. 1 1.128 9 0.288 0 1 5.574 6 0.018 2 7月、9月 July vs. Sept. 1 0.511 9 0.474 3 1 19.126 2 0.000 0 Le/T 5月、7月 May vs. July 1 1.025 5 0.311 2 1 4.046 0 0.044 3 5月、9月 May vs. Sept. 1 4.351 1 0.037 0 1 6.291 6 0.012 1 7月、9月 July vs. Sept. 1 3.208 5 0.073 3 1 22.295 6 0.000 0 La/T 5月、7月 May vs. July 1 2.631 8 0.104 7 1 6.833 4 0.008 9 5月、9月 May vs. Sept. 1 0.196 0 0.658 0 1 0.580 5 0.446 1 7月、9月 July vs. Sept. 1 - - 1 4.070 1 0.043 6
B, 老枝; La, 压条; Le, 叶片; R, 根; S, 新枝; T, 整株。LR , 似然比。 B, older stem; La, layering; Le, leaf; R, root; S, newborn stem; T, whole plant. LR , likelihood ratio. 新窗口打开 |
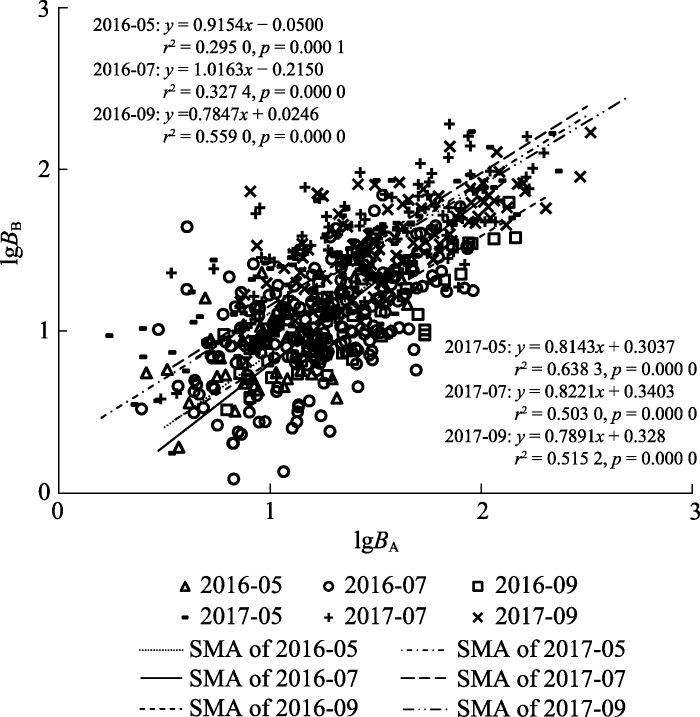
下载CSV 2.2 白刺地上部分与地下部分生物量的异速生长关系 各物候期白刺地上部分与地下部分生物量之间的异速生长关系如
图1 所示, 由图可知, 其斜率在连续两个生长季3个物候期均呈先增高后降低的趋势, 按照时间顺序依次为: 0.915 4、1.016 3、0.784 7、0.814 3、0.822 1、0.789 1。斜率间的两两似然比检验结果表明, 2016年盛果期(07-16)除与前一时期(05-16)差异不显著(
p = 0.552)外, 与09-16、05-17、07-17和09-17之间均差异显著(
p < 0.05), 而其他5个时期之间并无显著差异(
p > 0.05)。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1白刺不同物候期地上部分与地下部分生物量的异速生长关系。 B A , 地上生物量;
B B , 地下生物量。SMA, 标准化主轴回归。
Fig. 1Allometric relationship between aboveground and belowground biomass in different phenological phases of Nitraria tangutorum. B A , aboveground biomass;
B B , belowground biomass. SMA, standardized major axis regression.
2.3 白刺支持器官与同化器官生物量之间的异速生长关系 各物候期白刺支持器官(S、B、La)与同化器官(Le、R)生物量之间的异速生长关系如
图2 所示, 由图可知, 随着时间的推移, 其斜率除2017年盛花期到盛果期有回升趋势外, 其余均呈逐渐降低的趋势,按照时间顺序依次为: 1.316 7、1.239 9、1.190 1、0.847 8、1.095 6、0.897 7。似然比检验结果表明, 其斜率在2016年盛花期与营养生长期之间差异显著(
p < 0.05), 2017年3个物候期之间的差异均不显著(
p > 0.05), 而在连续两个生长季各物候期的对比中, 差异均达到显著水平(
p < 0.05)。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2白刺不同物候期支持器官与同化器官生物量的异速生长关系。 B AO , 同化器官生物量;
B SO , 支持器官生物量。SMA, 标准化主轴回归。
Fig. 2Allometric relationship between assimilating and supporting organs biomass in different phenological phases of Nitraria tangutorum. B AO , assimilating organs biomass;
B SO , supporting organs biomass. SMA, standardized major axis regression.
3 讨论和结论 3.1 依赖于个体大小的生物量分配对物候期的响应 异速生长的提出, 为我们研究依赖于个体大小的生物量分配方式提供了方法, 其能够揭示生物在生长过程中的变化规律(
Shipley & Meziane, 2002 ;
Komiyama et al ., 2008 ;
Peng & Yang, 2016 )。本文采用SMA分别拟合了白刺2016与2017年在盛花期、盛果期和营养生长期3个物候期不同器官的异速生长关系。结果显示, 白刺各器官的个体大小依赖性在各时期均极显著, 且相关性较高, 这进一步验证了
范高华等(2017) 的观点, 即植物生育期的变化, 对繁殖生物量与个体大小的关系具有显著影响。
对于各器官不同物候期间的差异显著性检验结果表明, 物候期对白刺生物量分配模式具有一定的影响, 且对不同器官分配模式影响存在差异, 这可能与不同器官的功能有关。有些研究也指出植物生物量分配模式在季节之间的变化似乎与其季节性光合作用和水的需求的变化同时发生(
Barnes, 2002 )。由于本文所研究的3个物候期为盛花期、盛果期与营养生长期, 它们之间的差异实质上反映的是繁殖分配和营养分配之间的差异。繁殖分配作为植物生命周期中的一个重要特征, 是植物进行资源分配的一个核心问题(
Tian et al ., 2012 ), 不同的繁殖分配反映了植物在自然选择过程中的生存策略(
Reekie & Bazzaz, 2011 ), 但根据本文的研究结果可知, 繁殖生长对生物量分配模式造成的影响主要体现在相对生物量分配比例(36.00%)而不是个体大小的依赖性程度上(16.67%), 而且对S的影响较大, 但其变化趋势不一致。繁殖生长增加对Le的相对生物量分配比例, 减少对B的相对生物量分配比例, 但并没有改变他们的大小依赖性程度, 进一步证实了植物会通过调节不同器官中的生物量分配, 来实现和维持其生长最大化(
郭丽珠等, 2019 )。同时, 植物大小也是影响生殖分配的重要因素(
Niklas & Enquist, 2002 ;
Cheplick, 2005 ;
Weiner et al ., 2009 )。随着植物的生长, 两种繁殖方式的有效性也会随之发生改变, 一般而言有性繁殖对植物大小有一个阈值要求, 对于克隆植物而言, 在达到阈值之前有性繁殖后代若不能存活下来, 此时克隆繁殖产生的新分株可能成为其唯一的后代并存留下来, 但一旦达到阈值, 伴随着克隆繁殖分配的减小, 有性繁殖分配将会呈现增加的趋势(
Aarssen, 2008 )。
3.2 白刺地上与地下部分、同化与吸收器官在生长过程中的生物量分配 在生物量分配的研究中, 地上部分与地下部分生物量之间的关系因可作为评估植物生长响应生态环境的重要指标而被广泛研究(
Niklas, 2005 )。本文通过比较白刺2016与2017年地上部分(
x 轴)与地下部分(
y 轴)生物量的SMA回归斜率发现, 白刺在生物量的累积过程中会优先将生物量分配给地下部分, 且其异速生长指数随着时间的推移具有减小趋势, 表明随着植株的生长, 白刺将生物量分配给地下部分的倾向更加明显, 这与
Bond (2000) 和
Niinemets (2004) 等的研究结果相似, 这些现象进一步说明了植物地上部分与地下部分生物量之间的异速生长指数并不总是符合等速理论的观点(
Chen et al ., 2019 )。同时, 在2016与2017年盛果期的SMA回归斜率均有所提升, 表明繁殖分配, 特别是盛果期会在一定程度上减弱这种倾向, 并将部分生物量分配给地上部分的繁殖器官, 以确保自身物候周期的完整性。
Husáková等(2018) 基于20种旱地植物的研究结果表明, 多年生植物随着生长的进行会分配更多的生物量到地上部分, 本文的研究结果与之不同, 这可能与植物自身的生活史有关, 生活史间的差异导致植物生物量分配模式的差异, 进而避免了干旱等环境因子对生长造成的影响(
Chen et al ., 2019 )。研究认为在干旱环境下, 植物会增加根系的生物量分配以获取更多的水资源(
Müller et al ., 2000 ;
Poorter et al ., 2012 ),
Eziz等(2017) 基于对164篇关于干旱胁迫与生物量分配模式文献的分析, 认为干旱胁迫会显著增加根的生物量分配而减少茎和叶的。同时, 当受到养分限制时, 植物同样会增加根系的生物量分配以更大范围地吸收养分(
Shipley & Meziane, 2002 ;
Poorter et al ., 2012 )。
对白刺连续两个生长季的地上同化器官(
x 轴)与支持器官(
y 轴)生物量的SMA回归斜率比较发现, 在2016年其斜率均大于1, 表明随着个体生物量的增大其生物量向支持器官分配的比率也越大, 但随着时间的推移, 斜率整体上呈下降趋势, 且在2017年的盛花期与营养生长期均低于1, 表明其支持器官的生物量相对于同化器官生物量的变化速率在不同年份之间存在差异, 本文中白刺植株随着生长时间的推移, 会更倾向于将生物量分配给同化吸收器官而不是支持器官。
Canham等(1996) 认为, 植物对环境因子的可塑性响应是为了在资源有限的情况下能够最大限度地增长和生存。因此, 如果受到土壤水分限制, 植物必须生成更多的根系并减少叶面积, 以保持植物体内水分吸收和消耗平衡(
Deng et al. , 2006 ;
Xu et al ., 2007 ), 但是茎秆等支持器官对大部分陆生植物而言, 可能会因为其对资源获取作用不大而在资源限制下无明显响应(
Poorter & Nagel, 2000 ;
Coleman et al ., 2004 ), 本文的研究结果与之相似。
本文对植物的生物量分配在不同器官之间, 以及不同物候期之间生物量分配的异速生长关系进行了探讨, 进一步明确了异速生长理论中生物量分配的个体大小依赖性在不同物候期的差异性。对于荒漠植物白刺而言, 尽管其生物量分配的个体大小依赖性在两个生长季间存在差异, 且随着个体生物量的累积, 地下部分和支持器官的生物量分配率逐渐增大, 但是相对于个体大小依赖性程度, 白刺的繁殖生长更加对相对生物量分配比例产生影响。 繁殖生长使新枝生物量分配的大小依赖性程度发生变化, 也对叶片和老枝的相对生物量分配比例分别产生增加和降低的影响, 而且在一定程度内减缓了地下部分生物量分配率随个体生物量累积的增大速率。研究结果有助于更好地了解和预测植物的生长模式及生活史策略。但是其中对于生物量分配中的个体发育漂变的规律还需要在时间和空间跨度上进行更深一步的研究, 且有待分析其个体发育漂变的机理, 以进一步拓展植物在面对不同生活环境(光、温、水、肥等)时的生物量分配规律。
[1] Aarssen LW 2008 ). Death without sex—The ‘problem of the small’ and selection for reproductive economy in flowering plantsEvolutionary Ecology 279 -298 . DOI:
10.1007/s10682-007-9170-z URL [本文引用: 1] Most of the resident plants within vegetation fail to leave descendants because of death without sex—i.e. sexual reproduction fails (zero fecundity), primarily because of relatively small plant size. I propose that this ‘problem of the small’ represents one of the principal driving forces of evolution by natural selection, and that the main product of this selection is ‘reproductive economy’, manifested by several plant traits that are widely distributed among angiosperms: sexual maturity at a relatively young age and small size, relatively small seed size, selfing (including through mixed mating), and of particular interest here, clonality. In non-clonal species, an offspring develops from a zygote into a single ‘rooted unit’, i.e. a distinct vascular transition point between live shoot and root tissue. Clonal species can produce an indeterminate number of these rooted unit offspring asexually, all as products of a single zygote. Clonality is a common strategy in angiosperms because it confers a potential two-fold fitness benefit—especially in relatively small species—by promoting longevity of the zygote product, while at the same time providing a fecundity supplement (through asexual multiplication of rooted units), thereby allowing offspring production economically, i.e. without requiring large adult size, and without even requiring the fertilization of ovules. The primary fitness benefit from clonality, therefore, is that the somatic product of a zygote can effectively avoid an intrinsic limitation predicted for all non-clonal plants: the trade-off between longevity and the potential rate of offspring/descendant production. These major fitness benefits of clonality are explored in considering why clonality is less common in larger species, why the largest species (trees) generally do not have the longest-lived zygote product, and in re-assessing traditional and recent views concerning the loss of sex in clonal plants, the predicted trade-off between the size and number of clonal offspring, and the predicted trade-off between sexual and asexual reproduction.]]>
[2] Barnes AD 2002 ). Effects of phenology, water availability and seed source on loblolly pine biomass partitioning and transpirationTree Physiology 733 -740 . [本文引用: 1] [3] Bond BJ 2000 ). Age-related changes in photosynthesis of woody plantsTrends in Plant Science 349 -353 . URL PMID:
10908880 [本文引用: 1] [4] Bonser SP Aarssen LW 2009 ). Interpreting reproductive allometry: individual strategies of allocation explain size- dependent reproduction in plant populationsPerspectives in Plant Ecology, Evolution and Systematics 31 -40 . [本文引用: 2] [5] Canham CD Berkowitz AR Kelly VR Lovett GM Ollinger SV Schnurr J 1996 ). Biomass allocation and multiple resource limitation in tree seedlingsCanadian Journal of Forest Research 1521 -1530 . [本文引用: 1] [6] Chen RF Ran JZ Huang H Dong LW Sun Y Ji MF Hu WG Yao SR Lu JL Gong HY Xie SB Du QJ Hou QQ Niklas KJ Deng JM 2019 ). Life history strategies drive size-dependent biomass allocation patterns of dryland ephemerals and shrubsEcosphere e02709 . DOI: 10.1002/ecs2.2709 . [本文引用: 2] [7] Cheplick GP 2005 ). Biomass partitioning and reproductive allocation in the invasive, cleistogamous grass Microstegium vimineum : influence of the light environmentJournal of the Torrey Botanical Society 214 -224 . [本文引用: 1] [8] Cheverud JM 1982 ). Relationships among ontogenetic, static, and evolutionary allometryAmerican Journal of Physical Anthropology 139 -149 . URL PMID:
7149015 [本文引用: 1] [9] Coleman MD Friend AL Kern CC 2004 ). Carbon allocation and nitrogen acquisition in a developing Populus deltoides plantationTree Physiology 1347 -1357 . URL PMID:
15465697 [本文引用: 1] [10] Deng JM Wang GX Morris EC Wei XP Li DX Chen BM Zhao CM Liu J Wang Y 2006 ). Plant mass-density relationship along a moisture gradient in north-west ChinaJournal of Ecology 953 -958 . [本文引用: 1] [11] Eziz A Yan ZB Tian D Han WX Tang ZY Fang JY 2017 ). Drought effect on plant biomass allocation: a meta- analysisEcology and Evolution 11002 -11010 . [本文引用: 1] [12] Fan GH Huang YX Zhao XY Shen XJ 2017 ). Effect of population density on the allometric growth of Agriophyllum squarrosum Acta Prataculturae Sinica 53 -64 . [本文引用: 1] [ 范高华 , 黄迎新 , 赵学勇 , 神祥金 (2017 ). 种群密度对沙米异速生长的影响草业学报 , 26, 53 -64 .] [本文引用: 1] [13] Fraterrigo JM Turner MG Pearson SM 2006 ). Previous land use alters plant allocation and growth in forest herbsJournal of Ecology 548 -557 . [本文引用: 1] [14] Fu YH Zhao HF Piao SL Peaucelle M Peng SS Zhou GZ Ciais P Huang MT Menzel A Pe?uelas J Song Y Vitasse Y Zeng ZZ Janssens IA 2015 ). Declining global warming effects on the phenology of spring leaf unfoldingNature 104 -107 . URL PMID:
26416746 [本文引用: 1] [15] Gedroc JJ McConnaughay KDM Coleman JS 1996 ). Plasticity in root/shoot partitioning: optimal, ontogenetic, or both?Functional Ecology 44 -50 . [本文引用: 1] [16] Guo LZ Huang D Zhang C Li JH Zhao H Wang K 2019 ). Analysis of biomass allocation and allometric growth of Stellera chamaejasme in degraded typical steppeChinese Journal of Grassland 53 -59 . [本文引用: 1] [ 郭丽珠 , 黄顶 , 张丛 , 李佳欢 , 赵欢 , 王堃 (2019 ). 退化典型草原狼毒生物量分配及异速生长分析中国草地学报 , 41, 53 -59 .] [本文引用: 1] [17] Husáková I Weiner J Münzbergová Z 2018 ). Species traits and shoot-root biomass allocation in 20 dry-grassland speciesJournal of Plant Ecology 273 -285 . [本文引用: 1] [18] Klinkhamer PGL Meelis E de Jong TJ Weiner J 1992 ). On the analysis of size-dependent reproductive output in plantsFunctional Ecology 308 -316 . [本文引用: 1] [19] Komiyama A Ong JE Poungparn S 2008 ). Allometry, biomass, and productivity of mangrove forests: a reviewAquatic Botany 128 -137 . [本文引用: 1] [20] Li QH Xin ZM Gao TT Wang SX Xu J Sun F 2012 ). Reproductive allocation in four desert species of the genus Nitraria LActa Ecologica Sinica 5054 -5061 . [本文引用: 1] [ 李清河 , 辛智鸣 , 高婷婷 , 王赛宵 , 徐军 , 孙非 (2012 ). 荒漠植物白刺属4个物种的生殖分配比较生态学报 , 32, 5054 -5061 .] [本文引用: 1] [21] Menzel A Sparks TH Estrella N Koch E Aasa A Ahas R Alm-Kübler K Bissolli P Braslavská O Briede A Chmielewski FM Crepinsek Z Curnel Y Dahl ? Defila C Donnelly A Filella Y Jatczak K M?ge F Mestre A Nordli ? Pe?uelas J Pirinen P Remi?ová V Scheifinger H Striz M Susnik A van Vliet AJH Wielgolaski FE Zach S Zust A 2006 ). European phenological response to climate change matches the warming patternGlobal Change Biology 1969 -1976 . DOI:
10.1111/gcb.2006.12.issue-10 URL [本文引用: 1] [22] McCarthy MC Enquist BJ 2007 ). Consistency between an allometric approach and optimal partitioning theory in global patterns of plant biomass allocationFunctional Ecology 713 -720 . [本文引用: 1] [23] McConnaughay KDM Coleman JS 1999 ). Biomass allocation in plants: ontogeny or optimality? A test along three resource gradientsEcology 2581 -2593 . [本文引用: 1] [24] Müller I Schmid B Weiner J 2000 ). The effect of nutrient availability on biomass allocation patterns in 27 species of herbaceous plantsPerspectives in Plant Ecology , Evolution and Systematics 115 -127 . [本文引用: 1] [25] Niinemets U 2004 ). Adaptive adjustments to light in foliage and whole-plant characteristics depend on relative age in the perennial herb Leontodon hispidus New Phytologist 683 -696 . [本文引用: 1] [26] Niklas KJ 2005 ). Modelling below- and above-ground biomass for non-woody and woody plantsAnnals of Botany 315 -321 . URL PMID:
15546927 [本文引用: 1] [27] Niklas KJ 2006 ). A phyletic perspective on the allometry of plant biomass-partitioning patterns and functionally equivalent organ-categoriesNew Phytologist 27 -40 . URL PMID:
16771980 [本文引用: 1] [28] Niklas KJ Enquist BJ 2002 ). On the vegetative biomass partitioning of seed plant leaves, stems, and rootsThe American Naturalist 482 -497 . URL PMID:
18707431 [本文引用: 1] [29] Peng YF Yang YH 2016 ). Allometric biomass partitioning under nitrogen enrichment: evidence from manipulative experiments around the worldScientific Reports 28918 . DOI: 10.1038/srep28918 . URL PMID:
27349584 [本文引用: 2] [30] Poorter H Nagel O 2000 ). The role of biomass allocation in the growth response of plants to different levels of light, CO2 , nutrients and water: a quantitative reviewAustralian Journal of Plant Physiology 595 -607 . [本文引用: 1] [31] Poorter H Niklas KJ Reich PB Oleksyn J Poot P Mommer L 2012 ). Biomass allocation to leaves, stems and roots: meta-analyses of interspecific variation and environmental controlNew Phytologist 30 -50 . DOI:
10.1111/j.1469-8137.2011.03952.x URL PMID:
22085245 [本文引用: 4] We quantified the biomass allocation patterns to leaves, stems and roots in vegetative plants, and how this is influenced by the growth environment, plant size, evolutionary history and competition. Dose-response curves of allocation were constructed by means of a meta-analysis from a wide array of experimental data. They show that the fraction of whole-plant mass represented by leaves (LMF) increases most strongly with nutrients and decreases most strongly with light. Correction for size-induced allocation patterns diminishes the LMF-response to light, but makes the effect of temperature on LMF more apparent. There is a clear phylogenetic effect on allocation, as eudicots invest relatively more than monocots in leaves, as do gymnosperms compared with woody angiosperms. Plants grown at high densities show a clear increase in the stem fraction. However, in most comparisons across species groups or environmental factors, the variation in LMF is smaller than the variation in one of the other components of the growth analysis equation: the leaf area : leaf mass ratio (SLA). In competitive situations, the stem mass fraction increases to a smaller extent than the specific stem length (stem length : stem mass). Thus, we conclude that plants generally are less able to adjust allocation than to alter organ morphology.
[32] Reekie EG Bazzaz FA 2011 ). Reproductive Allocation in Plants. Academic Press, New York. 1 -49 . [本文引用: 2] [33] Reich PB Luo YJ Bradford JB Poorter H Perry CH Oleksyn J 2014 ). Temperature drives global patterns in forest biomass distribution in leaves, stems, and rootsProceedings of the National Academy of Sciences of the United States of America 13721 -13726 . DOI:
10.1073/pnas.1216053111 URL PMID:
25225412 [本文引用: 1] Whether the fraction of total forest biomass distributed in roots, stems, or leaves varies systematically across geographic gradients remains unknown despite its importance for understanding forest ecology and modeling global carbon cycles. It has been hypothesized that plants should maintain proportionally more biomass in the organ that acquires the most limiting resource. Accordingly, we hypothesize greater biomass distribution in roots and less in stems and foliage in increasingly arid climates and in colder environments at high latitudes. Such a strategy would increase uptake of soil water in dry conditions and of soil nutrients in cold soils, where they are at low supply and are less mobile. We use a large global biomass dataset (>6,200 forests from 61 countries, across a 40 degrees C gradient in mean annual temperature) to address these questions. Climate metrics involving temperature were better predictors of biomass partitioning than those involving moisture availability, because, surprisingly, fractional distribution of biomass to roots or foliage was unrelated to aridity. In contrast, in increasingly cold climates, the proportion of total forest biomass in roots was greater and in foliage was smaller for both angiosperm and gymnosperm forests. These findings support hypotheses about adaptive strategies of forest trees to temperature and provide biogeographically explicit relationships to improve ecosystem and earth system models. They also will allow, for the first time to our knowledge, representations of root carbon pools that consider biogeographic differences, which are useful for quantifying whole-ecosystem carbon stocks and cycles and for assessing the impact of climate change on forest carbon dynamics.
[34] Shipley B Meziane D 2002 ). The balanced-growth hypothesis and the allometry of leaf and root biomass allocationFunctional Ecology 326 -331 . [本文引用: 3] [35] Tian DS Pan QM Simmons M Chaolu HD Du BH Bai YF Wang H Han XG 2012 ). Hierarchical reproductive allocation and allometry within a perennial bunchgrass after 11 years of nutrient additionPLOS ONE e42833 . DOI: 10.1371/journal.pone.0042833 . URL PMID:
22984408 [本文引用: 1] [36] Weiner J Campbell LG Pino J Echarte L 2009 ). The allometry of reproduction within plant populationsJournal of Ecology 1220 -1233 . [本文引用: 1] [37] Wolkovich EM Cook BI Allen JM Crimmins TM Betancourt JL Travers SE Pau S Regetz J Davies TJ Kraft NJB Ault TR Bolmgren K Mazer SJ McCabe GJ McGill BJ Parmesan C Salamin N Schwartz MD Cleland EE 2012 ). Warming experiments underpredict plant phenollogical responses to climate changeNature 494 -497 . URL PMID:
22622576 [本文引用: 1] [38] Xie JB Tang LS Wang ZY Xu GQ Li Y 2012 ). Distinguishing the biomass allocation variance resulting from ontogenetic drift or acclimation to soil texturePLOS ONE e41502 . DOI: 10.1371/journal.pone.0041502 . URL PMID:
22911802 [本文引用: 2] [39] Xu H Li Y Xu GQ Zou T 2007 ). Ecophysiological response and morphological adjustment of two Central Asian desert shrubs towards variation in summer precipitationPlant, Cell & Environment 399 -409 . URL PMID:
17324227 [本文引用: 1] Death without sex—The ‘problem of the small’ and selection for reproductive economy in flowering plants
1
2008
... 对于各器官不同物候期间的差异显著性检验结果表明, 物候期对白刺生物量分配模式具有一定的影响, 且对不同器官分配模式影响存在差异, 这可能与不同器官的功能有关.有些研究也指出植物生物量分配模式在季节之间的变化似乎与其季节性光合作用和水的需求的变化同时发生(
Barnes, 2002 ).由于本文所研究的3个物候期为盛花期、盛果期与营养生长期, 它们之间的差异实质上反映的是繁殖分配和营养分配之间的差异.繁殖分配作为植物生命周期中的一个重要特征, 是植物进行资源分配的一个核心问题(
Tian et al ., 2012 ), 不同的繁殖分配反映了植物在自然选择过程中的生存策略(
Reekie & Bazzaz, 2011 ), 但根据本文的研究结果可知, 繁殖生长对生物量分配模式造成的影响主要体现在相对生物量分配比例(36.00%)而不是个体大小的依赖性程度上(16.67%), 而且对S的影响较大, 但其变化趋势不一致.繁殖生长增加对Le的相对生物量分配比例, 减少对B的相对生物量分配比例, 但并没有改变他们的大小依赖性程度, 进一步证实了植物会通过调节不同器官中的生物量分配, 来实现和维持其生长最大化(
郭丽珠等, 2019 ).同时, 植物大小也是影响生殖分配的重要因素(
Niklas & Enquist, 2002 ;
Cheplick, 2005 ;
Weiner et al ., 2009 ).随着植物的生长, 两种繁殖方式的有效性也会随之发生改变, 一般而言有性繁殖对植物大小有一个阈值要求, 对于克隆植物而言, 在达到阈值之前有性繁殖后代若不能存活下来, 此时克隆繁殖产生的新分株可能成为其唯一的后代并存留下来, 但一旦达到阈值, 伴随着克隆繁殖分配的减小, 有性繁殖分配将会呈现增加的趋势(
Aarssen, 2008 ). ...
Effects of phenology, water availability and seed source on loblolly pine biomass partitioning and transpiration
1
2002
... 对于各器官不同物候期间的差异显著性检验结果表明, 物候期对白刺生物量分配模式具有一定的影响, 且对不同器官分配模式影响存在差异, 这可能与不同器官的功能有关.有些研究也指出植物生物量分配模式在季节之间的变化似乎与其季节性光合作用和水的需求的变化同时发生(
Barnes, 2002 ).由于本文所研究的3个物候期为盛花期、盛果期与营养生长期, 它们之间的差异实质上反映的是繁殖分配和营养分配之间的差异.繁殖分配作为植物生命周期中的一个重要特征, 是植物进行资源分配的一个核心问题(
Tian et al ., 2012 ), 不同的繁殖分配反映了植物在自然选择过程中的生存策略(
Reekie & Bazzaz, 2011 ), 但根据本文的研究结果可知, 繁殖生长对生物量分配模式造成的影响主要体现在相对生物量分配比例(36.00%)而不是个体大小的依赖性程度上(16.67%), 而且对S的影响较大, 但其变化趋势不一致.繁殖生长增加对Le的相对生物量分配比例, 减少对B的相对生物量分配比例, 但并没有改变他们的大小依赖性程度, 进一步证实了植物会通过调节不同器官中的生物量分配, 来实现和维持其生长最大化(
郭丽珠等, 2019 ).同时, 植物大小也是影响生殖分配的重要因素(
Niklas & Enquist, 2002 ;
Cheplick, 2005 ;
Weiner et al ., 2009 ).随着植物的生长, 两种繁殖方式的有效性也会随之发生改变, 一般而言有性繁殖对植物大小有一个阈值要求, 对于克隆植物而言, 在达到阈值之前有性繁殖后代若不能存活下来, 此时克隆繁殖产生的新分株可能成为其唯一的后代并存留下来, 但一旦达到阈值, 伴随着克隆繁殖分配的减小, 有性繁殖分配将会呈现增加的趋势(
Aarssen, 2008 ). ...
Age-related changes in photosynthesis of woody plants
1
2000
... 在生物量分配的研究中, 地上部分与地下部分生物量之间的关系因可作为评估植物生长响应生态环境的重要指标而被广泛研究(
Niklas, 2005 ).本文通过比较白刺2016与2017年地上部分(
x 轴)与地下部分(
y 轴)生物量的SMA回归斜率发现, 白刺在生物量的累积过程中会优先将生物量分配给地下部分, 且其异速生长指数随着时间的推移具有减小趋势, 表明随着植株的生长, 白刺将生物量分配给地下部分的倾向更加明显, 这与
Bond (2000) 和
Niinemets (2004) 等的研究结果相似, 这些现象进一步说明了植物地上部分与地下部分生物量之间的异速生长指数并不总是符合等速理论的观点(
Chen et al ., 2019 ).同时, 在2016与2017年盛果期的SMA回归斜率均有所提升, 表明繁殖分配, 特别是盛果期会在一定程度上减弱这种倾向, 并将部分生物量分配给地上部分的繁殖器官, 以确保自身物候周期的完整性.
Husáková等(2018) 基于20种旱地植物的研究结果表明, 多年生植物随着生长的进行会分配更多的生物量到地上部分, 本文的研究结果与之不同, 这可能与植物自身的生活史有关, 生活史间的差异导致植物生物量分配模式的差异, 进而避免了干旱等环境因子对生长造成的影响(
Chen et al ., 2019 ).研究认为在干旱环境下, 植物会增加根系的生物量分配以获取更多的水资源(
Müller et al ., 2000 ;
Poorter et al ., 2012 ),
Eziz等(2017) 基于对164篇关于干旱胁迫与生物量分配模式文献的分析, 认为干旱胁迫会显著增加根的生物量分配而减少茎和叶的.同时, 当受到养分限制时, 植物同样会增加根系的生物量分配以更大范围地吸收养分(
Shipley & Meziane, 2002 ;
Poorter et al ., 2012 ). ...
Interpreting reproductive allometry: individual strategies of allocation explain size- dependent reproduction in plant populations
2
2009
... 作为植物最基本的生态学特性和功能性状, 生物量累积是生态系统生产力的直观体现(
Klinkhamer et al ., 1992 ).植物生物量的分配模式是植物对环境适应的结果, 并伴随着植物生活史的每一个阶段, 与植物的生长和发育相互影响(
Bonser & Aarssen, 2009 ;
Poorter et al ., 2012 ).同时, 生物量分配模式实质上也反映出植物在各器官生物量分配中的权衡问题.关于植物生物量分配机制的探讨, 一种理论认为植物倾向于将生物量分配给能够获取稀缺资源的器官以最大化适应外界环境的变化(
Fraterrigo et al ., 2006 ;
Poorter et al ., 2012 ;
Reich et al ., 2014 ), 另一种理论认为生物量分配是由植物本身的生长模式可塑性主导的(
McCarthy & Enquist, 2007 ).但事实上, 它们并非对立而是相互补充的(
Niklas, 2006 ).但不可否认的是无论生物量分配模式是为了适应外界环境的变化还是由于生物量分配模式本身的可塑性所致, 每当生物量分配模式显示部分或全部发育的异速生长增长时, 生物量分配特征将显示个体发育漂变(即, 不管环境条件如何, 生物量分配模式将随植物生长和发育而变化), 只有当生物量积累在各个组成部分之间的函数关系表现为等距关系或线性时, 才可以忽略个体发育漂变(
Gedroc et al ., 1996 ;
McConnaughay & Coleman; 1999 ;
Xie et al ., 2012 ).但大多数研究基于集中在温带森林物种的数据, 而忽视了适应干旱条件的沙漠物种的数据.生活环境的差异也必然导致植物的生长规律发生适应性的变化(
Fu et al ., 2015 ), 而植物在不同物候期的生物量分配变化尚不明确. ...
... 近年来, 异速生长已成为研究生物量分配及权衡的重要方法.一些研究结果也表明植物的生物量分配规律可能符合异速分配理论(
Shipley & Meziane, 2002 ;
Peng & Yang, 2016 ), 表明不同器官生物量的分配比例是依赖于植物大小的.如以叶片为例, 假设其生物量分配是依赖于个体大小的, 那么随着其个体生物量的累积(
x ), 叶片的生物量累积(
y )形式已经明确, 并可以用函数ln
y =
c +
b ln
x 表示.而任何一个时期自然界的每一种植物都存在不同大小的个体, 因此, 可通过某一时期不同个体大小之间的拟合来将其异速生长方程表达出来(
Cheverud, 1982 ), 当然对于某些植物在不同年份之间的差异可归因于年龄大小造成的分配模式差异.同时, 植物在长期进化的过程中, 不同的物候期形成了一种响应环境季节性变化的生长发育阶段(如抽芽、开花、结果、落叶、休眠等), 这种生长阶段容易受到环境因子的影响(
Menzel et al ., 2006 ;
Wolkovich et al ., 2012 ), 但却是其本身生长模式对环境的一种适应.大量的研究也表明植物的不同生长发育期之间, 特别是营养生长期与繁殖生长期之间的生物量分配有着显著的差异(
Bonser & Aarssen, 2009 ;
Reekie & Bazzaz, 2011 ;
李清河等; 2012 ). ...
Biomass allocation and multiple resource limitation in tree seedlings
1
1996
... 对白刺连续两个生长季的地上同化器官(
x 轴)与支持器官(
y 轴)生物量的SMA回归斜率比较发现, 在2016年其斜率均大于1, 表明随着个体生物量的增大其生物量向支持器官分配的比率也越大, 但随着时间的推移, 斜率整体上呈下降趋势, 且在2017年的盛花期与营养生长期均低于1, 表明其支持器官的生物量相对于同化器官生物量的变化速率在不同年份之间存在差异, 本文中白刺植株随着生长时间的推移, 会更倾向于将生物量分配给同化吸收器官而不是支持器官.
Canham等(1996) 认为, 植物对环境因子的可塑性响应是为了在资源有限的情况下能够最大限度地增长和生存.因此, 如果受到土壤水分限制, 植物必须生成更多的根系并减少叶面积, 以保持植物体内水分吸收和消耗平衡(
Deng et al. , 2006 ;
Xu et al ., 2007 ), 但是茎秆等支持器官对大部分陆生植物而言, 可能会因为其对资源获取作用不大而在资源限制下无明显响应(
Poorter & Nagel, 2000 ;
Coleman et al ., 2004 ), 本文的研究结果与之相似. ...
Life history strategies drive size-dependent biomass allocation patterns of dryland ephemerals and shrubs
2
2019
... 在生物量分配的研究中, 地上部分与地下部分生物量之间的关系因可作为评估植物生长响应生态环境的重要指标而被广泛研究(
Niklas, 2005 ).本文通过比较白刺2016与2017年地上部分(
x 轴)与地下部分(
y 轴)生物量的SMA回归斜率发现, 白刺在生物量的累积过程中会优先将生物量分配给地下部分, 且其异速生长指数随着时间的推移具有减小趋势, 表明随着植株的生长, 白刺将生物量分配给地下部分的倾向更加明显, 这与
Bond (2000) 和
Niinemets (2004) 等的研究结果相似, 这些现象进一步说明了植物地上部分与地下部分生物量之间的异速生长指数并不总是符合等速理论的观点(
Chen et al ., 2019 ).同时, 在2016与2017年盛果期的SMA回归斜率均有所提升, 表明繁殖分配, 特别是盛果期会在一定程度上减弱这种倾向, 并将部分生物量分配给地上部分的繁殖器官, 以确保自身物候周期的完整性.
Husáková等(2018) 基于20种旱地植物的研究结果表明, 多年生植物随着生长的进行会分配更多的生物量到地上部分, 本文的研究结果与之不同, 这可能与植物自身的生活史有关, 生活史间的差异导致植物生物量分配模式的差异, 进而避免了干旱等环境因子对生长造成的影响(
Chen et al ., 2019 ).研究认为在干旱环境下, 植物会增加根系的生物量分配以获取更多的水资源(
Müller et al ., 2000 ;
Poorter et al ., 2012 ),
Eziz等(2017) 基于对164篇关于干旱胁迫与生物量分配模式文献的分析, 认为干旱胁迫会显著增加根的生物量分配而减少茎和叶的.同时, 当受到养分限制时, 植物同样会增加根系的生物量分配以更大范围地吸收养分(
Shipley & Meziane, 2002 ;
Poorter et al ., 2012 ). ...
... 基于20种旱地植物的研究结果表明, 多年生植物随着生长的进行会分配更多的生物量到地上部分, 本文的研究结果与之不同, 这可能与植物自身的生活史有关, 生活史间的差异导致植物生物量分配模式的差异, 进而避免了干旱等环境因子对生长造成的影响(
Chen et al ., 2019 ).研究认为在干旱环境下, 植物会增加根系的生物量分配以获取更多的水资源(
Müller et al ., 2000 ;
Poorter et al ., 2012 ),
Eziz等(2017) 基于对164篇关于干旱胁迫与生物量分配模式文献的分析, 认为干旱胁迫会显著增加根的生物量分配而减少茎和叶的.同时, 当受到养分限制时, 植物同样会增加根系的生物量分配以更大范围地吸收养分(
Shipley & Meziane, 2002 ;
Poorter et al ., 2012 ). ...
Biomass partitioning and reproductive allocation in the invasive, cleistogamous grass
Microstegium vimineum : influence of the light environment
1
2005
... 对于各器官不同物候期间的差异显著性检验结果表明, 物候期对白刺生物量分配模式具有一定的影响, 且对不同器官分配模式影响存在差异, 这可能与不同器官的功能有关.有些研究也指出植物生物量分配模式在季节之间的变化似乎与其季节性光合作用和水的需求的变化同时发生(
Barnes, 2002 ).由于本文所研究的3个物候期为盛花期、盛果期与营养生长期, 它们之间的差异实质上反映的是繁殖分配和营养分配之间的差异.繁殖分配作为植物生命周期中的一个重要特征, 是植物进行资源分配的一个核心问题(
Tian et al ., 2012 ), 不同的繁殖分配反映了植物在自然选择过程中的生存策略(
Reekie & Bazzaz, 2011 ), 但根据本文的研究结果可知, 繁殖生长对生物量分配模式造成的影响主要体现在相对生物量分配比例(36.00%)而不是个体大小的依赖性程度上(16.67%), 而且对S的影响较大, 但其变化趋势不一致.繁殖生长增加对Le的相对生物量分配比例, 减少对B的相对生物量分配比例, 但并没有改变他们的大小依赖性程度, 进一步证实了植物会通过调节不同器官中的生物量分配, 来实现和维持其生长最大化(
郭丽珠等, 2019 ).同时, 植物大小也是影响生殖分配的重要因素(
Niklas & Enquist, 2002 ;
Cheplick, 2005 ;
Weiner et al ., 2009 ).随着植物的生长, 两种繁殖方式的有效性也会随之发生改变, 一般而言有性繁殖对植物大小有一个阈值要求, 对于克隆植物而言, 在达到阈值之前有性繁殖后代若不能存活下来, 此时克隆繁殖产生的新分株可能成为其唯一的后代并存留下来, 但一旦达到阈值, 伴随着克隆繁殖分配的减小, 有性繁殖分配将会呈现增加的趋势(
Aarssen, 2008 ). ...
Relationships among ontogenetic, static, and evolutionary allometry
1
1982
... 近年来, 异速生长已成为研究生物量分配及权衡的重要方法.一些研究结果也表明植物的生物量分配规律可能符合异速分配理论(
Shipley & Meziane, 2002 ;
Peng & Yang, 2016 ), 表明不同器官生物量的分配比例是依赖于植物大小的.如以叶片为例, 假设其生物量分配是依赖于个体大小的, 那么随着其个体生物量的累积(
x ), 叶片的生物量累积(
y )形式已经明确, 并可以用函数ln
y =
c +
b ln
x 表示.而任何一个时期自然界的每一种植物都存在不同大小的个体, 因此, 可通过某一时期不同个体大小之间的拟合来将其异速生长方程表达出来(
Cheverud, 1982 ), 当然对于某些植物在不同年份之间的差异可归因于年龄大小造成的分配模式差异.同时, 植物在长期进化的过程中, 不同的物候期形成了一种响应环境季节性变化的生长发育阶段(如抽芽、开花、结果、落叶、休眠等), 这种生长阶段容易受到环境因子的影响(
Menzel et al ., 2006 ;
Wolkovich et al ., 2012 ), 但却是其本身生长模式对环境的一种适应.大量的研究也表明植物的不同生长发育期之间, 特别是营养生长期与繁殖生长期之间的生物量分配有着显著的差异(
Bonser & Aarssen, 2009 ;
Reekie & Bazzaz, 2011 ;
李清河等; 2012 ). ...
Carbon allocation and nitrogen acquisition in a developing
Populus deltoides plantation
1
2004
... 对白刺连续两个生长季的地上同化器官(
x 轴)与支持器官(
y 轴)生物量的SMA回归斜率比较发现, 在2016年其斜率均大于1, 表明随着个体生物量的增大其生物量向支持器官分配的比率也越大, 但随着时间的推移, 斜率整体上呈下降趋势, 且在2017年的盛花期与营养生长期均低于1, 表明其支持器官的生物量相对于同化器官生物量的变化速率在不同年份之间存在差异, 本文中白刺植株随着生长时间的推移, 会更倾向于将生物量分配给同化吸收器官而不是支持器官.
Canham等(1996) 认为, 植物对环境因子的可塑性响应是为了在资源有限的情况下能够最大限度地增长和生存.因此, 如果受到土壤水分限制, 植物必须生成更多的根系并减少叶面积, 以保持植物体内水分吸收和消耗平衡(
Deng et al. , 2006 ;
Xu et al ., 2007 ), 但是茎秆等支持器官对大部分陆生植物而言, 可能会因为其对资源获取作用不大而在资源限制下无明显响应(
Poorter & Nagel, 2000 ;
Coleman et al ., 2004 ), 本文的研究结果与之相似. ...
Plant mass-density relationship along a moisture gradient in north-west China
1
2006
... 对白刺连续两个生长季的地上同化器官(
x 轴)与支持器官(
y 轴)生物量的SMA回归斜率比较发现, 在2016年其斜率均大于1, 表明随着个体生物量的增大其生物量向支持器官分配的比率也越大, 但随着时间的推移, 斜率整体上呈下降趋势, 且在2017年的盛花期与营养生长期均低于1, 表明其支持器官的生物量相对于同化器官生物量的变化速率在不同年份之间存在差异, 本文中白刺植株随着生长时间的推移, 会更倾向于将生物量分配给同化吸收器官而不是支持器官.
Canham等(1996) 认为, 植物对环境因子的可塑性响应是为了在资源有限的情况下能够最大限度地增长和生存.因此, 如果受到土壤水分限制, 植物必须生成更多的根系并减少叶面积, 以保持植物体内水分吸收和消耗平衡(
Deng et al. , 2006 ;
Xu et al ., 2007 ), 但是茎秆等支持器官对大部分陆生植物而言, 可能会因为其对资源获取作用不大而在资源限制下无明显响应(
Poorter & Nagel, 2000 ;
Coleman et al ., 2004 ), 本文的研究结果与之相似. ...
Drought effect on plant biomass allocation: a meta- analysis
1
2017
... 在生物量分配的研究中, 地上部分与地下部分生物量之间的关系因可作为评估植物生长响应生态环境的重要指标而被广泛研究(
Niklas, 2005 ).本文通过比较白刺2016与2017年地上部分(
x 轴)与地下部分(
y 轴)生物量的SMA回归斜率发现, 白刺在生物量的累积过程中会优先将生物量分配给地下部分, 且其异速生长指数随着时间的推移具有减小趋势, 表明随着植株的生长, 白刺将生物量分配给地下部分的倾向更加明显, 这与
Bond (2000) 和
Niinemets (2004) 等的研究结果相似, 这些现象进一步说明了植物地上部分与地下部分生物量之间的异速生长指数并不总是符合等速理论的观点(
Chen et al ., 2019 ).同时, 在2016与2017年盛果期的SMA回归斜率均有所提升, 表明繁殖分配, 特别是盛果期会在一定程度上减弱这种倾向, 并将部分生物量分配给地上部分的繁殖器官, 以确保自身物候周期的完整性.
Husáková等(2018) 基于20种旱地植物的研究结果表明, 多年生植物随着生长的进行会分配更多的生物量到地上部分, 本文的研究结果与之不同, 这可能与植物自身的生活史有关, 生活史间的差异导致植物生物量分配模式的差异, 进而避免了干旱等环境因子对生长造成的影响(
Chen et al ., 2019 ).研究认为在干旱环境下, 植物会增加根系的生物量分配以获取更多的水资源(
Müller et al ., 2000 ;
Poorter et al ., 2012 ),
Eziz等(2017) 基于对164篇关于干旱胁迫与生物量分配模式文献的分析, 认为干旱胁迫会显著增加根的生物量分配而减少茎和叶的.同时, 当受到养分限制时, 植物同样会增加根系的生物量分配以更大范围地吸收养分(
Shipley & Meziane, 2002 ;
Poorter et al ., 2012 ). ...
种群密度对沙米异速生长的影响
1
2017
... 异速生长的提出, 为我们研究依赖于个体大小的生物量分配方式提供了方法, 其能够揭示生物在生长过程中的变化规律(
Shipley & Meziane, 2002 ;
Komiyama et al ., 2008 ;
Peng & Yang, 2016 ).本文采用SMA分别拟合了白刺2016与2017年在盛花期、盛果期和营养生长期3个物候期不同器官的异速生长关系.结果显示, 白刺各器官的个体大小依赖性在各时期均极显著, 且相关性较高, 这进一步验证了
范高华等(2017) 的观点, 即植物生育期的变化, 对繁殖生物量与个体大小的关系具有显著影响. ...
种群密度对沙米异速生长的影响
1
2017
... 异速生长的提出, 为我们研究依赖于个体大小的生物量分配方式提供了方法, 其能够揭示生物在生长过程中的变化规律(
Shipley & Meziane, 2002 ;
Komiyama et al ., 2008 ;
Peng & Yang, 2016 ).本文采用SMA分别拟合了白刺2016与2017年在盛花期、盛果期和营养生长期3个物候期不同器官的异速生长关系.结果显示, 白刺各器官的个体大小依赖性在各时期均极显著, 且相关性较高, 这进一步验证了
范高华等(2017) 的观点, 即植物生育期的变化, 对繁殖生物量与个体大小的关系具有显著影响. ...
Previous land use alters plant allocation and growth in forest herbs
1
2006
... 作为植物最基本的生态学特性和功能性状, 生物量累积是生态系统生产力的直观体现(
Klinkhamer et al ., 1992 ).植物生物量的分配模式是植物对环境适应的结果, 并伴随着植物生活史的每一个阶段, 与植物的生长和发育相互影响(
Bonser & Aarssen, 2009 ;
Poorter et al ., 2012 ).同时, 生物量分配模式实质上也反映出植物在各器官生物量分配中的权衡问题.关于植物生物量分配机制的探讨, 一种理论认为植物倾向于将生物量分配给能够获取稀缺资源的器官以最大化适应外界环境的变化(
Fraterrigo et al ., 2006 ;
Poorter et al ., 2012 ;
Reich et al ., 2014 ), 另一种理论认为生物量分配是由植物本身的生长模式可塑性主导的(
McCarthy & Enquist, 2007 ).但事实上, 它们并非对立而是相互补充的(
Niklas, 2006 ).但不可否认的是无论生物量分配模式是为了适应外界环境的变化还是由于生物量分配模式本身的可塑性所致, 每当生物量分配模式显示部分或全部发育的异速生长增长时, 生物量分配特征将显示个体发育漂变(即, 不管环境条件如何, 生物量分配模式将随植物生长和发育而变化), 只有当生物量积累在各个组成部分之间的函数关系表现为等距关系或线性时, 才可以忽略个体发育漂变(
Gedroc et al ., 1996 ;
McConnaughay & Coleman; 1999 ;
Xie et al ., 2012 ).但大多数研究基于集中在温带森林物种的数据, 而忽视了适应干旱条件的沙漠物种的数据.生活环境的差异也必然导致植物的生长规律发生适应性的变化(
Fu et al ., 2015 ), 而植物在不同物候期的生物量分配变化尚不明确. ...
Declining global warming effects on the phenology of spring leaf unfolding
1
2015
... 作为植物最基本的生态学特性和功能性状, 生物量累积是生态系统生产力的直观体现(
Klinkhamer et al ., 1992 ).植物生物量的分配模式是植物对环境适应的结果, 并伴随着植物生活史的每一个阶段, 与植物的生长和发育相互影响(
Bonser & Aarssen, 2009 ;
Poorter et al ., 2012 ).同时, 生物量分配模式实质上也反映出植物在各器官生物量分配中的权衡问题.关于植物生物量分配机制的探讨, 一种理论认为植物倾向于将生物量分配给能够获取稀缺资源的器官以最大化适应外界环境的变化(
Fraterrigo et al ., 2006 ;
Poorter et al ., 2012 ;
Reich et al ., 2014 ), 另一种理论认为生物量分配是由植物本身的生长模式可塑性主导的(
McCarthy & Enquist, 2007 ).但事实上, 它们并非对立而是相互补充的(
Niklas, 2006 ).但不可否认的是无论生物量分配模式是为了适应外界环境的变化还是由于生物量分配模式本身的可塑性所致, 每当生物量分配模式显示部分或全部发育的异速生长增长时, 生物量分配特征将显示个体发育漂变(即, 不管环境条件如何, 生物量分配模式将随植物生长和发育而变化), 只有当生物量积累在各个组成部分之间的函数关系表现为等距关系或线性时, 才可以忽略个体发育漂变(
Gedroc et al ., 1996 ;
McConnaughay & Coleman; 1999 ;
Xie et al ., 2012 ).但大多数研究基于集中在温带森林物种的数据, 而忽视了适应干旱条件的沙漠物种的数据.生活环境的差异也必然导致植物的生长规律发生适应性的变化(
Fu et al ., 2015 ), 而植物在不同物候期的生物量分配变化尚不明确. ...
Plasticity in root/shoot partitioning: optimal, ontogenetic, or both?
1
1996
... 作为植物最基本的生态学特性和功能性状, 生物量累积是生态系统生产力的直观体现(
Klinkhamer et al ., 1992 ).植物生物量的分配模式是植物对环境适应的结果, 并伴随着植物生活史的每一个阶段, 与植物的生长和发育相互影响(
Bonser & Aarssen, 2009 ;
Poorter et al ., 2012 ).同时, 生物量分配模式实质上也反映出植物在各器官生物量分配中的权衡问题.关于植物生物量分配机制的探讨, 一种理论认为植物倾向于将生物量分配给能够获取稀缺资源的器官以最大化适应外界环境的变化(
Fraterrigo et al ., 2006 ;
Poorter et al ., 2012 ;
Reich et al ., 2014 ), 另一种理论认为生物量分配是由植物本身的生长模式可塑性主导的(
McCarthy & Enquist, 2007 ).但事实上, 它们并非对立而是相互补充的(
Niklas, 2006 ).但不可否认的是无论生物量分配模式是为了适应外界环境的变化还是由于生物量分配模式本身的可塑性所致, 每当生物量分配模式显示部分或全部发育的异速生长增长时, 生物量分配特征将显示个体发育漂变(即, 不管环境条件如何, 生物量分配模式将随植物生长和发育而变化), 只有当生物量积累在各个组成部分之间的函数关系表现为等距关系或线性时, 才可以忽略个体发育漂变(
Gedroc et al ., 1996 ;
McConnaughay & Coleman; 1999 ;
Xie et al ., 2012 ).但大多数研究基于集中在温带森林物种的数据, 而忽视了适应干旱条件的沙漠物种的数据.生活环境的差异也必然导致植物的生长规律发生适应性的变化(
Fu et al ., 2015 ), 而植物在不同物候期的生物量分配变化尚不明确. ...
退化典型草原狼毒生物量分配及异速生长分析
1
2019
... 对于各器官不同物候期间的差异显著性检验结果表明, 物候期对白刺生物量分配模式具有一定的影响, 且对不同器官分配模式影响存在差异, 这可能与不同器官的功能有关.有些研究也指出植物生物量分配模式在季节之间的变化似乎与其季节性光合作用和水的需求的变化同时发生(
Barnes, 2002 ).由于本文所研究的3个物候期为盛花期、盛果期与营养生长期, 它们之间的差异实质上反映的是繁殖分配和营养分配之间的差异.繁殖分配作为植物生命周期中的一个重要特征, 是植物进行资源分配的一个核心问题(
Tian et al ., 2012 ), 不同的繁殖分配反映了植物在自然选择过程中的生存策略(
Reekie & Bazzaz, 2011 ), 但根据本文的研究结果可知, 繁殖生长对生物量分配模式造成的影响主要体现在相对生物量分配比例(36.00%)而不是个体大小的依赖性程度上(16.67%), 而且对S的影响较大, 但其变化趋势不一致.繁殖生长增加对Le的相对生物量分配比例, 减少对B的相对生物量分配比例, 但并没有改变他们的大小依赖性程度, 进一步证实了植物会通过调节不同器官中的生物量分配, 来实现和维持其生长最大化(
郭丽珠等, 2019 ).同时, 植物大小也是影响生殖分配的重要因素(
Niklas & Enquist, 2002 ;
Cheplick, 2005 ;
Weiner et al ., 2009 ).随着植物的生长, 两种繁殖方式的有效性也会随之发生改变, 一般而言有性繁殖对植物大小有一个阈值要求, 对于克隆植物而言, 在达到阈值之前有性繁殖后代若不能存活下来, 此时克隆繁殖产生的新分株可能成为其唯一的后代并存留下来, 但一旦达到阈值, 伴随着克隆繁殖分配的减小, 有性繁殖分配将会呈现增加的趋势(
Aarssen, 2008 ). ...
退化典型草原狼毒生物量分配及异速生长分析
1
2019
... 对于各器官不同物候期间的差异显著性检验结果表明, 物候期对白刺生物量分配模式具有一定的影响, 且对不同器官分配模式影响存在差异, 这可能与不同器官的功能有关.有些研究也指出植物生物量分配模式在季节之间的变化似乎与其季节性光合作用和水的需求的变化同时发生(
Barnes, 2002 ).由于本文所研究的3个物候期为盛花期、盛果期与营养生长期, 它们之间的差异实质上反映的是繁殖分配和营养分配之间的差异.繁殖分配作为植物生命周期中的一个重要特征, 是植物进行资源分配的一个核心问题(
Tian et al ., 2012 ), 不同的繁殖分配反映了植物在自然选择过程中的生存策略(
Reekie & Bazzaz, 2011 ), 但根据本文的研究结果可知, 繁殖生长对生物量分配模式造成的影响主要体现在相对生物量分配比例(36.00%)而不是个体大小的依赖性程度上(16.67%), 而且对S的影响较大, 但其变化趋势不一致.繁殖生长增加对Le的相对生物量分配比例, 减少对B的相对生物量分配比例, 但并没有改变他们的大小依赖性程度, 进一步证实了植物会通过调节不同器官中的生物量分配, 来实现和维持其生长最大化(
郭丽珠等, 2019 ).同时, 植物大小也是影响生殖分配的重要因素(
Niklas & Enquist, 2002 ;
Cheplick, 2005 ;
Weiner et al ., 2009 ).随着植物的生长, 两种繁殖方式的有效性也会随之发生改变, 一般而言有性繁殖对植物大小有一个阈值要求, 对于克隆植物而言, 在达到阈值之前有性繁殖后代若不能存活下来, 此时克隆繁殖产生的新分株可能成为其唯一的后代并存留下来, 但一旦达到阈值, 伴随着克隆繁殖分配的减小, 有性繁殖分配将会呈现增加的趋势(
Aarssen, 2008 ). ...
Species traits and shoot-root biomass allocation in 20 dry-grassland species
1
2018
... 在生物量分配的研究中, 地上部分与地下部分生物量之间的关系因可作为评估植物生长响应生态环境的重要指标而被广泛研究(
Niklas, 2005 ).本文通过比较白刺2016与2017年地上部分(
x 轴)与地下部分(
y 轴)生物量的SMA回归斜率发现, 白刺在生物量的累积过程中会优先将生物量分配给地下部分, 且其异速生长指数随着时间的推移具有减小趋势, 表明随着植株的生长, 白刺将生物量分配给地下部分的倾向更加明显, 这与
Bond (2000) 和
Niinemets (2004) 等的研究结果相似, 这些现象进一步说明了植物地上部分与地下部分生物量之间的异速生长指数并不总是符合等速理论的观点(
Chen et al ., 2019 ).同时, 在2016与2017年盛果期的SMA回归斜率均有所提升, 表明繁殖分配, 特别是盛果期会在一定程度上减弱这种倾向, 并将部分生物量分配给地上部分的繁殖器官, 以确保自身物候周期的完整性.
Husáková等(2018) 基于20种旱地植物的研究结果表明, 多年生植物随着生长的进行会分配更多的生物量到地上部分, 本文的研究结果与之不同, 这可能与植物自身的生活史有关, 生活史间的差异导致植物生物量分配模式的差异, 进而避免了干旱等环境因子对生长造成的影响(
Chen et al ., 2019 ).研究认为在干旱环境下, 植物会增加根系的生物量分配以获取更多的水资源(
Müller et al ., 2000 ;
Poorter et al ., 2012 ),
Eziz等(2017) 基于对164篇关于干旱胁迫与生物量分配模式文献的分析, 认为干旱胁迫会显著增加根的生物量分配而减少茎和叶的.同时, 当受到养分限制时, 植物同样会增加根系的生物量分配以更大范围地吸收养分(
Shipley & Meziane, 2002 ;
Poorter et al ., 2012 ). ...
On the analysis of size-dependent reproductive output in plants
1
1992
... 作为植物最基本的生态学特性和功能性状, 生物量累积是生态系统生产力的直观体现(
Klinkhamer et al ., 1992 ).植物生物量的分配模式是植物对环境适应的结果, 并伴随着植物生活史的每一个阶段, 与植物的生长和发育相互影响(
Bonser & Aarssen, 2009 ;
Poorter et al ., 2012 ).同时, 生物量分配模式实质上也反映出植物在各器官生物量分配中的权衡问题.关于植物生物量分配机制的探讨, 一种理论认为植物倾向于将生物量分配给能够获取稀缺资源的器官以最大化适应外界环境的变化(
Fraterrigo et al ., 2006 ;
Poorter et al ., 2012 ;
Reich et al ., 2014 ), 另一种理论认为生物量分配是由植物本身的生长模式可塑性主导的(
McCarthy & Enquist, 2007 ).但事实上, 它们并非对立而是相互补充的(
Niklas, 2006 ).但不可否认的是无论生物量分配模式是为了适应外界环境的变化还是由于生物量分配模式本身的可塑性所致, 每当生物量分配模式显示部分或全部发育的异速生长增长时, 生物量分配特征将显示个体发育漂变(即, 不管环境条件如何, 生物量分配模式将随植物生长和发育而变化), 只有当生物量积累在各个组成部分之间的函数关系表现为等距关系或线性时, 才可以忽略个体发育漂变(
Gedroc et al ., 1996 ;
McConnaughay & Coleman; 1999 ;
Xie et al ., 2012 ).但大多数研究基于集中在温带森林物种的数据, 而忽视了适应干旱条件的沙漠物种的数据.生活环境的差异也必然导致植物的生长规律发生适应性的变化(
Fu et al ., 2015 ), 而植物在不同物候期的生物量分配变化尚不明确. ...
Allometry, biomass, and productivity of mangrove forests: a review
1
2008
... 异速生长的提出, 为我们研究依赖于个体大小的生物量分配方式提供了方法, 其能够揭示生物在生长过程中的变化规律(
Shipley & Meziane, 2002 ;
Komiyama et al ., 2008 ;
Peng & Yang, 2016 ).本文采用SMA分别拟合了白刺2016与2017年在盛花期、盛果期和营养生长期3个物候期不同器官的异速生长关系.结果显示, 白刺各器官的个体大小依赖性在各时期均极显著, 且相关性较高, 这进一步验证了
范高华等(2017) 的观点, 即植物生育期的变化, 对繁殖生物量与个体大小的关系具有显著影响. ...
荒漠植物白刺属4个物种的生殖分配比较
1
2012
... 近年来, 异速生长已成为研究生物量分配及权衡的重要方法.一些研究结果也表明植物的生物量分配规律可能符合异速分配理论(
Shipley & Meziane, 2002 ;
Peng & Yang, 2016 ), 表明不同器官生物量的分配比例是依赖于植物大小的.如以叶片为例, 假设其生物量分配是依赖于个体大小的, 那么随着其个体生物量的累积(
x ), 叶片的生物量累积(
y )形式已经明确, 并可以用函数ln
y =
c +
b ln
x 表示.而任何一个时期自然界的每一种植物都存在不同大小的个体, 因此, 可通过某一时期不同个体大小之间的拟合来将其异速生长方程表达出来(
Cheverud, 1982 ), 当然对于某些植物在不同年份之间的差异可归因于年龄大小造成的分配模式差异.同时, 植物在长期进化的过程中, 不同的物候期形成了一种响应环境季节性变化的生长发育阶段(如抽芽、开花、结果、落叶、休眠等), 这种生长阶段容易受到环境因子的影响(
Menzel et al ., 2006 ;
Wolkovich et al ., 2012 ), 但却是其本身生长模式对环境的一种适应.大量的研究也表明植物的不同生长发育期之间, 特别是营养生长期与繁殖生长期之间的生物量分配有着显著的差异(
Bonser & Aarssen, 2009 ;
Reekie & Bazzaz, 2011 ;
李清河等; 2012 ). ...
荒漠植物白刺属4个物种的生殖分配比较
1
2012
... 近年来, 异速生长已成为研究生物量分配及权衡的重要方法.一些研究结果也表明植物的生物量分配规律可能符合异速分配理论(
Shipley & Meziane, 2002 ;
Peng & Yang, 2016 ), 表明不同器官生物量的分配比例是依赖于植物大小的.如以叶片为例, 假设其生物量分配是依赖于个体大小的, 那么随着其个体生物量的累积(
x ), 叶片的生物量累积(
y )形式已经明确, 并可以用函数ln
y =
c +
b ln
x 表示.而任何一个时期自然界的每一种植物都存在不同大小的个体, 因此, 可通过某一时期不同个体大小之间的拟合来将其异速生长方程表达出来(
Cheverud, 1982 ), 当然对于某些植物在不同年份之间的差异可归因于年龄大小造成的分配模式差异.同时, 植物在长期进化的过程中, 不同的物候期形成了一种响应环境季节性变化的生长发育阶段(如抽芽、开花、结果、落叶、休眠等), 这种生长阶段容易受到环境因子的影响(
Menzel et al ., 2006 ;
Wolkovich et al ., 2012 ), 但却是其本身生长模式对环境的一种适应.大量的研究也表明植物的不同生长发育期之间, 特别是营养生长期与繁殖生长期之间的生物量分配有着显著的差异(
Bonser & Aarssen, 2009 ;
Reekie & Bazzaz, 2011 ;
李清河等; 2012 ). ...
European phenological response to climate change matches the warming pattern
1
2006
... 近年来, 异速生长已成为研究生物量分配及权衡的重要方法.一些研究结果也表明植物的生物量分配规律可能符合异速分配理论(
Shipley & Meziane, 2002 ;
Peng & Yang, 2016 ), 表明不同器官生物量的分配比例是依赖于植物大小的.如以叶片为例, 假设其生物量分配是依赖于个体大小的, 那么随着其个体生物量的累积(
x ), 叶片的生物量累积(
y )形式已经明确, 并可以用函数ln
y =
c +
b ln
x 表示.而任何一个时期自然界的每一种植物都存在不同大小的个体, 因此, 可通过某一时期不同个体大小之间的拟合来将其异速生长方程表达出来(
Cheverud, 1982 ), 当然对于某些植物在不同年份之间的差异可归因于年龄大小造成的分配模式差异.同时, 植物在长期进化的过程中, 不同的物候期形成了一种响应环境季节性变化的生长发育阶段(如抽芽、开花、结果、落叶、休眠等), 这种生长阶段容易受到环境因子的影响(
Menzel et al ., 2006 ;
Wolkovich et al ., 2012 ), 但却是其本身生长模式对环境的一种适应.大量的研究也表明植物的不同生长发育期之间, 特别是营养生长期与繁殖生长期之间的生物量分配有着显著的差异(
Bonser & Aarssen, 2009 ;
Reekie & Bazzaz, 2011 ;
李清河等; 2012 ). ...
Consistency between an allometric approach and optimal partitioning theory in global patterns of plant biomass allocation
1
2007
... 作为植物最基本的生态学特性和功能性状, 生物量累积是生态系统生产力的直观体现(
Klinkhamer et al ., 1992 ).植物生物量的分配模式是植物对环境适应的结果, 并伴随着植物生活史的每一个阶段, 与植物的生长和发育相互影响(
Bonser & Aarssen, 2009 ;
Poorter et al ., 2012 ).同时, 生物量分配模式实质上也反映出植物在各器官生物量分配中的权衡问题.关于植物生物量分配机制的探讨, 一种理论认为植物倾向于将生物量分配给能够获取稀缺资源的器官以最大化适应外界环境的变化(
Fraterrigo et al ., 2006 ;
Poorter et al ., 2012 ;
Reich et al ., 2014 ), 另一种理论认为生物量分配是由植物本身的生长模式可塑性主导的(
McCarthy & Enquist, 2007 ).但事实上, 它们并非对立而是相互补充的(
Niklas, 2006 ).但不可否认的是无论生物量分配模式是为了适应外界环境的变化还是由于生物量分配模式本身的可塑性所致, 每当生物量分配模式显示部分或全部发育的异速生长增长时, 生物量分配特征将显示个体发育漂变(即, 不管环境条件如何, 生物量分配模式将随植物生长和发育而变化), 只有当生物量积累在各个组成部分之间的函数关系表现为等距关系或线性时, 才可以忽略个体发育漂变(
Gedroc et al ., 1996 ;
McConnaughay & Coleman; 1999 ;
Xie et al ., 2012 ).但大多数研究基于集中在温带森林物种的数据, 而忽视了适应干旱条件的沙漠物种的数据.生活环境的差异也必然导致植物的生长规律发生适应性的变化(
Fu et al ., 2015 ), 而植物在不同物候期的生物量分配变化尚不明确. ...
Biomass allocation in plants: ontogeny or optimality? A test along three resource gradients
1
1999
... 作为植物最基本的生态学特性和功能性状, 生物量累积是生态系统生产力的直观体现(
Klinkhamer et al ., 1992 ).植物生物量的分配模式是植物对环境适应的结果, 并伴随着植物生活史的每一个阶段, 与植物的生长和发育相互影响(
Bonser & Aarssen, 2009 ;
Poorter et al ., 2012 ).同时, 生物量分配模式实质上也反映出植物在各器官生物量分配中的权衡问题.关于植物生物量分配机制的探讨, 一种理论认为植物倾向于将生物量分配给能够获取稀缺资源的器官以最大化适应外界环境的变化(
Fraterrigo et al ., 2006 ;
Poorter et al ., 2012 ;
Reich et al ., 2014 ), 另一种理论认为生物量分配是由植物本身的生长模式可塑性主导的(
McCarthy & Enquist, 2007 ).但事实上, 它们并非对立而是相互补充的(
Niklas, 2006 ).但不可否认的是无论生物量分配模式是为了适应外界环境的变化还是由于生物量分配模式本身的可塑性所致, 每当生物量分配模式显示部分或全部发育的异速生长增长时, 生物量分配特征将显示个体发育漂变(即, 不管环境条件如何, 生物量分配模式将随植物生长和发育而变化), 只有当生物量积累在各个组成部分之间的函数关系表现为等距关系或线性时, 才可以忽略个体发育漂变(
Gedroc et al ., 1996 ;
McConnaughay & Coleman; 1999 ;
Xie et al ., 2012 ).但大多数研究基于集中在温带森林物种的数据, 而忽视了适应干旱条件的沙漠物种的数据.生活环境的差异也必然导致植物的生长规律发生适应性的变化(
Fu et al ., 2015 ), 而植物在不同物候期的生物量分配变化尚不明确. ...
The effect of nutrient availability on biomass allocation patterns in 27 species of herbaceous plants
1
2000
... 在生物量分配的研究中, 地上部分与地下部分生物量之间的关系因可作为评估植物生长响应生态环境的重要指标而被广泛研究(
Niklas, 2005 ).本文通过比较白刺2016与2017年地上部分(
x 轴)与地下部分(
y 轴)生物量的SMA回归斜率发现, 白刺在生物量的累积过程中会优先将生物量分配给地下部分, 且其异速生长指数随着时间的推移具有减小趋势, 表明随着植株的生长, 白刺将生物量分配给地下部分的倾向更加明显, 这与
Bond (2000) 和
Niinemets (2004) 等的研究结果相似, 这些现象进一步说明了植物地上部分与地下部分生物量之间的异速生长指数并不总是符合等速理论的观点(
Chen et al ., 2019 ).同时, 在2016与2017年盛果期的SMA回归斜率均有所提升, 表明繁殖分配, 特别是盛果期会在一定程度上减弱这种倾向, 并将部分生物量分配给地上部分的繁殖器官, 以确保自身物候周期的完整性.
Husáková等(2018) 基于20种旱地植物的研究结果表明, 多年生植物随着生长的进行会分配更多的生物量到地上部分, 本文的研究结果与之不同, 这可能与植物自身的生活史有关, 生活史间的差异导致植物生物量分配模式的差异, 进而避免了干旱等环境因子对生长造成的影响(
Chen et al ., 2019 ).研究认为在干旱环境下, 植物会增加根系的生物量分配以获取更多的水资源(
Müller et al ., 2000 ;
Poorter et al ., 2012 ),
Eziz等(2017) 基于对164篇关于干旱胁迫与生物量分配模式文献的分析, 认为干旱胁迫会显著增加根的生物量分配而减少茎和叶的.同时, 当受到养分限制时, 植物同样会增加根系的生物量分配以更大范围地吸收养分(
Shipley & Meziane, 2002 ;
Poorter et al ., 2012 ). ...
Adaptive adjustments to light in foliage and whole-plant characteristics depend on relative age in the perennial herb
Leontodon hispidus 1
2004
... 在生物量分配的研究中, 地上部分与地下部分生物量之间的关系因可作为评估植物生长响应生态环境的重要指标而被广泛研究(
Niklas, 2005 ).本文通过比较白刺2016与2017年地上部分(
x 轴)与地下部分(
y 轴)生物量的SMA回归斜率发现, 白刺在生物量的累积过程中会优先将生物量分配给地下部分, 且其异速生长指数随着时间的推移具有减小趋势, 表明随着植株的生长, 白刺将生物量分配给地下部分的倾向更加明显, 这与
Bond (2000) 和
Niinemets (2004) 等的研究结果相似, 这些现象进一步说明了植物地上部分与地下部分生物量之间的异速生长指数并不总是符合等速理论的观点(
Chen et al ., 2019 ).同时, 在2016与2017年盛果期的SMA回归斜率均有所提升, 表明繁殖分配, 特别是盛果期会在一定程度上减弱这种倾向, 并将部分生物量分配给地上部分的繁殖器官, 以确保自身物候周期的完整性.
Husáková等(2018) 基于20种旱地植物的研究结果表明, 多年生植物随着生长的进行会分配更多的生物量到地上部分, 本文的研究结果与之不同, 这可能与植物自身的生活史有关, 生活史间的差异导致植物生物量分配模式的差异, 进而避免了干旱等环境因子对生长造成的影响(
Chen et al ., 2019 ).研究认为在干旱环境下, 植物会增加根系的生物量分配以获取更多的水资源(
Müller et al ., 2000 ;
Poorter et al ., 2012 ),
Eziz等(2017) 基于对164篇关于干旱胁迫与生物量分配模式文献的分析, 认为干旱胁迫会显著增加根的生物量分配而减少茎和叶的.同时, 当受到养分限制时, 植物同样会增加根系的生物量分配以更大范围地吸收养分(
Shipley & Meziane, 2002 ;
Poorter et al ., 2012 ). ...
Modelling below- and above-ground biomass for non-woody and woody plants
1
2005
... 在生物量分配的研究中, 地上部分与地下部分生物量之间的关系因可作为评估植物生长响应生态环境的重要指标而被广泛研究(
Niklas, 2005 ).本文通过比较白刺2016与2017年地上部分(
x 轴)与地下部分(
y 轴)生物量的SMA回归斜率发现, 白刺在生物量的累积过程中会优先将生物量分配给地下部分, 且其异速生长指数随着时间的推移具有减小趋势, 表明随着植株的生长, 白刺将生物量分配给地下部分的倾向更加明显, 这与
Bond (2000) 和
Niinemets (2004) 等的研究结果相似, 这些现象进一步说明了植物地上部分与地下部分生物量之间的异速生长指数并不总是符合等速理论的观点(
Chen et al ., 2019 ).同时, 在2016与2017年盛果期的SMA回归斜率均有所提升, 表明繁殖分配, 特别是盛果期会在一定程度上减弱这种倾向, 并将部分生物量分配给地上部分的繁殖器官, 以确保自身物候周期的完整性.
Husáková等(2018) 基于20种旱地植物的研究结果表明, 多年生植物随着生长的进行会分配更多的生物量到地上部分, 本文的研究结果与之不同, 这可能与植物自身的生活史有关, 生活史间的差异导致植物生物量分配模式的差异, 进而避免了干旱等环境因子对生长造成的影响(
Chen et al ., 2019 ).研究认为在干旱环境下, 植物会增加根系的生物量分配以获取更多的水资源(
Müller et al ., 2000 ;
Poorter et al ., 2012 ),
Eziz等(2017) 基于对164篇关于干旱胁迫与生物量分配模式文献的分析, 认为干旱胁迫会显著增加根的生物量分配而减少茎和叶的.同时, 当受到养分限制时, 植物同样会增加根系的生物量分配以更大范围地吸收养分(
Shipley & Meziane, 2002 ;
Poorter et al ., 2012 ). ...
A phyletic perspective on the allometry of plant biomass-partitioning patterns and functionally equivalent organ-categories
1
2006
... 作为植物最基本的生态学特性和功能性状, 生物量累积是生态系统生产力的直观体现(
Klinkhamer et al ., 1992 ).植物生物量的分配模式是植物对环境适应的结果, 并伴随着植物生活史的每一个阶段, 与植物的生长和发育相互影响(
Bonser & Aarssen, 2009 ;
Poorter et al ., 2012 ).同时, 生物量分配模式实质上也反映出植物在各器官生物量分配中的权衡问题.关于植物生物量分配机制的探讨, 一种理论认为植物倾向于将生物量分配给能够获取稀缺资源的器官以最大化适应外界环境的变化(
Fraterrigo et al ., 2006 ;
Poorter et al ., 2012 ;
Reich et al ., 2014 ), 另一种理论认为生物量分配是由植物本身的生长模式可塑性主导的(
McCarthy & Enquist, 2007 ).但事实上, 它们并非对立而是相互补充的(
Niklas, 2006 ).但不可否认的是无论生物量分配模式是为了适应外界环境的变化还是由于生物量分配模式本身的可塑性所致, 每当生物量分配模式显示部分或全部发育的异速生长增长时, 生物量分配特征将显示个体发育漂变(即, 不管环境条件如何, 生物量分配模式将随植物生长和发育而变化), 只有当生物量积累在各个组成部分之间的函数关系表现为等距关系或线性时, 才可以忽略个体发育漂变(
Gedroc et al ., 1996 ;
McConnaughay & Coleman; 1999 ;
Xie et al ., 2012 ).但大多数研究基于集中在温带森林物种的数据, 而忽视了适应干旱条件的沙漠物种的数据.生活环境的差异也必然导致植物的生长规律发生适应性的变化(
Fu et al ., 2015 ), 而植物在不同物候期的生物量分配变化尚不明确. ...
On the vegetative biomass partitioning of seed plant leaves, stems, and roots
1
2002
... 对于各器官不同物候期间的差异显著性检验结果表明, 物候期对白刺生物量分配模式具有一定的影响, 且对不同器官分配模式影响存在差异, 这可能与不同器官的功能有关.有些研究也指出植物生物量分配模式在季节之间的变化似乎与其季节性光合作用和水的需求的变化同时发生(
Barnes, 2002 ).由于本文所研究的3个物候期为盛花期、盛果期与营养生长期, 它们之间的差异实质上反映的是繁殖分配和营养分配之间的差异.繁殖分配作为植物生命周期中的一个重要特征, 是植物进行资源分配的一个核心问题(
Tian et al ., 2012 ), 不同的繁殖分配反映了植物在自然选择过程中的生存策略(
Reekie & Bazzaz, 2011 ), 但根据本文的研究结果可知, 繁殖生长对生物量分配模式造成的影响主要体现在相对生物量分配比例(36.00%)而不是个体大小的依赖性程度上(16.67%), 而且对S的影响较大, 但其变化趋势不一致.繁殖生长增加对Le的相对生物量分配比例, 减少对B的相对生物量分配比例, 但并没有改变他们的大小依赖性程度, 进一步证实了植物会通过调节不同器官中的生物量分配, 来实现和维持其生长最大化(
郭丽珠等, 2019 ).同时, 植物大小也是影响生殖分配的重要因素(
Niklas & Enquist, 2002 ;
Cheplick, 2005 ;
Weiner et al ., 2009 ).随着植物的生长, 两种繁殖方式的有效性也会随之发生改变, 一般而言有性繁殖对植物大小有一个阈值要求, 对于克隆植物而言, 在达到阈值之前有性繁殖后代若不能存活下来, 此时克隆繁殖产生的新分株可能成为其唯一的后代并存留下来, 但一旦达到阈值, 伴随着克隆繁殖分配的减小, 有性繁殖分配将会呈现增加的趋势(
Aarssen, 2008 ). ...
Allometric biomass partitioning under nitrogen enrichment: evidence from manipulative experiments around the world
2
2016
... 近年来, 异速生长已成为研究生物量分配及权衡的重要方法.一些研究结果也表明植物的生物量分配规律可能符合异速分配理论(
Shipley & Meziane, 2002 ;
Peng & Yang, 2016 ), 表明不同器官生物量的分配比例是依赖于植物大小的.如以叶片为例, 假设其生物量分配是依赖于个体大小的, 那么随着其个体生物量的累积(
x ), 叶片的生物量累积(
y )形式已经明确, 并可以用函数ln
y =
c +
b ln
x 表示.而任何一个时期自然界的每一种植物都存在不同大小的个体, 因此, 可通过某一时期不同个体大小之间的拟合来将其异速生长方程表达出来(
Cheverud, 1982 ), 当然对于某些植物在不同年份之间的差异可归因于年龄大小造成的分配模式差异.同时, 植物在长期进化的过程中, 不同的物候期形成了一种响应环境季节性变化的生长发育阶段(如抽芽、开花、结果、落叶、休眠等), 这种生长阶段容易受到环境因子的影响(
Menzel et al ., 2006 ;
Wolkovich et al ., 2012 ), 但却是其本身生长模式对环境的一种适应.大量的研究也表明植物的不同生长发育期之间, 特别是营养生长期与繁殖生长期之间的生物量分配有着显著的差异(
Bonser & Aarssen, 2009 ;
Reekie & Bazzaz, 2011 ;
李清河等; 2012 ). ...
... 异速生长的提出, 为我们研究依赖于个体大小的生物量分配方式提供了方法, 其能够揭示生物在生长过程中的变化规律(
Shipley & Meziane, 2002 ;
Komiyama et al ., 2008 ;
Peng & Yang, 2016 ).本文采用SMA分别拟合了白刺2016与2017年在盛花期、盛果期和营养生长期3个物候期不同器官的异速生长关系.结果显示, 白刺各器官的个体大小依赖性在各时期均极显著, 且相关性较高, 这进一步验证了
范高华等(2017) 的观点, 即植物生育期的变化, 对繁殖生物量与个体大小的关系具有显著影响. ...
The role of biomass allocation in the growth response of plants to different levels of light, CO
2 , nutrients and water: a quantitative review
1
2000
... 对白刺连续两个生长季的地上同化器官(
x 轴)与支持器官(
y 轴)生物量的SMA回归斜率比较发现, 在2016年其斜率均大于1, 表明随着个体生物量的增大其生物量向支持器官分配的比率也越大, 但随着时间的推移, 斜率整体上呈下降趋势, 且在2017年的盛花期与营养生长期均低于1, 表明其支持器官的生物量相对于同化器官生物量的变化速率在不同年份之间存在差异, 本文中白刺植株随着生长时间的推移, 会更倾向于将生物量分配给同化吸收器官而不是支持器官.
Canham等(1996) 认为, 植物对环境因子的可塑性响应是为了在资源有限的情况下能够最大限度地增长和生存.因此, 如果受到土壤水分限制, 植物必须生成更多的根系并减少叶面积, 以保持植物体内水分吸收和消耗平衡(
Deng et al. , 2006 ;
Xu et al ., 2007 ), 但是茎秆等支持器官对大部分陆生植物而言, 可能会因为其对资源获取作用不大而在资源限制下无明显响应(
Poorter & Nagel, 2000 ;
Coleman et al ., 2004 ), 本文的研究结果与之相似. ...
Biomass allocation to leaves, stems and roots: meta-analyses of interspecific variation and environmental control
4
2012
... 作为植物最基本的生态学特性和功能性状, 生物量累积是生态系统生产力的直观体现(
Klinkhamer et al ., 1992 ).植物生物量的分配模式是植物对环境适应的结果, 并伴随着植物生活史的每一个阶段, 与植物的生长和发育相互影响(
Bonser & Aarssen, 2009 ;
Poorter et al ., 2012 ).同时, 生物量分配模式实质上也反映出植物在各器官生物量分配中的权衡问题.关于植物生物量分配机制的探讨, 一种理论认为植物倾向于将生物量分配给能够获取稀缺资源的器官以最大化适应外界环境的变化(
Fraterrigo et al ., 2006 ;
Poorter et al ., 2012 ;
Reich et al ., 2014 ), 另一种理论认为生物量分配是由植物本身的生长模式可塑性主导的(
McCarthy & Enquist, 2007 ).但事实上, 它们并非对立而是相互补充的(
Niklas, 2006 ).但不可否认的是无论生物量分配模式是为了适应外界环境的变化还是由于生物量分配模式本身的可塑性所致, 每当生物量分配模式显示部分或全部发育的异速生长增长时, 生物量分配特征将显示个体发育漂变(即, 不管环境条件如何, 生物量分配模式将随植物生长和发育而变化), 只有当生物量积累在各个组成部分之间的函数关系表现为等距关系或线性时, 才可以忽略个体发育漂变(
Gedroc et al ., 1996 ;
McConnaughay & Coleman; 1999 ;
Xie et al ., 2012 ).但大多数研究基于集中在温带森林物种的数据, 而忽视了适应干旱条件的沙漠物种的数据.生活环境的差异也必然导致植物的生长规律发生适应性的变化(
Fu et al ., 2015 ), 而植物在不同物候期的生物量分配变化尚不明确. ...
... ;
Poorter et al ., 2012 ;
Reich et al ., 2014 ), 另一种理论认为生物量分配是由植物本身的生长模式可塑性主导的(
McCarthy & Enquist, 2007 ).但事实上, 它们并非对立而是相互补充的(
Niklas, 2006 ).但不可否认的是无论生物量分配模式是为了适应外界环境的变化还是由于生物量分配模式本身的可塑性所致, 每当生物量分配模式显示部分或全部发育的异速生长增长时, 生物量分配特征将显示个体发育漂变(即, 不管环境条件如何, 生物量分配模式将随植物生长和发育而变化), 只有当生物量积累在各个组成部分之间的函数关系表现为等距关系或线性时, 才可以忽略个体发育漂变(
Gedroc et al ., 1996 ;
McConnaughay & Coleman; 1999 ;
Xie et al ., 2012 ).但大多数研究基于集中在温带森林物种的数据, 而忽视了适应干旱条件的沙漠物种的数据.生活环境的差异也必然导致植物的生长规律发生适应性的变化(
Fu et al ., 2015 ), 而植物在不同物候期的生物量分配变化尚不明确. ...
... 在生物量分配的研究中, 地上部分与地下部分生物量之间的关系因可作为评估植物生长响应生态环境的重要指标而被广泛研究(
Niklas, 2005 ).本文通过比较白刺2016与2017年地上部分(
x 轴)与地下部分(
y 轴)生物量的SMA回归斜率发现, 白刺在生物量的累积过程中会优先将生物量分配给地下部分, 且其异速生长指数随着时间的推移具有减小趋势, 表明随着植株的生长, 白刺将生物量分配给地下部分的倾向更加明显, 这与
Bond (2000) 和
Niinemets (2004) 等的研究结果相似, 这些现象进一步说明了植物地上部分与地下部分生物量之间的异速生长指数并不总是符合等速理论的观点(
Chen et al ., 2019 ).同时, 在2016与2017年盛果期的SMA回归斜率均有所提升, 表明繁殖分配, 特别是盛果期会在一定程度上减弱这种倾向, 并将部分生物量分配给地上部分的繁殖器官, 以确保自身物候周期的完整性.
Husáková等(2018) 基于20种旱地植物的研究结果表明, 多年生植物随着生长的进行会分配更多的生物量到地上部分, 本文的研究结果与之不同, 这可能与植物自身的生活史有关, 生活史间的差异导致植物生物量分配模式的差异, 进而避免了干旱等环境因子对生长造成的影响(
Chen et al ., 2019 ).研究认为在干旱环境下, 植物会增加根系的生物量分配以获取更多的水资源(
Müller et al ., 2000 ;
Poorter et al ., 2012 ),
Eziz等(2017) 基于对164篇关于干旱胁迫与生物量分配模式文献的分析, 认为干旱胁迫会显著增加根的生物量分配而减少茎和叶的.同时, 当受到养分限制时, 植物同样会增加根系的生物量分配以更大范围地吸收养分(
Shipley & Meziane, 2002 ;
Poorter et al ., 2012 ). ...
... ;
Poorter et al ., 2012 ). ...
2
2011
... 近年来, 异速生长已成为研究生物量分配及权衡的重要方法.一些研究结果也表明植物的生物量分配规律可能符合异速分配理论(
Shipley & Meziane, 2002 ;
Peng & Yang, 2016 ), 表明不同器官生物量的分配比例是依赖于植物大小的.如以叶片为例, 假设其生物量分配是依赖于个体大小的, 那么随着其个体生物量的累积(
x ), 叶片的生物量累积(
y )形式已经明确, 并可以用函数ln
y =
c +
b ln
x 表示.而任何一个时期自然界的每一种植物都存在不同大小的个体, 因此, 可通过某一时期不同个体大小之间的拟合来将其异速生长方程表达出来(
Cheverud, 1982 ), 当然对于某些植物在不同年份之间的差异可归因于年龄大小造成的分配模式差异.同时, 植物在长期进化的过程中, 不同的物候期形成了一种响应环境季节性变化的生长发育阶段(如抽芽、开花、结果、落叶、休眠等), 这种生长阶段容易受到环境因子的影响(
Menzel et al ., 2006 ;
Wolkovich et al ., 2012 ), 但却是其本身生长模式对环境的一种适应.大量的研究也表明植物的不同生长发育期之间, 特别是营养生长期与繁殖生长期之间的生物量分配有着显著的差异(
Bonser & Aarssen, 2009 ;
Reekie & Bazzaz, 2011 ;
李清河等; 2012 ). ...
... 对于各器官不同物候期间的差异显著性检验结果表明, 物候期对白刺生物量分配模式具有一定的影响, 且对不同器官分配模式影响存在差异, 这可能与不同器官的功能有关.有些研究也指出植物生物量分配模式在季节之间的变化似乎与其季节性光合作用和水的需求的变化同时发生(
Barnes, 2002 ).由于本文所研究的3个物候期为盛花期、盛果期与营养生长期, 它们之间的差异实质上反映的是繁殖分配和营养分配之间的差异.繁殖分配作为植物生命周期中的一个重要特征, 是植物进行资源分配的一个核心问题(
Tian et al ., 2012 ), 不同的繁殖分配反映了植物在自然选择过程中的生存策略(
Reekie & Bazzaz, 2011 ), 但根据本文的研究结果可知, 繁殖生长对生物量分配模式造成的影响主要体现在相对生物量分配比例(36.00%)而不是个体大小的依赖性程度上(16.67%), 而且对S的影响较大, 但其变化趋势不一致.繁殖生长增加对Le的相对生物量分配比例, 减少对B的相对生物量分配比例, 但并没有改变他们的大小依赖性程度, 进一步证实了植物会通过调节不同器官中的生物量分配, 来实现和维持其生长最大化(
郭丽珠等, 2019 ).同时, 植物大小也是影响生殖分配的重要因素(
Niklas & Enquist, 2002 ;
Cheplick, 2005 ;
Weiner et al ., 2009 ).随着植物的生长, 两种繁殖方式的有效性也会随之发生改变, 一般而言有性繁殖对植物大小有一个阈值要求, 对于克隆植物而言, 在达到阈值之前有性繁殖后代若不能存活下来, 此时克隆繁殖产生的新分株可能成为其唯一的后代并存留下来, 但一旦达到阈值, 伴随着克隆繁殖分配的减小, 有性繁殖分配将会呈现增加的趋势(
Aarssen, 2008 ). ...
Temperature drives global patterns in forest biomass distribution in leaves, stems, and roots
1
2014
... 作为植物最基本的生态学特性和功能性状, 生物量累积是生态系统生产力的直观体现(
Klinkhamer et al ., 1992 ).植物生物量的分配模式是植物对环境适应的结果, 并伴随着植物生活史的每一个阶段, 与植物的生长和发育相互影响(
Bonser & Aarssen, 2009 ;
Poorter et al ., 2012 ).同时, 生物量分配模式实质上也反映出植物在各器官生物量分配中的权衡问题.关于植物生物量分配机制的探讨, 一种理论认为植物倾向于将生物量分配给能够获取稀缺资源的器官以最大化适应外界环境的变化(
Fraterrigo et al ., 2006 ;
Poorter et al ., 2012 ;
Reich et al ., 2014 ), 另一种理论认为生物量分配是由植物本身的生长模式可塑性主导的(
McCarthy & Enquist, 2007 ).但事实上, 它们并非对立而是相互补充的(
Niklas, 2006 ).但不可否认的是无论生物量分配模式是为了适应外界环境的变化还是由于生物量分配模式本身的可塑性所致, 每当生物量分配模式显示部分或全部发育的异速生长增长时, 生物量分配特征将显示个体发育漂变(即, 不管环境条件如何, 生物量分配模式将随植物生长和发育而变化), 只有当生物量积累在各个组成部分之间的函数关系表现为等距关系或线性时, 才可以忽略个体发育漂变(
Gedroc et al ., 1996 ;
McConnaughay & Coleman; 1999 ;
Xie et al ., 2012 ).但大多数研究基于集中在温带森林物种的数据, 而忽视了适应干旱条件的沙漠物种的数据.生活环境的差异也必然导致植物的生长规律发生适应性的变化(
Fu et al ., 2015 ), 而植物在不同物候期的生物量分配变化尚不明确. ...
The balanced-growth hypothesis and the allometry of leaf and root biomass allocation
3
2002
... 近年来, 异速生长已成为研究生物量分配及权衡的重要方法.一些研究结果也表明植物的生物量分配规律可能符合异速分配理论(
Shipley & Meziane, 2002 ;
Peng & Yang, 2016 ), 表明不同器官生物量的分配比例是依赖于植物大小的.如以叶片为例, 假设其生物量分配是依赖于个体大小的, 那么随着其个体生物量的累积(
x ), 叶片的生物量累积(
y )形式已经明确, 并可以用函数ln
y =
c +
b ln
x 表示.而任何一个时期自然界的每一种植物都存在不同大小的个体, 因此, 可通过某一时期不同个体大小之间的拟合来将其异速生长方程表达出来(
Cheverud, 1982 ), 当然对于某些植物在不同年份之间的差异可归因于年龄大小造成的分配模式差异.同时, 植物在长期进化的过程中, 不同的物候期形成了一种响应环境季节性变化的生长发育阶段(如抽芽、开花、结果、落叶、休眠等), 这种生长阶段容易受到环境因子的影响(
Menzel et al ., 2006 ;
Wolkovich et al ., 2012 ), 但却是其本身生长模式对环境的一种适应.大量的研究也表明植物的不同生长发育期之间, 特别是营养生长期与繁殖生长期之间的生物量分配有着显著的差异(
Bonser & Aarssen, 2009 ;
Reekie & Bazzaz, 2011 ;
李清河等; 2012 ). ...
... 异速生长的提出, 为我们研究依赖于个体大小的生物量分配方式提供了方法, 其能够揭示生物在生长过程中的变化规律(
Shipley & Meziane, 2002 ;
Komiyama et al ., 2008 ;
Peng & Yang, 2016 ).本文采用SMA分别拟合了白刺2016与2017年在盛花期、盛果期和营养生长期3个物候期不同器官的异速生长关系.结果显示, 白刺各器官的个体大小依赖性在各时期均极显著, 且相关性较高, 这进一步验证了
范高华等(2017) 的观点, 即植物生育期的变化, 对繁殖生物量与个体大小的关系具有显著影响. ...
... 在生物量分配的研究中, 地上部分与地下部分生物量之间的关系因可作为评估植物生长响应生态环境的重要指标而被广泛研究(
Niklas, 2005 ).本文通过比较白刺2016与2017年地上部分(
x 轴)与地下部分(
y 轴)生物量的SMA回归斜率发现, 白刺在生物量的累积过程中会优先将生物量分配给地下部分, 且其异速生长指数随着时间的推移具有减小趋势, 表明随着植株的生长, 白刺将生物量分配给地下部分的倾向更加明显, 这与
Bond (2000) 和
Niinemets (2004) 等的研究结果相似, 这些现象进一步说明了植物地上部分与地下部分生物量之间的异速生长指数并不总是符合等速理论的观点(
Chen et al ., 2019 ).同时, 在2016与2017年盛果期的SMA回归斜率均有所提升, 表明繁殖分配, 特别是盛果期会在一定程度上减弱这种倾向, 并将部分生物量分配给地上部分的繁殖器官, 以确保自身物候周期的完整性.
Husáková等(2018) 基于20种旱地植物的研究结果表明, 多年生植物随着生长的进行会分配更多的生物量到地上部分, 本文的研究结果与之不同, 这可能与植物自身的生活史有关, 生活史间的差异导致植物生物量分配模式的差异, 进而避免了干旱等环境因子对生长造成的影响(
Chen et al ., 2019 ).研究认为在干旱环境下, 植物会增加根系的生物量分配以获取更多的水资源(
Müller et al ., 2000 ;
Poorter et al ., 2012 ),
Eziz等(2017) 基于对164篇关于干旱胁迫与生物量分配模式文献的分析, 认为干旱胁迫会显著增加根的生物量分配而减少茎和叶的.同时, 当受到养分限制时, 植物同样会增加根系的生物量分配以更大范围地吸收养分(
Shipley & Meziane, 2002 ;
Poorter et al ., 2012 ). ...
Hierarchical reproductive allocation and allometry within a perennial bunchgrass after 11 years of nutrient addition
1
2012
... 对于各器官不同物候期间的差异显著性检验结果表明, 物候期对白刺生物量分配模式具有一定的影响, 且对不同器官分配模式影响存在差异, 这可能与不同器官的功能有关.有些研究也指出植物生物量分配模式在季节之间的变化似乎与其季节性光合作用和水的需求的变化同时发生(
Barnes, 2002 ).由于本文所研究的3个物候期为盛花期、盛果期与营养生长期, 它们之间的差异实质上反映的是繁殖分配和营养分配之间的差异.繁殖分配作为植物生命周期中的一个重要特征, 是植物进行资源分配的一个核心问题(
Tian et al ., 2012 ), 不同的繁殖分配反映了植物在自然选择过程中的生存策略(
Reekie & Bazzaz, 2011 ), 但根据本文的研究结果可知, 繁殖生长对生物量分配模式造成的影响主要体现在相对生物量分配比例(36.00%)而不是个体大小的依赖性程度上(16.67%), 而且对S的影响较大, 但其变化趋势不一致.繁殖生长增加对Le的相对生物量分配比例, 减少对B的相对生物量分配比例, 但并没有改变他们的大小依赖性程度, 进一步证实了植物会通过调节不同器官中的生物量分配, 来实现和维持其生长最大化(
郭丽珠等, 2019 ).同时, 植物大小也是影响生殖分配的重要因素(
Niklas & Enquist, 2002 ;
Cheplick, 2005 ;
Weiner et al ., 2009 ).随着植物的生长, 两种繁殖方式的有效性也会随之发生改变, 一般而言有性繁殖对植物大小有一个阈值要求, 对于克隆植物而言, 在达到阈值之前有性繁殖后代若不能存活下来, 此时克隆繁殖产生的新分株可能成为其唯一的后代并存留下来, 但一旦达到阈值, 伴随着克隆繁殖分配的减小, 有性繁殖分配将会呈现增加的趋势(
Aarssen, 2008 ). ...
The allometry of reproduction within plant populations
1
2009
... 对于各器官不同物候期间的差异显著性检验结果表明, 物候期对白刺生物量分配模式具有一定的影响, 且对不同器官分配模式影响存在差异, 这可能与不同器官的功能有关.有些研究也指出植物生物量分配模式在季节之间的变化似乎与其季节性光合作用和水的需求的变化同时发生(
Barnes, 2002 ).由于本文所研究的3个物候期为盛花期、盛果期与营养生长期, 它们之间的差异实质上反映的是繁殖分配和营养分配之间的差异.繁殖分配作为植物生命周期中的一个重要特征, 是植物进行资源分配的一个核心问题(
Tian et al ., 2012 ), 不同的繁殖分配反映了植物在自然选择过程中的生存策略(
Reekie & Bazzaz, 2011 ), 但根据本文的研究结果可知, 繁殖生长对生物量分配模式造成的影响主要体现在相对生物量分配比例(36.00%)而不是个体大小的依赖性程度上(16.67%), 而且对S的影响较大, 但其变化趋势不一致.繁殖生长增加对Le的相对生物量分配比例, 减少对B的相对生物量分配比例, 但并没有改变他们的大小依赖性程度, 进一步证实了植物会通过调节不同器官中的生物量分配, 来实现和维持其生长最大化(
郭丽珠等, 2019 ).同时, 植物大小也是影响生殖分配的重要因素(
Niklas & Enquist, 2002 ;
Cheplick, 2005 ;
Weiner et al ., 2009 ).随着植物的生长, 两种繁殖方式的有效性也会随之发生改变, 一般而言有性繁殖对植物大小有一个阈值要求, 对于克隆植物而言, 在达到阈值之前有性繁殖后代若不能存活下来, 此时克隆繁殖产生的新分株可能成为其唯一的后代并存留下来, 但一旦达到阈值, 伴随着克隆繁殖分配的减小, 有性繁殖分配将会呈现增加的趋势(
Aarssen, 2008 ). ...
Warming experiments underpredict plant phenollogical responses to climate change
1
2012
... 近年来, 异速生长已成为研究生物量分配及权衡的重要方法.一些研究结果也表明植物的生物量分配规律可能符合异速分配理论(
Shipley & Meziane, 2002 ;
Peng & Yang, 2016 ), 表明不同器官生物量的分配比例是依赖于植物大小的.如以叶片为例, 假设其生物量分配是依赖于个体大小的, 那么随着其个体生物量的累积(
x ), 叶片的生物量累积(
y )形式已经明确, 并可以用函数ln
y =
c +
b ln
x 表示.而任何一个时期自然界的每一种植物都存在不同大小的个体, 因此, 可通过某一时期不同个体大小之间的拟合来将其异速生长方程表达出来(
Cheverud, 1982 ), 当然对于某些植物在不同年份之间的差异可归因于年龄大小造成的分配模式差异.同时, 植物在长期进化的过程中, 不同的物候期形成了一种响应环境季节性变化的生长发育阶段(如抽芽、开花、结果、落叶、休眠等), 这种生长阶段容易受到环境因子的影响(
Menzel et al ., 2006 ;
Wolkovich et al ., 2012 ), 但却是其本身生长模式对环境的一种适应.大量的研究也表明植物的不同生长发育期之间, 特别是营养生长期与繁殖生长期之间的生物量分配有着显著的差异(
Bonser & Aarssen, 2009 ;
Reekie & Bazzaz, 2011 ;
李清河等; 2012 ). ...
Distinguishing the biomass allocation variance resulting from ontogenetic drift or acclimation to soil texture
2
2012
... 作为植物最基本的生态学特性和功能性状, 生物量累积是生态系统生产力的直观体现(
Klinkhamer et al ., 1992 ).植物生物量的分配模式是植物对环境适应的结果, 并伴随着植物生活史的每一个阶段, 与植物的生长和发育相互影响(
Bonser & Aarssen, 2009 ;
Poorter et al ., 2012 ).同时, 生物量分配模式实质上也反映出植物在各器官生物量分配中的权衡问题.关于植物生物量分配机制的探讨, 一种理论认为植物倾向于将生物量分配给能够获取稀缺资源的器官以最大化适应外界环境的变化(
Fraterrigo et al ., 2006 ;
Poorter et al ., 2012 ;
Reich et al ., 2014 ), 另一种理论认为生物量分配是由植物本身的生长模式可塑性主导的(
McCarthy & Enquist, 2007 ).但事实上, 它们并非对立而是相互补充的(
Niklas, 2006 ).但不可否认的是无论生物量分配模式是为了适应外界环境的变化还是由于生物量分配模式本身的可塑性所致, 每当生物量分配模式显示部分或全部发育的异速生长增长时, 生物量分配特征将显示个体发育漂变(即, 不管环境条件如何, 生物量分配模式将随植物生长和发育而变化), 只有当生物量积累在各个组成部分之间的函数关系表现为等距关系或线性时, 才可以忽略个体发育漂变(
Gedroc et al ., 1996 ;
McConnaughay & Coleman; 1999 ;
Xie et al ., 2012 ).但大多数研究基于集中在温带森林物种的数据, 而忽视了适应干旱条件的沙漠物种的数据.生活环境的差异也必然导致植物的生长规律发生适应性的变化(
Fu et al ., 2015 ), 而植物在不同物候期的生物量分配变化尚不明确. ...
... 各器官以及整株生物量之间的异速生长关系采用常用对数(lg)转换后的值, 由R 3.5.2软件中的smatr包中的SMA函数进行标准化主轴回归(SMA)分析.该函数可根据需要对多组数据回归方程的斜率与截距进行似然比(
LR )检验与多重比较.对于不同时期SMA回归方程的斜率以及截距的显著性比较解释参照
Xie等(2012) : 若某一器官的异速生长SMA的斜率在不同时期之间具有显著性差异, 则认为其大小依赖性程度在不同时期产生了漂变, 即该器官在不同生长时期有着不同的生长轨迹.若其斜率之间不具有显著性差异, 则进一步对其截距进行差异显著性分析.若截距之间差异显著, 则认为不同时期其生物量分配的大小依赖性的变化率不变, 但相对生物量分配比例产生了变化(截距变大则相对增加了该器官的生物量分配比例, 反之, 截距变小则相对降低了该器官的生物量分配比例); 若截距之间的差异也不显著, 则认为不同时期生物量分配规律无显著变化.讨论地上部分和地下部分、同化器官和支持器官的异速生长关系时仅对比斜率之间的差异, 在此, 斜率可以表征生物量分配率, 说明在植物生物量累积的过程中, 某一器官生物量占比的变化速率.图表制作在Excel 2012中完成. ...
Ecophysiological response and morphological adjustment of two Central Asian desert shrubs towards variation in summer precipitation
1
2007
... 对白刺连续两个生长季的地上同化器官(
x 轴)与支持器官(
y 轴)生物量的SMA回归斜率比较发现, 在2016年其斜率均大于1, 表明随着个体生物量的增大其生物量向支持器官分配的比率也越大, 但随着时间的推移, 斜率整体上呈下降趋势, 且在2017年的盛花期与营养生长期均低于1, 表明其支持器官的生物量相对于同化器官生物量的变化速率在不同年份之间存在差异, 本文中白刺植株随着生长时间的推移, 会更倾向于将生物量分配给同化吸收器官而不是支持器官.
Canham等(1996) 认为, 植物对环境因子的可塑性响应是为了在资源有限的情况下能够最大限度地增长和生存.因此, 如果受到土壤水分限制, 植物必须生成更多的根系并减少叶面积, 以保持植物体内水分吸收和消耗平衡(
Deng et al. , 2006 ;
Xu et al ., 2007 ), 但是茎秆等支持器官对大部分陆生植物而言, 可能会因为其对资源获取作用不大而在资源限制下无明显响应(
Poorter & Nagel, 2000 ;
Coleman et al ., 2004 ), 本文的研究结果与之相似. ...
 ,
, ,1,2,*, 刘成功1,2, 李慧卿1,2, 孙高洁1,2
,1,2,*, 刘成功1,2, 李慧卿1,2, 孙高洁1,2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT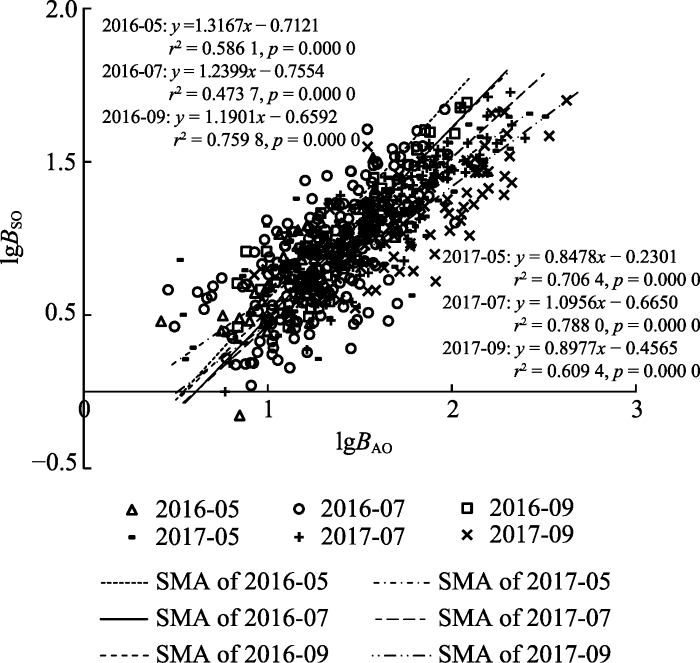 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}