Variation and correlation in functional traits of main woody plants in the Cyclobalanopsis glauca community in the karst hills of Guilin, southwest China
Run-Hong LIU1, Jin-Lian BAI2, Han BAO1,3, Juan-Li NONG1,3, Jia-Jia ZHAO1,3, Yong JIANG,1,3,*, Shi-Chu LIANG1,3, Yue-Juan LI1,31Key Laboratory of Wild Animal and Plant Ecology of Guangxi Colleges and Universities, Guangxi Normal University, Guilin, Guangxi 541006, China 2Wangfeng Experimental School, Zhongshan, Guangxi 542699, China 3College of Life Science, Guangxi Normal University, Guilin, Guangxi 541006, China
Abstract Aims Exploring the variation and the relationship between different functional traits of different growth forms and life forms woody species is helpful to understand the adaptation strategies of plants to the external environment, and is of great significance for understanding community assembly and biodiversity maintenance mechanisms. Methods We measured leaf chlorophyll content (CHL), leaf thickness (LTH), leaf area (LA), leaf dry mass (LDM), specific leaf area (SLA), leaf dry matter content (LDMC), leaf tissue density (LTD), twig dry matter content (TDMC) and twig tissue density (TTD) of 18 main woody species from the Cyclobalanopsis glauca community in karst hills of Guilin, southwest China. Traits variations among different plant functional types (growth form and life form) of woody species were analyzed by a series of methods, including the one-way analysis of variance (one-way ANOVA), and the linear mixed-effects model. In addition, the relationships between nine functional traits on individual and species levels were assessed by the Pearson’s correlation test and principal component analysis (PCA). Important findings The results showed that: (1) The nine functional traits had different degrees of variation. Specifically, LA and LDM had the maximum coefficient of intraspecific and interspecific variation, while the intraspecific and interspecific variation coefficients of TDMC and TTD were the lowest. (2) For different growth forms, there were significant differences in most functional traits between trees, shrubs and woody lianas. (3) For different life forms, except that the deciduous species showed significantly higher LA and SLA values than evergreen species, and for the other seven functional traits, evergreen species showed significantly higher values than those of deciduous species. (4) There were differences in the intraspecific and interspecific variation of functional traits between different growth forms and life forms plants. Except for some plant functional traits showing the intraspecific variation higher than interspecific variation, most of the other functional traits showed the interspecific variation was higher than intraspecific variation. (5) The relationships between nine functional traits are roughly the same at the individual level and the species level, while the significant correlation ratio at the individual level is higher than the species level. In conclusion, the interspecific variation of plant functional traits is basically higher than the intraspecific variation, but the intraspecific variation cannot be ignored. In addition, species with different growth forms and life forms adopt different ecological strategies to adapt to the karst habitat. Future research should be based on sampling at the individual level, and in combination with environmental factors to explore the variation and correlation in functional traits of different plant functional types at different scales. Keywords:karst hill;Cyclobalanopsis glauca community;plant functional traits;interspecific variability;intraspecific variability
PDF (1786KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 刘润红, 白金连, 包含, 农娟丽, 赵佳佳, 姜勇, 梁士楚, 李月娟. 桂林岩溶石山青冈群落主要木本植物功能性状变异与关联. 植物生态学报, 2020, 44(8): 828-841. DOI: 10.17521/cjpe.2019.0146 LIU Run-Hong, BAI Jin-Lian, BAO Han, NONG Juan-Li, ZHAO Jia-Jia, JIANG Yong, LIANG Shi-Chu, LI Yue-Juan. Variation and correlation in functional traits of main woody plants in the Cyclobalanopsis glauca community in the karst hills of Guilin, southwest China. Chinese Journal of Plant Ecology, 2020, 44(8): 828-841. DOI: 10.17521/cjpe.2019.0146
Table 1 表1 表1桂林岩溶石山青冈群落18种主要木本植物信息 Table 1Information about 18 main woody plant species of the Cyclobalanopsis glauca community in the karst hills of Guilin, southwest China
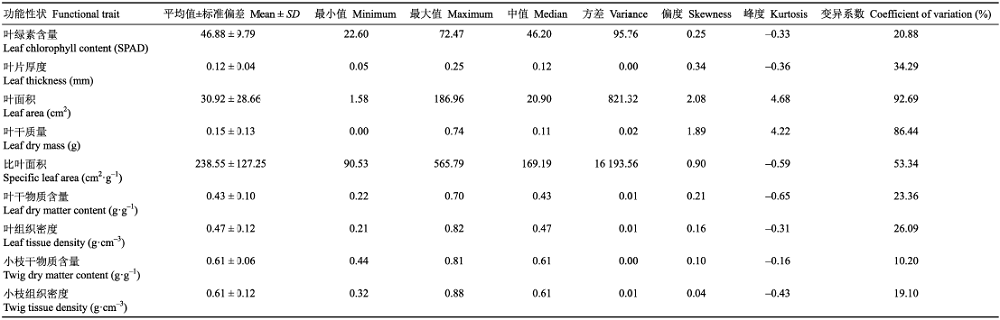
Table 2 表2 表2桂林岩溶石山青冈群落主要木本植物功能性状的描述性统计 Table 2Characteristics of plant functional traits of main woody plant species of the Cyclobalanopsis glauca community in the karst hlls of Guilin, southwest China
Table 3 表3 表3不同生长型和生活型的植物功能性状(平均值±标准偏差)及(种内/种间)变异系数 Table 3The mean (mean ± SD) and the variation coefficient (intraspecific/interspecific for plant functional traits on different growth forms and life forms
Table 4 表4 表4种内和种间变异对不同植物功能性状总体变异的贡献(%) Table 4Contributions of intraspecific and interspecific variations to the overall variation of different plant functional traits (%)
CHL
LTH
LA
LDM
SLA
LDMC
LTD
TDMC
TTD
种间 Interspecific
78.98
66.40
78.41
76.85
92.08
58.36
56.05
55.83
70.82
种内 Intraspecific
13.37
30.15
17.92
23.04
4.21
32.28
36.01
28.36
23.72
随机误差 Random error
7.65
3.45
3.67
0.11
3.71
9.36
7.94
15.81
5.46
表中各功能性状缩写同表3。 The abbreviations of plant functional traits are shown in Table 3.
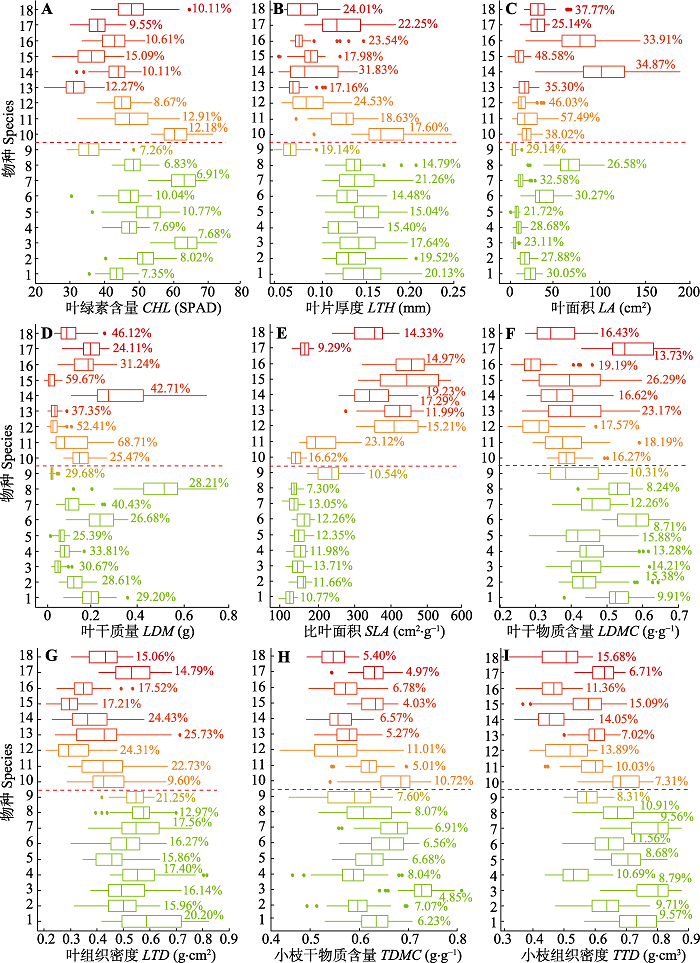
Fig. 1Boxplot of plant functional traits value for the main woody plant species of the Cyclobalanopsis glauca community in the karst hills of Guilin, southwest China. CHL, leaf chlorophyll content; LA, leaf area; LDM, leaf dry mass; LDMC, leaf dry matter content; LTD, leaf tissue density; LTH, leaf thickness; SLA, specific leaf area; TDMC, twig dry matter content; TTD, twig tissue density. The solid dots in the boxplot indicate the abnormal value; the percentage in the figure is the variation coefficient at the intraspecific level. The ordinate represents the name of the species, from the bottom to the top, the species number: 1, Cyclobalanopsis glauca; 2, Pittosporum planilobum; 3, Murraya paniculata; 4, Cinnamomum saxatile; 5, Syzygium championii; 6, Mallotus philippensis; 7, Tarenna depauperata; 8, Helicia reticulata; 9, Pterolobium punctatum; 10, Clausena dunniana; 11, Celtis biondii; 12, Radermachera sinica; 13, Fordia cauliflora; 14, Fordia cauliflora; 15, Flueggea suffruticosa; 16, Alchornea davidii; 17, Bauhinia championii; 18, Acacia concinna. 1 to 9 are evergreen species, and 10 to 18 are deciduous species.
新窗口打开|下载原图ZIP|生成PPT 图2桂林岩溶石山青冈群落主要木本植物功能性状在个体(A)和物种(B)水平上的Pearson相关系数。*, p < 0. 05; **, p < 0. 01; ***, p < 0. 001。图中各功能性状缩写同表3。
Fig. 2Pearson correlation coefficients at the individual (A) and species (B) levels between plant functional traits of the main woody plant species of the Cyclobalanopsis glauca community in the karst hills of Guilin, southwest China. The abbreviations of plant functional traits are shown in Table 3.
Fig. 3Principal component analysis (PCA) at the individual level (A) and species level (B) between plant functional traits of the main woody plant species of the Cyclobalanopsis glauca community in the karst hills of Guilin, southwest China. The abbreviations of plant functional traits are shown in Table 1. The radius of the equilibrium contribution circle (black circle in the figure) represents the average contribution rate of the variable’s vector length to the sorting space. The variables that have vectors longer than the radius of the equilibrium contribution circle are shown by red solid line, indicating that its contribution to the sorting space is greater than the average contribution of all variables.
3 讨论
3.1 植物功能性状的种内和种间变异特征
种内变异是指同种生物不同个体间的遗传或表型差异(Albert et al., 2010b)。从遗传和发育机制来看, 种内变异可能源于同种个体之间的遗传变异或不同环境条件下产生的表型可塑性(Jung et al., 2010)。本研究中各功能性状的总体种内变异幅度为6.76%- 48.30%, 种内变异平均达24.14%, 相比其他植被类型, 桂林岩溶石山青冈群落植物功能性状变异程度较低, 这与钟巧连等(2018)对黔中喀斯特木本植物功能性状变异的研究结果一致, 这说明植物功能性状变异幅度随不同植被类型及地理分布而存在较大差异, 即植物会随生境发生趋同或趋异适应, 这可能与植物对地理环境的适应策略及其资源利用能力有关, 是植物对其生存环境长期适应的结果。
Supplement I Loadings and explained variance for nine plant functional traits in the first four axes in principal component analysis (PCA) at the individual and species levels between plant functional traits of the main woody plant species of the Cyclobalanopsis glauca community in the karst hills of Guilin, southwest China
AckerlyDD, ReichPB (1999). Convergence and correlations among leaf size and function in seed plants: a comparative test using independent contrasts American Journal of Botany, 86, 1272-1281. URLPMID:10487815 Prior studies of a broad array of seed plants have reported strong correlations among leaf life span, specific leaf area, nitrogen concentration, and carbon assimilation rates, which have been interpreted as evidence of coordinated leaf physiological strategies. However, it is not known whether these relationships reflect patterns of evolutionary convergence, or whether they are due to contrasting characteristics of major seed plant lineages. We reevaluated a published data set for these seven traits measured in over 100 species, using phylogenetic independent contrasts calculated over a range of alternative seed plant phylogenies derived from recent molecular systematic analyses. In general, pairwise correlations among these seven traits were similar with and without consideration of phylogeny, and results were robust over a range of alternative phylogenies. We also evaluated relationships between these seven traits and lamina area, another important aspect of leaf function, and found moderate correlations with specific leaf area (0.64), mass-based photosynthesis (0.54), area-based nitrogen (-0.56), and leaf life span (-0.42). However, several of these correlations were markedly reduced using independent contrasts; for example, the correlation between leaf life span and lamina area was reduced to close to zero. This change reflects the large differences in both these traits between conifers and angiosperms and the absence of a relationship between the traits within these groups. This analysis illustrates that most interspecific relationships among leaf functional traits, considered across a broad range of seed plant taxa, reflect significant patterns of correlated evolutionary change, lending further support to the adaptive interpretation of these relationships.
AlbertCH, ThuillerW, YoccozNG, DouzetR, AubertS, LavorelS (2010a). A multi-trait approach reveals the structure and the relative importance of intra- vs. interspecific variability in plant traits Functional Ecology, 24, 1192-1201. [本文引用: 2]
AlbertCH, ThuillerW, YoccozNG, SoudantA, BoucherF, SacconeP, LavorelS (2010b). Intraspecific functional variability: extent, structure and sources of variation Journal of Ecology, 98, 604-613. [本文引用: 3]
AndereggWRL (2015). Spatial and temporal variation in plant hydraulic traits and their relevance for climate change impacts on vegetation New Phytologist, 205, 1008-1014. [本文引用: 1]
AugerS, ShipleyB (2013). Inter-specific and intra-specific trait variation along short environmental gradients in an old- growth temperate forest Journal of Vegetation Science, 24, 419-428.
BaralotoC, TimothyPaine CE, PoorterL, BeaucheneJ, BonalD, DomenachAM, HéraultB, Pati?oS, RoggyJC, ChaveJ (2010). Decoupled leaf and stem economics in rain forest trees Ecology Letters, 13, 1338-1347. DOI:10.1111/j.1461-0248.2010.01517.xURLPMID:20807232 [本文引用: 3] Cross-species analyses of plant functional traits have shed light on factors contributing to differences in performance and distribution, but to date most studies have focused on either leaves or stems. We extend these tissue-specific analyses of functional strategy towards a whole-plant approach by integrating data on functional traits for 13 448 leaves and wood tissues from 4672 trees representing 668 species of Neotropical trees. Strong correlations amongst traits previously defined as the leaf economics spectrum reflect a tradeoff between investments in productive leaves with rapid turnover vs. costly physical leaf structure with a long revenue stream. A second axis of variation, the 'stem economics spectrum', defines a similar tradeoff at the stem level: dense wood vs. high wood water content and thick bark. Most importantly, these two axes are orthogonal, suggesting that tradeoffs operate independently at the leaf and at the stem levels. By simplifying the multivariate ecological strategies of tropical trees into positions along these two spectra, our results provide a basis to improve global vegetation models predicting responses of tropical forests to global change.
BolmgrenK, CowanPD (2008). Time-size tradeoffs: a phylogenetic comparative study of flowering time, plant height and seed mass in a north-temperate flora Oikos, 117, 424-429. [本文引用: 1]
BucciSJ, GoldsteinG, MeinzerFC, ScholzFG, FrancoAC, BustamanteM (2004). Functional convergence in hydraulic architecture and water relations of tropical savanna trees: from leaf to whole plant Tree Physiology, 24, 891-899. DOI:10.1093/treephys/24.8.891URLPMID:15172839 [本文引用: 1] Functional convergence in hydraulic architecture and water relations, and potential trade-offs in resource allocation were investigated in six dominant neotropical savanna tree species from central Brazil during the peak of the dry season. Common relationships between wood density and several aspects of plant water relations and hydraulic architecture were observed. All species and individuals shared the same negative exponential relationship between sapwood saturated water content and wood density. Wood density was a good predictor of minimum (midday) leaf water potential and total daily transpiration, both of which decreased linearly with increasing wood density for all individuals and species. With respect to hydraulic architecture, specific and leaf-specific hydraulic conductivity decreased and the leaf:sapwood area ratio increased more than 5-fold as wood density increased from 0.37 to 0.71 g cm(-3) for all individuals and species. Wood density was also a good predictor of the temporal dynamics of water flow in stems, with the time of onset of sap flow in the morning and the maximum sap flow tending to occur progressively earlier in the day as wood density increased. Leaf properties associated with wood density included stomatal conductance, specific leaf area, and osmotic potential at the turgor loss point, which decreased linearly with increasing wood density. Wood density increased linearly with decreasing bulk soil water potential experienced by individual plants during the dry season, suggesting that wood density was greatest in individuals with mostly shallow roots, and therefore limited access to more abundant soil water at greater depths. Despite their taxonomic diversity and large intrapopulation differences in architectural traits, the six co-occurring species and their individuals shared similar functional relationships between all pairs of variables studied. Thus, rather than differing intrinsically in physiological responsiveness, the species and the individuals appeared to have distinct operating ranges along common physiological response curves dictated by plant architectural and structural features. The patterns of water uptake and access to soil water during the dry season appeared to be the main determinant of wood density, which constrained evolutionary options related to plant water economy and hydraulic architecture, leading to functional convergence in the neotropical savanna trees studied.
CadotteMW, ArnillasCA, LivingstoneSW, YasuiSLE (2015). Predicting communities from functional traits Trends in Ecology & Evolution, 30, 510-511. DOI:10.1016/j.tree.2015.07.001URLPMID:26190136 [本文引用: 1] Species traits influence where species live and how they interact. While there have been many advances in describing the functional composition and diversity of communities, only recently do researchers have the ability to predict community composition and diversity. This predictive ability can offer fundamental insights into ecosystem resilience and restoration.
Castro-DíeP, Montserrat-MartíG, CornelissenJHC (2003). Trade-offs between phenology, relative growth rate, life form and seed mass among 22 Mediterranean woody species Plant Ecology, 166, 117-129. [本文引用: 1]
ChaveJ, CoomesD, JansenS, LewisSL, SwensonNG, ZanneAE (2009). Towards a worldwide wood economics spectrum Ecology Letters, 12, 351-366. DOI:10.1111/j.1461-0248.2009.01285.xURLPMID:19243406 [本文引用: 2] Wood performs several essential functions in plants, including mechanically supporting aboveground tissue, storing water and other resources, and transporting sap. Woody tissues are likely to face physiological, structural and defensive trade-offs. How a plant optimizes among these competing functions can have major ecological implications, which have been under-appreciated by ecologists compared to the focus they have given to leaf function. To draw together our current understanding of wood function, we identify and collate data on the major wood functional traits, including the largest wood density database to date (8412 taxa), mechanical strength measures and anatomical features, as well as clade-specific features such as secondary chemistry. We then show how wood traits are related to one another, highlighting functional trade-offs, and to ecological and demographic plant features (growth form, growth rate, latitude, ecological setting). We suggest that, similar to the manifold that tree species leaf traits cluster around the 'leaf economics spectrum', a similar 'wood economics spectrum' may be defined. We then discuss the biogeography, evolution and biogeochemistry of the spectrum, and conclude by pointing out the major gaps in our current knowledge of wood functional traits.
ChengW, YuYH, XiongKN, ZhangY, XuM, TanDJ (2019). Leaf functional traits of dominant species in karst plateau- canyon areas Guihaia, 39, 1039-1049. [本文引用: 1]
dela Riva EG, TostoA, Pérez-RamosIM, Navarro-FernándezCM, OlmoM, AntenNPR, Mara?ónT, VillarR (2016). A plant economics spectrum in Mediterranean forests along environmental gradients: Is there coordination among leaf, stem and root traits? Journal of Vegetation Science, 27, 187-199. [本文引用: 1]
DíazS, KattgeJ, CornelissenJHC, WrightIJ, LavorelS, DrayS, ReuB, KleyerM, WirthC, ColinPrentice I, GarnierE, B?nischG, WestobyM, PoorterH, ReichPB, MolesAT, DickieJ, GillisonAN, ZanneAE, ChaveJ, JosephWright S, Sheremet?evSN, JactelH, BaralotoC, CeraboliniB, PierceS, ShipleyB, KirkupD, CasanovesF, JoswigJS, GüntherA, FalczukV, RügerN, MahechaMD, GornéLD (2016). The global spectrum of plant form and function Nature, 529, 167-171. DOI:10.1038/nature16489URLPMID:26700811 [本文引用: 2] Earth is home to a remarkable diversity of plant forms and life histories, yet comparatively few essential trait combinations have proved evolutionarily viable in today's terrestrial biosphere. By analysing worldwide variation in six major traits critical to growth, survival and reproduction within the largest sample of vascular plant species ever compiled, we found that occupancy of six-dimensional trait space is strongly concentrated, indicating coordination and trade-offs. Three-quarters of trait variation is captured in a two-dimensional global spectrum of plant form and function. One major dimension within this plane reflects the size of whole plants and their parts; the other represents the leaf economics spectrum, which balances leaf construction costs against growth potential. The global plant trait spectrum provides a backdrop for elucidating constraints on evolution, for functionally qualifying species and ecosystems, and for improving models that predict future vegetation based on continuous variation in plant form and function.
FunkJL, StandishRJ, StockWD, ValladaresF (2016). Plant functional traits of dominant native and invasive species in mediterranean-climate ecosystems Ecology, 97, 75-83. DOI:10.1890/15-0974.1URLPMID:27008777 [本文引用: 1] The idea that dominant invasive plant species outperform neighboring native species through higher rates of carbon assimilation and growth is supported by several analyses of global data sets. However, theory suggests that native and invasive species occurring in low-resource environments will be functionally similar, as environmental factors restrict the range of observed physiological and morphological trait values. We measured resource-use traits in native and invasive plant species across eight diverse vegetation communities distributed throughout the five mediterranean-climate regions, which are drought prone and increasingly threatened by human activities, including the introduction of exotic species. Traits differed strongly across the five regions. In regions with functional differences between native and invasive species groups, invasive species displayed traits consistent with high resource acquisition; however, these patterns were largely attributable to differences in life form. We found that species invading mediterranean-climate regions were more likely to be annual than perennial: three of the five regions were dominated by native woody species and invasive annuals. These results suggest that trait differences between native and invasive species are context dependent and will vary across vegetation communities. Native and invasive species within annual and perennial groups had similar patterns of carbon assimilation and resource use, which contradicts the widespread idea that invasive species optimize resource acquisition rather than resource conservation. .
GarnierE, LaurentG, BellmannA, DebainS, BerthelierP, DucoutB, RoumetC, NavasML (2001). Consistency of species ranking based on functional leaf traits New phytologist, 152, 69-83. [本文引用: 1]
GrimeJP (1977). Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory The American Naturalist, 111, 1169-1194. [本文引用: 1]
HeD (2016). Plant Functional Trait Variation and Community Assembly: A Case Study in A Subtropical Evergreen Forest. PhD dissertation, Sun Yat-Sen University, Guangzhou. [本文引用: 5]
JacksonBG, PeltzerDA, WardleDA (2013). The within- species leaf economic spectrum does not predict leaf litter decomposability at either the within-species or whole community levels Journal of Ecology, 101, 1409-1419. [本文引用: 1]
JiangY, ChenXB, MaJM, LiangSC, HuangJ, LiuRH, PanYF (2016). Interspecific and intraspecific variation in functional traits of subtropical evergreen and deciduous broadleaved mixed forests in karst topography, Guilin, Southwest China Tropical Conservation Science, 9, 194008291668021. DOI: 10.1177/1940082916680211. [本文引用: 2]
JungV, ViolleC, MondyC, HoffmannL, MullerS (2010). Intraspecific variability and trait-based community assembly Journal of Ecology, 98, 1134-1140. [本文引用: 2]
KangM, ChangSX, YanER, WangXH (2014). Trait variability differs between leaf and wood tissues across ecological scales in subtropical forests Journal of Vegetation Science, 25, 703-714.
LiDX, LiG, ShenZH, XuSD, HanQY, WangGF, TianFL (2017). Growth-form regulates the altitudinal variation of interspecific seed mass of woody plants in Mt. Dalaoling, the Three Gorges Region, China Chinese Journal of Plant Ecology, 41, 539-548. [本文引用: 1]
LiuHW, WangW, ZuoJ, TaoJP (2014). Leaf traits of main plants on limestone area in Zhongliang Mountain Journal of Southwest China Normal University (Natural Science Edition), 39, 50-55. [本文引用: 1]
LiuRH, LiangSC, HuangDL, HuangCY, LiJF, ChangB, JiangY (2019). Variation in functional traits of woody species across organizational scales in a riparian zone of Lijiang River, Southwest China Acta Ecologica Sinica, 39, 8038-8047. [本文引用: 2]
Méndez-AlonzoR, PazH, ZuluagaRC, RosellJA, OlsonME (2012). Coordinated evolution of leaf and stem economics in tropical dry forest trees Ecology, 93, 2397-2406. DOI:10.1890/11-1213.1URLPMID:23236911 [本文引用: 1] With data from 15 species in eight families of tropical dry forest trees, we provide evidence of coordination between the stem and leaf economic spectra. Species with low-density, flexible, breakable, hydraulically efficient but cavitationally vulnerable wood shed their leaves rapidly in response to drought and had low leaf mass per area and dry mass content. In contrast, species with the opposite xylem syndrome shed their costlier but more drought-resistant leaves late in the dry season. Our results explain variation in the timing of leaf shedding in tropical dry forests: selection eliminates combinations such as low-productivity leaves atop highly vulnerable xylem or water-greedy leaves supplied by xylem of low conductive efficiency. Across biomes, rather than a fundamental trade-off underlying a single axis of trait covariation, the relationship between leaf and stem economics is likely to occupy a wide space in which multiple combinations are possible.
MessierJ, McGillBJ, EnquistBJ, LechowiczMJ (2017). Trait variation and integration across scales: Is the leaf economic spectrum present at local scales? Ecography, 40, 685-697. [本文引用: 1]
MessierJ, McGillBJ, LechowiczMJ (2010). How do traits vary across ecological scales? A case for trait-based ecology Ecology Letters, 13, 838-848. DOI:10.1111/j.1461-0248.2010.01476.xURLPMID:20482582 [本文引用: 2] Despite the increasing importance of functional traits for the study of plant ecology, we do not know how variation in a given trait changes across ecological scales, which prevents us from assessing potential scale-dependent aspects of trait variation. To address this deficiency, we partitioned the variance in two key functional traits (leaf mass area and leaf dry matter content) across six nested ecological scales (site, plot, species, tree, strata and leaf) in lowland tropical rainforests. In both traits, the plot level shows virtually no variance despite high species turnover among plots and the size of within-species variation (leaf + strata + tree) is comparable with that of species level variation. The lack of variance at the plot level brings substantial support to the idea that trait-based environmental filtering plays a central role in plant community assembly. These results and the finding that the amount of within-species variation is comparable with interspecific variation support a shift of focus from species-based to trait-based ecology.
MouillotD, GrahamNAJ, VillégerS, MasonNWH, BellwoodDR (2013). A functional approach reveals community responses to disturbances Trends in Ecology & Evolution, 28, 167-177.
OsnasJLD, LichsteinJW, ReichPB, PacalaSW (2013). Global leaf trait relationships: mass, area, and the leaf economics spectrum Science, 340, 741-744. DOI:10.1126/science.1231574URLPMID:23539179 [本文引用: 1] The leaf economics spectrum (LES) describes multivariate correlations that constrain leaf traits of plant species primarily to a single axis of variation if data are normalized by leaf mass. We show that these traits are approximately distributed proportional to leaf area instead of mass, as expected for a light- and carbon dioxide-collecting organ. Much of the structure in the mass-normalized LES results from normalizing area-proportional traits by mass. Mass normalization induces strong correlations among area-proportional traits because of large variation among species in leaf mass per area (LMA). The high LMA variance likely reflects its functional relationship with leaf life span. A LES that is independent of mass- or area-normalization and LMA reveals physiological relationships that are inconsistent with those in global vegetation models designed to address climate change.
PangZQ, LuWL, JiangLS, JinK, QiZ (2019). Leaf traits of different growing plants in karst area of Shilin, China Guihaia, 39, 1126-1138. [本文引用: 1]
PoorterL, WrightSJ, PazH, AckerlyDD, ConditR, Ibarra-Manríouez G, HarmsKE, LiconaJC, Martínez-RamosM, MazerSJ, Muller-LandauHC, Pe?a-ClarosM, WebbCO, WrightIJ (2008). Are functional traits good predictors of demographic rates? Evidence from five neotropical forests Ecology, 89, 1908-1920. DOI:10.1890/07-0207.1URLPMID:18705377 [本文引用: 1] A central goal of comparative plant ecology is to understand how functional traits vary among species and to what extent this variation has adaptive value. Here we evaluate relationships between four functional traits (seed volume, specific leaf area, wood density, and adult stature) and two demographic attributes (diameter growth and tree mortality) for large trees of 240 tree species from five Neotropical forests. We evaluate how these key functional traits are related to survival and growth and whether similar relationships between traits and demography hold across different tropical forests. There was a tendency for a trade-off between growth and survival across rain forest tree species. Wood density, seed volume, and adult stature were significant predictors of growth and/or mortality. Both growth and mortality rates declined with an increase in wood density. This is consistent with greater construction costs and greater resistance to stem damage for denser wood. Growth and mortality rates also declined as seed volume increased. This is consistent with an adaptive syndrome in which species tolerant of low resource availability (in this case shade-tolerant species) have large seeds to establish successfully and low inherent growth and mortality rates. Growth increased and mortality decreased with an increase in adult stature, because taller species have a greater access to light and longer life spans. Specific leaf area was, surprisingly, only modestly informative for the performance of large trees and had ambiguous relationships with growth and survival. Single traits accounted for 9-55% of the interspecific variation in growth and mortality rates at individual sites. Significant correlations with demographic rates tended to be similar across forests and for phylogenetically independent contrasts as well as for cross-species analyses that treated each species as an independent observation. In combination, the morphological traits explained 41% of the variation in growth rate and 54% of the variation in mortality rate, with wood density being the best predictor of growth and mortality. Relationships between functional traits and demographic rates were statistically similar across a wide range of Neotropical forests. The consistency of these results strongly suggests that tropical rain forest species face similar trade-offs in different sites and converge on similar sets of solutions.
ReichPB, WaltersMB, EllsworthDS (1992). Leaf life-span in relation to leaf, plant, and stand characteristics among diverse ecosystems Ecological Monographs, 62, 365-392. [本文引用: 1]
RoweN, SpeckT (2005). Plant growth forms: an ecological and evolutionary perspective New Phytologist, 166, 61-72. [本文引用: 1]
SantiagoLS, WrightSJ (2007). Leaf functional traits of tropical forest plants in relation to growth form Functional Ecology, 21, 19-27. [本文引用: 1]
SiefertA, ViolleC, ChalmandrierL, AlbertCH, TaudiereA, FajardoA, AarssenLW, BaralotoC, CarlucciMB, CianciarusoMV, deL Dantas V, deBello F, DuarteLDS, FonsecaCR, FreschetGT, GaucherandS, GrossN, HikosakaK, JacksonB, JungV, KamiyamaC, KatabuchiM, KembelSW, KicheninE, KraftNJB, Lagerstr?mA, LeBagousse-Pinguet Y, LiY, MasonN, MessierJ, NakashizukaT, OvertonJM, PeltzerDA, Pérez-RamosIM, PillarVD, PrenticeHC, RichardsonS, SasakiT, SchampBS, Sch?bC, ShipleyB, SundqvistM, SykesMT, VandewalleM, WardleDA (2015). A global meta-analysis of the relative extent of intraspecific trait variation in plant communities Ecology Letters, 18, 1406-1419. DOI:10.1111/ele.12508URLPMID:26415616 [本文引用: 4] Recent studies have shown that accounting for intraspecific trait variation (ITV) may better address major questions in community ecology. However, a general picture of the relative extent of ITV compared to interspecific trait variation in plant communities is still missing. Here, we conducted a meta-analysis of the relative extent of ITV within and among plant communities worldwide, using a data set encompassing 629 communities (plots) and 36 functional traits. Overall, ITV accounted for 25% of the total trait variation within communities and 32% of the total trait variation among communities on average. The relative extent of ITV tended to be greater for whole-plant (e.g. plant height) vs. organ-level traits and for leaf chemical (e.g. leaf N and P concentration) vs. leaf morphological (e.g. leaf area and thickness) traits. The relative amount of ITV decreased with increasing species richness and spatial extent, but did not vary with plant growth form or climate. These results highlight global patterns in the relative importance of ITV in plant communities, providing practical guidelines for when researchers should include ITV in trait-based community and ecosystem studies.
SilvertownJ (2004). Plant coexistence and the niche Trends in Ecology & Evolution, 19, 605-611. [本文引用: 1]
SpasojevicMJ, TurnerBL, MyersJA (2016). When does intraspecific trait variation contribute to functional beta- diversity? Journal of Ecology, 104, 487-496. [本文引用: 1]
TangQQ, HuangYT, DingY, ZangRG (2016). Interspecific and intraspecific variation in functional traits of subtropical evergreen and deciduous broad-leaved mixed forests Biodiversity Science, 24, 262-270. DOI:10.17520/biods.2015200URL [本文引用: 2]
TomlinsonKW, PoorterL, BongersF, BorghettiF, JacobsL, vanLangevelde F (2014). Relative growth rate variation of evergreen and deciduous savanna tree species is driven by different traits Annals of Botany, 114, 315-324. DOI:10.1093/aob/mcu107URLPMID:24958787 [本文引用: 1] BACKGROUND AND AIMS: Plant relative growth rate (RGR) depends on biomass allocation to leaves (leaf mass fraction, LMF), efficient construction of leaf surface area (specific leaf area, SLA) and biomass growth per unit leaf area (net assimilation rate, NAR). Functional groups of species may differ in any of these traits, potentially resulting in (1) differences in mean RGR of groups, and (2) differences in the traits driving RGR variation within each group. We tested these predictions by comparing deciduous and evergreen savanna trees. METHODS: RGR, changes to biomass allocation and leaf morphology, and root non-structural carbohydrate reserves were evaluated for juveniles of 51 savanna species (34 deciduous, 17 evergreen) grown in a common garden experiment. It was anticipated that drivers of RGR would differ between leaf habit groups because deciduous species have to allocate carbohydrates to storage in roots to be able to flush leaves again, which directly compromises their LMF, whereas evergreen species are not subject to this constraint. KEY RESULTS: Evergreen species had greater LMF and RGR than deciduous species. Among deciduous species LMF explained 27 % of RGR variation (SLA 34 % and NAR 29 %), whereas among evergreen species LMF explained between 2 and 17 % of RGR variation (SLA 32-35 % and NAR 38-62 %). RGR and LMF were (negatively) related to carbohydrate storage only among deciduous species. CONCLUSIONS: Trade-offs between investment in carbohydrate reserves and growth occurred only among deciduous species, leading to differences in relative contribution made by the underlying components of RGR between the leaf habit groups. The results suggest that differences in drivers of RGR occur among savanna species because these have different selected strategies for coping with fire disturbance in savannas. It is expected that variation in the drivers of RGR will be found in other functional types that respond differently to particular disturbances.
ViolleC, EnquistBJ, McGillBJ, JiangL, AlbertCH, HulshofC, JungV, MessierJ (2012). The return of the variance: intraspecific variability in community ecology Trends in Ecology & Evolution, 27, 244-252. DOI:10.1016/j.tree.2011.11.014URLPMID:22244797 [本文引用: 1] Despite being recognized as a promoter of diversity and a condition for local coexistence decades ago, the importance of intraspecific variance has been neglected over time in community ecology. Recently, there has been a new emphasis on intraspecific variability. Indeed, recent developments in trait-based community ecology have underlined the need to integrate variation at both the intraspecific as well as interspecific level. We introduce new T-statistics ('T' for trait), based on the comparison of intraspecific and interspecific variances of functional traits across organizational levels, to operationally incorporate intraspecific variability into community ecology theory. We show that a focus on the distribution of traits at local and regional scales combined with original analytical tools can provide unique insights into the primary forces structuring communities.
ViolleC, NavasML, VileD, KazakouE, FortunelC, HummelI, GarnierE (2007). Let the concept of trait be functional! Oikos, 116, 882-892. [本文引用: 1]
WangKL, ChenHS, ZengFP, YueYM, ZhangW, FuZY (2018). Ecological research supports eco-environmental management and poverty alleviation in karst region of southwest China Bulletin of the Chinese Academy of Sciences, 33, 213-222. [本文引用: 1]
WeemstraM, MommerL, VisserEJW, vanRuijven J, KuyperTW, MohrenGMJ, SterckFJ (2016). Towards a multidimensional root trait framework: a tree root review New Phytologist, 211, 1159-1169. [本文引用: 1]
WestobyM (1998). A leaf-height-seed (LHS) plant ecology strategy scheme Plant and Soil, 199, 213-227. [本文引用: 1]
WrightIJ, AckerlyDD, BongersF, HarmsKE, Ibarra-ManriquezG, Martinez-RamosM, MazerSJ, Muller-LandauHC, PazH, PitmanNCA, PoorterL, SilmanMR, VriesendorpCF, WebbCO, WestobyM, WrightSJ (2007). Relationships among ecologically important dimensions of plant trait variation in seven Neotropical forests Annals of Botany, 99, 1003-1015. DOI:10.1093/aob/mcl066URLPMID:16595553 [本文引用: 2] BACKGROUND AND AIMS: When ecologically important plant traits are correlated they may be said to constitute an ecological 'strategy' dimension. Through identifying these dimensions and understanding their inter-relationships we gain insight into why particular trait combinations are favoured over others and into the implications of trait differences among species. Here we investigated relationships among several traits, and thus the strategy dimensions they represented, across 2134 woody species from seven Neotropical forests. METHODS: Six traits were studied: specific leaf area (SLA), the average size of leaves, seed and fruit, typical maximum plant height, and wood density (WD). Trait relationships were quantified across species at each individual forest as well as across the dataset as a whole. 'Phylogenetic' analyses were used to test for correlations among evolutionary trait-divergences and to ascertain whether interspecific relationships were biased by strong taxonomic patterning in the traits. KEY RESULTS: The interspecific and phylogenetic analyses yielded congruent results. Seed and fruit size were expected, and confirmed, to be tightly related. As expected, plant height was correlated with each of seed and fruit size, albeit weakly. Weak support was found for an expected positive relationship between leaf and fruit size. The prediction that SLA and WD would be negatively correlated was not supported. Otherwise the traits were predicted to be largely unrelated, being representatives of putatively independent strategy dimensions. This was indeed the case, although WD was consistently, negatively related to leaf size. CONCLUSIONS: The dimensions represented by SLA, seed/fruit size and leaf size were essentially independent and thus conveyed largely independent information about plant strategies. To a lesser extent the same was true for plant height and WD. Our tentative explanation for negative WD-leaf size relationships, now also known from other habitats, is that the traits are indirectly linked via plant hydraulics.
WrightIJ, FalsterDS, PickupM, WestobyM (2006). Cross- species patterns in the coordination between leaf and stem traits, and their implications for plant hydraulics Physiologia Plantarum, 127, 445-456. [本文引用: 1]
WrightIJ, ReichPB, WestobyM, AckerlyDD, BaruchZ, BongersF, Cavender-BaresJ, ChapinT, CornelissenJHC, DiemerM, FlexasJ, GarnierE, GroomPK, GuliasJ, HikosakaK, LamontBB, LeeT, LeeW, LuskC, MidgleyJJ, NavasML, Niinemetsü, OleksynJ, OsadaN, PoorterH, PootP, PriorL, PyankovVI, RoumetC, ThomasSC, TjoelkerMG, VeneklaasEJ, VillarR (2004). The worldwide leaf economics spectrum Nature, 428, 821-827. DOI:10.1038/nature02403URLPMID:15103368 [本文引用: 1] Bringing together leaf trait data spanning 2,548 species and 175 sites we describe, for the first time at global scale, a universal spectrum of leaf economics consisting of key chemical, structural and physiological properties. The spectrum runs from quick to slow return on investments of nutrients and dry mass in leaves, and operates largely independently of growth form, plant functional type or biome. Categories along the spectrum would, in general, describe leaf economic variation at the global scale better than plant functional types, because functional types overlap substantially in their leaf traits. Overall, modulation of leaf traits and trait relationships by climate is surprisingly modest, although some striking and significant patterns can be seen. Reliable quantification of the leaf economics spectrum and its interaction with climate will prove valuable for modelling nutrient fluxes and vegetation boundaries under changing land-use and climate.
ZhongQL, LiuLB, XuX, YangY, GuoYM, XuHY, CaiXL, NiJ (2018). Variations of plant functional traits and adaptive strategy of woody species in a karst forest of central Guizhou Province, Southwestern China Chinese Journal of Plant Ecology, 42, 562-572. [本文引用: 3]
Convergence and correlations among leaf size and function in seed plants: a comparative test using independent contrasts 1999
A multi-trait approach reveals the structure and the relative importance of intra- vs. interspecific variability in plant traits 2 2010
... 在自然群落中, 物种间性状的差异被认为是物种共存的重要前提(Díaz et al., 2004), 故先前大多数基于功能性状的群落生态学研究仅考虑种间性状变异, 即采用群落中少数个体的性状平均值来代表物种性状值, 而忽略种内变异, 这隐含假设条件: 种间性状变异程度远大于种内变异程度, 种内变异相比种间变异可以忽略不计(Díaz et al., 2004; Jackson et al., 2013).然而, 最近十余年的研究表明, 种内变异虽然通常小于种间变异, 但是其可达性状总体变异的28%-52%, 对植物群落内和群落间的总体变异具有重要作用(Albert et al., 2010b; Messier et al., 2010; Siefert et al., 2015).种内变异在功能性状研究中的重要性取决于所选择的生态系统、功能性状、物种和研究目的(Albert et al., 2010a), 以及研究的时间尺度(Anderegg, 2015)、空间尺度(Spasojevic et al., 2016; Chalmandrier et al., 2017)、生态尺度(Messier et al., 2017; 刘润红, 2018)等因素.具体来说, 随着空间尺度增大, 不同区域之间存在较大的环境差异导致群落中物种组成存在显著差异, 种间性状变异能够在很大的程度上解释物种多样性与环境因子间的关系, 种间性状变异相对于种内变异有可能逐渐凸显, 因此在区域尺度以上的大空间尺度研究中, 忽略种内变异可能是合理的(Violle et al., 2012; Díaz et al., 2016).但是在小尺度(如种群尺度等)或者局域环境下, 尤其是在物种丰富度较低的群落中, 种内性状变异对于群落物种共存具有重要意义, 不容忽略(Siefert et al., 2015).就不同功能性状而言, 叶片化学性状(如叶片氮和磷含量)的种内变异比叶片形态性状(如叶片面积和叶片厚度)的种内变异大; 植物整株性状(如株高等)的种内变异比器官水平性状(叶、茎和根性状等)的种内变异大(Siefert et al., 2015).此外, 不同生态系统的植物功能性状变异也存在一定的差异, 种内变异程度在受环境胁迫和物种数量较低的群落中更大(Siefert et al., 2015).因此, 只有结合种间和种内性状变异才能够准确地揭示群落构建和生物多样性维持机制(Albert et al., 2010b; Jung et al., 2010; Messier et al., 2010; 刘润红等, 2019). ...
... 植物在其漫长的历史进化和发展过程中, 通过内部不同功能性状之间的协同进化与权衡来适应变化的外界环境(Silvertown, 2004).当两个或多个重要植物功能性状在不同物种间都存在一致的相关性时, 这种性状组合关系可以认为是一个策略维度, 也称“生态策略”, 即植物沿一定的生态策略轴排列于最适应或最具竞争力的位置(Wright et al., 2006, 2007; 刘晓娟和马克平, 2015).因此, 许多研究在性状变异的多维空间里甄别出少数几个能够充分描述物种间功能性状变异的独立轴主轴来代表生态策略的分化方向, 如叶片-株高-种子(leaf-height-seed, LHS)权衡(Westoby, 1998)、叶经济型谱(leaf economic spectrum, LES)(Wright et al., 2004)、木材经济型谱(wood economic spectrum, WES)(Chave et al., 2009)和根经济型谱(root economic spectrum, RES)(Weemstra et al., 2016)等.值得注意的是, 上述功能性状的协变与权衡格局基本是在大尺度上确立的, 然而, 植物在进化过程中会面临自然选择或对环境胁迫的适应, 特定区域内不同性状之间的关系可能会与全球尺度或区域尺度的变化不一致, 反映出植物对特殊生境的适应(Santiago & Wright, 2007).因此, 还需要在不同尺度(如局域植物群落内部共存物种之间、种内或个体水平)上验证功能性状之间的这种协变与权衡关系是否仍然成立(Albert et al., 2010a; 何东, 2016). ...
Intraspecific functional variability: extent, structure and sources of variation 3 2010
... 在自然群落中, 物种间性状的差异被认为是物种共存的重要前提(Díaz et al., 2004), 故先前大多数基于功能性状的群落生态学研究仅考虑种间性状变异, 即采用群落中少数个体的性状平均值来代表物种性状值, 而忽略种内变异, 这隐含假设条件: 种间性状变异程度远大于种内变异程度, 种内变异相比种间变异可以忽略不计(Díaz et al., 2004; Jackson et al., 2013).然而, 最近十余年的研究表明, 种内变异虽然通常小于种间变异, 但是其可达性状总体变异的28%-52%, 对植物群落内和群落间的总体变异具有重要作用(Albert et al., 2010b; Messier et al., 2010; Siefert et al., 2015).种内变异在功能性状研究中的重要性取决于所选择的生态系统、功能性状、物种和研究目的(Albert et al., 2010a), 以及研究的时间尺度(Anderegg, 2015)、空间尺度(Spasojevic et al., 2016; Chalmandrier et al., 2017)、生态尺度(Messier et al., 2017; 刘润红, 2018)等因素.具体来说, 随着空间尺度增大, 不同区域之间存在较大的环境差异导致群落中物种组成存在显著差异, 种间性状变异能够在很大的程度上解释物种多样性与环境因子间的关系, 种间性状变异相对于种内变异有可能逐渐凸显, 因此在区域尺度以上的大空间尺度研究中, 忽略种内变异可能是合理的(Violle et al., 2012; Díaz et al., 2016).但是在小尺度(如种群尺度等)或者局域环境下, 尤其是在物种丰富度较低的群落中, 种内性状变异对于群落物种共存具有重要意义, 不容忽略(Siefert et al., 2015).就不同功能性状而言, 叶片化学性状(如叶片氮和磷含量)的种内变异比叶片形态性状(如叶片面积和叶片厚度)的种内变异大; 植物整株性状(如株高等)的种内变异比器官水平性状(叶、茎和根性状等)的种内变异大(Siefert et al., 2015).此外, 不同生态系统的植物功能性状变异也存在一定的差异, 种内变异程度在受环境胁迫和物种数量较低的群落中更大(Siefert et al., 2015).因此, 只有结合种间和种内性状变异才能够准确地揭示群落构建和生物多样性维持机制(Albert et al., 2010b; Jung et al., 2010; Messier et al., 2010; 刘润红等, 2019). ...
... ).因此, 只有结合种间和种内性状变异才能够准确地揭示群落构建和生物多样性维持机制(Albert et al., 2010b; Jung et al., 2010; Messier et al., 2010; 刘润红等, 2019). ...
... 种内变异是指同种生物不同个体间的遗传或表型差异(Albert et al., 2010b).从遗传和发育机制来看, 种内变异可能源于同种个体之间的遗传变异或不同环境条件下产生的表型可塑性(Jung et al., 2010).本研究中各功能性状的总体种内变异幅度为6.76%- 48.30%, 种内变异平均达24.14%, 相比其他植被类型, 桂林岩溶石山青冈群落植物功能性状变异程度较低, 这与钟巧连等(2018)对黔中喀斯特木本植物功能性状变异的研究结果一致, 这说明植物功能性状变异幅度随不同植被类型及地理分布而存在较大差异, 即植物会随生境发生趋同或趋异适应, 这可能与植物对地理环境的适应策略及其资源利用能力有关, 是植物对其生存环境长期适应的结果. ...
Spatial and temporal variation in plant hydraulic traits and their relevance for climate change impacts on vegetation 1 2015
... 在自然群落中, 物种间性状的差异被认为是物种共存的重要前提(Díaz et al., 2004), 故先前大多数基于功能性状的群落生态学研究仅考虑种间性状变异, 即采用群落中少数个体的性状平均值来代表物种性状值, 而忽略种内变异, 这隐含假设条件: 种间性状变异程度远大于种内变异程度, 种内变异相比种间变异可以忽略不计(Díaz et al., 2004; Jackson et al., 2013).然而, 最近十余年的研究表明, 种内变异虽然通常小于种间变异, 但是其可达性状总体变异的28%-52%, 对植物群落内和群落间的总体变异具有重要作用(Albert et al., 2010b; Messier et al., 2010; Siefert et al., 2015).种内变异在功能性状研究中的重要性取决于所选择的生态系统、功能性状、物种和研究目的(Albert et al., 2010a), 以及研究的时间尺度(Anderegg, 2015)、空间尺度(Spasojevic et al., 2016; Chalmandrier et al., 2017)、生态尺度(Messier et al., 2017; 刘润红, 2018)等因素.具体来说, 随着空间尺度增大, 不同区域之间存在较大的环境差异导致群落中物种组成存在显著差异, 种间性状变异能够在很大的程度上解释物种多样性与环境因子间的关系, 种间性状变异相对于种内变异有可能逐渐凸显, 因此在区域尺度以上的大空间尺度研究中, 忽略种内变异可能是合理的(Violle et al., 2012; Díaz et al., 2016).但是在小尺度(如种群尺度等)或者局域环境下, 尤其是在物种丰富度较低的群落中, 种内性状变异对于群落物种共存具有重要意义, 不容忽略(Siefert et al., 2015).就不同功能性状而言, 叶片化学性状(如叶片氮和磷含量)的种内变异比叶片形态性状(如叶片面积和叶片厚度)的种内变异大; 植物整株性状(如株高等)的种内变异比器官水平性状(叶、茎和根性状等)的种内变异大(Siefert et al., 2015).此外, 不同生态系统的植物功能性状变异也存在一定的差异, 种内变异程度在受环境胁迫和物种数量较低的群落中更大(Siefert et al., 2015).因此, 只有结合种间和种内性状变异才能够准确地揭示群落构建和生物多样性维持机制(Albert et al., 2010b; Jung et al., 2010; Messier et al., 2010; 刘润红等, 2019). ...
Inter-specific and intra-specific trait variation along short environmental gradients in an old- growth temperate forest 2013
Decoupled leaf and stem economics in rain forest trees 3 2010
... 本研究发现叶经济型谱性状SLA与LTH、LDMC及LTD呈显著负相关关系, 小枝性状TTD和TDMC则呈高度正相关关系, 而LA在一定程度与小枝性状TTD和TDMC呈负相关关系, 这同先前大多数研究(Bucci et al., 2004; Wright et al., 2007; Baraloto et al., 2010; de la Riva et al., 2016; Díaz et al., 2016; 何东, 2016)的结果相似.SLA反映了植物对光照、水分和养分等资源的获取和利用能力, 与植物的生存策略密切相关(Garnier et al., 2001).通常, SLA较低的植物, 其叶片光合作用产物多投资于构建保卫细胞或增加叶肉细胞密度, 即增加叶片内部水分向叶片表面扩散的距离或阻力(王瑞丽等, 2016), 其LTH、LDMC和LTD也会相应地增加, 以减少蒸腾造成的水分散失, 提高水分利用效率, 从而具有较强的抵抗干旱胁迫和土壤贫瘠生境的能力, 提高其对环境的适合度(程雯等, 2019; 庞志强等, 2019).而SLA与LA在物种尺度上无显著相关性, PCA分析也发现, LA和SLA正交, 这说明两个性状属于不同的生态策略维度. ...
Spatial scale and intraspecific trait variability mediate assembly rules in alpine grasslands 1 2017
... 在自然群落中, 物种间性状的差异被认为是物种共存的重要前提(Díaz et al., 2004), 故先前大多数基于功能性状的群落生态学研究仅考虑种间性状变异, 即采用群落中少数个体的性状平均值来代表物种性状值, 而忽略种内变异, 这隐含假设条件: 种间性状变异程度远大于种内变异程度, 种内变异相比种间变异可以忽略不计(Díaz et al., 2004; Jackson et al., 2013).然而, 最近十余年的研究表明, 种内变异虽然通常小于种间变异, 但是其可达性状总体变异的28%-52%, 对植物群落内和群落间的总体变异具有重要作用(Albert et al., 2010b; Messier et al., 2010; Siefert et al., 2015).种内变异在功能性状研究中的重要性取决于所选择的生态系统、功能性状、物种和研究目的(Albert et al., 2010a), 以及研究的时间尺度(Anderegg, 2015)、空间尺度(Spasojevic et al., 2016; Chalmandrier et al., 2017)、生态尺度(Messier et al., 2017; 刘润红, 2018)等因素.具体来说, 随着空间尺度增大, 不同区域之间存在较大的环境差异导致群落中物种组成存在显著差异, 种间性状变异能够在很大的程度上解释物种多样性与环境因子间的关系, 种间性状变异相对于种内变异有可能逐渐凸显, 因此在区域尺度以上的大空间尺度研究中, 忽略种内变异可能是合理的(Violle et al., 2012; Díaz et al., 2016).但是在小尺度(如种群尺度等)或者局域环境下, 尤其是在物种丰富度较低的群落中, 种内性状变异对于群落物种共存具有重要意义, 不容忽略(Siefert et al., 2015).就不同功能性状而言, 叶片化学性状(如叶片氮和磷含量)的种内变异比叶片形态性状(如叶片面积和叶片厚度)的种内变异大; 植物整株性状(如株高等)的种内变异比器官水平性状(叶、茎和根性状等)的种内变异大(Siefert et al., 2015).此外, 不同生态系统的植物功能性状变异也存在一定的差异, 种内变异程度在受环境胁迫和物种数量较低的群落中更大(Siefert et al., 2015).因此, 只有结合种间和种内性状变异才能够准确地揭示群落构建和生物多样性维持机制(Albert et al., 2010b; Jung et al., 2010; Messier et al., 2010; 刘润红等, 2019). ...
Towards a worldwide wood economics spectrum 2 2009
... 植物在其漫长的历史进化和发展过程中, 通过内部不同功能性状之间的协同进化与权衡来适应变化的外界环境(Silvertown, 2004).当两个或多个重要植物功能性状在不同物种间都存在一致的相关性时, 这种性状组合关系可以认为是一个策略维度, 也称“生态策略”, 即植物沿一定的生态策略轴排列于最适应或最具竞争力的位置(Wright et al., 2006, 2007; 刘晓娟和马克平, 2015).因此, 许多研究在性状变异的多维空间里甄别出少数几个能够充分描述物种间功能性状变异的独立轴主轴来代表生态策略的分化方向, 如叶片-株高-种子(leaf-height-seed, LHS)权衡(Westoby, 1998)、叶经济型谱(leaf economic spectrum, LES)(Wright et al., 2004)、木材经济型谱(wood economic spectrum, WES)(Chave et al., 2009)和根经济型谱(root economic spectrum, RES)(Weemstra et al., 2016)等.值得注意的是, 上述功能性状的协变与权衡格局基本是在大尺度上确立的, 然而, 植物在进化过程中会面临自然选择或对环境胁迫的适应, 特定区域内不同性状之间的关系可能会与全球尺度或区域尺度的变化不一致, 反映出植物对特殊生境的适应(Santiago & Wright, 2007).因此, 还需要在不同尺度(如局域植物群落内部共存物种之间、种内或个体水平)上验证功能性状之间的这种协变与权衡关系是否仍然成立(Albert et al., 2010a; 何东, 2016). ...
... 本研究发现叶经济型谱性状SLA与LTH、LDMC及LTD呈显著负相关关系, 小枝性状TTD和TDMC则呈高度正相关关系, 而LA在一定程度与小枝性状TTD和TDMC呈负相关关系, 这同先前大多数研究(Bucci et al., 2004; Wright et al., 2007; Baraloto et al., 2010; de la Riva et al., 2016; Díaz et al., 2016; 何东, 2016)的结果相似.SLA反映了植物对光照、水分和养分等资源的获取和利用能力, 与植物的生存策略密切相关(Garnier et al., 2001).通常, SLA较低的植物, 其叶片光合作用产物多投资于构建保卫细胞或增加叶肉细胞密度, 即增加叶片内部水分向叶片表面扩散的距离或阻力(王瑞丽等, 2016), 其LTH、LDMC和LTD也会相应地增加, 以减少蒸腾造成的水分散失, 提高水分利用效率, 从而具有较强的抵抗干旱胁迫和土壤贫瘠生境的能力, 提高其对环境的适合度(程雯等, 2019; 庞志强等, 2019).而SLA与LA在物种尺度上无显著相关性, PCA分析也发现, LA和SLA正交, 这说明两个性状属于不同的生态策略维度. ...
喀斯特高原峡谷优势种叶片功能性状分析 1 2019
... 本研究发现叶经济型谱性状SLA与LTH、LDMC及LTD呈显著负相关关系, 小枝性状TTD和TDMC则呈高度正相关关系, 而LA在一定程度与小枝性状TTD和TDMC呈负相关关系, 这同先前大多数研究(Bucci et al., 2004; Wright et al., 2007; Baraloto et al., 2010; de la Riva et al., 2016; Díaz et al., 2016; 何东, 2016)的结果相似.SLA反映了植物对光照、水分和养分等资源的获取和利用能力, 与植物的生存策略密切相关(Garnier et al., 2001).通常, SLA较低的植物, 其叶片光合作用产物多投资于构建保卫细胞或增加叶肉细胞密度, 即增加叶片内部水分向叶片表面扩散的距离或阻力(王瑞丽等, 2016), 其LTH、LDMC和LTD也会相应地增加, 以减少蒸腾造成的水分散失, 提高水分利用效率, 从而具有较强的抵抗干旱胁迫和土壤贫瘠生境的能力, 提高其对环境的适合度(程雯等, 2019; 庞志强等, 2019).而SLA与LA在物种尺度上无显著相关性, PCA分析也发现, LA和SLA正交, 这说明两个性状属于不同的生态策略维度. ...
A plant economics spectrum in Mediterranean forests along environmental gradients: Is there coordination among leaf, stem and root traits? 1 2016
... 本研究发现叶经济型谱性状SLA与LTH、LDMC及LTD呈显著负相关关系, 小枝性状TTD和TDMC则呈高度正相关关系, 而LA在一定程度与小枝性状TTD和TDMC呈负相关关系, 这同先前大多数研究(Bucci et al., 2004; Wright et al., 2007; Baraloto et al., 2010; de la Riva et al., 2016; Díaz et al., 2016; 何东, 2016)的结果相似.SLA反映了植物对光照、水分和养分等资源的获取和利用能力, 与植物的生存策略密切相关(Garnier et al., 2001).通常, SLA较低的植物, 其叶片光合作用产物多投资于构建保卫细胞或增加叶肉细胞密度, 即增加叶片内部水分向叶片表面扩散的距离或阻力(王瑞丽等, 2016), 其LTH、LDMC和LTD也会相应地增加, 以减少蒸腾造成的水分散失, 提高水分利用效率, 从而具有较强的抵抗干旱胁迫和土壤贫瘠生境的能力, 提高其对环境的适合度(程雯等, 2019; 庞志强等, 2019).而SLA与LA在物种尺度上无显著相关性, PCA分析也发现, LA和SLA正交, 这说明两个性状属于不同的生态策略维度. ...
The plant traits that drive ecosystems: evidence from three continents 2 2004
... 在自然群落中, 物种间性状的差异被认为是物种共存的重要前提(Díaz et al., 2004), 故先前大多数基于功能性状的群落生态学研究仅考虑种间性状变异, 即采用群落中少数个体的性状平均值来代表物种性状值, 而忽略种内变异, 这隐含假设条件: 种间性状变异程度远大于种内变异程度, 种内变异相比种间变异可以忽略不计(Díaz et al., 2004; Jackson et al., 2013).然而, 最近十余年的研究表明, 种内变异虽然通常小于种间变异, 但是其可达性状总体变异的28%-52%, 对植物群落内和群落间的总体变异具有重要作用(Albert et al., 2010b; Messier et al., 2010; Siefert et al., 2015).种内变异在功能性状研究中的重要性取决于所选择的生态系统、功能性状、物种和研究目的(Albert et al., 2010a), 以及研究的时间尺度(Anderegg, 2015)、空间尺度(Spasojevic et al., 2016; Chalmandrier et al., 2017)、生态尺度(Messier et al., 2017; 刘润红, 2018)等因素.具体来说, 随着空间尺度增大, 不同区域之间存在较大的环境差异导致群落中物种组成存在显著差异, 种间性状变异能够在很大的程度上解释物种多样性与环境因子间的关系, 种间性状变异相对于种内变异有可能逐渐凸显, 因此在区域尺度以上的大空间尺度研究中, 忽略种内变异可能是合理的(Violle et al., 2012; Díaz et al., 2016).但是在小尺度(如种群尺度等)或者局域环境下, 尤其是在物种丰富度较低的群落中, 种内性状变异对于群落物种共存具有重要意义, 不容忽略(Siefert et al., 2015).就不同功能性状而言, 叶片化学性状(如叶片氮和磷含量)的种内变异比叶片形态性状(如叶片面积和叶片厚度)的种内变异大; 植物整株性状(如株高等)的种内变异比器官水平性状(叶、茎和根性状等)的种内变异大(Siefert et al., 2015).此外, 不同生态系统的植物功能性状变异也存在一定的差异, 种内变异程度在受环境胁迫和物种数量较低的群落中更大(Siefert et al., 2015).因此, 只有结合种间和种内性状变异才能够准确地揭示群落构建和生物多样性维持机制(Albert et al., 2010b; Jung et al., 2010; Messier et al., 2010; 刘润红等, 2019). ...
... ), 故先前大多数基于功能性状的群落生态学研究仅考虑种间性状变异, 即采用群落中少数个体的性状平均值来代表物种性状值, 而忽略种内变异, 这隐含假设条件: 种间性状变异程度远大于种内变异程度, 种内变异相比种间变异可以忽略不计(Díaz et al., 2004; Jackson et al., 2013).然而, 最近十余年的研究表明, 种内变异虽然通常小于种间变异, 但是其可达性状总体变异的28%-52%, 对植物群落内和群落间的总体变异具有重要作用(Albert et al., 2010b; Messier et al., 2010; Siefert et al., 2015).种内变异在功能性状研究中的重要性取决于所选择的生态系统、功能性状、物种和研究目的(Albert et al., 2010a), 以及研究的时间尺度(Anderegg, 2015)、空间尺度(Spasojevic et al., 2016; Chalmandrier et al., 2017)、生态尺度(Messier et al., 2017; 刘润红, 2018)等因素.具体来说, 随着空间尺度增大, 不同区域之间存在较大的环境差异导致群落中物种组成存在显著差异, 种间性状变异能够在很大的程度上解释物种多样性与环境因子间的关系, 种间性状变异相对于种内变异有可能逐渐凸显, 因此在区域尺度以上的大空间尺度研究中, 忽略种内变异可能是合理的(Violle et al., 2012; Díaz et al., 2016).但是在小尺度(如种群尺度等)或者局域环境下, 尤其是在物种丰富度较低的群落中, 种内性状变异对于群落物种共存具有重要意义, 不容忽略(Siefert et al., 2015).就不同功能性状而言, 叶片化学性状(如叶片氮和磷含量)的种内变异比叶片形态性状(如叶片面积和叶片厚度)的种内变异大; 植物整株性状(如株高等)的种内变异比器官水平性状(叶、茎和根性状等)的种内变异大(Siefert et al., 2015).此外, 不同生态系统的植物功能性状变异也存在一定的差异, 种内变异程度在受环境胁迫和物种数量较低的群落中更大(Siefert et al., 2015).因此, 只有结合种间和种内性状变异才能够准确地揭示群落构建和生物多样性维持机制(Albert et al., 2010b; Jung et al., 2010; Messier et al., 2010; 刘润红等, 2019). ...
The global spectrum of plant form and function 2 2016
... 在自然群落中, 物种间性状的差异被认为是物种共存的重要前提(Díaz et al., 2004), 故先前大多数基于功能性状的群落生态学研究仅考虑种间性状变异, 即采用群落中少数个体的性状平均值来代表物种性状值, 而忽略种内变异, 这隐含假设条件: 种间性状变异程度远大于种内变异程度, 种内变异相比种间变异可以忽略不计(Díaz et al., 2004; Jackson et al., 2013).然而, 最近十余年的研究表明, 种内变异虽然通常小于种间变异, 但是其可达性状总体变异的28%-52%, 对植物群落内和群落间的总体变异具有重要作用(Albert et al., 2010b; Messier et al., 2010; Siefert et al., 2015).种内变异在功能性状研究中的重要性取决于所选择的生态系统、功能性状、物种和研究目的(Albert et al., 2010a), 以及研究的时间尺度(Anderegg, 2015)、空间尺度(Spasojevic et al., 2016; Chalmandrier et al., 2017)、生态尺度(Messier et al., 2017; 刘润红, 2018)等因素.具体来说, 随着空间尺度增大, 不同区域之间存在较大的环境差异导致群落中物种组成存在显著差异, 种间性状变异能够在很大的程度上解释物种多样性与环境因子间的关系, 种间性状变异相对于种内变异有可能逐渐凸显, 因此在区域尺度以上的大空间尺度研究中, 忽略种内变异可能是合理的(Violle et al., 2012; Díaz et al., 2016).但是在小尺度(如种群尺度等)或者局域环境下, 尤其是在物种丰富度较低的群落中, 种内性状变异对于群落物种共存具有重要意义, 不容忽略(Siefert et al., 2015).就不同功能性状而言, 叶片化学性状(如叶片氮和磷含量)的种内变异比叶片形态性状(如叶片面积和叶片厚度)的种内变异大; 植物整株性状(如株高等)的种内变异比器官水平性状(叶、茎和根性状等)的种内变异大(Siefert et al., 2015).此外, 不同生态系统的植物功能性状变异也存在一定的差异, 种内变异程度在受环境胁迫和物种数量较低的群落中更大(Siefert et al., 2015).因此, 只有结合种间和种内性状变异才能够准确地揭示群落构建和生物多样性维持机制(Albert et al., 2010b; Jung et al., 2010; Messier et al., 2010; 刘润红等, 2019). ...
... 本研究发现叶经济型谱性状SLA与LTH、LDMC及LTD呈显著负相关关系, 小枝性状TTD和TDMC则呈高度正相关关系, 而LA在一定程度与小枝性状TTD和TDMC呈负相关关系, 这同先前大多数研究(Bucci et al., 2004; Wright et al., 2007; Baraloto et al., 2010; de la Riva et al., 2016; Díaz et al., 2016; 何东, 2016)的结果相似.SLA反映了植物对光照、水分和养分等资源的获取和利用能力, 与植物的生存策略密切相关(Garnier et al., 2001).通常, SLA较低的植物, 其叶片光合作用产物多投资于构建保卫细胞或增加叶肉细胞密度, 即增加叶片内部水分向叶片表面扩散的距离或阻力(王瑞丽等, 2016), 其LTH、LDMC和LTD也会相应地增加, 以减少蒸腾造成的水分散失, 提高水分利用效率, 从而具有较强的抵抗干旱胁迫和土壤贫瘠生境的能力, 提高其对环境的适合度(程雯等, 2019; 庞志强等, 2019).而SLA与LA在物种尺度上无显著相关性, PCA分析也发现, LA和SLA正交, 这说明两个性状属于不同的生态策略维度. ...
Plant functional traits of dominant native and invasive species in mediterranean-climate ecosystems 1 2016
... 植物功能性状是指植物体具有的对其定植、存活、生长、繁殖、死亡和最终适合度存在显著影响的形态、生理和物候等属性(孟婷婷等, 2007; Violle et al., 2007; 刘晓娟和马克平, 2015).因此, 研究植物功能性状变异与关联, 不仅能客观地反映植物自身生理过程及其对外部环境适应策略的差异, 而且还能有效地将植物个体、环境和生态系统的结构、过程和功能等有机结合起来, 揭示生物对生态系统过程与功能的影响, 从而为研究植物群落构建和生物多样性维持机制等诸多生态学热点问题提供了重要的突破口(Poorter et al., 2008; Cadotte et al., 2015; Funk et al., 2016; 刘润红等, 2019). ...
Consistency of species ranking based on functional leaf traits 1 2001
... 本研究发现叶经济型谱性状SLA与LTH、LDMC及LTD呈显著负相关关系, 小枝性状TTD和TDMC则呈高度正相关关系, 而LA在一定程度与小枝性状TTD和TDMC呈负相关关系, 这同先前大多数研究(Bucci et al., 2004; Wright et al., 2007; Baraloto et al., 2010; de la Riva et al., 2016; Díaz et al., 2016; 何东, 2016)的结果相似.SLA反映了植物对光照、水分和养分等资源的获取和利用能力, 与植物的生存策略密切相关(Garnier et al., 2001).通常, SLA较低的植物, 其叶片光合作用产物多投资于构建保卫细胞或增加叶肉细胞密度, 即增加叶片内部水分向叶片表面扩散的距离或阻力(王瑞丽等, 2016), 其LTH、LDMC和LTD也会相应地增加, 以减少蒸腾造成的水分散失, 提高水分利用效率, 从而具有较强的抵抗干旱胁迫和土壤贫瘠生境的能力, 提高其对环境的适合度(程雯等, 2019; 庞志强等, 2019).而SLA与LA在物种尺度上无显著相关性, PCA分析也发现, LA和SLA正交, 这说明两个性状属于不同的生态策略维度. ...
Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory 1 1977
The within- species leaf economic spectrum does not predict leaf litter decomposability at either the within-species or whole community levels 1 2013
... 在自然群落中, 物种间性状的差异被认为是物种共存的重要前提(Díaz et al., 2004), 故先前大多数基于功能性状的群落生态学研究仅考虑种间性状变异, 即采用群落中少数个体的性状平均值来代表物种性状值, 而忽略种内变异, 这隐含假设条件: 种间性状变异程度远大于种内变异程度, 种内变异相比种间变异可以忽略不计(Díaz et al., 2004; Jackson et al., 2013).然而, 最近十余年的研究表明, 种内变异虽然通常小于种间变异, 但是其可达性状总体变异的28%-52%, 对植物群落内和群落间的总体变异具有重要作用(Albert et al., 2010b; Messier et al., 2010; Siefert et al., 2015).种内变异在功能性状研究中的重要性取决于所选择的生态系统、功能性状、物种和研究目的(Albert et al., 2010a), 以及研究的时间尺度(Anderegg, 2015)、空间尺度(Spasojevic et al., 2016; Chalmandrier et al., 2017)、生态尺度(Messier et al., 2017; 刘润红, 2018)等因素.具体来说, 随着空间尺度增大, 不同区域之间存在较大的环境差异导致群落中物种组成存在显著差异, 种间性状变异能够在很大的程度上解释物种多样性与环境因子间的关系, 种间性状变异相对于种内变异有可能逐渐凸显, 因此在区域尺度以上的大空间尺度研究中, 忽略种内变异可能是合理的(Violle et al., 2012; Díaz et al., 2016).但是在小尺度(如种群尺度等)或者局域环境下, 尤其是在物种丰富度较低的群落中, 种内性状变异对于群落物种共存具有重要意义, 不容忽略(Siefert et al., 2015).就不同功能性状而言, 叶片化学性状(如叶片氮和磷含量)的种内变异比叶片形态性状(如叶片面积和叶片厚度)的种内变异大; 植物整株性状(如株高等)的种内变异比器官水平性状(叶、茎和根性状等)的种内变异大(Siefert et al., 2015).此外, 不同生态系统的植物功能性状变异也存在一定的差异, 种内变异程度在受环境胁迫和物种数量较低的群落中更大(Siefert et al., 2015).因此, 只有结合种间和种内性状变异才能够准确地揭示群落构建和生物多样性维持机制(Albert et al., 2010b; Jung et al., 2010; Messier et al., 2010; 刘润红等, 2019). ...
Interspecific and intraspecific variation in functional traits of subtropical evergreen and deciduous broadleaved mixed forests in karst topography, Guilin, Southwest China 2 2016
Intraspecific variability and trait-based community assembly 2 2010
... 在自然群落中, 物种间性状的差异被认为是物种共存的重要前提(Díaz et al., 2004), 故先前大多数基于功能性状的群落生态学研究仅考虑种间性状变异, 即采用群落中少数个体的性状平均值来代表物种性状值, 而忽略种内变异, 这隐含假设条件: 种间性状变异程度远大于种内变异程度, 种内变异相比种间变异可以忽略不计(Díaz et al., 2004; Jackson et al., 2013).然而, 最近十余年的研究表明, 种内变异虽然通常小于种间变异, 但是其可达性状总体变异的28%-52%, 对植物群落内和群落间的总体变异具有重要作用(Albert et al., 2010b; Messier et al., 2010; Siefert et al., 2015).种内变异在功能性状研究中的重要性取决于所选择的生态系统、功能性状、物种和研究目的(Albert et al., 2010a), 以及研究的时间尺度(Anderegg, 2015)、空间尺度(Spasojevic et al., 2016; Chalmandrier et al., 2017)、生态尺度(Messier et al., 2017; 刘润红, 2018)等因素.具体来说, 随着空间尺度增大, 不同区域之间存在较大的环境差异导致群落中物种组成存在显著差异, 种间性状变异能够在很大的程度上解释物种多样性与环境因子间的关系, 种间性状变异相对于种内变异有可能逐渐凸显, 因此在区域尺度以上的大空间尺度研究中, 忽略种内变异可能是合理的(Violle et al., 2012; Díaz et al., 2016).但是在小尺度(如种群尺度等)或者局域环境下, 尤其是在物种丰富度较低的群落中, 种内性状变异对于群落物种共存具有重要意义, 不容忽略(Siefert et al., 2015).就不同功能性状而言, 叶片化学性状(如叶片氮和磷含量)的种内变异比叶片形态性状(如叶片面积和叶片厚度)的种内变异大; 植物整株性状(如株高等)的种内变异比器官水平性状(叶、茎和根性状等)的种内变异大(Siefert et al., 2015).此外, 不同生态系统的植物功能性状变异也存在一定的差异, 种内变异程度在受环境胁迫和物种数量较低的群落中更大(Siefert et al., 2015).因此, 只有结合种间和种内性状变异才能够准确地揭示群落构建和生物多样性维持机制(Albert et al., 2010b; Jung et al., 2010; Messier et al., 2010; 刘润红等, 2019). ...
... 种内变异是指同种生物不同个体间的遗传或表型差异(Albert et al., 2010b).从遗传和发育机制来看, 种内变异可能源于同种个体之间的遗传变异或不同环境条件下产生的表型可塑性(Jung et al., 2010).本研究中各功能性状的总体种内变异幅度为6.76%- 48.30%, 种内变异平均达24.14%, 相比其他植被类型, 桂林岩溶石山青冈群落植物功能性状变异程度较低, 这与钟巧连等(2018)对黔中喀斯特木本植物功能性状变异的研究结果一致, 这说明植物功能性状变异幅度随不同植被类型及地理分布而存在较大差异, 即植物会随生境发生趋同或趋异适应, 这可能与植物对地理环境的适应策略及其资源利用能力有关, 是植物对其生存环境长期适应的结果. ...
Trait variability differs between leaf and wood tissues across ecological scales in subtropical forests 2014
... 在自然群落中, 物种间性状的差异被认为是物种共存的重要前提(Díaz et al., 2004), 故先前大多数基于功能性状的群落生态学研究仅考虑种间性状变异, 即采用群落中少数个体的性状平均值来代表物种性状值, 而忽略种内变异, 这隐含假设条件: 种间性状变异程度远大于种内变异程度, 种内变异相比种间变异可以忽略不计(Díaz et al., 2004; Jackson et al., 2013).然而, 最近十余年的研究表明, 种内变异虽然通常小于种间变异, 但是其可达性状总体变异的28%-52%, 对植物群落内和群落间的总体变异具有重要作用(Albert et al., 2010b; Messier et al., 2010; Siefert et al., 2015).种内变异在功能性状研究中的重要性取决于所选择的生态系统、功能性状、物种和研究目的(Albert et al., 2010a), 以及研究的时间尺度(Anderegg, 2015)、空间尺度(Spasojevic et al., 2016; Chalmandrier et al., 2017)、生态尺度(Messier et al., 2017; 刘润红, 2018)等因素.具体来说, 随着空间尺度增大, 不同区域之间存在较大的环境差异导致群落中物种组成存在显著差异, 种间性状变异能够在很大的程度上解释物种多样性与环境因子间的关系, 种间性状变异相对于种内变异有可能逐渐凸显, 因此在区域尺度以上的大空间尺度研究中, 忽略种内变异可能是合理的(Violle et al., 2012; Díaz et al., 2016).但是在小尺度(如种群尺度等)或者局域环境下, 尤其是在物种丰富度较低的群落中, 种内性状变异对于群落物种共存具有重要意义, 不容忽略(Siefert et al., 2015).就不同功能性状而言, 叶片化学性状(如叶片氮和磷含量)的种内变异比叶片形态性状(如叶片面积和叶片厚度)的种内变异大; 植物整株性状(如株高等)的种内变异比器官水平性状(叶、茎和根性状等)的种内变异大(Siefert et al., 2015).此外, 不同生态系统的植物功能性状变异也存在一定的差异, 种内变异程度在受环境胁迫和物种数量较低的群落中更大(Siefert et al., 2015).因此, 只有结合种间和种内性状变异才能够准确地揭示群落构建和生物多样性维持机制(Albert et al., 2010b; Jung et al., 2010; Messier et al., 2010; 刘润红等, 2019). ...
1 2018
... 在自然群落中, 物种间性状的差异被认为是物种共存的重要前提(Díaz et al., 2004), 故先前大多数基于功能性状的群落生态学研究仅考虑种间性状变异, 即采用群落中少数个体的性状平均值来代表物种性状值, 而忽略种内变异, 这隐含假设条件: 种间性状变异程度远大于种内变异程度, 种内变异相比种间变异可以忽略不计(Díaz et al., 2004; Jackson et al., 2013).然而, 最近十余年的研究表明, 种内变异虽然通常小于种间变异, 但是其可达性状总体变异的28%-52%, 对植物群落内和群落间的总体变异具有重要作用(Albert et al., 2010b; Messier et al., 2010; Siefert et al., 2015).种内变异在功能性状研究中的重要性取决于所选择的生态系统、功能性状、物种和研究目的(Albert et al., 2010a), 以及研究的时间尺度(Anderegg, 2015)、空间尺度(Spasojevic et al., 2016; Chalmandrier et al., 2017)、生态尺度(Messier et al., 2017; 刘润红, 2018)等因素.具体来说, 随着空间尺度增大, 不同区域之间存在较大的环境差异导致群落中物种组成存在显著差异, 种间性状变异能够在很大的程度上解释物种多样性与环境因子间的关系, 种间性状变异相对于种内变异有可能逐渐凸显, 因此在区域尺度以上的大空间尺度研究中, 忽略种内变异可能是合理的(Violle et al., 2012; Díaz et al., 2016).但是在小尺度(如种群尺度等)或者局域环境下, 尤其是在物种丰富度较低的群落中, 种内性状变异对于群落物种共存具有重要意义, 不容忽略(Siefert et al., 2015).就不同功能性状而言, 叶片化学性状(如叶片氮和磷含量)的种内变异比叶片形态性状(如叶片面积和叶片厚度)的种内变异大; 植物整株性状(如株高等)的种内变异比器官水平性状(叶、茎和根性状等)的种内变异大(Siefert et al., 2015).此外, 不同生态系统的植物功能性状变异也存在一定的差异, 种内变异程度在受环境胁迫和物种数量较低的群落中更大(Siefert et al., 2015).因此, 只有结合种间和种内性状变异才能够准确地揭示群落构建和生物多样性维持机制(Albert et al., 2010b; Jung et al., 2010; Messier et al., 2010; 刘润红等, 2019). ...
漓江河岸带木本植物功能性状跨尺度变异研究 2 2019
... 植物功能性状是指植物体具有的对其定植、存活、生长、繁殖、死亡和最终适合度存在显著影响的形态、生理和物候等属性(孟婷婷等, 2007; Violle et al., 2007; 刘晓娟和马克平, 2015).因此, 研究植物功能性状变异与关联, 不仅能客观地反映植物自身生理过程及其对外部环境适应策略的差异, 而且还能有效地将植物个体、环境和生态系统的结构、过程和功能等有机结合起来, 揭示生物对生态系统过程与功能的影响, 从而为研究植物群落构建和生物多样性维持机制等诸多生态学热点问题提供了重要的突破口(Poorter et al., 2008; Cadotte et al., 2015; Funk et al., 2016; 刘润红等, 2019). ...
... 在自然群落中, 物种间性状的差异被认为是物种共存的重要前提(Díaz et al., 2004), 故先前大多数基于功能性状的群落生态学研究仅考虑种间性状变异, 即采用群落中少数个体的性状平均值来代表物种性状值, 而忽略种内变异, 这隐含假设条件: 种间性状变异程度远大于种内变异程度, 种内变异相比种间变异可以忽略不计(Díaz et al., 2004; Jackson et al., 2013).然而, 最近十余年的研究表明, 种内变异虽然通常小于种间变异, 但是其可达性状总体变异的28%-52%, 对植物群落内和群落间的总体变异具有重要作用(Albert et al., 2010b; Messier et al., 2010; Siefert et al., 2015).种内变异在功能性状研究中的重要性取决于所选择的生态系统、功能性状、物种和研究目的(Albert et al., 2010a), 以及研究的时间尺度(Anderegg, 2015)、空间尺度(Spasojevic et al., 2016; Chalmandrier et al., 2017)、生态尺度(Messier et al., 2017; 刘润红, 2018)等因素.具体来说, 随着空间尺度增大, 不同区域之间存在较大的环境差异导致群落中物种组成存在显著差异, 种间性状变异能够在很大的程度上解释物种多样性与环境因子间的关系, 种间性状变异相对于种内变异有可能逐渐凸显, 因此在区域尺度以上的大空间尺度研究中, 忽略种内变异可能是合理的(Violle et al., 2012; Díaz et al., 2016).但是在小尺度(如种群尺度等)或者局域环境下, 尤其是在物种丰富度较低的群落中, 种内性状变异对于群落物种共存具有重要意义, 不容忽略(Siefert et al., 2015).就不同功能性状而言, 叶片化学性状(如叶片氮和磷含量)的种内变异比叶片形态性状(如叶片面积和叶片厚度)的种内变异大; 植物整株性状(如株高等)的种内变异比器官水平性状(叶、茎和根性状等)的种内变异大(Siefert et al., 2015).此外, 不同生态系统的植物功能性状变异也存在一定的差异, 种内变异程度在受环境胁迫和物种数量较低的群落中更大(Siefert et al., 2015).因此, 只有结合种间和种内性状变异才能够准确地揭示群落构建和生物多样性维持机制(Albert et al., 2010b; Jung et al., 2010; Messier et al., 2010; 刘润红等, 2019). ...
漓江河岸带木本植物功能性状跨尺度变异研究 2 2019
... 植物功能性状是指植物体具有的对其定植、存活、生长、繁殖、死亡和最终适合度存在显著影响的形态、生理和物候等属性(孟婷婷等, 2007; Violle et al., 2007; 刘晓娟和马克平, 2015).因此, 研究植物功能性状变异与关联, 不仅能客观地反映植物自身生理过程及其对外部环境适应策略的差异, 而且还能有效地将植物个体、环境和生态系统的结构、过程和功能等有机结合起来, 揭示生物对生态系统过程与功能的影响, 从而为研究植物群落构建和生物多样性维持机制等诸多生态学热点问题提供了重要的突破口(Poorter et al., 2008; Cadotte et al., 2015; Funk et al., 2016; 刘润红等, 2019). ...
... 在自然群落中, 物种间性状的差异被认为是物种共存的重要前提(Díaz et al., 2004), 故先前大多数基于功能性状的群落生态学研究仅考虑种间性状变异, 即采用群落中少数个体的性状平均值来代表物种性状值, 而忽略种内变异, 这隐含假设条件: 种间性状变异程度远大于种内变异程度, 种内变异相比种间变异可以忽略不计(Díaz et al., 2004; Jackson et al., 2013).然而, 最近十余年的研究表明, 种内变异虽然通常小于种间变异, 但是其可达性状总体变异的28%-52%, 对植物群落内和群落间的总体变异具有重要作用(Albert et al., 2010b; Messier et al., 2010; Siefert et al., 2015).种内变异在功能性状研究中的重要性取决于所选择的生态系统、功能性状、物种和研究目的(Albert et al., 2010a), 以及研究的时间尺度(Anderegg, 2015)、空间尺度(Spasojevic et al., 2016; Chalmandrier et al., 2017)、生态尺度(Messier et al., 2017; 刘润红, 2018)等因素.具体来说, 随着空间尺度增大, 不同区域之间存在较大的环境差异导致群落中物种组成存在显著差异, 种间性状变异能够在很大的程度上解释物种多样性与环境因子间的关系, 种间性状变异相对于种内变异有可能逐渐凸显, 因此在区域尺度以上的大空间尺度研究中, 忽略种内变异可能是合理的(Violle et al., 2012; Díaz et al., 2016).但是在小尺度(如种群尺度等)或者局域环境下, 尤其是在物种丰富度较低的群落中, 种内性状变异对于群落物种共存具有重要意义, 不容忽略(Siefert et al., 2015).就不同功能性状而言, 叶片化学性状(如叶片氮和磷含量)的种内变异比叶片形态性状(如叶片面积和叶片厚度)的种内变异大; 植物整株性状(如株高等)的种内变异比器官水平性状(叶、茎和根性状等)的种内变异大(Siefert et al., 2015).此外, 不同生态系统的植物功能性状变异也存在一定的差异, 种内变异程度在受环境胁迫和物种数量较低的群落中更大(Siefert et al., 2015).因此, 只有结合种间和种内性状变异才能够准确地揭示群落构建和生物多样性维持机制(Albert et al., 2010b; Jung et al., 2010; Messier et al., 2010; 刘润红等, 2019). ...
植物功能性状研究进展 2 2015
... 植物功能性状是指植物体具有的对其定植、存活、生长、繁殖、死亡和最终适合度存在显著影响的形态、生理和物候等属性(孟婷婷等, 2007; Violle et al., 2007; 刘晓娟和马克平, 2015).因此, 研究植物功能性状变异与关联, 不仅能客观地反映植物自身生理过程及其对外部环境适应策略的差异, 而且还能有效地将植物个体、环境和生态系统的结构、过程和功能等有机结合起来, 揭示生物对生态系统过程与功能的影响, 从而为研究植物群落构建和生物多样性维持机制等诸多生态学热点问题提供了重要的突破口(Poorter et al., 2008; Cadotte et al., 2015; Funk et al., 2016; 刘润红等, 2019). ...
... 植物在其漫长的历史进化和发展过程中, 通过内部不同功能性状之间的协同进化与权衡来适应变化的外界环境(Silvertown, 2004).当两个或多个重要植物功能性状在不同物种间都存在一致的相关性时, 这种性状组合关系可以认为是一个策略维度, 也称“生态策略”, 即植物沿一定的生态策略轴排列于最适应或最具竞争力的位置(Wright et al., 2006, 2007; 刘晓娟和马克平, 2015).因此, 许多研究在性状变异的多维空间里甄别出少数几个能够充分描述物种间功能性状变异的独立轴主轴来代表生态策略的分化方向, 如叶片-株高-种子(leaf-height-seed, LHS)权衡(Westoby, 1998)、叶经济型谱(leaf economic spectrum, LES)(Wright et al., 2004)、木材经济型谱(wood economic spectrum, WES)(Chave et al., 2009)和根经济型谱(root economic spectrum, RES)(Weemstra et al., 2016)等.值得注意的是, 上述功能性状的协变与权衡格局基本是在大尺度上确立的, 然而, 植物在进化过程中会面临自然选择或对环境胁迫的适应, 特定区域内不同性状之间的关系可能会与全球尺度或区域尺度的变化不一致, 反映出植物对特殊生境的适应(Santiago & Wright, 2007).因此, 还需要在不同尺度(如局域植物群落内部共存物种之间、种内或个体水平)上验证功能性状之间的这种协变与权衡关系是否仍然成立(Albert et al., 2010a; 何东, 2016). ...
植物功能性状研究进展 2 2015
... 植物功能性状是指植物体具有的对其定植、存活、生长、繁殖、死亡和最终适合度存在显著影响的形态、生理和物候等属性(孟婷婷等, 2007; Violle et al., 2007; 刘晓娟和马克平, 2015).因此, 研究植物功能性状变异与关联, 不仅能客观地反映植物自身生理过程及其对外部环境适应策略的差异, 而且还能有效地将植物个体、环境和生态系统的结构、过程和功能等有机结合起来, 揭示生物对生态系统过程与功能的影响, 从而为研究植物群落构建和生物多样性维持机制等诸多生态学热点问题提供了重要的突破口(Poorter et al., 2008; Cadotte et al., 2015; Funk et al., 2016; 刘润红等, 2019). ...
... 植物在其漫长的历史进化和发展过程中, 通过内部不同功能性状之间的协同进化与权衡来适应变化的外界环境(Silvertown, 2004).当两个或多个重要植物功能性状在不同物种间都存在一致的相关性时, 这种性状组合关系可以认为是一个策略维度, 也称“生态策略”, 即植物沿一定的生态策略轴排列于最适应或最具竞争力的位置(Wright et al., 2006, 2007; 刘晓娟和马克平, 2015).因此, 许多研究在性状变异的多维空间里甄别出少数几个能够充分描述物种间功能性状变异的独立轴主轴来代表生态策略的分化方向, 如叶片-株高-种子(leaf-height-seed, LHS)权衡(Westoby, 1998)、叶经济型谱(leaf economic spectrum, LES)(Wright et al., 2004)、木材经济型谱(wood economic spectrum, WES)(Chave et al., 2009)和根经济型谱(root economic spectrum, RES)(Weemstra et al., 2016)等.值得注意的是, 上述功能性状的协变与权衡格局基本是在大尺度上确立的, 然而, 植物在进化过程中会面临自然选择或对环境胁迫的适应, 特定区域内不同性状之间的关系可能会与全球尺度或区域尺度的变化不一致, 反映出植物对特殊生境的适应(Santiago & Wright, 2007).因此, 还需要在不同尺度(如局域植物群落内部共存物种之间、种内或个体水平)上验证功能性状之间的这种协变与权衡关系是否仍然成立(Albert et al., 2010a; 何东, 2016). ...
Coordinated evolution of leaf and stem economics in tropical dry forest trees 1 2012
... 植物功能性状是指植物体具有的对其定植、存活、生长、繁殖、死亡和最终适合度存在显著影响的形态、生理和物候等属性(孟婷婷等, 2007; Violle et al., 2007; 刘晓娟和马克平, 2015).因此, 研究植物功能性状变异与关联, 不仅能客观地反映植物自身生理过程及其对外部环境适应策略的差异, 而且还能有效地将植物个体、环境和生态系统的结构、过程和功能等有机结合起来, 揭示生物对生态系统过程与功能的影响, 从而为研究植物群落构建和生物多样性维持机制等诸多生态学热点问题提供了重要的突破口(Poorter et al., 2008; Cadotte et al., 2015; Funk et al., 2016; 刘润红等, 2019). ...
植物功能性状与环境和生态系统功能 1 2007
... 植物功能性状是指植物体具有的对其定植、存活、生长、繁殖、死亡和最终适合度存在显著影响的形态、生理和物候等属性(孟婷婷等, 2007; Violle et al., 2007; 刘晓娟和马克平, 2015).因此, 研究植物功能性状变异与关联, 不仅能客观地反映植物自身生理过程及其对外部环境适应策略的差异, 而且还能有效地将植物个体、环境和生态系统的结构、过程和功能等有机结合起来, 揭示生物对生态系统过程与功能的影响, 从而为研究植物群落构建和生物多样性维持机制等诸多生态学热点问题提供了重要的突破口(Poorter et al., 2008; Cadotte et al., 2015; Funk et al., 2016; 刘润红等, 2019). ...
Trait variation and integration across scales: Is the leaf economic spectrum present at local scales? 1 2017
... 在自然群落中, 物种间性状的差异被认为是物种共存的重要前提(Díaz et al., 2004), 故先前大多数基于功能性状的群落生态学研究仅考虑种间性状变异, 即采用群落中少数个体的性状平均值来代表物种性状值, 而忽略种内变异, 这隐含假设条件: 种间性状变异程度远大于种内变异程度, 种内变异相比种间变异可以忽略不计(Díaz et al., 2004; Jackson et al., 2013).然而, 最近十余年的研究表明, 种内变异虽然通常小于种间变异, 但是其可达性状总体变异的28%-52%, 对植物群落内和群落间的总体变异具有重要作用(Albert et al., 2010b; Messier et al., 2010; Siefert et al., 2015).种内变异在功能性状研究中的重要性取决于所选择的生态系统、功能性状、物种和研究目的(Albert et al., 2010a), 以及研究的时间尺度(Anderegg, 2015)、空间尺度(Spasojevic et al., 2016; Chalmandrier et al., 2017)、生态尺度(Messier et al., 2017; 刘润红, 2018)等因素.具体来说, 随着空间尺度增大, 不同区域之间存在较大的环境差异导致群落中物种组成存在显著差异, 种间性状变异能够在很大的程度上解释物种多样性与环境因子间的关系, 种间性状变异相对于种内变异有可能逐渐凸显, 因此在区域尺度以上的大空间尺度研究中, 忽略种内变异可能是合理的(Violle et al., 2012; Díaz et al., 2016).但是在小尺度(如种群尺度等)或者局域环境下, 尤其是在物种丰富度较低的群落中, 种内性状变异对于群落物种共存具有重要意义, 不容忽略(Siefert et al., 2015).就不同功能性状而言, 叶片化学性状(如叶片氮和磷含量)的种内变异比叶片形态性状(如叶片面积和叶片厚度)的种内变异大; 植物整株性状(如株高等)的种内变异比器官水平性状(叶、茎和根性状等)的种内变异大(Siefert et al., 2015).此外, 不同生态系统的植物功能性状变异也存在一定的差异, 种内变异程度在受环境胁迫和物种数量较低的群落中更大(Siefert et al., 2015).因此, 只有结合种间和种内性状变异才能够准确地揭示群落构建和生物多样性维持机制(Albert et al., 2010b; Jung et al., 2010; Messier et al., 2010; 刘润红等, 2019). ...
How do traits vary across ecological scales? A case for trait-based ecology 2 2010
... 在自然群落中, 物种间性状的差异被认为是物种共存的重要前提(Díaz et al., 2004), 故先前大多数基于功能性状的群落生态学研究仅考虑种间性状变异, 即采用群落中少数个体的性状平均值来代表物种性状值, 而忽略种内变异, 这隐含假设条件: 种间性状变异程度远大于种内变异程度, 种内变异相比种间变异可以忽略不计(Díaz et al., 2004; Jackson et al., 2013).然而, 最近十余年的研究表明, 种内变异虽然通常小于种间变异, 但是其可达性状总体变异的28%-52%, 对植物群落内和群落间的总体变异具有重要作用(Albert et al., 2010b; Messier et al., 2010; Siefert et al., 2015).种内变异在功能性状研究中的重要性取决于所选择的生态系统、功能性状、物种和研究目的(Albert et al., 2010a), 以及研究的时间尺度(Anderegg, 2015)、空间尺度(Spasojevic et al., 2016; Chalmandrier et al., 2017)、生态尺度(Messier et al., 2017; 刘润红, 2018)等因素.具体来说, 随着空间尺度增大, 不同区域之间存在较大的环境差异导致群落中物种组成存在显著差异, 种间性状变异能够在很大的程度上解释物种多样性与环境因子间的关系, 种间性状变异相对于种内变异有可能逐渐凸显, 因此在区域尺度以上的大空间尺度研究中, 忽略种内变异可能是合理的(Violle et al., 2012; Díaz et al., 2016).但是在小尺度(如种群尺度等)或者局域环境下, 尤其是在物种丰富度较低的群落中, 种内性状变异对于群落物种共存具有重要意义, 不容忽略(Siefert et al., 2015).就不同功能性状而言, 叶片化学性状(如叶片氮和磷含量)的种内变异比叶片形态性状(如叶片面积和叶片厚度)的种内变异大; 植物整株性状(如株高等)的种内变异比器官水平性状(叶、茎和根性状等)的种内变异大(Siefert et al., 2015).此外, 不同生态系统的植物功能性状变异也存在一定的差异, 种内变异程度在受环境胁迫和物种数量较低的群落中更大(Siefert et al., 2015).因此, 只有结合种间和种内性状变异才能够准确地揭示群落构建和生物多样性维持机制(Albert et al., 2010b; Jung et al., 2010; Messier et al., 2010; 刘润红等, 2019). ...
... ; Messier et al., 2010; 刘润红等, 2019). ...
A functional approach reveals community responses to disturbances 2013
Leaf functional traits of tropical forest plants in relation to growth form 1 2007
... 植物在其漫长的历史进化和发展过程中, 通过内部不同功能性状之间的协同进化与权衡来适应变化的外界环境(Silvertown, 2004).当两个或多个重要植物功能性状在不同物种间都存在一致的相关性时, 这种性状组合关系可以认为是一个策略维度, 也称“生态策略”, 即植物沿一定的生态策略轴排列于最适应或最具竞争力的位置(Wright et al., 2006, 2007; 刘晓娟和马克平, 2015).因此, 许多研究在性状变异的多维空间里甄别出少数几个能够充分描述物种间功能性状变异的独立轴主轴来代表生态策略的分化方向, 如叶片-株高-种子(leaf-height-seed, LHS)权衡(Westoby, 1998)、叶经济型谱(leaf economic spectrum, LES)(Wright et al., 2004)、木材经济型谱(wood economic spectrum, WES)(Chave et al., 2009)和根经济型谱(root economic spectrum, RES)(Weemstra et al., 2016)等.值得注意的是, 上述功能性状的协变与权衡格局基本是在大尺度上确立的, 然而, 植物在进化过程中会面临自然选择或对环境胁迫的适应, 特定区域内不同性状之间的关系可能会与全球尺度或区域尺度的变化不一致, 反映出植物对特殊生境的适应(Santiago & Wright, 2007).因此, 还需要在不同尺度(如局域植物群落内部共存物种之间、种内或个体水平)上验证功能性状之间的这种协变与权衡关系是否仍然成立(Albert et al., 2010a; 何东, 2016). ...
A global meta-analysis of the relative extent of intraspecific trait variation in plant communities 4 2015
... 在自然群落中, 物种间性状的差异被认为是物种共存的重要前提(Díaz et al., 2004), 故先前大多数基于功能性状的群落生态学研究仅考虑种间性状变异, 即采用群落中少数个体的性状平均值来代表物种性状值, 而忽略种内变异, 这隐含假设条件: 种间性状变异程度远大于种内变异程度, 种内变异相比种间变异可以忽略不计(Díaz et al., 2004; Jackson et al., 2013).然而, 最近十余年的研究表明, 种内变异虽然通常小于种间变异, 但是其可达性状总体变异的28%-52%, 对植物群落内和群落间的总体变异具有重要作用(Albert et al., 2010b; Messier et al., 2010; Siefert et al., 2015).种内变异在功能性状研究中的重要性取决于所选择的生态系统、功能性状、物种和研究目的(Albert et al., 2010a), 以及研究的时间尺度(Anderegg, 2015)、空间尺度(Spasojevic et al., 2016; Chalmandrier et al., 2017)、生态尺度(Messier et al., 2017; 刘润红, 2018)等因素.具体来说, 随着空间尺度增大, 不同区域之间存在较大的环境差异导致群落中物种组成存在显著差异, 种间性状变异能够在很大的程度上解释物种多样性与环境因子间的关系, 种间性状变异相对于种内变异有可能逐渐凸显, 因此在区域尺度以上的大空间尺度研究中, 忽略种内变异可能是合理的(Violle et al., 2012; Díaz et al., 2016).但是在小尺度(如种群尺度等)或者局域环境下, 尤其是在物种丰富度较低的群落中, 种内性状变异对于群落物种共存具有重要意义, 不容忽略(Siefert et al., 2015).就不同功能性状而言, 叶片化学性状(如叶片氮和磷含量)的种内变异比叶片形态性状(如叶片面积和叶片厚度)的种内变异大; 植物整株性状(如株高等)的种内变异比器官水平性状(叶、茎和根性状等)的种内变异大(Siefert et al., 2015).此外, 不同生态系统的植物功能性状变异也存在一定的差异, 种内变异程度在受环境胁迫和物种数量较低的群落中更大(Siefert et al., 2015).因此, 只有结合种间和种内性状变异才能够准确地揭示群落构建和生物多样性维持机制(Albert et al., 2010b; Jung et al., 2010; Messier et al., 2010; 刘润红等, 2019). ...
... ).但是在小尺度(如种群尺度等)或者局域环境下, 尤其是在物种丰富度较低的群落中, 种内性状变异对于群落物种共存具有重要意义, 不容忽略(Siefert et al., 2015).就不同功能性状而言, 叶片化学性状(如叶片氮和磷含量)的种内变异比叶片形态性状(如叶片面积和叶片厚度)的种内变异大; 植物整株性状(如株高等)的种内变异比器官水平性状(叶、茎和根性状等)的种内变异大(Siefert et al., 2015).此外, 不同生态系统的植物功能性状变异也存在一定的差异, 种内变异程度在受环境胁迫和物种数量较低的群落中更大(Siefert et al., 2015).因此, 只有结合种间和种内性状变异才能够准确地揭示群落构建和生物多样性维持机制(Albert et al., 2010b; Jung et al., 2010; Messier et al., 2010; 刘润红等, 2019). ...
... ).就不同功能性状而言, 叶片化学性状(如叶片氮和磷含量)的种内变异比叶片形态性状(如叶片面积和叶片厚度)的种内变异大; 植物整株性状(如株高等)的种内变异比器官水平性状(叶、茎和根性状等)的种内变异大(Siefert et al., 2015).此外, 不同生态系统的植物功能性状变异也存在一定的差异, 种内变异程度在受环境胁迫和物种数量较低的群落中更大(Siefert et al., 2015).因此, 只有结合种间和种内性状变异才能够准确地揭示群落构建和生物多样性维持机制(Albert et al., 2010b; Jung et al., 2010; Messier et al., 2010; 刘润红等, 2019). ...
... ).此外, 不同生态系统的植物功能性状变异也存在一定的差异, 种内变异程度在受环境胁迫和物种数量较低的群落中更大(Siefert et al., 2015).因此, 只有结合种间和种内性状变异才能够准确地揭示群落构建和生物多样性维持机制(Albert et al., 2010b; Jung et al., 2010; Messier et al., 2010; 刘润红等, 2019). ...
Plant coexistence and the niche 1 2004
... 植物在其漫长的历史进化和发展过程中, 通过内部不同功能性状之间的协同进化与权衡来适应变化的外界环境(Silvertown, 2004).当两个或多个重要植物功能性状在不同物种间都存在一致的相关性时, 这种性状组合关系可以认为是一个策略维度, 也称“生态策略”, 即植物沿一定的生态策略轴排列于最适应或最具竞争力的位置(Wright et al., 2006, 2007; 刘晓娟和马克平, 2015).因此, 许多研究在性状变异的多维空间里甄别出少数几个能够充分描述物种间功能性状变异的独立轴主轴来代表生态策略的分化方向, 如叶片-株高-种子(leaf-height-seed, LHS)权衡(Westoby, 1998)、叶经济型谱(leaf economic spectrum, LES)(Wright et al., 2004)、木材经济型谱(wood economic spectrum, WES)(Chave et al., 2009)和根经济型谱(root economic spectrum, RES)(Weemstra et al., 2016)等.值得注意的是, 上述功能性状的协变与权衡格局基本是在大尺度上确立的, 然而, 植物在进化过程中会面临自然选择或对环境胁迫的适应, 特定区域内不同性状之间的关系可能会与全球尺度或区域尺度的变化不一致, 反映出植物对特殊生境的适应(Santiago & Wright, 2007).因此, 还需要在不同尺度(如局域植物群落内部共存物种之间、种内或个体水平)上验证功能性状之间的这种协变与权衡关系是否仍然成立(Albert et al., 2010a; 何东, 2016). ...
When does intraspecific trait variation contribute to functional beta- diversity? 1 2016
... 在自然群落中, 物种间性状的差异被认为是物种共存的重要前提(Díaz et al., 2004), 故先前大多数基于功能性状的群落生态学研究仅考虑种间性状变异, 即采用群落中少数个体的性状平均值来代表物种性状值, 而忽略种内变异, 这隐含假设条件: 种间性状变异程度远大于种内变异程度, 种内变异相比种间变异可以忽略不计(Díaz et al., 2004; Jackson et al., 2013).然而, 最近十余年的研究表明, 种内变异虽然通常小于种间变异, 但是其可达性状总体变异的28%-52%, 对植物群落内和群落间的总体变异具有重要作用(Albert et al., 2010b; Messier et al., 2010; Siefert et al., 2015).种内变异在功能性状研究中的重要性取决于所选择的生态系统、功能性状、物种和研究目的(Albert et al., 2010a), 以及研究的时间尺度(Anderegg, 2015)、空间尺度(Spasojevic et al., 2016; Chalmandrier et al., 2017)、生态尺度(Messier et al., 2017; 刘润红, 2018)等因素.具体来说, 随着空间尺度增大, 不同区域之间存在较大的环境差异导致群落中物种组成存在显著差异, 种间性状变异能够在很大的程度上解释物种多样性与环境因子间的关系, 种间性状变异相对于种内变异有可能逐渐凸显, 因此在区域尺度以上的大空间尺度研究中, 忽略种内变异可能是合理的(Violle et al., 2012; Díaz et al., 2016).但是在小尺度(如种群尺度等)或者局域环境下, 尤其是在物种丰富度较低的群落中, 种内性状变异对于群落物种共存具有重要意义, 不容忽略(Siefert et al., 2015).就不同功能性状而言, 叶片化学性状(如叶片氮和磷含量)的种内变异比叶片形态性状(如叶片面积和叶片厚度)的种内变异大; 植物整株性状(如株高等)的种内变异比器官水平性状(叶、茎和根性状等)的种内变异大(Siefert et al., 2015).此外, 不同生态系统的植物功能性状变异也存在一定的差异, 种内变异程度在受环境胁迫和物种数量较低的群落中更大(Siefert et al., 2015).因此, 只有结合种间和种内性状变异才能够准确地揭示群落构建和生物多样性维持机制(Albert et al., 2010b; Jung et al., 2010; Messier et al., 2010; 刘润红等, 2019). ...
The return of the variance: intraspecific variability in community ecology 1 2012
... 在自然群落中, 物种间性状的差异被认为是物种共存的重要前提(Díaz et al., 2004), 故先前大多数基于功能性状的群落生态学研究仅考虑种间性状变异, 即采用群落中少数个体的性状平均值来代表物种性状值, 而忽略种内变异, 这隐含假设条件: 种间性状变异程度远大于种内变异程度, 种内变异相比种间变异可以忽略不计(Díaz et al., 2004; Jackson et al., 2013).然而, 最近十余年的研究表明, 种内变异虽然通常小于种间变异, 但是其可达性状总体变异的28%-52%, 对植物群落内和群落间的总体变异具有重要作用(Albert et al., 2010b; Messier et al., 2010; Siefert et al., 2015).种内变异在功能性状研究中的重要性取决于所选择的生态系统、功能性状、物种和研究目的(Albert et al., 2010a), 以及研究的时间尺度(Anderegg, 2015)、空间尺度(Spasojevic et al., 2016; Chalmandrier et al., 2017)、生态尺度(Messier et al., 2017; 刘润红, 2018)等因素.具体来说, 随着空间尺度增大, 不同区域之间存在较大的环境差异导致群落中物种组成存在显著差异, 种间性状变异能够在很大的程度上解释物种多样性与环境因子间的关系, 种间性状变异相对于种内变异有可能逐渐凸显, 因此在区域尺度以上的大空间尺度研究中, 忽略种内变异可能是合理的(Violle et al., 2012; Díaz et al., 2016).但是在小尺度(如种群尺度等)或者局域环境下, 尤其是在物种丰富度较低的群落中, 种内性状变异对于群落物种共存具有重要意义, 不容忽略(Siefert et al., 2015).就不同功能性状而言, 叶片化学性状(如叶片氮和磷含量)的种内变异比叶片形态性状(如叶片面积和叶片厚度)的种内变异大; 植物整株性状(如株高等)的种内变异比器官水平性状(叶、茎和根性状等)的种内变异大(Siefert et al., 2015).此外, 不同生态系统的植物功能性状变异也存在一定的差异, 种内变异程度在受环境胁迫和物种数量较低的群落中更大(Siefert et al., 2015).因此, 只有结合种间和种内性状变异才能够准确地揭示群落构建和生物多样性维持机制(Albert et al., 2010b; Jung et al., 2010; Messier et al., 2010; 刘润红等, 2019). ...
Let the concept of trait be functional! 1 2007
... 植物功能性状是指植物体具有的对其定植、存活、生长、繁殖、死亡和最终适合度存在显著影响的形态、生理和物候等属性(孟婷婷等, 2007; Violle et al., 2007; 刘晓娟和马克平, 2015).因此, 研究植物功能性状变异与关联, 不仅能客观地反映植物自身生理过程及其对外部环境适应策略的差异, 而且还能有效地将植物个体、环境和生态系统的结构、过程和功能等有机结合起来, 揭示生物对生态系统过程与功能的影响, 从而为研究植物群落构建和生物多样性维持机制等诸多生态学热点问题提供了重要的突破口(Poorter et al., 2008; Cadotte et al., 2015; Funk et al., 2016; 刘润红等, 2019). ...
 ,1,3,*, 梁士楚1,3, 李月娟1,3
,1,3,*, 梁士楚1,3, 李月娟1,3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT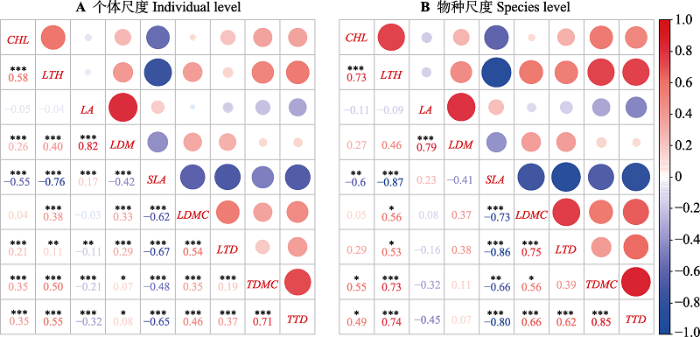 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT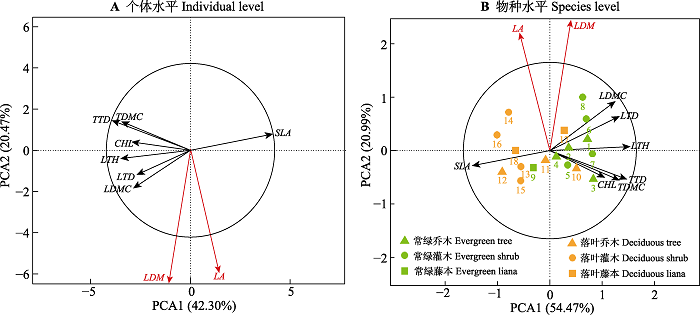 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}