Effects of simulated warming on biological soil crust-soil system respiration in alpine sandy lands
He-Ju ZHAO1,2, Yan-Peng YUE1,2, Xiao-Hong JIA,1,2,*, Long CHENG1,2, Bo WU1,2, Yuan-Shou LI3, Hong ZHOU1,2, Xue-Bin ZHAO41Institute of Desertification Studies, Chinese Academy of Forestry, Beijing 100091, China 2Key Laboratory of Desert Ecosystem and Global Change, State Administration of Forestry and Grassland, Beijing 100091, China 3Chinese Academy of Meteorological Sciences, Beijing 100081, China 4 Qinghai Gonghe Desert Ecosystem National Positioning Research Station, Gonghe, Qinghai 813000, China
Supported by the National Key R&D Program of China(2016YFC0500801) the Special Survey on Science and Technology Basic Resources(2017FY100206) the National Natural Science Foundation of China(41471096) the National Natural Science Foundation of China(41371093)
Abstract Aims Biological soil crust is an important type of surface cover in alpine sandy lands. Understanding of the effect of warming on respiration from the biological soil crust-soil system in alpine regions can provide theoretical reference to the assessment of the response and feedback of biological soil crusts to climate changes. Methods The moss and algae crusts in the artificial vegetation restoration areas were taken as the research objects. The open top chamber (OTC) was used as a passive warming device to simulate warming. The daily and growing season dynamics of respiration rates in two types of biological soil crust-soil systems were measured. The effects of warming on CO2 emission and its temperature sensitivity were discussed. Important findings Both the daily and the growing season dynamics of respiration rate of the moss and algae crust-soil system showed ��single-peak�� curves and were not affected by warming. The daily peaks appeared around 13:00, and the growing season peaks appeared around August. Warming changed the daily peak value of respiration rate of the biological soil crust-soil system. In the relatively dry year (2017), moderate warming increased cumulative CO2 emission from the two types of biological soil crust-soil system during growing season, but the increase declined under excessive warming. In the relatively wet year (2018), as warming got greater, CO2 emission from the two types of biological soil crust-soil system increased more. The relationship between respiration rate and temperature of two types of biological soil crust-soil system followed the exponential function. In the relatively dry year, more increase of temperature induced smaller temperature sensitivity of CO2 emission, and the temperature sensitivity varied from 1.47 to 1.61 and 1.60 to 1.95 in the moss and algae crust soil system respectively. In the relatively wet year, with the increase of temperature, temperature sensitivity of system respiration increased, and the temperature sensitivity varied from 1.44 to 1.68 and 1.44 to 1.76 in the moss and algae crust soil system respectively. This study shows that global warming has greatly increased the respiration of biological soil crust-soil system in alpine ecosystems. Therefore, we should fully consider the impact of climate warming on the wide spread biological soil crusts in this area for better evaluation of carbon cycling processes in alpine ecosystems. Keywords��alpine sandy area;biological soil crust;warming;respiration;temperature sensitivity
PDF (1183KB)Ԫ������ά�����������������EndNote|Ris|Bibtex�ղر��� ���ñ��� �ԺӾ�, ������, ������, ����, �Ⲩ, ��Ԫ��, �ܺ�, ��ѩ��. ģ�����¶Ըߺ�ɳ������������Ƥ-����ϵͳ������Ӱ��. ֲ����̬ѧ��, 2020, 44(9): 916-925. DOI: 10.17521/cjpe.2020.0018 ZHAO He-Ju, YUE Yan-Peng, JIA Xiao-Hong, CHENG Long, WU Bo, LI Yuan-Shou, ZHOU Hong, ZHAO Xue-Bin. Effects of simulated warming on biological soil crust-soil system respiration in alpine sandy lands. Chinese Journal of Plant Ecology, 2020, 44(9): 916-925. DOI: 10.17521/cjpe.2020.0018
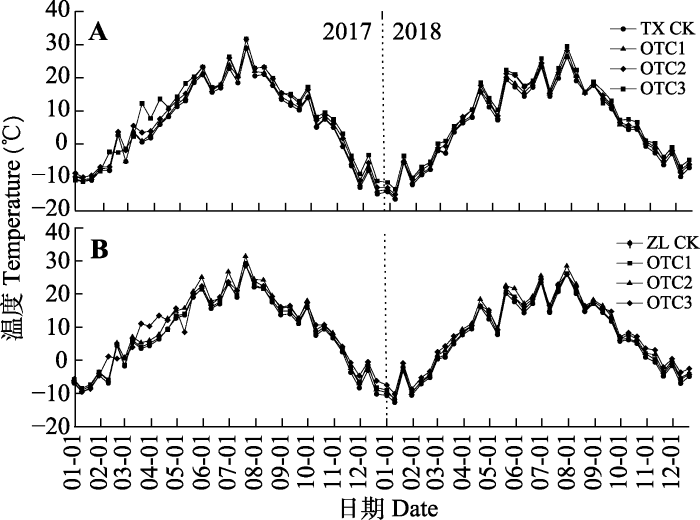
Fig. 1Changes of soil temperature (0-5 cm) under moss and algae crusts in open top chamber (OTC) a passive warming device with different specifications. A, Moss crust-soil system (TX). B, Algae crust-soil system (ZL). CK means control group; OTC1, OTC2, OTC3 represent different warming treatments.
Fig. 2Increase in soil temperature (0-5 cm) under moss and algae crusts in open top chamber (OTC) a passive warming device with different specifications. TX, moss crust-soil system; ZL, algae crust-soil system.
Fig. 3Daily dynamics of respiration rate of the biological soil crust-soil system under simulated warming. A, Moss crust-soil system (TX). B, Algae crust-soil system (ZL). CK means control group; OTC1, OTC2, OTC3 represent different warming treatments.
Fig. 4Growing season dynamics of respiration rate of the biological soil crust-soil system under simulated warming (mean �� SE). A, Moss crust-soil system (TX). B, Algae crust-soil system (ZL). CK means control group; OTC1, OTC2, OTC3 represent different warming treatments.
Table 3 ��3 ��3ģ������������������Ƥ-����ϵͳ�������ۻ�CO2�ͷ��� Table 3Cumulative CO2 emission from the biological soil crust-soil system in the growing season under simulated warming treatments
���� Type
��� Year
CO2�ͷ��� CO2 emission (g��m-2)
CK (CV, %)
OTC1 (CV, %)
OTC2 (CV, %)
OTC3 (CV, %)
̦ Moss
2017
120.71 (31.62)
239.69 (27.64)
223.83 (30.83)
207.96 (34.96)
2018
386.97 (30.91)
430.63 (33.54)
442.82 (32.31)
481.35 (33.53)
���� Algae
2017
79.03 (51.98)
115.87 (53.15)
105.86 (56.54)
94.94 (56.39)
2018
236.80 (46.93)
272.71 (45.30)
285.90 (43.74)
308.64 (42.13)
CV is the coefficient of variation. CK means control group; OTC1, OTC2, OTC3 represent different warming treatments. CVΪ����ϵ����CKΪ������, OTC1��OTC2��OTC3Ϊ��ͬ���´�����
Table 4 ��4 ��4ģ������������������Ƥ-����ϵͳ����������0-5 cm�������¶ȼ�Ļع��ϵ���¶�������(Q10)�Ƚ� Table 4Regression relationship between respiration rate of the biological soil crust-soil system and soil temperature in the 0-5 cm layer under simulated warming and comparison of temperature sensitivity (Q10) among different treatments
���� Type
���� Treatment
2017
2018
�ع鷽�� Regression equation
R2
Q10
p
�ع鷽�� Regression equation
R2
Q10
p
̦ Moss
CK
y = 0.389e0.0489x
0.36**
1.61
0.001
y = 1.131e0.0364x
0.37**
1.44
0.000
OTC1
y = 0.612e0.0422x
0.37**
1.53
0.008
y = 1.163e0.0419x
0.51**
1.52
0.001
OTC2
y = 0.546e0.0405x
0.53**
1.50
0.000
y = 0.901e0.047x
0.45**
1.60
0.000
OTC3
y = 0.502e0.0385x
0.66**
1.47
0.000
y = 1.009e0.0519x
0.58**
1.68
0.009
���� Algae
CK
y = 0.103e0.0667x
0.76**
1.95
0.001
y = 0.615e0.0363x
0.33**
1.44
0.000
OTC1
y = 0.090e0.064x
0.35**
1.90
0.005
y = 0.554e0.0445x
0.27**
1.56
0.004
OTC2
y = 0.149e0.0501x
0.66**
1.65
0.000
y = 0.361e0.0519x
0.40**
1.68
0.000
OTC3
y = 0.204e0.0467x
0.31**
1.60
0.000
y = 0.522e0.0567x
0.49**
1.76
0.000
* represents a significance level of 0.05; ** represents a significance level of 0.01. CK means control group; OTC1, OTC2, OTC3 represent different warming treatments. *�����ﵽ0.05��������ˮƽ; **�����ﵽ0.01��������ˮƽ��CKΪ������, OTC1��OTC2��OTC3Ϊ��ͬ���´�����
AllisonSD, TresederKK (2008). Warming and drying suppress microbial activity and carbon cycling in boreal forest soils .Global Change Biology, 14, 2898-2909. DOI:10.1111/j.1365-2486.2008.01716.xURL [��������: 1]
AlmagroM, L��pezJ, QuerejetaJI, Mart��nez-MenaM (2009). Temperature dependence of soil CO2 efflux is strongly modulated by seasonal patterns of moisture availability in a Mediterranean ecosystem .Soil Biology & Biochemistry, 41, 594-605. DOI:10.1016/j.soilbio.2008.12.021URL [��������: 1]
BokhorstS, BjerkeJW, MelilloJ, CallaghanTV, PhoenixGK (2010). Impacts of extreme winter warming events on litter decomposition in a sub-Arctic heathland .Soil Biology & Biochemistry, 42, 611-617. DOI:10.1016/j.soilbio.2009.12.011URL [��������: 1]
ChenH, ZhuQ, PengCH, WuN, WangYF, FangXQ, GaoYH, ZhuD, YangG, TianJQ, KangXM, PiaoSL, OuyangH, XiangWH, LuoZB, JiangH, SongXZ, ZhangY, YuGR, ZhaoXQ, GongP, YaoTD, WuJH (2013). The impacts of climate change and human activities on biogeochemical cycles on the Qinghai-Tibetan Plateau .Global Change Biology, 19, 2940-2955. DOI:10.1111/gcb.2013.19.issue-10URL [��������: 1]
ChenQS, LiLH, HanXG, YanZD, WangYF, ZhangY, XiongXG, ChenSP, ZhangLX, GaoYZ, TangF, YangJ, DongYS (2004). Temperature sensitivity of soil respiration in relation to soil moisture in 11 communities of typical temperate steppe in Inner Mongolia .Acta Ecologica Sinica, 24, 831-836.
ChenZF (2012). Effects of Simulated Warming and Nitrogen Addition on Gas Exchange in Desert Steppe Ecosystems. Master degree dissertation, Inner Mongolia Agricultural University Hohhot. [��������: 1]
DavidsonEA, JanssensIA (2006). Temperature sensitivity of soil carbon decomposition and feedbacks to climate change .Nature, 440, 165-173. DOI:10.1038/nature04514URL
DoreMHI (2005). Climate change and changes in global precipitation patterns: What do we know? Environment International, 31, 1167-1181. DOI:10.1016/j.envint.2005.03.004URL [��������: 1]
EldridgeDJ, GreeneRSB (1994). Microbiotic soil crusts��A review of their roles in soil and ecological processes in the rangelands of Australia .Australian Journal of Soil Research, 32, 389-415. [��������: 1]
FengW (2014). Photosynthetic Carbon Fixation of Biological Soil Crusts in MU US Desert and Their Impact on Soil Carbon Emission .PhD dissertation, Beijing Forestry University, Beijing. [��������: 1]
FuW, ZhangXY, ZhaoJ, DuSL, HouMT (2017). Effects of experimental warming on soil respiration during growing period in cropland in the black soil region of Northeast China .Chinese Journal of Ecology, 36, 601-608. [��������: 1]
GengXD, XuR, WeiD (2017). Response of greenhouse gases flux to multi-level warming in an alpine meadow of Tibetan Plateau .Ecology and Environment Sciences, 26, 445-452.
GuanC, ZhangP, LiXR (2017). Responses of soil respiration with biocrust cover to water and temperature in the southeastern edge of Tengger Desert, Northwest China . Chinese Journal of Plant Ecology, 41, 301-310. DOI:10.17521/cjpe.2016.0326URL
HanGX, ZhouGS, XuZZ, YangY, LiuJL, ShiKQ (2007). Soil temperature and biotic factors drive the seasonal variation of soil respiration in a maize (Zea mays L.) agricultural ecosystem Plant and Soil, 291, 15-26. DOI:10.1007/s11104-006-9170-8URL [��������: 1]
HanHY (2014). Biological Soil Crust Carbon Emission and Its Effects on Soil Respiration Alpine in Alpine Sandy Land PhD dissertation, Chinese Academy of Forestry, Beijing. [��������: 1]
IPCC (2013). Climate Change 2013: the Physical Science Basis. Contribution of Working Group I to fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, UK. [��������: 1]
JanssensIA, PilegaardK (2003). Large seasonal changes inQ10 of soil respiration in a beech forest Global Change Biology, 9, 911-918. DOI:10.1046/j.1365-2486.2003.00636.xURL
KlimekB, Choczy��skiM, JuszkiewiczA (2009). Scots pine (Pinus sylvestris L.) roots and soil moisture did not affect soil thermal sensitivity European Journal of Soil Biology, 45, 442-447. DOI:10.1016/j.ejsobi.2009.06.008URL [��������: 2]
LanSB, WuL, ZhangDL, HuCX (2012). Successional stages of biological soil crusts and their microstructure variability in Shapotou region (China) .Environmental Earth Sciences, 65, 77-88. DOI:10.1007/s12665-011-1066-0URL
LeifeldJ, FuhrerJ (2005). The temperature response of CO2 production from bulk soils and soil fractions is related to soil organic matter quality .Biogeochemistry, 75, 433-453. DOI:10.1007/s10533-005-2237-4URL [��������: 1]
LiXR, ZhangYM, ZhaoYG (2009). A study of biological soil crusts: recent development, trend and prospect .Advances in Earth Science, 24, 11-24. [��������: 1]
LinGH, RygiewiczPT, EhleringerJR, JohnsonMG, TingeyDT (2001). Time-dependent responses of soil CO2 efflux components to elevated atmospheric [CO2] and temperature in experimental forest mesocosms .Plant and Soil, 229, 259-270. DOI:10.1023/A:1004883221036URL [��������: 1]
LiuDJ (2012). Responses of Soil Respiration of Prickly Prickles Community to Rainfall Increase in Extreme Arid Area. PhD dissertation, Chinese Academy of Forestry, Beijing. [��������: 2]
LiuHS, LiuHJ, WangZP, XuM, HanXG, LiLH (2008). The temperature sensitivity of soil respiration .Progress in Geography, 27, 51-60. [��������: 1]
LuoCY, XuGP, ChaoZG, WangSP, LinXW, HuYG, ZhangZH, DuanJC, ChangXF, SuAL, LiYN, ZhaoXQ, DuMY, TangYH, KimballB (2010). Effect of warming and grazing on litter mass loss and temperature sensitivity of litter and dung mass loss on the Tibetan Plateau .Global Change Biology, 16, 1606-1617. DOI:10.1111/gcb.2010.16.issue-5URL [��������: 1]
MaestreFT, EscolarC, de GuevaraML, QueroJL, L��zaroR, Delgado-BaquerizoM, OchoaV, BerdugoM, GozaloB, GallardoA (2013). Changes in biocrust cover drive carbon cycle responses to climate change in drylands .Global Change Biology, 19, 3835-3847. DOI:10.1111/gcb.12306URL [��������: 1]
MarilleyL, HartwigUA, AragnoM (1999). Influence of an elevated atmospheric CO2 content on soil and rhizosphere bacterial communities beneathLolium perenne and Trifolium repens under field conditions Microbial Ecology, 38, 39-49. DOI:10.1007/s002489900155URL [��������: 1]
McCulleyRL, BouttonTW, ArcherSR (2007). Soil respiration in a subtropical savanna parkland: response to water additions .Soil Science Society of America Journal, 71, 820-828. DOI:10.2136/sssaj2006.0303URL
Nikli��skaM, Marya��skiM, LaskowskiR (1999). Effect of temperature on humus respiration rate and nitrogen mineralization: implications for global climate change .Biogeochemistry, 44, 239-257. [��������: 1]
PanXL, LinB, LiuQ (2008). Effects of elevated temperature on soil organic carbon and soil respiration under subalpine coniferous forest in western Sichuan Province, China .Chinese Journal of Applied Ecology,19, 1637-1643. [��������: 1]
QinY, YiSH, LiNJ, RenSL, WangXY, ChenJJ (2012). Advance in studies of carbon cycling on alpine grasslands of the Qinghai-Tibetan Plateau .Acta Prataculturae Sinica, 21, 275-285.
ReichsteinM, SubkeJA, AngeliAC, TenhuneneJD (2005). Does the temperature sensitivity of decomposition of soil organic matter depend upon water content, soil horizon, or incubation time? Global Change Biology, 11, 1754-1767. DOI:10.1111/gcb.2005.11.issue-10URL [��������: 1]
SongB, NiuSL (2016). Global change and terrestrial carbon cycle: a review .Journal of Southwest University for Nationalities (Natural Science Edition), 42, 14-23. [��������: 1]
SuYG, WuL, ZhouZB, LiuYB, ZhangYM (2013). Carbon flux in deserts depends on soil cover type: a case study in the Gurbantunggute desert, North China . Soil Biology �� Biochemistry, 58, 332-340. [��������: 2]
WanSQ, NorbyRJ, LedfordJ, NorbyRJ, LedfordJ, WeltzinJF (2007). Responses of soil respiration to elevated CO2, air warming, and changing soil water availability in a model old-field grassland .Global Change Biology, 13, 2411-2424. DOI:10.1111/gcb.2007.13.issue-11URL [��������: 1]
WangZ, ZhaoML, HanGD, GaoFG, HanX (2012). Response of soil respiration to simulated warming and N addition in the desert steppe .Journal of Arid Land Resources and Environment, 26, 98-103. [��������: 1]
XiaJ, HanY, ZhangZ, ZhangZ, WanS (2009). Effects of diurnal warming on soil respiration are not equal to the summed effects of day and night warming in a temperate steppe .Biogeosciences, 6, 1361-1370. DOI:10.5194/bg-6-1361-2009URL [��������: 1]
XiongP, XuZF, LinB, LiuQ (2010). Short-term response of winter soil respiration to simulated warming in aPinus armandii plantation in the upper reaches of the Minjiang River, China Chinese Journal of Plant Ecology,34, 1369-1376. DOI:10.3773/j.issn.1005-264x.2010.12.002URL [��������: 1] Aims Winter soil respiration is reported to be a significant component of the annual carbon budgets. In order to understand how projected global warming affects winter soil respiration and soil microbial properties, we conducted a warming experiment in a young Pinus armandii plantation during the winter period to assess short-term response of winter soil CO2 efflux and microbial properties to artificial warming. Methods We used an infrared heater and monitored soil temperature and relative air humidity in November 2009 and analyzed soil respiration, microbial biomass and inorganic nitrogen pools throughout the winter. Important findings The average daily air and soil temperatures were 2.1 and 6.7 �� higher in the warmed plots than in the control plots, respectively. Warming declined relative air humidity and soil water content by 15.2% and 7.2%, respectively. Across all measuring dates, simulated warming increased average soil CO2 efflux by 31.4%. Warming significantly enhanced the soil microbial biomass carbon (SMB-C) and nitrogen (SMB-N) by 23.2% and 22.7%, respectively, but did not affect the ratio of SMB-C to SMB-N, which indicated that elevated temperature tended to accelerate the growth of SMB, but did not change the community structure of SMB. Likewise, warming tended to increase soil inorganic nitrogen pools. Nitrate (NO3–-N) and ammonium (NH4+-N) in the warmed soil increased 38.5% and 12.3%, respectively, compared with control soil. Results indicate that winter soil respiration, microbial activities and nutrient mineralization in the coniferous forest soils could be sensitive to future global warming.]]> [����, �����, �ֲ�, ���� (2010). ẽ����λ�ɽ���ֶ�������������ģ�����µĶ�����Ӧ ֲ����̬ѧ��, 34, 1369-1376.] DOI:10.3773/j.issn.1005-264x.2010.12.002URL [��������: 1] Pinus armandii)�˹��ֶ����������������������������������ģ�����µ���Ӧ���������: �ڶ���(2009��11��–����3��), ģ�������������������������������, ƽ��������31.4%; ͬ��ģ������ʹ��������������̼�����ֱ�����23.2%��22.7%, ��������������̼����û��Ӱ��, �¶����������ٽ������������, ��û�иı������Ⱥ��ṹ; ��������������NO3–-N��NH4+-NŨ�Ƚ϶��շֱ�������38.5%��12.3%, ������������������Ŀ�����������������������, ������Ҷ�ֶ��������������������������ֿ�δ�������ů�dz����С�]]>
XuBX, HuYG, ZhangZS, ChenYL, ZhangP, LiG (2014). Effects of experimental warming on CO2, CH4 and N2O fluxes of biological soil crust and soil system in a desert region .Chinese Journal of Plant Ecology, 38, 809-820. DOI:10.3724/SP.J.1258.2014.00076URL
XuM, QiY (2001). Spatial and seasonal variations ofQ10 determined by soil respiration measurements at a Sierra Nevadan Forest Global Biogeochemical Cycles, 15, 687-696. DOI:10.1029/2000GB001365URL [��������: 1]
YangQP, XuM, LiuHS, WangJS, LiuLX, ChiYG, ZhengYP (2011). Impact factors and uncertainties of the temperature sensitivity of soil respiration .Acta Ecologica Sinica, 31, 2301-2311.
YangY, HuangM, LiuHS, LiuHJ (2011). The interrelation between temperature sensitivity and adaptability of soil respiration .Journal of Natural Resources, 26, 1811-1820. [��������: 1]
YusteJC, MaS, BaldocchiDD (2010). Plant-soil interactions and acclimation to temperature of microbial-mediated soil respiration may affect predictions of soil CO2 efflux .Biogeochemistry, 98, 127-138. DOI:10.1007/s10533-009-9381-1URL [��������: 1]
ZhaoJX, LuoTX, WeiHX, DengZH, LiX, LiRC, TangYH (2019). Increased precipitation offsets the negative effect of warming on plant biomass and ecosystem respiration in a Tibetan alpine steppe .Agricultural and Forest Meteorology, 279, 107761. DOI: 10.1016/j.agrformet.?2019.?107761. DOI:10.1016/j.agrformet.2019.107761URL
Warming and drying suppress microbial activity and carbon cycling in boreal forest soils 1 2008
... ����������Ҫ������������(ֲ���ϵ����)����������(����������������).�����������OTC��Զ��ֲ��2 m���������ܱ��ڹ����������������8100-104�н��вⶨ, �ʿɲ���������������Ƥ������ú�ֲ���ϵ��������������ɵ�Ӱ��.���о�����, ����������������������������Ƥ-����ϵͳ��CO2�ͷ���, ���·��Ȳ�ͬ, �����ӳ̶�Ҳ��ͬ, ����̦��Ƥ-����ϵͳ��CO2�ͷ������������Ƥ-����ϵͳ.�����������ԭ��������: ����, ����ͨ����������������Ƥ-����ϵͳ�е�������(Marilley et al., 1999), ֱ��Ӱ���������; ���, ���»���������������Ƥ-����ϵͳ������Ļ���(Bokhorst et al., 2010), �Ӷ��ٽ������ĺ�������; ���, �����ܹ���������������Ƥ-����ϵͳ��ø�Ļ���, ���ٺ����������ֽ���ѷֽ���ֵ�����, ��������������Ӱ��(Yuste et al., 2010).���ߵ����·���, ���������������Ƥ-����ϵͳ�е�������Ժ�ø���Բ�������, ͬʱ���ή������ˮ�ֵ���Ч��, �������������ĺ�������; ����ˮ��������Ϊ����ʱ, ˮ���ܹ���һ���̶��ϵ������߷������µ�����Ӱ��, �������������ֳ�����������(Zhao et al., 2019).����������Ƥ-����ϵͳ, ̦��Ƥ-����ϵͳ�����������������ࡢ�����ͻ��Ծ����ڸ��ѵ�״̬, ��̦��Ƥ�����ı�ˮ���ܸ���, �����ڼ�������������, ʹ̦��Ƥ-����ϵͳ��CO2�ͷ������������Ƥ-����ϵͳ(������, 2017).�����о�����, ���������ܹ��������������ĺ�������(Lin et al., 2001; Wan et al., 2007; Xia et al., 2009; ��־��, 2012; Maestre et al., 2013), ���о������֮һ��.�����ڸ���ɳĮ���о�ȴ�����¶����߶Ը���������������Ƥ-����ϵͳ��ƽ��CO2�ͷ���������Ӱ��(����ε�, 2014), ����������ڲ�ͬ�о���������ˮ�������IJ�ͬ�����µ�.����ڸߺ�ɳ��, �ڸ���ɳĮ�Ľ�ˮ��Ϊ�ѷ�, ƽ���꽵ˮ��186.55 mm, ���Ե����о���2017��226.7 mm��2018��372.4 mm�Ľ�ˮ��, ����ˮ�ֵ�����, ����������Ƥ-����ϵͳ�е�������Ժͺ����������Ч��Ҳ���ܵ�����, �����»��һ���Ӿ�����ˮ�ֵ�ȱʧ, ������ø���Բ�������(Allison & Treseder, 2008), ������º�CO2�ͷ����IJ��.Ҳ���о�����, ���º���������û�з������Ա仯��ԭ���������������ö��¶Ȳ�����Ӧ�ԵĽ��(��������, 2008; ����, 2017), ���Ҫ��һ�����ڵĹ۲�ȥ����˵��. ...
Temperature dependence of soil CO2 efflux is strongly modulated by seasonal patterns of moisture availability in a Mediterranean ecosystem 1 2009
... �����������¶��������ܹ���ӳ�����ů��ȫ����̬ϵͳ̼ѭ����Ĺ�ϵ, �¶ȵ������ܹ��������������¶������Բ���Ӱ��(Davidson & Janssens, 2006; Liu et al., 2008; Zhao et al., 2019).���о�����, �����������¶������Ի����¶ȵ����߶�����(Xu & Qi, 2001; Luo et al., 2010; �������, 2011), �����о�����, ���ʪ�����(2018), ��ˮ������, ����������������������������Ƥ-����ϵͳ�����������߶ȵ��¶�������, �����·���Խ��, ���ӳ̶�Խ��.�����������ˮ�������IJ�������˹��߷������µ�Ч��, ��������������������, �����������������¶�������(Zhao et al., 2019).ˮ�ֻ�ͨ��Ӱ�������л��ʵ���ɢ��������������Ӱ��, �������л�����ø�����䶼��Ҫ��Һ���н���, ���»ή��������ˮ�ֺ���, ���ͺ��������ø����ɢת��������Ļ���, �Ӷ������������������ĽӴ�����, Ӱ������������������; ����ˮ�ֽ�Ϊ����ʱ, �ʶ�����, ���ἤ������Ļ�����ø����, ���ٺ�������ֽ�, ��������������¶�������(�����, 2011).�����о�����, ����������Q10ֵ������ˮ�־���һ����������, ͨ�������, ����ˮ��ȱʧ�ᵼ��Q10�½�, ��һ���̶���������ˮ��������, Q10Ҳ����Ӧ����(Janssens & Pilegaard, 2003; Reichstein et al., 2005; Almagro et al., 2009).�����ȴ���ԭ��̬ϵͳ���о�����, ����ˮ�ֵ�����, �ܹ������������������Q10ֵ(McCulley et al., 2007), ���о������֮��ͬ.���ڱ��о�ֻ�ǶԶ������¶���������������������Ƥ-����ϵͳ�����¶�������Ӱ��ij���̽��, �й����¶����������¶�������Ӱ������ڻ����볤��ЧӦ, �������ǽ�һ�����ڹ۲��о�. ...
Impacts of extreme winter warming events on litter decomposition in a sub-Arctic heathland 1 2010
... ����������Ҫ������������(ֲ���ϵ����)����������(����������������).�����������OTC��Զ��ֲ��2 m���������ܱ��ڹ����������������8100-104�н��вⶨ, �ʿɲ���������������Ƥ������ú�ֲ���ϵ��������������ɵ�Ӱ��.���о�����, ����������������������������Ƥ-����ϵͳ��CO2�ͷ���, ���·��Ȳ�ͬ, �����ӳ̶�Ҳ��ͬ, ����̦��Ƥ-����ϵͳ��CO2�ͷ������������Ƥ-����ϵͳ.�����������ԭ��������: ����, ����ͨ����������������Ƥ-����ϵͳ�е�������(Marilley et al., 1999), ֱ��Ӱ���������; ���, ���»���������������Ƥ-����ϵͳ������Ļ���(Bokhorst et al., 2010), �Ӷ��ٽ������ĺ�������; ���, �����ܹ���������������Ƥ-����ϵͳ��ø�Ļ���, ���ٺ����������ֽ���ѷֽ���ֵ�����, ��������������Ӱ��(Yuste et al., 2010).���ߵ����·���, ���������������Ƥ-����ϵͳ�е�������Ժ�ø���Բ�������, ͬʱ���ή������ˮ�ֵ���Ч��, �������������ĺ�������; ����ˮ��������Ϊ����ʱ, ˮ���ܹ���һ���̶��ϵ������߷������µ�����Ӱ��, �������������ֳ�����������(Zhao et al., 2019).����������Ƥ-����ϵͳ, ̦��Ƥ-����ϵͳ�����������������ࡢ�����ͻ��Ծ����ڸ��ѵ�״̬, ��̦��Ƥ�����ı�ˮ���ܸ���, �����ڼ�������������, ʹ̦��Ƥ-����ϵͳ��CO2�ͷ������������Ƥ-����ϵͳ(������, 2017).�����о�����, ���������ܹ��������������ĺ�������(Lin et al., 2001; Wan et al., 2007; Xia et al., 2009; ��־��, 2012; Maestre et al., 2013), ���о������֮һ��.�����ڸ���ɳĮ���о�ȴ�����¶����߶Ը���������������Ƥ-����ϵͳ��ƽ��CO2�ͷ���������Ӱ��(����ε�, 2014), ����������ڲ�ͬ�о���������ˮ�������IJ�ͬ�����µ�.����ڸߺ�ɳ��, �ڸ���ɳĮ�Ľ�ˮ��Ϊ�ѷ�, ƽ���꽵ˮ��186.55 mm, ���Ե����о���2017��226.7 mm��2018��372.4 mm�Ľ�ˮ��, ����ˮ�ֵ�����, ����������Ƥ-����ϵͳ�е�������Ժͺ����������Ч��Ҳ���ܵ�����, �����»��һ���Ӿ�����ˮ�ֵ�ȱʧ, ������ø���Բ�������(Allison & Treseder, 2008), ������º�CO2�ͷ����IJ��.Ҳ���о�����, ���º���������û�з������Ա仯��ԭ���������������ö��¶Ȳ�����Ӧ�ԵĽ��(��������, 2008; ����, 2017), ���Ҫ��һ�����ڵĹ۲�ȥ����˵��. ...
The impacts of climate change and human activities on biogeochemical cycles on the Qinghai-Tibetan Plateau 1 2013
Changes in biocrust cover drive carbon cycle responses to climate change in drylands 1 2013
... ����������Ƥ�������塢���¡��������ֲ�����������ͨ����˿�塢�ٸ��ͷ�����Ƚ�����������ϸС�����γɵĸ�����(Eldridge & Greene, 1994; Lan et al., 2012).���Ǹɺ���ɺ�������Ҫ�ĵر���������, �ڻ�Į��̬ϵͳ�еĸ��Ƕ��ܴﵽ40%����(�����ٵ�, 2009), �ڷ����ɳ������ˮ������̬�ָ��ȷ��������Ҫ����̬����.����������Ƥ���ص������ṹʹ������������������, ���ܽ��й������, Ҳ�ܽ��к�������, ���������������Լ���̬ϵͳ̼ѭ��������ҪӰ��. ...
Influence of an elevated atmospheric CO2 content on soil and rhizosphere bacterial communities beneathLolium perenne and Trifolium repens under field conditions 1 1999
 ,1,2,*, ����1,2, �Ⲩ1,2, ��Ԫ��3, �ܺ�1,2, ��ѩ��4
,1,2,*, ����1,2, �Ⲩ1,2, ��Ԫ��3, �ܺ�1,2, ��ѩ��4
 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPT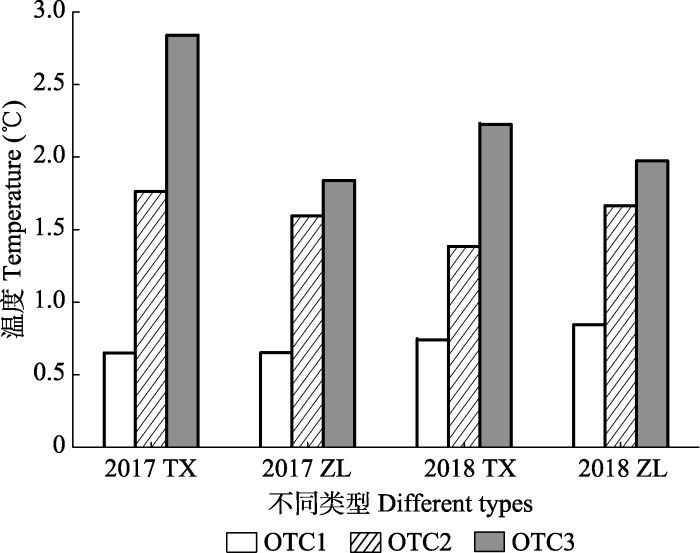 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPT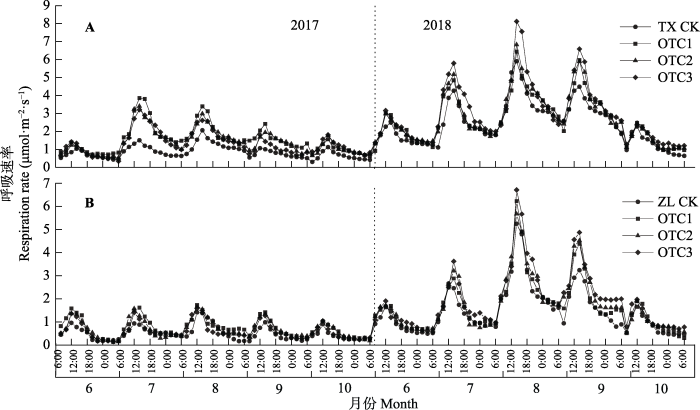 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPT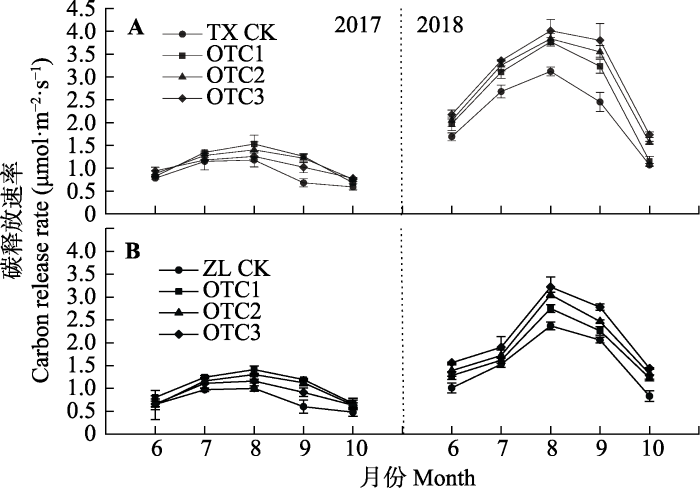 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}