Characteristics of soil carbon and nitrogen contents and enzyme activities in sub-alpine secondary forests with different successional stages in Western Sichuan, China
Zong-Da HU,1,*, Shi-Rong LIU3, Ming-Xia LUO1, Jing HU4, Xing-Liang LIU2, Ya-Fei LI1, Hao YU1, Ding-Hua OU11College of Resources, Sichuan Agricultural University, Chengdu 611130, China 2Ecological Restoration and Conservation on Forest and Wetland Key Laboratory of Sichuan Province, Sichuan Academy of Forestry, Chengdu 610081, China 3Research Institute of Forest Ecology, Environment and Protection, Chinese Academy of Forestry, Beijing 100091, China 4College of Resources and Environment, Hunan Agricultural University, Changsha 410128, China
Supported by Forest and Wetland Ecological Restoration and Conservation of Open Project from Key Laboratory of Sichuan Province(2019KFKT03) the National Key R&D Program of China(2017YFC0505004) the Environmental Governance and Ecological Protection Major Project of Sichuan Province (2018SZDZX0031)
Abstract Aims Regeneration of sub-alpine forests have an capacity to sequester carbon and nitrogen. Our objectives were to quantify variations of soil organic carbon and nitrogen content and enzyme activities at different successional stages of natural secondary forests, and to better understand the underlying mechanisms of carbon and nitrogen sequestration in these sub-alpine forests. Methods We used the space-for-time substitution method and selected four sub-alpine forests in Miyaluo forest of Western Sichuan, China. The secondary forests were at different successional stages with natural regeneration on cutting-blanks in 1960s (60-NSF), 1970s (70-NSF) and 1980s (80-NSF), and the Abies faxoniana primary forest was used as control (CK). The soil samples were taken from 0 to 20 cm depths in each forest in late July, 2019, and were transferred to the laboratory. Soil organic carbon (SOC), soil total nitrogen (TN), dissolved organic carbon (DOC) and nitrogen (DON), light fraction organic carbon (LFOC) content and soil enzyme activities were measured. The activities of five soil extracellular enzymes related to soil carbon and nitrogen cycling were determined to explain their relationships with soil physico-chemical properties. Important findings The contents of topsoil SOC, DOC, LFOC decreased with succession stage, whereas TN and DON were all in the order of 60-NSF < 80-NSF < 70-NSF, although 80-NSF and 70-NSF exhibited no significant difference. The topsoil organic carbon and nitrogen and their active fractions contents in natural secondary forests were lower than those in the primary forest, while no significant difference in DOC and DON contents was observed between 80-NSF and CK. The activities of soil ��-4-glucosidase (��G), ��-4-N-acetylglucosa- minidase (NAG) and polyphenol oxidase (PHO) in CK were significantly higher than those in natural secondary forests, whereas soil cellulose hydrolysis (CBH) and phenol oxidase (PEO) activities had no significant difference between secondary forests and CK. Activities of ��G and CBH in 60-NSF were significantly lower than those in 70-NSF and 80-NSF, but activities of NAG in 80-NSF were significantly higher than those in 60-NSFand 70-NSF. There was no significant difference in PEO activities among different types of forest. Both Pearson correlation analysis and redundancy analysis showed that soil enzyme activities were significantly correlated with soil TN, LFOC and DOC contents. TN content explained 65.4% of the variations in enzyme activity, which implied that change in soil nitrogen content might affect C-related hydrolytic enzyme activities (e.g. ��G, CBH and NAG). Meanwhile soil microorganisms prefer to use readily decomposable carbon and nitrogen. Therefore, activities of some soil enzymes such as ��G, CBH and NAG in natural secondary forests decreased due to the declines in soil TN, LFOC and DOC contents. We conclude that soil enzyme activities could be more favorable to C and N cycling in the Abies faxoniana primary forest than in the secondary forest at the early-successional stages (<60 a) in high-altitude sub-alpine forest ecosystems in Western Sichuan, China. Keywords��soil organic carbon;soil total nitrogen;enzyme activity;succession stage;natural secondary forest;soil carbon cycle;soil nitrogen cycle
PDF (1230KB)Ԫ������ά�����������������EndNote|Ris|Bibtex�ղر��� ���ñ��� ���ڴ�, ������, ����ϼ, ���Z, ������, ���Ƿ�, ���, ŷ����. �����Ǹ�ɽ��ͬ�������Ȼ����������̼��������ø��������. ֲ����̬ѧ��, 2020, 44(9): 973-985. DOI: 10.17521/cjpe.2020.0203 HU Zong-Da, LIU Shi-Rong, LUO Ming-Xia, HU Jing, LIU Xing-Liang, LI Ya-Fei, YU Hao, OU Ding-Hua. Characteristics of soil carbon and nitrogen contents and enzyme activities in sub-alpine secondary forests with different successional stages in Western Sichuan, China. Chinese Journal of Plant Ecology, 2020, 44(9): 973-985. DOI: 10.17521/cjpe.2020.0203
ɭ�������л�̼�����������Ƿ�ӳ����������ɭ�ֽ���״������Ҫָ��, ֱ�ӻ��ӵ�Ӱ�쵽ɭ���������������������о���ʾ, ɭ������̼����ԼΪȫ������̼������73% (Post & Kwon, 2000), �ڵ���ȫ��̼ƽ�ⷽ��������Ҫ���á���������Ϊ½��ֲ����̬ϵͳ����Ҫ���(Kauppi et al., 2006), ��ά����������Ժ�����̼��ѭ���а�������Ҫ��ɫ(Yang et al., 2011)��ɭ�ָ�������ı���ֲ��Ⱥ��ṹ��������ɼ����ܶȶ�Ӱ�쵽��������, ���������л�̼����������ֺ�����������Է����仯(Chandra et al., 2016; Feng et al., 2019)�����о���Ϊ, ɭ��ֲ�������������ڽ��������л�̼�ֽ����ʳ�������ֲ�������������л�̼�Ĺ̳�����, ʹ���京����Խϵ�(Ohtsuka et al., 2010; Milcu et al., 2011), Ҳ���о�����, ����̼����������ɭ������������ֳ�����(Deng et al., 2013; ��ѩ��, 2016; Liu et al., 2020)���仯������(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)���Ƚ�����(Tang et al., 2009)�ı仯����, ˵��ɭ�����������̼��������Ӱ���Ϊ���ӡ������л��ʽ����ܵ����������Ӱ��(Sinsabaugh, 2010; Liu et al., 2020), ������������Ҳ���ܵ�����̼����Ч�Ե���Լ(Kooch et al., 2018; Dove et al., 2019)��ɭ������ı�����������, ����������ɻ���֮�����仯, �����ı��������������(Zhang et al., 2017), �������Ҫ��������Դ�ڶ�ֲ�����ֽ���������������Լ�ֲ���ϵ������İ���ø�ϡ�����ø���л�̼���ֽ����ת����Ҫ����(Weintraub et al., 2007; Xu et al., 2017), ���������л�̼���Ļ�������������ø�Ŀֽ�����(Sinsabaugh et al., 2014; Mayer et al., 2020)�������л��ʿ������ܿ��ڰ���ø����, ����ˮ��ø��Ҫ�������̼���ֽ�ת��(�����ࡢ���ۡ���ά�ص�), ������-��������ø(��G)����ά��ˮ��ø(CBH)����-����������������ø(NAG)��; ����ø��Ҫ��������л�̼�����(��ľ����)�����ֽ��������ֳ�����̵�, �����������ø(PHO)����������ø(PEO)��(Luo et al., 2019), ���, ø���ԵĸߵͿɱ��������������������Լ������ֹ���ˮƽ֮��Ĺ�ϵ�����о���ʾ, ����ø�������������ֺ������������(Burns et al., 2013; Das & Mondal, 2019)�����������(Li et al., 2015; Feng et al., 2019)��ͬʱ, �����������Ч��Ҳ��Ӱ���л�̼�͵��ķֽ�ø����, ʹ������P�����л��ʱ��ֳ���ͬ�ķֽ�����(Kang et al., 2009; Zhou et al., 2020)������, ����ø������������ʪ�ȼ�pH�������й�(��ΰΰ��, 2019; Zhou et al., 2020), ˵������ø����������ͷ��������صĸ������йء���Ȼ��������ø�����������л�̼����ֺ�����Ĺ�ϵ�ѱ��㷺��ע, �����ڴ����Ǹ�ɽ��Ȼ�����������л�̼����������������ø����������ʶ������̡�
Table 1 ��1 ��12019��������������ͬ�������Ȼ�����ֺ�ẽ���ɼԭʼ����Ҫ���ͺ��������� Table 1Stand and site characteristics of natural secondary forests and Abies faxoniana primary forest at different stages of succession in 2019 in Miyaluo forest area
���� Forest type
�������� Site characteristic
�������� Stand characteristic
ƽ������ Mean altitude (m)
���� Slope aspect
ƽ���¶� Mean slope (��)
��λ Slope position
�ַ��ܶ� Stand density (tree��hm-2)
ƽ������ Mean height (m)
ƽ���ؾ� Mean DBH (cm)
���ն� Canopy density
�ݱ���Ƕ� Coverage of herb (%)
80-NSF
3 387
���� NE
22
���� Middle slope
2 375
6.35
10.83
0.60
65.2
70-NSF
3 312
���� NW
27
���� Middle slope
1 833
7.25
11.12
0.62
79.3
60-NSF
3 080
���� NE
24
���� Middle slope
1 583
6.83
14.17
0.68
40.5
CK
3 189
���� NE
25
������ Upper-middle slope
1 633
10.56
26.70
0.76
43.3
80-NSF, 70-NSF and 60-NSF indicate the natural secondary forest regenerated after logging of the primary forest in 1980s, 1970s and 1960s, respectively. CK indicates the Abies faxoniana primary forest; DBH indicates diameter at breast height. 80-NSF, 70-NSF��60-NSF�ֱ����20����80��70��60����������ص���Ȼ�����֡�CKΪẽ���ɼԭʼ�֡�
Table 3 ��3 ��3������������ͬ���͵���Ҫ�����������ʺ���Ч���ֺ���(ƽ��ֵ����ƫ��, n = 3) Table 3Main soil physical and chemical properties and soil available nutrients in different forest types in Miyaluo forest area (mean �� SD, n = 3)
80-NSF, 70-NSF and 60-NSF indicate the natural secondary forest regenerated after logging of the primary forest in 1980s, 1970s and 1960s, respectively; CK indicates the Abies faxoniana primary forest. Different lowercase letters indicate significant difference among different forest types (p < 0.05). 80-NSF, 70-NSF��60-NSF�ֱ����20����80��70��60����������ص���Ȼ������; CKΪẽ���ɼԭʼ�֡���ͬСд��ĸ������ͬ���ͼ�IJ�������(p < 0.05)��
Table 4 ��4 ��4������������ͬ���͵�����ø����(ƽ��ֵ����ƫ��, n = 3) Table 4Soil enzyme activities in different natural secondary forests and the primary Abies faxoniana forest in Miyaluo forest area (mean �� SD, n = 3)
���� Forest type
��G (nmol��g-1��h-1)
CBH (mg��g-1��h-1)
NAG (nmol��g-1��h-1)
PHO (mg��g-1��h-1)
PEO (mg��g-1��h-1)
80-NSF
22.64 �� 3.83a
26.15 �� 2.49a
9.76 �� 0.82a
38.83 �� 1.59a
36.19 �� 2.61a
70-NSF
22.30 �� 1.23a
25.66 �� 1.88a
4.01 �� 0.36b
35.09 �� 1.56a
31.24 �� 2.91a
60-NSF
6.20 �� 1.69b
18.53 �� 0.95b
2.37 �� 0.05b
34.82 �� 1.04a
36.69 �� 1.18a
CK
31.54 �� 5.51c
22.60 �� 1.03ab
12.40 �� 0.56c
46.02 �� 1.60b
37.81 �� 1.54a
80-NSF, 70-NSF and 60-NSF indicate the natural secondary forest regenerated after logging of the primary forest in 1980s, 1970s and 1960s, respectively; CK indicates the Abies faxoniana primary forest. ��G, ��-4-glucosidase; CBH, cellulose hydrolysis; NAG, ��-4-N-acetylglucosaminidase; PEO, peroxidase; PHO, phenol oxidase. Different lowercase letters indicate significant difference among different forest types (p < 0.05). 80-NSF, 70-NSF��60-NSF�ֱ����20����80��70��60����������ص���Ȼ������; CKΪẽ���ɼԭʼ�֡���G, ��-4-��������ø; CBH, ��ά��ˮ��ø; NAG, ��-4-N-������������������ø; PEO, ��������ø; PHO, �������ø����ͬСд��ĸ������ͬ���ͼ��������(p < 0.05)��
Table 5 ��5 ��5������������Ȼ�����ָ�����������б���������ָ�������� Table 5Correlation among the soil properties in the natural secondary forests in Miyaluo forest area
��Ŀ Item
DOC ���� DOC content
DON ���� DON content
SOC ���� SOC content
TN���� TN content
TP���� TP content
TK���� TK content
C:N
pH
���� ��ˮ�� SWC
H
D
J
E
�����л�̼���� LFOC content
0.853**
0.431
0.833**
0.805**
0.253
-0.189
-0.564
0.354
0.810**
0.085
0.137
0.204
0.208
�������л�̼���� DOC content
1
0.510
0.857**
0.770**
-0.079
-0.479
-0.456
-0.029
0.869**
0.235
0.242
0.248
0.240
������������� DON content
1
0.712**
0.754**
0.311
-0.596*
-0.837**
-0.451
0.659*
0.683*
0.594*
0.148
0.369
�л�̼���� SOC content
1
0.983**
0.194
-0.450
-0.799**
0.017
0.860**
0.332
0.302
0.111
0.295
ȫ������ TN content
1
0.329
-0.392
-0.885**
0.061
0.847**
0.386
0.334
0.052
0.342
ȫ���� TP content
1
0.444
-0.486
0.408
0.157
0.420
0.282
-0.203
0.632*
ȫ�غ��� TK content
1
0.425
0.744**
-0.483
-0.347
-0.390
-0.306
0.212
̼���� C:N
1
0.060
-0.710**
-0.519
-0.431
0.040
-0.307
����pH Soil pH
1
0.013
-0.421
-0.410
-0.247
0.082
������ˮ�� SWC
1
0.340
0.280
0.051
0.222
D, Simpson dominance index; E, Margalef richness index; H, Shannon-Wiener diversity index; J, Pielou evenness index. DOC, dissolved organic carbon; DON, dissolved organic nitrogen; LFOC, light group organic carbon; SOC, soil organic carbon; SWC, soil water content; TK, total potassium; TN, total nitrogen; TP, total phosphorus. * means correlation is significant at 0.05 level, ** means correlation is significant at 0.01 level. D, Simpsonָ��; E, Margalef�ḻ��ָ��; H, Shannon-Wienerָ��; J, Pielou���ȶ�ָ����*��ʾ�������(p < 0.05), **��ʾ���������(p < 0.01)��
Table 6 ��6 ��6��������������ø�����������������ʺ����ֶ����Ե������ Table 6Correlation among soil enzyme activity, soil physicochemical properties and species diversity in Miyaluo forest area
Fig. 2Redundancy analysis on the relationship between soil enzyme activity and soil physicochemical properties, dissolved organic carbon and nitrogen, light fraction organic carbon in different forest types in Miyaluo forest area. CK, Abies faxoniana primary forest; 80-NSF, 70-NSF and 60-NSF indicate the natural secondary forest regenerated after logging of the primary forest in 1980s, 1970s and 1960s, respectively. D, Simpson dominance index; H, Shannon-Wiener diversity index. C:N, soil carbon-nitrogen ratio; DOC, dissolved organic carbon; DON, dissolved organic nitrogen; LFOC, light fraction organic carbon; pH, soil acidity-alkalinity; SOC, soil organic carbon; SWC, soil water content; TN, total nitrogen; TK, total potassium. ��G, ��-4-glucosidase; CBH, cellulose hydrolysis; NAG, ��-4-N-acetylglucosaminidase; PEO, peroxidase; PHO, phenol oxidase.
3 ����
3.1 �����ֲ�ͬ����ζ�����̼��������Ӱ��
ɭ����Ȼ����ʹ��������ɡ������������Լ�ֲ���ϵ�����仯��Ӱ�쵽����̼��ѭ������(��Ծ�µ�, 2013; Dhillon & van Rees, 2017)�����о���Ϊ, ��������������ٽ�����������������̼�ֽ��ͷŶ�����������ЧӦ��, ����������ԭ�е��л��ʷֽ�(Saiya-Cork et al., 2002; Qiao et al., 2019), ����ɭ�ָ�������Ľ���, ���뵽�����ĵ�����������(��Զ����, 2019), �Ӷ���ʹ����ԭ�д���IJ����л��ʷֽ�, ��ֲ���ϵ��������������������л��ʷֽ�����(Adamczyk et al., 2019), ʹ�������л�̼��������ɭ�ָ�������������½�����, ��Ҫ��Ϊ�����л�̼�������ܵ�ֲ�������Լ�������������ɵ�Ӱ��(del Valle et al., 2020)�����о���, ԭʼ��ɼ��(CK)��SOC��TN������������3����Ȼ������(p < 0.05), ����3����Ȼ��������, SOC��������Ϊ80-NSF > 70-NSF > 60-NSF��TNΪ70-NSF > 80-NSF > 60-NSF, ����80-NSF��70-NSF���첻����(p > 0.05), һ�������ڲɷ�������Ȼ������������ֵ����ڽ�, ���������ı仯�������������ۻ����л��ʷֽ�����, �Լ���Ҷ�����е�̼��ľ���ض������Ҷ�����ҷֽ����ʽϿ�(���͵�, 2017); ��һ������Ȼ������������60��, �������ֽ����Ѷ�������ʹ���ֶ����Գ��ֽ�������(ͼ3), ����DON�������ٶ�Ӱ�쵽����SOC��TN����������, �ڴ����Ǹ�ɽ�ߺ���������������������̬��������������, ��ɭ�������ƻ���, �������������Իָ������о�����60���ڲ�ͬ����εĴ����ֽ����˹۲�, ������̼����̬�仯���Ƶ���������, ���о������ʾ, ���Ҫ�ָ����ɷ�ǰԭʼ�ֵ���������ˮƽ(��3)��Ҫ������ʱ�䡣
ͼ3
�´��ڴ�|����ԭͼZIP|����PPT ͼ3������������ͬ���������ֲ�����Ͷ�����ָ�� (ƽ��ֵ����ƫ��, n = 3)��80-NSF, 70-NSF��60-NSF�ֱ����20����80��70��60����������ص���Ȼ������, CKΪẽ���ɼԭʼ�֡�
Fig. 3Diversity index (mean �� SD) of the secondary forests with different successional stages in Miyaluo forest area. 80-NSF, 70-NSF and 60-NSF indicate the natural secondary forest regenerated after logging of the primary forest in 1980s, 1970s and 1960s, respectively. CK indicates the Abies faxoniana primary forest.
���������л�̼���������������л��ʵ���Ҫ���, ��Ҫ��Դ���½���֦��Ҷ����ֳ�ʡ���ϵ�������Լ��������(Kalbitz et al., 2000; ���µ�, 2018)��ɭ�ֲ�ͬ��Ȼ���»ָ���, ���ַ������������Իָ���������ɵIJ���, ���µر�������������������������ʪ�ȡ�������ɵȷ�����ͬ�̶ȱ仯, �Ӷ�Ӱ�쵽���������л�̼�������ı仯(��ѩ��, 2016; Hagedorn et al., 2019)�����о���, DOC��LFOC��DON�����������������������½�����(p < 0.05), ����70-NSF��80-NSF�в��첻����(��3), �����浽ẽ���ɼԭʼ�ֽ�, ����������̼��������������(p < 0.05), ����������˹�����������б���(0-20 cm)��������̼���������о��������(������, 2009), ����ͬ���ȴ�(���ܵ�, 2019)�����ȴ�(��Ծ�µ�, 2013)ɭ��������о����, ˵����ͬ��ʪ������ֲ���ַֽṹ�����������л�̼����ֺ�����Ӱ����ֳ���ͬ�ı仯����(Li et al., 2014; Taylor et al., 2019)������ԭ��, һ��������ɭ�ָ�������, ����̼������������(Mylliemngap et al., 2016), ����Ҷ�ֵĿ����ʸ�����Ҷ��(C?t�� et al., 2000); ��һ�������ڱ��������ܶ�����������, �������ž۶ȡ��ṹϵ�����ܿ�϶��ë�ܿ�϶������������������ֽ�������(��ѧ�µ�, 2003), �����л�̼����ȱ�������ž���������Ա���, �������������л�̼��DON�������͡�����, ẽ���ɼԭʼ�ֵ���������̼����ֺ���������Ȼ������(��3), ��������ַ���Ҫ���ֹ���(��Ҷ+��Ҷ����)��һ������, Դ�ڸ������������ֵ���Ҷ�ں���һ�����ڷֽ�(����÷��, 2015), ���ͷŵ����ֱ�������ֽ�����, �������������л�̼�������ӡ���Ҫ��������Ӱ�����ض����������л�̼������������, �����һ�������о�����ط�������, ��ͬ��Ȼ����������ֵر�������SOC��TN����������л�̼����ֺ�����DON�������ʼ���������ع�ϵ(p < 0.01), ˵�����������л�̼�͵�����������SOC��TN����֮���ϵ��Ϊ����, ���������о�(�����, 2016; ������, 2017) һ��, ˵����Ȼ�����������������ǵ�����̼�ֵ̳���Ҫ����(Li et al., 2012; Smal et al., 2019)��
3.2 ��ͬ��������δ���������ø���Եı仯����Ӱ������
ɭ�ָ�������Ļָ�������, �ַֽṹ��������ɡ����������Լ������������˲�ͬ�̶ȵı仯, ��������������̼��Դ��ͬ�����������л��ʷֽ����ʡ�������������Լ����л������仯(Wardle et al., 2004; Lucas-Borja & Delgado- Baquerizo, 2019), ����Ӱ�쵽����ø����(Adamczyk et al., 2015; Xu et al., 2017)������, ��ƽ�����¡���ˮ������������Ժ����ֶ�����Ҳ��Ӱ������ø���Ե���Ҫ����(�����, 2014; Zhou et al., 2020)�����о���, ��Ȼ�����ָ�����������(<60 a)��ͬ�εı�������ˮ��ø(��G��CBH��NAG)������������ẽ���ɼԭʼ��(��4), ������Ȼ����������̳�������������(80-NSF > 70-NSF > 60-NSF), ��ͬ�ڴ����Ǹ�ɽƽ���ذ�����ˮ��ø���Ը�����ɼ��(�ܺ��ٵ�, 2004)���´�����(Betula platyphylla)����������̱���Ϊ�����Ӻ͵ı仯����(���Ե�, 2013), һ����˵��ẽ���ɼԭʼ�ֵ���������ѭ��������Խϸ�, ���������ֹ�������; ��һ����˵����������Ȼ�������������, ���ϵ��������ӵ�������̼�����������ֺ������½�(Liu et al., 2017; Lyu et al., 2019), ʹ������������Խ���(�����, 2018), ������������ˮ��ø�����½�������, 80-NSF��70-NSF��60-NSF����������ø���Ա仯������, ��PHO�Ļ�����������CK, ��PEO�Ļ�����CK���첻����(��4), ��ԭ�������ẽ���ɼԭʼ���к��н�Ϊ�ḻ��ľ���ء����ʺ����ʵ��ѽ��⻯����, �ٽ��˽���ľ���ص�����Ⱥ������Ի�������������, ��������ָ�������εĽ���, ��ϵ��������ֲ��������������仯, ����������pH����, ��ʹPHO��������(Fujii et al., 2013)��
BaoY, GaoY, ZengXM, YuanP, SiYT, ChenYM, ChenYY ( 2018). Relationships between carbon and nitrogen contents and enzyme activities in soil of three typical subtropical forests in China Chinese Journal of Plant Ecology, 42, 508-516. DOI:10.17521/cjpe.2017.0311URL
BurnsRG, DeForestJL, MarxsenJ, SinsabaughRL, StrombergerME, WallensteinMD, WeintraubMN, ZoppiniA ( 2013). Soil enzymes in a changing environment: current knowledge and future directions Soil Biology & Biochemistry, 58, 216-234. DOI:10.1016/j.soilbio.2012.11.009URL [��������: 1]
ChandraLR, GuptaS, PandeV, SinghN ( 2016). Impact of forest vegetation on soil characteristics: a correlation between soil biological and physico-chemical properties 3 Biotech, 6, 188. DOI: 10.1007/s13205-016-0510-y. DOI:10.1007/s13205-016-0510-yURL [��������: 1]
ChenJ, LuoY, LiJ, ZhouX, CaoJ, WangR-W, WangY, SheltonS, JinZ, WalkerLM, FengZ, NiuS, FengW, JianS, ZhouL ( 2017). Costimulation of soil glycosidase activity and soil respiration by nitrogen addition Global Change Biology, 23, 1328-1337. DOI:10.1111/gcb.13402URL [��������: 1]
ChenQM, WangZX, LiuY, ZhengHF, LiHJ, WangLF, ChenYM, ChenX, TangSY ( 2019). Response of soil enzyme activity and stoichiometric ratio to simulated nitrogen deposition in subalpine coniferous forests of western Sichuan Chinese Journal of Applied & Environmental Biology, 25, 791-800.
C?t��L, BrownS, Par��D, FylesJ, BauhusJ ( 2000). Dynamics of carbon and nitrogen mineralization in relation to stand type, stand age and soil texture in the boreal mixedwood Soil Biology & Biochemistry, 32, 1079-1090. DOI:10.1016/S0038-0717(00)00017-1URL
DasC, MondalNK ( 2019). Temporal and vertical variation of selected extracellular enzyme activities on tree litter degradation of a subtropical forest Agricultural Research, 8, 84-91. DOI:10.1007/s40003-018-0353-3URL [��������: 1]
DeForestJL ( 2009). The influence of time, storage temperature, and substrate age on potential soil enzyme activity in acidic forest soils using MUB-linked substrates and l-DOPA Soil Biology & Biochemistry, 41, 1180-1186. DOI:10.1016/j.soilbio.2009.02.029URL [��������: 1]
del ValleI, WebsterTM, ChengHY, ThiesJE, KesslerA, MillerMK, BallZT, MacKenzieKR, MasielloCA, SilbergJJ, LehmannJ ( 2020). Soil organic matter attenuates the efficacy of flavonoid-based plant-microbe communication Science Advances, 6, eaax8254. DOI:10.1126/ sciadv.aax8254. [��������: 1]
DengL, WangKB, ChenML, ShangguanZP, SweeneyS ( 2013). Soil organic carbon storage capacity positively related to forest succession on the Loess Plateau, China Catena, 110, 1-7. DOI:10.1016/j.catena.2013.06.016URL [��������: 1]
DhillonGS, van ReesKCJ ( 2017). Distribution of soil organic carbon in the light and heavy fractions for six shelterbelt species and their adjacent agricultural fields in Saskatchewan Canadian Journal of Soil Science, 97, 732-744. [��������: 2]
DoveNC, StarkJM, NewmanGS, HartSC ( 2019). Carbon control on terrestrial ecosystem function across contrasting site productivities: the carbon connection revisited Ecology, 100, e02695. DOI: 10.1002/ecy.2695. [��������: 1]
FanYX, YangYS, YangZJ, XieJS, ChenGS, ZhongXJ, GuoJF ( 2013). Seasonal dynamics and content of soil labile organic carbon of mid-subtropical evergreen broadleaved forest during natural succession Acta Ecologica Sinica, 33, 5751-5759. DOI:10.5846/stxbURL [��������: 1]
FengC, MaYH, JinX, WangZ, MaY, FuSL, ChenHYH ( 2019). Soil enzyme activities increase following restoration of degraded subtropical forests Geoderma, 351, 180-187. DOI:10.1016/j.geoderma.2019.05.006URL [��������: 1]
FujiiK, UemuraM, HayakawaC, FunakawaS, KosakiT ( 2013). Environmental control of lignin peroxidase, manganese peroxidase, and laccase activities in forest floor layers in humid Asia Soil Biology & Biochemistry, 57, 109-115. DOI:10.1016/j.soilbio.2012.07.007URL
GongL, LiuGH, LiZS, YeX, WangH ( 2017). Altitudinal changes in nitrogen, organic carbon, and its labile fractions in different soil layers in an Abies faxoniana forest in Wolong Acta Ecologica Sinica, 37, 4696-4705. [��������: 2]
GuX, ZhangSJ, LiuZD, LiLD, ChenJL, WangLF, FangX ( 2018). Effects of vegetation restoration on soil organic carbon concentration and density in the mid-subtropical region of China Chinese Journal of Plant Ecology, 42, 595-608. DOI:10.17521/cjpe.2018.0021URL [��������: 1]
GuX, ZhangSJ, XiangWH, LiLD, LiuZD, SunWJ, FangX ( 2016). Seasonal dynamics of active soil organic carbon in four subtropical forests in Southern China Chinese Journal of Plant Ecology, 40, 1064-1076. DOI:10.17521/cjpe.2015.0412URL [��������: 1]
HagedornF, GavazovK, AlexanderJM ( 2019). Above- and belowground linkages shape responses of mountain vegetation to climate change Science, 365, 1119-1123. DOI:10.1126/science.aax4737URL
HuS, ZhangY, ShiRJ, HanSQ, LiH, XuH ( 2013). Temporal variations of soil microbial biomass and enzyme activities during the secondary succession of primary broadleaved- Pinus koraiensis forests in Changbai Mountains of Northeast. Chinese Journal of Applied Ecology, 24, 366-372. [��������: 1]
HuangYM, YangWQ, ZhangJ ( 2015). Process of leaf litter mass loss and the contributions of soil organisms in Picea aspoerata plantations of western Sichuan. Resources and Environment in the Yangtze Basin, 24, 676-683.
JiangFY, SunH, LinB, LiuQ ( 2009). Dynamic changes of topsoil organic carbon in subalpine spruce plantation at different succession stages in western Sichuan Province Chinese Journal of Applied Ecology, 20, 2581-2587. [��������: 1]
KalbitzK, SolingerS, ParkJH, MichalzikB, MatznerE ( 2000). Controls on the dynamics of dissolved organic matter in soils: a review Soil Science, 165, 277-304. DOI:10.1097/00010694-200004000-00001URL
KangH, KangS, LeeD ( 2009). Variations of soil enzyme activities in a temperate forest soil Ecological Research, 24, 1137-1143. DOI:10.1007/s11284-009-0594-5URL [��������: 1]
KauppiPE, AusubelJH, FangJ, MatherAS, SedjoRA, WaggonerPE ( 2006). Returning forests analyzed with the forest identity Proceedings of the National Academy of Sciences of the United States of America, 103, 17574-17579. [��������: 1]
KoochY, SanjiR, TabariM ( 2018). Increasing tree diversity enhances microbial and enzyme activities in temperate Iranian forests Trees, 32, 809-822. DOI:10.1007/s00468-018-1674-3URL
LiDJ, NiuSL, LuoYQ ( 2012). Global patterns of the dynamics of soil carbon and nitrogen stocks following afforestation: a meta-analysis New Phytologist, 195, 172-181. DOI:10.1111/j.1469-8137.2012.04150.xURL [��������: 1]
LiJJ, ZhouXM, YanJX, LiHJ, HeJZ ( 2015). Effects of regenerating vegetation on soil enzyme activity and microbial structure in reclaimed soils on a surface coal mine site Applied Soil Ecology, 87, 56-62. DOI:10.1016/j.apsoil.2014.11.010URL
LiLQ, WangD, LiuXY, ZhangB, LiuYZ, XieT, DuYX, PanGX ( 2014). Soil organic carbon fractions and microbial community and functions under changes in vegetation: a case of vegetation succession in karst forest Environmental Earth Sciences, 71, 3727-3735. DOI:10.1007/s12665-013-2767-3URL [��������: 1]
LiYY, ShaoMA ( 2004). The change of plant diversity during natural recovery process of vegetation in Ziwuling area Acta Ecologica Sinica, 24, 252-260.
Lucas-BorjaME, Delgado-BaquerizoM ( 2019). Plant diversity and soil stoichiometry regulates the changes in multifunctionality during pine temperate forest secondary succession Science of the Total Environment, 697, 134204. DOI: 10.1016/j.scitotenv.2019.134204. DOI:10.1016/j.scitotenv.2019.134204URL [��������: 1]
LuoXZ, HouEQ, ZhangLL, ZangXW, YiYF, ZhangGH, WenDZ ( 2019). Effects of forest conversion on carbon-degrading enzyme activities in subtropical China Science of the Total Environment, 696, 133968. DOI: 10.1016/ j.scitotenv.2019.133968. DOI:10.1016/j.scitotenv.2019.133968URL [��������: 1]
LyuM, LiX, XieJ, HomyakPM, UkonmaanahoL, YangZ, LiuX, RuanC, YangY ( 2019). Root-microbial interaction accelerates soil nitrogen depletion but not soil carbon after increasing litter inputs to a coniferous forest Plant and Soil, 444, 153-164. DOI:10.1007/s11104-019-04265-wURL [��������: 1]
MaWW, WangLX, LiN, ZhengDH, XieLL, LiuQ, YinCY ( 2019). Dynamic effects of nitrogen deposition on soil enzyme activities in soils with different moisture content Acta Ecologica Sinica, 39, 7218-7228. [��������: 1]
MayerM, PrescottCE, AbakerWEA, AugustoL, C��cillonL, FerreiraGWD, JamesJ, JandlR, KatzensteinerK, LaclauJ-P, Lagani��reJ, NouvellonY, Par��D, StanturfJA, VanguelovaEI, VesterdalL ( 2020). Tamm Review: Influence of forest management activities on soil organic carbon stocks: a knowledge synthesis Forest Ecology and Management, 466, 118127. DOI: 10.1016/j.foreco.2020.118127. [��������: 1]
MilcuA, HeimA, EllisRJ, ScheuS, ManningP ( 2011). Identification of general patterns of nutrient and labile carbon control on soil carbon dynamics across a successional gradient Ecosystems, 14, 710-719. DOI:10.1007/s10021-011-9440-zURL
MylliemngapW, NathD, BarikSK ( 2016). Changes in vegetation and nitrogen mineralization during recovery of a montane subtropical broadleaved forest in North-eastern India following anthropogenic disturbance Ecological Research, 31, 21-38. DOI:10.1007/s11284-015-1309-8URL [��������: 1]
OhtsukaT, ShizuY, NishiwakiA, YashiroY, KoizumiH ( 2010). Carbon cycling and net ecosystem production at an early stage of secondary succession in an abandoned coppice forest Journal of Plant Research, 123, 393-401. [��������: 1]
PangDB, CuiM, LiuYG, WangGZ, CaoJH, WangXR, DanXQ, ZhouJX ( 2019). Responses of soil labile organic carbon fractions and stocks to different vegetation restoration strategies in degraded karst ecosystems of southwest China Ecological Engineering, 138, 391-402. [��������: 1]
PangXY, LiuSQ, LiuQ, WuY, LinB, HeH, ZhangZJ ( 2003). Influence of plant community succession on soil physical properties during subalpine coniferous plantation rehabilitation in western Sichuan Journal of Soil and Water Conservation, 17(4), 42-45, 50.
PangXY, NingW, QingL, BaoWK ( 2009). The relation among soil microorganism, enzyme activity and soil nutrients under subalpine coniferous forest in Western Sichuan Acta Ecologica Sinica, 29, 286-292. DOI:10.1016/j.chnaes.2009.09.005URL
PushkarevaE, EckhardtKU, HotterV, FrossardA, LeinweberP, FreyB, KarstenU ( 2020). Chemical composition of soil organic matter and potential enzyme activity in the topsoil along a moisture gradient in the High Arctic (Svalbard) Geoderma, 368, 114304. DOI: 10.1016/j.geoderma. 2020. 114304. DOI:10.1016/j.geoderma.2020.114304URL [��������: 1]
QiaoN, WangJ, XuX, ShenY, LongX, HuY, SchaeferD, LiS, WangH, KuzyakovY ( 2019). Priming alters soil carbon dynamics during forest succession Biology and Fertility of Soils, 55, 339-350. DOI:10.1007/s00374-019-01351-0URL [��������: 1]
Saiya-CorkKR, SinsabaughRL, ZakDR ( 2002). The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil Soil Biology & Biochemistry, 34, 1309-1315. DOI:10.1016/S0038-0717(02)00074-3URL
SchedlbauerJL, KavanaghKL ( 2008). Soil carbon dynamics in a chronosequence of secondary forests in northeastern Costa Rica Forest Ecology and Management, 255, 1326-1335. [��������: 1]
ShenX, LiuY, TangSY, YangS, ChenYM, YangL, ZhengHF, LiHJ ( 2017). Characteristics of substrate quality variation at different litter decomposition stages in subalpine forest of Western Sichuan Acta Botanica Boreali- Occidentalia Sinica , 37, 586-594. [��������: 1]
SmalH, Lig?zaS, PranagalJ, UrbanD, Pietruczyk-Pop?awskaD ( 2019). Changes in the stocks of soil organic carbon, total nitrogen and phosphorus following afforestation of post-arable soils: a chronosequence study Forest Ecology and Management, 451, 117536. DOI: 10.1016/j.foreco.2019.117536. DOI:10.1016/j.foreco.2019.117536URL [��������: 1]
TangJ, BolstadPV, MartinJG ( 2009). Soil carbon fluxes and stocks in a Great Lakes forest chronosequence Global Change Biology, 15, 145-155. DOI:10.1111/gcb.2009.15.issue-1URL [��������: 1]
TaylorAR, WangJR, ChenHYH ( 2007). Carbon storage in a chronosequence of red spruce (Picea rubens) forests in central Nova Scotia, Canada Canadian Journal of Forest Research, 37, 2260-2269. DOI:10.1139/X07-080URL
TaylorBN, ChazdonRL, MengeDNL ( 2019). Successional dynamics of nitrogen fixation and forest growth in regenerating Costa Rican rainforests Ecology, 100, e02637. DOI: 10.1002/ecy.2637. [��������: 1]
WardleDA, BardgettRD, KlironomosJN, Set?l?H, van der PuttenWH, WallDH ( 2004). Ecological linkages between aboveground and belowground Biota Science, 304, 1629-1633. DOI:10.1126/science.1094875URL [��������: 1]
WeintraubMN, Scott-DentonLE, SchmidtSK, MonsonRK ( 2007). The effects of tree rhizodeposition on soil exoenzyme activity, dissolved organic carbon, and nutrient availability in a subalpine forest ecosystem Oecologia, 154, 327-338. DOI:10.1007/s00442-007-0804-1URL [��������: 1]
XiongHZ, WangKY, YangWQ ( 2004). Seasonal variations of soil enzyme activities in fir and birch forests in subalpine area of Western Sichuan Chinese Journal of Applied & Environmental Biology, 4, 416-420. [��������: 1]
XuZW, YuGR, ZhangXY, HeNP, WangQF, WangSZ, WangRL, ZhaoN, JiaYL, WangCY ( 2017). Soil enzyme activity and stoichiometry in forest ecosystems along the North-South Transect in eastern China (NSTEC) Soil Biology & Biochemistry, 104, 152-163. DOI:10.1016/j.soilbio.2016.10.020URL [��������: 1]
YangY, LuoY, FinziAC ( 2011). Carbon and nitrogen dynamics during forest stand development: a global synthesis New Phytologist, 190, 977-989. DOI:10.1111/nph.2011.190.issue-4URL [��������: 1]
YuY, ZouL, SunTT, GuoJ, ZhangGQ, TangQM ( 2014). Relationship between vegetation diversity and soil functional diversity in native pine forests Pratacultural Science, 31, 1241-1247. [��������: 1]
ZhangWW, LuZT, YangK, ZhuJJ ( 2017). Impacts of conversion from secondary forests to larch plantations on the structure and function of microbial communities Applied Soil Ecology, 111, 73-83. DOI:10.1016/j.apsoil.2016.11.019URL
ZhangX, HanSJ, WangSQ, GuY, YueLY, FengY, GengSC, ChenZJ ( 2016). Change of soil organic carbon fractions at different successional stages of Betula platyphylla forest in Changbai Mountains Chinese Journal of Ecology, 35, 282-289. URL [��������: 1] P<0.05) with the increasing soil layer depth. ROC/SOC and POC/SOC ranged between 12.91%-47.95% and 14.21%-69.46% in the five successional stages, respectively. The correlation analysis showed a significant positive correlation (P<0.01) between MBC, ROC, POC and SOC, and a significant positive correlation (P<0.01) between SOC, MBC, ROC, POC and total nitrogen, total phosphorus, carbon to nitrogen ratio. Our results provide data support for clarifying soil organic carbon stability and carbon sequestration potential during the succession of the birch forest.]]> [ ��ѩ, ��ʿ��, ������, ��Խ, ������, ����, ������, ��־�� ( 2016). ����ɽ�����ֲ�ͬ����������л�̼��ֵı仯 ��̬ѧ��־, 35, 282-289.] URL [��������: 1] P<0.05����5������������ROC/SOC��POC/SOC�ı仯��Χ�ֱ�Ϊ12.91%��47.95%��14.21%��69.46%����ط���������MBC��ROC��POC�������������л�̼(SOC)�����ʼ���������أ���P<0.01������SOC��MBC��ROC��POC������ȫ����ȫ��̼���ȳʼ����������(P<0.01)���о����Ϊ�˽����������������������л�̼���ȶ��Ա仯��̼DZ���ṩ����֧�֡�]]>
ZhangYD, LiuYC, GuFX, GuoMM, MiaoN, LiuSR ( 2019). Litter composition and its dynamic in five main forest types in subalpine areas of west Sichuan, China Acta Ecologica Sinica, 39, 502-508. [��������: 1]
ZhouYG, HaoKJ, LiXW, FanC, ChenLL, WangX, WangXH ( 2014). Effect of different land use patterns on seasonal dynamic of soil microbial biomass carbon in the subalpine forest of western Sichuan, China Journal of Natural Resources, 29, 1944-1956. [��������: 1]
Soil enzymes in a changing environment: current knowledge and future directions 1 2013
... ���������л�̼���������������л��ʵ���Ҫ���, ��Ҫ��Դ���½���֦��Ҷ����ֳ�ʡ���ϵ�������Լ��������(Kalbitz et al., 2000; ���µ�, 2018).ɭ�ֲ�ͬ��Ȼ���»ָ���, ���ַ������������Իָ���������ɵIJ���, ���µر�������������������������ʪ�ȡ�������ɵȷ�����ͬ�̶ȱ仯, �Ӷ�Ӱ�쵽���������л�̼�������ı仯(��ѩ��, 2016; Hagedorn et al., 2019).���о���, DOC��LFOC��DON�����������������������½�����(p < 0.05), ����70-NSF��80-NSF�в��첻����(��3), �����浽ẽ���ɼԭʼ�ֽ�, ����������̼��������������(p < 0.05), ����������˹�����������б���(0-20 cm)��������̼���������о��������(������, 2009), ����ͬ���ȴ�(���ܵ�, 2019)�����ȴ�(��Ծ�µ�, 2013)ɭ��������о����, ˵����ͬ��ʪ������ֲ���ַֽṹ�����������л�̼����ֺ�����Ӱ����ֳ���ͬ�ı仯����(Li et al., 2014; Taylor et al., 2019).����ԭ��, һ��������ɭ�ָ�������, ����̼������������(Mylliemngap et al., 2016), ����Ҷ�ֵĿ����ʸ�����Ҷ��(C?t�� et al., 2000); ��һ�������ڱ��������ܶ�����������, �������ž۶ȡ��ṹϵ�����ܿ�϶��ë�ܿ�϶������������������ֽ�������(��ѧ�µ�, 2003), �����л�̼����ȱ�������ž���������Ա���, �������������л�̼��DON��������.����, ẽ���ɼԭʼ�ֵ���������̼����ֺ���������Ȼ������(��3), ��������ַ���Ҫ���ֹ���(��Ҷ+��Ҷ����)��һ������, Դ�ڸ������������ֵ���Ҷ�ں���һ�����ڷֽ�(����÷��, 2015), ���ͷŵ����ֱ�������ֽ�����, �������������л�̼��������.��Ҫ��������Ӱ�����ض����������л�̼������������, �����һ�������о�.��ط�������, ��ͬ��Ȼ����������ֵر�������SOC��TN����������л�̼����ֺ�����DON�������ʼ���������ع�ϵ(p < 0.01), ˵�����������л�̼�͵�����������SOC��TN����֮���ϵ��Ϊ����, ���������о�(�����, 2016; ������, 2017) һ��, ˵����Ȼ�����������������ǵ�����̼�ֵ̳���Ҫ����(Li et al., 2012; Smal et al., 2019). ...
Impact of forest vegetation on soil characteristics: a correlation between soil biological and physico-chemical properties 1 2016
... ɭ�������л�̼�����������Ƿ�ӳ����������ɭ�ֽ���״������Ҫָ��, ֱ�ӻ��ӵ�Ӱ�쵽ɭ������������������.�о���ʾ, ɭ������̼����ԼΪȫ������̼������73% (Post & Kwon, 2000), �ڵ���ȫ��̼ƽ�ⷽ��������Ҫ����.��������Ϊ½��ֲ����̬ϵͳ����Ҫ���(Kauppi et al., 2006), ��ά����������Ժ�����̼��ѭ���а�������Ҫ��ɫ(Yang et al., 2011).ɭ�ָ�������ı���ֲ��Ⱥ��ṹ��������ɼ����ܶȶ�Ӱ�쵽��������, ���������л�̼����������ֺ�����������Է����仯(Chandra et al., 2016; Feng et al., 2019).���о���Ϊ, ɭ��ֲ�������������ڽ��������л�̼�ֽ����ʳ�������ֲ�������������л�̼�Ĺ̳�����, ʹ���京����Խϵ�(Ohtsuka et al., 2010; Milcu et al., 2011), Ҳ���о�����, ����̼����������ɭ������������ֳ�����(Deng et al., 2013; ��ѩ��, 2016; Liu et al., 2020)���仯������(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)���Ƚ�����(Tang et al., 2009)�ı仯����, ˵��ɭ�����������̼��������Ӱ���Ϊ����.�����л��ʽ����ܵ����������Ӱ��(Sinsabaugh, 2010; Liu et al., 2020), ������������Ҳ���ܵ�����̼����Ч�Ե���Լ(Kooch et al., 2018; Dove et al., 2019).ɭ������ı�����������, ����������ɻ���֮�����仯, �����ı��������������(Zhang et al., 2017), �������Ҫ��������Դ�ڶ�ֲ�����ֽ���������������Լ�ֲ���ϵ������İ���ø��.����ø���л�̼���ֽ����ת����Ҫ����(Weintraub et al., 2007; Xu et al., 2017), ���������л�̼���Ļ�������������ø�Ŀֽ�����(Sinsabaugh et al., 2014; Mayer et al., 2020).�����л��ʿ������ܿ��ڰ���ø����, ����ˮ��ø��Ҫ�������̼���ֽ�ת��(�����ࡢ���ۡ���ά�ص�), ������-��������ø(��G)����ά��ˮ��ø(CBH)����-����������������ø(NAG)��; ����ø��Ҫ��������л�̼�����(��ľ����)�����ֽ��������ֳ�����̵�, �����������ø(PHO)����������ø(PEO)��(Luo et al., 2019), ���, ø���ԵĸߵͿɱ��������������������Լ������ֹ���ˮƽ֮��Ĺ�ϵ.���о���ʾ, ����ø�������������ֺ������������(Burns et al., 2013; Das & Mondal, 2019)�����������(Li et al., 2015; Feng et al., 2019).ͬʱ, �����������Ч��Ҳ��Ӱ���л�̼�͵��ķֽ�ø����, ʹ������P�����л��ʱ��ֳ���ͬ�ķֽ�����(Kang et al., 2009; Zhou et al., 2020).����, ����ø������������ʪ�ȼ�pH�������й�(��ΰΰ��, 2019; Zhou et al., 2020), ˵������ø����������ͷ��������صĸ������й�.��Ȼ��������ø�����������л�̼����ֺ�����Ĺ�ϵ�ѱ��㷺��ע, �����ڴ����Ǹ�ɽ��Ȼ�����������л�̼����������������ø����������ʶ�������. ...
Costimulation of soil glycosidase activity and soil respiration by nitrogen addition 1 2017
... ɭ�������л�̼�����������Ƿ�ӳ����������ɭ�ֽ���״������Ҫָ��, ֱ�ӻ��ӵ�Ӱ�쵽ɭ������������������.�о���ʾ, ɭ������̼����ԼΪȫ������̼������73% (Post & Kwon, 2000), �ڵ���ȫ��̼ƽ�ⷽ��������Ҫ����.��������Ϊ½��ֲ����̬ϵͳ����Ҫ���(Kauppi et al., 2006), ��ά����������Ժ�����̼��ѭ���а�������Ҫ��ɫ(Yang et al., 2011).ɭ�ָ�������ı���ֲ��Ⱥ��ṹ��������ɼ����ܶȶ�Ӱ�쵽��������, ���������л�̼����������ֺ�����������Է����仯(Chandra et al., 2016; Feng et al., 2019).���о���Ϊ, ɭ��ֲ�������������ڽ��������л�̼�ֽ����ʳ�������ֲ�������������л�̼�Ĺ̳�����, ʹ���京����Խϵ�(Ohtsuka et al., 2010; Milcu et al., 2011), Ҳ���о�����, ����̼����������ɭ������������ֳ�����(Deng et al., 2013; ��ѩ��, 2016; Liu et al., 2020)���仯������(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)���Ƚ�����(Tang et al., 2009)�ı仯����, ˵��ɭ�����������̼��������Ӱ���Ϊ����.�����л��ʽ����ܵ����������Ӱ��(Sinsabaugh, 2010; Liu et al., 2020), ������������Ҳ���ܵ�����̼����Ч�Ե���Լ(Kooch et al., 2018; Dove et al., 2019).ɭ������ı�����������, ����������ɻ���֮�����仯, �����ı��������������(Zhang et al., 2017), �������Ҫ��������Դ�ڶ�ֲ�����ֽ���������������Լ�ֲ���ϵ������İ���ø��.����ø���л�̼���ֽ����ת����Ҫ����(Weintraub et al., 2007; Xu et al., 2017), ���������л�̼���Ļ�������������ø�Ŀֽ�����(Sinsabaugh et al., 2014; Mayer et al., 2020).�����л��ʿ������ܿ��ڰ���ø����, ����ˮ��ø��Ҫ�������̼���ֽ�ת��(�����ࡢ���ۡ���ά�ص�), ������-��������ø(��G)����ά��ˮ��ø(CBH)����-����������������ø(NAG)��; ����ø��Ҫ��������л�̼�����(��ľ����)�����ֽ��������ֳ�����̵�, �����������ø(PHO)����������ø(PEO)��(Luo et al., 2019), ���, ø���ԵĸߵͿɱ��������������������Լ������ֹ���ˮƽ֮��Ĺ�ϵ.���о���ʾ, ����ø�������������ֺ������������(Burns et al., 2013; Das & Mondal, 2019)�����������(Li et al., 2015; Feng et al., 2019).ͬʱ, �����������Ч��Ҳ��Ӱ���л�̼�͵��ķֽ�ø����, ʹ������P�����л��ʱ��ֳ���ͬ�ķֽ�����(Kang et al., 2009; Zhou et al., 2020).����, ����ø������������ʪ�ȼ�pH�������й�(��ΰΰ��, 2019; Zhou et al., 2020), ˵������ø����������ͷ��������صĸ������й�.��Ȼ��������ø�����������л�̼����ֺ�����Ĺ�ϵ�ѱ��㷺��ע, �����ڴ����Ǹ�ɽ��Ȼ�����������л�̼����������������ø����������ʶ�������. ...
�����Ǹ�ɽ��Ҷ������ø���仯ѧ�����ȶ�ģ���������Ӧ 2019
�����Ǹ�ɽ��Ҷ������ø���仯ѧ�����ȶ�ģ���������Ӧ 2019
Dynamics of carbon and nitrogen mineralization in relation to stand type, stand age and soil texture in the boreal mixedwood 2000
Temporal and vertical variation of selected extracellular enzyme activities on tree litter degradation of a subtropical forest 1 2019
The influence of time, storage temperature, and substrate age on potential soil enzyme activity in acidic forest soils using MUB-linked substrates and l-DOPA 1 2009
... ���������л�̼���������������л��ʵ���Ҫ���, ��Ҫ��Դ���½���֦��Ҷ����ֳ�ʡ���ϵ�������Լ��������(Kalbitz et al., 2000; ���µ�, 2018).ɭ�ֲ�ͬ��Ȼ���»ָ���, ���ַ������������Իָ���������ɵIJ���, ���µر�������������������������ʪ�ȡ�������ɵȷ�����ͬ�̶ȱ仯, �Ӷ�Ӱ�쵽���������л�̼�������ı仯(��ѩ��, 2016; Hagedorn et al., 2019).���о���, DOC��LFOC��DON�����������������������½�����(p < 0.05), ����70-NSF��80-NSF�в��첻����(��3), �����浽ẽ���ɼԭʼ�ֽ�, ����������̼��������������(p < 0.05), ����������˹�����������б���(0-20 cm)��������̼���������о��������(������, 2009), ����ͬ���ȴ�(���ܵ�, 2019)�����ȴ�(��Ծ�µ�, 2013)ɭ��������о����, ˵����ͬ��ʪ������ֲ���ַֽṹ�����������л�̼����ֺ�����Ӱ����ֳ���ͬ�ı仯����(Li et al., 2014; Taylor et al., 2019).����ԭ��, һ��������ɭ�ָ�������, ����̼������������(Mylliemngap et al., 2016), ����Ҷ�ֵĿ����ʸ�����Ҷ��(C?t�� et al., 2000); ��һ�������ڱ��������ܶ�����������, �������ž۶ȡ��ṹϵ�����ܿ�϶��ë�ܿ�϶������������������ֽ�������(��ѧ�µ�, 2003), �����л�̼����ȱ�������ž���������Ա���, �������������л�̼��DON��������.����, ẽ���ɼԭʼ�ֵ���������̼����ֺ���������Ȼ������(��3), ��������ַ���Ҫ���ֹ���(��Ҷ+��Ҷ����)��һ������, Դ�ڸ������������ֵ���Ҷ�ں���һ�����ڷֽ�(����÷��, 2015), ���ͷŵ����ֱ�������ֽ�����, �������������л�̼��������.��Ҫ��������Ӱ�����ض����������л�̼������������, �����һ�������о�.��ط�������, ��ͬ��Ȼ����������ֵر�������SOC��TN����������л�̼����ֺ�����DON�������ʼ���������ع�ϵ(p < 0.01), ˵�����������л�̼�͵�����������SOC��TN����֮���ϵ��Ϊ����, ���������о�(�����, 2016; ������, 2017) һ��, ˵����Ȼ�����������������ǵ�����̼�ֵ̳���Ҫ����(Li et al., 2012; Smal et al., 2019). ...
Soil organic matter attenuates the efficacy of flavonoid-based plant-microbe communication 1 2020
... ɭ�������л�̼�����������Ƿ�ӳ����������ɭ�ֽ���״������Ҫָ��, ֱ�ӻ��ӵ�Ӱ�쵽ɭ������������������.�о���ʾ, ɭ������̼����ԼΪȫ������̼������73% (Post & Kwon, 2000), �ڵ���ȫ��̼ƽ�ⷽ��������Ҫ����.��������Ϊ½��ֲ����̬ϵͳ����Ҫ���(Kauppi et al., 2006), ��ά����������Ժ�����̼��ѭ���а�������Ҫ��ɫ(Yang et al., 2011).ɭ�ָ�������ı���ֲ��Ⱥ��ṹ��������ɼ����ܶȶ�Ӱ�쵽��������, ���������л�̼����������ֺ�����������Է����仯(Chandra et al., 2016; Feng et al., 2019).���о���Ϊ, ɭ��ֲ�������������ڽ��������л�̼�ֽ����ʳ�������ֲ�������������л�̼�Ĺ̳�����, ʹ���京����Խϵ�(Ohtsuka et al., 2010; Milcu et al., 2011), Ҳ���о�����, ����̼����������ɭ������������ֳ�����(Deng et al., 2013; ��ѩ��, 2016; Liu et al., 2020)���仯������(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)���Ƚ�����(Tang et al., 2009)�ı仯����, ˵��ɭ�����������̼��������Ӱ���Ϊ����.�����л��ʽ����ܵ����������Ӱ��(Sinsabaugh, 2010; Liu et al., 2020), ������������Ҳ���ܵ�����̼����Ч�Ե���Լ(Kooch et al., 2018; Dove et al., 2019).ɭ������ı�����������, ����������ɻ���֮�����仯, �����ı��������������(Zhang et al., 2017), �������Ҫ��������Դ�ڶ�ֲ�����ֽ���������������Լ�ֲ���ϵ������İ���ø��.����ø���л�̼���ֽ����ת����Ҫ����(Weintraub et al., 2007; Xu et al., 2017), ���������л�̼���Ļ�������������ø�Ŀֽ�����(Sinsabaugh et al., 2014; Mayer et al., 2020).�����л��ʿ������ܿ��ڰ���ø����, ����ˮ��ø��Ҫ�������̼���ֽ�ת��(�����ࡢ���ۡ���ά�ص�), ������-��������ø(��G)����ά��ˮ��ø(CBH)����-����������������ø(NAG)��; ����ø��Ҫ��������л�̼�����(��ľ����)�����ֽ��������ֳ�����̵�, �����������ø(PHO)����������ø(PEO)��(Luo et al., 2019), ���, ø���ԵĸߵͿɱ��������������������Լ������ֹ���ˮƽ֮��Ĺ�ϵ.���о���ʾ, ����ø�������������ֺ������������(Burns et al., 2013; Das & Mondal, 2019)�����������(Li et al., 2015; Feng et al., 2019).ͬʱ, �����������Ч��Ҳ��Ӱ���л�̼�͵��ķֽ�ø����, ʹ������P�����л��ʱ��ֳ���ͬ�ķֽ�����(Kang et al., 2009; Zhou et al., 2020).����, ����ø������������ʪ�ȼ�pH�������й�(��ΰΰ��, 2019; Zhou et al., 2020), ˵������ø����������ͷ��������صĸ������й�.��Ȼ��������ø�����������л�̼����ֺ�����Ĺ�ϵ�ѱ��㷺��ע, �����ڴ����Ǹ�ɽ��Ȼ�����������л�̼����������������ø����������ʶ�������. ...
Soil organic carbon storage capacity positively related to forest succession on the Loess Plateau, China 1 2013
Distribution of soil organic carbon in the light and heavy fractions for six shelterbelt species and their adjacent agricultural fields in Saskatchewan 2 2017
Carbon control on terrestrial ecosystem function across contrasting site productivities: the carbon connection revisited 1 2019
... ɭ�������л�̼�����������Ƿ�ӳ����������ɭ�ֽ���״������Ҫָ��, ֱ�ӻ��ӵ�Ӱ�쵽ɭ������������������.�о���ʾ, ɭ������̼����ԼΪȫ������̼������73% (Post & Kwon, 2000), �ڵ���ȫ��̼ƽ�ⷽ��������Ҫ����.��������Ϊ½��ֲ����̬ϵͳ����Ҫ���(Kauppi et al., 2006), ��ά����������Ժ�����̼��ѭ���а�������Ҫ��ɫ(Yang et al., 2011).ɭ�ָ�������ı���ֲ��Ⱥ��ṹ��������ɼ����ܶȶ�Ӱ�쵽��������, ���������л�̼����������ֺ�����������Է����仯(Chandra et al., 2016; Feng et al., 2019).���о���Ϊ, ɭ��ֲ�������������ڽ��������л�̼�ֽ����ʳ�������ֲ�������������л�̼�Ĺ̳�����, ʹ���京����Խϵ�(Ohtsuka et al., 2010; Milcu et al., 2011), Ҳ���о�����, ����̼����������ɭ������������ֳ�����(Deng et al., 2013; ��ѩ��, 2016; Liu et al., 2020)���仯������(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)���Ƚ�����(Tang et al., 2009)�ı仯����, ˵��ɭ�����������̼��������Ӱ���Ϊ����.�����л��ʽ����ܵ����������Ӱ��(Sinsabaugh, 2010; Liu et al., 2020), ������������Ҳ���ܵ�����̼����Ч�Ե���Լ(Kooch et al., 2018; Dove et al., 2019).ɭ������ı�����������, ����������ɻ���֮�����仯, �����ı��������������(Zhang et al., 2017), �������Ҫ��������Դ�ڶ�ֲ�����ֽ���������������Լ�ֲ���ϵ������İ���ø��.����ø���л�̼���ֽ����ת����Ҫ����(Weintraub et al., 2007; Xu et al., 2017), ���������л�̼���Ļ�������������ø�Ŀֽ�����(Sinsabaugh et al., 2014; Mayer et al., 2020).�����л��ʿ������ܿ��ڰ���ø����, ����ˮ��ø��Ҫ�������̼���ֽ�ת��(�����ࡢ���ۡ���ά�ص�), ������-��������ø(��G)����ά��ˮ��ø(CBH)����-����������������ø(NAG)��; ����ø��Ҫ��������л�̼�����(��ľ����)�����ֽ��������ֳ�����̵�, �����������ø(PHO)����������ø(PEO)��(Luo et al., 2019), ���, ø���ԵĸߵͿɱ��������������������Լ������ֹ���ˮƽ֮��Ĺ�ϵ.���о���ʾ, ����ø�������������ֺ������������(Burns et al., 2013; Das & Mondal, 2019)�����������(Li et al., 2015; Feng et al., 2019).ͬʱ, �����������Ч��Ҳ��Ӱ���л�̼�͵��ķֽ�ø����, ʹ������P�����л��ʱ��ֳ���ͬ�ķֽ�����(Kang et al., 2009; Zhou et al., 2020).����, ����ø������������ʪ�ȼ�pH�������й�(��ΰΰ��, 2019; Zhou et al., 2020), ˵������ø����������ͷ��������صĸ������й�.��Ȼ��������ø�����������л�̼����ֺ�����Ĺ�ϵ�ѱ��㷺��ע, �����ڴ����Ǹ�ɽ��Ȼ�����������л�̼����������������ø����������ʶ�������. ...
Soil enzyme activities increase following restoration of degraded subtropical forests 1 2019
... ɭ�������л�̼�����������Ƿ�ӳ����������ɭ�ֽ���״������Ҫָ��, ֱ�ӻ��ӵ�Ӱ�쵽ɭ������������������.�о���ʾ, ɭ������̼����ԼΪȫ������̼������73% (Post & Kwon, 2000), �ڵ���ȫ��̼ƽ�ⷽ��������Ҫ����.��������Ϊ½��ֲ����̬ϵͳ����Ҫ���(Kauppi et al., 2006), ��ά����������Ժ�����̼��ѭ���а�������Ҫ��ɫ(Yang et al., 2011).ɭ�ָ�������ı���ֲ��Ⱥ��ṹ��������ɼ����ܶȶ�Ӱ�쵽��������, ���������л�̼����������ֺ�����������Է����仯(Chandra et al., 2016; Feng et al., 2019).���о���Ϊ, ɭ��ֲ�������������ڽ��������л�̼�ֽ����ʳ�������ֲ�������������л�̼�Ĺ̳�����, ʹ���京����Խϵ�(Ohtsuka et al., 2010; Milcu et al., 2011), Ҳ���о�����, ����̼����������ɭ������������ֳ�����(Deng et al., 2013; ��ѩ��, 2016; Liu et al., 2020)���仯������(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)���Ƚ�����(Tang et al., 2009)�ı仯����, ˵��ɭ�����������̼��������Ӱ���Ϊ����.�����л��ʽ����ܵ����������Ӱ��(Sinsabaugh, 2010; Liu et al., 2020), ������������Ҳ���ܵ�����̼����Ч�Ե���Լ(Kooch et al., 2018; Dove et al., 2019).ɭ������ı�����������, ����������ɻ���֮�����仯, �����ı��������������(Zhang et al., 2017), �������Ҫ��������Դ�ڶ�ֲ�����ֽ���������������Լ�ֲ���ϵ������İ���ø��.����ø���л�̼���ֽ����ת����Ҫ����(Weintraub et al., 2007; Xu et al., 2017), ���������л�̼���Ļ�������������ø�Ŀֽ�����(Sinsabaugh et al., 2014; Mayer et al., 2020).�����л��ʿ������ܿ��ڰ���ø����, ����ˮ��ø��Ҫ�������̼���ֽ�ת��(�����ࡢ���ۡ���ά�ص�), ������-��������ø(��G)����ά��ˮ��ø(CBH)����-����������������ø(NAG)��; ����ø��Ҫ��������л�̼�����(��ľ����)�����ֽ��������ֳ�����̵�, �����������ø(PHO)����������ø(PEO)��(Luo et al., 2019), ���, ø���ԵĸߵͿɱ��������������������Լ������ֹ���ˮƽ֮��Ĺ�ϵ.���о���ʾ, ����ø�������������ֺ������������(Burns et al., 2013; Das & Mondal, 2019)�����������(Li et al., 2015; Feng et al., 2019).ͬʱ, �����������Ч��Ҳ��Ӱ���л�̼�͵��ķֽ�ø����, ʹ������P�����л��ʱ��ֳ���ͬ�ķֽ�����(Kang et al., 2009; Zhou et al., 2020).����, ����ø������������ʪ�ȼ�pH�������й�(��ΰΰ��, 2019; Zhou et al., 2020), ˵������ø����������ͷ��������صĸ������й�.��Ȼ��������ø�����������л�̼����ֺ�����Ĺ�ϵ�ѱ��㷺��ע, �����ڴ����Ǹ�ɽ��Ȼ�����������л�̼����������������ø����������ʶ�������. ...
Environmental control of lignin peroxidase, manganese peroxidase, and laccase activities in forest floor layers in humid Asia 2013
... ���������л�̼���������������л��ʵ���Ҫ���, ��Ҫ��Դ���½���֦��Ҷ����ֳ�ʡ���ϵ�������Լ��������(Kalbitz et al., 2000; ���µ�, 2018).ɭ�ֲ�ͬ��Ȼ���»ָ���, ���ַ������������Իָ���������ɵIJ���, ���µر�������������������������ʪ�ȡ�������ɵȷ�����ͬ�̶ȱ仯, �Ӷ�Ӱ�쵽���������л�̼�������ı仯(��ѩ��, 2016; Hagedorn et al., 2019).���о���, DOC��LFOC��DON�����������������������½�����(p < 0.05), ����70-NSF��80-NSF�в��첻����(��3), �����浽ẽ���ɼԭʼ�ֽ�, ����������̼��������������(p < 0.05), ����������˹�����������б���(0-20 cm)��������̼���������о��������(������, 2009), ����ͬ���ȴ�(���ܵ�, 2019)�����ȴ�(��Ծ�µ�, 2013)ɭ��������о����, ˵����ͬ��ʪ������ֲ���ַֽṹ�����������л�̼����ֺ�����Ӱ����ֳ���ͬ�ı仯����(Li et al., 2014; Taylor et al., 2019).����ԭ��, һ��������ɭ�ָ�������, ����̼������������(Mylliemngap et al., 2016), ����Ҷ�ֵĿ����ʸ�����Ҷ��(C?t�� et al., 2000); ��һ�������ڱ��������ܶ�����������, �������ž۶ȡ��ṹϵ�����ܿ�϶��ë�ܿ�϶������������������ֽ�������(��ѧ�µ�, 2003), �����л�̼����ȱ�������ž���������Ա���, �������������л�̼��DON��������.����, ẽ���ɼԭʼ�ֵ���������̼����ֺ���������Ȼ������(��3), ��������ַ���Ҫ���ֹ���(��Ҷ+��Ҷ����)��һ������, Դ�ڸ������������ֵ���Ҷ�ں���һ�����ڷֽ�(����÷��, 2015), ���ͷŵ����ֱ�������ֽ�����, �������������л�̼��������.��Ҫ��������Ӱ�����ض����������л�̼������������, �����һ�������о�.��ط�������, ��ͬ��Ȼ����������ֵر�������SOC��TN����������л�̼����ֺ�����DON�������ʼ���������ع�ϵ(p < 0.01), ˵�����������л�̼�͵�����������SOC��TN����֮���ϵ��Ϊ����, ���������о�(�����, 2016; ������, 2017) һ��, ˵����Ȼ�����������������ǵ�����̼�ֵ̳���Ҫ����(Li et al., 2012; Smal et al., 2019). ...
�����ȴ�ֲ���ָ��������л�̼������̼�ܶȵ�Ӱ�� 1 2018
... ɭ�������л�̼�����������Ƿ�ӳ����������ɭ�ֽ���״������Ҫָ��, ֱ�ӻ��ӵ�Ӱ�쵽ɭ������������������.�о���ʾ, ɭ������̼����ԼΪȫ������̼������73% (Post & Kwon, 2000), �ڵ���ȫ��̼ƽ�ⷽ��������Ҫ����.��������Ϊ½��ֲ����̬ϵͳ����Ҫ���(Kauppi et al., 2006), ��ά����������Ժ�����̼��ѭ���а�������Ҫ��ɫ(Yang et al., 2011).ɭ�ָ�������ı���ֲ��Ⱥ��ṹ��������ɼ����ܶȶ�Ӱ�쵽��������, ���������л�̼����������ֺ�����������Է����仯(Chandra et al., 2016; Feng et al., 2019).���о���Ϊ, ɭ��ֲ�������������ڽ��������л�̼�ֽ����ʳ�������ֲ�������������л�̼�Ĺ̳�����, ʹ���京����Խϵ�(Ohtsuka et al., 2010; Milcu et al., 2011), Ҳ���о�����, ����̼����������ɭ������������ֳ�����(Deng et al., 2013; ��ѩ��, 2016; Liu et al., 2020)���仯������(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)���Ƚ�����(Tang et al., 2009)�ı仯����, ˵��ɭ�����������̼��������Ӱ���Ϊ����.�����л��ʽ����ܵ����������Ӱ��(Sinsabaugh, 2010; Liu et al., 2020), ������������Ҳ���ܵ�����̼����Ч�Ե���Լ(Kooch et al., 2018; Dove et al., 2019).ɭ������ı�����������, ����������ɻ���֮�����仯, �����ı��������������(Zhang et al., 2017), �������Ҫ��������Դ�ڶ�ֲ�����ֽ���������������Լ�ֲ���ϵ������İ���ø��.����ø���л�̼���ֽ����ת����Ҫ����(Weintraub et al., 2007; Xu et al., 2017), ���������л�̼���Ļ�������������ø�Ŀֽ�����(Sinsabaugh et al., 2014; Mayer et al., 2020).�����л��ʿ������ܿ��ڰ���ø����, ����ˮ��ø��Ҫ�������̼���ֽ�ת��(�����ࡢ���ۡ���ά�ص�), ������-��������ø(��G)����ά��ˮ��ø(CBH)����-����������������ø(NAG)��; ����ø��Ҫ��������л�̼�����(��ľ����)�����ֽ��������ֳ�����̵�, �����������ø(PHO)����������ø(PEO)��(Luo et al., 2019), ���, ø���ԵĸߵͿɱ��������������������Լ������ֹ���ˮƽ֮��Ĺ�ϵ.���о���ʾ, ����ø�������������ֺ������������(Burns et al., 2013; Das & Mondal, 2019)�����������(Li et al., 2015; Feng et al., 2019).ͬʱ, �����������Ч��Ҳ��Ӱ���л�̼�͵��ķֽ�ø����, ʹ������P�����л��ʱ��ֳ���ͬ�ķֽ�����(Kang et al., 2009; Zhou et al., 2020).����, ����ø������������ʪ�ȼ�pH�������й�(��ΰΰ��, 2019; Zhou et al., 2020), ˵������ø����������ͷ��������صĸ������й�.��Ȼ��������ø�����������л�̼����ֺ�����Ĺ�ϵ�ѱ��㷺��ע, �����ڴ����Ǹ�ɽ��Ȼ�����������л�̼����������������ø����������ʶ�������. ...
�����ȴ�ֲ���ָ��������л�̼������̼�ܶȵ�Ӱ�� 1 2018
... ɭ�������л�̼�����������Ƿ�ӳ����������ɭ�ֽ���״������Ҫָ��, ֱ�ӻ��ӵ�Ӱ�쵽ɭ������������������.�о���ʾ, ɭ������̼����ԼΪȫ������̼������73% (Post & Kwon, 2000), �ڵ���ȫ��̼ƽ�ⷽ��������Ҫ����.��������Ϊ½��ֲ����̬ϵͳ����Ҫ���(Kauppi et al., 2006), ��ά����������Ժ�����̼��ѭ���а�������Ҫ��ɫ(Yang et al., 2011).ɭ�ָ�������ı���ֲ��Ⱥ��ṹ��������ɼ����ܶȶ�Ӱ�쵽��������, ���������л�̼����������ֺ�����������Է����仯(Chandra et al., 2016; Feng et al., 2019).���о���Ϊ, ɭ��ֲ�������������ڽ��������л�̼�ֽ����ʳ�������ֲ�������������л�̼�Ĺ̳�����, ʹ���京����Խϵ�(Ohtsuka et al., 2010; Milcu et al., 2011), Ҳ���о�����, ����̼����������ɭ������������ֳ�����(Deng et al., 2013; ��ѩ��, 2016; Liu et al., 2020)���仯������(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)���Ƚ�����(Tang et al., 2009)�ı仯����, ˵��ɭ�����������̼��������Ӱ���Ϊ����.�����л��ʽ����ܵ����������Ӱ��(Sinsabaugh, 2010; Liu et al., 2020), ������������Ҳ���ܵ�����̼����Ч�Ե���Լ(Kooch et al., 2018; Dove et al., 2019).ɭ������ı�����������, ����������ɻ���֮�����仯, �����ı��������������(Zhang et al., 2017), �������Ҫ��������Դ�ڶ�ֲ�����ֽ���������������Լ�ֲ���ϵ������İ���ø��.����ø���л�̼���ֽ����ת����Ҫ����(Weintraub et al., 2007; Xu et al., 2017), ���������л�̼���Ļ�������������ø�Ŀֽ�����(Sinsabaugh et al., 2014; Mayer et al., 2020).�����л��ʿ������ܿ��ڰ���ø����, ����ˮ��ø��Ҫ�������̼���ֽ�ת��(�����ࡢ���ۡ���ά�ص�), ������-��������ø(��G)����ά��ˮ��ø(CBH)����-����������������ø(NAG)��; ����ø��Ҫ��������л�̼�����(��ľ����)�����ֽ��������ֳ�����̵�, �����������ø(PHO)����������ø(PEO)��(Luo et al., 2019), ���, ø���ԵĸߵͿɱ��������������������Լ������ֹ���ˮƽ֮��Ĺ�ϵ.���о���ʾ, ����ø�������������ֺ������������(Burns et al., 2013; Das & Mondal, 2019)�����������(Li et al., 2015; Feng et al., 2019).ͬʱ, �����������Ч��Ҳ��Ӱ���л�̼�͵��ķֽ�ø����, ʹ������P�����л��ʱ��ֳ���ͬ�ķֽ�����(Kang et al., 2009; Zhou et al., 2020).����, ����ø������������ʪ�ȼ�pH�������й�(��ΰΰ��, 2019; Zhou et al., 2020), ˵������ø����������ͷ��������صĸ������й�.��Ȼ��������ø�����������л�̼����ֺ�����Ĺ�ϵ�ѱ��㷺��ע, �����ڴ����Ǹ�ɽ��Ȼ�����������л�̼����������������ø����������ʶ�������. ...
�����ȴ�4��ɭ���������������л�̼�ļ��ڶ�̬���� 1 2016
... ɭ�ָ�������Ļָ�������, �ַֽṹ��������ɡ����������Լ������������˲�ͬ�̶ȵı仯, ��������������̼��Դ��ͬ�����������л��ʷֽ����ʡ�������������Լ����л������仯(Wardle et al., 2004; Lucas-Borja & Delgado- Baquerizo, 2019), ����Ӱ�쵽����ø����(Adamczyk et al., 2015; Xu et al., 2017).����, ��ƽ�����¡���ˮ������������Ժ����ֶ�����Ҳ��Ӱ������ø���Ե���Ҫ����(�����, 2014; Zhou et al., 2020).���о���, ��Ȼ�����ָ�����������(<60 a)��ͬ�εı�������ˮ��ø(��G��CBH��NAG)������������ẽ���ɼԭʼ��(��4), ������Ȼ����������̳�������������(80-NSF > 70-NSF > 60-NSF), ��ͬ�ڴ����Ǹ�ɽƽ���ذ�����ˮ��ø���Ը�����ɼ��(�ܺ��ٵ�, 2004)���´�����(Betula platyphylla)����������̱���Ϊ�����Ӻ͵ı仯����(���Ե�, 2013), һ����˵��ẽ���ɼԭʼ�ֵ���������ѭ��������Խϸ�, ���������ֹ�������; ��һ����˵����������Ȼ�������������, ���ϵ��������ӵ�������̼�����������ֺ������½�(Liu et al., 2017; Lyu et al., 2019), ʹ������������Խ���(�����, 2018), ������������ˮ��ø�����½�.����, 80-NSF��70-NSF��60-NSF����������ø���Ա仯������, ��PHO�Ļ�����������CK, ��PEO�Ļ�����CK���첻����(��4), ��ԭ�������ẽ���ɼԭʼ���к��н�Ϊ�ḻ��ľ���ء����ʺ����ʵ��ѽ��⻯����, �ٽ��˽���ľ���ص�����Ⱥ������Ի�������������, ��������ָ�������εĽ���, ��ϵ��������ֲ��������������仯, ����������pH����, ��ʹPHO��������(Fujii et al., 2013). ...
�����ȴ�4��ɭ���������������л�̼�ļ��ڶ�̬���� 1 2016
... ɭ�ָ�������Ļָ�������, �ַֽṹ��������ɡ����������Լ������������˲�ͬ�̶ȵı仯, ��������������̼��Դ��ͬ�����������л��ʷֽ����ʡ�������������Լ����л������仯(Wardle et al., 2004; Lucas-Borja & Delgado- Baquerizo, 2019), ����Ӱ�쵽����ø����(Adamczyk et al., 2015; Xu et al., 2017).����, ��ƽ�����¡���ˮ������������Ժ����ֶ�����Ҳ��Ӱ������ø���Ե���Ҫ����(�����, 2014; Zhou et al., 2020).���о���, ��Ȼ�����ָ�����������(<60 a)��ͬ�εı�������ˮ��ø(��G��CBH��NAG)������������ẽ���ɼԭʼ��(��4), ������Ȼ����������̳�������������(80-NSF > 70-NSF > 60-NSF), ��ͬ�ڴ����Ǹ�ɽƽ���ذ�����ˮ��ø���Ը�����ɼ��(�ܺ��ٵ�, 2004)���´�����(Betula platyphylla)����������̱���Ϊ�����Ӻ͵ı仯����(���Ե�, 2013), һ����˵��ẽ���ɼԭʼ�ֵ���������ѭ��������Խϸ�, ���������ֹ�������; ��һ����˵����������Ȼ�������������, ���ϵ��������ӵ�������̼�����������ֺ������½�(Liu et al., 2017; Lyu et al., 2019), ʹ������������Խ���(�����, 2018), ������������ˮ��ø�����½�.����, 80-NSF��70-NSF��60-NSF����������ø���Ա仯������, ��PHO�Ļ�����������CK, ��PEO�Ļ�����CK���첻����(��4), ��ԭ�������ẽ���ɼԭʼ���к��н�Ϊ�ḻ��ľ���ء����ʺ����ʵ��ѽ��⻯����, �ٽ��˽���ľ���ص�����Ⱥ������Ի�������������, ��������ָ�������εĽ���, ��ϵ��������ֲ��������������仯, ����������pH����, ��ʹPHO��������(Fujii et al., 2013). ...
Above- and belowground linkages shape responses of mountain vegetation to climate change 2019
... ���������л�̼���������������л��ʵ���Ҫ���, ��Ҫ��Դ���½���֦��Ҷ����ֳ�ʡ���ϵ�������Լ��������(Kalbitz et al., 2000; ���µ�, 2018).ɭ�ֲ�ͬ��Ȼ���»ָ���, ���ַ������������Իָ���������ɵIJ���, ���µر�������������������������ʪ�ȡ�������ɵȷ�����ͬ�̶ȱ仯, �Ӷ�Ӱ�쵽���������л�̼�������ı仯(��ѩ��, 2016; Hagedorn et al., 2019).���о���, DOC��LFOC��DON�����������������������½�����(p < 0.05), ����70-NSF��80-NSF�в��첻����(��3), �����浽ẽ���ɼԭʼ�ֽ�, ����������̼��������������(p < 0.05), ����������˹�����������б���(0-20 cm)��������̼���������о��������(������, 2009), ����ͬ���ȴ�(���ܵ�, 2019)�����ȴ�(��Ծ�µ�, 2013)ɭ��������о����, ˵����ͬ��ʪ������ֲ���ַֽṹ�����������л�̼����ֺ�����Ӱ����ֳ���ͬ�ı仯����(Li et al., 2014; Taylor et al., 2019).����ԭ��, һ��������ɭ�ָ�������, ����̼������������(Mylliemngap et al., 2016), ����Ҷ�ֵĿ����ʸ�����Ҷ��(C?t�� et al., 2000); ��һ�������ڱ��������ܶ�����������, �������ž۶ȡ��ṹϵ�����ܿ�϶��ë�ܿ�϶������������������ֽ�������(��ѧ�µ�, 2003), �����л�̼����ȱ�������ž���������Ա���, �������������л�̼��DON��������.����, ẽ���ɼԭʼ�ֵ���������̼����ֺ���������Ȼ������(��3), ��������ַ���Ҫ���ֹ���(��Ҷ+��Ҷ����)��һ������, Դ�ڸ������������ֵ���Ҷ�ں���һ�����ڷֽ�(����÷��, 2015), ���ͷŵ����ֱ�������ֽ�����, �������������л�̼��������.��Ҫ��������Ӱ�����ض����������л�̼������������, �����һ�������о�.��ط�������, ��ͬ��Ȼ����������ֵر�������SOC��TN����������л�̼����ֺ�����DON�������ʼ���������ع�ϵ(p < 0.01), ˵�����������л�̼�͵�����������SOC��TN����֮���ϵ��Ϊ����, ���������о�(�����, 2016; ������, 2017) һ��, ˵����Ȼ�����������������ǵ�����̼�ֵ̳���Ҫ����(Li et al., 2012; Smal et al., 2019). ...
�����Ǹ�ɽ��ɼҶ������������ʧ���̼�������������� 2015
�����Ǹ�ɽ��ɼҶ������������ʧ���̼�������������� 2015
�����Ǹ�ɽ��ɼ�˹��ָֻ������б�������̼��̬�仯 1 2009
... ɭ�ָ�������Ļָ�������, �ַֽṹ��������ɡ����������Լ������������˲�ͬ�̶ȵı仯, ��������������̼��Դ��ͬ�����������л��ʷֽ����ʡ�������������Լ����л������仯(Wardle et al., 2004; Lucas-Borja & Delgado- Baquerizo, 2019), ����Ӱ�쵽����ø����(Adamczyk et al., 2015; Xu et al., 2017).����, ��ƽ�����¡���ˮ������������Ժ����ֶ�����Ҳ��Ӱ������ø���Ե���Ҫ����(�����, 2014; Zhou et al., 2020).���о���, ��Ȼ�����ָ�����������(<60 a)��ͬ�εı�������ˮ��ø(��G��CBH��NAG)������������ẽ���ɼԭʼ��(��4), ������Ȼ����������̳�������������(80-NSF > 70-NSF > 60-NSF), ��ͬ�ڴ����Ǹ�ɽƽ���ذ�����ˮ��ø���Ը�����ɼ��(�ܺ��ٵ�, 2004)���´�����(Betula platyphylla)����������̱���Ϊ�����Ӻ͵ı仯����(���Ե�, 2013), һ����˵��ẽ���ɼԭʼ�ֵ���������ѭ��������Խϸ�, ���������ֹ�������; ��һ����˵����������Ȼ�������������, ���ϵ��������ӵ�������̼�����������ֺ������½�(Liu et al., 2017; Lyu et al., 2019), ʹ������������Խ���(�����, 2018), ������������ˮ��ø�����½�.����, 80-NSF��70-NSF��60-NSF����������ø���Ա仯������, ��PHO�Ļ�����������CK, ��PEO�Ļ�����CK���첻����(��4), ��ԭ�������ẽ���ɼԭʼ���к��н�Ϊ�ḻ��ľ���ء����ʺ����ʵ��ѽ��⻯����, �ٽ��˽���ľ���ص�����Ⱥ������Ի�������������, ��������ָ�������εĽ���, ��ϵ��������ֲ��������������仯, ����������pH����, ��ʹPHO��������(Fujii et al., 2013). ...
�����Ǹ�ɽ��ɼ�˹��ָֻ������б�������̼��̬�仯 1 2009
... ɭ�ָ�������Ļָ�������, �ַֽṹ��������ɡ����������Լ������������˲�ͬ�̶ȵı仯, ��������������̼��Դ��ͬ�����������л��ʷֽ����ʡ�������������Լ����л������仯(Wardle et al., 2004; Lucas-Borja & Delgado- Baquerizo, 2019), ����Ӱ�쵽����ø����(Adamczyk et al., 2015; Xu et al., 2017).����, ��ƽ�����¡���ˮ������������Ժ����ֶ�����Ҳ��Ӱ������ø���Ե���Ҫ����(�����, 2014; Zhou et al., 2020).���о���, ��Ȼ�����ָ�����������(<60 a)��ͬ�εı�������ˮ��ø(��G��CBH��NAG)������������ẽ���ɼԭʼ��(��4), ������Ȼ����������̳�������������(80-NSF > 70-NSF > 60-NSF), ��ͬ�ڴ����Ǹ�ɽƽ���ذ�����ˮ��ø���Ը�����ɼ��(�ܺ��ٵ�, 2004)���´�����(Betula platyphylla)����������̱���Ϊ�����Ӻ͵ı仯����(���Ե�, 2013), һ����˵��ẽ���ɼԭʼ�ֵ���������ѭ��������Խϸ�, ���������ֹ�������; ��һ����˵����������Ȼ�������������, ���ϵ��������ӵ�������̼�����������ֺ������½�(Liu et al., 2017; Lyu et al., 2019), ʹ������������Խ���(�����, 2018), ������������ˮ��ø�����½�.����, 80-NSF��70-NSF��60-NSF����������ø���Ա仯������, ��PHO�Ļ�����������CK, ��PEO�Ļ�����CK���첻����(��4), ��ԭ�������ẽ���ɼԭʼ���к��н�Ϊ�ḻ��ľ���ء����ʺ����ʵ��ѽ��⻯����, �ٽ��˽���ľ���ص�����Ⱥ������Ի�������������, ��������ָ�������εĽ���, ��ϵ��������ֲ��������������仯, ����������pH����, ��ʹPHO��������(Fujii et al., 2013). ...
Controls on the dynamics of dissolved organic matter in soils: a review 2000
Variations of soil enzyme activities in a temperate forest soil 1 2009
... ���������л�̼���������������л��ʵ���Ҫ���, ��Ҫ��Դ���½���֦��Ҷ����ֳ�ʡ���ϵ�������Լ��������(Kalbitz et al., 2000; ���µ�, 2018).ɭ�ֲ�ͬ��Ȼ���»ָ���, ���ַ������������Իָ���������ɵIJ���, ���µر�������������������������ʪ�ȡ�������ɵȷ�����ͬ�̶ȱ仯, �Ӷ�Ӱ�쵽���������л�̼�������ı仯(��ѩ��, 2016; Hagedorn et al., 2019).���о���, DOC��LFOC��DON�����������������������½�����(p < 0.05), ����70-NSF��80-NSF�в��첻����(��3), �����浽ẽ���ɼԭʼ�ֽ�, ����������̼��������������(p < 0.05), ����������˹�����������б���(0-20 cm)��������̼���������о��������(������, 2009), ����ͬ���ȴ�(���ܵ�, 2019)�����ȴ�(��Ծ�µ�, 2013)ɭ��������о����, ˵����ͬ��ʪ������ֲ���ַֽṹ�����������л�̼����ֺ�����Ӱ����ֳ���ͬ�ı仯����(Li et al., 2014; Taylor et al., 2019).����ԭ��, һ��������ɭ�ָ�������, ����̼������������(Mylliemngap et al., 2016), ����Ҷ�ֵĿ����ʸ�����Ҷ��(C?t�� et al., 2000); ��һ�������ڱ��������ܶ�����������, �������ž۶ȡ��ṹϵ�����ܿ�϶��ë�ܿ�϶������������������ֽ�������(��ѧ�µ�, 2003), �����л�̼����ȱ�������ž���������Ա���, �������������л�̼��DON��������.����, ẽ���ɼԭʼ�ֵ���������̼����ֺ���������Ȼ������(��3), ��������ַ���Ҫ���ֹ���(��Ҷ+��Ҷ����)��һ������, Դ�ڸ������������ֵ���Ҷ�ں���һ�����ڷֽ�(����÷��, 2015), ���ͷŵ����ֱ�������ֽ�����, �������������л�̼��������.��Ҫ��������Ӱ�����ض����������л�̼������������, �����һ�������о�.��ط�������, ��ͬ��Ȼ����������ֵر�������SOC��TN����������л�̼����ֺ�����DON�������ʼ���������ع�ϵ(p < 0.01), ˵�����������л�̼�͵�����������SOC��TN����֮���ϵ��Ϊ����, ���������о�(�����, 2016; ������, 2017) һ��, ˵����Ȼ�����������������ǵ�����̼�ֵ̳���Ҫ����(Li et al., 2012; Smal et al., 2019). ...
Returning forests analyzed with the forest identity 1 2006
... ���������л�̼���������������л��ʵ���Ҫ���, ��Ҫ��Դ���½���֦��Ҷ����ֳ�ʡ���ϵ�������Լ��������(Kalbitz et al., 2000; ���µ�, 2018).ɭ�ֲ�ͬ��Ȼ���»ָ���, ���ַ������������Իָ���������ɵIJ���, ���µر�������������������������ʪ�ȡ�������ɵȷ�����ͬ�̶ȱ仯, �Ӷ�Ӱ�쵽���������л�̼�������ı仯(��ѩ��, 2016; Hagedorn et al., 2019).���о���, DOC��LFOC��DON�����������������������½�����(p < 0.05), ����70-NSF��80-NSF�в��첻����(��3), �����浽ẽ���ɼԭʼ�ֽ�, ����������̼��������������(p < 0.05), ����������˹�����������б���(0-20 cm)��������̼���������о��������(������, 2009), ����ͬ���ȴ�(���ܵ�, 2019)�����ȴ�(��Ծ�µ�, 2013)ɭ��������о����, ˵����ͬ��ʪ������ֲ���ַֽṹ�����������л�̼����ֺ�����Ӱ����ֳ���ͬ�ı仯����(Li et al., 2014; Taylor et al., 2019).����ԭ��, һ��������ɭ�ָ�������, ����̼������������(Mylliemngap et al., 2016), ����Ҷ�ֵĿ����ʸ�����Ҷ��(C?t�� et al., 2000); ��һ�������ڱ��������ܶ�����������, �������ž۶ȡ��ṹϵ�����ܿ�϶��ë�ܿ�϶������������������ֽ�������(��ѧ�µ�, 2003), �����л�̼����ȱ�������ž���������Ա���, �������������л�̼��DON��������.����, ẽ���ɼԭʼ�ֵ���������̼����ֺ���������Ȼ������(��3), ��������ַ���Ҫ���ֹ���(��Ҷ+��Ҷ����)��һ������, Դ�ڸ������������ֵ���Ҷ�ں���һ�����ڷֽ�(����÷��, 2015), ���ͷŵ����ֱ�������ֽ�����, �������������л�̼��������.��Ҫ��������Ӱ�����ض����������л�̼������������, �����һ�������о�.��ط�������, ��ͬ��Ȼ����������ֵر�������SOC��TN����������л�̼����ֺ�����DON�������ʼ���������ع�ϵ(p < 0.01), ˵�����������л�̼�͵�����������SOC��TN����֮���ϵ��Ϊ����, ���������о�(�����, 2016; ������, 2017) һ��, ˵����Ȼ�����������������ǵ�����̼�ֵ̳���Ҫ����(Li et al., 2012; Smal et al., 2019). ...
Increasing tree diversity enhances microbial and enzyme activities in temperate Iranian forests 2018
Global patterns of the dynamics of soil carbon and nitrogen stocks following afforestation: a meta-analysis 1 2012
... ɭ�ָ�������Ļָ�������, �ַֽṹ��������ɡ����������Լ������������˲�ͬ�̶ȵı仯, ��������������̼��Դ��ͬ�����������л��ʷֽ����ʡ�������������Լ����л������仯(Wardle et al., 2004; Lucas-Borja & Delgado- Baquerizo, 2019), ����Ӱ�쵽����ø����(Adamczyk et al., 2015; Xu et al., 2017).����, ��ƽ�����¡���ˮ������������Ժ����ֶ�����Ҳ��Ӱ������ø���Ե���Ҫ����(�����, 2014; Zhou et al., 2020).���о���, ��Ȼ�����ָ�����������(<60 a)��ͬ�εı�������ˮ��ø(��G��CBH��NAG)������������ẽ���ɼԭʼ��(��4), ������Ȼ����������̳�������������(80-NSF > 70-NSF > 60-NSF), ��ͬ�ڴ����Ǹ�ɽƽ���ذ�����ˮ��ø���Ը�����ɼ��(�ܺ��ٵ�, 2004)���´�����(Betula platyphylla)����������̱���Ϊ�����Ӻ͵ı仯����(���Ե�, 2013), һ����˵��ẽ���ɼԭʼ�ֵ���������ѭ��������Խϸ�, ���������ֹ�������; ��һ����˵����������Ȼ�������������, ���ϵ��������ӵ�������̼�����������ֺ������½�(Liu et al., 2017; Lyu et al., 2019), ʹ������������Խ���(�����, 2018), ������������ˮ��ø�����½�.����, 80-NSF��70-NSF��60-NSF����������ø���Ա仯������, ��PHO�Ļ�����������CK, ��PEO�Ļ�����CK���첻����(��4), ��ԭ�������ẽ���ɼԭʼ���к��н�Ϊ�ḻ��ľ���ء����ʺ����ʵ��ѽ��⻯����, �ٽ��˽���ľ���ص�����Ⱥ������Ի�������������, ��������ָ�������εĽ���, ��ϵ��������ֲ��������������仯, ����������pH����, ��ʹPHO��������(Fujii et al., 2013). ...
Effects of regenerating vegetation on soil enzyme activity and microbial structure in reclaimed soils on a surface coal mine site 2015
Soil organic carbon fractions and microbial community and functions under changes in vegetation: a case of vegetation succession in karst forest 1 2014
... ���������л�̼���������������л��ʵ���Ҫ���, ��Ҫ��Դ���½���֦��Ҷ����ֳ�ʡ���ϵ�������Լ��������(Kalbitz et al., 2000; ���µ�, 2018).ɭ�ֲ�ͬ��Ȼ���»ָ���, ���ַ������������Իָ���������ɵIJ���, ���µر�������������������������ʪ�ȡ�������ɵȷ�����ͬ�̶ȱ仯, �Ӷ�Ӱ�쵽���������л�̼�������ı仯(��ѩ��, 2016; Hagedorn et al., 2019).���о���, DOC��LFOC��DON�����������������������½�����(p < 0.05), ����70-NSF��80-NSF�в��첻����(��3), �����浽ẽ���ɼԭʼ�ֽ�, ����������̼��������������(p < 0.05), ����������˹�����������б���(0-20 cm)��������̼���������о��������(������, 2009), ����ͬ���ȴ�(���ܵ�, 2019)�����ȴ�(��Ծ�µ�, 2013)ɭ��������о����, ˵����ͬ��ʪ������ֲ���ַֽṹ�����������л�̼����ֺ�����Ӱ����ֳ���ͬ�ı仯����(Li et al., 2014; Taylor et al., 2019).����ԭ��, һ��������ɭ�ָ�������, ����̼������������(Mylliemngap et al., 2016), ����Ҷ�ֵĿ����ʸ�����Ҷ��(C?t�� et al., 2000); ��һ�������ڱ��������ܶ�����������, �������ž۶ȡ��ṹϵ�����ܿ�϶��ë�ܿ�϶������������������ֽ�������(��ѧ�µ�, 2003), �����л�̼����ȱ�������ž���������Ա���, �������������л�̼��DON��������.����, ẽ���ɼԭʼ�ֵ���������̼����ֺ���������Ȼ������(��3), ��������ַ���Ҫ���ֹ���(��Ҷ+��Ҷ����)��һ������, Դ�ڸ������������ֵ���Ҷ�ں���һ�����ڷֽ�(����÷��, 2015), ���ͷŵ����ֱ�������ֽ�����, �������������л�̼��������.��Ҫ��������Ӱ�����ض����������л�̼������������, �����һ�������о�.��ط�������, ��ͬ��Ȼ����������ֵر�������SOC��TN����������л�̼����ֺ�����DON�������ʼ���������ع�ϵ(p < 0.01), ˵�����������л�̼�͵�����������SOC��TN����֮���ϵ��Ϊ����, ���������о�(�����, 2016; ������, 2017) һ��, ˵����Ȼ�����������������ǵ�����̼�ֵ̳���Ҫ����(Li et al., 2012; Smal et al., 2019). ...
������ֲ����Ȼ�ָ�������ֲ������Եı仯 2004
������ֲ����Ȼ�ָ�������ֲ������Եı仯 2004
Increased litter in subtropical forests boosts soil respiration in natural forests but not plantations of 2 2017
... ���������л�̼���������������л��ʵ���Ҫ���, ��Ҫ��Դ���½���֦��Ҷ����ֳ�ʡ���ϵ�������Լ��������(Kalbitz et al., 2000; ���µ�, 2018).ɭ�ֲ�ͬ��Ȼ���»ָ���, ���ַ������������Իָ���������ɵIJ���, ���µر�������������������������ʪ�ȡ�������ɵȷ�����ͬ�̶ȱ仯, �Ӷ�Ӱ�쵽���������л�̼�������ı仯(��ѩ��, 2016; Hagedorn et al., 2019).���о���, DOC��LFOC��DON�����������������������½�����(p < 0.05), ����70-NSF��80-NSF�в��첻����(��3), �����浽ẽ���ɼԭʼ�ֽ�, ����������̼��������������(p < 0.05), ����������˹�����������б���(0-20 cm)��������̼���������о��������(������, 2009), ����ͬ���ȴ�(���ܵ�, 2019)�����ȴ�(��Ծ�µ�, 2013)ɭ��������о����, ˵����ͬ��ʪ������ֲ���ַֽṹ�����������л�̼����ֺ�����Ӱ����ֳ���ͬ�ı仯����(Li et al., 2014; Taylor et al., 2019).����ԭ��, һ��������ɭ�ָ�������, ����̼������������(Mylliemngap et al., 2016), ����Ҷ�ֵĿ����ʸ�����Ҷ��(C?t�� et al., 2000); ��һ�������ڱ��������ܶ�����������, �������ž۶ȡ��ṹϵ�����ܿ�϶��ë�ܿ�϶������������������ֽ�������(��ѧ�µ�, 2003), �����л�̼����ȱ�������ž���������Ա���, �������������л�̼��DON��������.����, ẽ���ɼԭʼ�ֵ���������̼����ֺ���������Ȼ������(��3), ��������ַ���Ҫ���ֹ���(��Ҷ+��Ҷ����)��һ������, Դ�ڸ������������ֵ���Ҷ�ں���һ�����ڷֽ�(����÷��, 2015), ���ͷŵ����ֱ�������ֽ�����, �������������л�̼��������.��Ҫ��������Ӱ�����ض����������л�̼������������, �����һ�������о�.��ط�������, ��ͬ��Ȼ����������ֵر�������SOC��TN����������л�̼����ֺ�����DON�������ʼ���������ع�ϵ(p < 0.01), ˵�����������л�̼�͵�����������SOC��TN����֮���ϵ��Ϊ����, ���������о�(�����, 2016; ������, 2017) һ��, ˵����Ȼ�����������������ǵ�����̼�ֵ̳���Ҫ����(Li et al., 2012; Smal et al., 2019). ...
Long-term forest succession improves plant diversity and soil quality but not significantly increase soil microbial diversity: evidence from the Loess Plateau 1 2020
... ���������л�̼���������������л��ʵ���Ҫ���, ��Ҫ��Դ���½���֦��Ҷ����ֳ�ʡ���ϵ�������Լ��������(Kalbitz et al., 2000; ���µ�, 2018).ɭ�ֲ�ͬ��Ȼ���»ָ���, ���ַ������������Իָ���������ɵIJ���, ���µر�������������������������ʪ�ȡ�������ɵȷ�����ͬ�̶ȱ仯, �Ӷ�Ӱ�쵽���������л�̼�������ı仯(��ѩ��, 2016; Hagedorn et al., 2019).���о���, DOC��LFOC��DON�����������������������½�����(p < 0.05), ����70-NSF��80-NSF�в��첻����(��3), �����浽ẽ���ɼԭʼ�ֽ�, ����������̼��������������(p < 0.05), ����������˹�����������б���(0-20 cm)��������̼���������о��������(������, 2009), ����ͬ���ȴ�(���ܵ�, 2019)�����ȴ�(��Ծ�µ�, 2013)ɭ��������о����, ˵����ͬ��ʪ������ֲ���ַֽṹ�����������л�̼����ֺ�����Ӱ����ֳ���ͬ�ı仯����(Li et al., 2014; Taylor et al., 2019).����ԭ��, һ��������ɭ�ָ�������, ����̼������������(Mylliemngap et al., 2016), ����Ҷ�ֵĿ����ʸ�����Ҷ��(C?t�� et al., 2000); ��һ�������ڱ��������ܶ�����������, �������ž۶ȡ��ṹϵ�����ܿ�϶��ë�ܿ�϶������������������ֽ�������(��ѧ�µ�, 2003), �����л�̼����ȱ�������ž���������Ա���, �������������л�̼��DON��������.����, ẽ���ɼԭʼ�ֵ���������̼����ֺ���������Ȼ������(��3), ��������ַ���Ҫ���ֹ���(��Ҷ+��Ҷ����)��һ������, Դ�ڸ������������ֵ���Ҷ�ں���һ�����ڷֽ�(����÷��, 2015), ���ͷŵ����ֱ�������ֽ�����, �������������л�̼��������.��Ҫ��������Ӱ�����ض����������л�̼������������, �����һ�������о�.��ط�������, ��ͬ��Ȼ����������ֵر�������SOC��TN����������л�̼����ֺ�����DON�������ʼ���������ع�ϵ(p < 0.01), ˵�����������л�̼�͵�����������SOC��TN����֮���ϵ��Ϊ����, ���������о�(�����, 2016; ������, 2017) һ��, ˵����Ȼ�����������������ǵ�����̼�ֵ̳���Ҫ����(Li et al., 2012; Smal et al., 2019). ...
2 2002
... ɭ�������л�̼�����������Ƿ�ӳ����������ɭ�ֽ���״������Ҫָ��, ֱ�ӻ��ӵ�Ӱ�쵽ɭ������������������.�о���ʾ, ɭ������̼����ԼΪȫ������̼������73% (Post & Kwon, 2000), �ڵ���ȫ��̼ƽ�ⷽ��������Ҫ����.��������Ϊ½��ֲ����̬ϵͳ����Ҫ���(Kauppi et al., 2006), ��ά����������Ժ�����̼��ѭ���а�������Ҫ��ɫ(Yang et al., 2011).ɭ�ָ�������ı���ֲ��Ⱥ��ṹ��������ɼ����ܶȶ�Ӱ�쵽��������, ���������л�̼����������ֺ�����������Է����仯(Chandra et al., 2016; Feng et al., 2019).���о���Ϊ, ɭ��ֲ�������������ڽ��������л�̼�ֽ����ʳ�������ֲ�������������л�̼�Ĺ̳�����, ʹ���京����Խϵ�(Ohtsuka et al., 2010; Milcu et al., 2011), Ҳ���о�����, ����̼����������ɭ������������ֳ�����(Deng et al., 2013; ��ѩ��, 2016; Liu et al., 2020)���仯������(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)���Ƚ�����(Tang et al., 2009)�ı仯����, ˵��ɭ�����������̼��������Ӱ���Ϊ����.�����л��ʽ����ܵ����������Ӱ��(Sinsabaugh, 2010; Liu et al., 2020), ������������Ҳ���ܵ�����̼����Ч�Ե���Լ(Kooch et al., 2018; Dove et al., 2019).ɭ������ı�����������, ����������ɻ���֮�����仯, �����ı��������������(Zhang et al., 2017), �������Ҫ��������Դ�ڶ�ֲ�����ֽ���������������Լ�ֲ���ϵ������İ���ø��.����ø���л�̼���ֽ����ת����Ҫ����(Weintraub et al., 2007; Xu et al., 2017), ���������л�̼���Ļ�������������ø�Ŀֽ�����(Sinsabaugh et al., 2014; Mayer et al., 2020).�����л��ʿ������ܿ��ڰ���ø����, ����ˮ��ø��Ҫ�������̼���ֽ�ת��(�����ࡢ���ۡ���ά�ص�), ������-��������ø(��G)����ά��ˮ��ø(CBH)����-����������������ø(NAG)��; ����ø��Ҫ��������л�̼�����(��ľ����)�����ֽ��������ֳ�����̵�, �����������ø(PHO)����������ø(PEO)��(Luo et al., 2019), ���, ø���ԵĸߵͿɱ��������������������Լ������ֹ���ˮƽ֮��Ĺ�ϵ.���о���ʾ, ����ø�������������ֺ������������(Burns et al., 2013; Das & Mondal, 2019)�����������(Li et al., 2015; Feng et al., 2019).ͬʱ, �����������Ч��Ҳ��Ӱ���л�̼�͵��ķֽ�ø����, ʹ������P�����л��ʱ��ֳ���ͬ�ķֽ�����(Kang et al., 2009; Zhou et al., 2020).����, ����ø������������ʪ�ȼ�pH�������й�(��ΰΰ��, 2019; Zhou et al., 2020), ˵������ø����������ͷ��������صĸ������й�.��Ȼ��������ø�����������л�̼����ֺ�����Ĺ�ϵ�ѱ��㷺��ע, �����ڴ����Ǹ�ɽ��Ȼ�����������л�̼����������������ø����������ʶ�������. ...
... ɭ�������л�̼�����������Ƿ�ӳ����������ɭ�ֽ���״������Ҫָ��, ֱ�ӻ��ӵ�Ӱ�쵽ɭ������������������.�о���ʾ, ɭ������̼����ԼΪȫ������̼������73% (Post & Kwon, 2000), �ڵ���ȫ��̼ƽ�ⷽ��������Ҫ����.��������Ϊ½��ֲ����̬ϵͳ����Ҫ���(Kauppi et al., 2006), ��ά����������Ժ�����̼��ѭ���а�������Ҫ��ɫ(Yang et al., 2011).ɭ�ָ�������ı���ֲ��Ⱥ��ṹ��������ɼ����ܶȶ�Ӱ�쵽��������, ���������л�̼����������ֺ�����������Է����仯(Chandra et al., 2016; Feng et al., 2019).���о���Ϊ, ɭ��ֲ�������������ڽ��������л�̼�ֽ����ʳ�������ֲ�������������л�̼�Ĺ̳�����, ʹ���京����Խϵ�(Ohtsuka et al., 2010; Milcu et al., 2011), Ҳ���о�����, ����̼����������ɭ������������ֳ�����(Deng et al., 2013; ��ѩ��, 2016; Liu et al., 2020)���仯������(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)���Ƚ�����(Tang et al., 2009)�ı仯����, ˵��ɭ�����������̼��������Ӱ���Ϊ����.�����л��ʽ����ܵ����������Ӱ��(Sinsabaugh, 2010; Liu et al., 2020), ������������Ҳ���ܵ�����̼����Ч�Ե���Լ(Kooch et al., 2018; Dove et al., 2019).ɭ������ı�����������, ����������ɻ���֮�����仯, �����ı��������������(Zhang et al., 2017), �������Ҫ��������Դ�ڶ�ֲ�����ֽ���������������Լ�ֲ���ϵ������İ���ø��.����ø���л�̼���ֽ����ת����Ҫ����(Weintraub et al., 2007; Xu et al., 2017), ���������л�̼���Ļ�������������ø�Ŀֽ�����(Sinsabaugh et al., 2014; Mayer et al., 2020).�����л��ʿ������ܿ��ڰ���ø����, ����ˮ��ø��Ҫ�������̼���ֽ�ת��(�����ࡢ���ۡ���ά�ص�), ������-��������ø(��G)����ά��ˮ��ø(CBH)����-����������������ø(NAG)��; ����ø��Ҫ��������л�̼�����(��ľ����)�����ֽ��������ֳ�����̵�, �����������ø(PHO)����������ø(PEO)��(Luo et al., 2019), ���, ø���ԵĸߵͿɱ��������������������Լ������ֹ���ˮƽ֮��Ĺ�ϵ.���о���ʾ, ����ø�������������ֺ������������(Burns et al., 2013; Das & Mondal, 2019)�����������(Li et al., 2015; Feng et al., 2019).ͬʱ, �����������Ч��Ҳ��Ӱ���л�̼�͵��ķֽ�ø����, ʹ������P�����л��ʱ��ֳ���ͬ�ķֽ�����(Kang et al., 2009; Zhou et al., 2020).����, ����ø������������ʪ�ȼ�pH�������й�(��ΰΰ��, 2019; Zhou et al., 2020), ˵������ø����������ͷ��������صĸ������й�.��Ȼ��������ø�����������л�̼����ֺ�����Ĺ�ϵ�ѱ��㷺��ע, �����ڴ����Ǹ�ɽ��Ȼ�����������л�̼����������������ø����������ʶ�������. ...
Plant diversity and soil stoichiometry regulates the changes in multifunctionality during pine temperate forest secondary succession 1 2019
... ɭ�������л�̼�����������Ƿ�ӳ����������ɭ�ֽ���״������Ҫָ��, ֱ�ӻ��ӵ�Ӱ�쵽ɭ������������������.�о���ʾ, ɭ������̼����ԼΪȫ������̼������73% (Post & Kwon, 2000), �ڵ���ȫ��̼ƽ�ⷽ��������Ҫ����.��������Ϊ½��ֲ����̬ϵͳ����Ҫ���(Kauppi et al., 2006), ��ά����������Ժ�����̼��ѭ���а�������Ҫ��ɫ(Yang et al., 2011).ɭ�ָ�������ı���ֲ��Ⱥ��ṹ��������ɼ����ܶȶ�Ӱ�쵽��������, ���������л�̼����������ֺ�����������Է����仯(Chandra et al., 2016; Feng et al., 2019).���о���Ϊ, ɭ��ֲ�������������ڽ��������л�̼�ֽ����ʳ�������ֲ�������������л�̼�Ĺ̳�����, ʹ���京����Խϵ�(Ohtsuka et al., 2010; Milcu et al., 2011), Ҳ���о�����, ����̼����������ɭ������������ֳ�����(Deng et al., 2013; ��ѩ��, 2016; Liu et al., 2020)���仯������(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)���Ƚ�����(Tang et al., 2009)�ı仯����, ˵��ɭ�����������̼��������Ӱ���Ϊ����.�����л��ʽ����ܵ����������Ӱ��(Sinsabaugh, 2010; Liu et al., 2020), ������������Ҳ���ܵ�����̼����Ч�Ե���Լ(Kooch et al., 2018; Dove et al., 2019).ɭ������ı�����������, ����������ɻ���֮�����仯, �����ı��������������(Zhang et al., 2017), �������Ҫ��������Դ�ڶ�ֲ�����ֽ���������������Լ�ֲ���ϵ������İ���ø��.����ø���л�̼���ֽ����ת����Ҫ����(Weintraub et al., 2007; Xu et al., 2017), ���������л�̼���Ļ�������������ø�Ŀֽ�����(Sinsabaugh et al., 2014; Mayer et al., 2020).�����л��ʿ������ܿ��ڰ���ø����, ����ˮ��ø��Ҫ�������̼���ֽ�ת��(�����ࡢ���ۡ���ά�ص�), ������-��������ø(��G)����ά��ˮ��ø(CBH)����-����������������ø(NAG)��; ����ø��Ҫ��������л�̼�����(��ľ����)�����ֽ��������ֳ�����̵�, �����������ø(PHO)����������ø(PEO)��(Luo et al., 2019), ���, ø���ԵĸߵͿɱ��������������������Լ������ֹ���ˮƽ֮��Ĺ�ϵ.���о���ʾ, ����ø�������������ֺ������������(Burns et al., 2013; Das & Mondal, 2019)�����������(Li et al., 2015; Feng et al., 2019).ͬʱ, �����������Ч��Ҳ��Ӱ���л�̼�͵��ķֽ�ø����, ʹ������P�����л��ʱ��ֳ���ͬ�ķֽ�����(Kang et al., 2009; Zhou et al., 2020).����, ����ø������������ʪ�ȼ�pH�������й�(��ΰΰ��, 2019; Zhou et al., 2020), ˵������ø����������ͷ��������صĸ������й�.��Ȼ��������ø�����������л�̼����ֺ�����Ĺ�ϵ�ѱ��㷺��ע, �����ڴ����Ǹ�ɽ��Ȼ�����������л�̼����������������ø����������ʶ�������. ...
Effects of forest conversion on carbon-degrading enzyme activities in subtropical China 1 2019
... ɭ�������л�̼�����������Ƿ�ӳ����������ɭ�ֽ���״������Ҫָ��, ֱ�ӻ��ӵ�Ӱ�쵽ɭ������������������.�о���ʾ, ɭ������̼����ԼΪȫ������̼������73% (Post & Kwon, 2000), �ڵ���ȫ��̼ƽ�ⷽ��������Ҫ����.��������Ϊ½��ֲ����̬ϵͳ����Ҫ���(Kauppi et al., 2006), ��ά����������Ժ�����̼��ѭ���а�������Ҫ��ɫ(Yang et al., 2011).ɭ�ָ�������ı���ֲ��Ⱥ��ṹ��������ɼ����ܶȶ�Ӱ�쵽��������, ���������л�̼����������ֺ�����������Է����仯(Chandra et al., 2016; Feng et al., 2019).���о���Ϊ, ɭ��ֲ�������������ڽ��������л�̼�ֽ����ʳ�������ֲ�������������л�̼�Ĺ̳�����, ʹ���京����Խϵ�(Ohtsuka et al., 2010; Milcu et al., 2011), Ҳ���о�����, ����̼����������ɭ������������ֳ�����(Deng et al., 2013; ��ѩ��, 2016; Liu et al., 2020)���仯������(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)���Ƚ�����(Tang et al., 2009)�ı仯����, ˵��ɭ�����������̼��������Ӱ���Ϊ����.�����л��ʽ����ܵ����������Ӱ��(Sinsabaugh, 2010; Liu et al., 2020), ������������Ҳ���ܵ�����̼����Ч�Ե���Լ(Kooch et al., 2018; Dove et al., 2019).ɭ������ı�����������, ����������ɻ���֮�����仯, �����ı��������������(Zhang et al., 2017), �������Ҫ��������Դ�ڶ�ֲ�����ֽ���������������Լ�ֲ���ϵ������İ���ø��.����ø���л�̼���ֽ����ת����Ҫ����(Weintraub et al., 2007; Xu et al., 2017), ���������л�̼���Ļ�������������ø�Ŀֽ�����(Sinsabaugh et al., 2014; Mayer et al., 2020).�����л��ʿ������ܿ��ڰ���ø����, ����ˮ��ø��Ҫ�������̼���ֽ�ת��(�����ࡢ���ۡ���ά�ص�), ������-��������ø(��G)����ά��ˮ��ø(CBH)����-����������������ø(NAG)��; ����ø��Ҫ��������л�̼�����(��ľ����)�����ֽ��������ֳ�����̵�, �����������ø(PHO)����������ø(PEO)��(Luo et al., 2019), ���, ø���ԵĸߵͿɱ��������������������Լ������ֹ���ˮƽ֮��Ĺ�ϵ.���о���ʾ, ����ø�������������ֺ������������(Burns et al., 2013; Das & Mondal, 2019)�����������(Li et al., 2015; Feng et al., 2019).ͬʱ, �����������Ч��Ҳ��Ӱ���л�̼�͵��ķֽ�ø����, ʹ������P�����л��ʱ��ֳ���ͬ�ķֽ�����(Kang et al., 2009; Zhou et al., 2020).����, ����ø������������ʪ�ȼ�pH�������й�(��ΰΰ��, 2019; Zhou et al., 2020), ˵������ø����������ͷ��������صĸ������й�.��Ȼ��������ø�����������л�̼����ֺ�����Ĺ�ϵ�ѱ��㷺��ע, �����ڴ����Ǹ�ɽ��Ȼ�����������л�̼����������������ø����������ʶ�������. ...
Root-microbial interaction accelerates soil nitrogen depletion but not soil carbon after increasing litter inputs to a coniferous forest 1 2019
... ���������л�̼���������������л��ʵ���Ҫ���, ��Ҫ��Դ���½���֦��Ҷ����ֳ�ʡ���ϵ�������Լ��������(Kalbitz et al., 2000; ���µ�, 2018).ɭ�ֲ�ͬ��Ȼ���»ָ���, ���ַ������������Իָ���������ɵIJ���, ���µر�������������������������ʪ�ȡ�������ɵȷ�����ͬ�̶ȱ仯, �Ӷ�Ӱ�쵽���������л�̼�������ı仯(��ѩ��, 2016; Hagedorn et al., 2019).���о���, DOC��LFOC��DON�����������������������½�����(p < 0.05), ����70-NSF��80-NSF�в��첻����(��3), �����浽ẽ���ɼԭʼ�ֽ�, ����������̼��������������(p < 0.05), ����������˹�����������б���(0-20 cm)��������̼���������о��������(������, 2009), ����ͬ���ȴ�(���ܵ�, 2019)�����ȴ�(��Ծ�µ�, 2013)ɭ��������о����, ˵����ͬ��ʪ������ֲ���ַֽṹ�����������л�̼����ֺ�����Ӱ����ֳ���ͬ�ı仯����(Li et al., 2014; Taylor et al., 2019).����ԭ��, һ��������ɭ�ָ�������, ����̼������������(Mylliemngap et al., 2016), ����Ҷ�ֵĿ����ʸ�����Ҷ��(C?t�� et al., 2000); ��һ�������ڱ��������ܶ�����������, �������ž۶ȡ��ṹϵ�����ܿ�϶��ë�ܿ�϶������������������ֽ�������(��ѧ�µ�, 2003), �����л�̼����ȱ�������ž���������Ա���, �������������л�̼��DON��������.����, ẽ���ɼԭʼ�ֵ���������̼����ֺ���������Ȼ������(��3), ��������ַ���Ҫ���ֹ���(��Ҷ+��Ҷ����)��һ������, Դ�ڸ������������ֵ���Ҷ�ں���һ�����ڷֽ�(����÷��, 2015), ���ͷŵ����ֱ�������ֽ�����, �������������л�̼��������.��Ҫ��������Ӱ�����ض����������л�̼������������, �����һ�������о�.��ط�������, ��ͬ��Ȼ����������ֵر�������SOC��TN����������л�̼����ֺ�����DON�������ʼ���������ع�ϵ(p < 0.01), ˵�����������л�̼�͵�����������SOC��TN����֮���ϵ��Ϊ����, ���������о�(�����, 2016; ������, 2017) һ��, ˵����Ȼ�����������������ǵ�����̼�ֵ̳���Ҫ����(Li et al., 2012; Smal et al., 2019). ...
��ͬˮ��ˮƽ�Դ����Ǹ�ɽ�ֵ�����ø���Ե�Ӱ�� 1 2019
... ɭ�������л�̼�����������Ƿ�ӳ����������ɭ�ֽ���״������Ҫָ��, ֱ�ӻ��ӵ�Ӱ�쵽ɭ������������������.�о���ʾ, ɭ������̼����ԼΪȫ������̼������73% (Post & Kwon, 2000), �ڵ���ȫ��̼ƽ�ⷽ��������Ҫ����.��������Ϊ½��ֲ����̬ϵͳ����Ҫ���(Kauppi et al., 2006), ��ά����������Ժ�����̼��ѭ���а�������Ҫ��ɫ(Yang et al., 2011).ɭ�ָ�������ı���ֲ��Ⱥ��ṹ��������ɼ����ܶȶ�Ӱ�쵽��������, ���������л�̼����������ֺ�����������Է����仯(Chandra et al., 2016; Feng et al., 2019).���о���Ϊ, ɭ��ֲ�������������ڽ��������л�̼�ֽ����ʳ�������ֲ�������������л�̼�Ĺ̳�����, ʹ���京����Խϵ�(Ohtsuka et al., 2010; Milcu et al., 2011), Ҳ���о�����, ����̼����������ɭ������������ֳ�����(Deng et al., 2013; ��ѩ��, 2016; Liu et al., 2020)���仯������(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)���Ƚ�����(Tang et al., 2009)�ı仯����, ˵��ɭ�����������̼��������Ӱ���Ϊ����.�����л��ʽ����ܵ����������Ӱ��(Sinsabaugh, 2010; Liu et al., 2020), ������������Ҳ���ܵ�����̼����Ч�Ե���Լ(Kooch et al., 2018; Dove et al., 2019).ɭ������ı�����������, ����������ɻ���֮�����仯, �����ı��������������(Zhang et al., 2017), �������Ҫ��������Դ�ڶ�ֲ�����ֽ���������������Լ�ֲ���ϵ������İ���ø��.����ø���л�̼���ֽ����ת����Ҫ����(Weintraub et al., 2007; Xu et al., 2017), ���������л�̼���Ļ�������������ø�Ŀֽ�����(Sinsabaugh et al., 2014; Mayer et al., 2020).�����л��ʿ������ܿ��ڰ���ø����, ����ˮ��ø��Ҫ�������̼���ֽ�ת��(�����ࡢ���ۡ���ά�ص�), ������-��������ø(��G)����ά��ˮ��ø(CBH)����-����������������ø(NAG)��; ����ø��Ҫ��������л�̼�����(��ľ����)�����ֽ��������ֳ�����̵�, �����������ø(PHO)����������ø(PEO)��(Luo et al., 2019), ���, ø���ԵĸߵͿɱ��������������������Լ������ֹ���ˮƽ֮��Ĺ�ϵ.���о���ʾ, ����ø�������������ֺ������������(Burns et al., 2013; Das & Mondal, 2019)�����������(Li et al., 2015; Feng et al., 2019).ͬʱ, �����������Ч��Ҳ��Ӱ���л�̼�͵��ķֽ�ø����, ʹ������P�����л��ʱ��ֳ���ͬ�ķֽ�����(Kang et al., 2009; Zhou et al., 2020).����, ����ø������������ʪ�ȼ�pH�������й�(��ΰΰ��, 2019; Zhou et al., 2020), ˵������ø����������ͷ��������صĸ������й�.��Ȼ��������ø�����������л�̼����ֺ�����Ĺ�ϵ�ѱ��㷺��ע, �����ڴ����Ǹ�ɽ��Ȼ�����������л�̼����������������ø����������ʶ�������. ...
��ͬˮ��ˮƽ�Դ����Ǹ�ɽ�ֵ�����ø���Ե�Ӱ�� 1 2019
... ɭ�������л�̼�����������Ƿ�ӳ����������ɭ�ֽ���״������Ҫָ��, ֱ�ӻ��ӵ�Ӱ�쵽ɭ������������������.�о���ʾ, ɭ������̼����ԼΪȫ������̼������73% (Post & Kwon, 2000), �ڵ���ȫ��̼ƽ�ⷽ��������Ҫ����.��������Ϊ½��ֲ����̬ϵͳ����Ҫ���(Kauppi et al., 2006), ��ά����������Ժ�����̼��ѭ���а�������Ҫ��ɫ(Yang et al., 2011).ɭ�ָ�������ı���ֲ��Ⱥ��ṹ��������ɼ����ܶȶ�Ӱ�쵽��������, ���������л�̼����������ֺ�����������Է����仯(Chandra et al., 2016; Feng et al., 2019).���о���Ϊ, ɭ��ֲ�������������ڽ��������л�̼�ֽ����ʳ�������ֲ�������������л�̼�Ĺ̳�����, ʹ���京����Խϵ�(Ohtsuka et al., 2010; Milcu et al., 2011), Ҳ���о�����, ����̼����������ɭ������������ֳ�����(Deng et al., 2013; ��ѩ��, 2016; Liu et al., 2020)���仯������(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)���Ƚ�����(Tang et al., 2009)�ı仯����, ˵��ɭ�����������̼��������Ӱ���Ϊ����.�����л��ʽ����ܵ����������Ӱ��(Sinsabaugh, 2010; Liu et al., 2020), ������������Ҳ���ܵ�����̼����Ч�Ե���Լ(Kooch et al., 2018; Dove et al., 2019).ɭ������ı�����������, ����������ɻ���֮�����仯, �����ı��������������(Zhang et al., 2017), �������Ҫ��������Դ�ڶ�ֲ�����ֽ���������������Լ�ֲ���ϵ������İ���ø��.����ø���л�̼���ֽ����ת����Ҫ����(Weintraub et al., 2007; Xu et al., 2017), ���������л�̼���Ļ�������������ø�Ŀֽ�����(Sinsabaugh et al., 2014; Mayer et al., 2020).�����л��ʿ������ܿ��ڰ���ø����, ����ˮ��ø��Ҫ�������̼���ֽ�ת��(�����ࡢ���ۡ���ά�ص�), ������-��������ø(��G)����ά��ˮ��ø(CBH)����-����������������ø(NAG)��; ����ø��Ҫ��������л�̼�����(��ľ����)�����ֽ��������ֳ�����̵�, �����������ø(PHO)����������ø(PEO)��(Luo et al., 2019), ���, ø���ԵĸߵͿɱ��������������������Լ������ֹ���ˮƽ֮��Ĺ�ϵ.���о���ʾ, ����ø�������������ֺ������������(Burns et al., 2013; Das & Mondal, 2019)�����������(Li et al., 2015; Feng et al., 2019).ͬʱ, �����������Ч��Ҳ��Ӱ���л�̼�͵��ķֽ�ø����, ʹ������P�����л��ʱ��ֳ���ͬ�ķֽ�����(Kang et al., 2009; Zhou et al., 2020).����, ����ø������������ʪ�ȼ�pH�������й�(��ΰΰ��, 2019; Zhou et al., 2020), ˵������ø����������ͷ��������صĸ������й�.��Ȼ��������ø�����������л�̼����ֺ�����Ĺ�ϵ�ѱ��㷺��ע, �����ڴ����Ǹ�ɽ��Ȼ�����������л�̼����������������ø����������ʶ�������. ...
Tamm Review: Influence of forest management activities on soil organic carbon stocks: a knowledge synthesis 1 2020
... ���������л�̼���������������л��ʵ���Ҫ���, ��Ҫ��Դ���½���֦��Ҷ����ֳ�ʡ���ϵ�������Լ��������(Kalbitz et al., 2000; ���µ�, 2018).ɭ�ֲ�ͬ��Ȼ���»ָ���, ���ַ������������Իָ���������ɵIJ���, ���µر�������������������������ʪ�ȡ�������ɵȷ�����ͬ�̶ȱ仯, �Ӷ�Ӱ�쵽���������л�̼�������ı仯(��ѩ��, 2016; Hagedorn et al., 2019).���о���, DOC��LFOC��DON�����������������������½�����(p < 0.05), ����70-NSF��80-NSF�в��첻����(��3), �����浽ẽ���ɼԭʼ�ֽ�, ����������̼��������������(p < 0.05), ����������˹�����������б���(0-20 cm)��������̼���������о��������(������, 2009), ����ͬ���ȴ�(���ܵ�, 2019)�����ȴ�(��Ծ�µ�, 2013)ɭ��������о����, ˵����ͬ��ʪ������ֲ���ַֽṹ�����������л�̼����ֺ�����Ӱ����ֳ���ͬ�ı仯����(Li et al., 2014; Taylor et al., 2019).����ԭ��, һ��������ɭ�ָ�������, ����̼������������(Mylliemngap et al., 2016), ����Ҷ�ֵĿ����ʸ�����Ҷ��(C?t�� et al., 2000); ��һ�������ڱ��������ܶ�����������, �������ž۶ȡ��ṹϵ�����ܿ�϶��ë�ܿ�϶������������������ֽ�������(��ѧ�µ�, 2003), �����л�̼����ȱ�������ž���������Ա���, �������������л�̼��DON��������.����, ẽ���ɼԭʼ�ֵ���������̼����ֺ���������Ȼ������(��3), ��������ַ���Ҫ���ֹ���(��Ҷ+��Ҷ����)��һ������, Դ�ڸ������������ֵ���Ҷ�ں���һ�����ڷֽ�(����÷��, 2015), ���ͷŵ����ֱ�������ֽ�����, �������������л�̼��������.��Ҫ��������Ӱ�����ض����������л�̼������������, �����һ�������о�.��ط�������, ��ͬ��Ȼ����������ֵر�������SOC��TN����������л�̼����ֺ�����DON�������ʼ���������ع�ϵ(p < 0.01), ˵�����������л�̼�͵�����������SOC��TN����֮���ϵ��Ϊ����, ���������о�(�����, 2016; ������, 2017) һ��, ˵����Ȼ�����������������ǵ�����̼�ֵ̳���Ҫ����(Li et al., 2012; Smal et al., 2019). ...
Identification of general patterns of nutrient and labile carbon control on soil carbon dynamics across a successional gradient 2011
Changes in vegetation and nitrogen mineralization during recovery of a montane subtropical broadleaved forest in North-eastern India following anthropogenic disturbance 1 2016
Carbon cycling and net ecosystem production at an early stage of secondary succession in an abandoned coppice forest 1 2010
... ɭ�ָ�������Ļָ�������, �ַֽṹ��������ɡ����������Լ������������˲�ͬ�̶ȵı仯, ��������������̼��Դ��ͬ�����������л��ʷֽ����ʡ�������������Լ����л������仯(Wardle et al., 2004; Lucas-Borja & Delgado- Baquerizo, 2019), ����Ӱ�쵽����ø����(Adamczyk et al., 2015; Xu et al., 2017).����, ��ƽ�����¡���ˮ������������Ժ����ֶ�����Ҳ��Ӱ������ø���Ե���Ҫ����(�����, 2014; Zhou et al., 2020).���о���, ��Ȼ�����ָ�����������(<60 a)��ͬ�εı�������ˮ��ø(��G��CBH��NAG)������������ẽ���ɼԭʼ��(��4), ������Ȼ����������̳�������������(80-NSF > 70-NSF > 60-NSF), ��ͬ�ڴ����Ǹ�ɽƽ���ذ�����ˮ��ø���Ը�����ɼ��(�ܺ��ٵ�, 2004)���´�����(Betula platyphylla)����������̱���Ϊ�����Ӻ͵ı仯����(���Ե�, 2013), һ����˵��ẽ���ɼԭʼ�ֵ���������ѭ��������Խϸ�, ���������ֹ�������; ��һ����˵����������Ȼ�������������, ���ϵ��������ӵ�������̼�����������ֺ������½�(Liu et al., 2017; Lyu et al., 2019), ʹ������������Խ���(�����, 2018), ������������ˮ��ø�����½�.����, 80-NSF��70-NSF��60-NSF����������ø���Ա仯������, ��PHO�Ļ�����������CK, ��PEO�Ļ�����CK���첻����(��4), ��ԭ�������ẽ���ɼԭʼ���к��н�Ϊ�ḻ��ľ���ء����ʺ����ʵ��ѽ��⻯����, �ٽ��˽���ľ���ص�����Ⱥ������Ի�������������, ��������ָ�������εĽ���, ��ϵ��������ֲ��������������仯, ����������pH����, ��ʹPHO��������(Fujii et al., 2013). ...
Responses of soil labile organic carbon fractions and stocks to different vegetation restoration strategies in degraded karst ecosystems of southwest China 1 2019
... ɭ�������л�̼�����������Ƿ�ӳ����������ɭ�ֽ���״������Ҫָ��, ֱ�ӻ��ӵ�Ӱ�쵽ɭ������������������.�о���ʾ, ɭ������̼����ԼΪȫ������̼������73% (Post & Kwon, 2000), �ڵ���ȫ��̼ƽ�ⷽ��������Ҫ����.��������Ϊ½��ֲ����̬ϵͳ����Ҫ���(Kauppi et al., 2006), ��ά����������Ժ�����̼��ѭ���а�������Ҫ��ɫ(Yang et al., 2011).ɭ�ָ�������ı���ֲ��Ⱥ��ṹ��������ɼ����ܶȶ�Ӱ�쵽��������, ���������л�̼����������ֺ�����������Է����仯(Chandra et al., 2016; Feng et al., 2019).���о���Ϊ, ɭ��ֲ�������������ڽ��������л�̼�ֽ����ʳ�������ֲ�������������л�̼�Ĺ̳�����, ʹ���京����Խϵ�(Ohtsuka et al., 2010; Milcu et al., 2011), Ҳ���о�����, ����̼����������ɭ������������ֳ�����(Deng et al., 2013; ��ѩ��, 2016; Liu et al., 2020)���仯������(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)���Ƚ�����(Tang et al., 2009)�ı仯����, ˵��ɭ�����������̼��������Ӱ���Ϊ����.�����л��ʽ����ܵ����������Ӱ��(Sinsabaugh, 2010; Liu et al., 2020), ������������Ҳ���ܵ�����̼����Ч�Ե���Լ(Kooch et al., 2018; Dove et al., 2019).ɭ������ı�����������, ����������ɻ���֮�����仯, �����ı��������������(Zhang et al., 2017), �������Ҫ��������Դ�ڶ�ֲ�����ֽ���������������Լ�ֲ���ϵ������İ���ø��.����ø���л�̼���ֽ����ת����Ҫ����(Weintraub et al., 2007; Xu et al., 2017), ���������л�̼���Ļ�������������ø�Ŀֽ�����(Sinsabaugh et al., 2014; Mayer et al., 2020).�����л��ʿ������ܿ��ڰ���ø����, ����ˮ��ø��Ҫ�������̼���ֽ�ת��(�����ࡢ���ۡ���ά�ص�), ������-��������ø(��G)����ά��ˮ��ø(CBH)����-����������������ø(NAG)��; ����ø��Ҫ��������л�̼�����(��ľ����)�����ֽ��������ֳ�����̵�, �����������ø(PHO)����������ø(PEO)��(Luo et al., 2019), ���, ø���ԵĸߵͿɱ��������������������Լ������ֹ���ˮƽ֮��Ĺ�ϵ.���о���ʾ, ����ø�������������ֺ������������(Burns et al., 2013; Das & Mondal, 2019)�����������(Li et al., 2015; Feng et al., 2019).ͬʱ, �����������Ч��Ҳ��Ӱ���л�̼�͵��ķֽ�ø����, ʹ������P�����л��ʱ��ֳ���ͬ�ķֽ�����(Kang et al., 2009; Zhou et al., 2020).����, ����ø������������ʪ�ȼ�pH�������й�(��ΰΰ��, 2019; Zhou et al., 2020), ˵������ø����������ͷ��������صĸ������й�.��Ȼ��������ø�����������л�̼����ֺ�����Ĺ�ϵ�ѱ��㷺��ע, �����ڴ����Ǹ�ɽ��Ȼ�����������л�̼����������������ø����������ʶ�������. ...
�����Ǹ�ɽ��Ҷ��ֲ��Ⱥ��������������ʵ�Ӱ�� 2003
�����Ǹ�ɽ��Ҷ��ֲ��Ⱥ��������������ʵ�Ӱ�� 2003
The relation among soil microorganism, enzyme activity and soil nutrients under subalpine coniferous forest in Western Sichuan 2009
Soil carbon sequestration and land-use change: processes and potential 1 2000
... ɭ�ָ�������Ļָ�������, �ַֽṹ��������ɡ����������Լ������������˲�ͬ�̶ȵı仯, ��������������̼��Դ��ͬ�����������л��ʷֽ����ʡ�������������Լ����л������仯(Wardle et al., 2004; Lucas-Borja & Delgado- Baquerizo, 2019), ����Ӱ�쵽����ø����(Adamczyk et al., 2015; Xu et al., 2017).����, ��ƽ�����¡���ˮ������������Ժ����ֶ�����Ҳ��Ӱ������ø���Ե���Ҫ����(�����, 2014; Zhou et al., 2020).���о���, ��Ȼ�����ָ�����������(<60 a)��ͬ�εı�������ˮ��ø(��G��CBH��NAG)������������ẽ���ɼԭʼ��(��4), ������Ȼ����������̳�������������(80-NSF > 70-NSF > 60-NSF), ��ͬ�ڴ����Ǹ�ɽƽ���ذ�����ˮ��ø���Ը�����ɼ��(�ܺ��ٵ�, 2004)���´�����(Betula platyphylla)����������̱���Ϊ�����Ӻ͵ı仯����(���Ե�, 2013), һ����˵��ẽ���ɼԭʼ�ֵ���������ѭ��������Խϸ�, ���������ֹ�������; ��һ����˵����������Ȼ�������������, ���ϵ��������ӵ�������̼�����������ֺ������½�(Liu et al., 2017; Lyu et al., 2019), ʹ������������Խ���(�����, 2018), ������������ˮ��ø�����½�.����, 80-NSF��70-NSF��60-NSF����������ø���Ա仯������, ��PHO�Ļ�����������CK, ��PEO�Ļ�����CK���첻����(��4), ��ԭ�������ẽ���ɼԭʼ���к��н�Ϊ�ḻ��ľ���ء����ʺ����ʵ��ѽ��⻯����, �ٽ��˽���ľ���ص�����Ⱥ������Ի�������������, ��������ָ�������εĽ���, ��ϵ��������ֲ��������������仯, ����������pH����, ��ʹPHO��������(Fujii et al., 2013). ...
Chemical composition of soil organic matter and potential enzyme activity in the topsoil along a moisture gradient in the High Arctic (Svalbard) 1 2020
... ɭ�������л�̼�����������Ƿ�ӳ����������ɭ�ֽ���״������Ҫָ��, ֱ�ӻ��ӵ�Ӱ�쵽ɭ������������������.�о���ʾ, ɭ������̼����ԼΪȫ������̼������73% (Post & Kwon, 2000), �ڵ���ȫ��̼ƽ�ⷽ��������Ҫ����.��������Ϊ½��ֲ����̬ϵͳ����Ҫ���(Kauppi et al., 2006), ��ά����������Ժ�����̼��ѭ���а�������Ҫ��ɫ(Yang et al., 2011).ɭ�ָ�������ı���ֲ��Ⱥ��ṹ��������ɼ����ܶȶ�Ӱ�쵽��������, ���������л�̼����������ֺ�����������Է����仯(Chandra et al., 2016; Feng et al., 2019).���о���Ϊ, ɭ��ֲ�������������ڽ��������л�̼�ֽ����ʳ�������ֲ�������������л�̼�Ĺ̳�����, ʹ���京����Խϵ�(Ohtsuka et al., 2010; Milcu et al., 2011), Ҳ���о�����, ����̼����������ɭ������������ֳ�����(Deng et al., 2013; ��ѩ��, 2016; Liu et al., 2020)���仯������(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)���Ƚ�����(Tang et al., 2009)�ı仯����, ˵��ɭ�����������̼��������Ӱ���Ϊ����.�����л��ʽ����ܵ����������Ӱ��(Sinsabaugh, 2010; Liu et al., 2020), ������������Ҳ���ܵ�����̼����Ч�Ե���Լ(Kooch et al., 2018; Dove et al., 2019).ɭ������ı�����������, ����������ɻ���֮�����仯, �����ı��������������(Zhang et al., 2017), �������Ҫ��������Դ�ڶ�ֲ�����ֽ���������������Լ�ֲ���ϵ������İ���ø��.����ø���л�̼���ֽ����ת����Ҫ����(Weintraub et al., 2007; Xu et al., 2017), ���������л�̼���Ļ�������������ø�Ŀֽ�����(Sinsabaugh et al., 2014; Mayer et al., 2020).�����л��ʿ������ܿ��ڰ���ø����, ����ˮ��ø��Ҫ�������̼���ֽ�ת��(�����ࡢ���ۡ���ά�ص�), ������-��������ø(��G)����ά��ˮ��ø(CBH)����-����������������ø(NAG)��; ����ø��Ҫ��������л�̼�����(��ľ����)�����ֽ��������ֳ�����̵�, �����������ø(PHO)����������ø(PEO)��(Luo et al., 2019), ���, ø���ԵĸߵͿɱ��������������������Լ������ֹ���ˮƽ֮��Ĺ�ϵ.���о���ʾ, ����ø�������������ֺ������������(Burns et al., 2013; Das & Mondal, 2019)�����������(Li et al., 2015; Feng et al., 2019).ͬʱ, �����������Ч��Ҳ��Ӱ���л�̼�͵��ķֽ�ø����, ʹ������P�����л��ʱ��ֳ���ͬ�ķֽ�����(Kang et al., 2009; Zhou et al., 2020).����, ����ø������������ʪ�ȼ�pH�������й�(��ΰΰ��, 2019; Zhou et al., 2020), ˵������ø����������ͷ��������صĸ������й�.��Ȼ��������ø�����������л�̼����ֺ�����Ĺ�ϵ�ѱ��㷺��ע, �����ڴ����Ǹ�ɽ��Ȼ�����������л�̼����������������ø����������ʶ�������. ...
Priming alters soil carbon dynamics during forest succession 1 2019
... ɭ�ָ�������Ļָ�������, �ַֽṹ��������ɡ����������Լ������������˲�ͬ�̶ȵı仯, ��������������̼��Դ��ͬ�����������л��ʷֽ����ʡ�������������Լ����л������仯(Wardle et al., 2004; Lucas-Borja & Delgado- Baquerizo, 2019), ����Ӱ�쵽����ø����(Adamczyk et al., 2015; Xu et al., 2017).����, ��ƽ�����¡���ˮ������������Ժ����ֶ�����Ҳ��Ӱ������ø���Ե���Ҫ����(�����, 2014; Zhou et al., 2020).���о���, ��Ȼ�����ָ�����������(<60 a)��ͬ�εı�������ˮ��ø(��G��CBH��NAG)������������ẽ���ɼԭʼ��(��4), ������Ȼ����������̳�������������(80-NSF > 70-NSF > 60-NSF), ��ͬ�ڴ����Ǹ�ɽƽ���ذ�����ˮ��ø���Ը�����ɼ��(�ܺ��ٵ�, 2004)���´�����(Betula platyphylla)����������̱���Ϊ�����Ӻ͵ı仯����(���Ե�, 2013), һ����˵��ẽ���ɼԭʼ�ֵ���������ѭ��������Խϸ�, ���������ֹ�������; ��һ����˵����������Ȼ�������������, ���ϵ��������ӵ�������̼�����������ֺ������½�(Liu et al., 2017; Lyu et al., 2019), ʹ������������Խ���(�����, 2018), ������������ˮ��ø�����½�.����, 80-NSF��70-NSF��60-NSF����������ø���Ա仯������, ��PHO�Ļ�����������CK, ��PEO�Ļ�����CK���첻����(��4), ��ԭ�������ẽ���ɼԭʼ���к��н�Ϊ�ḻ��ľ���ء����ʺ����ʵ��ѽ��⻯����, �ٽ��˽���ľ���ص�����Ⱥ������Ի�������������, ��������ָ�������εĽ���, ��ϵ��������ֲ��������������仯, ����������pH����, ��ʹPHO��������(Fujii et al., 2013). ...
The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil 2002
Soil carbon dynamics in a chronosequence of secondary forests in northeastern Costa Rica 1 2008
... ɭ�������л�̼�����������Ƿ�ӳ����������ɭ�ֽ���״������Ҫָ��, ֱ�ӻ��ӵ�Ӱ�쵽ɭ������������������.�о���ʾ, ɭ������̼����ԼΪȫ������̼������73% (Post & Kwon, 2000), �ڵ���ȫ��̼ƽ�ⷽ��������Ҫ����.��������Ϊ½��ֲ����̬ϵͳ����Ҫ���(Kauppi et al., 2006), ��ά����������Ժ�����̼��ѭ���а�������Ҫ��ɫ(Yang et al., 2011).ɭ�ָ�������ı���ֲ��Ⱥ��ṹ��������ɼ����ܶȶ�Ӱ�쵽��������, ���������л�̼����������ֺ�����������Է����仯(Chandra et al., 2016; Feng et al., 2019).���о���Ϊ, ɭ��ֲ�������������ڽ��������л�̼�ֽ����ʳ�������ֲ�������������л�̼�Ĺ̳�����, ʹ���京����Խϵ�(Ohtsuka et al., 2010; Milcu et al., 2011), Ҳ���о�����, ����̼����������ɭ������������ֳ�����(Deng et al., 2013; ��ѩ��, 2016; Liu et al., 2020)���仯������(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)���Ƚ�����(Tang et al., 2009)�ı仯����, ˵��ɭ�����������̼��������Ӱ���Ϊ����.�����л��ʽ����ܵ����������Ӱ��(Sinsabaugh, 2010; Liu et al., 2020), ������������Ҳ���ܵ�����̼����Ч�Ե���Լ(Kooch et al., 2018; Dove et al., 2019).ɭ������ı�����������, ����������ɻ���֮�����仯, �����ı��������������(Zhang et al., 2017), �������Ҫ��������Դ�ڶ�ֲ�����ֽ���������������Լ�ֲ���ϵ������İ���ø��.����ø���л�̼���ֽ����ת����Ҫ����(Weintraub et al., 2007; Xu et al., 2017), ���������л�̼���Ļ�������������ø�Ŀֽ�����(Sinsabaugh et al., 2014; Mayer et al., 2020).�����л��ʿ������ܿ��ڰ���ø����, ����ˮ��ø��Ҫ�������̼���ֽ�ת��(�����ࡢ���ۡ���ά�ص�), ������-��������ø(��G)����ά��ˮ��ø(CBH)����-����������������ø(NAG)��; ����ø��Ҫ��������л�̼�����(��ľ����)�����ֽ��������ֳ�����̵�, �����������ø(PHO)����������ø(PEO)��(Luo et al., 2019), ���, ø���ԵĸߵͿɱ��������������������Լ������ֹ���ˮƽ֮��Ĺ�ϵ.���о���ʾ, ����ø�������������ֺ������������(Burns et al., 2013; Das & Mondal, 2019)�����������(Li et al., 2015; Feng et al., 2019).ͬʱ, �����������Ч��Ҳ��Ӱ���л�̼�͵��ķֽ�ø����, ʹ������P�����л��ʱ��ֳ���ͬ�ķֽ�����(Kang et al., 2009; Zhou et al., 2020).����, ����ø������������ʪ�ȼ�pH�������й�(��ΰΰ��, 2019; Zhou et al., 2020), ˵������ø����������ͷ��������صĸ������й�.��Ȼ��������ø�����������л�̼����ֺ�����Ĺ�ϵ�ѱ��㷺��ע, �����ڴ����Ǹ�ɽ��Ȼ�����������л�̼����������������ø����������ʶ�������. ...
�����Ǹ�ɽɭ�ֵ����ﲻͬ�ֽ�λ����������� 1 2017
... ɭ�������л�̼�����������Ƿ�ӳ����������ɭ�ֽ���״������Ҫָ��, ֱ�ӻ��ӵ�Ӱ�쵽ɭ������������������.�о���ʾ, ɭ������̼����ԼΪȫ������̼������73% (Post & Kwon, 2000), �ڵ���ȫ��̼ƽ�ⷽ��������Ҫ����.��������Ϊ½��ֲ����̬ϵͳ����Ҫ���(Kauppi et al., 2006), ��ά����������Ժ�����̼��ѭ���а�������Ҫ��ɫ(Yang et al., 2011).ɭ�ָ�������ı���ֲ��Ⱥ��ṹ��������ɼ����ܶȶ�Ӱ�쵽��������, ���������л�̼����������ֺ�����������Է����仯(Chandra et al., 2016; Feng et al., 2019).���о���Ϊ, ɭ��ֲ�������������ڽ��������л�̼�ֽ����ʳ�������ֲ�������������л�̼�Ĺ̳�����, ʹ���京����Խϵ�(Ohtsuka et al., 2010; Milcu et al., 2011), Ҳ���о�����, ����̼����������ɭ������������ֳ�����(Deng et al., 2013; ��ѩ��, 2016; Liu et al., 2020)���仯������(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)���Ƚ�����(Tang et al., 2009)�ı仯����, ˵��ɭ�����������̼��������Ӱ���Ϊ����.�����л��ʽ����ܵ����������Ӱ��(Sinsabaugh, 2010; Liu et al., 2020), ������������Ҳ���ܵ�����̼����Ч�Ե���Լ(Kooch et al., 2018; Dove et al., 2019).ɭ������ı�����������, ����������ɻ���֮�����仯, �����ı��������������(Zhang et al., 2017), �������Ҫ��������Դ�ڶ�ֲ�����ֽ���������������Լ�ֲ���ϵ������İ���ø��.����ø���л�̼���ֽ����ת����Ҫ����(Weintraub et al., 2007; Xu et al., 2017), ���������л�̼���Ļ�������������ø�Ŀֽ�����(Sinsabaugh et al., 2014; Mayer et al., 2020).�����л��ʿ������ܿ��ڰ���ø����, ����ˮ��ø��Ҫ�������̼���ֽ�ת��(�����ࡢ���ۡ���ά�ص�), ������-��������ø(��G)����ά��ˮ��ø(CBH)����-����������������ø(NAG)��; ����ø��Ҫ��������л�̼�����(��ľ����)�����ֽ��������ֳ�����̵�, �����������ø(PHO)����������ø(PEO)��(Luo et al., 2019), ���, ø���ԵĸߵͿɱ��������������������Լ������ֹ���ˮƽ֮��Ĺ�ϵ.���о���ʾ, ����ø�������������ֺ������������(Burns et al., 2013; Das & Mondal, 2019)�����������(Li et al., 2015; Feng et al., 2019).ͬʱ, �����������Ч��Ҳ��Ӱ���л�̼�͵��ķֽ�ø����, ʹ������P�����л��ʱ��ֳ���ͬ�ķֽ�����(Kang et al., 2009; Zhou et al., 2020).����, ����ø������������ʪ�ȼ�pH�������й�(��ΰΰ��, 2019; Zhou et al., 2020), ˵������ø����������ͷ��������صĸ������й�.��Ȼ��������ø�����������л�̼����ֺ�����Ĺ�ϵ�ѱ��㷺��ע, �����ڴ����Ǹ�ɽ��Ȼ�����������л�̼����������������ø����������ʶ�������. ...
�����Ǹ�ɽɭ�ֵ����ﲻͬ�ֽ�λ����������� 1 2017
... ɭ�������л�̼�����������Ƿ�ӳ����������ɭ�ֽ���״������Ҫָ��, ֱ�ӻ��ӵ�Ӱ�쵽ɭ������������������.�о���ʾ, ɭ������̼����ԼΪȫ������̼������73% (Post & Kwon, 2000), �ڵ���ȫ��̼ƽ�ⷽ��������Ҫ����.��������Ϊ½��ֲ����̬ϵͳ����Ҫ���(Kauppi et al., 2006), ��ά����������Ժ�����̼��ѭ���а�������Ҫ��ɫ(Yang et al., 2011).ɭ�ָ�������ı���ֲ��Ⱥ��ṹ��������ɼ����ܶȶ�Ӱ�쵽��������, ���������л�̼����������ֺ�����������Է����仯(Chandra et al., 2016; Feng et al., 2019).���о���Ϊ, ɭ��ֲ�������������ڽ��������л�̼�ֽ����ʳ�������ֲ�������������л�̼�Ĺ̳�����, ʹ���京����Խϵ�(Ohtsuka et al., 2010; Milcu et al., 2011), Ҳ���о�����, ����̼����������ɭ������������ֳ�����(Deng et al., 2013; ��ѩ��, 2016; Liu et al., 2020)���仯������(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)���Ƚ�����(Tang et al., 2009)�ı仯����, ˵��ɭ�����������̼��������Ӱ���Ϊ����.�����л��ʽ����ܵ����������Ӱ��(Sinsabaugh, 2010; Liu et al., 2020), ������������Ҳ���ܵ�����̼����Ч�Ե���Լ(Kooch et al., 2018; Dove et al., 2019).ɭ������ı�����������, ����������ɻ���֮�����仯, �����ı��������������(Zhang et al., 2017), �������Ҫ��������Դ�ڶ�ֲ�����ֽ���������������Լ�ֲ���ϵ������İ���ø��.����ø���л�̼���ֽ����ת����Ҫ����(Weintraub et al., 2007; Xu et al., 2017), ���������л�̼���Ļ�������������ø�Ŀֽ�����(Sinsabaugh et al., 2014; Mayer et al., 2020).�����л��ʿ������ܿ��ڰ���ø����, ����ˮ��ø��Ҫ�������̼���ֽ�ת��(�����ࡢ���ۡ���ά�ص�), ������-��������ø(��G)����ά��ˮ��ø(CBH)����-����������������ø(NAG)��; ����ø��Ҫ��������л�̼�����(��ľ����)�����ֽ��������ֳ�����̵�, �����������ø(PHO)����������ø(PEO)��(Luo et al., 2019), ���, ø���ԵĸߵͿɱ��������������������Լ������ֹ���ˮƽ֮��Ĺ�ϵ.���о���ʾ, ����ø�������������ֺ������������(Burns et al., 2013; Das & Mondal, 2019)�����������(Li et al., 2015; Feng et al., 2019).ͬʱ, �����������Ч��Ҳ��Ӱ���л�̼�͵��ķֽ�ø����, ʹ������P�����л��ʱ��ֳ���ͬ�ķֽ�����(Kang et al., 2009; Zhou et al., 2020).����, ����ø������������ʪ�ȼ�pH�������й�(��ΰΰ��, 2019; Zhou et al., 2020), ˵������ø����������ͷ��������صĸ������й�.��Ȼ��������ø�����������л�̼����ֺ�����Ĺ�ϵ�ѱ��㷺��ע, �����ڴ����Ǹ�ɽ��Ȼ�����������л�̼����������������ø����������ʶ�������. ...
Phenol oxidase, peroxidase and organic matter dynamics of soil 1 2010
... ɭ�������л�̼�����������Ƿ�ӳ����������ɭ�ֽ���״������Ҫָ��, ֱ�ӻ��ӵ�Ӱ�쵽ɭ������������������.�о���ʾ, ɭ������̼����ԼΪȫ������̼������73% (Post & Kwon, 2000), �ڵ���ȫ��̼ƽ�ⷽ��������Ҫ����.��������Ϊ½��ֲ����̬ϵͳ����Ҫ���(Kauppi et al., 2006), ��ά����������Ժ�����̼��ѭ���а�������Ҫ��ɫ(Yang et al., 2011).ɭ�ָ�������ı���ֲ��Ⱥ��ṹ��������ɼ����ܶȶ�Ӱ�쵽��������, ���������л�̼����������ֺ�����������Է����仯(Chandra et al., 2016; Feng et al., 2019).���о���Ϊ, ɭ��ֲ�������������ڽ��������л�̼�ֽ����ʳ�������ֲ�������������л�̼�Ĺ̳�����, ʹ���京����Խϵ�(Ohtsuka et al., 2010; Milcu et al., 2011), Ҳ���о�����, ����̼����������ɭ������������ֳ�����(Deng et al., 2013; ��ѩ��, 2016; Liu et al., 2020)���仯������(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)���Ƚ�����(Tang et al., 2009)�ı仯����, ˵��ɭ�����������̼��������Ӱ���Ϊ����.�����л��ʽ����ܵ����������Ӱ��(Sinsabaugh, 2010; Liu et al., 2020), ������������Ҳ���ܵ�����̼����Ч�Ե���Լ(Kooch et al., 2018; Dove et al., 2019).ɭ������ı�����������, ����������ɻ���֮�����仯, �����ı��������������(Zhang et al., 2017), �������Ҫ��������Դ�ڶ�ֲ�����ֽ���������������Լ�ֲ���ϵ������İ���ø��.����ø���л�̼���ֽ����ת����Ҫ����(Weintraub et al., 2007; Xu et al., 2017), ���������л�̼���Ļ�������������ø�Ŀֽ�����(Sinsabaugh et al., 2014; Mayer et al., 2020).�����л��ʿ������ܿ��ڰ���ø����, ����ˮ��ø��Ҫ�������̼���ֽ�ת��(�����ࡢ���ۡ���ά�ص�), ������-��������ø(��G)����ά��ˮ��ø(CBH)����-����������������ø(NAG)��; ����ø��Ҫ��������л�̼�����(��ľ����)�����ֽ��������ֳ�����̵�, �����������ø(PHO)����������ø(PEO)��(Luo et al., 2019), ���, ø���ԵĸߵͿɱ��������������������Լ������ֹ���ˮƽ֮��Ĺ�ϵ.���о���ʾ, ����ø�������������ֺ������������(Burns et al., 2013; Das & Mondal, 2019)�����������(Li et al., 2015; Feng et al., 2019).ͬʱ, �����������Ч��Ҳ��Ӱ���л�̼�͵��ķֽ�ø����, ʹ������P�����л��ʱ��ֳ���ͬ�ķֽ�����(Kang et al., 2009; Zhou et al., 2020).����, ����ø������������ʪ�ȼ�pH�������й�(��ΰΰ��, 2019; Zhou et al., 2020), ˵������ø����������ͷ��������صĸ������й�.��Ȼ��������ø�����������л�̼����ֺ�����Ĺ�ϵ�ѱ��㷺��ע, �����ڴ����Ǹ�ɽ��Ȼ�����������л�̼����������������ø����������ʶ�������. ...
Extracellular enzyme kinetics scale with resource availability 1 2014
... ���������л�̼���������������л��ʵ���Ҫ���, ��Ҫ��Դ���½���֦��Ҷ����ֳ�ʡ���ϵ�������Լ��������(Kalbitz et al., 2000; ���µ�, 2018).ɭ�ֲ�ͬ��Ȼ���»ָ���, ���ַ������������Իָ���������ɵIJ���, ���µر�������������������������ʪ�ȡ�������ɵȷ�����ͬ�̶ȱ仯, �Ӷ�Ӱ�쵽���������л�̼�������ı仯(��ѩ��, 2016; Hagedorn et al., 2019).���о���, DOC��LFOC��DON�����������������������½�����(p < 0.05), ����70-NSF��80-NSF�в��첻����(��3), �����浽ẽ���ɼԭʼ�ֽ�, ����������̼��������������(p < 0.05), ����������˹�����������б���(0-20 cm)��������̼���������о��������(������, 2009), ����ͬ���ȴ�(���ܵ�, 2019)�����ȴ�(��Ծ�µ�, 2013)ɭ��������о����, ˵����ͬ��ʪ������ֲ���ַֽṹ�����������л�̼����ֺ�����Ӱ����ֳ���ͬ�ı仯����(Li et al., 2014; Taylor et al., 2019).����ԭ��, һ��������ɭ�ָ�������, ����̼������������(Mylliemngap et al., 2016), ����Ҷ�ֵĿ����ʸ�����Ҷ��(C?t�� et al., 2000); ��һ�������ڱ��������ܶ�����������, �������ž۶ȡ��ṹϵ�����ܿ�϶��ë�ܿ�϶������������������ֽ�������(��ѧ�µ�, 2003), �����л�̼����ȱ�������ž���������Ա���, �������������л�̼��DON��������.����, ẽ���ɼԭʼ�ֵ���������̼����ֺ���������Ȼ������(��3), ��������ַ���Ҫ���ֹ���(��Ҷ+��Ҷ����)��һ������, Դ�ڸ������������ֵ���Ҷ�ں���һ�����ڷֽ�(����÷��, 2015), ���ͷŵ����ֱ�������ֽ�����, �������������л�̼��������.��Ҫ��������Ӱ�����ض����������л�̼������������, �����һ�������о�.��ط�������, ��ͬ��Ȼ����������ֵر�������SOC��TN����������л�̼����ֺ�����DON�������ʼ���������ع�ϵ(p < 0.01), ˵�����������л�̼�͵�����������SOC��TN����֮���ϵ��Ϊ����, ���������о�(�����, 2016; ������, 2017) һ��, ˵����Ȼ�����������������ǵ�����̼�ֵ̳���Ҫ����(Li et al., 2012; Smal et al., 2019). ...
Changes in the stocks of soil organic carbon, total nitrogen and phosphorus following afforestation of post-arable soils: a chronosequence study 1 2019
... ɭ�������л�̼�����������Ƿ�ӳ����������ɭ�ֽ���״������Ҫָ��, ֱ�ӻ��ӵ�Ӱ�쵽ɭ������������������.�о���ʾ, ɭ������̼����ԼΪȫ������̼������73% (Post & Kwon, 2000), �ڵ���ȫ��̼ƽ�ⷽ��������Ҫ����.��������Ϊ½��ֲ����̬ϵͳ����Ҫ���(Kauppi et al., 2006), ��ά����������Ժ�����̼��ѭ���а�������Ҫ��ɫ(Yang et al., 2011).ɭ�ָ�������ı���ֲ��Ⱥ��ṹ��������ɼ����ܶȶ�Ӱ�쵽��������, ���������л�̼����������ֺ�����������Է����仯(Chandra et al., 2016; Feng et al., 2019).���о���Ϊ, ɭ��ֲ�������������ڽ��������л�̼�ֽ����ʳ�������ֲ�������������л�̼�Ĺ̳�����, ʹ���京����Խϵ�(Ohtsuka et al., 2010; Milcu et al., 2011), Ҳ���о�����, ����̼����������ɭ������������ֳ�����(Deng et al., 2013; ��ѩ��, 2016; Liu et al., 2020)���仯������(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)���Ƚ�����(Tang et al., 2009)�ı仯����, ˵��ɭ�����������̼��������Ӱ���Ϊ����.�����л��ʽ����ܵ����������Ӱ��(Sinsabaugh, 2010; Liu et al., 2020), ������������Ҳ���ܵ�����̼����Ч�Ե���Լ(Kooch et al., 2018; Dove et al., 2019).ɭ������ı�����������, ����������ɻ���֮�����仯, �����ı��������������(Zhang et al., 2017), �������Ҫ��������Դ�ڶ�ֲ�����ֽ���������������Լ�ֲ���ϵ������İ���ø��.����ø���л�̼���ֽ����ת����Ҫ����(Weintraub et al., 2007; Xu et al., 2017), ���������л�̼���Ļ�������������ø�Ŀֽ�����(Sinsabaugh et al., 2014; Mayer et al., 2020).�����л��ʿ������ܿ��ڰ���ø����, ����ˮ��ø��Ҫ�������̼���ֽ�ת��(�����ࡢ���ۡ���ά�ص�), ������-��������ø(��G)����ά��ˮ��ø(CBH)����-����������������ø(NAG)��; ����ø��Ҫ��������л�̼�����(��ľ����)�����ֽ��������ֳ�����̵�, �����������ø(PHO)����������ø(PEO)��(Luo et al., 2019), ���, ø���ԵĸߵͿɱ��������������������Լ������ֹ���ˮƽ֮��Ĺ�ϵ.���о���ʾ, ����ø�������������ֺ������������(Burns et al., 2013; Das & Mondal, 2019)�����������(Li et al., 2015; Feng et al., 2019).ͬʱ, �����������Ч��Ҳ��Ӱ���л�̼�͵��ķֽ�ø����, ʹ������P�����л��ʱ��ֳ���ͬ�ķֽ�����(Kang et al., 2009; Zhou et al., 2020).����, ����ø������������ʪ�ȼ�pH�������й�(��ΰΰ��, 2019; Zhou et al., 2020), ˵������ø����������ͷ��������صĸ������й�.��Ȼ��������ø�����������л�̼����ֺ�����Ĺ�ϵ�ѱ��㷺��ע, �����ڴ����Ǹ�ɽ��Ȼ�����������л�̼����������������ø����������ʶ�������. ...
Soil carbon fluxes and stocks in a Great Lakes forest chronosequence 1 2009
The effects of tree rhizodeposition on soil exoenzyme activity, dissolved organic carbon, and nutrient availability in a subalpine forest ecosystem 1 2007
... ɭ�������л�̼�����������Ƿ�ӳ����������ɭ�ֽ���״������Ҫָ��, ֱ�ӻ��ӵ�Ӱ�쵽ɭ������������������.�о���ʾ, ɭ������̼����ԼΪȫ������̼������73% (Post & Kwon, 2000), �ڵ���ȫ��̼ƽ�ⷽ��������Ҫ����.��������Ϊ½��ֲ����̬ϵͳ����Ҫ���(Kauppi et al., 2006), ��ά����������Ժ�����̼��ѭ���а�������Ҫ��ɫ(Yang et al., 2011).ɭ�ָ�������ı���ֲ��Ⱥ��ṹ��������ɼ����ܶȶ�Ӱ�쵽��������, ���������л�̼����������ֺ�����������Է����仯(Chandra et al., 2016; Feng et al., 2019).���о���Ϊ, ɭ��ֲ�������������ڽ��������л�̼�ֽ����ʳ�������ֲ�������������л�̼�Ĺ̳�����, ʹ���京����Խϵ�(Ohtsuka et al., 2010; Milcu et al., 2011), Ҳ���о�����, ����̼����������ɭ������������ֳ�����(Deng et al., 2013; ��ѩ��, 2016; Liu et al., 2020)���仯������(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)���Ƚ�����(Tang et al., 2009)�ı仯����, ˵��ɭ�����������̼��������Ӱ���Ϊ����.�����л��ʽ����ܵ����������Ӱ��(Sinsabaugh, 2010; Liu et al., 2020), ������������Ҳ���ܵ�����̼����Ч�Ե���Լ(Kooch et al., 2018; Dove et al., 2019).ɭ������ı�����������, ����������ɻ���֮�����仯, �����ı��������������(Zhang et al., 2017), �������Ҫ��������Դ�ڶ�ֲ�����ֽ���������������Լ�ֲ���ϵ������İ���ø��.����ø���л�̼���ֽ����ת����Ҫ����(Weintraub et al., 2007; Xu et al., 2017), ���������л�̼���Ļ�������������ø�Ŀֽ�����(Sinsabaugh et al., 2014; Mayer et al., 2020).�����л��ʿ������ܿ��ڰ���ø����, ����ˮ��ø��Ҫ�������̼���ֽ�ת��(�����ࡢ���ۡ���ά�ص�), ������-��������ø(��G)����ά��ˮ��ø(CBH)����-����������������ø(NAG)��; ����ø��Ҫ��������л�̼�����(��ľ����)�����ֽ��������ֳ�����̵�, �����������ø(PHO)����������ø(PEO)��(Luo et al., 2019), ���, ø���ԵĸߵͿɱ��������������������Լ������ֹ���ˮƽ֮��Ĺ�ϵ.���о���ʾ, ����ø�������������ֺ������������(Burns et al., 2013; Das & Mondal, 2019)�����������(Li et al., 2015; Feng et al., 2019).ͬʱ, �����������Ч��Ҳ��Ӱ���л�̼�͵��ķֽ�ø����, ʹ������P�����л��ʱ��ֳ���ͬ�ķֽ�����(Kang et al., 2009; Zhou et al., 2020).����, ����ø������������ʪ�ȼ�pH�������й�(��ΰΰ��, 2019; Zhou et al., 2020), ˵������ø����������ͷ��������صĸ������й�.��Ȼ��������ø�����������л�̼����ֺ�����Ĺ�ϵ�ѱ��㷺��ע, �����ڴ����Ǹ�ɽ��Ȼ�����������л�̼����������������ø����������ʶ�������. ...
... ɭ�������л�̼�����������Ƿ�ӳ����������ɭ�ֽ���״������Ҫָ��, ֱ�ӻ��ӵ�Ӱ�쵽ɭ������������������.�о���ʾ, ɭ������̼����ԼΪȫ������̼������73% (Post & Kwon, 2000), �ڵ���ȫ��̼ƽ�ⷽ��������Ҫ����.��������Ϊ½��ֲ����̬ϵͳ����Ҫ���(Kauppi et al., 2006), ��ά����������Ժ�����̼��ѭ���а�������Ҫ��ɫ(Yang et al., 2011).ɭ�ָ�������ı���ֲ��Ⱥ��ṹ��������ɼ����ܶȶ�Ӱ�쵽��������, ���������л�̼����������ֺ�����������Է����仯(Chandra et al., 2016; Feng et al., 2019).���о���Ϊ, ɭ��ֲ�������������ڽ��������л�̼�ֽ����ʳ�������ֲ�������������л�̼�Ĺ̳�����, ʹ���京����Խϵ�(Ohtsuka et al., 2010; Milcu et al., 2011), Ҳ���о�����, ����̼����������ɭ������������ֳ�����(Deng et al., 2013; ��ѩ��, 2016; Liu et al., 2020)���仯������(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)���Ƚ�����(Tang et al., 2009)�ı仯����, ˵��ɭ�����������̼��������Ӱ���Ϊ����.�����л��ʽ����ܵ����������Ӱ��(Sinsabaugh, 2010; Liu et al., 2020), ������������Ҳ���ܵ�����̼����Ч�Ե���Լ(Kooch et al., 2018; Dove et al., 2019).ɭ������ı�����������, ����������ɻ���֮�����仯, �����ı��������������(Zhang et al., 2017), �������Ҫ��������Դ�ڶ�ֲ�����ֽ���������������Լ�ֲ���ϵ������İ���ø��.����ø���л�̼���ֽ����ת����Ҫ����(Weintraub et al., 2007; Xu et al., 2017), ���������л�̼���Ļ�������������ø�Ŀֽ�����(Sinsabaugh et al., 2014; Mayer et al., 2020).�����л��ʿ������ܿ��ڰ���ø����, ����ˮ��ø��Ҫ�������̼���ֽ�ת��(�����ࡢ���ۡ���ά�ص�), ������-��������ø(��G)����ά��ˮ��ø(CBH)����-����������������ø(NAG)��; ����ø��Ҫ��������л�̼�����(��ľ����)�����ֽ��������ֳ�����̵�, �����������ø(PHO)����������ø(PEO)��(Luo et al., 2019), ���, ø���ԵĸߵͿɱ��������������������Լ������ֹ���ˮƽ֮��Ĺ�ϵ.���о���ʾ, ����ø�������������ֺ������������(Burns et al., 2013; Das & Mondal, 2019)�����������(Li et al., 2015; Feng et al., 2019).ͬʱ, �����������Ч��Ҳ��Ӱ���л�̼�͵��ķֽ�ø����, ʹ������P�����л��ʱ��ֳ���ͬ�ķֽ�����(Kang et al., 2009; Zhou et al., 2020).����, ����ø������������ʪ�ȼ�pH�������й�(��ΰΰ��, 2019; Zhou et al., 2020), ˵������ø����������ͷ��������صĸ������й�.��Ȼ��������ø�����������л�̼����ֺ�����Ĺ�ϵ�ѱ��㷺��ע, �����ڴ����Ǹ�ɽ��Ȼ�����������л�̼����������������ø����������ʶ�������. ...
��Ȼ������ֲ�����������������ܶ����ԵĹ�ϵ 1 2014
... ɭ�������л�̼�����������Ƿ�ӳ����������ɭ�ֽ���״������Ҫָ��, ֱ�ӻ��ӵ�Ӱ�쵽ɭ������������������.�о���ʾ, ɭ������̼����ԼΪȫ������̼������73% (Post & Kwon, 2000), �ڵ���ȫ��̼ƽ�ⷽ��������Ҫ����.��������Ϊ½��ֲ����̬ϵͳ����Ҫ���(Kauppi et al., 2006), ��ά����������Ժ�����̼��ѭ���а�������Ҫ��ɫ(Yang et al., 2011).ɭ�ָ�������ı���ֲ��Ⱥ��ṹ��������ɼ����ܶȶ�Ӱ�쵽��������, ���������л�̼����������ֺ�����������Է����仯(Chandra et al., 2016; Feng et al., 2019).���о���Ϊ, ɭ��ֲ�������������ڽ��������л�̼�ֽ����ʳ�������ֲ�������������л�̼�Ĺ̳�����, ʹ���京����Խϵ�(Ohtsuka et al., 2010; Milcu et al., 2011), Ҳ���о�����, ����̼����������ɭ������������ֳ�����(Deng et al., 2013; ��ѩ��, 2016; Liu et al., 2020)���仯������(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)���Ƚ�����(Tang et al., 2009)�ı仯����, ˵��ɭ�����������̼��������Ӱ���Ϊ����.�����л��ʽ����ܵ����������Ӱ��(Sinsabaugh, 2010; Liu et al., 2020), ������������Ҳ���ܵ�����̼����Ч�Ե���Լ(Kooch et al., 2018; Dove et al., 2019).ɭ������ı�����������, ����������ɻ���֮�����仯, �����ı��������������(Zhang et al., 2017), �������Ҫ��������Դ�ڶ�ֲ�����ֽ���������������Լ�ֲ���ϵ������İ���ø��.����ø���л�̼���ֽ����ת����Ҫ����(Weintraub et al., 2007; Xu et al., 2017), ���������л�̼���Ļ�������������ø�Ŀֽ�����(Sinsabaugh et al., 2014; Mayer et al., 2020).�����л��ʿ������ܿ��ڰ���ø����, ����ˮ��ø��Ҫ�������̼���ֽ�ת��(�����ࡢ���ۡ���ά�ص�), ������-��������ø(��G)����ά��ˮ��ø(CBH)����-����������������ø(NAG)��; ����ø��Ҫ��������л�̼�����(��ľ����)�����ֽ��������ֳ�����̵�, �����������ø(PHO)����������ø(PEO)��(Luo et al., 2019), ���, ø���ԵĸߵͿɱ��������������������Լ������ֹ���ˮƽ֮��Ĺ�ϵ.���о���ʾ, ����ø�������������ֺ������������(Burns et al., 2013; Das & Mondal, 2019)�����������(Li et al., 2015; Feng et al., 2019).ͬʱ, �����������Ч��Ҳ��Ӱ���л�̼�͵��ķֽ�ø����, ʹ������P�����л��ʱ��ֳ���ͬ�ķֽ�����(Kang et al., 2009; Zhou et al., 2020).����, ����ø������������ʪ�ȼ�pH�������й�(��ΰΰ��, 2019; Zhou et al., 2020), ˵������ø����������ͷ��������صĸ������й�.��Ȼ��������ø�����������л�̼����ֺ�����Ĺ�ϵ�ѱ��㷺��ע, �����ڴ����Ǹ�ɽ��Ȼ�����������л�̼����������������ø����������ʶ�������. ...
Impacts of conversion from secondary forests to larch plantations on the structure and function of microbial communities 2017
... ɭ�������л�̼�����������Ƿ�ӳ����������ɭ�ֽ���״������Ҫָ��, ֱ�ӻ��ӵ�Ӱ�쵽ɭ������������������.�о���ʾ, ɭ������̼����ԼΪȫ������̼������73% (Post & Kwon, 2000), �ڵ���ȫ��̼ƽ�ⷽ��������Ҫ����.��������Ϊ½��ֲ����̬ϵͳ����Ҫ���(Kauppi et al., 2006), ��ά����������Ժ�����̼��ѭ���а�������Ҫ��ɫ(Yang et al., 2011).ɭ�ָ�������ı���ֲ��Ⱥ��ṹ��������ɼ����ܶȶ�Ӱ�쵽��������, ���������л�̼����������ֺ�����������Է����仯(Chandra et al., 2016; Feng et al., 2019).���о���Ϊ, ɭ��ֲ�������������ڽ��������л�̼�ֽ����ʳ�������ֲ�������������л�̼�Ĺ̳�����, ʹ���京����Խϵ�(Ohtsuka et al., 2010; Milcu et al., 2011), Ҳ���о�����, ����̼����������ɭ������������ֳ�����(Deng et al., 2013; ��ѩ��, 2016; Liu et al., 2020)���仯������(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)���Ƚ�����(Tang et al., 2009)�ı仯����, ˵��ɭ�����������̼��������Ӱ���Ϊ����.�����л��ʽ����ܵ����������Ӱ��(Sinsabaugh, 2010; Liu et al., 2020), ������������Ҳ���ܵ�����̼����Ч�Ե���Լ(Kooch et al., 2018; Dove et al., 2019).ɭ������ı�����������, ����������ɻ���֮�����仯, �����ı��������������(Zhang et al., 2017), �������Ҫ��������Դ�ڶ�ֲ�����ֽ���������������Լ�ֲ���ϵ������İ���ø��.����ø���л�̼���ֽ����ת����Ҫ����(Weintraub et al., 2007; Xu et al., 2017), ���������л�̼���Ļ�������������ø�Ŀֽ�����(Sinsabaugh et al., 2014; Mayer et al., 2020).�����л��ʿ������ܿ��ڰ���ø����, ����ˮ��ø��Ҫ�������̼���ֽ�ת��(�����ࡢ���ۡ���ά�ص�), ������-��������ø(��G)����ά��ˮ��ø(CBH)����-����������������ø(NAG)��; ����ø��Ҫ��������л�̼�����(��ľ����)�����ֽ��������ֳ�����̵�, �����������ø(PHO)����������ø(PEO)��(Luo et al., 2019), ���, ø���ԵĸߵͿɱ��������������������Լ������ֹ���ˮƽ֮��Ĺ�ϵ.���о���ʾ, ����ø�������������ֺ������������(Burns et al., 2013; Das & Mondal, 2019)�����������(Li et al., 2015; Feng et al., 2019).ͬʱ, �����������Ч��Ҳ��Ӱ���л�̼�͵��ķֽ�ø����, ʹ������P�����л��ʱ��ֳ���ͬ�ķֽ�����(Kang et al., 2009; Zhou et al., 2020).����, ����ø������������ʪ�ȼ�pH�������й�(��ΰΰ��, 2019; Zhou et al., 2020), ˵������ø����������ͷ��������صĸ������й�.��Ȼ��������ø�����������л�̼����ֺ�����Ĺ�ϵ�ѱ��㷺��ע, �����ڴ����Ǹ�ɽ��Ȼ�����������л�̼����������������ø����������ʶ�������. ...
�����Ǹ�ɽ������Ҫɭ�����͵�������ɼ���̬ 1 2019
... ɭ�������л�̼�����������Ƿ�ӳ����������ɭ�ֽ���״������Ҫָ��, ֱ�ӻ��ӵ�Ӱ�쵽ɭ������������������.�о���ʾ, ɭ������̼����ԼΪȫ������̼������73% (Post & Kwon, 2000), �ڵ���ȫ��̼ƽ�ⷽ��������Ҫ����.��������Ϊ½��ֲ����̬ϵͳ����Ҫ���(Kauppi et al., 2006), ��ά����������Ժ�����̼��ѭ���а�������Ҫ��ɫ(Yang et al., 2011).ɭ�ָ�������ı���ֲ��Ⱥ��ṹ��������ɼ����ܶȶ�Ӱ�쵽��������, ���������л�̼����������ֺ�����������Է����仯(Chandra et al., 2016; Feng et al., 2019).���о���Ϊ, ɭ��ֲ�������������ڽ��������л�̼�ֽ����ʳ�������ֲ�������������л�̼�Ĺ̳�����, ʹ���京����Խϵ�(Ohtsuka et al., 2010; Milcu et al., 2011), Ҳ���о�����, ����̼����������ɭ������������ֳ�����(Deng et al., 2013; ��ѩ��, 2016; Liu et al., 2020)���仯������(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)���Ƚ�����(Tang et al., 2009)�ı仯����, ˵��ɭ�����������̼��������Ӱ���Ϊ����.�����л��ʽ����ܵ����������Ӱ��(Sinsabaugh, 2010; Liu et al., 2020), ������������Ҳ���ܵ�����̼����Ч�Ե���Լ(Kooch et al., 2018; Dove et al., 2019).ɭ������ı�����������, ����������ɻ���֮�����仯, �����ı��������������(Zhang et al., 2017), �������Ҫ��������Դ�ڶ�ֲ�����ֽ���������������Լ�ֲ���ϵ������İ���ø��.����ø���л�̼���ֽ����ת����Ҫ����(Weintraub et al., 2007; Xu et al., 2017), ���������л�̼���Ļ�������������ø�Ŀֽ�����(Sinsabaugh et al., 2014; Mayer et al., 2020).�����л��ʿ������ܿ��ڰ���ø����, ����ˮ��ø��Ҫ�������̼���ֽ�ת��(�����ࡢ���ۡ���ά�ص�), ������-��������ø(��G)����ά��ˮ��ø(CBH)����-����������������ø(NAG)��; ����ø��Ҫ��������л�̼�����(��ľ����)�����ֽ��������ֳ�����̵�, �����������ø(PHO)����������ø(PEO)��(Luo et al., 2019), ���, ø���ԵĸߵͿɱ��������������������Լ������ֹ���ˮƽ֮��Ĺ�ϵ.���о���ʾ, ����ø�������������ֺ������������(Burns et al., 2013; Das & Mondal, 2019)�����������(Li et al., 2015; Feng et al., 2019).ͬʱ, �����������Ч��Ҳ��Ӱ���л�̼�͵��ķֽ�ø����, ʹ������P�����л��ʱ��ֳ���ͬ�ķֽ�����(Kang et al., 2009; Zhou et al., 2020).����, ����ø������������ʪ�ȼ�pH�������й�(��ΰΰ��, 2019; Zhou et al., 2020), ˵������ø����������ͷ��������صĸ������й�.��Ȼ��������ø�����������л�̼����ֺ�����Ĺ�ϵ�ѱ��㷺��ע, �����ڴ����Ǹ�ɽ��Ȼ�����������л�̼����������������ø����������ʶ�������. ...
Microarray-based analysis of changes in diversity of microbial genes involved in organic carbon decomposition following land use/cover changes 1 2007
... ɭ�������л�̼�����������Ƿ�ӳ����������ɭ�ֽ���״������Ҫָ��, ֱ�ӻ��ӵ�Ӱ�쵽ɭ������������������.�о���ʾ, ɭ������̼����ԼΪȫ������̼������73% (Post & Kwon, 2000), �ڵ���ȫ��̼ƽ�ⷽ��������Ҫ����.��������Ϊ½��ֲ����̬ϵͳ����Ҫ���(Kauppi et al., 2006), ��ά����������Ժ�����̼��ѭ���а�������Ҫ��ɫ(Yang et al., 2011).ɭ�ָ�������ı���ֲ��Ⱥ��ṹ��������ɼ����ܶȶ�Ӱ�쵽��������, ���������л�̼����������ֺ�����������Է����仯(Chandra et al., 2016; Feng et al., 2019).���о���Ϊ, ɭ��ֲ�������������ڽ��������л�̼�ֽ����ʳ�������ֲ�������������л�̼�Ĺ̳�����, ʹ���京����Խϵ�(Ohtsuka et al., 2010; Milcu et al., 2011), Ҳ���о�����, ����̼����������ɭ������������ֳ�����(Deng et al., 2013; ��ѩ��, 2016; Liu et al., 2020)���仯������(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)���Ƚ�����(Tang et al., 2009)�ı仯����, ˵��ɭ�����������̼��������Ӱ���Ϊ����.�����л��ʽ����ܵ����������Ӱ��(Sinsabaugh, 2010; Liu et al., 2020), ������������Ҳ���ܵ�����̼����Ч�Ե���Լ(Kooch et al., 2018; Dove et al., 2019).ɭ������ı�����������, ����������ɻ���֮�����仯, �����ı��������������(Zhang et al., 2017), �������Ҫ��������Դ�ڶ�ֲ�����ֽ���������������Լ�ֲ���ϵ������İ���ø��.����ø���л�̼���ֽ����ת����Ҫ����(Weintraub et al., 2007; Xu et al., 2017), ���������л�̼���Ļ�������������ø�Ŀֽ�����(Sinsabaugh et al., 2014; Mayer et al., 2020).�����л��ʿ������ܿ��ڰ���ø����, ����ˮ��ø��Ҫ�������̼���ֽ�ת��(�����ࡢ���ۡ���ά�ص�), ������-��������ø(��G)����ά��ˮ��ø(CBH)����-����������������ø(NAG)��; ����ø��Ҫ��������л�̼�����(��ľ����)�����ֽ��������ֳ�����̵�, �����������ø(PHO)����������ø(PEO)��(Luo et al., 2019), ���, ø���ԵĸߵͿɱ��������������������Լ������ֹ���ˮƽ֮��Ĺ�ϵ.���о���ʾ, ����ø�������������ֺ������������(Burns et al., 2013; Das & Mondal, 2019)�����������(Li et al., 2015; Feng et al., 2019).ͬʱ, �����������Ч��Ҳ��Ӱ���л�̼�͵��ķֽ�ø����, ʹ������P�����л��ʱ��ֳ���ͬ�ķֽ�����(Kang et al., 2009; Zhou et al., 2020).����, ����ø������������ʪ�ȼ�pH�������й�(��ΰΰ��, 2019; Zhou et al., 2020), ˵������ø����������ͷ��������صĸ������й�.��Ȼ��������ø�����������л�̼����ֺ�����Ĺ�ϵ�ѱ��㷺��ע, �����ڴ����Ǹ�ɽ��Ȼ�����������л�̼����������������ø����������ʶ�������. ...
�����������л�̼����˫�����ȴ�ɭ��Ⱥ���������Ӧ 1 2019
... ���������л�̼���������������л��ʵ���Ҫ���, ��Ҫ��Դ���½���֦��Ҷ����ֳ�ʡ���ϵ�������Լ��������(Kalbitz et al., 2000; ���µ�, 2018).ɭ�ֲ�ͬ��Ȼ���»ָ���, ���ַ������������Իָ���������ɵIJ���, ���µر�������������������������ʪ�ȡ�������ɵȷ�����ͬ�̶ȱ仯, �Ӷ�Ӱ�쵽���������л�̼�������ı仯(��ѩ��, 2016; Hagedorn et al., 2019).���о���, DOC��LFOC��DON�����������������������½�����(p < 0.05), ����70-NSF��80-NSF�в��첻����(��3), �����浽ẽ���ɼԭʼ�ֽ�, ����������̼��������������(p < 0.05), ����������˹�����������б���(0-20 cm)��������̼���������о��������(������, 2009), ����ͬ���ȴ�(���ܵ�, 2019)�����ȴ�(��Ծ�µ�, 2013)ɭ��������о����, ˵����ͬ��ʪ������ֲ���ַֽṹ�����������л�̼����ֺ�����Ӱ����ֳ���ͬ�ı仯����(Li et al., 2014; Taylor et al., 2019).����ԭ��, һ��������ɭ�ָ�������, ����̼������������(Mylliemngap et al., 2016), ����Ҷ�ֵĿ����ʸ�����Ҷ��(C?t�� et al., 2000); ��һ�������ڱ��������ܶ�����������, �������ž۶ȡ��ṹϵ�����ܿ�϶��ë�ܿ�϶������������������ֽ�������(��ѧ�µ�, 2003), �����л�̼����ȱ�������ž���������Ա���, �������������л�̼��DON��������.����, ẽ���ɼԭʼ�ֵ���������̼����ֺ���������Ȼ������(��3), ��������ַ���Ҫ���ֹ���(��Ҷ+��Ҷ����)��һ������, Դ�ڸ������������ֵ���Ҷ�ں���һ�����ڷֽ�(����÷��, 2015), ���ͷŵ����ֱ�������ֽ�����, �������������л�̼��������.��Ҫ��������Ӱ�����ض����������л�̼������������, �����һ�������о�.��ط�������, ��ͬ��Ȼ����������ֵر�������SOC��TN����������л�̼����ֺ�����DON�������ʼ���������ع�ϵ(p < 0.01), ˵�����������л�̼�͵�����������SOC��TN����֮���ϵ��Ϊ����, ���������о�(�����, 2016; ������, 2017) һ��, ˵����Ȼ�����������������ǵ�����̼�ֵ̳���Ҫ����(Li et al., 2012; Smal et al., 2019). ...
�����������л�̼����˫�����ȴ�ɭ��Ⱥ���������Ӧ 1 2019
... ���������л�̼���������������л��ʵ���Ҫ���, ��Ҫ��Դ���½���֦��Ҷ����ֳ�ʡ���ϵ�������Լ��������(Kalbitz et al., 2000; ���µ�, 2018).ɭ�ֲ�ͬ��Ȼ���»ָ���, ���ַ������������Իָ���������ɵIJ���, ���µر�������������������������ʪ�ȡ�������ɵȷ�����ͬ�̶ȱ仯, �Ӷ�Ӱ�쵽���������л�̼�������ı仯(��ѩ��, 2016; Hagedorn et al., 2019).���о���, DOC��LFOC��DON�����������������������½�����(p < 0.05), ����70-NSF��80-NSF�в��첻����(��3), �����浽ẽ���ɼԭʼ�ֽ�, ����������̼��������������(p < 0.05), ����������˹�����������б���(0-20 cm)��������̼���������о��������(������, 2009), ����ͬ���ȴ�(���ܵ�, 2019)�����ȴ�(��Ծ�µ�, 2013)ɭ��������о����, ˵����ͬ��ʪ������ֲ���ַֽṹ�����������л�̼����ֺ�����Ӱ����ֳ���ͬ�ı仯����(Li et al., 2014; Taylor et al., 2019).����ԭ��, һ��������ɭ�ָ�������, ����̼������������(Mylliemngap et al., 2016), ����Ҷ�ֵĿ����ʸ�����Ҷ��(C?t�� et al., 2000); ��һ�������ڱ��������ܶ�����������, �������ž۶ȡ��ṹϵ�����ܿ�϶��ë�ܿ�϶������������������ֽ�������(��ѧ�µ�, 2003), �����л�̼����ȱ�������ž���������Ա���, �������������л�̼��DON��������.����, ẽ���ɼԭʼ�ֵ���������̼����ֺ���������Ȼ������(��3), ��������ַ���Ҫ���ֹ���(��Ҷ+��Ҷ����)��һ������, Դ�ڸ������������ֵ���Ҷ�ں���һ�����ڷֽ�(����÷��, 2015), ���ͷŵ����ֱ�������ֽ�����, �������������л�̼��������.��Ҫ��������Ӱ�����ض����������л�̼������������, �����һ�������о�.��ط�������, ��ͬ��Ȼ����������ֵر�������SOC��TN����������л�̼����ֺ�����DON�������ʼ���������ع�ϵ(p < 0.01), ˵�����������л�̼�͵�����������SOC��TN����֮���ϵ��Ϊ����, ���������о�(�����, 2016; ������, 2017) һ��, ˵����Ȼ�����������������ǵ�����̼�ֵ̳���Ҫ����(Li et al., 2012; Smal et al., 2019). ...
Soil extracellular enzyme activity and stoichiometry in China��s forests 1 2020
... ɭ�������л�̼�����������Ƿ�ӳ����������ɭ�ֽ���״������Ҫָ��, ֱ�ӻ��ӵ�Ӱ�쵽ɭ������������������.�о���ʾ, ɭ������̼����ԼΪȫ������̼������73% (Post & Kwon, 2000), �ڵ���ȫ��̼ƽ�ⷽ��������Ҫ����.��������Ϊ½��ֲ����̬ϵͳ����Ҫ���(Kauppi et al., 2006), ��ά����������Ժ�����̼��ѭ���а�������Ҫ��ɫ(Yang et al., 2011).ɭ�ָ�������ı���ֲ��Ⱥ��ṹ��������ɼ����ܶȶ�Ӱ�쵽��������, ���������л�̼����������ֺ�����������Է����仯(Chandra et al., 2016; Feng et al., 2019).���о���Ϊ, ɭ��ֲ�������������ڽ��������л�̼�ֽ����ʳ�������ֲ�������������л�̼�Ĺ̳�����, ʹ���京����Խϵ�(Ohtsuka et al., 2010; Milcu et al., 2011), Ҳ���о�����, ����̼����������ɭ������������ֳ�����(Deng et al., 2013; ��ѩ��, 2016; Liu et al., 2020)���仯������(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)���Ƚ�����(Tang et al., 2009)�ı仯����, ˵��ɭ�����������̼��������Ӱ���Ϊ����.�����л��ʽ����ܵ����������Ӱ��(Sinsabaugh, 2010; Liu et al., 2020), ������������Ҳ���ܵ�����̼����Ч�Ե���Լ(Kooch et al., 2018; Dove et al., 2019).ɭ������ı�����������, ����������ɻ���֮�����仯, �����ı��������������(Zhang et al., 2017), �������Ҫ��������Դ�ڶ�ֲ�����ֽ���������������Լ�ֲ���ϵ������İ���ø��.����ø���л�̼���ֽ����ת����Ҫ����(Weintraub et al., 2007; Xu et al., 2017), ���������л�̼���Ļ�������������ø�Ŀֽ�����(Sinsabaugh et al., 2014; Mayer et al., 2020).�����л��ʿ������ܿ��ڰ���ø����, ����ˮ��ø��Ҫ�������̼���ֽ�ת��(�����ࡢ���ۡ���ά�ص�), ������-��������ø(��G)����ά��ˮ��ø(CBH)����-����������������ø(NAG)��; ����ø��Ҫ��������л�̼�����(��ľ����)�����ֽ��������ֳ�����̵�, �����������ø(PHO)����������ø(PEO)��(Luo et al., 2019), ���, ø���ԵĸߵͿɱ��������������������Լ������ֹ���ˮƽ֮��Ĺ�ϵ.���о���ʾ, ����ø�������������ֺ������������(Burns et al., 2013; Das & Mondal, 2019)�����������(Li et al., 2015; Feng et al., 2019).ͬʱ, �����������Ч��Ҳ��Ӱ���л�̼�͵��ķֽ�ø����, ʹ������P�����л��ʱ��ֳ���ͬ�ķֽ�����(Kang et al., 2009; Zhou et al., 2020).����, ����ø������������ʪ�ȼ�pH�������й�(��ΰΰ��, 2019; Zhou et al., 2020), ˵������ø����������ͷ��������صĸ������й�.��Ȼ��������ø�����������л�̼����ֺ�����Ĺ�ϵ�ѱ��㷺��ע, �����ڴ����Ǹ�ɽ��Ȼ�����������л�̼����������������ø����������ʶ�������. ...
... ɭ�������л�̼�����������Ƿ�ӳ����������ɭ�ֽ���״������Ҫָ��, ֱ�ӻ��ӵ�Ӱ�쵽ɭ������������������.�о���ʾ, ɭ������̼����ԼΪȫ������̼������73% (Post & Kwon, 2000), �ڵ���ȫ��̼ƽ�ⷽ��������Ҫ����.��������Ϊ½��ֲ����̬ϵͳ����Ҫ���(Kauppi et al., 2006), ��ά����������Ժ�����̼��ѭ���а�������Ҫ��ɫ(Yang et al., 2011).ɭ�ָ�������ı���ֲ��Ⱥ��ṹ��������ɼ����ܶȶ�Ӱ�쵽��������, ���������л�̼����������ֺ�����������Է����仯(Chandra et al., 2016; Feng et al., 2019).���о���Ϊ, ɭ��ֲ�������������ڽ��������л�̼�ֽ����ʳ�������ֲ�������������л�̼�Ĺ̳�����, ʹ���京����Խϵ�(Ohtsuka et al., 2010; Milcu et al., 2011), Ҳ���о�����, ����̼����������ɭ������������ֳ�����(Deng et al., 2013; ��ѩ��, 2016; Liu et al., 2020)���仯������(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)���Ƚ�����(Tang et al., 2009)�ı仯����, ˵��ɭ�����������̼��������Ӱ���Ϊ����.�����л��ʽ����ܵ����������Ӱ��(Sinsabaugh, 2010; Liu et al., 2020), ������������Ҳ���ܵ�����̼����Ч�Ե���Լ(Kooch et al., 2018; Dove et al., 2019).ɭ������ı�����������, ����������ɻ���֮�����仯, �����ı��������������(Zhang et al., 2017), �������Ҫ��������Դ�ڶ�ֲ�����ֽ���������������Լ�ֲ���ϵ������İ���ø��.����ø���л�̼���ֽ����ת����Ҫ����(Weintraub et al., 2007; Xu et al., 2017), ���������л�̼���Ļ�������������ø�Ŀֽ�����(Sinsabaugh et al., 2014; Mayer et al., 2020).�����л��ʿ������ܿ��ڰ���ø����, ����ˮ��ø��Ҫ�������̼���ֽ�ת��(�����ࡢ���ۡ���ά�ص�), ������-��������ø(��G)����ά��ˮ��ø(CBH)����-����������������ø(NAG)��; ����ø��Ҫ��������л�̼�����(��ľ����)�����ֽ��������ֳ�����̵�, �����������ø(PHO)����������ø(PEO)��(Luo et al., 2019), ���, ø���ԵĸߵͿɱ��������������������Լ������ֹ���ˮƽ֮��Ĺ�ϵ.���о���ʾ, ����ø�������������ֺ������������(Burns et al., 2013; Das & Mondal, 2019)�����������(Li et al., 2015; Feng et al., 2019).ͬʱ, �����������Ч��Ҳ��Ӱ���л�̼�͵��ķֽ�ø����, ʹ������P�����л��ʱ��ֳ���ͬ�ķֽ�����(Kang et al., 2009; Zhou et al., 2020).����, ����ø������������ʪ�ȼ�pH�������й�(��ΰΰ��, 2019; Zhou et al., 2020), ˵������ø����������ͷ��������صĸ������й�.��Ȼ��������ø�����������л�̼����ֺ�����Ĺ�ϵ�ѱ��㷺��ע, �����ڴ����Ǹ�ɽ��Ȼ�����������л�̼����������������ø����������ʶ�������. ...
... ɭ�������л�̼�����������Ƿ�ӳ����������ɭ�ֽ���״������Ҫָ��, ֱ�ӻ��ӵ�Ӱ�쵽ɭ������������������.�о���ʾ, ɭ������̼����ԼΪȫ������̼������73% (Post & Kwon, 2000), �ڵ���ȫ��̼ƽ�ⷽ��������Ҫ����.��������Ϊ½��ֲ����̬ϵͳ����Ҫ���(Kauppi et al., 2006), ��ά����������Ժ�����̼��ѭ���а�������Ҫ��ɫ(Yang et al., 2011).ɭ�ָ�������ı���ֲ��Ⱥ��ṹ��������ɼ����ܶȶ�Ӱ�쵽��������, ���������л�̼����������ֺ�����������Է����仯(Chandra et al., 2016; Feng et al., 2019).���о���Ϊ, ɭ��ֲ�������������ڽ��������л�̼�ֽ����ʳ�������ֲ�������������л�̼�Ĺ̳�����, ʹ���京����Խϵ�(Ohtsuka et al., 2010; Milcu et al., 2011), Ҳ���о�����, ����̼����������ɭ������������ֳ�����(Deng et al., 2013; ��ѩ��, 2016; Liu et al., 2020)���仯������(Taylor et al., 2007; Schedlbauer & Kavanagh, 2008)���Ƚ�����(Tang et al., 2009)�ı仯����, ˵��ɭ�����������̼��������Ӱ���Ϊ����.�����л��ʽ����ܵ����������Ӱ��(Sinsabaugh, 2010; Liu et al., 2020), ������������Ҳ���ܵ�����̼����Ч�Ե���Լ(Kooch et al., 2018; Dove et al., 2019).ɭ������ı�����������, ����������ɻ���֮�����仯, �����ı��������������(Zhang et al., 2017), �������Ҫ��������Դ�ڶ�ֲ�����ֽ���������������Լ�ֲ���ϵ������İ���ø��.����ø���л�̼���ֽ����ת����Ҫ����(Weintraub et al., 2007; Xu et al., 2017), ���������л�̼���Ļ�������������ø�Ŀֽ�����(Sinsabaugh et al., 2014; Mayer et al., 2020).�����л��ʿ������ܿ��ڰ���ø����, ����ˮ��ø��Ҫ�������̼���ֽ�ת��(�����ࡢ���ۡ���ά�ص�), ������-��������ø(��G)����ά��ˮ��ø(CBH)����-����������������ø(NAG)��; ����ø��Ҫ��������л�̼�����(��ľ����)�����ֽ��������ֳ�����̵�, �����������ø(PHO)����������ø(PEO)��(Luo et al., 2019), ���, ø���ԵĸߵͿɱ��������������������Լ������ֹ���ˮƽ֮��Ĺ�ϵ.���о���ʾ, ����ø�������������ֺ������������(Burns et al., 2013; Das & Mondal, 2019)�����������(Li et al., 2015; Feng et al., 2019).ͬʱ, �����������Ч��Ҳ��Ӱ���л�̼�͵��ķֽ�ø����, ʹ������P�����л��ʱ��ֳ���ͬ�ķֽ�����(Kang et al., 2009; Zhou et al., 2020).����, ����ø������������ʪ�ȼ�pH�������й�(��ΰΰ��, 2019; Zhou et al., 2020), ˵������ø����������ͷ��������صĸ������й�.��Ȼ��������ø�����������л�̼����ֺ�����Ĺ�ϵ�ѱ��㷺��ע, �����ڴ����Ǹ�ɽ��Ȼ�����������л�̼����������������ø����������ʶ�������. ...
 ,1,*, ������3, ����ϼ1, ���Z4, ������2, ���Ƿ�1, ���1, ŷ����1
,1,*, ������3, ����ϼ1, ���Z4, ������2, ���Ƿ�1, ���1, ŷ����1
 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPT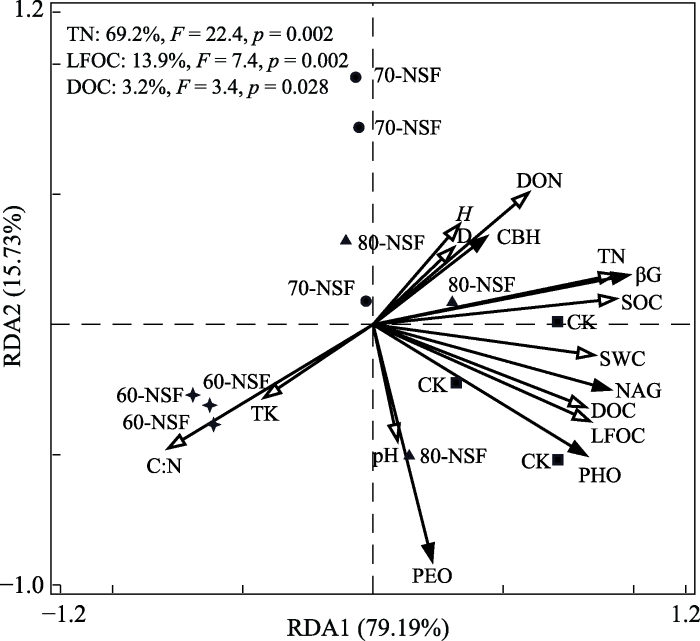 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPT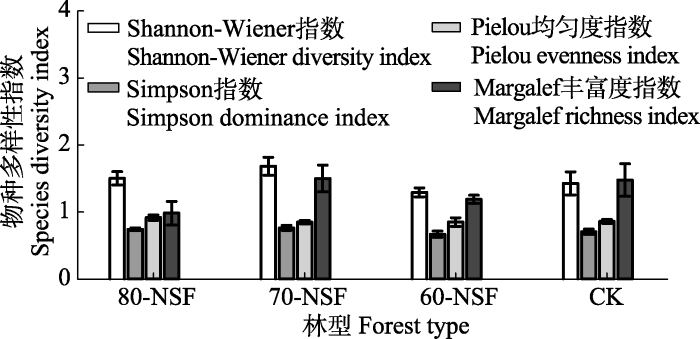 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}