Relationships of flower longevity with attractiveness traits and their effects on female fitness of alpine meadow plants on the Qinghai-Xizang Plateau, China
Yu-Xian WANG1, Meng HOU1, Yan-Yan XIE1, Zuo-Jun LIU2, Zhi-Gang ZHAO,,1,*, Ning-Na LU31School of Life Sciences, Lanzhou University, State Key Laboratory of Grassland Agro-Ecosystems, Lanzhou 730000 2School of Life Science and Engineering, Lanzhou University of Technology, Lanzhou 730050 3School of Life Sciences, Northwest Normal University, Lanzhou 730000
Supported by the National Key R&D Program of China(2017YFC0504801) the National Natural Science Foundation of China (31570229) the National Natural Science Foundation of China (31870411) the Key Research Program of Gansu(18YF1FA053)
Abstract Aims Flower longevity, the length of time that flowers remain open and functional, is an important floral trait that influences plant reproductive success. Prolonging floral longevity can provide reproductive assurance under unfavorable pollination environment, but may suffer some fitness costs. The theoretical models of flower longevity have suggested that there is a trade-off of resources allocation between the flower longevity and attractiveness traits. Flower longevity can adjust according to resource allocation. To understand the potential adaptive significance of flower longevity, it is necessary to disentangle the potential relationships between flower longevity and attractiveness traits (flower size and number) and their effects on female fitness of alpine meadow plants at the inter-species level on the Qinghai-Xizang Plateau. Methods In the alpine meadows of Qinghai-Xizang Plateau (2 900 m vs. 3 600 m), we assessed how the relationships between floral longevity and flower attractiveness traits of 11 species varied at the inter-species level under different pollination contexts. And we quantified the relative contribution of flower longevity and flower attractiveness traits to the female fitness of plants at the inter-species level at low and high habitats. Important findings Results show that there is a trade-off between flower longevity and flower number in both low and high altitude communities. And we found that both longer flower longevity and larger flower size increase the female fitness of plants in low-altitude communities, However, the female fitness of plants is only associated with flower longevity in high-altitude communities. Overall, our results suggest that flower longevity is more important for the female fitness in high-altitude plants than low-altitude plants. Keywords:Qinghai-Xizang Plateau;flower longevity;floral attractiveness traits;trade-off;reproductive success
PDF (1296KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 王玉贤, 侯盟, 谢言言, 刘左军, 赵志刚, 路宁娜. 青藏高原高寒草甸植物花寿命与花吸引特征的关系及其对雌性繁殖成功的影响. 植物生态学报, 2020, 44(9): 905-915. DOI: 10.17521/cjpe.2020.0130 WANG Yu-Xian, HOU Meng, XIE Yan-Yan, LIU Zuo-Jun, ZHAO Zhi-Gang, LU Ning-Na. Relationships of flower longevity with attractiveness traits and their effects on female fitness of alpine meadow plants on the Qinghai-Xizang Plateau, China. Chinese Journal of Plant Ecology, 2020, 44(9): 905-915. DOI: 10.17521/cjpe.2020.0130
本研究于2017年6-7月在兰州大学高寒草甸与湿地生态系统定位站的合作站(34.95° N, 102.88° E, 低海拔样地, 约2 900 m)与阿孜站(33.67° N, 101.87° E, 高海拔样地, 约3 600 m)进行。在两个站的高寒草甸的自然群落中, 分别选取一个50 m × 50 m的样地。
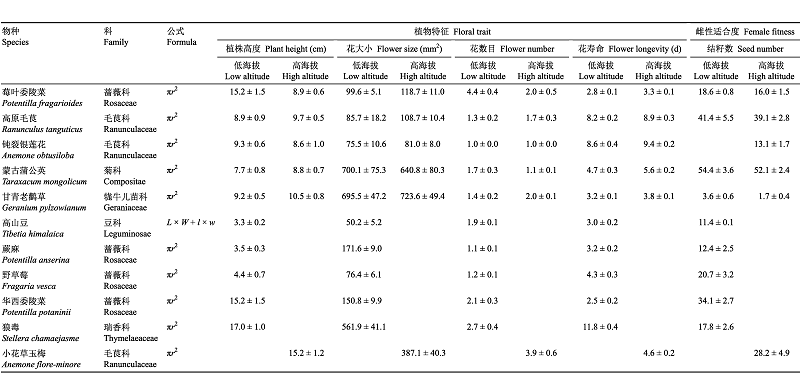
Table 1 Table 1Mean plant height, flower size, flower number, flower longevity and seeds number of 16 research populations at low (2 900 m) and high (3 600 m) altitudes (mean土SE)
对于具有圆形轮廓花的植物来说,r为花冠半径;对于豆科植物来说, L和W分别为旗瓣长和宽,和w分别为翼瓣长和宽。 For flowers with circular outine, r rssents the corolla radius. For the leguminous plants, L and W represent the length and width of banner, and I and w represent the length and width of wing, respetively.
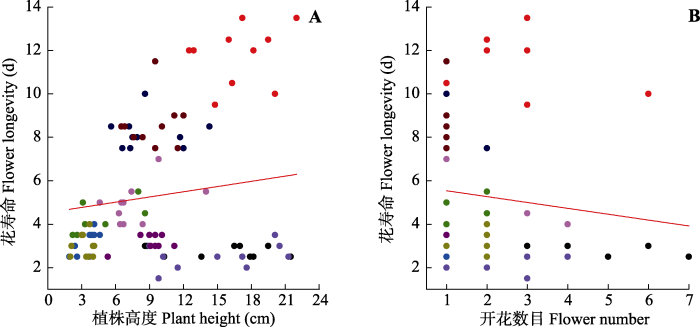
Table 2 Table 2Potential effects of plant height, flower size (log transform), and flower number on flower longevity of studied species in low (2 900 m) and high (3 600 m) altitude communities
海拔 Altitude
因素 Effect
估计值±标准误差 Estimate ± SE
t
p
低海拔 Low altitude
截距 Intercept
3.98 ± 1.69
2.353
0.023
植株高度 Plant height (cm)
0.07 ± 0.03
2.183
0.032
Log (花大小) log (flower size)
0.21 ± 0.28
0.751
0.454
开花数目 Flower number
-0.25 ± 0.11
-2.366
0.020
高海拔 High altitude
截距 Intercept
2.85 ± 1.77
1.609
0.123
植株高度 Plant height (cm)
0.06 ± 0.03
2.224
0.030
Log (花大小) log (flower size)
0.49 ± 0.24
2.038
0.046
开花数目 Flower number
-0.14 ± 0.07
-2.065
0.044
运用一般线性混合效应模型检验这些因素的显著性, 用粗体表示差异显著(p < 0.05)。 Significant differences are examined from general linear mixed effect models at p < 0.05 and indicated in bold.
Fig. 1Scatterplots of the relationships between flower longevity and plant height (A), and flower number (B) at low altitude plant communities. Data points represent individuals, and each colored point represents different species. Red line indicates the mean response across species.
Fig. 2Scatterplots of the relationships between flower longevity and plant height (A), log (flower size)(B), and flower number (C) at high altitude plant communities. Data points represent individuals, and each colored point represents different species. Red line indicates the mean response across species.
2.2 花寿命和花吸引特征对雌性适合度的相对贡献
在低海拔植物群落中, 结籽数在(3.6 ± 0.6)(甘青老鹳草)到(54.4 ± 3.6)(蒙古蒲公英)之间变化(表1), 平均结籽数为(24.6 ± 2.0)。结籽数与植物的花大小(z = 2.319, p = 0.020; 表3; 图3A)和花寿命(z = 2.213, p = 0.027; 表3; 图3B)均呈显著的正相关关系; 而结籽数与植物的开花数目(z = 0.101, p = 0.919; 表3)和植株高度(z = 0.856, p = 0.392; 表3)均没有显著的相关关系。
Table 3 Table 3Effects of plant height, flower size (log transform), flower longevity and flower number on female fitness (seeds number) of studied species in low (2 900 m) and high (3 600 m) communities
海拔 Altitude
因素 Effect
估计值±标准误差 Estimate ± SE
z
p
低海拔 Low altitude
截距 Intercept
1.62 ± 0.46
3.545
0.000
植株高度 Plant height (cm)
0.01 ± 0.01
0.856
0.392
Log (花大小) log (flower size)
0.18 ± 0.08
2.319
0.020
花寿命 Flower longevity (day)
0.07 ± 0.03
2.213
0.027
开花数目 Flower number
0.01 ± 0.03
0.101
0.919
高海拔 High altitude
截距 Intercept
1.89 ± 0.60
3.131
0.002
植株高度 Plant height (cm)
0.01 ± 0.01
1.094
0.274
Log (花大小) log (flower size) (mm2)
-0.03 ± 0.09
-0.340
0.734
花寿命 Flower longevity (day)
0.16 ± 0.05
3.380
0.001
开花数目 Flower number
-0.01 ± 0.03
-0.200
0.841
运用广义线性混合效应模型检验这些因素的显著性, 用粗体表示差异显著(p < 0.05)。 Significant differences are examined from generalized linear mixed effect models with Poisson distributions at p < 0.05 and indicated in bold.
Fig. 3Scatterplots of the relationships between female fitness (seed number) and log (flower size)(A), and flower longevity (B) at low altitude plant communities. Data points represent individuals, and each colored point represents different species. Red line indicates the mean response across species.
Fig. 4Relationships between seed set and flower longevity at high altitude plant communities. Data points represent individuals, and each colored point represents different species. Red line indicates the mean response across species.
Abdala-RobertsL, Parra-TablaV, NavarroJ ( 2007). Is floral longevity influenced by reproductive costs and pollination success in Cohniella ascendens (Orchidaceae)? Annals of Botany, 100, 1367-1371. DOI:10.1093/aob/mcm219URLPMID:17881335 [本文引用: 4] BACKGROUND AND AIMS: Although studies have shown that pollen addition and/or removal decreases floral longevity, less attention has been paid to the relationship between reproductive costs and floral longevity. In addition, the influence of reproductive costs on floral longevity responses to pollen addition and/or removal has not yet been evaluated. Here, the orchid Cohniella ascendens is used to answer the following questions. (a) Does experimental removal of flower buds in C. ascendens increase flower longevity? (b) Does pollen addition and/or removal decrease floral longevity, and does this response depend on plant reproductive resource status? METHODS: To study the effect of reproductive costs on floral longevity 21 plants were selected from which we removed 50 % of the developing flower buds on a marked inflorescence. Another 21 plants were not manipulated (controls). One month later, one of four flowers on each marked inflorescence received one of the following pollen manipulation treatments: control, pollinia removal, pollination without pollinia removal or pollination with pollinia removal. The response variable measured was the number of days each flower remained open (i.e. longevity). KEY RESULTS: The results showed significant flower bud removal and pollen manipulation effects on floral longevity; the interaction between these two factors was not significant. Flowers on inflorescences with previously removed flower buds remained open significantly longer than flowers on control inflorescences. On the other hand, pollinated flowers closed much faster than control and removed-pollinia flowers, the latter not closing significantly faster than control flowers, although this result was marginal. CONCLUSIONS: The results emphasize the strong relationship between floral longevity and pollination in orchids, as well as the influence of reproductive costs on the former.
AristaM, OrtizPL ( 2007). Differential gender selection on floral size: an experimental approach using Cistus salvifolius Journal of Ecology, 95, 973-982. DOI:10.1111/jec.2007.95.issue-5URL [本文引用: 3]
ArroyoMTK, ArmestoJJ, PrimackRB ( 1985). Community studies in pollination ecology in the high temperate Andes of central Chile II. Effect of temperature on visitation rates and pollination possibilities Plant Systematics and Evolution, 149, 187-203. DOI:10.1007/BF00983305URL [本文引用: 3]
ArroyoMTK, ArmestoJJ, VillagranC ( 1981). Plant phenological patterns in the high Andean cordillera of central Chile Journal of Ecology, 69, 205-223. DOI:10.2307/2259826URL [本文引用: 4]
ArroyoMTK, DudleyLS, JespersenG, PachecoDA, CavieresLA ( 2013). Temperature-driven flower longevity in a high-alpine species of Oxalis influences reproductive assurance New Phytologist, 200, 1260-1268. DOI:10.1111/nph.12443URL [本文引用: 10] Pollinator-excluded and hand-pollinated flowers were experimentally warmed with open-top chambers (OTCs) on a site at 3470m above sea level (asl). Flower-specific temperatures, and pollinator-excluded and open-pollination flower life-spans were measured at six alpine sites between 3100 and 3470m asl. Fruit set was analyzed in relation to inferred pollination time.Warming reduced potential FL. Variable thermal conditions across the alpine landscape predicted potential and actual FL; flower senescence was pollination-regulated. Actual FL and potential FL were coupled. Prolonged potential FL generally increased fruit set under cooler conditions.Plastic responses permit virgin flowers of O. compacta to remain open longer under cooler temperatures, thereby ameliorating slow pollination, and to close earlier when pollination tends to be faster under warmer conditions. Plastic potential FL provides adaptive advantages in the cold, thermally variable alpine habitat, and has important implications for reproductive success in alpine plants in a warming world.]]>
AshmanTL, SchoenDJ ( 1996). Floral longevity: fitness consequences and resource costs//Lloyd D, Barrett S. Floral Biology: Studies on Floral Evolution in Animal-Pollinated Plants. Springer New York.112-139. [本文引用: 9]
AshmanTL, SchoenDJ ( 1997). The cost of floral longevity in Clarkia tembloriensis: an experimental investigation Evolutionary Ecology, 11, 289-300. DOI:10.1023/A:1018416403530URL [本文引用: 4]
BauerAA, ClaytonMK, BrunetJ ( 2017). Floral traits influencing plant attractiveness to three bee species: consequences for plant reproductive success American Journal of Botany, 104, 772-781. DOI:10.3732/ajb.1600405URLPMID:28533203 [本文引用: 2] PREMISE OF THE STUDY: The ability to attract pollinators is crucial to plants that rely on insects for pollination. We contrasted the roles of floral display size and flower color in attracting three bee species and determined the relationships between plant attractiveness (number of pollinator visits) and seed set for each bee species. METHODS: We recorded pollinator visits to plants, measured plant traits, and quantified plant reproductive success. A zero-inflated Poisson regression model indicated plant traits associated with pollinator attraction. It identified traits that increased the number of bee visits and traits that increased the probability of a plant not receiving any visits. Different components of floral display size were examined and two models of flower color contrasted. Relationships between plant attractiveness and seed set were determined using regression analyses. KEY RESULTS: Plants with more racemes received more bee visits from all three bee species. Plants with few racemes were more likely not to receive any bee visits. The role of flower color varied with bee species and was influenced by the choice of the flower color model. Increasing bee visits increased seed set for all three bee species, with the steepest slope for leafcutting bees, followed by bumble bees, and finally honey bees. CONCLUSIONS: Floral display size influenced pollinator attraction more consistently than flower color. The same plant traits affected the probability of not being visited and the number of pollinator visits received. The impact of plant attractiveness on female reproductive success varied, together with pollinator effectiveness, by pollinator species.
CastroS, SilveiraP, NavarroL ( 2008). Effect of pollination on floral longevity and costs of delaying fertilization in the out-crossing Polygala vayredae Costa (Polygalaceae) Annals of Botany, 102, 1043-1048. DOI:10.1093/aob/mcn184URLPMID:18829587 [本文引用: 5] BACKGROUND AND AIMS: The effect of pollination on flower life span has been widely studied, but so far little attention has been paid to the reproductive consequences of delayed pollination in plants with long floral life spans. In the present study, Polygala vayredae was used to answer the following questions. (1) How does male and female success affect the floral longevity of individual flowers? (2) How does delaying fertilization affect the female fitness of this species? METHODS: Floral longevity was studied after experimental pollinations involving male and/or female accomplishment, bagging and open pollination. The reproductive costs of a delay in the moment of fertilization were evaluated through fruit set, seed-ovule ratio and seed weight, after pollination of flowers that had been bagged for 2-18 d. KEY RESULTS: Senescence of the flowers of P. vayredae was activated by pollen reception on the stigmatic papillae, while pollen removal had no effect on floral longevity. Nonetheless, a minimum longevity of 8 d was detected, even after successful pollination and pollen dissemination. This period may be involved with the enhancement of male accrual rates, as the female accomplishment is generally achieved after the first visit. Floral life span of open-pollinated flowers was variable and negatively correlated with pollinator visitation rates. Delayed pollination had a major impact on the reproductive success of the plant, with fruit set, seed-ovule ratio and seed weight being significantly diminished with the increase of flower age at the moment of fertilization. CONCLUSIONS: A strong relationship between pollination and floral longevity was observed. Flowers revealed the ability to extend or reduce their longevity, within some limits, in response to the abundance of efficient pollinators (i.e. reproductive fulfilment rates). Furthermore, with scarce or unpredictable pollinators, a long floral life span could maintain the opportunity for fertilization but would also have reproductive costs on production of offspring. Reduced female fitness late in the flower's life could shift the cost-benefit balance towards a shorter life span, partially counteracting the selection for longer floral life span potentially mediated by scarce pollination services.
DudashMR, HasslerC, StevensPM, FensterCB ( 2011). Experimental floral and inflorescence trait manipulations affect pollinator preference and function in a hummingbird- pollinated plant American Journal of Botany, 98, 275-282. DOI:10.3732/ajb.1000350URL [本文引用: 1] Methods : In single trait manipulative experiments we quantified hummingbird visitation preference and/or fluorescent dye (a pollen analog) donation as a function of number of flowers displayed (inflorescence size), height of the floral display (inflorescence height), floral color, and corolla tube length.Key results : Hummingbirds preferred to visit larger floral displays and floral displays at greater height, likely representing a general pollinator preference for larger, more visible signals and/or greater rewards. In addition, hummingbirds preferred to visit red flowers, and male function was greater in flowers manipulated to have longer corolla tubes.Conclusions : Selection pressures exerted by hummingbirds on S. virginica floral and inflorescence design through male reproductive success are consistent with the contemporary expression of floral traits of S. virginica relative to related Silene species with different pollinators, and they are consistent with the hummingbird syndrome of traits expressed by S. virginica]]>
DukasR, RealLA ( 1993). Learning constraints and floral choice behaviour in bumble bees Animal Behaviour, 46, 637-644. DOI:10.1006/anbe.1993.1240URL [本文引用: 1]
EvanhoeL, GallowayLF ( 2002). Floral longevity in Campanula americana (Campanulaceae): a comparison of morphological and functional gender phases American Journal of Botany, 89, 587-591. DOI:10.3732/ajb.89.4.587URLPMID:21665659 [本文引用: 1] Plastic responses to pollination and/or pollen removal may shift a flower's realized longevity closer to an optimal longevity that maximizes reproductive output per unit resource input. In particular, conditional responses to pollen removal and pollen deposition are expected in flowers of protandrous species in which the lengths of the male and female phases may be adjusted independently. We investigated plasticity in floral longevity in Campanula americana, a protandrous, insect-pollinated herb. In greenhouse studies, we found that the longevity of the morphological male phase was shortened by pollen removal and that the longevity of the morphological female phase was shortened by pollen deposition. In a natural population, male and female sexual functions saturated within a few hours of morphological gender phase onset. In contrast to theory, morphological gender phases did not terminate immediately upon saturation of sexual function. These findings are discussed in the context of current floral longevity theory.
GalenC, NewportMEA ( 1987). Bumble bee behavior and selection on flower size in the sky pilot, Polemonium viscosum Oecologia, 74, 20-23. DOI:10.1007/BF00377340URLPMID:28310409 [本文引用: 3] In alpine Polemonium viscosum, plants having sweet-scented flowers are primarily pollinated by queens of the bumble bee species, Bombus kirbyellus. In this paper we ask whether two aspects of the pollination effectiveness of bumble bees, visitation rate and pollination efficiency, vary significantly with flower size in sweet-flowered P. viscosum.(i) Bumble bees visited plants with large flowers on 80-90% of encounters, but visited those with smaller flowers on only 49% of encounters. (ii) However, the gain in pollination that large-flowered plants obtained via increased visitation was countered in part because bumble bees deposited fewer outcross pollen grains per visit on stigmas of large flowers than on those of small ones. When both visitation rate and pollination efficiency are taken into account, the predicted value of a single bumble bee encounter declines from 1.06 seeds for flowers larger than 18 mm in diameter to 0.55 seeds for flowers smaller than 12 mm in diameter. Our results suggest that bumble bee pollinators of P. viscosum prefer flower morphologies that are poorly suited for precise pollination. Such behavioral complexities are likely to place constraints on the evolution of
GaoJ, XiongYZ, HuangSQ ( 2015). Effects of floral sexual investment and dichogamy on floral longevity Journal of Plant Ecology, 8, 116-121. DOI:10.1093/jpe/rtv011URL [本文引用: 2] Aims Floral longevity, the duration that a flower remains open and functional, varies greatly among species. Variation in floral longevity has been considered to be optimal strategy for resource allocation under different ecological conditions, mainly determined by the rates of pollination and cost of flower maintenance. However, it is unclear whether an intrinsic factor, floral sexual investment, constrains evolution of floral longevity. The theoretical model also predicts that dichogamy favors long-lived flowers, but empirical studies to test this prediction remain unexplored.Methods To examine the effect of floral sexual investment on floral longevity, we measured flower size together with pollen and ovule production in 37 sympatric flowering plants in a natural community. The duration of the female and male phase in 21 protandrous species and floral longevity of the other 16 adichogamous species were documented in the field.Important findings Floral longevity varied from 1 day to 15 days, while pollen number per flower varied from 643 to 710880 and ovule number per flower from 1 to 426 in the 37 species. Flower size was correlated with pollen production as well as ovule production. Floral longevity was positively related to pollen production but not to ovule production. Consistent with the prediction that dichogamy favors long-lived flowers, we found the floral longevity of protandrous species was significantly longer than that of adichogamous species. In the protandrous species, pollen production per flower was observed to be positively related to male duration, while ovule production was not related to female duration. Our analyses of variation in floral longevity and sexual investment among different species suggest that the floral sexual investment could be an intrinsic factor contributing to the selected floral longevity, particularly the male phase, and that high pollen production could potentially increase pollen removal, i.e. male productive success.]]>
GiblinDE ( 2005). Variation in floral longevity between populations of Campanula rotundifolia (Campanulaceae) in response to fitness accrual rate manipulation American Journal of Botany, 92, 1714-1722. DOI:10.3732/ajb.92.10.1714URLPMID:21646089 [本文引用: 1] Floral longevity, the time between corolla expansion and senescence, contributes directly and indirectly to a plant's overall fitness. Though mating opportunities for insect-pollinated species often differ among populations, few empirical studies have addressed whether floral longevity varies in a manner consistent with these differences. I conducted experiments at thermally distinct sites to examine whether the prevailing floral longevity model predicted such variation between a montane and an alpine population of Campanula rotundifolia. Staminate phase duration was significantly shorter for montane vs. alpine C. rotundifolia flowers in the presence of pollinators, but significantly longer when pollinators were excluded. Montane flowers had a significantly higher female fitness accrual rate, significantly shorter total longevities, and, unlike alpine flowers, were not pollen-limited. Delaying pollinator access to pistillate phase flowers significantly increased total longevity in alpine flowers only. Significant differences in total longevity between populations resulted from an extended pistillate phase in alpine flowers. Overall, the prevailing model accurately predicted the total floral longevity trends found here. However, I provide novel evidence for geographic and gender-specific plasticity in the floral longevity response to fitness accrual rate variation, suggesting C. rotundifolia populations in this study may be attuned to local schedules of pollinator activity.
GoodwillieC, SargentRD, EckertCG, ElleE, GeberMA, JohnstonMO, KaliszS, MoellerDA, ReeRH, Vallejo- MarinM, WinnAA ( 2010). Correlated evolution of mating system and floral display traits in flowering plants and its implications for the distribution of mating system variation New Phytologist, 185, 311-321. DOI:10.1111/j.1469-8137.2009.03043.xURL [本文引用: 1]
HarderLD, JohnsonSD ( 2009). Darwin’s beautiful contrivances: evolutionary and functional evidence for floral adaptation New Phytologist, 183, 530-545. DOI:10.1111/nph.2009.183.issue-3URL [本文引用: 1]
HeYP, FeiSM, LiuJQ, ChenXM, WangP, JiangJM, HeF ( 2005). A preliminary review of studies of alpine plant breeding system Journal of Sichuan Forestry Science and Technology, 26(4), 43-49. [本文引用: 1]
HeglandSJ, Totland? ( 2005). Relationships between species’ floral traits and pollinator visitation in a temperate grassland Oecologia, 145, 586-594. DOI:10.1007/s00442-005-0165-6URL [本文引用: 3] Knowledge about plant–plant interactions for pollinator service at the plant community level is still scarce, although such interactions may be important to seed production and hence the population dynamics of individual plant species and the species compositions of communities. An important step towards a better understanding of pollination interactions at the community level is to assess if the variation in floral traits among plant species explain the variation in flower visitation frequency among those species. We investigated the relative importance of various floral traits for the visitation frequency of all insects, and bumblebees and flies separately, to plant species by measuring the visitation frequency to all insect-pollinated species in a community during an entire flowering season. Visitation frequency was identified to be strongly positive related to the visual display area and the date of peak flowering of plant species. Categorical variables, such as flower form and symmetry, were important to the visitation frequency of flies only. We constructed floral similarity measures based on the species’ floral traits and found that the floral similarity for all species’ traits combined and the continuous traits separately were positively related to individual visitation frequency. On the other hand, plant species with similar categorical floral traits did not have similar visitation frequencies. In conclusion, our results show that continuous traits, such as flower size and/or density, are more important for the variation in visitation frequency among plant species than thought earlier. Furthermore, differences in visitation frequency among pollinator groups give a poor support to the expectations derived from the classical pollination syndromes.]]>
HuC, LiuZJ, WuGQ, ZhaoZG ( 2013). Floral characteristic and breeding system of Anemone obtusiloba Acta Agrestia Sinica, 21, 783-788.
ItagakiT, SakaiS ( 2006). Relationship between floral longevity and sex allocation among flowers within inflorescences in Aquilegia buergeriana var. oxysepala (Ranunculaceae) American Journal of Botany, 93, 1320-1327. DOI:10.3732/ajb.93.9.1320URLPMID:21642197 [本文引用: 2] Understanding the fitness of plants with inflorescences requires examining variation in sex allocation among flowers within inflorescences. We examined whether differences in the duration of the male and female phases of flowering lead to variation in sex allocation and reproductive success among flowers within inflorescences. In 2002 and 2003, we quantified floral longevity, floral sex allocation, and reproductive success between the first and the second flowers within inflorescences in a protandrous species, Aquilegia buergeriana var. oxysepala. Floral longevity was greater in the first flowers than in the second ones in both years. The male phase lasted longer, and the initial number of pollen grains and the number of pollen grains removed were greater in the first flowers than in the second ones in both years. Within first flowers, the number of pollen grains removed was greater in flowers that had longer male phases, thus duration of the male phase may positively affect male reproductive success in the first flowers. The female phase lasted longer and the number of ovules was greater in the first flowers than in the second only in 2002. However, seed production per flower and female phase duration in both years were not significantly related. The variation in the number of pollen grains among flowers in this species may be caused by the variation in male phase duration.
KarronJD, MitchellRJ ( 2012). Effects of floral display size on male and female reproductive success in Mimulus ringens Annals of Botany, 109, 563-570. DOI:10.1093/aob/mcr193URL [本文引用: 1] To facilitate unambiguous assignment of paternity, we established four replicate (cloned) arrays of Mimulus ringens, each consisting of genets with unique combinations of homozygous marker genotypes. In each array, we trimmed displays to two, four, eight or 16 flowers. When fruits ripened, we counted the number of seeds per fruit and assigned paternity to 1935 progeny.Siring success per flower declined sharply with increasing display size, while female success per flower did not vary with display. The rate of self-fertilization increased for large floral displays, but siring losses due to geitonogamous pollen discounting were much greater than siring gains through increased self-fertilization. As display size increased, each additional seed sired through geitonogamous self-pollination was associated with a loss of 97 seeds sired through outcrossing.Although total fitness increased with floral display size, the marginal return on each additional flower declined steadily as display size increased. Therefore, a plant could maximize fitness by producing small displays over a long flowering period, rather than large displays over a brief flowering period.]]>
Lawrence VenableD ( 1992). Size-number trade-offs and the variation of seed size with plant resource status The American Naturalist, 140, 287-304. DOI:10.1086/285413URL [本文引用: 2]
LázaroA, JakobssonA, Totland? ( 2013). How do pollinator visitation rate and seed set relate to species’ floral traits and community context? Oecologia, 173, 881-893. DOI:10.1007/s00442-013-2652-5URL [本文引用: 2] Differences among plant species in visitation rate and seed set within a community may be explained both by the species' floral traits and the community context. Additionally, the importance of species' floral traits vs. community context on visitation rate and seed set may vary among communities. In communities where the pollinator-to-flower ratio is low, floral traits may be more important than community context, as pollinators may have the opportunity to be choosier when visiting plant species. In this study we investigated whether species' floral traits (flower shape, size and number, and flowering duration) and community context (conspecific and heterospecific flower density, and pollinator abundance) could explain among-species variation in visitation rate and seed set. For this, we used data on 47 plant species from two Norwegian plant communities differing in pollinator-to-flower ratio. Differences among species in visitation rate and seed set within a community could be explained by similar variables as those explaining visitation rate and seed set within species. As expected, we found floral traits to be more important than community context in the community with a lower pollinator-to-flower ratio; whereas in the community with a higher pollinator-to-flower ratio, community context played a bigger role. Our study gives significant insights into the relative importance of floral traits on species' visitation rate and seed set, and contributes to our understanding of the role of the community context on the fitness of plant species.
MengJL ( 2010). Reproductive Strategies of Three Common Species of Ranunculaceae at Alpine Meadow in Qinghai Tibetan Plateau. PhD dissertation, Lanzhou University Lanzhou. [本文引用: 7]
MullerI, SchmidB, WeinerJ ( 2000). The effect of nutrient availability on biomass allocation patterns in 27 species of herbaceous plants. Perspectives in Plant Ecology, Evolution and Systematics, 3, 115-127.
NiuT, ChenLX, ZhouZJ ( 2004). The characteristics of climate change over the Tibetan Plateau in the last 40 years and the detection of climatic jumps Advances in Atmospheric Sciences, 21, 193-203. DOI:10.1007/BF02915705URL [本文引用: 3] Through analyzing the yearly average data obtained from 123 regular meteorological observatories located in the Tibetan Plateau (T-P), this article studies the characteristics of climate change in T-P in the last 40 years. From the distribution of the linear trend, it can be concluded that the southeastern part of T-P becomes warmer and wetter, with an obvious increase of rainfall. The same characteristics are found in the southwestern part of T-P, but the shift is smaller. In the middle of T-P, temperature and humidity obviously increase with the center of the increase in Bangoin-Amdo. The south of the Tarim Basin also exhibits the same tendency. The reason for this area being humid is that it gets less sunshine and milder wind. The northeastern part of T-P turns warmer and drier. Qaidam Basin and its western and southern areas are the center of this shift, in which the living environment is deteriorating. Analyzing the characteristics of the regional average time series, it can be found that in the mid-1970s, a significant sudden change occurred to annual rainfall, yearly average snow-accumulation days and surface pressure in the eastern part of T-P. In the mid-1980s, another evident climatic jump happened to yearly average temperature, total cloud amount, surface pressure, relative humidity, and sunshine duration in the same area. That is, in the mid 1980s, the plateau experienced a climatic jump that is featured by the increase of temperature, snow-accumulation days, relative humidity, surface pressure, and by the decrease of sunshine duration and total cloud amount. The sudden climatic change of temperature in T-P is later than that of the global-mean temperature. From this paper it can be seen that in the middle of the 1980s, a climatic jump from warm-dry to warm-wet occurred in T-P.]]>
RathckeBJ ( 2003). Floral longevity and reproductive assurance: seasonal patterns and an experimental test with Kalmia latifolia (Ericaceae) American Journal of Botany, 90, 1328-1332. DOI:10.3732/ajb.90.9.1328URLPMID:21659232 [本文引用: 1] Floral longevity is assumed to reflect a balance between the benefit of increased pollination success and the cost of flower maintenance. Flowers of Kalmia latifolia (Ericaceae), mountain laurel, have a long duration and can remain viable up to 21 d if unpollinated. I experimentally tested whether this long duration increases pollination success by clipping stigmas to reduce functional floral longevity to 3-4 d. Clipping stigmas decreased fruit set from 65% to only 10%. Flowers with natural life spans were not pollination-limited, demonstrating that long floral duration ensured female reproductive success. The long floral duration of K. latifolia was unique in this site (the Great Swamp, Rhode Island, USA). Coflowering shrubs in summer had a mean floral life span of 3.4 d. Spring-flowering species had significantly longer mean floral durations (7.2 d). These duration differences may reflect seasonal variation in pollinator availability. However, K. latifolia flowers in summer, when its bumble bee pollinators are abundant but it is a poor competitor for bees because its flowers produce little nectar. The long floral duration allows K. latifolia to outlast coflowering competitors and attract sufficient pollinators. I hypothesize that the long floral duration of K. latifolia functions as a mechanism for competitive avoidance and reproductive assurance.
SchoenDJ, AshmanTL ( 1995). The evolution of floral longevity: resource allocation to maintenance versus construction of repeated parts in modular organisms Evolution, 49, 131-139. DOI:10.1111/j.1558-5646.1995.tb05965.xURLPMID:28593673 [本文引用: 4] The component parts of modular organisms often show interspecific variation in their longevity. In plants, the flower is an example of such a structure. Models are developed in this paper to predict optimal floral longevity (the optimal length of time that flowers should remain open and functional) under a variety of conditions. A tradeoff involving allocation of resources to floral construction versus floral maintenance is assumed. The main model variables are the rate at which pollen and seed fitness accrue over time (fitness-accrual rates) and the daily cost of maintaining an existing flower relative to the cost of constructing a new one (floral maintenance cost). Long-lived flowers are selected when fitness-accrual rates and floral maintenance costs are low, whereas short-lived flowers are selected when fitness-accrual rates and floral maintenance costs are high. Dichogamy favors longer-lived flowers relative to homogamy, whereas nonindependence among flowers in their attractiveness to pollinators (attraction to flower clusters) selects for shorter-lived flowers. Reduction in floral maintenance costs later on in the flower's life favors longer-lived flowers. Observations on the dissemination and receipt of pollen in individual flowers over time, together with measurements of corolla respiration and nectar sugar production rate are required to test the model quantitatively. The parameters important to the evolution of optimal floral longevity (i.e., maintenance and construction costs, and fitness-accrual rates) may be general features of evolution of optimal longevities of other repeated structures.
SpiglerRB ( 2017). Plasticity of floral longevity and floral display in the self-compatible biennial Sabatia angularis (Gentianaceae): untangling the role of multiple components of pollination Annals of Botany, 119, 167-176. DOI:10.1093/aob/mcw195URLPMID:28062510 [本文引用: 7] BACKGROUND AND AIMS: Plasticity of floral traits in response to pollination can enable plants to maximize opportunities for pollen import and export under poor pollination conditions, while minimizing costs under favourable ones. Both floral longevity and display are key traits influencing pollination. While pollination-induced flower wilting is widely documented, we lack an understanding of the multifactorial complexity of this response, including the influence of other pollination components, costs of extended longevity and subsequent impacts on floral display. METHODS: Plasticity of floral longevity was experimentally evaluated in Sabatia angularis in response to multiple pollination factors: pollen addition, removal, and source (self, single-donor outcross, multiple-donor outcross) and timing of pollination. Effects of pollen quantity were further evaluated by exploiting variation in autonomous self-pollen deposition. Delayed pollination costs were tested comparing seed set from early versus late pollinations. Finally, I compared floral display metrics (peak floral display, time to peak flower, flowering duration, mean flowering rate) between experimentally pollinated and control plants. KEY RESULTS: Floral longevity was highly plastic in response to pollen addition and its timing, and the response was dose-dependent but insensitive to pollen source. Pollen removal tended to extend floral longevity, but only insofar as it precluded pollination-induced wilting via autonomous self-pollination. Under delayed pollination, the wilting response was faster and no cost was detected. Pollination further led to reduced peak floral displays and condensed flowering periods. CONCLUSIONS: Floral longevity and display plasticity could optimize fitness in S. angularis, a species prone to pollen limitation and high inbreeding depression. Under pollinator scarcity, extended floral longevities offer greater opportunities for pollen receipt and export at no cost to seed set, reproductive assurance via autonomous self-pollination and larger, more attractive floral displays. Under high pollinator availability, shortened longevities lead to smaller displays that should lower the risk of geitonogamy.
SpiglerRB, WoodardAJ ( 2019). Context-dependency of resource allocation trade-offs highlights constraints to the evolution of floral longevity in a monocarpic herb New Phytologist, 221, 2298-2307. DOI:10.1111/nph.2019.221.issue-4URL [本文引用: 4]
SteinacherG, WagnerJ ( 2010). Flower longevity and duration of pistil receptivity in high mountain plants Flora, 205, 376-387. DOI:10.1016/j.flora.2009.12.012URL [本文引用: 1]
StrattonDA ( 1989). Longevity of individual flowers in a Costa Rican cloud forest: ecological correlates and phylogenetic constraints Biotropica, 21, 308-318. DOI:10.2307/2388281URL [本文引用: 3]
TeixidoAL, ValladaresF ( 2014). Disproportionate carbon and water maintenance costs of large corollas in hot Mediterranean ecosystems Perspectives in Plant Ecology Evolution and Systematics, 16, 83-92. DOI:10.1016/j.ppees.2014.02.002URL [本文引用: 1] Larger corollas increase the reproductive success of entomophilous plants, but are also associated with increased carbon and water costs, especially under hot and dry conditions. Minimizing floral carbon and water loss by reducing corolla size should be potentially advantageous for plants living in these environments. We quantify maintenance costs of corollas (water and carbon) in large-flowered rockroses (Cistus spp.) in a Mediterranean ecosystem. We performed field studies of two coflowering sympatric Cistus of contrasting corolla size to analyze water costs. Additionally, we used the larger-flowered species (C ladanifer) to analyze the effects of intraspecific variation in corolla size on floral net carbon exchange and transpiration rates. We also assessed the mean daily percentage of plant water and carbon consumed by corollas by comparing with that of leaves at the time of flowering in C ladanifer. Temperature and corolla area increased water maintenance costs, following an allometric relationship where transpiration rate per unit of area increased with corolla area. Larger flowers tended to heat less under strong irradiance than smaller ones in both species, especially in C ladanifer, demonstrating a stronger transpirational cooling effect on larger flowers. In terms of carbon, temperature significantly affected net carbon exchange rates, which were not affected by corolla size. Daily water and carbon expenses of corolla were ca. 50% of those of leaves on an organ surface area basis. Our results suggest that water and carbon maintenance costs Of large flowers in the Mediterranean impose significant constraints to corolla size, ecophysiologically favoring smaller-flowered individuals in these ecosystems. (C) 2014 Geobotanisches Institut ETH, Stiftung Ruebel. Published by Elsevier GmbH:
TeixidoAL, ValladaresF ( 2015). Temperature-limited floral longevity in the large-flowered Mediterranean shrub Cistus ladanifer (Cistaceae) International Journal of Plant Sciences, 176, 131-140. DOI:10.1086/679477URL [本文引用: 1]
TotlandR, SottocornolaM ( 2001). Pollen limitation of reproductive success in two sympatric alpine willows (Salicaceae) with contrasting pollination strategies American Journal of Botany, 88, 1011-1015. URLPMID:11410464 [本文引用: 1] We compared the extent of pollen limitation on female reproductive success of Salix lanata L., an entirely insect-pollinated willow, and S. lapponum L., which is 50 : 50% insect : wind pollinated (ambophilous). Supplemental hand-pollination significantly increased seed number per fruit by nearly 50% in the insect-pollinated willow, but had no significant impact on seed number in the dually pollinated species. Fruit set was not affected by the treatment in either of the species. These results demonstrate that pollen limitation on reproductive success is most pronounced in the species that depends entirely on insects for pollination. In general, pollinator visitation was highest to S. lapponum, but bumble bees were only observed on S. lanata, suggesting that the quantity and quality of pollinator visitation differed between the species. Our results empirically support the hypothesis that a dual pollination strategy is most effective in alpine environments with low and infrequent pollinator activity and high wind speeds.
TrunschkeJ, St?cklinJ ( 2017). Plasticity of flower longevity in alpine plants is increased in populations from high elevation compared to low elevation populations Alpine Botany, 127, 41-51. DOI:10.1007/s00035-016-0176-4URL [本文引用: 1]
WangYX, LiuZJ, ZhaoZG, HouM, ZhangXR, LüWL ( 2018). Responses of floral longevity to pollination environments in 11 species from two alpine meadows Biodiversity Science, 26, 510-518. DOI:10.17520/biods.2018065URL [本文引用: 3] Floral longevity, the length of time that a flower remains open and functional, varies greatly among species. The high plasticity of floral longevity can reflect an adaptation to variable pollination environments. In the alpine meadows of Qinghai-Tibetan Plateau (2,900 m vs. 3,600 m), we assessed how potential floral longevity, the shortest floral longevity and the actual floral longevity of 11 species varied under treatments of different pollination contexts. We modeled the response of floral longevity to pollinator exclusion (potential floral longevity minus actual floral longevity) and supplemental pollination (actual longevity minus shortest longevity), and the plasticity of floral longevity (potential longevity minus shortest longevity) at high and low altitude habitats. We found that the plants at the high-altitude community had longer potential floral longevity compared to the low-altitude community, while the shortest floral longevity was not significantly different. Furthermore, pollinator exclusion significantly increased flower longevity, while supplemental pollination significantly decreased floral longevity (i.e. the potential floral longevity > the actual floral longevity > the shortest floral longevity) in both high and low-altitude habitats. In comparison with the low-altitude community, high-altitude plants exhibited higher plasticity of floral longevity. Overall, our results suggest that greater plasticity of floral longevity may increase the opportunity for pollination and thus aid fitness at higher altitudes where pollinators are scarce and unpredictable. [ 王玉贤, 刘左军, 赵志刚, 侯盟, 张小瑞, 吕婉灵 ( 2018). 青藏高原高寒草甸植物花寿命对传粉环境的响应 生物多样性, 26, 510-518.] DOI:10.17520/biods.2018065URL [本文引用: 3] 花寿命是指花保持开放且具有功能的时间长度。不同物种的花寿命具有显著的差异, 但一定程度上的可塑性反映了植物对传粉环境变化的适应性。本研究以青藏高原高寒草甸不同海拔(2,900 m和3,600 m)的11种开花植物为研究对象, 通过套袋、补充授粉以及自然授粉处理分别测量了植物的潜在花寿命、最短花寿命以及实际花寿命。分析了不同海拔植物花寿命的变异, 以及对套袋处理(潜在花寿命与实际花寿命之差)和补充授粉处理(实际花寿命与最短花寿命之差)的响应及其可塑性(潜在花寿命与最短花寿命之差)的变异。结果表明: 无论是高海拔还是低海拔, 套袋显著延长了花寿命, 而补充授粉显著缩短了花寿命, 即, 潜在花寿命>实际花寿命>最短花寿命。高海拔群落中植物的实际花寿命和潜在花寿命均显著延长, 而最短花寿命在海拔间差异不显著。相对于低海拔群落, 高海拔植物的花寿命对补充授粉处理更敏感, 花寿命的可塑性更大。本研究说明传粉者稀少的高寒环境中, 植物通过提高花寿命及其可塑性来增加授粉机会, 获得更高的适合度。
ZhangFP, YangYJ, YangQY, ZhangW, BrodribbTJ, HaoGY, HuH, ZhangSB ( 2017). Floral mass per area and water maintenance traits are correlated with floral longevity in Paphiopedilum (Orchidaceae) Frontiers in Plant Science, 8, 501. DOI: 10.3389/fpls.2017.00501. DOI:10.3389/fpls.2017.00501URLPMID:28439283 [本文引用: 1] Floral longevity (FL) determines the balance between pollination success and flower maintenance. While a longer floral duration enhances the ability of plants to attract pollinators, it can be detrimental if it negatively affects overall plant fitness. Longer-lived leaves display a positive correlation with their dry mass per unit area, which influences leaf construction costs and physiological functions. However, little is known about the association among FL and floral dry mass per unit area (FMA) and water maintenance traits. We investigated whether increased FL might incur similar costs. Our assessment of 11 species of Paphiopedilum (slipper orchids) considered the impact of FMA and flower water-maintenance characteristics on FL. We found a positive relationship between FL and FMA. Floral longevity showed significant correlations with osmotic potential at the turgor loss and bulk modulus of elasticity but not with FA. Neither the size nor the mass per area was correlated between leaves and flowers, indicating that flower and leaf economic traits evolved independently. Therefore, our findings demonstrate a clear relationship between FL and the capacity to maintain water status in the flower. These economic constraints also indicate that extending the flower life span can have a high physiological cost in Paphiopedilum.
ZhangJ ( 2013). Study on Reproductive Biology of Taraxacum species in Northeast China. PhD dissertation, Shenyang Agricultural University Shenyang.
[ 张建 ( 2013). 蒲公英属植物繁殖生物学研究 博士学位论文, 沈阳农业大学, 沈阳.]
ZhangZQ, ZhangYH, SunH ( 2011). The reproductive biology of Stellera chamaejasme (Thymelaeaceae): a self-incompatible weed with specialized flowers Flora, 206, 567-574. DOI:10.1016/j.flora.2011.01.008URL [本文引用: 3] It has profound implications for species invasions and fitness, how weeds achieve reproductive success. We present the first study on the reproductive biology of Stellera chamaejasme, a toxic perennial weed which is abundant in the alpine meadows on the eastern Tibetan Plateau of China since the 1960s. S. chamaejasme has synchronogamy and herkogamous flowers, which last for about 11 days. Flowers can be yellow, white or reddish. Our results suggest that this species is self-incompatible and depends entirely on pollen vectors for seed production. All the pollinators observed were Lepidopteran species; however, the pollinator assemblage was significantly different among populations and likely independent on floral colour. Reproductive success and invasion of the weed might be a complex function of floral traits, breeding system and pollination ecology. Generally, plants with generalized pollination system are more invasive. However, our results suggest that specialized flowers and self-incompatibility in the present case may not prevent invasion of the weed. (C) 2011 Published by Elsevier GmbH.
ZhaoZG, DuGZ ( 2003). Characteristics of the mating system and strategies for resource allocation in Ranunculaceae Journal of Lanzhou University (Natural Science), 39, 70-74. [本文引用: 1]
ZhaoZG, HouM, WangYJ, DuGZ ( 2020). Phenological variation of flower longevity and duration of sex phases in a protandrous alpine plant: potential causes and fitness significance BMC Plant Biology, 20, 137. DOI:10.1186/s12870-020-02356-wURLPMID:32245411 BACKGROUND: Flower longevity plays an important role in pollen dispersal and reproductive success in plants. In dichogamous plants, the duration of anthesis as well as the time allocated to male and female functions can vary in response to intrinsic factors (e.g., flowering time and resource allocation) and pollination context along a growth season. However, the fitness consequences of phenological dynamics have rarely been examined. This study aims to unravel the potential causes driving variation in flower longevity, duration of sex phases, and phenotypic sex during a flowering season of strongly protandrous Aconitum gymnandrum, and particularly reproductive consequences of the phenological pattern. RESULTS: Population floral sex ratio shifted from completely male at the beginning to completely female at the end of the season, as is common in other protandrous plants. Phenological dynamics of the floral sex ratio and the duration of sex phases caused a shift from femaleness to maleness in the mean phenotypic sex over the whole season. Floral longevity was negatively correlated with flower size and positively affected by temperature. Early flowers within inflorescences rather than early-flowering individuals emphasized the duration of female over male phase. Owing to the dominance of male-phase flowers, early flowering for individual flowers and plants, or female-biased sex resulted in higher pollen deposition per flower and seed set. At the flower level, flower longevity positively affected female reproductive success, while the effect of flower size was negative. By contrast, plant-level female reproductive success was negatively affected by flower longevity but positively correlated to flower size. CONCLUSIONS: The major result of this study lies in elucidating the relationship between variation in phenological sex expression and floral longevity and their fitness consequences of protandrous A. gymnandrum. The contrasting results on female fitness for individual flowers and plants contribute to our current understanding of the adaptive significance of floral longevity.
ZhaoZG, WangYK ( 2015). Selection by pollinators on floral traits in generalized Trollius ranunculoides (Ranunculaceae) along altitudinal gradients PLOS ONE, 10, e0118299. DOI: 10.1371/journal.pone.0118299. DOI:10.1371/journal.pone.0118299URLPMID:25692295 [本文引用: 2] Abundance and visitation of pollinator assemblages tend to decrease with altitude, leading to an increase in pollen limitation. Thus increased competition for pollinators may generate stronger selection on attractive traits of flowers at high elevations and cause floral adaptive evolution. Few studies have related geographically variable selection from pollinators and intraspecific floral differentiation. We investigated the variation of Trollius ranunculoides flowers and its pollinators along an altitudinal gradient on the eastern Qinghai-Tibet Plateau, and measured phenotypic selection by pollinators on floral traits across populations. The results showed significant decline of visitation rate of bees along altitudinal gradients, while flies was unchanged. When fitness is estimated by the visitation rate rather than the seed number per plant, phenotypic selection on the sepal length and width shows a significant correlation between the selection strength and the altitude, with stronger selection at higher altitudes. However, significant decreases in the sepal length and width of T. ranunculoides along the altitudinal gradient did not correspond to stronger selection of pollinators. In contrast to the pollinator visitation, mean annual precipitation negatively affected the sepal length and width, and contributed more to geographical variation in measured floral traits than the visitation rate of pollinators. Therefore, the sepal size may have been influenced by conflicting selection pressures from biotic and abiotic selective agents. This study supports the hypothesis that lower pollinator availability at high altitude can intensify selection on flower attractive traits, but abiotic selection is preventing a response to selection from pollinators.
ZhuWQ, ChenLX, ZhouZJ (2001). Several characteristics of contemporary climate change in the Tibetan Plateau Science in China Series D: Earth Sciences, 44, 410-420. [本文引用: 1]
ZuurAF, IenoEN, WalkerN, SavelievAA, SmithGM (2009). Mixed Effects Models and Extensions in Ecology with R Springer, New York.
Is floral longevity influenced by reproductive costs and pollination success in Cohniella ascendens (Orchidaceae)? 4 2007
Community studies in pollination ecology in the high temperate Andes of central Chile II. Effect of temperature on visitation rates and pollination possibilities 3 1985
... 青藏高原恶劣的环境条件(如低温、高辐射以及气候多变等; Zhu et al., 2001; Niu et al., 2004)使得传粉昆虫的数量稀少且不可预测(Arroyo et al., 1985).这严重限制了植物的生存与繁殖, 尤其是依赖传粉者传粉的植物(Arroyo et al., 1985; Totland & Sottocornola, 2001).青藏高原东部高寒草甸植物的繁殖受到严重的花粉限制(孟金柳, 2010), 这使该地区成为研究延长花寿命作为繁殖保障策略的一个理想的生态系统(Arroyo et al., 1981, 2013).尽管王玉贤等(2018)已经证明在青藏高原东部的高寒草甸环境中, 植物通过提高花寿命及其可塑性来增加授粉机会, 获得更高的适合度; 且相对于低海拔生境, 高海拔生境中的花寿命的可塑性更大.但这仅仅是基于不同海拔的传粉环境(传粉者资源)的差异得出的结论, 在很大程度上忽略了延长花寿命会增加花的维持成本和繁殖成本(Ashman & Schoen, 1994, 1996).基于这一研究结果和资源分配策略, 我们调查了青藏高原东部高寒草甸中不同海拔的两个植物群落, 主要回答以下问题: (1)这两个研究群落中, 在物种水平上, 植物的花寿命与花吸引特征(花大小和花数目)之间是否存在资源权衡?(2)在不同的传粉环境中, 植物的花寿命与花吸引特征对植物雌性繁殖成功的相对重要性是否发生改变?并进一步预测在传粉者更为稀少的高海拔群落中, 花寿命对植物雌性适合度更为重要. ...
Correlated evolution of mating system and floral display traits in flowering plants and its implications for the distribution of mating system variation 1 2010
Relationship between floral longevity and sex allocation among flowers within inflorescences in Aquilegia buergeriana var. oxysepala (Ranunculaceae) 2 2006
... 植物资源状况与花寿命的关系是影响植物繁殖保障和总体适合度的一种重要机制(Primack, 1985; Rathcke, 2003), 这在传粉者访问频率低和/或繁殖成本高的物种中显得更为重要.然而很多研究却忽略了花寿命与资源状况之间的联系(Ashman & Schoen, 1997; Abdala-Roberts et al., 2007).研究结果显示, 无论是低海拔还是高海拔环境, 植物的花寿命与植物的植株高度均呈显著的正相关关系.这与Abdala-Roberts等(2007)关于Cohniella ascendens花寿命与资源水平的研究结果是一致的.在传粉者资源稀少或不确定的高寒环境中, 植物繁殖成功通常受到严重的花粉限制(Arroyo et al., 2013), 植物往往通过可塑性地延长花寿命来增加接受传粉者访问的机会, 进而保障植物的繁殖(Arroyo et al., 2013; 王玉贤等, 2018).尽管延长花寿命需要消耗资源, 但相对于构建新花, 延长花寿命的资源投入较少(Arroyo et al., 2013).因此, 对于高寒环境的植物来说, 可利用的资源越多, 植物将有更多的资源用于花寿命的延长, 以期在传粉者资源不足的情况下, 增加传粉者访问的机会, 并使植物获得更大的适合度(Rathcke, 2003; Abdala-Roberts et al., 2007). ...
Plasticity of floral longevity and floral display in the self-compatible biennial Sabatia angularis (Gentianaceae): untangling the role of multiple components of pollination 7 2017
 ,
, ,1,*, 路宁娜3
,1,*, 路宁娜3

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT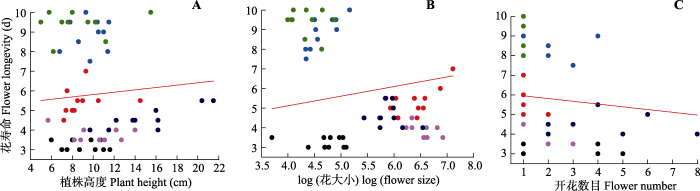 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT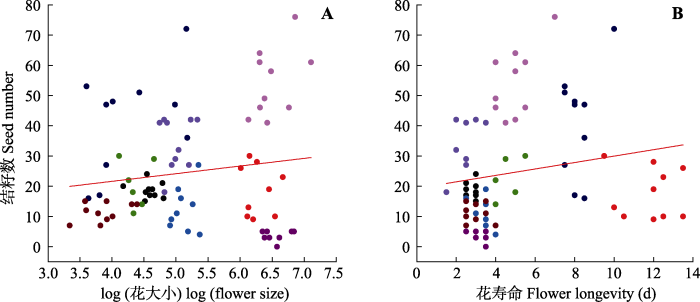 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT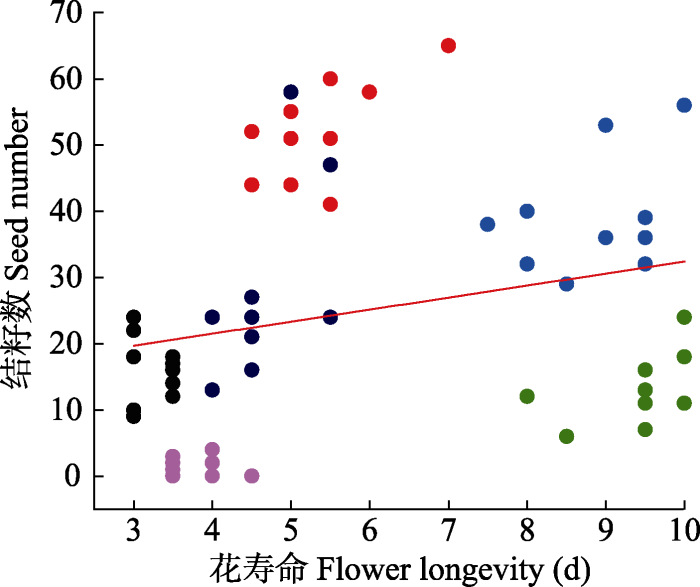 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}