Supported by the National Natural Science Foundation of China(31971636) the Yong Elite Scientists Sponsorship Program by CAST(31971636) the Fundamental Research Fund for the Central Universities(31971636)
Abstract Aims Variations of many branch traits are affected by plant size, branch age and environment factors, but the relative importance of these factors to intraspecies variations of branch traits has rarely been evaluated simultaneously. Methods In this study, we took Pinus koraiensis as the research object, to explore the effects of plant size (diameter at breast height (DBH) or tree height), branch age and environmental factors (light intensity, soil nutrient content and water availability) on branch traits, by measuring morphological traits, chemical traits and anatomical traits in different branch ages of 69 individuals with DBH in the range of 0.3-100.0 cm. Important findings Our results showed that: (1) DBH and tree height had different effects on branch traits: wood density (WD), the xylem area-to-total cross-sectional area ratio (RXA), the phloem area-to-total cross-sectional area ratio (RPHA) and the pith area-to-total cross-sectional area ratio (RPA) were more sensitive to DBH, while the total resin canal area-to-total cross-sectional area ratio (RRC) and wood nitrogen content (WN) were more affected by the tree height; (2) branch age was the most important factor in driving intra-specific variations of branch traits of P. koraiensis, followed by plant size, while the impact of environment factors was minimal; (3) WD and RPHA were significantly positively correlated with DBH, while RPA was significantly negatively correlated with DBH; and RRC and WN were significantly positively correlated with tree height. Except for WN, the relationships between branch traits and branch age were significant, and as tree growth, the rate of RRC decreasing with branch age was enhanced, but the rate of RPA decreasing with branch age was weakened. The results of our study are helpful to understand the driving factors of intraspecific variation of branch traits at the local scale and the adaptation mechanism of branches to cope with environmental changes. Keywords:branch age;diameter at breast height;tree height;environment;wood density;anatomical trait;wood nitrogen content
PDF (1979KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 于青含, 金光泽, 刘志理. 植株大小、枝龄和环境共同驱动红松枝性状的变异. 植物生态学报, 2020, 44(9): 939-950. DOI: 10.17521/cjpe.2020.0173 YU Qing-Han, JIN Guang-Ze, LIU Zhi-Li. Plant size, branch age and environment factors co-drive variations of branch traits of Pinus koraiensis. Chinese Journal of Plant Ecology, 2020, 44(9): 939-950. DOI: 10.17521/cjpe.2020.0173
Fig. 1Schematic diagram of anatomical structure of branches of different ages under an optical microscope for Pinus koraiensis. A, Current-year branch. B, Two-years-old branch. C, Three-years-old branch. D, Four-years-old branch.
1.4 环境因子的测定
针对每株样树, 采集样枝前利用半球摄影法(带有180°鱼眼镜头的Nikon Coolpix 4500数码相机, Nikon, Tokyo, Japan)采集半球图片。通过Gap Light Analyzer ver. 2.0软件计算每张半球图片0-60°天顶角范围内的总入射辐射(mol·m-2·d-1), 以该值表征光照强度(Liu et al., 2020)。
对于每株样树, 于树干底部0-10 cm的土层范围内使用土壤取芯器采集土壤子样本, 重复3次且任意两个采样方向之间的角度约为120°, 将这3个子样本进行混合并剔除明显的根和凋落物等杂质(Xu et al., 1987; Yang et al., 2019a; Liu et al., 2020)。然后, 利用烘干法测定每个样品的土壤水含量(g·g-1); 采用AQ400自动间断化学分析仪(SEAL Analytical, Mequon, USA)测量全氮含量(mg·g-1)和全磷含量(mg·g-1)。
Table 1 表1 表1红松胸径(DBH)或树高与枝性状回归分析的赤池信息量准则(AIC)值 Table 1Akaike information criterion (AIC) values in the regressions of diameter at breast height (DBH) or tree height against each branch traits of Pinus koraiensis
性状 Trait
DBH (cm)
树高 Tree height (m)
木质密度 WD (g·cm-3)
-1 025
-1 022
枝氮含量 WN (mg·g-1)
1 282
1 272
木质部面积占比 RXA
-327
-325
韧皮部面积占比 RPHA
-1 090
-1 086
髓面积占比 RPA
-1 364
-1 362
树脂道总面积占比 RRC
-1 129
-1 142
Parameters with bold were used to represent plant size. RPA, pith area-to-total cross-sectional area ratio; RPHA, phloem area-to-total cross-sectional area ratio; RRC, total resin canal area-to-total cross-sectional area ratio; RXA, xylem area-to-total cross-sectional area ratio; WD, wood density; WN, wood nitrogen content. 利用加粗值对应的参数来表征植株大小。
Table 2 表2 表2木质密度(WD)、枝氮含量(WN)、木质部面积占比(RXA)、韧皮部面积占比(RPHA)、髓面积占比(RPA)以及树脂道总面积占比(RRC)的统计信息 Table 2Statistical information of wood density (WD), wood nitrogen content (WN), the xylem area-to-total cross-sectional area ratio (RXA), the phloem area-to-total cross-sectional area ratio (RPHA), the pith area-to-total cross-sectional area ratio (RPA) and the total resin canal area-to-total cross-sectional area ratio (RRC)
新窗口打开|下载原图ZIP|生成PPT 图2红松当年生和多年生枝性状随植株大小(胸径或树高)的变异。*, p > 0.05; **, p > 0.01; ***, p > 0.001。
Fig. 2Variations of branch traits in current-year and old branches with plant size (DBH or tree height) for Pinus koraiensis. DBH, diameter at breast height; RPA, pith area-to-total cross-sectional area ratio; RPHA, phloem area-to-total cross-sectional area ratio; RRC, total resin canal area-to-total cross-sectional area ratio; RXA, xylem area-to-total cross-sectional area ratio; WD, wood density; WN, wood nitrogen content. *, p > 0.05; **, p > 0.01; ***, p > 0.001.
Table 3 表3 表3红松枝性状与植株大小、枝龄和环境因素(光照强度、土壤含水量、土壤氮含量和磷含量)之间的广义线性模型(GLM) Table 3Generalized linear models (GLM) among branch traits, tree size, branch age and environment factors (light availability, soil water content, soil nitrogen content, soil phosphorus content) for Pinus koraiensis
性状 Trait
胸径 DBH (cm)
树高 Tree height (m)
枝龄 Branch age (year)
光照强度 Light intensity (mol·m-2·d-1)
土壤含水量 Soil water content (g·g-1)
土壤氮含量 Soil nitrogen content (mg·g-1)
土壤磷含量 Soil phosphorus content (mg·g-1)
截距 Intercept
木质密度 WD (g·cm-3)
0.007**
0.009***
0.003
0.003
-0.001
-0.001
0.393***
枝氮含量 WN (mg·g-1)
1.732***
-0.373
0.121
-0.062
0.302
-0.224
8.881***
木质部面积占比 RXA
-0.006
0.090***
-0.009
0.005
0.020**
-0.019*
0.310***
韧皮部面积占比 RPHA
0.006***
0.007***
-0.005**
0.005*
-0.001
-0.003
0.087***
髓面积占比 RPA
-0.003***
-0.008***
>0.001
>0.001
-0.001
>0.001
0.033***
树脂道总面积占比 RRC
0.011***
-0.008***
0.002
>0.003
-0.003
-0.001
0.067***
DBH, diameter at breast height; RPA, pith area-to-total cross-sectional area ratio; RPHA, phloem area-to-total cross-sectional area ratio; RRC, total resin canal area-to-total cross-sectional area ratio; RXA, xylem area-to-total cross-sectional area ratio; WD, wood density; WN, wood nitrogen content. *, p > 0.05; **, p > 0.01; ***, p > 0.001.
新窗口打开|下载原图ZIP|生成PPT 图3不同植株大小(胸径(DBH)或树高)红松枝性状随枝龄的变异。*, p > 0.05; **, p > 0.01; ***, p > 0.001。
Fig. 3Variations of branch traits of different plant sizes (DBH or tree height) with branch age for Pinus koraiensis. DBH, diameter at breast height; RPA, pith area-to-total cross-sectional area ratio; RPHA, phloem area-to-total cross-sectional area ratio; RRC, total resin canal area-to-total cross-sectional area ratio; RXA, xylem area-to-total cross-sectional area ratio; WD, wood density; WN, wood nitrogen content. *, p > 0.05; **, p > 0.01; ***, p > 0.001.
新窗口打开|下载原图ZIP|生成PPT 图4红松枝性状随枝龄的变化斜率随不同植株大小(DBH或树高)的变异趋势。RPA, 髓面积占比; RPHA, 韧皮部面积占比; RRC, 树脂道总面积占比; RXA, 木质部面积占比; WD, 木质密度。*, p > 0.05; **, p > 0.01。
Fig. 4Variation trend of the slope of branch traits against branch age with different plant sizes (DBH or tree height) for Pinus koraiensis. DBH, diameter at breast height; RPA, pith area-to-total cross-sectional area ratio; RPHA, phloem area-to-total cross-sectional area ratio; RRC, total resin canal area-to-total cross-sectional area ratio; RXA, xylem area-to-total cross-sectional area ratio; WD, wood density. *, p > 0.05; **, p > 0.01.
3 讨论
以往研究表明植株大小、枝龄及环境因素均是引起枝性状变异的主要因素(Rosell et al., 2017; Ka?par et al., 2019), 但很少有研究同时评价这些因素对枝性状的影响程度及差异。本研究表明植株大小(DBH或树高)、枝龄和环境均能独立影响枝性状的变异, 但其影响程度在不同枝性状间存在明显差异。整体而言, 枝龄、植株大小及环境对枝性状的影响依次减小。
AertsR, ChapinFS ( 1999). The mineral nutrition of wild plants revisited: a re-evaluation of processes and patterns Advances in Ecological Research, 30, 1-67. [本文引用: 1]
AgustíJ, BlázquezMA ( 2020). Plant vascular development: mechanisms and environmental regulation Cellular and Molecular Life Sciences. DOI: 10.1007/s00018-020- 03496-w. DOI:10.1007/s00018-020-03714-5URLPMID:33263776 [本文引用: 3] Neurodevelopmental disorders (NDDs), including intellectual disability (ID) and autism spectrum disorders (ASD), are a large group of disorders in which early insults during brain development result in a wide and heterogeneous spectrum of clinical diagnoses. Mutations in genes coding for chromatin remodelers are overrepresented in NDD cohorts, pointing towards epigenetics as a convergent pathogenic pathway between these disorders. In this review we detail the role of NDD-associated chromatin remodelers during the developmental continuum of progenitor expansion, differentiation, cell-type specification, migration and maturation. We discuss how defects in chromatin remodelling during these early developmental time points compound over time and result in impaired brain circuit establishment. In particular, we focus on their role in the three largest cell populations: glutamatergic neurons, GABAergic neurons, and glia cells. An in-depth understanding of the spatiotemporal role of chromatin remodelers during neurodevelopment can contribute to the identification of molecular targets for treatment strategies.
BazotS, FresneauC, DamesinC, BarthesL ( 2016). Contribution of previous year’s leaf N and soil N uptake to current year’s leaf growth in sessile oak Biogeosciences, 13, 3475-3484. DOI:10.5194/bg-13-3475-2016URL [本文引用: 1]
BloomfieldKJ, CernusakLA, EamusD, EllsworthDS, PrenticeIC, WrightIJ, BoerMM, BradfordMG, CaleP, CleverlyJ, EgertonJJG, EvansBJ, HayesLS, HutchinsonMF, LiddellMJ, MacfarlaneC, MeyerWS, ProberSM, TogashiHF, WardlawT, ZhuL, AtkinOK ( 2018). A continental- scale assessment of variabilityin leaf traits: within species, across sites and between seasons Functional Ecology, 32, 1492-1506. DOI:10.1111/fec.2018.32.issue-6URL [本文引用: 1]
BorchertR ( 1994). Soil and stem water storage determine phenology and distribution of tropical dry forest trees Ecology, 75, 1437-1449. DOI:10.2307/1937467URL [本文引用: 1]
BrodribbTJ, FeildTS ( 2000). Stem hydraulic supply is linked to leaf photosynthetic capacity: evidence from New Caledonian and Tasmanian rainforests Plant, Cell & Environment, 23, 1381-1388. [本文引用: 1]
CavaleriMA, OberbauerSF, ClarkDB, ClarkDA, RyanMG ( 2010). Height is more important than light in determining leaf morphology in a tropical forest Ecology, 91, 1730-1739. DOI:10.1890/09-1326.1URLPMID:20583714 [本文引用: 2] Both within and between species, leaf physiological parameters are strongly related to leaf dry mass per area (LMA, g/m2), which has been found to increase from forest floor to canopy top in every forest where it has been measured. Although vertical LMA gradients in forests have historically been attributed to a direct phenotypic response to light, an increasing number of recent studies have provided evidence that water limitation in the upper canopy can constrain foliar morphological adaptations to higher light levels. We measured height, light, and LMA of all species encountered along 45 vertical canopy transects across a Costa Rican tropical rain forest. LMA was correlated with light levels in the lower canopy until approximately 18 m sample height and 22% diffuse transmittance. Height showed a remarkably linear relationship with LMA throughout the entire vertical canopy profile for all species pooled and for each functional group individually (except epiphytes), possibly through the influence of gravity on leaf water potential and turgor pressure. Models of forest function may be greatly simplified by estimating LMA-correlated leaf physiological parameters solely from foliage height profiles, which in turn can be assessed with satellite- and aircraft-based remote sensing.
CorcueraL, CamareroJJ, Gil-PelegrínE ( 2004). Effects of a severe drought on Quercus ilex radial growth and xylem anatomy Trees, 18, 83-92. DOI:10.1007/s00468-003-0284-9URL [本文引用: 1]
DengCY, ZhengJM, ZhangWC, GuoSZ, XueQH, YeLY, SunJW ( 2015). Ecological wood anatomy of Rhizophora stylosa Chinese Journal of Plant Ecology, 39, 604-615. DOI:10.17521/cjpe.2015.0058URL AimsMangrove forest is desirable for studying variations in wood structure along an ecological gradient because mangroves are subjected to considerable habitat changes apart from salt stress within a small area. To elucidate the adaptive capability of wood structures towards fluctuating environmental conditions, variations in wood structures were investigated in 18 individuals of Rhizophora stylosa representing 6 populations along a natural soil physicochemical gradient in the National Nature Reserve of Dongzhai Harbor, Hainan Province.MethodsSoil physicochemical properties were determined at the sites of 18 sampling trees in six R. stylosa populations. The anatomical characteristics of the secondary xylem were studied in details in the 18 trees by means of light microscopy, laser scanning confocal microscopy, scanning electron microscopy and transmission electron microscopy. Variations in the quantitative wood anatomical features in R. stylosa were assessed in details. Relationships between soil physicochemical variables and the quantitative wood anatomical features were analyzed by means of statistical methods.Important findingsSome common specialized wood structures were observed in R. stylosa growing in different habitats, suggesting that these features may function for safely conducting water under high negative pressure and are thus adaptive to intertidal habitats. These common features include the occurrence of: 1) some fibriform vessel elements and a few vasicentric tracheids; 2) vesturing in pits of vessels and helical structures on vessel walls; 3) growth rings; 4) starch grains in ray cells and septate fibers. The quantitative anatomical characteristics have great plasticity in response to heterogeneous habitats. Stepwise regression analyses revealed that the total salt, contents of Mn2+, Na+, Cl-, Ca2+, organic matters, and total phosphorus of soils, and soil pH all have significant effects on quantitative wood anatomical features. Variations in the quantitative wood anatomical features in R. stylosa growing at different sites are adaptive to fluctuating environmental conditions in the intertidal areas.]]> [ 邓传远, 郑俊鸣, 张万超, 郭素枝, 薛秋华, 叶露莹, 孙建文 ( 2015). 红海榄木材结构的生态解剖 植物生态学报, 39, 604-615.] DOI:10.17521/cjpe.2015.0058URL Rhizophora stylosa)是我国红树林生态系统的建群种之一。为了揭示红海榄次生木质部解剖特征可塑性的生态适应意义, 该文测定了海南省东寨港红树林自然保护区6个红海榄种群18个采样点的土壤理化性质, 应用显微镜和电镜详细观测了各采样点生长的18株红海榄植株次生木质部的形态解剖特征, 并测量了红海榄次生木质部的数量解剖特征。不同样地的红海榄次生木质部都具有纤维状导管和环管管胞、螺旋雕纹和附物、生长轮、薄壁细胞(含淀粉粒)等结构。这些特化结构具有生态适应意义, 在潮间带高盐生境中能促进水分输导的安全性。不同生境中红海榄次生木质部数量解剖特征可塑性大, 利用逐步回归分析方法分析了土壤理化因子与红海榄次生木质部数量特征的关系。结果表明: 土壤全盐含量、土壤Mn2+含量、土壤Na+含量、土壤Cl-含量、土壤Ca2+含量、土壤有机质含量、土壤全磷含量和土壤pH值对次生木质部数量特征的影响达到极显著水平, 不同样地红海榄次生木质部数量特征的变化是红海榄适应异质生境的结果。]]>
DomecJC, LachenbruchB, MeinzerFC, WoodruffDR, WarrenJM, McCullohKA ( 2008). Maximum height in a conifer is associated with conflicting requirements for xylem design Proceedings of the National Academy of Sciences of the United States of America, 105, 12069-12074. [本文引用: 2]
EwersBE, OrenR, KimHS, BohrerG, LaiCT ( 2007). Effects of hydraulic architecture and spatial variation in light on mean stomatal conductance of tree branches and crowns. Plant, Cell & Environment, 30, 483-496. [本文引用: 2]
FalsterDS, WestobyM ( 2005). Alternative height strategies among 45 dicot rain forest species from tropical Queensland, Australia Journal of Ecology, 93, 521-535. DOI:10.1890/11-0930.1URLPMID:22624207 [本文引用: 1] Despite the likely importance of inter-year dynamics of plant production and consumer biota for driving community- and ecosystem-level processes, very few studies have explored how and why these dynamics vary across contrasting ecosystems. We utilized a well-characterized system of 30 lake islands in the boreal forest zone of northern Sweden across which soil fertility and productivity vary considerably, with larger islands being more fertile and productive than smaller ones. In this system we assessed the inter-year dynamics of several measures of plant production and the soil microbial community (primary consumers in the decomposer food web) for each of nine years, and soil microfaunal groups (secondary and tertiary consumers) for each of six of those years. We found that, for measures of plant production and each of the three consumer trophic levels, inter-year dynamics were strongly affected by island size. Further, many variables were strongly affected by island size (and thus bottom-up regulation by soil fertility and resources) in some years, but not in other years, most likely due to inter-year variation in climatic conditions. For each of the plant and microbial variables for which we had nine years of data, we also determined the inter-year coefficient of variation (CV), an inverse measure of stability. We found that CVs of some measures of plant productivity were greater on large islands, whereas those of other measures were greater on smaller islands; CVs of microbial variables were unresponsive to island size. We also found that the effects of island size on the temporal dynamics of some variables were related to inter-year variability of macroclimatic variables. As such, our results show that the inter-year dynamics of both plant productivity and decomposer biota across each of three trophic levels, as well as the inter-year stability of plant productivity, differ greatly across contrasting ecosystems, with potentially important but largely overlooked implications for community and ecosystem processes.
FangSZ, SunDY, ShangXL, FuXX, YangWX ( 2020). Variation in radial growth and wood density of Cyclocarya paliurus across its natural distribution New Forests, 51, 453-467. DOI:10.1007/s11056-019-09742-9URL [本文引用: 1]
GebauerR, AlbrechtováP, PlichtaR, Vola?íkD ( 2019). The comparative xylem structure and function of petioles and twigs of mistletoe Loranthus europaeus and its host Quercus pubescence Trees, 33, 933-942. DOI:10.1007/s00468-019-01829-2URL [本文引用: 4]
GleasonSM, WestobyM, JansenS, ChoatB, HackeUG, PrattRB, BhaskarR, BrodribbTJ, BucciSJ, CaoKF, CochardH, DelzonS, DomecJC, FanZX, FeildTS, JacobsenAL, JohnsonDM, LensF, MaheraliH, Martínez-VilaltaJ, MayrS, McCullohKA, MencucciniM, MitchellPJ, MorrisH, NardiniA, PittermannJ, PlavcováL, SchreiberSG, SperryJS, WrightIJ, ZanneAE ( 2016). Weak tradeoff between xylem safety and xylem-specific hydraulic efficiency across the world’s woody plant species New Phytologist, 209, 123-136. DOI:10.1111/nph.13646URL [本文引用: 4]
HackeUG, SperryJS, WheelerJK, CastroL ( 2006). Scaling of angiosperm xylem structure with safety and efficiency Tree Physiology, 26, 689-701. DOI:10.1093/treephys/26.6.689URLPMID:16510385 [本文引用: 1] We tested the hypothesis that greater cavitation resistance correlates with less total inter-vessel pit area per vessel (the pit area hypothesis) and evaluated a trade-off between cavitation safety and transport efficiency. Fourteen species of diverse growth form (vine, ring- and diffuse-porous tree, shrub) and family affinity were added to published data predominately from the Rosaceae (29 species total). Two types of vulnerability-to-cavitation curves were found. Ring-porous trees and vines showed an abrupt drop in hydraulic conductivity with increasing negative pressure, whereas hydraulic conductivity in diffuse-porous species generally decreased gradually. The ring-porous type curve was not an artifact of the centrifuge method because it was obtained also with the air-injection technique. A safety versus efficiency trade-off was evident when curves were compared across species: for a given pressure, there was a limited range of optimal vulnerability curves. The pit area hypothesis was supported by a strong relationship (r2 = 0.77) between increasing cavitation resistance and diminishing pit membrane area per vessel (A(P)). Small A(P) was associated with small vessel surface area and hence narrow vessel diameter (D) and short vessel length (L)--consistent with an increase in vessel flow resistance with cavitation resistance. This trade-off was amplified at the tissue level by an increase in xylem/vessel area ratio with cavitation resistance. Ring-porous species were more efficient than diffuse-porous species on a vessel basis but not on a xylem basis owing to higher xylem/vessel area ratios in ring-porous anatomy. Across four orders of magnitude, lumen and end-wall resistivities maintained a relatively tight proportionality with a near-optimal mean of 56% of the total vessel resistivity residing in the end-wall. This was consistent with an underlying scaling of L to D(3/2) across species. Pit flow resistance did not increase with cavitation safety, suggesting that cavitation pressure was not related to mean pit membrane porosity.
HajekP, HertelD, LeuschnerC ( 2013). Intraspecific variation in root and leaf traits and leaf-root trait linkages in eight aspen demes (Populus tremula and P. tremuloides) Frontiers in Plant Science, 4, 415. DOI: 10.3389/fpls.2013.00415. DOI:10.3389/fpls.2013.00415URLPMID:24155751 [本文引用: 1] Leaf and fine root morphology and physiology have been found to vary considerably among tree species, but not much is known about intraspecific variation in root traits and their relatedness to leaf traits. Various aspen progenies (Populus tremula and P. tremuloides) with different growth performance are used in short-rotation forestry. Hence, a better understanding of the link between root trait syndromes and the adaptation of a deme to a particular environment is essential in order to improve the match between planted varieties and their growth conditions. We examined the between-deme (genetic) and within-deme (mostly environmental) variation in important fine root traits [mean root diameter, specific root area (SRA) and specific root length (SRL), root tissue density (RTD), root tip abundance, root N concentration] and their co-variation with leaf traits [specific leaf area (SLA), leaf size, leaf N concentration] in eight genetically distinct P. tremula and P. tremuloides demes. Five of the six root traits varied significantly between the demes with largest genotypic variation in root tip abundance and lowest in mean root diameter and RTD (no significant difference). Within-deme variation in root morphology was as large as between-deme variation suggesting a relatively low genetic control. Significant relationships existed neither between SLA and SRA nor between leaf N and root N concentration in a plant. Contrary to expectation, high aboveground relative growth rates (RGR) were associated with large, and not small, fine root diameters with low SRA and SRL. Compared to leaf traits, the influence of root traits on RGR was generally low. We conclude that aspen exhibits large intraspecific variation in leaf and also in root morphological traits which is only partly explained by genetic distances. A root order-related analysis might give deeper insights into intraspecific root trait variation.
HeNP, LiuCC, PiaoSL, SackL, XuL, LuoYQ, HeJS, HanXG, ZhouGS, ZhouXH, LinY, YuQ, LiuSR, SunW, NiuSL, LiSG, ZhangJH, YuGR ( 2019). Ecosystem traits linking functional traits to macroecology Trends in Ecology and Evolution, 34, 200-210. DOI:10.1016/j.tree.2018.11.004URLPMID:30527959 [本文引用: 1] As the range of studies on macroecology and functional traits expands, integration of traits into higher-level approaches offers new opportunities to improve clarification of larger-scale patterns and their mechanisms and predictions using models. Here, we propose a framework for quantifying 'ecosystem traits' and means to address the challenges of broadening the applicability of functional traits to macroecology. Ecosystem traits are traits or quantitative characteristics of organisms (plants, animals, and microbes) at the community level expressed as the intensity (or density) normalized per unit land area. Ecosystem traits can inter-relate and integrate data from field trait surveys, eddy-flux observation, remote sensing, and ecological models, and thereby provide new resolution of the responses and feedback at regional to global scale.
HikosakaK, Niinemetsü, AntenNPR ( 2016). Canopy Photosynthesis: from Basics to Applications. Springer, Dordrecht Netherlands. [本文引用: 1]
IsaacME, MartinAR, de Melo Virginio Fiho E, RapidelB, RoupsardO, van den MeerscheK ( 2017). Intraspecific trait variation and coordination: root and leaf economics spectra in coffee across environmental gradients Frontiers in Plant Science, 8, 1196. DOI: 10.3389/fpls.2017.01196. DOI:10.3389/fpls.2017.01196URLPMID:28747919 [本文引用: 1] Hypotheses on the existence of a universal
Ka?parJ, AnfodilloT, TremlV ( 2019). Tree size mostly drives the variation of xylem traits at the treeline ecotone Trees, 33, 1657-1665. DOI:10.1007/s00468-019-01887-6URL [本文引用: 1]
KongGH, LiuSZ, WuT, HuangJ, LinZF, ChenZD, ZhangJZ ( 2006). Differences in photosynthesis and plant growth of 26 woody species at the oil shale waste dump Journal of Tropical and Subtropical Botany, 14, 467-476. [本文引用: 7]
LachenbruchB, McCullohKA ( 2014). Traits, properties, and performance: How woody plants combine hydraulic and mechanical functions in a cell, tissue, or whole plant? New Phytologist, 204, 747-764. DOI:10.1111/nph.13035URL This review presents a framework for evaluating how cells, tissues, organs, and whole plants perform both hydraulic and mechanical functions. The morphological alterations that affect dual functionality are varied: individual cells can have altered morphology; tissues can have altered partitioning to functions or altered cell alignment; and organs and whole plants can differ in their allocation to different tissues, or in the geometric distribution of the tissues they have. A hierarchical model emphasizes that morphological traits influence the hydraulic or mechanical properties; the properties, combined with the plant unit's environment, then influence the performance of that plant unit. As a special case, we discuss the mechanisms by which the proxy property wood density has strong correlations to performance but without direct causality. Traits and properties influence multiple aspects of performance, and there can be mutual compensations such that similar performance occurs. This compensation emphasizes that natural selection acts on, and a plant's viability is determined by, its performance, rather than its contributing traits and properties. Continued research on the relationships among traits, and on their effects on multiple aspects of performance, will help us better predict, manage, and select plant material for success under multiple stresses in the future.
LeytonRBL ( 1972). Reviews: Trees: Structure and Function New Phytologist, 71, 769-770. [本文引用: 1]
LiYN, YangDM, SunSC, GaoXM ( 2008). Effects of twig size on biomass allocation within twigs and on Lamina area supporting efficiency in Rhododendron: allometric scaling analyses Journal of Plant Ecology (Chinese version), 32, 1175-1183. [本文引用: 1]
LillesEB, AstrupR, Lefran?oisML, David CoatesK ( 2014). Sapling leaf trait responses to light, tree height and soil nutrients for three conifer species of contrasting shade tolerance Tree Physiology, 34, 1334-1347. URLPMID:25422385 [本文引用: 1]
LinJX, SampsonD, CeulemansR ( 2001). The effect of crown position and tree age on resin-canal density in Scots pine (Pinus sylvestris L.) needles Canadian Journal of Botany, 79, 1257-1261. DOI:10.1139/b01-108URL
LiuXJ, MaKP ( 2015). Plant functional traits-concepts, applications, and future directions Scientia Sinica Vitae, 45, 325-339. DOI:10.1360/N052014-00244URL [本文引用: 1]
LiuZL, ChenJM, JinGZ, QiYJ ( 2015). Estimating seasonal variations of leaf area index using litterfall collection and optical methods in four mixed evergreen-deciduous forests Agricultural and Forest Meteorology, 209, 36-48. [本文引用: 1]
LiuZL, HikosakaK, LiFR, JinGZ ( 2020). Variations in leaf economics spectrum traits for an evergreen coniferous species: tree size dominates over environment factors Functional Ecology, 34, 458-467. DOI:10.1111/fec.v34.2URL [本文引用: 2]
Martínez-CabreraHI, JonesCS, EspinoS, SchenkHJ ( 2009). Wood anatomy and wood density in shrubs: responses to varying aridity along transcontinental transects American Journal of Botany, 96, 1388-1398. DOI:10.3732/ajb.0800237URLPMID:21628286 [本文引用: 2] Wood density plays a key role in ecological strategies and life history variation in woody plants, but little is known about its anatomical basis in shrubs. We quantified the relationships between wood density, anatomy, and climate in 61 shrub species from eight field sites along latitudinal belts between 31 degrees and 35 degrees in North and South America. Measurements included cell dimensions, transverse areas of each xylem cell type and percentage contact between different cell types and vessels. Wood density was more significantly correlated with precipitation and aridity than with temperature. High wood density was achieved through reductions in cell size and increases in the proportion of wall relative to lumen. Wood density was independent of vessel traits, suggesting that this trait does not impose conduction limitations in shrubs. The proportion of fibers in direct contact with vessels decreased with and was independent of wood density, indicating that the number of fiber-vessel contacts does not explain the previously observed correlation between wood density and implosion resistance. Axial and radial parenchyma each had a significant but opposite association with wood density. Fiber size and wall thickness link wood density, life history, and ecological strategies by controlling the proportion of carbon invested per unit stem volume.
Martinez-CabreraHI, SchenkHJ, Cevallos-FerrizSRS, JonesCS ( 2011). Integration of vessel traits, wood density, and height in angiosperm shrubs and trees American Journal of Botany, 98, 915-922. DOI:10.3732/ajb.1000335URL [本文引用: 3] . Method: In a sample of 200 woody plant species (65 shrubs and 135 trees) from Argentina, Mexico, and the United States, standardized major axis (SMA) regression, correlation analyses, and ANOVA were used to determine whether relationships among traits differed between growth forms. The influence of phylogenetic relationships was examined with a phylogenetic ANOVA and phylogenetically independent contrasts (PICs). A principal component analysis was conducted to determine whether trees and shrubs occupy different portions of multivariate trait space.. Key results: Wood density did not differ between shrubs and trees, but there were significant differences in vessel diameter, vessel density, theoretical conductivity, and as expected, height. In addition, relationships between vessel traits and wood density differed between growth forms. Trees showed coordination among vessel traits, wood density, and height, but in shrubs, wood density and vessel traits were independent. These results hold when phylogenetic relationships were considered. In the multivariate analyses, these differences translated as significantly different positions in multivariate trait space occupied by shrubs and trees.. Conclusions: Differences in trait integration between growth forms suggest that evolution of growth form in some lineages might be associated with the degree of trait interrelation.]]>
MeinzerFC, LachenbruchB, DawsonTE ( 2011). Size- and Age-related Changes in Tree Structure and Function. Springer, Dordrecht Netherlands.
MillardP, GreletGA ( 2010). Nitrogen storage and remobilization by trees: ecophysiological relevance in a changing world Tree Physiology, 30, 1083-1095. URLPMID:20551251 [本文引用: 6]
MiyataR, KuboT, NabeshimaE, KohyamaTS ( 2011). Common allometric response of open-grown leader shoots to tree height in co-occurring deciduous broadleaved trees Annals of Botany, 108, 1279-1286. DOI:10.1093/aob/mcr228URLPMID:21914698 BACKGROUND AND AIMS: Morphology of crown shoots changes with tree height. The height of forest trees is usually correlated with the light environment and this makes it difficult to separate the effects of tree size and of light conditions on the morphological plasticity of crown shoots. This paper addresses the tree-height dependence of shoot traits under full-light conditions where a tree crown is not shaded by other crowns. METHODS: Focus is given to relationships between tree height and top-shoot traits, which include the shoot's leaf-blades and non-leafy mass, its total leaf-blade area and the length and basal diameter of the shoot's stem. We examine the allometric characteristics of open-grown current-year leader shoots at the tops of forest tree crowns up to 24 m high and quantify their responses to tree height in 13 co-occurring deciduous hardwood species in a cool-temperate forest in northern Japan. KEY RESULTS: Dry mass allocated to leaf blades in a leader shoot increased with tree height in all 13 species. Specific leaf area decreased with tree height. Stem basal area was almost proportional to total leaf area in a leader shoot, where the proportionality constant did not depend on tree height, irrespective of species. Stem length for a given stem diameter decreased with tree height. CONCLUSIONS: In the 13 species observed, height-dependent changes in allometry of leader shoots were convergent. This finding suggests that there is a common functional constraint in tree-height development. Under full-light conditions, leader shoots of tall trees naturally experience more severe water stress than those of short trees. We hypothesize that the height dependence of shoot allometry detected reflects an integrated response to height-associated water stress, which contributes to successful crown expansion and height gain.
MolesAT, WartonDI, WarmanL, SwensonNG, LaffanSW, ZanneAE, PitmanAJ, HemmingsFA, LeishmanMR ( 2009). Global patterns in plant height Journal of Ecology, 97, 923-932. DOI:10.1111/jec.2009.97.issue-5URL [本文引用: 2]
Morot-GaudryJF (1997). Assimilation de l’ azote chez les plantes: aspects physiologique, biochimique et moléculaire. INRA Press, Paris. [本文引用: 5]
OlsonME, SorianoD, RosellJA, AnfodilloT, DonoghueMJ, EdwardsEJ, León-GómezC, DawsonT, Camarero MartínezJJ, CastorenaM, EcheverríaA, EspinosaCI, FajardoA, GazolA, IsnardS, LimaRS, MarcatiCR, Méndez-AlonzoR ( 2018). Plant height and hydraulic vulnerability to drought and cold Proceedings of the National Academy of Sciences of the United States of America, 115, 7551-7556. [本文引用: 2]
OsadaN ( 2011). Height-dependent changes in shoot structure and tree allometry in relation to maximum height in four deciduous tree species Functional Ecology, 25, 777-786. DOI:10.1111/j.1365-2435.2011.01833.xURL [本文引用: 3] 2. I investigated height-dependent changes in structure and biomass allocation patterns in current-year shoots of four coexisting tree species differing in H(max) in a cool-temperate forest in Japan. The relative importance of total biomass, biomass allocation, shoot allometry, and shoot angle to vertical growth was quantified and compared with tree allometry.3. Height-dependent changes in total biomass of current-year shoots varied across species. In contrast, stem length per unit mass, shoot angle, and total leaf area per unit stem cross-sectional area decreased, and leaf mass per unit area increased with height in all species. Vertical growth rate consequently declined with increasing height in all species. Sensitivity analyses revealed that the primary determinant of declining vertical growth rate was change in stem length per unit mass; shoot angle was a secondary determinant. In contrast, increases in total shoot mass with height modulated declining vertical growth rates.4. Vertical growth rate was greater in two canopy species than in two sub-canopy species at given heights at the shoot level, and this pattern coincided with allometry between tree height and trunk diameter. In contrast, vertical growth rate was greater in sub-canopy species than in canopy species near their maximum heights. These patterns suggest that allometric differences between species may be useful for evaluating crown-development patterns, but not for estimating H(max) of species.]]>
OsunkoyaOO, ShengTK, MahmudN, DamitN ( 2007). Variation in wood density, wood water content, stem growth and mortality among twenty-seven tree species in a tropical rainforest on Borneo Island Austral Ecology, 32, 191-201. DOI:10.1111/aec.2007.32.issue-2URL [本文引用: 1]
PeltoniemiMS, DuursmaRA, MedlynBE ( 2012). Co-optimal distribution of leaf nitrogen and hydraulic conductance in plant canopies Tree Physiology, 32, 510-519. DOI:10.1093/treephys/tps023URLPMID:22491524 Leaf properties vary significantly within plant canopies, due to the strong gradient in light availability through the canopy, and the need for plants to use resources efficiently. At high light, photosynthesis is maximized when leaves have a high nitrogen content and water supply, whereas at low light leaves have a lower requirement for both nitrogen and water. Studies of the distribution of leaf nitrogen (N) within canopies have shown that, if water supply is ignored, the optimal distribution is that where N is proportional to light, but that the gradient of N in real canopies is shallower than the optimal distribution. We extend this work by considering the optimal co-allocation of nitrogen and water supply within plant canopies. We developed a simple 'toy' two-leaf canopy model and optimized the distribution of N and hydraulic conductance (K) between the two leaves. We asked whether hydraulic constraints to water supply can explain shallow N gradients in canopies. We found that the optimal N distribution within plant canopies is proportional to the light distribution only if hydraulic conductance, K, is also optimally distributed. The optimal distribution of K is that where K and N are both proportional to incident light, such that optimal K is highest to the upper canopy. If the plant is constrained in its ability to construct higher K to sun-exposed leaves, the optimal N distribution does not follow the gradient in light within canopies, but instead follows a shallower gradient. We therefore hypothesize that measured deviations from the predicted optimal distribution of N could be explained by constraints on the distribution of K within canopies. Further empirical research is required on the extent to which plants can construct optimal K distributions, and whether shallow within-canopy N distributions can be explained by sub-optimal K distributions.
PhillipsNG, RyanMG, BondBJ, McDowellNG, HinckleyTM, ?ermákJ ( 2003). Reliance on stored water increases with tree size in three species in the Pacific Northwest Tree Physiology, 23, 237-245. DOI:10.1093/treephys/23.4.237URLPMID:12566259 In tall old forests, limitations to water transport may limit maximum tree height and reduce photosynthesis and carbon sequestration. We evaluated the degree to which tall trees could potentially compensate for hydraulic limitations to water transport by increased use of water stored in xylem. Using sap flux measurements in three tree species of the Pacific Northwest, we showed that reliance on stored water increases with tree size and estimated that use of stored water increases photosynthesis. For Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco), water stored in xylem accounted for 20 to 25% of total daily water use in 60-m trees, whereas stored water comprised 7% of daily water use in 15-m trees. For Oregon white oak (Quercus garryana Dougl. ex Hook.), water stored in xylem accounted for 10 to 23% of total daily water use in 25-m trees, whereas stored water comprised 9 to 13% of daily water use in 10-m trees. For ponderosa pine (Pinus ponderosa Dougl. ex Laws.), water stored in xylem accounted for 4 to 20% of total daily water use in 36-m trees, whereas stored water comprised 2 to 4% of daily water use in 12-m trees. In 60-m Douglas-fir trees, we estimated that use of stored water supported 18% more photosynthesis on a daily basis than would occur if no stored water were used, whereas 15-m Douglas-fir trees gained 10% greater daily photosynthesis from use of stored water. We conclude that water storage plays a significant role in the water and carbon economy of tall trees and old forests.
PoorterL, LianesE, HerasMDL, ZavalaMA ( 2012). Architecture of Iberian canopy tree species in relation to wood density, shade tolerance and climate Plant Ecology, 213, 707-722. DOI:10.1007/s11258-012-0032-6URL [本文引用: 2] Tree architecture has important consequences for tree performance as it determines resource capture, mechanical stability and dominance over competitors. We analyzed architectural relationships between stem and crown dimensions for 13 dominant Iberian canopy tree species belonging to the Pinaceae (six Pinus species) and Fagaceae (six Quercus species and Fagus sylvatica) and related these architectural traits to wood density, shade tolerance and climatic factors. Fagaceae had, compared with Pinaceae, denser wood, saplings with wider crowns and adults with larger maximal crown size but smaller maximal height. In combination, these traits enhance light acquisition and persistence in shaded environments; thus, contributing to their shade tolerance. Pinaceae species, in contrast, had low-density wood, allocate more resources to the formation of the central trunk rather than to branches and attained taller maximal heights, allowing them to grow rapidly in height and compete for light following disturbances; thus, contributing to their high light requirements. Wood density had a strong relationship with tree architecture, with dense-wooded species having smaller maximum height and wider crowns, probably because of cheaper expansion costs for producing biomechanically stable branches. Species from arid environments had shorter stems and shallower crowns for a given stem diameter, probably to reduce hydraulic path length and assure water transport. Wood density is an important correlate of variation in tree architecture between species and the two dominant families, with potentially large implications for their resource foraging strategies and successional dynamics.
PoorterL, McDonaldI, AlarcónA, FichtlerE, LiconaJC, Pe?a-ClarosM, SterckF, VillegasZ, Sass-Klaassenü ( 2010). The importance of wood traits and hydraulic conductance for the performance and life history strategies of 42 rainforest tree species New Phytologist, 185, 481-492. DOI:10.1111/j.1469-8137.2009.03092.xURL [本文引用: 6]
PoorterL, RozendaalDMA, BongersF, Almeida-CortezJS, Almeyda ZambranoAM, álvarezFS, AndradeJL, VillaLFA, BalvaneraP, BecknellJM, BentosTV, BhaskarR, BoukiliV, BrancalionPHS, BroadbentEN , et al. ( 2019). Wet and dry tropical forests show opposite successional pathways in wood density but converge over time Nature Ecology & Evolution, 3, 928-934. DOI:10.1038/s41559-019-0882-6URLPMID:31011177 [本文引用: 1] Tropical forests are converted at an alarming rate for agricultural use and pastureland, but also regrow naturally through secondary succession. For successful forest restoration, it is essential to understand the mechanisms of secondary succession. These mechanisms may vary across forest types, but analyses across broad spatial scales are lacking. Here, we analyse forest recovery using 1,403 plots that differ in age since agricultural abandonment from 50 sites across the Neotropics. We analyse changes in community composition using species-specific stem wood density (WD), which is a key trait for plant growth, survival and forest carbon storage. In wet forest, succession proceeds from low towards high community WD (acquisitive towards conservative trait values), in line with standard successional theory. However, in dry forest, succession proceeds from high towards low community WD (conservative towards acquisitive trait values), probably because high WD reflects drought tolerance in harsh early successional environments. Dry season intensity drives WD recovery by influencing the start and trajectory of succession, resulting in convergence of the community WD over time as vegetation cover builds up. These ecological insights can be used to improve species selection for reforestation. Reforestation species selected to establish a first protective canopy layer should, among other criteria, ideally have a similar WD to the early successional communities that dominate under the prevailing macroclimatic conditions.
PrattRB, BlackRA ( 2006). Do invasive trees have a hydraulic advantage over native trees? Biological Invasions, 8, 1331-1341. DOI:10.1007/s10530-005-0422-yURL [本文引用: 1] The hypothesis was tested that invasive trees have hydraulic traits that contribute to their invasive nature. Five pairs of co-occurring invasive and native trees, in mesic habitats, were selected: (1) Tamarix ramosissima and Salix amygdaloides; (2) Robinia pseudoacacia and Alnus rhombifolia (3) Schinus terebinthifolius and Myrica cerifera; (4) Ligustrum sinense and Acer negundo; and (5) Sapium sebiferum and Diospyros virginiana, respectively. Resistance to cavitation (the water potential [Ψ x ] at 75% loss of hydraulic conductivity [Ψ75]) was not consistently greater for invasive compared to native species (Ψ75=−1.91 and −1.67MPa, respectively). Xylem specific conductivity (K s), a measure of xylem efficiency, was not different between native and invasive species (K s = 3.50 and 3.70kgs−1MPa−1m−1, respectively). The lack of difference for resistance to cavitation among invasive and native species suggests that the sampled invaders are not more tolerant to water stress than co-occurring native species. Apparently the spread and invasive nature of the sampled species cannot be explained by hydraulic traits alone.]]>
PrattRB, JacobsenAL ( 2017). Conflicting demands on angiosperm xylem: tradeoffs among storage, transport and biomechanics. Plant, Cell & Environment, 40, 897-913. DOI:10.1016/0092-8674(85)90349-6URL [本文引用: 4]
PrattRB, JacobsenAL, EwersFW, DavisSD ( 2007). Relationships among xylem transport, biomechanics and storage in stems and roots of nine Rhamnaceae species of the California chaparral New Phytologist, 174, 787-798. DOI:10.1111/nph.2007.174.issue-4URL [本文引用: 2]
PrestonKA, CornwellWK, DenoyerJL ( 2006). Wood density and vessel traits as distinct correlates of ecological strategy in 51 California coast range angiosperms New Phytologist, 170, 807-818. DOI:10.1111/nph.2006.170.issue-4URL [本文引用: 2]
Pulido-RodríguezE, López-CamachoR, TórresJ, VelascoE, Salgado-NegretB ( 2020). Traits and trade-offs of wood anatomy between trunks and branches in tropical dry forest species Trees, 34, 497-505. DOI:10.1007/s00468-019-01931-5URL [本文引用: 2]
R CoreTeam ( 2018). R: a Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna Austria. [本文引用: 4]
RichardC, Lyons-SobaskiS, WiseR ( 2018). Periderm: a concept- based approach to the structure of seed plants//Richard C, Lyons-Sobaski S, Wise R. Plant Anatomy Springer, Cham, Switzerland. [本文引用: 2]
RijkersT, PonsTL, BongersF ( 2000). The effect of tree height and light availability on photosynthetic leaf traits of four neotropical species differing in shade tolerance Functional Ecology, 14, 77-86. DOI:10.1046/j.1365-2435.2000.00395.xURL [本文引用: 2]
Rodriguez-ZaccaroFD, Valdovinos-AyalaJ, PercollaMI, VenturasMD, PrattRB, JacobsenAL ( 2019). Wood structure and function change with maturity: age of the vascular cambium is associated with xylem changes in current-year growth Plant, Cell & Environment, 42, 1816-1831. DOI:10.1111/pce.13528URLPMID:30707440 [本文引用: 3] Xylem vessel structure changes as trees grow and mature. Age- and development-related changes in xylem structure are likely related to changes in hydraulic function. We examined whether hydraulic function, including hydraulic conductivity and vulnerability to water-stress-induced xylem embolism, changed over the course of cambial development in the stems of 17 tree species. We compared current-year growth of young (1-4 years), intermediate (2-7 years), and older (3-10 years) stems occurring in series along branches. Diffuse and ring porous species were examined, but nearly all species produced only diffuse porous xylem in the distal branches that were examined irrespective of their mature xylem porosity type. Vessel diameter and length increased with cambial age. Xylem became both more conductive and more cavitation resistant with cambial age. Ring porous species had longer and wider vessels and xylem that had higher conductivity and was more vulnerable to cavitation; however, these differences between porosity types were not present in young stem samples. Understanding plant hydraulic function and architecture requires the sampling of multiple-aged tissues because plants may vary considerably in their xylem structural and functional traits throughout the plant body, even over relatively short distances and closely aged tissues.
RosellJA, OlsonME, AnfodilloT ( 2017). Scaling of xylem vessel diameter with plant size: causes, predictions, and outstanding questions Current Forestry Reports, 3, 46-59. [本文引用: 1]
SchumannK, LeuschnerC, SchuldtB ( 2019). Xylem hydraulic safety and efficiency in relation to leaf and wood traits in three temperate Acer species differing in habitat preferences Trees, 33, 1475-1490. [本文引用: 1]
SperryJS, MeinzerFC, McCullohKA ( 2008). Safety and efficiency conflicts in hydraulic architecture: scaling from tissues to trees. Plant, Cell & Environment, 31, 632-645. [本文引用: 1]
SterckFJ, BongersF ( 2001). Crown development in tropical rain forest trees: patterns with tree height and light availability Journal of Ecology, 89, 1-13. [本文引用: 3]
StrattonL, GoldsteinG, MeinzerFC ( 2000). Stem water storage capacity and efficiency of water transport: their functional significance in a Hawaiian dry forest. Plant, Cell & Environment, 23, 99-106. [本文引用: 4]
SuzukiM, HiuraT ( 2000). Allometric differences between current-year shoots and large branches of deciduous broad-leaved tree species Tree Physiology, 20, 203-209. DOI:10.1093/treephys/20.3.203URLPMID:12651473 [本文引用: 2] Current-year shoots are mostly made of primary tissues, whereas first-order branches comprise mainly secondary tissues. Differences in tissue composition of these units reflect differences in functional design. We compared the allometry of current-year shoots and first-order branches in eight deciduous broad-leaved tree species and examined the functional differences underlying the design of current-year shoots and first-order branches. Allometric relationships of first-order branches tended to be compatible with predictions of the pipe model and elastic similarity model. That is, allometric constants of the relationships between leaf mass and stem diameter at the branch base and between stem diameter and stem mass were 2.0 and 0.33-0.38, respectively, indicating that the functional regulation of stem form of first-order branches can be predicted by the two models. However, allometric relationships of current-year shoots were not compatible with the predictions of the pipe and elastic similarity models. Thus, the allometric constant of the relationship between leaf mass and stem diameter at the base of current-year shoots was larger than 2.0, and the allometric constant of the relationship between stem length and stem diameter of shoots was larger than 1.0 in all species examined. However, current-year shoots had an allometric constant of leaf mass against stem length that was less than 1.0, suggesting a functional demand on shoot design to reduce self-shading. Also, allometric constants of stem length against stem diameter at the shoot base were larger in monopodial species than in sympodial species, whereas allometric constants of leaf mass per shoot against stem length were smaller in monopodial species than in sympodial species. We propose that the allometries of current-year shoots reflect their function as disposable units for temporary leaf arrangement.
TakashimaT, HikosakaK, HiroseT ( 2004). Photosynthesis or persistence: nitrogen allocation in leaves of evergreen and deciduous Quercus species. Plant, Cell & Environment, 27, 1047-1054. [本文引用: 1]
TarelkinY, HufkensK, HahnS, BulckeJ, BastinJF, IlondeaBA, DebeirO, AckerJ, BeeckmanH, CannièreC ( 2019). Wood anatomy variability under contrasted environmental conditions of common deciduous and evergreen species from central African forests Trees, 33, 893-909. DOI:10.1007/s00468-019-01826-5URL
Virgulino-JúniorPCC, GardunhoDCL, SilvaDNC, FernandesMEB ( 2020). Wood density in mangrove forests on the Brazilian Amazon Coast Trees, 34, 51-60. [本文引用: 4]
WangGL, FaheyTJ, XueS, LiuF ( 2013). Root morphology and architecture respond to N addition in Pinus tabuliformis, west China Oecologia, 171, 583-590. [本文引用: 1]
WangGL, LiuF, XueS ( 2017). Nitrogen addition enhanced water uptake by affecting fine root morphology and coarse root anatomy of Chinese pine seedlings Plant and Soil, 418, 177-189. [本文引用: 2]
WellsteinC, ChelliS, CampetellaG, BarthaS, GalièM, SpadaF, CanulloR ( 2013). Intraspecific phenotypic variability of plant functional traits in contrasting mountain grasslands habitats Biodiversity and Conservation, 22, 2353-2374. [本文引用: 1]
WillGM ( 1971). Nitrogen supply, apical dominance and branch growth in Pinus radiata Plant and Soil, 34, 515-517. [本文引用: 1]
WoodcockDW, ShierAD ( 2003). Does canopy position affect wood specific gravity in temperate forest trees? Annals of Botany, 91, 529-537. [本文引用: 1]
XiangS, WuN, SunSC ( 2009). Within-twig biomass allocation in subtropical evergreen broad-leaved species along an altitudinal gradient: allometric scaling analysis Trees, 23, 637-647. [本文引用: 1]
XiaoWF, XuDY, LiuSR, HanJJ ( 2002). The spatial and temporal characteristics of photosynthesis and transpiration of needles of Chinese fir Scientia Silvae Sinicae, 38(5), 38-46. [本文引用: 1]
XiaoY ( 1995). Horizontal and vertical fluctuation of number and longevity of leaf populations in henry pine (Pinus hbnryi) forest Acta Phytoecologica Sinica, 19, 43-54. [本文引用: 2]
XuZB, LiX, DaiHC, ZhangYP, GuoXF ( 1987). Root distributions of dominant trees in broad-leaved Korean pine forest of Changbai Mountain Chinese Journal of Ecology, 6, 19-24. [本文引用: 1]
YangDM, ZhangJJ, ZhouD, QianMJ, ZhengY, JinLM ( 2012). Leaf and twig functional traits of woody plants and their relationships with environmental change: a review Chinese Journal of Ecology, 31, 702-713. [本文引用: 1]
YangDX, SongL, JinGZ ( 2019a). The soil C:N stoichiometry is more sensitive than the leaf C:N stoichiometry to nitrogen addition: a four-year nitrogen addition experiment in a Pinus koraiensis plantation Plant and Soil, 442, 183-198. [本文引用: 1]
YangSJ, MaoJ, ZuoCW, TianFJ, LiWF, DawudaMM, MaZH, ChenBH ( 2019b). Branch age and angle as crucial drivers of leaf photosynthetic performance and fruiting in high-density planting: a study case in spur-type apple “Vallee Spur” (Malus domestica) Scientia Horticulturae, 246, 898-906. [本文引用: 2]
YangXD, YanER, ChangSX, WangXH, ZhaoYT, ShiQR ( 2014). Twig-leaf size relationships in woody plants vary intraspecifically along a soil moisture gradient Acta Oecologica, 60, 17-25.
ZhangXS, JinGZ, LiuZL ( 2019). Contribution of leaf anatomical traits to leaf mass per area among canopy layers for five coexisting broadleaf species across shade tolerances at a regional scale Forest Ecology and Management, 452, 117569. DOI: 10.1016/j.foreco.2019.117569. [本文引用: 2]
ZhaoPY, FengM, JiaoPP, LiZJ ( 2016). Relationship between morphological or anatomical features of leaves and trunk diameter at breast height at different growing stages of Populus euphratica Arid Zone Research, 33, 1071-1080. [本文引用: 1]
ZiacoE, LiangEY ( 2019). New perspectives on sub-seasonal xylem anatomical responses to climatic variability Trees, 33, 973-975. [本文引用: 2]
ZwienieckiMA, MelcherPJ, FeildTS, HolbrookNM ( 2004). A potential role for xylem-phloem interactions in the hydraulic architecture of trees: effects of phloem girdling on xylem hydraulic conductance Tree Physiology, 24, 911-917.
The mineral nutrition of wild plants revisited: a re-evaluation of processes and patterns 1 1999
... DBH和树高均是表征植株大小的重要变量(Rosell et al., 2017; Ka?par et al., 2019; Fang et al., 2020), 但对枝性状变异的解释程度因性状而异.本研究中WD、RXA、RPHA和RPA对DBH变化更敏感, 而RRC和WN受树高影响更大(表1).树脂道作为存在于针叶树中特有的组织成分, 不仅能够产生及运输树脂, 还具有防御作用(Lin et al., 2001).氮通常是森林生态系统中影响植物生长的限制因子之一(Aerts & Chapin, 2000), 也是决定植被生产力的主要因素(Hikosaka et al., 2016).在温度、降水相对均匀的地区, 光有效性和土壤养分是影响叶性状的主要因素(Lilles et al., 2014), 叶片氮含量的分配是应对光环境的一种重要适应策略, 植物光合作用与叶片氮含量密切相关(Takashima et al., 2004).氮在植物体内是通过根吸收, 通过树干转移至树枝, 进而运输至叶片, 为其提供光合作用所必需的氮(Will, 1971).树高与光有效性密切相关(Sterck & Bongers, 2010; Miyata et al., 2011), 通常随树高增大, 光有效性增强(Woodcock & Shier, 2003; Cavaleri et al., 2010; Liu et al., 2020), 分枝的伸展程度增大(Sterck & Bongers, 2010), 叶片光合作用增强(Rijkers et al., 2000), 因此需要更多的WN供给至叶片以合成所需的光合作用酶(Takashima et al., 2004; Millard & Grelet, 2010).WN和RRC的变化均与垂直运输有关, 因此对树高变化更敏感(表1). ...
Plant vascular development: mechanisms and environmental regulation 3 2020
... 枝是重要的木质器官, 不仅起到机械支撑作用, 还具有水分、养分的运输和存贮功能(Pratt et al., 2007; Poorter et al., 2010; Meinzer et al., 2011; Ziaco & Liang, 2018).性状不同, 其生态功能也存在差异.枝的解剖学研究中, 木质部代表水分运输能力(Pratt & Jacobsen, 2017; Ziaco & Liang, 2018; Gebauer et al., 2019; Schumann et al., 2019); 韧皮部代表光合产物运输能力(Agustí & Blázquez, 2020); 髓是存储养分和水分的重要场所; 树脂道不仅能够产生、运输树脂, 还具有防御功能(Lin et al., 2001).相对于细根和树干, 枝能够更快地向叶片提供养分(Stratton et al., 2000; Pratt & Black, 2006).此外, 枝的解剖性状对于预测树木对环境变化的适应策略具有重要意义(Tarelkin et al., 2019), 然而, 目前关于枝的解剖性状的种内变异规律及影响因素的研究较少(Poorter et al., 2010). ...
... 枝在组织分配中遵循一定的权衡策略(Pratt et al., 2007; 刘晓娟和马克平, 2015; Pratt & Jacobsen, 2017; Pulido-Rodríguez et al., 2020), 不同的组织成分承载的功能不同, 不同组织比例随植株大小的变化反映枝在不同时期的生活史策略.叶片在光合作用过程中需要水来维持气孔导度和CO2吸收(Gleason et al., 2016), 因此, 枝中木质部与叶片光合作用能力显著相关(Brodribb & Feild, 2000).木质部和韧皮部是主要的长距离液体运输通道(Zwieniecki et al., 2004), 分别具有水分运输能力(Pratt & Jacobsen, 2017; Gebauer et al., 2019; Ka?par et al., 2019; Ziaco & Liang, 2019)和光合产物运输能力(Agustí & Blázquez, 2020).水和养分的存储是影响树木健康和生活史策略的重要因素(Pratt & Black, 2006), 髓作为存储养分和水分的重要场所(Stratton et al., 2000), 髓面积大小及髓腔与木质部之间的水力运输能力显著影响叶片水平衡, 进而影响气孔导度限制植物的碳吸收(Phillips et al., 2003).此外, 髓腔中存储的水分比土壤水更靠近水分蒸发流失的位置, 相对于根和树干, 枝能够更快地将水和养分提供至叶片(Stratton et al., 2000).然而DBH作为观测木本植物生长状况的重要指标, 常被用于比较植株的生长速度(孔国辉等, 2006).随DBH增大, WD与RPHA增大, 而RPA减小(图2).WD越高, 植株生长速度减缓, 针叶的光合作用增强(赵鹏宇等, 2016), 其运输光合产物能力增强, 储水能力减弱, 这与以往研究结果一致(Poorter et al., 2019). ...
... 大多数枝性状与枝龄显著相关(p > 0.001, 表3): RPA和RRC随枝龄增大而减小, 在小树中RPA随枝龄增大而减小的速率更大; 相反, 在小树中RRC随枝龄增大而减小的速率更小; WD、RXA和RPHA随枝龄增大而增大(图3).随着枝龄增大, 结构支撑能力增强, 针叶光合能力减弱, 水分和养分存储能力减弱, RXA和RPHA增大, 更好地向形态学上端为不同年龄针叶的光合作用提供所需水分(Gebauer et al., 2019; Schumann et al., 2019), 以及向下运输不同年龄针叶的光合产物(Agustí & Blázquez, 2020).WD和RPHA随DBH增大而增大, 当年生枝的变化速率显著高于多年生枝(图2), 且WD随枝龄增大而增大, 而RPHA随枝龄增大而逐渐减小(图3).当年生针叶的光合作用强于多年生针叶, 随枝龄增长, 枝逐渐从运输器官转变为以机械支撑为主的器官.RXA受DBH大小变化的影响不显著(图2), RXA仅在小树(DBH > 10 cm)中随枝龄增大而增大.以往关于枝的木质部运输能力的研究表明, 相对于导管, 管胞更具有保证水力运输安全性的能力, 并且具有较强的抗空化性, 但水分输送效率较低(Hacke et al., 2006).RXA与植株大小、枝龄无显著相关性; 此外, 以往研究还表明枝的水力运输能力与土壤含水率无相关性(Domec et al., 2008; Sperry et al., 2008; Peltoniemi et al., 2012), 因此, 对于影响RXA的因素仍有待进一步研究. ...
Contribution of previous year’s leaf N and soil N uptake to current year’s leaf growth in sessile oak 1 2016
A continental- scale assessment of variabilityin leaf traits: within species, across sites and between seasons 1 2018
... 掌握植物功能性状的种内变异规律及影响因素, 对深入了解植物的存活、生长和死亡等生态学过程至关重要(Wright et al., 2004; 刘晓娟和马克平, 2015), 也有助于揭示植物的资源利用策略及植物对气候变化的响应机制(He et al., 2019).然而, 以往研究更关注叶性状和细根性状的变异(Hajek et al., 2013; Wellstein et al., 2013; Isaac et al., 2017; Bloomfield et al., 2018), 而对于枝性状种内变异的研究较少. ...
Soil and stem water storage determine phenology and distribution of tropical dry forest trees 1 1994
Stem hydraulic supply is linked to leaf photosynthetic capacity: evidence from New Caledonian and Tasmanian rainforests 1 2000
... 枝在组织分配中遵循一定的权衡策略(Pratt et al., 2007; 刘晓娟和马克平, 2015; Pratt & Jacobsen, 2017; Pulido-Rodríguez et al., 2020), 不同的组织成分承载的功能不同, 不同组织比例随植株大小的变化反映枝在不同时期的生活史策略.叶片在光合作用过程中需要水来维持气孔导度和CO2吸收(Gleason et al., 2016), 因此, 枝中木质部与叶片光合作用能力显著相关(Brodribb & Feild, 2000).木质部和韧皮部是主要的长距离液体运输通道(Zwieniecki et al., 2004), 分别具有水分运输能力(Pratt & Jacobsen, 2017; Gebauer et al., 2019; Ka?par et al., 2019; Ziaco & Liang, 2019)和光合产物运输能力(Agustí & Blázquez, 2020).水和养分的存储是影响树木健康和生活史策略的重要因素(Pratt & Black, 2006), 髓作为存储养分和水分的重要场所(Stratton et al., 2000), 髓面积大小及髓腔与木质部之间的水力运输能力显著影响叶片水平衡, 进而影响气孔导度限制植物的碳吸收(Phillips et al., 2003).此外, 髓腔中存储的水分比土壤水更靠近水分蒸发流失的位置, 相对于根和树干, 枝能够更快地将水和养分提供至叶片(Stratton et al., 2000).然而DBH作为观测木本植物生长状况的重要指标, 常被用于比较植株的生长速度(孔国辉等, 2006).随DBH增大, WD与RPHA增大, 而RPA减小(图2).WD越高, 植株生长速度减缓, 针叶的光合作用增强(赵鹏宇等, 2016), 其运输光合产物能力增强, 储水能力减弱, 这与以往研究结果一致(Poorter et al., 2019). ...
Height is more important than light in determining leaf morphology in a tropical forest 2 2010
... 光照强度、土壤养分和土壤水分等环境因素与许多枝性状的变异也密切相关.例如, WD随光照强度的增加而显著增加(Poorter et al., 2012), 而随土壤含水量的增加而减小(Preston et al., 2006; Yang et al., 2014).自然条件下, 植物所处环境通常随个体大小的变化而发生变化(Moles et al., 2009).例如, 树高通常与光有效性密切相关(Miyata et al., 2011), 即随树高增大, 光有效性呈增加趋势(Woodcock & Shier, 2003; Ewers et al., 2007; Cavaleri et al., 2010; Liu et al., 2020); 植株个体越大, 其根系系统越发达, 具有更强的土壤养分固定能力, 从而增加其土壤养分, 如土壤氮含量(Wang et al., 2013, 2017; Liu et al., 2020).可见, 植株大小(Osunkoya et al., 2007; Meinzer et al., 2011; Rosell et al., 2017; Ka?par et al., 2019)、枝龄(Rodriguez-Zaccaro et al., 2019)及环境(Preston et al., 2006; Poorter et al., 2012)均可能对枝性状的变异产生影响, 但这些因素是否能单独对枝性状产生影响以及对枝性状变异的相对贡献率尚不清晰. ...
... DBH和树高均是表征植株大小的重要变量(Rosell et al., 2017; Ka?par et al., 2019; Fang et al., 2020), 但对枝性状变异的解释程度因性状而异.本研究中WD、RXA、RPHA和RPA对DBH变化更敏感, 而RRC和WN受树高影响更大(表1).树脂道作为存在于针叶树中特有的组织成分, 不仅能够产生及运输树脂, 还具有防御作用(Lin et al., 2001).氮通常是森林生态系统中影响植物生长的限制因子之一(Aerts & Chapin, 2000), 也是决定植被生产力的主要因素(Hikosaka et al., 2016).在温度、降水相对均匀的地区, 光有效性和土壤养分是影响叶性状的主要因素(Lilles et al., 2014), 叶片氮含量的分配是应对光环境的一种重要适应策略, 植物光合作用与叶片氮含量密切相关(Takashima et al., 2004).氮在植物体内是通过根吸收, 通过树干转移至树枝, 进而运输至叶片, 为其提供光合作用所必需的氮(Will, 1971).树高与光有效性密切相关(Sterck & Bongers, 2010; Miyata et al., 2011), 通常随树高增大, 光有效性增强(Woodcock & Shier, 2003; Cavaleri et al., 2010; Liu et al., 2020), 分枝的伸展程度增大(Sterck & Bongers, 2010), 叶片光合作用增强(Rijkers et al., 2000), 因此需要更多的WN供给至叶片以合成所需的光合作用酶(Takashima et al., 2004; Millard & Grelet, 2010).WN和RRC的变化均与垂直运输有关, 因此对树高变化更敏感(表1). ...
Effects of a severe drought on Quercus ilex radial growth and xylem anatomy 1 2004
... 以往关于枝性状变异的研究中多侧重于当年生枝(Xiang et al., 2009; Miyata et al., 2011; Osada, 2011; Rosell et al., 2017; Gebauer et al., 2019).然而, 当年生枝的养分元素、水分和碳水化合物含量等与多年生枝差异显著(杨冬梅等, 2012), 在枝的解剖学研究中, 当年生枝主要由初生组织组成, 而多年生枝主要是次生生长的产物(Leyton, 1972), 当年生枝的初生结构与多年生枝的次生结构在养分元素、结构和功能上存在差异(Suzuki & Hiura, 2000; Richard et al., 2018; Yang et al., 2019b), 但不同年龄枝的次生结构差异不显著.不仅如此, 木质部结构和功能性状在不同枝龄间也存在很大差异(Rodriguez- Zaccaro et al., 2019).被子植物中导管直径随枝龄增大而增大(Corcuera et al., 2004; Rodriguez- Zaccaro et al., 2019), 而裸子植物枝性状随枝龄的变异规律尚不清晰.此外, 对于枝性状的研究通常关注某一生活史阶段(植株大小), 如幼苗、幼树或成年树(Pulido-Rodríguez et al., 2020), 然而, 枝性状在整个树木发育阶段是如何变化的, 以及枝性状随枝龄的变异规律是否受植株大小的影响仍有待检验. ...
红海榄木材结构的生态解剖 2015
红海榄木材结构的生态解剖 2015
Maximum height in a conifer is associated with conflicting requirements for xylem design 2 2008
Effects of hydraulic architecture and spatial variation in light on mean stomatal conductance of tree branches and crowns. Plant, 2 2007
... 植株大小是影响植物功能性状种内变异的主要因素.以往研究表明, 枝性状与植株大小之间存在显著相关性(Falster & Westoby, 2005; Domec et al., 2008; Meinzer et al., 2011; Ka?par et al., 2019).例如, 木质密度(WD)随树高增大而减小(Osunkoya et al., 2007; Martinez-Cabrera et al., 2011; Poorter et al., 2012; Fang et al., 2020); 随着树高增加, 当年生枝的茎长显著减小, 茎质量分数普遍提高(Osada, 2011).一些木质部性状(如导管面积)被认为对环境变化相对不敏感, 但与植株大小密切相关(Ka?par et al., 2019).此外, 以往研究通常采用胸径(DBH)或树高来表征植株大小(Osunkoya et al., 2007; Meinzer et al., 2011; Ka?par et al., 2019; Fang et al., 2020), 但Liu等(2020)的研究表明DBH和树高对红松(Pinus koraiensis)针叶性状的影响存在显著差异, 而DBH和树高对枝性状的影响是否也存在差异还有待验证. ...
... 大多数枝性状与枝龄显著相关(p > 0.001, 表3): RPA和RRC随枝龄增大而减小, 在小树中RPA随枝龄增大而减小的速率更大; 相反, 在小树中RRC随枝龄增大而减小的速率更小; WD、RXA和RPHA随枝龄增大而增大(图3).随着枝龄增大, 结构支撑能力增强, 针叶光合能力减弱, 水分和养分存储能力减弱, RXA和RPHA增大, 更好地向形态学上端为不同年龄针叶的光合作用提供所需水分(Gebauer et al., 2019; Schumann et al., 2019), 以及向下运输不同年龄针叶的光合产物(Agustí & Blázquez, 2020).WD和RPHA随DBH增大而增大, 当年生枝的变化速率显著高于多年生枝(图2), 且WD随枝龄增大而增大, 而RPHA随枝龄增大而逐渐减小(图3).当年生针叶的光合作用强于多年生针叶, 随枝龄增长, 枝逐渐从运输器官转变为以机械支撑为主的器官.RXA受DBH大小变化的影响不显著(图2), RXA仅在小树(DBH > 10 cm)中随枝龄增大而增大.以往关于枝的木质部运输能力的研究表明, 相对于导管, 管胞更具有保证水力运输安全性的能力, 并且具有较强的抗空化性, 但水分输送效率较低(Hacke et al., 2006).RXA与植株大小、枝龄无显著相关性; 此外, 以往研究还表明枝的水力运输能力与土壤含水率无相关性(Domec et al., 2008; Sperry et al., 2008; Peltoniemi et al., 2012), 因此, 对于影响RXA的因素仍有待进一步研究. ...
Alternative height strategies among 45 dicot rain forest species from tropical Queensland, Australia 1 2005
... 光照强度、土壤养分和土壤水分等环境因素与许多枝性状的变异也密切相关.例如, WD随光照强度的增加而显著增加(Poorter et al., 2012), 而随土壤含水量的增加而减小(Preston et al., 2006; Yang et al., 2014).自然条件下, 植物所处环境通常随个体大小的变化而发生变化(Moles et al., 2009).例如, 树高通常与光有效性密切相关(Miyata et al., 2011), 即随树高增大, 光有效性呈增加趋势(Woodcock & Shier, 2003; Ewers et al., 2007; Cavaleri et al., 2010; Liu et al., 2020); 植株个体越大, 其根系系统越发达, 具有更强的土壤养分固定能力, 从而增加其土壤养分, 如土壤氮含量(Wang et al., 2013, 2017; Liu et al., 2020).可见, 植株大小(Osunkoya et al., 2007; Meinzer et al., 2011; Rosell et al., 2017; Ka?par et al., 2019)、枝龄(Rodriguez-Zaccaro et al., 2019)及环境(Preston et al., 2006; Poorter et al., 2012)均可能对枝性状的变异产生影响, 但这些因素是否能单独对枝性状产生影响以及对枝性状变异的相对贡献率尚不清晰. ...
Variation in radial growth and wood density of Cyclocarya paliurus across its natural distribution 1 2020
... 植株大小是影响植物功能性状种内变异的主要因素.以往研究表明, 枝性状与植株大小之间存在显著相关性(Falster & Westoby, 2005; Domec et al., 2008; Meinzer et al., 2011; Ka?par et al., 2019).例如, 木质密度(WD)随树高增大而减小(Osunkoya et al., 2007; Martinez-Cabrera et al., 2011; Poorter et al., 2012; Fang et al., 2020); 随着树高增加, 当年生枝的茎长显著减小, 茎质量分数普遍提高(Osada, 2011).一些木质部性状(如导管面积)被认为对环境变化相对不敏感, 但与植株大小密切相关(Ka?par et al., 2019).此外, 以往研究通常采用胸径(DBH)或树高来表征植株大小(Osunkoya et al., 2007; Meinzer et al., 2011; Ka?par et al., 2019; Fang et al., 2020), 但Liu等(2020)的研究表明DBH和树高对红松(Pinus koraiensis)针叶性状的影响存在显著差异, 而DBH和树高对枝性状的影响是否也存在差异还有待验证. ...
The comparative xylem structure and function of petioles and twigs of mistletoe Loranthus europaeus and its host Quercus pubescence 4 2019
... 植株大小是影响植物功能性状种内变异的主要因素.以往研究表明, 枝性状与植株大小之间存在显著相关性(Falster & Westoby, 2005; Domec et al., 2008; Meinzer et al., 2011; Ka?par et al., 2019).例如, 木质密度(WD)随树高增大而减小(Osunkoya et al., 2007; Martinez-Cabrera et al., 2011; Poorter et al., 2012; Fang et al., 2020); 随着树高增加, 当年生枝的茎长显著减小, 茎质量分数普遍提高(Osada, 2011).一些木质部性状(如导管面积)被认为对环境变化相对不敏感, 但与植株大小密切相关(Ka?par et al., 2019).此外, 以往研究通常采用胸径(DBH)或树高来表征植株大小(Osunkoya et al., 2007; Meinzer et al., 2011; Ka?par et al., 2019; Fang et al., 2020), 但Liu等(2020)的研究表明DBH和树高对红松(Pinus koraiensis)针叶性状的影响存在显著差异, 而DBH和树高对枝性状的影响是否也存在差异还有待验证. ...
... ; Fang et al., 2020), 但Liu等(2020)的研究表明DBH和树高对红松(Pinus koraiensis)针叶性状的影响存在显著差异, 而DBH和树高对枝性状的影响是否也存在差异还有待验证. ...
... DBH和树高均是表征植株大小的重要变量(Rosell et al., 2017; Ka?par et al., 2019; Fang et al., 2020), 但对枝性状变异的解释程度因性状而异.本研究中WD、RXA、RPHA和RPA对DBH变化更敏感, 而RRC和WN受树高影响更大(表1).树脂道作为存在于针叶树中特有的组织成分, 不仅能够产生及运输树脂, 还具有防御作用(Lin et al., 2001).氮通常是森林生态系统中影响植物生长的限制因子之一(Aerts & Chapin, 2000), 也是决定植被生产力的主要因素(Hikosaka et al., 2016).在温度、降水相对均匀的地区, 光有效性和土壤养分是影响叶性状的主要因素(Lilles et al., 2014), 叶片氮含量的分配是应对光环境的一种重要适应策略, 植物光合作用与叶片氮含量密切相关(Takashima et al., 2004).氮在植物体内是通过根吸收, 通过树干转移至树枝, 进而运输至叶片, 为其提供光合作用所必需的氮(Will, 1971).树高与光有效性密切相关(Sterck & Bongers, 2010; Miyata et al., 2011), 通常随树高增大, 光有效性增强(Woodcock & Shier, 2003; Cavaleri et al., 2010; Liu et al., 2020), 分枝的伸展程度增大(Sterck & Bongers, 2010), 叶片光合作用增强(Rijkers et al., 2000), 因此需要更多的WN供给至叶片以合成所需的光合作用酶(Takashima et al., 2004; Millard & Grelet, 2010).WN和RRC的变化均与垂直运输有关, 因此对树高变化更敏感(表1). ...
... 本研究表明WD、RPHA与DBH显著正相关, RPA与DBH显著负相关, 而RRC、WN与树高显著正相关(表3; 图2).以往关于树干WD的研究表明, WD受树高影响程度较大(Osunkoya et al., 2007; Martinez- Cabrera et al., 2011; Poorter et al., 2012; Fang et al., 2020), 且与DBH显著负相关(Virgulino-Júnior et al., 2020).然而WD受DBH的影响大于树高(表1).随着DBH增大, 针叶总数量增加(肖瑜, 1995), 枝的WD显著增大(表3), 枝的结构支撑能力提高, 进一步证明相对于树干, 枝更侧重于对WD的投资以提供水平悬挂的结构支撑, 避免下垂或破损(Pratt & Jacobsen, 2017; Pulido- Rodríguez et al., 2020). ...
Weak tradeoff between xylem safety and xylem-specific hydraulic efficiency across the world’s woody plant species 4 2016
... 枝是重要的木质器官, 不仅起到机械支撑作用, 还具有水分、养分的运输和存贮功能(Pratt et al., 2007; Poorter et al., 2010; Meinzer et al., 2011; Ziaco & Liang, 2018).性状不同, 其生态功能也存在差异.枝的解剖学研究中, 木质部代表水分运输能力(Pratt & Jacobsen, 2017; Ziaco & Liang, 2018; Gebauer et al., 2019; Schumann et al., 2019); 韧皮部代表光合产物运输能力(Agustí & Blázquez, 2020); 髓是存储养分和水分的重要场所; 树脂道不仅能够产生、运输树脂, 还具有防御功能(Lin et al., 2001).相对于细根和树干, 枝能够更快地向叶片提供养分(Stratton et al., 2000; Pratt & Black, 2006).此外, 枝的解剖性状对于预测树木对环境变化的适应策略具有重要意义(Tarelkin et al., 2019), 然而, 目前关于枝的解剖性状的种内变异规律及影响因素的研究较少(Poorter et al., 2010). ...
... 以往关于枝性状变异的研究中多侧重于当年生枝(Xiang et al., 2009; Miyata et al., 2011; Osada, 2011; Rosell et al., 2017; Gebauer et al., 2019).然而, 当年生枝的养分元素、水分和碳水化合物含量等与多年生枝差异显著(杨冬梅等, 2012), 在枝的解剖学研究中, 当年生枝主要由初生组织组成, 而多年生枝主要是次生生长的产物(Leyton, 1972), 当年生枝的初生结构与多年生枝的次生结构在养分元素、结构和功能上存在差异(Suzuki & Hiura, 2000; Richard et al., 2018; Yang et al., 2019b), 但不同年龄枝的次生结构差异不显著.不仅如此, 木质部结构和功能性状在不同枝龄间也存在很大差异(Rodriguez- Zaccaro et al., 2019).被子植物中导管直径随枝龄增大而增大(Corcuera et al., 2004; Rodriguez- Zaccaro et al., 2019), 而裸子植物枝性状随枝龄的变异规律尚不清晰.此外, 对于枝性状的研究通常关注某一生活史阶段(植株大小), 如幼苗、幼树或成年树(Pulido-Rodríguez et al., 2020), 然而, 枝性状在整个树木发育阶段是如何变化的, 以及枝性状随枝龄的变异规律是否受植株大小的影响仍有待检验. ...
... 枝在组织分配中遵循一定的权衡策略(Pratt et al., 2007; 刘晓娟和马克平, 2015; Pratt & Jacobsen, 2017; Pulido-Rodríguez et al., 2020), 不同的组织成分承载的功能不同, 不同组织比例随植株大小的变化反映枝在不同时期的生活史策略.叶片在光合作用过程中需要水来维持气孔导度和CO2吸收(Gleason et al., 2016), 因此, 枝中木质部与叶片光合作用能力显著相关(Brodribb & Feild, 2000).木质部和韧皮部是主要的长距离液体运输通道(Zwieniecki et al., 2004), 分别具有水分运输能力(Pratt & Jacobsen, 2017; Gebauer et al., 2019; Ka?par et al., 2019; Ziaco & Liang, 2019)和光合产物运输能力(Agustí & Blázquez, 2020).水和养分的存储是影响树木健康和生活史策略的重要因素(Pratt & Black, 2006), 髓作为存储养分和水分的重要场所(Stratton et al., 2000), 髓面积大小及髓腔与木质部之间的水力运输能力显著影响叶片水平衡, 进而影响气孔导度限制植物的碳吸收(Phillips et al., 2003).此外, 髓腔中存储的水分比土壤水更靠近水分蒸发流失的位置, 相对于根和树干, 枝能够更快地将水和养分提供至叶片(Stratton et al., 2000).然而DBH作为观测木本植物生长状况的重要指标, 常被用于比较植株的生长速度(孔国辉等, 2006).随DBH增大, WD与RPHA增大, 而RPA减小(图2).WD越高, 植株生长速度减缓, 针叶的光合作用增强(赵鹏宇等, 2016), 其运输光合产物能力增强, 储水能力减弱, 这与以往研究结果一致(Poorter et al., 2019). ...
... 大多数枝性状与枝龄显著相关(p > 0.001, 表3): RPA和RRC随枝龄增大而减小, 在小树中RPA随枝龄增大而减小的速率更大; 相反, 在小树中RRC随枝龄增大而减小的速率更小; WD、RXA和RPHA随枝龄增大而增大(图3).随着枝龄增大, 结构支撑能力增强, 针叶光合能力减弱, 水分和养分存储能力减弱, RXA和RPHA增大, 更好地向形态学上端为不同年龄针叶的光合作用提供所需水分(Gebauer et al., 2019; Schumann et al., 2019), 以及向下运输不同年龄针叶的光合产物(Agustí & Blázquez, 2020).WD和RPHA随DBH增大而增大, 当年生枝的变化速率显著高于多年生枝(图2), 且WD随枝龄增大而增大, 而RPHA随枝龄增大而逐渐减小(图3).当年生针叶的光合作用强于多年生针叶, 随枝龄增长, 枝逐渐从运输器官转变为以机械支撑为主的器官.RXA受DBH大小变化的影响不显著(图2), RXA仅在小树(DBH > 10 cm)中随枝龄增大而增大.以往关于枝的木质部运输能力的研究表明, 相对于导管, 管胞更具有保证水力运输安全性的能力, 并且具有较强的抗空化性, 但水分输送效率较低(Hacke et al., 2006).RXA与植株大小、枝龄无显著相关性; 此外, 以往研究还表明枝的水力运输能力与土壤含水率无相关性(Domec et al., 2008; Sperry et al., 2008; Peltoniemi et al., 2012), 因此, 对于影响RXA的因素仍有待进一步研究. ...
Scaling of angiosperm xylem structure with safety and efficiency 1 2006
... 枝在组织分配中遵循一定的权衡策略(Pratt et al., 2007; 刘晓娟和马克平, 2015; Pratt & Jacobsen, 2017; Pulido-Rodríguez et al., 2020), 不同的组织成分承载的功能不同, 不同组织比例随植株大小的变化反映枝在不同时期的生活史策略.叶片在光合作用过程中需要水来维持气孔导度和CO2吸收(Gleason et al., 2016), 因此, 枝中木质部与叶片光合作用能力显著相关(Brodribb & Feild, 2000).木质部和韧皮部是主要的长距离液体运输通道(Zwieniecki et al., 2004), 分别具有水分运输能力(Pratt & Jacobsen, 2017; Gebauer et al., 2019; Ka?par et al., 2019; Ziaco & Liang, 2019)和光合产物运输能力(Agustí & Blázquez, 2020).水和养分的存储是影响树木健康和生活史策略的重要因素(Pratt & Black, 2006), 髓作为存储养分和水分的重要场所(Stratton et al., 2000), 髓面积大小及髓腔与木质部之间的水力运输能力显著影响叶片水平衡, 进而影响气孔导度限制植物的碳吸收(Phillips et al., 2003).此外, 髓腔中存储的水分比土壤水更靠近水分蒸发流失的位置, 相对于根和树干, 枝能够更快地将水和养分提供至叶片(Stratton et al., 2000).然而DBH作为观测木本植物生长状况的重要指标, 常被用于比较植株的生长速度(孔国辉等, 2006).随DBH增大, WD与RPHA增大, 而RPA减小(图2).WD越高, 植株生长速度减缓, 针叶的光合作用增强(赵鹏宇等, 2016), 其运输光合产物能力增强, 储水能力减弱, 这与以往研究结果一致(Poorter et al., 2019). ...
Intraspecific variation in root and leaf traits and leaf-root trait linkages in eight aspen demes (Populus tremula and P. tremuloides) 1 2013
... 大多数枝性状与枝龄显著相关(p > 0.001, 表3): RPA和RRC随枝龄增大而减小, 在小树中RPA随枝龄增大而减小的速率更大; 相反, 在小树中RRC随枝龄增大而减小的速率更小; WD、RXA和RPHA随枝龄增大而增大(图3).随着枝龄增大, 结构支撑能力增强, 针叶光合能力减弱, 水分和养分存储能力减弱, RXA和RPHA增大, 更好地向形态学上端为不同年龄针叶的光合作用提供所需水分(Gebauer et al., 2019; Schumann et al., 2019), 以及向下运输不同年龄针叶的光合产物(Agustí & Blázquez, 2020).WD和RPHA随DBH增大而增大, 当年生枝的变化速率显著高于多年生枝(图2), 且WD随枝龄增大而增大, 而RPHA随枝龄增大而逐渐减小(图3).当年生针叶的光合作用强于多年生针叶, 随枝龄增长, 枝逐渐从运输器官转变为以机械支撑为主的器官.RXA受DBH大小变化的影响不显著(图2), RXA仅在小树(DBH > 10 cm)中随枝龄增大而增大.以往关于枝的木质部运输能力的研究表明, 相对于导管, 管胞更具有保证水力运输安全性的能力, 并且具有较强的抗空化性, 但水分输送效率较低(Hacke et al., 2006).RXA与植株大小、枝龄无显著相关性; 此外, 以往研究还表明枝的水力运输能力与土壤含水率无相关性(Domec et al., 2008; Sperry et al., 2008; Peltoniemi et al., 2012), 因此, 对于影响RXA的因素仍有待进一步研究. ...
Ecosystem traits linking functional traits to macroecology 1 2019
... 掌握植物功能性状的种内变异规律及影响因素, 对深入了解植物的存活、生长和死亡等生态学过程至关重要(Wright et al., 2004; 刘晓娟和马克平, 2015), 也有助于揭示植物的资源利用策略及植物对气候变化的响应机制(He et al., 2019).然而, 以往研究更关注叶性状和细根性状的变异(Hajek et al., 2013; Wellstein et al., 2013; Isaac et al., 2017; Bloomfield et al., 2018), 而对于枝性状种内变异的研究较少. ...
Canopy Photosynthesis: from Basics to Applications. Springer, Dordrecht 1 2016
... 掌握植物功能性状的种内变异规律及影响因素, 对深入了解植物的存活、生长和死亡等生态学过程至关重要(Wright et al., 2004; 刘晓娟和马克平, 2015), 也有助于揭示植物的资源利用策略及植物对气候变化的响应机制(He et al., 2019).然而, 以往研究更关注叶性状和细根性状的变异(Hajek et al., 2013; Wellstein et al., 2013; Isaac et al., 2017; Bloomfield et al., 2018), 而对于枝性状种内变异的研究较少. ...
Intraspecific trait variation and coordination: root and leaf economics spectra in coffee across environmental gradients 1 2017
... DBH和树高均是表征植株大小的重要变量(Rosell et al., 2017; Ka?par et al., 2019; Fang et al., 2020), 但对枝性状变异的解释程度因性状而异.本研究中WD、RXA、RPHA和RPA对DBH变化更敏感, 而RRC和WN受树高影响更大(表1).树脂道作为存在于针叶树中特有的组织成分, 不仅能够产生及运输树脂, 还具有防御作用(Lin et al., 2001).氮通常是森林生态系统中影响植物生长的限制因子之一(Aerts & Chapin, 2000), 也是决定植被生产力的主要因素(Hikosaka et al., 2016).在温度、降水相对均匀的地区, 光有效性和土壤养分是影响叶性状的主要因素(Lilles et al., 2014), 叶片氮含量的分配是应对光环境的一种重要适应策略, 植物光合作用与叶片氮含量密切相关(Takashima et al., 2004).氮在植物体内是通过根吸收, 通过树干转移至树枝, 进而运输至叶片, 为其提供光合作用所必需的氮(Will, 1971).树高与光有效性密切相关(Sterck & Bongers, 2010; Miyata et al., 2011), 通常随树高增大, 光有效性增强(Woodcock & Shier, 2003; Cavaleri et al., 2010; Liu et al., 2020), 分枝的伸展程度增大(Sterck & Bongers, 2010), 叶片光合作用增强(Rijkers et al., 2000), 因此需要更多的WN供给至叶片以合成所需的光合作用酶(Takashima et al., 2004; Millard & Grelet, 2010).WN和RRC的变化均与垂直运输有关, 因此对树高变化更敏感(表1). ...
Tree size mostly drives the variation of xylem traits at the treeline ecotone 1 2019
... 掌握植物功能性状的种内变异规律及影响因素, 对深入了解植物的存活、生长和死亡等生态学过程至关重要(Wright et al., 2004; 刘晓娟和马克平, 2015), 也有助于揭示植物的资源利用策略及植物对气候变化的响应机制(He et al., 2019).然而, 以往研究更关注叶性状和细根性状的变异(Hajek et al., 2013; Wellstein et al., 2013; Isaac et al., 2017; Bloomfield et al., 2018), 而对于枝性状种内变异的研究较少. ...
油页岩废渣场26种木本植物光合作用和生长的差异 7 2006
... 植株大小是影响植物功能性状种内变异的主要因素.以往研究表明, 枝性状与植株大小之间存在显著相关性(Falster & Westoby, 2005; Domec et al., 2008; Meinzer et al., 2011; Ka?par et al., 2019).例如, 木质密度(WD)随树高增大而减小(Osunkoya et al., 2007; Martinez-Cabrera et al., 2011; Poorter et al., 2012; Fang et al., 2020); 随着树高增加, 当年生枝的茎长显著减小, 茎质量分数普遍提高(Osada, 2011).一些木质部性状(如导管面积)被认为对环境变化相对不敏感, 但与植株大小密切相关(Ka?par et al., 2019).此外, 以往研究通常采用胸径(DBH)或树高来表征植株大小(Osunkoya et al., 2007; Meinzer et al., 2011; Ka?par et al., 2019; Fang et al., 2020), 但Liu等(2020)的研究表明DBH和树高对红松(Pinus koraiensis)针叶性状的影响存在显著差异, 而DBH和树高对枝性状的影响是否也存在差异还有待验证. ...
... ).一些木质部性状(如导管面积)被认为对环境变化相对不敏感, 但与植株大小密切相关(Ka?par et al., 2019).此外, 以往研究通常采用胸径(DBH)或树高来表征植株大小(Osunkoya et al., 2007; Meinzer et al., 2011; Ka?par et al., 2019; Fang et al., 2020), 但Liu等(2020)的研究表明DBH和树高对红松(Pinus koraiensis)针叶性状的影响存在显著差异, 而DBH和树高对枝性状的影响是否也存在差异还有待验证. ...
... ; Ka?par et al., 2019; Fang et al., 2020), 但Liu等(2020)的研究表明DBH和树高对红松(Pinus koraiensis)针叶性状的影响存在显著差异, 而DBH和树高对枝性状的影响是否也存在差异还有待验证. ...
... 光照强度、土壤养分和土壤水分等环境因素与许多枝性状的变异也密切相关.例如, WD随光照强度的增加而显著增加(Poorter et al., 2012), 而随土壤含水量的增加而减小(Preston et al., 2006; Yang et al., 2014).自然条件下, 植物所处环境通常随个体大小的变化而发生变化(Moles et al., 2009).例如, 树高通常与光有效性密切相关(Miyata et al., 2011), 即随树高增大, 光有效性呈增加趋势(Woodcock & Shier, 2003; Ewers et al., 2007; Cavaleri et al., 2010; Liu et al., 2020); 植株个体越大, 其根系系统越发达, 具有更强的土壤养分固定能力, 从而增加其土壤养分, 如土壤氮含量(Wang et al., 2013, 2017; Liu et al., 2020).可见, 植株大小(Osunkoya et al., 2007; Meinzer et al., 2011; Rosell et al., 2017; Ka?par et al., 2019)、枝龄(Rodriguez-Zaccaro et al., 2019)及环境(Preston et al., 2006; Poorter et al., 2012)均可能对枝性状的变异产生影响, 但这些因素是否能单独对枝性状产生影响以及对枝性状变异的相对贡献率尚不清晰. ...
... 以往研究表明植株大小、枝龄及环境因素均是引起枝性状变异的主要因素(Rosell et al., 2017; Ka?par et al., 2019), 但很少有研究同时评价这些因素对枝性状的影响程度及差异.本研究表明植株大小(DBH或树高)、枝龄和环境均能独立影响枝性状的变异, 但其影响程度在不同枝性状间存在明显差异.整体而言, 枝龄、植株大小及环境对枝性状的影响依次减小. ...
... DBH和树高均是表征植株大小的重要变量(Rosell et al., 2017; Ka?par et al., 2019; Fang et al., 2020), 但对枝性状变异的解释程度因性状而异.本研究中WD、RXA、RPHA和RPA对DBH变化更敏感, 而RRC和WN受树高影响更大(表1).树脂道作为存在于针叶树中特有的组织成分, 不仅能够产生及运输树脂, 还具有防御作用(Lin et al., 2001).氮通常是森林生态系统中影响植物生长的限制因子之一(Aerts & Chapin, 2000), 也是决定植被生产力的主要因素(Hikosaka et al., 2016).在温度、降水相对均匀的地区, 光有效性和土壤养分是影响叶性状的主要因素(Lilles et al., 2014), 叶片氮含量的分配是应对光环境的一种重要适应策略, 植物光合作用与叶片氮含量密切相关(Takashima et al., 2004).氮在植物体内是通过根吸收, 通过树干转移至树枝, 进而运输至叶片, 为其提供光合作用所必需的氮(Will, 1971).树高与光有效性密切相关(Sterck & Bongers, 2010; Miyata et al., 2011), 通常随树高增大, 光有效性增强(Woodcock & Shier, 2003; Cavaleri et al., 2010; Liu et al., 2020), 分枝的伸展程度增大(Sterck & Bongers, 2010), 叶片光合作用增强(Rijkers et al., 2000), 因此需要更多的WN供给至叶片以合成所需的光合作用酶(Takashima et al., 2004; Millard & Grelet, 2010).WN和RRC的变化均与垂直运输有关, 因此对树高变化更敏感(表1). ...
... 枝在组织分配中遵循一定的权衡策略(Pratt et al., 2007; 刘晓娟和马克平, 2015; Pratt & Jacobsen, 2017; Pulido-Rodríguez et al., 2020), 不同的组织成分承载的功能不同, 不同组织比例随植株大小的变化反映枝在不同时期的生活史策略.叶片在光合作用过程中需要水来维持气孔导度和CO2吸收(Gleason et al., 2016), 因此, 枝中木质部与叶片光合作用能力显著相关(Brodribb & Feild, 2000).木质部和韧皮部是主要的长距离液体运输通道(Zwieniecki et al., 2004), 分别具有水分运输能力(Pratt & Jacobsen, 2017; Gebauer et al., 2019; Ka?par et al., 2019; Ziaco & Liang, 2019)和光合产物运输能力(Agustí & Blázquez, 2020).水和养分的存储是影响树木健康和生活史策略的重要因素(Pratt & Black, 2006), 髓作为存储养分和水分的重要场所(Stratton et al., 2000), 髓面积大小及髓腔与木质部之间的水力运输能力显著影响叶片水平衡, 进而影响气孔导度限制植物的碳吸收(Phillips et al., 2003).此外, 髓腔中存储的水分比土壤水更靠近水分蒸发流失的位置, 相对于根和树干, 枝能够更快地将水和养分提供至叶片(Stratton et al., 2000).然而DBH作为观测木本植物生长状况的重要指标, 常被用于比较植株的生长速度(孔国辉等, 2006).随DBH增大, WD与RPHA增大, 而RPA减小(图2).WD越高, 植株生长速度减缓, 针叶的光合作用增强(赵鹏宇等, 2016), 其运输光合产物能力增强, 储水能力减弱, 这与以往研究结果一致(Poorter et al., 2019). ...
油页岩废渣场26种木本植物光合作用和生长的差异 7 2006
... 植株大小是影响植物功能性状种内变异的主要因素.以往研究表明, 枝性状与植株大小之间存在显著相关性(Falster & Westoby, 2005; Domec et al., 2008; Meinzer et al., 2011; Ka?par et al., 2019).例如, 木质密度(WD)随树高增大而减小(Osunkoya et al., 2007; Martinez-Cabrera et al., 2011; Poorter et al., 2012; Fang et al., 2020); 随着树高增加, 当年生枝的茎长显著减小, 茎质量分数普遍提高(Osada, 2011).一些木质部性状(如导管面积)被认为对环境变化相对不敏感, 但与植株大小密切相关(Ka?par et al., 2019).此外, 以往研究通常采用胸径(DBH)或树高来表征植株大小(Osunkoya et al., 2007; Meinzer et al., 2011; Ka?par et al., 2019; Fang et al., 2020), 但Liu等(2020)的研究表明DBH和树高对红松(Pinus koraiensis)针叶性状的影响存在显著差异, 而DBH和树高对枝性状的影响是否也存在差异还有待验证. ...
... ).一些木质部性状(如导管面积)被认为对环境变化相对不敏感, 但与植株大小密切相关(Ka?par et al., 2019).此外, 以往研究通常采用胸径(DBH)或树高来表征植株大小(Osunkoya et al., 2007; Meinzer et al., 2011; Ka?par et al., 2019; Fang et al., 2020), 但Liu等(2020)的研究表明DBH和树高对红松(Pinus koraiensis)针叶性状的影响存在显著差异, 而DBH和树高对枝性状的影响是否也存在差异还有待验证. ...
... ; Ka?par et al., 2019; Fang et al., 2020), 但Liu等(2020)的研究表明DBH和树高对红松(Pinus koraiensis)针叶性状的影响存在显著差异, 而DBH和树高对枝性状的影响是否也存在差异还有待验证. ...
... 光照强度、土壤养分和土壤水分等环境因素与许多枝性状的变异也密切相关.例如, WD随光照强度的增加而显著增加(Poorter et al., 2012), 而随土壤含水量的增加而减小(Preston et al., 2006; Yang et al., 2014).自然条件下, 植物所处环境通常随个体大小的变化而发生变化(Moles et al., 2009).例如, 树高通常与光有效性密切相关(Miyata et al., 2011), 即随树高增大, 光有效性呈增加趋势(Woodcock & Shier, 2003; Ewers et al., 2007; Cavaleri et al., 2010; Liu et al., 2020); 植株个体越大, 其根系系统越发达, 具有更强的土壤养分固定能力, 从而增加其土壤养分, 如土壤氮含量(Wang et al., 2013, 2017; Liu et al., 2020).可见, 植株大小(Osunkoya et al., 2007; Meinzer et al., 2011; Rosell et al., 2017; Ka?par et al., 2019)、枝龄(Rodriguez-Zaccaro et al., 2019)及环境(Preston et al., 2006; Poorter et al., 2012)均可能对枝性状的变异产生影响, 但这些因素是否能单独对枝性状产生影响以及对枝性状变异的相对贡献率尚不清晰. ...
... 以往研究表明植株大小、枝龄及环境因素均是引起枝性状变异的主要因素(Rosell et al., 2017; Ka?par et al., 2019), 但很少有研究同时评价这些因素对枝性状的影响程度及差异.本研究表明植株大小(DBH或树高)、枝龄和环境均能独立影响枝性状的变异, 但其影响程度在不同枝性状间存在明显差异.整体而言, 枝龄、植株大小及环境对枝性状的影响依次减小. ...
... DBH和树高均是表征植株大小的重要变量(Rosell et al., 2017; Ka?par et al., 2019; Fang et al., 2020), 但对枝性状变异的解释程度因性状而异.本研究中WD、RXA、RPHA和RPA对DBH变化更敏感, 而RRC和WN受树高影响更大(表1).树脂道作为存在于针叶树中特有的组织成分, 不仅能够产生及运输树脂, 还具有防御作用(Lin et al., 2001).氮通常是森林生态系统中影响植物生长的限制因子之一(Aerts & Chapin, 2000), 也是决定植被生产力的主要因素(Hikosaka et al., 2016).在温度、降水相对均匀的地区, 光有效性和土壤养分是影响叶性状的主要因素(Lilles et al., 2014), 叶片氮含量的分配是应对光环境的一种重要适应策略, 植物光合作用与叶片氮含量密切相关(Takashima et al., 2004).氮在植物体内是通过根吸收, 通过树干转移至树枝, 进而运输至叶片, 为其提供光合作用所必需的氮(Will, 1971).树高与光有效性密切相关(Sterck & Bongers, 2010; Miyata et al., 2011), 通常随树高增大, 光有效性增强(Woodcock & Shier, 2003; Cavaleri et al., 2010; Liu et al., 2020), 分枝的伸展程度增大(Sterck & Bongers, 2010), 叶片光合作用增强(Rijkers et al., 2000), 因此需要更多的WN供给至叶片以合成所需的光合作用酶(Takashima et al., 2004; Millard & Grelet, 2010).WN和RRC的变化均与垂直运输有关, 因此对树高变化更敏感(表1). ...
... 枝在组织分配中遵循一定的权衡策略(Pratt et al., 2007; 刘晓娟和马克平, 2015; Pratt & Jacobsen, 2017; Pulido-Rodríguez et al., 2020), 不同的组织成分承载的功能不同, 不同组织比例随植株大小的变化反映枝在不同时期的生活史策略.叶片在光合作用过程中需要水来维持气孔导度和CO2吸收(Gleason et al., 2016), 因此, 枝中木质部与叶片光合作用能力显著相关(Brodribb & Feild, 2000).木质部和韧皮部是主要的长距离液体运输通道(Zwieniecki et al., 2004), 分别具有水分运输能力(Pratt & Jacobsen, 2017; Gebauer et al., 2019; Ka?par et al., 2019; Ziaco & Liang, 2019)和光合产物运输能力(Agustí & Blázquez, 2020).水和养分的存储是影响树木健康和生活史策略的重要因素(Pratt & Black, 2006), 髓作为存储养分和水分的重要场所(Stratton et al., 2000), 髓面积大小及髓腔与木质部之间的水力运输能力显著影响叶片水平衡, 进而影响气孔导度限制植物的碳吸收(Phillips et al., 2003).此外, 髓腔中存储的水分比土壤水更靠近水分蒸发流失的位置, 相对于根和树干, 枝能够更快地将水和养分提供至叶片(Stratton et al., 2000).然而DBH作为观测木本植物生长状况的重要指标, 常被用于比较植株的生长速度(孔国辉等, 2006).随DBH增大, WD与RPHA增大, 而RPA减小(图2).WD越高, 植株生长速度减缓, 针叶的光合作用增强(赵鹏宇等, 2016), 其运输光合产物能力增强, 储水能力减弱, 这与以往研究结果一致(Poorter et al., 2019). ...
Traits, properties, and performance: How woody plants combine hydraulic and mechanical functions in a cell, tissue, or whole plant? 2014
Reviews: Trees: Structure and Function 1 1972
... 枝在组织分配中遵循一定的权衡策略(Pratt et al., 2007; 刘晓娟和马克平, 2015; Pratt & Jacobsen, 2017; Pulido-Rodríguez et al., 2020), 不同的组织成分承载的功能不同, 不同组织比例随植株大小的变化反映枝在不同时期的生活史策略.叶片在光合作用过程中需要水来维持气孔导度和CO2吸收(Gleason et al., 2016), 因此, 枝中木质部与叶片光合作用能力显著相关(Brodribb & Feild, 2000).木质部和韧皮部是主要的长距离液体运输通道(Zwieniecki et al., 2004), 分别具有水分运输能力(Pratt & Jacobsen, 2017; Gebauer et al., 2019; Ka?par et al., 2019; Ziaco & Liang, 2019)和光合产物运输能力(Agustí & Blázquez, 2020).水和养分的存储是影响树木健康和生活史策略的重要因素(Pratt & Black, 2006), 髓作为存储养分和水分的重要场所(Stratton et al., 2000), 髓面积大小及髓腔与木质部之间的水力运输能力显著影响叶片水平衡, 进而影响气孔导度限制植物的碳吸收(Phillips et al., 2003).此外, 髓腔中存储的水分比土壤水更靠近水分蒸发流失的位置, 相对于根和树干, 枝能够更快地将水和养分提供至叶片(Stratton et al., 2000).然而DBH作为观测木本植物生长状况的重要指标, 常被用于比较植株的生长速度(孔国辉等, 2006).随DBH增大, WD与RPHA增大, 而RPA减小(图2).WD越高, 植株生长速度减缓, 针叶的光合作用增强(赵鹏宇等, 2016), 其运输光合产物能力增强, 储水能力减弱, 这与以往研究结果一致(Poorter et al., 2019). ...
Integration of vessel traits, wood density, and height in angiosperm shrubs and trees 3 2011
... 掌握植物功能性状的种内变异规律及影响因素, 对深入了解植物的存活、生长和死亡等生态学过程至关重要(Wright et al., 2004; 刘晓娟和马克平, 2015), 也有助于揭示植物的资源利用策略及植物对气候变化的响应机制(He et al., 2019).然而, 以往研究更关注叶性状和细根性状的变异(Hajek et al., 2013; Wellstein et al., 2013; Isaac et al., 2017; Bloomfield et al., 2018), 而对于枝性状种内变异的研究较少. ...
... 枝在组织分配中遵循一定的权衡策略(Pratt et al., 2007; 刘晓娟和马克平, 2015; Pratt & Jacobsen, 2017; Pulido-Rodríguez et al., 2020), 不同的组织成分承载的功能不同, 不同组织比例随植株大小的变化反映枝在不同时期的生活史策略.叶片在光合作用过程中需要水来维持气孔导度和CO2吸收(Gleason et al., 2016), 因此, 枝中木质部与叶片光合作用能力显著相关(Brodribb & Feild, 2000).木质部和韧皮部是主要的长距离液体运输通道(Zwieniecki et al., 2004), 分别具有水分运输能力(Pratt & Jacobsen, 2017; Gebauer et al., 2019; Ka?par et al., 2019; Ziaco & Liang, 2019)和光合产物运输能力(Agustí & Blázquez, 2020).水和养分的存储是影响树木健康和生活史策略的重要因素(Pratt & Black, 2006), 髓作为存储养分和水分的重要场所(Stratton et al., 2000), 髓面积大小及髓腔与木质部之间的水力运输能力显著影响叶片水平衡, 进而影响气孔导度限制植物的碳吸收(Phillips et al., 2003).此外, 髓腔中存储的水分比土壤水更靠近水分蒸发流失的位置, 相对于根和树干, 枝能够更快地将水和养分提供至叶片(Stratton et al., 2000).然而DBH作为观测木本植物生长状况的重要指标, 常被用于比较植株的生长速度(孔国辉等, 2006).随DBH增大, WD与RPHA增大, 而RPA减小(图2).WD越高, 植株生长速度减缓, 针叶的光合作用增强(赵鹏宇等, 2016), 其运输光合产物能力增强, 储水能力减弱, 这与以往研究结果一致(Poorter et al., 2019). ...
Size- and Age-related Changes in Tree Structure and Function. Springer, Dordrecht 2011
Nitrogen storage and remobilization by trees: ecophysiological relevance in a changing world 6 2010
... 光照强度、土壤养分和土壤水分等环境因素与许多枝性状的变异也密切相关.例如, WD随光照强度的增加而显著增加(Poorter et al., 2012), 而随土壤含水量的增加而减小(Preston et al., 2006; Yang et al., 2014).自然条件下, 植物所处环境通常随个体大小的变化而发生变化(Moles et al., 2009).例如, 树高通常与光有效性密切相关(Miyata et al., 2011), 即随树高增大, 光有效性呈增加趋势(Woodcock & Shier, 2003; Ewers et al., 2007; Cavaleri et al., 2010; Liu et al., 2020); 植株个体越大, 其根系系统越发达, 具有更强的土壤养分固定能力, 从而增加其土壤养分, 如土壤氮含量(Wang et al., 2013, 2017; Liu et al., 2020).可见, 植株大小(Osunkoya et al., 2007; Meinzer et al., 2011; Rosell et al., 2017; Ka?par et al., 2019)、枝龄(Rodriguez-Zaccaro et al., 2019)及环境(Preston et al., 2006; Poorter et al., 2012)均可能对枝性状的变异产生影响, 但这些因素是否能单独对枝性状产生影响以及对枝性状变异的相对贡献率尚不清晰. ...
... ; Liu et al., 2020).可见, 植株大小(Osunkoya et al., 2007; Meinzer et al., 2011; Rosell et al., 2017; Ka?par et al., 2019)、枝龄(Rodriguez-Zaccaro et al., 2019)及环境(Preston et al., 2006; Poorter et al., 2012)均可能对枝性状的变异产生影响, 但这些因素是否能单独对枝性状产生影响以及对枝性状变异的相对贡献率尚不清晰. ...
... 针对每株样树, 采集样枝前利用半球摄影法(带有180°鱼眼镜头的Nikon Coolpix 4500数码相机, Nikon, Tokyo, Japan)采集半球图片.通过Gap Light Analyzer ver. 2.0软件计算每张半球图片0-60°天顶角范围内的总入射辐射(mol·m-2·d-1), 以该值表征光照强度(Liu et al., 2020). ...
... 对于每株样树, 于树干底部0-10 cm的土层范围内使用土壤取芯器采集土壤子样本, 重复3次且任意两个采样方向之间的角度约为120°, 将这3个子样本进行混合并剔除明显的根和凋落物等杂质(Xu et al., 1987; Yang et al., 2019a; Liu et al., 2020).然后, 利用烘干法测定每个样品的土壤水含量(g·g-1); 采用AQ400自动间断化学分析仪(SEAL Analytical, Mequon, USA)测量全氮含量(mg·g-1)和全磷含量(mg·g-1). ...
... DBH和树高均是表征植株大小的重要变量(Rosell et al., 2017; Ka?par et al., 2019; Fang et al., 2020), 但对枝性状变异的解释程度因性状而异.本研究中WD、RXA、RPHA和RPA对DBH变化更敏感, 而RRC和WN受树高影响更大(表1).树脂道作为存在于针叶树中特有的组织成分, 不仅能够产生及运输树脂, 还具有防御作用(Lin et al., 2001).氮通常是森林生态系统中影响植物生长的限制因子之一(Aerts & Chapin, 2000), 也是决定植被生产力的主要因素(Hikosaka et al., 2016).在温度、降水相对均匀的地区, 光有效性和土壤养分是影响叶性状的主要因素(Lilles et al., 2014), 叶片氮含量的分配是应对光环境的一种重要适应策略, 植物光合作用与叶片氮含量密切相关(Takashima et al., 2004).氮在植物体内是通过根吸收, 通过树干转移至树枝, 进而运输至叶片, 为其提供光合作用所必需的氮(Will, 1971).树高与光有效性密切相关(Sterck & Bongers, 2010; Miyata et al., 2011), 通常随树高增大, 光有效性增强(Woodcock & Shier, 2003; Cavaleri et al., 2010; Liu et al., 2020), 分枝的伸展程度增大(Sterck & Bongers, 2010), 叶片光合作用增强(Rijkers et al., 2000), 因此需要更多的WN供给至叶片以合成所需的光合作用酶(Takashima et al., 2004; Millard & Grelet, 2010).WN和RRC的变化均与垂直运输有关, 因此对树高变化更敏感(表1). ...
Common allometric response of open-grown leader shoots to tree height in co-occurring deciduous broadleaved trees 2011
Global patterns in plant height 2 2009
... 植株大小是影响植物功能性状种内变异的主要因素.以往研究表明, 枝性状与植株大小之间存在显著相关性(Falster & Westoby, 2005; Domec et al., 2008; Meinzer et al., 2011; Ka?par et al., 2019).例如, 木质密度(WD)随树高增大而减小(Osunkoya et al., 2007; Martinez-Cabrera et al., 2011; Poorter et al., 2012; Fang et al., 2020); 随着树高增加, 当年生枝的茎长显著减小, 茎质量分数普遍提高(Osada, 2011).一些木质部性状(如导管面积)被认为对环境变化相对不敏感, 但与植株大小密切相关(Ka?par et al., 2019).此外, 以往研究通常采用胸径(DBH)或树高来表征植株大小(Osunkoya et al., 2007; Meinzer et al., 2011; Ka?par et al., 2019; Fang et al., 2020), 但Liu等(2020)的研究表明DBH和树高对红松(Pinus koraiensis)针叶性状的影响存在显著差异, 而DBH和树高对枝性状的影响是否也存在差异还有待验证. ...
... 本研究表明WD、RPHA与DBH显著正相关, RPA与DBH显著负相关, 而RRC、WN与树高显著正相关(表3; 图2).以往关于树干WD的研究表明, WD受树高影响程度较大(Osunkoya et al., 2007; Martinez- Cabrera et al., 2011; Poorter et al., 2012; Fang et al., 2020), 且与DBH显著负相关(Virgulino-Júnior et al., 2020).然而WD受DBH的影响大于树高(表1).随着DBH增大, 针叶总数量增加(肖瑜, 1995), 枝的WD显著增大(表3), 枝的结构支撑能力提高, 进一步证明相对于树干, 枝更侧重于对WD的投资以提供水平悬挂的结构支撑, 避免下垂或破损(Pratt & Jacobsen, 2017; Pulido- Rodríguez et al., 2020). ...
5 1997
... 枝是重要的木质器官, 不仅起到机械支撑作用, 还具有水分、养分的运输和存贮功能(Pratt et al., 2007; Poorter et al., 2010; Meinzer et al., 2011; Ziaco & Liang, 2018).性状不同, 其生态功能也存在差异.枝的解剖学研究中, 木质部代表水分运输能力(Pratt & Jacobsen, 2017; Ziaco & Liang, 2018; Gebauer et al., 2019; Schumann et al., 2019); 韧皮部代表光合产物运输能力(Agustí & Blázquez, 2020); 髓是存储养分和水分的重要场所; 树脂道不仅能够产生、运输树脂, 还具有防御功能(Lin et al., 2001).相对于细根和树干, 枝能够更快地向叶片提供养分(Stratton et al., 2000; Pratt & Black, 2006).此外, 枝的解剖性状对于预测树木对环境变化的适应策略具有重要意义(Tarelkin et al., 2019), 然而, 目前关于枝的解剖性状的种内变异规律及影响因素的研究较少(Poorter et al., 2010). ...
... 植株大小是影响植物功能性状种内变异的主要因素.以往研究表明, 枝性状与植株大小之间存在显著相关性(Falster & Westoby, 2005; Domec et al., 2008; Meinzer et al., 2011; Ka?par et al., 2019).例如, 木质密度(WD)随树高增大而减小(Osunkoya et al., 2007; Martinez-Cabrera et al., 2011; Poorter et al., 2012; Fang et al., 2020); 随着树高增加, 当年生枝的茎长显著减小, 茎质量分数普遍提高(Osada, 2011).一些木质部性状(如导管面积)被认为对环境变化相对不敏感, 但与植株大小密切相关(Ka?par et al., 2019).此外, 以往研究通常采用胸径(DBH)或树高来表征植株大小(Osunkoya et al., 2007; Meinzer et al., 2011; Ka?par et al., 2019; Fang et al., 2020), 但Liu等(2020)的研究表明DBH和树高对红松(Pinus koraiensis)针叶性状的影响存在显著差异, 而DBH和树高对枝性状的影响是否也存在差异还有待验证. ...
... ; Meinzer et al., 2011; Ka?par et al., 2019; Fang et al., 2020), 但Liu等(2020)的研究表明DBH和树高对红松(Pinus koraiensis)针叶性状的影响存在显著差异, 而DBH和树高对枝性状的影响是否也存在差异还有待验证. ...
... 光照强度、土壤养分和土壤水分等环境因素与许多枝性状的变异也密切相关.例如, WD随光照强度的增加而显著增加(Poorter et al., 2012), 而随土壤含水量的增加而减小(Preston et al., 2006; Yang et al., 2014).自然条件下, 植物所处环境通常随个体大小的变化而发生变化(Moles et al., 2009).例如, 树高通常与光有效性密切相关(Miyata et al., 2011), 即随树高增大, 光有效性呈增加趋势(Woodcock & Shier, 2003; Ewers et al., 2007; Cavaleri et al., 2010; Liu et al., 2020); 植株个体越大, 其根系系统越发达, 具有更强的土壤养分固定能力, 从而增加其土壤养分, 如土壤氮含量(Wang et al., 2013, 2017; Liu et al., 2020).可见, 植株大小(Osunkoya et al., 2007; Meinzer et al., 2011; Rosell et al., 2017; Ka?par et al., 2019)、枝龄(Rodriguez-Zaccaro et al., 2019)及环境(Preston et al., 2006; Poorter et al., 2012)均可能对枝性状的变异产生影响, 但这些因素是否能单独对枝性状产生影响以及对枝性状变异的相对贡献率尚不清晰. ...
Height-dependent changes in shoot structure and tree allometry in relation to maximum height in four deciduous tree species 3 2011
... 以往关于枝性状变异的研究中多侧重于当年生枝(Xiang et al., 2009; Miyata et al., 2011; Osada, 2011; Rosell et al., 2017; Gebauer et al., 2019).然而, 当年生枝的养分元素、水分和碳水化合物含量等与多年生枝差异显著(杨冬梅等, 2012), 在枝的解剖学研究中, 当年生枝主要由初生组织组成, 而多年生枝主要是次生生长的产物(Leyton, 1972), 当年生枝的初生结构与多年生枝的次生结构在养分元素、结构和功能上存在差异(Suzuki & Hiura, 2000; Richard et al., 2018; Yang et al., 2019b), 但不同年龄枝的次生结构差异不显著.不仅如此, 木质部结构和功能性状在不同枝龄间也存在很大差异(Rodriguez- Zaccaro et al., 2019).被子植物中导管直径随枝龄增大而增大(Corcuera et al., 2004; Rodriguez- Zaccaro et al., 2019), 而裸子植物枝性状随枝龄的变异规律尚不清晰.此外, 对于枝性状的研究通常关注某一生活史阶段(植株大小), 如幼苗、幼树或成年树(Pulido-Rodríguez et al., 2020), 然而, 枝性状在整个树木发育阶段是如何变化的, 以及枝性状随枝龄的变异规律是否受植株大小的影响仍有待检验. ...
... 光照强度、土壤养分和土壤水分等环境因素与许多枝性状的变异也密切相关.例如, WD随光照强度的增加而显著增加(Poorter et al., 2012), 而随土壤含水量的增加而减小(Preston et al., 2006; Yang et al., 2014).自然条件下, 植物所处环境通常随个体大小的变化而发生变化(Moles et al., 2009).例如, 树高通常与光有效性密切相关(Miyata et al., 2011), 即随树高增大, 光有效性呈增加趋势(Woodcock & Shier, 2003; Ewers et al., 2007; Cavaleri et al., 2010; Liu et al., 2020); 植株个体越大, 其根系系统越发达, 具有更强的土壤养分固定能力, 从而增加其土壤养分, 如土壤氮含量(Wang et al., 2013, 2017; Liu et al., 2020).可见, 植株大小(Osunkoya et al., 2007; Meinzer et al., 2011; Rosell et al., 2017; Ka?par et al., 2019)、枝龄(Rodriguez-Zaccaro et al., 2019)及环境(Preston et al., 2006; Poorter et al., 2012)均可能对枝性状的变异产生影响, 但这些因素是否能单独对枝性状产生影响以及对枝性状变异的相对贡献率尚不清晰. ...
... DBH和树高均是表征植株大小的重要变量(Rosell et al., 2017; Ka?par et al., 2019; Fang et al., 2020), 但对枝性状变异的解释程度因性状而异.本研究中WD、RXA、RPHA和RPA对DBH变化更敏感, 而RRC和WN受树高影响更大(表1).树脂道作为存在于针叶树中特有的组织成分, 不仅能够产生及运输树脂, 还具有防御作用(Lin et al., 2001).氮通常是森林生态系统中影响植物生长的限制因子之一(Aerts & Chapin, 2000), 也是决定植被生产力的主要因素(Hikosaka et al., 2016).在温度、降水相对均匀的地区, 光有效性和土壤养分是影响叶性状的主要因素(Lilles et al., 2014), 叶片氮含量的分配是应对光环境的一种重要适应策略, 植物光合作用与叶片氮含量密切相关(Takashima et al., 2004).氮在植物体内是通过根吸收, 通过树干转移至树枝, 进而运输至叶片, 为其提供光合作用所必需的氮(Will, 1971).树高与光有效性密切相关(Sterck & Bongers, 2010; Miyata et al., 2011), 通常随树高增大, 光有效性增强(Woodcock & Shier, 2003; Cavaleri et al., 2010; Liu et al., 2020), 分枝的伸展程度增大(Sterck & Bongers, 2010), 叶片光合作用增强(Rijkers et al., 2000), 因此需要更多的WN供给至叶片以合成所需的光合作用酶(Takashima et al., 2004; Millard & Grelet, 2010).WN和RRC的变化均与垂直运输有关, 因此对树高变化更敏感(表1). ...
Variation in wood density, wood water content, stem growth and mortality among twenty-seven tree species in a tropical rainforest on Borneo Island 1 2007
... 光照强度、土壤养分和土壤水分等环境因素与许多枝性状的变异也密切相关.例如, WD随光照强度的增加而显著增加(Poorter et al., 2012), 而随土壤含水量的增加而减小(Preston et al., 2006; Yang et al., 2014).自然条件下, 植物所处环境通常随个体大小的变化而发生变化(Moles et al., 2009).例如, 树高通常与光有效性密切相关(Miyata et al., 2011), 即随树高增大, 光有效性呈增加趋势(Woodcock & Shier, 2003; Ewers et al., 2007; Cavaleri et al., 2010; Liu et al., 2020); 植株个体越大, 其根系系统越发达, 具有更强的土壤养分固定能力, 从而增加其土壤养分, 如土壤氮含量(Wang et al., 2013, 2017; Liu et al., 2020).可见, 植株大小(Osunkoya et al., 2007; Meinzer et al., 2011; Rosell et al., 2017; Ka?par et al., 2019)、枝龄(Rodriguez-Zaccaro et al., 2019)及环境(Preston et al., 2006; Poorter et al., 2012)均可能对枝性状的变异产生影响, 但这些因素是否能单独对枝性状产生影响以及对枝性状变异的相对贡献率尚不清晰. ...
Co-optimal distribution of leaf nitrogen and hydraulic conductance in plant canopies 2012
Reliance on stored water increases with tree size in three species in the Pacific Northwest 2003
Architecture of Iberian canopy tree species in relation to wood density, shade tolerance and climate 2 2012
... 植株大小是影响植物功能性状种内变异的主要因素.以往研究表明, 枝性状与植株大小之间存在显著相关性(Falster & Westoby, 2005; Domec et al., 2008; Meinzer et al., 2011; Ka?par et al., 2019).例如, 木质密度(WD)随树高增大而减小(Osunkoya et al., 2007; Martinez-Cabrera et al., 2011; Poorter et al., 2012; Fang et al., 2020); 随着树高增加, 当年生枝的茎长显著减小, 茎质量分数普遍提高(Osada, 2011).一些木质部性状(如导管面积)被认为对环境变化相对不敏感, 但与植株大小密切相关(Ka?par et al., 2019).此外, 以往研究通常采用胸径(DBH)或树高来表征植株大小(Osunkoya et al., 2007; Meinzer et al., 2011; Ka?par et al., 2019; Fang et al., 2020), 但Liu等(2020)的研究表明DBH和树高对红松(Pinus koraiensis)针叶性状的影响存在显著差异, 而DBH和树高对枝性状的影响是否也存在差异还有待验证. ...
... 以往关于枝性状变异的研究中多侧重于当年生枝(Xiang et al., 2009; Miyata et al., 2011; Osada, 2011; Rosell et al., 2017; Gebauer et al., 2019).然而, 当年生枝的养分元素、水分和碳水化合物含量等与多年生枝差异显著(杨冬梅等, 2012), 在枝的解剖学研究中, 当年生枝主要由初生组织组成, 而多年生枝主要是次生生长的产物(Leyton, 1972), 当年生枝的初生结构与多年生枝的次生结构在养分元素、结构和功能上存在差异(Suzuki & Hiura, 2000; Richard et al., 2018; Yang et al., 2019b), 但不同年龄枝的次生结构差异不显著.不仅如此, 木质部结构和功能性状在不同枝龄间也存在很大差异(Rodriguez- Zaccaro et al., 2019).被子植物中导管直径随枝龄增大而增大(Corcuera et al., 2004; Rodriguez- Zaccaro et al., 2019), 而裸子植物枝性状随枝龄的变异规律尚不清晰.此外, 对于枝性状的研究通常关注某一生活史阶段(植株大小), 如幼苗、幼树或成年树(Pulido-Rodríguez et al., 2020), 然而, 枝性状在整个树木发育阶段是如何变化的, 以及枝性状随枝龄的变异规律是否受植株大小的影响仍有待检验. ...
The importance of wood traits and hydraulic conductance for the performance and life history strategies of 42 rainforest tree species 6 2010
... 植株大小是影响植物功能性状种内变异的主要因素.以往研究表明, 枝性状与植株大小之间存在显著相关性(Falster & Westoby, 2005; Domec et al., 2008; Meinzer et al., 2011; Ka?par et al., 2019).例如, 木质密度(WD)随树高增大而减小(Osunkoya et al., 2007; Martinez-Cabrera et al., 2011; Poorter et al., 2012; Fang et al., 2020); 随着树高增加, 当年生枝的茎长显著减小, 茎质量分数普遍提高(Osada, 2011).一些木质部性状(如导管面积)被认为对环境变化相对不敏感, 但与植株大小密切相关(Ka?par et al., 2019).此外, 以往研究通常采用胸径(DBH)或树高来表征植株大小(Osunkoya et al., 2007; Meinzer et al., 2011; Ka?par et al., 2019; Fang et al., 2020), 但Liu等(2020)的研究表明DBH和树高对红松(Pinus koraiensis)针叶性状的影响存在显著差异, 而DBH和树高对枝性状的影响是否也存在差异还有待验证. ...
... )或树高来表征植株大小(Osunkoya et al., 2007; Meinzer et al., 2011; Ka?par et al., 2019; Fang et al., 2020), 但Liu等(2020)的研究表明DBH和树高对红松(Pinus koraiensis)针叶性状的影响存在显著差异, 而DBH和树高对枝性状的影响是否也存在差异还有待验证. ...
... 光照强度、土壤养分和土壤水分等环境因素与许多枝性状的变异也密切相关.例如, WD随光照强度的增加而显著增加(Poorter et al., 2012), 而随土壤含水量的增加而减小(Preston et al., 2006; Yang et al., 2014).自然条件下, 植物所处环境通常随个体大小的变化而发生变化(Moles et al., 2009).例如, 树高通常与光有效性密切相关(Miyata et al., 2011), 即随树高增大, 光有效性呈增加趋势(Woodcock & Shier, 2003; Ewers et al., 2007; Cavaleri et al., 2010; Liu et al., 2020); 植株个体越大, 其根系系统越发达, 具有更强的土壤养分固定能力, 从而增加其土壤养分, 如土壤氮含量(Wang et al., 2013, 2017; Liu et al., 2020).可见, 植株大小(Osunkoya et al., 2007; Meinzer et al., 2011; Rosell et al., 2017; Ka?par et al., 2019)、枝龄(Rodriguez-Zaccaro et al., 2019)及环境(Preston et al., 2006; Poorter et al., 2012)均可能对枝性状的变异产生影响, 但这些因素是否能单独对枝性状产生影响以及对枝性状变异的相对贡献率尚不清晰. ...
... DBH和树高均是表征植株大小的重要变量(Rosell et al., 2017; Ka?par et al., 2019; Fang et al., 2020), 但对枝性状变异的解释程度因性状而异.本研究中WD、RXA、RPHA和RPA对DBH变化更敏感, 而RRC和WN受树高影响更大(表1).树脂道作为存在于针叶树中特有的组织成分, 不仅能够产生及运输树脂, 还具有防御作用(Lin et al., 2001).氮通常是森林生态系统中影响植物生长的限制因子之一(Aerts & Chapin, 2000), 也是决定植被生产力的主要因素(Hikosaka et al., 2016).在温度、降水相对均匀的地区, 光有效性和土壤养分是影响叶性状的主要因素(Lilles et al., 2014), 叶片氮含量的分配是应对光环境的一种重要适应策略, 植物光合作用与叶片氮含量密切相关(Takashima et al., 2004).氮在植物体内是通过根吸收, 通过树干转移至树枝, 进而运输至叶片, 为其提供光合作用所必需的氮(Will, 1971).树高与光有效性密切相关(Sterck & Bongers, 2010; Miyata et al., 2011), 通常随树高增大, 光有效性增强(Woodcock & Shier, 2003; Cavaleri et al., 2010; Liu et al., 2020), 分枝的伸展程度增大(Sterck & Bongers, 2010), 叶片光合作用增强(Rijkers et al., 2000), 因此需要更多的WN供给至叶片以合成所需的光合作用酶(Takashima et al., 2004; Millard & Grelet, 2010).WN和RRC的变化均与垂直运输有关, 因此对树高变化更敏感(表1). ...
... ), 分枝的伸展程度增大(Sterck & Bongers, 2010), 叶片光合作用增强(Rijkers et al., 2000), 因此需要更多的WN供给至叶片以合成所需的光合作用酶(Takashima et al., 2004; Millard & Grelet, 2010).WN和RRC的变化均与垂直运输有关, 因此对树高变化更敏感(表1). ...
... 本研究表明WD、RPHA与DBH显著正相关, RPA与DBH显著负相关, 而RRC、WN与树高显著正相关(表3; 图2).以往关于树干WD的研究表明, WD受树高影响程度较大(Osunkoya et al., 2007; Martinez- Cabrera et al., 2011; Poorter et al., 2012; Fang et al., 2020), 且与DBH显著负相关(Virgulino-Júnior et al., 2020).然而WD受DBH的影响大于树高(表1).随着DBH增大, 针叶总数量增加(肖瑜, 1995), 枝的WD显著增大(表3), 枝的结构支撑能力提高, 进一步证明相对于树干, 枝更侧重于对WD的投资以提供水平悬挂的结构支撑, 避免下垂或破损(Pratt & Jacobsen, 2017; Pulido- Rodríguez et al., 2020). ...
Wet and dry tropical forests show opposite successional pathways in wood density but converge over time 1 2019
... 大多数枝性状与枝龄显著相关(p > 0.001, 表3): RPA和RRC随枝龄增大而减小, 在小树中RPA随枝龄增大而减小的速率更大; 相反, 在小树中RRC随枝龄增大而减小的速率更小; WD、RXA和RPHA随枝龄增大而增大(图3).随着枝龄增大, 结构支撑能力增强, 针叶光合能力减弱, 水分和养分存储能力减弱, RXA和RPHA增大, 更好地向形态学上端为不同年龄针叶的光合作用提供所需水分(Gebauer et al., 2019; Schumann et al., 2019), 以及向下运输不同年龄针叶的光合产物(Agustí & Blázquez, 2020).WD和RPHA随DBH增大而增大, 当年生枝的变化速率显著高于多年生枝(图2), 且WD随枝龄增大而增大, 而RPHA随枝龄增大而逐渐减小(图3).当年生针叶的光合作用强于多年生针叶, 随枝龄增长, 枝逐渐从运输器官转变为以机械支撑为主的器官.RXA受DBH大小变化的影响不显著(图2), RXA仅在小树(DBH > 10 cm)中随枝龄增大而增大.以往关于枝的木质部运输能力的研究表明, 相对于导管, 管胞更具有保证水力运输安全性的能力, 并且具有较强的抗空化性, 但水分输送效率较低(Hacke et al., 2006).RXA与植株大小、枝龄无显著相关性; 此外, 以往研究还表明枝的水力运输能力与土壤含水率无相关性(Domec et al., 2008; Sperry et al., 2008; Peltoniemi et al., 2012), 因此, 对于影响RXA的因素仍有待进一步研究. ...
Do invasive trees have a hydraulic advantage over native trees? 1 2006
... 枝在组织分配中遵循一定的权衡策略(Pratt et al., 2007; 刘晓娟和马克平, 2015; Pratt & Jacobsen, 2017; Pulido-Rodríguez et al., 2020), 不同的组织成分承载的功能不同, 不同组织比例随植株大小的变化反映枝在不同时期的生活史策略.叶片在光合作用过程中需要水来维持气孔导度和CO2吸收(Gleason et al., 2016), 因此, 枝中木质部与叶片光合作用能力显著相关(Brodribb & Feild, 2000).木质部和韧皮部是主要的长距离液体运输通道(Zwieniecki et al., 2004), 分别具有水分运输能力(Pratt & Jacobsen, 2017; Gebauer et al., 2019; Ka?par et al., 2019; Ziaco & Liang, 2019)和光合产物运输能力(Agustí & Blázquez, 2020).水和养分的存储是影响树木健康和生活史策略的重要因素(Pratt & Black, 2006), 髓作为存储养分和水分的重要场所(Stratton et al., 2000), 髓面积大小及髓腔与木质部之间的水力运输能力显著影响叶片水平衡, 进而影响气孔导度限制植物的碳吸收(Phillips et al., 2003).此外, 髓腔中存储的水分比土壤水更靠近水分蒸发流失的位置, 相对于根和树干, 枝能够更快地将水和养分提供至叶片(Stratton et al., 2000).然而DBH作为观测木本植物生长状况的重要指标, 常被用于比较植株的生长速度(孔国辉等, 2006).随DBH增大, WD与RPHA增大, 而RPA减小(图2).WD越高, 植株生长速度减缓, 针叶的光合作用增强(赵鹏宇等, 2016), 其运输光合产物能力增强, 储水能力减弱, 这与以往研究结果一致(Poorter et al., 2019). ...
Conflicting demands on angiosperm xylem: tradeoffs among storage, transport and biomechanics. Plant, 4 2017
... 植株大小是影响植物功能性状种内变异的主要因素.以往研究表明, 枝性状与植株大小之间存在显著相关性(Falster & Westoby, 2005; Domec et al., 2008; Meinzer et al., 2011; Ka?par et al., 2019).例如, 木质密度(WD)随树高增大而减小(Osunkoya et al., 2007; Martinez-Cabrera et al., 2011; Poorter et al., 2012; Fang et al., 2020); 随着树高增加, 当年生枝的茎长显著减小, 茎质量分数普遍提高(Osada, 2011).一些木质部性状(如导管面积)被认为对环境变化相对不敏感, 但与植株大小密切相关(Ka?par et al., 2019).此外, 以往研究通常采用胸径(DBH)或树高来表征植株大小(Osunkoya et al., 2007; Meinzer et al., 2011; Ka?par et al., 2019; Fang et al., 2020), 但Liu等(2020)的研究表明DBH和树高对红松(Pinus koraiensis)针叶性状的影响存在显著差异, 而DBH和树高对枝性状的影响是否也存在差异还有待验证. ...
... 光照强度、土壤养分和土壤水分等环境因素与许多枝性状的变异也密切相关.例如, WD随光照强度的增加而显著增加(Poorter et al., 2012), 而随土壤含水量的增加而减小(Preston et al., 2006; Yang et al., 2014).自然条件下, 植物所处环境通常随个体大小的变化而发生变化(Moles et al., 2009).例如, 树高通常与光有效性密切相关(Miyata et al., 2011), 即随树高增大, 光有效性呈增加趋势(Woodcock & Shier, 2003; Ewers et al., 2007; Cavaleri et al., 2010; Liu et al., 2020); 植株个体越大, 其根系系统越发达, 具有更强的土壤养分固定能力, 从而增加其土壤养分, 如土壤氮含量(Wang et al., 2013, 2017; Liu et al., 2020).可见, 植株大小(Osunkoya et al., 2007; Meinzer et al., 2011; Rosell et al., 2017; Ka?par et al., 2019)、枝龄(Rodriguez-Zaccaro et al., 2019)及环境(Preston et al., 2006; Poorter et al., 2012)均可能对枝性状的变异产生影响, 但这些因素是否能单独对枝性状产生影响以及对枝性状变异的相对贡献率尚不清晰. ...
... ; Poorter et al., 2012)均可能对枝性状的变异产生影响, 但这些因素是否能单独对枝性状产生影响以及对枝性状变异的相对贡献率尚不清晰. ...
... 本研究表明WD、RPHA与DBH显著正相关, RPA与DBH显著负相关, 而RRC、WN与树高显著正相关(表3; 图2).以往关于树干WD的研究表明, WD受树高影响程度较大(Osunkoya et al., 2007; Martinez- Cabrera et al., 2011; Poorter et al., 2012; Fang et al., 2020), 且与DBH显著负相关(Virgulino-Júnior et al., 2020).然而WD受DBH的影响大于树高(表1).随着DBH增大, 针叶总数量增加(肖瑜, 1995), 枝的WD显著增大(表3), 枝的结构支撑能力提高, 进一步证明相对于树干, 枝更侧重于对WD的投资以提供水平悬挂的结构支撑, 避免下垂或破损(Pratt & Jacobsen, 2017; Pulido- Rodríguez et al., 2020). ...
Relationships among xylem transport, biomechanics and storage in stems and roots of nine Rhamnaceae species of the California chaparral 2 2007
... 枝是重要的木质器官, 不仅起到机械支撑作用, 还具有水分、养分的运输和存贮功能(Pratt et al., 2007; Poorter et al., 2010; Meinzer et al., 2011; Ziaco & Liang, 2018).性状不同, 其生态功能也存在差异.枝的解剖学研究中, 木质部代表水分运输能力(Pratt & Jacobsen, 2017; Ziaco & Liang, 2018; Gebauer et al., 2019; Schumann et al., 2019); 韧皮部代表光合产物运输能力(Agustí & Blázquez, 2020); 髓是存储养分和水分的重要场所; 树脂道不仅能够产生、运输树脂, 还具有防御功能(Lin et al., 2001).相对于细根和树干, 枝能够更快地向叶片提供养分(Stratton et al., 2000; Pratt & Black, 2006).此外, 枝的解剖性状对于预测树木对环境变化的适应策略具有重要意义(Tarelkin et al., 2019), 然而, 目前关于枝的解剖性状的种内变异规律及影响因素的研究较少(Poorter et al., 2010). ...
... ), 然而, 目前关于枝的解剖性状的种内变异规律及影响因素的研究较少(Poorter et al., 2010). ...
Wood density and vessel traits as distinct correlates of ecological strategy in 51 California coast range angiosperms 2 2006
... 枝在组织分配中遵循一定的权衡策略(Pratt et al., 2007; 刘晓娟和马克平, 2015; Pratt & Jacobsen, 2017; Pulido-Rodríguez et al., 2020), 不同的组织成分承载的功能不同, 不同组织比例随植株大小的变化反映枝在不同时期的生活史策略.叶片在光合作用过程中需要水来维持气孔导度和CO2吸收(Gleason et al., 2016), 因此, 枝中木质部与叶片光合作用能力显著相关(Brodribb & Feild, 2000).木质部和韧皮部是主要的长距离液体运输通道(Zwieniecki et al., 2004), 分别具有水分运输能力(Pratt & Jacobsen, 2017; Gebauer et al., 2019; Ka?par et al., 2019; Ziaco & Liang, 2019)和光合产物运输能力(Agustí & Blázquez, 2020).水和养分的存储是影响树木健康和生活史策略的重要因素(Pratt & Black, 2006), 髓作为存储养分和水分的重要场所(Stratton et al., 2000), 髓面积大小及髓腔与木质部之间的水力运输能力显著影响叶片水平衡, 进而影响气孔导度限制植物的碳吸收(Phillips et al., 2003).此外, 髓腔中存储的水分比土壤水更靠近水分蒸发流失的位置, 相对于根和树干, 枝能够更快地将水和养分提供至叶片(Stratton et al., 2000).然而DBH作为观测木本植物生长状况的重要指标, 常被用于比较植株的生长速度(孔国辉等, 2006).随DBH增大, WD与RPHA增大, 而RPA减小(图2).WD越高, 植株生长速度减缓, 针叶的光合作用增强(赵鹏宇等, 2016), 其运输光合产物能力增强, 储水能力减弱, 这与以往研究结果一致(Poorter et al., 2019). ...
Traits and trade-offs of wood anatomy between trunks and branches in tropical dry forest species 2 2020
... 枝是重要的木质器官, 不仅起到机械支撑作用, 还具有水分、养分的运输和存贮功能(Pratt et al., 2007; Poorter et al., 2010; Meinzer et al., 2011; Ziaco & Liang, 2018).性状不同, 其生态功能也存在差异.枝的解剖学研究中, 木质部代表水分运输能力(Pratt & Jacobsen, 2017; Ziaco & Liang, 2018; Gebauer et al., 2019; Schumann et al., 2019); 韧皮部代表光合产物运输能力(Agustí & Blázquez, 2020); 髓是存储养分和水分的重要场所; 树脂道不仅能够产生、运输树脂, 还具有防御功能(Lin et al., 2001).相对于细根和树干, 枝能够更快地向叶片提供养分(Stratton et al., 2000; Pratt & Black, 2006).此外, 枝的解剖性状对于预测树木对环境变化的适应策略具有重要意义(Tarelkin et al., 2019), 然而, 目前关于枝的解剖性状的种内变异规律及影响因素的研究较少(Poorter et al., 2010). ...
... 枝在组织分配中遵循一定的权衡策略(Pratt et al., 2007; 刘晓娟和马克平, 2015; Pratt & Jacobsen, 2017; Pulido-Rodríguez et al., 2020), 不同的组织成分承载的功能不同, 不同组织比例随植株大小的变化反映枝在不同时期的生活史策略.叶片在光合作用过程中需要水来维持气孔导度和CO2吸收(Gleason et al., 2016), 因此, 枝中木质部与叶片光合作用能力显著相关(Brodribb & Feild, 2000).木质部和韧皮部是主要的长距离液体运输通道(Zwieniecki et al., 2004), 分别具有水分运输能力(Pratt & Jacobsen, 2017; Gebauer et al., 2019; Ka?par et al., 2019; Ziaco & Liang, 2019)和光合产物运输能力(Agustí & Blázquez, 2020).水和养分的存储是影响树木健康和生活史策略的重要因素(Pratt & Black, 2006), 髓作为存储养分和水分的重要场所(Stratton et al., 2000), 髓面积大小及髓腔与木质部之间的水力运输能力显著影响叶片水平衡, 进而影响气孔导度限制植物的碳吸收(Phillips et al., 2003).此外, 髓腔中存储的水分比土壤水更靠近水分蒸发流失的位置, 相对于根和树干, 枝能够更快地将水和养分提供至叶片(Stratton et al., 2000).然而DBH作为观测木本植物生长状况的重要指标, 常被用于比较植株的生长速度(孔国辉等, 2006).随DBH增大, WD与RPHA增大, 而RPA减小(图2).WD越高, 植株生长速度减缓, 针叶的光合作用增强(赵鹏宇等, 2016), 其运输光合产物能力增强, 储水能力减弱, 这与以往研究结果一致(Poorter et al., 2019). ...
R: a Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna 4 2018
... 枝是重要的木质器官, 不仅起到机械支撑作用, 还具有水分、养分的运输和存贮功能(Pratt et al., 2007; Poorter et al., 2010; Meinzer et al., 2011; Ziaco & Liang, 2018).性状不同, 其生态功能也存在差异.枝的解剖学研究中, 木质部代表水分运输能力(Pratt & Jacobsen, 2017; Ziaco & Liang, 2018; Gebauer et al., 2019; Schumann et al., 2019); 韧皮部代表光合产物运输能力(Agustí & Blázquez, 2020); 髓是存储养分和水分的重要场所; 树脂道不仅能够产生、运输树脂, 还具有防御功能(Lin et al., 2001).相对于细根和树干, 枝能够更快地向叶片提供养分(Stratton et al., 2000; Pratt & Black, 2006).此外, 枝的解剖性状对于预测树木对环境变化的适应策略具有重要意义(Tarelkin et al., 2019), 然而, 目前关于枝的解剖性状的种内变异规律及影响因素的研究较少(Poorter et al., 2010). ...
... 本研究表明WD、RPHA与DBH显著正相关, RPA与DBH显著负相关, 而RRC、WN与树高显著正相关(表3; 图2).以往关于树干WD的研究表明, WD受树高影响程度较大(Osunkoya et al., 2007; Martinez- Cabrera et al., 2011; Poorter et al., 2012; Fang et al., 2020), 且与DBH显著负相关(Virgulino-Júnior et al., 2020).然而WD受DBH的影响大于树高(表1).随着DBH增大, 针叶总数量增加(肖瑜, 1995), 枝的WD显著增大(表3), 枝的结构支撑能力提高, 进一步证明相对于树干, 枝更侧重于对WD的投资以提供水平悬挂的结构支撑, 避免下垂或破损(Pratt & Jacobsen, 2017; Pulido- Rodríguez et al., 2020). ...
... 枝在组织分配中遵循一定的权衡策略(Pratt et al., 2007; 刘晓娟和马克平, 2015; Pratt & Jacobsen, 2017; Pulido-Rodríguez et al., 2020), 不同的组织成分承载的功能不同, 不同组织比例随植株大小的变化反映枝在不同时期的生活史策略.叶片在光合作用过程中需要水来维持气孔导度和CO2吸收(Gleason et al., 2016), 因此, 枝中木质部与叶片光合作用能力显著相关(Brodribb & Feild, 2000).木质部和韧皮部是主要的长距离液体运输通道(Zwieniecki et al., 2004), 分别具有水分运输能力(Pratt & Jacobsen, 2017; Gebauer et al., 2019; Ka?par et al., 2019; Ziaco & Liang, 2019)和光合产物运输能力(Agustí & Blázquez, 2020).水和养分的存储是影响树木健康和生活史策略的重要因素(Pratt & Black, 2006), 髓作为存储养分和水分的重要场所(Stratton et al., 2000), 髓面积大小及髓腔与木质部之间的水力运输能力显著影响叶片水平衡, 进而影响气孔导度限制植物的碳吸收(Phillips et al., 2003).此外, 髓腔中存储的水分比土壤水更靠近水分蒸发流失的位置, 相对于根和树干, 枝能够更快地将水和养分提供至叶片(Stratton et al., 2000).然而DBH作为观测木本植物生长状况的重要指标, 常被用于比较植株的生长速度(孔国辉等, 2006).随DBH增大, WD与RPHA增大, 而RPA减小(图2).WD越高, 植株生长速度减缓, 针叶的光合作用增强(赵鹏宇等, 2016), 其运输光合产物能力增强, 储水能力减弱, 这与以往研究结果一致(Poorter et al., 2019). ...
... ), 分别具有水分运输能力(Pratt & Jacobsen, 2017; Gebauer et al., 2019; Ka?par et al., 2019; Ziaco & Liang, 2019)和光合产物运输能力(Agustí & Blázquez, 2020).水和养分的存储是影响树木健康和生活史策略的重要因素(Pratt & Black, 2006), 髓作为存储养分和水分的重要场所(Stratton et al., 2000), 髓面积大小及髓腔与木质部之间的水力运输能力显著影响叶片水平衡, 进而影响气孔导度限制植物的碳吸收(Phillips et al., 2003).此外, 髓腔中存储的水分比土壤水更靠近水分蒸发流失的位置, 相对于根和树干, 枝能够更快地将水和养分提供至叶片(Stratton et al., 2000).然而DBH作为观测木本植物生长状况的重要指标, 常被用于比较植株的生长速度(孔国辉等, 2006).随DBH增大, WD与RPHA增大, 而RPA减小(图2).WD越高, 植株生长速度减缓, 针叶的光合作用增强(赵鹏宇等, 2016), 其运输光合产物能力增强, 储水能力减弱, 这与以往研究结果一致(Poorter et al., 2019). ...
Periderm: a concept- based approach to the structure of seed plants//Richard C, Lyons-Sobaski S, Wise R. Plant Anatomy 2 2018
... 枝是重要的木质器官, 不仅起到机械支撑作用, 还具有水分、养分的运输和存贮功能(Pratt et al., 2007; Poorter et al., 2010; Meinzer et al., 2011; Ziaco & Liang, 2018).性状不同, 其生态功能也存在差异.枝的解剖学研究中, 木质部代表水分运输能力(Pratt & Jacobsen, 2017; Ziaco & Liang, 2018; Gebauer et al., 2019; Schumann et al., 2019); 韧皮部代表光合产物运输能力(Agustí & Blázquez, 2020); 髓是存储养分和水分的重要场所; 树脂道不仅能够产生、运输树脂, 还具有防御功能(Lin et al., 2001).相对于细根和树干, 枝能够更快地向叶片提供养分(Stratton et al., 2000; Pratt & Black, 2006).此外, 枝的解剖性状对于预测树木对环境变化的适应策略具有重要意义(Tarelkin et al., 2019), 然而, 目前关于枝的解剖性状的种内变异规律及影响因素的研究较少(Poorter et al., 2010). ...
... 枝在组织分配中遵循一定的权衡策略(Pratt et al., 2007; 刘晓娟和马克平, 2015; Pratt & Jacobsen, 2017; Pulido-Rodríguez et al., 2020), 不同的组织成分承载的功能不同, 不同组织比例随植株大小的变化反映枝在不同时期的生活史策略.叶片在光合作用过程中需要水来维持气孔导度和CO2吸收(Gleason et al., 2016), 因此, 枝中木质部与叶片光合作用能力显著相关(Brodribb & Feild, 2000).木质部和韧皮部是主要的长距离液体运输通道(Zwieniecki et al., 2004), 分别具有水分运输能力(Pratt & Jacobsen, 2017; Gebauer et al., 2019; Ka?par et al., 2019; Ziaco & Liang, 2019)和光合产物运输能力(Agustí & Blázquez, 2020).水和养分的存储是影响树木健康和生活史策略的重要因素(Pratt & Black, 2006), 髓作为存储养分和水分的重要场所(Stratton et al., 2000), 髓面积大小及髓腔与木质部之间的水力运输能力显著影响叶片水平衡, 进而影响气孔导度限制植物的碳吸收(Phillips et al., 2003).此外, 髓腔中存储的水分比土壤水更靠近水分蒸发流失的位置, 相对于根和树干, 枝能够更快地将水和养分提供至叶片(Stratton et al., 2000).然而DBH作为观测木本植物生长状况的重要指标, 常被用于比较植株的生长速度(孔国辉等, 2006).随DBH增大, WD与RPHA增大, 而RPA减小(图2).WD越高, 植株生长速度减缓, 针叶的光合作用增强(赵鹏宇等, 2016), 其运输光合产物能力增强, 储水能力减弱, 这与以往研究结果一致(Poorter et al., 2019). ...
The effect of tree height and light availability on photosynthetic leaf traits of four neotropical species differing in shade tolerance 2 2000
... 光照强度、土壤养分和土壤水分等环境因素与许多枝性状的变异也密切相关.例如, WD随光照强度的增加而显著增加(Poorter et al., 2012), 而随土壤含水量的增加而减小(Preston et al., 2006; Yang et al., 2014).自然条件下, 植物所处环境通常随个体大小的变化而发生变化(Moles et al., 2009).例如, 树高通常与光有效性密切相关(Miyata et al., 2011), 即随树高增大, 光有效性呈增加趋势(Woodcock & Shier, 2003; Ewers et al., 2007; Cavaleri et al., 2010; Liu et al., 2020); 植株个体越大, 其根系系统越发达, 具有更强的土壤养分固定能力, 从而增加其土壤养分, 如土壤氮含量(Wang et al., 2013, 2017; Liu et al., 2020).可见, 植株大小(Osunkoya et al., 2007; Meinzer et al., 2011; Rosell et al., 2017; Ka?par et al., 2019)、枝龄(Rodriguez-Zaccaro et al., 2019)及环境(Preston et al., 2006; Poorter et al., 2012)均可能对枝性状的变异产生影响, 但这些因素是否能单独对枝性状产生影响以及对枝性状变异的相对贡献率尚不清晰. ...
... )及环境(Preston et al., 2006; Poorter et al., 2012)均可能对枝性状的变异产生影响, 但这些因素是否能单独对枝性状产生影响以及对枝性状变异的相对贡献率尚不清晰. ...
Wood structure and function change with maturity: age of the vascular cambium is associated with xylem changes in current-year growth 3 2019
... 以往关于枝性状变异的研究中多侧重于当年生枝(Xiang et al., 2009; Miyata et al., 2011; Osada, 2011; Rosell et al., 2017; Gebauer et al., 2019).然而, 当年生枝的养分元素、水分和碳水化合物含量等与多年生枝差异显著(杨冬梅等, 2012), 在枝的解剖学研究中, 当年生枝主要由初生组织组成, 而多年生枝主要是次生生长的产物(Leyton, 1972), 当年生枝的初生结构与多年生枝的次生结构在养分元素、结构和功能上存在差异(Suzuki & Hiura, 2000; Richard et al., 2018; Yang et al., 2019b), 但不同年龄枝的次生结构差异不显著.不仅如此, 木质部结构和功能性状在不同枝龄间也存在很大差异(Rodriguez- Zaccaro et al., 2019).被子植物中导管直径随枝龄增大而增大(Corcuera et al., 2004; Rodriguez- Zaccaro et al., 2019), 而裸子植物枝性状随枝龄的变异规律尚不清晰.此外, 对于枝性状的研究通常关注某一生活史阶段(植株大小), 如幼苗、幼树或成年树(Pulido-Rodríguez et al., 2020), 然而, 枝性状在整个树木发育阶段是如何变化的, 以及枝性状随枝龄的变异规律是否受植株大小的影响仍有待检验. ...
... 本研究表明WD、RPHA与DBH显著正相关, RPA与DBH显著负相关, 而RRC、WN与树高显著正相关(表3; 图2).以往关于树干WD的研究表明, WD受树高影响程度较大(Osunkoya et al., 2007; Martinez- Cabrera et al., 2011; Poorter et al., 2012; Fang et al., 2020), 且与DBH显著负相关(Virgulino-Júnior et al., 2020).然而WD受DBH的影响大于树高(表1).随着DBH增大, 针叶总数量增加(肖瑜, 1995), 枝的WD显著增大(表3), 枝的结构支撑能力提高, 进一步证明相对于树干, 枝更侧重于对WD的投资以提供水平悬挂的结构支撑, 避免下垂或破损(Pratt & Jacobsen, 2017; Pulido- Rodríguez et al., 2020). ...
... 枝在组织分配中遵循一定的权衡策略(Pratt et al., 2007; 刘晓娟和马克平, 2015; Pratt & Jacobsen, 2017; Pulido-Rodríguez et al., 2020), 不同的组织成分承载的功能不同, 不同组织比例随植株大小的变化反映枝在不同时期的生活史策略.叶片在光合作用过程中需要水来维持气孔导度和CO2吸收(Gleason et al., 2016), 因此, 枝中木质部与叶片光合作用能力显著相关(Brodribb & Feild, 2000).木质部和韧皮部是主要的长距离液体运输通道(Zwieniecki et al., 2004), 分别具有水分运输能力(Pratt & Jacobsen, 2017; Gebauer et al., 2019; Ka?par et al., 2019; Ziaco & Liang, 2019)和光合产物运输能力(Agustí & Blázquez, 2020).水和养分的存储是影响树木健康和生活史策略的重要因素(Pratt & Black, 2006), 髓作为存储养分和水分的重要场所(Stratton et al., 2000), 髓面积大小及髓腔与木质部之间的水力运输能力显著影响叶片水平衡, 进而影响气孔导度限制植物的碳吸收(Phillips et al., 2003).此外, 髓腔中存储的水分比土壤水更靠近水分蒸发流失的位置, 相对于根和树干, 枝能够更快地将水和养分提供至叶片(Stratton et al., 2000).然而DBH作为观测木本植物生长状况的重要指标, 常被用于比较植株的生长速度(孔国辉等, 2006).随DBH增大, WD与RPHA增大, 而RPA减小(图2).WD越高, 植株生长速度减缓, 针叶的光合作用增强(赵鹏宇等, 2016), 其运输光合产物能力增强, 储水能力减弱, 这与以往研究结果一致(Poorter et al., 2019). ...
Scaling of xylem vessel diameter with plant size: causes, predictions, and outstanding questions 1 2017
Xylem hydraulic safety and efficiency in relation to leaf and wood traits in three temperate Acer species differing in habitat preferences 1 2019
... 以往关于枝性状变异的研究中多侧重于当年生枝(Xiang et al., 2009; Miyata et al., 2011; Osada, 2011; Rosell et al., 2017; Gebauer et al., 2019).然而, 当年生枝的养分元素、水分和碳水化合物含量等与多年生枝差异显著(杨冬梅等, 2012), 在枝的解剖学研究中, 当年生枝主要由初生组织组成, 而多年生枝主要是次生生长的产物(Leyton, 1972), 当年生枝的初生结构与多年生枝的次生结构在养分元素、结构和功能上存在差异(Suzuki & Hiura, 2000; Richard et al., 2018; Yang et al., 2019b), 但不同年龄枝的次生结构差异不显著.不仅如此, 木质部结构和功能性状在不同枝龄间也存在很大差异(Rodriguez- Zaccaro et al., 2019).被子植物中导管直径随枝龄增大而增大(Corcuera et al., 2004; Rodriguez- Zaccaro et al., 2019), 而裸子植物枝性状随枝龄的变异规律尚不清晰.此外, 对于枝性状的研究通常关注某一生活史阶段(植株大小), 如幼苗、幼树或成年树(Pulido-Rodríguez et al., 2020), 然而, 枝性状在整个树木发育阶段是如何变化的, 以及枝性状随枝龄的变异规律是否受植株大小的影响仍有待检验. ...
Safety and efficiency conflicts in hydraulic architecture: scaling from tissues to trees. Plant, 1 2008
... DBH和树高均是表征植株大小的重要变量(Rosell et al., 2017; Ka?par et al., 2019; Fang et al., 2020), 但对枝性状变异的解释程度因性状而异.本研究中WD、RXA、RPHA和RPA对DBH变化更敏感, 而RRC和WN受树高影响更大(表1).树脂道作为存在于针叶树中特有的组织成分, 不仅能够产生及运输树脂, 还具有防御作用(Lin et al., 2001).氮通常是森林生态系统中影响植物生长的限制因子之一(Aerts & Chapin, 2000), 也是决定植被生产力的主要因素(Hikosaka et al., 2016).在温度、降水相对均匀的地区, 光有效性和土壤养分是影响叶性状的主要因素(Lilles et al., 2014), 叶片氮含量的分配是应对光环境的一种重要适应策略, 植物光合作用与叶片氮含量密切相关(Takashima et al., 2004).氮在植物体内是通过根吸收, 通过树干转移至树枝, 进而运输至叶片, 为其提供光合作用所必需的氮(Will, 1971).树高与光有效性密切相关(Sterck & Bongers, 2010; Miyata et al., 2011), 通常随树高增大, 光有效性增强(Woodcock & Shier, 2003; Cavaleri et al., 2010; Liu et al., 2020), 分枝的伸展程度增大(Sterck & Bongers, 2010), 叶片光合作用增强(Rijkers et al., 2000), 因此需要更多的WN供给至叶片以合成所需的光合作用酶(Takashima et al., 2004; Millard & Grelet, 2010).WN和RRC的变化均与垂直运输有关, 因此对树高变化更敏感(表1). ...
Crown development in tropical rain forest trees: patterns with tree height and light availability 3 2001
... 以往关于枝性状变异的研究中多侧重于当年生枝(Xiang et al., 2009; Miyata et al., 2011; Osada, 2011; Rosell et al., 2017; Gebauer et al., 2019).然而, 当年生枝的养分元素、水分和碳水化合物含量等与多年生枝差异显著(杨冬梅等, 2012), 在枝的解剖学研究中, 当年生枝主要由初生组织组成, 而多年生枝主要是次生生长的产物(Leyton, 1972), 当年生枝的初生结构与多年生枝的次生结构在养分元素、结构和功能上存在差异(Suzuki & Hiura, 2000; Richard et al., 2018; Yang et al., 2019b), 但不同年龄枝的次生结构差异不显著.不仅如此, 木质部结构和功能性状在不同枝龄间也存在很大差异(Rodriguez- Zaccaro et al., 2019).被子植物中导管直径随枝龄增大而增大(Corcuera et al., 2004; Rodriguez- Zaccaro et al., 2019), 而裸子植物枝性状随枝龄的变异规律尚不清晰.此外, 对于枝性状的研究通常关注某一生活史阶段(植株大小), 如幼苗、幼树或成年树(Pulido-Rodríguez et al., 2020), 然而, 枝性状在整个树木发育阶段是如何变化的, 以及枝性状随枝龄的变异规律是否受植株大小的影响仍有待检验. ...
... ; Rodriguez- Zaccaro et al., 2019), 而裸子植物枝性状随枝龄的变异规律尚不清晰.此外, 对于枝性状的研究通常关注某一生活史阶段(植株大小), 如幼苗、幼树或成年树(Pulido-Rodríguez et al., 2020), 然而, 枝性状在整个树木发育阶段是如何变化的, 以及枝性状随枝龄的变异规律是否受植株大小的影响仍有待检验. ...
... 光照强度、土壤养分和土壤水分等环境因素与许多枝性状的变异也密切相关.例如, WD随光照强度的增加而显著增加(Poorter et al., 2012), 而随土壤含水量的增加而减小(Preston et al., 2006; Yang et al., 2014).自然条件下, 植物所处环境通常随个体大小的变化而发生变化(Moles et al., 2009).例如, 树高通常与光有效性密切相关(Miyata et al., 2011), 即随树高增大, 光有效性呈增加趋势(Woodcock & Shier, 2003; Ewers et al., 2007; Cavaleri et al., 2010; Liu et al., 2020); 植株个体越大, 其根系系统越发达, 具有更强的土壤养分固定能力, 从而增加其土壤养分, 如土壤氮含量(Wang et al., 2013, 2017; Liu et al., 2020).可见, 植株大小(Osunkoya et al., 2007; Meinzer et al., 2011; Rosell et al., 2017; Ka?par et al., 2019)、枝龄(Rodriguez-Zaccaro et al., 2019)及环境(Preston et al., 2006; Poorter et al., 2012)均可能对枝性状的变异产生影响, 但这些因素是否能单独对枝性状产生影响以及对枝性状变异的相对贡献率尚不清晰. ...
Stem water storage capacity and efficiency of water transport: their functional significance in a Hawaiian dry forest. 4 2000
... 以往关于枝性状变异的研究中多侧重于当年生枝(Xiang et al., 2009; Miyata et al., 2011; Osada, 2011; Rosell et al., 2017; Gebauer et al., 2019).然而, 当年生枝的养分元素、水分和碳水化合物含量等与多年生枝差异显著(杨冬梅等, 2012), 在枝的解剖学研究中, 当年生枝主要由初生组织组成, 而多年生枝主要是次生生长的产物(Leyton, 1972), 当年生枝的初生结构与多年生枝的次生结构在养分元素、结构和功能上存在差异(Suzuki & Hiura, 2000; Richard et al., 2018; Yang et al., 2019b), 但不同年龄枝的次生结构差异不显著.不仅如此, 木质部结构和功能性状在不同枝龄间也存在很大差异(Rodriguez- Zaccaro et al., 2019).被子植物中导管直径随枝龄增大而增大(Corcuera et al., 2004; Rodriguez- Zaccaro et al., 2019), 而裸子植物枝性状随枝龄的变异规律尚不清晰.此外, 对于枝性状的研究通常关注某一生活史阶段(植株大小), 如幼苗、幼树或成年树(Pulido-Rodríguez et al., 2020), 然而, 枝性状在整个树木发育阶段是如何变化的, 以及枝性状随枝龄的变异规律是否受植株大小的影响仍有待检验. ...
... 光照强度、土壤养分和土壤水分等环境因素与许多枝性状的变异也密切相关.例如, WD随光照强度的增加而显著增加(Poorter et al., 2012), 而随土壤含水量的增加而减小(Preston et al., 2006; Yang et al., 2014).自然条件下, 植物所处环境通常随个体大小的变化而发生变化(Moles et al., 2009).例如, 树高通常与光有效性密切相关(Miyata et al., 2011), 即随树高增大, 光有效性呈增加趋势(Woodcock & Shier, 2003; Ewers et al., 2007; Cavaleri et al., 2010; Liu et al., 2020); 植株个体越大, 其根系系统越发达, 具有更强的土壤养分固定能力, 从而增加其土壤养分, 如土壤氮含量(Wang et al., 2013, 2017; Liu et al., 2020).可见, 植株大小(Osunkoya et al., 2007; Meinzer et al., 2011; Rosell et al., 2017; Ka?par et al., 2019)、枝龄(Rodriguez-Zaccaro et al., 2019)及环境(Preston et al., 2006; Poorter et al., 2012)均可能对枝性状的变异产生影响, 但这些因素是否能单独对枝性状产生影响以及对枝性状变异的相对贡献率尚不清晰. ...
... 以往研究表明植株大小、枝龄及环境因素均是引起枝性状变异的主要因素(Rosell et al., 2017; Ka?par et al., 2019), 但很少有研究同时评价这些因素对枝性状的影响程度及差异.本研究表明植株大小(DBH或树高)、枝龄和环境均能独立影响枝性状的变异, 但其影响程度在不同枝性状间存在明显差异.整体而言, 枝龄、植株大小及环境对枝性状的影响依次减小. ...
... DBH和树高均是表征植株大小的重要变量(Rosell et al., 2017; Ka?par et al., 2019; Fang et al., 2020), 但对枝性状变异的解释程度因性状而异.本研究中WD、RXA、RPHA和RPA对DBH变化更敏感, 而RRC和WN受树高影响更大(表1).树脂道作为存在于针叶树中特有的组织成分, 不仅能够产生及运输树脂, 还具有防御作用(Lin et al., 2001).氮通常是森林生态系统中影响植物生长的限制因子之一(Aerts & Chapin, 2000), 也是决定植被生产力的主要因素(Hikosaka et al., 2016).在温度、降水相对均匀的地区, 光有效性和土壤养分是影响叶性状的主要因素(Lilles et al., 2014), 叶片氮含量的分配是应对光环境的一种重要适应策略, 植物光合作用与叶片氮含量密切相关(Takashima et al., 2004).氮在植物体内是通过根吸收, 通过树干转移至树枝, 进而运输至叶片, 为其提供光合作用所必需的氮(Will, 1971).树高与光有效性密切相关(Sterck & Bongers, 2010; Miyata et al., 2011), 通常随树高增大, 光有效性增强(Woodcock & Shier, 2003; Cavaleri et al., 2010; Liu et al., 2020), 分枝的伸展程度增大(Sterck & Bongers, 2010), 叶片光合作用增强(Rijkers et al., 2000), 因此需要更多的WN供给至叶片以合成所需的光合作用酶(Takashima et al., 2004; Millard & Grelet, 2010).WN和RRC的变化均与垂直运输有关, 因此对树高变化更敏感(表1). ...
Allometric differences between current-year shoots and large branches of deciduous broad-leaved tree species 2 2000
... 枝是重要的木质器官, 不仅起到机械支撑作用, 还具有水分、养分的运输和存贮功能(Pratt et al., 2007; Poorter et al., 2010; Meinzer et al., 2011; Ziaco & Liang, 2018).性状不同, 其生态功能也存在差异.枝的解剖学研究中, 木质部代表水分运输能力(Pratt & Jacobsen, 2017; Ziaco & Liang, 2018; Gebauer et al., 2019; Schumann et al., 2019); 韧皮部代表光合产物运输能力(Agustí & Blázquez, 2020); 髓是存储养分和水分的重要场所; 树脂道不仅能够产生、运输树脂, 还具有防御功能(Lin et al., 2001).相对于细根和树干, 枝能够更快地向叶片提供养分(Stratton et al., 2000; Pratt & Black, 2006).此外, 枝的解剖性状对于预测树木对环境变化的适应策略具有重要意义(Tarelkin et al., 2019), 然而, 目前关于枝的解剖性状的种内变异规律及影响因素的研究较少(Poorter et al., 2010). ...
... 大多数枝性状与枝龄显著相关(p > 0.001, 表3): RPA和RRC随枝龄增大而减小, 在小树中RPA随枝龄增大而减小的速率更大; 相反, 在小树中RRC随枝龄增大而减小的速率更小; WD、RXA和RPHA随枝龄增大而增大(图3).随着枝龄增大, 结构支撑能力增强, 针叶光合能力减弱, 水分和养分存储能力减弱, RXA和RPHA增大, 更好地向形态学上端为不同年龄针叶的光合作用提供所需水分(Gebauer et al., 2019; Schumann et al., 2019), 以及向下运输不同年龄针叶的光合产物(Agustí & Blázquez, 2020).WD和RPHA随DBH增大而增大, 当年生枝的变化速率显著高于多年生枝(图2), 且WD随枝龄增大而增大, 而RPHA随枝龄增大而逐渐减小(图3).当年生针叶的光合作用强于多年生针叶, 随枝龄增长, 枝逐渐从运输器官转变为以机械支撑为主的器官.RXA受DBH大小变化的影响不显著(图2), RXA仅在小树(DBH > 10 cm)中随枝龄增大而增大.以往关于枝的木质部运输能力的研究表明, 相对于导管, 管胞更具有保证水力运输安全性的能力, 并且具有较强的抗空化性, 但水分输送效率较低(Hacke et al., 2006).RXA与植株大小、枝龄无显著相关性; 此外, 以往研究还表明枝的水力运输能力与土壤含水率无相关性(Domec et al., 2008; Sperry et al., 2008; Peltoniemi et al., 2012), 因此, 对于影响RXA的因素仍有待进一步研究. ...
Photosynthesis or persistence: nitrogen allocation in leaves of evergreen and deciduous Quercus species. Plant, 1 2004
... 大多数枝性状与枝龄显著相关(p > 0.001, 表3): RPA和RRC随枝龄增大而减小, 在小树中RPA随枝龄增大而减小的速率更大; 相反, 在小树中RRC随枝龄增大而减小的速率更小; WD、RXA和RPHA随枝龄增大而增大(图3).随着枝龄增大, 结构支撑能力增强, 针叶光合能力减弱, 水分和养分存储能力减弱, RXA和RPHA增大, 更好地向形态学上端为不同年龄针叶的光合作用提供所需水分(Gebauer et al., 2019; Schumann et al., 2019), 以及向下运输不同年龄针叶的光合产物(Agustí & Blázquez, 2020).WD和RPHA随DBH增大而增大, 当年生枝的变化速率显著高于多年生枝(图2), 且WD随枝龄增大而增大, 而RPHA随枝龄增大而逐渐减小(图3).当年生针叶的光合作用强于多年生针叶, 随枝龄增长, 枝逐渐从运输器官转变为以机械支撑为主的器官.RXA受DBH大小变化的影响不显著(图2), RXA仅在小树(DBH > 10 cm)中随枝龄增大而增大.以往关于枝的木质部运输能力的研究表明, 相对于导管, 管胞更具有保证水力运输安全性的能力, 并且具有较强的抗空化性, 但水分输送效率较低(Hacke et al., 2006).RXA与植株大小、枝龄无显著相关性; 此外, 以往研究还表明枝的水力运输能力与土壤含水率无相关性(Domec et al., 2008; Sperry et al., 2008; Peltoniemi et al., 2012), 因此, 对于影响RXA的因素仍有待进一步研究. ...
Wood anatomy variability under contrasted environmental conditions of common deciduous and evergreen species from central African forests 2019
Wood density in mangrove forests on the Brazilian Amazon Coast 4 2020
... 枝是重要的木质器官, 不仅起到机械支撑作用, 还具有水分、养分的运输和存贮功能(Pratt et al., 2007; Poorter et al., 2010; Meinzer et al., 2011; Ziaco & Liang, 2018).性状不同, 其生态功能也存在差异.枝的解剖学研究中, 木质部代表水分运输能力(Pratt & Jacobsen, 2017; Ziaco & Liang, 2018; Gebauer et al., 2019; Schumann et al., 2019); 韧皮部代表光合产物运输能力(Agustí & Blázquez, 2020); 髓是存储养分和水分的重要场所; 树脂道不仅能够产生、运输树脂, 还具有防御功能(Lin et al., 2001).相对于细根和树干, 枝能够更快地向叶片提供养分(Stratton et al., 2000; Pratt & Black, 2006).此外, 枝的解剖性状对于预测树木对环境变化的适应策略具有重要意义(Tarelkin et al., 2019), 然而, 目前关于枝的解剖性状的种内变异规律及影响因素的研究较少(Poorter et al., 2010). ...
... 枝在组织分配中遵循一定的权衡策略(Pratt et al., 2007; 刘晓娟和马克平, 2015; Pratt & Jacobsen, 2017; Pulido-Rodríguez et al., 2020), 不同的组织成分承载的功能不同, 不同组织比例随植株大小的变化反映枝在不同时期的生活史策略.叶片在光合作用过程中需要水来维持气孔导度和CO2吸收(Gleason et al., 2016), 因此, 枝中木质部与叶片光合作用能力显著相关(Brodribb & Feild, 2000).木质部和韧皮部是主要的长距离液体运输通道(Zwieniecki et al., 2004), 分别具有水分运输能力(Pratt & Jacobsen, 2017; Gebauer et al., 2019; Ka?par et al., 2019; Ziaco & Liang, 2019)和光合产物运输能力(Agustí & Blázquez, 2020).水和养分的存储是影响树木健康和生活史策略的重要因素(Pratt & Black, 2006), 髓作为存储养分和水分的重要场所(Stratton et al., 2000), 髓面积大小及髓腔与木质部之间的水力运输能力显著影响叶片水平衡, 进而影响气孔导度限制植物的碳吸收(Phillips et al., 2003).此外, 髓腔中存储的水分比土壤水更靠近水分蒸发流失的位置, 相对于根和树干, 枝能够更快地将水和养分提供至叶片(Stratton et al., 2000).然而DBH作为观测木本植物生长状况的重要指标, 常被用于比较植株的生长速度(孔国辉等, 2006).随DBH增大, WD与RPHA增大, 而RPA减小(图2).WD越高, 植株生长速度减缓, 针叶的光合作用增强(赵鹏宇等, 2016), 其运输光合产物能力增强, 储水能力减弱, 这与以往研究结果一致(Poorter et al., 2019). ...
Intraspecific phenotypic variability of plant functional traits in contrasting mountain grasslands habitats 1 2013
... 枝是重要的木质器官, 不仅起到机械支撑作用, 还具有水分、养分的运输和存贮功能(Pratt et al., 2007; Poorter et al., 2010; Meinzer et al., 2011; Ziaco & Liang, 2018).性状不同, 其生态功能也存在差异.枝的解剖学研究中, 木质部代表水分运输能力(Pratt & Jacobsen, 2017; Ziaco & Liang, 2018; Gebauer et al., 2019; Schumann et al., 2019); 韧皮部代表光合产物运输能力(Agustí & Blázquez, 2020); 髓是存储养分和水分的重要场所; 树脂道不仅能够产生、运输树脂, 还具有防御功能(Lin et al., 2001).相对于细根和树干, 枝能够更快地向叶片提供养分(Stratton et al., 2000; Pratt & Black, 2006).此外, 枝的解剖性状对于预测树木对环境变化的适应策略具有重要意义(Tarelkin et al., 2019), 然而, 目前关于枝的解剖性状的种内变异规律及影响因素的研究较少(Poorter et al., 2010). ...
Nitrogen supply, apical dominance and branch growth in Pinus radiata 1 1971
... 本研究表明WD、RPHA与DBH显著正相关, RPA与DBH显著负相关, 而RRC、WN与树高显著正相关(表3; 图2).以往关于树干WD的研究表明, WD受树高影响程度较大(Osunkoya et al., 2007; Martinez- Cabrera et al., 2011; Poorter et al., 2012; Fang et al., 2020), 且与DBH显著负相关(Virgulino-Júnior et al., 2020).然而WD受DBH的影响大于树高(表1).随着DBH增大, 针叶总数量增加(肖瑜, 1995), 枝的WD显著增大(表3), 枝的结构支撑能力提高, 进一步证明相对于树干, 枝更侧重于对WD的投资以提供水平悬挂的结构支撑, 避免下垂或破损(Pratt & Jacobsen, 2017; Pulido- Rodríguez et al., 2020). ...
Does canopy position affect wood specific gravity in temperate forest trees? 1 2003
... 光照强度、土壤养分和土壤水分等环境因素与许多枝性状的变异也密切相关.例如, WD随光照强度的增加而显著增加(Poorter et al., 2012), 而随土壤含水量的增加而减小(Preston et al., 2006; Yang et al., 2014).自然条件下, 植物所处环境通常随个体大小的变化而发生变化(Moles et al., 2009).例如, 树高通常与光有效性密切相关(Miyata et al., 2011), 即随树高增大, 光有效性呈增加趋势(Woodcock & Shier, 2003; Ewers et al., 2007; Cavaleri et al., 2010; Liu et al., 2020); 植株个体越大, 其根系系统越发达, 具有更强的土壤养分固定能力, 从而增加其土壤养分, 如土壤氮含量(Wang et al., 2013, 2017; Liu et al., 2020).可见, 植株大小(Osunkoya et al., 2007; Meinzer et al., 2011; Rosell et al., 2017; Ka?par et al., 2019)、枝龄(Rodriguez-Zaccaro et al., 2019)及环境(Preston et al., 2006; Poorter et al., 2012)均可能对枝性状的变异产生影响, 但这些因素是否能单独对枝性状产生影响以及对枝性状变异的相对贡献率尚不清晰. ...
The worldwide leaf economics spectrum 1 2004
... 光照强度、土壤养分和土壤水分等环境因素与许多枝性状的变异也密切相关.例如, WD随光照强度的增加而显著增加(Poorter et al., 2012), 而随土壤含水量的增加而减小(Preston et al., 2006; Yang et al., 2014).自然条件下, 植物所处环境通常随个体大小的变化而发生变化(Moles et al., 2009).例如, 树高通常与光有效性密切相关(Miyata et al., 2011), 即随树高增大, 光有效性呈增加趋势(Woodcock & Shier, 2003; Ewers et al., 2007; Cavaleri et al., 2010; Liu et al., 2020); 植株个体越大, 其根系系统越发达, 具有更强的土壤养分固定能力, 从而增加其土壤养分, 如土壤氮含量(Wang et al., 2013, 2017; Liu et al., 2020).可见, 植株大小(Osunkoya et al., 2007; Meinzer et al., 2011; Rosell et al., 2017; Ka?par et al., 2019)、枝龄(Rodriguez-Zaccaro et al., 2019)及环境(Preston et al., 2006; Poorter et al., 2012)均可能对枝性状的变异产生影响, 但这些因素是否能单独对枝性状产生影响以及对枝性状变异的相对贡献率尚不清晰. ...
Within-twig biomass allocation in subtropical evergreen broad-leaved species along an altitudinal gradient: allometric scaling analysis 1 2009
... 掌握植物功能性状的种内变异规律及影响因素, 对深入了解植物的存活、生长和死亡等生态学过程至关重要(Wright et al., 2004; 刘晓娟和马克平, 2015), 也有助于揭示植物的资源利用策略及植物对气候变化的响应机制(He et al., 2019).然而, 以往研究更关注叶性状和细根性状的变异(Hajek et al., 2013; Wellstein et al., 2013; Isaac et al., 2017; Bloomfield et al., 2018), 而对于枝性状种内变异的研究较少. ...
杉木人工林针叶光合与蒸腾作用的时空特征 1 2002
... DBH和树高均是表征植株大小的重要变量(Rosell et al., 2017; Ka?par et al., 2019; Fang et al., 2020), 但对枝性状变异的解释程度因性状而异.本研究中WD、RXA、RPHA和RPA对DBH变化更敏感, 而RRC和WN受树高影响更大(表1).树脂道作为存在于针叶树中特有的组织成分, 不仅能够产生及运输树脂, 还具有防御作用(Lin et al., 2001).氮通常是森林生态系统中影响植物生长的限制因子之一(Aerts & Chapin, 2000), 也是决定植被生产力的主要因素(Hikosaka et al., 2016).在温度、降水相对均匀的地区, 光有效性和土壤养分是影响叶性状的主要因素(Lilles et al., 2014), 叶片氮含量的分配是应对光环境的一种重要适应策略, 植物光合作用与叶片氮含量密切相关(Takashima et al., 2004).氮在植物体内是通过根吸收, 通过树干转移至树枝, 进而运输至叶片, 为其提供光合作用所必需的氮(Will, 1971).树高与光有效性密切相关(Sterck & Bongers, 2010; Miyata et al., 2011), 通常随树高增大, 光有效性增强(Woodcock & Shier, 2003; Cavaleri et al., 2010; Liu et al., 2020), 分枝的伸展程度增大(Sterck & Bongers, 2010), 叶片光合作用增强(Rijkers et al., 2000), 因此需要更多的WN供给至叶片以合成所需的光合作用酶(Takashima et al., 2004; Millard & Grelet, 2010).WN和RRC的变化均与垂直运输有关, 因此对树高变化更敏感(表1). ...
杉木人工林针叶光合与蒸腾作用的时空特征 1 2002
... DBH和树高均是表征植株大小的重要变量(Rosell et al., 2017; Ka?par et al., 2019; Fang et al., 2020), 但对枝性状变异的解释程度因性状而异.本研究中WD、RXA、RPHA和RPA对DBH变化更敏感, 而RRC和WN受树高影响更大(表1).树脂道作为存在于针叶树中特有的组织成分, 不仅能够产生及运输树脂, 还具有防御作用(Lin et al., 2001).氮通常是森林生态系统中影响植物生长的限制因子之一(Aerts & Chapin, 2000), 也是决定植被生产力的主要因素(Hikosaka et al., 2016).在温度、降水相对均匀的地区, 光有效性和土壤养分是影响叶性状的主要因素(Lilles et al., 2014), 叶片氮含量的分配是应对光环境的一种重要适应策略, 植物光合作用与叶片氮含量密切相关(Takashima et al., 2004).氮在植物体内是通过根吸收, 通过树干转移至树枝, 进而运输至叶片, 为其提供光合作用所必需的氮(Will, 1971).树高与光有效性密切相关(Sterck & Bongers, 2010; Miyata et al., 2011), 通常随树高增大, 光有效性增强(Woodcock & Shier, 2003; Cavaleri et al., 2010; Liu et al., 2020), 分枝的伸展程度增大(Sterck & Bongers, 2010), 叶片光合作用增强(Rijkers et al., 2000), 因此需要更多的WN供给至叶片以合成所需的光合作用酶(Takashima et al., 2004; Millard & Grelet, 2010).WN和RRC的变化均与垂直运输有关, 因此对树高变化更敏感(表1). ...
巴山松针叶群体数量和寿命的水平和垂直变化趋势分析 2 1995
... 光照强度、土壤养分和土壤水分等环境因素与许多枝性状的变异也密切相关.例如, WD随光照强度的增加而显著增加(Poorter et al., 2012), 而随土壤含水量的增加而减小(Preston et al., 2006; Yang et al., 2014).自然条件下, 植物所处环境通常随个体大小的变化而发生变化(Moles et al., 2009).例如, 树高通常与光有效性密切相关(Miyata et al., 2011), 即随树高增大, 光有效性呈增加趋势(Woodcock & Shier, 2003; Ewers et al., 2007; Cavaleri et al., 2010; Liu et al., 2020); 植株个体越大, 其根系系统越发达, 具有更强的土壤养分固定能力, 从而增加其土壤养分, 如土壤氮含量(Wang et al., 2013, 2017; Liu et al., 2020).可见, 植株大小(Osunkoya et al., 2007; Meinzer et al., 2011; Rosell et al., 2017; Ka?par et al., 2019)、枝龄(Rodriguez-Zaccaro et al., 2019)及环境(Preston et al., 2006; Poorter et al., 2012)均可能对枝性状的变异产生影响, 但这些因素是否能单独对枝性状产生影响以及对枝性状变异的相对贡献率尚不清晰. ...
... DBH和树高均是表征植株大小的重要变量(Rosell et al., 2017; Ka?par et al., 2019; Fang et al., 2020), 但对枝性状变异的解释程度因性状而异.本研究中WD、RXA、RPHA和RPA对DBH变化更敏感, 而RRC和WN受树高影响更大(表1).树脂道作为存在于针叶树中特有的组织成分, 不仅能够产生及运输树脂, 还具有防御作用(Lin et al., 2001).氮通常是森林生态系统中影响植物生长的限制因子之一(Aerts & Chapin, 2000), 也是决定植被生产力的主要因素(Hikosaka et al., 2016).在温度、降水相对均匀的地区, 光有效性和土壤养分是影响叶性状的主要因素(Lilles et al., 2014), 叶片氮含量的分配是应对光环境的一种重要适应策略, 植物光合作用与叶片氮含量密切相关(Takashima et al., 2004).氮在植物体内是通过根吸收, 通过树干转移至树枝, 进而运输至叶片, 为其提供光合作用所必需的氮(Will, 1971).树高与光有效性密切相关(Sterck & Bongers, 2010; Miyata et al., 2011), 通常随树高增大, 光有效性增强(Woodcock & Shier, 2003; Cavaleri et al., 2010; Liu et al., 2020), 分枝的伸展程度增大(Sterck & Bongers, 2010), 叶片光合作用增强(Rijkers et al., 2000), 因此需要更多的WN供给至叶片以合成所需的光合作用酶(Takashima et al., 2004; Millard & Grelet, 2010).WN和RRC的变化均与垂直运输有关, 因此对树高变化更敏感(表1). ...
巴山松针叶群体数量和寿命的水平和垂直变化趋势分析 2 1995
... 光照强度、土壤养分和土壤水分等环境因素与许多枝性状的变异也密切相关.例如, WD随光照强度的增加而显著增加(Poorter et al., 2012), 而随土壤含水量的增加而减小(Preston et al., 2006; Yang et al., 2014).自然条件下, 植物所处环境通常随个体大小的变化而发生变化(Moles et al., 2009).例如, 树高通常与光有效性密切相关(Miyata et al., 2011), 即随树高增大, 光有效性呈增加趋势(Woodcock & Shier, 2003; Ewers et al., 2007; Cavaleri et al., 2010; Liu et al., 2020); 植株个体越大, 其根系系统越发达, 具有更强的土壤养分固定能力, 从而增加其土壤养分, 如土壤氮含量(Wang et al., 2013, 2017; Liu et al., 2020).可见, 植株大小(Osunkoya et al., 2007; Meinzer et al., 2011; Rosell et al., 2017; Ka?par et al., 2019)、枝龄(Rodriguez-Zaccaro et al., 2019)及环境(Preston et al., 2006; Poorter et al., 2012)均可能对枝性状的变异产生影响, 但这些因素是否能单独对枝性状产生影响以及对枝性状变异的相对贡献率尚不清晰. ...
... DBH和树高均是表征植株大小的重要变量(Rosell et al., 2017; Ka?par et al., 2019; Fang et al., 2020), 但对枝性状变异的解释程度因性状而异.本研究中WD、RXA、RPHA和RPA对DBH变化更敏感, 而RRC和WN受树高影响更大(表1).树脂道作为存在于针叶树中特有的组织成分, 不仅能够产生及运输树脂, 还具有防御作用(Lin et al., 2001).氮通常是森林生态系统中影响植物生长的限制因子之一(Aerts & Chapin, 2000), 也是决定植被生产力的主要因素(Hikosaka et al., 2016).在温度、降水相对均匀的地区, 光有效性和土壤养分是影响叶性状的主要因素(Lilles et al., 2014), 叶片氮含量的分配是应对光环境的一种重要适应策略, 植物光合作用与叶片氮含量密切相关(Takashima et al., 2004).氮在植物体内是通过根吸收, 通过树干转移至树枝, 进而运输至叶片, 为其提供光合作用所必需的氮(Will, 1971).树高与光有效性密切相关(Sterck & Bongers, 2010; Miyata et al., 2011), 通常随树高增大, 光有效性增强(Woodcock & Shier, 2003; Cavaleri et al., 2010; Liu et al., 2020), 分枝的伸展程度增大(Sterck & Bongers, 2010), 叶片光合作用增强(Rijkers et al., 2000), 因此需要更多的WN供给至叶片以合成所需的光合作用酶(Takashima et al., 2004; Millard & Grelet, 2010).WN和RRC的变化均与垂直运输有关, 因此对树高变化更敏感(表1). ...
长白山阔叶红松林主要树种根系分布规律的研究 1 1987
... 掌握植物功能性状的种内变异规律及影响因素, 对深入了解植物的存活、生长和死亡等生态学过程至关重要(Wright et al., 2004; 刘晓娟和马克平, 2015), 也有助于揭示植物的资源利用策略及植物对气候变化的响应机制(He et al., 2019).然而, 以往研究更关注叶性状和细根性状的变异(Hajek et al., 2013; Wellstein et al., 2013; Isaac et al., 2017; Bloomfield et al., 2018), 而对于枝性状种内变异的研究较少. ...
长白山阔叶红松林主要树种根系分布规律的研究 1 1987
... 掌握植物功能性状的种内变异规律及影响因素, 对深入了解植物的存活、生长和死亡等生态学过程至关重要(Wright et al., 2004; 刘晓娟和马克平, 2015), 也有助于揭示植物的资源利用策略及植物对气候变化的响应机制(He et al., 2019).然而, 以往研究更关注叶性状和细根性状的变异(Hajek et al., 2013; Wellstein et al., 2013; Isaac et al., 2017; Bloomfield et al., 2018), 而对于枝性状种内变异的研究较少. ...
木本植物茎叶功能性状及其关系随环境变化的研究进展 1 2012
... 以往关于枝性状变异的研究中多侧重于当年生枝(Xiang et al., 2009; Miyata et al., 2011; Osada, 2011; Rosell et al., 2017; Gebauer et al., 2019).然而, 当年生枝的养分元素、水分和碳水化合物含量等与多年生枝差异显著(杨冬梅等, 2012), 在枝的解剖学研究中, 当年生枝主要由初生组织组成, 而多年生枝主要是次生生长的产物(Leyton, 1972), 当年生枝的初生结构与多年生枝的次生结构在养分元素、结构和功能上存在差异(Suzuki & Hiura, 2000; Richard et al., 2018; Yang et al., 2019b), 但不同年龄枝的次生结构差异不显著.不仅如此, 木质部结构和功能性状在不同枝龄间也存在很大差异(Rodriguez- Zaccaro et al., 2019).被子植物中导管直径随枝龄增大而增大(Corcuera et al., 2004; Rodriguez- Zaccaro et al., 2019), 而裸子植物枝性状随枝龄的变异规律尚不清晰.此外, 对于枝性状的研究通常关注某一生活史阶段(植株大小), 如幼苗、幼树或成年树(Pulido-Rodríguez et al., 2020), 然而, 枝性状在整个树木发育阶段是如何变化的, 以及枝性状随枝龄的变异规律是否受植株大小的影响仍有待检验. ...
木本植物茎叶功能性状及其关系随环境变化的研究进展 1 2012
... 以往关于枝性状变异的研究中多侧重于当年生枝(Xiang et al., 2009; Miyata et al., 2011; Osada, 2011; Rosell et al., 2017; Gebauer et al., 2019).然而, 当年生枝的养分元素、水分和碳水化合物含量等与多年生枝差异显著(杨冬梅等, 2012), 在枝的解剖学研究中, 当年生枝主要由初生组织组成, 而多年生枝主要是次生生长的产物(Leyton, 1972), 当年生枝的初生结构与多年生枝的次生结构在养分元素、结构和功能上存在差异(Suzuki & Hiura, 2000; Richard et al., 2018; Yang et al., 2019b), 但不同年龄枝的次生结构差异不显著.不仅如此, 木质部结构和功能性状在不同枝龄间也存在很大差异(Rodriguez- Zaccaro et al., 2019).被子植物中导管直径随枝龄增大而增大(Corcuera et al., 2004; Rodriguez- Zaccaro et al., 2019), 而裸子植物枝性状随枝龄的变异规律尚不清晰.此外, 对于枝性状的研究通常关注某一生活史阶段(植株大小), 如幼苗、幼树或成年树(Pulido-Rodríguez et al., 2020), 然而, 枝性状在整个树木发育阶段是如何变化的, 以及枝性状随枝龄的变异规律是否受植株大小的影响仍有待检验. ...
The soil C:N stoichiometry is more sensitive than the leaf C:N stoichiometry to nitrogen addition: a four-year nitrogen addition experiment in a Pinus koraiensis plantation 1 2019
... 对于每株样树, 于树干底部0-10 cm的土层范围内使用土壤取芯器采集土壤子样本, 重复3次且任意两个采样方向之间的角度约为120°, 将这3个子样本进行混合并剔除明显的根和凋落物等杂质(Xu et al., 1987; Yang et al., 2019a; Liu et al., 2020).然后, 利用烘干法测定每个样品的土壤水含量(g·g-1); 采用AQ400自动间断化学分析仪(SEAL Analytical, Mequon, USA)测量全氮含量(mg·g-1)和全磷含量(mg·g-1). ...
Branch age and angle as crucial drivers of leaf photosynthetic performance and fruiting in high-density planting: a study case in spur-type apple “Vallee Spur” (Malus domestica) 2 2019
... 以往关于枝性状变异的研究中多侧重于当年生枝(Xiang et al., 2009; Miyata et al., 2011; Osada, 2011; Rosell et al., 2017; Gebauer et al., 2019).然而, 当年生枝的养分元素、水分和碳水化合物含量等与多年生枝差异显著(杨冬梅等, 2012), 在枝的解剖学研究中, 当年生枝主要由初生组织组成, 而多年生枝主要是次生生长的产物(Leyton, 1972), 当年生枝的初生结构与多年生枝的次生结构在养分元素、结构和功能上存在差异(Suzuki & Hiura, 2000; Richard et al., 2018; Yang et al., 2019b), 但不同年龄枝的次生结构差异不显著.不仅如此, 木质部结构和功能性状在不同枝龄间也存在很大差异(Rodriguez- Zaccaro et al., 2019).被子植物中导管直径随枝龄增大而增大(Corcuera et al., 2004; Rodriguez- Zaccaro et al., 2019), 而裸子植物枝性状随枝龄的变异规律尚不清晰.此外, 对于枝性状的研究通常关注某一生活史阶段(植株大小), 如幼苗、幼树或成年树(Pulido-Rodríguez et al., 2020), 然而, 枝性状在整个树木发育阶段是如何变化的, 以及枝性状随枝龄的变异规律是否受植株大小的影响仍有待检验. ...
Twig-leaf size relationships in woody plants vary intraspecifically along a soil moisture gradient 2014
Contribution of leaf anatomical traits to leaf mass per area among canopy layers for five coexisting broadleaf species across shade tolerances at a regional scale 2 2019
... 本研究表明WD、RPHA与DBH显著正相关, RPA与DBH显著负相关, 而RRC、WN与树高显著正相关(表3; 图2).以往关于树干WD的研究表明, WD受树高影响程度较大(Osunkoya et al., 2007; Martinez- Cabrera et al., 2011; Poorter et al., 2012; Fang et al., 2020), 且与DBH显著负相关(Virgulino-Júnior et al., 2020).然而WD受DBH的影响大于树高(表1).随着DBH增大, 针叶总数量增加(肖瑜, 1995), 枝的WD显著增大(表3), 枝的结构支撑能力提高, 进一步证明相对于树干, 枝更侧重于对WD的投资以提供水平悬挂的结构支撑, 避免下垂或破损(Pratt & Jacobsen, 2017; Pulido- Rodríguez et al., 2020). ...
胡杨不同发育阶段叶片形态解剖学特征及其与胸径的关系 1 2016
... 对于每株样树, 于树干底部0-10 cm的土层范围内使用土壤取芯器采集土壤子样本, 重复3次且任意两个采样方向之间的角度约为120°, 将这3个子样本进行混合并剔除明显的根和凋落物等杂质(Xu et al., 1987; Yang et al., 2019a; Liu et al., 2020).然后, 利用烘干法测定每个样品的土壤水含量(g·g-1); 采用AQ400自动间断化学分析仪(SEAL Analytical, Mequon, USA)测量全氮含量(mg·g-1)和全磷含量(mg·g-1). ...
胡杨不同发育阶段叶片形态解剖学特征及其与胸径的关系 1 2016
... 对于每株样树, 于树干底部0-10 cm的土层范围内使用土壤取芯器采集土壤子样本, 重复3次且任意两个采样方向之间的角度约为120°, 将这3个子样本进行混合并剔除明显的根和凋落物等杂质(Xu et al., 1987; Yang et al., 2019a; Liu et al., 2020).然后, 利用烘干法测定每个样品的土壤水含量(g·g-1); 采用AQ400自动间断化学分析仪(SEAL Analytical, Mequon, USA)测量全氮含量(mg·g-1)和全磷含量(mg·g-1). ...
New perspectives on sub-seasonal xylem anatomical responses to climatic variability 2 2019
... 枝是重要的木质器官, 不仅起到机械支撑作用, 还具有水分、养分的运输和存贮功能(Pratt et al., 2007; Poorter et al., 2010; Meinzer et al., 2011; Ziaco & Liang, 2018).性状不同, 其生态功能也存在差异.枝的解剖学研究中, 木质部代表水分运输能力(Pratt & Jacobsen, 2017; Ziaco & Liang, 2018; Gebauer et al., 2019; Schumann et al., 2019); 韧皮部代表光合产物运输能力(Agustí & Blázquez, 2020); 髓是存储养分和水分的重要场所; 树脂道不仅能够产生、运输树脂, 还具有防御功能(Lin et al., 2001).相对于细根和树干, 枝能够更快地向叶片提供养分(Stratton et al., 2000; Pratt & Black, 2006).此外, 枝的解剖性状对于预测树木对环境变化的适应策略具有重要意义(Tarelkin et al., 2019), 然而, 目前关于枝的解剖性状的种内变异规律及影响因素的研究较少(Poorter et al., 2010). ...
... ; Ziaco & Liang, 2018; Gebauer et al., 2019; Schumann et al., 2019); 韧皮部代表光合产物运输能力(Agustí & Blázquez, 2020); 髓是存储养分和水分的重要场所; 树脂道不仅能够产生、运输树脂, 还具有防御功能(Lin et al., 2001).相对于细根和树干, 枝能够更快地向叶片提供养分(Stratton et al., 2000; Pratt & Black, 2006).此外, 枝的解剖性状对于预测树木对环境变化的适应策略具有重要意义(Tarelkin et al., 2019), 然而, 目前关于枝的解剖性状的种内变异规律及影响因素的研究较少(Poorter et al., 2010). ...
A potential role for xylem-phloem interactions in the hydraulic architecture of trees: effects of phloem girdling on xylem hydraulic conductance 2004
 ,*东北林业大学森林生态系统可持续经营教育部重点实验室, 哈尔滨 150040
,*东北林业大学森林生态系统可持续经营教育部重点实验室, 哈尔滨 150040
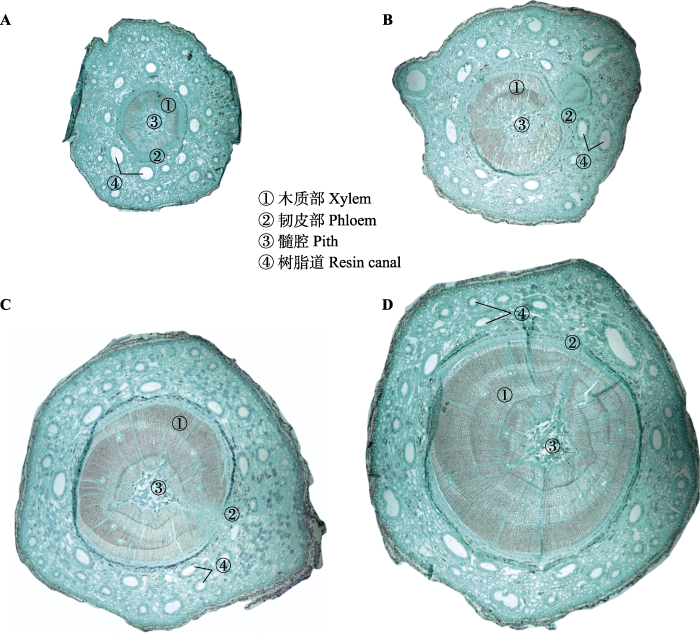 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT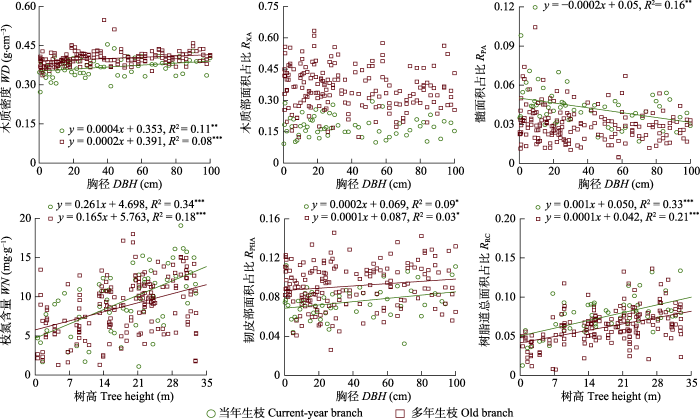 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT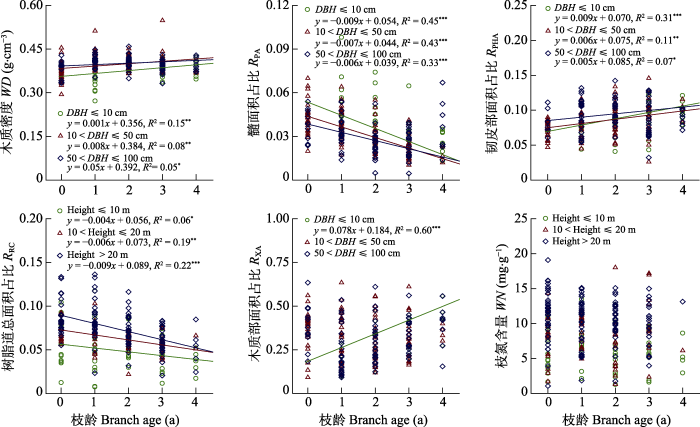 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT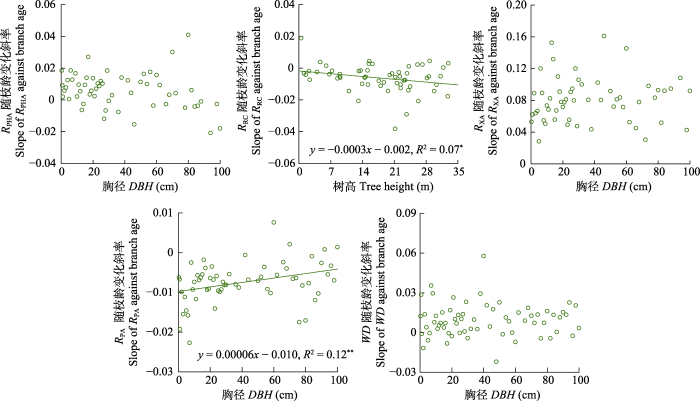 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}