Effects of waterlogging and increased soil nutrients on growth and reproduction of Polygonum hydropiper in the hydro-fluctuation belt of the Three Gorges Reservoir Region
Yu-Han CHEN1, Yi-Fu LUO1, Xin-Sheng SUN1, Guan-Wen WEI1, Wen-Jun HUANG2, Fang-Li LUO,1,*, Fei-Hai YU31School of Ecology and Nature Conservation, Beijing Forestry University, Beijing 100083, China 2Sichuan Academy of Forestry, Chengdu 610081, China 3Institute of Wetland Ecology & Clone Ecology/Zhejiang Provincial Key Laboratory of Plant Evolutionary Ecology and Conservation, Taizhou University, Taizhou, Zhejiang 318000, China
National Key R&D Program of China(2017YFC0505903) National Natural Science Foundation of China(31670428) the Fundamental Research Funds for the Central Universities(2015ZCQ-BH-01)
Abstract Aims Flooding and soil nutrients are the main environmental factors that affect plant growth in the hydro-fluctuation belt of the Three Gorges Reservoir Region (TGRR). Flooding intensity and concentration of soil nutrients experienced by riparian plants at different elevations of the hydro-fluctuation belt are different; therefore, we hypothesized that growth and reproductive responses of plants of the same species from different elevations to waterlogging and increased soil nutrients are also different. Methods In this study, the riparian species Polygonum hydropiper, which is widely distributed at low and high elevations of the hydro-fluctuation belt of the TGRR, was selected and its seeds were collected from natural populations. Effects of waterlogging and soil nutrients on growth and reproductive traits of P. hydropiper from high- and low-elevation areas were studied in a common-garden greenhouse experiment. Important findings Waterlogging significantly, or with marginal significance, decreased length and width of functional leaves, total branch number, leaf biomass, flower biomass, and total biomass of plants; low nutrient treatment significantly or marginal significantly decreased total node number, total branch number, root biomass, flower biomass, and total biomass of plants, indicating that both waterlogging and low soil nutrients inhibited growth and reproduction of P. hydropiper. Moreover, the interaction between waterlogging and soil nutrients significantly affected root biomass, showing higher root biomass accumulation at high soil nutrient conditions upon waterlogging. The high-elevation plants had significantly or marginal significantly higher leaf and root biomass than those from low elevation; however, flowering time of the low-elevation plants was significantly earlier, and reproduction allocation was higher than the high-elevation plants, indicating that resource allocation strategy was different between the high- and the low-elevation plants. The results indicate that growth and reproduction of P. hydropiper are inhibited by both waterlogging and soil nutrients, and this species has high adaptability at high soil nutrient conditions to waterlogging; meanwhile, low-elevation plants can adjust their growth and reproductive characteristics to improve their fitness under environmental stress. Keywords:Three Gorges Reservoir Region;waterlogging;soil nutrients;elevation;wetland plant;reproduction allocation
PDF (1285KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 陈禹含, 罗亦夫, 孙鑫晟, 魏冠文, 黄文军, 罗芳丽, 于飞海. 根部水淹和土壤养分提升对三峡库区消落带水蓼生长和繁殖特性的影响. 植物生态学报, 2020, 44(11): 1184-1194. DOI: 10.17521/cjpe.2020.0159 CHEN Yu-Han, LUO Yi-Fu, SUN Xin-Sheng, WEI Guan-Wen, HUANG Wen-Jun, LUO Fang-Li, YU Fei-Hai. Effects of waterlogging and increased soil nutrients on growth and reproduction of Polygonum hydropiper in the hydro-fluctuation belt of the Three Gorges Reservoir Region. Chinese Journal of Plant Ecology, 2020, 44(11): 1184-1194. DOI: 10.17521/cjpe.2020.0159
河(湖)岸带的土壤水分和养分含量往往沿垂直分布高度的不同而存在差异, 湿地或两栖植物如Ranunculus reptans和Solanum dulcamara分布在不同区域的植株通过种内分化, 表现出不同的形态适应特征以较好地适应所在生境(Lenssen et al., 2004; Zhang et al., 2016); 同时, 也有研究发现某些物种的种内分化即使在较小空间尺度下也能发生(Lenssen et al., 2004)。由于分布在消落带不同高程的水蓼植株长期经历不同的淹水强度和土壤养分环境, 因此, 我们假设来自不同高程的水蓼植株可能产生了性状分化, 从而形成不同的生长和繁殖策略以适应不同的淹水和土壤养分条件。为了验证以上假设, 本研究以三峡库区消落带高低高程区域的水蓼为研究对象, 在温室同质园条件下, 研究了不同的淹水和土壤养分处理对来源于高低高程水蓼植株的形态、生物量和繁殖性状的影响, 以期能明确不同高程水蓼植株对根部水淹和土壤养分提升的生长和繁殖响应, 为消落带植被的保护与恢复提供一定的理论依据。
1 材料和方法
1.1 实验材料与地点
在三峡库区消落带重庆北碚至湖北宜昌区域选取了3个样地, 在每个样地各选取高低高程的2个水蓼自然种群, 分别为: 重庆奉节县草堂镇梨木咀的高高程种群(173 m; 31.07° N, 109.60° E)和低高程种群(153 m; 31.07° N, 109.60° E), 重庆巴东县渡河镇老屋场的高高程种群(172 m; 31.20° N, 110.30° E)和低高程种群(156 m; 31.20° N, 110.30° E), 湖北秭归县归州镇官庄坪村乔家坝的高高程种群(166 m; 31.00° N, 110.76° E)和低高程种群(155 m; 31.00° N, 110.76° E)。在每个种群随机采集9株成熟水蓼植株的种子, 相邻植株间距至少2 m。种子自然风干并混合后, 用于温室实验。
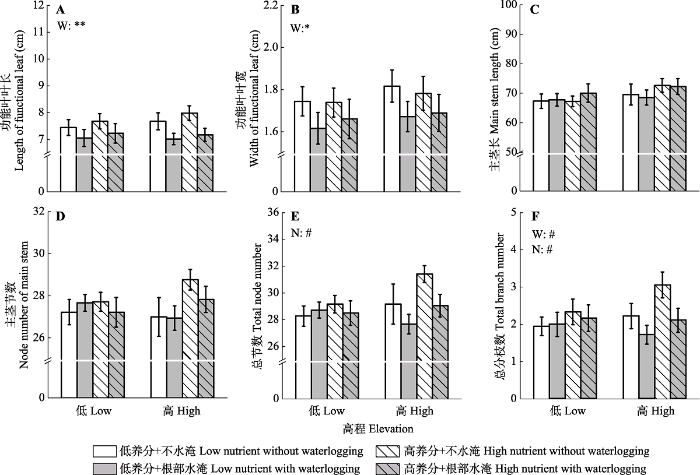
新窗口打开|下载原图ZIP|生成PPT 图1不同根部水淹(W)和土壤养分(N)处理下高低高程水蓼植株的功能叶叶长(A)、叶宽(B)、主茎长(C)?主茎节数(D)?总节数(E)和总分枝数(F)(平均值±标准误差)。**, p < 0.01; *, p < 0.05; #, 0.05 ≤ p < 0.1。
Fig. 1Length (A) and width (B) of functional leaf, main stem length (C), node number of main stem (D), total node number (E), and total branch number (F) of Polygonum hydropiper from high and low elevations under different waterlogging (W) and soil nutrients (N) treatments (mean ± SE). **, p < 0.01; *, p < 0.05; #, 0.05 ≤ p < 0.1.
Table 1 表1 表1根部水淹、土壤养分和高程对水蓼形态、生物量和繁殖性状的影响 Table 1Effects of waterlogging, soil nutrients, and elevation on morphological, biomass, and reproductive traits of Polygonum hydropiper
性状 Trait
根部水淹 Waterlogging (W)
养分 Nutrient (N)
高程 Elevation (E)
E × W
E × N
W × N
W × N × E
形态性状 Morphological trait
功能叶长 Length of functional leaf
7.91**
0.99
0.45
0.31
0.01
0.11
0.01
功能叶宽 Width of functional leaf
3.97*
0.01
0.80
0.02
0.07
0.21
0.00
主茎长 Main stem length
0.07
1.45
1.98
0.39
0.38
0.16
0.05
主茎节数 Node number of main stem
0.36
2.41
0.17
0.28
2.22
1.09
0.00
总节数 Total node number
2.74
3.04#
1.14
2.17
1.45
0.64
0.01
总分枝数 Total branch number
2.97#
3.87#
0.55
2.18
0.55
0.55
0.06
生物量性状 Biomass trait
根生物量 Root biomass
2.58
5.08*
3.03#
0.84
0.20
4.28*
0.71
茎生物量 Stem biomass
1.88
0.86
1.27
2.71
0.34
2.26
0.05
叶生物量 Leaf biomass
3.55#
0.74
5.44*
0.72
0.19
2.70
0.00
总生物量 Total biomass
6.74*
2.87#
0.46
2.43
0.52
1.20
0.14
繁殖性状 Reproductive trait
始花时间 First flowering time
0.37
0.09
4.05*
0.00
0.58
0.33
0.17
花生物量 Flower biomass
10.44**
2.98#
2.59
0.14
1.57
1.47
0.16
繁殖分配 Reproduction allocation
2.36
0.44
6.75*
0.50
0.23
2.48
0.30
表中数据为F值。方差检验的显著性水平: ***, p < 0.001; **, p < 0.01; *, p < 0.05; #, 0.05 ≤ p < 0.1; 无标注即为p ≥ 0.1。当p < 0.05时, F值加粗显示。始花时间、花生物量和繁殖分配的自由度为(1, 133), 其余指标的自由度均为(1, 135)。 Values are F and symbols show p (***, p < 0.001; **, p < 0.01; *, p < 0.05; #, 0.05 ≤ p < 0.1, and no symbols p ≥ 0.1). Values of p < 0.05 are in bold. Degree of freedom is (1, 133) for first flowering time, flower biomass and reproduction allocation, and degree of freedom is (1, 135) for other indexes.
新窗口打开|下载原图ZIP|生成PPT 图2不同根部水淹(W)和土壤养分(N)处理下高低高程(E)水蓼植株的根生物量(A)?茎生物量(B)?叶生物量(C)和总生物量(D)(平均值±标准误差)。*, p < 0.05; #, 0.05 ≤ p < 0.1。
Fig. 2Root biomass (A), stem biomass (B), leaf biomass (C), and total biomass (D) of Polygonum hydropiper from high and low elevations (E) under different waterlogging (W) and soil nutrients (N) treatments (mean ± SE). *, p < 0.05; #, 0.05 ≤ p < 0.1.
新窗口打开|下载原图ZIP|生成PPT 图3不同根部水淹(W)和土壤养分(N)处理下高低高程(E)水蓼植株的始花时间(A)?花生物量(B)和繁殖分配(C)(平均值±标准误差)。**, p < 0.01; *, p < 0.05; #, 0.05 ≤ p < 0.1。
Fig. 3First flowering time (A), flower biomass (B), and reproduction allocation (C) of Polygonum hydropiper from high and low elevation (E) under different waterlogging (W) and soil nutrients (N) treatments (mean ± SE). **, p < 0.01; *, p < 0.05; #, 0.05 ≤ p < 0.1.
Supplement I 附录I 附录I三峡水库2009-2018年蓄水期间高流量引起的水位上涨速率(m·d-1) Supplement IWater-level rise rate caused by the high flow of the Three Gorges Reservoir from 2009 to 2018 (m·d-1)
2009
2010
2011
2012
2013
2014
2015
2016
2017
2018
1.00
2.65
2.46
2.34
1.32
1.31
1.54
1.51
1.91
1.23
1.38
3.04
2.49
2.46
1.50
1.34
1.55
1.31
1.65
1.06
1.91
4.00
2.70
3.15
1.62
3.80
2.47
1.25
1.48
1.02
数据为每年水库蓄水期间水位上涨最快3天的数据。水位上涨速率根据2009-2018年全国水雨情信息网(http://xxfb.mwr.cn/sq_djdh.html)每日水位数据计算得出。 Data shown are the water-level rise rate of the fastest three days during the impoundment of the reservoir each year. The water level rise rate was calculated based on the daily water level data of the National Water and Rain Information Network (http://xxfb.mwr.cn/sq_djdh.html) from 2009 to 2018.
AbbottJM, StachowiczJJ (2016). The relative importance of trait vs. genetic differentiation for the outcome of interactions among plant genotypes Ecology, 97, 84-94. URLPMID:27008778 [本文引用: 1]
AyiQL, ZengB, YangK, LinF, ZhangXP, van BodegomPM, CornelissenJHC (2019). Similar growth performance but contrasting biomass allocation of root-flooded terrestrial plant Alternanthera philoxeroides (Mart.) Griseb. in response to nutrient versus dissolved oxygen stress Frontiers in Plant Science, 10, 111. DOI: 10.3389/fpls.2019.00111. URLPMID:30792729 [本文引用: 1]
BarkerHL, HoleskiLM, LindrothRL (2019). Independent and interactive effects of plant genotype and environment on plant traits and insect herbivore performance: a meta-analysis with Salicaceae Functional Ecology, 33, 422-435. [本文引用: 1]
ChenFQ, GuanSP, MaYR, XieZQ, LüK, HuangYW, JiaGM (2019a). Impact of regulated water level fluctuations on the sexual reproduction of remnant Myricaria laxiflora populations Global Ecology and Conservation, 18, e00628. DOI: 10.1016/j.gecco.2019.e00628. [本文引用: 1]
ChenFQ, LiY, QieGW, XuWN (2008). The morphological responses and endurance of Polygonum hydropiper to flooding stress Journal of Wuhan Botanical Research, 26, 142-146. [本文引用: 3]
ChenFQ, XieZQ (2007). Reproductive allocation, seed dispersal and germination of Myricaria laxiflora, an endangered species in the Three Gorges Reservoir area Plant Ecology, 191, 67-75. [本文引用: 1]
ChenFQ, XieZQ (2011). Ecophysiological Response of Two Herbaceous Species to Flooding Implication for Ecological Restoration of Vegetation on Water-level-fluctuating Zone. International Conference on Electronics, Communications and Control (ICECC). Ningbo, China. 4260-4263. [本文引用: 3]
ChenFQ, ZhangM, WuY, HuangYW (2020). Seed rain and seed bank of a draw-down zone and their similarities to vegetation under the regulated water-level fluctuation in Xiangxi River Journal of Freshwater Ecology, 35, 57-71. [本文引用: 2]
ChenXS, DengZM, XieYH, LiF, LiX (2014). Differential growth and vegetative reproduction by two co-occurring emergent macrophytes along a water table gradient Pakistan Journal of Botany, 46, 881-886. [本文引用: 1]
ChenXS, LiYF, CaiYH, XieYH, DengZM, LiF, HouZY (2019b). Differential strategies to tolerate flooding in Polygonum hydropiper plants originating from low- and high-elevation habitats Frontiers in Plant Science, 9, 1970. DOI: 10.3389/fpls.2018.01970. URLPMID:30687365 [本文引用: 4]
ChengRM, LiuZB, XiaoWF, WangN, WangXR, ShenYF (2017). Changes of soil chemical properties in typical hydro-fluctuation belt of Three Gorges Reservoir Scientia Silvae Sinicae, 53(2), 19-25. [本文引用: 1]
ColmerTD, VoesenekLACJ (2009). Flooding tolerance: suites of plant traits in variable environments Functional Plant Biology, 36, 665-681. URLPMID:32688679 [本文引用: 1]
DuH, ZhangXP, ZengB (2016). Dissolved oxygen in water affects the tolerance of two terrestrial plants, Alternanthera philoxeroides and Hemarthria altissima, to complete submergence Acta Ecologica Sinica, 36, 7562-7569. [本文引用: 1]
FanDY, XiongGM, ZhangAY, LiuX, XieZQ, LiZJ (2015). Effect of water-lever regulation on species selection for ecological restoration practice in the water-level fluctuation zone of Three Gorges Reservoir Chinese Journal of Plant Ecology, 39, 416-432. [本文引用: 3]
FanSF, YuHH, LiuCH, YuD, HanYQ, WangLG (2015). The effects of complete submergence on the morphological and biomass allocation response of the invasive plant Alternanthera philoxeroides Hydrobiologia, 746, 159-169.
FanXL, YangTH, GaoHY, ZhangZS, YangC, LiuMJ (2014). Effect of oxygen on the damage of photosynthetic apparatus in plant leaves induced by dark-submergence Plant Physiology Journal, 50, 542-548. [本文引用: 1]
GuYB, PanYW, ChenFY, LouYJ, TangZH (2019). Effects of water level and nitrogen concentration on growth and biomass allocation of Scirpus nipponicus seedlings Chinese Journal of Ecology, 38, 2302-2309. [本文引用: 1]
GuoY, YangS, ShenYF, XiaoWF, ChengRM (2019). Study on the natural distribution characteristics and community species diversity of existing plants in the Three Gorges Reservoir Acta Ecologica Sinica, 39, 4255-4265. [本文引用: 2]
HeRT, YangK, ZengB, LiR, NiuHG, ShiSH, AyiQL, SuXL (2020). Distribution pattern of vegetation in water-level fluctuation zone of the Three Gorges Reservoir as affected by differential flooding regimes Acta Ecologica Sinica, 40, 834-842. [本文引用: 1]
HeydelF, EngelsJG, FeigsJT, VásquezE, RudolphB, RohwerJG, JensenK (2017). Adaptation to tidal flooding and rapid genetic divergence between a narrow endemic grass species and its widespread congener lead to an early stage of ecological speciation Perspectives in Plant Ecology, Evolution and Systematics, 27, 57-67. [本文引用: 1]
HongM, GuoQS, NieBH, KangY, PeiSX, JinJQ, WangXF (2011). Responses of Cynodon dactylon population in hydro-fluctuation belt of Three Gorges Reservoir area to flooding-drying habitat change Chinese Journal of Applied Ecology, 22, 2829-2835. URLPMID:22303657 [本文引用: 1]
HuaJF, HanLW, WangZQ, GuCS, YinYL (2017). Morpho-anatomical and photosynthetic responses of Taxodium hybrid ‘Zhongshanshan’ 406 to prolonged flooding Flora, 231, 29-37. [本文引用: 1]
LeiB, WangYC, YouYF, ZhangS, YangCH (2014). Diversity and structure of herbaceous plant community in typical water-level-fluctuation zone with different spacing elevations in Three Gorges Reservior Journal of Lake Sciences, 26, 600-606. [本文引用: 1]
LenssenJPM, van KleunenM, FischerM, de KroonH (2004). Local adaptation of the clonal plant Ranunculus reptans to flooding along a small-scale gradient Journal of Ecology, 92, 696-706. [本文引用: 2]
LiQH, LiuSP, ZhiCY, LiXF, ChenFF, ZengQK (2013). Adaptation mechanism of three herbs in the water-level- fluctuation-zone of reservoir to complete submergence Journal of Tropical and Subtropical Botany, 21, 459-465. [本文引用: 1]
LiXX, LiCX, SongH, YuanZX (2019). Effects of flooding and planting density on the photosynthesis of Hemarthria compressa and Cynodon dactylon cottage seedlings Acta Prataculturae Sinica, 28, 197-206. [本文引用: 2]
LiYF, ChenXS, XiangWH, XieYH (2016). Effects of water levels on the growth and reproductive characteristics of Carex brevicuspis growing on sites with different elevations Acta Ecologica Sinica, 36, 1959-1966. [本文引用: 2]
LiuWW, WangJ, WangY, YangF (2012). The differences of plant community diversity among the different altitudes in the water-level-fluctuating zone of the Three Gorges Reservoir Acta Ecologica Sinica, 32, 5454-5466. [本文引用: 1]
LiuZB, ChengRM, XiaoWF, WangRL, FengXH, WangXR (2013). Effect of waterlogging on photosynthetic and physioecological characteristics of plants World Forestry Research, 26, 33-38. [本文引用: 3]
LuoMJ, ZhangSG, CuiLJ, TanFL, HuangYR (2012). Response of growth and biomass allocation of Aegiceras corniculatum to waterlogging stress Journal of Zhejiang Forestry Science and Technology, 32, 15-19. [本文引用: 1]
MonyC, MercierE, BonisA, BouzilléJB (2010). Reproductive strategies may explain plant tolerance to inundation: a mesocosm experiment using six marsh species Aquatic Botany, 92, 99-104. [本文引用: 1]
NamJM, KimJH, KimJG (2017). Effects of light intensity and plant density on growth and reproduction of the amphicarpic annual Persicaria thunbergii Aquatic Botany, 142, 119-122. [本文引用: 1]
PanY, XieYH, DengZM, TangY, PanDD (2014). High water level impedes the adaptation of Polygonum hydropiper to deep burial: responses of biomass allocation and root morphology Scientific Reports, 4, 5612. DOI: 10.1038/srep05612. URLPMID:25002329 [本文引用: 1]
ParkH, KimJG (2020). Temporal and spatial variations of vegetation in a riparian zone of South Korea Journal of Ecology and Environment, 44, 9. DOI: 10.1186/s41610-020-00152-z. [本文引用: 1]
PhukanUJ, MishraS, ShuklaRK (2016). Waterlogging and submergence stress: affects and acclimation Critical Reviews in Biotechnology, 36, 956-966. URLPMID:26177332 [本文引用: 1]
QinHW, GaoF, LiuZX, LiHL, ZhengLD, SuHY, MengJM (2017). Effects of light intensity on the growth and physiology of Cynodon dactylon seedlings under water submerged environment Acta Agrestia Sinica, 25, 675-679. [本文引用: 2]
SchaffSD, PezeshkiSR, ShieldsFD (2003). Effects of soil conditions on survival and growth of black willow cuttings Environmental Management, 31, 748-763. URLPMID:14565695 [本文引用: 1]
SoininenEM, Br?thenKA, JusdadoJGH, ReidingerS, HartleySE (2013). More than herbivory: levels of silica-based defences in grasses vary with plant species, genotype and location Oikos, 122, 30-41. DOI:10.1111/more.2013.122.issue-1URL [本文引用: 1]
StrikerGG, ManzurME, GrimoldiAA (2011). Increasing defoliation frequency constrains regrowth of the forage legume Lotus tenuis under flooding. The role of crown reserves Plant and Soil, 343, 261-272. [本文引用: 1]
SuXL, NilssonC, PilottoF, LiuSP, ShiSH, ZengB (2017). Soil erosion and deposition in the new shorelines of the Three Gorges Reservoir Science of the Total Environment, 599-600, 1485-1492. [本文引用: 1]
SuXL, ZengB, HuangWJ, XuSJ, LeiST (2012). Effects of the Three Gorges Dam on preupland and preriparian drawdown zones vegetation in the upper watershed of the Yangtze River, P. R. China Ecological Engineering, 44, 123-127. [本文引用: 1]
SuXL, ZengB, LinF, QiaoP, AyiQL, HuangWJ (2016). How does long-term complete submergence influence sex ratio and resource allocation of a dioecious shrub, Salix variegata Franch.? Ecological Engineering, 87, 218-223. [本文引用: 1]
SuXL, ZengB, QiaoP, AyiQL, HuangWJ (2010). The effects of winter water submergence on flowering phenology and reproductive allocation of Salix variegata Franch. in Three Gorges reservoir region Acta Ecologica Sinica, 30, 2585-2592. [本文引用: 2]
TaoM, BaoDC, JiangMX (2011). Effects of submergence on seed germination of nine annual plant species in the Three Gorges Reservoir region and their implication to vegetation restoration Acta Ecologica Sinica, 31, 906-913. [本文引用: 1]
VillellasJ, GarcíaMB (2018). Life-history trade-offs vary with resource availability across the geographic range of a widespread plant Plant Biology, 20, 483-489. URLPMID:29247581 [本文引用: 1]
VoesenekLACJ, ColmerTD, PierikR, MillenaarFF, PeetersAJM (2006). How plants cope with complete submergence New Phytologist, 170, 213-226. [本文引用: 1]
WangC, FangF, YuanZY, ZhangR, ZhangW, GuoJS (2020). Spatial variations of soil phosphorus forms and the risks of phosphorus release in the water-level fluctuation zone in a tributary of the Three Gorges Reservoir Science of the Total Environment, 699, 134124. DOI: 10.1016/j.scitotenv. 2019.134124. [本文引用: 1]
WangP, ZhangQ, XuYS, YuFH (2016). Effects of water level fluctuation on the growth of submerged macrophyte communities Flora, 223, 83-89. [本文引用: 1]
WangYC, LeiB, ZhangS (2012). Differences in vegetation and soil characteristics at different water-level altitudes in the drawdown areas of Three Gorges Reservoir area Journal of Lake Sciences, 24, 206-212. [本文引用: 1]
WangYJ, ChenFQ, ZhangM, ChenSH (2016a). Response of soil nutrient levels and spatial distribution to water-level fluctuation on the shanmu riverbanks in the Three Gorges Reservoir area Journal of Hydroecology, 37, 56-61. [本文引用: 2]
WangYJ, ChenFQ, ZhangM, WuY, ChenSH (2016b). Characteristics of soil nutrient and spatial distribution on riparian zone restored by different vegetation restoration methods at Wanzhou section in the Three Gorges Reservoir area, China Journal of Agricultural Resources and Environment, 33, 127-133. [本文引用: 2]
WeiGW, SunXS, ChenYH, LuoFL, YuFH (2020). Growth and reproductive responses of Polygonum hydropiper populations to elevational difference associated with flooding Global Ecology and Conservation, 23, e01156. DOI: 10.1016/j.gecco.2020.e01156. URLPMID:32292803 [本文引用: 1]
WuJG, HuangJH, HanXG, GaoXM, HeFL, JiangMX, JiangZG, PrimackRB, ShenZH (2004). The Three Gorges Dam: an ecological perspective Frontiers in Ecology and the Environment, 2, 241-248. [本文引用: 1]
XuJP, ZhangXP, ZengB, YuanSH, LiuJH, LiuMZ (2014). Effects of light and dissolved oxygen on the phenotypic plasticity of Alternanthera philoxeroides in submergence conditions Acta Ecologica Sinica, 34, 258-268. [本文引用: 2]
XuJY, ChenHM, WangXL (2016). A review on water depth effect on the growth and reproduction of plants in the wetlands Wetland Science, 14, 725-732. [本文引用: 1]
YanH, LiuRQ, LiuZN, WangX, LuoWB, ShengLX (2015). Growth and physiological responses to water depths in Carex schmidtii meinsh PLOS ONE, 10, e0128176. DOI: 10.1371/journal.pone.0128176. URLPMID:26720417 [本文引用: 1]
YeC, ChenCR, ButlerOM, RashtiMR, EsfandbodM, DuM, ZhangQF (2019). Spatial and temporal dynamics of nutrients in riparian soils after nine years of operation of the Three Gorges Reservoir, China Science of the Total Environment, 664, 841-850. [本文引用: 1]
YeXQ, ZengB, MengJL, WuM, ZhangXP (2018). Responses in shoot elongation, carbohydrate utilization and growth recovery of an invasive species to submergence at different water temperatures Scientific Reports, 8, 306. DOI: 10.1038/s41598-017-18735-7. URLPMID:29321607 [本文引用: 1]
YouYF, YangCH, LeiB, ZhangS, WangYC, LiuJH (2017). Effect of water level regulation on vegetation characteristics in the water-level-fluctuation zone of the Three Gorges Reservoir Chinese Journal of Applied and Environmental Biology, 23, 1103-1109. [本文引用: 1]
YuanSH, ZengB, SuXL, XuJP (2014). Effect of water-level fluctuation discrepancy on the composition of different annuals in Three Gorges Reservoir drawdown zone Acta Ecologica Sinica, 34, 6481-6488. [本文引用: 1]
ZhangAY, XiongGM, FanDY, YangD, XieZQ (2018). Effects of Three Gorges Dam on riparian vascular plants of the main stream of Yangtze River Resources and Environment in the Yangtze Basin, 27, 145-156. [本文引用: 1]
ZhangQ, PetersJL, VisserEJW, de KroonH, HuberH (2016). Hydrologically contrasting environments induce genetic but not phenotypic differentiation in Solanum dulcamara Journal of Ecology, 104, 1649-1661. [本文引用: 1]
ZhangY, CaiJG, SunOW, ShiJJ (2019). Research on photosynthetic responses mechanisms of Hydrangea macrophylla under waterlogging stress Journal of Nuclear Agricultural Sciences, 33, 808-815. [本文引用: 2]
ZhangZY, ChengYC, ChengL, WanCY, LiJB (2016). Characteristics of vegetation and soil in the water level fluctuation zone of the Wanzhou region of Three Gorges Reservoir Journal of Hydroecology, 37, 24-33. [本文引用: 1]
ZhaoQ, ChenJB (2018). Study and practice on ecological restoration strategy of the hydro-fluctuation belt in the Three Gorges Reservoir area Journal of Anhui Agricultural Sciences, 46, 5-7. [本文引用: 1]
ZhouWG, ChenF, MengYJ, ChandrasekaranU, LuoXF, YangWY, ShuK (2020). Plant waterlogging/flooding stress responses: from seed germination to maturation Plant Physiology and Biochemistry, 148, 228-236. DOI:10.1016/j.plaphy.2020.01.020URLPMID:31981875 [本文引用: 1] Global climate change is strongly associated with variations in precipitation and flooding events. Flooding usually causes submergence- or partial submergence stress in plants, which significantly has a negative influence on agricultural production, from seed germination to vegetative and reproductive growth. Flooding stress results in crop growth under low oxygen conditions and thus, negatively affects the developmental periods of plant lifecycle. The survival strategies of different plant species under this stressful condition are distinct, whereas the perception pathways associated with flooding stress are similar at the molecular level. Plants respond to flooding stress by mediating changes in their architecture, energy metabolism, photosynthesis, respiration and endogenous phytohormone biosynthesis/signaling, because aerobic respiration is inhibited under flooding stress, the decrease of energy metabolism further constrains plant development. Consequently, to acclimate under these unfavorable conditions, the anaerobic respiration cascade must be promoted. In this updated review, we primarily focus on recent advances in our understanding of the mechanisms underlying plant responses to flooding stress. We summarize the functions of the flooding response factors involved in energy metabolism and phytohormone biosynthesis/signaling cascades. Finally, the current understanding of how plants circumvent flooding stress, and the potential challenges for future research, are discussed.
ZhuQ, ZhangZY, HuHQ, WanCY, HuL, LiuH (2014). Soil property changes under drain-flooding condition in Xiaojiang water-level-fluctuating belt of the Three-Gorge- Reservoir region Soils, 46, 927-932. [本文引用: 1]
Similar growth performance but contrasting biomass allocation of root-flooded terrestrial plant Alternanthera philoxeroides (Mart.) Griseb. in response to nutrient versus dissolved oxygen stress 1 2019
Independent and interactive effects of plant genotype and environment on plant traits and insect herbivore performance: a meta-analysis with Salicaceae 1 2019
Impact of regulated water level fluctuations on the sexual reproduction of remnant Myricaria laxiflora populations 1 2019
... 三峡工程是世界上最大的水利水电枢纽工程(Wu et al., 2004).三峡水库夏季水位降至145 m, 冬季蓄水至175 m, 形成了与自然河流相反的“冬蓄夏排”周期性水位变化节律(王业春等, 2012; 赵琴和陈教斌, 2018).由于大坝的截流作用, 水库内水流速度非常缓慢, 接近于静止水体(Su et al., 2017).水位周期性变化形成大面积的消落带区域, 其高低高程水位落差最大可达30 m (雷波等, 2014; 郭燕等, 2019).三峡库区高高程区域的植物经历的水淹时间较短且深度较浅, 而低高程区域的植物经历的水淹时间较长且深度较深(刘维暐等, 2012; Chen et al., 2019b).除了三峡库区以外, 国内外河流自然消落区的高程落差很少能达到10 m以上, 通过控制实验就更难实现较大的高程差(Schaff et al., 2003; Park & Kim, 2020).因此, 目前针对高程对河岸带植物生长和繁殖特性影响的研究仍较为缺乏.仅有少数****对高程的效应进行了初步的探讨, 但其研究样地海拔落差仍不超过5 m (Chen et al., 2019a, 2019b). ...
水蓼对水淹胁迫的耐受能力和形态学响应 3 2008
... 水蓼(Polygonum hydropiper)是蓼科蓼属(Polygonum)的一年生草本植物, 自然生长在河(湖)岸边, 在我国分布较为广泛(李安仁, 1998).同时, 该物种也具有非常发达的地下根状茎, 节处可着生细根和幼芽(Chen et al., 2019b).植株可以以地下根状茎越冬, 翌年春季萌发形成新的分株.水蓼对湿地自然水位变化具有较强的适应能力, 能适应陆生和水生生境, 是很多湿地的优势植物群落类型(Chen et al., 2020).已有研究表明水蓼对水淹具有一定的适应性(陈芳清等, 2008; Chen & Xie, 2011).在淹水条件下, 植株可以通过改变叶片形状、形成不定根以及改变生物量分配等方式来适应淹水环境(Pan et al., 2014).该物种是三峡库区消落带的常见物种, 在消落带的不同高程具有大面积的自然分布(Chen et al., 2020). ...
Differential strategies to tolerate flooding in Polygonum hydropiper plants originating from low- and high-elevation habitats 4 2019
... 三峡工程是世界上最大的水利水电枢纽工程(Wu et al., 2004).三峡水库夏季水位降至145 m, 冬季蓄水至175 m, 形成了与自然河流相反的“冬蓄夏排”周期性水位变化节律(王业春等, 2012; 赵琴和陈教斌, 2018).由于大坝的截流作用, 水库内水流速度非常缓慢, 接近于静止水体(Su et al., 2017).水位周期性变化形成大面积的消落带区域, 其高低高程水位落差最大可达30 m (雷波等, 2014; 郭燕等, 2019).三峡库区高高程区域的植物经历的水淹时间较短且深度较浅, 而低高程区域的植物经历的水淹时间较长且深度较深(刘维暐等, 2012; Chen et al., 2019b).除了三峡库区以外, 国内外河流自然消落区的高程落差很少能达到10 m以上, 通过控制实验就更难实现较大的高程差(Schaff et al., 2003; Park & Kim, 2020).因此, 目前针对高程对河岸带植物生长和繁殖特性影响的研究仍较为缺乏.仅有少数****对高程的效应进行了初步的探讨, 但其研究样地海拔落差仍不超过5 m (Chen et al., 2019a, 2019b). ...
... , 2019b). ...
... 水蓼(Polygonum hydropiper)是蓼科蓼属(Polygonum)的一年生草本植物, 自然生长在河(湖)岸边, 在我国分布较为广泛(李安仁, 1998).同时, 该物种也具有非常发达的地下根状茎, 节处可着生细根和幼芽(Chen et al., 2019b).植株可以以地下根状茎越冬, 翌年春季萌发形成新的分株.水蓼对湿地自然水位变化具有较强的适应能力, 能适应陆生和水生生境, 是很多湿地的优势植物群落类型(Chen et al., 2020).已有研究表明水蓼对水淹具有一定的适应性(陈芳清等, 2008; Chen & Xie, 2011).在淹水条件下, 植株可以通过改变叶片形状、形成不定根以及改变生物量分配等方式来适应淹水环境(Pan et al., 2014).该物种是三峡库区消落带的常见物种, 在消落带的不同高程具有大面积的自然分布(Chen et al., 2020). ...
... 高高程水蓼植株的叶和根生物量显著或趋于显著高于低高程植株(图2), 表明高高程植株将更多的生物量投入到营养生长.根据三峡水库的水位调度方案, 与低高程植株相比, 每年消落带高高程区域被水淹的时间较晚而退水的时间较早(Su et al., 2012; Fan et al., 2015; 樊大勇等, 2015; 由永飞等, 2017).因此, 高高程植株水淹间歇的恢复时间比低高程植株更长; 植株将更多的生物量分配到叶和根的生长, 有利于植株光合产物的积累和对土壤养分的吸收(Yan et al., 2015; 徐金英等, 2016; Chen et al., 2019b).已有的研究也发现在相对较浅的水淹环境下河岸带植物如狗牙根、三江藨草(Scirpus nipponicus)和桐花树(Aegiceras corniculatum)等具有较高的叶和根生物量分配(洪明等, 2011; 罗美娟等, 2012; 古勇波等, 2019). ...
... 三峡工程是世界上最大的水利水电枢纽工程(Wu et al., 2004).三峡水库夏季水位降至145 m, 冬季蓄水至175 m, 形成了与自然河流相反的“冬蓄夏排”周期性水位变化节律(王业春等, 2012; 赵琴和陈教斌, 2018).由于大坝的截流作用, 水库内水流速度非常缓慢, 接近于静止水体(Su et al., 2017).水位周期性变化形成大面积的消落带区域, 其高低高程水位落差最大可达30 m (雷波等, 2014; 郭燕等, 2019).三峡库区高高程区域的植物经历的水淹时间较短且深度较浅, 而低高程区域的植物经历的水淹时间较长且深度较深(刘维暐等, 2012; Chen et al., 2019b).除了三峡库区以外, 国内外河流自然消落区的高程落差很少能达到10 m以上, 通过控制实验就更难实现较大的高程差(Schaff et al., 2003; Park & Kim, 2020).因此, 目前针对高程对河岸带植物生长和繁殖特性影响的研究仍较为缺乏.仅有少数****对高程的效应进行了初步的探讨, 但其研究样地海拔落差仍不超过5 m (Chen et al., 2019a, 2019b). ...
... 三峡工程是世界上最大的水利水电枢纽工程(Wu et al., 2004).三峡水库夏季水位降至145 m, 冬季蓄水至175 m, 形成了与自然河流相反的“冬蓄夏排”周期性水位变化节律(王业春等, 2012; 赵琴和陈教斌, 2018).由于大坝的截流作用, 水库内水流速度非常缓慢, 接近于静止水体(Su et al., 2017).水位周期性变化形成大面积的消落带区域, 其高低高程水位落差最大可达30 m (雷波等, 2014; 郭燕等, 2019).三峡库区高高程区域的植物经历的水淹时间较短且深度较浅, 而低高程区域的植物经历的水淹时间较长且深度较深(刘维暐等, 2012; Chen et al., 2019b).除了三峡库区以外, 国内外河流自然消落区的高程落差很少能达到10 m以上, 通过控制实验就更难实现较大的高程差(Schaff et al., 2003; Park & Kim, 2020).因此, 目前针对高程对河岸带植物生长和繁殖特性影响的研究仍较为缺乏.仅有少数****对高程的效应进行了初步的探讨, 但其研究样地海拔落差仍不超过5 m (Chen et al., 2019a, 2019b). ...
Adaptation to tidal flooding and rapid genetic divergence between a narrow endemic grass species and its widespread congener lead to an early stage of ecological speciation 1 2017
... 应对周期性的自然水淹, 河岸带植物表现出形态性状上的适应性, 比如通气组织的形成、叶片变薄、茎伸长生长以及气生根的生成等(Phukan et al., 2016; Wang et al., 2016; Heydel et al., 2017; Hua et al., 2017); 同时, 河岸带植物的繁殖特性也随着淹水而发生变化, 水淹耐受能力较强的物种往往能有效地利用退水或两次淹水中间的间隙快速地完成其繁殖过程; 这些植物可以通过调整繁殖期、缩短花期以降低或避免淹水对繁殖过程的影响, 例如稗(Echinochloa crus-galli)的种子可在水位下降后迅速萌发以提高其子代的存活率(Colmer & Voesenek, 2009; 陶敏等, 2011; 樊大勇等, 2015; Su et al., 2016).河岸带不同高程的植物所经历的水淹胁迫往往是不同的, 分布在高高程的植物经历的淹水频率较低, 时长更短; 而分布在低高程的植物经历的淹水频率更高且时长更长(Chen & Xie, 2007; 袁慎鸿等, 2014).已有研究表明不同高程的植物群落特征如物种数和优势种存在较大差异, 物种数随高程的升高表现出增加的趋势(张爱英等, 2018).由于长期遭受不同的淹水强度, 河岸带不同高程植物所受到环境选择压力也是不同的; 因此, 同一物种分布在不同高程的植株的生长和繁殖特性也可能存在差异. ...
三峡库区消落带狗牙根种群对水陆生境变化的响应 1 2011
... 高高程水蓼植株的叶和根生物量显著或趋于显著高于低高程植株(图2), 表明高高程植株将更多的生物量投入到营养生长.根据三峡水库的水位调度方案, 与低高程植株相比, 每年消落带高高程区域被水淹的时间较晚而退水的时间较早(Su et al., 2012; Fan et al., 2015; 樊大勇等, 2015; 由永飞等, 2017).因此, 高高程植株水淹间歇的恢复时间比低高程植株更长; 植株将更多的生物量分配到叶和根的生长, 有利于植株光合产物的积累和对土壤养分的吸收(Yan et al., 2015; 徐金英等, 2016; Chen et al., 2019b).已有的研究也发现在相对较浅的水淹环境下河岸带植物如狗牙根、三江藨草(Scirpus nipponicus)和桐花树(Aegiceras corniculatum)等具有较高的叶和根生物量分配(洪明等, 2011; 罗美娟等, 2012; 古勇波等, 2019). ...
三峡库区消落带狗牙根种群对水陆生境变化的响应 1 2011
... 高高程水蓼植株的叶和根生物量显著或趋于显著高于低高程植株(图2), 表明高高程植株将更多的生物量投入到营养生长.根据三峡水库的水位调度方案, 与低高程植株相比, 每年消落带高高程区域被水淹的时间较晚而退水的时间较早(Su et al., 2012; Fan et al., 2015; 樊大勇等, 2015; 由永飞等, 2017).因此, 高高程植株水淹间歇的恢复时间比低高程植株更长; 植株将更多的生物量分配到叶和根的生长, 有利于植株光合产物的积累和对土壤养分的吸收(Yan et al., 2015; 徐金英等, 2016; Chen et al., 2019b).已有的研究也发现在相对较浅的水淹环境下河岸带植物如狗牙根、三江藨草(Scirpus nipponicus)和桐花树(Aegiceras corniculatum)等具有较高的叶和根生物量分配(洪明等, 2011; 罗美娟等, 2012; 古勇波等, 2019). ...
Morpho-anatomical and photosynthetic responses of Taxodium hybrid ‘Zhongshanshan’ 406 to prolonged flooding 1 2017
... 应对周期性的自然水淹, 河岸带植物表现出形态性状上的适应性, 比如通气组织的形成、叶片变薄、茎伸长生长以及气生根的生成等(Phukan et al., 2016; Wang et al., 2016; Heydel et al., 2017; Hua et al., 2017); 同时, 河岸带植物的繁殖特性也随着淹水而发生变化, 水淹耐受能力较强的物种往往能有效地利用退水或两次淹水中间的间隙快速地完成其繁殖过程; 这些植物可以通过调整繁殖期、缩短花期以降低或避免淹水对繁殖过程的影响, 例如稗(Echinochloa crus-galli)的种子可在水位下降后迅速萌发以提高其子代的存活率(Colmer & Voesenek, 2009; 陶敏等, 2011; 樊大勇等, 2015; Su et al., 2016).河岸带不同高程的植物所经历的水淹胁迫往往是不同的, 分布在高高程的植物经历的淹水频率较低, 时长更短; 而分布在低高程的植物经历的淹水频率更高且时长更长(Chen & Xie, 2007; 袁慎鸿等, 2014).已有研究表明不同高程的植物群落特征如物种数和优势种存在较大差异, 物种数随高程的升高表现出增加的趋势(张爱英等, 2018).由于长期遭受不同的淹水强度, 河岸带不同高程植物所受到环境选择压力也是不同的; 因此, 同一物种分布在不同高程的植株的生长和繁殖特性也可能存在差异. ...
三峡水库不同间距高程消落带草本植物群落物种多样性与结构特征 1 2014
... 三峡工程是世界上最大的水利水电枢纽工程(Wu et al., 2004).三峡水库夏季水位降至145 m, 冬季蓄水至175 m, 形成了与自然河流相反的“冬蓄夏排”周期性水位变化节律(王业春等, 2012; 赵琴和陈教斌, 2018).由于大坝的截流作用, 水库内水流速度非常缓慢, 接近于静止水体(Su et al., 2017).水位周期性变化形成大面积的消落带区域, 其高低高程水位落差最大可达30 m (雷波等, 2014; 郭燕等, 2019).三峡库区高高程区域的植物经历的水淹时间较短且深度较浅, 而低高程区域的植物经历的水淹时间较长且深度较深(刘维暐等, 2012; Chen et al., 2019b).除了三峡库区以外, 国内外河流自然消落区的高程落差很少能达到10 m以上, 通过控制实验就更难实现较大的高程差(Schaff et al., 2003; Park & Kim, 2020).因此, 目前针对高程对河岸带植物生长和繁殖特性影响的研究仍较为缺乏.仅有少数****对高程的效应进行了初步的探讨, 但其研究样地海拔落差仍不超过5 m (Chen et al., 2019a, 2019b). ...
三峡水库不同间距高程消落带草本植物群落物种多样性与结构特征 1 2014
... 三峡工程是世界上最大的水利水电枢纽工程(Wu et al., 2004).三峡水库夏季水位降至145 m, 冬季蓄水至175 m, 形成了与自然河流相反的“冬蓄夏排”周期性水位变化节律(王业春等, 2012; 赵琴和陈教斌, 2018).由于大坝的截流作用, 水库内水流速度非常缓慢, 接近于静止水体(Su et al., 2017).水位周期性变化形成大面积的消落带区域, 其高低高程水位落差最大可达30 m (雷波等, 2014; 郭燕等, 2019).三峡库区高高程区域的植物经历的水淹时间较短且深度较浅, 而低高程区域的植物经历的水淹时间较长且深度较深(刘维暐等, 2012; Chen et al., 2019b).除了三峡库区以外, 国内外河流自然消落区的高程落差很少能达到10 m以上, 通过控制实验就更难实现较大的高程差(Schaff et al., 2003; Park & Kim, 2020).因此, 目前针对高程对河岸带植物生长和繁殖特性影响的研究仍较为缺乏.仅有少数****对高程的效应进行了初步的探讨, 但其研究样地海拔落差仍不超过5 m (Chen et al., 2019a, 2019b). ...
Local adaptation of the clonal plant Ranunculus reptans to flooding along a small-scale gradient 2 2004
... 河(湖)岸带的土壤水分和养分含量往往沿垂直分布高度的不同而存在差异, 湿地或两栖植物如Ranunculus reptans和Solanum dulcamara分布在不同区域的植株通过种内分化, 表现出不同的形态适应特征以较好地适应所在生境(Lenssen et al., 2004; Zhang et al., 2016); 同时, 也有研究发现某些物种的种内分化即使在较小空间尺度下也能发生(Lenssen et al., 2004).由于分布在消落带不同高程的水蓼植株长期经历不同的淹水强度和土壤养分环境, 因此, 我们假设来自不同高程的水蓼植株可能产生了性状分化, 从而形成不同的生长和繁殖策略以适应不同的淹水和土壤养分条件.为了验证以上假设, 本研究以三峡库区消落带高低高程区域的水蓼为研究对象, 在温室同质园条件下, 研究了不同的淹水和土壤养分处理对来源于高低高程水蓼植株的形态、生物量和繁殖性状的影响, 以期能明确不同高程水蓼植株对根部水淹和土壤养分提升的生长和繁殖响应, 为消落带植被的保护与恢复提供一定的理论依据. ...
... 三峡工程是世界上最大的水利水电枢纽工程(Wu et al., 2004).三峡水库夏季水位降至145 m, 冬季蓄水至175 m, 形成了与自然河流相反的“冬蓄夏排”周期性水位变化节律(王业春等, 2012; 赵琴和陈教斌, 2018).由于大坝的截流作用, 水库内水流速度非常缓慢, 接近于静止水体(Su et al., 2017).水位周期性变化形成大面积的消落带区域, 其高低高程水位落差最大可达30 m (雷波等, 2014; 郭燕等, 2019).三峡库区高高程区域的植物经历的水淹时间较短且深度较浅, 而低高程区域的植物经历的水淹时间较长且深度较深(刘维暐等, 2012; Chen et al., 2019b).除了三峡库区以外, 国内外河流自然消落区的高程落差很少能达到10 m以上, 通过控制实验就更难实现较大的高程差(Schaff et al., 2003; Park & Kim, 2020).因此, 目前针对高程对河岸带植物生长和繁殖特性影响的研究仍较为缺乏.仅有少数****对高程的效应进行了初步的探讨, 但其研究样地海拔落差仍不超过5 m (Chen et al., 2019a, 2019b). ...
三峡水库消落区不同海拔高度的植物群落多样性差异 1 2012
... 三峡工程是世界上最大的水利水电枢纽工程(Wu et al., 2004).三峡水库夏季水位降至145 m, 冬季蓄水至175 m, 形成了与自然河流相反的“冬蓄夏排”周期性水位变化节律(王业春等, 2012; 赵琴和陈教斌, 2018).由于大坝的截流作用, 水库内水流速度非常缓慢, 接近于静止水体(Su et al., 2017).水位周期性变化形成大面积的消落带区域, 其高低高程水位落差最大可达30 m (雷波等, 2014; 郭燕等, 2019).三峡库区高高程区域的植物经历的水淹时间较短且深度较浅, 而低高程区域的植物经历的水淹时间较长且深度较深(刘维暐等, 2012; Chen et al., 2019b).除了三峡库区以外, 国内外河流自然消落区的高程落差很少能达到10 m以上, 通过控制实验就更难实现较大的高程差(Schaff et al., 2003; Park & Kim, 2020).因此, 目前针对高程对河岸带植物生长和繁殖特性影响的研究仍较为缺乏.仅有少数****对高程的效应进行了初步的探讨, 但其研究样地海拔落差仍不超过5 m (Chen et al., 2019a, 2019b). ...
High water level impedes the adaptation of Polygonum hydropiper to deep burial: responses of biomass allocation and root morphology 1 2014
... 水蓼(Polygonum hydropiper)是蓼科蓼属(Polygonum)的一年生草本植物, 自然生长在河(湖)岸边, 在我国分布较为广泛(李安仁, 1998).同时, 该物种也具有非常发达的地下根状茎, 节处可着生细根和幼芽(Chen et al., 2019b).植株可以以地下根状茎越冬, 翌年春季萌发形成新的分株.水蓼对湿地自然水位变化具有较强的适应能力, 能适应陆生和水生生境, 是很多湿地的优势植物群落类型(Chen et al., 2020).已有研究表明水蓼对水淹具有一定的适应性(陈芳清等, 2008; Chen & Xie, 2011).在淹水条件下, 植株可以通过改变叶片形状、形成不定根以及改变生物量分配等方式来适应淹水环境(Pan et al., 2014).该物种是三峡库区消落带的常见物种, 在消落带的不同高程具有大面积的自然分布(Chen et al., 2020). ...
Temporal and spatial variations of vegetation in a riparian zone of South Korea 1 2020
... 三峡工程是世界上最大的水利水电枢纽工程(Wu et al., 2004).三峡水库夏季水位降至145 m, 冬季蓄水至175 m, 形成了与自然河流相反的“冬蓄夏排”周期性水位变化节律(王业春等, 2012; 赵琴和陈教斌, 2018).由于大坝的截流作用, 水库内水流速度非常缓慢, 接近于静止水体(Su et al., 2017).水位周期性变化形成大面积的消落带区域, 其高低高程水位落差最大可达30 m (雷波等, 2014; 郭燕等, 2019).三峡库区高高程区域的植物经历的水淹时间较短且深度较浅, 而低高程区域的植物经历的水淹时间较长且深度较深(刘维暐等, 2012; Chen et al., 2019b).除了三峡库区以外, 国内外河流自然消落区的高程落差很少能达到10 m以上, 通过控制实验就更难实现较大的高程差(Schaff et al., 2003; Park & Kim, 2020).因此, 目前针对高程对河岸带植物生长和繁殖特性影响的研究仍较为缺乏.仅有少数****对高程的效应进行了初步的探讨, 但其研究样地海拔落差仍不超过5 m (Chen et al., 2019a, 2019b). ...
Waterlogging and submergence stress: affects and acclimation 1 2016
... 应对周期性的自然水淹, 河岸带植物表现出形态性状上的适应性, 比如通气组织的形成、叶片变薄、茎伸长生长以及气生根的生成等(Phukan et al., 2016; Wang et al., 2016; Heydel et al., 2017; Hua et al., 2017); 同时, 河岸带植物的繁殖特性也随着淹水而发生变化, 水淹耐受能力较强的物种往往能有效地利用退水或两次淹水中间的间隙快速地完成其繁殖过程; 这些植物可以通过调整繁殖期、缩短花期以降低或避免淹水对繁殖过程的影响, 例如稗(Echinochloa crus-galli)的种子可在水位下降后迅速萌发以提高其子代的存活率(Colmer & Voesenek, 2009; 陶敏等, 2011; 樊大勇等, 2015; Su et al., 2016).河岸带不同高程的植物所经历的水淹胁迫往往是不同的, 分布在高高程的植物经历的淹水频率较低, 时长更短; 而分布在低高程的植物经历的淹水频率更高且时长更长(Chen & Xie, 2007; 袁慎鸿等, 2014).已有研究表明不同高程的植物群落特征如物种数和优势种存在较大差异, 物种数随高程的升高表现出增加的趋势(张爱英等, 2018).由于长期遭受不同的淹水强度, 河岸带不同高程植物所受到环境选择压力也是不同的; 因此, 同一物种分布在不同高程的植株的生长和繁殖特性也可能存在差异. ...
Effects of soil conditions on survival and growth of black willow cuttings 1 2003
... 三峡工程是世界上最大的水利水电枢纽工程(Wu et al., 2004).三峡水库夏季水位降至145 m, 冬季蓄水至175 m, 形成了与自然河流相反的“冬蓄夏排”周期性水位变化节律(王业春等, 2012; 赵琴和陈教斌, 2018).由于大坝的截流作用, 水库内水流速度非常缓慢, 接近于静止水体(Su et al., 2017).水位周期性变化形成大面积的消落带区域, 其高低高程水位落差最大可达30 m (雷波等, 2014; 郭燕等, 2019).三峡库区高高程区域的植物经历的水淹时间较短且深度较浅, 而低高程区域的植物经历的水淹时间较长且深度较深(刘维暐等, 2012; Chen et al., 2019b).除了三峡库区以外, 国内外河流自然消落区的高程落差很少能达到10 m以上, 通过控制实验就更难实现较大的高程差(Schaff et al., 2003; Park & Kim, 2020).因此, 目前针对高程对河岸带植物生长和繁殖特性影响的研究仍较为缺乏.仅有少数****对高程的效应进行了初步的探讨, 但其研究样地海拔落差仍不超过5 m (Chen et al., 2019a, 2019b). ...
More than herbivory: levels of silica-based defences in grasses vary with plant species, genotype and location 1 2013
Increasing defoliation frequency constrains regrowth of the forage legume Lotus tenuis under flooding. The role of crown reserves 1 2011
... 在三峡库区消落带区域, 淹水和土壤养分往往共同影响河岸带植物的生长和发育(张志永等, 2016; Zhou et al., 2020).本研究结果表明根部水淹抑制了水蓼植株的叶生长和分株的形成, 降低了其花生物量和总生物量(表1; 图1-3).这与已有的研究结果一致——淹水显著降低了虉草(Phalaris arundinacea)、牛鞭草(Hemarthria altissima)、狗牙根(Cynodon dactylon)等植物的生物量积累(李秋华等, 2013).这可能与植物在水淹条件下光合能力受到抑制有关(刘泽彬等, 2013; 李晓雪等, 2019; 章毅等, 2019); 同时, 也有研究发现植物在水淹条件下可通过落叶以降低其呼吸消耗, 提高其对水淹的适应性(Striker et al., 2011; Ye et al., 2018). ...
Soil erosion and deposition in the new shorelines of the Three Gorges Reservoir 1 2017
... 三峡工程是世界上最大的水利水电枢纽工程(Wu et al., 2004).三峡水库夏季水位降至145 m, 冬季蓄水至175 m, 形成了与自然河流相反的“冬蓄夏排”周期性水位变化节律(王业春等, 2012; 赵琴和陈教斌, 2018).由于大坝的截流作用, 水库内水流速度非常缓慢, 接近于静止水体(Su et al., 2017).水位周期性变化形成大面积的消落带区域, 其高低高程水位落差最大可达30 m (雷波等, 2014; 郭燕等, 2019).三峡库区高高程区域的植物经历的水淹时间较短且深度较浅, 而低高程区域的植物经历的水淹时间较长且深度较深(刘维暐等, 2012; Chen et al., 2019b).除了三峡库区以外, 国内外河流自然消落区的高程落差很少能达到10 m以上, 通过控制实验就更难实现较大的高程差(Schaff et al., 2003; Park & Kim, 2020).因此, 目前针对高程对河岸带植物生长和繁殖特性影响的研究仍较为缺乏.仅有少数****对高程的效应进行了初步的探讨, 但其研究样地海拔落差仍不超过5 m (Chen et al., 2019a, 2019b). ...
Effects of the Three Gorges Dam on preupland and preriparian drawdown zones vegetation in the upper watershed of the Yangtze River, P. R. China 1 2012
... 高高程水蓼植株的叶和根生物量显著或趋于显著高于低高程植株(图2), 表明高高程植株将更多的生物量投入到营养生长.根据三峡水库的水位调度方案, 与低高程植株相比, 每年消落带高高程区域被水淹的时间较晚而退水的时间较早(Su et al., 2012; Fan et al., 2015; 樊大勇等, 2015; 由永飞等, 2017).因此, 高高程植株水淹间歇的恢复时间比低高程植株更长; 植株将更多的生物量分配到叶和根的生长, 有利于植株光合产物的积累和对土壤养分的吸收(Yan et al., 2015; 徐金英等, 2016; Chen et al., 2019b).已有的研究也发现在相对较浅的水淹环境下河岸带植物如狗牙根、三江藨草(Scirpus nipponicus)和桐花树(Aegiceras corniculatum)等具有较高的叶和根生物量分配(洪明等, 2011; 罗美娟等, 2012; 古勇波等, 2019). ...
How does long-term complete submergence influence sex ratio and resource allocation of a dioecious shrub, Salix variegata Franch.? 1 2016
... 应对周期性的自然水淹, 河岸带植物表现出形态性状上的适应性, 比如通气组织的形成、叶片变薄、茎伸长生长以及气生根的生成等(Phukan et al., 2016; Wang et al., 2016; Heydel et al., 2017; Hua et al., 2017); 同时, 河岸带植物的繁殖特性也随着淹水而发生变化, 水淹耐受能力较强的物种往往能有效地利用退水或两次淹水中间的间隙快速地完成其繁殖过程; 这些植物可以通过调整繁殖期、缩短花期以降低或避免淹水对繁殖过程的影响, 例如稗(Echinochloa crus-galli)的种子可在水位下降后迅速萌发以提高其子代的存活率(Colmer & Voesenek, 2009; 陶敏等, 2011; 樊大勇等, 2015; Su et al., 2016).河岸带不同高程的植物所经历的水淹胁迫往往是不同的, 分布在高高程的植物经历的淹水频率较低, 时长更短; 而分布在低高程的植物经历的淹水频率更高且时长更长(Chen & Xie, 2007; 袁慎鸿等, 2014).已有研究表明不同高程的植物群落特征如物种数和优势种存在较大差异, 物种数随高程的升高表现出增加的趋势(张爱英等, 2018).由于长期遭受不同的淹水强度, 河岸带不同高程植物所受到环境选择压力也是不同的; 因此, 同一物种分布在不同高程的植株的生长和繁殖特性也可能存在差异. ...
Spatial variations of soil phosphorus forms and the risks of phosphorus release in the water-level fluctuation zone in a tributary of the Three Gorges Reservoir 1 2020
... 2018年6月, 每个种群选择500粒成熟饱满的种子, 放在河沙为基质的培养皿中进行萌发.每个种群随机选取24株大小高度一致的实生苗移栽于塑料花盆(盆口直径13.1 cm, 盆底直径9.2 cm, 盆高11.3 cm)中, 共144株植株, 具体实验设计为: 6种群× 2个淹水处理× 2个土壤养分处理× 6重复.淹水处理包括不水淹(正常浇水)和根部水淹处理(从花盆底部向上淹水5 cm).土壤养分处理包括高养分和低养分处理, 两者的草炭:沙:壤土的体积比分别为: 1:1:3和1:3:1; 养分含量分别为: N, (1.51 ± 0.03) mg·g-1, P, (1.46 ± 0.03) mg·g-1; N, (0.48 ± 0.03) mg·g-1, P, (0.55 ± 0.04) mg·g-1.土壤养分梯度设置依据我们前期对三峡库区消落带高低高程土壤养分含量的测定结果, 以及近期发表的相关文献中该区域的土壤养分含量的变化范围: N, (0.40-1.92) mg·g-1; P, (0.43-2.47) mg·g-1 (Ye et al., 2019; Wang et al., 2020; Wei et al., 2020)进行设定.实验采用裂区实验设计, 高养分和低养分处理的72株植株分别放置于12个塑料箱中(长41 cm, 宽31 cm, 高14.5 cm), 每个塑料箱中放置6株来自不同样地不同高程的植株.从高养分和低养分处理中随机选择6个塑料箱, 对其中的植株进行根部水淹处理, 另外6个塑料箱中的植株无水淹处理.当大部分植株进入繁殖期后, 对植株进行收获.实验时间共154天.实验在北京林业大学园林实验教学示范中心开展. ...
Effects of water level fluctuation on the growth of submerged macrophyte communities 1 2016
... 应对周期性的自然水淹, 河岸带植物表现出形态性状上的适应性, 比如通气组织的形成、叶片变薄、茎伸长生长以及气生根的生成等(Phukan et al., 2016; Wang et al., 2016; Heydel et al., 2017; Hua et al., 2017); 同时, 河岸带植物的繁殖特性也随着淹水而发生变化, 水淹耐受能力较强的物种往往能有效地利用退水或两次淹水中间的间隙快速地完成其繁殖过程; 这些植物可以通过调整繁殖期、缩短花期以降低或避免淹水对繁殖过程的影响, 例如稗(Echinochloa crus-galli)的种子可在水位下降后迅速萌发以提高其子代的存活率(Colmer & Voesenek, 2009; 陶敏等, 2011; 樊大勇等, 2015; Su et al., 2016).河岸带不同高程的植物所经历的水淹胁迫往往是不同的, 分布在高高程的植物经历的淹水频率较低, 时长更短; 而分布在低高程的植物经历的淹水频率更高且时长更长(Chen & Xie, 2007; 袁慎鸿等, 2014).已有研究表明不同高程的植物群落特征如物种数和优势种存在较大差异, 物种数随高程的升高表现出增加的趋势(张爱英等, 2018).由于长期遭受不同的淹水强度, 河岸带不同高程植物所受到环境选择压力也是不同的; 因此, 同一物种分布在不同高程的植株的生长和繁殖特性也可能存在差异. ...
三峡库区消落带不同水位高程植被和土壤特征差异 1 2012
... 三峡工程是世界上最大的水利水电枢纽工程(Wu et al., 2004).三峡水库夏季水位降至145 m, 冬季蓄水至175 m, 形成了与自然河流相反的“冬蓄夏排”周期性水位变化节律(王业春等, 2012; 赵琴和陈教斌, 2018).由于大坝的截流作用, 水库内水流速度非常缓慢, 接近于静止水体(Su et al., 2017).水位周期性变化形成大面积的消落带区域, 其高低高程水位落差最大可达30 m (雷波等, 2014; 郭燕等, 2019).三峡库区高高程区域的植物经历的水淹时间较短且深度较浅, 而低高程区域的植物经历的水淹时间较长且深度较深(刘维暐等, 2012; Chen et al., 2019b).除了三峡库区以外, 国内外河流自然消落区的高程落差很少能达到10 m以上, 通过控制实验就更难实现较大的高程差(Schaff et al., 2003; Park & Kim, 2020).因此, 目前针对高程对河岸带植物生长和繁殖特性影响的研究仍较为缺乏.仅有少数****对高程的效应进行了初步的探讨, 但其研究样地海拔落差仍不超过5 m (Chen et al., 2019a, 2019b). ...
三峡库区消落带不同水位高程植被和土壤特征差异 1 2012
... 三峡工程是世界上最大的水利水电枢纽工程(Wu et al., 2004).三峡水库夏季水位降至145 m, 冬季蓄水至175 m, 形成了与自然河流相反的“冬蓄夏排”周期性水位变化节律(王业春等, 2012; 赵琴和陈教斌, 2018).由于大坝的截流作用, 水库内水流速度非常缓慢, 接近于静止水体(Su et al., 2017).水位周期性变化形成大面积的消落带区域, 其高低高程水位落差最大可达30 m (雷波等, 2014; 郭燕等, 2019).三峡库区高高程区域的植物经历的水淹时间较短且深度较浅, 而低高程区域的植物经历的水淹时间较长且深度较深(刘维暐等, 2012; Chen et al., 2019b).除了三峡库区以外, 国内外河流自然消落区的高程落差很少能达到10 m以上, 通过控制实验就更难实现较大的高程差(Schaff et al., 2003; Park & Kim, 2020).因此, 目前针对高程对河岸带植物生长和繁殖特性影响的研究仍较为缺乏.仅有少数****对高程的效应进行了初步的探讨, 但其研究样地海拔落差仍不超过5 m (Chen et al., 2019a, 2019b). ...
Growth and reproductive responses of Polygonum hydropiper populations to elevational difference associated with flooding 1 2020
... 2018年6月, 每个种群选择500粒成熟饱满的种子, 放在河沙为基质的培养皿中进行萌发.每个种群随机选取24株大小高度一致的实生苗移栽于塑料花盆(盆口直径13.1 cm, 盆底直径9.2 cm, 盆高11.3 cm)中, 共144株植株, 具体实验设计为: 6种群× 2个淹水处理× 2个土壤养分处理× 6重复.淹水处理包括不水淹(正常浇水)和根部水淹处理(从花盆底部向上淹水5 cm).土壤养分处理包括高养分和低养分处理, 两者的草炭:沙:壤土的体积比分别为: 1:1:3和1:3:1; 养分含量分别为: N, (1.51 ± 0.03) mg·g-1, P, (1.46 ± 0.03) mg·g-1; N, (0.48 ± 0.03) mg·g-1, P, (0.55 ± 0.04) mg·g-1.土壤养分梯度设置依据我们前期对三峡库区消落带高低高程土壤养分含量的测定结果, 以及近期发表的相关文献中该区域的土壤养分含量的变化范围: N, (0.40-1.92) mg·g-1; P, (0.43-2.47) mg·g-1 (Ye et al., 2019; Wang et al., 2020; Wei et al., 2020)进行设定.实验采用裂区实验设计, 高养分和低养分处理的72株植株分别放置于12个塑料箱中(长41 cm, 宽31 cm, 高14.5 cm), 每个塑料箱中放置6株来自不同样地不同高程的植株.从高养分和低养分处理中随机选择6个塑料箱, 对其中的植株进行根部水淹处理, 另外6个塑料箱中的植株无水淹处理.当大部分植株进入繁殖期后, 对植株进行收获.实验时间共154天.实验在北京林业大学园林实验教学示范中心开展. ...
The Three Gorges Dam: an ecological perspective 1 2004
... 三峡工程是世界上最大的水利水电枢纽工程(Wu et al., 2004).三峡水库夏季水位降至145 m, 冬季蓄水至175 m, 形成了与自然河流相反的“冬蓄夏排”周期性水位变化节律(王业春等, 2012; 赵琴和陈教斌, 2018).由于大坝的截流作用, 水库内水流速度非常缓慢, 接近于静止水体(Su et al., 2017).水位周期性变化形成大面积的消落带区域, 其高低高程水位落差最大可达30 m (雷波等, 2014; 郭燕等, 2019).三峡库区高高程区域的植物经历的水淹时间较短且深度较浅, 而低高程区域的植物经历的水淹时间较长且深度较深(刘维暐等, 2012; Chen et al., 2019b).除了三峡库区以外, 国内外河流自然消落区的高程落差很少能达到10 m以上, 通过控制实验就更难实现较大的高程差(Schaff et al., 2003; Park & Kim, 2020).因此, 目前针对高程对河岸带植物生长和繁殖特性影响的研究仍较为缺乏.仅有少数****对高程的效应进行了初步的探讨, 但其研究样地海拔落差仍不超过5 m (Chen et al., 2019a, 2019b). ...
... 高高程水蓼植株的叶和根生物量显著或趋于显著高于低高程植株(图2), 表明高高程植株将更多的生物量投入到营养生长.根据三峡水库的水位调度方案, 与低高程植株相比, 每年消落带高高程区域被水淹的时间较晚而退水的时间较早(Su et al., 2012; Fan et al., 2015; 樊大勇等, 2015; 由永飞等, 2017).因此, 高高程植株水淹间歇的恢复时间比低高程植株更长; 植株将更多的生物量分配到叶和根的生长, 有利于植株光合产物的积累和对土壤养分的吸收(Yan et al., 2015; 徐金英等, 2016; Chen et al., 2019b).已有的研究也发现在相对较浅的水淹环境下河岸带植物如狗牙根、三江藨草(Scirpus nipponicus)和桐花树(Aegiceras corniculatum)等具有较高的叶和根生物量分配(洪明等, 2011; 罗美娟等, 2012; 古勇波等, 2019). ...
水深对湿地植物生长和繁殖影响研究进展 1 2016
... 高高程水蓼植株的叶和根生物量显著或趋于显著高于低高程植株(图2), 表明高高程植株将更多的生物量投入到营养生长.根据三峡水库的水位调度方案, 与低高程植株相比, 每年消落带高高程区域被水淹的时间较晚而退水的时间较早(Su et al., 2012; Fan et al., 2015; 樊大勇等, 2015; 由永飞等, 2017).因此, 高高程植株水淹间歇的恢复时间比低高程植株更长; 植株将更多的生物量分配到叶和根的生长, 有利于植株光合产物的积累和对土壤养分的吸收(Yan et al., 2015; 徐金英等, 2016; Chen et al., 2019b).已有的研究也发现在相对较浅的水淹环境下河岸带植物如狗牙根、三江藨草(Scirpus nipponicus)和桐花树(Aegiceras corniculatum)等具有较高的叶和根生物量分配(洪明等, 2011; 罗美娟等, 2012; 古勇波等, 2019). ...
Growth and physiological responses to water depths in Carex schmidtii meinsh 1 2015
... 高高程水蓼植株的叶和根生物量显著或趋于显著高于低高程植株(图2), 表明高高程植株将更多的生物量投入到营养生长.根据三峡水库的水位调度方案, 与低高程植株相比, 每年消落带高高程区域被水淹的时间较晚而退水的时间较早(Su et al., 2012; Fan et al., 2015; 樊大勇等, 2015; 由永飞等, 2017).因此, 高高程植株水淹间歇的恢复时间比低高程植株更长; 植株将更多的生物量分配到叶和根的生长, 有利于植株光合产物的积累和对土壤养分的吸收(Yan et al., 2015; 徐金英等, 2016; Chen et al., 2019b).已有的研究也发现在相对较浅的水淹环境下河岸带植物如狗牙根、三江藨草(Scirpus nipponicus)和桐花树(Aegiceras corniculatum)等具有较高的叶和根生物量分配(洪明等, 2011; 罗美娟等, 2012; 古勇波等, 2019). ...
Spatial and temporal dynamics of nutrients in riparian soils after nine years of operation of the Three Gorges Reservoir, China 1 2019
... 2018年6月, 每个种群选择500粒成熟饱满的种子, 放在河沙为基质的培养皿中进行萌发.每个种群随机选取24株大小高度一致的实生苗移栽于塑料花盆(盆口直径13.1 cm, 盆底直径9.2 cm, 盆高11.3 cm)中, 共144株植株, 具体实验设计为: 6种群× 2个淹水处理× 2个土壤养分处理× 6重复.淹水处理包括不水淹(正常浇水)和根部水淹处理(从花盆底部向上淹水5 cm).土壤养分处理包括高养分和低养分处理, 两者的草炭:沙:壤土的体积比分别为: 1:1:3和1:3:1; 养分含量分别为: N, (1.51 ± 0.03) mg·g-1, P, (1.46 ± 0.03) mg·g-1; N, (0.48 ± 0.03) mg·g-1, P, (0.55 ± 0.04) mg·g-1.土壤养分梯度设置依据我们前期对三峡库区消落带高低高程土壤养分含量的测定结果, 以及近期发表的相关文献中该区域的土壤养分含量的变化范围: N, (0.40-1.92) mg·g-1; P, (0.43-2.47) mg·g-1 (Ye et al., 2019; Wang et al., 2020; Wei et al., 2020)进行设定.实验采用裂区实验设计, 高养分和低养分处理的72株植株分别放置于12个塑料箱中(长41 cm, 宽31 cm, 高14.5 cm), 每个塑料箱中放置6株来自不同样地不同高程的植株.从高养分和低养分处理中随机选择6个塑料箱, 对其中的植株进行根部水淹处理, 另外6个塑料箱中的植株无水淹处理.当大部分植株进入繁殖期后, 对植株进行收获.实验时间共154天.实验在北京林业大学园林实验教学示范中心开展. ...
Responses in shoot elongation, carbohydrate utilization and growth recovery of an invasive species to submergence at different water temperatures 1 2018
... 在三峡库区消落带区域, 淹水和土壤养分往往共同影响河岸带植物的生长和发育(张志永等, 2016; Zhou et al., 2020).本研究结果表明根部水淹抑制了水蓼植株的叶生长和分株的形成, 降低了其花生物量和总生物量(表1; 图1-3).这与已有的研究结果一致——淹水显著降低了虉草(Phalaris arundinacea)、牛鞭草(Hemarthria altissima)、狗牙根(Cynodon dactylon)等植物的生物量积累(李秋华等, 2013).这可能与植物在水淹条件下光合能力受到抑制有关(刘泽彬等, 2013; 李晓雪等, 2019; 章毅等, 2019); 同时, 也有研究发现植物在水淹条件下可通过落叶以降低其呼吸消耗, 提高其对水淹的适应性(Striker et al., 2011; Ye et al., 2018). ...
水位调节对三峡水库消落带植被群落特征的影响 1 2017
... 高高程水蓼植株的叶和根生物量显著或趋于显著高于低高程植株(图2), 表明高高程植株将更多的生物量投入到营养生长.根据三峡水库的水位调度方案, 与低高程植株相比, 每年消落带高高程区域被水淹的时间较晚而退水的时间较早(Su et al., 2012; Fan et al., 2015; 樊大勇等, 2015; 由永飞等, 2017).因此, 高高程植株水淹间歇的恢复时间比低高程植株更长; 植株将更多的生物量分配到叶和根的生长, 有利于植株光合产物的积累和对土壤养分的吸收(Yan et al., 2015; 徐金英等, 2016; Chen et al., 2019b).已有的研究也发现在相对较浅的水淹环境下河岸带植物如狗牙根、三江藨草(Scirpus nipponicus)和桐花树(Aegiceras corniculatum)等具有较高的叶和根生物量分配(洪明等, 2011; 罗美娟等, 2012; 古勇波等, 2019). ...
水位调节对三峡水库消落带植被群落特征的影响 1 2017
... 高高程水蓼植株的叶和根生物量显著或趋于显著高于低高程植株(图2), 表明高高程植株将更多的生物量投入到营养生长.根据三峡水库的水位调度方案, 与低高程植株相比, 每年消落带高高程区域被水淹的时间较晚而退水的时间较早(Su et al., 2012; Fan et al., 2015; 樊大勇等, 2015; 由永飞等, 2017).因此, 高高程植株水淹间歇的恢复时间比低高程植株更长; 植株将更多的生物量分配到叶和根的生长, 有利于植株光合产物的积累和对土壤养分的吸收(Yan et al., 2015; 徐金英等, 2016; Chen et al., 2019b).已有的研究也发现在相对较浅的水淹环境下河岸带植物如狗牙根、三江藨草(Scirpus nipponicus)和桐花树(Aegiceras corniculatum)等具有较高的叶和根生物量分配(洪明等, 2011; 罗美娟等, 2012; 古勇波等, 2019). ...
水位节律差异对三峡水库消落区不同物候类型1年生植物物种构成的影响 1 2014
... 应对周期性的自然水淹, 河岸带植物表现出形态性状上的适应性, 比如通气组织的形成、叶片变薄、茎伸长生长以及气生根的生成等(Phukan et al., 2016; Wang et al., 2016; Heydel et al., 2017; Hua et al., 2017); 同时, 河岸带植物的繁殖特性也随着淹水而发生变化, 水淹耐受能力较强的物种往往能有效地利用退水或两次淹水中间的间隙快速地完成其繁殖过程; 这些植物可以通过调整繁殖期、缩短花期以降低或避免淹水对繁殖过程的影响, 例如稗(Echinochloa crus-galli)的种子可在水位下降后迅速萌发以提高其子代的存活率(Colmer & Voesenek, 2009; 陶敏等, 2011; 樊大勇等, 2015; Su et al., 2016).河岸带不同高程的植物所经历的水淹胁迫往往是不同的, 分布在高高程的植物经历的淹水频率较低, 时长更短; 而分布在低高程的植物经历的淹水频率更高且时长更长(Chen & Xie, 2007; 袁慎鸿等, 2014).已有研究表明不同高程的植物群落特征如物种数和优势种存在较大差异, 物种数随高程的升高表现出增加的趋势(张爱英等, 2018).由于长期遭受不同的淹水强度, 河岸带不同高程植物所受到环境选择压力也是不同的; 因此, 同一物种分布在不同高程的植株的生长和繁殖特性也可能存在差异. ...
水位节律差异对三峡水库消落区不同物候类型1年生植物物种构成的影响 1 2014
... 应对周期性的自然水淹, 河岸带植物表现出形态性状上的适应性, 比如通气组织的形成、叶片变薄、茎伸长生长以及气生根的生成等(Phukan et al., 2016; Wang et al., 2016; Heydel et al., 2017; Hua et al., 2017); 同时, 河岸带植物的繁殖特性也随着淹水而发生变化, 水淹耐受能力较强的物种往往能有效地利用退水或两次淹水中间的间隙快速地完成其繁殖过程; 这些植物可以通过调整繁殖期、缩短花期以降低或避免淹水对繁殖过程的影响, 例如稗(Echinochloa crus-galli)的种子可在水位下降后迅速萌发以提高其子代的存活率(Colmer & Voesenek, 2009; 陶敏等, 2011; 樊大勇等, 2015; Su et al., 2016).河岸带不同高程的植物所经历的水淹胁迫往往是不同的, 分布在高高程的植物经历的淹水频率较低, 时长更短; 而分布在低高程的植物经历的淹水频率更高且时长更长(Chen & Xie, 2007; 袁慎鸿等, 2014).已有研究表明不同高程的植物群落特征如物种数和优势种存在较大差异, 物种数随高程的升高表现出增加的趋势(张爱英等, 2018).由于长期遭受不同的淹水强度, 河岸带不同高程植物所受到环境选择压力也是不同的; 因此, 同一物种分布在不同高程的植株的生长和繁殖特性也可能存在差异. ...
三峡水库蓄水对长江干流河岸植物组成的影响 1 2018
... 应对周期性的自然水淹, 河岸带植物表现出形态性状上的适应性, 比如通气组织的形成、叶片变薄、茎伸长生长以及气生根的生成等(Phukan et al., 2016; Wang et al., 2016; Heydel et al., 2017; Hua et al., 2017); 同时, 河岸带植物的繁殖特性也随着淹水而发生变化, 水淹耐受能力较强的物种往往能有效地利用退水或两次淹水中间的间隙快速地完成其繁殖过程; 这些植物可以通过调整繁殖期、缩短花期以降低或避免淹水对繁殖过程的影响, 例如稗(Echinochloa crus-galli)的种子可在水位下降后迅速萌发以提高其子代的存活率(Colmer & Voesenek, 2009; 陶敏等, 2011; 樊大勇等, 2015; Su et al., 2016).河岸带不同高程的植物所经历的水淹胁迫往往是不同的, 分布在高高程的植物经历的淹水频率较低, 时长更短; 而分布在低高程的植物经历的淹水频率更高且时长更长(Chen & Xie, 2007; 袁慎鸿等, 2014).已有研究表明不同高程的植物群落特征如物种数和优势种存在较大差异, 物种数随高程的升高表现出增加的趋势(张爱英等, 2018).由于长期遭受不同的淹水强度, 河岸带不同高程植物所受到环境选择压力也是不同的; 因此, 同一物种分布在不同高程的植株的生长和繁殖特性也可能存在差异. ...
三峡水库蓄水对长江干流河岸植物组成的影响 1 2018
... 应对周期性的自然水淹, 河岸带植物表现出形态性状上的适应性, 比如通气组织的形成、叶片变薄、茎伸长生长以及气生根的生成等(Phukan et al., 2016; Wang et al., 2016; Heydel et al., 2017; Hua et al., 2017); 同时, 河岸带植物的繁殖特性也随着淹水而发生变化, 水淹耐受能力较强的物种往往能有效地利用退水或两次淹水中间的间隙快速地完成其繁殖过程; 这些植物可以通过调整繁殖期、缩短花期以降低或避免淹水对繁殖过程的影响, 例如稗(Echinochloa crus-galli)的种子可在水位下降后迅速萌发以提高其子代的存活率(Colmer & Voesenek, 2009; 陶敏等, 2011; 樊大勇等, 2015; Su et al., 2016).河岸带不同高程的植物所经历的水淹胁迫往往是不同的, 分布在高高程的植物经历的淹水频率较低, 时长更短; 而分布在低高程的植物经历的淹水频率更高且时长更长(Chen & Xie, 2007; 袁慎鸿等, 2014).已有研究表明不同高程的植物群落特征如物种数和优势种存在较大差异, 物种数随高程的升高表现出增加的趋势(张爱英等, 2018).由于长期遭受不同的淹水强度, 河岸带不同高程植物所受到环境选择压力也是不同的; 因此, 同一物种分布在不同高程的植株的生长和繁殖特性也可能存在差异. ...
Hydrologically contrasting environments induce genetic but not phenotypic differentiation in Solanum dulcamara 1 2016
... 河(湖)岸带的土壤水分和养分含量往往沿垂直分布高度的不同而存在差异, 湿地或两栖植物如Ranunculus reptans和Solanum dulcamara分布在不同区域的植株通过种内分化, 表现出不同的形态适应特征以较好地适应所在生境(Lenssen et al., 2004; Zhang et al., 2016); 同时, 也有研究发现某些物种的种内分化即使在较小空间尺度下也能发生(Lenssen et al., 2004).由于分布在消落带不同高程的水蓼植株长期经历不同的淹水强度和土壤养分环境, 因此, 我们假设来自不同高程的水蓼植株可能产生了性状分化, 从而形成不同的生长和繁殖策略以适应不同的淹水和土壤养分条件.为了验证以上假设, 本研究以三峡库区消落带高低高程区域的水蓼为研究对象, 在温室同质园条件下, 研究了不同的淹水和土壤养分处理对来源于高低高程水蓼植株的形态、生物量和繁殖性状的影响, 以期能明确不同高程水蓼植株对根部水淹和土壤养分提升的生长和繁殖响应, 为消落带植被的保护与恢复提供一定的理论依据. ...
... 在三峡库区消落带区域, 淹水和土壤养分往往共同影响河岸带植物的生长和发育(张志永等, 2016; Zhou et al., 2020).本研究结果表明根部水淹抑制了水蓼植株的叶生长和分株的形成, 降低了其花生物量和总生物量(表1; 图1-3).这与已有的研究结果一致——淹水显著降低了虉草(Phalaris arundinacea)、牛鞭草(Hemarthria altissima)、狗牙根(Cynodon dactylon)等植物的生物量积累(李秋华等, 2013).这可能与植物在水淹条件下光合能力受到抑制有关(刘泽彬等, 2013; 李晓雪等, 2019; 章毅等, 2019); 同时, 也有研究发现植物在水淹条件下可通过落叶以降低其呼吸消耗, 提高其对水淹的适应性(Striker et al., 2011; Ye et al., 2018). ...
三峡库区万州段消落带植被及土壤理化特征分析 1 2016
... 在三峡库区消落带区域, 淹水和土壤养分往往共同影响河岸带植物的生长和发育(张志永等, 2016; Zhou et al., 2020).本研究结果表明根部水淹抑制了水蓼植株的叶生长和分株的形成, 降低了其花生物量和总生物量(表1; 图1-3).这与已有的研究结果一致——淹水显著降低了虉草(Phalaris arundinacea)、牛鞭草(Hemarthria altissima)、狗牙根(Cynodon dactylon)等植物的生物量积累(李秋华等, 2013).这可能与植物在水淹条件下光合能力受到抑制有关(刘泽彬等, 2013; 李晓雪等, 2019; 章毅等, 2019); 同时, 也有研究发现植物在水淹条件下可通过落叶以降低其呼吸消耗, 提高其对水淹的适应性(Striker et al., 2011; Ye et al., 2018). ...
三峡库区万州段消落带植被及土壤理化特征分析 1 2016
... 在三峡库区消落带区域, 淹水和土壤养分往往共同影响河岸带植物的生长和发育(张志永等, 2016; Zhou et al., 2020).本研究结果表明根部水淹抑制了水蓼植株的叶生长和分株的形成, 降低了其花生物量和总生物量(表1; 图1-3).这与已有的研究结果一致——淹水显著降低了虉草(Phalaris arundinacea)、牛鞭草(Hemarthria altissima)、狗牙根(Cynodon dactylon)等植物的生物量积累(李秋华等, 2013).这可能与植物在水淹条件下光合能力受到抑制有关(刘泽彬等, 2013; 李晓雪等, 2019; 章毅等, 2019); 同时, 也有研究发现植物在水淹条件下可通过落叶以降低其呼吸消耗, 提高其对水淹的适应性(Striker et al., 2011; Ye et al., 2018). ...
三峡库区消落带生态修复策略研究与实践 1 2018
... 三峡工程是世界上最大的水利水电枢纽工程(Wu et al., 2004).三峡水库夏季水位降至145 m, 冬季蓄水至175 m, 形成了与自然河流相反的“冬蓄夏排”周期性水位变化节律(王业春等, 2012; 赵琴和陈教斌, 2018).由于大坝的截流作用, 水库内水流速度非常缓慢, 接近于静止水体(Su et al., 2017).水位周期性变化形成大面积的消落带区域, 其高低高程水位落差最大可达30 m (雷波等, 2014; 郭燕等, 2019).三峡库区高高程区域的植物经历的水淹时间较短且深度较浅, 而低高程区域的植物经历的水淹时间较长且深度较深(刘维暐等, 2012; Chen et al., 2019b).除了三峡库区以外, 国内外河流自然消落区的高程落差很少能达到10 m以上, 通过控制实验就更难实现较大的高程差(Schaff et al., 2003; Park & Kim, 2020).因此, 目前针对高程对河岸带植物生长和繁殖特性影响的研究仍较为缺乏.仅有少数****对高程的效应进行了初步的探讨, 但其研究样地海拔落差仍不超过5 m (Chen et al., 2019a, 2019b). ...
三峡库区消落带生态修复策略研究与实践 1 2018
... 三峡工程是世界上最大的水利水电枢纽工程(Wu et al., 2004).三峡水库夏季水位降至145 m, 冬季蓄水至175 m, 形成了与自然河流相反的“冬蓄夏排”周期性水位变化节律(王业春等, 2012; 赵琴和陈教斌, 2018).由于大坝的截流作用, 水库内水流速度非常缓慢, 接近于静止水体(Su et al., 2017).水位周期性变化形成大面积的消落带区域, 其高低高程水位落差最大可达30 m (雷波等, 2014; 郭燕等, 2019).三峡库区高高程区域的植物经历的水淹时间较短且深度较浅, 而低高程区域的植物经历的水淹时间较长且深度较深(刘维暐等, 2012; Chen et al., 2019b).除了三峡库区以外, 国内外河流自然消落区的高程落差很少能达到10 m以上, 通过控制实验就更难实现较大的高程差(Schaff et al., 2003; Park & Kim, 2020).因此, 目前针对高程对河岸带植物生长和繁殖特性影响的研究仍较为缺乏.仅有少数****对高程的效应进行了初步的探讨, 但其研究样地海拔落差仍不超过5 m (Chen et al., 2019a, 2019b). ...
Plant waterlogging/flooding stress responses: from seed germination to maturation 1 2020
... 在三峡库区消落带区域, 淹水和土壤养分往往共同影响河岸带植物的生长和发育(张志永等, 2016; Zhou et al., 2020).本研究结果表明根部水淹抑制了水蓼植株的叶生长和分株的形成, 降低了其花生物量和总生物量(表1; 图1-3).这与已有的研究结果一致——淹水显著降低了虉草(Phalaris arundinacea)、牛鞭草(Hemarthria altissima)、狗牙根(Cynodon dactylon)等植物的生物量积累(李秋华等, 2013).这可能与植物在水淹条件下光合能力受到抑制有关(刘泽彬等, 2013; 李晓雪等, 2019; 章毅等, 2019); 同时, 也有研究发现植物在水淹条件下可通过落叶以降低其呼吸消耗, 提高其对水淹的适应性(Striker et al., 2011; Ye et al., 2018). ...
 ,1,*, 于飞海3
,1,*, 于飞海3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT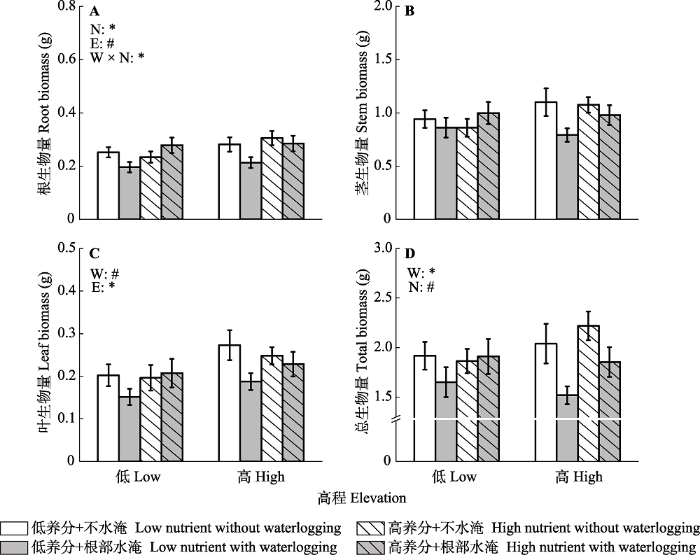 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT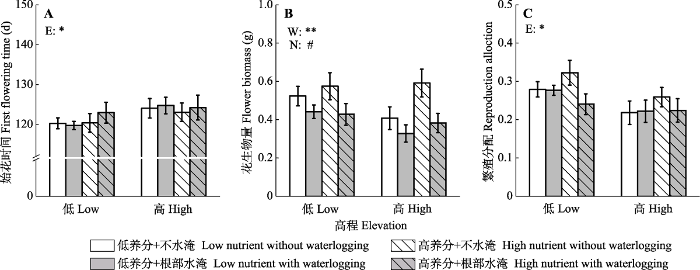 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}