Separation of phylogeny and ecological behaviors between evergreen and deciduous woody angiosperms in the subtropical forest dynamics plots of China
Jian CHE1,3, Jie ZHENG1,3, Ya JIANG3, Yi JIN,,1,3,*, Yin YI,1,2,*1Key Laboratory of National Forestry and Grassland Administration on Biodiversity Conservation in Karst Mountainous Areas of Southwestern China, Guizhou Normal University, Guiyang 550025, China 2Key Laboratory of Plant Physiology and Developmental Regulation of Guizhou Province, Guizhou Normal University, Guiyang 550025, China 3School of Life Sciences, Guizhou Normal University, Guiyang 550025, China
Corresponding authors: *ORCID: 0000-0003-4961-5172 Jin Y:codon@126.com;Yi Y:gzklppdr@gznu.edu.cn Received:2020-05-29Accepted:2020-08-10Online:2020-10-20
Fund supported:
Joint Fund of the National Natural Science Foundation of China and the Karst Science Research Center of Guizhou Province(U1812401) Natural Science and Technology Foundation of Guizhou Province(黔科合基础[2020]1Z013)
Abstract Aims Evergreen (EBL) and deciduous broad-leaved (DBL) woody angiosperms are two major plant groups in the subtropical broad-leaved forests of eastern Asia. Exploring the separation between these two groups in ecological niche, will shed light on the biodiversity maintenance mechanisms of subtropical broad-leaved forests. Methods Adopting statistical methods including the linear regression model and the multiple regression method of Mantel test, we compared the phylogeny and ecological behaviors of the two plant groups in eight forest dynamics plots in China. Important findings We found that (1) leaf habit, be either EBL or DBL, was phylogenetically conserved in the 788 study angiosperm taxa. EBLs and DBLs differed in ecological behaviors towards light, temperature, water, soil reaction and soil fertility. EBLs prefer low light and soil pH, high temperature, water and soil fertility; while the opposite was true for DBLs. (2) Within plot, DBLs were more clustered in phylogenetic dispersion, but more overdispersed in ecological behavior, compared with EBLs; similarly, between plots, DBLs were less diverse in phylogenetic composition, but more diverse in ecological behaviors, than EBLs. On the other hand, divergence in phylogenetic composition of DBLs between plots increased with difference in mean annual temperature (MAT). Further, we found that (3) the ratio of DBLs to EBLs in species richness decreased with MAT increased, but not with dry season length or annual precipitation. The findings show that EBLs and DBLs of the eight study plots differ in both phylogeny and ecological behaviors, and imply that niche separation may be a major mechanism that maintains the biodiversity of subtropical broad-leaved forests. Keywords:leaf habit;ecological niche;biodiversity;subtropical forest;Chinese Forest Biodiversity Monitoring Network
PDF (2313KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 车俭, 郑洁, 蒋娅, 金毅, 乙引. 中国亚热带森林动态监测样地常绿和落叶木本被子植物谱系结构及生态习性差异. 植物生态学报, 2020, 44(10): 1007-1014. DOI: 10.17521/cjpe.2020.0174 CHE Jian, ZHENG Jie, JIANG Ya, JIN Yi, YI Yin. Separation of phylogeny and ecological behaviors between evergreen and deciduous woody angiosperms in the subtropical forest dynamics plots of China. Chinese Journal of Plant Ecology, 2020, 44(10): 1007-1014. DOI: 10.17521/cjpe.2020.0174
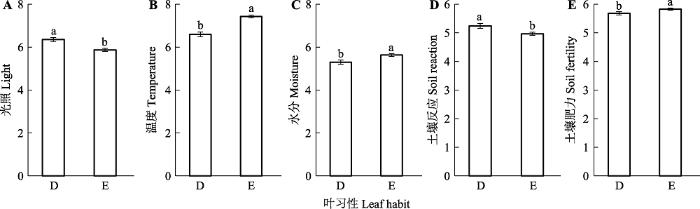
新窗口打开|下载原图ZIP|生成PPT 图1常绿和落叶木本被子植物的各项生态习性对比(平均值±标准误差)。D, 落叶木本被子植物; E, 常绿木本被子植物。柱形上方字母不同, 说明两者间有显著差异(p < 0.05), p值由谱系线性回归模型估计(附录II)。
Fig. 1Difference in each ecological behavior between evergreen and deciduous plants (mean ± SE). D, deciduous woody angiosperms; E, evergreen woody angiosperms. Different letters above the bars indicate significant difference (p < 0.05). p-value is estimated by the phylogenetic linear regression model (Supplement II).
新窗口打开|下载原图ZIP|生成PPT 图2样地常绿和落叶物种的谱系(A)和生态习性聚集情况(B)。D, 落叶木本被子植物; E, 常绿木本被子植物。箱线图上方字母不同说明两者间有显著差异(p < 0.05, Wilcoxon符合秩检验)。
Fig. 2Phylogenetic (A) and ecological behavior dispersions (B) of evergreen and deciduous plants of the eight plots. D, deciduous woody angiosperms; E, evergreen woody angiosperms. NRI, net relatedness index; NFRI, net functional relatedness index. Different lowercase letters above the boxes indicate significant difference (p < 0.05, Wilcoxon signed- rank test with p-value adjusted by false discovery rate).
图3
新窗口打开|下载原图ZIP|生成PPT 图3样地常绿和落叶木本被子植物类群的谱系(A)和生态习性组成差异情况(B)。D, 落叶木本被子植物; E, 常绿木本被子植物。箱线图上方字母不同说明两者间有显著差异(p < 0.05, Wilcoxon秩和检验, p值经过伪发现率法校正)。
Fig. 3Phylogenetic (A) and ecological behavior compositional differences (B) of evergreen and deciduous woody angiosperms of the eight plots. D, deciduous woody angiosperms; E, evergreen woody angiosperms. Different letters above the boxes indicate significant difference (p < 0.05, Wilcoxon signed-rank test with p-value adjusted by false discovery rate). βNRI, inter-sample net relatedness index; βNFRI, inter-sample net functional relatedness index.
2.2 常绿和落叶木本被子植物的谱系、生态习性及物种丰富度比例的驱动环境因子
针对8个样地落叶木本被子植物NRI的最优线性拟合模型不包含任何自变量, 提示各环境因子对落叶植物谱系结构收敛程度的影响不明显; 落叶植物NFRI的最优拟合模型虽然包含了降水量, 但作用也不显著(效应强度0.118, p = 0.536)。针对8个样地常绿木本被子植物NRI的最优拟合模型仅包含样地面积(效应强度-0.769, p = 0.016), 表明随面积增大, 谱系结构更加发散; 常绿植物NFRI的最优线性模型结果不包含任何自变量, 提示各环境因子对落叶植物的谱系结构影响不明显。
Table 1 表1 表1中国亚热带8个森林样地间常绿和落叶木本被子植物谱系组成差异指数(βNRI)和生态习性组成差异指数(βNFRI)的Mantel检验多元回归手段分析结果 Table 1Results of the multiple regression method of Mantel test of net relatedness index among samples (βNRI) and net functional relatedness index among samples (βNFRI) of deciduous and evergreen woody angiosperms among the eight forest plots
影响因子
落叶木本被子植物 D
常绿木本被子植物 E
Predictor variable
βNRI
βNFRI
βNRI
βNFRI
样地面积 A
0.225
0.303
0.063
0.716
岛屿 I
0.211
0.783
-0.617
2.154
年降水量 AP
0.156
0.574
0.425
-0.465
旱季时间 DS
0.090
0.168
-0.035
0.667
年平均气温 MAT
-0.222*
-0.217
-0.018
-0.332
D, deciduous woody angiosperms; E, evergreen woody angiosperms. A, area; AP, annual precipitation; DS, dry season length; I, island; MAT, mean annual temperature. *, p < 0.01.
Fig. 4Variation in the ratio of deciduous: evergreen woody angiosperm richness with mean annual temperature. MAT, mean annual temperature; D:E, the ratio of deciduous and evergreen woody angiosperms in species richness. BDGS, Badagongshan; BSZ, Baishanzu; DHS, Dinghushan; FS, Fushan; GTS, Gutianshan; HSD, Heishiding; LHC, Lienhuachi; TT, Tiantong. Each soild black circle represents a plot, the dark grey line represents the mean of deciduous: evergreen plants richness ratio, and the grey area represents 95% confidence interval of the mean, as fitted by the best supported linear regression model (Supplement III). Adjusted r-square of the linear regression model is R2adj = 0.707.
Supplement II Results of difference between evergreen and deciduous woody angiosperms in each ecological behavior as fitted by the phylogenetic linear regression model in eight subtropical forest dynamics plots of China
Supplement III Results of comparison of linear model of environmental factors effects on deciduous versus evergreen woody angiosperms species richness ratio of each plot in eight subtropical forest dynamics plots of China
Axelrod (1966). Origin of deciduous and evergreen habits in temperate forests Evolution, 20, 1-15. [本文引用: 1]
BartońK (2016). MuMIn: Multi-Model Inference. R package version 1.15.6. [2019-12-21] https://CRAN.R-project.org/ package=MuMIn. URL [本文引用: 1]
BewsJW (1927). Studies in the ecological evolution of angiosperms The New Phytologist, 26(2), 65-84. [本文引用: 1]
CadotteMW, DaviesTJ (2016). Phylogenies in Ecology. Princeton University Press, Princeton, USA. [本文引用: 1]
CadotteMW, DaviesTJ, PeresnetoPR (2017). Why phylogenies do not always predict ecological differences Ecological Monographs, 87, 535-551. [本文引用: 1]
DuanR, HuangM, KongX, WangZ, FanW (2015). Ecophysiological responses to different forest patch type of two codominant tree seedlings Ecology and Evolution, 5, 265-274. [本文引用: 1]
EllenbergH, WeberHE, DuellR, WirthV, WernerW (2001). Zeigerwerte der Gefasspflanzen Mitteleuropas (3 Aufl) Scripta Geobotanica, 18, 1-262. [本文引用: 2]
EscuderoA, del ArcoJM (1987). Ecological significance of the phenology of leaf abscission Oikos, 49, 11-14. [本文引用: 1]
FangXF, ShenGC, YangQS, LiuHM, MaZP, DeaneDC, WangXH (2017). Habitat heterogeneity explains mosaics of evergreen and deciduous trees at local-scales in a subtropical evergreen broad-leaved forest Journal of Vegetation Science, 28, 379-388. [本文引用: 1]
FangXF, YangQS, LiuHM, MaZP, DongS, CaoY, YuanMJ, FeiXY, SunXY, WangXH (2016). Distribution of species abundance of evergreen and deciduous woody plants in the evergreen broad-leaved forests at Tiantong, Zhejiang Biodiversity Science, 24, 629-638. [本文引用: 1]
GeJL, XieZQ (2017). Geographical and climatic gradients of evergreen versus deciduous broad-leaved tree species in subtropical China: implications for the definition of the mixed forest Ecology and Evolution, 7, 3636-3644. [本文引用: 2]
GivnishTJ (1979). On the adaptive significance of leaf form//Solbrig OT, Jain S, Johnson GB, Raven PH Topics in Plant Population Biology. Palgrave, London, 375-407. [本文引用: 1]
GivnishTJ (2002). Adaptive signifi cance of evergreen vs. deciduous leaves: solving the triple paradox Silva Fennica, 36, 703-743. [本文引用: 3]
GosleeSC, UrbanDL (2007). The ecodist package for dissimilarity-based analysis of ecological data Journal of Statistical Software, 22, 1-19. [本文引用: 1]
HuangYT, YaoL, AiXR, LüSA, DingY (2015). Quantitative classification of the subtropical evergreen-deciduous broadleaved mixed forest and the deciduous and evergreen species composition structure across two national nature reserves in the southwest of Hubei, China Chinese Journal of Plant Ecology, 39, 990-1002. [本文引用: 1]
JinY, RussoSE, YuM (2018). Effects of light and topography on regeneration and coexistence of evergreen and deciduous tree species in a Chinese subtropical forest Journal of Ecology, 106, 1634-1645. [本文引用: 2]
KiraT (1991). Forest ecosystems of east and southeast Asia in a global perspective Ecological Research, 6, 185-200. [本文引用: 5]
KraftNJ, GodoyO, LevineJM (2015). Plant functional traits and the multidimensional nature of species coexistence Proceedings of the National Academy of Sciences of the United States of America, 112, 797-802. [本文引用: 1]
MiuraM, ManabeT, NishimuraN, YamamotoS (2001). Forest canopy and community dynamics in a temperate old- growth evergreen broad-leaved forest, south-western Japan: a 7-year study of a 4-ha plot Journal of Ecology, 89, 841-849. [本文引用: 2]
OksanenJF, BlanchetG, FriendlyM, KindtR, LegendreP, McGlinnD, MinchinPR, O?HaraRB, SimpsonGL, SolymosP, StevensMHH, SzoecsE, WagnerH (2017). vegan: Community Ecology Package. R package version 2.4-3. [2020-04-09]. https://CRAN.R-project.org/package=vegan. URL [本文引用: 1]
ParadisE, ClaudeJ, StrimmerK (2004). Analyses of phylogenetics and evolution in R language Bioinformatics, 20, 289-290. [本文引用: 1]
PennellMW, EastmanJM, SlaterGJ, BrownJW, UyedaJC, FitzjohnRG, AlfaroME, HarmonLJ (2014). geiger v2.0: an expanded suite of methods for fitting macroevolutionary models to phylogenetic trees Bioinformatics, 30, 2216-2218. [本文引用: 1]
QianH, JinY (2016). An updated megaphylogeny of plants, a tool for generating plant phylogenies and an analysis of phylogenetic community structure Journal of Plant Ecology, 9, 233-239. [本文引用: 1]
R Core Team (2016). R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. [2020-04-12]. https://www.R-project.org/. URL [本文引用: 1]
ReichPB (2014). The world-wide “fast-slow” plant economics spectrum: a traits manifesto Journal of Ecology, 102, 275-301. [本文引用: 1]
RicklefsRE, HeF (2016). Region effects influence local tree species diversity Proceedings of the National Academy of Sciences of the United States of America, 113, 674-679. [本文引用: 4]
SongK, KohyamaT, DaL (2014). Transition patterns across an evergreen-deciduous broad-leaved forest ecotone: the effect of topographies Journal of Vegetation Science, 25, 1257-1266. [本文引用: 1]
SongYC (1999). Perspective of the vegetation zonafion of forest region in eastern China Acta Botanica Sinica, 41, 541-552. [本文引用: 1]
TangCQ, OhsawaM (2002). Coexistence mechanisms of evergreen, deciduous and coniferous trees in a mid-montane mixed forest on Mt. Emei, Sichuan, China Plant Ecology, 161, 215-230. [本文引用: 1]
WebbCO, AckerlyDD, KembelSW (2008). Phylocom: software for the analysis of phylogenetic community structure and trait evolution Bioinformatics, 24, 2098-2100. [本文引用: 5]
WuZY (1980). Vegetation of China. Science Press, Beijing. [本文引用: 1]
ZhangL, LuoT, ZhuH, DalyC, DengK (2009). Leaf life span as a simple predictor of evergreen forest zonation in China Journal of Biogeography, 37, 27-36. [本文引用: 1]
ZhangTT, WangX, JinY, YuJP, RenHB, QianHY, ChenSW, MaKP, YuMJ (2020). Comparative study on species composition between Gutianshan and other eastern typical evergreen broad-leaved forests in China Guihaia, 40, 1061-1070. [本文引用: 2]
ZhuY, ZhaoGF, ZhangLW, ShenGC, MiXC, RenHB, YuMJ, ChenJH, ChenSW, FangT, MaKP (2008). Community composition and structure of Gutianshan forest dynamic plot in a mid-subtropical evergreen broad-leaved forest, East China Chinese Journal of Plant Ecology (Chinese Version), 32, 262-273. DOI:10.3773/j.issn.1005-264x.2008.02.004URL [本文引用: 1] Aims Mainly distributed in China, subtropical evergreen broad-leaved forest is one of important vegetation types in the world. Here we report preliminary results of floristic characteristics, community composition, vertical structure, size class structure, and spatial structure of Gutianshan(GTS) forest plot. Methods We established a 24-hm2 (600 m×400 m) forest permanent plot from November, 2004 to September, 2005 in mid- subtropical evergreen broad-leaved forest of Gutianshan Nature Reserve, China. Following the standard census procedure of the Centre for Tropical Forest Science (CTFS), all free-standing trees ≥1 cm in diameter at breast height (DBH) in the forest were mapped, tagged and identified to species. We employed software R 2.6.0 to analyze our data. Important findings The results of floristic characteristics indicates that the tropical elements are more than temperate elements. At family level, the proportion of the pantropic type is the greatest (28.6%), the number of the tropic elements are more than temperate ones (24/13). At genus level,there are 53 tropic genera and 44 temperate ones. As for community composition, there are 159 species, 103 genera and 49 families, 140 700 individuals in total. The evergreen tree species in community are dominant (i.e. 91 species, total relative dominance is 90 .6%, importance value is 85.6%, accounts for 85.9% of the total abundance). GTS forest plot is typical mid-subtropical evergreen broad-leaved forest, which displays characteristics of both temperate deciduous broad-leaved forest and tropical rain forest. On the one hand, community composition has obvious dominant species, which is similar to temperate deciduous broad-leaved forest. There are 3 mostly dominant species, Castanopsis eyrei, Schima superba and Pinus massoniana. Large numbers of rare species (59 rare species, equal to or less than one tree per hm2) in the community account for 37.1% species richness, which is similar to tropical rain forest. Vertical structure is composed of canopy layer (63 species ), sub-uree layer (70 species), shrub layer (26 species).The structure of DBH size class of all species in the plot generally appears reverse `J’ shape, which indicates successful community regeneration. Spatial distribution of several dominant species, from small to adult tree or old tree, shifts from closer aggregation to looser aggregation, and shows different habitat preference. Finally, we compare the large plot approach with conventional sampling method.]]> [ 祝燕, 赵谷风, 张俪文, 沈国春, 米湘成, 任海保, 于明坚, 陈建华, 陈声文, 方腾, 马克平 (2008). 古田山中亚热带常绿阔叶林动态监测样地——群落组成与结构 植物生态学报, 32, 262-273.] [本文引用: 1]
Origin of deciduous and evergreen habits in temperate forests 1 1966
Ecological significance of the phenology of leaf abscission 1 1987
... 常绿和落叶木本被子植物各项生态习性均有差异, 提示阔叶林内的两类植物存在着明显生态位分化.其中, 落叶植物偏好高光照但凉爽、土壤干燥贫瘠的环境; 而常绿植物则位于低光照但温暖、土壤湿润肥沃的环境.落叶植物对于高光照的偏好, 也与以往研究(Miura et al., 2001; Duan et al., 2015; Jin et al., 2018)所发现的常绿阔叶林林窗内的高光照环境更有利于落叶植物更新相一致, 也与叶经济谱假说(Reich, 2014)的预测一致.而落叶植物对于凉爽和土壤贫瘠环境的偏好, 则可能与落叶性状相关, 在此类环境(例如, 群落演替早期, 较高纬度或高海拔地区)中, 可通过落叶避免受到季节性低温或干旱的伤害, 从而形成对常绿植物的竞争优势(Givnish, 1979, 2002).而在低光照、温暖、土壤湿润而肥沃的低纬度常绿阔叶林环境中, 则是可以全年进行光合作用的常绿植物处于竞争优势地位(Escudero & del Arco, 1987; Givnish, 2002). ...
Habitat heterogeneity explains mosaics of evergreen and deciduous trees at local-scales in a subtropical evergreen broad-leaved forest 1 2017
... 在群落尺度上, 常绿和落叶被子植物通常在亚热带阔叶林内共同存在(祝燕等, 2008; 方晓峰等, 2016).两类植物之所以能够在阔叶林群落内共存, 被认为与其对于地形和土壤条件(Tang & Ohsawa, 2002; Fang et al., 2017), 以及光照(Miura et al., 2001; Jin et al., 2018)等环境因子需求的差异有关.但是, 由于物种共存涉及到生态位的多个维度(Kraft et al., 2015), 以往研究缺乏对于常绿和落叶被子植物生态位总体差异的综合分析.了解常绿和落叶被子植物的谱系差异(Cadotte et al., 2017), 结合两者对光照、温度、水分和土壤条件等主要环境因子的生态习性(Ellenberg et al., 2001; 宋永昌, 2013)差异, 可进一步加深对两者生态位差异特点的认识. ...
天童常绿阔叶林中常绿与落叶物种的物种多度分布格局 1 2016
... 在群落尺度上, 常绿和落叶被子植物通常在亚热带阔叶林内共同存在(祝燕等, 2008; 方晓峰等, 2016).两类植物之所以能够在阔叶林群落内共存, 被认为与其对于地形和土壤条件(Tang & Ohsawa, 2002; Fang et al., 2017), 以及光照(Miura et al., 2001; Jin et al., 2018)等环境因子需求的差异有关.但是, 由于物种共存涉及到生态位的多个维度(Kraft et al., 2015), 以往研究缺乏对于常绿和落叶被子植物生态位总体差异的综合分析.了解常绿和落叶被子植物的谱系差异(Cadotte et al., 2017), 结合两者对光照、温度、水分和土壤条件等主要环境因子的生态习性(Ellenberg et al., 2001; 宋永昌, 2013)差异, 可进一步加深对两者生态位差异特点的认识. ...
天童常绿阔叶林中常绿与落叶物种的物种多度分布格局 1 2016
... 在群落尺度上, 常绿和落叶被子植物通常在亚热带阔叶林内共同存在(祝燕等, 2008; 方晓峰等, 2016).两类植物之所以能够在阔叶林群落内共存, 被认为与其对于地形和土壤条件(Tang & Ohsawa, 2002; Fang et al., 2017), 以及光照(Miura et al., 2001; Jin et al., 2018)等环境因子需求的差异有关.但是, 由于物种共存涉及到生态位的多个维度(Kraft et al., 2015), 以往研究缺乏对于常绿和落叶被子植物生态位总体差异的综合分析.了解常绿和落叶被子植物的谱系差异(Cadotte et al., 2017), 结合两者对光照、温度、水分和土壤条件等主要环境因子的生态习性(Ellenberg et al., 2001; 宋永昌, 2013)差异, 可进一步加深对两者生态位差异特点的认识. ...
Geographical and climatic gradients of evergreen versus deciduous broad-leaved tree species in subtropical China: implications for the definition of the mixed forest 2 2017
Forest canopy and community dynamics in a temperate old- growth evergreen broad-leaved forest, south-western Japan: a 7-year study of a 4-ha plot 2 2001
... 在群落尺度上, 常绿和落叶被子植物通常在亚热带阔叶林内共同存在(祝燕等, 2008; 方晓峰等, 2016).两类植物之所以能够在阔叶林群落内共存, 被认为与其对于地形和土壤条件(Tang & Ohsawa, 2002; Fang et al., 2017), 以及光照(Miura et al., 2001; Jin et al., 2018)等环境因子需求的差异有关.但是, 由于物种共存涉及到生态位的多个维度(Kraft et al., 2015), 以往研究缺乏对于常绿和落叶被子植物生态位总体差异的综合分析.了解常绿和落叶被子植物的谱系差异(Cadotte et al., 2017), 结合两者对光照、温度、水分和土壤条件等主要环境因子的生态习性(Ellenberg et al., 2001; 宋永昌, 2013)差异, 可进一步加深对两者生态位差异特点的认识. ...
... 常绿和落叶木本被子植物各项生态习性均有差异, 提示阔叶林内的两类植物存在着明显生态位分化.其中, 落叶植物偏好高光照但凉爽、土壤干燥贫瘠的环境; 而常绿植物则位于低光照但温暖、土壤湿润肥沃的环境.落叶植物对于高光照的偏好, 也与以往研究(Miura et al., 2001; Duan et al., 2015; Jin et al., 2018)所发现的常绿阔叶林林窗内的高光照环境更有利于落叶植物更新相一致, 也与叶经济谱假说(Reich, 2014)的预测一致.而落叶植物对于凉爽和土壤贫瘠环境的偏好, 则可能与落叶性状相关, 在此类环境(例如, 群落演替早期, 较高纬度或高海拔地区)中, 可通过落叶避免受到季节性低温或干旱的伤害, 从而形成对常绿植物的竞争优势(Givnish, 1979, 2002).而在低光照、温暖、土壤湿润而肥沃的低纬度常绿阔叶林环境中, 则是可以全年进行光合作用的常绿植物处于竞争优势地位(Escudero & del Arco, 1987; Givnish, 2002). ...
1 2017
... 针对生态习性树的构建, 首先是采用vegan软件包(Oksanen et al., 2017)中“decostand”函数的“standardize”参数标准化各个生态习性指标值, 方法为减去平均值, 然后除以单位方差, 目的是将各变量的变异控制在相同的数量尺度, 以便进行比较; 然后用“dist”函数的“euclidean”距离参数计算种间总体生态习性的欧氏距离矩阵; 然后用“hclust”函数进行聚类分析, 聚类方法采用非加权组平均法(UPGMA); 最后采用ape软件包(Paradis et al., 2004)中“as.phylo”函数转换为树状数据格式(即phylo). ...
Analyses of phylogenetics and evolution in R language 1 2004
... 针对生态习性树的构建, 首先是采用vegan软件包(Oksanen et al., 2017)中“decostand”函数的“standardize”参数标准化各个生态习性指标值, 方法为减去平均值, 然后除以单位方差, 目的是将各变量的变异控制在相同的数量尺度, 以便进行比较; 然后用“dist”函数的“euclidean”距离参数计算种间总体生态习性的欧氏距离矩阵; 然后用“hclust”函数进行聚类分析, 聚类方法采用非加权组平均法(UPGMA); 最后采用ape软件包(Paradis et al., 2004)中“as.phylo”函数转换为树状数据格式(即phylo). ...
geiger v2.0: an expanded suite of methods for fitting macroevolutionary models to phylogenetic trees 1 2014
 ,
, ,1,3,*, 乙引
,1,3,*, 乙引
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT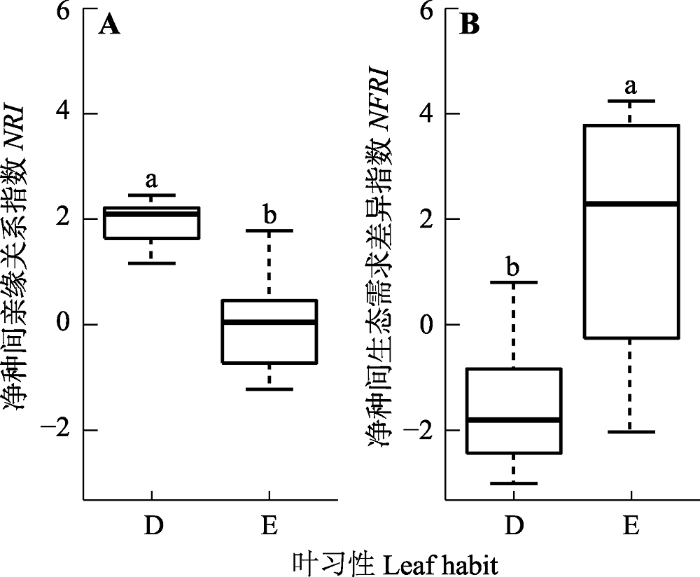 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT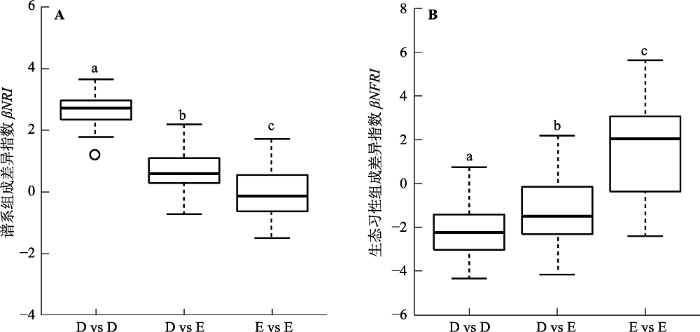 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT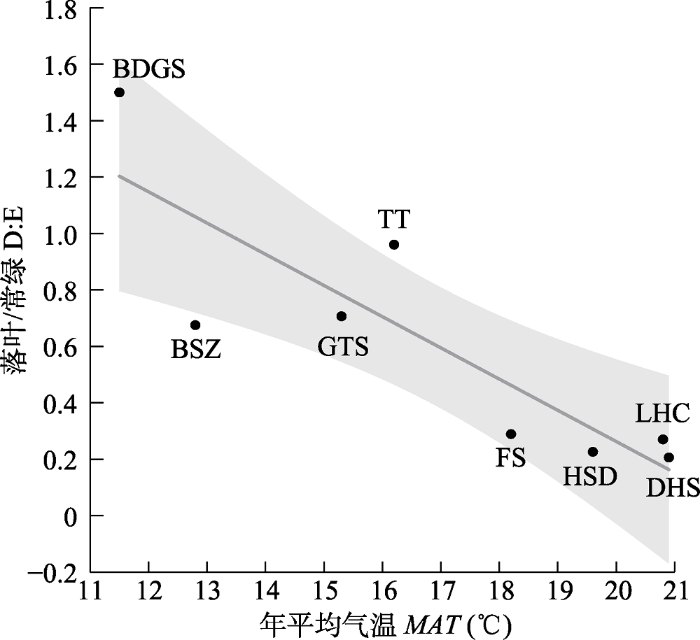 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}