Effects of microhabitat changes on seedling establishment of native plants in a dry valley
Hui HU1,2, Yu YANG1,2, Wei-Kai BAO1, Xin LIU1, Fang-Lan LI,1,*1CAS Key Laboratory of Mountain Ecological Restoration and Bioresource Utilization & Ecological Restoration and Biodiversity Conservation Key Laboratory of Sichuan Province, Chengdu Institute of Biology, Chinese Academy of Sciences, Chengdu 610041, China 2University of Chinese Academy of Sciences, Beijing 100049, China
National Key R&D Program of China(2017YFC0505105) Science and Technology Major Project of Sichuan Province(2018SZDZX0033) Science and Technology Major Project of Sichuan Province(2018SZDZX0035)
Abstract Aims The patchy distribution patterns of vegetation result in heterogeneous microhabitats and thus affect seed germination and seedling establishment in arid ecosystems. The impact of changes in microhabitat conditions on the colonization of native plants and the specificity need to be understood for restoring vegetation by adopting microhabitat regulation measures in arid ecosystems where vegetation degradations are exacerbated by climate change and increasing anthropogenic disturbances. Methods Three types of microhabitat conditions, including vegetation cover of shrubs and semi-shrubs, respectively, and bare lands, were selected in an dry valley in Minjiang River basin. The effects of microhabitat conditions on seedling establishment were studied in Bauhinia brachycarpa by measuring the survival rate and the height and biomass of seedlings. The microhabitat effects on seed germination and seedling establishment were also studied in six other native plant species on an extremely degraded roadside slope. Important findings The results show that B. brachycarpa seedlings had significantly greater survival rate and total biomass, as well as greater biomass of roots and shoots on bare lands than in habitats with shrub and semi-shrub covers; whereas the specific leaf area and leaf mass fraction of seedlings were smaller on bare lands than in shrub and semi-shrub patches. Humus addition treatment significantly increased the total biomass of seedlings in bare land microhabitat. Moreover, non-woven geotextile cover and humus addition treatment increased the germination rate in herbaceous plants, resulting in high herb density four months after treatment. The results suggest that humus addition and geotextile cover are effective in regulating microhabitat for improvement of seedling colonization and maintenance of the community structure stability. In conclusion, uses of native plants are important in the vegetation restoration of extremely degraded sites in arid regions due to their strong adaptability to road slope habitats and capability of forming a shrub-grass mosaic community structures. Keywords:ecological restoration;interspecific variation;community structure;roadside;microhabitat;seedling establishment
PDF (1335KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 胡慧, 杨雨, 包维楷, 刘鑫, 李芳兰. 干旱河谷微生境变化对乡土植物幼苗定植的影响. 植物生态学报, 2020, 44(10): 1028-1039. DOI: 10.17521/cjpe.2020.0216 HU Hui, YANG Yu, BAO Wei-Kai, LIU Xin, LI Fang-Lan. Effects of microhabitat changes on seedling establishment of native plants in a dry valley. Chinese Journal of Plant Ecology, 2020, 44(10): 1028-1039. DOI: 10.17521/cjpe.2020.0216
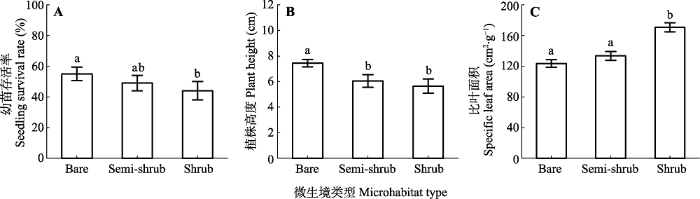
Fig. 1Survival rate, plant height and specific leaf area of two-year old Bauhinia brachycarpa seedlings in different microhabitats in the dry valley studied (means ± SE). Bare, bare land microhabitats; Semi-shrub, semi-shrub microhabitats; Shrub, shrub microhabitats. Different lowercase letters indicate significant differences among the microhabitats (n = 15; p < 0.05).
新窗口打开|下载原图ZIP|生成PPT 图2干旱河谷不同微生境条件下二年生鞍叶羊蹄甲幼苗生物量(平均值±标准误差)。Bare, 裸地微生境; Semi-shrub, 半灌木微生境; Shrub, 灌木微生境。相同施肥条件下不同小写字母指示不同微生境之间差异显著(n = 15; p < 0.05)。*, 同一微生境类型中施肥效果显著(n = 15; p < 0.05); ns, 施肥效果不显著(n = 15; p > 0.05)。
Fig. 2Biomass (means ± SE) of two-year old Bauhinia brachycarpa seedlings in different microhabitats in the dry valley studied. Bare, bare land microhabitats; Semi-shrub, semi-shrub microhabitats; Shrub, shrub microhabitats. Different lowercase letters within a fertile condition indicate significant differences among the microhabitats (n = 15; p < 0.05). *, significant difference between fertilized and unfertilized treatments (n = 15; p < 0.05); ns, non-significant difference between fertilized and unfertilized treatments (n = 15; p > 0.05).
Fig. 3Biomass allocation (means ± SE) of two-year old Bauhinia brachycarpa seedlings in different microhabitats in the dry valley studied. Bare, bare land microhabitats; Semi-shrub, semi-shrub microhabitats; Shrub, shrub microhabitats. Different lowercase letters indicate significant differences among the microhabitats (n = 15; p < 0.05).
Table 2 表2 表2干旱河谷道路边坡不同乡土植物种子发芽率、萌发率和幼苗保存率(平均值±标准误差) Table 2Seed germination rate, seedling emergence and seedling survival rate (means ± SE) of different native plants on roadside slope of the dry valley studied
生活型 Life form
物种 Species
种子发芽率 Seed germination rate (%)
出苗率 Seedling emergence rate (%)
保存率 Seedling survival rate (%)
草本和半灌木 Herb and semi-shrub
早熟禾 Poa annua
61.6 ± 5.5a
56.8 ± 12.40a
-
狗尾草 Setaria viridis
72.3 ± 12.6ab
68.2 ± 8.11a
-
毛莲蒿 Artemisia vestit
80.2 ± 9.1b
23.9 ± 6.52b
-
灌木 Shrub
白刺花 Sophora davidii
61.2 ± 5.4a
43.8 ± 5.49a
77.0 ± 6.2a
鞍叶羊蹄甲 Bauhinia brachycarpa
98.0 ± 0.3b
68.1 ± 9.53a
75.7 ± 7.5a
四川黄栌 Cotinus szechuanensis
34.6 ± 2.6c
10.3 ± 2.34b
79.9 ± 8.2a
小叶杭子梢 Campylotropis wilsonii
76.7 ± 3.8ab
58.5 ± 13.20a
76.4 ± 5.5a
同列不同小写字母表示不同物种之间差异显著(n = 3; p < 0.05)。 Different lowercase letters within a column indicate significant differences among species (n = 3; p < 0.05).
新窗口打开|下载原图ZIP|生成PPT 图4干旱河谷道路边坡不同微生境处理下群落结构参数(平均值±标准误差)。Bare, 裸露播种; Bare + N, 播种+施加养分; Cov + N, 播种+施加养分+纤维毯覆盖; Control, 裸露不播种。不同小写字母表示不同处理之间差异显著(n = 3; p < 0.05)。
Fig. 4Community structures (means ± SE) under varying microhabitat treatments on roadside slope of the dry valley studied. Bare, bare land only with seedling; Bare + N, bare land with seedling and nutrient; Cov + N, covered with seedling and nutrient; Control, bare land without seedling. Different lowercase letters indicate significant differences among treatments (n = 3; p < 0.05).
图5
新窗口打开|下载原图ZIP|生成PPT 图5干旱河谷道路边坡不同微生境处理下群落密度和根系分布深度(平均值±标准误差)。Bare, 裸露播种; Bare + N, 播种+施加养分; Cov + N, 播种+施加养分+纤维毯覆盖; Control, 裸露不播种。不同小写字母表示不同处理之间差异显著(n = 3; p < 0.05)。
Fig. 5Community density and rooting depth (means ± SE) under varying microhabitat treatments on roadside slope of the dry valley studied. Control, bare land without seedling; Bare, bare land only with seedling; Bare + N, bare land with seedling and nutrient; Cov + N, covered with seedling and nutrient. Different lowercase letters indicate significant differences among treatments (n = 3; p < 0.05).
新窗口打开|下载原图ZIP|生成PPT 图6干旱河谷道路边坡不同微生境处理下群落密度的动态变化(平均值±标准误差)。Bare, 裸露播种; Bare + N, 播种+施加养分; Cov + N, 播种+施加养分+纤维毯覆盖。
Fig. 6Dynamics of community density (means ± SE) under varying microhabitat treatments on roadside slope of the dry valley studied. Bare, bare land only with seedling; Bare + N, bare land with seedling and nutrient; Cov + N, covered with seedling and nutrient.
Table 3 表3 表3生长周期与微生境处理两因素对道路边坡上群落生物量及盖度的交互作用 Table 3Responses of community coverage and biomass to growing duration and varying microhabitat treatments and their interaction
变量 Variable
生物量 Biomass (g·m-2)
盖度 Coverage (%)
F
p
F
p
生长年份 Growing duration (Gd)
46.874
<0.001
16.971
0.001
微生境处理 Microhabitat treatments (Mt)
12.191
0.001
8.276
0.006
Gd × Mt
3.926
0.049
0.062
0.941
R2
0.868
0.737
通过两因素方差分析检验F和p的差异。 Differences in F and p were assessed with two-way ANOVA.
新窗口打开|下载原图ZIP|生成PPT 图7干旱河谷道路边坡上不同微生境处理下群落盖度和生物量的年际间变化(平均值±标准误差)。Bare, 裸露播种; Bare + N, 播种+施加养分; Cov + N, 播种+施加养分+纤维毯覆盖; Control, 裸露不播种。**和*表示同一微生境类型年份之间差异显著(n = 15; 分别为p < 0.01 和p < 0.05), ns表示年份之间差异不显著(n = 15; p > 0.05)。
Fig. 7Inter-annual dynamics of community density and biomass (means ± SE) under varying microhabitat treatments on roadside slope of the dry valley studied. Bare, bare land only with seedling; Bare + N, bare land with seedling and nutrient; Cov + N, covered with seedling and nutrient; Control, bare land without seedling. ** and * indicate significant differences between the two observational years at p < 0.01 and p < 0.05 levels, respectively (n = 15); ns indicate non-significant differences between the two observational years (n = 15; p > 0.05).
BaoWK, PangXY, LiFL, ZhouZQ (2012). A Study of Ecological Restoration and Sustainable Management of the Arid Minjiang River Valley, China. Science Press, Beijing. [本文引用: 6]
Castro-DíezP, PuyravaudJP, CornelissenJHC (2000). Leaf structure and anatomy as related to leaf mass per area variation in seedlings of a wide range of woody plant species and types Oecologia, 124, 476-486. [本文引用: 1]
CertiniG, CampbellCD, EdwardsAC (2004). Rock fragments in soil support a different microbial community from the fine earth Soil Biology & Biochemistry, 36, 1119-1128. [本文引用: 1]
ChartzoulakisK, NoitsakisB, TheriosI (1993). Photosynthesis, plant growth and dry matter distribution in kiwifruit as influenced by water deficits Irrigation Science, 14, 1-5. [本文引用: 2]
de KroonH, VisserEJW (2003). Root Ecology. Springer-Verlag, New York [本文引用: 1]
EngelbrechtBMJ, KursarTA, TyreeMT (2005). Drought effects on seedling survival in a tropical moist forest Trees, 19, 312-321. [本文引用: 1]
Estrada-MedinaH, GrahamRC, AllenMF, Jiménez-OsornioJJ, Robles-CasolcoS (2013). The importance of limestone bedrock and dissolution karst features on tree root distribution in northern Yucatán, México Plant and Soil, 362, 37-50. [本文引用: 1]
GindabaJ, RozanovA, NegashL (2004). Response of seedlings of two Eucalyptus and three deciduous tree species from Ethiopia to severe water stress Forest Ecology and Management, 201, 119-129. [本文引用: 1]
HanAR, KimHJ, JungJB, ParkPS (2018). Seed germination and initial seedling survival of the subalpine tree species, Picea jezoensis, on different forest floor substrates under elevated temperature Forest Ecology and Management, 429, 579-588. [本文引用: 1]
LeckMA, ParkerVT, SimpsonRL (2008). Seedling Ecology and Evolution. Cambridge University Press, Cambridge. [本文引用: 3]
LehouckV, SpanhoveT, GonsamoA, CordeiroNJ, LensL (2009). Spatial and temporal effects on recruitment of an Afromontane forest tree in a threatened fragmented ecosystem Biological Conservation, 142, 518-528. [本文引用: 4]
LiFL, BaoWK, PangXY, LengL (2009). Seedling emergence, survival and growth of five endemic species in the dry valley of Minjiang River Acta Ecologica Sinica, 29, 2219-2230. [本文引用: 7]
LiFL, BaoWK, WuN (2009). Effects of water stress on growth, dry matter allocation and water-use efficiency of a leguminous species, Sophora davidii Agroforestry Systems, 77, 193-201.
LiJ, ZhaoCY, ZhuH, WangF (2007). Species effect of Tamarix spp. and Haloxylon ammodendronon shrub “fertile island” Acta Ecologica Sinica, 27, 5138-5348. [本文引用: 3]
LiY, BaoWK, WuN (2011). Spatial patterns of the soil seed bank and extant vegetation across the dry Minjiang River valley in southwest China Journal of Arid Environments, 75, 1083-1089. [本文引用: 1]
LiuCX, HanLB (2007). Review of researches in vegetation restoration of freeway slopes Acta Ecologica Sinica, 27, 2090-2098. [本文引用: 1]
LuoH, ZhaoTY, PengXF, GuoY, LiangC (2013). Effectiveness of soil and water conservation of greening mulch of roadside slope Transactions of the Chinese Society of Agricultural Engineering, 29(5), 63-70. [本文引用: 1]
LynchJP (2019). Root phenotypes for improved nutrient capture: an underexploited opportunity for global agriculture New Phytologist, 223, 548-564. [本文引用: 1]
MaWB, JiHJ, SuYM, LiuXL, HeJS, ZhangL (2013). Characteristics and research application of vegetation blanket slope protection Soil and Water Conservation in China, 1(9), 30-33. [本文引用: 1]
MiMX, ShaoMG, LiuBX (2016). Effect of rock fragments content on water consumption, biomass and water-use efficiency of plants under different water conditions Ecological Engineering, 94, 574-582. [本文引用: 1]
MilbauA, ScheerlinckL, ReheulD, de CauwerB, NijsI (2005). Ecophysiological and morphological parameters related to survival in grass species exposed to an extreme climatic event Physiologia Plantarum, 125, 500-512. [本文引用: 1]
PaquetteA, BouchardA, CogliastroA (2006). Survival and growth of under-planted trees: a meta-analysis across four biomes Ecological Applications, 16, 1575-1589. [本文引用: 1]
PoorterH, NiklasKJ, ReichPB, OleksynJ, PootP, MommerL (2012). Biomass allocation to leaves, stems and roots: meta-analyses of interspecific variation and environmental control New Phytologist, 193, 30-50. [本文引用: 1]
QuLY, WangZB, HuangYY, ZhangYX, SongCJ, MaKM (2017). Effects of plant coverage on shrub fertile islands in the Upper Minjiang River Valley Science China: Life Sciences, 61, 340-347. [本文引用: 8]
SackL, GrubbPJ, Mara?ónT (2003). The functional morphology of juvenile plants tolerant of strong summer drought in shaded forest understories in southern Spain Plant Ecology, 168, 139-163. [本文引用: 3]
SunH, TangY, HuangXJ, HuangCM (2005). Present situations and its R & D of dry valleys in the Hengduan Mountains of SW China World Sci-Tech R & D, 27, 54-61. [本文引用: 1]
TeteganM, de ForgesACR, VerbequeB, NicoullaudB, DesbourdesC, BouthierA, ArrouaysD, CousinI (2015). The effect of soil stoniness on the estimation of water retention properties of soils: a case study from central France Catena, 129, 95-102. [本文引用: 1]
WuFZ, BaoWK, ZhouZQ, LiFL (2012). Appropriate nitrogen supply could improve soil microbial and chemical characteristics with Sophora davidii seedlings cultivated in water stress conditions Acta Agriculturae Scandinavica, Section B: Soil & Plant Science, 62, 49-58. [本文引用: 4]
XuXL, MaKM, FuBJ, SongCJ, LiuW (2008). Influence of three plant species with different morphologies on water runoff and soil loss in a dry-warm river valley, SW China Forest Ecology and Management, 256, 656-663. [本文引用: 1]
ZhangRZ (1992). The Dry Valleys of the Hengduan Mountains Region. Science Press, Beijing. 1-211. [本文引用: 1]
ZhouZQ, BaoWK (2011). Levels of physiological dormancy and methods for improving seed germination of four rose species Scientia Horticulturae, 129, 818-824. [本文引用: 1]
ZhuLH, BaoWK, HeBH (2009). Assessment on ecological restoration effect of afforestation with Cupressus chengiana seedlings in the dry Minjiang River valley, southwestern China Chinese Journal of Applied and Environmental Biology, 15, 774-780. [本文引用: 1]
... 干旱胁迫与土壤贫瘠共同制约了干旱河谷区植物生长与植被发育(Li et al., 2008; Wu et al., 2012).由于幼苗对环境胁迫的忍受能力弱, 幼苗死亡率高是绝大多数生态系统中种群更新的“瓶颈” (Engelbrecht et al., 2005; Han et al., 2018).在干旱河谷区的调查也发现, 自然生态系统中种子萌发率低, 而幼苗死亡率很高, 导致幼苗稀少, 植被更新困难(李芳兰等, 2009; Li et al., 2011; 包维楷等, 2012).干旱河谷的自然植被存在灌丛“肥岛”效应(Qu et al., 2017).土壤资源、光照、温度等环境因子在植被斑块与斑块及裸地之间差异明显(李君等, 2007; Qu et al., 2017).幼苗的可塑性较强, 其通过改变生长速率、生物量分配和生理特征适应异质环境(Lehouck et al., 2008).因此, 理解幼苗定植能力的微尺度空间变化及其限制因子, 突破环境胁迫对幼苗定植的限制问题, 将为自然植被更新和群落结构完善提供理论依据(Lehouck et al., 2008).此外, 在典型生态退化地段采用微生境调控技术促进乡土植物幼苗定植、群落构建和稳定性维持, 能够缓解严酷环境对区域植被恢复的限制.虽然我们前期系统揭示了干旱河谷植物自然更新潜力与空间分布特点, 以及种子与幼苗对干旱和土壤养分胁迫的响应机制(Li et al., 2008; Zhou & Bao, 2011; Wu et al., 2012), 但是对幼苗定植过程中灌丛“肥岛”的效应解释较少. ...
... 干旱胁迫与土壤贫瘠共同制约了干旱河谷区植物生长与植被发育(Li et al., 2008; Wu et al., 2012).由于幼苗对环境胁迫的忍受能力弱, 幼苗死亡率高是绝大多数生态系统中种群更新的“瓶颈” (Engelbrecht et al., 2005; Han et al., 2018).在干旱河谷区的调查也发现, 自然生态系统中种子萌发率低, 而幼苗死亡率很高, 导致幼苗稀少, 植被更新困难(李芳兰等, 2009; Li et al., 2011; 包维楷等, 2012).干旱河谷的自然植被存在灌丛“肥岛”效应(Qu et al., 2017).土壤资源、光照、温度等环境因子在植被斑块与斑块及裸地之间差异明显(李君等, 2007; Qu et al., 2017).幼苗的可塑性较强, 其通过改变生长速率、生物量分配和生理特征适应异质环境(Lehouck et al., 2008).因此, 理解幼苗定植能力的微尺度空间变化及其限制因子, 突破环境胁迫对幼苗定植的限制问题, 将为自然植被更新和群落结构完善提供理论依据(Lehouck et al., 2008).此外, 在典型生态退化地段采用微生境调控技术促进乡土植物幼苗定植、群落构建和稳定性维持, 能够缓解严酷环境对区域植被恢复的限制.虽然我们前期系统揭示了干旱河谷植物自然更新潜力与空间分布特点, 以及种子与幼苗对干旱和土壤养分胁迫的响应机制(Li et al., 2008; Zhou & Bao, 2011; Wu et al., 2012), 但是对幼苗定植过程中灌丛“肥岛”的效应解释较少. ...
Leaf structure and anatomy as related to leaf mass per area variation in seedlings of a wide range of woody plant species and types 1 2000
... 不同微生境之间幼苗生物量分配和叶片功能性状的差异体现了植物适应环境变化的功能策略.Sack等(2003)认为植物通过减少资源需求适应环境变化, 例如干旱胁迫下植株通常变矮, 便于水分从根向叶片传输.另外, 单叶片面积和数量减小均降低蒸腾, 是植物对干旱胁迫的适应方式(Chartzoulakis et al., 1993; Gindaba et al., 2004; Wright et al., 2017).从不同器官生物量分配格局来看, 裸地的植物为了减少资源需求, 将更多的碳投入到支撑器官(根和茎), 因此其生物量分配比例增加(图3).相反, 在植被覆盖斑块中, 光、热等资源受到限制, 植物将更多的碳投入到地上部分去捕获更多的光热资源, 从而导致叶片或者枝条生物量的比例增加(图3), 此结果证实了功能平衡假说(Poorter et al., 2012).此外, 叶片性状对光照和水分最敏感(Chartzoulakis et al., 1993).在裸地上, 幼苗比叶面积较小, 可能是在于强光环境下叶片具有较少细胞或单个细胞有较大生物量, 或者叶片较厚(Casrto-Diez et al., 2000), 因为比叶面积较小的个体具有较强的支持和抵御功能(Wright et al., 2017).幼苗在植被斑块中具有较大的比叶面积, 主要在于叶片较薄, 这对适应冠层下弱光条件至关重要(Milbau et al., 2005).Sack等(2003)等对林下植物观测数据表明弱光条件下, 叶片N含量较高, 寿命较短, 比叶面积较大.我们的结论与此一致. ...
Rock fragments in soil support a different microbial community from the fine earth 1 2004
... 在极度退化的干旱生态系统中, 土壤干旱、贫瘠和根系穿透阻力大等环境胁迫并存(Certini et al., 2004; 刘春霞和韩烈保, 2007; Estrada-Medina et al., 2013; Lynch, 2019), 其中干旱胁迫和土壤结构退化往往相互促进(de Kroon & Visser, 2003; Tetegan et al., 2015; Mi et al., 2016).我们针对干旱河谷区幼苗萌发和生长的关键制约因子, 进行不同处理的对比, 发现地表覆盖和施肥是提高道路边坡乡土植物幼苗定植和维持群落结构稳定的有效措施.试验结果显示, 土壤施肥显著提高了植株密度(图6A), 这与荒坡不同微生境斑块研究结论一致: 在干旱河谷地区及工程扰动区退化生态地段的植被恢复中适度施肥不仅能够提高乡土植物生长和存活, 而且能够维持群落稳定发展.我们前期采用水分和施氮两因素控制试验发现, 适度施氮能够提高灌木幼苗的水分利用效率和抗旱能力(Wu et al., 2012), 为本试验对自然条件下植物群落结构的观测结论提供了相关生理生态学证据. ...
Photosynthesis, plant growth and dry matter distribution in kiwifruit as influenced by water deficits 2 1993
... 不同微生境之间幼苗生物量分配和叶片功能性状的差异体现了植物适应环境变化的功能策略.Sack等(2003)认为植物通过减少资源需求适应环境变化, 例如干旱胁迫下植株通常变矮, 便于水分从根向叶片传输.另外, 单叶片面积和数量减小均降低蒸腾, 是植物对干旱胁迫的适应方式(Chartzoulakis et al., 1993; Gindaba et al., 2004; Wright et al., 2017).从不同器官生物量分配格局来看, 裸地的植物为了减少资源需求, 将更多的碳投入到支撑器官(根和茎), 因此其生物量分配比例增加(图3).相反, 在植被覆盖斑块中, 光、热等资源受到限制, 植物将更多的碳投入到地上部分去捕获更多的光热资源, 从而导致叶片或者枝条生物量的比例增加(图3), 此结果证实了功能平衡假说(Poorter et al., 2012).此外, 叶片性状对光照和水分最敏感(Chartzoulakis et al., 1993).在裸地上, 幼苗比叶面积较小, 可能是在于强光环境下叶片具有较少细胞或单个细胞有较大生物量, 或者叶片较厚(Casrto-Diez et al., 2000), 因为比叶面积较小的个体具有较强的支持和抵御功能(Wright et al., 2017).幼苗在植被斑块中具有较大的比叶面积, 主要在于叶片较薄, 这对适应冠层下弱光条件至关重要(Milbau et al., 2005).Sack等(2003)等对林下植物观测数据表明弱光条件下, 叶片N含量较高, 寿命较短, 比叶面积较大.我们的结论与此一致. ...
... ).此外, 叶片性状对光照和水分最敏感(Chartzoulakis et al., 1993).在裸地上, 幼苗比叶面积较小, 可能是在于强光环境下叶片具有较少细胞或单个细胞有较大生物量, 或者叶片较厚(Casrto-Diez et al., 2000), 因为比叶面积较小的个体具有较强的支持和抵御功能(Wright et al., 2017).幼苗在植被斑块中具有较大的比叶面积, 主要在于叶片较薄, 这对适应冠层下弱光条件至关重要(Milbau et al., 2005).Sack等(2003)等对林下植物观测数据表明弱光条件下, 叶片N含量较高, 寿命较短, 比叶面积较大.我们的结论与此一致. ...
Root Ecology. 1 2003
... 在极度退化的干旱生态系统中, 土壤干旱、贫瘠和根系穿透阻力大等环境胁迫并存(Certini et al., 2004; 刘春霞和韩烈保, 2007; Estrada-Medina et al., 2013; Lynch, 2019), 其中干旱胁迫和土壤结构退化往往相互促进(de Kroon & Visser, 2003; Tetegan et al., 2015; Mi et al., 2016).我们针对干旱河谷区幼苗萌发和生长的关键制约因子, 进行不同处理的对比, 发现地表覆盖和施肥是提高道路边坡乡土植物幼苗定植和维持群落结构稳定的有效措施.试验结果显示, 土壤施肥显著提高了植株密度(图6A), 这与荒坡不同微生境斑块研究结论一致: 在干旱河谷地区及工程扰动区退化生态地段的植被恢复中适度施肥不仅能够提高乡土植物生长和存活, 而且能够维持群落稳定发展.我们前期采用水分和施氮两因素控制试验发现, 适度施氮能够提高灌木幼苗的水分利用效率和抗旱能力(Wu et al., 2012), 为本试验对自然条件下植物群落结构的观测结论提供了相关生理生态学证据. ...
Drought effects on seedling survival in a tropical moist forest 1 2005
... 干旱胁迫与土壤贫瘠共同制约了干旱河谷区植物生长与植被发育(Li et al., 2008; Wu et al., 2012).由于幼苗对环境胁迫的忍受能力弱, 幼苗死亡率高是绝大多数生态系统中种群更新的“瓶颈” (Engelbrecht et al., 2005; Han et al., 2018).在干旱河谷区的调查也发现, 自然生态系统中种子萌发率低, 而幼苗死亡率很高, 导致幼苗稀少, 植被更新困难(李芳兰等, 2009; Li et al., 2011; 包维楷等, 2012).干旱河谷的自然植被存在灌丛“肥岛”效应(Qu et al., 2017).土壤资源、光照、温度等环境因子在植被斑块与斑块及裸地之间差异明显(李君等, 2007; Qu et al., 2017).幼苗的可塑性较强, 其通过改变生长速率、生物量分配和生理特征适应异质环境(Lehouck et al., 2008).因此, 理解幼苗定植能力的微尺度空间变化及其限制因子, 突破环境胁迫对幼苗定植的限制问题, 将为自然植被更新和群落结构完善提供理论依据(Lehouck et al., 2008).此外, 在典型生态退化地段采用微生境调控技术促进乡土植物幼苗定植、群落构建和稳定性维持, 能够缓解严酷环境对区域植被恢复的限制.虽然我们前期系统揭示了干旱河谷植物自然更新潜力与空间分布特点, 以及种子与幼苗对干旱和土壤养分胁迫的响应机制(Li et al., 2008; Zhou & Bao, 2011; Wu et al., 2012), 但是对幼苗定植过程中灌丛“肥岛”的效应解释较少. ...
The importance of limestone bedrock and dissolution karst features on tree root distribution in northern Yucatán, México 1 2013
... 在极度退化的干旱生态系统中, 土壤干旱、贫瘠和根系穿透阻力大等环境胁迫并存(Certini et al., 2004; 刘春霞和韩烈保, 2007; Estrada-Medina et al., 2013; Lynch, 2019), 其中干旱胁迫和土壤结构退化往往相互促进(de Kroon & Visser, 2003; Tetegan et al., 2015; Mi et al., 2016).我们针对干旱河谷区幼苗萌发和生长的关键制约因子, 进行不同处理的对比, 发现地表覆盖和施肥是提高道路边坡乡土植物幼苗定植和维持群落结构稳定的有效措施.试验结果显示, 土壤施肥显著提高了植株密度(图6A), 这与荒坡不同微生境斑块研究结论一致: 在干旱河谷地区及工程扰动区退化生态地段的植被恢复中适度施肥不仅能够提高乡土植物生长和存活, 而且能够维持群落稳定发展.我们前期采用水分和施氮两因素控制试验发现, 适度施氮能够提高灌木幼苗的水分利用效率和抗旱能力(Wu et al., 2012), 为本试验对自然条件下植物群落结构的观测结论提供了相关生理生态学证据. ...
Response of seedlings of two Eucalyptus and three deciduous tree species from Ethiopia to severe water stress 1 2004
... 不同微生境之间幼苗生物量分配和叶片功能性状的差异体现了植物适应环境变化的功能策略.Sack等(2003)认为植物通过减少资源需求适应环境变化, 例如干旱胁迫下植株通常变矮, 便于水分从根向叶片传输.另外, 单叶片面积和数量减小均降低蒸腾, 是植物对干旱胁迫的适应方式(Chartzoulakis et al., 1993; Gindaba et al., 2004; Wright et al., 2017).从不同器官生物量分配格局来看, 裸地的植物为了减少资源需求, 将更多的碳投入到支撑器官(根和茎), 因此其生物量分配比例增加(图3).相反, 在植被覆盖斑块中, 光、热等资源受到限制, 植物将更多的碳投入到地上部分去捕获更多的光热资源, 从而导致叶片或者枝条生物量的比例增加(图3), 此结果证实了功能平衡假说(Poorter et al., 2012).此外, 叶片性状对光照和水分最敏感(Chartzoulakis et al., 1993).在裸地上, 幼苗比叶面积较小, 可能是在于强光环境下叶片具有较少细胞或单个细胞有较大生物量, 或者叶片较厚(Casrto-Diez et al., 2000), 因为比叶面积较小的个体具有较强的支持和抵御功能(Wright et al., 2017).幼苗在植被斑块中具有较大的比叶面积, 主要在于叶片较薄, 这对适应冠层下弱光条件至关重要(Milbau et al., 2005).Sack等(2003)等对林下植物观测数据表明弱光条件下, 叶片N含量较高, 寿命较短, 比叶面积较大.我们的结论与此一致. ...
Seed germination and initial seedling survival of the subalpine tree species, Picea jezoensis, on different forest floor substrates under elevated temperature 1 2018
... 干旱胁迫与土壤贫瘠共同制约了干旱河谷区植物生长与植被发育(Li et al., 2008; Wu et al., 2012).由于幼苗对环境胁迫的忍受能力弱, 幼苗死亡率高是绝大多数生态系统中种群更新的“瓶颈” (Engelbrecht et al., 2005; Han et al., 2018).在干旱河谷区的调查也发现, 自然生态系统中种子萌发率低, 而幼苗死亡率很高, 导致幼苗稀少, 植被更新困难(李芳兰等, 2009; Li et al., 2011; 包维楷等, 2012).干旱河谷的自然植被存在灌丛“肥岛”效应(Qu et al., 2017).土壤资源、光照、温度等环境因子在植被斑块与斑块及裸地之间差异明显(李君等, 2007; Qu et al., 2017).幼苗的可塑性较强, 其通过改变生长速率、生物量分配和生理特征适应异质环境(Lehouck et al., 2008).因此, 理解幼苗定植能力的微尺度空间变化及其限制因子, 突破环境胁迫对幼苗定植的限制问题, 将为自然植被更新和群落结构完善提供理论依据(Lehouck et al., 2008).此外, 在典型生态退化地段采用微生境调控技术促进乡土植物幼苗定植、群落构建和稳定性维持, 能够缓解严酷环境对区域植被恢复的限制.虽然我们前期系统揭示了干旱河谷植物自然更新潜力与空间分布特点, 以及种子与幼苗对干旱和土壤养分胁迫的响应机制(Li et al., 2008; Zhou & Bao, 2011; Wu et al., 2012), 但是对幼苗定植过程中灌丛“肥岛”的效应解释较少. ...
... 虽然不同斑块类型之间土壤有机质和N含量等存在一定的差异(表1), 但是我们发现植被斑块中土壤添加养分(消除养分限制)对幼苗生物量未产生显著影响; 而裸地上添加养分显著增加了幼苗生物量(图2), 进一步证明养分限制不是幼苗生物量变化的主导因子.前期研究发现不同时空尺度上100 cm土层深度的土壤储水量在36.2-151.2 mm之间剧烈变化(包维楷等, 2012), 干旱胁迫是制约区域植被发育的关键.裸地上, 光、热资源充足(Leck et al., 2008; Qu et al., 2017), 植物种间或者种内资源竞争较小, 对小降雨事件响应敏感(Leck et al., 2008), 有利于幼苗生长.相反灌木覆盖对土壤水分含量具有负面影响(Qu et al., 2017), 这可能限制幼苗生长. ...
... ), 植物种间或者种内资源竞争较小, 对小降雨事件响应敏感(Leck et al., 2008), 有利于幼苗生长.相反灌木覆盖对土壤水分含量具有负面影响(Qu et al., 2017), 这可能限制幼苗生长. ...
Spatial and temporal effects on recruitment of an Afromontane forest tree in a threatened fragmented ecosystem 4 2009
... 干旱胁迫与土壤贫瘠共同制约了干旱河谷区植物生长与植被发育(Li et al., 2008; Wu et al., 2012).由于幼苗对环境胁迫的忍受能力弱, 幼苗死亡率高是绝大多数生态系统中种群更新的“瓶颈” (Engelbrecht et al., 2005; Han et al., 2018).在干旱河谷区的调查也发现, 自然生态系统中种子萌发率低, 而幼苗死亡率很高, 导致幼苗稀少, 植被更新困难(李芳兰等, 2009; Li et al., 2011; 包维楷等, 2012).干旱河谷的自然植被存在灌丛“肥岛”效应(Qu et al., 2017).土壤资源、光照、温度等环境因子在植被斑块与斑块及裸地之间差异明显(李君等, 2007; Qu et al., 2017).幼苗的可塑性较强, 其通过改变生长速率、生物量分配和生理特征适应异质环境(Lehouck et al., 2008).因此, 理解幼苗定植能力的微尺度空间变化及其限制因子, 突破环境胁迫对幼苗定植的限制问题, 将为自然植被更新和群落结构完善提供理论依据(Lehouck et al., 2008).此外, 在典型生态退化地段采用微生境调控技术促进乡土植物幼苗定植、群落构建和稳定性维持, 能够缓解严酷环境对区域植被恢复的限制.虽然我们前期系统揭示了干旱河谷植物自然更新潜力与空间分布特点, 以及种子与幼苗对干旱和土壤养分胁迫的响应机制(Li et al., 2008; Zhou & Bao, 2011; Wu et al., 2012), 但是对幼苗定植过程中灌丛“肥岛”的效应解释较少. ...
... ).因此, 理解幼苗定植能力的微尺度空间变化及其限制因子, 突破环境胁迫对幼苗定植的限制问题, 将为自然植被更新和群落结构完善提供理论依据(Lehouck et al., 2008).此外, 在典型生态退化地段采用微生境调控技术促进乡土植物幼苗定植、群落构建和稳定性维持, 能够缓解严酷环境对区域植被恢复的限制.虽然我们前期系统揭示了干旱河谷植物自然更新潜力与空间分布特点, 以及种子与幼苗对干旱和土壤养分胁迫的响应机制(Li et al., 2008; Zhou & Bao, 2011; Wu et al., 2012), 但是对幼苗定植过程中灌丛“肥岛”的效应解释较少. ...
... 干旱胁迫与土壤贫瘠共同制约了干旱河谷区植物生长与植被发育(Li et al., 2008; Wu et al., 2012).由于幼苗对环境胁迫的忍受能力弱, 幼苗死亡率高是绝大多数生态系统中种群更新的“瓶颈” (Engelbrecht et al., 2005; Han et al., 2018).在干旱河谷区的调查也发现, 自然生态系统中种子萌发率低, 而幼苗死亡率很高, 导致幼苗稀少, 植被更新困难(李芳兰等, 2009; Li et al., 2011; 包维楷等, 2012).干旱河谷的自然植被存在灌丛“肥岛”效应(Qu et al., 2017).土壤资源、光照、温度等环境因子在植被斑块与斑块及裸地之间差异明显(李君等, 2007; Qu et al., 2017).幼苗的可塑性较强, 其通过改变生长速率、生物量分配和生理特征适应异质环境(Lehouck et al., 2008).因此, 理解幼苗定植能力的微尺度空间变化及其限制因子, 突破环境胁迫对幼苗定植的限制问题, 将为自然植被更新和群落结构完善提供理论依据(Lehouck et al., 2008).此外, 在典型生态退化地段采用微生境调控技术促进乡土植物幼苗定植、群落构建和稳定性维持, 能够缓解严酷环境对区域植被恢复的限制.虽然我们前期系统揭示了干旱河谷植物自然更新潜力与空间分布特点, 以及种子与幼苗对干旱和土壤养分胁迫的响应机制(Li et al., 2008; Zhou & Bao, 2011; Wu et al., 2012), 但是对幼苗定植过程中灌丛“肥岛”的效应解释较少. ...
... 干旱胁迫与土壤贫瘠共同制约了干旱河谷区植物生长与植被发育(Li et al., 2008; Wu et al., 2012).由于幼苗对环境胁迫的忍受能力弱, 幼苗死亡率高是绝大多数生态系统中种群更新的“瓶颈” (Engelbrecht et al., 2005; Han et al., 2018).在干旱河谷区的调查也发现, 自然生态系统中种子萌发率低, 而幼苗死亡率很高, 导致幼苗稀少, 植被更新困难(李芳兰等, 2009; Li et al., 2011; 包维楷等, 2012).干旱河谷的自然植被存在灌丛“肥岛”效应(Qu et al., 2017).土壤资源、光照、温度等环境因子在植被斑块与斑块及裸地之间差异明显(李君等, 2007; Qu et al., 2017).幼苗的可塑性较强, 其通过改变生长速率、生物量分配和生理特征适应异质环境(Lehouck et al., 2008).因此, 理解幼苗定植能力的微尺度空间变化及其限制因子, 突破环境胁迫对幼苗定植的限制问题, 将为自然植被更新和群落结构完善提供理论依据(Lehouck et al., 2008).此外, 在典型生态退化地段采用微生境调控技术促进乡土植物幼苗定植、群落构建和稳定性维持, 能够缓解严酷环境对区域植被恢复的限制.虽然我们前期系统揭示了干旱河谷植物自然更新潜力与空间分布特点, 以及种子与幼苗对干旱和土壤养分胁迫的响应机制(Li et al., 2008; Zhou & Bao, 2011; Wu et al., 2012), 但是对幼苗定植过程中灌丛“肥岛”的效应解释较少. ...
... 干旱胁迫与土壤贫瘠共同制约了干旱河谷区植物生长与植被发育(Li et al., 2008; Wu et al., 2012).由于幼苗对环境胁迫的忍受能力弱, 幼苗死亡率高是绝大多数生态系统中种群更新的“瓶颈” (Engelbrecht et al., 2005; Han et al., 2018).在干旱河谷区的调查也发现, 自然生态系统中种子萌发率低, 而幼苗死亡率很高, 导致幼苗稀少, 植被更新困难(李芳兰等, 2009; Li et al., 2011; 包维楷等, 2012).干旱河谷的自然植被存在灌丛“肥岛”效应(Qu et al., 2017).土壤资源、光照、温度等环境因子在植被斑块与斑块及裸地之间差异明显(李君等, 2007; Qu et al., 2017).幼苗的可塑性较强, 其通过改变生长速率、生物量分配和生理特征适应异质环境(Lehouck et al., 2008).因此, 理解幼苗定植能力的微尺度空间变化及其限制因子, 突破环境胁迫对幼苗定植的限制问题, 将为自然植被更新和群落结构完善提供理论依据(Lehouck et al., 2008).此外, 在典型生态退化地段采用微生境调控技术促进乡土植物幼苗定植、群落构建和稳定性维持, 能够缓解严酷环境对区域植被恢复的限制.虽然我们前期系统揭示了干旱河谷植物自然更新潜力与空间分布特点, 以及种子与幼苗对干旱和土壤养分胁迫的响应机制(Li et al., 2008; Zhou & Bao, 2011; Wu et al., 2012), 但是对幼苗定植过程中灌丛“肥岛”的效应解释较少. ...
... 干旱胁迫与土壤贫瘠共同制约了干旱河谷区植物生长与植被发育(Li et al., 2008; Wu et al., 2012).由于幼苗对环境胁迫的忍受能力弱, 幼苗死亡率高是绝大多数生态系统中种群更新的“瓶颈” (Engelbrecht et al., 2005; Han et al., 2018).在干旱河谷区的调查也发现, 自然生态系统中种子萌发率低, 而幼苗死亡率很高, 导致幼苗稀少, 植被更新困难(李芳兰等, 2009; Li et al., 2011; 包维楷等, 2012).干旱河谷的自然植被存在灌丛“肥岛”效应(Qu et al., 2017).土壤资源、光照、温度等环境因子在植被斑块与斑块及裸地之间差异明显(李君等, 2007; Qu et al., 2017).幼苗的可塑性较强, 其通过改变生长速率、生物量分配和生理特征适应异质环境(Lehouck et al., 2008).因此, 理解幼苗定植能力的微尺度空间变化及其限制因子, 突破环境胁迫对幼苗定植的限制问题, 将为自然植被更新和群落结构完善提供理论依据(Lehouck et al., 2008).此外, 在典型生态退化地段采用微生境调控技术促进乡土植物幼苗定植、群落构建和稳定性维持, 能够缓解严酷环境对区域植被恢复的限制.虽然我们前期系统揭示了干旱河谷植物自然更新潜力与空间分布特点, 以及种子与幼苗对干旱和土壤养分胁迫的响应机制(Li et al., 2008; Zhou & Bao, 2011; Wu et al., 2012), 但是对幼苗定植过程中灌丛“肥岛”的效应解释较少. ...
Fertile islands under Artemisia ordosica in inland dunes of northern China: effects of habitats and plant developmental stages 4 2008
... 干旱胁迫与土壤贫瘠共同制约了干旱河谷区植物生长与植被发育(Li et al., 2008; Wu et al., 2012).由于幼苗对环境胁迫的忍受能力弱, 幼苗死亡率高是绝大多数生态系统中种群更新的“瓶颈” (Engelbrecht et al., 2005; Han et al., 2018).在干旱河谷区的调查也发现, 自然生态系统中种子萌发率低, 而幼苗死亡率很高, 导致幼苗稀少, 植被更新困难(李芳兰等, 2009; Li et al., 2011; 包维楷等, 2012).干旱河谷的自然植被存在灌丛“肥岛”效应(Qu et al., 2017).土壤资源、光照、温度等环境因子在植被斑块与斑块及裸地之间差异明显(李君等, 2007; Qu et al., 2017).幼苗的可塑性较强, 其通过改变生长速率、生物量分配和生理特征适应异质环境(Lehouck et al., 2008).因此, 理解幼苗定植能力的微尺度空间变化及其限制因子, 突破环境胁迫对幼苗定植的限制问题, 将为自然植被更新和群落结构完善提供理论依据(Lehouck et al., 2008).此外, 在典型生态退化地段采用微生境调控技术促进乡土植物幼苗定植、群落构建和稳定性维持, 能够缓解严酷环境对区域植被恢复的限制.虽然我们前期系统揭示了干旱河谷植物自然更新潜力与空间分布特点, 以及种子与幼苗对干旱和土壤养分胁迫的响应机制(Li et al., 2008; Zhou & Bao, 2011; Wu et al., 2012), 但是对幼苗定植过程中灌丛“肥岛”的效应解释较少. ...
... ).此外, 在典型生态退化地段采用微生境调控技术促进乡土植物幼苗定植、群落构建和稳定性维持, 能够缓解严酷环境对区域植被恢复的限制.虽然我们前期系统揭示了干旱河谷植物自然更新潜力与空间分布特点, 以及种子与幼苗对干旱和土壤养分胁迫的响应机制(Li et al., 2008; Zhou & Bao, 2011; Wu et al., 2012), 但是对幼苗定植过程中灌丛“肥岛”的效应解释较少. ...
Spatial patterns of the soil seed bank and extant vegetation across the dry Minjiang River valley in southwest China 1 2011
... 干旱胁迫与土壤贫瘠共同制约了干旱河谷区植物生长与植被发育(Li et al., 2008; Wu et al., 2012).由于幼苗对环境胁迫的忍受能力弱, 幼苗死亡率高是绝大多数生态系统中种群更新的“瓶颈” (Engelbrecht et al., 2005; Han et al., 2018).在干旱河谷区的调查也发现, 自然生态系统中种子萌发率低, 而幼苗死亡率很高, 导致幼苗稀少, 植被更新困难(李芳兰等, 2009; Li et al., 2011; 包维楷等, 2012).干旱河谷的自然植被存在灌丛“肥岛”效应(Qu et al., 2017).土壤资源、光照、温度等环境因子在植被斑块与斑块及裸地之间差异明显(李君等, 2007; Qu et al., 2017).幼苗的可塑性较强, 其通过改变生长速率、生物量分配和生理特征适应异质环境(Lehouck et al., 2008).因此, 理解幼苗定植能力的微尺度空间变化及其限制因子, 突破环境胁迫对幼苗定植的限制问题, 将为自然植被更新和群落结构完善提供理论依据(Lehouck et al., 2008).此外, 在典型生态退化地段采用微生境调控技术促进乡土植物幼苗定植、群落构建和稳定性维持, 能够缓解严酷环境对区域植被恢复的限制.虽然我们前期系统揭示了干旱河谷植物自然更新潜力与空间分布特点, 以及种子与幼苗对干旱和土壤养分胁迫的响应机制(Li et al., 2008; Zhou & Bao, 2011; Wu et al., 2012), 但是对幼苗定植过程中灌丛“肥岛”的效应解释较少. ...
高速公路边坡植被恢复研究进展 1 2007
... 在极度退化的干旱生态系统中, 土壤干旱、贫瘠和根系穿透阻力大等环境胁迫并存(Certini et al., 2004; 刘春霞和韩烈保, 2007; Estrada-Medina et al., 2013; Lynch, 2019), 其中干旱胁迫和土壤结构退化往往相互促进(de Kroon & Visser, 2003; Tetegan et al., 2015; Mi et al., 2016).我们针对干旱河谷区幼苗萌发和生长的关键制约因子, 进行不同处理的对比, 发现地表覆盖和施肥是提高道路边坡乡土植物幼苗定植和维持群落结构稳定的有效措施.试验结果显示, 土壤施肥显著提高了植株密度(图6A), 这与荒坡不同微生境斑块研究结论一致: 在干旱河谷地区及工程扰动区退化生态地段的植被恢复中适度施肥不仅能够提高乡土植物生长和存活, 而且能够维持群落稳定发展.我们前期采用水分和施氮两因素控制试验发现, 适度施氮能够提高灌木幼苗的水分利用效率和抗旱能力(Wu et al., 2012), 为本试验对自然条件下植物群落结构的观测结论提供了相关生理生态学证据. ...
高速公路边坡植被恢复研究进展 1 2007
... 在极度退化的干旱生态系统中, 土壤干旱、贫瘠和根系穿透阻力大等环境胁迫并存(Certini et al., 2004; 刘春霞和韩烈保, 2007; Estrada-Medina et al., 2013; Lynch, 2019), 其中干旱胁迫和土壤结构退化往往相互促进(de Kroon & Visser, 2003; Tetegan et al., 2015; Mi et al., 2016).我们针对干旱河谷区幼苗萌发和生长的关键制约因子, 进行不同处理的对比, 发现地表覆盖和施肥是提高道路边坡乡土植物幼苗定植和维持群落结构稳定的有效措施.试验结果显示, 土壤施肥显著提高了植株密度(图6A), 这与荒坡不同微生境斑块研究结论一致: 在干旱河谷地区及工程扰动区退化生态地段的植被恢复中适度施肥不仅能够提高乡土植物生长和存活, 而且能够维持群落稳定发展.我们前期采用水分和施氮两因素控制试验发现, 适度施氮能够提高灌木幼苗的水分利用效率和抗旱能力(Wu et al., 2012), 为本试验对自然条件下植物群落结构的观测结论提供了相关生理生态学证据. ...
... 干旱胁迫与土壤贫瘠共同制约了干旱河谷区植物生长与植被发育(Li et al., 2008; Wu et al., 2012).由于幼苗对环境胁迫的忍受能力弱, 幼苗死亡率高是绝大多数生态系统中种群更新的“瓶颈” (Engelbrecht et al., 2005; Han et al., 2018).在干旱河谷区的调查也发现, 自然生态系统中种子萌发率低, 而幼苗死亡率很高, 导致幼苗稀少, 植被更新困难(李芳兰等, 2009; Li et al., 2011; 包维楷等, 2012).干旱河谷的自然植被存在灌丛“肥岛”效应(Qu et al., 2017).土壤资源、光照、温度等环境因子在植被斑块与斑块及裸地之间差异明显(李君等, 2007; Qu et al., 2017).幼苗的可塑性较强, 其通过改变生长速率、生物量分配和生理特征适应异质环境(Lehouck et al., 2008).因此, 理解幼苗定植能力的微尺度空间变化及其限制因子, 突破环境胁迫对幼苗定植的限制问题, 将为自然植被更新和群落结构完善提供理论依据(Lehouck et al., 2008).此外, 在典型生态退化地段采用微生境调控技术促进乡土植物幼苗定植、群落构建和稳定性维持, 能够缓解严酷环境对区域植被恢复的限制.虽然我们前期系统揭示了干旱河谷植物自然更新潜力与空间分布特点, 以及种子与幼苗对干旱和土壤养分胁迫的响应机制(Li et al., 2008; Zhou & Bao, 2011; Wu et al., 2012), 但是对幼苗定植过程中灌丛“肥岛”的效应解释较少. ...
... ; Qu et al., 2017).幼苗的可塑性较强, 其通过改变生长速率、生物量分配和生理特征适应异质环境(Lehouck et al., 2008).因此, 理解幼苗定植能力的微尺度空间变化及其限制因子, 突破环境胁迫对幼苗定植的限制问题, 将为自然植被更新和群落结构完善提供理论依据(Lehouck et al., 2008).此外, 在典型生态退化地段采用微生境调控技术促进乡土植物幼苗定植、群落构建和稳定性维持, 能够缓解严酷环境对区域植被恢复的限制.虽然我们前期系统揭示了干旱河谷植物自然更新潜力与空间分布特点, 以及种子与幼苗对干旱和土壤养分胁迫的响应机制(Li et al., 2008; Zhou & Bao, 2011; Wu et al., 2012), 但是对幼苗定植过程中灌丛“肥岛”的效应解释较少. ...
The effect of soil stoniness on the estimation of water retention properties of soils: a case study from central France 1 2015
... 在极度退化的干旱生态系统中, 土壤干旱、贫瘠和根系穿透阻力大等环境胁迫并存(Certini et al., 2004; 刘春霞和韩烈保, 2007; Estrada-Medina et al., 2013; Lynch, 2019), 其中干旱胁迫和土壤结构退化往往相互促进(de Kroon & Visser, 2003; Tetegan et al., 2015; Mi et al., 2016).我们针对干旱河谷区幼苗萌发和生长的关键制约因子, 进行不同处理的对比, 发现地表覆盖和施肥是提高道路边坡乡土植物幼苗定植和维持群落结构稳定的有效措施.试验结果显示, 土壤施肥显著提高了植株密度(图6A), 这与荒坡不同微生境斑块研究结论一致: 在干旱河谷地区及工程扰动区退化生态地段的植被恢复中适度施肥不仅能够提高乡土植物生长和存活, 而且能够维持群落稳定发展.我们前期采用水分和施氮两因素控制试验发现, 适度施氮能够提高灌木幼苗的水分利用效率和抗旱能力(Wu et al., 2012), 为本试验对自然条件下植物群落结构的观测结论提供了相关生理生态学证据. ...
Global climatic drivers of leaf size 2 2017
... 不同微生境之间幼苗生物量分配和叶片功能性状的差异体现了植物适应环境变化的功能策略.Sack等(2003)认为植物通过减少资源需求适应环境变化, 例如干旱胁迫下植株通常变矮, 便于水分从根向叶片传输.另外, 单叶片面积和数量减小均降低蒸腾, 是植物对干旱胁迫的适应方式(Chartzoulakis et al., 1993; Gindaba et al., 2004; Wright et al., 2017).从不同器官生物量分配格局来看, 裸地的植物为了减少资源需求, 将更多的碳投入到支撑器官(根和茎), 因此其生物量分配比例增加(图3).相反, 在植被覆盖斑块中, 光、热等资源受到限制, 植物将更多的碳投入到地上部分去捕获更多的光热资源, 从而导致叶片或者枝条生物量的比例增加(图3), 此结果证实了功能平衡假说(Poorter et al., 2012).此外, 叶片性状对光照和水分最敏感(Chartzoulakis et al., 1993).在裸地上, 幼苗比叶面积较小, 可能是在于强光环境下叶片具有较少细胞或单个细胞有较大生物量, 或者叶片较厚(Casrto-Diez et al., 2000), 因为比叶面积较小的个体具有较强的支持和抵御功能(Wright et al., 2017).幼苗在植被斑块中具有较大的比叶面积, 主要在于叶片较薄, 这对适应冠层下弱光条件至关重要(Milbau et al., 2005).Sack等(2003)等对林下植物观测数据表明弱光条件下, 叶片N含量较高, 寿命较短, 比叶面积较大.我们的结论与此一致. ...
... ), 因为比叶面积较小的个体具有较强的支持和抵御功能(Wright et al., 2017).幼苗在植被斑块中具有较大的比叶面积, 主要在于叶片较薄, 这对适应冠层下弱光条件至关重要(Milbau et al., 2005).Sack等(2003)等对林下植物观测数据表明弱光条件下, 叶片N含量较高, 寿命较短, 比叶面积较大.我们的结论与此一致. ...
Appropriate nitrogen supply could improve soil microbial and chemical characteristics with Sophora davidii seedlings cultivated in water stress conditions 4 2012
... 干旱胁迫与土壤贫瘠共同制约了干旱河谷区植物生长与植被发育(Li et al., 2008; Wu et al., 2012).由于幼苗对环境胁迫的忍受能力弱, 幼苗死亡率高是绝大多数生态系统中种群更新的“瓶颈” (Engelbrecht et al., 2005; Han et al., 2018).在干旱河谷区的调查也发现, 自然生态系统中种子萌发率低, 而幼苗死亡率很高, 导致幼苗稀少, 植被更新困难(李芳兰等, 2009; Li et al., 2011; 包维楷等, 2012).干旱河谷的自然植被存在灌丛“肥岛”效应(Qu et al., 2017).土壤资源、光照、温度等环境因子在植被斑块与斑块及裸地之间差异明显(李君等, 2007; Qu et al., 2017).幼苗的可塑性较强, 其通过改变生长速率、生物量分配和生理特征适应异质环境(Lehouck et al., 2008).因此, 理解幼苗定植能力的微尺度空间变化及其限制因子, 突破环境胁迫对幼苗定植的限制问题, 将为自然植被更新和群落结构完善提供理论依据(Lehouck et al., 2008).此外, 在典型生态退化地段采用微生境调控技术促进乡土植物幼苗定植、群落构建和稳定性维持, 能够缓解严酷环境对区域植被恢复的限制.虽然我们前期系统揭示了干旱河谷植物自然更新潜力与空间分布特点, 以及种子与幼苗对干旱和土壤养分胁迫的响应机制(Li et al., 2008; Zhou & Bao, 2011; Wu et al., 2012), 但是对幼苗定植过程中灌丛“肥岛”的效应解释较少. ...
... ; Wu et al., 2012), 但是对幼苗定植过程中灌丛“肥岛”的效应解释较少. ...
... 6种乡土灌木、半灌木和草本植物对干旱河谷区土石混杂道路边坡环境具有较强的适应能力, 多数物种如早熟禾、狗尾草、毛莲蒿、小叶杭子梢4种植物出苗率较高, 大于60%, 幼苗保存率达到50%以上; 其中灌木幼苗保存率均大于75% (图1), 植株密度随着时间变化保持稳定(图6B).这些物种通过种子直播方式种植, 不仅成本较低、操作方便, 而且生长稳定.白刺花和四川黄栌种子出苗率较低, 小于40%, 但是其苗木具有较强抗旱能力(Li et al., 2009).近期研究发现, 这两种幼苗能够很好适应不同结构的土壤环境, 适宜采用幼苗移栽的种植方式去提高苗木定植能力.前期的植物生态适应性理论研究(Li et al., 2009; Wu et al., 2012)和本次野外播种试验结果证明, 6种乡土灌木和草本植物是干旱区极端退化的生态系统植被恢复的优良材料, 具有很好的推广应用价值. ...
... 在极度退化的干旱生态系统中, 土壤干旱、贫瘠和根系穿透阻力大等环境胁迫并存(Certini et al., 2004; 刘春霞和韩烈保, 2007; Estrada-Medina et al., 2013; Lynch, 2019), 其中干旱胁迫和土壤结构退化往往相互促进(de Kroon & Visser, 2003; Tetegan et al., 2015; Mi et al., 2016).我们针对干旱河谷区幼苗萌发和生长的关键制约因子, 进行不同处理的对比, 发现地表覆盖和施肥是提高道路边坡乡土植物幼苗定植和维持群落结构稳定的有效措施.试验结果显示, 土壤施肥显著提高了植株密度(图6A), 这与荒坡不同微生境斑块研究结论一致: 在干旱河谷地区及工程扰动区退化生态地段的植被恢复中适度施肥不仅能够提高乡土植物生长和存活, 而且能够维持群落稳定发展.我们前期采用水分和施氮两因素控制试验发现, 适度施氮能够提高灌木幼苗的水分利用效率和抗旱能力(Wu et al., 2012), 为本试验对自然条件下植物群落结构的观测结论提供了相关生理生态学证据. ...
Influence of three plant species with different morphologies on water runoff and soil loss in a dry-warm river valley, SW China 1 2008
Levels of physiological dormancy and methods for improving seed germination of four rose species 1 2011
... 干旱胁迫与土壤贫瘠共同制约了干旱河谷区植物生长与植被发育(Li et al., 2008; Wu et al., 2012).由于幼苗对环境胁迫的忍受能力弱, 幼苗死亡率高是绝大多数生态系统中种群更新的“瓶颈” (Engelbrecht et al., 2005; Han et al., 2018).在干旱河谷区的调查也发现, 自然生态系统中种子萌发率低, 而幼苗死亡率很高, 导致幼苗稀少, 植被更新困难(李芳兰等, 2009; Li et al., 2011; 包维楷等, 2012).干旱河谷的自然植被存在灌丛“肥岛”效应(Qu et al., 2017).土壤资源、光照、温度等环境因子在植被斑块与斑块及裸地之间差异明显(李君等, 2007; Qu et al., 2017).幼苗的可塑性较强, 其通过改变生长速率、生物量分配和生理特征适应异质环境(Lehouck et al., 2008).因此, 理解幼苗定植能力的微尺度空间变化及其限制因子, 突破环境胁迫对幼苗定植的限制问题, 将为自然植被更新和群落结构完善提供理论依据(Lehouck et al., 2008).此外, 在典型生态退化地段采用微生境调控技术促进乡土植物幼苗定植、群落构建和稳定性维持, 能够缓解严酷环境对区域植被恢复的限制.虽然我们前期系统揭示了干旱河谷植物自然更新潜力与空间分布特点, 以及种子与幼苗对干旱和土壤养分胁迫的响应机制(Li et al., 2008; Zhou & Bao, 2011; Wu et al., 2012), 但是对幼苗定植过程中灌丛“肥岛”的效应解释较少. ...
 ,1,*
,1,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT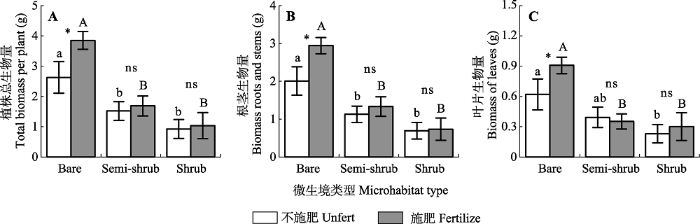 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT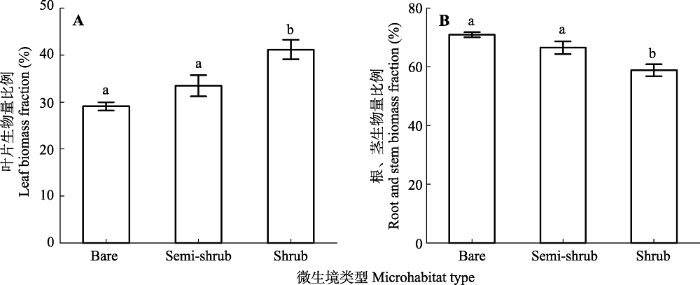 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT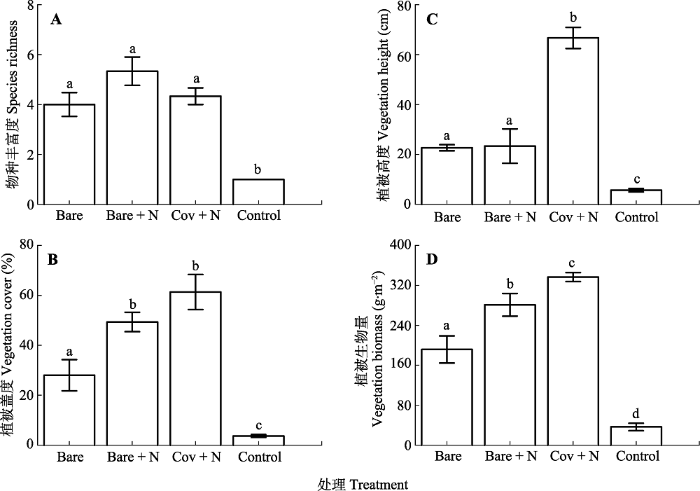 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT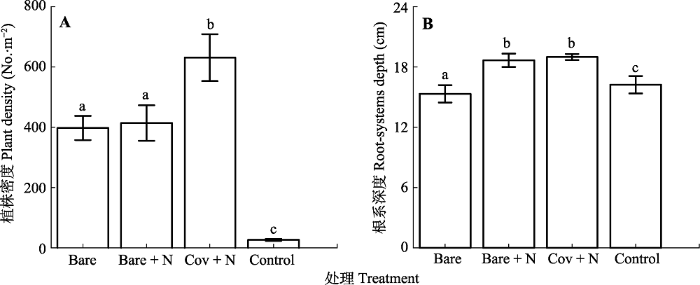 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT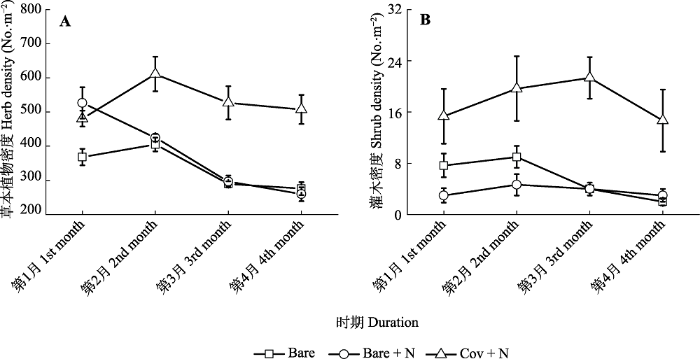 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT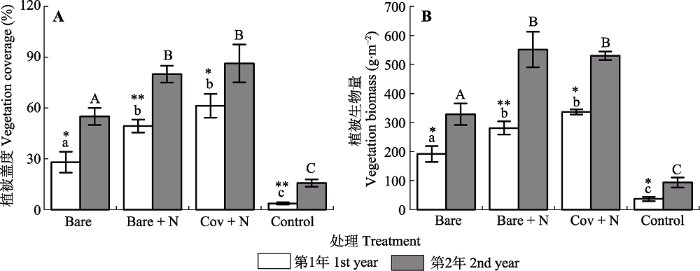 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}