莫丹, 王振孟, 左有璐, 向双
, , * 中国科学院成都生物研究所, 中国科学院山地生态恢复与生物资源利用重点实验室, 生态恢复与生物多样性保育四川省重点实验室, 成都 610041; 中国科学院大学, 北京 100049
Trade-off between shooting and leaf developing of woody species saplings in subtropical evergreen broad-leaved forests Dan MO, Zhen-Meng WANG, You-Lu ZUO, Shuang XIANG
, , * CAS Key Laboratory of Mountain Ecological Restoration and Bioresource Utilization & Ecological Restoration and Biodiversity Conservation Key Laboratory of Sichuan Province, Chengdu Institute of Biology, Chinese Academy of Sciences, Chengdu 610041, China; and University of Chinese Academy of Sciences, Beijing 100049, China
通讯作者: *向双:ORCID:0000-0002-7361-9723,
xiangshuang@cib.ac.cn 编委: 曾波
责任编辑: 李敏
收稿日期: 2020-05-11
接受日期: 2020-09-17
网络出版日期: 2020-10-20
基金资助: 国家自然科学基金 (31370594 )国家重点研发计划 (2017YFC0505000 )
Corresponding authors: *ORCID:0000-0002-7361-9723,
xiangshuang@cib.ac.cn Received: 2020-05-11
Accepted: 2020-09-17
Online: 2020-10-20
Fund supported: National Natural Science Foundation of China (31370594 )National Key R&D Program of China (2017YFC0505000 )
摘要 亚热带常绿阔叶林植物幼树阶段适应林内生境并开枝散叶是其长成大树的一个重要过程, 植物一年内多次抽枝的现象及其在抽枝展叶过程中小枝伸长、枝茎增粗与叶面积的增加优先顺序及其内在驱动机制还有待进一步研究。该研究对青城山常绿阔叶林木本植物多次抽枝发生比例进行了调查, 并以茶(
Camellia sinensis )、细枝柃(
Eurya loquaiana )、短刺米槠(
Castanopsis carlesii var.
spinulosa )、润楠(
Machilus nanmu )和大叶山矾(
Symplocos grandis ) 5种植物的幼树为研究对象, 比较分析了植物在多次抽枝中小枝和叶片生长动态及适应策略的差异。结果显示: 1)一、二次抽枝分别开始于春季(4月)和夏末(8月下旬), 小枝水平上二次抽枝率乔木小于灌木, 常绿植物小于落叶植物。2)一次抽枝小枝枝长、单叶面积, 小枝直径和叶片数量(除大叶山矾外)均高于二次抽枝, 但二次抽枝单叶面积相对生长速率均高于一次抽枝, 二次抽枝叶片比叶质量(
LMA )的增长速率高于一次抽枝。3)一次抽枝小枝枝长、叶片数量、小枝直径(除细枝柃和短刺米槠外)和总叶面积(除短刺米槠外)最大相对生长速率均高于二次抽枝, 且大部分物种最大相对生长速率出现在抽枝开始的第一、二周。4)两次抽枝中, 物种先侧重于叶片的生长, 其次是小枝枝长的生长, 最后是小枝直径的增粗。单叶面积和总叶面积皆随着小枝枝长和小枝直径的增加呈显著的异速生长关系, 表明叶片的增长速度大于小枝。单叶面积与叶片数呈显著大于1的异速生长关系, 暗示单叶面积的增长速度大于叶片数的增加速度。小枝枝长与小枝直径也呈显著大于1的异速生长关系, 揭示小枝枝长的增长速度大于小枝直径。综上所述, 两次抽枝过程中, 植物枝叶的优先生长顺序反映了植物为获取更多的资源(尤其是光源)而形成特定的抽枝展叶策略; 二次抽枝单叶面积相对生长速率和
LMA 增长速率高于一次抽枝, 这可能与植物即将面临的昆虫取食和气温降低压力有关。因此, 了解植物抽枝策略对于理解物种生态适应机制, 揭示物种生活史过程中存在的权衡关系具有重要的理论意义。
关键词: 抽枝 ;
二次抽枝 ;
相对生长速率 ;
异速生长 ;
权衡 ;
亚热带常绿阔叶林 Abstract Aims Own to their genetic characteristics and long-term adaptation to the understory environment, the saplings of subtropical evergreen broad-leaved forest woody species could shoot twigs and develop new leaves several times a year, in which how they survive in the habitats and ultimately grow to adult trees are vital. The objective of this study was to examine what specific adaptation strategies have woody plants taken in the face of environmental pressure in subtropical evergreen broad-leaved forests, especially the growth priorities of twig elongation, stem thickening and leaf area increasing during the shooting process.
Methods The study site is located in Qingcheng Mountain scenic area, Sichuan Province. We chose five species with high second shooting rate in similar habits. Five plants with no disease and insect pests of each species were randomly selected for tree height measurement and long-term shooting observation. Each plant was randomly labeled with three young current-year twigs from different direction outside of tree canopy, and the twig length, twig diameter, leaf number and area were recorded when the twigs burst. The scaling relationships between traits of twig and leaf were analyzed by standardized major axis estimation (SMA).
Important findings 1) The first and second shootings started from spring (April) and late summer (the late August) respectively. The proportion of the second shooting of trees was lower than that of shrubs, and the proportion of second shooting of evergreen plants was lower than that of deciduous counterparts respectively. 2) The twig length and individual leaf area, the twig diameter and leaf number (expect for the
Symplocos grandis ) of the first shooting were all higher than those of the second shooting, but the relative growth rate of individual area and leaf mass per area of the first shooting were lower than those of the second shooting. 3) The maximum relative growth rates of the twig length and leaf number, the twig diameter (expect for
Eurya loquaiana and
Castanopsis carlesii var.
spinulosa ) and total leaf area (expect for
Castanopsis carlesii var.
spinulosa ) of the first shooting were all higher than those of the second shooting, and appeared in the first and second weeks of shooting. 4) In the two shootings, all the plants put priorities on the growth of leaf area and number, then on the growth of twig length, and finally on the twig diameter. The individual and total leaf area showed a significant allometric scaling relationship with the increasing of twig length and diameter, indicating that the growth rate of leaves was higher than that of twigs. The relationship between the individual leaf area and the leaf number was significantly greater than 1, suggesting that the growth rate of the individual leaf area was higher than that of the leaf number. Besides, the relationship between the twig length and diameter was also significantly greater than 1, suggesting that the growth rate of the twig length was higher than that of twig diameter. The study reveals the shooting strategies formed by plants improved their survival and reproductive ability by obtaining more resources (especially intercept the light resources).
Keywords: shooting ;
second shooting ;
relative growth rate ;
allometry ;
trade-off ;
subtropical evergreen broad-leaved forest PDF (1495KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 莫丹, 王振孟, 左有璐, 向双. 亚热带常绿阔叶林木本植物幼树阶段抽枝展叶的权衡关系. 植物生态学报, 2020, 44(10): 995-1006. DOI:
10.17521/cjpe.2020.0143 MO Dan, WANG Zhen-Meng, ZUO You-Lu, XIANG Shuang.
Trade-off between shooting and leaf developing of woody species saplings in subtropical evergreen broad-leaved forests .
Chinese Journal of Plant Ecology , 2020, 44(10): 995-1006. DOI:
10.17521/cjpe.2020.0143 抽枝是植物生长过程中普遍存在的现象, 一般指小芽结构抽出新枝条并展开幼叶片的动态过程。新枝条长出后继而伸长增粗, 达到一定营养条件时, 长出新的叶片(数量和面积的增加)。展叶过程包括出叶、展叶和叶的增厚等阶段(
Dale, 1982 )。以植物每年抽枝的次数为依据, 可将植物抽枝分为一次抽枝、二次抽枝、多次抽枝等抽枝类型。植物在抽枝过程中, 枝条和叶片会尽可能地在空间上权衡分布, 增加同化和吸收表面, 以充分利用外界有利资源, 达到提高自身营养物质储存的目的, 从而在适应外界环境的同时, 强大自身的繁殖能力(陆时万等, 2011)。而二次抽枝和多次抽枝使植物增加了叶片数量和面积, 为植物获取更多的光源, 积累更多的碳提供了有利条件, 是对一次抽枝期间碳损失进行的一种补偿(
Mizumachi et al ., 2006 ); 同时, 通过二次抽枝和多次抽枝, 植物可以积累更多的生物量以扩大冠层结构, 增加在群落中的生长竞争优势(
Hutchings & de Kroon, 1994 ;
Sultan, 2000 )。二次和多次抽枝是植物生长过程中响应多变的生境, 提高空间资源利用能力的一种生态策略(
Ozawa et al ., 2000 ;
史元春等, 2015 ;
龙嘉翼等, 2018 ), 也是对体内积累的生物量资源权衡分配的结果。因此, 研究植物抽枝策略对于揭示物种的生活史特征以及森林植物物种共存有着重要的理论价值(
Chabot & Hicks, 1982 )。
目前, 除了对二次抽枝现象及二次抽枝发生比例有很多观察报道外, 国内外关于木本植物抽枝展叶的研究主要集中在两次抽枝展叶动态的对比分析、抽枝展叶特征与昆虫取食和叶片形态的关系两方面(
夏洋洁等, 2013 ;
王瑞雪, 2015 ;
时盼, 2016 ), 但关于植物在两次抽枝过程中小枝和叶片间的异速生长关系、各功能部分先后生长顺序等生长策略与内在权衡仍缺乏研究报道。因此, 本文以四川省都江堰市青城山风景区亚热带常绿阔叶林为研究地点, 通过长期定位跟踪观测, 调查了常绿阔叶林下木本植物抽枝进程发生比例, 以具有典型二次抽枝现象的5种木本植物幼树为研究对象, 分析了每个物种在两次抽枝中的展叶动态、功能性状以及枝叶异速生长关系, 试图回答以下科学问题: 1)常绿阔叶林木本植物两次抽枝过程中小枝和叶片分别具有怎样的形态特征差异?2)两次抽枝过程中小枝和叶片相对生长速率的动态变化是否一致?3)抽枝期间叶片数量、叶片面积、小枝枝长和小枝直径四者之间是否存在权衡?通过对上述科学问题的研究, 试图揭示常绿阔叶林木本植物抽枝展叶所采取的对策, 理解植物在林下生境中忍耐、存活与生长的过程机制。
1 材料和方法 1.1 研究区概况 研究地区位于四川省都江堰市青城山风景区(30.90° N, 103.58° E, 海拔800-1 200 m), 该地区属于亚热带湿润气候, 全年温暖湿润, 年平均气温15.2 ℃, 最热月和最冷月的平均气温分别为24.7 ℃ (7月)和4.6 ℃ (1月); 年降水量约1 243.8 mm
, 平均相对湿度81%, 无霜期280天, 全年日照时间1 016.9 h。主要植被类型为亚热带常绿阔叶林和常绿落叶阔叶混交林, 暖性针叶林少有分布。生物多样性丰富, 根据《中国都江堰市植物名录》, 青城山景区内维管植物约346种, 其中蕨类植物51种, 种子植物295种(
马丹炜等, 2002 )。土壤类型为山地黄壤, 是亚热带温湿型气候条件下形成的地带性土壤, 母岩为侏罗纪紫色砂岩、泥岩和砾岩的坡积物。
1.2 样地植被与组成 具体研究地点位于“天然图画”景点附近, 海拔900 m, 坡度32°, 坡向为149°。植被类型为以樟科、山茶科、山矾科和壳斗科为主的典型亚热带常绿阔叶林, 郁闭度在0.9左右。植物种类丰富, 其中灌木层物种多样性最高, 乔木层次之, 草本层最低。群落水平结构上呈现斑块状镶嵌分布格局, 垂直结构上植被分层现象明显, 乔木层主要以楠木(
Phoebe zhennan )、润楠(
Machilus nanmu )、栲(
Castanopsis fargesii )、杉木(
Cunninghamia lanceolata )等植物为主; 灌木层以山茶科的柃木属(
Eurya )、山茶属(
Camellia )、山矾属(
Symplocos )植物为主; 草本层主要由鸢尾科的扁竹兰(
Iris confusa )和一些蕨类植物组成。
1.3 研究方法 1.3.1 植物标记与测定
经过长期观察, 青城山常绿阔叶林部分木本植物每年发生两次抽枝, 一次抽枝和二次抽枝开始时间分别为4月和8月下旬。因此, 选择2017年3月底以及2017年8月初在林内人为干扰较少且植物群落典型的地点, 对随机遇到的植物做抽枝调查, 并计算小枝水平上的二次抽枝率。结合样地内物种二次抽枝现象, 选择二次抽枝率较高的茶(
Camellia sinensis )、细枝柃(
Eurya loquaiana )、润楠、短刺米槠(
Castanopsis carlesii var.
spinulosa )以及大叶山矾(
Symplocos grandis ) 5种代表性植物进行研究, 于2017年3月中下旬植物爆芽之前在样地内相似的生境中每个物种随机选取5株长势良好且无病虫害的植株进行树高测量并长期观测。每株随机标记不同方向的3个当年生小枝, 从小枝爆芽开始记录小枝的长度、直径、叶片数量、叶片大小以及展叶时间。长度测量采用直尺和电子游标卡尺(IP54, 上海美耐特实业有限公司, 上海)。在进行叶片数量和大小观测时, 叶片长度达到0.50 cm时计入新叶数目并测量长宽。展叶早期每隔5-7天观测一次, 以后观测间隔逐渐拉长, 直到连续三次测量叶片面积无明显的增长为止, 第二次抽枝观测方式与第一次相同(
朱旭斌等, 2005 )。在抽枝过程中, 每次观测后采集相似生境中植株个体上的完好健康叶片, 每个物种叶片采集30枚左右。每个物种共选取200枚包含展叶过程各个阶段大小的叶片, 通过佳能Lide 300扫描仪(Canon, Hanoi, Vietnam)扫描叶片。
1.3.2 数据处理与分析
使用Image J软件计算扫描后的每个叶片长、宽及相对应的面积, 采用以下公式拟合各个物种的叶面积:
LA = α × LL × LW 。公式中
LA 为叶片面积,
LL 为叶片长,
LW 为叶片宽,
α 为物种相关的常数。将野外观测的叶片长宽数据进行转换, 所有观测物种方程拟合
R 2 > 0.90。增长量随时间的动态变化转化成百分比计算, 即:
At = at /a max 。
at 为时间
t 时小枝的单叶面积、枝长以及直径的增长量,
a max 为小枝总的增长量,
At 为在时间为
t 的时候小枝各性状增长量占其总增长量的比例。单叶面积相对生长速率用方程
A = A max / (1
+ exp(
β -
α × t ))求得, 其中
A 为叶面积,
A max 为最大单叶面积,
t 为儒略日(本文中指距离2017年1月1日的天数),
α 为相对生长速率,
β 为物种相关常数。定义展叶开始的时间为单叶面积达到最大单叶面积的10%, 记为
T (10) , 展叶结束的时间为单叶面积达到最大面积的90%, 记为
T (90) , 展叶持续时间为
T (90) 和
T (10) 的差值(
夏洋洁等, 2013 )。新生小枝枝长和直径及总叶面积相对生长速率(
RGR )的公式是:
RGR = (ln
L 1 - ln
L 2 )/(
T 1 -
T 2 )。公式中,
T 1 以及
T 2 分别是两次测定的时间,
L 1 和
L 2 分别是
T 1 和
T 2 时的枝长、直径和总叶面积; ln是自然对数(
Aanderud et al ., 2003 )。
所有性状数据均为3次重复的平均值(先求小枝内的平均值, 然后求个体内的平均值, 最后求物种内的平均值)。将物种平均值转化成百分比(同上述), 物种功能性状分析采用线性函数
y = ax + b ,
y 、
x 代表特定的性状,
b 为线性关系的截距(异速生长常数),
a 为线性关系的斜率(异速生长指数), 当
a = 1为等速生长,
a ≠ 1时为异速生长。对相关关系显著的功能性状进行标准化主轴分析(SMA), 由(S)MATR Version 2.0 (
Falster et al ., 2006 )完成。线性回归后, 对斜率进行同质性检验并计算共同斜率和截距差异(
Warton & Weber, 2002 )。用SPSS 21.0数据分析软件对每个物种第一次抽枝与第二次抽枝之间现叶时间、展叶速率、叶片数量、单叶大小、小枝长度、小枝直径等是否存在显著差异进行双样本
t 检验分析, 并用作图软件Origin 9.0完成图表制作。
2 结果 2.1 亚热带常绿阔叶林木本植物二次抽枝比例 经过前期长时间的观察, 发现青城山常绿阔叶林中大部分木本植物可发生一次抽枝, 而少部分木本植物有二次抽枝现象。因此, 在二次抽枝期间, 我们调查了408株个体, 涉及87个物种。其中包含乔木植物43种, 灌木植物44种; 常绿植物52种, 落叶植物35种。调查结果显示: 在小枝水平上, 不同生长型乔木植物二次抽枝率(20.97%)小于灌木植物(47.72%); 不同生活型常绿植物二次抽枝率(31.39%)小于落叶植物(37.39%)(
表1 )。
Table 1 表1 表1 青城山常绿阔叶林木本植物二次抽枝率
Table 1
Second shooting proportion of evergreen broad-leaved forest woody species of Qingcheng Mountain 生长型/生活型 个体数 物种数 二次抽枝率SE ) 乔木 Tree 198 43 20.97 ± 0.042a 灌木 Shrub 210 44 47.72 ± 0.053b 常绿 Evergreen 281 52 31.39 ± 0.044a 落叶 Deciduous 127 35 37.39 ± 0.062b
不同小写字母表示差异显著(p < 0.05)。 Different lowercase letters indicate significance difference at p < 0.05 level. 新窗口打开 |
下载CSV 2.2 亚热带常绿阔叶林木本植物抽枝展叶动态 第一次和第二次抽枝分别开始于4月和8月下旬, 相同抽枝批次不同物种间展叶开始时间差异较小。细枝柃和短刺米槠在两次抽枝中分别表现为出叶最早和最晚(
图1 )。茶和细枝柃在第一、二次抽枝持续时间上差异显著(
p < 0.05), 而大叶山矾、润楠、短刺米槠在两次抽枝持续时间上差异均不显著(
p > 0.05)(
表2 )。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1青城山常绿阔叶林木本植物第一、二次抽枝时间历程(平均值±标准误差)。 Fig. 1Specific duration of the first and second shooting of evergreen broad-leaved forest woody species of Qingcheng Mountain (mean ± SE ). Table 2 表2 表2 青城山常绿阔叶林木本植物第一 、二次抽枝时间、叶片大小和小枝大小的比较(平均值±标准误差)
Table 2
Comparison of timing, leaf size and twig size between the first and second shooting of evergreen broad-leaved forest woody species of Qingcheng Mountain (mean±SE) 最大树高数据参考于《中国植物志》(中国科学院中国植物志编辑委员会,1980 )。相同小写字母表示第一、二次抽枝之间差异不显著(p > 0.05);不同小写字母表示两者间差异显著(p < 0.05)。 The data in the column of maximum tree height are from Flora Reipublicae Popularis Sinicae (The Editorial Committee of Flora Reipubulicae Popularis Simicae, Chinese Academy of Sciences, 1980 ). The same lowercase letters indicate that there is no significant difference between the first and second shooting (p > 0.05), different lowercase letters indicate significance at p < 0.05 level. 新窗口打开 |
下载CSV 一次抽枝小枝枝长、小枝直径(除大叶山矾外)均大于二次抽枝。其中, 茶、细枝柃和润楠小枝枝长差异在两次抽枝之间达到显著水平(
p < 0.05), 短刺米槠和大叶山矾差异不显著(
p > 0.05)。细枝柃、润楠以及短刺米槠小枝直径差异在两次抽枝之间达到显著水平(
p < 0.05), 而茶和大叶山矾差异不显著(
p > 0.05)。一次抽枝叶片数量(除大叶山矾外)大于二次抽枝, 润楠和短刺米槠叶片数量差异在两次抽枝中达到显著水平(
p < 0.05), 其他3个物种差异不显著(
p > 0.05)。5个物种中, 一次抽枝的单叶面积均显著高于二次抽枝(
p < 0.05), 而二次抽枝的单叶面积相对生长速率均高于一次抽枝, 其中细枝柃和短刺米槠达到显著水平(
p < 0.05), 短刺米槠在两次抽枝中单叶面积相对生长速率均表现为最大, 分别是0.233和0.330 cm
2 ·d
-1 (
表2 )。
一次抽枝展叶前期, 叶面积增长缓慢, 各物种比叶质量总体上均呈下降趋势(茶和细枝柃的比叶质量先上升后下降)并达到最小值; 而展叶中后期, 叶片逐渐成熟, 各物种比叶质量呈先快速升高而后轻微下降的趋势(润楠一直呈上升趋势)。整个抽枝过程中, 茶、细枝柃、润楠、短刺米槠和大叶山矾的最大比叶质量分别为: 61.909、61.639、64.156、73.538、88.290 g·m
-2 。二次抽枝的茶、润楠和大叶山矾的比叶质量总体呈升高趋势, 最大值分别为: 63.790、96.544、74.231 g·m
-2 ; 细枝柃和短刺米槠的比叶质量变化趋势相似, 升高后下降再轻微上升, 最大值分别为70.023和72.745 g·m
-2 (
图2 )。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2青城山常绿阔叶林木本植物第一、二次抽枝叶片比叶质量(LMA )的动态变化。 Fig. 2Dynamic changes of newly developed leaf mass per area (LMA ) of the first and second shooting of evergreen broad-leaved forest woody species of Qingcheng Mountain. 2.3 亚热带常绿阔叶林木本植物小枝和叶片生长速率 二次抽枝小枝枝长最大相对生长速率小于一次抽枝。一次抽枝过程中, 茶、细枝柃和短刺米槠的相对生长速率在抽枝发生的第二周(4月14日)达到最大, 分别为0.090、0.078和0.113 cm·d
-1 , 而润楠和大叶山矾枝长最大相对生长速率分别与细枝柃和短刺米槠相同, 但出现的时间在抽枝开始的第一周(4月6日)且随抽枝发生时间的增加而下降; 二次抽枝过程中, 茶、细枝柃、润楠、短刺米槠和大叶山矾5个物种相对生长速率最大值(分别为: 0.036、0.066、0.043、0.100、0.051 cm·d
-1 )均出现在抽枝开始的第一周(8月29日)且均随时间的增加最终呈下降趋势(
图3A )。一次抽枝过程中, 短刺米槠小枝直径的相对生长速率最大值(0.023 cm·d
-1 )出现在抽枝开始的第六周左右(5月14日), 而茶、细枝柃、润楠和大叶山矾均在抽枝开始的第一周(4月6日)有最大值, 分别为: 0.034、0.007、0.036、0.054 cm·d
-1 ; 二次抽枝过程中, 5个物种直径的相对生长速率与二次抽枝中枝长的生长速率规律相同, 最大值分别为: 0.024、0.017、0.013、0.033、0.051 cm·d
-1 。两次抽枝相比, 除细枝柃和短刺米槠外, 所有物种一次抽枝直径的相对生长速率的最大值均高于二次抽枝, 且大叶山矾直径的相对生长速率最大值高于其他物种(
图3B )。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3青城山常绿阔叶林木本植物第一、二次抽枝小枝各构件的相对生长速率。 Fig. 3Relative growth rate of twig components for both the first and second shooting of evergreen broad-leaved forest woody species of Qingcheng Mountain. 除短刺米槠外, 其他物种一次抽枝总叶面积最大相对生长速率大于二次抽枝。一次抽枝中, 茶、细枝柃、润楠和大叶山矾的相对生长速率在抽枝发生的第一周(4月6日)有最大值, 分别为0.500、0.140、0.270和0.155 cm
2 ·d
-1 , 而短刺米槠在第四周左右(4月29日)有最大相对生长速率0.200 cm
2 ·d
-1 。二次抽枝中, 5个物种相对生长速率最大值(分别为: 0.052、0.058、0.054、0.266和0.105 cm
2 ·d
-1 )均出现在抽枝开始的第一周(
图3C )。一次抽枝叶片数量相对生长速率最大值大于二次抽枝。除短刺米槠外, 茶、细枝柃、润楠和大叶山矾一次抽枝均在抽枝发生的第一周有相对生长速率最大值, 分别为0.457、0.091、0.014和0.084片·d
-1 , 而短刺米槠在第四周有最大相对生长速率0.141片·d
-1 。二次抽枝中, 短刺米槠(0.048片·d
-1 )和大叶山矾(0.078片·d
-1 )在抽枝的第一周相对生长速率达到最大, 茶、细枝柃、润楠则在抽枝的第二周有最大相对生长速率, 分别为: 0.004、0.003、0.001片·d
-1 (
图3D )。
2.4 木本植物抽枝与展叶间的权衡关系 第一次、第二次抽枝单叶面积和总叶面积与小枝长和小枝直径分别存在极显著的正相关关系, 单叶面积与叶片数、小枝长与小枝直径也存在极显著的正相关关系(
p < 0.001,
表3 )。
Table 3 表3 表3 青城山常绿阔叶林木本植物第一、二次抽枝间小枝功能特征的标准化主轴(SMA)分析
Table 3
Standardized major axis (SMA) regression analysis on the scaling relationships between the twig functional traits of the first (n = 50) and second (n = 35) shooting of evergreen broad-leaved forest woody species of Qingcheng Mountain y -x 抽枝批次 回归系数 p 分组斜率(置信区间)CI s) 截距(置信区间)CI s) 共同斜率(置信区间)CI s) p 0 截距 单叶面积-小枝长 I 0.741 <0.001 1.358 (1.172, 1.573) -0.455 (-0.619, -0.291) 1.347 (1.198, 1.515) 0.855 -0.446a II 0.679 <0.001 1.327 (1.087, 1.620) -0.309 (-0.545, -0.074) -0.326b 单叶面积-小枝直径 I 0.447 <0.001 1.930 (1.558, 2.390) -1.037 (-1.398, -0.676) 2.206 (1.942, 2.487) 0.136 -1.268a II 0.826 <0.001 2.342 (2.021, 2.713) -1.274 (-1.588, -0.960) -1.151b 单叶面积-叶片数 I 0.633 <0.001 1.334 (1.120, 1.589) -0.426 (-0.618, -0.234) 1.570 0.001 II 0.443 <0.001 2.317 (1.784, 3.010) -1.361 (-1.947, -0.775) 小枝长-小枝直径 I 0.645 <0.001 1.421 (1.196, 1.688) -0.429 (-0.642, -0.216) 1.554 (1.355, 1.778) 0.111 -0.540a II 0.653 <0.001 1.764 (1.434, 2.170) -0.727 (-1.061, -0.393) -0.537a 总叶面积-小枝长 I 0.720 <0.001 1.369 (1.175, 1.596) -0.488 (-0.660, -0.315) 1.350 (1.205, 1.512) 0.794 -0.473a II 0.761 <0.001 1.327 (1.117, 1.576) -0.312 (-0.515, -0.109) -0.332b 总叶面积-小枝直径 I 0.427 <0.001 1.945 (1.565, 2.419) -1.075 (-1.446, -0.704) 2.207 (1.937, 2.498) 0.168 -1.294a
不同小写字母表示第一、二次抽枝间差异显著(p < 0.05)。 Different lowercase letters indicate significance between the first and second shooting at p < 0.05 level. 新窗口打开 |
下载CSV 单叶面积与小枝长的共同斜率为1.347 (95%
CI = 1.198-1.515,
p = 0.855,
图4A ), 单叶面积与小枝直径的共同斜率为2.206 (95%
CI = 1.942-2.487,
p = 0.136,
图4B ); 总叶面积与小枝长的共同斜率为1.350 (95%
CI = 1.205-1.512,
p = 0.794), 总叶面积与小枝直径的共同斜率为2.207 (95%
CI = 1.937- 2.498,
p = 0.168)。四个共同斜率均显著大于1, 表明单叶面积和总叶面积皆随着小枝长和小枝直径的增加呈显著的异速生长关系, 在抽枝过程中, 叶片的增长速度大于小枝(
表3 ), 且一次抽枝单叶面积与小枝长和小枝直径的共同斜率的
y 轴截距大于二次抽枝, 表明在给定的小枝长和小枝直径时, 一次抽枝的单叶面积小于二次抽枝, 即二次抽枝叶片生长速率快于一次抽枝。
图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4青城山常绿阔叶林木本植物第一、二次抽枝小枝各构件属性间的异速生长关系。 Fig. 4Scaling relationships of twig component traits between the first and second shooting of evergreen broad-leaved forest woody species of Qingcheng Mountain. 单叶面积与叶片数的共同斜率为1.570 (
p = 0.001,
图4C ), 表明单叶面积与叶片数呈显著大于1 (
p < 0.01)的异速生长关系, 单叶面积的增长速度大于叶片数的增加速度; 且一次抽枝单叶面积与叶片数的共同斜率的
y 轴截距大于二次抽枝, 表明在给定的叶片数时, 一次抽枝的单叶面积大于二次抽枝。
小枝长与小枝直径的共同斜率为1.554 (95%
CI = 1.355-1.778,
p = 0.111,
图4D ), 表明小枝长与小枝直径呈显著大于1 (
p < 0.01)的异速生长关系, 小枝长的增加速度大于小枝直径。且一次抽枝小枝长与小枝直径的共同斜率的
y 轴截距等于二次抽枝, 表明在给定的小枝直径时小枝长生长速度, 一次抽枝与二次抽枝无显著差异。
3 讨论 3.1 小枝水平上抽枝展叶间的权衡关系 小枝是植物最有活力的构件, 可为叶片的着生提供节点, 行使物理支撑与传导水分和养分的功能, 在植物生长发育过程中起着至关重要的作用; 叶片一般着生在小枝上, 主要行使截获光源并进行光合作用的功能。植物为应对环境的异质性, 可以权衡枝和叶的生物量分配(
Xiang et al ., 2009 ;
Messier et al ., 2017 ), 由此产生一系列相对适合的资源投入分配模式。在枝和叶的投资权衡上, 植物能够找到最适的收益/代价关系, 并以最经济的方式将养分等资源分配给光合结构(
Dombroskie & Aarssen, 2012 )。植物枝叶并不一定是同时发育和生长, 资源也不一定被平均分配到枝叶上。抽枝展叶过程中, 植物的枝叶生长是否同步反映了植物生长过程中碳收支策略的差异(
Lohier et al ., 2014 )。应对不同的环境条件, 植物如何在各个器官间权衡分配光合产物暂无一致的结论。我们通过本研究, 发现抽枝展叶初期, 不但在小枝和叶片的生长对比中, 两次抽枝的叶片生长速度快于枝条生长速度; 在小枝和叶片的内部(枝长-直径, 叶片大小-数量)也存在这样的生长关系, 枝长生长快于枝径, 叶片大小生长快于叶片数量。表明两次抽枝过程中, 植物均优先侧重于资源获取器官——叶片大小的生长, 以实现短时期内叶片截获光能的最大效率并尽可能快地获得光合回报(
Meng et al ., 2015 )。而植物在小枝伸长过程中优先枝条伸长, 也是植物尽快拓展空间, 为最大化展叶提供支持的佐证, 同时也可能是植物为减少虫食的一种生长对策(
朱旭斌, 2005 )。通常小枝越长越易受风力、枝条的机械强度影响, 所以需要增粗枝径, 以加强机械抗力。小枝水平上的抽枝展叶权衡可能与物种个体发育阶段的不同和物种遗传特征有关, 这种权衡关系影响着植物抽枝展叶的分配模式, 进而影响着植物的结构和功能(
Yan et al ., 2013 )。小枝水平上枝叶生长动态方面, 小枝枝长、小枝直径和叶片数量相对生长速率一次抽枝高于二次抽枝, 而单叶面积相对生长速率和
LMA 的增长速率二次抽枝高于一次抽枝, 这与植物小枝经过权衡选择快速增加生长速率、减少叶片投资的生长方式有关。二次抽枝植物的快速生长伴随着大量的能量消耗导致小枝对叶片面积增长投入不足, 最终形成的叶片面积小于一次抽枝。因此, 小枝水平上抽枝展叶的权衡关系是植物对林下光资源有限生境下的适应策略, 可帮助我们理解特定生境和生活史阶段枝和叶的资源投资权衡关系。
3.2 影响二次抽枝发生的因素 通过前期对青城山常绿阔叶林木本植物抽枝现象进行调查, 发现一次抽枝率高于二次抽枝率, 除了与植物本身遗传特性相关外, 一次抽枝所处的适宜气候条件以及更长的生长时间也是重要影响因素。一次抽枝后, 部分植物能量耗损严重, 无法满足二次抽枝的需要(
Ralhan et al ., 1985 ), 外界环境因素如二次抽枝后气温很快降低, 降水大量减少, 以及昆虫的取食压力等(
Zvereva & Kozlov, 2006 )也会造成二次抽枝率减少。青城山地区为湿润性亚热带常绿阔叶林分布区, 气候温暖湿润, 特别是夏末秋初的温度在16-22 ℃左右, 平均湿度约为95% (
王振孟, 2018 ), 与春季抽枝阶段的气候相似。外界适宜环境条件加之植物自身碳积累量富余是部分物种发生二次抽枝的重要原因(
杨芳等, 2019 )。二次抽枝阶段也是植食性昆虫生长繁殖的好时机, 展叶期高营养和高水分的新生叶片使昆虫得以大量繁殖, 昆虫取食引起叶片组织损伤, 削弱植物的生长(
Weed et al ., 2011 ), 给植物二次抽枝造成了很大的压力。有研究表明, 出叶较早的新生叶片比出叶较晚的新生叶片具有更强的耐霜冻能力(
Vitasse et al ., 2014 ), 因此, 多数发生二次抽枝的植物选择快速完成抽枝展叶的策略。除此以外, 植物二次抽枝的发生与其生活型和生长型存在一定的联系。本研究指出不同生长型的植物二次抽枝率在小枝水平上灌木大于乔木, 这与
夏洋洁等(2013) 以及
时盼(2016) 的研究结果相同。气候环境条件, 尤其是气温对乔木层和灌木层物种抽枝出叶存在很大的影响, 而3个研究地区的气温差异较小, 夏洋洁等的研究地区年平均气温为16.2 ℃; 时盼的研究地区年平均气温为15.4 ℃; 而本研究地区年平均气温为15.2 ℃。另外, 林内灌木层光环境较弱, 光照面积小, 更多的灌木通过二次抽枝不断分叉, 侧向生长以促进小枝长度和叶片数量的增加, 扩大冠层深度, 进一步最大化地获取光资源促进碳的积累, 从而维持自身的生存繁衍。不同生活型的植物在小枝水平上落叶植物二次抽枝率大于常绿植物, 落叶植物比常绿植物具有更大的小枝长度、小枝直径和茎生物量, 且落叶植物小枝水平上支撑的总叶面积和单叶面积显著高于常绿植物(
Sun et al ., 2006 ), 这些均与落叶植物的高的二次抽枝率存在必然联系。相对而言, 落叶植物二次抽枝的发生是与其快速的叶片周转效率相关, 而常绿植物尤其是林内幼树阶段的二次抽枝则与其对耐阴环境的长期适应对策相关。
3.3 物种特性对抽枝策略的影响 本研究的5个物种为长期耐阴物种如灌木(茶、细枝柃)和乔木的幼树阶段(润楠、短刺米槠、大叶山矾)。研究所选的样本植物高度均在1-2 m范围内, 茶和细枝柃两种灌木在最适宜的生长环境条件下能达到的最大树高分别为6和10 m, 短刺米槠和大叶山矾最大树高也不超过20 m。因此, 本研究的物种幼树或成树阶段几乎均生长在林下层或中层, 树冠结构多呈由较大叶片构成的开阔型树冠, 叶片大多较平整且均匀地排列在小枝上以最大限度地捕获光资源。林窗数量少且面积小导致植物接受到的光照较弱, 长期以来就形成了较强的耐阴特征, 在林内光资源有限的逆境下这些物种能够进行多次抽枝增加碳获取是促进其生活史成功的重要因素。除大叶山矾外, 其余研究物种一次抽枝展叶持续时间、小枝直径和叶片数量均高于二次抽枝, 大叶山矾在两次抽枝中, 小枝直径和单叶面积均大于其他物种。这可能和大叶山矾本身的物种特性有关, 5个研究物种中大叶山矾植株个体最高, 获取资源的能力更强; 同时与
Corner (1949) 在70年前的发现一致, 不同物种较粗的当年生小枝通常具有较大的叶片, 这一观点也被广泛证实(
Westoby & Wright, 2003 ;
Sun et al ., 2006 )。大叶山矾的粗枝和大叶使其在叶片伸展方面更具有优势, 能截获更多的光资源(
Pickup et al ., 2005 )积累更多的碳, 同时叶片的大小反映了植物生长能力的强弱, 对其应对环境条件变化所采取的适应策略有着显著影响(
Conesa et al ., 2019 )。茶和细枝柃两种灌木植株个体较矮, 尤其是茶, 物种本身的遗传特性决定了其二次抽枝的高抽枝率, 两者
LMA 在两次抽枝中均小于润楠、短刺米槠和大叶山矾3种幼树阶段的乔木, 投入相同资源时, 茶和细枝柃的叶面积生长更大, 比起其他物种耐阴性也更强。除了短刺米槠和大叶山矾外, 其余3个研究物种二次抽枝最大
LMA 均大于一次抽枝, 原因可能是短刺米槠和大叶山矾幼树阶段对光资源的需求更加强烈, 而二次抽枝期间光照强度的减弱使得这两个物种的二次抽枝最大
LMA 小于一次抽枝; 茶、细枝柃和润楠在二次抽枝过程中通过快速增加
LMA , 促进叶片厚度和硬度的增长以抵抗植食性昆虫的大量取食,
LMA 较高的叶片含水量和N含量较低, 昆虫食用后会出现生长缓慢、患病甚至死亡的现象(
邹钟琳, 1980 ;
柳凤娟, 2010 ;
杨芳等, 2019 ), 这在一定程度上反映了植物应对昆虫取食所表现出的适应对策。因此, 植物抽枝策略的差异与物种特性, 如遗传特征、生活型、耐阴特性、植株高矮等息息相关, 植物通过形成不同的抽枝策略与环境变化相协调, 这对于提高外界资源的利用效率, 快速占领生态位, 扩大群落竞争优势有着重要意义。
致谢 感谢青城山风景区管理局、都江堰市林业局对本研究实验样地设置和维护给予大力支持, 本实验室何蕾蕾、魏雪等在实验观测与采样中给予协助!
附录I 青城山常绿阔叶林二次抽枝物种资料 Supplement I Data of species for the second shooting of evergreen broad-leaved forest in Qingcheng Mountain https://www.plant-ecology.com/fileup/1005-264X/PDF/cjpe.2020.0143-S1.pdf [1] Aanderud ZT Bledsoe CS Richards JH 2003 ). Contribution of relative growth rate to root foraging by annual and perennial grasses from California oak woodlandsOecologia, 424 -430 . DOI:
10.1007/s00442-003-1275-7 URL [本文引用: 1] [2] Calle Z Schlumpberger BO Piedrahita L Leftin A Hammer SA Tye A Borchert R 2010 ). Seasonal variation in daily insolation induces synchronous bud break and flowering in the tropicsTrees, 865 -877 . [3] Chabot BF Hicks DJ 1982 ). The ecology of leaf life spansAnnual Review of Ecology and Systematics, 229 -259 . [本文引用: 1] [4] Chmielewski FM R?tzer T 2001 ). Response of tree phenology to climate change across EuropeAgricultural and Forest Meteorology, 101 -112 . [5] Conesa Mà Mus M Galmés J 2019 ). Leaf size as a key determinant of contrasting growth patterns in closely related Limonium (Plumbaginaceae) speciesJournal of Plant Physiology, 152984 . DOI: 10.1016/j.jplph.2019.05.011 . [本文引用: 1] [6] Corner EJH 1949 ). The durian theory or the origin of the modern treeAnnals of Botany, 367 -414 . [本文引用: 1] [7] Dale JE 1982 ). The Growth of Leaves Edward Arnold Limited, London . 31 -41 . [本文引用: 1] [8] Doi H Gordo O Katano I 2008 ). Heterogeneous intra-annual climatic changes drive different phenological responses at two trophic levelsClimate Research, 181 -190 . [9] Dombroskie SL Aarssen LW 2012 ). The leaf size/number trade-off within species and within plants for woody angiospermsPlant Ecology and Evolution, 38 -45 . [本文引用: 1] [10] Falster DS Warton DI Wright IJ 2006 ). User’s Guide to SMATR: Standardised Major Axis Tests & Routines Version 2.0 URL [本文引用: 1] [11] Guo QX Chai J Qian F Zhang TD Liu WD Tao JP 2013 ). Leaf and stem traits of current-year twigs vary with different functional types of woody plantChinese Journal of Ecology, 1465 -1470 . [ 郭庆学 , 柴捷 , 钱凤 , 张腾达 , 刘文丹 , 陶建平 (2013 ). 不同木本植物功能型当年生小枝功能性状差异生态学杂志 , 32, 1465 -1470 .] [12] He WM Zhong ZC 2000 ). Morphological and growth responses of the climbing plant, Gynostemma pentaphyllum seedlings to varying light intensityActa Phytoecologica Sinica, 375 -378 . [ 何维明 , 钟章成 (2000 ). 攀援植物绞股蓝幼苗对光照强度的形态和生长反应植物生态学报 , 24, 375 -378 .] [13] Hutchings MJ de Kroon H 1994 ). Foraging in plants: the role of morphological plasticity in resource acquisitionAdvances in Ecological Research, 159 -238 . [本文引用: 1] [14] Koriba K 1958 ). On the periodicity of the tree-growth in the tropics, with reference to the mode of branching, the leaf-fall, and the formation of the resting budGardens Bulletin, 11 -81 . [15] le Hir R Leduc N Jeannette E Viemont JD Pelleschi-Travier S 2006 ). Variations in sucrose and ABA concentrations are concomitant with heteroblastic leaf shape changes in a rhythmically growing species (Quercus robur )Tree Physiology, 229 -238 . [16] Liu FJ Xiang S Yang XC Sun SC 2010 ). Comparison of leaf mass per area, photosynthetic capacity and chemical defense traits of four evergreen broad-leaved tree species under contrasting light conditionsChinese Journal of Applied and Environmental Biology, 462 -467 . [本文引用: 1] [ 柳凤娟 , 向双 , 阳小成 , 孙书存 (2010 ). 两种光照生境下4种常绿阔叶树的单位叶面积干重、光合能力与化学防御物质含量比较应用与环境生物学报 , 16, 462 -467 .] [本文引用: 1] [17] Lohier T Jabot F Meziane D Shipley B Reich PB Deffuant G 2014 ). Explaining ontogenetic shifts in root-shoot scaling with transient dynamicsAnnals of Botany, 513 -524 . [本文引用: 1] [18] Long JY Zhao YM Kong XQ Chen ZY Wang XS Zhao K Cao R Huang LS Lü J Cui Y Yu YL Xu CY 2018 ). Trade-offs between twig and leaf traits of ornamental shrubs grown in shadeActa Ecologica Sinica, 8022 -8030 . [本文引用: 1] [ 龙嘉翼 , 赵宇萌 , 孔祥琦 , 陈治羊 , 王秀松 , 赵凯 , 曹然 , 黄丽莎 , 吕娇 , 崔义 , 余玉磊 , 徐程扬 (2018 ). 观赏灌木小枝和叶性状在林下庇荫环境中的权衡关系生态学报 , 38, 8022 -8030 .] [本文引用: 1] [19] Lu E Tsai C Lin J Yang S 2012 ). Leaf emergence, shedding, and lifespan of dominant hardwood species in Chitou, central TaiwanBotanical Studies, 255 -264 . [20] Ma DW Zhang G Wang YH Wang DM 2002 ). Studies on species diversity of forest vegetation on Qingcheng MountainJournal of Sichuan University (Natural Science Edition )115 -123 . [本文引用: 1] [ 马丹炜 , 张果 , 王跃华 , 王道模 (2002 ). 青城山森林植被物种多样性的研究四川大学学报(自然科学版) , 39, 115 -123 .] [本文引用: 1] [21] Martinez KA Fridley JD 2018 ). Acclimation of leaf traits in seasonal light environments: Are non-native species more plastic?Journal of Ecology, 2019 -2030 . [22] Meng FQ Zhang GF Li XC Niklas KJ Sun SC 2015 ). Growth synchrony between leaves and stems during twig development differs among plant functional types of subtropical rainforest woody speciesTree Physiology, 621 -631 . [本文引用: 1] [23] Messier J Lechowicz MJ McGill BJ Violle C Enquist BJ 2017 ). Interspecific integration of trait dimensions at local scales: the plant phenotype as an integrated networkJournal of Ecology, 1775 -1790 . [本文引用: 1] [24] Mizumachi E Mori A Osawa N Akiyama R Tokuchi N 2006 ). Shoot development and extension of Quercus serrata saplings in response to insect damage and nutrient conditionsAnnals of Botany, 219 -226 . [本文引用: 1] [25] Moles AT Westoby M 2000 ). Do small leaves expand faster than large leaves, and do shorter expansion times reduce herbivore damage?Oikos, 517 -524 . [26] Nitta I Ohsawa M 1997 ). Leaf dynamics and shoot phenology of eleven warm-temperate evergreen broad-leaved trees near their northern limit in central JapanPlant Ecology, 71 -88 . [27] Ozawa H Itoh K Hori Y 2000 ). Shoot structure and dynamics of saplings and canopies of three deciduous broad-leaved trees of a coppice forest in central JapanTrees, 206 -214 . [本文引用: 1] [28] Pichon C Hladik A Hladik M Tarnaud L Roger E 2015 ). Leaf phenological patterns of trees, shrubs and lianas in a dry semi-deciduous forest of northwestern Madagascar: functional types and adaptive significanceRevue D Ecologie, 197 -212 . [29] Pickup M Westoby M Basden A 2005 ). Dry mass costs of deploying leaf area in relation to leaf sizeFunctional Ecology, 88 -97 . [本文引用: 1] [30] Ralhan PK Khanna RK Singh SP Singh JS 1985 ). Certain phenological characters of the shrub layer of Kumaun Himalayan forestsVegetatio, 113 -119 . [本文引用: 1] [31] Rathcke B Lacey EP 1985 ). Patterns of terrestrial plantsAnnual Review of Ecology and Systematics, 179 -214 . [32] Sultan SE 2000 ). Phenotypic plasticity for plant development, function and life historyTrends in Plant Science, 537 -542 . [本文引用: 1] [33] Shi P 2016 ). Proportion and Dynamic of the Second Shooting in Woody Species from Deciduous Broad-leaved Forest in Xianlin, Nanjing Master degree dissertation, Nanjing Normal University, Nanjing [本文引用: 2] [ 时盼 (2016 ). 南京仙林地区落叶阔叶林中木本植物二次抽枝的发生比例及动态研究硕士学位论文, 南京师范大学, 南京 .] [本文引用: 2] [34] Shi YC Zhao CZ Song QH Du J Chen J Wang JW 2015 ). Slope-related variations in twig and leaf traits of Robinia pseudoacacia in the northern mountains of LanzhouChinese Journal of Plant Ecology, 362 -370 . [本文引用: 1] [ 史元春 , 赵成章 , 宋清华 , 杜晶 , 陈静 , 王继伟 (2015 ). 兰州北山刺槐枝叶性状的坡向差异性植物生态学报 , 39, 362 -370 .] [本文引用: 1] [35] Sun M Tian K Zhang Y Wang H Guan DX Yue HT 2017 ). Research on leaf functional traits and their environmental adaptationPlant Science Journal, 940 -949 . [ 孙梅 , 田昆 , 张贇 , 王行 , 管东旭 , 岳海涛 (2017 ). 植物叶片功能性状及其环境适应研究植物科学学报 , 35, 940 -949 .] [36] Sun SC Jin DM Shi PL 2006 ). The leaf size-twig size spectrum of temperate woody species along an altitudinal gradient: an invariant allometric scaling relationshipAnnals of Botany, 97 -107 . [本文引用: 2] [37] The Editorial Committee of Flora Reipubulicae Popularis Sinicae (1980 ). Flora Reipubulicae Popularis Sinicae. [本文引用: 2] [ 中国科学院中国植物志编辑委员会 (1980 ). 中国植物志 . 科学出版社, 北京.] [本文引用: 2] [38] Vitasse Y Lenz A Hoch G K?rner C 2014 ). Earlier leaf-out rather than difference in freezing resistance puts juvenile trees at greater risk of damage than adult treesJournal of Ecology, 981 -988 . [本文引用: 1] [39] Volkenburgh EV 1999 ). Leaf expansion: an integrating plant behaviourPlant, Cell & Environment, 1463 -1473 . [40] Wang RX 2015 ). Leaf Population Dynamic of Main Plant Species from Deciduous Broad-leaved Forest in Xianlin, Nanjing. Master degree dissertation, Nanjing Normal University, Nanjing . [本文引用: 1] [ 王瑞雪 (2015 ). 南京仙林落叶阔叶林中主要植物的叶群体动态硕士学位论文, 南京师范大学, 南京 .] [本文引用: 1] [41] Wang ZM 2018 ). The Strategies of Woody Saplings Adapting to the Light and Temperature Environments in the Subtropical Evergreen Broad-leaved Forests. Master degree dissertation, Chengdu Institute of Biology, Chinese Academy of Science, Chengdu . [本文引用: 1] [ 王振孟 (2018 ). 亚热带常绿阔叶林木本植物幼树阶段光温适应对策研究硕士学位论文, 中国科学院成都生物研究所, 成都 .] [本文引用: 1] [42] Warton DI Weber NC 2002 ). Common slope tests for bivariate errors-in-variables modelsBiometrical Journal, 161 -174 . [本文引用: 1] [43] Weed AS Gassmann A Casagrande RA 2011 ). Effects of leaf and root herbivory by potential insect biological control agents on the performance of invasive Vincetoxicum sppBiological Control, 50 -58 . [本文引用: 1] [44] Westoby M Wright IJ 2003 ). The leaf size-twig size spectrum and its relationship to other important spectra of variation among speciesOecologia, 621 -628 . [本文引用: 1] [45] Xia YJ Tang JQ Zhang GF Huang C Meng FQ Sun SC 2013 ). First and second sets of shoots in five evergreen woody species from Tiantong National Forest Park of Zhejiang, ChinaChinese Journal of Plant Ecology, 220 -229 . DOI:
10.3724/SP.J.1258.2013.00022 URL [本文引用: 3] [ 夏洋洁 , 唐坚强 , 张光富 , 黄超 , 蒙凤群 , 孙书存 (2013 ). 浙江天童国家森林公园5种常绿阔叶植物的一次和二次抽枝进程植物生态学报 , 37, 220 -229 .] [本文引用: 3] [46] Xiang S Wu N Sun SC 2009 ). Within-twig biomass allocation in subtropical evergreen broad-leaved species along an altitudinal gradient: allometric scaling analysisTrees, 637 -647 . [本文引用: 1] [47] Xu GX Luo SX Guo QS Pei SX Shi ZM Zhu L Zhu NN 2014 ). Responses of leaf unfolding and flowering to climate change in 12 tropical evergreen broadleaf tree species in Jianfengling, Hainan IslandChinese Journal of Plant Ecology, 585 -598 . [ 许格希 , 罗水兴 , 郭泉水 , 裴顺祥 , 史作民 , 朱莉 , 朱妮妮 (2014 ). 海南岛尖峰岭12种热带常绿阔叶乔木展叶期与开花期对气候变化的响应植物生态学报 , 38, 585 -598 .] [48] Yan ER Wang XH Chang SX He FL 2013 ). Scaling relationships among twig size, leaf size and leafing intensity in a successional series of subtropical forestsTree Physiology, 609 -617 . [本文引用: 1] [49] Yang F Wang ZM Zhu DH Yang XC Xiang S 2019 ). Dynamic characteristics of non-structural carbohydrates in leaves of six woody plants from an evergreen broad-leaved forestChinese Journal of Applied and Environmental Biology, 1075 -1083 . [本文引用: 2] [ 杨芳 , 王振孟 , 朱大海 , 阳小成 , 向双 (2019 ). 常绿阔叶林林下6种木本植物叶片非结构性碳水化合物的动态特征应用与环境生物学报 , 25, 1075 -1083 .] [本文引用: 2] [50] Zhu XB Liu YM Sun SC 2005 ). Leaf expansion of the dominant woody species of three deciduous oak forests in Nanjing, East ChinaActa Phytoecologica Sinica, 128 -136 . [本文引用: 2] [ 朱旭斌 , 刘娅梅 , 孙书存 (2005 ). 南京地区落叶栎林主要木本植物的展叶动态研究植物生态学报 , 29, 128 -136 .] [本文引用: 2] [51] Zou ZL 1980 ). Insect Ecology. [本文引用: 1] [ 邹钟琳 (1980 ). 昆虫生态学 . 上海科学技术出版社, 上海.] [本文引用: 1] [52] Zvereva EL Kozlov MV 2006 ). Consequences of simultaneous elevation of carbon dioxide and temperature for plant-herbivore interactions: a meta analysisGlobal Change Biology, 27 -41 . [本文引用: 1] Contribution of relative growth rate to root foraging by annual and perennial grasses from California oak woodlands
1
2003
... 使用Image J软件计算扫描后的每个叶片长、宽及相对应的面积, 采用以下公式拟合各个物种的叶面积:
LA = α × LL × LW .公式中
LA 为叶片面积,
LL 为叶片长,
LW 为叶片宽,
α 为物种相关的常数.将野外观测的叶片长宽数据进行转换, 所有观测物种方程拟合
R 2 > 0.90.增长量随时间的动态变化转化成百分比计算, 即:
At = at /a max .
at 为时间
t 时小枝的单叶面积、枝长以及直径的增长量,
a max 为小枝总的增长量,
At 为在时间为
t 的时候小枝各性状增长量占其总增长量的比例.单叶面积相对生长速率用方程
A = A max / (1
+ exp(
β -
α × t ))求得, 其中
A 为叶面积,
A max 为最大单叶面积,
t 为儒略日(本文中指距离2017年1月1日的天数),
α 为相对生长速率,
β 为物种相关常数.定义展叶开始的时间为单叶面积达到最大单叶面积的10%, 记为
T (10) , 展叶结束的时间为单叶面积达到最大面积的90%, 记为
T (90) , 展叶持续时间为
T (90) 和
T (10) 的差值(
夏洋洁等, 2013 ).新生小枝枝长和直径及总叶面积相对生长速率(
RGR )的公式是:
RGR = (ln
L 1 - ln
L 2 )/(
T 1 -
T 2 ).公式中,
T 1 以及
T 2 分别是两次测定的时间,
L 1 和
L 2 分别是
T 1 和
T 2 时的枝长、直径和总叶面积; ln是自然对数(
Aanderud et al ., 2003 ). ...
Seasonal variation in daily insolation induces synchronous bud break and flowering in the tropics
2010
The ecology of leaf life spans
1
1982
... 抽枝是植物生长过程中普遍存在的现象, 一般指小芽结构抽出新枝条并展开幼叶片的动态过程.新枝条长出后继而伸长增粗, 达到一定营养条件时, 长出新的叶片(数量和面积的增加).展叶过程包括出叶、展叶和叶的增厚等阶段(
Dale, 1982 ).以植物每年抽枝的次数为依据, 可将植物抽枝分为一次抽枝、二次抽枝、多次抽枝等抽枝类型.植物在抽枝过程中, 枝条和叶片会尽可能地在空间上权衡分布, 增加同化和吸收表面, 以充分利用外界有利资源, 达到提高自身营养物质储存的目的, 从而在适应外界环境的同时, 强大自身的繁殖能力(陆时万等, 2011).而二次抽枝和多次抽枝使植物增加了叶片数量和面积, 为植物获取更多的光源, 积累更多的碳提供了有利条件, 是对一次抽枝期间碳损失进行的一种补偿(
Mizumachi et al ., 2006 ); 同时, 通过二次抽枝和多次抽枝, 植物可以积累更多的生物量以扩大冠层结构, 增加在群落中的生长竞争优势(
Hutchings & de Kroon, 1994 ;
Sultan, 2000 ).二次和多次抽枝是植物生长过程中响应多变的生境, 提高空间资源利用能力的一种生态策略(
Ozawa et al ., 2000 ;
史元春等, 2015 ;
龙嘉翼等, 2018 ), 也是对体内积累的生物量资源权衡分配的结果.因此, 研究植物抽枝策略对于揭示物种的生活史特征以及森林植物物种共存有着重要的理论价值(
Chabot & Hicks, 1982 ). ...
Response of tree phenology to climate change across Europe
2001
Leaf size as a key determinant of contrasting growth patterns in closely related
Limonium (Plumbaginaceae) species
1
2019
... 本研究的5个物种为长期耐阴物种如灌木(茶、细枝柃)和乔木的幼树阶段(润楠、短刺米槠、大叶山矾).研究所选的样本植物高度均在1-2 m范围内, 茶和细枝柃两种灌木在最适宜的生长环境条件下能达到的最大树高分别为6和10 m, 短刺米槠和大叶山矾最大树高也不超过20 m.因此, 本研究的物种幼树或成树阶段几乎均生长在林下层或中层, 树冠结构多呈由较大叶片构成的开阔型树冠, 叶片大多较平整且均匀地排列在小枝上以最大限度地捕获光资源.林窗数量少且面积小导致植物接受到的光照较弱, 长期以来就形成了较强的耐阴特征, 在林内光资源有限的逆境下这些物种能够进行多次抽枝增加碳获取是促进其生活史成功的重要因素.除大叶山矾外, 其余研究物种一次抽枝展叶持续时间、小枝直径和叶片数量均高于二次抽枝, 大叶山矾在两次抽枝中, 小枝直径和单叶面积均大于其他物种.这可能和大叶山矾本身的物种特性有关, 5个研究物种中大叶山矾植株个体最高, 获取资源的能力更强; 同时与
Corner (1949) 在70年前的发现一致, 不同物种较粗的当年生小枝通常具有较大的叶片, 这一观点也被广泛证实(
Westoby & Wright, 2003 ;
Sun et al ., 2006 ).大叶山矾的粗枝和大叶使其在叶片伸展方面更具有优势, 能截获更多的光资源(
Pickup et al ., 2005 )积累更多的碳, 同时叶片的大小反映了植物生长能力的强弱, 对其应对环境条件变化所采取的适应策略有着显著影响(
Conesa et al ., 2019 ).茶和细枝柃两种灌木植株个体较矮, 尤其是茶, 物种本身的遗传特性决定了其二次抽枝的高抽枝率, 两者
LMA 在两次抽枝中均小于润楠、短刺米槠和大叶山矾3种幼树阶段的乔木, 投入相同资源时, 茶和细枝柃的叶面积生长更大, 比起其他物种耐阴性也更强.除了短刺米槠和大叶山矾外, 其余3个研究物种二次抽枝最大
LMA 均大于一次抽枝, 原因可能是短刺米槠和大叶山矾幼树阶段对光资源的需求更加强烈, 而二次抽枝期间光照强度的减弱使得这两个物种的二次抽枝最大
LMA 小于一次抽枝; 茶、细枝柃和润楠在二次抽枝过程中通过快速增加
LMA , 促进叶片厚度和硬度的增长以抵抗植食性昆虫的大量取食,
LMA 较高的叶片含水量和N含量较低, 昆虫食用后会出现生长缓慢、患病甚至死亡的现象(
邹钟琳, 1980 ;
柳凤娟, 2010 ;
杨芳等, 2019 ), 这在一定程度上反映了植物应对昆虫取食所表现出的适应对策.因此, 植物抽枝策略的差异与物种特性, 如遗传特征、生活型、耐阴特性、植株高矮等息息相关, 植物通过形成不同的抽枝策略与环境变化相协调, 这对于提高外界资源的利用效率, 快速占领生态位, 扩大群落竞争优势有着重要意义. ...
The durian theory or the origin of the modern tree
1
1949
... 本研究的5个物种为长期耐阴物种如灌木(茶、细枝柃)和乔木的幼树阶段(润楠、短刺米槠、大叶山矾).研究所选的样本植物高度均在1-2 m范围内, 茶和细枝柃两种灌木在最适宜的生长环境条件下能达到的最大树高分别为6和10 m, 短刺米槠和大叶山矾最大树高也不超过20 m.因此, 本研究的物种幼树或成树阶段几乎均生长在林下层或中层, 树冠结构多呈由较大叶片构成的开阔型树冠, 叶片大多较平整且均匀地排列在小枝上以最大限度地捕获光资源.林窗数量少且面积小导致植物接受到的光照较弱, 长期以来就形成了较强的耐阴特征, 在林内光资源有限的逆境下这些物种能够进行多次抽枝增加碳获取是促进其生活史成功的重要因素.除大叶山矾外, 其余研究物种一次抽枝展叶持续时间、小枝直径和叶片数量均高于二次抽枝, 大叶山矾在两次抽枝中, 小枝直径和单叶面积均大于其他物种.这可能和大叶山矾本身的物种特性有关, 5个研究物种中大叶山矾植株个体最高, 获取资源的能力更强; 同时与
Corner (1949) 在70年前的发现一致, 不同物种较粗的当年生小枝通常具有较大的叶片, 这一观点也被广泛证实(
Westoby & Wright, 2003 ;
Sun et al ., 2006 ).大叶山矾的粗枝和大叶使其在叶片伸展方面更具有优势, 能截获更多的光资源(
Pickup et al ., 2005 )积累更多的碳, 同时叶片的大小反映了植物生长能力的强弱, 对其应对环境条件变化所采取的适应策略有着显著影响(
Conesa et al ., 2019 ).茶和细枝柃两种灌木植株个体较矮, 尤其是茶, 物种本身的遗传特性决定了其二次抽枝的高抽枝率, 两者
LMA 在两次抽枝中均小于润楠、短刺米槠和大叶山矾3种幼树阶段的乔木, 投入相同资源时, 茶和细枝柃的叶面积生长更大, 比起其他物种耐阴性也更强.除了短刺米槠和大叶山矾外, 其余3个研究物种二次抽枝最大
LMA 均大于一次抽枝, 原因可能是短刺米槠和大叶山矾幼树阶段对光资源的需求更加强烈, 而二次抽枝期间光照强度的减弱使得这两个物种的二次抽枝最大
LMA 小于一次抽枝; 茶、细枝柃和润楠在二次抽枝过程中通过快速增加
LMA , 促进叶片厚度和硬度的增长以抵抗植食性昆虫的大量取食,
LMA 较高的叶片含水量和N含量较低, 昆虫食用后会出现生长缓慢、患病甚至死亡的现象(
邹钟琳, 1980 ;
柳凤娟, 2010 ;
杨芳等, 2019 ), 这在一定程度上反映了植物应对昆虫取食所表现出的适应对策.因此, 植物抽枝策略的差异与物种特性, 如遗传特征、生活型、耐阴特性、植株高矮等息息相关, 植物通过形成不同的抽枝策略与环境变化相协调, 这对于提高外界资源的利用效率, 快速占领生态位, 扩大群落竞争优势有着重要意义. ...
The Growth of Leaves 1
1982
... 抽枝是植物生长过程中普遍存在的现象, 一般指小芽结构抽出新枝条并展开幼叶片的动态过程.新枝条长出后继而伸长增粗, 达到一定营养条件时, 长出新的叶片(数量和面积的增加).展叶过程包括出叶、展叶和叶的增厚等阶段(
Dale, 1982 ).以植物每年抽枝的次数为依据, 可将植物抽枝分为一次抽枝、二次抽枝、多次抽枝等抽枝类型.植物在抽枝过程中, 枝条和叶片会尽可能地在空间上权衡分布, 增加同化和吸收表面, 以充分利用外界有利资源, 达到提高自身营养物质储存的目的, 从而在适应外界环境的同时, 强大自身的繁殖能力(陆时万等, 2011).而二次抽枝和多次抽枝使植物增加了叶片数量和面积, 为植物获取更多的光源, 积累更多的碳提供了有利条件, 是对一次抽枝期间碳损失进行的一种补偿(
Mizumachi et al ., 2006 ); 同时, 通过二次抽枝和多次抽枝, 植物可以积累更多的生物量以扩大冠层结构, 增加在群落中的生长竞争优势(
Hutchings & de Kroon, 1994 ;
Sultan, 2000 ).二次和多次抽枝是植物生长过程中响应多变的生境, 提高空间资源利用能力的一种生态策略(
Ozawa et al ., 2000 ;
史元春等, 2015 ;
龙嘉翼等, 2018 ), 也是对体内积累的生物量资源权衡分配的结果.因此, 研究植物抽枝策略对于揭示物种的生活史特征以及森林植物物种共存有着重要的理论价值(
Chabot & Hicks, 1982 ). ...
Heterogeneous intra-annual climatic changes drive different phenological responses at two trophic levels
2008
The leaf size/number trade-off within species and within plants for woody angiosperms
1
2012
... 小枝是植物最有活力的构件, 可为叶片的着生提供节点, 行使物理支撑与传导水分和养分的功能, 在植物生长发育过程中起着至关重要的作用; 叶片一般着生在小枝上, 主要行使截获光源并进行光合作用的功能.植物为应对环境的异质性, 可以权衡枝和叶的生物量分配(
Xiang et al ., 2009 ;
Messier et al ., 2017 ), 由此产生一系列相对适合的资源投入分配模式.在枝和叶的投资权衡上, 植物能够找到最适的收益/代价关系, 并以最经济的方式将养分等资源分配给光合结构(
Dombroskie & Aarssen, 2012 ).植物枝叶并不一定是同时发育和生长, 资源也不一定被平均分配到枝叶上.抽枝展叶过程中, 植物的枝叶生长是否同步反映了植物生长过程中碳收支策略的差异(
Lohier et al ., 2014 ).应对不同的环境条件, 植物如何在各个器官间权衡分配光合产物暂无一致的结论.我们通过本研究, 发现抽枝展叶初期, 不但在小枝和叶片的生长对比中, 两次抽枝的叶片生长速度快于枝条生长速度; 在小枝和叶片的内部(枝长-直径, 叶片大小-数量)也存在这样的生长关系, 枝长生长快于枝径, 叶片大小生长快于叶片数量.表明两次抽枝过程中, 植物均优先侧重于资源获取器官——叶片大小的生长, 以实现短时期内叶片截获光能的最大效率并尽可能快地获得光合回报(
Meng et al ., 2015 ).而植物在小枝伸长过程中优先枝条伸长, 也是植物尽快拓展空间, 为最大化展叶提供支持的佐证, 同时也可能是植物为减少虫食的一种生长对策(
朱旭斌, 2005 ).通常小枝越长越易受风力、枝条的机械强度影响, 所以需要增粗枝径, 以加强机械抗力.小枝水平上的抽枝展叶权衡可能与物种个体发育阶段的不同和物种遗传特征有关, 这种权衡关系影响着植物抽枝展叶的分配模式, 进而影响着植物的结构和功能(
Yan et al ., 2013 ).小枝水平上枝叶生长动态方面, 小枝枝长、小枝直径和叶片数量相对生长速率一次抽枝高于二次抽枝, 而单叶面积相对生长速率和
LMA 的增长速率二次抽枝高于一次抽枝, 这与植物小枝经过权衡选择快速增加生长速率、减少叶片投资的生长方式有关.二次抽枝植物的快速生长伴随着大量的能量消耗导致小枝对叶片面积增长投入不足, 最终形成的叶片面积小于一次抽枝.因此, 小枝水平上抽枝展叶的权衡关系是植物对林下光资源有限生境下的适应策略, 可帮助我们理解特定生境和生活史阶段枝和叶的资源投资权衡关系. ...
1
2006
... 所有性状数据均为3次重复的平均值(先求小枝内的平均值, 然后求个体内的平均值, 最后求物种内的平均值).将物种平均值转化成百分比(同上述), 物种功能性状分析采用线性函数
y = ax + b ,
y 、
x 代表特定的性状,
b 为线性关系的截距(异速生长常数),
a 为线性关系的斜率(异速生长指数), 当
a = 1为等速生长,
a ≠ 1时为异速生长.对相关关系显著的功能性状进行标准化主轴分析(SMA), 由(S)MATR Version 2.0 (
Falster et al ., 2006 )完成.线性回归后, 对斜率进行同质性检验并计算共同斜率和截距差异(
Warton & Weber, 2002 ).用SPSS 21.0数据分析软件对每个物种第一次抽枝与第二次抽枝之间现叶时间、展叶速率、叶片数量、单叶大小、小枝长度、小枝直径等是否存在显著差异进行双样本
t 检验分析, 并用作图软件Origin 9.0完成图表制作. ...
不同木本植物功能型当年生小枝功能性状差异
2013
不同木本植物功能型当年生小枝功能性状差异
2013
攀援植物绞股蓝幼苗对光照强度的形态和生长反应
2000
攀援植物绞股蓝幼苗对光照强度的形态和生长反应
2000
Foraging in plants: the role of morphological plasticity in resource acquisition
1
1994
... 抽枝是植物生长过程中普遍存在的现象, 一般指小芽结构抽出新枝条并展开幼叶片的动态过程.新枝条长出后继而伸长增粗, 达到一定营养条件时, 长出新的叶片(数量和面积的增加).展叶过程包括出叶、展叶和叶的增厚等阶段(
Dale, 1982 ).以植物每年抽枝的次数为依据, 可将植物抽枝分为一次抽枝、二次抽枝、多次抽枝等抽枝类型.植物在抽枝过程中, 枝条和叶片会尽可能地在空间上权衡分布, 增加同化和吸收表面, 以充分利用外界有利资源, 达到提高自身营养物质储存的目的, 从而在适应外界环境的同时, 强大自身的繁殖能力(陆时万等, 2011).而二次抽枝和多次抽枝使植物增加了叶片数量和面积, 为植物获取更多的光源, 积累更多的碳提供了有利条件, 是对一次抽枝期间碳损失进行的一种补偿(
Mizumachi et al ., 2006 ); 同时, 通过二次抽枝和多次抽枝, 植物可以积累更多的生物量以扩大冠层结构, 增加在群落中的生长竞争优势(
Hutchings & de Kroon, 1994 ;
Sultan, 2000 ).二次和多次抽枝是植物生长过程中响应多变的生境, 提高空间资源利用能力的一种生态策略(
Ozawa et al ., 2000 ;
史元春等, 2015 ;
龙嘉翼等, 2018 ), 也是对体内积累的生物量资源权衡分配的结果.因此, 研究植物抽枝策略对于揭示物种的生活史特征以及森林植物物种共存有着重要的理论价值(
Chabot & Hicks, 1982 ). ...
On the periodicity of the tree-growth in the tropics, with reference to the mode of branching, the leaf-fall, and the formation of the resting bud
1958
Variations in sucrose and ABA concentrations are concomitant with heteroblastic leaf shape changes in a rhythmically growing species (
Quercus robur )
2006
两种光照生境下4种常绿阔叶树的单位叶面积干重、光合能力与化学防御物质含量比较
1
2010
... 本研究的5个物种为长期耐阴物种如灌木(茶、细枝柃)和乔木的幼树阶段(润楠、短刺米槠、大叶山矾).研究所选的样本植物高度均在1-2 m范围内, 茶和细枝柃两种灌木在最适宜的生长环境条件下能达到的最大树高分别为6和10 m, 短刺米槠和大叶山矾最大树高也不超过20 m.因此, 本研究的物种幼树或成树阶段几乎均生长在林下层或中层, 树冠结构多呈由较大叶片构成的开阔型树冠, 叶片大多较平整且均匀地排列在小枝上以最大限度地捕获光资源.林窗数量少且面积小导致植物接受到的光照较弱, 长期以来就形成了较强的耐阴特征, 在林内光资源有限的逆境下这些物种能够进行多次抽枝增加碳获取是促进其生活史成功的重要因素.除大叶山矾外, 其余研究物种一次抽枝展叶持续时间、小枝直径和叶片数量均高于二次抽枝, 大叶山矾在两次抽枝中, 小枝直径和单叶面积均大于其他物种.这可能和大叶山矾本身的物种特性有关, 5个研究物种中大叶山矾植株个体最高, 获取资源的能力更强; 同时与
Corner (1949) 在70年前的发现一致, 不同物种较粗的当年生小枝通常具有较大的叶片, 这一观点也被广泛证实(
Westoby & Wright, 2003 ;
Sun et al ., 2006 ).大叶山矾的粗枝和大叶使其在叶片伸展方面更具有优势, 能截获更多的光资源(
Pickup et al ., 2005 )积累更多的碳, 同时叶片的大小反映了植物生长能力的强弱, 对其应对环境条件变化所采取的适应策略有着显著影响(
Conesa et al ., 2019 ).茶和细枝柃两种灌木植株个体较矮, 尤其是茶, 物种本身的遗传特性决定了其二次抽枝的高抽枝率, 两者
LMA 在两次抽枝中均小于润楠、短刺米槠和大叶山矾3种幼树阶段的乔木, 投入相同资源时, 茶和细枝柃的叶面积生长更大, 比起其他物种耐阴性也更强.除了短刺米槠和大叶山矾外, 其余3个研究物种二次抽枝最大
LMA 均大于一次抽枝, 原因可能是短刺米槠和大叶山矾幼树阶段对光资源的需求更加强烈, 而二次抽枝期间光照强度的减弱使得这两个物种的二次抽枝最大
LMA 小于一次抽枝; 茶、细枝柃和润楠在二次抽枝过程中通过快速增加
LMA , 促进叶片厚度和硬度的增长以抵抗植食性昆虫的大量取食,
LMA 较高的叶片含水量和N含量较低, 昆虫食用后会出现生长缓慢、患病甚至死亡的现象(
邹钟琳, 1980 ;
柳凤娟, 2010 ;
杨芳等, 2019 ), 这在一定程度上反映了植物应对昆虫取食所表现出的适应对策.因此, 植物抽枝策略的差异与物种特性, 如遗传特征、生活型、耐阴特性、植株高矮等息息相关, 植物通过形成不同的抽枝策略与环境变化相协调, 这对于提高外界资源的利用效率, 快速占领生态位, 扩大群落竞争优势有着重要意义. ...
两种光照生境下4种常绿阔叶树的单位叶面积干重、光合能力与化学防御物质含量比较
1
2010
... 本研究的5个物种为长期耐阴物种如灌木(茶、细枝柃)和乔木的幼树阶段(润楠、短刺米槠、大叶山矾).研究所选的样本植物高度均在1-2 m范围内, 茶和细枝柃两种灌木在最适宜的生长环境条件下能达到的最大树高分别为6和10 m, 短刺米槠和大叶山矾最大树高也不超过20 m.因此, 本研究的物种幼树或成树阶段几乎均生长在林下层或中层, 树冠结构多呈由较大叶片构成的开阔型树冠, 叶片大多较平整且均匀地排列在小枝上以最大限度地捕获光资源.林窗数量少且面积小导致植物接受到的光照较弱, 长期以来就形成了较强的耐阴特征, 在林内光资源有限的逆境下这些物种能够进行多次抽枝增加碳获取是促进其生活史成功的重要因素.除大叶山矾外, 其余研究物种一次抽枝展叶持续时间、小枝直径和叶片数量均高于二次抽枝, 大叶山矾在两次抽枝中, 小枝直径和单叶面积均大于其他物种.这可能和大叶山矾本身的物种特性有关, 5个研究物种中大叶山矾植株个体最高, 获取资源的能力更强; 同时与
Corner (1949) 在70年前的发现一致, 不同物种较粗的当年生小枝通常具有较大的叶片, 这一观点也被广泛证实(
Westoby & Wright, 2003 ;
Sun et al ., 2006 ).大叶山矾的粗枝和大叶使其在叶片伸展方面更具有优势, 能截获更多的光资源(
Pickup et al ., 2005 )积累更多的碳, 同时叶片的大小反映了植物生长能力的强弱, 对其应对环境条件变化所采取的适应策略有着显著影响(
Conesa et al ., 2019 ).茶和细枝柃两种灌木植株个体较矮, 尤其是茶, 物种本身的遗传特性决定了其二次抽枝的高抽枝率, 两者
LMA 在两次抽枝中均小于润楠、短刺米槠和大叶山矾3种幼树阶段的乔木, 投入相同资源时, 茶和细枝柃的叶面积生长更大, 比起其他物种耐阴性也更强.除了短刺米槠和大叶山矾外, 其余3个研究物种二次抽枝最大
LMA 均大于一次抽枝, 原因可能是短刺米槠和大叶山矾幼树阶段对光资源的需求更加强烈, 而二次抽枝期间光照强度的减弱使得这两个物种的二次抽枝最大
LMA 小于一次抽枝; 茶、细枝柃和润楠在二次抽枝过程中通过快速增加
LMA , 促进叶片厚度和硬度的增长以抵抗植食性昆虫的大量取食,
LMA 较高的叶片含水量和N含量较低, 昆虫食用后会出现生长缓慢、患病甚至死亡的现象(
邹钟琳, 1980 ;
柳凤娟, 2010 ;
杨芳等, 2019 ), 这在一定程度上反映了植物应对昆虫取食所表现出的适应对策.因此, 植物抽枝策略的差异与物种特性, 如遗传特征、生活型、耐阴特性、植株高矮等息息相关, 植物通过形成不同的抽枝策略与环境变化相协调, 这对于提高外界资源的利用效率, 快速占领生态位, 扩大群落竞争优势有着重要意义. ...
Explaining ontogenetic shifts in root-shoot scaling with transient dynamics
1
2014
... 小枝是植物最有活力的构件, 可为叶片的着生提供节点, 行使物理支撑与传导水分和养分的功能, 在植物生长发育过程中起着至关重要的作用; 叶片一般着生在小枝上, 主要行使截获光源并进行光合作用的功能.植物为应对环境的异质性, 可以权衡枝和叶的生物量分配(
Xiang et al ., 2009 ;
Messier et al ., 2017 ), 由此产生一系列相对适合的资源投入分配模式.在枝和叶的投资权衡上, 植物能够找到最适的收益/代价关系, 并以最经济的方式将养分等资源分配给光合结构(
Dombroskie & Aarssen, 2012 ).植物枝叶并不一定是同时发育和生长, 资源也不一定被平均分配到枝叶上.抽枝展叶过程中, 植物的枝叶生长是否同步反映了植物生长过程中碳收支策略的差异(
Lohier et al ., 2014 ).应对不同的环境条件, 植物如何在各个器官间权衡分配光合产物暂无一致的结论.我们通过本研究, 发现抽枝展叶初期, 不但在小枝和叶片的生长对比中, 两次抽枝的叶片生长速度快于枝条生长速度; 在小枝和叶片的内部(枝长-直径, 叶片大小-数量)也存在这样的生长关系, 枝长生长快于枝径, 叶片大小生长快于叶片数量.表明两次抽枝过程中, 植物均优先侧重于资源获取器官——叶片大小的生长, 以实现短时期内叶片截获光能的最大效率并尽可能快地获得光合回报(
Meng et al ., 2015 ).而植物在小枝伸长过程中优先枝条伸长, 也是植物尽快拓展空间, 为最大化展叶提供支持的佐证, 同时也可能是植物为减少虫食的一种生长对策(
朱旭斌, 2005 ).通常小枝越长越易受风力、枝条的机械强度影响, 所以需要增粗枝径, 以加强机械抗力.小枝水平上的抽枝展叶权衡可能与物种个体发育阶段的不同和物种遗传特征有关, 这种权衡关系影响着植物抽枝展叶的分配模式, 进而影响着植物的结构和功能(
Yan et al ., 2013 ).小枝水平上枝叶生长动态方面, 小枝枝长、小枝直径和叶片数量相对生长速率一次抽枝高于二次抽枝, 而单叶面积相对生长速率和
LMA 的增长速率二次抽枝高于一次抽枝, 这与植物小枝经过权衡选择快速增加生长速率、减少叶片投资的生长方式有关.二次抽枝植物的快速生长伴随着大量的能量消耗导致小枝对叶片面积增长投入不足, 最终形成的叶片面积小于一次抽枝.因此, 小枝水平上抽枝展叶的权衡关系是植物对林下光资源有限生境下的适应策略, 可帮助我们理解特定生境和生活史阶段枝和叶的资源投资权衡关系. ...
观赏灌木小枝和叶性状在林下庇荫环境中的权衡关系
1
2018
... 抽枝是植物生长过程中普遍存在的现象, 一般指小芽结构抽出新枝条并展开幼叶片的动态过程.新枝条长出后继而伸长增粗, 达到一定营养条件时, 长出新的叶片(数量和面积的增加).展叶过程包括出叶、展叶和叶的增厚等阶段(
Dale, 1982 ).以植物每年抽枝的次数为依据, 可将植物抽枝分为一次抽枝、二次抽枝、多次抽枝等抽枝类型.植物在抽枝过程中, 枝条和叶片会尽可能地在空间上权衡分布, 增加同化和吸收表面, 以充分利用外界有利资源, 达到提高自身营养物质储存的目的, 从而在适应外界环境的同时, 强大自身的繁殖能力(陆时万等, 2011).而二次抽枝和多次抽枝使植物增加了叶片数量和面积, 为植物获取更多的光源, 积累更多的碳提供了有利条件, 是对一次抽枝期间碳损失进行的一种补偿(
Mizumachi et al ., 2006 ); 同时, 通过二次抽枝和多次抽枝, 植物可以积累更多的生物量以扩大冠层结构, 增加在群落中的生长竞争优势(
Hutchings & de Kroon, 1994 ;
Sultan, 2000 ).二次和多次抽枝是植物生长过程中响应多变的生境, 提高空间资源利用能力的一种生态策略(
Ozawa et al ., 2000 ;
史元春等, 2015 ;
龙嘉翼等, 2018 ), 也是对体内积累的生物量资源权衡分配的结果.因此, 研究植物抽枝策略对于揭示物种的生活史特征以及森林植物物种共存有着重要的理论价值(
Chabot & Hicks, 1982 ). ...
观赏灌木小枝和叶性状在林下庇荫环境中的权衡关系
1
2018
... 抽枝是植物生长过程中普遍存在的现象, 一般指小芽结构抽出新枝条并展开幼叶片的动态过程.新枝条长出后继而伸长增粗, 达到一定营养条件时, 长出新的叶片(数量和面积的增加).展叶过程包括出叶、展叶和叶的增厚等阶段(
Dale, 1982 ).以植物每年抽枝的次数为依据, 可将植物抽枝分为一次抽枝、二次抽枝、多次抽枝等抽枝类型.植物在抽枝过程中, 枝条和叶片会尽可能地在空间上权衡分布, 增加同化和吸收表面, 以充分利用外界有利资源, 达到提高自身营养物质储存的目的, 从而在适应外界环境的同时, 强大自身的繁殖能力(陆时万等, 2011).而二次抽枝和多次抽枝使植物增加了叶片数量和面积, 为植物获取更多的光源, 积累更多的碳提供了有利条件, 是对一次抽枝期间碳损失进行的一种补偿(
Mizumachi et al ., 2006 ); 同时, 通过二次抽枝和多次抽枝, 植物可以积累更多的生物量以扩大冠层结构, 增加在群落中的生长竞争优势(
Hutchings & de Kroon, 1994 ;
Sultan, 2000 ).二次和多次抽枝是植物生长过程中响应多变的生境, 提高空间资源利用能力的一种生态策略(
Ozawa et al ., 2000 ;
史元春等, 2015 ;
龙嘉翼等, 2018 ), 也是对体内积累的生物量资源权衡分配的结果.因此, 研究植物抽枝策略对于揭示物种的生活史特征以及森林植物物种共存有着重要的理论价值(
Chabot & Hicks, 1982 ). ...
Leaf emergence, shedding, and lifespan of dominant hardwood species in Chitou, central Taiwan
2012
青城山森林植被物种多样性的研究
1
2002
... 研究地区位于四川省都江堰市青城山风景区(30.90° N, 103.58° E, 海拔800-1 200 m), 该地区属于亚热带湿润气候, 全年温暖湿润, 年平均气温15.2 ℃, 最热月和最冷月的平均气温分别为24.7 ℃ (7月)和4.6 ℃ (1月); 年降水量约1 243.8 mm
, 平均相对湿度81%, 无霜期280天, 全年日照时间1 016.9 h.主要植被类型为亚热带常绿阔叶林和常绿落叶阔叶混交林, 暖性针叶林少有分布.生物多样性丰富, 根据《中国都江堰市植物名录》, 青城山景区内维管植物约346种, 其中蕨类植物51种, 种子植物295种(
马丹炜等, 2002 ).土壤类型为山地黄壤, 是亚热带温湿型气候条件下形成的地带性土壤, 母岩为侏罗纪紫色砂岩、泥岩和砾岩的坡积物. ...
青城山森林植被物种多样性的研究
1
2002
... 研究地区位于四川省都江堰市青城山风景区(30.90° N, 103.58° E, 海拔800-1 200 m), 该地区属于亚热带湿润气候, 全年温暖湿润, 年平均气温15.2 ℃, 最热月和最冷月的平均气温分别为24.7 ℃ (7月)和4.6 ℃ (1月); 年降水量约1 243.8 mm
, 平均相对湿度81%, 无霜期280天, 全年日照时间1 016.9 h.主要植被类型为亚热带常绿阔叶林和常绿落叶阔叶混交林, 暖性针叶林少有分布.生物多样性丰富, 根据《中国都江堰市植物名录》, 青城山景区内维管植物约346种, 其中蕨类植物51种, 种子植物295种(
马丹炜等, 2002 ).土壤类型为山地黄壤, 是亚热带温湿型气候条件下形成的地带性土壤, 母岩为侏罗纪紫色砂岩、泥岩和砾岩的坡积物. ...
Acclimation of leaf traits in seasonal light environments: Are non-native species more plastic?
2018
Growth synchrony between leaves and stems during twig development differs among plant functional types of subtropical rainforest woody species
1
2015
... 小枝是植物最有活力的构件, 可为叶片的着生提供节点, 行使物理支撑与传导水分和养分的功能, 在植物生长发育过程中起着至关重要的作用; 叶片一般着生在小枝上, 主要行使截获光源并进行光合作用的功能.植物为应对环境的异质性, 可以权衡枝和叶的生物量分配(
Xiang et al ., 2009 ;
Messier et al ., 2017 ), 由此产生一系列相对适合的资源投入分配模式.在枝和叶的投资权衡上, 植物能够找到最适的收益/代价关系, 并以最经济的方式将养分等资源分配给光合结构(
Dombroskie & Aarssen, 2012 ).植物枝叶并不一定是同时发育和生长, 资源也不一定被平均分配到枝叶上.抽枝展叶过程中, 植物的枝叶生长是否同步反映了植物生长过程中碳收支策略的差异(
Lohier et al ., 2014 ).应对不同的环境条件, 植物如何在各个器官间权衡分配光合产物暂无一致的结论.我们通过本研究, 发现抽枝展叶初期, 不但在小枝和叶片的生长对比中, 两次抽枝的叶片生长速度快于枝条生长速度; 在小枝和叶片的内部(枝长-直径, 叶片大小-数量)也存在这样的生长关系, 枝长生长快于枝径, 叶片大小生长快于叶片数量.表明两次抽枝过程中, 植物均优先侧重于资源获取器官——叶片大小的生长, 以实现短时期内叶片截获光能的最大效率并尽可能快地获得光合回报(
Meng et al ., 2015 ).而植物在小枝伸长过程中优先枝条伸长, 也是植物尽快拓展空间, 为最大化展叶提供支持的佐证, 同时也可能是植物为减少虫食的一种生长对策(
朱旭斌, 2005 ).通常小枝越长越易受风力、枝条的机械强度影响, 所以需要增粗枝径, 以加强机械抗力.小枝水平上的抽枝展叶权衡可能与物种个体发育阶段的不同和物种遗传特征有关, 这种权衡关系影响着植物抽枝展叶的分配模式, 进而影响着植物的结构和功能(
Yan et al ., 2013 ).小枝水平上枝叶生长动态方面, 小枝枝长、小枝直径和叶片数量相对生长速率一次抽枝高于二次抽枝, 而单叶面积相对生长速率和
LMA 的增长速率二次抽枝高于一次抽枝, 这与植物小枝经过权衡选择快速增加生长速率、减少叶片投资的生长方式有关.二次抽枝植物的快速生长伴随着大量的能量消耗导致小枝对叶片面积增长投入不足, 最终形成的叶片面积小于一次抽枝.因此, 小枝水平上抽枝展叶的权衡关系是植物对林下光资源有限生境下的适应策略, 可帮助我们理解特定生境和生活史阶段枝和叶的资源投资权衡关系. ...
Interspecific integration of trait dimensions at local scales: the plant phenotype as an integrated network
1
2017
... 小枝是植物最有活力的构件, 可为叶片的着生提供节点, 行使物理支撑与传导水分和养分的功能, 在植物生长发育过程中起着至关重要的作用; 叶片一般着生在小枝上, 主要行使截获光源并进行光合作用的功能.植物为应对环境的异质性, 可以权衡枝和叶的生物量分配(
Xiang et al ., 2009 ;
Messier et al ., 2017 ), 由此产生一系列相对适合的资源投入分配模式.在枝和叶的投资权衡上, 植物能够找到最适的收益/代价关系, 并以最经济的方式将养分等资源分配给光合结构(
Dombroskie & Aarssen, 2012 ).植物枝叶并不一定是同时发育和生长, 资源也不一定被平均分配到枝叶上.抽枝展叶过程中, 植物的枝叶生长是否同步反映了植物生长过程中碳收支策略的差异(
Lohier et al ., 2014 ).应对不同的环境条件, 植物如何在各个器官间权衡分配光合产物暂无一致的结论.我们通过本研究, 发现抽枝展叶初期, 不但在小枝和叶片的生长对比中, 两次抽枝的叶片生长速度快于枝条生长速度; 在小枝和叶片的内部(枝长-直径, 叶片大小-数量)也存在这样的生长关系, 枝长生长快于枝径, 叶片大小生长快于叶片数量.表明两次抽枝过程中, 植物均优先侧重于资源获取器官——叶片大小的生长, 以实现短时期内叶片截获光能的最大效率并尽可能快地获得光合回报(
Meng et al ., 2015 ).而植物在小枝伸长过程中优先枝条伸长, 也是植物尽快拓展空间, 为最大化展叶提供支持的佐证, 同时也可能是植物为减少虫食的一种生长对策(
朱旭斌, 2005 ).通常小枝越长越易受风力、枝条的机械强度影响, 所以需要增粗枝径, 以加强机械抗力.小枝水平上的抽枝展叶权衡可能与物种个体发育阶段的不同和物种遗传特征有关, 这种权衡关系影响着植物抽枝展叶的分配模式, 进而影响着植物的结构和功能(
Yan et al ., 2013 ).小枝水平上枝叶生长动态方面, 小枝枝长、小枝直径和叶片数量相对生长速率一次抽枝高于二次抽枝, 而单叶面积相对生长速率和
LMA 的增长速率二次抽枝高于一次抽枝, 这与植物小枝经过权衡选择快速增加生长速率、减少叶片投资的生长方式有关.二次抽枝植物的快速生长伴随着大量的能量消耗导致小枝对叶片面积增长投入不足, 最终形成的叶片面积小于一次抽枝.因此, 小枝水平上抽枝展叶的权衡关系是植物对林下光资源有限生境下的适应策略, 可帮助我们理解特定生境和生活史阶段枝和叶的资源投资权衡关系. ...
Shoot development and extension of
Quercus serrata saplings in response to insect damage and nutrient conditions
1
2006
... 抽枝是植物生长过程中普遍存在的现象, 一般指小芽结构抽出新枝条并展开幼叶片的动态过程.新枝条长出后继而伸长增粗, 达到一定营养条件时, 长出新的叶片(数量和面积的增加).展叶过程包括出叶、展叶和叶的增厚等阶段(
Dale, 1982 ).以植物每年抽枝的次数为依据, 可将植物抽枝分为一次抽枝、二次抽枝、多次抽枝等抽枝类型.植物在抽枝过程中, 枝条和叶片会尽可能地在空间上权衡分布, 增加同化和吸收表面, 以充分利用外界有利资源, 达到提高自身营养物质储存的目的, 从而在适应外界环境的同时, 强大自身的繁殖能力(陆时万等, 2011).而二次抽枝和多次抽枝使植物增加了叶片数量和面积, 为植物获取更多的光源, 积累更多的碳提供了有利条件, 是对一次抽枝期间碳损失进行的一种补偿(
Mizumachi et al ., 2006 ); 同时, 通过二次抽枝和多次抽枝, 植物可以积累更多的生物量以扩大冠层结构, 增加在群落中的生长竞争优势(
Hutchings & de Kroon, 1994 ;
Sultan, 2000 ).二次和多次抽枝是植物生长过程中响应多变的生境, 提高空间资源利用能力的一种生态策略(
Ozawa et al ., 2000 ;
史元春等, 2015 ;
龙嘉翼等, 2018 ), 也是对体内积累的生物量资源权衡分配的结果.因此, 研究植物抽枝策略对于揭示物种的生活史特征以及森林植物物种共存有着重要的理论价值(
Chabot & Hicks, 1982 ). ...
Do small leaves expand faster than large leaves, and do shorter expansion times reduce herbivore damage?
2000
Leaf dynamics and shoot phenology of eleven warm-temperate evergreen broad-leaved trees near their northern limit in central Japan
1997
Shoot structure and dynamics of saplings and canopies of three deciduous broad-leaved trees of a coppice forest in central Japan
1
2000
... 抽枝是植物生长过程中普遍存在的现象, 一般指小芽结构抽出新枝条并展开幼叶片的动态过程.新枝条长出后继而伸长增粗, 达到一定营养条件时, 长出新的叶片(数量和面积的增加).展叶过程包括出叶、展叶和叶的增厚等阶段(
Dale, 1982 ).以植物每年抽枝的次数为依据, 可将植物抽枝分为一次抽枝、二次抽枝、多次抽枝等抽枝类型.植物在抽枝过程中, 枝条和叶片会尽可能地在空间上权衡分布, 增加同化和吸收表面, 以充分利用外界有利资源, 达到提高自身营养物质储存的目的, 从而在适应外界环境的同时, 强大自身的繁殖能力(陆时万等, 2011).而二次抽枝和多次抽枝使植物增加了叶片数量和面积, 为植物获取更多的光源, 积累更多的碳提供了有利条件, 是对一次抽枝期间碳损失进行的一种补偿(
Mizumachi et al ., 2006 ); 同时, 通过二次抽枝和多次抽枝, 植物可以积累更多的生物量以扩大冠层结构, 增加在群落中的生长竞争优势(
Hutchings & de Kroon, 1994 ;
Sultan, 2000 ).二次和多次抽枝是植物生长过程中响应多变的生境, 提高空间资源利用能力的一种生态策略(
Ozawa et al ., 2000 ;
史元春等, 2015 ;
龙嘉翼等, 2018 ), 也是对体内积累的生物量资源权衡分配的结果.因此, 研究植物抽枝策略对于揭示物种的生活史特征以及森林植物物种共存有着重要的理论价值(
Chabot & Hicks, 1982 ). ...
Leaf phenological patterns of trees, shrubs and lianas in a dry semi-deciduous forest of northwestern Madagascar: functional types and adaptive significance
2015
Dry mass costs of deploying leaf area in relation to leaf size
1
2005
... 本研究的5个物种为长期耐阴物种如灌木(茶、细枝柃)和乔木的幼树阶段(润楠、短刺米槠、大叶山矾).研究所选的样本植物高度均在1-2 m范围内, 茶和细枝柃两种灌木在最适宜的生长环境条件下能达到的最大树高分别为6和10 m, 短刺米槠和大叶山矾最大树高也不超过20 m.因此, 本研究的物种幼树或成树阶段几乎均生长在林下层或中层, 树冠结构多呈由较大叶片构成的开阔型树冠, 叶片大多较平整且均匀地排列在小枝上以最大限度地捕获光资源.林窗数量少且面积小导致植物接受到的光照较弱, 长期以来就形成了较强的耐阴特征, 在林内光资源有限的逆境下这些物种能够进行多次抽枝增加碳获取是促进其生活史成功的重要因素.除大叶山矾外, 其余研究物种一次抽枝展叶持续时间、小枝直径和叶片数量均高于二次抽枝, 大叶山矾在两次抽枝中, 小枝直径和单叶面积均大于其他物种.这可能和大叶山矾本身的物种特性有关, 5个研究物种中大叶山矾植株个体最高, 获取资源的能力更强; 同时与
Corner (1949) 在70年前的发现一致, 不同物种较粗的当年生小枝通常具有较大的叶片, 这一观点也被广泛证实(
Westoby & Wright, 2003 ;
Sun et al ., 2006 ).大叶山矾的粗枝和大叶使其在叶片伸展方面更具有优势, 能截获更多的光资源(
Pickup et al ., 2005 )积累更多的碳, 同时叶片的大小反映了植物生长能力的强弱, 对其应对环境条件变化所采取的适应策略有着显著影响(
Conesa et al ., 2019 ).茶和细枝柃两种灌木植株个体较矮, 尤其是茶, 物种本身的遗传特性决定了其二次抽枝的高抽枝率, 两者
LMA 在两次抽枝中均小于润楠、短刺米槠和大叶山矾3种幼树阶段的乔木, 投入相同资源时, 茶和细枝柃的叶面积生长更大, 比起其他物种耐阴性也更强.除了短刺米槠和大叶山矾外, 其余3个研究物种二次抽枝最大
LMA 均大于一次抽枝, 原因可能是短刺米槠和大叶山矾幼树阶段对光资源的需求更加强烈, 而二次抽枝期间光照强度的减弱使得这两个物种的二次抽枝最大
LMA 小于一次抽枝; 茶、细枝柃和润楠在二次抽枝过程中通过快速增加
LMA , 促进叶片厚度和硬度的增长以抵抗植食性昆虫的大量取食,
LMA 较高的叶片含水量和N含量较低, 昆虫食用后会出现生长缓慢、患病甚至死亡的现象(
邹钟琳, 1980 ;
柳凤娟, 2010 ;
杨芳等, 2019 ), 这在一定程度上反映了植物应对昆虫取食所表现出的适应对策.因此, 植物抽枝策略的差异与物种特性, 如遗传特征、生活型、耐阴特性、植株高矮等息息相关, 植物通过形成不同的抽枝策略与环境变化相协调, 这对于提高外界资源的利用效率, 快速占领生态位, 扩大群落竞争优势有着重要意义. ...
Certain phenological characters of the shrub layer of Kumaun Himalayan forests
1
1985
... 通过前期对青城山常绿阔叶林木本植物抽枝现象进行调查, 发现一次抽枝率高于二次抽枝率, 除了与植物本身遗传特性相关外, 一次抽枝所处的适宜气候条件以及更长的生长时间也是重要影响因素.一次抽枝后, 部分植物能量耗损严重, 无法满足二次抽枝的需要(
Ralhan et al ., 1985 ), 外界环境因素如二次抽枝后气温很快降低, 降水大量减少, 以及昆虫的取食压力等(
Zvereva & Kozlov, 2006 )也会造成二次抽枝率减少.青城山地区为湿润性亚热带常绿阔叶林分布区, 气候温暖湿润, 特别是夏末秋初的温度在16-22 ℃左右, 平均湿度约为95% (
王振孟, 2018 ), 与春季抽枝阶段的气候相似.外界适宜环境条件加之植物自身碳积累量富余是部分物种发生二次抽枝的重要原因(
杨芳等, 2019 ).二次抽枝阶段也是植食性昆虫生长繁殖的好时机, 展叶期高营养和高水分的新生叶片使昆虫得以大量繁殖, 昆虫取食引起叶片组织损伤, 削弱植物的生长(
Weed et al ., 2011 ), 给植物二次抽枝造成了很大的压力.有研究表明, 出叶较早的新生叶片比出叶较晚的新生叶片具有更强的耐霜冻能力(
Vitasse et al ., 2014 ), 因此, 多数发生二次抽枝的植物选择快速完成抽枝展叶的策略.除此以外, 植物二次抽枝的发生与其生活型和生长型存在一定的联系.本研究指出不同生长型的植物二次抽枝率在小枝水平上灌木大于乔木, 这与
夏洋洁等(2013) 以及
时盼(2016) 的研究结果相同.气候环境条件, 尤其是气温对乔木层和灌木层物种抽枝出叶存在很大的影响, 而3个研究地区的气温差异较小, 夏洋洁等的研究地区年平均气温为16.2 ℃; 时盼的研究地区年平均气温为15.4 ℃; 而本研究地区年平均气温为15.2 ℃.另外, 林内灌木层光环境较弱, 光照面积小, 更多的灌木通过二次抽枝不断分叉, 侧向生长以促进小枝长度和叶片数量的增加, 扩大冠层深度, 进一步最大化地获取光资源促进碳的积累, 从而维持自身的生存繁衍.不同生活型的植物在小枝水平上落叶植物二次抽枝率大于常绿植物, 落叶植物比常绿植物具有更大的小枝长度、小枝直径和茎生物量, 且落叶植物小枝水平上支撑的总叶面积和单叶面积显著高于常绿植物(
Sun et al ., 2006 ), 这些均与落叶植物的高的二次抽枝率存在必然联系.相对而言, 落叶植物二次抽枝的发生是与其快速的叶片周转效率相关, 而常绿植物尤其是林内幼树阶段的二次抽枝则与其对耐阴环境的长期适应对策相关. ...
Patterns of terrestrial plants
1985
Phenotypic plasticity for plant development, function and life history
1
2000
... 抽枝是植物生长过程中普遍存在的现象, 一般指小芽结构抽出新枝条并展开幼叶片的动态过程.新枝条长出后继而伸长增粗, 达到一定营养条件时, 长出新的叶片(数量和面积的增加).展叶过程包括出叶、展叶和叶的增厚等阶段(
Dale, 1982 ).以植物每年抽枝的次数为依据, 可将植物抽枝分为一次抽枝、二次抽枝、多次抽枝等抽枝类型.植物在抽枝过程中, 枝条和叶片会尽可能地在空间上权衡分布, 增加同化和吸收表面, 以充分利用外界有利资源, 达到提高自身营养物质储存的目的, 从而在适应外界环境的同时, 强大自身的繁殖能力(陆时万等, 2011).而二次抽枝和多次抽枝使植物增加了叶片数量和面积, 为植物获取更多的光源, 积累更多的碳提供了有利条件, 是对一次抽枝期间碳损失进行的一种补偿(
Mizumachi et al ., 2006 ); 同时, 通过二次抽枝和多次抽枝, 植物可以积累更多的生物量以扩大冠层结构, 增加在群落中的生长竞争优势(
Hutchings & de Kroon, 1994 ;
Sultan, 2000 ).二次和多次抽枝是植物生长过程中响应多变的生境, 提高空间资源利用能力的一种生态策略(
Ozawa et al ., 2000 ;
史元春等, 2015 ;
龙嘉翼等, 2018 ), 也是对体内积累的生物量资源权衡分配的结果.因此, 研究植物抽枝策略对于揭示物种的生活史特征以及森林植物物种共存有着重要的理论价值(
Chabot & Hicks, 1982 ). ...
南京仙林地区落叶阔叶林中木本植物二次抽枝的发生比例及动态研究
2
2016
... 目前, 除了对二次抽枝现象及二次抽枝发生比例有很多观察报道外, 国内外关于木本植物抽枝展叶的研究主要集中在两次抽枝展叶动态的对比分析、抽枝展叶特征与昆虫取食和叶片形态的关系两方面(
夏洋洁等, 2013 ;
王瑞雪, 2015 ;
时盼, 2016 ), 但关于植物在两次抽枝过程中小枝和叶片间的异速生长关系、各功能部分先后生长顺序等生长策略与内在权衡仍缺乏研究报道.因此, 本文以四川省都江堰市青城山风景区亚热带常绿阔叶林为研究地点, 通过长期定位跟踪观测, 调查了常绿阔叶林下木本植物抽枝进程发生比例, 以具有典型二次抽枝现象的5种木本植物幼树为研究对象, 分析了每个物种在两次抽枝中的展叶动态、功能性状以及枝叶异速生长关系, 试图回答以下科学问题: 1)常绿阔叶林木本植物两次抽枝过程中小枝和叶片分别具有怎样的形态特征差异?2)两次抽枝过程中小枝和叶片相对生长速率的动态变化是否一致?3)抽枝期间叶片数量、叶片面积、小枝枝长和小枝直径四者之间是否存在权衡?通过对上述科学问题的研究, 试图揭示常绿阔叶林木本植物抽枝展叶所采取的对策, 理解植物在林下生境中忍耐、存活与生长的过程机制. ...
... 通过前期对青城山常绿阔叶林木本植物抽枝现象进行调查, 发现一次抽枝率高于二次抽枝率, 除了与植物本身遗传特性相关外, 一次抽枝所处的适宜气候条件以及更长的生长时间也是重要影响因素.一次抽枝后, 部分植物能量耗损严重, 无法满足二次抽枝的需要(
Ralhan et al ., 1985 ), 外界环境因素如二次抽枝后气温很快降低, 降水大量减少, 以及昆虫的取食压力等(
Zvereva & Kozlov, 2006 )也会造成二次抽枝率减少.青城山地区为湿润性亚热带常绿阔叶林分布区, 气候温暖湿润, 特别是夏末秋初的温度在16-22 ℃左右, 平均湿度约为95% (
王振孟, 2018 ), 与春季抽枝阶段的气候相似.外界适宜环境条件加之植物自身碳积累量富余是部分物种发生二次抽枝的重要原因(
杨芳等, 2019 ).二次抽枝阶段也是植食性昆虫生长繁殖的好时机, 展叶期高营养和高水分的新生叶片使昆虫得以大量繁殖, 昆虫取食引起叶片组织损伤, 削弱植物的生长(
Weed et al ., 2011 ), 给植物二次抽枝造成了很大的压力.有研究表明, 出叶较早的新生叶片比出叶较晚的新生叶片具有更强的耐霜冻能力(
Vitasse et al ., 2014 ), 因此, 多数发生二次抽枝的植物选择快速完成抽枝展叶的策略.除此以外, 植物二次抽枝的发生与其生活型和生长型存在一定的联系.本研究指出不同生长型的植物二次抽枝率在小枝水平上灌木大于乔木, 这与
夏洋洁等(2013) 以及
时盼(2016) 的研究结果相同.气候环境条件, 尤其是气温对乔木层和灌木层物种抽枝出叶存在很大的影响, 而3个研究地区的气温差异较小, 夏洋洁等的研究地区年平均气温为16.2 ℃; 时盼的研究地区年平均气温为15.4 ℃; 而本研究地区年平均气温为15.2 ℃.另外, 林内灌木层光环境较弱, 光照面积小, 更多的灌木通过二次抽枝不断分叉, 侧向生长以促进小枝长度和叶片数量的增加, 扩大冠层深度, 进一步最大化地获取光资源促进碳的积累, 从而维持自身的生存繁衍.不同生活型的植物在小枝水平上落叶植物二次抽枝率大于常绿植物, 落叶植物比常绿植物具有更大的小枝长度、小枝直径和茎生物量, 且落叶植物小枝水平上支撑的总叶面积和单叶面积显著高于常绿植物(
Sun et al ., 2006 ), 这些均与落叶植物的高的二次抽枝率存在必然联系.相对而言, 落叶植物二次抽枝的发生是与其快速的叶片周转效率相关, 而常绿植物尤其是林内幼树阶段的二次抽枝则与其对耐阴环境的长期适应对策相关. ...
南京仙林地区落叶阔叶林中木本植物二次抽枝的发生比例及动态研究
2
2016
... 目前, 除了对二次抽枝现象及二次抽枝发生比例有很多观察报道外, 国内外关于木本植物抽枝展叶的研究主要集中在两次抽枝展叶动态的对比分析、抽枝展叶特征与昆虫取食和叶片形态的关系两方面(
夏洋洁等, 2013 ;
王瑞雪, 2015 ;
时盼, 2016 ), 但关于植物在两次抽枝过程中小枝和叶片间的异速生长关系、各功能部分先后生长顺序等生长策略与内在权衡仍缺乏研究报道.因此, 本文以四川省都江堰市青城山风景区亚热带常绿阔叶林为研究地点, 通过长期定位跟踪观测, 调查了常绿阔叶林下木本植物抽枝进程发生比例, 以具有典型二次抽枝现象的5种木本植物幼树为研究对象, 分析了每个物种在两次抽枝中的展叶动态、功能性状以及枝叶异速生长关系, 试图回答以下科学问题: 1)常绿阔叶林木本植物两次抽枝过程中小枝和叶片分别具有怎样的形态特征差异?2)两次抽枝过程中小枝和叶片相对生长速率的动态变化是否一致?3)抽枝期间叶片数量、叶片面积、小枝枝长和小枝直径四者之间是否存在权衡?通过对上述科学问题的研究, 试图揭示常绿阔叶林木本植物抽枝展叶所采取的对策, 理解植物在林下生境中忍耐、存活与生长的过程机制. ...
... 通过前期对青城山常绿阔叶林木本植物抽枝现象进行调查, 发现一次抽枝率高于二次抽枝率, 除了与植物本身遗传特性相关外, 一次抽枝所处的适宜气候条件以及更长的生长时间也是重要影响因素.一次抽枝后, 部分植物能量耗损严重, 无法满足二次抽枝的需要(
Ralhan et al ., 1985 ), 外界环境因素如二次抽枝后气温很快降低, 降水大量减少, 以及昆虫的取食压力等(
Zvereva & Kozlov, 2006 )也会造成二次抽枝率减少.青城山地区为湿润性亚热带常绿阔叶林分布区, 气候温暖湿润, 特别是夏末秋初的温度在16-22 ℃左右, 平均湿度约为95% (
王振孟, 2018 ), 与春季抽枝阶段的气候相似.外界适宜环境条件加之植物自身碳积累量富余是部分物种发生二次抽枝的重要原因(
杨芳等, 2019 ).二次抽枝阶段也是植食性昆虫生长繁殖的好时机, 展叶期高营养和高水分的新生叶片使昆虫得以大量繁殖, 昆虫取食引起叶片组织损伤, 削弱植物的生长(
Weed et al ., 2011 ), 给植物二次抽枝造成了很大的压力.有研究表明, 出叶较早的新生叶片比出叶较晚的新生叶片具有更强的耐霜冻能力(
Vitasse et al ., 2014 ), 因此, 多数发生二次抽枝的植物选择快速完成抽枝展叶的策略.除此以外, 植物二次抽枝的发生与其生活型和生长型存在一定的联系.本研究指出不同生长型的植物二次抽枝率在小枝水平上灌木大于乔木, 这与
夏洋洁等(2013) 以及
时盼(2016) 的研究结果相同.气候环境条件, 尤其是气温对乔木层和灌木层物种抽枝出叶存在很大的影响, 而3个研究地区的气温差异较小, 夏洋洁等的研究地区年平均气温为16.2 ℃; 时盼的研究地区年平均气温为15.4 ℃; 而本研究地区年平均气温为15.2 ℃.另外, 林内灌木层光环境较弱, 光照面积小, 更多的灌木通过二次抽枝不断分叉, 侧向生长以促进小枝长度和叶片数量的增加, 扩大冠层深度, 进一步最大化地获取光资源促进碳的积累, 从而维持自身的生存繁衍.不同生活型的植物在小枝水平上落叶植物二次抽枝率大于常绿植物, 落叶植物比常绿植物具有更大的小枝长度、小枝直径和茎生物量, 且落叶植物小枝水平上支撑的总叶面积和单叶面积显著高于常绿植物(
Sun et al ., 2006 ), 这些均与落叶植物的高的二次抽枝率存在必然联系.相对而言, 落叶植物二次抽枝的发生是与其快速的叶片周转效率相关, 而常绿植物尤其是林内幼树阶段的二次抽枝则与其对耐阴环境的长期适应对策相关. ...
兰州北山刺槐枝叶性状的坡向差异性
1
2015
... 抽枝是植物生长过程中普遍存在的现象, 一般指小芽结构抽出新枝条并展开幼叶片的动态过程.新枝条长出后继而伸长增粗, 达到一定营养条件时, 长出新的叶片(数量和面积的增加).展叶过程包括出叶、展叶和叶的增厚等阶段(
Dale, 1982 ).以植物每年抽枝的次数为依据, 可将植物抽枝分为一次抽枝、二次抽枝、多次抽枝等抽枝类型.植物在抽枝过程中, 枝条和叶片会尽可能地在空间上权衡分布, 增加同化和吸收表面, 以充分利用外界有利资源, 达到提高自身营养物质储存的目的, 从而在适应外界环境的同时, 强大自身的繁殖能力(陆时万等, 2011).而二次抽枝和多次抽枝使植物增加了叶片数量和面积, 为植物获取更多的光源, 积累更多的碳提供了有利条件, 是对一次抽枝期间碳损失进行的一种补偿(
Mizumachi et al ., 2006 ); 同时, 通过二次抽枝和多次抽枝, 植物可以积累更多的生物量以扩大冠层结构, 增加在群落中的生长竞争优势(
Hutchings & de Kroon, 1994 ;
Sultan, 2000 ).二次和多次抽枝是植物生长过程中响应多变的生境, 提高空间资源利用能力的一种生态策略(
Ozawa et al ., 2000 ;
史元春等, 2015 ;
龙嘉翼等, 2018 ), 也是对体内积累的生物量资源权衡分配的结果.因此, 研究植物抽枝策略对于揭示物种的生活史特征以及森林植物物种共存有着重要的理论价值(
Chabot & Hicks, 1982 ). ...
兰州北山刺槐枝叶性状的坡向差异性
1
2015
... 抽枝是植物生长过程中普遍存在的现象, 一般指小芽结构抽出新枝条并展开幼叶片的动态过程.新枝条长出后继而伸长增粗, 达到一定营养条件时, 长出新的叶片(数量和面积的增加).展叶过程包括出叶、展叶和叶的增厚等阶段(
Dale, 1982 ).以植物每年抽枝的次数为依据, 可将植物抽枝分为一次抽枝、二次抽枝、多次抽枝等抽枝类型.植物在抽枝过程中, 枝条和叶片会尽可能地在空间上权衡分布, 增加同化和吸收表面, 以充分利用外界有利资源, 达到提高自身营养物质储存的目的, 从而在适应外界环境的同时, 强大自身的繁殖能力(陆时万等, 2011).而二次抽枝和多次抽枝使植物增加了叶片数量和面积, 为植物获取更多的光源, 积累更多的碳提供了有利条件, 是对一次抽枝期间碳损失进行的一种补偿(
Mizumachi et al ., 2006 ); 同时, 通过二次抽枝和多次抽枝, 植物可以积累更多的生物量以扩大冠层结构, 增加在群落中的生长竞争优势(
Hutchings & de Kroon, 1994 ;
Sultan, 2000 ).二次和多次抽枝是植物生长过程中响应多变的生境, 提高空间资源利用能力的一种生态策略(
Ozawa et al ., 2000 ;
史元春等, 2015 ;
龙嘉翼等, 2018 ), 也是对体内积累的生物量资源权衡分配的结果.因此, 研究植物抽枝策略对于揭示物种的生活史特征以及森林植物物种共存有着重要的理论价值(
Chabot & Hicks, 1982 ). ...
植物叶片功能性状及其环境适应研究
2017
植物叶片功能性状及其环境适应研究
2017
The leaf size-twig size spectrum of temperate woody species along an altitudinal gradient: an invariant allometric scaling relationship
2
2006
... 通过前期对青城山常绿阔叶林木本植物抽枝现象进行调查, 发现一次抽枝率高于二次抽枝率, 除了与植物本身遗传特性相关外, 一次抽枝所处的适宜气候条件以及更长的生长时间也是重要影响因素.一次抽枝后, 部分植物能量耗损严重, 无法满足二次抽枝的需要(
Ralhan et al ., 1985 ), 外界环境因素如二次抽枝后气温很快降低, 降水大量减少, 以及昆虫的取食压力等(
Zvereva & Kozlov, 2006 )也会造成二次抽枝率减少.青城山地区为湿润性亚热带常绿阔叶林分布区, 气候温暖湿润, 特别是夏末秋初的温度在16-22 ℃左右, 平均湿度约为95% (
王振孟, 2018 ), 与春季抽枝阶段的气候相似.外界适宜环境条件加之植物自身碳积累量富余是部分物种发生二次抽枝的重要原因(
杨芳等, 2019 ).二次抽枝阶段也是植食性昆虫生长繁殖的好时机, 展叶期高营养和高水分的新生叶片使昆虫得以大量繁殖, 昆虫取食引起叶片组织损伤, 削弱植物的生长(
Weed et al ., 2011 ), 给植物二次抽枝造成了很大的压力.有研究表明, 出叶较早的新生叶片比出叶较晚的新生叶片具有更强的耐霜冻能力(
Vitasse et al ., 2014 ), 因此, 多数发生二次抽枝的植物选择快速完成抽枝展叶的策略.除此以外, 植物二次抽枝的发生与其生活型和生长型存在一定的联系.本研究指出不同生长型的植物二次抽枝率在小枝水平上灌木大于乔木, 这与
夏洋洁等(2013) 以及
时盼(2016) 的研究结果相同.气候环境条件, 尤其是气温对乔木层和灌木层物种抽枝出叶存在很大的影响, 而3个研究地区的气温差异较小, 夏洋洁等的研究地区年平均气温为16.2 ℃; 时盼的研究地区年平均气温为15.4 ℃; 而本研究地区年平均气温为15.2 ℃.另外, 林内灌木层光环境较弱, 光照面积小, 更多的灌木通过二次抽枝不断分叉, 侧向生长以促进小枝长度和叶片数量的增加, 扩大冠层深度, 进一步最大化地获取光资源促进碳的积累, 从而维持自身的生存繁衍.不同生活型的植物在小枝水平上落叶植物二次抽枝率大于常绿植物, 落叶植物比常绿植物具有更大的小枝长度、小枝直径和茎生物量, 且落叶植物小枝水平上支撑的总叶面积和单叶面积显著高于常绿植物(
Sun et al ., 2006 ), 这些均与落叶植物的高的二次抽枝率存在必然联系.相对而言, 落叶植物二次抽枝的发生是与其快速的叶片周转效率相关, 而常绿植物尤其是林内幼树阶段的二次抽枝则与其对耐阴环境的长期适应对策相关. ...
... 本研究的5个物种为长期耐阴物种如灌木(茶、细枝柃)和乔木的幼树阶段(润楠、短刺米槠、大叶山矾).研究所选的样本植物高度均在1-2 m范围内, 茶和细枝柃两种灌木在最适宜的生长环境条件下能达到的最大树高分别为6和10 m, 短刺米槠和大叶山矾最大树高也不超过20 m.因此, 本研究的物种幼树或成树阶段几乎均生长在林下层或中层, 树冠结构多呈由较大叶片构成的开阔型树冠, 叶片大多较平整且均匀地排列在小枝上以最大限度地捕获光资源.林窗数量少且面积小导致植物接受到的光照较弱, 长期以来就形成了较强的耐阴特征, 在林内光资源有限的逆境下这些物种能够进行多次抽枝增加碳获取是促进其生活史成功的重要因素.除大叶山矾外, 其余研究物种一次抽枝展叶持续时间、小枝直径和叶片数量均高于二次抽枝, 大叶山矾在两次抽枝中, 小枝直径和单叶面积均大于其他物种.这可能和大叶山矾本身的物种特性有关, 5个研究物种中大叶山矾植株个体最高, 获取资源的能力更强; 同时与
Corner (1949) 在70年前的发现一致, 不同物种较粗的当年生小枝通常具有较大的叶片, 这一观点也被广泛证实(
Westoby & Wright, 2003 ;
Sun et al ., 2006 ).大叶山矾的粗枝和大叶使其在叶片伸展方面更具有优势, 能截获更多的光资源(
Pickup et al ., 2005 )积累更多的碳, 同时叶片的大小反映了植物生长能力的强弱, 对其应对环境条件变化所采取的适应策略有着显著影响(
Conesa et al ., 2019 ).茶和细枝柃两种灌木植株个体较矮, 尤其是茶, 物种本身的遗传特性决定了其二次抽枝的高抽枝率, 两者
LMA 在两次抽枝中均小于润楠、短刺米槠和大叶山矾3种幼树阶段的乔木, 投入相同资源时, 茶和细枝柃的叶面积生长更大, 比起其他物种耐阴性也更强.除了短刺米槠和大叶山矾外, 其余3个研究物种二次抽枝最大
LMA 均大于一次抽枝, 原因可能是短刺米槠和大叶山矾幼树阶段对光资源的需求更加强烈, 而二次抽枝期间光照强度的减弱使得这两个物种的二次抽枝最大
LMA 小于一次抽枝; 茶、细枝柃和润楠在二次抽枝过程中通过快速增加
LMA , 促进叶片厚度和硬度的增长以抵抗植食性昆虫的大量取食,
LMA 较高的叶片含水量和N含量较低, 昆虫食用后会出现生长缓慢、患病甚至死亡的现象(
邹钟琳, 1980 ;
柳凤娟, 2010 ;
杨芳等, 2019 ), 这在一定程度上反映了植物应对昆虫取食所表现出的适应对策.因此, 植物抽枝策略的差异与物种特性, 如遗传特征、生活型、耐阴特性、植株高矮等息息相关, 植物通过形成不同的抽枝策略与环境变化相协调, 这对于提高外界资源的利用效率, 快速占领生态位, 扩大群落竞争优势有着重要意义. ...
2
1980
... 最大树高数据参考于《中国植物志》(
中国科学院中国植物志编辑委员会,1980 ).相同小写字母表示第一、二次抽枝之间差异不显著(p > 0.05);不同小写字母表示两者间差异显著(p < 0.05). ...
... The data in the column of maximum tree height are from
Flora Reipublicae Popularis Sinicae (
The Editorial Committee of Flora Reipubulicae Popularis Simicae, Chinese Academy of Sciences, 1980 ). The same lowercase letters indicate that there is no significant difference between the first and second shooting (p > 0.05), different lowercase letters indicate significance at p < 0.05 level. ...
2
1980
... 最大树高数据参考于《中国植物志》(
中国科学院中国植物志编辑委员会,1980 ).相同小写字母表示第一、二次抽枝之间差异不显著(p > 0.05);不同小写字母表示两者间差异显著(p < 0.05). ...
... The data in the column of maximum tree height are from
Flora Reipublicae Popularis Sinicae (
The Editorial Committee of Flora Reipubulicae Popularis Simicae, Chinese Academy of Sciences, 1980 ). The same lowercase letters indicate that there is no significant difference between the first and second shooting (p > 0.05), different lowercase letters indicate significance at p < 0.05 level. ...
Earlier leaf-out rather than difference in freezing resistance puts juvenile trees at greater risk of damage than adult trees
1
2014
... 通过前期对青城山常绿阔叶林木本植物抽枝现象进行调查, 发现一次抽枝率高于二次抽枝率, 除了与植物本身遗传特性相关外, 一次抽枝所处的适宜气候条件以及更长的生长时间也是重要影响因素.一次抽枝后, 部分植物能量耗损严重, 无法满足二次抽枝的需要(
Ralhan et al ., 1985 ), 外界环境因素如二次抽枝后气温很快降低, 降水大量减少, 以及昆虫的取食压力等(
Zvereva & Kozlov, 2006 )也会造成二次抽枝率减少.青城山地区为湿润性亚热带常绿阔叶林分布区, 气候温暖湿润, 特别是夏末秋初的温度在16-22 ℃左右, 平均湿度约为95% (
王振孟, 2018 ), 与春季抽枝阶段的气候相似.外界适宜环境条件加之植物自身碳积累量富余是部分物种发生二次抽枝的重要原因(
杨芳等, 2019 ).二次抽枝阶段也是植食性昆虫生长繁殖的好时机, 展叶期高营养和高水分的新生叶片使昆虫得以大量繁殖, 昆虫取食引起叶片组织损伤, 削弱植物的生长(
Weed et al ., 2011 ), 给植物二次抽枝造成了很大的压力.有研究表明, 出叶较早的新生叶片比出叶较晚的新生叶片具有更强的耐霜冻能力(
Vitasse et al ., 2014 ), 因此, 多数发生二次抽枝的植物选择快速完成抽枝展叶的策略.除此以外, 植物二次抽枝的发生与其生活型和生长型存在一定的联系.本研究指出不同生长型的植物二次抽枝率在小枝水平上灌木大于乔木, 这与
夏洋洁等(2013) 以及
时盼(2016) 的研究结果相同.气候环境条件, 尤其是气温对乔木层和灌木层物种抽枝出叶存在很大的影响, 而3个研究地区的气温差异较小, 夏洋洁等的研究地区年平均气温为16.2 ℃; 时盼的研究地区年平均气温为15.4 ℃; 而本研究地区年平均气温为15.2 ℃.另外, 林内灌木层光环境较弱, 光照面积小, 更多的灌木通过二次抽枝不断分叉, 侧向生长以促进小枝长度和叶片数量的增加, 扩大冠层深度, 进一步最大化地获取光资源促进碳的积累, 从而维持自身的生存繁衍.不同生活型的植物在小枝水平上落叶植物二次抽枝率大于常绿植物, 落叶植物比常绿植物具有更大的小枝长度、小枝直径和茎生物量, 且落叶植物小枝水平上支撑的总叶面积和单叶面积显著高于常绿植物(
Sun et al ., 2006 ), 这些均与落叶植物的高的二次抽枝率存在必然联系.相对而言, 落叶植物二次抽枝的发生是与其快速的叶片周转效率相关, 而常绿植物尤其是林内幼树阶段的二次抽枝则与其对耐阴环境的长期适应对策相关. ...
Leaf expansion: an integrating plant behaviour
1999
南京仙林落叶阔叶林中主要植物的叶群体动态
1
2015
... 目前, 除了对二次抽枝现象及二次抽枝发生比例有很多观察报道外, 国内外关于木本植物抽枝展叶的研究主要集中在两次抽枝展叶动态的对比分析、抽枝展叶特征与昆虫取食和叶片形态的关系两方面(
夏洋洁等, 2013 ;
王瑞雪, 2015 ;
时盼, 2016 ), 但关于植物在两次抽枝过程中小枝和叶片间的异速生长关系、各功能部分先后生长顺序等生长策略与内在权衡仍缺乏研究报道.因此, 本文以四川省都江堰市青城山风景区亚热带常绿阔叶林为研究地点, 通过长期定位跟踪观测, 调查了常绿阔叶林下木本植物抽枝进程发生比例, 以具有典型二次抽枝现象的5种木本植物幼树为研究对象, 分析了每个物种在两次抽枝中的展叶动态、功能性状以及枝叶异速生长关系, 试图回答以下科学问题: 1)常绿阔叶林木本植物两次抽枝过程中小枝和叶片分别具有怎样的形态特征差异?2)两次抽枝过程中小枝和叶片相对生长速率的动态变化是否一致?3)抽枝期间叶片数量、叶片面积、小枝枝长和小枝直径四者之间是否存在权衡?通过对上述科学问题的研究, 试图揭示常绿阔叶林木本植物抽枝展叶所采取的对策, 理解植物在林下生境中忍耐、存活与生长的过程机制. ...
南京仙林落叶阔叶林中主要植物的叶群体动态
1
2015
... 目前, 除了对二次抽枝现象及二次抽枝发生比例有很多观察报道外, 国内外关于木本植物抽枝展叶的研究主要集中在两次抽枝展叶动态的对比分析、抽枝展叶特征与昆虫取食和叶片形态的关系两方面(
夏洋洁等, 2013 ;
王瑞雪, 2015 ;
时盼, 2016 ), 但关于植物在两次抽枝过程中小枝和叶片间的异速生长关系、各功能部分先后生长顺序等生长策略与内在权衡仍缺乏研究报道.因此, 本文以四川省都江堰市青城山风景区亚热带常绿阔叶林为研究地点, 通过长期定位跟踪观测, 调查了常绿阔叶林下木本植物抽枝进程发生比例, 以具有典型二次抽枝现象的5种木本植物幼树为研究对象, 分析了每个物种在两次抽枝中的展叶动态、功能性状以及枝叶异速生长关系, 试图回答以下科学问题: 1)常绿阔叶林木本植物两次抽枝过程中小枝和叶片分别具有怎样的形态特征差异?2)两次抽枝过程中小枝和叶片相对生长速率的动态变化是否一致?3)抽枝期间叶片数量、叶片面积、小枝枝长和小枝直径四者之间是否存在权衡?通过对上述科学问题的研究, 试图揭示常绿阔叶林木本植物抽枝展叶所采取的对策, 理解植物在林下生境中忍耐、存活与生长的过程机制. ...
亚热带常绿阔叶林木本植物幼树阶段光温适应对策研究
1
2018
... 通过前期对青城山常绿阔叶林木本植物抽枝现象进行调查, 发现一次抽枝率高于二次抽枝率, 除了与植物本身遗传特性相关外, 一次抽枝所处的适宜气候条件以及更长的生长时间也是重要影响因素.一次抽枝后, 部分植物能量耗损严重, 无法满足二次抽枝的需要(
Ralhan et al ., 1985 ), 外界环境因素如二次抽枝后气温很快降低, 降水大量减少, 以及昆虫的取食压力等(
Zvereva & Kozlov, 2006 )也会造成二次抽枝率减少.青城山地区为湿润性亚热带常绿阔叶林分布区, 气候温暖湿润, 特别是夏末秋初的温度在16-22 ℃左右, 平均湿度约为95% (
王振孟, 2018 ), 与春季抽枝阶段的气候相似.外界适宜环境条件加之植物自身碳积累量富余是部分物种发生二次抽枝的重要原因(
杨芳等, 2019 ).二次抽枝阶段也是植食性昆虫生长繁殖的好时机, 展叶期高营养和高水分的新生叶片使昆虫得以大量繁殖, 昆虫取食引起叶片组织损伤, 削弱植物的生长(
Weed et al ., 2011 ), 给植物二次抽枝造成了很大的压力.有研究表明, 出叶较早的新生叶片比出叶较晚的新生叶片具有更强的耐霜冻能力(
Vitasse et al ., 2014 ), 因此, 多数发生二次抽枝的植物选择快速完成抽枝展叶的策略.除此以外, 植物二次抽枝的发生与其生活型和生长型存在一定的联系.本研究指出不同生长型的植物二次抽枝率在小枝水平上灌木大于乔木, 这与
夏洋洁等(2013) 以及
时盼(2016) 的研究结果相同.气候环境条件, 尤其是气温对乔木层和灌木层物种抽枝出叶存在很大的影响, 而3个研究地区的气温差异较小, 夏洋洁等的研究地区年平均气温为16.2 ℃; 时盼的研究地区年平均气温为15.4 ℃; 而本研究地区年平均气温为15.2 ℃.另外, 林内灌木层光环境较弱, 光照面积小, 更多的灌木通过二次抽枝不断分叉, 侧向生长以促进小枝长度和叶片数量的增加, 扩大冠层深度, 进一步最大化地获取光资源促进碳的积累, 从而维持自身的生存繁衍.不同生活型的植物在小枝水平上落叶植物二次抽枝率大于常绿植物, 落叶植物比常绿植物具有更大的小枝长度、小枝直径和茎生物量, 且落叶植物小枝水平上支撑的总叶面积和单叶面积显著高于常绿植物(
Sun et al ., 2006 ), 这些均与落叶植物的高的二次抽枝率存在必然联系.相对而言, 落叶植物二次抽枝的发生是与其快速的叶片周转效率相关, 而常绿植物尤其是林内幼树阶段的二次抽枝则与其对耐阴环境的长期适应对策相关. ...
亚热带常绿阔叶林木本植物幼树阶段光温适应对策研究
1
2018
... 通过前期对青城山常绿阔叶林木本植物抽枝现象进行调查, 发现一次抽枝率高于二次抽枝率, 除了与植物本身遗传特性相关外, 一次抽枝所处的适宜气候条件以及更长的生长时间也是重要影响因素.一次抽枝后, 部分植物能量耗损严重, 无法满足二次抽枝的需要(
Ralhan et al ., 1985 ), 外界环境因素如二次抽枝后气温很快降低, 降水大量减少, 以及昆虫的取食压力等(
Zvereva & Kozlov, 2006 )也会造成二次抽枝率减少.青城山地区为湿润性亚热带常绿阔叶林分布区, 气候温暖湿润, 特别是夏末秋初的温度在16-22 ℃左右, 平均湿度约为95% (
王振孟, 2018 ), 与春季抽枝阶段的气候相似.外界适宜环境条件加之植物自身碳积累量富余是部分物种发生二次抽枝的重要原因(
杨芳等, 2019 ).二次抽枝阶段也是植食性昆虫生长繁殖的好时机, 展叶期高营养和高水分的新生叶片使昆虫得以大量繁殖, 昆虫取食引起叶片组织损伤, 削弱植物的生长(
Weed et al ., 2011 ), 给植物二次抽枝造成了很大的压力.有研究表明, 出叶较早的新生叶片比出叶较晚的新生叶片具有更强的耐霜冻能力(
Vitasse et al ., 2014 ), 因此, 多数发生二次抽枝的植物选择快速完成抽枝展叶的策略.除此以外, 植物二次抽枝的发生与其生活型和生长型存在一定的联系.本研究指出不同生长型的植物二次抽枝率在小枝水平上灌木大于乔木, 这与
夏洋洁等(2013) 以及
时盼(2016) 的研究结果相同.气候环境条件, 尤其是气温对乔木层和灌木层物种抽枝出叶存在很大的影响, 而3个研究地区的气温差异较小, 夏洋洁等的研究地区年平均气温为16.2 ℃; 时盼的研究地区年平均气温为15.4 ℃; 而本研究地区年平均气温为15.2 ℃.另外, 林内灌木层光环境较弱, 光照面积小, 更多的灌木通过二次抽枝不断分叉, 侧向生长以促进小枝长度和叶片数量的增加, 扩大冠层深度, 进一步最大化地获取光资源促进碳的积累, 从而维持自身的生存繁衍.不同生活型的植物在小枝水平上落叶植物二次抽枝率大于常绿植物, 落叶植物比常绿植物具有更大的小枝长度、小枝直径和茎生物量, 且落叶植物小枝水平上支撑的总叶面积和单叶面积显著高于常绿植物(
Sun et al ., 2006 ), 这些均与落叶植物的高的二次抽枝率存在必然联系.相对而言, 落叶植物二次抽枝的发生是与其快速的叶片周转效率相关, 而常绿植物尤其是林内幼树阶段的二次抽枝则与其对耐阴环境的长期适应对策相关. ...
Common slope tests for bivariate errors-in-variables models
1
2002
... 所有性状数据均为3次重复的平均值(先求小枝内的平均值, 然后求个体内的平均值, 最后求物种内的平均值).将物种平均值转化成百分比(同上述), 物种功能性状分析采用线性函数
y = ax + b ,
y 、
x 代表特定的性状,
b 为线性关系的截距(异速生长常数),
a 为线性关系的斜率(异速生长指数), 当
a = 1为等速生长,
a ≠ 1时为异速生长.对相关关系显著的功能性状进行标准化主轴分析(SMA), 由(S)MATR Version 2.0 (
Falster et al ., 2006 )完成.线性回归后, 对斜率进行同质性检验并计算共同斜率和截距差异(
Warton & Weber, 2002 ).用SPSS 21.0数据分析软件对每个物种第一次抽枝与第二次抽枝之间现叶时间、展叶速率、叶片数量、单叶大小、小枝长度、小枝直径等是否存在显著差异进行双样本
t 检验分析, 并用作图软件Origin 9.0完成图表制作. ...
Effects of leaf and root herbivory by potential insect biological control agents on the performance of invasive
Vincetoxicum spp
1
2011
... 通过前期对青城山常绿阔叶林木本植物抽枝现象进行调查, 发现一次抽枝率高于二次抽枝率, 除了与植物本身遗传特性相关外, 一次抽枝所处的适宜气候条件以及更长的生长时间也是重要影响因素.一次抽枝后, 部分植物能量耗损严重, 无法满足二次抽枝的需要(
Ralhan et al ., 1985 ), 外界环境因素如二次抽枝后气温很快降低, 降水大量减少, 以及昆虫的取食压力等(
Zvereva & Kozlov, 2006 )也会造成二次抽枝率减少.青城山地区为湿润性亚热带常绿阔叶林分布区, 气候温暖湿润, 特别是夏末秋初的温度在16-22 ℃左右, 平均湿度约为95% (
王振孟, 2018 ), 与春季抽枝阶段的气候相似.外界适宜环境条件加之植物自身碳积累量富余是部分物种发生二次抽枝的重要原因(
杨芳等, 2019 ).二次抽枝阶段也是植食性昆虫生长繁殖的好时机, 展叶期高营养和高水分的新生叶片使昆虫得以大量繁殖, 昆虫取食引起叶片组织损伤, 削弱植物的生长(
Weed et al ., 2011 ), 给植物二次抽枝造成了很大的压力.有研究表明, 出叶较早的新生叶片比出叶较晚的新生叶片具有更强的耐霜冻能力(
Vitasse et al ., 2014 ), 因此, 多数发生二次抽枝的植物选择快速完成抽枝展叶的策略.除此以外, 植物二次抽枝的发生与其生活型和生长型存在一定的联系.本研究指出不同生长型的植物二次抽枝率在小枝水平上灌木大于乔木, 这与
夏洋洁等(2013) 以及
时盼(2016) 的研究结果相同.气候环境条件, 尤其是气温对乔木层和灌木层物种抽枝出叶存在很大的影响, 而3个研究地区的气温差异较小, 夏洋洁等的研究地区年平均气温为16.2 ℃; 时盼的研究地区年平均气温为15.4 ℃; 而本研究地区年平均气温为15.2 ℃.另外, 林内灌木层光环境较弱, 光照面积小, 更多的灌木通过二次抽枝不断分叉, 侧向生长以促进小枝长度和叶片数量的增加, 扩大冠层深度, 进一步最大化地获取光资源促进碳的积累, 从而维持自身的生存繁衍.不同生活型的植物在小枝水平上落叶植物二次抽枝率大于常绿植物, 落叶植物比常绿植物具有更大的小枝长度、小枝直径和茎生物量, 且落叶植物小枝水平上支撑的总叶面积和单叶面积显著高于常绿植物(
Sun et al ., 2006 ), 这些均与落叶植物的高的二次抽枝率存在必然联系.相对而言, 落叶植物二次抽枝的发生是与其快速的叶片周转效率相关, 而常绿植物尤其是林内幼树阶段的二次抽枝则与其对耐阴环境的长期适应对策相关. ...
The leaf size-twig size spectrum and its relationship to other important spectra of variation among species
1
2003
... 本研究的5个物种为长期耐阴物种如灌木(茶、细枝柃)和乔木的幼树阶段(润楠、短刺米槠、大叶山矾).研究所选的样本植物高度均在1-2 m范围内, 茶和细枝柃两种灌木在最适宜的生长环境条件下能达到的最大树高分别为6和10 m, 短刺米槠和大叶山矾最大树高也不超过20 m.因此, 本研究的物种幼树或成树阶段几乎均生长在林下层或中层, 树冠结构多呈由较大叶片构成的开阔型树冠, 叶片大多较平整且均匀地排列在小枝上以最大限度地捕获光资源.林窗数量少且面积小导致植物接受到的光照较弱, 长期以来就形成了较强的耐阴特征, 在林内光资源有限的逆境下这些物种能够进行多次抽枝增加碳获取是促进其生活史成功的重要因素.除大叶山矾外, 其余研究物种一次抽枝展叶持续时间、小枝直径和叶片数量均高于二次抽枝, 大叶山矾在两次抽枝中, 小枝直径和单叶面积均大于其他物种.这可能和大叶山矾本身的物种特性有关, 5个研究物种中大叶山矾植株个体最高, 获取资源的能力更强; 同时与
Corner (1949) 在70年前的发现一致, 不同物种较粗的当年生小枝通常具有较大的叶片, 这一观点也被广泛证实(
Westoby & Wright, 2003 ;
Sun et al ., 2006 ).大叶山矾的粗枝和大叶使其在叶片伸展方面更具有优势, 能截获更多的光资源(
Pickup et al ., 2005 )积累更多的碳, 同时叶片的大小反映了植物生长能力的强弱, 对其应对环境条件变化所采取的适应策略有着显著影响(
Conesa et al ., 2019 ).茶和细枝柃两种灌木植株个体较矮, 尤其是茶, 物种本身的遗传特性决定了其二次抽枝的高抽枝率, 两者
LMA 在两次抽枝中均小于润楠、短刺米槠和大叶山矾3种幼树阶段的乔木, 投入相同资源时, 茶和细枝柃的叶面积生长更大, 比起其他物种耐阴性也更强.除了短刺米槠和大叶山矾外, 其余3个研究物种二次抽枝最大
LMA 均大于一次抽枝, 原因可能是短刺米槠和大叶山矾幼树阶段对光资源的需求更加强烈, 而二次抽枝期间光照强度的减弱使得这两个物种的二次抽枝最大
LMA 小于一次抽枝; 茶、细枝柃和润楠在二次抽枝过程中通过快速增加
LMA , 促进叶片厚度和硬度的增长以抵抗植食性昆虫的大量取食,
LMA 较高的叶片含水量和N含量较低, 昆虫食用后会出现生长缓慢、患病甚至死亡的现象(
邹钟琳, 1980 ;
柳凤娟, 2010 ;
杨芳等, 2019 ), 这在一定程度上反映了植物应对昆虫取食所表现出的适应对策.因此, 植物抽枝策略的差异与物种特性, 如遗传特征、生活型、耐阴特性、植株高矮等息息相关, 植物通过形成不同的抽枝策略与环境变化相协调, 这对于提高外界资源的利用效率, 快速占领生态位, 扩大群落竞争优势有着重要意义. ...
浙江天童国家森林公园5种常绿阔叶植物的一次和二次抽枝进程
3
2013
... 目前, 除了对二次抽枝现象及二次抽枝发生比例有很多观察报道外, 国内外关于木本植物抽枝展叶的研究主要集中在两次抽枝展叶动态的对比分析、抽枝展叶特征与昆虫取食和叶片形态的关系两方面(
夏洋洁等, 2013 ;
王瑞雪, 2015 ;
时盼, 2016 ), 但关于植物在两次抽枝过程中小枝和叶片间的异速生长关系、各功能部分先后生长顺序等生长策略与内在权衡仍缺乏研究报道.因此, 本文以四川省都江堰市青城山风景区亚热带常绿阔叶林为研究地点, 通过长期定位跟踪观测, 调查了常绿阔叶林下木本植物抽枝进程发生比例, 以具有典型二次抽枝现象的5种木本植物幼树为研究对象, 分析了每个物种在两次抽枝中的展叶动态、功能性状以及枝叶异速生长关系, 试图回答以下科学问题: 1)常绿阔叶林木本植物两次抽枝过程中小枝和叶片分别具有怎样的形态特征差异?2)两次抽枝过程中小枝和叶片相对生长速率的动态变化是否一致?3)抽枝期间叶片数量、叶片面积、小枝枝长和小枝直径四者之间是否存在权衡?通过对上述科学问题的研究, 试图揭示常绿阔叶林木本植物抽枝展叶所采取的对策, 理解植物在林下生境中忍耐、存活与生长的过程机制. ...
... 使用Image J软件计算扫描后的每个叶片长、宽及相对应的面积, 采用以下公式拟合各个物种的叶面积:
LA = α × LL × LW .公式中
LA 为叶片面积,
LL 为叶片长,
LW 为叶片宽,
α 为物种相关的常数.将野外观测的叶片长宽数据进行转换, 所有观测物种方程拟合
R 2 > 0.90.增长量随时间的动态变化转化成百分比计算, 即:
At = at /a max .
at 为时间
t 时小枝的单叶面积、枝长以及直径的增长量,
a max 为小枝总的增长量,
At 为在时间为
t 的时候小枝各性状增长量占其总增长量的比例.单叶面积相对生长速率用方程
A = A max / (1
+ exp(
β -
α × t ))求得, 其中
A 为叶面积,
A max 为最大单叶面积,
t 为儒略日(本文中指距离2017年1月1日的天数),
α 为相对生长速率,
β 为物种相关常数.定义展叶开始的时间为单叶面积达到最大单叶面积的10%, 记为
T (10) , 展叶结束的时间为单叶面积达到最大面积的90%, 记为
T (90) , 展叶持续时间为
T (90) 和
T (10) 的差值(
夏洋洁等, 2013 ).新生小枝枝长和直径及总叶面积相对生长速率(
RGR )的公式是:
RGR = (ln
L 1 - ln
L 2 )/(
T 1 -
T 2 ).公式中,
T 1 以及
T 2 分别是两次测定的时间,
L 1 和
L 2 分别是
T 1 和
T 2 时的枝长、直径和总叶面积; ln是自然对数(
Aanderud et al ., 2003 ). ...
... 通过前期对青城山常绿阔叶林木本植物抽枝现象进行调查, 发现一次抽枝率高于二次抽枝率, 除了与植物本身遗传特性相关外, 一次抽枝所处的适宜气候条件以及更长的生长时间也是重要影响因素.一次抽枝后, 部分植物能量耗损严重, 无法满足二次抽枝的需要(
Ralhan et al ., 1985 ), 外界环境因素如二次抽枝后气温很快降低, 降水大量减少, 以及昆虫的取食压力等(
Zvereva & Kozlov, 2006 )也会造成二次抽枝率减少.青城山地区为湿润性亚热带常绿阔叶林分布区, 气候温暖湿润, 特别是夏末秋初的温度在16-22 ℃左右, 平均湿度约为95% (
王振孟, 2018 ), 与春季抽枝阶段的气候相似.外界适宜环境条件加之植物自身碳积累量富余是部分物种发生二次抽枝的重要原因(
杨芳等, 2019 ).二次抽枝阶段也是植食性昆虫生长繁殖的好时机, 展叶期高营养和高水分的新生叶片使昆虫得以大量繁殖, 昆虫取食引起叶片组织损伤, 削弱植物的生长(
Weed et al ., 2011 ), 给植物二次抽枝造成了很大的压力.有研究表明, 出叶较早的新生叶片比出叶较晚的新生叶片具有更强的耐霜冻能力(
Vitasse et al ., 2014 ), 因此, 多数发生二次抽枝的植物选择快速完成抽枝展叶的策略.除此以外, 植物二次抽枝的发生与其生活型和生长型存在一定的联系.本研究指出不同生长型的植物二次抽枝率在小枝水平上灌木大于乔木, 这与
夏洋洁等(2013) 以及
时盼(2016) 的研究结果相同.气候环境条件, 尤其是气温对乔木层和灌木层物种抽枝出叶存在很大的影响, 而3个研究地区的气温差异较小, 夏洋洁等的研究地区年平均气温为16.2 ℃; 时盼的研究地区年平均气温为15.4 ℃; 而本研究地区年平均气温为15.2 ℃.另外, 林内灌木层光环境较弱, 光照面积小, 更多的灌木通过二次抽枝不断分叉, 侧向生长以促进小枝长度和叶片数量的增加, 扩大冠层深度, 进一步最大化地获取光资源促进碳的积累, 从而维持自身的生存繁衍.不同生活型的植物在小枝水平上落叶植物二次抽枝率大于常绿植物, 落叶植物比常绿植物具有更大的小枝长度、小枝直径和茎生物量, 且落叶植物小枝水平上支撑的总叶面积和单叶面积显著高于常绿植物(
Sun et al ., 2006 ), 这些均与落叶植物的高的二次抽枝率存在必然联系.相对而言, 落叶植物二次抽枝的发生是与其快速的叶片周转效率相关, 而常绿植物尤其是林内幼树阶段的二次抽枝则与其对耐阴环境的长期适应对策相关. ...
浙江天童国家森林公园5种常绿阔叶植物的一次和二次抽枝进程
3
2013
... 目前, 除了对二次抽枝现象及二次抽枝发生比例有很多观察报道外, 国内外关于木本植物抽枝展叶的研究主要集中在两次抽枝展叶动态的对比分析、抽枝展叶特征与昆虫取食和叶片形态的关系两方面(
夏洋洁等, 2013 ;
王瑞雪, 2015 ;
时盼, 2016 ), 但关于植物在两次抽枝过程中小枝和叶片间的异速生长关系、各功能部分先后生长顺序等生长策略与内在权衡仍缺乏研究报道.因此, 本文以四川省都江堰市青城山风景区亚热带常绿阔叶林为研究地点, 通过长期定位跟踪观测, 调查了常绿阔叶林下木本植物抽枝进程发生比例, 以具有典型二次抽枝现象的5种木本植物幼树为研究对象, 分析了每个物种在两次抽枝中的展叶动态、功能性状以及枝叶异速生长关系, 试图回答以下科学问题: 1)常绿阔叶林木本植物两次抽枝过程中小枝和叶片分别具有怎样的形态特征差异?2)两次抽枝过程中小枝和叶片相对生长速率的动态变化是否一致?3)抽枝期间叶片数量、叶片面积、小枝枝长和小枝直径四者之间是否存在权衡?通过对上述科学问题的研究, 试图揭示常绿阔叶林木本植物抽枝展叶所采取的对策, 理解植物在林下生境中忍耐、存活与生长的过程机制. ...
... 使用Image J软件计算扫描后的每个叶片长、宽及相对应的面积, 采用以下公式拟合各个物种的叶面积:
LA = α × LL × LW .公式中
LA 为叶片面积,
LL 为叶片长,
LW 为叶片宽,
α 为物种相关的常数.将野外观测的叶片长宽数据进行转换, 所有观测物种方程拟合
R 2 > 0.90.增长量随时间的动态变化转化成百分比计算, 即:
At = at /a max .
at 为时间
t 时小枝的单叶面积、枝长以及直径的增长量,
a max 为小枝总的增长量,
At 为在时间为
t 的时候小枝各性状增长量占其总增长量的比例.单叶面积相对生长速率用方程
A = A max / (1
+ exp(
β -
α × t ))求得, 其中
A 为叶面积,
A max 为最大单叶面积,
t 为儒略日(本文中指距离2017年1月1日的天数),
α 为相对生长速率,
β 为物种相关常数.定义展叶开始的时间为单叶面积达到最大单叶面积的10%, 记为
T (10) , 展叶结束的时间为单叶面积达到最大面积的90%, 记为
T (90) , 展叶持续时间为
T (90) 和
T (10) 的差值(
夏洋洁等, 2013 ).新生小枝枝长和直径及总叶面积相对生长速率(
RGR )的公式是:
RGR = (ln
L 1 - ln
L 2 )/(
T 1 -
T 2 ).公式中,
T 1 以及
T 2 分别是两次测定的时间,
L 1 和
L 2 分别是
T 1 和
T 2 时的枝长、直径和总叶面积; ln是自然对数(
Aanderud et al ., 2003 ). ...
... 通过前期对青城山常绿阔叶林木本植物抽枝现象进行调查, 发现一次抽枝率高于二次抽枝率, 除了与植物本身遗传特性相关外, 一次抽枝所处的适宜气候条件以及更长的生长时间也是重要影响因素.一次抽枝后, 部分植物能量耗损严重, 无法满足二次抽枝的需要(
Ralhan et al ., 1985 ), 外界环境因素如二次抽枝后气温很快降低, 降水大量减少, 以及昆虫的取食压力等(
Zvereva & Kozlov, 2006 )也会造成二次抽枝率减少.青城山地区为湿润性亚热带常绿阔叶林分布区, 气候温暖湿润, 特别是夏末秋初的温度在16-22 ℃左右, 平均湿度约为95% (
王振孟, 2018 ), 与春季抽枝阶段的气候相似.外界适宜环境条件加之植物自身碳积累量富余是部分物种发生二次抽枝的重要原因(
杨芳等, 2019 ).二次抽枝阶段也是植食性昆虫生长繁殖的好时机, 展叶期高营养和高水分的新生叶片使昆虫得以大量繁殖, 昆虫取食引起叶片组织损伤, 削弱植物的生长(
Weed et al ., 2011 ), 给植物二次抽枝造成了很大的压力.有研究表明, 出叶较早的新生叶片比出叶较晚的新生叶片具有更强的耐霜冻能力(
Vitasse et al ., 2014 ), 因此, 多数发生二次抽枝的植物选择快速完成抽枝展叶的策略.除此以外, 植物二次抽枝的发生与其生活型和生长型存在一定的联系.本研究指出不同生长型的植物二次抽枝率在小枝水平上灌木大于乔木, 这与
夏洋洁等(2013) 以及
时盼(2016) 的研究结果相同.气候环境条件, 尤其是气温对乔木层和灌木层物种抽枝出叶存在很大的影响, 而3个研究地区的气温差异较小, 夏洋洁等的研究地区年平均气温为16.2 ℃; 时盼的研究地区年平均气温为15.4 ℃; 而本研究地区年平均气温为15.2 ℃.另外, 林内灌木层光环境较弱, 光照面积小, 更多的灌木通过二次抽枝不断分叉, 侧向生长以促进小枝长度和叶片数量的增加, 扩大冠层深度, 进一步最大化地获取光资源促进碳的积累, 从而维持自身的生存繁衍.不同生活型的植物在小枝水平上落叶植物二次抽枝率大于常绿植物, 落叶植物比常绿植物具有更大的小枝长度、小枝直径和茎生物量, 且落叶植物小枝水平上支撑的总叶面积和单叶面积显著高于常绿植物(
Sun et al ., 2006 ), 这些均与落叶植物的高的二次抽枝率存在必然联系.相对而言, 落叶植物二次抽枝的发生是与其快速的叶片周转效率相关, 而常绿植物尤其是林内幼树阶段的二次抽枝则与其对耐阴环境的长期适应对策相关. ...
Within-twig biomass allocation in subtropical evergreen broad-leaved species along an altitudinal gradient: allometric scaling analysis
1
2009
... 小枝是植物最有活力的构件, 可为叶片的着生提供节点, 行使物理支撑与传导水分和养分的功能, 在植物生长发育过程中起着至关重要的作用; 叶片一般着生在小枝上, 主要行使截获光源并进行光合作用的功能.植物为应对环境的异质性, 可以权衡枝和叶的生物量分配(
Xiang et al ., 2009 ;
Messier et al ., 2017 ), 由此产生一系列相对适合的资源投入分配模式.在枝和叶的投资权衡上, 植物能够找到最适的收益/代价关系, 并以最经济的方式将养分等资源分配给光合结构(
Dombroskie & Aarssen, 2012 ).植物枝叶并不一定是同时发育和生长, 资源也不一定被平均分配到枝叶上.抽枝展叶过程中, 植物的枝叶生长是否同步反映了植物生长过程中碳收支策略的差异(
Lohier et al ., 2014 ).应对不同的环境条件, 植物如何在各个器官间权衡分配光合产物暂无一致的结论.我们通过本研究, 发现抽枝展叶初期, 不但在小枝和叶片的生长对比中, 两次抽枝的叶片生长速度快于枝条生长速度; 在小枝和叶片的内部(枝长-直径, 叶片大小-数量)也存在这样的生长关系, 枝长生长快于枝径, 叶片大小生长快于叶片数量.表明两次抽枝过程中, 植物均优先侧重于资源获取器官——叶片大小的生长, 以实现短时期内叶片截获光能的最大效率并尽可能快地获得光合回报(
Meng et al ., 2015 ).而植物在小枝伸长过程中优先枝条伸长, 也是植物尽快拓展空间, 为最大化展叶提供支持的佐证, 同时也可能是植物为减少虫食的一种生长对策(
朱旭斌, 2005 ).通常小枝越长越易受风力、枝条的机械强度影响, 所以需要增粗枝径, 以加强机械抗力.小枝水平上的抽枝展叶权衡可能与物种个体发育阶段的不同和物种遗传特征有关, 这种权衡关系影响着植物抽枝展叶的分配模式, 进而影响着植物的结构和功能(
Yan et al ., 2013 ).小枝水平上枝叶生长动态方面, 小枝枝长、小枝直径和叶片数量相对生长速率一次抽枝高于二次抽枝, 而单叶面积相对生长速率和
LMA 的增长速率二次抽枝高于一次抽枝, 这与植物小枝经过权衡选择快速增加生长速率、减少叶片投资的生长方式有关.二次抽枝植物的快速生长伴随着大量的能量消耗导致小枝对叶片面积增长投入不足, 最终形成的叶片面积小于一次抽枝.因此, 小枝水平上抽枝展叶的权衡关系是植物对林下光资源有限生境下的适应策略, 可帮助我们理解特定生境和生活史阶段枝和叶的资源投资权衡关系. ...
海南岛尖峰岭12种热带常绿阔叶乔木展叶期与开花期对气候变化的响应
2014
海南岛尖峰岭12种热带常绿阔叶乔木展叶期与开花期对气候变化的响应
2014
Scaling relationships among twig size, leaf size and leafing intensity in a successional series of subtropical forests
1
2013
... 小枝是植物最有活力的构件, 可为叶片的着生提供节点, 行使物理支撑与传导水分和养分的功能, 在植物生长发育过程中起着至关重要的作用; 叶片一般着生在小枝上, 主要行使截获光源并进行光合作用的功能.植物为应对环境的异质性, 可以权衡枝和叶的生物量分配(
Xiang et al ., 2009 ;
Messier et al ., 2017 ), 由此产生一系列相对适合的资源投入分配模式.在枝和叶的投资权衡上, 植物能够找到最适的收益/代价关系, 并以最经济的方式将养分等资源分配给光合结构(
Dombroskie & Aarssen, 2012 ).植物枝叶并不一定是同时发育和生长, 资源也不一定被平均分配到枝叶上.抽枝展叶过程中, 植物的枝叶生长是否同步反映了植物生长过程中碳收支策略的差异(
Lohier et al ., 2014 ).应对不同的环境条件, 植物如何在各个器官间权衡分配光合产物暂无一致的结论.我们通过本研究, 发现抽枝展叶初期, 不但在小枝和叶片的生长对比中, 两次抽枝的叶片生长速度快于枝条生长速度; 在小枝和叶片的内部(枝长-直径, 叶片大小-数量)也存在这样的生长关系, 枝长生长快于枝径, 叶片大小生长快于叶片数量.表明两次抽枝过程中, 植物均优先侧重于资源获取器官——叶片大小的生长, 以实现短时期内叶片截获光能的最大效率并尽可能快地获得光合回报(
Meng et al ., 2015 ).而植物在小枝伸长过程中优先枝条伸长, 也是植物尽快拓展空间, 为最大化展叶提供支持的佐证, 同时也可能是植物为减少虫食的一种生长对策(
朱旭斌, 2005 ).通常小枝越长越易受风力、枝条的机械强度影响, 所以需要增粗枝径, 以加强机械抗力.小枝水平上的抽枝展叶权衡可能与物种个体发育阶段的不同和物种遗传特征有关, 这种权衡关系影响着植物抽枝展叶的分配模式, 进而影响着植物的结构和功能(
Yan et al ., 2013 ).小枝水平上枝叶生长动态方面, 小枝枝长、小枝直径和叶片数量相对生长速率一次抽枝高于二次抽枝, 而单叶面积相对生长速率和
LMA 的增长速率二次抽枝高于一次抽枝, 这与植物小枝经过权衡选择快速增加生长速率、减少叶片投资的生长方式有关.二次抽枝植物的快速生长伴随着大量的能量消耗导致小枝对叶片面积增长投入不足, 最终形成的叶片面积小于一次抽枝.因此, 小枝水平上抽枝展叶的权衡关系是植物对林下光资源有限生境下的适应策略, 可帮助我们理解特定生境和生活史阶段枝和叶的资源投资权衡关系. ...
常绿阔叶林林下6种木本植物叶片非结构性碳水化合物的动态特征
2
2019
... 通过前期对青城山常绿阔叶林木本植物抽枝现象进行调查, 发现一次抽枝率高于二次抽枝率, 除了与植物本身遗传特性相关外, 一次抽枝所处的适宜气候条件以及更长的生长时间也是重要影响因素.一次抽枝后, 部分植物能量耗损严重, 无法满足二次抽枝的需要(
Ralhan et al ., 1985 ), 外界环境因素如二次抽枝后气温很快降低, 降水大量减少, 以及昆虫的取食压力等(
Zvereva & Kozlov, 2006 )也会造成二次抽枝率减少.青城山地区为湿润性亚热带常绿阔叶林分布区, 气候温暖湿润, 特别是夏末秋初的温度在16-22 ℃左右, 平均湿度约为95% (
王振孟, 2018 ), 与春季抽枝阶段的气候相似.外界适宜环境条件加之植物自身碳积累量富余是部分物种发生二次抽枝的重要原因(
杨芳等, 2019 ).二次抽枝阶段也是植食性昆虫生长繁殖的好时机, 展叶期高营养和高水分的新生叶片使昆虫得以大量繁殖, 昆虫取食引起叶片组织损伤, 削弱植物的生长(
Weed et al ., 2011 ), 给植物二次抽枝造成了很大的压力.有研究表明, 出叶较早的新生叶片比出叶较晚的新生叶片具有更强的耐霜冻能力(
Vitasse et al ., 2014 ), 因此, 多数发生二次抽枝的植物选择快速完成抽枝展叶的策略.除此以外, 植物二次抽枝的发生与其生活型和生长型存在一定的联系.本研究指出不同生长型的植物二次抽枝率在小枝水平上灌木大于乔木, 这与
夏洋洁等(2013) 以及
时盼(2016) 的研究结果相同.气候环境条件, 尤其是气温对乔木层和灌木层物种抽枝出叶存在很大的影响, 而3个研究地区的气温差异较小, 夏洋洁等的研究地区年平均气温为16.2 ℃; 时盼的研究地区年平均气温为15.4 ℃; 而本研究地区年平均气温为15.2 ℃.另外, 林内灌木层光环境较弱, 光照面积小, 更多的灌木通过二次抽枝不断分叉, 侧向生长以促进小枝长度和叶片数量的增加, 扩大冠层深度, 进一步最大化地获取光资源促进碳的积累, 从而维持自身的生存繁衍.不同生活型的植物在小枝水平上落叶植物二次抽枝率大于常绿植物, 落叶植物比常绿植物具有更大的小枝长度、小枝直径和茎生物量, 且落叶植物小枝水平上支撑的总叶面积和单叶面积显著高于常绿植物(
Sun et al ., 2006 ), 这些均与落叶植物的高的二次抽枝率存在必然联系.相对而言, 落叶植物二次抽枝的发生是与其快速的叶片周转效率相关, 而常绿植物尤其是林内幼树阶段的二次抽枝则与其对耐阴环境的长期适应对策相关. ...
... 本研究的5个物种为长期耐阴物种如灌木(茶、细枝柃)和乔木的幼树阶段(润楠、短刺米槠、大叶山矾).研究所选的样本植物高度均在1-2 m范围内, 茶和细枝柃两种灌木在最适宜的生长环境条件下能达到的最大树高分别为6和10 m, 短刺米槠和大叶山矾最大树高也不超过20 m.因此, 本研究的物种幼树或成树阶段几乎均生长在林下层或中层, 树冠结构多呈由较大叶片构成的开阔型树冠, 叶片大多较平整且均匀地排列在小枝上以最大限度地捕获光资源.林窗数量少且面积小导致植物接受到的光照较弱, 长期以来就形成了较强的耐阴特征, 在林内光资源有限的逆境下这些物种能够进行多次抽枝增加碳获取是促进其生活史成功的重要因素.除大叶山矾外, 其余研究物种一次抽枝展叶持续时间、小枝直径和叶片数量均高于二次抽枝, 大叶山矾在两次抽枝中, 小枝直径和单叶面积均大于其他物种.这可能和大叶山矾本身的物种特性有关, 5个研究物种中大叶山矾植株个体最高, 获取资源的能力更强; 同时与
Corner (1949) 在70年前的发现一致, 不同物种较粗的当年生小枝通常具有较大的叶片, 这一观点也被广泛证实(
Westoby & Wright, 2003 ;
Sun et al ., 2006 ).大叶山矾的粗枝和大叶使其在叶片伸展方面更具有优势, 能截获更多的光资源(
Pickup et al ., 2005 )积累更多的碳, 同时叶片的大小反映了植物生长能力的强弱, 对其应对环境条件变化所采取的适应策略有着显著影响(
Conesa et al ., 2019 ).茶和细枝柃两种灌木植株个体较矮, 尤其是茶, 物种本身的遗传特性决定了其二次抽枝的高抽枝率, 两者
LMA 在两次抽枝中均小于润楠、短刺米槠和大叶山矾3种幼树阶段的乔木, 投入相同资源时, 茶和细枝柃的叶面积生长更大, 比起其他物种耐阴性也更强.除了短刺米槠和大叶山矾外, 其余3个研究物种二次抽枝最大
LMA 均大于一次抽枝, 原因可能是短刺米槠和大叶山矾幼树阶段对光资源的需求更加强烈, 而二次抽枝期间光照强度的减弱使得这两个物种的二次抽枝最大
LMA 小于一次抽枝; 茶、细枝柃和润楠在二次抽枝过程中通过快速增加
LMA , 促进叶片厚度和硬度的增长以抵抗植食性昆虫的大量取食,
LMA 较高的叶片含水量和N含量较低, 昆虫食用后会出现生长缓慢、患病甚至死亡的现象(
邹钟琳, 1980 ;
柳凤娟, 2010 ;
杨芳等, 2019 ), 这在一定程度上反映了植物应对昆虫取食所表现出的适应对策.因此, 植物抽枝策略的差异与物种特性, 如遗传特征、生活型、耐阴特性、植株高矮等息息相关, 植物通过形成不同的抽枝策略与环境变化相协调, 这对于提高外界资源的利用效率, 快速占领生态位, 扩大群落竞争优势有着重要意义. ...
常绿阔叶林林下6种木本植物叶片非结构性碳水化合物的动态特征
2
2019
... 通过前期对青城山常绿阔叶林木本植物抽枝现象进行调查, 发现一次抽枝率高于二次抽枝率, 除了与植物本身遗传特性相关外, 一次抽枝所处的适宜气候条件以及更长的生长时间也是重要影响因素.一次抽枝后, 部分植物能量耗损严重, 无法满足二次抽枝的需要(
Ralhan et al ., 1985 ), 外界环境因素如二次抽枝后气温很快降低, 降水大量减少, 以及昆虫的取食压力等(
Zvereva & Kozlov, 2006 )也会造成二次抽枝率减少.青城山地区为湿润性亚热带常绿阔叶林分布区, 气候温暖湿润, 特别是夏末秋初的温度在16-22 ℃左右, 平均湿度约为95% (
王振孟, 2018 ), 与春季抽枝阶段的气候相似.外界适宜环境条件加之植物自身碳积累量富余是部分物种发生二次抽枝的重要原因(
杨芳等, 2019 ).二次抽枝阶段也是植食性昆虫生长繁殖的好时机, 展叶期高营养和高水分的新生叶片使昆虫得以大量繁殖, 昆虫取食引起叶片组织损伤, 削弱植物的生长(
Weed et al ., 2011 ), 给植物二次抽枝造成了很大的压力.有研究表明, 出叶较早的新生叶片比出叶较晚的新生叶片具有更强的耐霜冻能力(
Vitasse et al ., 2014 ), 因此, 多数发生二次抽枝的植物选择快速完成抽枝展叶的策略.除此以外, 植物二次抽枝的发生与其生活型和生长型存在一定的联系.本研究指出不同生长型的植物二次抽枝率在小枝水平上灌木大于乔木, 这与
夏洋洁等(2013) 以及
时盼(2016) 的研究结果相同.气候环境条件, 尤其是气温对乔木层和灌木层物种抽枝出叶存在很大的影响, 而3个研究地区的气温差异较小, 夏洋洁等的研究地区年平均气温为16.2 ℃; 时盼的研究地区年平均气温为15.4 ℃; 而本研究地区年平均气温为15.2 ℃.另外, 林内灌木层光环境较弱, 光照面积小, 更多的灌木通过二次抽枝不断分叉, 侧向生长以促进小枝长度和叶片数量的增加, 扩大冠层深度, 进一步最大化地获取光资源促进碳的积累, 从而维持自身的生存繁衍.不同生活型的植物在小枝水平上落叶植物二次抽枝率大于常绿植物, 落叶植物比常绿植物具有更大的小枝长度、小枝直径和茎生物量, 且落叶植物小枝水平上支撑的总叶面积和单叶面积显著高于常绿植物(
Sun et al ., 2006 ), 这些均与落叶植物的高的二次抽枝率存在必然联系.相对而言, 落叶植物二次抽枝的发生是与其快速的叶片周转效率相关, 而常绿植物尤其是林内幼树阶段的二次抽枝则与其对耐阴环境的长期适应对策相关. ...
... 本研究的5个物种为长期耐阴物种如灌木(茶、细枝柃)和乔木的幼树阶段(润楠、短刺米槠、大叶山矾).研究所选的样本植物高度均在1-2 m范围内, 茶和细枝柃两种灌木在最适宜的生长环境条件下能达到的最大树高分别为6和10 m, 短刺米槠和大叶山矾最大树高也不超过20 m.因此, 本研究的物种幼树或成树阶段几乎均生长在林下层或中层, 树冠结构多呈由较大叶片构成的开阔型树冠, 叶片大多较平整且均匀地排列在小枝上以最大限度地捕获光资源.林窗数量少且面积小导致植物接受到的光照较弱, 长期以来就形成了较强的耐阴特征, 在林内光资源有限的逆境下这些物种能够进行多次抽枝增加碳获取是促进其生活史成功的重要因素.除大叶山矾外, 其余研究物种一次抽枝展叶持续时间、小枝直径和叶片数量均高于二次抽枝, 大叶山矾在两次抽枝中, 小枝直径和单叶面积均大于其他物种.这可能和大叶山矾本身的物种特性有关, 5个研究物种中大叶山矾植株个体最高, 获取资源的能力更强; 同时与
Corner (1949) 在70年前的发现一致, 不同物种较粗的当年生小枝通常具有较大的叶片, 这一观点也被广泛证实(
Westoby & Wright, 2003 ;
Sun et al ., 2006 ).大叶山矾的粗枝和大叶使其在叶片伸展方面更具有优势, 能截获更多的光资源(
Pickup et al ., 2005 )积累更多的碳, 同时叶片的大小反映了植物生长能力的强弱, 对其应对环境条件变化所采取的适应策略有着显著影响(
Conesa et al ., 2019 ).茶和细枝柃两种灌木植株个体较矮, 尤其是茶, 物种本身的遗传特性决定了其二次抽枝的高抽枝率, 两者
LMA 在两次抽枝中均小于润楠、短刺米槠和大叶山矾3种幼树阶段的乔木, 投入相同资源时, 茶和细枝柃的叶面积生长更大, 比起其他物种耐阴性也更强.除了短刺米槠和大叶山矾外, 其余3个研究物种二次抽枝最大
LMA 均大于一次抽枝, 原因可能是短刺米槠和大叶山矾幼树阶段对光资源的需求更加强烈, 而二次抽枝期间光照强度的减弱使得这两个物种的二次抽枝最大
LMA 小于一次抽枝; 茶、细枝柃和润楠在二次抽枝过程中通过快速增加
LMA , 促进叶片厚度和硬度的增长以抵抗植食性昆虫的大量取食,
LMA 较高的叶片含水量和N含量较低, 昆虫食用后会出现生长缓慢、患病甚至死亡的现象(
邹钟琳, 1980 ;
柳凤娟, 2010 ;
杨芳等, 2019 ), 这在一定程度上反映了植物应对昆虫取食所表现出的适应对策.因此, 植物抽枝策略的差异与物种特性, 如遗传特征、生活型、耐阴特性、植株高矮等息息相关, 植物通过形成不同的抽枝策略与环境变化相协调, 这对于提高外界资源的利用效率, 快速占领生态位, 扩大群落竞争优势有着重要意义. ...
南京地区落叶栎林主要木本植物的展叶动态研究
2
2005
... 经过长期观察, 青城山常绿阔叶林部分木本植物每年发生两次抽枝, 一次抽枝和二次抽枝开始时间分别为4月和8月下旬.因此, 选择2017年3月底以及2017年8月初在林内人为干扰较少且植物群落典型的地点, 对随机遇到的植物做抽枝调查, 并计算小枝水平上的二次抽枝率.结合样地内物种二次抽枝现象, 选择二次抽枝率较高的茶(
Camellia sinensis )、细枝柃(
Eurya loquaiana )、润楠、短刺米槠(
Castanopsis carlesii var.
spinulosa )以及大叶山矾(
Symplocos grandis ) 5种代表性植物进行研究, 于2017年3月中下旬植物爆芽之前在样地内相似的生境中每个物种随机选取5株长势良好且无病虫害的植株进行树高测量并长期观测.每株随机标记不同方向的3个当年生小枝, 从小枝爆芽开始记录小枝的长度、直径、叶片数量、叶片大小以及展叶时间.长度测量采用直尺和电子游标卡尺(IP54, 上海美耐特实业有限公司, 上海).在进行叶片数量和大小观测时, 叶片长度达到0.50 cm时计入新叶数目并测量长宽.展叶早期每隔5-7天观测一次, 以后观测间隔逐渐拉长, 直到连续三次测量叶片面积无明显的增长为止, 第二次抽枝观测方式与第一次相同(
朱旭斌等, 2005 ).在抽枝过程中, 每次观测后采集相似生境中植株个体上的完好健康叶片, 每个物种叶片采集30枚左右.每个物种共选取200枚包含展叶过程各个阶段大小的叶片, 通过佳能Lide 300扫描仪(Canon, Hanoi, Vietnam)扫描叶片. ...
... 小枝是植物最有活力的构件, 可为叶片的着生提供节点, 行使物理支撑与传导水分和养分的功能, 在植物生长发育过程中起着至关重要的作用; 叶片一般着生在小枝上, 主要行使截获光源并进行光合作用的功能.植物为应对环境的异质性, 可以权衡枝和叶的生物量分配(
Xiang et al ., 2009 ;
Messier et al ., 2017 ), 由此产生一系列相对适合的资源投入分配模式.在枝和叶的投资权衡上, 植物能够找到最适的收益/代价关系, 并以最经济的方式将养分等资源分配给光合结构(
Dombroskie & Aarssen, 2012 ).植物枝叶并不一定是同时发育和生长, 资源也不一定被平均分配到枝叶上.抽枝展叶过程中, 植物的枝叶生长是否同步反映了植物生长过程中碳收支策略的差异(
Lohier et al ., 2014 ).应对不同的环境条件, 植物如何在各个器官间权衡分配光合产物暂无一致的结论.我们通过本研究, 发现抽枝展叶初期, 不但在小枝和叶片的生长对比中, 两次抽枝的叶片生长速度快于枝条生长速度; 在小枝和叶片的内部(枝长-直径, 叶片大小-数量)也存在这样的生长关系, 枝长生长快于枝径, 叶片大小生长快于叶片数量.表明两次抽枝过程中, 植物均优先侧重于资源获取器官——叶片大小的生长, 以实现短时期内叶片截获光能的最大效率并尽可能快地获得光合回报(
Meng et al ., 2015 ).而植物在小枝伸长过程中优先枝条伸长, 也是植物尽快拓展空间, 为最大化展叶提供支持的佐证, 同时也可能是植物为减少虫食的一种生长对策(
朱旭斌, 2005 ).通常小枝越长越易受风力、枝条的机械强度影响, 所以需要增粗枝径, 以加强机械抗力.小枝水平上的抽枝展叶权衡可能与物种个体发育阶段的不同和物种遗传特征有关, 这种权衡关系影响着植物抽枝展叶的分配模式, 进而影响着植物的结构和功能(
Yan et al ., 2013 ).小枝水平上枝叶生长动态方面, 小枝枝长、小枝直径和叶片数量相对生长速率一次抽枝高于二次抽枝, 而单叶面积相对生长速率和
LMA 的增长速率二次抽枝高于一次抽枝, 这与植物小枝经过权衡选择快速增加生长速率、减少叶片投资的生长方式有关.二次抽枝植物的快速生长伴随着大量的能量消耗导致小枝对叶片面积增长投入不足, 最终形成的叶片面积小于一次抽枝.因此, 小枝水平上抽枝展叶的权衡关系是植物对林下光资源有限生境下的适应策略, 可帮助我们理解特定生境和生活史阶段枝和叶的资源投资权衡关系. ...
南京地区落叶栎林主要木本植物的展叶动态研究
2
2005
... 经过长期观察, 青城山常绿阔叶林部分木本植物每年发生两次抽枝, 一次抽枝和二次抽枝开始时间分别为4月和8月下旬.因此, 选择2017年3月底以及2017年8月初在林内人为干扰较少且植物群落典型的地点, 对随机遇到的植物做抽枝调查, 并计算小枝水平上的二次抽枝率.结合样地内物种二次抽枝现象, 选择二次抽枝率较高的茶(
Camellia sinensis )、细枝柃(
Eurya loquaiana )、润楠、短刺米槠(
Castanopsis carlesii var.
spinulosa )以及大叶山矾(
Symplocos grandis ) 5种代表性植物进行研究, 于2017年3月中下旬植物爆芽之前在样地内相似的生境中每个物种随机选取5株长势良好且无病虫害的植株进行树高测量并长期观测.每株随机标记不同方向的3个当年生小枝, 从小枝爆芽开始记录小枝的长度、直径、叶片数量、叶片大小以及展叶时间.长度测量采用直尺和电子游标卡尺(IP54, 上海美耐特实业有限公司, 上海).在进行叶片数量和大小观测时, 叶片长度达到0.50 cm时计入新叶数目并测量长宽.展叶早期每隔5-7天观测一次, 以后观测间隔逐渐拉长, 直到连续三次测量叶片面积无明显的增长为止, 第二次抽枝观测方式与第一次相同(
朱旭斌等, 2005 ).在抽枝过程中, 每次观测后采集相似生境中植株个体上的完好健康叶片, 每个物种叶片采集30枚左右.每个物种共选取200枚包含展叶过程各个阶段大小的叶片, 通过佳能Lide 300扫描仪(Canon, Hanoi, Vietnam)扫描叶片. ...
... 小枝是植物最有活力的构件, 可为叶片的着生提供节点, 行使物理支撑与传导水分和养分的功能, 在植物生长发育过程中起着至关重要的作用; 叶片一般着生在小枝上, 主要行使截获光源并进行光合作用的功能.植物为应对环境的异质性, 可以权衡枝和叶的生物量分配(
Xiang et al ., 2009 ;
Messier et al ., 2017 ), 由此产生一系列相对适合的资源投入分配模式.在枝和叶的投资权衡上, 植物能够找到最适的收益/代价关系, 并以最经济的方式将养分等资源分配给光合结构(
Dombroskie & Aarssen, 2012 ).植物枝叶并不一定是同时发育和生长, 资源也不一定被平均分配到枝叶上.抽枝展叶过程中, 植物的枝叶生长是否同步反映了植物生长过程中碳收支策略的差异(
Lohier et al ., 2014 ).应对不同的环境条件, 植物如何在各个器官间权衡分配光合产物暂无一致的结论.我们通过本研究, 发现抽枝展叶初期, 不但在小枝和叶片的生长对比中, 两次抽枝的叶片生长速度快于枝条生长速度; 在小枝和叶片的内部(枝长-直径, 叶片大小-数量)也存在这样的生长关系, 枝长生长快于枝径, 叶片大小生长快于叶片数量.表明两次抽枝过程中, 植物均优先侧重于资源获取器官——叶片大小的生长, 以实现短时期内叶片截获光能的最大效率并尽可能快地获得光合回报(
Meng et al ., 2015 ).而植物在小枝伸长过程中优先枝条伸长, 也是植物尽快拓展空间, 为最大化展叶提供支持的佐证, 同时也可能是植物为减少虫食的一种生长对策(
朱旭斌, 2005 ).通常小枝越长越易受风力、枝条的机械强度影响, 所以需要增粗枝径, 以加强机械抗力.小枝水平上的抽枝展叶权衡可能与物种个体发育阶段的不同和物种遗传特征有关, 这种权衡关系影响着植物抽枝展叶的分配模式, 进而影响着植物的结构和功能(
Yan et al ., 2013 ).小枝水平上枝叶生长动态方面, 小枝枝长、小枝直径和叶片数量相对生长速率一次抽枝高于二次抽枝, 而单叶面积相对生长速率和
LMA 的增长速率二次抽枝高于一次抽枝, 这与植物小枝经过权衡选择快速增加生长速率、减少叶片投资的生长方式有关.二次抽枝植物的快速生长伴随着大量的能量消耗导致小枝对叶片面积增长投入不足, 最终形成的叶片面积小于一次抽枝.因此, 小枝水平上抽枝展叶的权衡关系是植物对林下光资源有限生境下的适应策略, 可帮助我们理解特定生境和生活史阶段枝和叶的资源投资权衡关系. ...
1
1980
... 本研究的5个物种为长期耐阴物种如灌木(茶、细枝柃)和乔木的幼树阶段(润楠、短刺米槠、大叶山矾).研究所选的样本植物高度均在1-2 m范围内, 茶和细枝柃两种灌木在最适宜的生长环境条件下能达到的最大树高分别为6和10 m, 短刺米槠和大叶山矾最大树高也不超过20 m.因此, 本研究的物种幼树或成树阶段几乎均生长在林下层或中层, 树冠结构多呈由较大叶片构成的开阔型树冠, 叶片大多较平整且均匀地排列在小枝上以最大限度地捕获光资源.林窗数量少且面积小导致植物接受到的光照较弱, 长期以来就形成了较强的耐阴特征, 在林内光资源有限的逆境下这些物种能够进行多次抽枝增加碳获取是促进其生活史成功的重要因素.除大叶山矾外, 其余研究物种一次抽枝展叶持续时间、小枝直径和叶片数量均高于二次抽枝, 大叶山矾在两次抽枝中, 小枝直径和单叶面积均大于其他物种.这可能和大叶山矾本身的物种特性有关, 5个研究物种中大叶山矾植株个体最高, 获取资源的能力更强; 同时与
Corner (1949) 在70年前的发现一致, 不同物种较粗的当年生小枝通常具有较大的叶片, 这一观点也被广泛证实(
Westoby & Wright, 2003 ;
Sun et al ., 2006 ).大叶山矾的粗枝和大叶使其在叶片伸展方面更具有优势, 能截获更多的光资源(
Pickup et al ., 2005 )积累更多的碳, 同时叶片的大小反映了植物生长能力的强弱, 对其应对环境条件变化所采取的适应策略有着显著影响(
Conesa et al ., 2019 ).茶和细枝柃两种灌木植株个体较矮, 尤其是茶, 物种本身的遗传特性决定了其二次抽枝的高抽枝率, 两者
LMA 在两次抽枝中均小于润楠、短刺米槠和大叶山矾3种幼树阶段的乔木, 投入相同资源时, 茶和细枝柃的叶面积生长更大, 比起其他物种耐阴性也更强.除了短刺米槠和大叶山矾外, 其余3个研究物种二次抽枝最大
LMA 均大于一次抽枝, 原因可能是短刺米槠和大叶山矾幼树阶段对光资源的需求更加强烈, 而二次抽枝期间光照强度的减弱使得这两个物种的二次抽枝最大
LMA 小于一次抽枝; 茶、细枝柃和润楠在二次抽枝过程中通过快速增加
LMA , 促进叶片厚度和硬度的增长以抵抗植食性昆虫的大量取食,
LMA 较高的叶片含水量和N含量较低, 昆虫食用后会出现生长缓慢、患病甚至死亡的现象(
邹钟琳, 1980 ;
柳凤娟, 2010 ;
杨芳等, 2019 ), 这在一定程度上反映了植物应对昆虫取食所表现出的适应对策.因此, 植物抽枝策略的差异与物种特性, 如遗传特征、生活型、耐阴特性、植株高矮等息息相关, 植物通过形成不同的抽枝策略与环境变化相协调, 这对于提高外界资源的利用效率, 快速占领生态位, 扩大群落竞争优势有着重要意义. ...
1
1980
... 本研究的5个物种为长期耐阴物种如灌木(茶、细枝柃)和乔木的幼树阶段(润楠、短刺米槠、大叶山矾).研究所选的样本植物高度均在1-2 m范围内, 茶和细枝柃两种灌木在最适宜的生长环境条件下能达到的最大树高分别为6和10 m, 短刺米槠和大叶山矾最大树高也不超过20 m.因此, 本研究的物种幼树或成树阶段几乎均生长在林下层或中层, 树冠结构多呈由较大叶片构成的开阔型树冠, 叶片大多较平整且均匀地排列在小枝上以最大限度地捕获光资源.林窗数量少且面积小导致植物接受到的光照较弱, 长期以来就形成了较强的耐阴特征, 在林内光资源有限的逆境下这些物种能够进行多次抽枝增加碳获取是促进其生活史成功的重要因素.除大叶山矾外, 其余研究物种一次抽枝展叶持续时间、小枝直径和叶片数量均高于二次抽枝, 大叶山矾在两次抽枝中, 小枝直径和单叶面积均大于其他物种.这可能和大叶山矾本身的物种特性有关, 5个研究物种中大叶山矾植株个体最高, 获取资源的能力更强; 同时与
Corner (1949) 在70年前的发现一致, 不同物种较粗的当年生小枝通常具有较大的叶片, 这一观点也被广泛证实(
Westoby & Wright, 2003 ;
Sun et al ., 2006 ).大叶山矾的粗枝和大叶使其在叶片伸展方面更具有优势, 能截获更多的光资源(
Pickup et al ., 2005 )积累更多的碳, 同时叶片的大小反映了植物生长能力的强弱, 对其应对环境条件变化所采取的适应策略有着显著影响(
Conesa et al ., 2019 ).茶和细枝柃两种灌木植株个体较矮, 尤其是茶, 物种本身的遗传特性决定了其二次抽枝的高抽枝率, 两者
LMA 在两次抽枝中均小于润楠、短刺米槠和大叶山矾3种幼树阶段的乔木, 投入相同资源时, 茶和细枝柃的叶面积生长更大, 比起其他物种耐阴性也更强.除了短刺米槠和大叶山矾外, 其余3个研究物种二次抽枝最大
LMA 均大于一次抽枝, 原因可能是短刺米槠和大叶山矾幼树阶段对光资源的需求更加强烈, 而二次抽枝期间光照强度的减弱使得这两个物种的二次抽枝最大
LMA 小于一次抽枝; 茶、细枝柃和润楠在二次抽枝过程中通过快速增加
LMA , 促进叶片厚度和硬度的增长以抵抗植食性昆虫的大量取食,
LMA 较高的叶片含水量和N含量较低, 昆虫食用后会出现生长缓慢、患病甚至死亡的现象(
邹钟琳, 1980 ;
柳凤娟, 2010 ;
杨芳等, 2019 ), 这在一定程度上反映了植物应对昆虫取食所表现出的适应对策.因此, 植物抽枝策略的差异与物种特性, 如遗传特征、生活型、耐阴特性、植株高矮等息息相关, 植物通过形成不同的抽枝策略与环境变化相协调, 这对于提高外界资源的利用效率, 快速占领生态位, 扩大群落竞争优势有着重要意义. ...
Consequences of simultaneous elevation of carbon dioxide and temperature for plant-herbivore interactions: a meta analysis
1
2006
... 通过前期对青城山常绿阔叶林木本植物抽枝现象进行调查, 发现一次抽枝率高于二次抽枝率, 除了与植物本身遗传特性相关外, 一次抽枝所处的适宜气候条件以及更长的生长时间也是重要影响因素.一次抽枝后, 部分植物能量耗损严重, 无法满足二次抽枝的需要(
Ralhan et al ., 1985 ), 外界环境因素如二次抽枝后气温很快降低, 降水大量减少, 以及昆虫的取食压力等(
Zvereva & Kozlov, 2006 )也会造成二次抽枝率减少.青城山地区为湿润性亚热带常绿阔叶林分布区, 气候温暖湿润, 特别是夏末秋初的温度在16-22 ℃左右, 平均湿度约为95% (
王振孟, 2018 ), 与春季抽枝阶段的气候相似.外界适宜环境条件加之植物自身碳积累量富余是部分物种发生二次抽枝的重要原因(
杨芳等, 2019 ).二次抽枝阶段也是植食性昆虫生长繁殖的好时机, 展叶期高营养和高水分的新生叶片使昆虫得以大量繁殖, 昆虫取食引起叶片组织损伤, 削弱植物的生长(
Weed et al ., 2011 ), 给植物二次抽枝造成了很大的压力.有研究表明, 出叶较早的新生叶片比出叶较晚的新生叶片具有更强的耐霜冻能力(
Vitasse et al ., 2014 ), 因此, 多数发生二次抽枝的植物选择快速完成抽枝展叶的策略.除此以外, 植物二次抽枝的发生与其生活型和生长型存在一定的联系.本研究指出不同生长型的植物二次抽枝率在小枝水平上灌木大于乔木, 这与
夏洋洁等(2013) 以及
时盼(2016) 的研究结果相同.气候环境条件, 尤其是气温对乔木层和灌木层物种抽枝出叶存在很大的影响, 而3个研究地区的气温差异较小, 夏洋洁等的研究地区年平均气温为16.2 ℃; 时盼的研究地区年平均气温为15.4 ℃; 而本研究地区年平均气温为15.2 ℃.另外, 林内灌木层光环境较弱, 光照面积小, 更多的灌木通过二次抽枝不断分叉, 侧向生长以促进小枝长度和叶片数量的增加, 扩大冠层深度, 进一步最大化地获取光资源促进碳的积累, 从而维持自身的生存繁衍.不同生活型的植物在小枝水平上落叶植物二次抽枝率大于常绿植物, 落叶植物比常绿植物具有更大的小枝长度、小枝直径和茎生物量, 且落叶植物小枝水平上支撑的总叶面积和单叶面积显著高于常绿植物(
Sun et al ., 2006 ), 这些均与落叶植物的高的二次抽枝率存在必然联系.相对而言, 落叶植物二次抽枝的发生是与其快速的叶片周转效率相关, 而常绿植物尤其是林内幼树阶段的二次抽枝则与其对耐阴环境的长期适应对策相关. ...
 ,
, ,*中国科学院成都生物研究所, 中国科学院山地生态恢复与生物资源利用重点实验室, 生态恢复与生物多样性保育四川省重点实验室, 成都 610041; 中国科学院大学, 北京 100049
,*中国科学院成都生物研究所, 中国科学院山地生态恢复与生物资源利用重点实验室, 生态恢复与生物多样性保育四川省重点实验室, 成都 610041; 中国科学院大学, 北京 100049
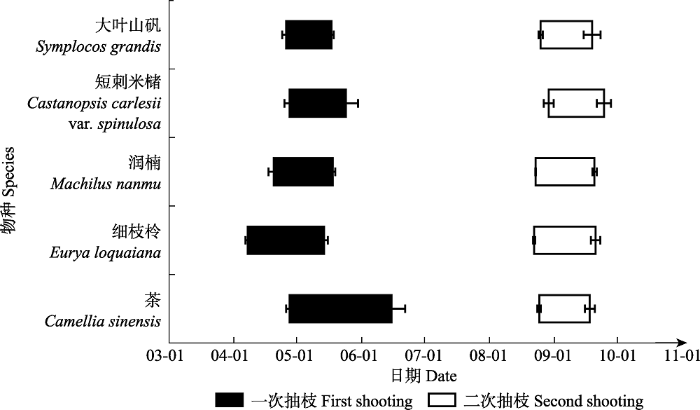 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT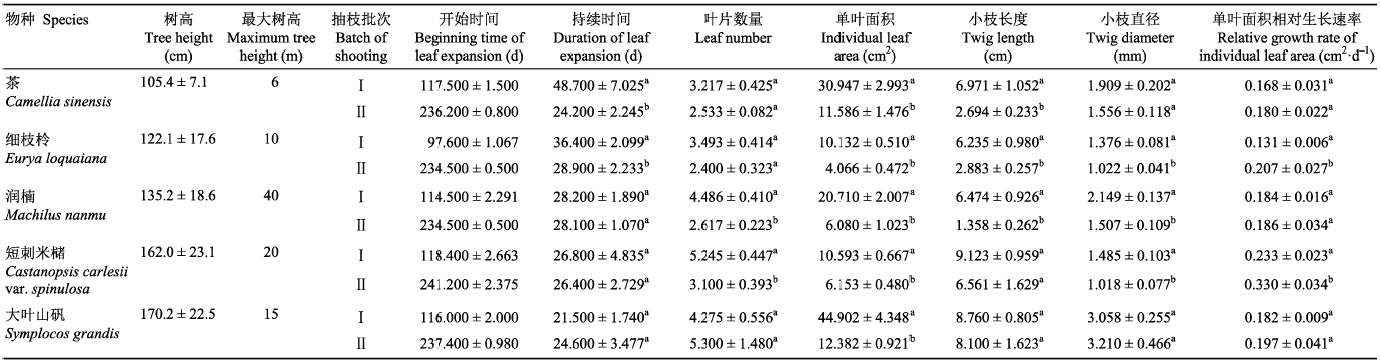
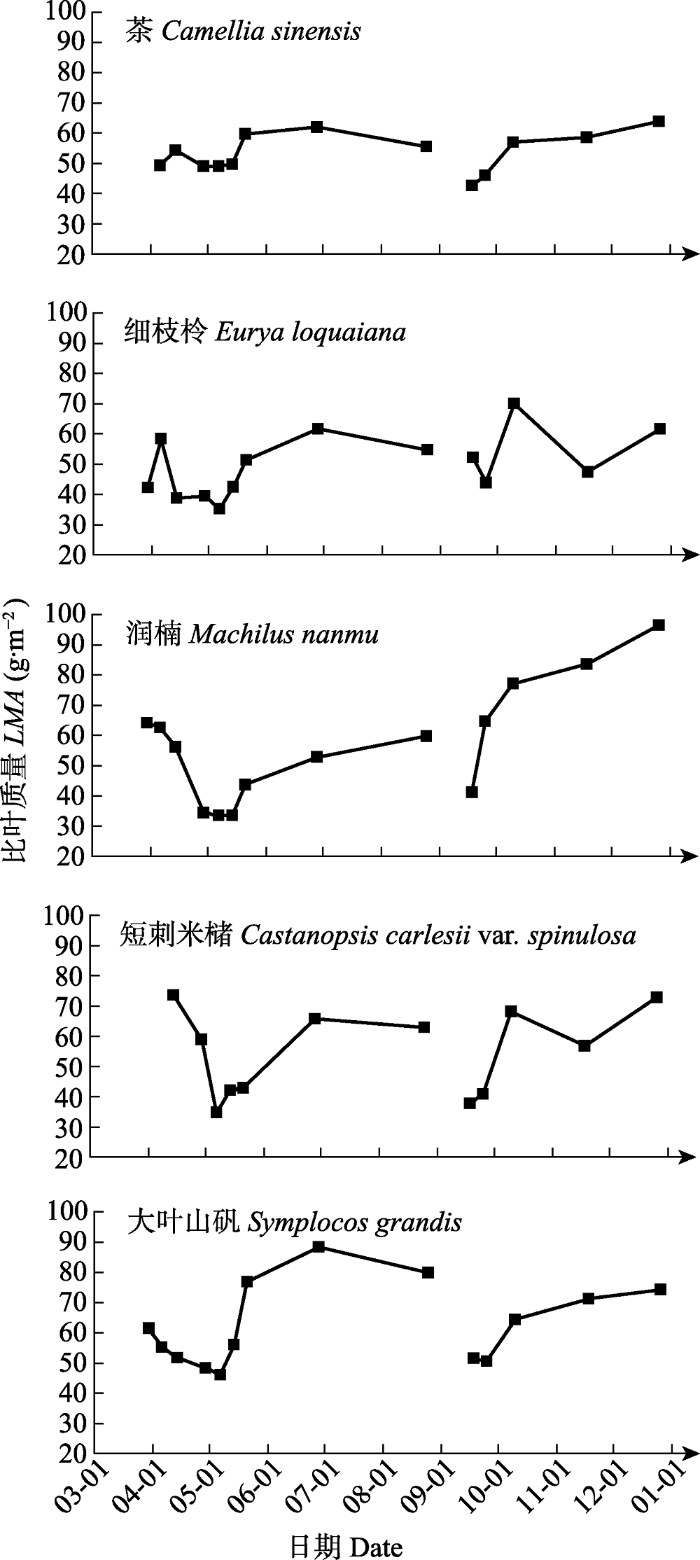 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT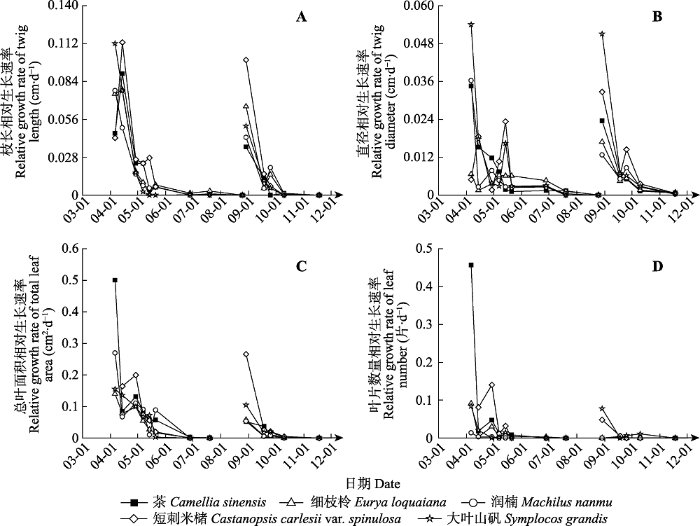 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT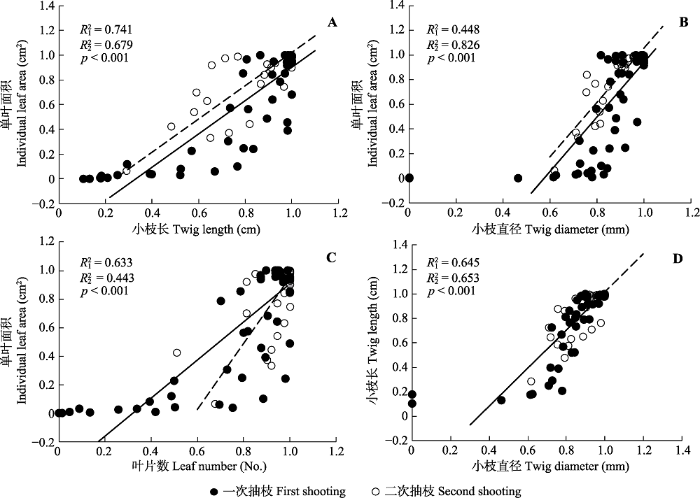 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}