<script type="text/javascript" src="https://cdn.bootcss.com/mathjax/2.7.2-beta.0/MathJax.js?config=TeX-AMS-MML_HTMLorMML"></script>
<script type='text/x-mathjax-config'>
MathJax.Hub.Config({
extensions: ["tex2jax.js"],
jax: ["input/TeX", "output/HTML-CSS"],
tex2jax: {inlineMath: [ ['$','$'], ["\\(","\\)"] ],displayMath: [ ['$$','$$'], ["\\[","\\]"] ],processEscapes: true},
"HTML-CSS": { availableFonts: ["TeX"] },
TeX: {equationNumbers: {autoNumber: ["none"], useLabelIds: true}},
"HTML-CSS": {linebreaks: {automatic: true}},
SVG: {linebreaks: {automatic: true}}
});
</script>
张亚洲
1 , 2 , * , 王淞伟
1 , 3 , * , 何小芳
1 , 2 , 杨扬
1 , 陈建国
, , 1 , ** , 孙航
, 1 , ** 1 中国科学院昆明植物研究所, 中国科学院东亚植物多样性和生物地理学重点实验室, 昆明 6502012 中国科学院大学, 北京 1000493 云南大学生命科学学院, 昆明 650091Altitudinal variation in flowering area and position and their ecological significances of an alpine cushion Arenaria polytrichoides, a gynodioecious herb Ya-Zhou ZHANG
1 , 2 , * , Song-Wei WANG
1 , 3 , * , Xiao-Fang HE
1 , 2 , Yang YANG
1 , Jian-Guo CHEN
, , 1 , ** , Hang SUN
, 1 , ** 1 CAS Key Laboratory for Plant Biodiversity and Biogeography of East Asia, Kunming Institute of Botany, Chinese Academy of Sciences, Kunming 650201, China2 University of Chinese Academy of Sciences, Beijing 100049, China3 School of Life Science, Yunnan University, Kunming 650091, China通讯作者: **陈建国: ORCID:0000-0003-3062-9881,Chen JG:
chenjianguo@mail.kib.ac.cn ;Sun H:
sunhang@mail.kib.ac.cn *同等贡献 编委: 黄双全(特邀)
责任编辑: 李敏
收稿日期: 2020-04-27
接受日期: 2020-09-12
网络出版日期: 2020-11-20
基金资助: 国家重点研发计划 (2017YFC0505200 )国家自然科学基金-云南省联合基金 (U1802232 )第二次青藏高原综合科学考察研究 (2019QZKK0502 )云南省应用基础研究计划 (202001AT070060 )云南省**** (YNWR-QNBJ-2018-318 )
Corresponding authors: **ORCID:0000-0003-3062-9881,Chen JG:
chenjianguo@mail.kib.ac.cn ;Sun H:
sunhang@mail.kib.ac.cn *Contributed equally to this work. Received: 2020-04-27
Accepted: 2020-09-12
Online: 2020-11-20
Fund supported: National Key R&D Program of China (2017YFC0505200 )Key Projects of the Joint Fund of the National Natural Science Foundation of China (U1802232 )Second Tibetan Plateau Scientific Expedition and Research Program (2019QZKK0502 )Yunnan Applied Basic Research Project (202001AT070060 )Ten-thousand Talents Program of Yunnan Province (YNWR-QNBJ-2018-318 )
摘要 作为高山生态系统中的奠基种(foundation species), 垫状植物自身种群的繁殖与扩张, 对高山生态系统功能稳定性起着关键作用。但是, 垫状植物如何在极端环境条件下实现资源的有效利用与分配, 达到繁殖最优化, 至今鲜为人知。该研究在滇西北白马雪山沿海拔梯度选择具有不同坡度及坡向的5个团状福禄草(
Arenaria polytrichoides )种群, 调查、比较种群内、种群间以及具有不同性系统的植株个体之间的开花面积比、开花方位, 并分析不同生态因子对其开花特性的影响。结果表明: 随着海拔的升高, 团状福禄草个体变小, 其分配到开花的资源比例总体上随海拔上升呈现下降的趋势, 说明团状福禄草的繁殖分配受到由海拔所引起的生态因子的调控。但是, 部分低海拔种群内植物个体的繁殖分配显著低于部分高海拔种群, 说明海拔并非控制植物繁殖分配的唯一因素。此外, 植株开花总面积随植株个体增大而增加, 但开花面积比却随个体增大而变小, 说明植株分配到开花的资源增长速率可能低于植株个体的增长速率。在性别差异方面, 两性植株对开花的资源分配比例要显著高于雌性植株, 但是, 其差异程度受到海拔因素的影响。最后, 在同一种群内, 团状福禄草在冠层表面不同方位上的开花面积比存在显著差异性, 这种差异性在不同种群之间又具有不同的表现形式。
关键词: 高山植物 ;
开花面积 ;
繁殖分配 ;
资源限制 Abstract Aims As foundation species in the alpine ecosystems, the reproduction and recruitment of alpine cushion plants are very important for sustaining the alpine ecosystem functions. However, it still remains unclear that how cushion plants effectively allocate resources to optimize reproductive fitness.
Methods Here we selected five populations of a gynodioecious herb
Arenaria polytrichoides with different exposures and slopes along an altitudinal gradient on the Baima snow mountain in northwest Yunnan, southwest China, to investigate and compare flowering area and positions, within and among populations and between female and hermaphroditic morphs. By doing so, we further discuss how the environmental stresses affect the cushion’s flowering attributes thus the population-level reproduction.
Important findings The results showed that, individual plant size and resources allocated to flowering (flowering area %) both decreased with increasing elevation, indicating that the reproductive allocation strategy was significantly affected by elevation. However, a population at lower elevation showed lower reproductive investment than higher populations, suggesting that elevation was not the only factor affecting the cushion’s reproductive allocation. In addition, absolute flowering area increased with increasing individual size, but the flowering area ratio decreased, indicating that the increases in reproductive allocation are fewer than that in vegetative allocation. Hermaphroditic individuals invested more resources to flowering than females did, but again, such effect was affected by elevation. Moreover, within a single population, the flowering areas were significantly different among the four directions (east, south, west and north) within one single individual canopy, but such differences varied in different populations.
Keywords: alpine plant ;
flowering area ;
reproductive allocation ;
resource limitation PDF (1833KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 张亚洲, 王淞伟, 何小芳, 杨扬, 陈建国, 孙航. 高山垫状植物团状福禄草开花面积与方位随海拔的变化及其适应性. 植物生态学报, 2020, 44(11): 1154-1163. DOI:
10.17521/cjpe.2020.0124 ZHANG Ya-Zhou, WANG Song-Wei, HE Xiao-Fang, YANG Yang, CHEN Jian-Guo, SUN Hang.
Altitudinal variation in flowering area and position and their ecological significances of an alpine cushion Arenaria polytrichoides, a gynodioecious herb .
Chinese Journal of Plant Ecology , 2020, 44(11): 1154-1163. DOI:
10.17521/cjpe.2020.0124 随着海拔逐渐上升, 环境条件变得更加严酷, 如: 海拔每上升100 m, 气温约下降0.6 ℃, 气压下降8-9 hPa (
K?rner, 2003 ), 土壤养分含量也随之减少(
曹舰艇等, 2019 )。面对高海拔地区低温、土壤养分贫瘠等恶劣环境因子, 高山植物在长期的进化过程中形成了丰富多样的特化适应性状或进化策略(
Sun et al ., 2014 ;
杨扬等, 2019 ), 种类丰富的植物多样性对维持高山生态系统的稳定性起到了关键作用(
Hooper et al ., 2005 )。滇西北处于横断山高山植物多样性中心区域(
郭辉军和龙春林, 1998 ;
Zhang et al. , 2015 )。长期以来, 青藏高原及横断山高海拔地区植物对极端环境因子的适应机制, 备受植物学家和生态学家的关注, 相关研究的深入开展, 加深了对该地区植物多样性形成及其进化适应机制的认识(
Sun et al ., 2014 ;
杨扬等, 2019 )。
温度、资源(包括土壤养分、降水资源及太阳辐射量等)是限制高山植物生长与繁殖的关键因素(
Bloom et al ., 1985 ;
Chen et al ., 2017b )。为了应对高海拔地区贫瘠的土壤条件, 植物在自身资源获取及分配方面进化出了众多与环境相适应的资源分配策略。其中, 繁殖分配(即植物分配到繁殖器官的资源量)是植物适合度的关键表现(
Bazzaz et al ., 1987 ;
Weiner, 1988 ;
Hartnett, 1990 ;
Vega et al ., 2000 )。研究表明, 植物繁殖分配同时受到众多非生物与生物因子的控制(
Reekie & Bazzaz, 2005 ;
张婵等, 2020 )。非生物因子方面, 温度、水分、土壤营养等对植物的繁殖分配起着重要作用(
Reekie & Bazzaz, 2005 ;
Zhang et al ., 2014 )。而在生物因子方面, 种间竞争、传粉者特性以及植物自身性系统特征(
Ashman, 1994 ;
Case & Ashman, 2005 ;
Brzyski et al ., 2014 ;
Chen et al ., 2017a ,
2017b )对其繁殖分配策略都起着重要的影响。例如, 具有雌全异株性系统特征(即同一种植物同时存在雌性个体和两性个体)的植物, 其同一种群内的雌性个体与两性个体在繁殖分配策略上存在较大差异, 表现为雌性个体主要将资源分配到雌性功能(如胚珠及种子生产), 而两性个体则主要将资源分配到雄性功能(如花粉生产以及传粉者吸引)(
Ashman, 1994 ;
Chen et al ., 2017a ,
2017b ), 并且这种分配模式受到环境因子的强烈影响(
Chen et al ., 2017b )。为了应对资源的不足, 植物往往需要在营养分配(即分配到营养器官的资源量)与繁殖分配之间做出权衡, 以达到最优的繁殖适合度(
Reekie & Avila-Sakar, 2005 )。开花是植物繁殖的最初阶段, 植物开花的数目, 直接反映出植物在特定环境条件下的繁殖分配策略。另一方面, 开花数目的多少也可直接决定着植物未来的繁殖输出量(如果实及种子产量;
Reekie & Bazzaz, 2005 )。
垫状植物是一类具有球形或半球形结构的植物的总称, 是植物经过多年辐射性生长而形成的小枝紧密簇生结构(
图1 ;
Gibson & Hope, 1986 ), 因其适应世界上大多数山地和极地极端干旱和低温环境的能力而备受植物学家的关注(
Aubert et al ., 2014 )。并因其具有生态系统工程师的功能, 对其他植物具有“护理作用”, 可提高和维持生态系统中物种多样性(
Butterfield et al ., 2013 ;
Cavieres et al ., 2014 ;
Chen et al ., 2015a ,
2015b ,
2019 ), 而被认为是所在生态系统当中的奠基种(foundation species;
Kikvidze et al ., 2015 )。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1研究区域及材料。A, 团状福禄草及开花朝向。B, 野外调查方法。C, 垫状植物内部结构。D, 研究区域及群落景观。 Fig. 1Study area and materials. A, Arenaria polytrichoides and flowering aspect of one individual. B, Field investigation protocol. C, Structure of the cushion plant. D, Study area and community landscape. 团状福禄草(
Arenaria polytrichoides )为石竹科无心菜属(
Arenaria )多年生垫状草本植物, 是青藏高原特有种, 沿喜马拉雅—横断山高海拔(3 500-5 300 m)地区分布(
Wu et al ., 2013 )。营养生长自冰雪消融开始, 花期主要在6-7月, 不同分布区域及海拔的种群的花期存在一定的差异(
Chen et al ., 2017a )。并且, 团状福禄草具有雌全异株的性二态系统, 即同一种群中同时存在雌性植株和两性植株, 不同性别的个体在繁殖产出(果实及种子产量)方面存在显著差异, 并且受到海拔的影响(
Chen et al ., 2017a )。表现为较高海拔的雌性比例高于较低海拔的, 两性植株开花数目大于雌性植株但结实率显著低于雌性植株, 同时, 较低海拔种群内个体单位面积的果实与种子产量高于较高海拔种群的个体(
Chen et al ., 2017a )。此外, 团状福禄草被证实为横断山高山生态系统当中的关键种, 能有效改良微环境条件(温度、土壤营养及水分), 从而为其他低耐受物种提供适宜的微生境, 进而调节种群动态、提升生态系统物种多样性及地面生产力(
Yang et al ., 2010 ,
2017 ;
Chen et al ., 2015a ,
2019 )。然而, 团状福禄草种群的更新却受到种间竞争的强烈影响(
Chen et al ., 2020 )。研究者在近十年内对团状福禄草做了深入调查与研究(
Yang et al ., 2010 ,
2017 ;
Chen et al ., 2015a ,
2015b ,
2017a ,
2019 ), 但对其种群的繁殖与更新机制, 却直到最近才予以关注(
Chen et al ., 2017a ,
2020 )。理论上讲, 团状福禄草的每一个小分枝(
图1C )在生长季节都能开出一朵花, 然而, 我们在多年的野外研究中发现, 实际上, 其每年的开花数量极为有限, 只在其冠层表面的部分区域开出一定面积的花(
图1A 、1B)。这一现象说明, 垫状植物的开花可能受到了资源的限制, 并有可能同时受到其他环境因子的影响。例如, 据野外观察, 团状福禄草开花方位(只在其冠层表面特定区域开放,
图1A 、1B)呈现出一定的规律性, 且在不同的种群之间存在差异。为了获取充足的太阳辐射能量从而满足自身繁殖需求, 一些高山植物的开花特性受到太阳辐射的强烈影响(
Pati?o et al ., 2002 ;
Galen, 2006 ;
Zhang et al ., 2010 ), 因此, 我们猜测团状福禄草的开花数量和方位可能受到资源和太阳辐射的双重影响。但其繁殖分配策略, 即个体对开花的资源分配策略对环境胁迫因子的响应方式, 目前却不为人知。深入探究高山生态系统中这一关键类群的种群更新与维持机制, 对揭示其种群繁殖的初始阶段(开花特性)对环境因子的响应机制以及这种响应机制在不同性别个体之间的差异性具有重要意义。
1 材料和方法 1.1 研究区域及实验材料 研究区域位于滇西北德钦县白马雪山, 地理坐标为28.48° N, 98.99° E, 海拔范围为4 360?4 920 m, 生境类型从低至高依次为灌草甸、草甸-流石滩交错带和流石滩(
图1D )。滇西北处于横断山高山植物多样性中心区域(
郭辉军和龙春林, 1998 ;
Zhang et al ., 2015 ), 具有典型的季节性夏季季风气候, 以冷雨或雨夹雪为特征, 并伴有短时间的强烈太阳辐射, 年降水量680-790 mm, 年平均气温-1 ℃ (
张荣祖等, 1997 )。植物生长季节通常开始于5月中旬的融雪期, 结束于9月中下旬的夏季风停息期。我们前期的测量数据显示, 2015年6月中旬至9月下旬, 白马雪山海拔4 900 m处, 距离地面15 cm处的平均气温和湿度分别为(4.87 ± 0.04) ℃ 和(89.9 ± 0.24)%, 绝对最高气温和最低气温分别为17.80和-1.95 ℃, 昼夜温差大于10 ℃, 最高达16 ℃ (陈建国等, 未发表数据)。此外, 土壤养分含量随海拔升高而普遍降低(
Chen et al ., 2015b ,
2019 )。
团状福禄草在研究区域的海拔范围内呈连续分布, 但受特定微生境特征及种间竞争强度的影响, 不同海拔带上的种群特征(大小、密度、结构)存在一定差异(
Chen et al ., 2020 )。团状福禄草是群落的奠基种, 粗糙红景天(
Rhodiola coccinea subsp.
scabrida )是群落的另一优势种, 群落中的常见物种还包括狭叶委陵菜(
Potentilla stenophylla )、七叶龙胆(
Gentiana arethusae var.
delicatula )、钉柱委陵菜(
Potentilla saundersiana )、镰萼喉毛花(
Comastoma falcatum )、圆穗蓼(
Polygonum macrophyllum )、滇边大黄(
Rheum delavayi )及早熟禾(
Poa spp.)等。
1.2 野外实验 于2017年6月下旬在研究区域内沿海拔梯度选取5个种群, 种群的选择考虑野外研究的可达性, 同时考虑不同种群的微生境差异性(坡向、坡度)。种群选定后, 精确测量其坡向和坡度。根据种群大小, 在每个种群内随机选择一定数量的个体, 对其开花特征进行调查和记录。各种群地理信息、生境特征及调查个体数量详见
表1 。对于随机选定的植物个体, 首先鉴定并记录其性系统特征(雌性或两性), 其次使用罗盘仪在其冠层表面标定东南西北4个方位(
图1B ), 然后, 使用孔径大小为 3 cm × 3 cm的尼龙网覆盖冠层表面, 尽量调整网格位置, 使其保持正方形。最后, 分别统计每个植株个体表面的总网格数及东南西北4个方向各自的开花网格数。具体操作方法: 垫状植物表面为球形或椭球形, 无论球形或椭球形, 都有一个“中心点(长、短轴交叉点)”, 以“中心点”为“原点”, 沿东西和南北方向各“画”一条线(即长、短轴), 可将其表面分割为4个“半球”, 即南北“半球”和东西“半球”, 然后计数各个“半球”的网格数。少部分网格里仅有部分面积开花, 为了使结果更客观, 对于这样的网格采取如下的方法计数: 开花面积小于网格面积1/3的, 不计入开花网格, 大于1/3的, 计入开花网格。
Table 1 表1 表1 团状福禄草种群地理信息、生境类型及各种群内个体调查数量
Table 1
Geographical information of the studied populations, habitat type and number of individuals investigated in each population of Arenaria polytrichoides 种群编号 生境类型 海拔 地理坐标 坡向 坡度 调查个体数 1 草甸 Meadow 4 360 28.45° N, 99.00° E 西 West 15 98 (41) 2 草甸 Meadow 4 450 28.45° N, 99.00° E 西 West 45 90 (36) 3 草甸 Meadow 4 520 28.46° N, 99.00° E 东南 SE 40 100 (56) 4 草甸-流石滩交错带 Meadow-screes ecotone 4 720 28.48° N, 99.00° E 东南 SE 40 100 (51) 5 流石滩 Screes 4 920 28.48° N, 99.01° E 山顶 Peak 0 100 (47)
括号内数字为雌性个体数。 Numbers in parentheses present the numbers of female individuals investigated in each population. 新窗口打开 |
下载CSV 1.3 数据分析 首先, 将各方向上开花网格数相加, 得到各植株的总开花网格(面积), 其次, 分别将总开花网格数和每个方向的开花网格数都除以总网格数, 得到各个植株总的开花面积比(为了论述方便, 本研究中, 必要时将开花面积比称为“繁殖分配”)以及单个方位上的开花面积比, 并采用这些数据进行后续的比较分析。不同植株的大小(年龄)会对其资源分配策略产生影响, 即可能存在个体效应(
St?cklin & Favre, 1994 ;
Sakai & Sakai, 1995 ), 同时, 垫状植物冠层表面积大小通常与其年龄呈正相关关系(
Chen et al ., 2017a ), 因此, 计数了每个调查个体的总网格数(即表面积大小), 将其作为植株大小的替代来进行后续分析。首先, 为了探究不同种群以及不同性别植株的总开花面积与其个体大小之间的关系, 对植株总开花面积与其个体大小做了回归分析。其次, 为了探究海拔对植株大小的影响, 采用混合效应模型比较了不同海拔种群间植株的大小, 植株大小为因变量, 海拔为主因素, 植株个体编号为随机因素。最后, 为了探究开花面积比(即繁殖分配)如何受到海拔、性别和植株大小的影响, 使用混合效应模型对上述关系进行拟合。模型中, 开花面积比为因变量, 海拔、性别和植株大小为主因素, 植株个体编号为随机因素。此外, 由于自变量与因变量间存在多样的组合形式, 基于赤池信息量准则(AIC)权重选择最优模型, 最优模型具有最大的AIC权重, 因子在每个模型的AIC权重被用来计算模型平均系数, 而每个因子在所有模型中的AIC值权重之和(
w )被用来描述该因子的统计支持。同时, 评估了考虑交互作用的模型与上述模型的优劣, 校准AIC (
AICc )结果显示两种模型没有差异(
AICc = -402.1 vs.
AICc = -401.4), 但是交互作用不显著, 因而以上述的模型结果为准。由于各组数据不完全符合正态分布及方差齐性原则, 使用Kruskal-Wallis非参数检验分别比较同一种群内不同性别、不同方位之间开花面积比以及不同种群之间开花面积比的差异。所有统计分析及画图都通过R 3.5.1 (
R Core Team, 2018 )实现, Kruskal-Wallis非参数检验使用“agricolae”包(
de Felipe, 2019 ), 混合效应模型使用“coxme”包(
Therneau, 2018 ), 模型选择使用“MuMIn”包(
Bartoń, 2019 )。
2 结果 随着海拔升高, 种群内植物个体大小呈现减小的趋势(混合效应模型系数= -0.131,
p < 0.001), 个体总开花面积比同样呈现下降趋势(
图2 ), 最小值同时出现在种群2 (次低海拔)和种群5 (最高海拔), 说明种群2除了受到海拔影响外, 可能还受到其他环境因子的强烈影响。植株个体大小与其总开花面积之间呈显著正相关关系(
图3A ), 说明随着植株年龄增长, 植物分配到开花的资源量也随之增长。但是, 植株越大, 总开花面积比越小(
表2 ;
图3B ), 说明植株分配到开花的资源增长速率可能低于植株个体的增长速率。种群1和种群2坡向相同, 坡度相差达30° (
表1 ), 个体总开花面积比差异显著(
图2 A)。种群3和种群4坡向、坡度均一致, 个体总开花面积比具有显著差异性(高海拔开花总比例低于低海)。此外, 种群5 (最高海拔)具有最低的总开花面积比, 说明海拔对个体繁殖分配具有强烈影响(
表2 )。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2团状福禄草种群间(A)及研究区域内不同性别间(B)开花面积比(平均值±标准误差)。不同的小写字母表示差异显著(p ≤ 0.05)。 Fig. 2Comparison of ratios of flowering area among populations (A) and between different gender morphs (B) of Arenaria polytrichoides in the study area (mean ± SE ). Different lowercase letters indicate significant difference (p ≤ 0.05). 图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3团状福禄草植株个体大小与其总开花面积及其面积比的关系。A、B, 所有种群。C、D, 种群1。E、F, 种群2。G、H, 种群3。I、J, 种群4。K、L, 种群5。回归线: 普通最小二乘线性回归线; Coef, 回归系数。所有数据被标准化至0-1之间。相关显著性: *, p ≤ 0.05; **, p ≤ 0.01; ***, p ≤ 0.001。 Fig. 3Relationship between plant size and the total flowering area and the relevant ratio of flowering area of Arenaria polytrichoides . A, B, All populations. C, D, Population 1. E, F, Population 2. G, H, Population 3. I, J, Population 4. K, L, Population 5. Regression line: ordinary least squares linear regression lines; coef, regression coefficient. All data are normalized to between 0 and 1. Significant relationship: *, p ≤ 0.05; **, p ≤ 0.01; ***, p ≤ 0.001. Table 2 表2 表2 植株开花面积与影响因子间作用的混合效应模型及模型选择结果
Table 2
Results of mixed effect models and selecting models of cushion flowering area and the explainary variables 模型系数 Value 标准误差 SE z p w AICc 截距 Intercept 1.919 1 0.186 3 10.300 0 <0.001 - 海拔 Altitude -0.000 3 0.000 0 -8.020 0 <0.001 1 -401.4 性别 Gender 0.035 4 0.014 4 2.460 0 0.014 0.88 植株大小 Plant size -0.001 2 0.000 2 -5.210 0 <0.001 1
该模型为最优模型: 开花面积=海拔+性别+植株大小+随机因素(植株个体编号)。AICc, 校准赤池信息量准则(AIC); w, 每一个变量在所有模型中的AIC权重之和。 The best model: Flowering area = Altitude + Gender + Plant size + Random factor (individual ID). AICc, corrected Akaike’s information criterion (AIC); w, the sum of the AIC weights of each predictor in all models. 新窗口打开 |
下载CSV 不同性别个体的植株在繁殖分配策略上存在较大差异。两性植株的总开花面积比显著大于雌性植株的总开花面积比(
图2B )。同时, 海拔对两性个体间的这种差异有一定影响, 表现为低海拔种群内两性个体总开花面积比显著大于雌性个体, 这种差异性随海拔升高而逐渐变小, 直至最高海拔种群处, 两性个体总开花面积比甚至略小于(不显著)雌性个体(
图4 )。
图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4团状福禄草各种群内不同性别植物个体开花面积比(平均值±标准误差)。不同小写字母表示组间差异显著(p ≤ 0.05)。 Fig. 4Ratio of flowering area (mean ± SE ) of different gender morphs in the five studied populations of Arenaria polytrichoides . Different lowercase letters indicate significant differences (p ≤ 0.05). 不同坡向及坡度上, 种群内垫状植物表面的开花方位存在着较大差异。具体表现为: 西向、坡度较小的种群1, 北面开花面积比最高, 西面最低, 东、南面无差异; 而同样为西向但坡度较大的种群2, 其南面开花面积最大, 其次为西面, 北面和东面随其后(
图5 )。东南向、坡度一致的种群3和种群4, 东面开花面积最大, 其他方位上开花面积无差异(
图5 )。位于山顶海拔最高(坡度为0)的种群5, 开花面积东面最大, 西面最小, 依次为东>北>南>西(
图5 )。
图5 新窗口打开 |
下载原图ZIP |
生成PPT 图5团状福禄草种群内不同方位的开花面积比(平均值±标准误差)比较。不同小写字母表示组间差异显著(p ≤ 0.05)。 Fig. 5Comparison of ratios of flowering areas (mean ± SE ) in different aspects within populations of Arenaria polytrichoides . Different lowercase letters indicate significant differences (p ≤ 0.05). 3 讨论 横断山高海拔地区绝大多数植物种群的更新与扩张都依赖有性繁殖(
Peng et al ., 2012 )。植物对繁殖功能的资源分配, 在很大程度上决定了其种群未来的扩张速率。而作为植物实现繁殖功能的重要环节, 植物对开花的资源配置决定了植物的最终繁殖产出(
Reekie & Bazzaz, 2005 )。不同的环境因子在植物的每一个繁殖阶段都起着重要作用, 甚至一些因子(如资源量)直接决定着植物的繁殖分配及其产出(
Delph & Carroll, 2001 )。在我们的研究区域内, 环境胁迫程度(如土壤养分胁迫及低温胁迫)随海拔升高而增强(
Chen et al ., 2015b ,
2019 )。我们的研究结果显示, 随着海拔升高, 种群内植株个体呈现减小的趋势。植株大小对开花特性具有显著影响。一方面, 植株个体越大, 其总开花面积也越大, 但另一方面, 植株越大, 其总开花面积比越小(
表2 ), 并且这种趋势在不同性别植株间具有高度一致性(
图3 ), 说明资源限制是真实存在的, 即, 植物从环境中获取并分配给开花的资源量有限。此外, 该结果也说明植株分配到开花的资源增长速率可能低于植株个体的增长速率, 即繁殖分配的增加速率低于营养分配的增加速率。相应结果进一步暗示, 在面临资源限制或环境胁迫时, 团状福禄草的资源分配策略以保证植株存活(即增加营养分配)为首要目的。此外, 具有不同坡度及坡向的种群间开花特性也存在一定的变化,说明由坡向及坡度变化而引起的环境因子变化, 如太阳辐射(
田辉等, 2007 )和土壤养分(
曹舰艇等, 2019 ), 可能也同时对其繁殖分配策略具有一定的影响。
前期研究发现, 团状福禄草植物个体表面单位面积的果实及种子产量随海拔上升而呈现总体下降趋势(
Chen et al ., 2017a )。但是, 该研究并未考虑团状福禄草植物整体的繁殖分配对环境因子的响应, 而仅是比较了其特定开花区域内单位面积的繁殖产出。本研究结果显示, 团状福禄草在开花阶段的繁殖分配在不同海拔种群间存在显著差异, 总体上随海拔上升而减少, 表明高海拔更加贫瘠的土壤条件(
Chen et al ., 2015b ,
2019 )可能限制了植物总体的资源获取量, 植物需要将有限的资源在营养生长与繁殖生长两方面之间做权衡(
Reekie & Avila-Sakar, 2005 ), 从而导致其在繁殖功能上的总体分配比例的下降。但是, 我们也发现, 部分低海拔种群(种群2)内植物个体的繁殖分配显著低于部分高海拔种群(种群3、4), 这表明, 海拔并非控制植物繁殖分配的唯一因素, 植物繁殖分配是多种生态因子共同作用的结果(
张婵等, 2020 )。种群2与种群1海拔相差较小, 但坡度相差较大, 其与种群3坡度相近, 但具有不同坡向(
表1 )。因此, 我们推测除了海拔因素, 坡度与坡向可能同时对其繁殖分配产生影响。一方面坡度大可能导致土壤养分流失更严重(
Bruce-Okine & Lal, 1975 ;
Donker & Damen, 1984 ), 从而引起资源胁迫的加剧, 并进而导致繁殖分配的减少。另一方面, 坡度和坡向通常会引起太阳辐射特征的变化, 如太阳入射角、入射时间在不同坡度和坡向上会发生变化(
田辉等, 2007 ), 从而引起植物可积累和利用的有效太阳辐射能量的变化。在高海拔地区, 太阳辐射对植物的生长与繁殖策略都起着重要的影响。如, 为了充分利用太阳辐射, 一些植物花器官进化出了“向日性”功能(
Zhang et al ., 2010 )。又如, 安第斯山脉中部高海拔岩石斜坡典型垫状植物
Azorella compacta 为了最大化利用太阳辐射, 其个体偏爱生长在向北的(南半球)坡度约为20°的斜坡(
Kleier et al ., 2015 )。因此, 我们推测由坡度与坡向变化引起的土壤养分状况及太阳辐射特征的变化, 可能同时对团状福禄草的繁殖分配起着重要的影响。
此外, 团状福禄草具有性二态系统, 即雌性植株与两性植株同时存在于同一种群中。不同性别的植物个体获得繁殖适合度的方式不同, 雌性个体主要通过胚珠及种子产出获得繁殖适合度, 而两性植株主要通过花粉与种子产出获得繁殖适合度(
Charlesworth & Charlesworth, 1978 ;
Charlesworth & Morgan, 1991 ;
Ashman, 1994 )。因此, 不同性别的植株个体在繁殖分配策略上存在一定的差异性, 这种差异性直接反映了不同性别个体对特定环境因子的适应策略(
Case & Ashman, 2005 ;
Ashman, 2006 ;
Chen et al ., 2017b )。例如, 同样分布于滇西北高海拔地区, 且同样具有雌全异株性系统的细叶蓝钟花(
Cyananthus delavayi ), 在面对环境因子胁迫时, 雌性植株主要将资源分配到种子生产, 而两性植株则主要将资源分配到花粉生产以及对传粉昆虫的吸引功能方面(花瓣)(
Chen et al ., 2017b )。我们的研究结果表明, 总体而言, 两性植株对开花的资源分配比例要显著高于雌性植株(
图2B ), 但是, 其差异程度受到海拔因素的影响。具体而言, 低海拔种群两性植株分配给开花的资源比例显著高于雌性植株, 而高海拔种群不同性别植株对开花的资源分配差异性不显著(
图4 )。
Chen等(2017a) 研究发现, 在任何海拔条件下, 团状福禄草两性植株单位面积的开花数量始终大于雌性植株, 但其真实繁殖产出(单位面积果实和种子产量)却始终低于雌性植株。但是, 本研究发现, 尽管不同性别的个体之间在低海拔处的总繁殖分配比例存在差异性, 但是随着海拔升高, 环境胁迫程度增加, 这种差异性趋于消失(
图4 )。我们猜测, 在高海拔环境胁迫加剧条件下, 不同性别的植物个体在资源权衡分配方面采取了相同的策略。另一方面, 除了物理胁迫的加剧, 生物胁迫也同时加剧, 如, 传粉昆虫多样性及多度的下降(
Arroyo et al ., 1982 ;
Medan et al ., 2002 )可能引起两性植株雄性适合度输出(花粉传播)的降低, 从而引起其分配到开花的资源随之减少。
如前所述, 太阳辐射特征可能对植物开花特性具有一定的调节功能。一方面表现在开花数量的多少, 另一方也可能表现在开花方位上。我们研究发现, 在同一种群内, 团状福禄草在冠层表面不同方位上的开花面积比存在显著差异, 但是, 这种差异在不同种群(即不同环境因子条件)之间又具有不同的表现形式。如, 最低海拔的种群1, 北面开花面积比最高, 东、南面次之且差异不显著, 而西面开花面积比最低; 最高海拔处的种群5, 其开花面积比呈现出东、北、南、西递减趋势, 且各方位上差异均显著(
图5 )。不同海拔及坡向上的种群, 可能由于坡度、坡向的差异而导致太阳辐射时间乃至辐射强度的变化(
田辉等, 2007 )。植物为了充分获取太阳辐射能量, 而尽可能将花开放在有利的方位上, 以满足果实及种子成熟过程中对太阳辐射能量的需求。
综上所述, 本研究证明了高山垫状植物团状福禄草的繁殖分配受到海拔因素的强烈影响, 并且不同性别的植株对海拔因子的响应机制存在显著差异。此外, 由海拔、坡度及坡向变化所引起的其他环境因子(诸如土壤养分、太阳辐射等)的变化, 可能同时对植物繁殖分配策略起着调节作用。植物在特定环境条件下, 通过调节自身的繁殖分配策略, 达到最优繁殖适合度, 保证了个体及种群的繁衍与扩张。然而, 本研究的不足之处在于, 我们无法准确回答坡度和坡向究竟如何影响土壤养分及太阳辐射等环境因子, 以及这些环境因子如何精确影响团状福禄草对繁殖资源的分配策略。未来更深入的研究, 需要更精确地设计研究实验, 定量分析各环境因子对垫状植物繁殖分配策略的影响, 以便更好地揭示其种群的繁衍与更新机制并预测其种群发展动态。
致谢 感谢中国科学院昆明植物研究所宋波、马祥光在文稿修改中给予的帮助。
[1] Arroyo MTK Primack R Armesto J 1982 ). Community studies in pollination ecology in the high temperate Andes of central Chile. I. Pollination mechanisms and altitudinal variationAmerican Journal of Botany 82 -97 . [本文引用: 1] [2] Ashman TL 1994 ). Reproductive allocation in hermaphrodite and female plants of Sidalcea oregana ssp. spicata (Malvaceae) using four currenciesAmerican Journal of Botany 433 -438 . [本文引用: 3] [3] Ashman TL 2006 ). The evolution of separate sexes: a focus on the ecological context//Harder LD, Barrett SCHEcology and Evolution of Flowers 204 -219 . [本文引用: 1] [4] Aubert S Boucher F Lavergne S Renaud J Choler P 2014 ). 1914-2014: a revised worldwide catalogue of cushion plants 100 years after Hauri and Schr?terAlpine Botany 59 -70 . [本文引用: 1] [5] Bartoń K 2019 ). MuMIn: Multi-Model Inference URL [本文引用: 1] [6] Bazzaz FA Chiariello NR Coley PD Pitelka LF 1987 ). Allocating resources to reproduction and defenseBioScience 58 -67 . [本文引用: 1] [7] Bloom AJ Chapin FS Mooney HA 1985 ). Resource limitation in plants — An economic analogyAnnual Review of Ecology and Systematics 363 -392 . [本文引用: 1] [8] Bruce-Okine E Lal R 1975 ). Soil erodibility as determined by raindrop techniqueSoil Science 149 -157 . [本文引用: 1] [9] Brzyski JR Taylor W McLetchie DN 2014 ). Reproductive allocation between the sexes, across natural and novel habitats, and its impact on genetic diversityEvolutionary Ecology 247 -261 . [本文引用: 1] [10] Butterfield BJ Cavieres LA Callaway RM Cook BJ Kikvidze Z Lortie CJ Michalet R Pugnaire FI Sch?b C Xiao S Zaitchek B Anthelme F Bj?rk RG Dickinson K Gavilán R et al 2013 ). Alpine cushion plants inhibit the loss of phylogenetic diversity in severe environmentsEcology Letters 478 -486 . [本文引用: 1] [11] Cao JT Yang H Cui ZZ Zhang XJ 2019 ). Distribution characteristics of soil nutrient contents at different altitudes in the eastern and western slopes of Sejila Mountain, Southeastern TibetJournal of Plateau Agriculture 1 ), 19 -27, 35 . [本文引用: 2] [ 曹舰艇 , 杨红 , 崔在忠 , 张新军 (2019 ). 藏东南色季拉山东西坡不同海拔高度土壤养分含量分布特征高原农业 , 3(1 ), 19 -27, 35 .] [本文引用: 2] [12] Case AL Ashman TL 2005 ). Sex-specific physiology and its implications for the cost of reproduction//Reekie E, Bazzaz FAReproductive Allocation in Plants 126 -154 . [本文引用: 2] [13] Cavieres LA Brooker RW Butterfield BJ Cook BJ Kikvidze Z Lortie CJ Michalet R Pugnaire FI Schob C Xiao S Anthelme F Bjork RG Dickinson KJM Cranston BH Gavilan R et al 2014 ). Facilitative plant interactions and climate simultaneously drive alpine plant diversityEcology Letters 193 -202 . URL PMID:
24238015 [本文引用: 1] [14] Charlesworth B Charlesworth D 1978 ). A model for the evolution of dioecy and gynodioecyThe American Naturalist 975 -997 . [本文引用: 1] [15] Charlesworth D Morgan MT 1991 ). Allocation of resources to sex functions in flowering plantsPhilosophical Transactions of the Royal Society of London Series B: Biological Sciences 91 -102 [本文引用: 1] [16] Chen JG He XF Wang SW Yang Y Sun H 2019 ). Cushion and shrub ecosystem engineers contribute differently to diversity and functions in alpine ecosystemsJournal of Vegetation Science 362 -374 . [本文引用: 6] [17] Chen JG Li YB Yang Y Sun H 2017 a). How cushion communities are maintained in alpine ecosystems: a review and case study on alpine cushion plant reproductionPlant Diversity 221 -228 . DOI:
10.1016/j.pld.2017.07.002 URL PMID:
30159515 [本文引用: 10] Cushion species occur in nearly all alpine environments worldwide. In past decades, the adaptive and ecosystem-engineering roles of such highly specialized life forms have been well studied. However, the adaptive strategies responsible for cushion species reproductive success and maintenance in severe alpine habitats remain largely unclear. In this study, we reviewed the current understanding of reproductive strategies and population persistence in alpine cushion species. We then present a preliminary case study on the sexual reproduction of Arenaria polytrichoides (Caryophyllaceae), a typical cushion species inhabiting high elevations of the Himalaya Hengduan Mountains, which is a hotspot for diversification of cushion species. Finally, we highlight the limitations of our current understanding of alpine cushion species reproduction and propose future directions for study.
[18] Chen JG Niu Y Li ZM Yang Y Sun H 2017 b). Sex allocation in gynodioecious Cyananthus delavayi differs between gender morphs and soil qualityPlant Reproduction 107 -117 . DOI:
10.1007/s00497-017-0303-4 URL PMID:
28597166 [本文引用: 6] KEY MESSAGE: Sex allocation in Cyananthus delavayi. Gynodioecy, where females and hermaphrodites coexist in the same natural population, is particularly suitable for predicting the ecological pressures that drive the stability of gender polymorphism. Since females have a disadvantage in that they only contribute to the next generation via ovules, they should gain an advantage via other means, of which resource allocation is an important component. Thus, to study their sex allocation is very helpful to understand how the dimorphic sexual system is maintained in natural systems. We studied the sex allocation patterns and reproductive output of the gynodioecious Cyananthus delavayi in three populations with different soil qualities (organic matter, N, P and K). The hermaphroditic flowers and pistils were much larger than those of female individuals. Although both gender morphs invested similar biomass in the pistils, females allocated more of their resource pool to the seed production, while hermaphrodites allocated more to pollinator advertisement. The pollen production of hermaphrodites did not differ between populations, suggesting that pollen production by hermaphrodites was not limited by soil nutrients. Fruit set of females, but not hermaphrodites, decreased with declining soil quality, whereas seeds per fruit of both females and hermaphrodites were highest in poor soils. Overall, this study shows that females achieve greater reproductive success by allocating more of their resource pool to enhancing seed production, which should favor their presence in gynodioecious populations. The hermaphrodites achieve reproductive success from both pollen and seed production, and unnecessarily reduce their allocation to pollen production. Soil quality should explain, at least partially, the sexual allocation patterns. Furthermore, some of our findings contradict previous hypotheses, thus adding a new example to the body of research on plant sex allocation and the development of future theories.
[19] Chen JG Sch?b C Zhou Z Gong Q Li X Yang Y Li Z Sun H 2015 a). Cushion plants can have a positive effect on diversity at high elevations in the Himalayan Hengduan MountainsJournal of Vegetation Science 768 -777 . [本文引用: 3] [20] Chen JG Yang Y St?cklin J Cavieres LA Peng D Li Z Sun H 2015 b). Soil nutrient availability determines the facilitative effects of cushion plants on other plant species at high elevations in the south-eastern HimalayasPlant Ecology & Diversity 199 -210 . [本文引用: 5] [21] Chen JG Yang Y Wang SW St?cklin J Peng DL Sun H 2020 ). Recruitment of the high elevation cushion plant Arenaria polytrichoides is limited by competition, thus threatened by currently established vegetationJournal of Systematics and Evolution 59 -68 . [本文引用: 3] [22] de Felipe M 2019 ). agricolae: Statistical Procedures for Agricultural Research URL [本文引用: 1] [23] Delph LF Carroll SB 2001 ). Factors affecting relative seed fitness and female frequency in a gynodioecious species, Silene acaulis Evolutionary Ecology Research 465 -476 . [本文引用: 1] [24] Donker NHW Damen MCJ 1984 ). Gully system development and an assessment of gully initiation risk in Miocene deposits near Daroca: SpainZeitschrift Fur Geomorphologie 37 -50 . [本文引用: 1] [25] Galen C 2006 ). Solar furnaces or swamp coolers: costs and benefits of water use by solar-tracking flowers of the alpine snow buttercup, Ranunculus adoneus Oecologia 195 -201 . URL PMID:
16465542 [本文引用: 1] [26] Gihson N Hope G 1986 ). On the origin of and evolution of Australasian alpine cushion plants//Barrow BAFlora and Fauna of Alpine Australasia 63 -81 . [本文引用: 1] [27] Guo HJ Long CL 1998 ). Yunnan’s Biodiversity [本文引用: 2] [ 郭辉军 , 龙春林 (1998 ). 云南的生物多样性. 云南科技出版社, 昆明 .] [本文引用: 2] [28] Hartnett DC 1990 ). Size-dependent allocation to sexual and vegetative reproduction in four clonal compositesOecologia 254 -259 . URL PMID:
28312762 [本文引用: 1] [29] Hooper D Chapin F Ewel JJ Hector A Inchausti P Lavorel S Lawton JH Lodge DM Loreau M Naeem S Schmid B Set?t? H Symstad AJ Vandermeer J Wardle DA 2005 ). Effects of biodiversity on ecosystem functioning: a consensus of current knowledgeEcological Monographs 1 ), 3 -35 . [本文引用: 1] [30] Kikvidze Z Brooker RW Butterfield BJ Callaway RM Cavieres LA Cook BJ Lortie CJ Michalet R Pugnaire FI Xiao S Anthelme F Bj?rk RG Cranston BH Gavilan RG Kanka R et al 2015 ). The effects of foundation species on community assembly: a global study on alpine cushion plant communitiesEcology 2064 -2069 . [本文引用: 1] [31] Kleier C Trenary T Graham EA Stenzel W Rundel PW 2015 ). Size class structure, growth rates, and orientation of the central Andean cushion Azorella compacta PeerJ 10.7717/peerj.843 . URL PMID:
33717695 [本文引用: 1] [32] K?rner C 2003 ). Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems [本文引用: 1] [33] Medan D Montaldo NH Devoto M Maniese A Vasellati V Roitman GG Bartoloni NH 2002 ). Plant-pollinator relationships at two altitudes in the Andes of Mendoza, ArgentinaArctic, Antarctic, and Alpine Research 233 -241 . [本文引用: 1] [34] Pati?o S Jeffree C Grace J 2002 ). The ecological role of orientation in tropical convolvulaceous flowersOecologia 373 -379 . URL PMID:
28547043 [本文引用: 1] [35] Peng DL Zhang ZQ Xu B Li ZM Sun H 2012 ). Patterns of flower morphology and sexual systems in the subnival belt of the Hengduan Mountains, SW ChinaAlpine Botany 65 -73 . [本文引用: 1] [36] R Core Team 2018 ). R: A Language and Environment for Statistical Computing . R Foundation for Statistical Computing, Vienna, Austria. [本文引用: 1] [37] Reekie EG Avila-Sakar G 2005 ). The shape of the trade-off function between reproduction and growth//Reekie EG, Bazzaz FAReproductive Allocation in Plants . Elsevier, Amsterdam189 -214 . [本文引用: 2] [38] Reekie EG Bazzaz FA 2005 ). Reproductive Allocation in Plants . Elsevier, Amsterdam. URL PMID:
27135522 [本文引用: 4] [39] Sakai S Sakai A 1995 ). Flower size-dependent variation in seed size: theory and a testThe American Naturalist 918 -934 . [本文引用: 1] [40] St?cklin J Favre P 1994 ). Effects of plant size and morphological constraints on variation in reproductive components in 2 related species of Epilobium Journal of Ecology 735 -746 . [本文引用: 1] [41] Sun H Niu Y Chen YS Song B Liu CQ Peng DL Chen JG Yang Y 2014 ). Survival and reproduction of plant species in the Qinghai-Tibet PlateauJournal of Systematics and Evolution 378 -396 . [本文引用: 2] [42] Therneau TM 2018 ). coxme: mixed effects cox models URL [本文引用: 1] [43] Tian H Wen J Ma YM Su ZB Wei ZG Wang JM Zhang TT Liu R Hu X 2007 ). Estimation of solar radiation over the complex terrain of the Heihe River BasinPlateau Meteorology 666 -676 . [本文引用: 3] [ 田辉 , 文军 , 马耀明 , 苏中波 , 韦志刚 , 王介民 , 张堂堂 , 刘蓉 , 胡晓 (2007 ). 复杂地形下黑河流域的太阳辐射计算高原气象 , 26, 666 -676 .] [本文引用: 3] [44] Vega CRC Sadras VO Andrade FH Uhart SA 2000 ). Reproductive allometry in soybean, maize and sunflowerAnnals of Botany 461 -468 . [本文引用: 1] [45] Weiner J 1988 ). The influence of competition on plant reproduction//Lovett Doust J, Lovett Doust LPlant Reproductive Ecology: Patterns and Strategies. Oxford University Press, New York228 -245 . [本文引用: 1] [46] Wu CY Raven PH Hong DY 2013 ). Flora of China . Science Press & Missouri Botanical Garden Press, Beijing & St. Louis [本文引用: 1] [47] Yang Y Chen JG Sch?b C Sun H 2017 ). Size-mediated interaction between a cushion species and other non- cushion species at high elevations of the Hengduan Mountains, SW ChinaFrontiers in Plant Science 465 . DOI: 10.3389/fpls.2017.00465 . URL PMID:
28424726 [本文引用: 2] [48] Yang Y Chen JG Song B Niu Y Peng DL Zhang JW Deng T Luo D Ma XG Zhou Z Sun H 2019 ). Advances in the studies of plant diversity and ecological adaptation in the subnival ecosystem of the Qinghai-Tibet PlateauChinese Science Bulletin 2856 -2864 . [本文引用: 2] [ 杨扬 , 陈建国 , 宋波 , 牛洋 , 彭德力 , 张建文 , 邓涛 , 罗冬 , 马祥光 , 周卓 , 孙航 (2019 ). 青藏高原冰缘植物多样性适应机制研究进展科学通报 , 64, 2856 -2864 .] [本文引用: 2] [49] Yang Y Niu Y Cavieres LA Sun H 2010 ). Positive associations between the cushion plant Arenaria polytrichoides (Caryophyllaceae) and other alpine plant species increase with altitude in the Sino-HimalayasJournal of Vegetation Science 1048 -1057 . [本文引用: 2] [50] Zhang C An YM J?schke Y Wang LL Zhou ZL Wang LP Yang YP Duan YW 2020 ). Processes on reproductive ecology of plant species in the Qinghai-Xizang Plateau and adjacent highlandsChinese Journal of Plant Ecology 1 -21 . [本文引用: 2] [ 张婵 , 安宇梦 , J?schke Y 王林林 , 周知里 , 王力平 , 杨永平 , 段元文 (2020 ). 青藏高原及周边高山地区的植物繁殖生态学研究进展植物生态学报 , 44, 1 -21 .] [本文引用: 2] [51] Zhang LH Zhang YW Zhao XN Huang SJ Zhao JM Yang YF 2014 ). Effects of different nutrient sources on plasticity of reproductive strategies in a monoecious species, Sagittaria graminea (Alismataceae)Journal of Systematics and Evolution 84 -91 . [本文引用: 1] [52] Zhang RZ Zheng D Yang QY Liu YH 1997 ). The Physical Geography of the Hengduan Mountains [本文引用: 1] [ 张荣祖 , 郑度 , 杨勤业 , 刘燕华 (1997 ). 横断山区自然地理. 科学出版社, 北京 .] [本文引用: 1] [53] Zhang S Ai HL Yu WB Wang H Li DZ 2010 ). Flower heliotropism of Anemone rivularis (Ranunculaceae) in the Himalayas: effects on floral temperature and reproductive fitnessPlant Ecology 301 -312 . [本文引用: 2] [54] Zhang ZJ He JS Li JS Tang ZY 2015 ). Distribution and conservation of threatened plants in ChinaBiological Conservation 454 -460 . [本文引用: 2] Community studies in pollination ecology in the high temperate Andes of central Chile. I. Pollination mechanisms and altitudinal variation
1
1982
... 此外, 团状福禄草具有性二态系统, 即雌性植株与两性植株同时存在于同一种群中.不同性别的植物个体获得繁殖适合度的方式不同, 雌性个体主要通过胚珠及种子产出获得繁殖适合度, 而两性植株主要通过花粉与种子产出获得繁殖适合度(
Charlesworth & Charlesworth, 1978 ;
Charlesworth & Morgan, 1991 ;
Ashman, 1994 ).因此, 不同性别的植株个体在繁殖分配策略上存在一定的差异性, 这种差异性直接反映了不同性别个体对特定环境因子的适应策略(
Case & Ashman, 2005 ;
Ashman, 2006 ;
Chen et al ., 2017b ).例如, 同样分布于滇西北高海拔地区, 且同样具有雌全异株性系统的细叶蓝钟花(
Cyananthus delavayi ), 在面对环境因子胁迫时, 雌性植株主要将资源分配到种子生产, 而两性植株则主要将资源分配到花粉生产以及对传粉昆虫的吸引功能方面(花瓣)(
Chen et al ., 2017b ).我们的研究结果表明, 总体而言, 两性植株对开花的资源分配比例要显著高于雌性植株(
图2B ), 但是, 其差异程度受到海拔因素的影响.具体而言, 低海拔种群两性植株分配给开花的资源比例显著高于雌性植株, 而高海拔种群不同性别植株对开花的资源分配差异性不显著(
图4 ).
Chen等(2017a) 研究发现, 在任何海拔条件下, 团状福禄草两性植株单位面积的开花数量始终大于雌性植株, 但其真实繁殖产出(单位面积果实和种子产量)却始终低于雌性植株.但是, 本研究发现, 尽管不同性别的个体之间在低海拔处的总繁殖分配比例存在差异性, 但是随着海拔升高, 环境胁迫程度增加, 这种差异性趋于消失(
图4 ).我们猜测, 在高海拔环境胁迫加剧条件下, 不同性别的植物个体在资源权衡分配方面采取了相同的策略.另一方面, 除了物理胁迫的加剧, 生物胁迫也同时加剧, 如, 传粉昆虫多样性及多度的下降(
Arroyo et al ., 1982 ;
Medan et al ., 2002 )可能引起两性植株雄性适合度输出(花粉传播)的降低, 从而引起其分配到开花的资源随之减少. ...
Reproductive allocation in hermaphrodite and female plants of
Sidalcea oregana ssp.
spicata (Malvaceae) using four currencies
3
1994
... 温度、资源(包括土壤养分、降水资源及太阳辐射量等)是限制高山植物生长与繁殖的关键因素(
Bloom et al ., 1985 ;
Chen et al ., 2017b ).为了应对高海拔地区贫瘠的土壤条件, 植物在自身资源获取及分配方面进化出了众多与环境相适应的资源分配策略.其中, 繁殖分配(即植物分配到繁殖器官的资源量)是植物适合度的关键表现(
Bazzaz et al ., 1987 ;
Weiner, 1988 ;
Hartnett, 1990 ;
Vega et al ., 2000 ).研究表明, 植物繁殖分配同时受到众多非生物与生物因子的控制(
Reekie & Bazzaz, 2005 ;
张婵等, 2020 ).非生物因子方面, 温度、水分、土壤营养等对植物的繁殖分配起着重要作用(
Reekie & Bazzaz, 2005 ;
Zhang et al ., 2014 ).而在生物因子方面, 种间竞争、传粉者特性以及植物自身性系统特征(
Ashman, 1994 ;
Case & Ashman, 2005 ;
Brzyski et al ., 2014 ;
Chen et al ., 2017a ,
2017b )对其繁殖分配策略都起着重要的影响.例如, 具有雌全异株性系统特征(即同一种植物同时存在雌性个体和两性个体)的植物, 其同一种群内的雌性个体与两性个体在繁殖分配策略上存在较大差异, 表现为雌性个体主要将资源分配到雌性功能(如胚珠及种子生产), 而两性个体则主要将资源分配到雄性功能(如花粉生产以及传粉者吸引)(
Ashman, 1994 ;
Chen et al ., 2017a ,
2017b ), 并且这种分配模式受到环境因子的强烈影响(
Chen et al ., 2017b ).为了应对资源的不足, 植物往往需要在营养分配(即分配到营养器官的资源量)与繁殖分配之间做出权衡, 以达到最优的繁殖适合度(
Reekie & Avila-Sakar, 2005 ).开花是植物繁殖的最初阶段, 植物开花的数目, 直接反映出植物在特定环境条件下的繁殖分配策略.另一方面, 开花数目的多少也可直接决定着植物未来的繁殖输出量(如果实及种子产量;
Reekie & Bazzaz, 2005 ). ...
... )对其繁殖分配策略都起着重要的影响.例如, 具有雌全异株性系统特征(即同一种植物同时存在雌性个体和两性个体)的植物, 其同一种群内的雌性个体与两性个体在繁殖分配策略上存在较大差异, 表现为雌性个体主要将资源分配到雌性功能(如胚珠及种子生产), 而两性个体则主要将资源分配到雄性功能(如花粉生产以及传粉者吸引)(
Ashman, 1994 ;
Chen et al ., 2017a ,
2017b ), 并且这种分配模式受到环境因子的强烈影响(
Chen et al ., 2017b ).为了应对资源的不足, 植物往往需要在营养分配(即分配到营养器官的资源量)与繁殖分配之间做出权衡, 以达到最优的繁殖适合度(
Reekie & Avila-Sakar, 2005 ).开花是植物繁殖的最初阶段, 植物开花的数目, 直接反映出植物在特定环境条件下的繁殖分配策略.另一方面, 开花数目的多少也可直接决定着植物未来的繁殖输出量(如果实及种子产量;
Reekie & Bazzaz, 2005 ). ...
... 此外, 团状福禄草具有性二态系统, 即雌性植株与两性植株同时存在于同一种群中.不同性别的植物个体获得繁殖适合度的方式不同, 雌性个体主要通过胚珠及种子产出获得繁殖适合度, 而两性植株主要通过花粉与种子产出获得繁殖适合度(
Charlesworth & Charlesworth, 1978 ;
Charlesworth & Morgan, 1991 ;
Ashman, 1994 ).因此, 不同性别的植株个体在繁殖分配策略上存在一定的差异性, 这种差异性直接反映了不同性别个体对特定环境因子的适应策略(
Case & Ashman, 2005 ;
Ashman, 2006 ;
Chen et al ., 2017b ).例如, 同样分布于滇西北高海拔地区, 且同样具有雌全异株性系统的细叶蓝钟花(
Cyananthus delavayi ), 在面对环境因子胁迫时, 雌性植株主要将资源分配到种子生产, 而两性植株则主要将资源分配到花粉生产以及对传粉昆虫的吸引功能方面(花瓣)(
Chen et al ., 2017b ).我们的研究结果表明, 总体而言, 两性植株对开花的资源分配比例要显著高于雌性植株(
图2B ), 但是, 其差异程度受到海拔因素的影响.具体而言, 低海拔种群两性植株分配给开花的资源比例显著高于雌性植株, 而高海拔种群不同性别植株对开花的资源分配差异性不显著(
图4 ).
Chen等(2017a) 研究发现, 在任何海拔条件下, 团状福禄草两性植株单位面积的开花数量始终大于雌性植株, 但其真实繁殖产出(单位面积果实和种子产量)却始终低于雌性植株.但是, 本研究发现, 尽管不同性别的个体之间在低海拔处的总繁殖分配比例存在差异性, 但是随着海拔升高, 环境胁迫程度增加, 这种差异性趋于消失(
图4 ).我们猜测, 在高海拔环境胁迫加剧条件下, 不同性别的植物个体在资源权衡分配方面采取了相同的策略.另一方面, 除了物理胁迫的加剧, 生物胁迫也同时加剧, 如, 传粉昆虫多样性及多度的下降(
Arroyo et al ., 1982 ;
Medan et al ., 2002 )可能引起两性植株雄性适合度输出(花粉传播)的降低, 从而引起其分配到开花的资源随之减少. ...
The evolution of separate sexes: a focus on the ecological context//Harder LD, Barrett SCH
1
2006
... 此外, 团状福禄草具有性二态系统, 即雌性植株与两性植株同时存在于同一种群中.不同性别的植物个体获得繁殖适合度的方式不同, 雌性个体主要通过胚珠及种子产出获得繁殖适合度, 而两性植株主要通过花粉与种子产出获得繁殖适合度(
Charlesworth & Charlesworth, 1978 ;
Charlesworth & Morgan, 1991 ;
Ashman, 1994 ).因此, 不同性别的植株个体在繁殖分配策略上存在一定的差异性, 这种差异性直接反映了不同性别个体对特定环境因子的适应策略(
Case & Ashman, 2005 ;
Ashman, 2006 ;
Chen et al ., 2017b ).例如, 同样分布于滇西北高海拔地区, 且同样具有雌全异株性系统的细叶蓝钟花(
Cyananthus delavayi ), 在面对环境因子胁迫时, 雌性植株主要将资源分配到种子生产, 而两性植株则主要将资源分配到花粉生产以及对传粉昆虫的吸引功能方面(花瓣)(
Chen et al ., 2017b ).我们的研究结果表明, 总体而言, 两性植株对开花的资源分配比例要显著高于雌性植株(
图2B ), 但是, 其差异程度受到海拔因素的影响.具体而言, 低海拔种群两性植株分配给开花的资源比例显著高于雌性植株, 而高海拔种群不同性别植株对开花的资源分配差异性不显著(
图4 ).
Chen等(2017a) 研究发现, 在任何海拔条件下, 团状福禄草两性植株单位面积的开花数量始终大于雌性植株, 但其真实繁殖产出(单位面积果实和种子产量)却始终低于雌性植株.但是, 本研究发现, 尽管不同性别的个体之间在低海拔处的总繁殖分配比例存在差异性, 但是随着海拔升高, 环境胁迫程度增加, 这种差异性趋于消失(
图4 ).我们猜测, 在高海拔环境胁迫加剧条件下, 不同性别的植物个体在资源权衡分配方面采取了相同的策略.另一方面, 除了物理胁迫的加剧, 生物胁迫也同时加剧, 如, 传粉昆虫多样性及多度的下降(
Arroyo et al ., 1982 ;
Medan et al ., 2002 )可能引起两性植株雄性适合度输出(花粉传播)的降低, 从而引起其分配到开花的资源随之减少. ...
1914-2014: a revised worldwide catalogue of cushion plants 100 years after Hauri and Schr?ter
1
2014
... 垫状植物是一类具有球形或半球形结构的植物的总称, 是植物经过多年辐射性生长而形成的小枝紧密簇生结构(
图1 ;
Gibson & Hope, 1986 ), 因其适应世界上大多数山地和极地极端干旱和低温环境的能力而备受植物学家的关注(
Aubert et al ., 2014 ).并因其具有生态系统工程师的功能, 对其他植物具有“护理作用”, 可提高和维持生态系统中物种多样性(
Butterfield et al ., 2013 ;
Cavieres et al ., 2014 ;
Chen et al ., 2015a ,
2015b ,
2019 ), 而被认为是所在生态系统当中的奠基种(foundation species;
Kikvidze et al ., 2015 ). ...
1
2019
... 首先, 将各方向上开花网格数相加, 得到各植株的总开花网格(面积), 其次, 分别将总开花网格数和每个方向的开花网格数都除以总网格数, 得到各个植株总的开花面积比(为了论述方便, 本研究中, 必要时将开花面积比称为“繁殖分配”)以及单个方位上的开花面积比, 并采用这些数据进行后续的比较分析.不同植株的大小(年龄)会对其资源分配策略产生影响, 即可能存在个体效应(
St?cklin & Favre, 1994 ;
Sakai & Sakai, 1995 ), 同时, 垫状植物冠层表面积大小通常与其年龄呈正相关关系(
Chen et al ., 2017a ), 因此, 计数了每个调查个体的总网格数(即表面积大小), 将其作为植株大小的替代来进行后续分析.首先, 为了探究不同种群以及不同性别植株的总开花面积与其个体大小之间的关系, 对植株总开花面积与其个体大小做了回归分析.其次, 为了探究海拔对植株大小的影响, 采用混合效应模型比较了不同海拔种群间植株的大小, 植株大小为因变量, 海拔为主因素, 植株个体编号为随机因素.最后, 为了探究开花面积比(即繁殖分配)如何受到海拔、性别和植株大小的影响, 使用混合效应模型对上述关系进行拟合.模型中, 开花面积比为因变量, 海拔、性别和植株大小为主因素, 植株个体编号为随机因素.此外, 由于自变量与因变量间存在多样的组合形式, 基于赤池信息量准则(AIC)权重选择最优模型, 最优模型具有最大的AIC权重, 因子在每个模型的AIC权重被用来计算模型平均系数, 而每个因子在所有模型中的AIC值权重之和(
w )被用来描述该因子的统计支持.同时, 评估了考虑交互作用的模型与上述模型的优劣, 校准AIC (
AICc )结果显示两种模型没有差异(
AICc = -402.1 vs.
AICc = -401.4), 但是交互作用不显著, 因而以上述的模型结果为准.由于各组数据不完全符合正态分布及方差齐性原则, 使用Kruskal-Wallis非参数检验分别比较同一种群内不同性别、不同方位之间开花面积比以及不同种群之间开花面积比的差异.所有统计分析及画图都通过R 3.5.1 (
R Core Team, 2018 )实现, Kruskal-Wallis非参数检验使用“agricolae”包(
de Felipe, 2019 ), 混合效应模型使用“coxme”包(
Therneau, 2018 ), 模型选择使用“MuMIn”包(
Bartoń, 2019 ). ...
Allocating resources to reproduction and defense
1
1987
... 温度、资源(包括土壤养分、降水资源及太阳辐射量等)是限制高山植物生长与繁殖的关键因素(
Bloom et al ., 1985 ;
Chen et al ., 2017b ).为了应对高海拔地区贫瘠的土壤条件, 植物在自身资源获取及分配方面进化出了众多与环境相适应的资源分配策略.其中, 繁殖分配(即植物分配到繁殖器官的资源量)是植物适合度的关键表现(
Bazzaz et al ., 1987 ;
Weiner, 1988 ;
Hartnett, 1990 ;
Vega et al ., 2000 ).研究表明, 植物繁殖分配同时受到众多非生物与生物因子的控制(
Reekie & Bazzaz, 2005 ;
张婵等, 2020 ).非生物因子方面, 温度、水分、土壤营养等对植物的繁殖分配起着重要作用(
Reekie & Bazzaz, 2005 ;
Zhang et al ., 2014 ).而在生物因子方面, 种间竞争、传粉者特性以及植物自身性系统特征(
Ashman, 1994 ;
Case & Ashman, 2005 ;
Brzyski et al ., 2014 ;
Chen et al ., 2017a ,
2017b )对其繁殖分配策略都起着重要的影响.例如, 具有雌全异株性系统特征(即同一种植物同时存在雌性个体和两性个体)的植物, 其同一种群内的雌性个体与两性个体在繁殖分配策略上存在较大差异, 表现为雌性个体主要将资源分配到雌性功能(如胚珠及种子生产), 而两性个体则主要将资源分配到雄性功能(如花粉生产以及传粉者吸引)(
Ashman, 1994 ;
Chen et al ., 2017a ,
2017b ), 并且这种分配模式受到环境因子的强烈影响(
Chen et al ., 2017b ).为了应对资源的不足, 植物往往需要在营养分配(即分配到营养器官的资源量)与繁殖分配之间做出权衡, 以达到最优的繁殖适合度(
Reekie & Avila-Sakar, 2005 ).开花是植物繁殖的最初阶段, 植物开花的数目, 直接反映出植物在特定环境条件下的繁殖分配策略.另一方面, 开花数目的多少也可直接决定着植物未来的繁殖输出量(如果实及种子产量;
Reekie & Bazzaz, 2005 ). ...
Resource limitation in plants
— An economic analogy
1
1985
... 温度、资源(包括土壤养分、降水资源及太阳辐射量等)是限制高山植物生长与繁殖的关键因素(
Bloom et al ., 1985 ;
Chen et al ., 2017b ).为了应对高海拔地区贫瘠的土壤条件, 植物在自身资源获取及分配方面进化出了众多与环境相适应的资源分配策略.其中, 繁殖分配(即植物分配到繁殖器官的资源量)是植物适合度的关键表现(
Bazzaz et al ., 1987 ;
Weiner, 1988 ;
Hartnett, 1990 ;
Vega et al ., 2000 ).研究表明, 植物繁殖分配同时受到众多非生物与生物因子的控制(
Reekie & Bazzaz, 2005 ;
张婵等, 2020 ).非生物因子方面, 温度、水分、土壤营养等对植物的繁殖分配起着重要作用(
Reekie & Bazzaz, 2005 ;
Zhang et al ., 2014 ).而在生物因子方面, 种间竞争、传粉者特性以及植物自身性系统特征(
Ashman, 1994 ;
Case & Ashman, 2005 ;
Brzyski et al ., 2014 ;
Chen et al ., 2017a ,
2017b )对其繁殖分配策略都起着重要的影响.例如, 具有雌全异株性系统特征(即同一种植物同时存在雌性个体和两性个体)的植物, 其同一种群内的雌性个体与两性个体在繁殖分配策略上存在较大差异, 表现为雌性个体主要将资源分配到雌性功能(如胚珠及种子生产), 而两性个体则主要将资源分配到雄性功能(如花粉生产以及传粉者吸引)(
Ashman, 1994 ;
Chen et al ., 2017a ,
2017b ), 并且这种分配模式受到环境因子的强烈影响(
Chen et al ., 2017b ).为了应对资源的不足, 植物往往需要在营养分配(即分配到营养器官的资源量)与繁殖分配之间做出权衡, 以达到最优的繁殖适合度(
Reekie & Avila-Sakar, 2005 ).开花是植物繁殖的最初阶段, 植物开花的数目, 直接反映出植物在特定环境条件下的繁殖分配策略.另一方面, 开花数目的多少也可直接决定着植物未来的繁殖输出量(如果实及种子产量;
Reekie & Bazzaz, 2005 ). ...
Soil erodibility as determined by raindrop technique
1
1975
... 前期研究发现, 团状福禄草植物个体表面单位面积的果实及种子产量随海拔上升而呈现总体下降趋势(
Chen et al ., 2017a ).但是, 该研究并未考虑团状福禄草植物整体的繁殖分配对环境因子的响应, 而仅是比较了其特定开花区域内单位面积的繁殖产出.本研究结果显示, 团状福禄草在开花阶段的繁殖分配在不同海拔种群间存在显著差异, 总体上随海拔上升而减少, 表明高海拔更加贫瘠的土壤条件(
Chen et al ., 2015b ,
2019 )可能限制了植物总体的资源获取量, 植物需要将有限的资源在营养生长与繁殖生长两方面之间做权衡(
Reekie & Avila-Sakar, 2005 ), 从而导致其在繁殖功能上的总体分配比例的下降.但是, 我们也发现, 部分低海拔种群(种群2)内植物个体的繁殖分配显著低于部分高海拔种群(种群3、4), 这表明, 海拔并非控制植物繁殖分配的唯一因素, 植物繁殖分配是多种生态因子共同作用的结果(
张婵等, 2020 ).种群2与种群1海拔相差较小, 但坡度相差较大, 其与种群3坡度相近, 但具有不同坡向(
表1 ).因此, 我们推测除了海拔因素, 坡度与坡向可能同时对其繁殖分配产生影响.一方面坡度大可能导致土壤养分流失更严重(
Bruce-Okine & Lal, 1975 ;
Donker & Damen, 1984 ), 从而引起资源胁迫的加剧, 并进而导致繁殖分配的减少.另一方面, 坡度和坡向通常会引起太阳辐射特征的变化, 如太阳入射角、入射时间在不同坡度和坡向上会发生变化(
田辉等, 2007 ), 从而引起植物可积累和利用的有效太阳辐射能量的变化.在高海拔地区, 太阳辐射对植物的生长与繁殖策略都起着重要的影响.如, 为了充分利用太阳辐射, 一些植物花器官进化出了“向日性”功能(
Zhang et al ., 2010 ).又如, 安第斯山脉中部高海拔岩石斜坡典型垫状植物
Azorella compacta 为了最大化利用太阳辐射, 其个体偏爱生长在向北的(南半球)坡度约为20°的斜坡(
Kleier et al ., 2015 ).因此, 我们推测由坡度与坡向变化引起的土壤养分状况及太阳辐射特征的变化, 可能同时对团状福禄草的繁殖分配起着重要的影响. ...
Reproductive allocation between the sexes, across natural and novel habitats, and its impact on genetic diversity
1
2014
... 温度、资源(包括土壤养分、降水资源及太阳辐射量等)是限制高山植物生长与繁殖的关键因素(
Bloom et al ., 1985 ;
Chen et al ., 2017b ).为了应对高海拔地区贫瘠的土壤条件, 植物在自身资源获取及分配方面进化出了众多与环境相适应的资源分配策略.其中, 繁殖分配(即植物分配到繁殖器官的资源量)是植物适合度的关键表现(
Bazzaz et al ., 1987 ;
Weiner, 1988 ;
Hartnett, 1990 ;
Vega et al ., 2000 ).研究表明, 植物繁殖分配同时受到众多非生物与生物因子的控制(
Reekie & Bazzaz, 2005 ;
张婵等, 2020 ).非生物因子方面, 温度、水分、土壤营养等对植物的繁殖分配起着重要作用(
Reekie & Bazzaz, 2005 ;
Zhang et al ., 2014 ).而在生物因子方面, 种间竞争、传粉者特性以及植物自身性系统特征(
Ashman, 1994 ;
Case & Ashman, 2005 ;
Brzyski et al ., 2014 ;
Chen et al ., 2017a ,
2017b )对其繁殖分配策略都起着重要的影响.例如, 具有雌全异株性系统特征(即同一种植物同时存在雌性个体和两性个体)的植物, 其同一种群内的雌性个体与两性个体在繁殖分配策略上存在较大差异, 表现为雌性个体主要将资源分配到雌性功能(如胚珠及种子生产), 而两性个体则主要将资源分配到雄性功能(如花粉生产以及传粉者吸引)(
Ashman, 1994 ;
Chen et al ., 2017a ,
2017b ), 并且这种分配模式受到环境因子的强烈影响(
Chen et al ., 2017b ).为了应对资源的不足, 植物往往需要在营养分配(即分配到营养器官的资源量)与繁殖分配之间做出权衡, 以达到最优的繁殖适合度(
Reekie & Avila-Sakar, 2005 ).开花是植物繁殖的最初阶段, 植物开花的数目, 直接反映出植物在特定环境条件下的繁殖分配策略.另一方面, 开花数目的多少也可直接决定着植物未来的繁殖输出量(如果实及种子产量;
Reekie & Bazzaz, 2005 ). ...
Alpine cushion plants inhibit the loss of phylogenetic diversity in severe environments
1
2013
... 垫状植物是一类具有球形或半球形结构的植物的总称, 是植物经过多年辐射性生长而形成的小枝紧密簇生结构(
图1 ;
Gibson & Hope, 1986 ), 因其适应世界上大多数山地和极地极端干旱和低温环境的能力而备受植物学家的关注(
Aubert et al ., 2014 ).并因其具有生态系统工程师的功能, 对其他植物具有“护理作用”, 可提高和维持生态系统中物种多样性(
Butterfield et al ., 2013 ;
Cavieres et al ., 2014 ;
Chen et al ., 2015a ,
2015b ,
2019 ), 而被认为是所在生态系统当中的奠基种(foundation species;
Kikvidze et al ., 2015 ). ...
藏东南色季拉山东西坡不同海拔高度土壤养分含量分布特征
2
2019
... 随着海拔逐渐上升, 环境条件变得更加严酷, 如: 海拔每上升100 m, 气温约下降0.6 ℃, 气压下降8-9 hPa (
K?rner, 2003 ), 土壤养分含量也随之减少(
曹舰艇等, 2019 ).面对高海拔地区低温、土壤养分贫瘠等恶劣环境因子, 高山植物在长期的进化过程中形成了丰富多样的特化适应性状或进化策略(
Sun et al ., 2014 ;
杨扬等, 2019 ), 种类丰富的植物多样性对维持高山生态系统的稳定性起到了关键作用(
Hooper et al ., 2005 ).滇西北处于横断山高山植物多样性中心区域(
郭辉军和龙春林, 1998 ;
Zhang et al. , 2015 ).长期以来, 青藏高原及横断山高海拔地区植物对极端环境因子的适应机制, 备受植物学家和生态学家的关注, 相关研究的深入开展, 加深了对该地区植物多样性形成及其进化适应机制的认识(
Sun et al ., 2014 ;
杨扬等, 2019 ). ...
... 横断山高海拔地区绝大多数植物种群的更新与扩张都依赖有性繁殖(
Peng et al ., 2012 ).植物对繁殖功能的资源分配, 在很大程度上决定了其种群未来的扩张速率.而作为植物实现繁殖功能的重要环节, 植物对开花的资源配置决定了植物的最终繁殖产出(
Reekie & Bazzaz, 2005 ).不同的环境因子在植物的每一个繁殖阶段都起着重要作用, 甚至一些因子(如资源量)直接决定着植物的繁殖分配及其产出(
Delph & Carroll, 2001 ).在我们的研究区域内, 环境胁迫程度(如土壤养分胁迫及低温胁迫)随海拔升高而增强(
Chen et al ., 2015b ,
2019 ).我们的研究结果显示, 随着海拔升高, 种群内植株个体呈现减小的趋势.植株大小对开花特性具有显著影响.一方面, 植株个体越大, 其总开花面积也越大, 但另一方面, 植株越大, 其总开花面积比越小(
表2 ), 并且这种趋势在不同性别植株间具有高度一致性(
图3 ), 说明资源限制是真实存在的, 即, 植物从环境中获取并分配给开花的资源量有限.此外, 该结果也说明植株分配到开花的资源增长速率可能低于植株个体的增长速率, 即繁殖分配的增加速率低于营养分配的增加速率.相应结果进一步暗示, 在面临资源限制或环境胁迫时, 团状福禄草的资源分配策略以保证植株存活(即增加营养分配)为首要目的.此外, 具有不同坡度及坡向的种群间开花特性也存在一定的变化,说明由坡向及坡度变化而引起的环境因子变化, 如太阳辐射(
田辉等, 2007 )和土壤养分(
曹舰艇等, 2019 ), 可能也同时对其繁殖分配策略具有一定的影响. ...
藏东南色季拉山东西坡不同海拔高度土壤养分含量分布特征
2
2019
... 随着海拔逐渐上升, 环境条件变得更加严酷, 如: 海拔每上升100 m, 气温约下降0.6 ℃, 气压下降8-9 hPa (
K?rner, 2003 ), 土壤养分含量也随之减少(
曹舰艇等, 2019 ).面对高海拔地区低温、土壤养分贫瘠等恶劣环境因子, 高山植物在长期的进化过程中形成了丰富多样的特化适应性状或进化策略(
Sun et al ., 2014 ;
杨扬等, 2019 ), 种类丰富的植物多样性对维持高山生态系统的稳定性起到了关键作用(
Hooper et al ., 2005 ).滇西北处于横断山高山植物多样性中心区域(
郭辉军和龙春林, 1998 ;
Zhang et al. , 2015 ).长期以来, 青藏高原及横断山高海拔地区植物对极端环境因子的适应机制, 备受植物学家和生态学家的关注, 相关研究的深入开展, 加深了对该地区植物多样性形成及其进化适应机制的认识(
Sun et al ., 2014 ;
杨扬等, 2019 ). ...
... 横断山高海拔地区绝大多数植物种群的更新与扩张都依赖有性繁殖(
Peng et al ., 2012 ).植物对繁殖功能的资源分配, 在很大程度上决定了其种群未来的扩张速率.而作为植物实现繁殖功能的重要环节, 植物对开花的资源配置决定了植物的最终繁殖产出(
Reekie & Bazzaz, 2005 ).不同的环境因子在植物的每一个繁殖阶段都起着重要作用, 甚至一些因子(如资源量)直接决定着植物的繁殖分配及其产出(
Delph & Carroll, 2001 ).在我们的研究区域内, 环境胁迫程度(如土壤养分胁迫及低温胁迫)随海拔升高而增强(
Chen et al ., 2015b ,
2019 ).我们的研究结果显示, 随着海拔升高, 种群内植株个体呈现减小的趋势.植株大小对开花特性具有显著影响.一方面, 植株个体越大, 其总开花面积也越大, 但另一方面, 植株越大, 其总开花面积比越小(
表2 ), 并且这种趋势在不同性别植株间具有高度一致性(
图3 ), 说明资源限制是真实存在的, 即, 植物从环境中获取并分配给开花的资源量有限.此外, 该结果也说明植株分配到开花的资源增长速率可能低于植株个体的增长速率, 即繁殖分配的增加速率低于营养分配的增加速率.相应结果进一步暗示, 在面临资源限制或环境胁迫时, 团状福禄草的资源分配策略以保证植株存活(即增加营养分配)为首要目的.此外, 具有不同坡度及坡向的种群间开花特性也存在一定的变化,说明由坡向及坡度变化而引起的环境因子变化, 如太阳辐射(
田辉等, 2007 )和土壤养分(
曹舰艇等, 2019 ), 可能也同时对其繁殖分配策略具有一定的影响. ...
Sex-specific physiology and its implications for the cost of reproduction//Reekie E, Bazzaz FA
2
2005
... 温度、资源(包括土壤养分、降水资源及太阳辐射量等)是限制高山植物生长与繁殖的关键因素(
Bloom et al ., 1985 ;
Chen et al ., 2017b ).为了应对高海拔地区贫瘠的土壤条件, 植物在自身资源获取及分配方面进化出了众多与环境相适应的资源分配策略.其中, 繁殖分配(即植物分配到繁殖器官的资源量)是植物适合度的关键表现(
Bazzaz et al ., 1987 ;
Weiner, 1988 ;
Hartnett, 1990 ;
Vega et al ., 2000 ).研究表明, 植物繁殖分配同时受到众多非生物与生物因子的控制(
Reekie & Bazzaz, 2005 ;
张婵等, 2020 ).非生物因子方面, 温度、水分、土壤营养等对植物的繁殖分配起着重要作用(
Reekie & Bazzaz, 2005 ;
Zhang et al ., 2014 ).而在生物因子方面, 种间竞争、传粉者特性以及植物自身性系统特征(
Ashman, 1994 ;
Case & Ashman, 2005 ;
Brzyski et al ., 2014 ;
Chen et al ., 2017a ,
2017b )对其繁殖分配策略都起着重要的影响.例如, 具有雌全异株性系统特征(即同一种植物同时存在雌性个体和两性个体)的植物, 其同一种群内的雌性个体与两性个体在繁殖分配策略上存在较大差异, 表现为雌性个体主要将资源分配到雌性功能(如胚珠及种子生产), 而两性个体则主要将资源分配到雄性功能(如花粉生产以及传粉者吸引)(
Ashman, 1994 ;
Chen et al ., 2017a ,
2017b ), 并且这种分配模式受到环境因子的强烈影响(
Chen et al ., 2017b ).为了应对资源的不足, 植物往往需要在营养分配(即分配到营养器官的资源量)与繁殖分配之间做出权衡, 以达到最优的繁殖适合度(
Reekie & Avila-Sakar, 2005 ).开花是植物繁殖的最初阶段, 植物开花的数目, 直接反映出植物在特定环境条件下的繁殖分配策略.另一方面, 开花数目的多少也可直接决定着植物未来的繁殖输出量(如果实及种子产量;
Reekie & Bazzaz, 2005 ). ...
... 此外, 团状福禄草具有性二态系统, 即雌性植株与两性植株同时存在于同一种群中.不同性别的植物个体获得繁殖适合度的方式不同, 雌性个体主要通过胚珠及种子产出获得繁殖适合度, 而两性植株主要通过花粉与种子产出获得繁殖适合度(
Charlesworth & Charlesworth, 1978 ;
Charlesworth & Morgan, 1991 ;
Ashman, 1994 ).因此, 不同性别的植株个体在繁殖分配策略上存在一定的差异性, 这种差异性直接反映了不同性别个体对特定环境因子的适应策略(
Case & Ashman, 2005 ;
Ashman, 2006 ;
Chen et al ., 2017b ).例如, 同样分布于滇西北高海拔地区, 且同样具有雌全异株性系统的细叶蓝钟花(
Cyananthus delavayi ), 在面对环境因子胁迫时, 雌性植株主要将资源分配到种子生产, 而两性植株则主要将资源分配到花粉生产以及对传粉昆虫的吸引功能方面(花瓣)(
Chen et al ., 2017b ).我们的研究结果表明, 总体而言, 两性植株对开花的资源分配比例要显著高于雌性植株(
图2B ), 但是, 其差异程度受到海拔因素的影响.具体而言, 低海拔种群两性植株分配给开花的资源比例显著高于雌性植株, 而高海拔种群不同性别植株对开花的资源分配差异性不显著(
图4 ).
Chen等(2017a) 研究发现, 在任何海拔条件下, 团状福禄草两性植株单位面积的开花数量始终大于雌性植株, 但其真实繁殖产出(单位面积果实和种子产量)却始终低于雌性植株.但是, 本研究发现, 尽管不同性别的个体之间在低海拔处的总繁殖分配比例存在差异性, 但是随着海拔升高, 环境胁迫程度增加, 这种差异性趋于消失(
图4 ).我们猜测, 在高海拔环境胁迫加剧条件下, 不同性别的植物个体在资源权衡分配方面采取了相同的策略.另一方面, 除了物理胁迫的加剧, 生物胁迫也同时加剧, 如, 传粉昆虫多样性及多度的下降(
Arroyo et al ., 1982 ;
Medan et al ., 2002 )可能引起两性植株雄性适合度输出(花粉传播)的降低, 从而引起其分配到开花的资源随之减少. ...
Facilitative plant interactions and climate simultaneously drive alpine plant diversity
1
2014
... 垫状植物是一类具有球形或半球形结构的植物的总称, 是植物经过多年辐射性生长而形成的小枝紧密簇生结构(
图1 ;
Gibson & Hope, 1986 ), 因其适应世界上大多数山地和极地极端干旱和低温环境的能力而备受植物学家的关注(
Aubert et al ., 2014 ).并因其具有生态系统工程师的功能, 对其他植物具有“护理作用”, 可提高和维持生态系统中物种多样性(
Butterfield et al ., 2013 ;
Cavieres et al ., 2014 ;
Chen et al ., 2015a ,
2015b ,
2019 ), 而被认为是所在生态系统当中的奠基种(foundation species;
Kikvidze et al ., 2015 ). ...
A model for the evolution of dioecy and gynodioecy
1
1978
... 此外, 团状福禄草具有性二态系统, 即雌性植株与两性植株同时存在于同一种群中.不同性别的植物个体获得繁殖适合度的方式不同, 雌性个体主要通过胚珠及种子产出获得繁殖适合度, 而两性植株主要通过花粉与种子产出获得繁殖适合度(
Charlesworth & Charlesworth, 1978 ;
Charlesworth & Morgan, 1991 ;
Ashman, 1994 ).因此, 不同性别的植株个体在繁殖分配策略上存在一定的差异性, 这种差异性直接反映了不同性别个体对特定环境因子的适应策略(
Case & Ashman, 2005 ;
Ashman, 2006 ;
Chen et al ., 2017b ).例如, 同样分布于滇西北高海拔地区, 且同样具有雌全异株性系统的细叶蓝钟花(
Cyananthus delavayi ), 在面对环境因子胁迫时, 雌性植株主要将资源分配到种子生产, 而两性植株则主要将资源分配到花粉生产以及对传粉昆虫的吸引功能方面(花瓣)(
Chen et al ., 2017b ).我们的研究结果表明, 总体而言, 两性植株对开花的资源分配比例要显著高于雌性植株(
图2B ), 但是, 其差异程度受到海拔因素的影响.具体而言, 低海拔种群两性植株分配给开花的资源比例显著高于雌性植株, 而高海拔种群不同性别植株对开花的资源分配差异性不显著(
图4 ).
Chen等(2017a) 研究发现, 在任何海拔条件下, 团状福禄草两性植株单位面积的开花数量始终大于雌性植株, 但其真实繁殖产出(单位面积果实和种子产量)却始终低于雌性植株.但是, 本研究发现, 尽管不同性别的个体之间在低海拔处的总繁殖分配比例存在差异性, 但是随着海拔升高, 环境胁迫程度增加, 这种差异性趋于消失(
图4 ).我们猜测, 在高海拔环境胁迫加剧条件下, 不同性别的植物个体在资源权衡分配方面采取了相同的策略.另一方面, 除了物理胁迫的加剧, 生物胁迫也同时加剧, 如, 传粉昆虫多样性及多度的下降(
Arroyo et al ., 1982 ;
Medan et al ., 2002 )可能引起两性植株雄性适合度输出(花粉传播)的降低, 从而引起其分配到开花的资源随之减少. ...
Allocation of resources to sex functions in flowering plants
1
1991
... 此外, 团状福禄草具有性二态系统, 即雌性植株与两性植株同时存在于同一种群中.不同性别的植物个体获得繁殖适合度的方式不同, 雌性个体主要通过胚珠及种子产出获得繁殖适合度, 而两性植株主要通过花粉与种子产出获得繁殖适合度(
Charlesworth & Charlesworth, 1978 ;
Charlesworth & Morgan, 1991 ;
Ashman, 1994 ).因此, 不同性别的植株个体在繁殖分配策略上存在一定的差异性, 这种差异性直接反映了不同性别个体对特定环境因子的适应策略(
Case & Ashman, 2005 ;
Ashman, 2006 ;
Chen et al ., 2017b ).例如, 同样分布于滇西北高海拔地区, 且同样具有雌全异株性系统的细叶蓝钟花(
Cyananthus delavayi ), 在面对环境因子胁迫时, 雌性植株主要将资源分配到种子生产, 而两性植株则主要将资源分配到花粉生产以及对传粉昆虫的吸引功能方面(花瓣)(
Chen et al ., 2017b ).我们的研究结果表明, 总体而言, 两性植株对开花的资源分配比例要显著高于雌性植株(
图2B ), 但是, 其差异程度受到海拔因素的影响.具体而言, 低海拔种群两性植株分配给开花的资源比例显著高于雌性植株, 而高海拔种群不同性别植株对开花的资源分配差异性不显著(
图4 ).
Chen等(2017a) 研究发现, 在任何海拔条件下, 团状福禄草两性植株单位面积的开花数量始终大于雌性植株, 但其真实繁殖产出(单位面积果实和种子产量)却始终低于雌性植株.但是, 本研究发现, 尽管不同性别的个体之间在低海拔处的总繁殖分配比例存在差异性, 但是随着海拔升高, 环境胁迫程度增加, 这种差异性趋于消失(
图4 ).我们猜测, 在高海拔环境胁迫加剧条件下, 不同性别的植物个体在资源权衡分配方面采取了相同的策略.另一方面, 除了物理胁迫的加剧, 生物胁迫也同时加剧, 如, 传粉昆虫多样性及多度的下降(
Arroyo et al ., 1982 ;
Medan et al ., 2002 )可能引起两性植株雄性适合度输出(花粉传播)的降低, 从而引起其分配到开花的资源随之减少. ...
Cushion and shrub ecosystem engineers contribute differently to diversity and functions in alpine ecosystems
6
2019
... 垫状植物是一类具有球形或半球形结构的植物的总称, 是植物经过多年辐射性生长而形成的小枝紧密簇生结构(
图1 ;
Gibson & Hope, 1986 ), 因其适应世界上大多数山地和极地极端干旱和低温环境的能力而备受植物学家的关注(
Aubert et al ., 2014 ).并因其具有生态系统工程师的功能, 对其他植物具有“护理作用”, 可提高和维持生态系统中物种多样性(
Butterfield et al ., 2013 ;
Cavieres et al ., 2014 ;
Chen et al ., 2015a ,
2015b ,
2019 ), 而被认为是所在生态系统当中的奠基种(foundation species;
Kikvidze et al ., 2015 ). ...
... 团状福禄草(
Arenaria polytrichoides )为石竹科无心菜属(
Arenaria )多年生垫状草本植物, 是青藏高原特有种, 沿喜马拉雅—横断山高海拔(3 500-5 300 m)地区分布(
Wu et al ., 2013 ).营养生长自冰雪消融开始, 花期主要在6-7月, 不同分布区域及海拔的种群的花期存在一定的差异(
Chen et al ., 2017a ).并且, 团状福禄草具有雌全异株的性二态系统, 即同一种群中同时存在雌性植株和两性植株, 不同性别的个体在繁殖产出(果实及种子产量)方面存在显著差异, 并且受到海拔的影响(
Chen et al ., 2017a ).表现为较高海拔的雌性比例高于较低海拔的, 两性植株开花数目大于雌性植株但结实率显著低于雌性植株, 同时, 较低海拔种群内个体单位面积的果实与种子产量高于较高海拔种群的个体(
Chen et al ., 2017a ).此外, 团状福禄草被证实为横断山高山生态系统当中的关键种, 能有效改良微环境条件(温度、土壤营养及水分), 从而为其他低耐受物种提供适宜的微生境, 进而调节种群动态、提升生态系统物种多样性及地面生产力(
Yang et al ., 2010 ,
2017 ;
Chen et al ., 2015a ,
2019 ).然而, 团状福禄草种群的更新却受到种间竞争的强烈影响(
Chen et al ., 2020 ).研究者在近十年内对团状福禄草做了深入调查与研究(
Yang et al ., 2010 ,
2017 ;
Chen et al ., 2015a ,
2015b ,
2017a ,
2019 ), 但对其种群的繁殖与更新机制, 却直到最近才予以关注(
Chen et al ., 2017a ,
2020 ).理论上讲, 团状福禄草的每一个小分枝(
图1C )在生长季节都能开出一朵花, 然而, 我们在多年的野外研究中发现, 实际上, 其每年的开花数量极为有限, 只在其冠层表面的部分区域开出一定面积的花(
图1A 、1B).这一现象说明, 垫状植物的开花可能受到了资源的限制, 并有可能同时受到其他环境因子的影响.例如, 据野外观察, 团状福禄草开花方位(只在其冠层表面特定区域开放,
图1A 、1B)呈现出一定的规律性, 且在不同的种群之间存在差异.为了获取充足的太阳辐射能量从而满足自身繁殖需求, 一些高山植物的开花特性受到太阳辐射的强烈影响(
Pati?o et al ., 2002 ;
Galen, 2006 ;
Zhang et al ., 2010 ), 因此, 我们猜测团状福禄草的开花数量和方位可能受到资源和太阳辐射的双重影响.但其繁殖分配策略, 即个体对开花的资源分配策略对环境胁迫因子的响应方式, 目前却不为人知.深入探究高山生态系统中这一关键类群的种群更新与维持机制, 对揭示其种群繁殖的初始阶段(开花特性)对环境因子的响应机制以及这种响应机制在不同性别个体之间的差异性具有重要意义. ...
... ,
2019 ), 但对其种群的繁殖与更新机制, 却直到最近才予以关注(
Chen et al ., 2017a ,
2020 ).理论上讲, 团状福禄草的每一个小分枝(
图1C )在生长季节都能开出一朵花, 然而, 我们在多年的野外研究中发现, 实际上, 其每年的开花数量极为有限, 只在其冠层表面的部分区域开出一定面积的花(
图1A 、1B).这一现象说明, 垫状植物的开花可能受到了资源的限制, 并有可能同时受到其他环境因子的影响.例如, 据野外观察, 团状福禄草开花方位(只在其冠层表面特定区域开放,
图1A 、1B)呈现出一定的规律性, 且在不同的种群之间存在差异.为了获取充足的太阳辐射能量从而满足自身繁殖需求, 一些高山植物的开花特性受到太阳辐射的强烈影响(
Pati?o et al ., 2002 ;
Galen, 2006 ;
Zhang et al ., 2010 ), 因此, 我们猜测团状福禄草的开花数量和方位可能受到资源和太阳辐射的双重影响.但其繁殖分配策略, 即个体对开花的资源分配策略对环境胁迫因子的响应方式, 目前却不为人知.深入探究高山生态系统中这一关键类群的种群更新与维持机制, 对揭示其种群繁殖的初始阶段(开花特性)对环境因子的响应机制以及这种响应机制在不同性别个体之间的差异性具有重要意义. ...
... 研究区域位于滇西北德钦县白马雪山, 地理坐标为28.48° N, 98.99° E, 海拔范围为4 360?4 920 m, 生境类型从低至高依次为灌草甸、草甸-流石滩交错带和流石滩(
图1D ).滇西北处于横断山高山植物多样性中心区域(
郭辉军和龙春林, 1998 ;
Zhang et al ., 2015 ), 具有典型的季节性夏季季风气候, 以冷雨或雨夹雪为特征, 并伴有短时间的强烈太阳辐射, 年降水量680-790 mm, 年平均气温-1 ℃ (
张荣祖等, 1997 ).植物生长季节通常开始于5月中旬的融雪期, 结束于9月中下旬的夏季风停息期.我们前期的测量数据显示, 2015年6月中旬至9月下旬, 白马雪山海拔4 900 m处, 距离地面15 cm处的平均气温和湿度分别为(4.87 ± 0.04) ℃ 和(89.9 ± 0.24)%, 绝对最高气温和最低气温分别为17.80和-1.95 ℃, 昼夜温差大于10 ℃, 最高达16 ℃ (陈建国等, 未发表数据).此外, 土壤养分含量随海拔升高而普遍降低(
Chen et al ., 2015b ,
2019 ). ...
... 横断山高海拔地区绝大多数植物种群的更新与扩张都依赖有性繁殖(
Peng et al ., 2012 ).植物对繁殖功能的资源分配, 在很大程度上决定了其种群未来的扩张速率.而作为植物实现繁殖功能的重要环节, 植物对开花的资源配置决定了植物的最终繁殖产出(
Reekie & Bazzaz, 2005 ).不同的环境因子在植物的每一个繁殖阶段都起着重要作用, 甚至一些因子(如资源量)直接决定着植物的繁殖分配及其产出(
Delph & Carroll, 2001 ).在我们的研究区域内, 环境胁迫程度(如土壤养分胁迫及低温胁迫)随海拔升高而增强(
Chen et al ., 2015b ,
2019 ).我们的研究结果显示, 随着海拔升高, 种群内植株个体呈现减小的趋势.植株大小对开花特性具有显著影响.一方面, 植株个体越大, 其总开花面积也越大, 但另一方面, 植株越大, 其总开花面积比越小(
表2 ), 并且这种趋势在不同性别植株间具有高度一致性(
图3 ), 说明资源限制是真实存在的, 即, 植物从环境中获取并分配给开花的资源量有限.此外, 该结果也说明植株分配到开花的资源增长速率可能低于植株个体的增长速率, 即繁殖分配的增加速率低于营养分配的增加速率.相应结果进一步暗示, 在面临资源限制或环境胁迫时, 团状福禄草的资源分配策略以保证植株存活(即增加营养分配)为首要目的.此外, 具有不同坡度及坡向的种群间开花特性也存在一定的变化,说明由坡向及坡度变化而引起的环境因子变化, 如太阳辐射(
田辉等, 2007 )和土壤养分(
曹舰艇等, 2019 ), 可能也同时对其繁殖分配策略具有一定的影响. ...
... 前期研究发现, 团状福禄草植物个体表面单位面积的果实及种子产量随海拔上升而呈现总体下降趋势(
Chen et al ., 2017a ).但是, 该研究并未考虑团状福禄草植物整体的繁殖分配对环境因子的响应, 而仅是比较了其特定开花区域内单位面积的繁殖产出.本研究结果显示, 团状福禄草在开花阶段的繁殖分配在不同海拔种群间存在显著差异, 总体上随海拔上升而减少, 表明高海拔更加贫瘠的土壤条件(
Chen et al ., 2015b ,
2019 )可能限制了植物总体的资源获取量, 植物需要将有限的资源在营养生长与繁殖生长两方面之间做权衡(
Reekie & Avila-Sakar, 2005 ), 从而导致其在繁殖功能上的总体分配比例的下降.但是, 我们也发现, 部分低海拔种群(种群2)内植物个体的繁殖分配显著低于部分高海拔种群(种群3、4), 这表明, 海拔并非控制植物繁殖分配的唯一因素, 植物繁殖分配是多种生态因子共同作用的结果(
张婵等, 2020 ).种群2与种群1海拔相差较小, 但坡度相差较大, 其与种群3坡度相近, 但具有不同坡向(
表1 ).因此, 我们推测除了海拔因素, 坡度与坡向可能同时对其繁殖分配产生影响.一方面坡度大可能导致土壤养分流失更严重(
Bruce-Okine & Lal, 1975 ;
Donker & Damen, 1984 ), 从而引起资源胁迫的加剧, 并进而导致繁殖分配的减少.另一方面, 坡度和坡向通常会引起太阳辐射特征的变化, 如太阳入射角、入射时间在不同坡度和坡向上会发生变化(
田辉等, 2007 ), 从而引起植物可积累和利用的有效太阳辐射能量的变化.在高海拔地区, 太阳辐射对植物的生长与繁殖策略都起着重要的影响.如, 为了充分利用太阳辐射, 一些植物花器官进化出了“向日性”功能(
Zhang et al ., 2010 ).又如, 安第斯山脉中部高海拔岩石斜坡典型垫状植物
Azorella compacta 为了最大化利用太阳辐射, 其个体偏爱生长在向北的(南半球)坡度约为20°的斜坡(
Kleier et al ., 2015 ).因此, 我们推测由坡度与坡向变化引起的土壤养分状况及太阳辐射特征的变化, 可能同时对团状福禄草的繁殖分配起着重要的影响. ...
How cushion communities are maintained in alpine ecosystems: a review and case study on alpine cushion plant reproduction
10
2017
... 温度、资源(包括土壤养分、降水资源及太阳辐射量等)是限制高山植物生长与繁殖的关键因素(
Bloom et al ., 1985 ;
Chen et al ., 2017b ).为了应对高海拔地区贫瘠的土壤条件, 植物在自身资源获取及分配方面进化出了众多与环境相适应的资源分配策略.其中, 繁殖分配(即植物分配到繁殖器官的资源量)是植物适合度的关键表现(
Bazzaz et al ., 1987 ;
Weiner, 1988 ;
Hartnett, 1990 ;
Vega et al ., 2000 ).研究表明, 植物繁殖分配同时受到众多非生物与生物因子的控制(
Reekie & Bazzaz, 2005 ;
张婵等, 2020 ).非生物因子方面, 温度、水分、土壤营养等对植物的繁殖分配起着重要作用(
Reekie & Bazzaz, 2005 ;
Zhang et al ., 2014 ).而在生物因子方面, 种间竞争、传粉者特性以及植物自身性系统特征(
Ashman, 1994 ;
Case & Ashman, 2005 ;
Brzyski et al ., 2014 ;
Chen et al ., 2017a ,
2017b )对其繁殖分配策略都起着重要的影响.例如, 具有雌全异株性系统特征(即同一种植物同时存在雌性个体和两性个体)的植物, 其同一种群内的雌性个体与两性个体在繁殖分配策略上存在较大差异, 表现为雌性个体主要将资源分配到雌性功能(如胚珠及种子生产), 而两性个体则主要将资源分配到雄性功能(如花粉生产以及传粉者吸引)(
Ashman, 1994 ;
Chen et al ., 2017a ,
2017b ), 并且这种分配模式受到环境因子的强烈影响(
Chen et al ., 2017b ).为了应对资源的不足, 植物往往需要在营养分配(即分配到营养器官的资源量)与繁殖分配之间做出权衡, 以达到最优的繁殖适合度(
Reekie & Avila-Sakar, 2005 ).开花是植物繁殖的最初阶段, 植物开花的数目, 直接反映出植物在特定环境条件下的繁殖分配策略.另一方面, 开花数目的多少也可直接决定着植物未来的繁殖输出量(如果实及种子产量;
Reekie & Bazzaz, 2005 ). ...
... ;
Chen et al ., 2017a ,
2017b ), 并且这种分配模式受到环境因子的强烈影响(
Chen et al ., 2017b ).为了应对资源的不足, 植物往往需要在营养分配(即分配到营养器官的资源量)与繁殖分配之间做出权衡, 以达到最优的繁殖适合度(
Reekie & Avila-Sakar, 2005 ).开花是植物繁殖的最初阶段, 植物开花的数目, 直接反映出植物在特定环境条件下的繁殖分配策略.另一方面, 开花数目的多少也可直接决定着植物未来的繁殖输出量(如果实及种子产量;
Reekie & Bazzaz, 2005 ). ...
... 团状福禄草(
Arenaria polytrichoides )为石竹科无心菜属(
Arenaria )多年生垫状草本植物, 是青藏高原特有种, 沿喜马拉雅—横断山高海拔(3 500-5 300 m)地区分布(
Wu et al ., 2013 ).营养生长自冰雪消融开始, 花期主要在6-7月, 不同分布区域及海拔的种群的花期存在一定的差异(
Chen et al ., 2017a ).并且, 团状福禄草具有雌全异株的性二态系统, 即同一种群中同时存在雌性植株和两性植株, 不同性别的个体在繁殖产出(果实及种子产量)方面存在显著差异, 并且受到海拔的影响(
Chen et al ., 2017a ).表现为较高海拔的雌性比例高于较低海拔的, 两性植株开花数目大于雌性植株但结实率显著低于雌性植株, 同时, 较低海拔种群内个体单位面积的果实与种子产量高于较高海拔种群的个体(
Chen et al ., 2017a ).此外, 团状福禄草被证实为横断山高山生态系统当中的关键种, 能有效改良微环境条件(温度、土壤营养及水分), 从而为其他低耐受物种提供适宜的微生境, 进而调节种群动态、提升生态系统物种多样性及地面生产力(
Yang et al ., 2010 ,
2017 ;
Chen et al ., 2015a ,
2019 ).然而, 团状福禄草种群的更新却受到种间竞争的强烈影响(
Chen et al ., 2020 ).研究者在近十年内对团状福禄草做了深入调查与研究(
Yang et al ., 2010 ,
2017 ;
Chen et al ., 2015a ,
2015b ,
2017a ,
2019 ), 但对其种群的繁殖与更新机制, 却直到最近才予以关注(
Chen et al ., 2017a ,
2020 ).理论上讲, 团状福禄草的每一个小分枝(
图1C )在生长季节都能开出一朵花, 然而, 我们在多年的野外研究中发现, 实际上, 其每年的开花数量极为有限, 只在其冠层表面的部分区域开出一定面积的花(
图1A 、1B).这一现象说明, 垫状植物的开花可能受到了资源的限制, 并有可能同时受到其他环境因子的影响.例如, 据野外观察, 团状福禄草开花方位(只在其冠层表面特定区域开放,
图1A 、1B)呈现出一定的规律性, 且在不同的种群之间存在差异.为了获取充足的太阳辐射能量从而满足自身繁殖需求, 一些高山植物的开花特性受到太阳辐射的强烈影响(
Pati?o et al ., 2002 ;
Galen, 2006 ;
Zhang et al ., 2010 ), 因此, 我们猜测团状福禄草的开花数量和方位可能受到资源和太阳辐射的双重影响.但其繁殖分配策略, 即个体对开花的资源分配策略对环境胁迫因子的响应方式, 目前却不为人知.深入探究高山生态系统中这一关键类群的种群更新与维持机制, 对揭示其种群繁殖的初始阶段(开花特性)对环境因子的响应机制以及这种响应机制在不同性别个体之间的差异性具有重要意义. ...
... ).并且, 团状福禄草具有雌全异株的性二态系统, 即同一种群中同时存在雌性植株和两性植株, 不同性别的个体在繁殖产出(果实及种子产量)方面存在显著差异, 并且受到海拔的影响(
Chen et al ., 2017a ).表现为较高海拔的雌性比例高于较低海拔的, 两性植株开花数目大于雌性植株但结实率显著低于雌性植株, 同时, 较低海拔种群内个体单位面积的果实与种子产量高于较高海拔种群的个体(
Chen et al ., 2017a ).此外, 团状福禄草被证实为横断山高山生态系统当中的关键种, 能有效改良微环境条件(温度、土壤营养及水分), 从而为其他低耐受物种提供适宜的微生境, 进而调节种群动态、提升生态系统物种多样性及地面生产力(
Yang et al ., 2010 ,
2017 ;
Chen et al ., 2015a ,
2019 ).然而, 团状福禄草种群的更新却受到种间竞争的强烈影响(
Chen et al ., 2020 ).研究者在近十年内对团状福禄草做了深入调查与研究(
Yang et al ., 2010 ,
2017 ;
Chen et al ., 2015a ,
2015b ,
2017a ,
2019 ), 但对其种群的繁殖与更新机制, 却直到最近才予以关注(
Chen et al ., 2017a ,
2020 ).理论上讲, 团状福禄草的每一个小分枝(
图1C )在生长季节都能开出一朵花, 然而, 我们在多年的野外研究中发现, 实际上, 其每年的开花数量极为有限, 只在其冠层表面的部分区域开出一定面积的花(
图1A 、1B).这一现象说明, 垫状植物的开花可能受到了资源的限制, 并有可能同时受到其他环境因子的影响.例如, 据野外观察, 团状福禄草开花方位(只在其冠层表面特定区域开放,
图1A 、1B)呈现出一定的规律性, 且在不同的种群之间存在差异.为了获取充足的太阳辐射能量从而满足自身繁殖需求, 一些高山植物的开花特性受到太阳辐射的强烈影响(
Pati?o et al ., 2002 ;
Galen, 2006 ;
Zhang et al ., 2010 ), 因此, 我们猜测团状福禄草的开花数量和方位可能受到资源和太阳辐射的双重影响.但其繁殖分配策略, 即个体对开花的资源分配策略对环境胁迫因子的响应方式, 目前却不为人知.深入探究高山生态系统中这一关键类群的种群更新与维持机制, 对揭示其种群繁殖的初始阶段(开花特性)对环境因子的响应机制以及这种响应机制在不同性别个体之间的差异性具有重要意义. ...
... ).表现为较高海拔的雌性比例高于较低海拔的, 两性植株开花数目大于雌性植株但结实率显著低于雌性植株, 同时, 较低海拔种群内个体单位面积的果实与种子产量高于较高海拔种群的个体(
Chen et al ., 2017a ).此外, 团状福禄草被证实为横断山高山生态系统当中的关键种, 能有效改良微环境条件(温度、土壤营养及水分), 从而为其他低耐受物种提供适宜的微生境, 进而调节种群动态、提升生态系统物种多样性及地面生产力(
Yang et al ., 2010 ,
2017 ;
Chen et al ., 2015a ,
2019 ).然而, 团状福禄草种群的更新却受到种间竞争的强烈影响(
Chen et al ., 2020 ).研究者在近十年内对团状福禄草做了深入调查与研究(
Yang et al ., 2010 ,
2017 ;
Chen et al ., 2015a ,
2015b ,
2017a ,
2019 ), 但对其种群的繁殖与更新机制, 却直到最近才予以关注(
Chen et al ., 2017a ,
2020 ).理论上讲, 团状福禄草的每一个小分枝(
图1C )在生长季节都能开出一朵花, 然而, 我们在多年的野外研究中发现, 实际上, 其每年的开花数量极为有限, 只在其冠层表面的部分区域开出一定面积的花(
图1A 、1B).这一现象说明, 垫状植物的开花可能受到了资源的限制, 并有可能同时受到其他环境因子的影响.例如, 据野外观察, 团状福禄草开花方位(只在其冠层表面特定区域开放,
图1A 、1B)呈现出一定的规律性, 且在不同的种群之间存在差异.为了获取充足的太阳辐射能量从而满足自身繁殖需求, 一些高山植物的开花特性受到太阳辐射的强烈影响(
Pati?o et al ., 2002 ;
Galen, 2006 ;
Zhang et al ., 2010 ), 因此, 我们猜测团状福禄草的开花数量和方位可能受到资源和太阳辐射的双重影响.但其繁殖分配策略, 即个体对开花的资源分配策略对环境胁迫因子的响应方式, 目前却不为人知.深入探究高山生态系统中这一关键类群的种群更新与维持机制, 对揭示其种群繁殖的初始阶段(开花特性)对环境因子的响应机制以及这种响应机制在不同性别个体之间的差异性具有重要意义. ...
... ,
2017a ,
2019 ), 但对其种群的繁殖与更新机制, 却直到最近才予以关注(
Chen et al ., 2017a ,
2020 ).理论上讲, 团状福禄草的每一个小分枝(
图1C )在生长季节都能开出一朵花, 然而, 我们在多年的野外研究中发现, 实际上, 其每年的开花数量极为有限, 只在其冠层表面的部分区域开出一定面积的花(
图1A 、1B).这一现象说明, 垫状植物的开花可能受到了资源的限制, 并有可能同时受到其他环境因子的影响.例如, 据野外观察, 团状福禄草开花方位(只在其冠层表面特定区域开放,
图1A 、1B)呈现出一定的规律性, 且在不同的种群之间存在差异.为了获取充足的太阳辐射能量从而满足自身繁殖需求, 一些高山植物的开花特性受到太阳辐射的强烈影响(
Pati?o et al ., 2002 ;
Galen, 2006 ;
Zhang et al ., 2010 ), 因此, 我们猜测团状福禄草的开花数量和方位可能受到资源和太阳辐射的双重影响.但其繁殖分配策略, 即个体对开花的资源分配策略对环境胁迫因子的响应方式, 目前却不为人知.深入探究高山生态系统中这一关键类群的种群更新与维持机制, 对揭示其种群繁殖的初始阶段(开花特性)对环境因子的响应机制以及这种响应机制在不同性别个体之间的差异性具有重要意义. ...
... ), 但对其种群的繁殖与更新机制, 却直到最近才予以关注(
Chen et al ., 2017a ,
2020 ).理论上讲, 团状福禄草的每一个小分枝(
图1C )在生长季节都能开出一朵花, 然而, 我们在多年的野外研究中发现, 实际上, 其每年的开花数量极为有限, 只在其冠层表面的部分区域开出一定面积的花(
图1A 、1B).这一现象说明, 垫状植物的开花可能受到了资源的限制, 并有可能同时受到其他环境因子的影响.例如, 据野外观察, 团状福禄草开花方位(只在其冠层表面特定区域开放,
图1A 、1B)呈现出一定的规律性, 且在不同的种群之间存在差异.为了获取充足的太阳辐射能量从而满足自身繁殖需求, 一些高山植物的开花特性受到太阳辐射的强烈影响(
Pati?o et al ., 2002 ;
Galen, 2006 ;
Zhang et al ., 2010 ), 因此, 我们猜测团状福禄草的开花数量和方位可能受到资源和太阳辐射的双重影响.但其繁殖分配策略, 即个体对开花的资源分配策略对环境胁迫因子的响应方式, 目前却不为人知.深入探究高山生态系统中这一关键类群的种群更新与维持机制, 对揭示其种群繁殖的初始阶段(开花特性)对环境因子的响应机制以及这种响应机制在不同性别个体之间的差异性具有重要意义. ...
... 首先, 将各方向上开花网格数相加, 得到各植株的总开花网格(面积), 其次, 分别将总开花网格数和每个方向的开花网格数都除以总网格数, 得到各个植株总的开花面积比(为了论述方便, 本研究中, 必要时将开花面积比称为“繁殖分配”)以及单个方位上的开花面积比, 并采用这些数据进行后续的比较分析.不同植株的大小(年龄)会对其资源分配策略产生影响, 即可能存在个体效应(
St?cklin & Favre, 1994 ;
Sakai & Sakai, 1995 ), 同时, 垫状植物冠层表面积大小通常与其年龄呈正相关关系(
Chen et al ., 2017a ), 因此, 计数了每个调查个体的总网格数(即表面积大小), 将其作为植株大小的替代来进行后续分析.首先, 为了探究不同种群以及不同性别植株的总开花面积与其个体大小之间的关系, 对植株总开花面积与其个体大小做了回归分析.其次, 为了探究海拔对植株大小的影响, 采用混合效应模型比较了不同海拔种群间植株的大小, 植株大小为因变量, 海拔为主因素, 植株个体编号为随机因素.最后, 为了探究开花面积比(即繁殖分配)如何受到海拔、性别和植株大小的影响, 使用混合效应模型对上述关系进行拟合.模型中, 开花面积比为因变量, 海拔、性别和植株大小为主因素, 植株个体编号为随机因素.此外, 由于自变量与因变量间存在多样的组合形式, 基于赤池信息量准则(AIC)权重选择最优模型, 最优模型具有最大的AIC权重, 因子在每个模型的AIC权重被用来计算模型平均系数, 而每个因子在所有模型中的AIC值权重之和(
w )被用来描述该因子的统计支持.同时, 评估了考虑交互作用的模型与上述模型的优劣, 校准AIC (
AICc )结果显示两种模型没有差异(
AICc = -402.1 vs.
AICc = -401.4), 但是交互作用不显著, 因而以上述的模型结果为准.由于各组数据不完全符合正态分布及方差齐性原则, 使用Kruskal-Wallis非参数检验分别比较同一种群内不同性别、不同方位之间开花面积比以及不同种群之间开花面积比的差异.所有统计分析及画图都通过R 3.5.1 (
R Core Team, 2018 )实现, Kruskal-Wallis非参数检验使用“agricolae”包(
de Felipe, 2019 ), 混合效应模型使用“coxme”包(
Therneau, 2018 ), 模型选择使用“MuMIn”包(
Bartoń, 2019 ). ...
... 前期研究发现, 团状福禄草植物个体表面单位面积的果实及种子产量随海拔上升而呈现总体下降趋势(
Chen et al ., 2017a ).但是, 该研究并未考虑团状福禄草植物整体的繁殖分配对环境因子的响应, 而仅是比较了其特定开花区域内单位面积的繁殖产出.本研究结果显示, 团状福禄草在开花阶段的繁殖分配在不同海拔种群间存在显著差异, 总体上随海拔上升而减少, 表明高海拔更加贫瘠的土壤条件(
Chen et al ., 2015b ,
2019 )可能限制了植物总体的资源获取量, 植物需要将有限的资源在营养生长与繁殖生长两方面之间做权衡(
Reekie & Avila-Sakar, 2005 ), 从而导致其在繁殖功能上的总体分配比例的下降.但是, 我们也发现, 部分低海拔种群(种群2)内植物个体的繁殖分配显著低于部分高海拔种群(种群3、4), 这表明, 海拔并非控制植物繁殖分配的唯一因素, 植物繁殖分配是多种生态因子共同作用的结果(
张婵等, 2020 ).种群2与种群1海拔相差较小, 但坡度相差较大, 其与种群3坡度相近, 但具有不同坡向(
表1 ).因此, 我们推测除了海拔因素, 坡度与坡向可能同时对其繁殖分配产生影响.一方面坡度大可能导致土壤养分流失更严重(
Bruce-Okine & Lal, 1975 ;
Donker & Damen, 1984 ), 从而引起资源胁迫的加剧, 并进而导致繁殖分配的减少.另一方面, 坡度和坡向通常会引起太阳辐射特征的变化, 如太阳入射角、入射时间在不同坡度和坡向上会发生变化(
田辉等, 2007 ), 从而引起植物可积累和利用的有效太阳辐射能量的变化.在高海拔地区, 太阳辐射对植物的生长与繁殖策略都起着重要的影响.如, 为了充分利用太阳辐射, 一些植物花器官进化出了“向日性”功能(
Zhang et al ., 2010 ).又如, 安第斯山脉中部高海拔岩石斜坡典型垫状植物
Azorella compacta 为了最大化利用太阳辐射, 其个体偏爱生长在向北的(南半球)坡度约为20°的斜坡(
Kleier et al ., 2015 ).因此, 我们推测由坡度与坡向变化引起的土壤养分状况及太阳辐射特征的变化, 可能同时对团状福禄草的繁殖分配起着重要的影响. ...
... 此外, 团状福禄草具有性二态系统, 即雌性植株与两性植株同时存在于同一种群中.不同性别的植物个体获得繁殖适合度的方式不同, 雌性个体主要通过胚珠及种子产出获得繁殖适合度, 而两性植株主要通过花粉与种子产出获得繁殖适合度(
Charlesworth & Charlesworth, 1978 ;
Charlesworth & Morgan, 1991 ;
Ashman, 1994 ).因此, 不同性别的植株个体在繁殖分配策略上存在一定的差异性, 这种差异性直接反映了不同性别个体对特定环境因子的适应策略(
Case & Ashman, 2005 ;
Ashman, 2006 ;
Chen et al ., 2017b ).例如, 同样分布于滇西北高海拔地区, 且同样具有雌全异株性系统的细叶蓝钟花(
Cyananthus delavayi ), 在面对环境因子胁迫时, 雌性植株主要将资源分配到种子生产, 而两性植株则主要将资源分配到花粉生产以及对传粉昆虫的吸引功能方面(花瓣)(
Chen et al ., 2017b ).我们的研究结果表明, 总体而言, 两性植株对开花的资源分配比例要显著高于雌性植株(
图2B ), 但是, 其差异程度受到海拔因素的影响.具体而言, 低海拔种群两性植株分配给开花的资源比例显著高于雌性植株, 而高海拔种群不同性别植株对开花的资源分配差异性不显著(
图4 ).
Chen等(2017a) 研究发现, 在任何海拔条件下, 团状福禄草两性植株单位面积的开花数量始终大于雌性植株, 但其真实繁殖产出(单位面积果实和种子产量)却始终低于雌性植株.但是, 本研究发现, 尽管不同性别的个体之间在低海拔处的总繁殖分配比例存在差异性, 但是随着海拔升高, 环境胁迫程度增加, 这种差异性趋于消失(
图4 ).我们猜测, 在高海拔环境胁迫加剧条件下, 不同性别的植物个体在资源权衡分配方面采取了相同的策略.另一方面, 除了物理胁迫的加剧, 生物胁迫也同时加剧, 如, 传粉昆虫多样性及多度的下降(
Arroyo et al ., 1982 ;
Medan et al ., 2002 )可能引起两性植株雄性适合度输出(花粉传播)的降低, 从而引起其分配到开花的资源随之减少. ...
Sex allocation in gynodioecious
Cyananthus delavayi differs between gender morphs and soil quality
6
2017
... 温度、资源(包括土壤养分、降水资源及太阳辐射量等)是限制高山植物生长与繁殖的关键因素(
Bloom et al ., 1985 ;
Chen et al ., 2017b ).为了应对高海拔地区贫瘠的土壤条件, 植物在自身资源获取及分配方面进化出了众多与环境相适应的资源分配策略.其中, 繁殖分配(即植物分配到繁殖器官的资源量)是植物适合度的关键表现(
Bazzaz et al ., 1987 ;
Weiner, 1988 ;
Hartnett, 1990 ;
Vega et al ., 2000 ).研究表明, 植物繁殖分配同时受到众多非生物与生物因子的控制(
Reekie & Bazzaz, 2005 ;
张婵等, 2020 ).非生物因子方面, 温度、水分、土壤营养等对植物的繁殖分配起着重要作用(
Reekie & Bazzaz, 2005 ;
Zhang et al ., 2014 ).而在生物因子方面, 种间竞争、传粉者特性以及植物自身性系统特征(
Ashman, 1994 ;
Case & Ashman, 2005 ;
Brzyski et al ., 2014 ;
Chen et al ., 2017a ,
2017b )对其繁殖分配策略都起着重要的影响.例如, 具有雌全异株性系统特征(即同一种植物同时存在雌性个体和两性个体)的植物, 其同一种群内的雌性个体与两性个体在繁殖分配策略上存在较大差异, 表现为雌性个体主要将资源分配到雌性功能(如胚珠及种子生产), 而两性个体则主要将资源分配到雄性功能(如花粉生产以及传粉者吸引)(
Ashman, 1994 ;
Chen et al ., 2017a ,
2017b ), 并且这种分配模式受到环境因子的强烈影响(
Chen et al ., 2017b ).为了应对资源的不足, 植物往往需要在营养分配(即分配到营养器官的资源量)与繁殖分配之间做出权衡, 以达到最优的繁殖适合度(
Reekie & Avila-Sakar, 2005 ).开花是植物繁殖的最初阶段, 植物开花的数目, 直接反映出植物在特定环境条件下的繁殖分配策略.另一方面, 开花数目的多少也可直接决定着植物未来的繁殖输出量(如果实及种子产量;
Reekie & Bazzaz, 2005 ). ...
... ,
2017b )对其繁殖分配策略都起着重要的影响.例如, 具有雌全异株性系统特征(即同一种植物同时存在雌性个体和两性个体)的植物, 其同一种群内的雌性个体与两性个体在繁殖分配策略上存在较大差异, 表现为雌性个体主要将资源分配到雌性功能(如胚珠及种子生产), 而两性个体则主要将资源分配到雄性功能(如花粉生产以及传粉者吸引)(
Ashman, 1994 ;
Chen et al ., 2017a ,
2017b ), 并且这种分配模式受到环境因子的强烈影响(
Chen et al ., 2017b ).为了应对资源的不足, 植物往往需要在营养分配(即分配到营养器官的资源量)与繁殖分配之间做出权衡, 以达到最优的繁殖适合度(
Reekie & Avila-Sakar, 2005 ).开花是植物繁殖的最初阶段, 植物开花的数目, 直接反映出植物在特定环境条件下的繁殖分配策略.另一方面, 开花数目的多少也可直接决定着植物未来的繁殖输出量(如果实及种子产量;
Reekie & Bazzaz, 2005 ). ...
... ,
2017b ), 并且这种分配模式受到环境因子的强烈影响(
Chen et al ., 2017b ).为了应对资源的不足, 植物往往需要在营养分配(即分配到营养器官的资源量)与繁殖分配之间做出权衡, 以达到最优的繁殖适合度(
Reekie & Avila-Sakar, 2005 ).开花是植物繁殖的最初阶段, 植物开花的数目, 直接反映出植物在特定环境条件下的繁殖分配策略.另一方面, 开花数目的多少也可直接决定着植物未来的繁殖输出量(如果实及种子产量;
Reekie & Bazzaz, 2005 ). ...
... ), 并且这种分配模式受到环境因子的强烈影响(
Chen et al ., 2017b ).为了应对资源的不足, 植物往往需要在营养分配(即分配到营养器官的资源量)与繁殖分配之间做出权衡, 以达到最优的繁殖适合度(
Reekie & Avila-Sakar, 2005 ).开花是植物繁殖的最初阶段, 植物开花的数目, 直接反映出植物在特定环境条件下的繁殖分配策略.另一方面, 开花数目的多少也可直接决定着植物未来的繁殖输出量(如果实及种子产量;
Reekie & Bazzaz, 2005 ). ...
... 此外, 团状福禄草具有性二态系统, 即雌性植株与两性植株同时存在于同一种群中.不同性别的植物个体获得繁殖适合度的方式不同, 雌性个体主要通过胚珠及种子产出获得繁殖适合度, 而两性植株主要通过花粉与种子产出获得繁殖适合度(
Charlesworth & Charlesworth, 1978 ;
Charlesworth & Morgan, 1991 ;
Ashman, 1994 ).因此, 不同性别的植株个体在繁殖分配策略上存在一定的差异性, 这种差异性直接反映了不同性别个体对特定环境因子的适应策略(
Case & Ashman, 2005 ;
Ashman, 2006 ;
Chen et al ., 2017b ).例如, 同样分布于滇西北高海拔地区, 且同样具有雌全异株性系统的细叶蓝钟花(
Cyananthus delavayi ), 在面对环境因子胁迫时, 雌性植株主要将资源分配到种子生产, 而两性植株则主要将资源分配到花粉生产以及对传粉昆虫的吸引功能方面(花瓣)(
Chen et al ., 2017b ).我们的研究结果表明, 总体而言, 两性植株对开花的资源分配比例要显著高于雌性植株(
图2B ), 但是, 其差异程度受到海拔因素的影响.具体而言, 低海拔种群两性植株分配给开花的资源比例显著高于雌性植株, 而高海拔种群不同性别植株对开花的资源分配差异性不显著(
图4 ).
Chen等(2017a) 研究发现, 在任何海拔条件下, 团状福禄草两性植株单位面积的开花数量始终大于雌性植株, 但其真实繁殖产出(单位面积果实和种子产量)却始终低于雌性植株.但是, 本研究发现, 尽管不同性别的个体之间在低海拔处的总繁殖分配比例存在差异性, 但是随着海拔升高, 环境胁迫程度增加, 这种差异性趋于消失(
图4 ).我们猜测, 在高海拔环境胁迫加剧条件下, 不同性别的植物个体在资源权衡分配方面采取了相同的策略.另一方面, 除了物理胁迫的加剧, 生物胁迫也同时加剧, 如, 传粉昆虫多样性及多度的下降(
Arroyo et al ., 1982 ;
Medan et al ., 2002 )可能引起两性植株雄性适合度输出(花粉传播)的降低, 从而引起其分配到开花的资源随之减少. ...
... ), 在面对环境因子胁迫时, 雌性植株主要将资源分配到种子生产, 而两性植株则主要将资源分配到花粉生产以及对传粉昆虫的吸引功能方面(花瓣)(
Chen et al ., 2017b ).我们的研究结果表明, 总体而言, 两性植株对开花的资源分配比例要显著高于雌性植株(
图2B ), 但是, 其差异程度受到海拔因素的影响.具体而言, 低海拔种群两性植株分配给开花的资源比例显著高于雌性植株, 而高海拔种群不同性别植株对开花的资源分配差异性不显著(
图4 ).
Chen等(2017a) 研究发现, 在任何海拔条件下, 团状福禄草两性植株单位面积的开花数量始终大于雌性植株, 但其真实繁殖产出(单位面积果实和种子产量)却始终低于雌性植株.但是, 本研究发现, 尽管不同性别的个体之间在低海拔处的总繁殖分配比例存在差异性, 但是随着海拔升高, 环境胁迫程度增加, 这种差异性趋于消失(
图4 ).我们猜测, 在高海拔环境胁迫加剧条件下, 不同性别的植物个体在资源权衡分配方面采取了相同的策略.另一方面, 除了物理胁迫的加剧, 生物胁迫也同时加剧, 如, 传粉昆虫多样性及多度的下降(
Arroyo et al ., 1982 ;
Medan et al ., 2002 )可能引起两性植株雄性适合度输出(花粉传播)的降低, 从而引起其分配到开花的资源随之减少. ...
Cushion plants can have a positive effect on diversity at high elevations in the Himalayan Hengduan Mountains
3
2015
... 垫状植物是一类具有球形或半球形结构的植物的总称, 是植物经过多年辐射性生长而形成的小枝紧密簇生结构(
图1 ;
Gibson & Hope, 1986 ), 因其适应世界上大多数山地和极地极端干旱和低温环境的能力而备受植物学家的关注(
Aubert et al ., 2014 ).并因其具有生态系统工程师的功能, 对其他植物具有“护理作用”, 可提高和维持生态系统中物种多样性(
Butterfield et al ., 2013 ;
Cavieres et al ., 2014 ;
Chen et al ., 2015a ,
2015b ,
2019 ), 而被认为是所在生态系统当中的奠基种(foundation species;
Kikvidze et al ., 2015 ). ...
... 团状福禄草(
Arenaria polytrichoides )为石竹科无心菜属(
Arenaria )多年生垫状草本植物, 是青藏高原特有种, 沿喜马拉雅—横断山高海拔(3 500-5 300 m)地区分布(
Wu et al ., 2013 ).营养生长自冰雪消融开始, 花期主要在6-7月, 不同分布区域及海拔的种群的花期存在一定的差异(
Chen et al ., 2017a ).并且, 团状福禄草具有雌全异株的性二态系统, 即同一种群中同时存在雌性植株和两性植株, 不同性别的个体在繁殖产出(果实及种子产量)方面存在显著差异, 并且受到海拔的影响(
Chen et al ., 2017a ).表现为较高海拔的雌性比例高于较低海拔的, 两性植株开花数目大于雌性植株但结实率显著低于雌性植株, 同时, 较低海拔种群内个体单位面积的果实与种子产量高于较高海拔种群的个体(
Chen et al ., 2017a ).此外, 团状福禄草被证实为横断山高山生态系统当中的关键种, 能有效改良微环境条件(温度、土壤营养及水分), 从而为其他低耐受物种提供适宜的微生境, 进而调节种群动态、提升生态系统物种多样性及地面生产力(
Yang et al ., 2010 ,
2017 ;
Chen et al ., 2015a ,
2019 ).然而, 团状福禄草种群的更新却受到种间竞争的强烈影响(
Chen et al ., 2020 ).研究者在近十年内对团状福禄草做了深入调查与研究(
Yang et al ., 2010 ,
2017 ;
Chen et al ., 2015a ,
2015b ,
2017a ,
2019 ), 但对其种群的繁殖与更新机制, 却直到最近才予以关注(
Chen et al ., 2017a ,
2020 ).理论上讲, 团状福禄草的每一个小分枝(
图1C )在生长季节都能开出一朵花, 然而, 我们在多年的野外研究中发现, 实际上, 其每年的开花数量极为有限, 只在其冠层表面的部分区域开出一定面积的花(
图1A 、1B).这一现象说明, 垫状植物的开花可能受到了资源的限制, 并有可能同时受到其他环境因子的影响.例如, 据野外观察, 团状福禄草开花方位(只在其冠层表面特定区域开放,
图1A 、1B)呈现出一定的规律性, 且在不同的种群之间存在差异.为了获取充足的太阳辐射能量从而满足自身繁殖需求, 一些高山植物的开花特性受到太阳辐射的强烈影响(
Pati?o et al ., 2002 ;
Galen, 2006 ;
Zhang et al ., 2010 ), 因此, 我们猜测团状福禄草的开花数量和方位可能受到资源和太阳辐射的双重影响.但其繁殖分配策略, 即个体对开花的资源分配策略对环境胁迫因子的响应方式, 目前却不为人知.深入探究高山生态系统中这一关键类群的种群更新与维持机制, 对揭示其种群繁殖的初始阶段(开花特性)对环境因子的响应机制以及这种响应机制在不同性别个体之间的差异性具有重要意义. ...
... ;
Chen et al ., 2015a ,
2015b ,
2017a ,
2019 ), 但对其种群的繁殖与更新机制, 却直到最近才予以关注(
Chen et al ., 2017a ,
2020 ).理论上讲, 团状福禄草的每一个小分枝(
图1C )在生长季节都能开出一朵花, 然而, 我们在多年的野外研究中发现, 实际上, 其每年的开花数量极为有限, 只在其冠层表面的部分区域开出一定面积的花(
图1A 、1B).这一现象说明, 垫状植物的开花可能受到了资源的限制, 并有可能同时受到其他环境因子的影响.例如, 据野外观察, 团状福禄草开花方位(只在其冠层表面特定区域开放,
图1A 、1B)呈现出一定的规律性, 且在不同的种群之间存在差异.为了获取充足的太阳辐射能量从而满足自身繁殖需求, 一些高山植物的开花特性受到太阳辐射的强烈影响(
Pati?o et al ., 2002 ;
Galen, 2006 ;
Zhang et al ., 2010 ), 因此, 我们猜测团状福禄草的开花数量和方位可能受到资源和太阳辐射的双重影响.但其繁殖分配策略, 即个体对开花的资源分配策略对环境胁迫因子的响应方式, 目前却不为人知.深入探究高山生态系统中这一关键类群的种群更新与维持机制, 对揭示其种群繁殖的初始阶段(开花特性)对环境因子的响应机制以及这种响应机制在不同性别个体之间的差异性具有重要意义. ...
Soil nutrient availability determines the facilitative effects of cushion plants on other plant species at high elevations in the south-eastern Himalayas
5
2015
... 垫状植物是一类具有球形或半球形结构的植物的总称, 是植物经过多年辐射性生长而形成的小枝紧密簇生结构(
图1 ;
Gibson & Hope, 1986 ), 因其适应世界上大多数山地和极地极端干旱和低温环境的能力而备受植物学家的关注(
Aubert et al ., 2014 ).并因其具有生态系统工程师的功能, 对其他植物具有“护理作用”, 可提高和维持生态系统中物种多样性(
Butterfield et al ., 2013 ;
Cavieres et al ., 2014 ;
Chen et al ., 2015a ,
2015b ,
2019 ), 而被认为是所在生态系统当中的奠基种(foundation species;
Kikvidze et al ., 2015 ). ...
... 团状福禄草(
Arenaria polytrichoides )为石竹科无心菜属(
Arenaria )多年生垫状草本植物, 是青藏高原特有种, 沿喜马拉雅—横断山高海拔(3 500-5 300 m)地区分布(
Wu et al ., 2013 ).营养生长自冰雪消融开始, 花期主要在6-7月, 不同分布区域及海拔的种群的花期存在一定的差异(
Chen et al ., 2017a ).并且, 团状福禄草具有雌全异株的性二态系统, 即同一种群中同时存在雌性植株和两性植株, 不同性别的个体在繁殖产出(果实及种子产量)方面存在显著差异, 并且受到海拔的影响(
Chen et al ., 2017a ).表现为较高海拔的雌性比例高于较低海拔的, 两性植株开花数目大于雌性植株但结实率显著低于雌性植株, 同时, 较低海拔种群内个体单位面积的果实与种子产量高于较高海拔种群的个体(
Chen et al ., 2017a ).此外, 团状福禄草被证实为横断山高山生态系统当中的关键种, 能有效改良微环境条件(温度、土壤营养及水分), 从而为其他低耐受物种提供适宜的微生境, 进而调节种群动态、提升生态系统物种多样性及地面生产力(
Yang et al ., 2010 ,
2017 ;
Chen et al ., 2015a ,
2019 ).然而, 团状福禄草种群的更新却受到种间竞争的强烈影响(
Chen et al ., 2020 ).研究者在近十年内对团状福禄草做了深入调查与研究(
Yang et al ., 2010 ,
2017 ;
Chen et al ., 2015a ,
2015b ,
2017a ,
2019 ), 但对其种群的繁殖与更新机制, 却直到最近才予以关注(
Chen et al ., 2017a ,
2020 ).理论上讲, 团状福禄草的每一个小分枝(
图1C )在生长季节都能开出一朵花, 然而, 我们在多年的野外研究中发现, 实际上, 其每年的开花数量极为有限, 只在其冠层表面的部分区域开出一定面积的花(
图1A 、1B).这一现象说明, 垫状植物的开花可能受到了资源的限制, 并有可能同时受到其他环境因子的影响.例如, 据野外观察, 团状福禄草开花方位(只在其冠层表面特定区域开放,
图1A 、1B)呈现出一定的规律性, 且在不同的种群之间存在差异.为了获取充足的太阳辐射能量从而满足自身繁殖需求, 一些高山植物的开花特性受到太阳辐射的强烈影响(
Pati?o et al ., 2002 ;
Galen, 2006 ;
Zhang et al ., 2010 ), 因此, 我们猜测团状福禄草的开花数量和方位可能受到资源和太阳辐射的双重影响.但其繁殖分配策略, 即个体对开花的资源分配策略对环境胁迫因子的响应方式, 目前却不为人知.深入探究高山生态系统中这一关键类群的种群更新与维持机制, 对揭示其种群繁殖的初始阶段(开花特性)对环境因子的响应机制以及这种响应机制在不同性别个体之间的差异性具有重要意义. ...
... 研究区域位于滇西北德钦县白马雪山, 地理坐标为28.48° N, 98.99° E, 海拔范围为4 360?4 920 m, 生境类型从低至高依次为灌草甸、草甸-流石滩交错带和流石滩(
图1D ).滇西北处于横断山高山植物多样性中心区域(
郭辉军和龙春林, 1998 ;
Zhang et al ., 2015 ), 具有典型的季节性夏季季风气候, 以冷雨或雨夹雪为特征, 并伴有短时间的强烈太阳辐射, 年降水量680-790 mm, 年平均气温-1 ℃ (
张荣祖等, 1997 ).植物生长季节通常开始于5月中旬的融雪期, 结束于9月中下旬的夏季风停息期.我们前期的测量数据显示, 2015年6月中旬至9月下旬, 白马雪山海拔4 900 m处, 距离地面15 cm处的平均气温和湿度分别为(4.87 ± 0.04) ℃ 和(89.9 ± 0.24)%, 绝对最高气温和最低气温分别为17.80和-1.95 ℃, 昼夜温差大于10 ℃, 最高达16 ℃ (陈建国等, 未发表数据).此外, 土壤养分含量随海拔升高而普遍降低(
Chen et al ., 2015b ,
2019 ). ...
... 横断山高海拔地区绝大多数植物种群的更新与扩张都依赖有性繁殖(
Peng et al ., 2012 ).植物对繁殖功能的资源分配, 在很大程度上决定了其种群未来的扩张速率.而作为植物实现繁殖功能的重要环节, 植物对开花的资源配置决定了植物的最终繁殖产出(
Reekie & Bazzaz, 2005 ).不同的环境因子在植物的每一个繁殖阶段都起着重要作用, 甚至一些因子(如资源量)直接决定着植物的繁殖分配及其产出(
Delph & Carroll, 2001 ).在我们的研究区域内, 环境胁迫程度(如土壤养分胁迫及低温胁迫)随海拔升高而增强(
Chen et al ., 2015b ,
2019 ).我们的研究结果显示, 随着海拔升高, 种群内植株个体呈现减小的趋势.植株大小对开花特性具有显著影响.一方面, 植株个体越大, 其总开花面积也越大, 但另一方面, 植株越大, 其总开花面积比越小(
表2 ), 并且这种趋势在不同性别植株间具有高度一致性(
图3 ), 说明资源限制是真实存在的, 即, 植物从环境中获取并分配给开花的资源量有限.此外, 该结果也说明植株分配到开花的资源增长速率可能低于植株个体的增长速率, 即繁殖分配的增加速率低于营养分配的增加速率.相应结果进一步暗示, 在面临资源限制或环境胁迫时, 团状福禄草的资源分配策略以保证植株存活(即增加营养分配)为首要目的.此外, 具有不同坡度及坡向的种群间开花特性也存在一定的变化,说明由坡向及坡度变化而引起的环境因子变化, 如太阳辐射(
田辉等, 2007 )和土壤养分(
曹舰艇等, 2019 ), 可能也同时对其繁殖分配策略具有一定的影响. ...
... 前期研究发现, 团状福禄草植物个体表面单位面积的果实及种子产量随海拔上升而呈现总体下降趋势(
Chen et al ., 2017a ).但是, 该研究并未考虑团状福禄草植物整体的繁殖分配对环境因子的响应, 而仅是比较了其特定开花区域内单位面积的繁殖产出.本研究结果显示, 团状福禄草在开花阶段的繁殖分配在不同海拔种群间存在显著差异, 总体上随海拔上升而减少, 表明高海拔更加贫瘠的土壤条件(
Chen et al ., 2015b ,
2019 )可能限制了植物总体的资源获取量, 植物需要将有限的资源在营养生长与繁殖生长两方面之间做权衡(
Reekie & Avila-Sakar, 2005 ), 从而导致其在繁殖功能上的总体分配比例的下降.但是, 我们也发现, 部分低海拔种群(种群2)内植物个体的繁殖分配显著低于部分高海拔种群(种群3、4), 这表明, 海拔并非控制植物繁殖分配的唯一因素, 植物繁殖分配是多种生态因子共同作用的结果(
张婵等, 2020 ).种群2与种群1海拔相差较小, 但坡度相差较大, 其与种群3坡度相近, 但具有不同坡向(
表1 ).因此, 我们推测除了海拔因素, 坡度与坡向可能同时对其繁殖分配产生影响.一方面坡度大可能导致土壤养分流失更严重(
Bruce-Okine & Lal, 1975 ;
Donker & Damen, 1984 ), 从而引起资源胁迫的加剧, 并进而导致繁殖分配的减少.另一方面, 坡度和坡向通常会引起太阳辐射特征的变化, 如太阳入射角、入射时间在不同坡度和坡向上会发生变化(
田辉等, 2007 ), 从而引起植物可积累和利用的有效太阳辐射能量的变化.在高海拔地区, 太阳辐射对植物的生长与繁殖策略都起着重要的影响.如, 为了充分利用太阳辐射, 一些植物花器官进化出了“向日性”功能(
Zhang et al ., 2010 ).又如, 安第斯山脉中部高海拔岩石斜坡典型垫状植物
Azorella compacta 为了最大化利用太阳辐射, 其个体偏爱生长在向北的(南半球)坡度约为20°的斜坡(
Kleier et al ., 2015 ).因此, 我们推测由坡度与坡向变化引起的土壤养分状况及太阳辐射特征的变化, 可能同时对团状福禄草的繁殖分配起着重要的影响. ...
Recruitment of the high elevation cushion plant Arenaria polytrichoides is limited by competition, thus threatened by currently established vegetation
3
2020
... 团状福禄草(
Arenaria polytrichoides )为石竹科无心菜属(
Arenaria )多年生垫状草本植物, 是青藏高原特有种, 沿喜马拉雅—横断山高海拔(3 500-5 300 m)地区分布(
Wu et al ., 2013 ).营养生长自冰雪消融开始, 花期主要在6-7月, 不同分布区域及海拔的种群的花期存在一定的差异(
Chen et al ., 2017a ).并且, 团状福禄草具有雌全异株的性二态系统, 即同一种群中同时存在雌性植株和两性植株, 不同性别的个体在繁殖产出(果实及种子产量)方面存在显著差异, 并且受到海拔的影响(
Chen et al ., 2017a ).表现为较高海拔的雌性比例高于较低海拔的, 两性植株开花数目大于雌性植株但结实率显著低于雌性植株, 同时, 较低海拔种群内个体单位面积的果实与种子产量高于较高海拔种群的个体(
Chen et al ., 2017a ).此外, 团状福禄草被证实为横断山高山生态系统当中的关键种, 能有效改良微环境条件(温度、土壤营养及水分), 从而为其他低耐受物种提供适宜的微生境, 进而调节种群动态、提升生态系统物种多样性及地面生产力(
Yang et al ., 2010 ,
2017 ;
Chen et al ., 2015a ,
2019 ).然而, 团状福禄草种群的更新却受到种间竞争的强烈影响(
Chen et al ., 2020 ).研究者在近十年内对团状福禄草做了深入调查与研究(
Yang et al ., 2010 ,
2017 ;
Chen et al ., 2015a ,
2015b ,
2017a ,
2019 ), 但对其种群的繁殖与更新机制, 却直到最近才予以关注(
Chen et al ., 2017a ,
2020 ).理论上讲, 团状福禄草的每一个小分枝(
图1C )在生长季节都能开出一朵花, 然而, 我们在多年的野外研究中发现, 实际上, 其每年的开花数量极为有限, 只在其冠层表面的部分区域开出一定面积的花(
图1A 、1B).这一现象说明, 垫状植物的开花可能受到了资源的限制, 并有可能同时受到其他环境因子的影响.例如, 据野外观察, 团状福禄草开花方位(只在其冠层表面特定区域开放,
图1A 、1B)呈现出一定的规律性, 且在不同的种群之间存在差异.为了获取充足的太阳辐射能量从而满足自身繁殖需求, 一些高山植物的开花特性受到太阳辐射的强烈影响(
Pati?o et al ., 2002 ;
Galen, 2006 ;
Zhang et al ., 2010 ), 因此, 我们猜测团状福禄草的开花数量和方位可能受到资源和太阳辐射的双重影响.但其繁殖分配策略, 即个体对开花的资源分配策略对环境胁迫因子的响应方式, 目前却不为人知.深入探究高山生态系统中这一关键类群的种群更新与维持机制, 对揭示其种群繁殖的初始阶段(开花特性)对环境因子的响应机制以及这种响应机制在不同性别个体之间的差异性具有重要意义. ...
... ,
2020 ).理论上讲, 团状福禄草的每一个小分枝(
图1C )在生长季节都能开出一朵花, 然而, 我们在多年的野外研究中发现, 实际上, 其每年的开花数量极为有限, 只在其冠层表面的部分区域开出一定面积的花(
图1A 、1B).这一现象说明, 垫状植物的开花可能受到了资源的限制, 并有可能同时受到其他环境因子的影响.例如, 据野外观察, 团状福禄草开花方位(只在其冠层表面特定区域开放,
图1A 、1B)呈现出一定的规律性, 且在不同的种群之间存在差异.为了获取充足的太阳辐射能量从而满足自身繁殖需求, 一些高山植物的开花特性受到太阳辐射的强烈影响(
Pati?o et al ., 2002 ;
Galen, 2006 ;
Zhang et al ., 2010 ), 因此, 我们猜测团状福禄草的开花数量和方位可能受到资源和太阳辐射的双重影响.但其繁殖分配策略, 即个体对开花的资源分配策略对环境胁迫因子的响应方式, 目前却不为人知.深入探究高山生态系统中这一关键类群的种群更新与维持机制, 对揭示其种群繁殖的初始阶段(开花特性)对环境因子的响应机制以及这种响应机制在不同性别个体之间的差异性具有重要意义. ...
... 团状福禄草在研究区域的海拔范围内呈连续分布, 但受特定微生境特征及种间竞争强度的影响, 不同海拔带上的种群特征(大小、密度、结构)存在一定差异(
Chen et al ., 2020 ).团状福禄草是群落的奠基种, 粗糙红景天(
Rhodiola coccinea subsp.
scabrida )是群落的另一优势种, 群落中的常见物种还包括狭叶委陵菜(
Potentilla stenophylla )、七叶龙胆(
Gentiana arethusae var.
delicatula )、钉柱委陵菜(
Potentilla saundersiana )、镰萼喉毛花(
Comastoma falcatum )、圆穗蓼(
Polygonum macrophyllum )、滇边大黄(
Rheum delavayi )及早熟禾(
Poa spp.)等. ...
1
2019
... 首先, 将各方向上开花网格数相加, 得到各植株的总开花网格(面积), 其次, 分别将总开花网格数和每个方向的开花网格数都除以总网格数, 得到各个植株总的开花面积比(为了论述方便, 本研究中, 必要时将开花面积比称为“繁殖分配”)以及单个方位上的开花面积比, 并采用这些数据进行后续的比较分析.不同植株的大小(年龄)会对其资源分配策略产生影响, 即可能存在个体效应(
St?cklin & Favre, 1994 ;
Sakai & Sakai, 1995 ), 同时, 垫状植物冠层表面积大小通常与其年龄呈正相关关系(
Chen et al ., 2017a ), 因此, 计数了每个调查个体的总网格数(即表面积大小), 将其作为植株大小的替代来进行后续分析.首先, 为了探究不同种群以及不同性别植株的总开花面积与其个体大小之间的关系, 对植株总开花面积与其个体大小做了回归分析.其次, 为了探究海拔对植株大小的影响, 采用混合效应模型比较了不同海拔种群间植株的大小, 植株大小为因变量, 海拔为主因素, 植株个体编号为随机因素.最后, 为了探究开花面积比(即繁殖分配)如何受到海拔、性别和植株大小的影响, 使用混合效应模型对上述关系进行拟合.模型中, 开花面积比为因变量, 海拔、性别和植株大小为主因素, 植株个体编号为随机因素.此外, 由于自变量与因变量间存在多样的组合形式, 基于赤池信息量准则(AIC)权重选择最优模型, 最优模型具有最大的AIC权重, 因子在每个模型的AIC权重被用来计算模型平均系数, 而每个因子在所有模型中的AIC值权重之和(
w )被用来描述该因子的统计支持.同时, 评估了考虑交互作用的模型与上述模型的优劣, 校准AIC (
AICc )结果显示两种模型没有差异(
AICc = -402.1 vs.
AICc = -401.4), 但是交互作用不显著, 因而以上述的模型结果为准.由于各组数据不完全符合正态分布及方差齐性原则, 使用Kruskal-Wallis非参数检验分别比较同一种群内不同性别、不同方位之间开花面积比以及不同种群之间开花面积比的差异.所有统计分析及画图都通过R 3.5.1 (
R Core Team, 2018 )实现, Kruskal-Wallis非参数检验使用“agricolae”包(
de Felipe, 2019 ), 混合效应模型使用“coxme”包(
Therneau, 2018 ), 模型选择使用“MuMIn”包(
Bartoń, 2019 ). ...
Factors affecting relative seed fitness and female frequency in a gynodioecious species,
Silene acaulis 1
2001
... 横断山高海拔地区绝大多数植物种群的更新与扩张都依赖有性繁殖(
Peng et al ., 2012 ).植物对繁殖功能的资源分配, 在很大程度上决定了其种群未来的扩张速率.而作为植物实现繁殖功能的重要环节, 植物对开花的资源配置决定了植物的最终繁殖产出(
Reekie & Bazzaz, 2005 ).不同的环境因子在植物的每一个繁殖阶段都起着重要作用, 甚至一些因子(如资源量)直接决定着植物的繁殖分配及其产出(
Delph & Carroll, 2001 ).在我们的研究区域内, 环境胁迫程度(如土壤养分胁迫及低温胁迫)随海拔升高而增强(
Chen et al ., 2015b ,
2019 ).我们的研究结果显示, 随着海拔升高, 种群内植株个体呈现减小的趋势.植株大小对开花特性具有显著影响.一方面, 植株个体越大, 其总开花面积也越大, 但另一方面, 植株越大, 其总开花面积比越小(
表2 ), 并且这种趋势在不同性别植株间具有高度一致性(
图3 ), 说明资源限制是真实存在的, 即, 植物从环境中获取并分配给开花的资源量有限.此外, 该结果也说明植株分配到开花的资源增长速率可能低于植株个体的增长速率, 即繁殖分配的增加速率低于营养分配的增加速率.相应结果进一步暗示, 在面临资源限制或环境胁迫时, 团状福禄草的资源分配策略以保证植株存活(即增加营养分配)为首要目的.此外, 具有不同坡度及坡向的种群间开花特性也存在一定的变化,说明由坡向及坡度变化而引起的环境因子变化, 如太阳辐射(
田辉等, 2007 )和土壤养分(
曹舰艇等, 2019 ), 可能也同时对其繁殖分配策略具有一定的影响. ...
Gully system development and an assessment of gully initiation risk in Miocene deposits near Daroca: Spain
1
1984
... 前期研究发现, 团状福禄草植物个体表面单位面积的果实及种子产量随海拔上升而呈现总体下降趋势(
Chen et al ., 2017a ).但是, 该研究并未考虑团状福禄草植物整体的繁殖分配对环境因子的响应, 而仅是比较了其特定开花区域内单位面积的繁殖产出.本研究结果显示, 团状福禄草在开花阶段的繁殖分配在不同海拔种群间存在显著差异, 总体上随海拔上升而减少, 表明高海拔更加贫瘠的土壤条件(
Chen et al ., 2015b ,
2019 )可能限制了植物总体的资源获取量, 植物需要将有限的资源在营养生长与繁殖生长两方面之间做权衡(
Reekie & Avila-Sakar, 2005 ), 从而导致其在繁殖功能上的总体分配比例的下降.但是, 我们也发现, 部分低海拔种群(种群2)内植物个体的繁殖分配显著低于部分高海拔种群(种群3、4), 这表明, 海拔并非控制植物繁殖分配的唯一因素, 植物繁殖分配是多种生态因子共同作用的结果(
张婵等, 2020 ).种群2与种群1海拔相差较小, 但坡度相差较大, 其与种群3坡度相近, 但具有不同坡向(
表1 ).因此, 我们推测除了海拔因素, 坡度与坡向可能同时对其繁殖分配产生影响.一方面坡度大可能导致土壤养分流失更严重(
Bruce-Okine & Lal, 1975 ;
Donker & Damen, 1984 ), 从而引起资源胁迫的加剧, 并进而导致繁殖分配的减少.另一方面, 坡度和坡向通常会引起太阳辐射特征的变化, 如太阳入射角、入射时间在不同坡度和坡向上会发生变化(
田辉等, 2007 ), 从而引起植物可积累和利用的有效太阳辐射能量的变化.在高海拔地区, 太阳辐射对植物的生长与繁殖策略都起着重要的影响.如, 为了充分利用太阳辐射, 一些植物花器官进化出了“向日性”功能(
Zhang et al ., 2010 ).又如, 安第斯山脉中部高海拔岩石斜坡典型垫状植物
Azorella compacta 为了最大化利用太阳辐射, 其个体偏爱生长在向北的(南半球)坡度约为20°的斜坡(
Kleier et al ., 2015 ).因此, 我们推测由坡度与坡向变化引起的土壤养分状况及太阳辐射特征的变化, 可能同时对团状福禄草的繁殖分配起着重要的影响. ...
Solar furnaces or swamp coolers: costs and benefits of water use by solar-tracking flowers of the alpine snow buttercup,
Ranunculus adoneus 1
2006
... 团状福禄草(
Arenaria polytrichoides )为石竹科无心菜属(
Arenaria )多年生垫状草本植物, 是青藏高原特有种, 沿喜马拉雅—横断山高海拔(3 500-5 300 m)地区分布(
Wu et al ., 2013 ).营养生长自冰雪消融开始, 花期主要在6-7月, 不同分布区域及海拔的种群的花期存在一定的差异(
Chen et al ., 2017a ).并且, 团状福禄草具有雌全异株的性二态系统, 即同一种群中同时存在雌性植株和两性植株, 不同性别的个体在繁殖产出(果实及种子产量)方面存在显著差异, 并且受到海拔的影响(
Chen et al ., 2017a ).表现为较高海拔的雌性比例高于较低海拔的, 两性植株开花数目大于雌性植株但结实率显著低于雌性植株, 同时, 较低海拔种群内个体单位面积的果实与种子产量高于较高海拔种群的个体(
Chen et al ., 2017a ).此外, 团状福禄草被证实为横断山高山生态系统当中的关键种, 能有效改良微环境条件(温度、土壤营养及水分), 从而为其他低耐受物种提供适宜的微生境, 进而调节种群动态、提升生态系统物种多样性及地面生产力(
Yang et al ., 2010 ,
2017 ;
Chen et al ., 2015a ,
2019 ).然而, 团状福禄草种群的更新却受到种间竞争的强烈影响(
Chen et al ., 2020 ).研究者在近十年内对团状福禄草做了深入调查与研究(
Yang et al ., 2010 ,
2017 ;
Chen et al ., 2015a ,
2015b ,
2017a ,
2019 ), 但对其种群的繁殖与更新机制, 却直到最近才予以关注(
Chen et al ., 2017a ,
2020 ).理论上讲, 团状福禄草的每一个小分枝(
图1C )在生长季节都能开出一朵花, 然而, 我们在多年的野外研究中发现, 实际上, 其每年的开花数量极为有限, 只在其冠层表面的部分区域开出一定面积的花(
图1A 、1B).这一现象说明, 垫状植物的开花可能受到了资源的限制, 并有可能同时受到其他环境因子的影响.例如, 据野外观察, 团状福禄草开花方位(只在其冠层表面特定区域开放,
图1A 、1B)呈现出一定的规律性, 且在不同的种群之间存在差异.为了获取充足的太阳辐射能量从而满足自身繁殖需求, 一些高山植物的开花特性受到太阳辐射的强烈影响(
Pati?o et al ., 2002 ;
Galen, 2006 ;
Zhang et al ., 2010 ), 因此, 我们猜测团状福禄草的开花数量和方位可能受到资源和太阳辐射的双重影响.但其繁殖分配策略, 即个体对开花的资源分配策略对环境胁迫因子的响应方式, 目前却不为人知.深入探究高山生态系统中这一关键类群的种群更新与维持机制, 对揭示其种群繁殖的初始阶段(开花特性)对环境因子的响应机制以及这种响应机制在不同性别个体之间的差异性具有重要意义. ...
On the origin of and evolution of Australasian alpine cushion plants//Barrow BA
1
1986
... 垫状植物是一类具有球形或半球形结构的植物的总称, 是植物经过多年辐射性生长而形成的小枝紧密簇生结构(
图1 ;
Gibson & Hope, 1986 ), 因其适应世界上大多数山地和极地极端干旱和低温环境的能力而备受植物学家的关注(
Aubert et al ., 2014 ).并因其具有生态系统工程师的功能, 对其他植物具有“护理作用”, 可提高和维持生态系统中物种多样性(
Butterfield et al ., 2013 ;
Cavieres et al ., 2014 ;
Chen et al ., 2015a ,
2015b ,
2019 ), 而被认为是所在生态系统当中的奠基种(foundation species;
Kikvidze et al ., 2015 ). ...
2
1998
... 随着海拔逐渐上升, 环境条件变得更加严酷, 如: 海拔每上升100 m, 气温约下降0.6 ℃, 气压下降8-9 hPa (
K?rner, 2003 ), 土壤养分含量也随之减少(
曹舰艇等, 2019 ).面对高海拔地区低温、土壤养分贫瘠等恶劣环境因子, 高山植物在长期的进化过程中形成了丰富多样的特化适应性状或进化策略(
Sun et al ., 2014 ;
杨扬等, 2019 ), 种类丰富的植物多样性对维持高山生态系统的稳定性起到了关键作用(
Hooper et al ., 2005 ).滇西北处于横断山高山植物多样性中心区域(
郭辉军和龙春林, 1998 ;
Zhang et al. , 2015 ).长期以来, 青藏高原及横断山高海拔地区植物对极端环境因子的适应机制, 备受植物学家和生态学家的关注, 相关研究的深入开展, 加深了对该地区植物多样性形成及其进化适应机制的认识(
Sun et al ., 2014 ;
杨扬等, 2019 ). ...
... 研究区域位于滇西北德钦县白马雪山, 地理坐标为28.48° N, 98.99° E, 海拔范围为4 360?4 920 m, 生境类型从低至高依次为灌草甸、草甸-流石滩交错带和流石滩(
图1D ).滇西北处于横断山高山植物多样性中心区域(
郭辉军和龙春林, 1998 ;
Zhang et al ., 2015 ), 具有典型的季节性夏季季风气候, 以冷雨或雨夹雪为特征, 并伴有短时间的强烈太阳辐射, 年降水量680-790 mm, 年平均气温-1 ℃ (
张荣祖等, 1997 ).植物生长季节通常开始于5月中旬的融雪期, 结束于9月中下旬的夏季风停息期.我们前期的测量数据显示, 2015年6月中旬至9月下旬, 白马雪山海拔4 900 m处, 距离地面15 cm处的平均气温和湿度分别为(4.87 ± 0.04) ℃ 和(89.9 ± 0.24)%, 绝对最高气温和最低气温分别为17.80和-1.95 ℃, 昼夜温差大于10 ℃, 最高达16 ℃ (陈建国等, 未发表数据).此外, 土壤养分含量随海拔升高而普遍降低(
Chen et al ., 2015b ,
2019 ). ...
2
1998
... 随着海拔逐渐上升, 环境条件变得更加严酷, 如: 海拔每上升100 m, 气温约下降0.6 ℃, 气压下降8-9 hPa (
K?rner, 2003 ), 土壤养分含量也随之减少(
曹舰艇等, 2019 ).面对高海拔地区低温、土壤养分贫瘠等恶劣环境因子, 高山植物在长期的进化过程中形成了丰富多样的特化适应性状或进化策略(
Sun et al ., 2014 ;
杨扬等, 2019 ), 种类丰富的植物多样性对维持高山生态系统的稳定性起到了关键作用(
Hooper et al ., 2005 ).滇西北处于横断山高山植物多样性中心区域(
郭辉军和龙春林, 1998 ;
Zhang et al. , 2015 ).长期以来, 青藏高原及横断山高海拔地区植物对极端环境因子的适应机制, 备受植物学家和生态学家的关注, 相关研究的深入开展, 加深了对该地区植物多样性形成及其进化适应机制的认识(
Sun et al ., 2014 ;
杨扬等, 2019 ). ...
... 研究区域位于滇西北德钦县白马雪山, 地理坐标为28.48° N, 98.99° E, 海拔范围为4 360?4 920 m, 生境类型从低至高依次为灌草甸、草甸-流石滩交错带和流石滩(
图1D ).滇西北处于横断山高山植物多样性中心区域(
郭辉军和龙春林, 1998 ;
Zhang et al ., 2015 ), 具有典型的季节性夏季季风气候, 以冷雨或雨夹雪为特征, 并伴有短时间的强烈太阳辐射, 年降水量680-790 mm, 年平均气温-1 ℃ (
张荣祖等, 1997 ).植物生长季节通常开始于5月中旬的融雪期, 结束于9月中下旬的夏季风停息期.我们前期的测量数据显示, 2015年6月中旬至9月下旬, 白马雪山海拔4 900 m处, 距离地面15 cm处的平均气温和湿度分别为(4.87 ± 0.04) ℃ 和(89.9 ± 0.24)%, 绝对最高气温和最低气温分别为17.80和-1.95 ℃, 昼夜温差大于10 ℃, 最高达16 ℃ (陈建国等, 未发表数据).此外, 土壤养分含量随海拔升高而普遍降低(
Chen et al ., 2015b ,
2019 ). ...
Size-dependent allocation to sexual and vegetative reproduction in four clonal composites
1
1990
... 温度、资源(包括土壤养分、降水资源及太阳辐射量等)是限制高山植物生长与繁殖的关键因素(
Bloom et al ., 1985 ;
Chen et al ., 2017b ).为了应对高海拔地区贫瘠的土壤条件, 植物在自身资源获取及分配方面进化出了众多与环境相适应的资源分配策略.其中, 繁殖分配(即植物分配到繁殖器官的资源量)是植物适合度的关键表现(
Bazzaz et al ., 1987 ;
Weiner, 1988 ;
Hartnett, 1990 ;
Vega et al ., 2000 ).研究表明, 植物繁殖分配同时受到众多非生物与生物因子的控制(
Reekie & Bazzaz, 2005 ;
张婵等, 2020 ).非生物因子方面, 温度、水分、土壤营养等对植物的繁殖分配起着重要作用(
Reekie & Bazzaz, 2005 ;
Zhang et al ., 2014 ).而在生物因子方面, 种间竞争、传粉者特性以及植物自身性系统特征(
Ashman, 1994 ;
Case & Ashman, 2005 ;
Brzyski et al ., 2014 ;
Chen et al ., 2017a ,
2017b )对其繁殖分配策略都起着重要的影响.例如, 具有雌全异株性系统特征(即同一种植物同时存在雌性个体和两性个体)的植物, 其同一种群内的雌性个体与两性个体在繁殖分配策略上存在较大差异, 表现为雌性个体主要将资源分配到雌性功能(如胚珠及种子生产), 而两性个体则主要将资源分配到雄性功能(如花粉生产以及传粉者吸引)(
Ashman, 1994 ;
Chen et al ., 2017a ,
2017b ), 并且这种分配模式受到环境因子的强烈影响(
Chen et al ., 2017b ).为了应对资源的不足, 植物往往需要在营养分配(即分配到营养器官的资源量)与繁殖分配之间做出权衡, 以达到最优的繁殖适合度(
Reekie & Avila-Sakar, 2005 ).开花是植物繁殖的最初阶段, 植物开花的数目, 直接反映出植物在特定环境条件下的繁殖分配策略.另一方面, 开花数目的多少也可直接决定着植物未来的繁殖输出量(如果实及种子产量;
Reekie & Bazzaz, 2005 ). ...
Effects of biodiversity on ecosystem functioning: a consensus of current knowledge
1
2005
... 随着海拔逐渐上升, 环境条件变得更加严酷, 如: 海拔每上升100 m, 气温约下降0.6 ℃, 气压下降8-9 hPa (
K?rner, 2003 ), 土壤养分含量也随之减少(
曹舰艇等, 2019 ).面对高海拔地区低温、土壤养分贫瘠等恶劣环境因子, 高山植物在长期的进化过程中形成了丰富多样的特化适应性状或进化策略(
Sun et al ., 2014 ;
杨扬等, 2019 ), 种类丰富的植物多样性对维持高山生态系统的稳定性起到了关键作用(
Hooper et al ., 2005 ).滇西北处于横断山高山植物多样性中心区域(
郭辉军和龙春林, 1998 ;
Zhang et al. , 2015 ).长期以来, 青藏高原及横断山高海拔地区植物对极端环境因子的适应机制, 备受植物学家和生态学家的关注, 相关研究的深入开展, 加深了对该地区植物多样性形成及其进化适应机制的认识(
Sun et al ., 2014 ;
杨扬等, 2019 ). ...
The effects of foundation species on community assembly: a global study on alpine cushion plant communities
1
2015
... 垫状植物是一类具有球形或半球形结构的植物的总称, 是植物经过多年辐射性生长而形成的小枝紧密簇生结构(
图1 ;
Gibson & Hope, 1986 ), 因其适应世界上大多数山地和极地极端干旱和低温环境的能力而备受植物学家的关注(
Aubert et al ., 2014 ).并因其具有生态系统工程师的功能, 对其他植物具有“护理作用”, 可提高和维持生态系统中物种多样性(
Butterfield et al ., 2013 ;
Cavieres et al ., 2014 ;
Chen et al ., 2015a ,
2015b ,
2019 ), 而被认为是所在生态系统当中的奠基种(foundation species;
Kikvidze et al ., 2015 ). ...
Size class structure, growth rates, and orientation of the central Andean cushion
Azorella compacta 1
2015
... 前期研究发现, 团状福禄草植物个体表面单位面积的果实及种子产量随海拔上升而呈现总体下降趋势(
Chen et al ., 2017a ).但是, 该研究并未考虑团状福禄草植物整体的繁殖分配对环境因子的响应, 而仅是比较了其特定开花区域内单位面积的繁殖产出.本研究结果显示, 团状福禄草在开花阶段的繁殖分配在不同海拔种群间存在显著差异, 总体上随海拔上升而减少, 表明高海拔更加贫瘠的土壤条件(
Chen et al ., 2015b ,
2019 )可能限制了植物总体的资源获取量, 植物需要将有限的资源在营养生长与繁殖生长两方面之间做权衡(
Reekie & Avila-Sakar, 2005 ), 从而导致其在繁殖功能上的总体分配比例的下降.但是, 我们也发现, 部分低海拔种群(种群2)内植物个体的繁殖分配显著低于部分高海拔种群(种群3、4), 这表明, 海拔并非控制植物繁殖分配的唯一因素, 植物繁殖分配是多种生态因子共同作用的结果(
张婵等, 2020 ).种群2与种群1海拔相差较小, 但坡度相差较大, 其与种群3坡度相近, 但具有不同坡向(
表1 ).因此, 我们推测除了海拔因素, 坡度与坡向可能同时对其繁殖分配产生影响.一方面坡度大可能导致土壤养分流失更严重(
Bruce-Okine & Lal, 1975 ;
Donker & Damen, 1984 ), 从而引起资源胁迫的加剧, 并进而导致繁殖分配的减少.另一方面, 坡度和坡向通常会引起太阳辐射特征的变化, 如太阳入射角、入射时间在不同坡度和坡向上会发生变化(
田辉等, 2007 ), 从而引起植物可积累和利用的有效太阳辐射能量的变化.在高海拔地区, 太阳辐射对植物的生长与繁殖策略都起着重要的影响.如, 为了充分利用太阳辐射, 一些植物花器官进化出了“向日性”功能(
Zhang et al ., 2010 ).又如, 安第斯山脉中部高海拔岩石斜坡典型垫状植物
Azorella compacta 为了最大化利用太阳辐射, 其个体偏爱生长在向北的(南半球)坡度约为20°的斜坡(
Kleier et al ., 2015 ).因此, 我们推测由坡度与坡向变化引起的土壤养分状况及太阳辐射特征的变化, 可能同时对团状福禄草的繁殖分配起着重要的影响. ...
1
2003
... 随着海拔逐渐上升, 环境条件变得更加严酷, 如: 海拔每上升100 m, 气温约下降0.6 ℃, 气压下降8-9 hPa (
K?rner, 2003 ), 土壤养分含量也随之减少(
曹舰艇等, 2019 ).面对高海拔地区低温、土壤养分贫瘠等恶劣环境因子, 高山植物在长期的进化过程中形成了丰富多样的特化适应性状或进化策略(
Sun et al ., 2014 ;
杨扬等, 2019 ), 种类丰富的植物多样性对维持高山生态系统的稳定性起到了关键作用(
Hooper et al ., 2005 ).滇西北处于横断山高山植物多样性中心区域(
郭辉军和龙春林, 1998 ;
Zhang et al. , 2015 ).长期以来, 青藏高原及横断山高海拔地区植物对极端环境因子的适应机制, 备受植物学家和生态学家的关注, 相关研究的深入开展, 加深了对该地区植物多样性形成及其进化适应机制的认识(
Sun et al ., 2014 ;
杨扬等, 2019 ). ...
Plant-pollinator relationships at two altitudes in the Andes of Mendoza, Argentina
1
2002
... 此外, 团状福禄草具有性二态系统, 即雌性植株与两性植株同时存在于同一种群中.不同性别的植物个体获得繁殖适合度的方式不同, 雌性个体主要通过胚珠及种子产出获得繁殖适合度, 而两性植株主要通过花粉与种子产出获得繁殖适合度(
Charlesworth & Charlesworth, 1978 ;
Charlesworth & Morgan, 1991 ;
Ashman, 1994 ).因此, 不同性别的植株个体在繁殖分配策略上存在一定的差异性, 这种差异性直接反映了不同性别个体对特定环境因子的适应策略(
Case & Ashman, 2005 ;
Ashman, 2006 ;
Chen et al ., 2017b ).例如, 同样分布于滇西北高海拔地区, 且同样具有雌全异株性系统的细叶蓝钟花(
Cyananthus delavayi ), 在面对环境因子胁迫时, 雌性植株主要将资源分配到种子生产, 而两性植株则主要将资源分配到花粉生产以及对传粉昆虫的吸引功能方面(花瓣)(
Chen et al ., 2017b ).我们的研究结果表明, 总体而言, 两性植株对开花的资源分配比例要显著高于雌性植株(
图2B ), 但是, 其差异程度受到海拔因素的影响.具体而言, 低海拔种群两性植株分配给开花的资源比例显著高于雌性植株, 而高海拔种群不同性别植株对开花的资源分配差异性不显著(
图4 ).
Chen等(2017a) 研究发现, 在任何海拔条件下, 团状福禄草两性植株单位面积的开花数量始终大于雌性植株, 但其真实繁殖产出(单位面积果实和种子产量)却始终低于雌性植株.但是, 本研究发现, 尽管不同性别的个体之间在低海拔处的总繁殖分配比例存在差异性, 但是随着海拔升高, 环境胁迫程度增加, 这种差异性趋于消失(
图4 ).我们猜测, 在高海拔环境胁迫加剧条件下, 不同性别的植物个体在资源权衡分配方面采取了相同的策略.另一方面, 除了物理胁迫的加剧, 生物胁迫也同时加剧, 如, 传粉昆虫多样性及多度的下降(
Arroyo et al ., 1982 ;
Medan et al ., 2002 )可能引起两性植株雄性适合度输出(花粉传播)的降低, 从而引起其分配到开花的资源随之减少. ...
The ecological role of orientation in tropical convolvulaceous flowers
1
2002
... 团状福禄草(
Arenaria polytrichoides )为石竹科无心菜属(
Arenaria )多年生垫状草本植物, 是青藏高原特有种, 沿喜马拉雅—横断山高海拔(3 500-5 300 m)地区分布(
Wu et al ., 2013 ).营养生长自冰雪消融开始, 花期主要在6-7月, 不同分布区域及海拔的种群的花期存在一定的差异(
Chen et al ., 2017a ).并且, 团状福禄草具有雌全异株的性二态系统, 即同一种群中同时存在雌性植株和两性植株, 不同性别的个体在繁殖产出(果实及种子产量)方面存在显著差异, 并且受到海拔的影响(
Chen et al ., 2017a ).表现为较高海拔的雌性比例高于较低海拔的, 两性植株开花数目大于雌性植株但结实率显著低于雌性植株, 同时, 较低海拔种群内个体单位面积的果实与种子产量高于较高海拔种群的个体(
Chen et al ., 2017a ).此外, 团状福禄草被证实为横断山高山生态系统当中的关键种, 能有效改良微环境条件(温度、土壤营养及水分), 从而为其他低耐受物种提供适宜的微生境, 进而调节种群动态、提升生态系统物种多样性及地面生产力(
Yang et al ., 2010 ,
2017 ;
Chen et al ., 2015a ,
2019 ).然而, 团状福禄草种群的更新却受到种间竞争的强烈影响(
Chen et al ., 2020 ).研究者在近十年内对团状福禄草做了深入调查与研究(
Yang et al ., 2010 ,
2017 ;
Chen et al ., 2015a ,
2015b ,
2017a ,
2019 ), 但对其种群的繁殖与更新机制, 却直到最近才予以关注(
Chen et al ., 2017a ,
2020 ).理论上讲, 团状福禄草的每一个小分枝(
图1C )在生长季节都能开出一朵花, 然而, 我们在多年的野外研究中发现, 实际上, 其每年的开花数量极为有限, 只在其冠层表面的部分区域开出一定面积的花(
图1A 、1B).这一现象说明, 垫状植物的开花可能受到了资源的限制, 并有可能同时受到其他环境因子的影响.例如, 据野外观察, 团状福禄草开花方位(只在其冠层表面特定区域开放,
图1A 、1B)呈现出一定的规律性, 且在不同的种群之间存在差异.为了获取充足的太阳辐射能量从而满足自身繁殖需求, 一些高山植物的开花特性受到太阳辐射的强烈影响(
Pati?o et al ., 2002 ;
Galen, 2006 ;
Zhang et al ., 2010 ), 因此, 我们猜测团状福禄草的开花数量和方位可能受到资源和太阳辐射的双重影响.但其繁殖分配策略, 即个体对开花的资源分配策略对环境胁迫因子的响应方式, 目前却不为人知.深入探究高山生态系统中这一关键类群的种群更新与维持机制, 对揭示其种群繁殖的初始阶段(开花特性)对环境因子的响应机制以及这种响应机制在不同性别个体之间的差异性具有重要意义. ...
Patterns of flower morphology and sexual systems in the subnival belt of the Hengduan Mountains, SW China
1
2012
... 横断山高海拔地区绝大多数植物种群的更新与扩张都依赖有性繁殖(
Peng et al ., 2012 ).植物对繁殖功能的资源分配, 在很大程度上决定了其种群未来的扩张速率.而作为植物实现繁殖功能的重要环节, 植物对开花的资源配置决定了植物的最终繁殖产出(
Reekie & Bazzaz, 2005 ).不同的环境因子在植物的每一个繁殖阶段都起着重要作用, 甚至一些因子(如资源量)直接决定着植物的繁殖分配及其产出(
Delph & Carroll, 2001 ).在我们的研究区域内, 环境胁迫程度(如土壤养分胁迫及低温胁迫)随海拔升高而增强(
Chen et al ., 2015b ,
2019 ).我们的研究结果显示, 随着海拔升高, 种群内植株个体呈现减小的趋势.植株大小对开花特性具有显著影响.一方面, 植株个体越大, 其总开花面积也越大, 但另一方面, 植株越大, 其总开花面积比越小(
表2 ), 并且这种趋势在不同性别植株间具有高度一致性(
图3 ), 说明资源限制是真实存在的, 即, 植物从环境中获取并分配给开花的资源量有限.此外, 该结果也说明植株分配到开花的资源增长速率可能低于植株个体的增长速率, 即繁殖分配的增加速率低于营养分配的增加速率.相应结果进一步暗示, 在面临资源限制或环境胁迫时, 团状福禄草的资源分配策略以保证植株存活(即增加营养分配)为首要目的.此外, 具有不同坡度及坡向的种群间开花特性也存在一定的变化,说明由坡向及坡度变化而引起的环境因子变化, 如太阳辐射(
田辉等, 2007 )和土壤养分(
曹舰艇等, 2019 ), 可能也同时对其繁殖分配策略具有一定的影响. ...
1
2018
... 首先, 将各方向上开花网格数相加, 得到各植株的总开花网格(面积), 其次, 分别将总开花网格数和每个方向的开花网格数都除以总网格数, 得到各个植株总的开花面积比(为了论述方便, 本研究中, 必要时将开花面积比称为“繁殖分配”)以及单个方位上的开花面积比, 并采用这些数据进行后续的比较分析.不同植株的大小(年龄)会对其资源分配策略产生影响, 即可能存在个体效应(
St?cklin & Favre, 1994 ;
Sakai & Sakai, 1995 ), 同时, 垫状植物冠层表面积大小通常与其年龄呈正相关关系(
Chen et al ., 2017a ), 因此, 计数了每个调查个体的总网格数(即表面积大小), 将其作为植株大小的替代来进行后续分析.首先, 为了探究不同种群以及不同性别植株的总开花面积与其个体大小之间的关系, 对植株总开花面积与其个体大小做了回归分析.其次, 为了探究海拔对植株大小的影响, 采用混合效应模型比较了不同海拔种群间植株的大小, 植株大小为因变量, 海拔为主因素, 植株个体编号为随机因素.最后, 为了探究开花面积比(即繁殖分配)如何受到海拔、性别和植株大小的影响, 使用混合效应模型对上述关系进行拟合.模型中, 开花面积比为因变量, 海拔、性别和植株大小为主因素, 植株个体编号为随机因素.此外, 由于自变量与因变量间存在多样的组合形式, 基于赤池信息量准则(AIC)权重选择最优模型, 最优模型具有最大的AIC权重, 因子在每个模型的AIC权重被用来计算模型平均系数, 而每个因子在所有模型中的AIC值权重之和(
w )被用来描述该因子的统计支持.同时, 评估了考虑交互作用的模型与上述模型的优劣, 校准AIC (
AICc )结果显示两种模型没有差异(
AICc = -402.1 vs.
AICc = -401.4), 但是交互作用不显著, 因而以上述的模型结果为准.由于各组数据不完全符合正态分布及方差齐性原则, 使用Kruskal-Wallis非参数检验分别比较同一种群内不同性别、不同方位之间开花面积比以及不同种群之间开花面积比的差异.所有统计分析及画图都通过R 3.5.1 (
R Core Team, 2018 )实现, Kruskal-Wallis非参数检验使用“agricolae”包(
de Felipe, 2019 ), 混合效应模型使用“coxme”包(
Therneau, 2018 ), 模型选择使用“MuMIn”包(
Bartoń, 2019 ). ...
The shape of the trade-off function between reproduction and growth//Reekie EG, Bazzaz FA
2
2005
... 温度、资源(包括土壤养分、降水资源及太阳辐射量等)是限制高山植物生长与繁殖的关键因素(
Bloom et al ., 1985 ;
Chen et al ., 2017b ).为了应对高海拔地区贫瘠的土壤条件, 植物在自身资源获取及分配方面进化出了众多与环境相适应的资源分配策略.其中, 繁殖分配(即植物分配到繁殖器官的资源量)是植物适合度的关键表现(
Bazzaz et al ., 1987 ;
Weiner, 1988 ;
Hartnett, 1990 ;
Vega et al ., 2000 ).研究表明, 植物繁殖分配同时受到众多非生物与生物因子的控制(
Reekie & Bazzaz, 2005 ;
张婵等, 2020 ).非生物因子方面, 温度、水分、土壤营养等对植物的繁殖分配起着重要作用(
Reekie & Bazzaz, 2005 ;
Zhang et al ., 2014 ).而在生物因子方面, 种间竞争、传粉者特性以及植物自身性系统特征(
Ashman, 1994 ;
Case & Ashman, 2005 ;
Brzyski et al ., 2014 ;
Chen et al ., 2017a ,
2017b )对其繁殖分配策略都起着重要的影响.例如, 具有雌全异株性系统特征(即同一种植物同时存在雌性个体和两性个体)的植物, 其同一种群内的雌性个体与两性个体在繁殖分配策略上存在较大差异, 表现为雌性个体主要将资源分配到雌性功能(如胚珠及种子生产), 而两性个体则主要将资源分配到雄性功能(如花粉生产以及传粉者吸引)(
Ashman, 1994 ;
Chen et al ., 2017a ,
2017b ), 并且这种分配模式受到环境因子的强烈影响(
Chen et al ., 2017b ).为了应对资源的不足, 植物往往需要在营养分配(即分配到营养器官的资源量)与繁殖分配之间做出权衡, 以达到最优的繁殖适合度(
Reekie & Avila-Sakar, 2005 ).开花是植物繁殖的最初阶段, 植物开花的数目, 直接反映出植物在特定环境条件下的繁殖分配策略.另一方面, 开花数目的多少也可直接决定着植物未来的繁殖输出量(如果实及种子产量;
Reekie & Bazzaz, 2005 ). ...
... 前期研究发现, 团状福禄草植物个体表面单位面积的果实及种子产量随海拔上升而呈现总体下降趋势(
Chen et al ., 2017a ).但是, 该研究并未考虑团状福禄草植物整体的繁殖分配对环境因子的响应, 而仅是比较了其特定开花区域内单位面积的繁殖产出.本研究结果显示, 团状福禄草在开花阶段的繁殖分配在不同海拔种群间存在显著差异, 总体上随海拔上升而减少, 表明高海拔更加贫瘠的土壤条件(
Chen et al ., 2015b ,
2019 )可能限制了植物总体的资源获取量, 植物需要将有限的资源在营养生长与繁殖生长两方面之间做权衡(
Reekie & Avila-Sakar, 2005 ), 从而导致其在繁殖功能上的总体分配比例的下降.但是, 我们也发现, 部分低海拔种群(种群2)内植物个体的繁殖分配显著低于部分高海拔种群(种群3、4), 这表明, 海拔并非控制植物繁殖分配的唯一因素, 植物繁殖分配是多种生态因子共同作用的结果(
张婵等, 2020 ).种群2与种群1海拔相差较小, 但坡度相差较大, 其与种群3坡度相近, 但具有不同坡向(
表1 ).因此, 我们推测除了海拔因素, 坡度与坡向可能同时对其繁殖分配产生影响.一方面坡度大可能导致土壤养分流失更严重(
Bruce-Okine & Lal, 1975 ;
Donker & Damen, 1984 ), 从而引起资源胁迫的加剧, 并进而导致繁殖分配的减少.另一方面, 坡度和坡向通常会引起太阳辐射特征的变化, 如太阳入射角、入射时间在不同坡度和坡向上会发生变化(
田辉等, 2007 ), 从而引起植物可积累和利用的有效太阳辐射能量的变化.在高海拔地区, 太阳辐射对植物的生长与繁殖策略都起着重要的影响.如, 为了充分利用太阳辐射, 一些植物花器官进化出了“向日性”功能(
Zhang et al ., 2010 ).又如, 安第斯山脉中部高海拔岩石斜坡典型垫状植物
Azorella compacta 为了最大化利用太阳辐射, 其个体偏爱生长在向北的(南半球)坡度约为20°的斜坡(
Kleier et al ., 2015 ).因此, 我们推测由坡度与坡向变化引起的土壤养分状况及太阳辐射特征的变化, 可能同时对团状福禄草的繁殖分配起着重要的影响. ...
4
2005
... 温度、资源(包括土壤养分、降水资源及太阳辐射量等)是限制高山植物生长与繁殖的关键因素(
Bloom et al ., 1985 ;
Chen et al ., 2017b ).为了应对高海拔地区贫瘠的土壤条件, 植物在自身资源获取及分配方面进化出了众多与环境相适应的资源分配策略.其中, 繁殖分配(即植物分配到繁殖器官的资源量)是植物适合度的关键表现(
Bazzaz et al ., 1987 ;
Weiner, 1988 ;
Hartnett, 1990 ;
Vega et al ., 2000 ).研究表明, 植物繁殖分配同时受到众多非生物与生物因子的控制(
Reekie & Bazzaz, 2005 ;
张婵等, 2020 ).非生物因子方面, 温度、水分、土壤营养等对植物的繁殖分配起着重要作用(
Reekie & Bazzaz, 2005 ;
Zhang et al ., 2014 ).而在生物因子方面, 种间竞争、传粉者特性以及植物自身性系统特征(
Ashman, 1994 ;
Case & Ashman, 2005 ;
Brzyski et al ., 2014 ;
Chen et al ., 2017a ,
2017b )对其繁殖分配策略都起着重要的影响.例如, 具有雌全异株性系统特征(即同一种植物同时存在雌性个体和两性个体)的植物, 其同一种群内的雌性个体与两性个体在繁殖分配策略上存在较大差异, 表现为雌性个体主要将资源分配到雌性功能(如胚珠及种子生产), 而两性个体则主要将资源分配到雄性功能(如花粉生产以及传粉者吸引)(
Ashman, 1994 ;
Chen et al ., 2017a ,
2017b ), 并且这种分配模式受到环境因子的强烈影响(
Chen et al ., 2017b ).为了应对资源的不足, 植物往往需要在营养分配(即分配到营养器官的资源量)与繁殖分配之间做出权衡, 以达到最优的繁殖适合度(
Reekie & Avila-Sakar, 2005 ).开花是植物繁殖的最初阶段, 植物开花的数目, 直接反映出植物在特定环境条件下的繁殖分配策略.另一方面, 开花数目的多少也可直接决定着植物未来的繁殖输出量(如果实及种子产量;
Reekie & Bazzaz, 2005 ). ...
... ).非生物因子方面, 温度、水分、土壤营养等对植物的繁殖分配起着重要作用(
Reekie & Bazzaz, 2005 ;
Zhang et al ., 2014 ).而在生物因子方面, 种间竞争、传粉者特性以及植物自身性系统特征(
Ashman, 1994 ;
Case & Ashman, 2005 ;
Brzyski et al ., 2014 ;
Chen et al ., 2017a ,
2017b )对其繁殖分配策略都起着重要的影响.例如, 具有雌全异株性系统特征(即同一种植物同时存在雌性个体和两性个体)的植物, 其同一种群内的雌性个体与两性个体在繁殖分配策略上存在较大差异, 表现为雌性个体主要将资源分配到雌性功能(如胚珠及种子生产), 而两性个体则主要将资源分配到雄性功能(如花粉生产以及传粉者吸引)(
Ashman, 1994 ;
Chen et al ., 2017a ,
2017b ), 并且这种分配模式受到环境因子的强烈影响(
Chen et al ., 2017b ).为了应对资源的不足, 植物往往需要在营养分配(即分配到营养器官的资源量)与繁殖分配之间做出权衡, 以达到最优的繁殖适合度(
Reekie & Avila-Sakar, 2005 ).开花是植物繁殖的最初阶段, 植物开花的数目, 直接反映出植物在特定环境条件下的繁殖分配策略.另一方面, 开花数目的多少也可直接决定着植物未来的繁殖输出量(如果实及种子产量;
Reekie & Bazzaz, 2005 ). ...
... ).开花是植物繁殖的最初阶段, 植物开花的数目, 直接反映出植物在特定环境条件下的繁殖分配策略.另一方面, 开花数目的多少也可直接决定着植物未来的繁殖输出量(如果实及种子产量;
Reekie & Bazzaz, 2005 ). ...
... 横断山高海拔地区绝大多数植物种群的更新与扩张都依赖有性繁殖(
Peng et al ., 2012 ).植物对繁殖功能的资源分配, 在很大程度上决定了其种群未来的扩张速率.而作为植物实现繁殖功能的重要环节, 植物对开花的资源配置决定了植物的最终繁殖产出(
Reekie & Bazzaz, 2005 ).不同的环境因子在植物的每一个繁殖阶段都起着重要作用, 甚至一些因子(如资源量)直接决定着植物的繁殖分配及其产出(
Delph & Carroll, 2001 ).在我们的研究区域内, 环境胁迫程度(如土壤养分胁迫及低温胁迫)随海拔升高而增强(
Chen et al ., 2015b ,
2019 ).我们的研究结果显示, 随着海拔升高, 种群内植株个体呈现减小的趋势.植株大小对开花特性具有显著影响.一方面, 植株个体越大, 其总开花面积也越大, 但另一方面, 植株越大, 其总开花面积比越小(
表2 ), 并且这种趋势在不同性别植株间具有高度一致性(
图3 ), 说明资源限制是真实存在的, 即, 植物从环境中获取并分配给开花的资源量有限.此外, 该结果也说明植株分配到开花的资源增长速率可能低于植株个体的增长速率, 即繁殖分配的增加速率低于营养分配的增加速率.相应结果进一步暗示, 在面临资源限制或环境胁迫时, 团状福禄草的资源分配策略以保证植株存活(即增加营养分配)为首要目的.此外, 具有不同坡度及坡向的种群间开花特性也存在一定的变化,说明由坡向及坡度变化而引起的环境因子变化, 如太阳辐射(
田辉等, 2007 )和土壤养分(
曹舰艇等, 2019 ), 可能也同时对其繁殖分配策略具有一定的影响. ...
Flower size-dependent variation in seed size: theory and a test
1
1995
... 首先, 将各方向上开花网格数相加, 得到各植株的总开花网格(面积), 其次, 分别将总开花网格数和每个方向的开花网格数都除以总网格数, 得到各个植株总的开花面积比(为了论述方便, 本研究中, 必要时将开花面积比称为“繁殖分配”)以及单个方位上的开花面积比, 并采用这些数据进行后续的比较分析.不同植株的大小(年龄)会对其资源分配策略产生影响, 即可能存在个体效应(
St?cklin & Favre, 1994 ;
Sakai & Sakai, 1995 ), 同时, 垫状植物冠层表面积大小通常与其年龄呈正相关关系(
Chen et al ., 2017a ), 因此, 计数了每个调查个体的总网格数(即表面积大小), 将其作为植株大小的替代来进行后续分析.首先, 为了探究不同种群以及不同性别植株的总开花面积与其个体大小之间的关系, 对植株总开花面积与其个体大小做了回归分析.其次, 为了探究海拔对植株大小的影响, 采用混合效应模型比较了不同海拔种群间植株的大小, 植株大小为因变量, 海拔为主因素, 植株个体编号为随机因素.最后, 为了探究开花面积比(即繁殖分配)如何受到海拔、性别和植株大小的影响, 使用混合效应模型对上述关系进行拟合.模型中, 开花面积比为因变量, 海拔、性别和植株大小为主因素, 植株个体编号为随机因素.此外, 由于自变量与因变量间存在多样的组合形式, 基于赤池信息量准则(AIC)权重选择最优模型, 最优模型具有最大的AIC权重, 因子在每个模型的AIC权重被用来计算模型平均系数, 而每个因子在所有模型中的AIC值权重之和(
w )被用来描述该因子的统计支持.同时, 评估了考虑交互作用的模型与上述模型的优劣, 校准AIC (
AICc )结果显示两种模型没有差异(
AICc = -402.1 vs.
AICc = -401.4), 但是交互作用不显著, 因而以上述的模型结果为准.由于各组数据不完全符合正态分布及方差齐性原则, 使用Kruskal-Wallis非参数检验分别比较同一种群内不同性别、不同方位之间开花面积比以及不同种群之间开花面积比的差异.所有统计分析及画图都通过R 3.5.1 (
R Core Team, 2018 )实现, Kruskal-Wallis非参数检验使用“agricolae”包(
de Felipe, 2019 ), 混合效应模型使用“coxme”包(
Therneau, 2018 ), 模型选择使用“MuMIn”包(
Bartoń, 2019 ). ...
Effects of plant size and morphological constraints on variation in reproductive components in 2 related species of
Epilobium 1
1994
... 首先, 将各方向上开花网格数相加, 得到各植株的总开花网格(面积), 其次, 分别将总开花网格数和每个方向的开花网格数都除以总网格数, 得到各个植株总的开花面积比(为了论述方便, 本研究中, 必要时将开花面积比称为“繁殖分配”)以及单个方位上的开花面积比, 并采用这些数据进行后续的比较分析.不同植株的大小(年龄)会对其资源分配策略产生影响, 即可能存在个体效应(
St?cklin & Favre, 1994 ;
Sakai & Sakai, 1995 ), 同时, 垫状植物冠层表面积大小通常与其年龄呈正相关关系(
Chen et al ., 2017a ), 因此, 计数了每个调查个体的总网格数(即表面积大小), 将其作为植株大小的替代来进行后续分析.首先, 为了探究不同种群以及不同性别植株的总开花面积与其个体大小之间的关系, 对植株总开花面积与其个体大小做了回归分析.其次, 为了探究海拔对植株大小的影响, 采用混合效应模型比较了不同海拔种群间植株的大小, 植株大小为因变量, 海拔为主因素, 植株个体编号为随机因素.最后, 为了探究开花面积比(即繁殖分配)如何受到海拔、性别和植株大小的影响, 使用混合效应模型对上述关系进行拟合.模型中, 开花面积比为因变量, 海拔、性别和植株大小为主因素, 植株个体编号为随机因素.此外, 由于自变量与因变量间存在多样的组合形式, 基于赤池信息量准则(AIC)权重选择最优模型, 最优模型具有最大的AIC权重, 因子在每个模型的AIC权重被用来计算模型平均系数, 而每个因子在所有模型中的AIC值权重之和(
w )被用来描述该因子的统计支持.同时, 评估了考虑交互作用的模型与上述模型的优劣, 校准AIC (
AICc )结果显示两种模型没有差异(
AICc = -402.1 vs.
AICc = -401.4), 但是交互作用不显著, 因而以上述的模型结果为准.由于各组数据不完全符合正态分布及方差齐性原则, 使用Kruskal-Wallis非参数检验分别比较同一种群内不同性别、不同方位之间开花面积比以及不同种群之间开花面积比的差异.所有统计分析及画图都通过R 3.5.1 (
R Core Team, 2018 )实现, Kruskal-Wallis非参数检验使用“agricolae”包(
de Felipe, 2019 ), 混合效应模型使用“coxme”包(
Therneau, 2018 ), 模型选择使用“MuMIn”包(
Bartoń, 2019 ). ...
Survival and reproduction of plant species in the Qinghai-Tibet Plateau
2
2014
... 随着海拔逐渐上升, 环境条件变得更加严酷, 如: 海拔每上升100 m, 气温约下降0.6 ℃, 气压下降8-9 hPa (
K?rner, 2003 ), 土壤养分含量也随之减少(
曹舰艇等, 2019 ).面对高海拔地区低温、土壤养分贫瘠等恶劣环境因子, 高山植物在长期的进化过程中形成了丰富多样的特化适应性状或进化策略(
Sun et al ., 2014 ;
杨扬等, 2019 ), 种类丰富的植物多样性对维持高山生态系统的稳定性起到了关键作用(
Hooper et al ., 2005 ).滇西北处于横断山高山植物多样性中心区域(
郭辉军和龙春林, 1998 ;
Zhang et al. , 2015 ).长期以来, 青藏高原及横断山高海拔地区植物对极端环境因子的适应机制, 备受植物学家和生态学家的关注, 相关研究的深入开展, 加深了对该地区植物多样性形成及其进化适应机制的认识(
Sun et al ., 2014 ;
杨扬等, 2019 ). ...
... ).长期以来, 青藏高原及横断山高海拔地区植物对极端环境因子的适应机制, 备受植物学家和生态学家的关注, 相关研究的深入开展, 加深了对该地区植物多样性形成及其进化适应机制的认识(
Sun et al ., 2014 ;
杨扬等, 2019 ). ...
1
2018
... 首先, 将各方向上开花网格数相加, 得到各植株的总开花网格(面积), 其次, 分别将总开花网格数和每个方向的开花网格数都除以总网格数, 得到各个植株总的开花面积比(为了论述方便, 本研究中, 必要时将开花面积比称为“繁殖分配”)以及单个方位上的开花面积比, 并采用这些数据进行后续的比较分析.不同植株的大小(年龄)会对其资源分配策略产生影响, 即可能存在个体效应(
St?cklin & Favre, 1994 ;
Sakai & Sakai, 1995 ), 同时, 垫状植物冠层表面积大小通常与其年龄呈正相关关系(
Chen et al ., 2017a ), 因此, 计数了每个调查个体的总网格数(即表面积大小), 将其作为植株大小的替代来进行后续分析.首先, 为了探究不同种群以及不同性别植株的总开花面积与其个体大小之间的关系, 对植株总开花面积与其个体大小做了回归分析.其次, 为了探究海拔对植株大小的影响, 采用混合效应模型比较了不同海拔种群间植株的大小, 植株大小为因变量, 海拔为主因素, 植株个体编号为随机因素.最后, 为了探究开花面积比(即繁殖分配)如何受到海拔、性别和植株大小的影响, 使用混合效应模型对上述关系进行拟合.模型中, 开花面积比为因变量, 海拔、性别和植株大小为主因素, 植株个体编号为随机因素.此外, 由于自变量与因变量间存在多样的组合形式, 基于赤池信息量准则(AIC)权重选择最优模型, 最优模型具有最大的AIC权重, 因子在每个模型的AIC权重被用来计算模型平均系数, 而每个因子在所有模型中的AIC值权重之和(
w )被用来描述该因子的统计支持.同时, 评估了考虑交互作用的模型与上述模型的优劣, 校准AIC (
AICc )结果显示两种模型没有差异(
AICc = -402.1 vs.
AICc = -401.4), 但是交互作用不显著, 因而以上述的模型结果为准.由于各组数据不完全符合正态分布及方差齐性原则, 使用Kruskal-Wallis非参数检验分别比较同一种群内不同性别、不同方位之间开花面积比以及不同种群之间开花面积比的差异.所有统计分析及画图都通过R 3.5.1 (
R Core Team, 2018 )实现, Kruskal-Wallis非参数检验使用“agricolae”包(
de Felipe, 2019 ), 混合效应模型使用“coxme”包(
Therneau, 2018 ), 模型选择使用“MuMIn”包(
Bartoń, 2019 ). ...
复杂地形下黑河流域的太阳辐射计算
3
2007
... 横断山高海拔地区绝大多数植物种群的更新与扩张都依赖有性繁殖(
Peng et al ., 2012 ).植物对繁殖功能的资源分配, 在很大程度上决定了其种群未来的扩张速率.而作为植物实现繁殖功能的重要环节, 植物对开花的资源配置决定了植物的最终繁殖产出(
Reekie & Bazzaz, 2005 ).不同的环境因子在植物的每一个繁殖阶段都起着重要作用, 甚至一些因子(如资源量)直接决定着植物的繁殖分配及其产出(
Delph & Carroll, 2001 ).在我们的研究区域内, 环境胁迫程度(如土壤养分胁迫及低温胁迫)随海拔升高而增强(
Chen et al ., 2015b ,
2019 ).我们的研究结果显示, 随着海拔升高, 种群内植株个体呈现减小的趋势.植株大小对开花特性具有显著影响.一方面, 植株个体越大, 其总开花面积也越大, 但另一方面, 植株越大, 其总开花面积比越小(
表2 ), 并且这种趋势在不同性别植株间具有高度一致性(
图3 ), 说明资源限制是真实存在的, 即, 植物从环境中获取并分配给开花的资源量有限.此外, 该结果也说明植株分配到开花的资源增长速率可能低于植株个体的增长速率, 即繁殖分配的增加速率低于营养分配的增加速率.相应结果进一步暗示, 在面临资源限制或环境胁迫时, 团状福禄草的资源分配策略以保证植株存活(即增加营养分配)为首要目的.此外, 具有不同坡度及坡向的种群间开花特性也存在一定的变化,说明由坡向及坡度变化而引起的环境因子变化, 如太阳辐射(
田辉等, 2007 )和土壤养分(
曹舰艇等, 2019 ), 可能也同时对其繁殖分配策略具有一定的影响. ...
... 前期研究发现, 团状福禄草植物个体表面单位面积的果实及种子产量随海拔上升而呈现总体下降趋势(
Chen et al ., 2017a ).但是, 该研究并未考虑团状福禄草植物整体的繁殖分配对环境因子的响应, 而仅是比较了其特定开花区域内单位面积的繁殖产出.本研究结果显示, 团状福禄草在开花阶段的繁殖分配在不同海拔种群间存在显著差异, 总体上随海拔上升而减少, 表明高海拔更加贫瘠的土壤条件(
Chen et al ., 2015b ,
2019 )可能限制了植物总体的资源获取量, 植物需要将有限的资源在营养生长与繁殖生长两方面之间做权衡(
Reekie & Avila-Sakar, 2005 ), 从而导致其在繁殖功能上的总体分配比例的下降.但是, 我们也发现, 部分低海拔种群(种群2)内植物个体的繁殖分配显著低于部分高海拔种群(种群3、4), 这表明, 海拔并非控制植物繁殖分配的唯一因素, 植物繁殖分配是多种生态因子共同作用的结果(
张婵等, 2020 ).种群2与种群1海拔相差较小, 但坡度相差较大, 其与种群3坡度相近, 但具有不同坡向(
表1 ).因此, 我们推测除了海拔因素, 坡度与坡向可能同时对其繁殖分配产生影响.一方面坡度大可能导致土壤养分流失更严重(
Bruce-Okine & Lal, 1975 ;
Donker & Damen, 1984 ), 从而引起资源胁迫的加剧, 并进而导致繁殖分配的减少.另一方面, 坡度和坡向通常会引起太阳辐射特征的变化, 如太阳入射角、入射时间在不同坡度和坡向上会发生变化(
田辉等, 2007 ), 从而引起植物可积累和利用的有效太阳辐射能量的变化.在高海拔地区, 太阳辐射对植物的生长与繁殖策略都起着重要的影响.如, 为了充分利用太阳辐射, 一些植物花器官进化出了“向日性”功能(
Zhang et al ., 2010 ).又如, 安第斯山脉中部高海拔岩石斜坡典型垫状植物
Azorella compacta 为了最大化利用太阳辐射, 其个体偏爱生长在向北的(南半球)坡度约为20°的斜坡(
Kleier et al ., 2015 ).因此, 我们推测由坡度与坡向变化引起的土壤养分状况及太阳辐射特征的变化, 可能同时对团状福禄草的繁殖分配起着重要的影响. ...
... 如前所述, 太阳辐射特征可能对植物开花特性具有一定的调节功能.一方面表现在开花数量的多少, 另一方也可能表现在开花方位上.我们研究发现, 在同一种群内, 团状福禄草在冠层表面不同方位上的开花面积比存在显著差异, 但是, 这种差异在不同种群(即不同环境因子条件)之间又具有不同的表现形式.如, 最低海拔的种群1, 北面开花面积比最高, 东、南面次之且差异不显著, 而西面开花面积比最低; 最高海拔处的种群5, 其开花面积比呈现出东、北、南、西递减趋势, 且各方位上差异均显著(
图5 ).不同海拔及坡向上的种群, 可能由于坡度、坡向的差异而导致太阳辐射时间乃至辐射强度的变化(
田辉等, 2007 ).植物为了充分获取太阳辐射能量, 而尽可能将花开放在有利的方位上, 以满足果实及种子成熟过程中对太阳辐射能量的需求. ...
复杂地形下黑河流域的太阳辐射计算
3
2007
... 横断山高海拔地区绝大多数植物种群的更新与扩张都依赖有性繁殖(
Peng et al ., 2012 ).植物对繁殖功能的资源分配, 在很大程度上决定了其种群未来的扩张速率.而作为植物实现繁殖功能的重要环节, 植物对开花的资源配置决定了植物的最终繁殖产出(
Reekie & Bazzaz, 2005 ).不同的环境因子在植物的每一个繁殖阶段都起着重要作用, 甚至一些因子(如资源量)直接决定着植物的繁殖分配及其产出(
Delph & Carroll, 2001 ).在我们的研究区域内, 环境胁迫程度(如土壤养分胁迫及低温胁迫)随海拔升高而增强(
Chen et al ., 2015b ,
2019 ).我们的研究结果显示, 随着海拔升高, 种群内植株个体呈现减小的趋势.植株大小对开花特性具有显著影响.一方面, 植株个体越大, 其总开花面积也越大, 但另一方面, 植株越大, 其总开花面积比越小(
表2 ), 并且这种趋势在不同性别植株间具有高度一致性(
图3 ), 说明资源限制是真实存在的, 即, 植物从环境中获取并分配给开花的资源量有限.此外, 该结果也说明植株分配到开花的资源增长速率可能低于植株个体的增长速率, 即繁殖分配的增加速率低于营养分配的增加速率.相应结果进一步暗示, 在面临资源限制或环境胁迫时, 团状福禄草的资源分配策略以保证植株存活(即增加营养分配)为首要目的.此外, 具有不同坡度及坡向的种群间开花特性也存在一定的变化,说明由坡向及坡度变化而引起的环境因子变化, 如太阳辐射(
田辉等, 2007 )和土壤养分(
曹舰艇等, 2019 ), 可能也同时对其繁殖分配策略具有一定的影响. ...
... 前期研究发现, 团状福禄草植物个体表面单位面积的果实及种子产量随海拔上升而呈现总体下降趋势(
Chen et al ., 2017a ).但是, 该研究并未考虑团状福禄草植物整体的繁殖分配对环境因子的响应, 而仅是比较了其特定开花区域内单位面积的繁殖产出.本研究结果显示, 团状福禄草在开花阶段的繁殖分配在不同海拔种群间存在显著差异, 总体上随海拔上升而减少, 表明高海拔更加贫瘠的土壤条件(
Chen et al ., 2015b ,
2019 )可能限制了植物总体的资源获取量, 植物需要将有限的资源在营养生长与繁殖生长两方面之间做权衡(
Reekie & Avila-Sakar, 2005 ), 从而导致其在繁殖功能上的总体分配比例的下降.但是, 我们也发现, 部分低海拔种群(种群2)内植物个体的繁殖分配显著低于部分高海拔种群(种群3、4), 这表明, 海拔并非控制植物繁殖分配的唯一因素, 植物繁殖分配是多种生态因子共同作用的结果(
张婵等, 2020 ).种群2与种群1海拔相差较小, 但坡度相差较大, 其与种群3坡度相近, 但具有不同坡向(
表1 ).因此, 我们推测除了海拔因素, 坡度与坡向可能同时对其繁殖分配产生影响.一方面坡度大可能导致土壤养分流失更严重(
Bruce-Okine & Lal, 1975 ;
Donker & Damen, 1984 ), 从而引起资源胁迫的加剧, 并进而导致繁殖分配的减少.另一方面, 坡度和坡向通常会引起太阳辐射特征的变化, 如太阳入射角、入射时间在不同坡度和坡向上会发生变化(
田辉等, 2007 ), 从而引起植物可积累和利用的有效太阳辐射能量的变化.在高海拔地区, 太阳辐射对植物的生长与繁殖策略都起着重要的影响.如, 为了充分利用太阳辐射, 一些植物花器官进化出了“向日性”功能(
Zhang et al ., 2010 ).又如, 安第斯山脉中部高海拔岩石斜坡典型垫状植物
Azorella compacta 为了最大化利用太阳辐射, 其个体偏爱生长在向北的(南半球)坡度约为20°的斜坡(
Kleier et al ., 2015 ).因此, 我们推测由坡度与坡向变化引起的土壤养分状况及太阳辐射特征的变化, 可能同时对团状福禄草的繁殖分配起着重要的影响. ...
... 如前所述, 太阳辐射特征可能对植物开花特性具有一定的调节功能.一方面表现在开花数量的多少, 另一方也可能表现在开花方位上.我们研究发现, 在同一种群内, 团状福禄草在冠层表面不同方位上的开花面积比存在显著差异, 但是, 这种差异在不同种群(即不同环境因子条件)之间又具有不同的表现形式.如, 最低海拔的种群1, 北面开花面积比最高, 东、南面次之且差异不显著, 而西面开花面积比最低; 最高海拔处的种群5, 其开花面积比呈现出东、北、南、西递减趋势, 且各方位上差异均显著(
图5 ).不同海拔及坡向上的种群, 可能由于坡度、坡向的差异而导致太阳辐射时间乃至辐射强度的变化(
田辉等, 2007 ).植物为了充分获取太阳辐射能量, 而尽可能将花开放在有利的方位上, 以满足果实及种子成熟过程中对太阳辐射能量的需求. ...
Reproductive allometry in soybean, maize and sunflower
1
2000
... 温度、资源(包括土壤养分、降水资源及太阳辐射量等)是限制高山植物生长与繁殖的关键因素(
Bloom et al ., 1985 ;
Chen et al ., 2017b ).为了应对高海拔地区贫瘠的土壤条件, 植物在自身资源获取及分配方面进化出了众多与环境相适应的资源分配策略.其中, 繁殖分配(即植物分配到繁殖器官的资源量)是植物适合度的关键表现(
Bazzaz et al ., 1987 ;
Weiner, 1988 ;
Hartnett, 1990 ;
Vega et al ., 2000 ).研究表明, 植物繁殖分配同时受到众多非生物与生物因子的控制(
Reekie & Bazzaz, 2005 ;
张婵等, 2020 ).非生物因子方面, 温度、水分、土壤营养等对植物的繁殖分配起着重要作用(
Reekie & Bazzaz, 2005 ;
Zhang et al ., 2014 ).而在生物因子方面, 种间竞争、传粉者特性以及植物自身性系统特征(
Ashman, 1994 ;
Case & Ashman, 2005 ;
Brzyski et al ., 2014 ;
Chen et al ., 2017a ,
2017b )对其繁殖分配策略都起着重要的影响.例如, 具有雌全异株性系统特征(即同一种植物同时存在雌性个体和两性个体)的植物, 其同一种群内的雌性个体与两性个体在繁殖分配策略上存在较大差异, 表现为雌性个体主要将资源分配到雌性功能(如胚珠及种子生产), 而两性个体则主要将资源分配到雄性功能(如花粉生产以及传粉者吸引)(
Ashman, 1994 ;
Chen et al ., 2017a ,
2017b ), 并且这种分配模式受到环境因子的强烈影响(
Chen et al ., 2017b ).为了应对资源的不足, 植物往往需要在营养分配(即分配到营养器官的资源量)与繁殖分配之间做出权衡, 以达到最优的繁殖适合度(
Reekie & Avila-Sakar, 2005 ).开花是植物繁殖的最初阶段, 植物开花的数目, 直接反映出植物在特定环境条件下的繁殖分配策略.另一方面, 开花数目的多少也可直接决定着植物未来的繁殖输出量(如果实及种子产量;
Reekie & Bazzaz, 2005 ). ...
The influence of competition on plant reproduction//Lovett Doust J, Lovett Doust L
1
1988
... 温度、资源(包括土壤养分、降水资源及太阳辐射量等)是限制高山植物生长与繁殖的关键因素(
Bloom et al ., 1985 ;
Chen et al ., 2017b ).为了应对高海拔地区贫瘠的土壤条件, 植物在自身资源获取及分配方面进化出了众多与环境相适应的资源分配策略.其中, 繁殖分配(即植物分配到繁殖器官的资源量)是植物适合度的关键表现(
Bazzaz et al ., 1987 ;
Weiner, 1988 ;
Hartnett, 1990 ;
Vega et al ., 2000 ).研究表明, 植物繁殖分配同时受到众多非生物与生物因子的控制(
Reekie & Bazzaz, 2005 ;
张婵等, 2020 ).非生物因子方面, 温度、水分、土壤营养等对植物的繁殖分配起着重要作用(
Reekie & Bazzaz, 2005 ;
Zhang et al ., 2014 ).而在生物因子方面, 种间竞争、传粉者特性以及植物自身性系统特征(
Ashman, 1994 ;
Case & Ashman, 2005 ;
Brzyski et al ., 2014 ;
Chen et al ., 2017a ,
2017b )对其繁殖分配策略都起着重要的影响.例如, 具有雌全异株性系统特征(即同一种植物同时存在雌性个体和两性个体)的植物, 其同一种群内的雌性个体与两性个体在繁殖分配策略上存在较大差异, 表现为雌性个体主要将资源分配到雌性功能(如胚珠及种子生产), 而两性个体则主要将资源分配到雄性功能(如花粉生产以及传粉者吸引)(
Ashman, 1994 ;
Chen et al ., 2017a ,
2017b ), 并且这种分配模式受到环境因子的强烈影响(
Chen et al ., 2017b ).为了应对资源的不足, 植物往往需要在营养分配(即分配到营养器官的资源量)与繁殖分配之间做出权衡, 以达到最优的繁殖适合度(
Reekie & Avila-Sakar, 2005 ).开花是植物繁殖的最初阶段, 植物开花的数目, 直接反映出植物在特定环境条件下的繁殖分配策略.另一方面, 开花数目的多少也可直接决定着植物未来的繁殖输出量(如果实及种子产量;
Reekie & Bazzaz, 2005 ). ...
1
2013
... 团状福禄草(
Arenaria polytrichoides )为石竹科无心菜属(
Arenaria )多年生垫状草本植物, 是青藏高原特有种, 沿喜马拉雅—横断山高海拔(3 500-5 300 m)地区分布(
Wu et al ., 2013 ).营养生长自冰雪消融开始, 花期主要在6-7月, 不同分布区域及海拔的种群的花期存在一定的差异(
Chen et al ., 2017a ).并且, 团状福禄草具有雌全异株的性二态系统, 即同一种群中同时存在雌性植株和两性植株, 不同性别的个体在繁殖产出(果实及种子产量)方面存在显著差异, 并且受到海拔的影响(
Chen et al ., 2017a ).表现为较高海拔的雌性比例高于较低海拔的, 两性植株开花数目大于雌性植株但结实率显著低于雌性植株, 同时, 较低海拔种群内个体单位面积的果实与种子产量高于较高海拔种群的个体(
Chen et al ., 2017a ).此外, 团状福禄草被证实为横断山高山生态系统当中的关键种, 能有效改良微环境条件(温度、土壤营养及水分), 从而为其他低耐受物种提供适宜的微生境, 进而调节种群动态、提升生态系统物种多样性及地面生产力(
Yang et al ., 2010 ,
2017 ;
Chen et al ., 2015a ,
2019 ).然而, 团状福禄草种群的更新却受到种间竞争的强烈影响(
Chen et al ., 2020 ).研究者在近十年内对团状福禄草做了深入调查与研究(
Yang et al ., 2010 ,
2017 ;
Chen et al ., 2015a ,
2015b ,
2017a ,
2019 ), 但对其种群的繁殖与更新机制, 却直到最近才予以关注(
Chen et al ., 2017a ,
2020 ).理论上讲, 团状福禄草的每一个小分枝(
图1C )在生长季节都能开出一朵花, 然而, 我们在多年的野外研究中发现, 实际上, 其每年的开花数量极为有限, 只在其冠层表面的部分区域开出一定面积的花(
图1A 、1B).这一现象说明, 垫状植物的开花可能受到了资源的限制, 并有可能同时受到其他环境因子的影响.例如, 据野外观察, 团状福禄草开花方位(只在其冠层表面特定区域开放,
图1A 、1B)呈现出一定的规律性, 且在不同的种群之间存在差异.为了获取充足的太阳辐射能量从而满足自身繁殖需求, 一些高山植物的开花特性受到太阳辐射的强烈影响(
Pati?o et al ., 2002 ;
Galen, 2006 ;
Zhang et al ., 2010 ), 因此, 我们猜测团状福禄草的开花数量和方位可能受到资源和太阳辐射的双重影响.但其繁殖分配策略, 即个体对开花的资源分配策略对环境胁迫因子的响应方式, 目前却不为人知.深入探究高山生态系统中这一关键类群的种群更新与维持机制, 对揭示其种群繁殖的初始阶段(开花特性)对环境因子的响应机制以及这种响应机制在不同性别个体之间的差异性具有重要意义. ...
Size-mediated interaction between a cushion species and other non- cushion species at high elevations of the Hengduan Mountains, SW China
2
2017
... 团状福禄草(
Arenaria polytrichoides )为石竹科无心菜属(
Arenaria )多年生垫状草本植物, 是青藏高原特有种, 沿喜马拉雅—横断山高海拔(3 500-5 300 m)地区分布(
Wu et al ., 2013 ).营养生长自冰雪消融开始, 花期主要在6-7月, 不同分布区域及海拔的种群的花期存在一定的差异(
Chen et al ., 2017a ).并且, 团状福禄草具有雌全异株的性二态系统, 即同一种群中同时存在雌性植株和两性植株, 不同性别的个体在繁殖产出(果实及种子产量)方面存在显著差异, 并且受到海拔的影响(
Chen et al ., 2017a ).表现为较高海拔的雌性比例高于较低海拔的, 两性植株开花数目大于雌性植株但结实率显著低于雌性植株, 同时, 较低海拔种群内个体单位面积的果实与种子产量高于较高海拔种群的个体(
Chen et al ., 2017a ).此外, 团状福禄草被证实为横断山高山生态系统当中的关键种, 能有效改良微环境条件(温度、土壤营养及水分), 从而为其他低耐受物种提供适宜的微生境, 进而调节种群动态、提升生态系统物种多样性及地面生产力(
Yang et al ., 2010 ,
2017 ;
Chen et al ., 2015a ,
2019 ).然而, 团状福禄草种群的更新却受到种间竞争的强烈影响(
Chen et al ., 2020 ).研究者在近十年内对团状福禄草做了深入调查与研究(
Yang et al ., 2010 ,
2017 ;
Chen et al ., 2015a ,
2015b ,
2017a ,
2019 ), 但对其种群的繁殖与更新机制, 却直到最近才予以关注(
Chen et al ., 2017a ,
2020 ).理论上讲, 团状福禄草的每一个小分枝(
图1C )在生长季节都能开出一朵花, 然而, 我们在多年的野外研究中发现, 实际上, 其每年的开花数量极为有限, 只在其冠层表面的部分区域开出一定面积的花(
图1A 、1B).这一现象说明, 垫状植物的开花可能受到了资源的限制, 并有可能同时受到其他环境因子的影响.例如, 据野外观察, 团状福禄草开花方位(只在其冠层表面特定区域开放,
图1A 、1B)呈现出一定的规律性, 且在不同的种群之间存在差异.为了获取充足的太阳辐射能量从而满足自身繁殖需求, 一些高山植物的开花特性受到太阳辐射的强烈影响(
Pati?o et al ., 2002 ;
Galen, 2006 ;
Zhang et al ., 2010 ), 因此, 我们猜测团状福禄草的开花数量和方位可能受到资源和太阳辐射的双重影响.但其繁殖分配策略, 即个体对开花的资源分配策略对环境胁迫因子的响应方式, 目前却不为人知.深入探究高山生态系统中这一关键类群的种群更新与维持机制, 对揭示其种群繁殖的初始阶段(开花特性)对环境因子的响应机制以及这种响应机制在不同性别个体之间的差异性具有重要意义. ...
... ,
2017 ;
Chen et al ., 2015a ,
2015b ,
2017a ,
2019 ), 但对其种群的繁殖与更新机制, 却直到最近才予以关注(
Chen et al ., 2017a ,
2020 ).理论上讲, 团状福禄草的每一个小分枝(
图1C )在生长季节都能开出一朵花, 然而, 我们在多年的野外研究中发现, 实际上, 其每年的开花数量极为有限, 只在其冠层表面的部分区域开出一定面积的花(
图1A 、1B).这一现象说明, 垫状植物的开花可能受到了资源的限制, 并有可能同时受到其他环境因子的影响.例如, 据野外观察, 团状福禄草开花方位(只在其冠层表面特定区域开放,
图1A 、1B)呈现出一定的规律性, 且在不同的种群之间存在差异.为了获取充足的太阳辐射能量从而满足自身繁殖需求, 一些高山植物的开花特性受到太阳辐射的强烈影响(
Pati?o et al ., 2002 ;
Galen, 2006 ;
Zhang et al ., 2010 ), 因此, 我们猜测团状福禄草的开花数量和方位可能受到资源和太阳辐射的双重影响.但其繁殖分配策略, 即个体对开花的资源分配策略对环境胁迫因子的响应方式, 目前却不为人知.深入探究高山生态系统中这一关键类群的种群更新与维持机制, 对揭示其种群繁殖的初始阶段(开花特性)对环境因子的响应机制以及这种响应机制在不同性别个体之间的差异性具有重要意义. ...
青藏高原冰缘植物多样性适应机制研究进展
2
2019
... 随着海拔逐渐上升, 环境条件变得更加严酷, 如: 海拔每上升100 m, 气温约下降0.6 ℃, 气压下降8-9 hPa (
K?rner, 2003 ), 土壤养分含量也随之减少(
曹舰艇等, 2019 ).面对高海拔地区低温、土壤养分贫瘠等恶劣环境因子, 高山植物在长期的进化过程中形成了丰富多样的特化适应性状或进化策略(
Sun et al ., 2014 ;
杨扬等, 2019 ), 种类丰富的植物多样性对维持高山生态系统的稳定性起到了关键作用(
Hooper et al ., 2005 ).滇西北处于横断山高山植物多样性中心区域(
郭辉军和龙春林, 1998 ;
Zhang et al. , 2015 ).长期以来, 青藏高原及横断山高海拔地区植物对极端环境因子的适应机制, 备受植物学家和生态学家的关注, 相关研究的深入开展, 加深了对该地区植物多样性形成及其进化适应机制的认识(
Sun et al ., 2014 ;
杨扬等, 2019 ). ...
... ;
杨扬等, 2019 ). ...
青藏高原冰缘植物多样性适应机制研究进展
2
2019
... 随着海拔逐渐上升, 环境条件变得更加严酷, 如: 海拔每上升100 m, 气温约下降0.6 ℃, 气压下降8-9 hPa (
K?rner, 2003 ), 土壤养分含量也随之减少(
曹舰艇等, 2019 ).面对高海拔地区低温、土壤养分贫瘠等恶劣环境因子, 高山植物在长期的进化过程中形成了丰富多样的特化适应性状或进化策略(
Sun et al ., 2014 ;
杨扬等, 2019 ), 种类丰富的植物多样性对维持高山生态系统的稳定性起到了关键作用(
Hooper et al ., 2005 ).滇西北处于横断山高山植物多样性中心区域(
郭辉军和龙春林, 1998 ;
Zhang et al. , 2015 ).长期以来, 青藏高原及横断山高海拔地区植物对极端环境因子的适应机制, 备受植物学家和生态学家的关注, 相关研究的深入开展, 加深了对该地区植物多样性形成及其进化适应机制的认识(
Sun et al ., 2014 ;
杨扬等, 2019 ). ...
... ;
杨扬等, 2019 ). ...
Positive associations between the cushion plant
Arenaria polytrichoides (Caryophyllaceae) and other alpine plant species increase with altitude in the Sino-Himalayas
2
2010
... 团状福禄草(
Arenaria polytrichoides )为石竹科无心菜属(
Arenaria )多年生垫状草本植物, 是青藏高原特有种, 沿喜马拉雅—横断山高海拔(3 500-5 300 m)地区分布(
Wu et al ., 2013 ).营养生长自冰雪消融开始, 花期主要在6-7月, 不同分布区域及海拔的种群的花期存在一定的差异(
Chen et al ., 2017a ).并且, 团状福禄草具有雌全异株的性二态系统, 即同一种群中同时存在雌性植株和两性植株, 不同性别的个体在繁殖产出(果实及种子产量)方面存在显著差异, 并且受到海拔的影响(
Chen et al ., 2017a ).表现为较高海拔的雌性比例高于较低海拔的, 两性植株开花数目大于雌性植株但结实率显著低于雌性植株, 同时, 较低海拔种群内个体单位面积的果实与种子产量高于较高海拔种群的个体(
Chen et al ., 2017a ).此外, 团状福禄草被证实为横断山高山生态系统当中的关键种, 能有效改良微环境条件(温度、土壤营养及水分), 从而为其他低耐受物种提供适宜的微生境, 进而调节种群动态、提升生态系统物种多样性及地面生产力(
Yang et al ., 2010 ,
2017 ;
Chen et al ., 2015a ,
2019 ).然而, 团状福禄草种群的更新却受到种间竞争的强烈影响(
Chen et al ., 2020 ).研究者在近十年内对团状福禄草做了深入调查与研究(
Yang et al ., 2010 ,
2017 ;
Chen et al ., 2015a ,
2015b ,
2017a ,
2019 ), 但对其种群的繁殖与更新机制, 却直到最近才予以关注(
Chen et al ., 2017a ,
2020 ).理论上讲, 团状福禄草的每一个小分枝(
图1C )在生长季节都能开出一朵花, 然而, 我们在多年的野外研究中发现, 实际上, 其每年的开花数量极为有限, 只在其冠层表面的部分区域开出一定面积的花(
图1A 、1B).这一现象说明, 垫状植物的开花可能受到了资源的限制, 并有可能同时受到其他环境因子的影响.例如, 据野外观察, 团状福禄草开花方位(只在其冠层表面特定区域开放,
图1A 、1B)呈现出一定的规律性, 且在不同的种群之间存在差异.为了获取充足的太阳辐射能量从而满足自身繁殖需求, 一些高山植物的开花特性受到太阳辐射的强烈影响(
Pati?o et al ., 2002 ;
Galen, 2006 ;
Zhang et al ., 2010 ), 因此, 我们猜测团状福禄草的开花数量和方位可能受到资源和太阳辐射的双重影响.但其繁殖分配策略, 即个体对开花的资源分配策略对环境胁迫因子的响应方式, 目前却不为人知.深入探究高山生态系统中这一关键类群的种群更新与维持机制, 对揭示其种群繁殖的初始阶段(开花特性)对环境因子的响应机制以及这种响应机制在不同性别个体之间的差异性具有重要意义. ...
... ).研究者在近十年内对团状福禄草做了深入调查与研究(
Yang et al ., 2010 ,
2017 ;
Chen et al ., 2015a ,
2015b ,
2017a ,
2019 ), 但对其种群的繁殖与更新机制, 却直到最近才予以关注(
Chen et al ., 2017a ,
2020 ).理论上讲, 团状福禄草的每一个小分枝(
图1C )在生长季节都能开出一朵花, 然而, 我们在多年的野外研究中发现, 实际上, 其每年的开花数量极为有限, 只在其冠层表面的部分区域开出一定面积的花(
图1A 、1B).这一现象说明, 垫状植物的开花可能受到了资源的限制, 并有可能同时受到其他环境因子的影响.例如, 据野外观察, 团状福禄草开花方位(只在其冠层表面特定区域开放,
图1A 、1B)呈现出一定的规律性, 且在不同的种群之间存在差异.为了获取充足的太阳辐射能量从而满足自身繁殖需求, 一些高山植物的开花特性受到太阳辐射的强烈影响(
Pati?o et al ., 2002 ;
Galen, 2006 ;
Zhang et al ., 2010 ), 因此, 我们猜测团状福禄草的开花数量和方位可能受到资源和太阳辐射的双重影响.但其繁殖分配策略, 即个体对开花的资源分配策略对环境胁迫因子的响应方式, 目前却不为人知.深入探究高山生态系统中这一关键类群的种群更新与维持机制, 对揭示其种群繁殖的初始阶段(开花特性)对环境因子的响应机制以及这种响应机制在不同性别个体之间的差异性具有重要意义. ...
青藏高原及周边高山地区的植物繁殖生态学研究进展
2
2020
... 温度、资源(包括土壤养分、降水资源及太阳辐射量等)是限制高山植物生长与繁殖的关键因素(
Bloom et al ., 1985 ;
Chen et al ., 2017b ).为了应对高海拔地区贫瘠的土壤条件, 植物在自身资源获取及分配方面进化出了众多与环境相适应的资源分配策略.其中, 繁殖分配(即植物分配到繁殖器官的资源量)是植物适合度的关键表现(
Bazzaz et al ., 1987 ;
Weiner, 1988 ;
Hartnett, 1990 ;
Vega et al ., 2000 ).研究表明, 植物繁殖分配同时受到众多非生物与生物因子的控制(
Reekie & Bazzaz, 2005 ;
张婵等, 2020 ).非生物因子方面, 温度、水分、土壤营养等对植物的繁殖分配起着重要作用(
Reekie & Bazzaz, 2005 ;
Zhang et al ., 2014 ).而在生物因子方面, 种间竞争、传粉者特性以及植物自身性系统特征(
Ashman, 1994 ;
Case & Ashman, 2005 ;
Brzyski et al ., 2014 ;
Chen et al ., 2017a ,
2017b )对其繁殖分配策略都起着重要的影响.例如, 具有雌全异株性系统特征(即同一种植物同时存在雌性个体和两性个体)的植物, 其同一种群内的雌性个体与两性个体在繁殖分配策略上存在较大差异, 表现为雌性个体主要将资源分配到雌性功能(如胚珠及种子生产), 而两性个体则主要将资源分配到雄性功能(如花粉生产以及传粉者吸引)(
Ashman, 1994 ;
Chen et al ., 2017a ,
2017b ), 并且这种分配模式受到环境因子的强烈影响(
Chen et al ., 2017b ).为了应对资源的不足, 植物往往需要在营养分配(即分配到营养器官的资源量)与繁殖分配之间做出权衡, 以达到最优的繁殖适合度(
Reekie & Avila-Sakar, 2005 ).开花是植物繁殖的最初阶段, 植物开花的数目, 直接反映出植物在特定环境条件下的繁殖分配策略.另一方面, 开花数目的多少也可直接决定着植物未来的繁殖输出量(如果实及种子产量;
Reekie & Bazzaz, 2005 ). ...
... 前期研究发现, 团状福禄草植物个体表面单位面积的果实及种子产量随海拔上升而呈现总体下降趋势(
Chen et al ., 2017a ).但是, 该研究并未考虑团状福禄草植物整体的繁殖分配对环境因子的响应, 而仅是比较了其特定开花区域内单位面积的繁殖产出.本研究结果显示, 团状福禄草在开花阶段的繁殖分配在不同海拔种群间存在显著差异, 总体上随海拔上升而减少, 表明高海拔更加贫瘠的土壤条件(
Chen et al ., 2015b ,
2019 )可能限制了植物总体的资源获取量, 植物需要将有限的资源在营养生长与繁殖生长两方面之间做权衡(
Reekie & Avila-Sakar, 2005 ), 从而导致其在繁殖功能上的总体分配比例的下降.但是, 我们也发现, 部分低海拔种群(种群2)内植物个体的繁殖分配显著低于部分高海拔种群(种群3、4), 这表明, 海拔并非控制植物繁殖分配的唯一因素, 植物繁殖分配是多种生态因子共同作用的结果(
张婵等, 2020 ).种群2与种群1海拔相差较小, 但坡度相差较大, 其与种群3坡度相近, 但具有不同坡向(
表1 ).因此, 我们推测除了海拔因素, 坡度与坡向可能同时对其繁殖分配产生影响.一方面坡度大可能导致土壤养分流失更严重(
Bruce-Okine & Lal, 1975 ;
Donker & Damen, 1984 ), 从而引起资源胁迫的加剧, 并进而导致繁殖分配的减少.另一方面, 坡度和坡向通常会引起太阳辐射特征的变化, 如太阳入射角、入射时间在不同坡度和坡向上会发生变化(
田辉等, 2007 ), 从而引起植物可积累和利用的有效太阳辐射能量的变化.在高海拔地区, 太阳辐射对植物的生长与繁殖策略都起着重要的影响.如, 为了充分利用太阳辐射, 一些植物花器官进化出了“向日性”功能(
Zhang et al ., 2010 ).又如, 安第斯山脉中部高海拔岩石斜坡典型垫状植物
Azorella compacta 为了最大化利用太阳辐射, 其个体偏爱生长在向北的(南半球)坡度约为20°的斜坡(
Kleier et al ., 2015 ).因此, 我们推测由坡度与坡向变化引起的土壤养分状况及太阳辐射特征的变化, 可能同时对团状福禄草的繁殖分配起着重要的影响. ...
青藏高原及周边高山地区的植物繁殖生态学研究进展
2
2020
... 温度、资源(包括土壤养分、降水资源及太阳辐射量等)是限制高山植物生长与繁殖的关键因素(
Bloom et al ., 1985 ;
Chen et al ., 2017b ).为了应对高海拔地区贫瘠的土壤条件, 植物在自身资源获取及分配方面进化出了众多与环境相适应的资源分配策略.其中, 繁殖分配(即植物分配到繁殖器官的资源量)是植物适合度的关键表现(
Bazzaz et al ., 1987 ;
Weiner, 1988 ;
Hartnett, 1990 ;
Vega et al ., 2000 ).研究表明, 植物繁殖分配同时受到众多非生物与生物因子的控制(
Reekie & Bazzaz, 2005 ;
张婵等, 2020 ).非生物因子方面, 温度、水分、土壤营养等对植物的繁殖分配起着重要作用(
Reekie & Bazzaz, 2005 ;
Zhang et al ., 2014 ).而在生物因子方面, 种间竞争、传粉者特性以及植物自身性系统特征(
Ashman, 1994 ;
Case & Ashman, 2005 ;
Brzyski et al ., 2014 ;
Chen et al ., 2017a ,
2017b )对其繁殖分配策略都起着重要的影响.例如, 具有雌全异株性系统特征(即同一种植物同时存在雌性个体和两性个体)的植物, 其同一种群内的雌性个体与两性个体在繁殖分配策略上存在较大差异, 表现为雌性个体主要将资源分配到雌性功能(如胚珠及种子生产), 而两性个体则主要将资源分配到雄性功能(如花粉生产以及传粉者吸引)(
Ashman, 1994 ;
Chen et al ., 2017a ,
2017b ), 并且这种分配模式受到环境因子的强烈影响(
Chen et al ., 2017b ).为了应对资源的不足, 植物往往需要在营养分配(即分配到营养器官的资源量)与繁殖分配之间做出权衡, 以达到最优的繁殖适合度(
Reekie & Avila-Sakar, 2005 ).开花是植物繁殖的最初阶段, 植物开花的数目, 直接反映出植物在特定环境条件下的繁殖分配策略.另一方面, 开花数目的多少也可直接决定着植物未来的繁殖输出量(如果实及种子产量;
Reekie & Bazzaz, 2005 ). ...
... 前期研究发现, 团状福禄草植物个体表面单位面积的果实及种子产量随海拔上升而呈现总体下降趋势(
Chen et al ., 2017a ).但是, 该研究并未考虑团状福禄草植物整体的繁殖分配对环境因子的响应, 而仅是比较了其特定开花区域内单位面积的繁殖产出.本研究结果显示, 团状福禄草在开花阶段的繁殖分配在不同海拔种群间存在显著差异, 总体上随海拔上升而减少, 表明高海拔更加贫瘠的土壤条件(
Chen et al ., 2015b ,
2019 )可能限制了植物总体的资源获取量, 植物需要将有限的资源在营养生长与繁殖生长两方面之间做权衡(
Reekie & Avila-Sakar, 2005 ), 从而导致其在繁殖功能上的总体分配比例的下降.但是, 我们也发现, 部分低海拔种群(种群2)内植物个体的繁殖分配显著低于部分高海拔种群(种群3、4), 这表明, 海拔并非控制植物繁殖分配的唯一因素, 植物繁殖分配是多种生态因子共同作用的结果(
张婵等, 2020 ).种群2与种群1海拔相差较小, 但坡度相差较大, 其与种群3坡度相近, 但具有不同坡向(
表1 ).因此, 我们推测除了海拔因素, 坡度与坡向可能同时对其繁殖分配产生影响.一方面坡度大可能导致土壤养分流失更严重(
Bruce-Okine & Lal, 1975 ;
Donker & Damen, 1984 ), 从而引起资源胁迫的加剧, 并进而导致繁殖分配的减少.另一方面, 坡度和坡向通常会引起太阳辐射特征的变化, 如太阳入射角、入射时间在不同坡度和坡向上会发生变化(
田辉等, 2007 ), 从而引起植物可积累和利用的有效太阳辐射能量的变化.在高海拔地区, 太阳辐射对植物的生长与繁殖策略都起着重要的影响.如, 为了充分利用太阳辐射, 一些植物花器官进化出了“向日性”功能(
Zhang et al ., 2010 ).又如, 安第斯山脉中部高海拔岩石斜坡典型垫状植物
Azorella compacta 为了最大化利用太阳辐射, 其个体偏爱生长在向北的(南半球)坡度约为20°的斜坡(
Kleier et al ., 2015 ).因此, 我们推测由坡度与坡向变化引起的土壤养分状况及太阳辐射特征的变化, 可能同时对团状福禄草的繁殖分配起着重要的影响. ...
Effects of different nutrient sources on plasticity of reproductive strategies in a monoecious species,
Sagittaria graminea (Alismataceae)
1
2014
... 温度、资源(包括土壤养分、降水资源及太阳辐射量等)是限制高山植物生长与繁殖的关键因素(
Bloom et al ., 1985 ;
Chen et al ., 2017b ).为了应对高海拔地区贫瘠的土壤条件, 植物在自身资源获取及分配方面进化出了众多与环境相适应的资源分配策略.其中, 繁殖分配(即植物分配到繁殖器官的资源量)是植物适合度的关键表现(
Bazzaz et al ., 1987 ;
Weiner, 1988 ;
Hartnett, 1990 ;
Vega et al ., 2000 ).研究表明, 植物繁殖分配同时受到众多非生物与生物因子的控制(
Reekie & Bazzaz, 2005 ;
张婵等, 2020 ).非生物因子方面, 温度、水分、土壤营养等对植物的繁殖分配起着重要作用(
Reekie & Bazzaz, 2005 ;
Zhang et al ., 2014 ).而在生物因子方面, 种间竞争、传粉者特性以及植物自身性系统特征(
Ashman, 1994 ;
Case & Ashman, 2005 ;
Brzyski et al ., 2014 ;
Chen et al ., 2017a ,
2017b )对其繁殖分配策略都起着重要的影响.例如, 具有雌全异株性系统特征(即同一种植物同时存在雌性个体和两性个体)的植物, 其同一种群内的雌性个体与两性个体在繁殖分配策略上存在较大差异, 表现为雌性个体主要将资源分配到雌性功能(如胚珠及种子生产), 而两性个体则主要将资源分配到雄性功能(如花粉生产以及传粉者吸引)(
Ashman, 1994 ;
Chen et al ., 2017a ,
2017b ), 并且这种分配模式受到环境因子的强烈影响(
Chen et al ., 2017b ).为了应对资源的不足, 植物往往需要在营养分配(即分配到营养器官的资源量)与繁殖分配之间做出权衡, 以达到最优的繁殖适合度(
Reekie & Avila-Sakar, 2005 ).开花是植物繁殖的最初阶段, 植物开花的数目, 直接反映出植物在特定环境条件下的繁殖分配策略.另一方面, 开花数目的多少也可直接决定着植物未来的繁殖输出量(如果实及种子产量;
Reekie & Bazzaz, 2005 ). ...
1
1997
... 研究区域位于滇西北德钦县白马雪山, 地理坐标为28.48° N, 98.99° E, 海拔范围为4 360?4 920 m, 生境类型从低至高依次为灌草甸、草甸-流石滩交错带和流石滩(
图1D ).滇西北处于横断山高山植物多样性中心区域(
郭辉军和龙春林, 1998 ;
Zhang et al ., 2015 ), 具有典型的季节性夏季季风气候, 以冷雨或雨夹雪为特征, 并伴有短时间的强烈太阳辐射, 年降水量680-790 mm, 年平均气温-1 ℃ (
张荣祖等, 1997 ).植物生长季节通常开始于5月中旬的融雪期, 结束于9月中下旬的夏季风停息期.我们前期的测量数据显示, 2015年6月中旬至9月下旬, 白马雪山海拔4 900 m处, 距离地面15 cm处的平均气温和湿度分别为(4.87 ± 0.04) ℃ 和(89.9 ± 0.24)%, 绝对最高气温和最低气温分别为17.80和-1.95 ℃, 昼夜温差大于10 ℃, 最高达16 ℃ (陈建国等, 未发表数据).此外, 土壤养分含量随海拔升高而普遍降低(
Chen et al ., 2015b ,
2019 ). ...
1
1997
... 研究区域位于滇西北德钦县白马雪山, 地理坐标为28.48° N, 98.99° E, 海拔范围为4 360?4 920 m, 生境类型从低至高依次为灌草甸、草甸-流石滩交错带和流石滩(
图1D ).滇西北处于横断山高山植物多样性中心区域(
郭辉军和龙春林, 1998 ;
Zhang et al ., 2015 ), 具有典型的季节性夏季季风气候, 以冷雨或雨夹雪为特征, 并伴有短时间的强烈太阳辐射, 年降水量680-790 mm, 年平均气温-1 ℃ (
张荣祖等, 1997 ).植物生长季节通常开始于5月中旬的融雪期, 结束于9月中下旬的夏季风停息期.我们前期的测量数据显示, 2015年6月中旬至9月下旬, 白马雪山海拔4 900 m处, 距离地面15 cm处的平均气温和湿度分别为(4.87 ± 0.04) ℃ 和(89.9 ± 0.24)%, 绝对最高气温和最低气温分别为17.80和-1.95 ℃, 昼夜温差大于10 ℃, 最高达16 ℃ (陈建国等, 未发表数据).此外, 土壤养分含量随海拔升高而普遍降低(
Chen et al ., 2015b ,
2019 ). ...
Flower heliotropism of
Anemone rivularis (Ranunculaceae) in the Himalayas: effects on floral temperature and reproductive fitness
2
2010
... 团状福禄草(
Arenaria polytrichoides )为石竹科无心菜属(
Arenaria )多年生垫状草本植物, 是青藏高原特有种, 沿喜马拉雅—横断山高海拔(3 500-5 300 m)地区分布(
Wu et al ., 2013 ).营养生长自冰雪消融开始, 花期主要在6-7月, 不同分布区域及海拔的种群的花期存在一定的差异(
Chen et al ., 2017a ).并且, 团状福禄草具有雌全异株的性二态系统, 即同一种群中同时存在雌性植株和两性植株, 不同性别的个体在繁殖产出(果实及种子产量)方面存在显著差异, 并且受到海拔的影响(
Chen et al ., 2017a ).表现为较高海拔的雌性比例高于较低海拔的, 两性植株开花数目大于雌性植株但结实率显著低于雌性植株, 同时, 较低海拔种群内个体单位面积的果实与种子产量高于较高海拔种群的个体(
Chen et al ., 2017a ).此外, 团状福禄草被证实为横断山高山生态系统当中的关键种, 能有效改良微环境条件(温度、土壤营养及水分), 从而为其他低耐受物种提供适宜的微生境, 进而调节种群动态、提升生态系统物种多样性及地面生产力(
Yang et al ., 2010 ,
2017 ;
Chen et al ., 2015a ,
2019 ).然而, 团状福禄草种群的更新却受到种间竞争的强烈影响(
Chen et al ., 2020 ).研究者在近十年内对团状福禄草做了深入调查与研究(
Yang et al ., 2010 ,
2017 ;
Chen et al ., 2015a ,
2015b ,
2017a ,
2019 ), 但对其种群的繁殖与更新机制, 却直到最近才予以关注(
Chen et al ., 2017a ,
2020 ).理论上讲, 团状福禄草的每一个小分枝(
图1C )在生长季节都能开出一朵花, 然而, 我们在多年的野外研究中发现, 实际上, 其每年的开花数量极为有限, 只在其冠层表面的部分区域开出一定面积的花(
图1A 、1B).这一现象说明, 垫状植物的开花可能受到了资源的限制, 并有可能同时受到其他环境因子的影响.例如, 据野外观察, 团状福禄草开花方位(只在其冠层表面特定区域开放,
图1A 、1B)呈现出一定的规律性, 且在不同的种群之间存在差异.为了获取充足的太阳辐射能量从而满足自身繁殖需求, 一些高山植物的开花特性受到太阳辐射的强烈影响(
Pati?o et al ., 2002 ;
Galen, 2006 ;
Zhang et al ., 2010 ), 因此, 我们猜测团状福禄草的开花数量和方位可能受到资源和太阳辐射的双重影响.但其繁殖分配策略, 即个体对开花的资源分配策略对环境胁迫因子的响应方式, 目前却不为人知.深入探究高山生态系统中这一关键类群的种群更新与维持机制, 对揭示其种群繁殖的初始阶段(开花特性)对环境因子的响应机制以及这种响应机制在不同性别个体之间的差异性具有重要意义. ...
... 前期研究发现, 团状福禄草植物个体表面单位面积的果实及种子产量随海拔上升而呈现总体下降趋势(
Chen et al ., 2017a ).但是, 该研究并未考虑团状福禄草植物整体的繁殖分配对环境因子的响应, 而仅是比较了其特定开花区域内单位面积的繁殖产出.本研究结果显示, 团状福禄草在开花阶段的繁殖分配在不同海拔种群间存在显著差异, 总体上随海拔上升而减少, 表明高海拔更加贫瘠的土壤条件(
Chen et al ., 2015b ,
2019 )可能限制了植物总体的资源获取量, 植物需要将有限的资源在营养生长与繁殖生长两方面之间做权衡(
Reekie & Avila-Sakar, 2005 ), 从而导致其在繁殖功能上的总体分配比例的下降.但是, 我们也发现, 部分低海拔种群(种群2)内植物个体的繁殖分配显著低于部分高海拔种群(种群3、4), 这表明, 海拔并非控制植物繁殖分配的唯一因素, 植物繁殖分配是多种生态因子共同作用的结果(
张婵等, 2020 ).种群2与种群1海拔相差较小, 但坡度相差较大, 其与种群3坡度相近, 但具有不同坡向(
表1 ).因此, 我们推测除了海拔因素, 坡度与坡向可能同时对其繁殖分配产生影响.一方面坡度大可能导致土壤养分流失更严重(
Bruce-Okine & Lal, 1975 ;
Donker & Damen, 1984 ), 从而引起资源胁迫的加剧, 并进而导致繁殖分配的减少.另一方面, 坡度和坡向通常会引起太阳辐射特征的变化, 如太阳入射角、入射时间在不同坡度和坡向上会发生变化(
田辉等, 2007 ), 从而引起植物可积累和利用的有效太阳辐射能量的变化.在高海拔地区, 太阳辐射对植物的生长与繁殖策略都起着重要的影响.如, 为了充分利用太阳辐射, 一些植物花器官进化出了“向日性”功能(
Zhang et al ., 2010 ).又如, 安第斯山脉中部高海拔岩石斜坡典型垫状植物
Azorella compacta 为了最大化利用太阳辐射, 其个体偏爱生长在向北的(南半球)坡度约为20°的斜坡(
Kleier et al ., 2015 ).因此, 我们推测由坡度与坡向变化引起的土壤养分状况及太阳辐射特征的变化, 可能同时对团状福禄草的繁殖分配起着重要的影响. ...
Distribution and conservation of threatened plants in China
2
2015
... 随着海拔逐渐上升, 环境条件变得更加严酷, 如: 海拔每上升100 m, 气温约下降0.6 ℃, 气压下降8-9 hPa (
K?rner, 2003 ), 土壤养分含量也随之减少(
曹舰艇等, 2019 ).面对高海拔地区低温、土壤养分贫瘠等恶劣环境因子, 高山植物在长期的进化过程中形成了丰富多样的特化适应性状或进化策略(
Sun et al ., 2014 ;
杨扬等, 2019 ), 种类丰富的植物多样性对维持高山生态系统的稳定性起到了关键作用(
Hooper et al ., 2005 ).滇西北处于横断山高山植物多样性中心区域(
郭辉军和龙春林, 1998 ;
Zhang et al. , 2015 ).长期以来, 青藏高原及横断山高海拔地区植物对极端环境因子的适应机制, 备受植物学家和生态学家的关注, 相关研究的深入开展, 加深了对该地区植物多样性形成及其进化适应机制的认识(
Sun et al ., 2014 ;
杨扬等, 2019 ). ...
... 研究区域位于滇西北德钦县白马雪山, 地理坐标为28.48° N, 98.99° E, 海拔范围为4 360?4 920 m, 生境类型从低至高依次为灌草甸、草甸-流石滩交错带和流石滩(
图1D ).滇西北处于横断山高山植物多样性中心区域(
郭辉军和龙春林, 1998 ;
Zhang et al ., 2015 ), 具有典型的季节性夏季季风气候, 以冷雨或雨夹雪为特征, 并伴有短时间的强烈太阳辐射, 年降水量680-790 mm, 年平均气温-1 ℃ (
张荣祖等, 1997 ).植物生长季节通常开始于5月中旬的融雪期, 结束于9月中下旬的夏季风停息期.我们前期的测量数据显示, 2015年6月中旬至9月下旬, 白马雪山海拔4 900 m处, 距离地面15 cm处的平均气温和湿度分别为(4.87 ± 0.04) ℃ 和(89.9 ± 0.24)%, 绝对最高气温和最低气温分别为17.80和-1.95 ℃, 昼夜温差大于10 ℃, 最高达16 ℃ (陈建国等, 未发表数据).此外, 土壤养分含量随海拔升高而普遍降低(
Chen et al ., 2015b ,
2019 ). ...
 ,
, ,1,**, 孙航
,1,**, 孙航
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT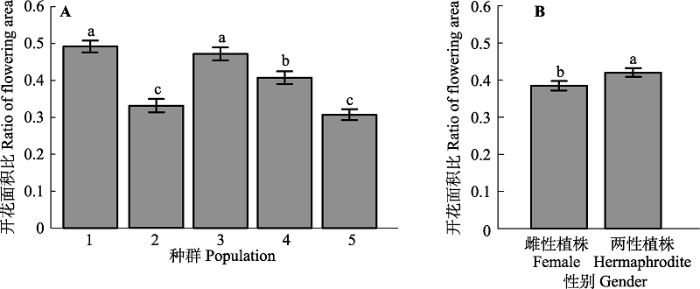 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT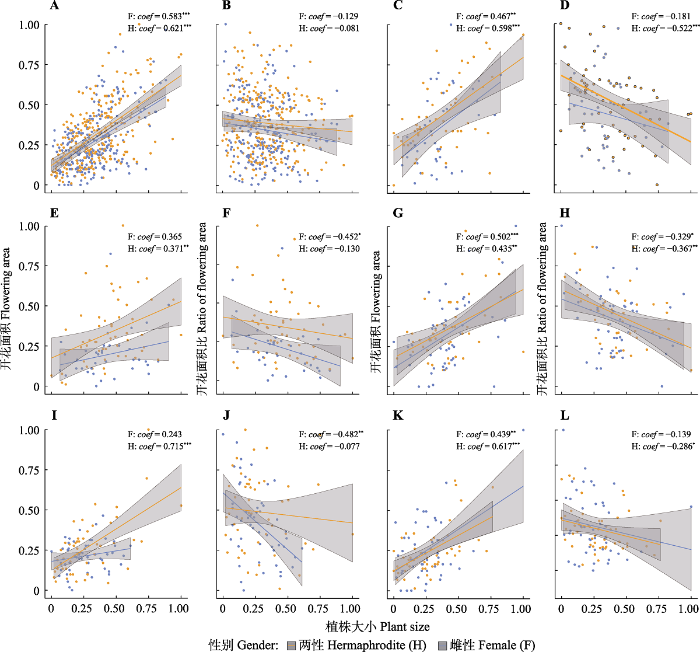 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT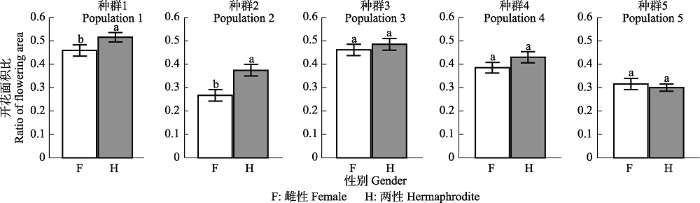 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT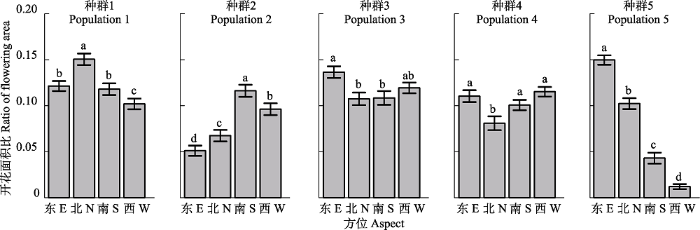 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}