 ,1,*, 孔德良,2,*
,1,*, 孔德良,2,*A review on the effects of invasive plants on mycorrhizal fungi of native plants and their underlying mechanisms
Jin-Qi TANG1, Xiao-Cheng GUO1, Xin-Yu LU1, Ming-Chao LIU1, Hai-Yan ZHANG1, Yu-Long FENG,1,*, De-Liang KONG,2,*通讯作者: *Feng YL:fyl@syau.edu.cn;Kong DL:deliangkong1999@126.com
编委: 何维明
责任编辑: 赵航
收稿日期:2020-05-25接受日期:2020-07-14网络出版日期:2020-11-20
| 基金资助: |
Corresponding authors: *Feng YL:fyl@syau.edu.cn;Kong DL:deliangkong1999@126.com
Received:2020-05-25Accepted:2020-07-14Online:2020-11-20
| Fund supported: |

摘要
菌根真菌共生是植物吸收养分的一个重要策略。外来植物可以干扰本地植物与菌根真菌的共生关系从而抑制本地植物生长, 这是近年来被发现的一种重要入侵机制, 在研究中得到日益广泛的关注。该文从以下几个方面着重综述这种入侵机制: 1)外来植物对本地植物菌根真菌的影响, 包括菌根真菌侵染率、菌根内部结构、根外菌丝的量、菌根真菌的群落组成、非菌根真菌的影响及网络结构; 2)外来植物对本地植物菌根真菌上述影响的机制, 包括资源竞争、化感作用和土壤肥力等生态机制以及相关的分子机制; 3)上述两个方面随入侵时间的变化格局。尽管干扰本地植物菌根真菌是一种重要的入侵机制, 但相对其他的入侵机制(例如天敌逃逸、新武器假说等)来说, 这类机制的研究目前仍很匮乏。鉴于此, 该文提出了未来需要重点关注的几个方面: 1)全球变化背景下, 入侵植物对本地植物菌根真菌的影响如何变化; 2)包括这种菌根机制在内的多种入侵机制之间的关系; 3)深入探究入侵的这种菌根机制在大的时空尺度上的变化规律。
关键词:
Abstract
Mycorrhizal fungi symbiosis is an important strategy for plant to uptake soil nutrients. Alien plants could thwart the symbiotic relationship between native plants and mycorrhizal fungus, and thus suppress the growth of native plants, which is an important mechanism for alien plant invasion, and has been increasingly emphasized in recent studies. In the present review, we summarized several key aspects of such mycorrhizal-related mechanism of plant invasion: 1) the impacts of alien plants on mycorrhizal fungi of native plants (i.e. mycorrhizal colonization rate, internal structures of mycorrhizal hyphae, amount of external hyphae, mycorrhizal and non-mycorrhizal composition, and mycorrhizal network); 2) mechanisms of alien-plant impacts on mycorrhizal fungi of native plants, including ecological mechanisms such as resource competition, allelopathy and edaphic fertility, as well as their molecular mechanisms; 3) variations of the above mentioned alien-plant impacts and related mechanisms at different durations of plant invasion. Despite thwarting mycorrhizal fungi of native plants is an important mechanism for alien plant invasion, researches on such mechanism are still scarce comparing to other mechanisms such as natural enemy release and new weapon hypothesis. Therefore, we proposal several research areas that need to be focused on in future studies: 1) how do global changes affect the alien-plant-invasion impacts on mycorrhizal fungi of native plants; 2) what are the relationships among different mechanisms including the mycorrhizal-related mechanism; 3) how does the mycorrhizae-related mechanism change at large spatio-temporal scales.
Keywords:
PDF (2216KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
唐金琦, 郭小城, 鲁新瑜, 刘明超, 张海艳, 冯玉龙, 孔德良. 外来入侵植物对本地植物菌根真菌的影响及其机制. 植物生态学报, 2020, 44(11): 1095-1112. DOI: 10.17521/cjpe.2020.0166
TANG Jin-Qi, GUO Xiao-Cheng, LU Xin-Yu, LIU Ming-Chao, ZHANG Hai-Yan, FENG Yu-Long, KONG De-Liang.
随着全球社会经济活动的日益紧密, 越来越多的植物被带入非原产地环境, 快速扩散成为入侵植物。全球变化背景下, 入侵植物已对当地生物多样性、社会经济以及人类健康造成重大威胁(万方浩等, 2011; 鞠瑞亭等, 2012; van Kleunen et al., 2015; Pathak et al., 2019)。探求外来植物成功入侵的机制是近几十年来生态和进化领域的一个重要议题, 同时也是进行入侵防控的前提(Wolfe et al., 2008; Catford et al., 2009; Lau & Schultheis, 2015; Wan et al., 2019)。外来植物可以通过多种途径成功入侵当地生态系统(Catford et al., 2009)。例如, 在入侵地由于缺乏有效天敌, 入侵种可以将原来分配到天敌防御上的资源(如碳和氮)用于生长繁殖而促进入侵(Blossey & Notzold, 1995; Feng et al., 2009; Zhao et al., 2020); 入侵种也会具有较高的表型可塑性, 能更好地适应环境资源的波动(Richards et al., 2006); 它们还会产生一些特殊的化感物质, 抑制本地植物生长(Callaway et al., 2008; Zheng et al., 2015)。
除了这些被广泛关注的途径, 近年来还发现一类重要的间接途径, 即入侵植物能通过干扰本地物种的共生关系(如植物-传粉昆虫和根系-菌根真菌等)而抑制本地植物的生长(Mitchell et al., 2006; Pringle et al., 2009; 张桂花等, 2009; Wilson et al., 2012)。陆地生态系统中80%以上的植物都与菌根真菌共生, 这是陆地上最古老和最重要的共生关系之一(Martin et al., 2017; Brundrett & Tedersoo, 2018; Steidinger et al., 2019; Tedersoo et al., 2020)。在这种共生关系中, 植物为菌根真菌提供碳源, 菌根真菌为宿主植物提供稀缺养分(如磷和氮等), 并帮助宿主防御病害, 适应胁迫环境(如干旱和重金属污染等)(Parniske, 2008; Bonfante & Genre, 2010; Kohler et al., 2015; Delavaux et al., 2017)。因此, 如果外来植物能干扰或破坏本地植物根系与菌根真菌的共生关系, 就可以抑制本地植物生长, 促进自身入侵。近年来, 一些研究报道了入侵植物这方面的作用(Stinson et al., 2006; Callaway et al., 2008; Vogelsang & Bever, 2009; Dickie et al., 2017), 表明对本地植物共生菌根真菌的干扰是外来植物一种重要的入侵机制。
本文围绕入侵植物对本地植物菌根真菌的影响和机制, 综述近年来国内外在这方面的进展, 并做了相关的评论和展望。我们首先分析入侵植物对本地植物菌根真菌侵染程度、菌根内部结构、根外菌丝特征、真菌群落组成及真菌网络的影响; 之后综合分析这些影响背后的生态和分子机制, 涉及资源竞争、土壤养分、化感作用、信号识别及转导等; 最后探讨外来植物入侵时间或强度的效应。
1 入侵植物对本地植物菌根真菌的影响
1.1 本地植物菌根真菌侵染程度
早在21世纪初, Stinson等(2006)就注意到了外来入侵植物对本地植物菌根真菌侵染的影响。他们发现在原产地欧洲, 葱芥(Alliaria petiolata)对其邻近植物菌根真菌侵染率的影响不显著; 但在入侵地北美显著降低了本地植物Acer saccharum、红枫(Acer rubrum)和Fraxinus americana丛枝菌根真菌(AMF)的侵染率, 下降幅度分别是30%、14%和42% (Stinson et al., 2006; Callaway et al., 2008)。Stinson等(2006)的研究引起了广泛关注, 他们的工作被认为是一种潜在的外来植物入侵的新机制。之后人们选取不同的入侵植物和本地植物体系来检验这种入侵机制的广泛性。例如, 生长在英国的本地植物长叶车前(Plantago lanceolata)、百脉根(Lotus corniculatus)和红车轴草(Trifolium pratense)也会受到入侵植物类似的影响, 在虎杖(Fallopia japonica)入侵时菌根真菌侵染率分别下降了20%、24%和20%, 而外来植物凤仙花(Impatiens glandulifera)对本地植物侵染率抑制程度弱一些, 下降幅度分别为12%、11%和19% (Tanner & Gange, 2013)。狗牙根(Cynodon dactylon)的入侵使匈牙利的小花蝇子草(Stipa borysthenica)的AMF侵染率降低25% (Endresz et al., 2013)。在我们检索到的国内外32篇相关文献中(AMF文献有27篇), 有44%的文献报道入侵植物显著降低了本地植物AMF侵染率(降低18.9%), 也有一少部分文献报道本地植物AMF侵染率未有变化甚至是增加的现象(增加9.5%)(表1)。例如, 海岸松(Pinus pinaster)和绢毛荣桦(Hakea sericea)入侵到夏威夷后当地植物群落的AMF侵染率均未发生改变(Allsopp & Holmes, 2001); 而乳浆大戟(Euphorbia esula)的入侵使本地植物AMF侵染率由原来的40%提高到79%, 入侵植物Centaurea stoebe使本地植物侵染率提升了1.2倍(Lekberg et al., 2013)。Tab.1
表1
表1入侵植物对本地植物菌根真菌侵染率和根外菌丝的影响
Tab.1
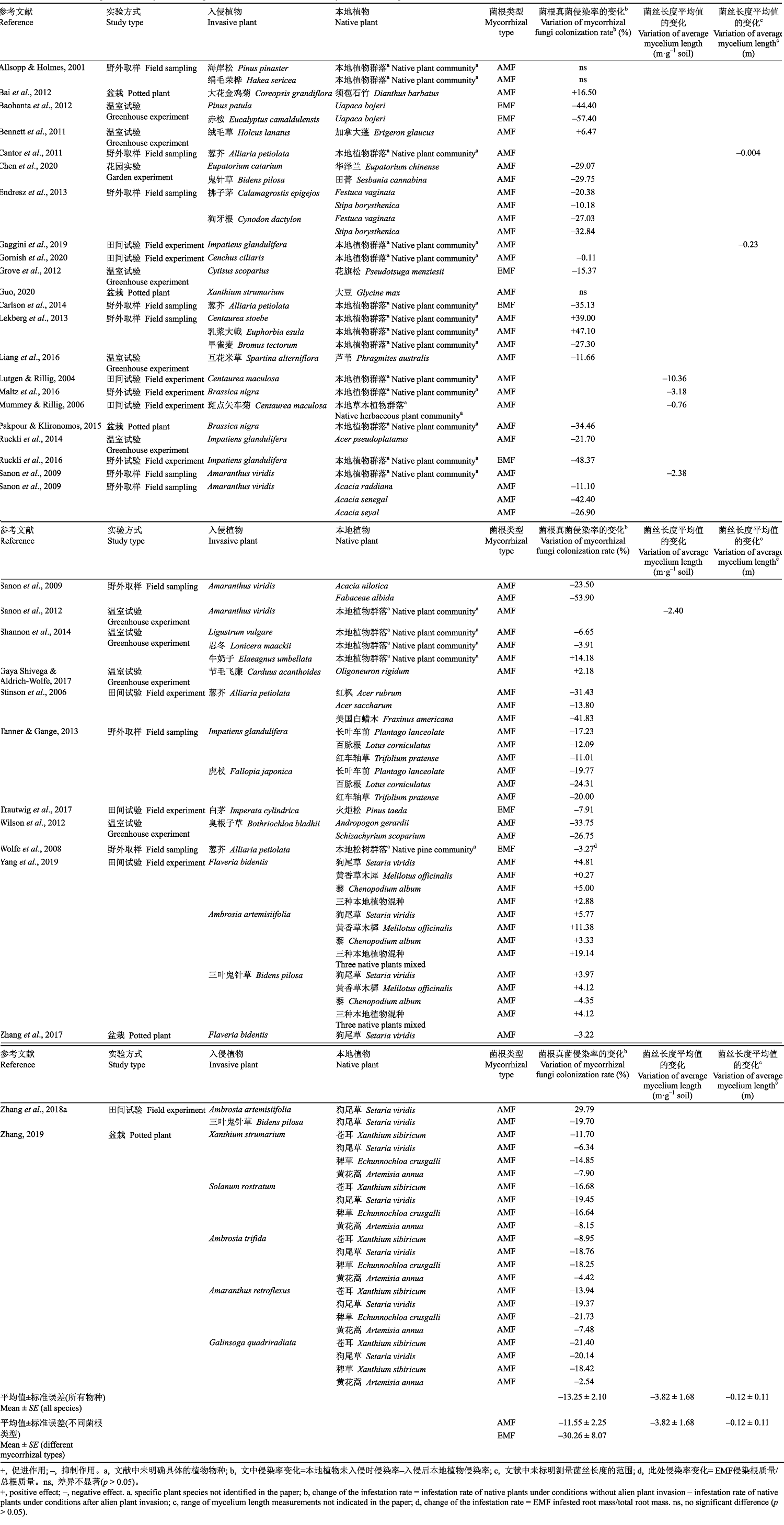 |
新窗口打开|下载CSV
相对于AMF, 外生菌根真菌(EMF)对外来干扰的抵抗能力更强(Chen et al., 2020), 但研究表明外来入侵植物能抑制本地植物EMF的侵染程度(Wolfe et al., 2008)。例如, 葱芥入侵后本地植物群落(红松(Pinus resinosa)和白松(Pinus strobus))的EMF侵染率降低约75% (Carlson et al., 2014)。我们整理搜集文献的相关数据发现, 入侵植物对EMF侵染率平均降低30%, 相对高于入侵植物对AMF的平均抑制程度(11.6%)(表1)。这表明菌根类型是影响入侵植物对本地植物菌根真菌干扰程度的重要因素。不过, 我们也注意到, 相比于AMF, 涉及入侵植物对本地植物EMF共生关系影响的研究较少。Grove等(2017a)综述的112篇相关文章中, 82%的文章阐述了入侵植物对本地AMF植物的影响, 只有18%的文章探讨了入侵植物对本地EMF植物的影响, 而且这种影响的程度在不同研究中有较大的差异。因此, 未来需要扩大对EMF植物的关注, 以检验入侵植物对本地植物EMF的抑制效果是否普遍高于对本地植物AMF的抑制性。截至目前, 除AMF和EMF这两种常见的菌根类型外, 我们还未见到入侵植物对其他一些生长在特殊环境中的重要菌根类型, 如杜鹃类菌根(ERM)、兰科菌根(ORM)等影响的报道。这可能是由于这些菌根类型生长在特殊的环境中, 例如杜鹃花科植物多生长于土壤贫瘠和干旱等恶劣环境中, 外来植物很难成功入侵到类似环境; 而兰科植物是常见的观赏花卉, 入侵植物在其生存环境中经常被园艺工人拔除, 导致研究者们忽略了入侵生物的存在。因此, 未来研究需要探讨不同菌根类型对入侵植物响应的差异和机制。
尽管入侵植物对本地植物菌根真菌影响的这些差异性被报道, 但是很少有研究者关注这些差异背后的原因。我们推测这可能至少与以下几个方面的原因有关: 1)本地植物和菌根真菌共生关系的强弱。例如, 有两个本地植物A和B, 如果A的菌根真菌侵染率远高于B (或者说A对菌根真菌的依赖性远超过B)(Eissenstat et al., 2015; Bergmann et al., 2020), 那么入侵植物对A的菌根真菌侵染率的影响要超过B。2)入侵植物的竞争力和化感作用的大小。例如, 张海艳(2019)研究发现竞争力较强的入侵种(Xanthium strumarium、Solanum rostratum、Ambrosia trifida、Amaranthus retroflexus和Galinsoga quadriradiata)对本地植物苍耳(Xanthium sibiricum)菌根真菌的影响反而较小, 这可能是由于入侵种的化感物质较弱。3)土壤养分的高低。例如, 张海艳(2019)研究发现入侵植物S. rostratum在低养分的情况下对本地植物菌根真菌的影响更显著, 这可能是低养分情况下本地植物的菌根真菌对入侵植物的竞争力和化感作用更敏感所致。
以往研究大多只关注具有单共生关系的本地植物(只受菌根真菌侵染), 而对具有双共生关系的本地植物(根瘤菌和AMF共同侵染, 如豆科植物)的研究很少。我们团队的研究发现, 入侵植物X. strumarium抑制了本地植物苍耳(单共生关系)的AMF侵染率(张海艳, 2019), 但没有改变本地植物大豆(Glycine max)(双共生关系)的AMF侵染率(郭小城, 2020)。这可能是物种本身的差异所致, 但也不排除根瘤的存在改变了菌根真菌对入侵植物的响应, 需要进一步研究。这里只涉及一种豆科植物, 我们还不清楚别的豆科植物以及其他双共生体系中菌根真菌对外来入侵植物的响应是否有差异。
1.2 本地植物菌根内部结构和功能
外来植物入侵后, 本地植物菌根真菌侵染率有时没有受到显著影响(Allison et al., 2006), 但这并不等于外来入侵植物不影响本地植物与菌根真菌的共生关系, 菌根的内部结构和功能可能发生了显著变化。丛枝菌根内部存在多种结构, 包括泡囊、丛枝、菌丝环以及根内菌丝(图1)。这些不同真菌结构具有不同的功能, 例如泡囊可以存储真菌脂质及其他成分, 丛枝和菌丝环是植物与菌根真菌之间进行碳和养分交换的部位, 根内菌丝可以帮助真菌在根内定殖并在菌丝内部运输养分(Johnson et al., 2003; Smith & Read, 2008)。菌根真菌的这些内部侵染结构会受到环境的影响。Lugli等(2019)研究表明, 土壤中磷的多少会改变丛枝菌根内部结构, 进而影响菌根功能。入侵植物可以提高入侵地的土壤养分含量(Pattison et al., 2016; Zhang et al., 2018a), 例如Liao等(2008)的meta分析表明, 入侵植物可以使入侵地的土壤养分平均增加30%。入侵地土壤养分的提升可能会导致本地植物菌根真菌侵染的改变, 包括侵染率和菌根内部结构两个方面。然而, 目前有关外来入侵植物对本地植物菌根真菌内部结构影响的研究很少, 在未来研究中需要加强。图1
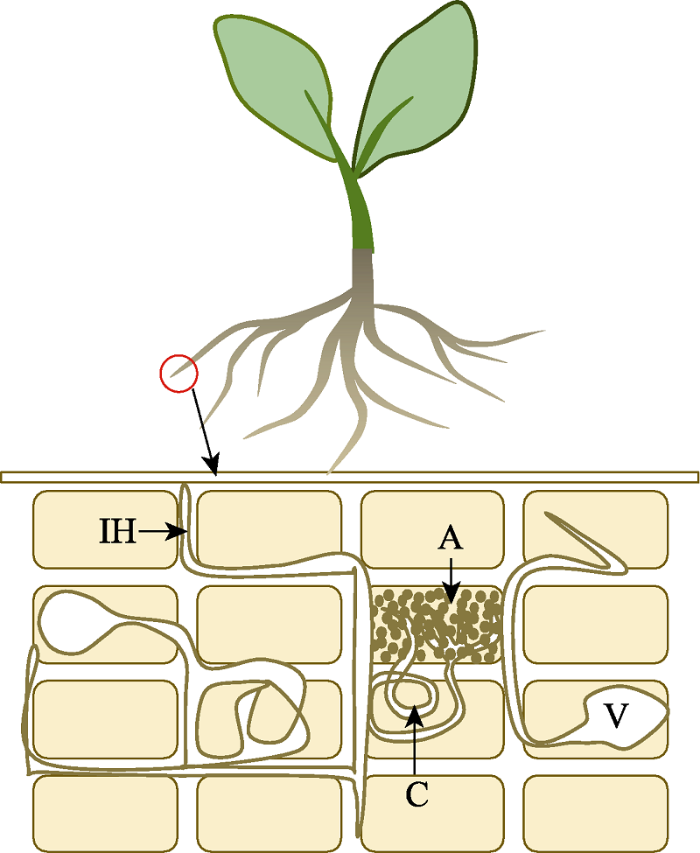 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1丛枝菌根内部结构概念图(改自Johnson等(2003))。A, 丛枝; C, 菌丝环; IH, 根内菌丝; V, 泡囊。
Fig. 1A conceptual diagram of the internal structures of arbuscular mycorrhizae in root tissues (adapted from Johnson et al., 2003). A, arbuscules; C, coils; IH, intraradical hyphae; V, vesicles.
1.3 本地植物根外菌丝特征
菌根的根内菌丝延伸至根外形成根外菌丝, 根外菌丝直接参与土壤养分的吸收。外来入侵植物在对本地植物菌根真菌侵染率、真菌内部结构产生影响的同时, 也会对根外菌丝产生影响(Lutgen & Rillig, 2004; Luo et al., 2019)(表1)。例如, Mummey和Rillig (2006)研究发现, 外来植物斑点矢车菊(Centaurea maculosa)的入侵使本地植物的AMF根外菌丝长度降低24%; Gaggini等(2019)发现, 凤仙花入侵一年使本地植物根外AMF菌丝生物量降低30%-80%。总体来说, 相对于菌根真菌侵染率, 有关根外菌丝特征对入侵植物响应的研究较少, 且相关研究中对根外菌丝的量化标准也不统一, 有的使用菌丝长度, 有的则使用菌丝生物量, 不利于将来进行整合分析。我们推荐使用菌丝长度这一指标, 因为其易于测定且成本低。入侵植物除了影响本地植物根外菌丝的长度和生物量外, 还可能会进一步影响根外菌丝与吸收功能密切相关的一些生理特征。例如, 1)根外菌丝分泌蛋白水解酶的能力, 这些水解酶往往参与分解土壤有机质和生物残体, 释放其中的养分(Martin et al., 2008; Zhang et al., 2019)。2)转运蛋白将土壤养分转运至菌丝内的效率(Bonfante & Genre, 2010; 田磊等, 2016; Jiang et al., 2017; Zhang et al., 2018b)。1.4 本地植物菌根真菌群落组成
入侵植物除了影响本地植物菌根真菌的侵染率、根内和根外菌丝的结构与功能外, 还可能改变其菌根真菌的组成。有研究发现, 外来植物入侵会使本地植物原有的AMF被其他种类的AMF替代(Hawkes et al., 2006; Barto et al., 2011; Jordan et al., 2011), 或有部分AMF被替代(Burke, 2008)。Zhang等(2018c)研究发现, 外来入侵植物提高了本地植物根际土微生物群落中AMF所占比例, 这可能会提高本地植物菌根真菌中AMF比例。但也有研究发现, 外来植物入侵未改变本地植物的菌根真菌群落组成(Koch et al., 2011; Meinhardt & Gehring, 2012), 如凤仙花入侵一年后本地植物菌根真菌组成未发生变化(Gaggini et al., 2018)。张海艳(2019)研究发现, 一方面同一本地植物菌根真菌群落组成对不同入侵植物的响应不一致, 如入侵植物X. strumarium改变了黄花蒿根内菌根真菌群落组成, 而入侵植物S. rostratum则没有影响; 另一方面同一入侵植物对不同本地植物菌根真菌群落组成的影响也不一致, 如入侵植物X. strumarium改变了黄花蒿的菌根真菌群落组成, 但未改变本地植物狗尾草的菌根真菌群落组成。这表明本地植物菌根真菌群落组成对入侵植物的响应既与本地植物自身特性有关, 也与入侵植物种类有关。但目前有关入侵植物对本地植物菌根真菌群落组成影响的研究大多数集中在AMF植物, 我们对本地植物EMF和其他类型菌根真菌群落组成如何响应入侵植物的认识十分有限。上述这些研究中, 本地植物菌根真菌群落组成都是通过变性梯度凝胶电泳(DGGE)(肖龙敏, 2018)、磷脂脂肪酸(PLFA)(席浩, 2019)和高通量测序(蒋玉玲等, 2019)等方法测定的。除此之外, 菌根真菌也可以按照其功能进行分类。例如, Kiers等(2011)将菌根真菌分为合作型真菌和寡合作型真菌, 合作型真菌Glomus intraradices对宿主的养分回报效率(真菌转运给植物的养分/真菌接受植物的碳之比)高于寡合作型真菌Glomus aggregatum。由于合作型真菌对宿主具有更高的养分回报率, 我们推测共生真菌中以合作型真菌为主的本地植物可能受外来植物的影响更大。尽管这种推测有待验证, 但我们认为这种基于真菌功能的真菌群落组成研究是未来研究中值得关注的一个方向, 这将有助于我们深入了解入侵植物对菌根真菌的影响及外来植物入侵机制。
1.5 本地植物菌根真菌网络
同种植物常常能与多种菌根真菌共生, 即使是单个根尖也往往被多个特定的菌根真菌类群侵染(Sepp et al., 2019)。这些菌根真菌之间由于不同强度和方向(包括正和负)的相互作用形成复杂的菌根真菌网络, 该网络的连接性、模块性往往用来评判真菌网络结构的稳定性(图2)(Bennett et al., 2013; de Vries & Wallenstein, 2017)。Sepp等(2019)将植物分为专性菌根植物(只依赖菌根吸收养分)和兼性菌根植物(除菌根外还通过其他方式吸收养分, 如豆科植物的根瘤), 他们发现专性菌根植物真菌网络的连接性更强。理论上, 不同的菌根真菌可能构成不同的网络结构, 而相同的菌根真菌群落组成也可能构成不同的网络结构。目前研究者往往关注胁迫环境对菌根真菌网络的影响以及同一环境下不同菌根类型的真菌网络之间的差异(Toju et al., 2018; 杨思琪等, 2019)。例如, de Vries等(2018)发现, 干旱情况下真菌共生网络的复杂性和稳定性均有所降低; 在AMF和EMF植物共存的系统中, AMF网络与宿主植物的连接性更强, EMF网络与宿主植物的专一性(共生伙伴偏好性)更强(杨思琪等, 2019)。菌根真菌网络的研究方兴未艾, 而菌根真菌网络对入侵植物响应的研究尚属空白。由于入侵植物会显著改变本地植物菌根真菌的组成和相对多度(Zhang et al., 2018c; 张海艳, 2019), 我们推断这些都会影响到真菌网络的连接性和模块性; 由于EMF对环境干扰的抗性强于AMF, 我们预测入侵植物对AMF网络结构的影响高于EMF网络。当然这里的预测仅仅是一些初步的想法, 未来还需要全面和深入的实验验证。图2
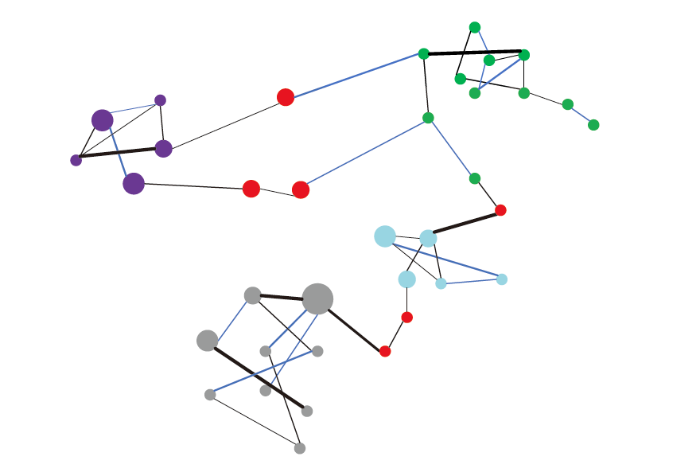 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2菌根真菌网络结构概念图(改自de Vries和Wallenstein (2017))。不同颜色的圆圈表示不同的模块(如不同种类的丛枝菌根真菌), 各模块的连接点用红色圆圈表示, 线粗细表示连接性强弱, 黑线表示正相关, 蓝线表示负相关。
Fig. 2A conceptual diagram of the mycorrhizal fungal network structure (adapted from de Vries & Wallenstein, 2017). Circles with different colors indicate different modules (i.e., different mycorrhizal fungal communities). Module connections are indicated by red circles. Thicker lines indicate stronger mycorrhizal fungi connections. The black lines indicate positive correlations, and the blue lines indicate negative correlations.
1.6 本地植物非菌根真菌
外来植物入侵不仅会影响本地植物菌根真菌, 也会影响其非菌根真菌(NMF), 包括致病真菌、寄生真菌、一些对植物可能无害的内生真菌以及在极端环境下可以帮助植物生存的有益内生菌(Hawkes et al., 2006; Meinhardt & Gehring, 2012; Hill et al., 2019)。有研究根据微生物之间的互作关系将本地植物根内微生物分为两类: 促进型和拮抗型。促进型微生物的存在能为其他微生物甚至是病原菌的侵染提供便利条件, 而拮抗型微生物不仅会抑制互惠微生物, 还会抑制致病菌的入侵(Li et al., 2019)。菌根真菌与NMF之间也存在拮抗关系(Bufford et al., 2019; Sarkar et al., 2019)。例如, 根中AMF侵染增加会使NMF的侵染下降(Maherali & Klironomos, 2007; Liang et al., 2015; Barnes et al., 2018); EMF也能有效抵抗病原菌的侵染(Chen et al., 2019)。Hawkes等(2006)发现, 外来植物旱雀麦(Bromus tectorum)入侵使两种本地植物的AMF丰富度降低, 而NMF所占比例增加。如果这些NMF属于致病菌, 那么旱雀麦入侵将使本地植物的生长受到双重抑制: 一是AMF丰富度减少带来植物养分吸收的降低; 二是致病真菌所占比例增加对植物的危害(Meinhardt & Gehring, 2012)。这说明入侵植物可以通过影响本地植物NMF间接影响菌根真菌与本地植物生长间的关系(图3)。因此, 入侵植物不仅可以直接影响本地植物菌根真菌的组成和丰富度等, 还可以通过改变NMF中有益真菌与有害真菌的组成或/和比例, 进而间接影响本地植物菌根真菌的侵染率、菌根内部真菌结构、根外菌丝、真菌群落组成及真菌网络等(图3)。在研究入侵植物对本地植物菌根真菌影响时, 我们应充分考虑到NMF的变化, 并查明这些NMF的“身份” (致病真菌、寄生真菌、无害真菌, 还是有益真菌)。
图3
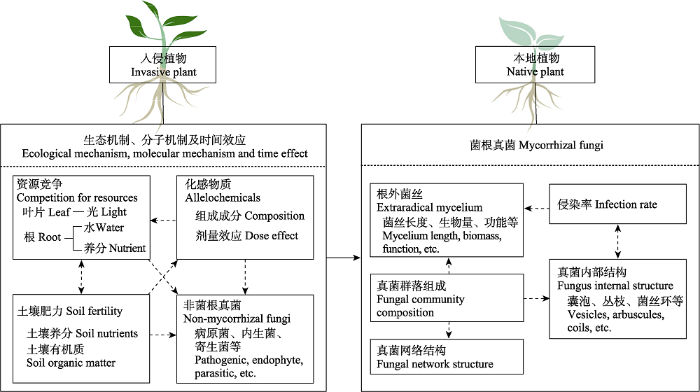 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3外来入侵植物对本地植物菌根真菌影响的概念图(改自Grove等(2017a))。实线箭头表示改变; 虚线箭头表示存在未知的关系。
Fig. 3Conceptual illustration showing how invasive plants affect the symbiotic mycorrhizal fungi in native plant roots (adapted from Grove et al., 2017a). Solid arrows indicate change; dotted arrows indicate possible relationships.
2 入侵植物影响本地植物菌根真菌的机制
2.1 生态机制
入侵植物可以通过多种生态机制影响本地植物的菌根真菌(Grove et al., 2017a), 目前国内外相关研究主要集中在物种竞争、土壤养分变化和化感作用3个方面。如表2所示, 入侵植物往往具有较强的资源(光和养分等)竞争能力。例如, Solanum carolinense有着强大的无性繁殖能力可以快速扩散(魏雪苹等, 2020)。入侵植物还可以抑制本地植物光合作用(Funk & Vitousek, 2007; van Kleunen et al., 2010; Huang et al., 2020), 光合产物的减少可能会抑制本地植物菌根真菌侵染(Fellbaum et al., 2012) (图3)。入侵植物往往具有较强的养分竞争力和大量的根系分泌物(Liao et al., 2008; Grove et al., 2017b; 韩雪等, 2020; Huang et al., 2020)(表2)。入侵植物还可以通过凋落物和根系分泌物等改善和提高土壤养分(Zhang et al., 2018b; Pérez Castro et al., 2019), 形成植物和土壤之间的正反馈(Zhang et al., 2018c, 2020; Zhao et al., 2019, 2020), 进而降低本地植物(如Solidago decurrens和Andropogon gerardii)菌根真菌的侵染率, 改变菌根真菌的群落组成(陆建忠等, 2005; Johnson et al., 2010)(图3)。入侵植物还可以通过化感作用对本地植物菌根真菌产生直接影响(Thorpe et al., 2009; Soudzilovskaia et al., 2015)(图3)。入侵植物产生的化感物质(如类黄酮和芥子油苷等)能抑制菌根真菌孢子萌发、生长和侵染, 从而抑制依赖这些真菌的本地植物的生长(Callaway & Aschehoug, 2000; Callaway & Ridenour, 2004; Prati & Bossdorf, 2004; Callaway et al., 2008)。这些化感物质也可能通过抑制本地植物的生长(Zheng et al., 2015), 从而降低植物对菌根真菌碳的供应, 进而抑制菌根真菌侵染。此外, 外来植物所分泌的化感物质可能也会影响本地植物的非菌根真菌, 进而对本地植物菌根真菌产生间接影响(图3)。实际上, 本地植物菌根真菌的变化可能是多种机制共同作用的结果(Grove et al., 2017a, 2017b)。不同入侵植物对本地植物菌根真菌的影响机制可能不同, 而不同本地植物菌根真菌对每种机制的响应可能也有差异, 这使得不同外来植物入侵对不同本地植物菌根真菌的影响可能呈现出不同的趋势。Table 2
表2
表2外来植物入侵的生态机制
Table 2
| 生态机制 Mechanism of ecology | 举例 Example | 参考文献 Reference |
|---|---|---|
| 物种竞争 Species competition | 与农作物(如高粱(Sorghum bicolor)、玉米(Zea mays))相比, 入侵植物Parthenium hysterophorus具有快速早期生长的能力。 Compared with crops (such as Sorghum bicolor and Zea mays), the invasive plant Parthenium hysterophorus can grow much more rapidly in their early stage. | Bajwa et al., 2016 |
| 比较夏威夷19对入侵植物和本地植物的资源利用效率, 发现入侵植物具有较高的碳同化率、光利用效率、瞬时氮利用效率和瞬时能量利用效率。 Comparing the resource utilization efficiency for 19 paired invasive-native plants in Hawaii, invasive plants are observed to have higher carbon assimilation rate, light use efficiency, instantaneous nitrogen and energy use efficiency. | Funk & Vitousek, 2007 | |
| 比较47对入侵植物和非入侵植物对草食性动物的抵抗能力及养分含量发现, 入侵植物拥有更高的叶氮含量, 受到食草动物的损伤较小。 Comparisons among 47 paired invasive and non-invasive plants for leaf herbivore-resistance and nutrient content show that invasive plants have higher leaf nitrogen content and less damage by herbivores. | Huang et al., 2020 | |
| 比对125种入侵植物和196种非入侵植物的生理性状发现, 入侵植物在生长率、养分分配及抗逆性更有优势。 Comparing physiological traits of 125 invasive with plants 196 non-invasive plants show that invasive plants have more advantages in growth rate, nutrient allocation and stress resistance. | van Kleunen et al., 2010 | |
| 外来植物Solanum carolinense具有较强的抗寒能力和无性繁殖能力。 Exotic plant, Solanum carolinense, has strong ability for cold resistance and asexual reproduction. | Wei et al., 2020 | |
| 气候变暖及氮沉降的增加可以增强入侵植物Solidago canadensis叶片的资源获取能力。 Climate warming and increased nitrogen deposition can enhance leaf resource acquisition of an invasive plant, Solidago canadensis. | Zhou et al., 2019 | |
| 化感物质 Allelochemical | 入侵植物白羊草(Bothriochloa ischaemum)的渗滤液抑制本地植物Andropogon gerardii和Schizachyrium scoparium发芽及生长。 The leachate of an invasive plant, Bothriochloa ischaemum, inhibits germination and growth of the native plants, Andropogon gerardii and Schizachyrium scoparium. | Greer et al., 2014 |
| 入侵植物水烛(Typha angustifolia)根系分泌的酚类物质抑制了本地植物Bolboschoenus fluviatilis的生长发育。 Phenolic compounds secreted by roots of an invasive plant, Typha angustifolia, inhibit growth and development of a native plant, Bolboschoenus fluviatilis. | Jarchow & Cook, 2009 | |
| Solidago canadensis入侵分泌的化感物质抑制了本地植物莴苣(Lactuca sativa)种子萌发和幼苗生长。 Allelochemicals secreted by the invasion of Solidago canadensis inhibit seed germination and seedling growth of a native plant, Lactuca sativa. | Hu et al., 2020 | |
| 外来植物Solanum carolinense含有龙葵碱及草酸盐结晶。 The exotic plant Solanum carolinense contains solanine and oxalate crystals. | Wei et al., 2020 | |
| Parthenium hysterophorus可以分泌帕台单宁、咖啡酸、香草酸等化感物质帮助自身入侵。 Parthenium hysterophorus can secrete parthenin, caffeic acid, vanillic acid and other allelochemicals to facilitate its invasion. | Shi & Adkins, 2018 | |
| 土壤养分变化 Soil nutrient variation | 氮沉降会增强S. canadensis入侵初期分泌的化感物质对本地植物莴苣种子萌发和幼苗生长的抑制程度。 Nitrogen deposition can enhance the inhibition of the allelochemicals secreted by S. canadensis at its initial invasive stage on seed germination and seedling growth of a native plant, Lactuca sativa. | Hu et al., 2020 |
| 分析94篇文章发现, 入侵植物增大了植物、土壤和土壤微生物中的碳和氮库大小。 Analysis of 94 published papers show that invasive plants increase the size of carbon and nitrogen pools in plants, soils and soil microorganisms. | Liao et al., 2008 | |
| 与本地植物Artemisia californica、Salvia mellifera、Salvia apiana、锦鸡儿(Calochortus splendens)和Dichelostemma capitatum相比, 外来植物Centaurea stoebe、Hirschfeldia incana和Bromus madritensis可以通过加快凋落物分解改善土壤养分。 Compared with native plants such as Artemisia californica, Salvia mellifera, Salvia apiana, Calochortus splendens and Dichelostemma capitatum, exotic plants, like Centaurea stoebe, Hirschfeldia incana and Bromus madritensis, can increase soil nutrient level by increasing litter decomposition. | Pérez Castro et al., 2019 | |
| 入侵植物Mikania micrantha分泌的化感物质绿原酸和β-石竹烯加快了土壤有机氮的矿化过程。 Allelochemicals secreted by the invasive plant, Mikania micrantha, such as chlorogenic acid and β-caryophyllene accelerate mineralization of soil organic nitrogen. | Yu et al., 2020 | |
| 入侵植物通过凋落物改变了根际微生物土壤群落和养分循环, 形成植物-土壤正反馈调节。 Invasive plants can form a positive plant-soil feedback via its litter decomposition which could change rhizosphere soil microbial communities and nutrient cycling. | Zhang et al., 2018c | |
| 杂草Conyza canadensis入侵的土壤中有效氮和有机质含量增加。 The content of available nitrogen and organic matter increases in the soil invaded by Conyza canadensis. | Zhang et al., 2020 | |
| 与非入侵地土壤相比, Eupatorium catarium入侵后土壤氮含量提高4.32 mg·kg-1。 Compared with non-invasive soils, soil nitrogen content increases by 4.32 mg·kg-1 after invasion by Eupatorium catarium. | Zhao et al., 2019 | |
| Ambrosia trifida在入侵地的土壤-植物反馈高于原产地。 Ambrosia trifida has a greater plant-soil feedback in invaded ranges than that in its native ranges. | Zhao et al., 2020 |
新窗口打开|下载CSV
2.2 分子机制
植物与菌根真菌形成共生关系是一个复杂的信号转导过程。目前已经证实, 丛枝菌根的形成始于根系与AMF之间的信号交流(Gutjahr & Parniske, 2013; Tian et al., 2019)。首先, 根系(如, 莲刺参(Lotus japonicus))分泌独脚金内酯等信号分子, 促进AMF孢子萌发和菌丝分枝, 增加菌丝与根系接触的概率(Akiyama et al., 2005)。之后, 从枝菌根真菌分泌多种信号分子(即菌根因子, 如LCOs、CO4/ CO5和PsMiSSP10b), 被根系相应的受体识别后激活钙离子信号通路, 进而形成侵染线, 激活菌根真菌的侵染过程(Maillet et al., 2011; Martin et al., 2017; Plett et al., 2020)(图4)。例如, 水稻AMF分泌脂壳寡糖(LCOs)和短链壳寡糖(CO4/CO5)因子, 能被水稻中LysM受体样激酶的异源体(包括OsMYR1/ OsLYK2和OsCERK1)识别(He et al., 2019)。但在豆科植物中, AMF释放的LCOs和CO4/CO5因子, 被NFR1/LYK3识别(Zhang et al., 2015)。在上述过程中, 菌根真菌除了产生小分子分泌蛋白(MiSSPs)来识别宿主之外, 也会产生植物细胞壁分解酶(PCWDEs) (Martin et al., 2008, 2017; Kohler et al., 2015)来破坏宿主植物的细胞壁, 为共生关系的建立扫除“障碍”。而根与菌根真菌之间关系的核心在于两者对于碳(脂类和糖)和养分的交换(Jiang et al., 2017), 这种物质交换过程涉及一系列脂类合成酶以及养分转运蛋白。例如, 丝裂原活化蛋白激酶(MAPKs)在植物和AMF信号沟通中起到重要作用; STR转运蛋白负责紫花苜蓿(Medicago truncatula)中脂肪酸的转运(Bonfante & Genre, 2010; 田磊等, 2016; Bravo et al., 2017)。入侵植物可以通过影响酶和这些转运蛋白干扰本地植物菌根真菌侵染进而影响本地植物生长。如Bravo等(2017)发现, 紫花苜蓿根系脂质合成酶(FatM和RAM2)突变后, AMF的侵染率降低, 这两个酶调控植物对菌根真菌所需脂质的供应。因此, 对根与菌根真菌物质交换过程的影响也可能是入侵植物干扰本地植物菌根真菌的一个重要的分子机制, 是未来研究中值得进一步关注的领域。图4
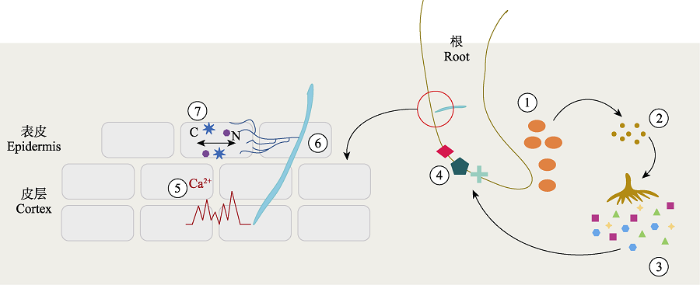 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4植物与菌根真菌共生关系分子模式图(参考Martin等(2017))。根系分泌信号分子(①), 促进土壤中丛枝孢子(②)的萌发和菌丝分枝; AMF分泌的菌根因子(③)被根系上的受体(④)识别并激活钙离子信号通路(⑤), 进而形成侵染线(⑥); 菌根真菌菌丝侵入根细胞后, 与根系进行碳-养分交换, 这中间涉及一系列酶和转运蛋白(⑦)的共同作用。
Fig. 4Conceptual model for the symbiotic relationship between plants and mycorrhizal fungi (adapted from Martin et al., 2017). Roots secrete signaling molecules (①) promote germination of AMF spores (②) and mycelium branching in the soil; mycorrhizal factors secreted by AMF (③) are recognized by receptor proteins in root cells (④), and then activate the calcium signaling pathway (⑤) to initiate formation of invasion lines of the mycorrhizal fungi (⑥). After that, the carbon and nutrient exchange between mycorrhizae and root cells also requires a series of enzymes and transport proteins in the root-mycorrhiza interface (⑦).
3 入侵时间的影响
已有研究证实外来植物的成功入侵受入侵时间的影响(Catford et al., 2019)。在入侵植物对本地植物菌根真菌影响的相关研究中, 大多数研究都是在某个特定的入侵时间或入侵阶段进行的, 不同入侵时间和阶段的对比还较缺乏(Grove et al., 2017a), 更不了解外来植物入侵对本地植物菌根真菌的影响是否也具有时间效应。另一方面, 相关研究中入侵时间序列的跨度较短。例如, Ruckli等(2016)比较研究了I. glandulifera入侵3和15个月时, 本地EMF植物青冈(Fagus sylvatica)的菌根真菌侵染率分别降低33%和66%。入侵植物的这种影响在一定入侵年限之后可能会有较大改变(Lankau et al., 2009; Dostál et al., 2013)。Grove等(2017b)研究发现, 入侵植物Cytisus scoparius对本地植物花旗松(Pseudotsuga menziesii)菌根真菌的影响在入侵3-31年间保持稳定。不过研究者也承认, 入侵更长时间之后C. scoparius的这种影响也可能减弱。随着入侵时间延长, 本地植物与土壤微生物会对入侵植物做出适应性响应, 从而消除入侵植物带来的负面影响(类延宝等, 2010)。我们推测, 随着入侵时间的持续, 外来植物对本地植物菌根真菌的影响会减弱, 且入侵植物影响菌根真菌的机制, 如前述的植物竞争(Hawkes, 2007; Yelenik & D’Antonio, 2013)、土壤养分(Grove et al., 2017b)及化感作用(Lankau et al., 2009; Li et al., 2017; 胡文杰等, 2020)等均会随入侵时间的延长而发生变化, 这些变化的方向和程度的差异, 最终可能会导致入侵的时间效应在入侵种间出现差异。我们建议通过连年栽培入侵植物, 驯化土壤以构建入侵时间序列(Flory & D’Antonio, 2015)。这种连年栽培实验, 一方面可以直接观测入侵植物种群的长期动态; 更重要的是, 我们可以系统地检验入侵植物对本地植物菌根真菌影响的上述几种机制(竞争、化感以及对土壤养分的调节等)的机制的作用及动态变化。这些研究能够为不同入侵阶段的外来植物的防控和管理提供针对性的措施。4 展望
外来植物通过干扰本地植物菌根真菌获得竞争优势是一种重要的入侵机制。尽管如此, 相对于其他入侵机理, 我们对外来植物入侵与本地植物菌根真菌间关系的认识仍然十分匮乏(Jordan et al., 2011; Gaggini et al., 2018; 孙建茹等, 2019)。这主要表现在: 1)目前入侵植物对本地植物菌根真菌影响的研究结果存在较大差异。一方面可能是由于所选植物类群有限选材范围较为狭窄(Allsopp & Holmes, 2001; Endresz et al., 2013; Lekberg et al., 2013); 另一方面菌根内部结构、根外菌丝相关功能蛋白和真菌网络结构的研究相对较少, 还无法得到普遍结论(Toju et al., 2018; Lugli et al., 2019; Luo et al., 2019); 此外研究者们经常忽略非菌根真菌(如病原菌)在外来植物入侵中起到的作用(Hawkes et al., 2006; 初亚男等, 2018)。2)相对于生态机制, 入侵植物如何影响本地植物菌根真菌与宿主根系的相互识别、侵染及养分交换的分子机制研究较少(He et al., 2019; Stefanowicz et al., 2019)。3)我们还不清楚外来植物长时间入侵时本地植物菌根真菌的响应(Dombrowski et al., 2017; Grove et al., 2017b)。此外, 我们认为在入侵植物影响本地植物菌根真菌相关研究中, 未来还应该重点关注(但不仅仅局限于)以下几个方面: 1)全球变化(如全球变暖、氮沉降、干旱、火灾及虫害等)如何调节外来植物对本地植物菌根真菌的影响, 尤其需要关注多种全球变化因子在多个水平时的交互作用。2)同时关注入侵植物对本地植物菌根真菌影响的多种生态机制(资源竞争、化感作用和土壤养分调节等)和分子机制(菌丝对土壤养分的吸收、转运以及和植物之间进行的碳-养分交换), 以及这些机制之间的相互关系。3)以往的研究往往局限于有限的空间范围和较短的时间, 未来的研究更需要探究在较大的时间(十数年甚至上百年)和空间(热带、亚热带和温带)尺度上研究入侵植物对本地植物菌根真菌影响的变化趋势以及相应的生态和分子机制。
致谢
感谢河南农业大学引进人才启动经费(30500854)的资助。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1038/nature03608URLPMID:15944706 [本文引用: 1]

Arbuscular mycorrhizal (AM) fungi form mutualistic, symbiotic associations with the roots of more than 80% of land plants. The fungi are incapable of completing their life cycle in the absence of a host root. Their spores can germinate and grow in the absence of a host, but their hyphal growth is very limited. Little is known about the molecular mechanisms that govern signalling and recognition between AM fungi and their host plants. In one of the first stages of host recognition, the hyphae of AM fungi show extensive branching in the vicinity of host roots before formation of the appressorium, the structure used to penetrate the plant root. Host roots are known to release signalling molecules that trigger hyphal branching, but these branching factors have not been isolated. Here we have isolated a branching factor from the root exudates of Lotus japonicus and used spectroscopic analysis and chemical synthesis to identify it as a strigolactone, 5-deoxy-strigol. Strigolactones are a group of sesquiterpene lactones, previously isolated as seed-germination stimulants for the parasitic weeds Striga and Orobanche. The natural strigolactones 5-deoxy-strigol, sorgolactone and strigol, and a synthetic analogue, GR24, induced extensive hyphal branching in germinating spores of the AM fungus Gigaspora margarita at very low concentrations.
DOI:10.1016/j.soilbio.2005.11.008URL [本文引用: 1]
DOI:10.1016/S0254-6299(15)31113-3URL [本文引用: 2]
DOI:10.1007/s00425-016-2510-xURLPMID:27056056 [本文引用: 1]
MAIN CONCLUSION: This review provides an insight into alien plant invasion taking into account the invasion mechanism of parthenium weed ( Parthenium hysterophorus L.). A multi-lateral understanding of the invasion biology of this weed has pragmatic implications for weed ecology and management. Biological invasions are one of the major drivers of restructuring and malfunctioning of ecosystems. Invasive plant species not only change the dynamics of species composition and biodiversity but also hinder the system productivity and efficiency in invaded regions. Parthenium weed, a well-known noxious invasive species, has invaded diverse climatic and biogeographic regions in more than 40 countries across five continents. Efforts are under way to minimize the parthenium weed-induced environmental, agricultural, social, and economic impacts. However, insufficient information regarding its invasion mechanism and interference with ecosystem stability is available. It is hard to devise effective management strategies without understanding the invasion process. Here, we reviewed the mechanism of parthenium weed invasion. Our main conclusions are: (1) morphological advantages, unique reproductive biology, competitive ability, escape from natural enemies in non-native regions, and a C3/C4 photosynthesis are all likely to be involved in parthenium weed invasiveness. (2) Tolerance to abiotic stresses and ability to grow in wide range of edaphic conditions are thought to be additional invasion tools on a physiological front. (3) An allelopathic potential of parthenium weed against crop, weed and pasture species, with multiple modes of allelochemical expression, may also be responsible for its invasion success. Moreover, the release of novel allelochemicals in non-native environments might have a pivotal role in parthenium weed invasion. (4) Genetic diversity found among different populations and biotypes of parthenium weed, based on geographic, edaphic, climatic, and ecological ranges, might also be a strong contributor towards its invasion success. (5) Rising temperatures and atmospheric carbon dioxide (CO2) concentrations and changing rainfall patterns, all within the present day climate change prediction range are favorable for parthenium weed growth, its reproductive output, and therefore its future spread and infestation. (6) Parthenium weed invasion in South Asia depicts the relative and overlapping contribution of all the above-mentioned mechanisms. Such an understanding of the core phenomena regulating the invasion biology has pragmatic implications for its management. A better understanding of the interaction of physiological processes, ecological functions, and genetic makeup within a range of environments may help to devise appropriate management strategies for parthenium weed.
DOI:10.1007/s10530-012-0238-5URL
The objectives of this study were to determine the impact of two exotic tree species (pine and eucalypts) on the early growth of Uapaca bojeri (an endemic tree species from Madagascar) via their influence on soil chemical, microbial characteristics, on ectomycorrhizal fungal community structures in a Madagascarian highland forest and to test the ability of an early-successional ectomycorrhizal shrub, Leptolena bojeriana, to mitigate the impacts of these exotic species. Finally, we hypothesized that L. bojeriana could act as a natural provider for ectomycorrhizal propagules. Soil bioassays were conducted with U. bojeri seedlings grown in soils collected under the native tree species (U. bojeri and L. bojeriana) and two exotic tree species (Eucalyptus camaldulensis and Pinus patula) and in the same soils but previously cultured by L. bojeriana seedlings. This study clearly shows that (1) the introduction of exotic tree species induces significant changes in soil biotic and abiotic characteristics, (2) exotic-invaded soil significantly reduces the early growth and ectomycorrhization of U. bojeri seedlings and (3) L. bojeriana decreased these negative effects of the exotic tree species by facilitating ectomycorrhizal establishment and consequently improved the U. bojeri early growth. This study provides evidence that L. bojeriana can facilitate the ectomycorrhizal infection of U. bojeri and mitigates the negative effects of the introduction of exotic tree species on the early growth of the native tree species. From a practical point of view, the use of ectotrophic early-successional shrub species should be considered to improve forest resaturation after exotic invasion.
DOI:10.1111/nph.14990URL [本文引用: 1]
DOI:10.1007/s10530-011-9945-6URL [本文引用: 1]
Garlic mustard (Alliaria petiolata) is a Eurasian native that has become invasive in North America. The invasive success of A. petiolata has been partly attributed to its production of allelopathic compounds that can limit the growth of arbuscular mycorrhizal fungi (AMF). Although such effects are well known, specific effects on the richness and community composition of AMF associated with woody species have not been explored. We collected sugar maple (Acer saccharum) seedlings from eight natural forest sites in Ohio and Massachusetts, containing areas either invaded or uninvaded by A. petiolata. We measured AMF root colonization of seedlings, isolated DNA from the roots and performed PCR-TRFLP analysis to assess the richness and community composition of AMF. As expected, we found reduced AMF colonization in A. petiolata invaded patches. A. petiolata did not alter the detected TRF richness, but was associated with significant changes in the composition of AMF communities in half of the sites monitored in each region. Our results suggest that although AMF colonization was reduced in A. petiolata patches, many indigenous AMF communities include AMF that are tolerant to allelopathic effects of A. petiolata.
DOI:10.1371/journal.pone.0085857URLPMID:24392031 [本文引用: 1]
DOI:10.3732/ajb.1000177URL
Methods : We examine four mechanisms that potentially contribute to the success of invasive velvetgrass Holcus lanatus : direct competition, indirect competition mediated by mammalian herbivores, interference competition via allelopathy, and indirect competition mediated by changes in the soil community. Using a combination of field and greenhouse approaches, we focus on the effects of H. lanatus on a common species in California coastal prairies, Erigeron glaucus, where the invasion is most intense.Key results : We found that H. lanatus had the strongest effects on E. glaucus via direct competition, but it also influenced the soil community in ways that feed back to negatively influence E. glaucus and other native species after H. lanatus removal.Conclusions : This approach provided evidence for multiple mechanisms contributing to negative effects of invasive species, and it identified when particular strategies were most likely to be important. These mechanisms can be applied to eradication of H. lanatus and conservation of California coastal prairie systems, and they illustrate the utility of an integrated set of experiments for determining the potential mechanisms of invasive species' success.]]>
[本文引用: 1]
DOI:10.2307/2261425URL [本文引用: 1]
DOI:10.1038/ncomms1046URLPMID:20975705 [本文引用: 3]
Mycorrhizal fungi are a heterogeneous group of diverse fungal taxa, associated with the roots of over 90% of all plant species. Recently, state-of-the-art molecular and genetic tools, coupled to high-throughput sequencing and advanced microscopy, have led to the genome and transcriptome analysis of several symbionts. Signalling pathways between plants and fungi have now been described and the identification of several novel nutrient transporters has revealed some of the cellular processes that underlie symbiosis. Thus, the contributions of each partner in a mycorrhizal association are starting to be unravelled. This new knowledge is now available for use in agricultural practices.
DOI:10.1111/nph.14533URL [本文引用: 2]
DOI:10.1111/nph.14976URL [本文引用: 1]
DOI:10.1111/jec.v108.2URL [本文引用: 1]
DOI:10.3732/ajb.0800184URLPMID:21628149 [本文引用: 1]
Herbaceous plant species are important components of forest ecosystems, and their persistence in forests may be affected by invasive plant species that reduce mycorrhizal colonization of plant roots. I examined the effect of the invasive plant Alliaria petiolata on arbuscular mycorrhizal fungi (AMF) colonizing the roots of three forest plant species. AMF root colonization and community structure was examined from plants that were growing either in the absence or presence of Alliaria under natural forest conditions. AMF root colonization varied among the plant species but was not significantly affected by Alliaria. With molecular methods, approximately 12 different taxa of AMF could be distinguished among the root samples, and these taxa belonged to the genera Acaulospora and Glomus, with Glomus dominating AMF communities. There were significant differences between the community of AMF colonizing roots of Maianthemum racemosum and Trillium grandiflorum, but only AMF communities of Maianthemum roots were significantly affected by Alliaria. Indicator species analysis found that an Acaulospora species type was a significant indicator of Maianthemum plants grown in the absence of Alliaria. These results suggest invasive plants like Alliaria may selectively suppress AMF fungi, and this suppression can affect AMF communities colonizing the roots of some native plant species.
DOI:10.1126/science.290.5491.521URLPMID:11039934 [本文引用: 1]
Invading exotic plants are thought to succeed primarily because they have escaped their natural enemies, not because of novel interactions with their new neighbors. However, we find that Centaurea diffusa, a noxious weed in North America, has much stronger negative effects on grass species from North America than on closely related grass species from communities to which Centaurea is native. Centaurea's advantage against North American species appears to be due to differences in the effects of its root exudates and how these root exudates affect competition for resources. Our results may help to explain why some exotic species so successfully invade natural plant communities.
DOI:10.1890/07-0370.1URLPMID:18481529 [本文引用: 4]
Why some invasive plant species transmogrify from weak competitors at home to strong competitors abroad remains one of the most elusive questions in ecology. Some evidence suggests that disproportionately high densities of some invaders are due to the release of biochemicals that are novel, and therefore harmful, to naive organisms in their new range. So far, such evidence has been restricted to the direct phytotoxic effects of plants on other plants. Here we found that one of North America's most aggressive invaders of undisturbed forest understories, Alliaria petiolata (garlic mustard) and a plant that inhibits mycorrhizal fungal mutualists of North American native plants, has far stronger inhibitory effects on mycorrhizas in invaded North American soils than on mycorrhizas in European soils where A. petiolata is native. This antifungal effect appears to be due to specific flavonoid fractions in A. petiolata extracts. Furthermore, we found that suppression of North American mycorrhizal fungi by A. petiolata corresponds with severe inhibition of North American plant species that rely on these fungi, whereas congeneric European plants are weakly affected. These results indicate that phytochemicals, benign to resistant mycorrhizal symbionts in the home range, may be lethal to naive native mutualists in the introduced range and indirectly suppress the plants that rely on them.
DOI:10.1890/1540-9295(2004)002[0436:NWISAT]2.0.CO;2URL [本文引用: 1]
DOI:10.1007/s10530-011-9986-xURL
Garlic mustard's (Alliaria petiolata, Brassicaceae) invasive success is attributed in part to its release of allyl isothiocyanate (AITC) into the soil. AITC can disrupt beneficial arbuscular mycorrhizal fungi (AMF) associated with native plant roots, which limits their soil resource uptake. However, AITC and its precursor, sinigrin, have never been detected in garlic mustard-invaded forest soils. Here, we use high performance liquid chromatography (HPLC) and gas chromatography-mass spectrometry (GC-MS) to assess the concentration and bioactivity of these putative allelochemicals in paired forest plots uninvaded or invaded by garlic mustard. Our methods detected AITC and sinigrin only where garlic mustard was present and our recovery of AITC/sinigrin coincided with adult senescence. A bioassay of in situ fungal hyphae abundance revealed significantly reduced hyphal abundance in the presence of garlic mustard relative to uninvaded soils. Finally, the lowest concentration of AITC measured in the field (similar to 0.001 mM) is highly inhibitory to the spore germination of a forest AMF species, Glomus clarum. Together, our data provide the first direct evidence of garlic mustard-produced sinigrin and AITC in forest soils and demonstrate that even low levels of these chemicals have the potential to significantly suppress AMF growth and spore germination, strengthening their status as allelopathic novel weapons.
[本文引用: 1]
DOI:10.1111/ddi.2009.15.issue-1URL [本文引用: 2]
DOI:10.1111/ele.13220URLPMID:30779414 [本文引用: 1]
Much uncertainty remains about traits linked with successful invasion - the establishment and spread of non-resident species into existing communities. Using a 20-year experiment, where 50 non-resident (but mostly native) grassland plant species were sown into savannah plots, we ask how traits linked with invasion depend on invasion stage (establishment, spread), indicator of invasion success (occupancy, relative abundance), time, environmental conditions, propagule rain, and traits of invaders and invaded communities. Trait data for 164 taxa showed that invader occupancy was primarily associated with traits of invaders, traits of recipient communities, and invader-community interactions. Invader abundance was more strongly associated with community traits (e.g. proportion legume) and trait differences between invaders and the most similar resident species. Annuals and invaders with high-specific leaf area were only successful early in stand development, whereas invaders with conservative carbon capture strategies persisted long-term. Our results indicate that invasion is context-dependent and long-term experiments are required to comprehensively understand invasions.
DOI:10.1111/nph.v226.2URL [本文引用: 1]
DOI:10.1126/science.aau1361URLPMID:31604314 [本文引用: 1]
The mechanisms underlying interspecific variation in conspecific negative density dependence (CNDD) are poorly understood. Using a multilevel modeling approach, we combined long-term seedling demographic data from a subtropical forest plot with soil fungal community data by means of DNA sequencing to address the feedback of various guilds of soil fungi on the density dependence of trees. We show that mycorrhizal type mediates tree neighborhood interactions at the community level, and much of the interspecific variation in CNDD is explained by how tree species differ in their fungal density accumulation rates as they grow. Species with higher accumulation rates of pathogenic fungi suffered more from CNDD, whereas species with lower CNDD had higher accumulation rates of ectomycorrhizal fungi, suggesting that mutualistic and pathogenic fungi play important but opposing roles.
DOI:10.13287/j.1001-9332.201801.001URLPMID:29692043 [本文引用: 1]
As a kind of globally widely distributed soil microorganism, arbuscular mycorrhizal (AM) fungi form the symbiosis with the majority of land plants, which is important for plant nutrition and ecosystem functioning. Few vascular species are considered to be nonmycorrhizal, especially those within the families Amaranthaceae, Chenopodiaceae, Carophyllaceae and Brassicaceae. At present, the interactions between these non-host plants and AM fungi are few and scattered, lacking systematic summary. In this paper, the type of non-host plants, the reason of low mycotrophy, and the effect of AM network formed by neighbor plants on AM fungi colonization on the non-host, and the possible interaction between AM fungi and non-host, as well as the material exchange between plants and AM fungi and their possible ecological functions were reviewed in order to give some new ideas on the function of the nonmycorrhizal plants in the fragile ecosystems.
PMID:29692043 [本文引用: 1]
DOI:10.1038/s41467-018-05516-7URLPMID:30072764 [本文引用: 1]
Soil microbial communities play a crucial role in ecosystem functioning, but it is unknown how co-occurrence networks within these communities respond to disturbances such as climate extremes. This represents an important knowledge gap because changes in microbial networks could have implications for their functioning and vulnerability to future disturbances. Here, we show in grassland mesocosms that drought promotes destabilising properties in soil bacterial, but not fungal, co-occurrence networks, and that changes in bacterial communities link more strongly to soil functioning during recovery than do changes in fungal communities. Moreover, we reveal that drought has a prolonged effect on bacterial communities and their co-occurrence networks via changes in vegetation composition and resultant reductions in soil moisture. Our results provide new insight in the mechanisms through which drought alters soil microbial communities with potential long-term consequences, including future plant community composition and the ability of aboveground and belowground communities to withstand future disturbances.
DOI:10.1111/jec.2017.105.issue-4URL [本文引用: 3]
DOI:10.1002/ecy.1892URLPMID:28500779 [本文引用: 1]
Arbuscular mycorrhizal fungi (AMF) can increase plant fitness under certain environmental conditions. Among the mechanisms that may drive this mutualism, the most studied is provisioning of nutrients by AMF in exchange for carbon from plant hosts. However, AMF may also provide a suite of non-nutritional benefits to plants including improved water uptake, disease resistance, plant chemical defense, soil aggregation, and allelochemical transport and protection. Here, we use a meta-analysis of 93 studies to assess the relative effect of AMF on nutritional and non-nutritional factors that may influence plant fitness. We find that the positive effects of AMF on soil aggregation, water flow and disease resistance are equal to the effect of AMF on plant nitrogen and phosphorus uptake. However, AMF had no effect on the uptake of other nutrients, plant water content, allelopathic transport or production of chemical defense compounds. We suggest future research directions, including experimentally assessing the relative contribution on plant fitness of AMF interactions by untangling the independence of alternative benefits of AMF from an increase in nutrient uptake. This will lead to a more holistic view of the mycorrhizal-plant association and a more accurate picture of the net impact on the plant or plant community in question.
DOI:10.1111/nph.2017.215.issue-4URL [本文引用: 1]
DOI:10.1038/ismej.2016.109URLPMID:27482927 [本文引用: 1]
Recent field and laboratory experiments with perennial Boechera stricta and annual Arabidopsis thaliana suggest that the root microbiota influences flowering time. Here we examined in long-term time-course experiments the bacterial root microbiota of the arctic-alpine perennial Arabis alpina in natural and controlled environments by 16S rRNA gene profiling. We identified soil type and residence time of plants in soil as major determinants explaining up to 15% of root microbiota variation, whereas environmental conditions and host genotype explain maximally 11% of variation. When grown in the same soil, the root microbiota composition of perennial A. alpina is largely similar to those of its annual relatives A. thaliana and Cardamine hirsuta. Non-flowering wild-type A. alpina and flowering pep1 mutant plants assemble an essentially indistinguishable root microbiota, thereby uncoupling flowering time from plant residence time-dependent microbiota changes. This reveals the robustness of the root microbiota against the onset and perpetual flowering of A. alpina. Together with previous studies, this implies a model in which parts of the root microbiota modulate flowering time, whereas, after microbiota acquisition during vegetative growth, the established root-associated bacterial assemblage is structurally robust to perturbations caused by flowering and drastic changes in plant stature.
DOI:10.1111/ele.12166URL [本文引用: 1]
;Many exotic plant invaders pose a serious threat to native communities, but little is known about the dynamics of their impacts over time. In this study, we explored the impact of an invasive plant Heracleum mantegazzianum (giant hogweed) at 24 grassland sites invaded for different periods of time (from 11 to 48 years). Native species' richness and productivity were initially reduced by hogweed invasion but tended to recover after ~30 years of hogweed residence at the sites. Hogweed cover declined over the whole period assessed. A complementary common garden experiment suggested that the dynamics observed in the field were due to a negative plant-soil feedback; hogweed survival and biomass, and its competitive ability were lower when growing in soil inocula collected from earlier-invaded grasslands. Our results provide evidence that the initial dominance of an invasive plant species and its negative impact can later be reversed by stabilising processes.
DOI:10.1111/nph.2015.208.issue-1URL [本文引用: 1]
DOI:10.1556/ComEc.14.2013.1.8URL [本文引用: 2]
Recent research indicates that the soil microbial community, particularly arbuscular mycorrhizal fungi (AMF), can influence plant invasion in several ways. We tested if 1) invasive species are colonised by AMF to a lower degree than resident native species, and 2) AMF colonisation of native plants is lower in a community inhabited by an invasive species than in an uninvaded resident community. The two tests were run in semiarid temperate grasslands on grass (Poaceae) species, and the frequency and intensity of mycorrhizal colonisation, and the proportion of arbuscules and vesicles in plant roots have been measured. In the first test, grasses representing three classes of invasiveness were included: invasive species, resident species becoming abundant upon disturbance, and non-invasive native species. Each class contained one C3 and one C4 species. The AMF colonisation of the invasive Calamagrostis epigejos and Cynodon dactylon was consistently lower than that of the non-invasive native Chrysopogon gryllus and Bromus inermis, and contained fewer arbuscules than the post-disturbance dominant resident grasses Bothriochloa ischaemum and Brachypodium pinnatum. The C3 and C4 grasses behaved alike despite their displaced phenologies in these habitats. The second test compared AMF colonisation for sand grassland dominant grasses Festuca vaginata and Stipa borysthenica in stands invaded by either C. epigejos or C. dactylon, and in the uninvaded natural community. Resident grasses showed lower degree of AMF colonisation in the invaded stand compared to the uninvaded natural community with F. vaginata responding so to both invaders, while S. borysthenica responding to C. dactylon only. These results indicate that invasive grasses supposedly less reliant on AMF symbionts have the capacity of altering the soil mycorrhizal community in such a way that resident native species can establish a considerably reduced extent of the beneficial AMF associations, hence their growth, reproduction and ultimately abundance may decline. Accumulating evidence suggests that such indirect influences of invasive alien plants on resident native species mediated by AMF or other members of the soil biota is probably more the rule than the exception.
DOI:10.1073/pnas.1118650109URL [本文引用: 1]
The arbuscular mycorrhizal (AM) symbiosis, formed between the majority of land plants and ubiquitous soil fungi of the phylum Glomeromycota, is responsible for massive nutrient transfer and global carbon sequestration. AM fungi take up nutrients from the soil and exchange them against photosynthetically fixed carbon (C) from the host. Recent studies have demonstrated that reciprocal reward strategies by plant and fungal partners guarantee a "fair trade" of phosphorus against C between partners [Kiers ET, et al. (2011) Science 333: 880-882], but whether a similar reward mechanism also controls nitrogen (N) flux in the AM symbiosis is not known. Using mycorrhizal root organ cultures, we manipulated the C supply to the host and fungus and followed the uptake and transport of N sources in the AM symbiosis, the enzymatic activities of arginase and urease, and fungal gene expression in the extraradical and intraradical mycelium. We found that the C supply of the host plant triggers the uptake and transport of N in the symbiosis, and that the increase in N transport is orchestrated by changes in fungal gene expression. N transport in the symbiosis is stimulated only when the C is delivered by the host across the mycorrhizal interface, not when C is supplied directly to the fungal extraradical mycelium in the form of acetate. These findings support the importance of C flux from the root to the fungus as a key trigger for N uptake and transport and provide insight into the N transport regulation in the AM symbiosis.
URLPMID:19171910 [本文引用: 1]
URLPMID:26101408 [本文引用: 1]
DOI:10.1038/nature05719URLPMID:17460672 [本文引用: 2]
No species can maximize growth, reproduction and competitive ability across all environments, so the success of invasive species is habitat-dependent. Nutrient-rich habitats often experience more invasion than resource-poor habitats, a pattern consistent with traits generally associated with successful invaders (high growth rates, early reproduction and many offspring). However, invaders do colonize resource-poor environments, and the mechanisms that allow their success in these systems are poorly understood. Traits associated with resource conservation are widespread among species adapted to resource-poor environments, and invasive species may succeed in low-resource environments by employing resource conservation traits such as high resource-use efficiency (RUE; carbon assimilation per unit of resource). We investigated RUE in invasive and native species from three habitats in Hawaii where light, water or nutrient availability was limiting to plant growth. Here we show that across multiple growth forms and broad taxonomic diversity invasive species were generally more efficient than native species at using limiting resources on short timescales and were similarly efficient when RUE measures were integrated over leaf lifespans. Our data challenge the idea that native species generally outperform invasive species under conditions of low resource availability, and suggest that managing resource levels is not always an effective strategy for invasive species control.
DOI:10.1016/j.apsoil.2017.11.021URL [本文引用: 2]
DOI:10.1016/j.funeco.2018.12.004URL [本文引用: 1]
DOI:10.1093/aobpla/plaa058URLPMID:33408845
The Malaysian state of Sabah on the Island of Borneo has recently emerged as a global hotspot of nickel hyperaccumulator plants. This study focuses on the tissue-level distribution of nickel and other physiologically relevant elements in hyperaccumulator plants with distinct phylogenetical affinities. The roots, old stems, young stems and leaves of Flacourtia kinabaluensis (Salicaceae), Actephila alanbakeri (Phyllanthaceae), Psychotria sarmentosa (Rubiaceae) and young stems and leaves of Glochidion brunneum (Phyllanthaceae) were studied using nuclear microprobe (micro-PIXE and micro-BS) analysis. The tissue-level distribution of nickel found in these species has the same overall pattern as in most other hyperaccumulator plants studied previously, with substantial enrichment in the epidermal cells and in the phloem. This study also revealed enrichment of potassium in the spongy and palisade mesophyll of the studied species. Calcium, chlorine, manganese and cobalt were found to be enriched in the phloem and also concentrated in the epidermis and cortex of the studied species. Although hyperaccumulation ostensibly evolved numerous times independently, the basic mechanisms inferred from tissue elemental localization are convergent in these tropical woody species from Borneo Island.
[本文引用: 1]
[本文引用: 6]
[本文引用: 5]
Master degree dissertation, Shenyang Agricultural University, Shenyang.
[本文引用: 1]
[本文引用: 1]
URLPMID:24099088 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1086/522842URLPMID:18171166 [本文引用: 1]
Successful plant invasions are often attributed to increased plant size, reproduction, or release from natural enemies, but the generality and persistence of these patterns remains widely debated. Meta-analysis was used to quantitatively assess invasive plant performance and release from enemy damage and how these change with residence time and geographic distribution. Invasive plants were compared either in their introduced and home ranges or with native congeners in the introduced range. Invasive plants in the introduced range were generally larger, allocated more to reproduction, and had lower levels of herbivore damage compared with conspecifics in the home range; pathogen attack, however, varied widely. In congener comparisons, invasive and native plants did not differ in size or herbivory, but invaders did allocate less to reproduction and had lower levels of pathogen damage. Time since introduction was a significant nonlinear predictor of enemy release for both herbivores and pathogens, with initial release in recently arrived species and little to no release after 50 to 200 years. Geographic distribution was also a significant nonlinear predictor of enemy release. The observed nonlinear relationships are consistent with dynamic invasions and may define targets for eradication efforts if these patterns hold up for individual species.
[本文引用: 4]
DOI:10.1016/j.molp.2019.10.015URLPMID:31706032 [本文引用: 2]
Symbiotic microorganisms improve nutrient uptake by plants. To initiate mutualistic symbiosis with arbuscular mycorrhizal (AM) fungi, plants perceive Myc factors, including lipochitooligosaccharides (LCOs) and short-chain chitooligosaccharides (CO4/CO5), secreted by AM fungi. However, the molecular mechanism of Myc factor perception remains elusive. In this study, we identified a heteromer of LysM receptor-like kinases consisting of OsMYR1/OsLYK2 and OsCERK1 that mediates the perception of AM fungi in rice. CO4 directly binds to OsMYR1, promoting the dimerization and phosphorylation of this receptor complex. Compared with control plants, Osmyr1 and Oscerk1 mutant rice plants are less sensitive to Myc factors and show decreased AM colonization. We engineered transgenic rice by expressing chimeric receptors that respectively replaced the ectodomains of OsMYR1 and OsCERK1 with those from the homologous Nod factor receptors MtNFP and MtLYK3 of Medicago truncatula. Transgenic plants displayed increased calcium oscillations in response to Nod factors compared with control rice. Our study provides significant mechanistic insights into AM symbiotic signal perception in rice. Expression of chimeric Nod/Myc receptors achieves a potentially important step toward generating cereals that host nitrogen-fixing bacteria.
URLPMID:31621153 [本文引用: 1]
[本文引用: 3]
[本文引用: 3]
[本文引用: 3]
[本文引用: 1]
URLPMID:28596307 [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
DOI:10.1073/pnas.0906710107URLPMID:20133855 [本文引用: 1]
Symbioses may be important mechanisms of plant adaptation to their environment. We conducted a reciprocal inoculation experiment to test the hypothesis that soil fertility is a key driver of local adaptation in arbuscular mycorrhizal (AM) symbioses. Ecotypes of Andropogon gerardii from phosphorus-limited and nitrogen-limited grasslands were grown with all possible
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
DOI:10.1126/science.1208473URLPMID:21836016 [本文引用: 1]
Plants and their arbuscular mycorrhizal fungal symbionts interact in complex underground networks involving multiple partners. This increases the potential for exploitation and defection by individuals, raising the question of how partners maintain a fair, two-way transfer of resources. We manipulated cooperation in plants and fungal partners to show that plants can detect, discriminate, and reward the best fungal partners with more carbohydrates. In turn, their fungal partners enforce cooperation by increasing nutrient transfer only to those roots providing more carbohydrates. On the basis of these observations we conclude that, unlike many other mutualisms, the symbiont cannot be
[本文引用: 1]
URLPMID:25706625 [本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:23486251 [本文引用: 2]
DOI:10.1111/ele.13177URLPMID:30460736 [本文引用: 1]
While several studies have established a positive correlation between community diversity and invasion resistance, it is less clear how species interactions within resident communities shape this process. Here, we experimentally tested how antagonistic and facilitative pairwise interactions within resident model microbial communities predict invasion by the plant-pathogenic bacterium Ralstonia solanacearum. We found that facilitative resident community interactions promoted and antagonistic interactions suppressed invasions both in the lab and in the tomato plant rhizosphere. Crucially, pairwise interactions reliably explained observed invasion outcomes also in multispecies communities, and mechanistically, this was linked to direct inhibition of the invader by antagonistic communities (antibiosis), and to a lesser degree by resource competition between members of the resident community and the invader. Together, our findings suggest that the type and strength of pairwise interactions can reliably predict the outcome of invasions in more complex multispecies communities.
DOI:10.1111/1365-2664.12878URL [本文引用: 1]
DOI:10.1890/14-0871.1URLPMID:26240876 [本文引用: 1]
Soilborne pathogens can contribute to diversity maintenance in tree communities through the Janzen-Connell effect, whereby the pathogenic reduction of seedling performance attenuates with distance from conspecifics. By contrast, arbuscular mycorrhizal fungi (AMF) have been reported to promote seedling performance; however, it is unknown whether this is also distance dependent. Here, we investigate the distance dependence of seedling performance in the presence of both pathogens and AMF. In a subtropical forest in south China, we conducted a four-year field census of four species with relatively large phylogenetic distances and found no distance-dependent mortality for newly germinated seedlings. By experimentally separating the effects of AMF and pathogens on seedling performance of six subtropical tree species in a shade house, we found that soil pathogens significantly inhibited seedling survival and growth while AMF largely promoted seedling growth, and these effects were host specific and declined with increasing conspecific distance. Together, our field and experimental results suggest that AMF can neutralize the negative effect of pathogens and that the Janzen-Connell effect may play a less prominent role in explaining diversity of nondominant tree species than previously thought.
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s11104-019-03963-9URL [本文引用: 2]
DOI:10.1674/0003-0031-182.1.12URL [本文引用: 2]
[本文引用: 1]
DOI:10.1126/science.1143082URLPMID:17588930 [本文引用: 1]
DOI:10.1038/nature09622URLPMID:21209659 [本文引用: 1]
Arbuscular mycorrhiza (AM) is a root endosymbiosis between plants and glomeromycete fungi. It is the most widespread terrestrial plant symbiosis, improving plant uptake of water and mineral nutrients. Yet, despite its crucial role in land ecosystems, molecular mechanisms leading to its formation are just beginning to be unravelled. Recent evidence suggests that AM fungi produce diffusible symbiotic signals. Here we show that Glomus intraradices secretes symbiotic signals that are a mixture of sulphated and non-sulphated simple lipochitooligosaccharides (LCOs), which stimulate formation of AM in plant species of diverse families (Fabaceae, Asteraceae and Umbelliferae). In the legume Medicago truncatula these signals stimulate root growth and branching by the symbiotic DMI signalling pathway. These findings provide a better understanding of the evolution of signalling mechanisms involved in plant root endosymbioses and will greatly facilitate their molecular dissection. They also open the way to using these natural and very active molecules in agriculture.
DOI:10.1038/nature06556URLPMID:18322534 [本文引用: 2]
Mycorrhizal symbioses--the union of roots and soil fungi--are universal in terrestrial ecosystems and may have been fundamental to land colonization by plants. Boreal, temperate and montane forests all depend on ectomycorrhizae. Identification of the primary factors that regulate symbiotic development and metabolic activity will therefore open the door to understanding the role of ectomycorrhizae in plant development and physiology, allowing the full ecological significance of this symbiosis to be explored. Here we report the genome sequence of the ectomycorrhizal basidiomycete Laccaria bicolor (Fig. 1) and highlight gene sets involved in rhizosphere colonization and symbiosis. This 65-megabase genome assembly contains approximately 20,000 predicted protein-encoding genes and a very large number of transposons and repeated sequences. We detected unexpected genomic features, most notably a battery of effector-type small secreted proteins (SSPs) with unknown function, several of which are only expressed in symbiotic tissues. The most highly expressed SSP accumulates in the proliferating hyphae colonizing the host root. The ectomycorrhizae-specific SSPs probably have a decisive role in the establishment of the symbiosis. The unexpected observation that the genome of L. bicolor lacks carbohydrate-active enzymes involved in degradation of plant cell walls, but maintains the ability to degrade non-plant cell wall polysaccharides, reveals the dual saprotrophic and biotrophic lifestyle of the mycorrhizal fungus that enables it to grow within both soil and living plant roots. The predicted gene inventory of the L. bicolor genome, therefore, points to previously unknown mechanisms of symbiosis operating in biotrophic mycorrhizal fungi. The availability of this genome provides an unparalleled opportunity to develop a deeper understanding of the processes by which symbionts interact with plants within their ecosystem to perform vital functions in the carbon and nitrogen cycles that are fundamental to sustainable plant productivity.
DOI:10.1126/science.371.6534.1089URLPMID:33707246 [本文引用: 5]
DOI:10.1890/11-1247.1URLPMID:22611852 [本文引用: 3]
The disruption of mutualisms between plants and mycorrhizal fungi is a potentially powerful mechanism by which invasives can negatively impact native species, yet our understanding of this mechanism's role in exotic species invasion is still in its infancy. Here, we provide several lines of evidence indicating that invasive tamarisk (Tamarix sp.) negatively affects native cottonwoods (Populus fremontii) by disrupting their associations with arbuscular mycorrhizal (AM) and ectomycorrhizal (EM) fungi. At a field site in the early stages of tamarisk invasion, cottonwoods with tamarisk neighbors had reduced EM colonization and altered EM fungal community composition relative to cottonwoods with native neighbors, leading to reductions in EM propagule abundance in the soil beneath tamarisk. Similarly, AM colonization of cottonwoods was reduced with a tamarisk neighbor, but there were no significant changes in AM fungal spore communities or propagule abundance. Root colonization by nonmycorrhizal fungi, including potential pathogens, was higher in cottonwoods with tamarisk neighbors. A greenhouse experiment in which AM and EM inoculation and plant neighbor were manipulated in a fully factorial design showed that cottonwoods benefited from mycorrhizas, especially EM, in terms of shoot biomass when grown with a conspecific, but shoot biomass was similar to that of nonmycorrhizal controls when cottonwoods were grown with a tamarisk neighbor. These results are partially explained by a reduction in EM but not AM colonization of cottonwoods by a tamarisk neighbor. Tamarisk neighbors negatively affected cottonwood specific leaf area, but not chlorophyll content, in the field. To pinpoint a mechanism for these changes, we measured soil chemistry in the field and the growth response of an EM fungus (Hebeloma crustuliniforme) to salt-amended media in the laboratory. Tamarisk increased both NO3- concentrations and electrical conductivity 2.5-fold beneath neighboring cottonwoods in the field. Salt-amended media did not affect the growth of H. crustuliniforme. Our findings demonstrate that a nonnative species, even in the early stages of invasion, can negatively affect a native species by disrupting its mycorrhizal symbioses. Some of these changes in mycorrhizal fungal communities may remain as legacy effects of invasives, even after their removal, and should be considered in management and restoration efforts.
DOI:10.1111/ele.2006.9.issue-6URL [本文引用: 1]
DOI:10.1007/s11104-006-9091-6URL [本文引用: 1]
While several recent studies have described changes in microbial communities associated with exotic plant invasion, how arbuscular mycorrhizal fungi (AMF) communities respond to exotic plant invasion is not well known, despite the salient role of this group in plant interactions. Here, we use molecular methods (terminal restriction fragment length polymorphism analyses based on the large subunit of the rRNA gene) to examine AMF community structure in sites dominated by the invasive mycorrhizal forb, Centaurea maculosa Lam. (spotted knapweed), and in adjacent native grassland sites. Our results indicate that significant AMF community alteration occurs following C. maculosa invasion. Moreover, a significant reduction in the number of restriction fragment sizes was found for samples collected in C. maculosa-dominated areas, suggesting reduced AMF diversity. Extraradical hyphal lengths exhibited a significant, on average 24%, reduction in C. maculosa-versus native grass-dominated sites. As both AMF community composition and abundance were altered by C.maculosa invasion, these data are strongly suggestive of potential impacts on AMF-mediated ecosystem processes. Given that the composition of AMF communities has the potential to differentially influence different plant species, our results may have important implications for site restoration after weed invasion.]]>
DOI:10.1098/rsos.150322URLPMID:27019723
Millimetre-waves offer the possibility of wide bandwidth and consequently high data rate for wireless communications. For both uni- and dual-polarized systems, signals sent over a link may suffer severe degradation due to antenna misalignment. Orientation robustness may be enhanced by the use of mutual orthogonality in three dimensions. Multiple-input multiple-output polarization diversity offers a way of improving signal reception without the limitations associated with spatial diversity. Scattering effects often assist propagation through multipath. However, high path loss at millimetre-wave frequencies may limit any reception enhancement through scattering. We show that the inclusion of a third orthogonal dipole provides orientation robustness in this setting, as well as in a rich scattering environment, by means of a Rician fading channel model covering all orientations for a millimetre-wave, tri-orthogonal, half-wave dipole transmitter and receiver employing polarization diversity. Our simulation extends the analysis into three dimensions, fully exploiting individual sub-channel paths. In both the presence and absence of multipath effects, capacity is observed to be higher than that of a dual-polarized system over the majority of a field of view.
DOI:10.1038/nrmicro1987URLPMID:18794914 [本文引用: 1]
Arbuscular mycorrhiza (AM), a symbiosis between plants and members of an ancient phylum of fungi, the Glomeromycota, improves the supply of water and nutrients, such as phosphate and nitrogen, to the host plant. In return, up to 20% of plant-fixed carbon is transferred to the fungus. Nutrient transport occurs through symbiotic structures inside plant root cells known as arbuscules. AM development is accompanied by an exchange of signalling molecules between the symbionts. A novel class of plant hormones known as strigolactones are exuded by the plant roots. On the one hand, strigolactones stimulate fungal metabolism and branching. On the other hand, they also trigger seed germination of parasitic plants. Fungi release signalling molecules, in the form of 'Myc factors' that trigger symbiotic root responses. Plant genes required for AM development have been characterized. During evolution, the genetic programme for AM has been recruited for other plant root symbioses: functional adaptation of a plant receptor kinase that is essential for AM symbiosis paved the way for nitrogen-fixing bacteria to form intracellular symbioses with plant cells.
[本文引用: 1]
[本文引用: 1]
DOI:10.1038/s41396-019-0389-9URLPMID:30872806 [本文引用: 2]
Significant gaps in our understanding of how global change drivers interact to affect the resistance and functioning of microbial communities hinders our ability to model ecosystem responses and feedbacks to co-occurring global stressors. Here, we investigated the effects of extreme drought and exotic plants, two of the most significant threats to Mediterranean-type ecosystems, on soil microbial community composition and carbon metabolic genes within a four-year field rainfall manipulation experiment. We combined measurements of bulk microbial and soil properties with high-throughput microbial community analyses to elucidate microbial responses and microbial-mediated alterations to carbon cycling. While microbial responses to experimental droughts were weak, scant rainfall periods resulted in decreased microbial biomass and activity, and relative abundances of bacterial groups such as Proteobacteria, Verrucomicrobia, and Acidobacteria decreased concomitantly with increases in Actinobacteria, Chloroflexi, and Firmicutes abundance. Soils under exotic plants had increased temperatures, enhanced infiltration during rainfall events, and decreased water retention and labile carbon in comparison to soils under native plants. Higher peaks and more seasonally variable microbial activity were found under exotic plants and, like drought periods, the microbial community shifted towards osmotic stress life-strategies. Relationships found between microbial taxonomic groups and carbon metabolic genes support the interpretation that exotic plants change microbial carbon cycling by altering the soil microclimate and supplying easily decomposed high-quality litter. Soil microbial community responses to drought and exotic plants could potentially impact ecosystem C storage by producing a smaller, more vulnerable C pool of microbial biomass that is prone to increased pulses of heterotrophic respiration.
DOI:10.1111/nph.v228.2URL [本文引用: 1]
DOI:10.3732/ajb.91.2.285URLPMID:21653384 [本文引用: 1]
Garlic mustard (Alliaria petiolata, Brassicaceae) is an invasive, nonindigenous species currently invading the understory of North American woodlands where it is a serious threat to the native flora. Part of this success might be due to allelopathic interference by garlic mustard. Two congeneric species, the European Geum urbanum and the North American Geum laciniatum, were tested for allelopathic inhibition of germination by garlic mustard. Seeds were germinated either on substrate contaminated by garlic mustard or on substrate with contamination neutralized by activated carbon. Allelopathic effects of native European and invasive North American garlic mustard populations were also compared. Activated carbon increased germination by 14%, indicating that garlic mustard contaminated the substrate through root exudates. Activated carbon in turn counteracted this effect. The two test species differed in their sensitivity to allelopathic interference. North American G. laciniatum had a much stronger increase in germination when activated carbon was added to the substrate, independent of the origin of garlic mustard. In contrast, the European G. urbanum germinated better in substrate precultivated with North American garlic mustard, whereas activated carbon increased its germination only in substrate precultivated with European garlic mustard.
[本文引用: 1]
DOI:10.1111/j.1461-0248.2006.00950.xURLPMID:16913942 [本文引用: 1]
Invasion biologists often suggest that phenotypic plasticity plays an important role in successful plant invasions. Assuming that plasticity enhances ecological niche breadth and therefore confers a fitness advantage, recent studies have posed two main hypotheses: (1) invasive species are more plastic than non-invasive or native ones; (2) populations in the introduced range of an invasive species have evolved greater plasticity than populations in the native range. These two hypotheses largely reflect the disparate interests of ecologists and evolutionary biologists. Because these sciences are typically interested in different temporal and spatial scales, we describe what is required to assess phenotypic plasticity at different levels. We explore the inevitable tradeoffs of experiments conducted at the genotype vs. species level, outline components of experimental design required to identify plasticity at different levels, and review some examples from the recent literature. Moreover, we suggest that a successful invader may benefit from plasticity as either (1) a Jack-of-all-trades, better able to maintain fitness in unfavourable environments; (2) a Master-of-some, better able to increase fitness in favourable environments; or (3) a Jack-and-master that combines some level of both abilities. This new framework can be applied when testing both ecological or evolutionary oriented hypotheses, and therefore promises to bridge the gap between the two perspectives.
[本文引用: 1]
DOI:10.1111/j.1574-6941.2009.00740.xURLPMID:19656191
The objectives of this study were to determine whether the invasive plant Amaranthus viridis influenced soil microbial and chemical properties and to assess the consequences of these modifications on native plant growth. The experiment was conducted in Senegal at two sites: one invaded by A. viridis and the other covered by other plant species. Soil nutrient contents as well as microbial community density, diversity and functions were measured. Additionally, five sahelian Acacia species were grown in (1) soil disinfected or not collected from both sites, (2) uninvaded soil exposed to an A. viridis plant aqueous extract and (3) soil collected from invaded and uninvaded sites and inoculated or not with the arbuscular mycorrhizal (AM) fungus Glomus intraradices. The results showed that the invasion of A. viridis increased soil nutrient availability, bacterial abundance and microbial activities. In contrast, AM fungi and rhizobial development and the growth of Acacia species were severely reduced in A. viridis-invaded soil. Amaranthus viridis aqueous extract also exhibited an inhibitory effect on rhizobial growth, indicating an antibacterial activity of this plant extract. However, the inoculation of G. intraradices was highly beneficial to the growth and nodulation of Acacia species. These results highlight the role of AM symbiosis in the processes involved in plant coexistence and in ecosystem management programs that target preservation of native plant diversity.
DOI:10.1016/j.jenvman.2011.01.025URLPMID:21342746
Plant species (exotic invasive vs native non-invasive) colonization pattern and the relation with the soil nutrient availability and AM fungi abundance, was investigated. Soil samples were collected from two sites: one invaded by the exotic plant, Amaranthus viridis, and one uninvaded site for chemical and AM propagules density analyses. Additionally, we grew five Sahelian Acacia species in soil from the two sites, sterilized or not, to test the involvement of soil biota in the invasion process. While nutrient availability was significantly higher in soil samples from the invaded sites, a drastic reduction in AM fungal community density, was observed. Moreover, Acacia seedlings' growth was severely reduced in soils invaded by Amaranthus and this effect was similar to that of sterilized soil of both origins. The observed growth inhibition was accompanied by reduction of AM colonization and nodulation of the roots. Finally, the influence of soil chemistry and AM symbiosis on exotic plants' invasion processes is discussed.
[本文引用: 1]
DOI:10.1111/mec.14924URLPMID:30403423 [本文引用: 2]
Arbuscular mycorrhizal (AM) fungi are obligate plant symbionts that have important functions in most terrestrial ecosystems, but there remains an incomplete understanding of host-fungus specificity and the relationships between species and functional groups of plants and AM fungi. Here, we aimed to provide a comprehensive description of plant-AM fungal interactions in a biodiverse semi-natural grassland. We sampled all plant species in a 1,000-m(2) homogeneous plot of dry calcareous grassland in two seasons (summer and autumn) and identified root-colonizing AM fungi by SSU rDNA sequencing. In the network of 33 plant and 100 AM fungal species, we found a significant effect of both host plant species and host plant functional group on AM fungal richness and community composition. Comparison with network null models revealed a larger-than-random degree of partner selectivity among plants. Grasses harboured a larger number of AM fungal partners and were more generalist in partner selection, compared with forbs. More generalist partner association and lower specialization were apparent among obligately, compared with facultatively, mycorrhizal plant species and among locally more abundant plant species. This study provides the most complete data set of co-occurring plant and AM fungal taxa to date, showing that at this particular site, the interaction network is assembled non-randomly, with moderate selectivity in associations between plant species and functional groups and their fungal symbionts.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1038/s41586-019-1128-0URLPMID:31092941 [本文引用: 1]
The identity of the dominant root-associated microbial symbionts in a forest determines the ability of trees to access limiting nutrients from atmospheric or soil pools(1,2), sequester carbon(3,4) and withstand the effects of climate change(5,6). Characterizing the global distribution of these symbioses and identifying the factors that control this distribution are thus integral to understanding the present and future functioning of forest ecosystems. Here we generate a spatially explicit global map of the symbiotic status of forests, using a database of over 1.1 million forest inventory plots that collectively contain over 28,000 tree species. Our analyses indicate that climate variables-in particular, climatically controlled variation in the rate of decomposition-are the primary drivers of the global distribution of major symbioses. We estimate that ectomycorrhizal trees, which represent only 2% of all plant species(7), constitute approximately 60% of tree stems on Earth. Ectomycorrhizal symbiosis dominates forests in which seasonally cold and dry climates inhibit decomposition, and is the predominant form of symbiosis at high latitudes and elevation. By contrast, arbuscular mycorrhizal trees dominate in aseasonal, warm tropical forests, and occur with ectomycorrhizal trees in temperate biomes in which seasonally warm-and-wet climates enhance decomposition. Continental transitions between forests dominated by ectomycorrhizal or arbuscular mycorrhizal trees occur relatively abruptly along climate-driven decomposition gradients; these transitions are probably caused by positive feedback effects between plants and microorganisms. Symbiotic nitrogen fixers-which are insensitive to climatic controls on decomposition (compared with mycorrhizal fungi)-are most abundant in arid biomes with alkaline soils and high maximum temperatures. The climatically driven global symbiosis gradient that we document provides a spatially explicit quantitative understanding of microbial symbioses at the global scale, and demonstrates the critical role of microbial mutualisms in shaping the distribution of plant species.
DOI:10.1371/journal.pbio.0040140URLPMID:16623597 [本文引用: 4]
The impact of exotic species on native organisms is widely acknowledged, but poorly understood. Very few studies have empirically investigated how invading plants may alter delicate ecological interactions among resident species in the invaded range. We present novel evidence that antifungal phytochemistry of the invasive plant, Alliaria petiolata, a European invader of North American forests, suppresses native plant growth by disrupting mutualistic associations between native canopy tree seedlings and belowground arbuscular mycorrhizal fungi. Our results elucidate an indirect mechanism by which invasive plants can impact native flora, and may help explain how this plant successfully invades relatively undisturbed forest habitat.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1126/science.371.6534.1089URLPMID:33707246 [本文引用: 1]
DOI:10.1111/jec.2009.97.issue-4URL [本文引用: 1]
DOI:10.1093/aob/mcz119URLPMID:31304965 [本文引用: 1]
BACKGROUND AND AIMS: Arbuscular mycorrhizal (AM) symbiosis begins with molecular signal communication (MSC) between AM fungi and the roots of the host plant. We aimed to test the hypothesis that the transcriptional profiles of wheat roots can be changed significantly by AM symbiotic signals, without direct contact. METHODS: Non-mycorrhizal (NM) and MSC treatments involved burying filter membrane bags containing sterilized and un-sterilized inoculum of the AM fungus Rhizophagus irregularis, respectively. The bags physically separated roots and AM structures but allowed molecular signals to pass through. Extracted RNA from wheat roots was sequenced by high-throughput sequencing. RESULTS: Shoot total nitrogen and phosphorus content of wheat plants was decreased by the MSC treatment. A total of 2360 differentially expressed genes (DEGs), including 1888 up-regulated DEGs and 472 down-regulated DEGs, were found dominantly distributed on chromosomes 2A, 2B, 2D, 3B, 5B and 5D. The expression of 59 and 121 genes was greatly up- and down-regulated, respectively. Only a portion of DEGs could be enriched into known terms during gene ontology analysis, and were mostly annotated to 'catalytic activity', 'protein metabolic process' and 'membrane' in the molecular function, biological process and cellular component ontology categories, respectively. More than 120 genes that may be involved in key processes during AM symbiosis development were regulated at the pre-physical contact stages. CONCLUSIONS: The transcriptional profiles of wheat roots can be changed dramatically by MSC. Much of the information provided by our study is of great importance for understanding the mechanisms underlying the development of AM symbiosis.
URLPMID:27305777 [本文引用: 2]
Arbuscular mycorrhizal (AM) fungi play an important role in energy flow and nutrient cycling, besides their wide distribution in the cosystem. With a long co-evolution, AM fungi and host plant have formed a symbiotic relationship, and fungal lipid metabolism may be the key point to find the symbiotic mechanism in arbusculart mycorrhiza. Here, we reviewed the most recent progress on the interaction between AM fungal lipid metabolism and symbiotic signaling networks, especially the response of AM fungal lipid metabolism to symbiotic signals. Furthermore, we discussed the response of AM fungal lipid storage and release to symbiotic or non-symbiotic status, and the correlation between fungal lipid metabolism and nutrient transfer in mycorrhiza. In addition, we explored the feedback of the lipolysis process to molecular signals during the establishment of symbiosis, and the corresponding material conversion and energy metabolism besides the crosstalk of fungal lipid metabolism and signaling networks. This review will help understand symbiotic mechanism of arbuscular mycorrhiza fungi and further application in ecosystem.
PMID:27305777 [本文引用: 2]
DOI:10.1186/s12870-018-1500-5URLPMID:30463525 [本文引用: 2]
BACKGROUND: Below-ground linkage between plant and fungal communities is one of the major drivers of terrestrial ecosystem dynamics. However, we still have limited knowledge of how such plant-fungus associations vary in their community-scale properties depending on fungal functional groups and geographic locations. METHODS: By compiling a high-throughput sequencing dataset of root-associated fungi in eight forests along the Japanese Archipelago, we performed a comparative analysis of arbuscular mycorrhizal, ectomycorrhizal, and saprotrophic/endophytic associations across a latitudinal gradient from cool-temperate to subtropical regions. RESULTS: In most of the plant-fungus networks analyzed, host-symbiont associations were significantly specialized but lacked
DOI:10.1038/nature14910URLPMID:26287466 [本文引用: 1]
All around the globe, humans have greatly altered the abiotic and biotic environment with ever-increasing speed. One defining feature of the Anthropocene epoch is the erosion of biogeographical barriers by human-mediated dispersal of species into new regions, where they can naturalize and cause ecological, economic and social damage. So far, no comprehensive analysis of the global accumulation and exchange of alien plant species between continents has been performed, primarily because of a lack of data. Here we bridge this knowledge gap by using a unique global database on the occurrences of naturalized alien plant species in 481 mainland and 362 island regions. In total, 13,168 plant species, corresponding to 3.9% of the extant global vascular flora, or approximately the size of the native European flora, have become naturalized somewhere on the globe as a result of human activity. North America has accumulated the largest number of naturalized species, whereas the Pacific Islands show the fastest increase in species numbers with respect to their land area. Continents in the Northern Hemisphere have been the major donors of naturalized alien species to all other continents. Our results quantify for the first time the extent of plant naturalizations worldwide, and illustrate the urgent need for globally integrated efforts to control, manage and understand the spread of alien species.
DOI:10.1111/j.1461-0248.2009.01418.xURLPMID:20002494 [本文引用: 2]
A major aim in ecology is identifying determinants of invasiveness. We performed a meta-analysis of 117 field or experimental-garden studies that measured pair-wise trait differences of a total of 125 invasive and 196 non-invasive plant species in the invasive range of the invasive species. We tested whether invasiveness is associated with performance-related traits (physiology, leaf-area allocation, shoot allocation, growth rate, size and fitness), and whether such associations depend on type of study and on biogeographical or biological factors. Overall, invasive species had significantly higher values than non-invasive species for all six trait categories. More trait differences were significant for invasive vs. native comparisons than for invasive vs. non-invasive alien comparisons. Moreover, for comparisons between invasive species and native species that themselves are invasive elsewhere, no trait differences were significant. Differences in physiology and growth rate were larger in tropical regions than in temperate regions. Trait differences did not depend on whether the invasive alien species originates from Europe, nor did they depend on the test environment. We conclude that invasive alien species had higher values for those traits related to performance than non-invasive species. This suggests that it might become possible to predict future plant invasions from species traits.
DOI:10.1890/07-2144.1URLPMID:19323224 [本文引用: 1]
Belowground interactions between herbaceous native species and nonnative species is a poorly understood but emerging area of interest to invasive-species researchers. Positive feedback dynamics are commonly observed in many invaded systems and have been suspected in California grasslands, where native plants associate strongly with soil mutualists such as arbuscular mycorrhizal fungi. In response to disturbance, invading nonnative plants proliferate, and to the degree these species associate weakly with soil mutualists, we would expect mutualist efficacy to degrade over time. Degraded mutualist efficacy would negatively impact mutualist-dependent native species or their recruitment following a disturbance. We investigated the feedback dynamics of soil conditioned both with native and nonnative herbaceous communities of southern California grasslands to test this degraded mutualist hypothesis. Using a mesocosm approach, we inoculated each community with live soil originating from a remnant native grassland and varied the plant communities (i.e., native or nonnative) along a plant-species-richness gradient. After one year, we then used this conditioned soil for reciprocal feedback tests on a native and nonnative indicator species. We show that a native herbaceous forb (Gnaphalium californicum) grows best in soil conditioned by a diverse mix of other native species that includes G. californicum but is inhibited by soil conditioned by a diverse mix of nonnative species. We also show that an invasive, nonnative herbaceous forb (Carduus pycnocephalus) exhibits strong growth in soil lacking arbuscular mycorrhizal fungi and in soil conditioned by a diverse mix of nonnative species that include C. pycnocephalus, and that it is inhibited by the same soil that best promotes the native, G. californicum. Separate bioassays for mycorrhizal density show a reduction of arbuscular mycorrhizal fungi in the nonnative-conditioned soil relative to the native-conditioned soil, which suggests that nonnative species do not promote the growth of mycorrhizal fungi in the same way that native species do. The growth patterns resulting from the vegetative history of these distinct soil communities provide evidence of a biotic feedback mechanism that may account for the maintenance of persistent communities of nonnative (and often invasive) plants ubiquitous throughout California grasslands.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 3]
DOI:10.1007/s00572-011-0407-xURLPMID:21845465 [本文引用: 1]
Soil organisms play important roles in regulating ecosystem-level processes and the association of arbuscular mycorrhizal (AM) fungi with a plant species can be a central force shaping plant species' ecology. Understanding how mycorrhizal associations are affected by plant invasions may be a critical aspect of the conservation and restoration of native ecosystems. We examined the competitive ability of old world bluestem, a non-native grass (Caucasian bluestem [Bothriochloa bladhii]), and the influence of B. bladhii competition on AM root colonization of native warm-season prairie grasses (Andropogon gerardii or Schizachyrium scoparium), using a substitutive design greenhouse competition experiment. Competition by the non-native resulted in significantly reduced biomass production and AM colonization of the native grasses. To assess plant-soil feedbacks of B. bladhii and Bothriochloa ischaemum, we conducted a second greenhouse study which examined soil alterations indirectly by assessing biomass production and AM colonization of native warm-season grasses planted into soil collected beneath Bothriochloa spp. This study was conducted using soil from four replicate prairie sites throughout Kansas and Oklahoma, USA. Our results indicate that a major mechanism in plant growth suppression following invasion by Bothriochloa spp. is the alteration in soil microbial communities. Plant growth was tightly correlated with AM root colonization demonstrating that mycorrhizae play an important role in the invasion of these systems by Bothriochloa spp. and indicating that the restoration of native AM fungal communities may be a fundamental consideration for the successful establishment of native grasses into invaded sites.
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
DOI:10.1038/nature12798URLPMID:24256723 [本文引用: 1]
Returning native species to habitats degraded by biological invasions is a critical conservation goal. A leading hypothesis poses that exotic plant dominance is self-reinforced by impacts on ecosystem processes, leading to persistent stable states. Invaders have been documented to modify fire regimes, alter soil nutrients or shift microbial communities in ways that feed back to benefit themselves over competitors. However, few studies have followed invasions through time to ask whether ecosystem impacts and feedbacks persist. Here we return to woodland sites in Hawai'i Volcanoes National Park that were invaded by exotic C4 grasses in the 1960s, the ecosystem impacts of which were studied intensively in the 1990s. We show that positive feedbacks between exotic grasses and soil nitrogen cycling have broken down, but rather than facilitating native vegetation, the weakening feedbacks facilitate new exotic species. Data from the 1990s showed that exotic grasses increased nitrogen-mineralization rates by two- to fourfold, but were nitrogen-limited. Thus, the impacts of the invader created a positive feedback early in the invasion. We now show that annual net soil nitrogen mineralization has since dropped to pre-invasion levels. In addition, a seedling outplanting experiment that varied soil nitrogen and grass competition demonstrates that the changing impacts of grasses do not favour native species re-establishment. Instead, decreased nitrogen availability most benefits another aggressive invader, the nitrogen-fixing tree Morella faya. Long-term studies of invasions may reveal that ecosystem impacts and feedbacks shift over time, but that this may not benefit native species recovery.
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s00572-018-0866-4URLPMID:30220052 [本文引用: 1]
Invasive species often cause enormous economic and ecological damage, and this is especially true for invasive plants in the Asteraceae family. Arbuscular mycorrhizal fungi (AMF) play an important role in the successful invasion by exotic plant species because of their ability to promote growth and influence interspecific competition. However, few studies have evaluated the effects of invasive Asteraceae species on AMF diversity and how feedback mechanisms during competition with native species subsequently affect the accumulation of nutrient resources. Two exotic Asteraceae, Ambrosia artemisiifolia and Bidens pilosa, were monitored during competition with a native grass species, Setaria viridis, which is being replaced by these exotic species in natural areas around the study site. From these species continuously maintained in a field plot for 5 years, we collected the rhizosphere soil and cloned and identified soil AMF. Furthermore, AM fungal spores were isolated from rhizosphere soil of the two invasive species and used as inoculum in greenhouse experiments, to compare growth and nutrient accumulation during competition. The results indicate that although the AMF diversity in the rhizosphere soil of A. artemisiifolia and B. pilosa differed, the three most abundant species (Septoglomus viscosum, Septoglomus constrictum, Glomus perpusillum) were identical. The addition of AMF inoculum changed the competition between the plants, increasing the competitive ability of the invasives and decreasing that of the native. The results show a similar AMF community composition between A. artemisiifolia and B. pilosa, increased AMF root colonization of the invasive species during competition, AMF-enhanced N accumulation, and AMF-facilitated competitive growth of the invasive species.
[本文引用: 1]
[本文引用: 1]
[本文引用: 6]
[本文引用: 6]
[本文引用: 2]
DOI:10.1038/s41396-018-0171-4URLPMID:29899507 [本文引用: 2]
Cooperation is a prevalent phenomenon in nature and how it originates and maintains is a fundamental question in ecology. Many efforts have been made to understand cooperation between individuals in the same species, while the mechanisms enabling cooperation between different species are less understood. Here, we investigated under strict in vitro culture conditions if the exchange of carbon and phosphorus is pivotal to the cooperation between the arbuscular mycorrhizal fungus (AMF) Rhizophagus irregularis and the phosphate solubilizing bacterium (PSB) Rahnella aquatilis. We observed that fructose exuded by the AMF stimulated the expression of phosphatase genes in the bacterium as well as the rate of phosphatase release into the growth medium by regulating its protein secretory system. The phosphatase activity was subsequently increased, promoting the mineralization of organic phosphorus (i.e., phytate) into inorganic phosphorus, stimulating simultaneously the processes involved in phosphorus uptake by the AMF. Our results demonstrated for the first time that fructose not only is a carbon source, but also plays a role as a signal molecule triggering bacteria-mediated organic phosphorus mineralization processes. These results highlighted the molecular mechanisms by which the hyphal exudates play a role in maintaining the cooperation between AMF and bacteria.
[本文引用: 4]
DOI:10.1111/tpj.12723URLPMID:25399831 [本文引用: 1]
The establishment of symbiotic interactions between mycorrhizal fungi, rhizobial bacteria and their legume hosts involves a common symbiosis signalling pathway. This signalling pathway is activated by Nod factors produced by rhizobia and these are recognised by the Nod factor receptors NFR1/LYK3 and NFR5/NFP. Mycorrhizal fungi produce lipochitooligosaccharides (LCOs) similar to Nod factors, as well as short-chain chitin oligomers (CO4/5), implying commonalities in signalling during mycorrhizal and rhizobial associations. Here we show that NFR1/LYK3, but not NFR5/NFP, is required for the establishment of the mycorrhizal interaction in legumes. NFR1/LYK3 is necessary for the recognition of mycorrhizal fungi and the activation of the symbiosis signalling pathway leading to induction of calcium oscillations and gene expression. Chitin oligosaccharides also act as microbe associated molecular patterns that promote plant immunity via similar LysM receptor-like kinases. CERK1 in rice has the highest homology to NFR1 and we show that this gene is also necessary for the establishment of the mycorrhizal interaction as well as for resistance to the rice blast fungus. Our results demonstrate that NFR1/LYK3/OsCERK1 represents a common receptor for chitooligosaccharide-based signals produced by mycorrhizal fungi, rhizobial bacteria (in legumes) and fungal pathogens. It would appear that mycorrhizal recognition has been conserved in multiple receptors across plant species, but additional diversification in certain plant species has defined other signals that this class of receptors can perceive.
DOI:10.1016/j.scitotenv.2019.04.330URL [本文引用: 2]
[本文引用: 3]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
Plant sesquiterpenes induce hyphal branching in arbuscular mycorrhizal fungi
1
2005
... 植物与菌根真菌形成共生关系是一个复杂的信号转导过程.目前已经证实, 丛枝菌根的形成始于根系与AMF之间的信号交流(
Elevated enzyme activities in soils under the invasive nitrogen-fixing tree Falcataria moluccana.
1
2006
... 外来植物入侵后, 本地植物菌根真菌侵染率有时没有受到显著影响(
The impact of alien plant invasion on mycorrhizas in mountain fynbos vegetation
2
2001
... 早在21世纪初,
... 外来植物通过干扰本地植物菌根真菌获得竞争优势是一种重要的入侵机制.尽管如此, 相对于其他入侵机理, 我们对外来植物入侵与本地植物菌根真菌间关系的认识仍然十分匮乏(
入侵植物大花金鸡菊与伴生植物须苞石竹的互作效应及其对AM真菌的影响
2012
入侵植物大花金鸡菊与伴生植物须苞石竹的互作效应及其对AM真菌的影响
2012
What do we really know about alien plant invasion? A review of the invasion mechanism of one of the world’s worst weeds
1
2016
... Ecological mechanisms of alien plant invasion
| 生态机制 Mechanism of ecology | 举例 Example | 参考文献 Reference |
|---|---|---|
| 物种竞争 Species competition | 与农作物(如高粱(Sorghum bicolor)、玉米(Zea mays))相比, 入侵植物Parthenium hysterophorus具有快速早期生长的能力. Compared with crops (such as Sorghum bicolor and Zea mays), the invasive plant Parthenium hysterophorus can grow much more rapidly in their early stage. | |
| 比较夏威夷19对入侵植物和本地植物的资源利用效率, 发现入侵植物具有较高的碳同化率、光利用效率、瞬时氮利用效率和瞬时能量利用效率. Comparing the resource utilization efficiency for 19 paired invasive-native plants in Hawaii, invasive plants are observed to have higher carbon assimilation rate, light use efficiency, instantaneous nitrogen and energy use efficiency. | ||
| 比较47对入侵植物和非入侵植物对草食性动物的抵抗能力及养分含量发现, 入侵植物拥有更高的叶氮含量, 受到食草动物的损伤较小. Comparisons among 47 paired invasive and non-invasive plants for leaf herbivore-resistance and nutrient content show that invasive plants have higher leaf nitrogen content and less damage by herbivores. | ||
| 比对125种入侵植物和196种非入侵植物的生理性状发现, 入侵植物在生长率、养分分配及抗逆性更有优势. Comparing physiological traits of 125 invasive with plants 196 non-invasive plants show that invasive plants have more advantages in growth rate, nutrient allocation and stress resistance. | ||
| 外来植物Solanum carolinense具有较强的抗寒能力和无性繁殖能力. Exotic plant, Solanum carolinense, has strong ability for cold resistance and asexual reproduction. | ||
| 气候变暖及氮沉降的增加可以增强入侵植物Solidago canadensis叶片的资源获取能力. Climate warming and increased nitrogen deposition can enhance leaf resource acquisition of an invasive plant, Solidago canadensis. | ||
| 化感物质 Allelochemical | 入侵植物白羊草(Bothriochloa ischaemum)的渗滤液抑制本地植物Andropogon gerardii和Schizachyrium scoparium发芽及生长. The leachate of an invasive plant, Bothriochloa ischaemum, inhibits germination and growth of the native plants, Andropogon gerardii and Schizachyrium scoparium. | |
| 入侵植物水烛(Typha angustifolia)根系分泌的酚类物质抑制了本地植物Bolboschoenus fluviatilis的生长发育. Phenolic compounds secreted by roots of an invasive plant, Typha angustifolia, inhibit growth and development of a native plant, Bolboschoenus fluviatilis. | ||
| Solidago canadensis入侵分泌的化感物质抑制了本地植物莴苣(Lactuca sativa)种子萌发和幼苗生长. Allelochemicals secreted by the invasion of Solidago canadensis inhibit seed germination and seedling growth of a native plant, Lactuca sativa. | ||
| 外来植物Solanum carolinense含有龙葵碱及草酸盐结晶. The exotic plant Solanum carolinense contains solanine and oxalate crystals. | ||
| Parthenium hysterophorus可以分泌帕台单宁、咖啡酸、香草酸等化感物质帮助自身入侵. Parthenium hysterophorus can secrete parthenin, caffeic acid, vanillic acid and other allelochemicals to facilitate its invasion. | ||
| 土壤养分变化 Soil nutrient variation | 氮沉降会增强S. canadensis入侵初期分泌的化感物质对本地植物莴苣种子萌发和幼苗生长的抑制程度. Nitrogen deposition can enhance the inhibition of the allelochemicals secreted by S. canadensis at its initial invasive stage on seed germination and seedling growth of a native plant, Lactuca sativa. | |
| 分析94篇文章发现, 入侵植物增大了植物、土壤和土壤微生物中的碳和氮库大小. Analysis of 94 published papers show that invasive plants increase the size of carbon and nitrogen pools in plants, soils and soil microorganisms. | ||
| 与本地植物Artemisia californica、Salvia mellifera、Salvia apiana、锦鸡儿(Calochortus splendens)和Dichelostemma capitatum相比, 外来植物Centaurea stoebe、Hirschfeldia incana和Bromus madritensis可以通过加快凋落物分解改善土壤养分. Compared with native plants such as Artemisia californica, Salvia mellifera, Salvia apiana, Calochortus splendens and Dichelostemma capitatum, exotic plants, like Centaurea stoebe, Hirschfeldia incana and Bromus madritensis, can increase soil nutrient level by increasing litter decomposition. | ||
| 入侵植物Mikania micrantha分泌的化感物质绿原酸和β-石竹烯加快了土壤有机氮的矿化过程. Allelochemicals secreted by the invasive plant, Mikania micrantha, such as chlorogenic acid and β-caryophyllene accelerate mineralization of soil organic nitrogen. | ||
| 入侵植物通过凋落物改变了根际微生物土壤群落和养分循环, 形成植物-土壤正反馈调节. Invasive plants can form a positive plant-soil feedback via its litter decomposition which could change rhizosphere soil microbial communities and nutrient cycling. | ||
| 杂草Conyza canadensis入侵的土壤中有效氮和有机质含量增加. The content of available nitrogen and organic matter increases in the soil invaded by Conyza canadensis. | ||
| 与非入侵地土壤相比, Eupatorium catarium入侵后土壤氮含量提高4.32 mg·kg-1. Compared with non-invasive soils, soil nitrogen content increases by 4.32 mg·kg-1 after invasion by Eupatorium catarium. | ||
| Ambrosia trifida在入侵地的土壤-植物反馈高于原产地. Ambrosia trifida has a greater plant-soil feedback in invaded ranges than that in its native ranges. |
Restoring native forest ecosystems after exotic tree plantation in Madagascar: combination of the local ectotrophic species Leptolena bojeriana and Uapaca bojeri mitigates the negative influence of the exotic species Eucalyptus camaldulensis and Pinus patula
2012
Extreme rainfall affects assembly of the root-associated fungal community
1
2018
... 外来植物入侵不仅会影响本地植物菌根真菌, 也会影响其非菌根真菌(NMF), 包括致病真菌、寄生真菌、一些对植物可能无害的内生真菌以及在极端环境下可以帮助植物生存的有益内生菌(
Differences in arbuscular mycorrhizal fungal communities associated with sugar maple seedlings in and outside of invaded garlic mustard forest patches
1
2011
... 入侵植物除了影响本地植物菌根真菌的侵染率、根内和根外菌丝的结构与功能外, 还可能改变其菌根真菌的组成.有研究发现, 外来植物入侵会使本地植物原有的AMF被其他种类的AMF替代(
Arbuscular mycorrhizal fungal networks vary throughout the growing season and between successional stages
1
2013
... 同种植物常常能与多种菌根真菌共生, 即使是单个根尖也往往被多个特定的菌根真菌类群侵染(
Multiple mechanisms enable invasive species to suppress native species
2011
The fungal collaboration gradient dominates the root economics space in plants
1
2020
... 尽管入侵植物对本地植物菌根真菌影响的这些差异性被报道, 但是很少有研究者关注这些差异背后的原因.我们推测这可能至少与以下几个方面的原因有关: 1)本地植物和菌根真菌共生关系的强弱.例如, 有两个本地植物A和B, 如果A的菌根真菌侵染率远高于B (或者说A对菌根真菌的依赖性远超过B)(
Evolution of increased competitive ability in invasive nonindigenous plants, a hypothesis
1
1995
... 随着全球社会经济活动的日益紧密, 越来越多的植物被带入非原产地环境, 快速扩散成为入侵植物.全球变化背景下, 入侵植物已对当地生物多样性、社会经济以及人类健康造成重大威胁(
Mechanisms underlying beneficial plant-fungus interactions in mycorrhizal symbiosis
3
2010
... 除了这些被广泛关注的途径, 近年来还发现一类重要的间接途径, 即入侵植物能通过干扰本地物种的共生关系(如植物-传粉昆虫和根系-菌根真菌等)而抑制本地植物的生长(
... 菌根的根内菌丝延伸至根外形成根外菌丝, 根外菌丝直接参与土壤养分的吸收.外来入侵植物在对本地植物菌根真菌侵染率、真菌内部结构产生影响的同时, 也会对根外菌丝产生影响(
... 植物与菌根真菌形成共生关系是一个复杂的信号转导过程.目前已经证实, 丛枝菌根的形成始于根系与AMF之间的信号交流(
Arbuscular mycorrhiza-specific enzymes FatM and RAM2 fine-tune lipid biosynthesis to promote development of arbuscular mycorrhiza
2
2017
... 植物与菌根真菌形成共生关系是一个复杂的信号转导过程.目前已经证实, 丛枝菌根的形成始于根系与AMF之间的信号交流(
... ).入侵植物可以通过影响酶和这些转运蛋白干扰本地植物菌根真菌侵染进而影响本地植物生长.如
Evolutionary history of mycorrhizal symbioses and global host plant diversity
1
2018
... 除了这些被广泛关注的途径, 近年来还发现一类重要的间接途径, 即入侵植物能通过干扰本地物种的共生关系(如植物-传粉昆虫和根系-菌根真菌等)而抑制本地植物的生长(
Novel interactions between alien pathogens and native plants increase plant-pathogen network connectance and decrease specialization
1
2019
... 外来植物入侵不仅会影响本地植物菌根真菌, 也会影响其非菌根真菌(NMF), 包括致病真菌、寄生真菌、一些对植物可能无害的内生真菌以及在极端环境下可以帮助植物生存的有益内生菌(
Effects of Alliaria petiolata (garlic mustard; Brassicaceae) on mycorrhizal colonization and community structure in three herbaceous plants in a mixed deciduous forest
1
2008
... 入侵植物除了影响本地植物菌根真菌的侵染率、根内和根外菌丝的结构与功能外, 还可能改变其菌根真菌的组成.有研究发现, 外来植物入侵会使本地植物原有的AMF被其他种类的AMF替代(
Invasive plants versus their new and old neighbors: a mechanism for exotic invasion
1
2000
... 入侵植物可以通过多种生态机制影响本地植物的菌根真菌(
Novel weapons: invasive plant suppresses fungal mutualists in America but not in its native Europe
4
2008
... 随着全球社会经济活动的日益紧密, 越来越多的植物被带入非原产地环境, 快速扩散成为入侵植物.全球变化背景下, 入侵植物已对当地生物多样性、社会经济以及人类健康造成重大威胁(
... 除了这些被广泛关注的途径, 近年来还发现一类重要的间接途径, 即入侵植物能通过干扰本地物种的共生关系(如植物-传粉昆虫和根系-菌根真菌等)而抑制本地植物的生长(
... 早在21世纪初,
... 入侵植物可以通过多种生态机制影响本地植物的菌根真菌(
Novel weapons: invasive success and the evolution of increased competitive ability
1
2004
... 入侵植物可以通过多种生态机制影响本地植物的菌根真菌(
Low allelochemical concentrations detected in Garlic mustard- invaded forest soils inhibit fungal growth and AMF spore germination
2011
Effect of garlic mustard invasion on ectomycorrhizae in mature pine trees and pine seedlings//Groninger JW, Holzmueller EJ, Nielsen CK, Dey DC
1
2014
... 相对于AMF, 外生菌根真菌(EMF)对外来干扰的抵抗能力更强(
Reducing redundancy in invasion ecology by integrating hypotheses into a single theoretical framework
2
2009
... 随着全球社会经济活动的日益紧密, 越来越多的植物被带入非原产地环境, 快速扩散成为入侵植物.全球变化背景下, 入侵植物已对当地生物多样性、社会经济以及人类健康造成重大威胁(
... ).外来植物可以通过多种途径成功入侵当地生态系统(
Traits linked with species invasiveness and community invasibility vary with time, stage and indicator of invasion in a long-term grassland experiment
1
2019
... 已有研究证实外来植物的成功入侵受入侵时间的影响(
Arbuscular mycorrhizal fungi are a double-edged sword in plant invasion controlled by phosphorus concentration
1
2020
... 相对于AMF, 外生菌根真菌(EMF)对外来干扰的抵抗能力更强(
Differential soil fungus accumulation and density dependence of trees in a subtropical forest
1
2019
... 外来植物入侵不仅会影响本地植物菌根真菌, 也会影响其非菌根真菌(NMF), 包括致病真菌、寄生真菌、一些对植物可能无害的内生真菌以及在极端环境下可以帮助植物生存的有益内生菌(
AM真菌与非菌根植物的相互作用关系
1
2018
... 外来植物通过干扰本地植物菌根真菌获得竞争优势是一种重要的入侵机制.尽管如此, 相对于其他入侵机理, 我们对外来植物入侵与本地植物菌根真菌间关系的认识仍然十分匮乏(
AM真菌与非菌根植物的相互作用关系
1
2018
... 外来植物通过干扰本地植物菌根真菌获得竞争优势是一种重要的入侵机制.尽管如此, 相对于其他入侵机理, 我们对外来植物入侵与本地植物菌根真菌间关系的认识仍然十分匮乏(
Soil bacterial networks are less stable under drought than fungal networks
1
2018
... 同种植物常常能与多种菌根真菌共生, 即使是单个根尖也往往被多个特定的菌根真菌类群侵染(
Below-ground connections underlying above-ground food production: a framework for optimising ecological connections in the rhizosphere
3
2017
... 同种植物常常能与多种菌根真菌共生, 即使是单个根尖也往往被多个特定的菌根真菌类群侵染(
... 菌根真菌网络结构概念图(改自
... A conceptual diagram of the mycorrhizal fungal network structure (adapted from
Beyond nutrients, a meta-analysis of the diverse effects of arbuscular mycorrhizal fungi on plants and soils
1
2017
... 除了这些被广泛关注的途径, 近年来还发现一类重要的间接途径, 即入侵植物能通过干扰本地物种的共生关系(如植物-传粉昆虫和根系-菌根真菌等)而抑制本地植物的生长(
The emerging science of linked plant-fungal invasions
1
2017
... 除了这些被广泛关注的途径, 近年来还发现一类重要的间接途径, 即入侵植物能通过干扰本地物种的共生关系(如植物-传粉昆虫和根系-菌根真菌等)而抑制本地植物的生长(
Root microbiota dynamics of perennial Arabis alpina are dependent on soil residence time but independent of flowering time
1
2017
... 外来植物通过干扰本地植物菌根真菌获得竞争优势是一种重要的入侵机制.尽管如此, 相对于其他入侵机理, 我们对外来植物入侵与本地植物菌根真菌间关系的认识仍然十分匮乏(
The impact of an invasive plant changes over time
1
2013
... 已有研究证实外来植物的成功入侵受入侵时间的影响(
Linking root traits to nutrient foraging in arbuscular mycorrhizal trees in a temperate forest
1
2015
... 尽管入侵植物对本地植物菌根真菌影响的这些差异性被报道, 但是很少有研究者关注这些差异背后的原因.我们推测这可能至少与以下几个方面的原因有关: 1)本地植物和菌根真菌共生关系的强弱.例如, 有两个本地植物A和B, 如果A的菌根真菌侵染率远高于B (或者说A对菌根真菌的依赖性远超过B)(
Arbuscular mycorrhizal colonisation of roots of grass species differing in invasiveness
2
2013
... 早在21世纪初,
... 外来植物通过干扰本地植物菌根真菌获得竞争优势是一种重要的入侵机制.尽管如此, 相对于其他入侵机理, 我们对外来植物入侵与本地植物菌根真菌间关系的认识仍然十分匮乏(
Carbon availability triggers fungal nitrogen uptake and transport in arbuscular mycorrhizal symbiosis
1
2012
... 入侵植物可以通过多种生态机制影响本地植物的菌根真菌(
Evolutionary tradeoffs for nitrogen allocation to photosynthesis versus cell walls in an invasive plant
1
2009
... 随着全球社会经济活动的日益紧密, 越来越多的植物被带入非原产地环境, 快速扩散成为入侵植物.全球变化背景下, 入侵植物已对当地生物多样性、社会经济以及人类健康造成重大威胁(
Taking the long view on the ecological effects of plant invasions
1
2015
... 已有研究证实外来植物的成功入侵受入侵时间的影响(
Resource-use efficiency and plant invasion in low-resource systems
2
2007
... 入侵植物可以通过多种生态机制影响本地植物的菌根真菌(
... Ecological mechanisms of alien plant invasion
| 生态机制 Mechanism of ecology | 举例 Example | 参考文献 Reference |
|---|---|---|
| 物种竞争 Species competition | 与农作物(如高粱(Sorghum bicolor)、玉米(Zea mays))相比, 入侵植物Parthenium hysterophorus具有快速早期生长的能力. Compared with crops (such as Sorghum bicolor and Zea mays), the invasive plant Parthenium hysterophorus can grow much more rapidly in their early stage. | |
| 比较夏威夷19对入侵植物和本地植物的资源利用效率, 发现入侵植物具有较高的碳同化率、光利用效率、瞬时氮利用效率和瞬时能量利用效率. Comparing the resource utilization efficiency for 19 paired invasive-native plants in Hawaii, invasive plants are observed to have higher carbon assimilation rate, light use efficiency, instantaneous nitrogen and energy use efficiency. | ||
| 比较47对入侵植物和非入侵植物对草食性动物的抵抗能力及养分含量发现, 入侵植物拥有更高的叶氮含量, 受到食草动物的损伤较小. Comparisons among 47 paired invasive and non-invasive plants for leaf herbivore-resistance and nutrient content show that invasive plants have higher leaf nitrogen content and less damage by herbivores. | ||
| 比对125种入侵植物和196种非入侵植物的生理性状发现, 入侵植物在生长率、养分分配及抗逆性更有优势. Comparing physiological traits of 125 invasive with plants 196 non-invasive plants show that invasive plants have more advantages in growth rate, nutrient allocation and stress resistance. | ||
| 外来植物Solanum carolinense具有较强的抗寒能力和无性繁殖能力. Exotic plant, Solanum carolinense, has strong ability for cold resistance and asexual reproduction. | ||
| 气候变暖及氮沉降的增加可以增强入侵植物Solidago canadensis叶片的资源获取能力. Climate warming and increased nitrogen deposition can enhance leaf resource acquisition of an invasive plant, Solidago canadensis. | ||
| 化感物质 Allelochemical | 入侵植物白羊草(Bothriochloa ischaemum)的渗滤液抑制本地植物Andropogon gerardii和Schizachyrium scoparium发芽及生长. The leachate of an invasive plant, Bothriochloa ischaemum, inhibits germination and growth of the native plants, Andropogon gerardii and Schizachyrium scoparium. | |
| 入侵植物水烛(Typha angustifolia)根系分泌的酚类物质抑制了本地植物Bolboschoenus fluviatilis的生长发育. Phenolic compounds secreted by roots of an invasive plant, Typha angustifolia, inhibit growth and development of a native plant, Bolboschoenus fluviatilis. | ||
| Solidago canadensis入侵分泌的化感物质抑制了本地植物莴苣(Lactuca sativa)种子萌发和幼苗生长. Allelochemicals secreted by the invasion of Solidago canadensis inhibit seed germination and seedling growth of a native plant, Lactuca sativa. | ||
| 外来植物Solanum carolinense含有龙葵碱及草酸盐结晶. The exotic plant Solanum carolinense contains solanine and oxalate crystals. | ||
| Parthenium hysterophorus可以分泌帕台单宁、咖啡酸、香草酸等化感物质帮助自身入侵. Parthenium hysterophorus can secrete parthenin, caffeic acid, vanillic acid and other allelochemicals to facilitate its invasion. | ||
| 土壤养分变化 Soil nutrient variation | 氮沉降会增强S. canadensis入侵初期分泌的化感物质对本地植物莴苣种子萌发和幼苗生长的抑制程度. Nitrogen deposition can enhance the inhibition of the allelochemicals secreted by S. canadensis at its initial invasive stage on seed germination and seedling growth of a native plant, Lactuca sativa. | |
| 分析94篇文章发现, 入侵植物增大了植物、土壤和土壤微生物中的碳和氮库大小. Analysis of 94 published papers show that invasive plants increase the size of carbon and nitrogen pools in plants, soils and soil microorganisms. | ||
| 与本地植物Artemisia californica、Salvia mellifera、Salvia apiana、锦鸡儿(Calochortus splendens)和Dichelostemma capitatum相比, 外来植物Centaurea stoebe、Hirschfeldia incana和Bromus madritensis可以通过加快凋落物分解改善土壤养分. Compared with native plants such as Artemisia californica, Salvia mellifera, Salvia apiana, Calochortus splendens and Dichelostemma capitatum, exotic plants, like Centaurea stoebe, Hirschfeldia incana and Bromus madritensis, can increase soil nutrient level by increasing litter decomposition. | ||
| 入侵植物Mikania micrantha分泌的化感物质绿原酸和β-石竹烯加快了土壤有机氮的矿化过程. Allelochemicals secreted by the invasive plant, Mikania micrantha, such as chlorogenic acid and β-caryophyllene accelerate mineralization of soil organic nitrogen. | ||
| 入侵植物通过凋落物改变了根际微生物土壤群落和养分循环, 形成植物-土壤正反馈调节. Invasive plants can form a positive plant-soil feedback via its litter decomposition which could change rhizosphere soil microbial communities and nutrient cycling. | ||
| 杂草Conyza canadensis入侵的土壤中有效氮和有机质含量增加. The content of available nitrogen and organic matter increases in the soil invaded by Conyza canadensis. | ||
| 与非入侵地土壤相比, Eupatorium catarium入侵后土壤氮含量提高4.32 mg·kg-1. Compared with non-invasive soils, soil nitrogen content increases by 4.32 mg·kg-1 after invasion by Eupatorium catarium. | ||
| Ambrosia trifida在入侵地的土壤-植物反馈高于原产地. Ambrosia trifida has a greater plant-soil feedback in invaded ranges than that in its native ranges. |
The invasive plant Impatiens glandulifera affects soil fungal diversity and the bacterial community in forests
2
2018
... 入侵植物除了影响本地植物菌根真菌的侵染率、根内和根外菌丝的结构与功能外, 还可能改变其菌根真菌的组成.有研究发现, 外来植物入侵会使本地植物原有的AMF被其他种类的AMF替代(
... 外来植物通过干扰本地植物菌根真菌获得竞争优势是一种重要的入侵机制.尽管如此, 相对于其他入侵机理, 我们对外来植物入侵与本地植物菌根真菌间关系的认识仍然十分匮乏(
The annual invasive plant Impatiens glandulifera reduces hyphal biomass of soil fungi in deciduous forests
1
2019
... 菌根的根内菌丝延伸至根外形成根外菌丝, 根外菌丝直接参与土壤养分的吸收.外来入侵植物在对本地植物菌根真菌侵染率、真菌内部结构产生影响的同时, 也会对根外菌丝产生影响(
Native plants fare better against an introduced competitor with native microbes and lower nitrogen availability
2017
Buffelgrass invasion and glyphosate effects on desert soil microbiome communities
2020
Experimental evidence that invasive grasses use allelopathic biochemicals as a potential mechanism for invasion: chemical warfare in nature
1
2014
... Ecological mechanisms of alien plant invasion
| 生态机制 Mechanism of ecology | 举例 Example | 参考文献 Reference |
|---|---|---|
| 物种竞争 Species competition | 与农作物(如高粱(Sorghum bicolor)、玉米(Zea mays))相比, 入侵植物Parthenium hysterophorus具有快速早期生长的能力. Compared with crops (such as Sorghum bicolor and Zea mays), the invasive plant Parthenium hysterophorus can grow much more rapidly in their early stage. | |
| 比较夏威夷19对入侵植物和本地植物的资源利用效率, 发现入侵植物具有较高的碳同化率、光利用效率、瞬时氮利用效率和瞬时能量利用效率. Comparing the resource utilization efficiency for 19 paired invasive-native plants in Hawaii, invasive plants are observed to have higher carbon assimilation rate, light use efficiency, instantaneous nitrogen and energy use efficiency. | ||
| 比较47对入侵植物和非入侵植物对草食性动物的抵抗能力及养分含量发现, 入侵植物拥有更高的叶氮含量, 受到食草动物的损伤较小. Comparisons among 47 paired invasive and non-invasive plants for leaf herbivore-resistance and nutrient content show that invasive plants have higher leaf nitrogen content and less damage by herbivores. | ||
| 比对125种入侵植物和196种非入侵植物的生理性状发现, 入侵植物在生长率、养分分配及抗逆性更有优势. Comparing physiological traits of 125 invasive with plants 196 non-invasive plants show that invasive plants have more advantages in growth rate, nutrient allocation and stress resistance. | ||
| 外来植物Solanum carolinense具有较强的抗寒能力和无性繁殖能力. Exotic plant, Solanum carolinense, has strong ability for cold resistance and asexual reproduction. | ||
| 气候变暖及氮沉降的增加可以增强入侵植物Solidago canadensis叶片的资源获取能力. Climate warming and increased nitrogen deposition can enhance leaf resource acquisition of an invasive plant, Solidago canadensis. | ||
| 化感物质 Allelochemical | 入侵植物白羊草(Bothriochloa ischaemum)的渗滤液抑制本地植物Andropogon gerardii和Schizachyrium scoparium发芽及生长. The leachate of an invasive plant, Bothriochloa ischaemum, inhibits germination and growth of the native plants, Andropogon gerardii and Schizachyrium scoparium. | |
| 入侵植物水烛(Typha angustifolia)根系分泌的酚类物质抑制了本地植物Bolboschoenus fluviatilis的生长发育. Phenolic compounds secreted by roots of an invasive plant, Typha angustifolia, inhibit growth and development of a native plant, Bolboschoenus fluviatilis. | ||
| Solidago canadensis入侵分泌的化感物质抑制了本地植物莴苣(Lactuca sativa)种子萌发和幼苗生长. Allelochemicals secreted by the invasion of Solidago canadensis inhibit seed germination and seedling growth of a native plant, Lactuca sativa. | ||
| 外来植物Solanum carolinense含有龙葵碱及草酸盐结晶. The exotic plant Solanum carolinense contains solanine and oxalate crystals. | ||
| Parthenium hysterophorus可以分泌帕台单宁、咖啡酸、香草酸等化感物质帮助自身入侵. Parthenium hysterophorus can secrete parthenin, caffeic acid, vanillic acid and other allelochemicals to facilitate its invasion. | ||
| 土壤养分变化 Soil nutrient variation | 氮沉降会增强S. canadensis入侵初期分泌的化感物质对本地植物莴苣种子萌发和幼苗生长的抑制程度. Nitrogen deposition can enhance the inhibition of the allelochemicals secreted by S. canadensis at its initial invasive stage on seed germination and seedling growth of a native plant, Lactuca sativa. | |
| 分析94篇文章发现, 入侵植物增大了植物、土壤和土壤微生物中的碳和氮库大小. Analysis of 94 published papers show that invasive plants increase the size of carbon and nitrogen pools in plants, soils and soil microorganisms. | ||
| 与本地植物Artemisia californica、Salvia mellifera、Salvia apiana、锦鸡儿(Calochortus splendens)和Dichelostemma capitatum相比, 外来植物Centaurea stoebe、Hirschfeldia incana和Bromus madritensis可以通过加快凋落物分解改善土壤养分. Compared with native plants such as Artemisia californica, Salvia mellifera, Salvia apiana, Calochortus splendens and Dichelostemma capitatum, exotic plants, like Centaurea stoebe, Hirschfeldia incana and Bromus madritensis, can increase soil nutrient level by increasing litter decomposition. | ||
| 入侵植物Mikania micrantha分泌的化感物质绿原酸和β-石竹烯加快了土壤有机氮的矿化过程. Allelochemicals secreted by the invasive plant, Mikania micrantha, such as chlorogenic acid and β-caryophyllene accelerate mineralization of soil organic nitrogen. | ||
| 入侵植物通过凋落物改变了根际微生物土壤群落和养分循环, 形成植物-土壤正反馈调节. Invasive plants can form a positive plant-soil feedback via its litter decomposition which could change rhizosphere soil microbial communities and nutrient cycling. | ||
| 杂草Conyza canadensis入侵的土壤中有效氮和有机质含量增加. The content of available nitrogen and organic matter increases in the soil invaded by Conyza canadensis. | ||
| 与非入侵地土壤相比, Eupatorium catarium入侵后土壤氮含量提高4.32 mg·kg-1. Compared with non-invasive soils, soil nitrogen content increases by 4.32 mg·kg-1 after invasion by Eupatorium catarium. | ||
| Ambrosia trifida在入侵地的土壤-植物反馈高于原产地. Ambrosia trifida has a greater plant-soil feedback in invaded ranges than that in its native ranges. |
Mycorrhizae, invasions, and the temporal dynamics of mutualism disruption
6
2017a
... 相对于AMF, 外生菌根真菌(EMF)对外来干扰的抵抗能力更强(
...
... Conceptual illustration showing how invasive plants affect the symbiotic mycorrhizal fungi in native plant roots (adapted from
... 入侵植物可以通过多种生态机制影响本地植物的菌根真菌(
... ).实际上, 本地植物菌根真菌的变化可能是多种机制共同作用的结果(
... 已有研究证实外来植物的成功入侵受入侵时间的影响(
Direct and indirect effects of allelopathy in the soil legacy of an exotic plant invasion
2012
Do impacts of an invasive nitrogen-fixing shrub on Douglas-fir and its ectomycorrhizal mutualism change over time following invasion?
5
2017b
... 入侵植物可以通过多种生态机制影响本地植物的菌根真菌(
... ,
... 已有研究证实外来植物的成功入侵受入侵时间的影响(
... )、土壤养分(
... 外来植物通过干扰本地植物菌根真菌获得竞争优势是一种重要的入侵机制.尽管如此, 相对于其他入侵机理, 我们对外来植物入侵与本地植物菌根真菌间关系的认识仍然十分匮乏(
不同光环境下入侵植物瘤突苍耳对大豆菌根真菌和根瘤菌的影响
1
2020
... 以往研究大多只关注具有单共生关系的本地植物(只受菌根真菌侵染), 而对具有双共生关系的本地植物(根瘤菌和AMF共同侵染, 如豆科植物)的研究很少.我们团队的研究发现, 入侵植物X. strumarium抑制了本地植物苍耳(单共生关系)的AMF侵染率(
不同光环境下入侵植物瘤突苍耳对大豆菌根真菌和根瘤菌的影响
1
2020
... 以往研究大多只关注具有单共生关系的本地植物(只受菌根真菌侵染), 而对具有双共生关系的本地植物(根瘤菌和AMF共同侵染, 如豆科植物)的研究很少.我们团队的研究发现, 入侵植物X. strumarium抑制了本地植物苍耳(单共生关系)的AMF侵染率(
Cell and developmental biology of arbuscular mycorrhiza symbiosis
1
2013
... 植物与菌根真菌形成共生关系是一个复杂的信号转导过程.目前已经证实, 丛枝菌根的形成始于根系与AMF之间的信号交流(
外来入侵植物的根系觅养行为研究进展
1
2020
... 入侵植物可以通过多种生态机制影响本地植物的菌根真菌(
外来入侵植物的根系觅养行为研究进展
1
2020
... 入侵植物可以通过多种生态机制影响本地植物的菌根真菌(
Are invaders moving targets? The generality and persistence of advantages in size, reproduction, and enemy release in invasive plant species with time since introduction
1
2007
... 已有研究证实外来植物的成功入侵受入侵时间的影响(
Arbuscular mycorrhizal assemblages in native plant roots change in the presence of invasive exotic grasses
4
2006
... 入侵植物除了影响本地植物菌根真菌的侵染率、根内和根外菌丝的结构与功能外, 还可能改变其菌根真菌的组成.有研究发现, 外来植物入侵会使本地植物原有的AMF被其他种类的AMF替代(
... 外来植物入侵不仅会影响本地植物菌根真菌, 也会影响其非菌根真菌(NMF), 包括致病真菌、寄生真菌、一些对植物可能无害的内生真菌以及在极端环境下可以帮助植物生存的有益内生菌(
...
... 外来植物通过干扰本地植物菌根真菌获得竞争优势是一种重要的入侵机制.尽管如此, 相对于其他入侵机理, 我们对外来植物入侵与本地植物菌根真菌间关系的认识仍然十分匮乏(
A LysM receptor heteromer mediates perception of arbuscular mycorrhizal symbiotic signal in rice
2
2019
... 植物与菌根真菌形成共生关系是一个复杂的信号转导过程.目前已经证实, 丛枝菌根的形成始于根系与AMF之间的信号交流(
... 外来植物通过干扰本地植物菌根真菌获得竞争优势是一种重要的入侵机制.尽管如此, 相对于其他入侵机理, 我们对外来植物入侵与本地植物菌根真菌间关系的认识仍然十分匮乏(
Angiosperm symbioses with non-mycorrhizal fungal partners enhance N acquisition from ancient organic matter in a warming maritime Antarctic
1
2019
... 外来植物入侵不仅会影响本地植物菌根真菌, 也会影响其非菌根真菌(NMF), 包括致病真菌、寄生真菌、一些对植物可能无害的内生真菌以及在极端环境下可以帮助植物生存的有益内生菌(
氮沉降对不同入侵程度加拿大一枝黄花化感作用的影响
3
2020
... Ecological mechanisms of alien plant invasion
| 生态机制 Mechanism of ecology | 举例 Example | 参考文献 Reference |
|---|---|---|
| 物种竞争 Species competition | 与农作物(如高粱(Sorghum bicolor)、玉米(Zea mays))相比, 入侵植物Parthenium hysterophorus具有快速早期生长的能力. Compared with crops (such as Sorghum bicolor and Zea mays), the invasive plant Parthenium hysterophorus can grow much more rapidly in their early stage. | |
| 比较夏威夷19对入侵植物和本地植物的资源利用效率, 发现入侵植物具有较高的碳同化率、光利用效率、瞬时氮利用效率和瞬时能量利用效率. Comparing the resource utilization efficiency for 19 paired invasive-native plants in Hawaii, invasive plants are observed to have higher carbon assimilation rate, light use efficiency, instantaneous nitrogen and energy use efficiency. | ||
| 比较47对入侵植物和非入侵植物对草食性动物的抵抗能力及养分含量发现, 入侵植物拥有更高的叶氮含量, 受到食草动物的损伤较小. Comparisons among 47 paired invasive and non-invasive plants for leaf herbivore-resistance and nutrient content show that invasive plants have higher leaf nitrogen content and less damage by herbivores. | ||
| 比对125种入侵植物和196种非入侵植物的生理性状发现, 入侵植物在生长率、养分分配及抗逆性更有优势. Comparing physiological traits of 125 invasive with plants 196 non-invasive plants show that invasive plants have more advantages in growth rate, nutrient allocation and stress resistance. | ||
| 外来植物Solanum carolinense具有较强的抗寒能力和无性繁殖能力. Exotic plant, Solanum carolinense, has strong ability for cold resistance and asexual reproduction. | ||
| 气候变暖及氮沉降的增加可以增强入侵植物Solidago canadensis叶片的资源获取能力. Climate warming and increased nitrogen deposition can enhance leaf resource acquisition of an invasive plant, Solidago canadensis. | ||
| 化感物质 Allelochemical | 入侵植物白羊草(Bothriochloa ischaemum)的渗滤液抑制本地植物Andropogon gerardii和Schizachyrium scoparium发芽及生长. The leachate of an invasive plant, Bothriochloa ischaemum, inhibits germination and growth of the native plants, Andropogon gerardii and Schizachyrium scoparium. | |
| 入侵植物水烛(Typha angustifolia)根系分泌的酚类物质抑制了本地植物Bolboschoenus fluviatilis的生长发育. Phenolic compounds secreted by roots of an invasive plant, Typha angustifolia, inhibit growth and development of a native plant, Bolboschoenus fluviatilis. | ||
| Solidago canadensis入侵分泌的化感物质抑制了本地植物莴苣(Lactuca sativa)种子萌发和幼苗生长. Allelochemicals secreted by the invasion of Solidago canadensis inhibit seed germination and seedling growth of a native plant, Lactuca sativa. | ||
| 外来植物Solanum carolinense含有龙葵碱及草酸盐结晶. The exotic plant Solanum carolinense contains solanine and oxalate crystals. | ||
| Parthenium hysterophorus可以分泌帕台单宁、咖啡酸、香草酸等化感物质帮助自身入侵. Parthenium hysterophorus can secrete parthenin, caffeic acid, vanillic acid and other allelochemicals to facilitate its invasion. | ||
| 土壤养分变化 Soil nutrient variation | 氮沉降会增强S. canadensis入侵初期分泌的化感物质对本地植物莴苣种子萌发和幼苗生长的抑制程度. Nitrogen deposition can enhance the inhibition of the allelochemicals secreted by S. canadensis at its initial invasive stage on seed germination and seedling growth of a native plant, Lactuca sativa. | |
| 分析94篇文章发现, 入侵植物增大了植物、土壤和土壤微生物中的碳和氮库大小. Analysis of 94 published papers show that invasive plants increase the size of carbon and nitrogen pools in plants, soils and soil microorganisms. | ||
| 与本地植物Artemisia californica、Salvia mellifera、Salvia apiana、锦鸡儿(Calochortus splendens)和Dichelostemma capitatum相比, 外来植物Centaurea stoebe、Hirschfeldia incana和Bromus madritensis可以通过加快凋落物分解改善土壤养分. Compared with native plants such as Artemisia californica, Salvia mellifera, Salvia apiana, Calochortus splendens and Dichelostemma capitatum, exotic plants, like Centaurea stoebe, Hirschfeldia incana and Bromus madritensis, can increase soil nutrient level by increasing litter decomposition. | ||
| 入侵植物Mikania micrantha分泌的化感物质绿原酸和β-石竹烯加快了土壤有机氮的矿化过程. Allelochemicals secreted by the invasive plant, Mikania micrantha, such as chlorogenic acid and β-caryophyllene accelerate mineralization of soil organic nitrogen. | ||
| 入侵植物通过凋落物改变了根际微生物土壤群落和养分循环, 形成植物-土壤正反馈调节. Invasive plants can form a positive plant-soil feedback via its litter decomposition which could change rhizosphere soil microbial communities and nutrient cycling. | ||
| 杂草Conyza canadensis入侵的土壤中有效氮和有机质含量增加. The content of available nitrogen and organic matter increases in the soil invaded by Conyza canadensis. | ||
| 与非入侵地土壤相比, Eupatorium catarium入侵后土壤氮含量提高4.32 mg·kg-1. Compared with non-invasive soils, soil nitrogen content increases by 4.32 mg·kg-1 after invasion by Eupatorium catarium. | ||
| Ambrosia trifida在入侵地的土壤-植物反馈高于原产地. Ambrosia trifida has a greater plant-soil feedback in invaded ranges than that in its native ranges. |
...
Analysis of 94 published papers show that invasive plants increase the size of carbon and nitrogen pools in plants, soils and soil microorganisms.
Compared with native plants such as Artemisia californica, Salvia mellifera, Salvia apiana, Calochortus splendens and Dichelostemma capitatum, exotic plants, like Centaurea stoebe, Hirschfeldia incana and Bromus madritensis, can increase soil nutrient level by increasing litter decomposition.
Allelochemicals secreted by the invasive plant, Mikania micrantha, such as chlorogenic acid and β-caryophyllene accelerate mineralization of soil organic nitrogen.
Invasive plants can form a positive plant-soil feedback via its litter decomposition which could change rhizosphere soil microbial communities and nutrient cycling.
The content of available nitrogen and organic matter increases in the soil invaded by Conyza canadensis.
Compared with non-invasive soils, soil nitrogen content increases by 4.32 mg·kg-1 after invasion by Eupatorium catarium.
Ambrosia trifida has a greater plant-soil feedback in invaded ranges than that in its native ranges.
... 已有研究证实外来植物的成功入侵受入侵时间的影响(
氮沉降对不同入侵程度加拿大一枝黄花化感作用的影响
3
2020
... Ecological mechanisms of alien plant invasion
| 生态机制 Mechanism of ecology | 举例 Example | 参考文献 Reference |
|---|---|---|
| 物种竞争 Species competition | 与农作物(如高粱(Sorghum bicolor)、玉米(Zea mays))相比, 入侵植物Parthenium hysterophorus具有快速早期生长的能力. Compared with crops (such as Sorghum bicolor and Zea mays), the invasive plant Parthenium hysterophorus can grow much more rapidly in their early stage. | |
| 比较夏威夷19对入侵植物和本地植物的资源利用效率, 发现入侵植物具有较高的碳同化率、光利用效率、瞬时氮利用效率和瞬时能量利用效率. Comparing the resource utilization efficiency for 19 paired invasive-native plants in Hawaii, invasive plants are observed to have higher carbon assimilation rate, light use efficiency, instantaneous nitrogen and energy use efficiency. | ||
| 比较47对入侵植物和非入侵植物对草食性动物的抵抗能力及养分含量发现, 入侵植物拥有更高的叶氮含量, 受到食草动物的损伤较小. Comparisons among 47 paired invasive and non-invasive plants for leaf herbivore-resistance and nutrient content show that invasive plants have higher leaf nitrogen content and less damage by herbivores. | ||
| 比对125种入侵植物和196种非入侵植物的生理性状发现, 入侵植物在生长率、养分分配及抗逆性更有优势. Comparing physiological traits of 125 invasive with plants 196 non-invasive plants show that invasive plants have more advantages in growth rate, nutrient allocation and stress resistance. | ||
| 外来植物Solanum carolinense具有较强的抗寒能力和无性繁殖能力. Exotic plant, Solanum carolinense, has strong ability for cold resistance and asexual reproduction. | ||
| 气候变暖及氮沉降的增加可以增强入侵植物Solidago canadensis叶片的资源获取能力. Climate warming and increased nitrogen deposition can enhance leaf resource acquisition of an invasive plant, Solidago canadensis. | ||
| 化感物质 Allelochemical | 入侵植物白羊草(Bothriochloa ischaemum)的渗滤液抑制本地植物Andropogon gerardii和Schizachyrium scoparium发芽及生长. The leachate of an invasive plant, Bothriochloa ischaemum, inhibits germination and growth of the native plants, Andropogon gerardii and Schizachyrium scoparium. | |
| 入侵植物水烛(Typha angustifolia)根系分泌的酚类物质抑制了本地植物Bolboschoenus fluviatilis的生长发育. Phenolic compounds secreted by roots of an invasive plant, Typha angustifolia, inhibit growth and development of a native plant, Bolboschoenus fluviatilis. | ||
| Solidago canadensis入侵分泌的化感物质抑制了本地植物莴苣(Lactuca sativa)种子萌发和幼苗生长. Allelochemicals secreted by the invasion of Solidago canadensis inhibit seed germination and seedling growth of a native plant, Lactuca sativa. | ||
| 外来植物Solanum carolinense含有龙葵碱及草酸盐结晶. The exotic plant Solanum carolinense contains solanine and oxalate crystals. | ||
| Parthenium hysterophorus可以分泌帕台单宁、咖啡酸、香草酸等化感物质帮助自身入侵. Parthenium hysterophorus can secrete parthenin, caffeic acid, vanillic acid and other allelochemicals to facilitate its invasion. | ||
| 土壤养分变化 Soil nutrient variation | 氮沉降会增强S. canadensis入侵初期分泌的化感物质对本地植物莴苣种子萌发和幼苗生长的抑制程度. Nitrogen deposition can enhance the inhibition of the allelochemicals secreted by S. canadensis at its initial invasive stage on seed germination and seedling growth of a native plant, Lactuca sativa. | |
| 分析94篇文章发现, 入侵植物增大了植物、土壤和土壤微生物中的碳和氮库大小. Analysis of 94 published papers show that invasive plants increase the size of carbon and nitrogen pools in plants, soils and soil microorganisms. | ||
| 与本地植物Artemisia californica、Salvia mellifera、Salvia apiana、锦鸡儿(Calochortus splendens)和Dichelostemma capitatum相比, 外来植物Centaurea stoebe、Hirschfeldia incana和Bromus madritensis可以通过加快凋落物分解改善土壤养分. Compared with native plants such as Artemisia californica, Salvia mellifera, Salvia apiana, Calochortus splendens and Dichelostemma capitatum, exotic plants, like Centaurea stoebe, Hirschfeldia incana and Bromus madritensis, can increase soil nutrient level by increasing litter decomposition. | ||
| 入侵植物Mikania micrantha分泌的化感物质绿原酸和β-石竹烯加快了土壤有机氮的矿化过程. Allelochemicals secreted by the invasive plant, Mikania micrantha, such as chlorogenic acid and β-caryophyllene accelerate mineralization of soil organic nitrogen. | ||
| 入侵植物通过凋落物改变了根际微生物土壤群落和养分循环, 形成植物-土壤正反馈调节. Invasive plants can form a positive plant-soil feedback via its litter decomposition which could change rhizosphere soil microbial communities and nutrient cycling. | ||
| 杂草Conyza canadensis入侵的土壤中有效氮和有机质含量增加. The content of available nitrogen and organic matter increases in the soil invaded by Conyza canadensis. | ||
| 与非入侵地土壤相比, Eupatorium catarium入侵后土壤氮含量提高4.32 mg·kg-1. Compared with non-invasive soils, soil nitrogen content increases by 4.32 mg·kg-1 after invasion by Eupatorium catarium. | ||
| Ambrosia trifida在入侵地的土壤-植物反馈高于原产地. Ambrosia trifida has a greater plant-soil feedback in invaded ranges than that in its native ranges. |
...
Analysis of 94 published papers show that invasive plants increase the size of carbon and nitrogen pools in plants, soils and soil microorganisms.
Compared with native plants such as Artemisia californica, Salvia mellifera, Salvia apiana, Calochortus splendens and Dichelostemma capitatum, exotic plants, like Centaurea stoebe, Hirschfeldia incana and Bromus madritensis, can increase soil nutrient level by increasing litter decomposition.
Allelochemicals secreted by the invasive plant, Mikania micrantha, such as chlorogenic acid and β-caryophyllene accelerate mineralization of soil organic nitrogen.
Invasive plants can form a positive plant-soil feedback via its litter decomposition which could change rhizosphere soil microbial communities and nutrient cycling.
The content of available nitrogen and organic matter increases in the soil invaded by Conyza canadensis.
Compared with non-invasive soils, soil nitrogen content increases by 4.32 mg·kg-1 after invasion by Eupatorium catarium.
Ambrosia trifida has a greater plant-soil feedback in invaded ranges than that in its native ranges.
... 已有研究证实外来植物的成功入侵受入侵时间的影响(
Lesser leaf herbivore damage and structural defense and greater nutrient concentrations for invasive alien plants: evidence from 47 pairs of invasive and non-invasive plants
3
2020
... 入侵植物可以通过多种生态机制影响本地植物的菌根真菌(
... ;
... Ecological mechanisms of alien plant invasion
| 生态机制 Mechanism of ecology | 举例 Example | 参考文献 Reference |
|---|---|---|
| 物种竞争 Species competition | 与农作物(如高粱(Sorghum bicolor)、玉米(Zea mays))相比, 入侵植物Parthenium hysterophorus具有快速早期生长的能力. Compared with crops (such as Sorghum bicolor and Zea mays), the invasive plant Parthenium hysterophorus can grow much more rapidly in their early stage. | |
| 比较夏威夷19对入侵植物和本地植物的资源利用效率, 发现入侵植物具有较高的碳同化率、光利用效率、瞬时氮利用效率和瞬时能量利用效率. Comparing the resource utilization efficiency for 19 paired invasive-native plants in Hawaii, invasive plants are observed to have higher carbon assimilation rate, light use efficiency, instantaneous nitrogen and energy use efficiency. | ||
| 比较47对入侵植物和非入侵植物对草食性动物的抵抗能力及养分含量发现, 入侵植物拥有更高的叶氮含量, 受到食草动物的损伤较小. Comparisons among 47 paired invasive and non-invasive plants for leaf herbivore-resistance and nutrient content show that invasive plants have higher leaf nitrogen content and less damage by herbivores. | ||
| 比对125种入侵植物和196种非入侵植物的生理性状发现, 入侵植物在生长率、养分分配及抗逆性更有优势. Comparing physiological traits of 125 invasive with plants 196 non-invasive plants show that invasive plants have more advantages in growth rate, nutrient allocation and stress resistance. | ||
| 外来植物Solanum carolinense具有较强的抗寒能力和无性繁殖能力. Exotic plant, Solanum carolinense, has strong ability for cold resistance and asexual reproduction. | ||
| 气候变暖及氮沉降的增加可以增强入侵植物Solidago canadensis叶片的资源获取能力. Climate warming and increased nitrogen deposition can enhance leaf resource acquisition of an invasive plant, Solidago canadensis. | ||
| 化感物质 Allelochemical | 入侵植物白羊草(Bothriochloa ischaemum)的渗滤液抑制本地植物Andropogon gerardii和Schizachyrium scoparium发芽及生长. The leachate of an invasive plant, Bothriochloa ischaemum, inhibits germination and growth of the native plants, Andropogon gerardii and Schizachyrium scoparium. | |
| 入侵植物水烛(Typha angustifolia)根系分泌的酚类物质抑制了本地植物Bolboschoenus fluviatilis的生长发育. Phenolic compounds secreted by roots of an invasive plant, Typha angustifolia, inhibit growth and development of a native plant, Bolboschoenus fluviatilis. | ||
| Solidago canadensis入侵分泌的化感物质抑制了本地植物莴苣(Lactuca sativa)种子萌发和幼苗生长. Allelochemicals secreted by the invasion of Solidago canadensis inhibit seed germination and seedling growth of a native plant, Lactuca sativa. | ||
| 外来植物Solanum carolinense含有龙葵碱及草酸盐结晶. The exotic plant Solanum carolinense contains solanine and oxalate crystals. | ||
| Parthenium hysterophorus可以分泌帕台单宁、咖啡酸、香草酸等化感物质帮助自身入侵. Parthenium hysterophorus can secrete parthenin, caffeic acid, vanillic acid and other allelochemicals to facilitate its invasion. | ||
| 土壤养分变化 Soil nutrient variation | 氮沉降会增强S. canadensis入侵初期分泌的化感物质对本地植物莴苣种子萌发和幼苗生长的抑制程度. Nitrogen deposition can enhance the inhibition of the allelochemicals secreted by S. canadensis at its initial invasive stage on seed germination and seedling growth of a native plant, Lactuca sativa. | |
| 分析94篇文章发现, 入侵植物增大了植物、土壤和土壤微生物中的碳和氮库大小. Analysis of 94 published papers show that invasive plants increase the size of carbon and nitrogen pools in plants, soils and soil microorganisms. | ||
| 与本地植物Artemisia californica、Salvia mellifera、Salvia apiana、锦鸡儿(Calochortus splendens)和Dichelostemma capitatum相比, 外来植物Centaurea stoebe、Hirschfeldia incana和Bromus madritensis可以通过加快凋落物分解改善土壤养分. Compared with native plants such as Artemisia californica, Salvia mellifera, Salvia apiana, Calochortus splendens and Dichelostemma capitatum, exotic plants, like Centaurea stoebe, Hirschfeldia incana and Bromus madritensis, can increase soil nutrient level by increasing litter decomposition. | ||
| 入侵植物Mikania micrantha分泌的化感物质绿原酸和β-石竹烯加快了土壤有机氮的矿化过程. Allelochemicals secreted by the invasive plant, Mikania micrantha, such as chlorogenic acid and β-caryophyllene accelerate mineralization of soil organic nitrogen. | ||
| 入侵植物通过凋落物改变了根际微生物土壤群落和养分循环, 形成植物-土壤正反馈调节. Invasive plants can form a positive plant-soil feedback via its litter decomposition which could change rhizosphere soil microbial communities and nutrient cycling. | ||
| 杂草Conyza canadensis入侵的土壤中有效氮和有机质含量增加. The content of available nitrogen and organic matter increases in the soil invaded by Conyza canadensis. | ||
| 与非入侵地土壤相比, Eupatorium catarium入侵后土壤氮含量提高4.32 mg·kg-1. Compared with non-invasive soils, soil nitrogen content increases by 4.32 mg·kg-1 after invasion by Eupatorium catarium. | ||
| Ambrosia trifida在入侵地的土壤-植物反馈高于原产地. Ambrosia trifida has a greater plant-soil feedback in invaded ranges than that in its native ranges. |
Allelopathy as a mechanism for the invasion of Typha angustifolia
1
2009
... Ecological mechanisms of alien plant invasion
| 生态机制 Mechanism of ecology | 举例 Example | 参考文献 Reference |
|---|---|---|
| 物种竞争 Species competition | 与农作物(如高粱(Sorghum bicolor)、玉米(Zea mays))相比, 入侵植物Parthenium hysterophorus具有快速早期生长的能力. Compared with crops (such as Sorghum bicolor and Zea mays), the invasive plant Parthenium hysterophorus can grow much more rapidly in their early stage. | |
| 比较夏威夷19对入侵植物和本地植物的资源利用效率, 发现入侵植物具有较高的碳同化率、光利用效率、瞬时氮利用效率和瞬时能量利用效率. Comparing the resource utilization efficiency for 19 paired invasive-native plants in Hawaii, invasive plants are observed to have higher carbon assimilation rate, light use efficiency, instantaneous nitrogen and energy use efficiency. | ||
| 比较47对入侵植物和非入侵植物对草食性动物的抵抗能力及养分含量发现, 入侵植物拥有更高的叶氮含量, 受到食草动物的损伤较小. Comparisons among 47 paired invasive and non-invasive plants for leaf herbivore-resistance and nutrient content show that invasive plants have higher leaf nitrogen content and less damage by herbivores. | ||
| 比对125种入侵植物和196种非入侵植物的生理性状发现, 入侵植物在生长率、养分分配及抗逆性更有优势. Comparing physiological traits of 125 invasive with plants 196 non-invasive plants show that invasive plants have more advantages in growth rate, nutrient allocation and stress resistance. | ||
| 外来植物Solanum carolinense具有较强的抗寒能力和无性繁殖能力. Exotic plant, Solanum carolinense, has strong ability for cold resistance and asexual reproduction. | ||
| 气候变暖及氮沉降的增加可以增强入侵植物Solidago canadensis叶片的资源获取能力. Climate warming and increased nitrogen deposition can enhance leaf resource acquisition of an invasive plant, Solidago canadensis. | ||
| 化感物质 Allelochemical | 入侵植物白羊草(Bothriochloa ischaemum)的渗滤液抑制本地植物Andropogon gerardii和Schizachyrium scoparium发芽及生长. The leachate of an invasive plant, Bothriochloa ischaemum, inhibits germination and growth of the native plants, Andropogon gerardii and Schizachyrium scoparium. | |
| 入侵植物水烛(Typha angustifolia)根系分泌的酚类物质抑制了本地植物Bolboschoenus fluviatilis的生长发育. Phenolic compounds secreted by roots of an invasive plant, Typha angustifolia, inhibit growth and development of a native plant, Bolboschoenus fluviatilis. | ||
| Solidago canadensis入侵分泌的化感物质抑制了本地植物莴苣(Lactuca sativa)种子萌发和幼苗生长. Allelochemicals secreted by the invasion of Solidago canadensis inhibit seed germination and seedling growth of a native plant, Lactuca sativa. | ||
| 外来植物Solanum carolinense含有龙葵碱及草酸盐结晶. The exotic plant Solanum carolinense contains solanine and oxalate crystals. | ||
| Parthenium hysterophorus可以分泌帕台单宁、咖啡酸、香草酸等化感物质帮助自身入侵. Parthenium hysterophorus can secrete parthenin, caffeic acid, vanillic acid and other allelochemicals to facilitate its invasion. | ||
| 土壤养分变化 Soil nutrient variation | 氮沉降会增强S. canadensis入侵初期分泌的化感物质对本地植物莴苣种子萌发和幼苗生长的抑制程度. Nitrogen deposition can enhance the inhibition of the allelochemicals secreted by S. canadensis at its initial invasive stage on seed germination and seedling growth of a native plant, Lactuca sativa. | |
| 分析94篇文章发现, 入侵植物增大了植物、土壤和土壤微生物中的碳和氮库大小. Analysis of 94 published papers show that invasive plants increase the size of carbon and nitrogen pools in plants, soils and soil microorganisms. | ||
| 与本地植物Artemisia californica、Salvia mellifera、Salvia apiana、锦鸡儿(Calochortus splendens)和Dichelostemma capitatum相比, 外来植物Centaurea stoebe、Hirschfeldia incana和Bromus madritensis可以通过加快凋落物分解改善土壤养分. Compared with native plants such as Artemisia californica, Salvia mellifera, Salvia apiana, Calochortus splendens and Dichelostemma capitatum, exotic plants, like Centaurea stoebe, Hirschfeldia incana and Bromus madritensis, can increase soil nutrient level by increasing litter decomposition. | ||
| 入侵植物Mikania micrantha分泌的化感物质绿原酸和β-石竹烯加快了土壤有机氮的矿化过程. Allelochemicals secreted by the invasive plant, Mikania micrantha, such as chlorogenic acid and β-caryophyllene accelerate mineralization of soil organic nitrogen. | ||
| 入侵植物通过凋落物改变了根际微生物土壤群落和养分循环, 形成植物-土壤正反馈调节. Invasive plants can form a positive plant-soil feedback via its litter decomposition which could change rhizosphere soil microbial communities and nutrient cycling. | ||
| 杂草Conyza canadensis入侵的土壤中有效氮和有机质含量增加. The content of available nitrogen and organic matter increases in the soil invaded by Conyza canadensis. | ||
| 与非入侵地土壤相比, Eupatorium catarium入侵后土壤氮含量提高4.32 mg·kg-1. Compared with non-invasive soils, soil nitrogen content increases by 4.32 mg·kg-1 after invasion by Eupatorium catarium. | ||
| Ambrosia trifida在入侵地的土壤-植物反馈高于原产地. Ambrosia trifida has a greater plant-soil feedback in invaded ranges than that in its native ranges. |
Plants transfer lipids to sustain colonization by mutualistic mycorrhizal and parasitic fungi
2
2017
... 菌根的根内菌丝延伸至根外形成根外菌丝, 根外菌丝直接参与土壤养分的吸收.外来入侵植物在对本地植物菌根真菌侵染率、真菌内部结构产生影响的同时, 也会对根外菌丝产生影响(
... 植物与菌根真菌形成共生关系是一个复杂的信号转导过程.目前已经证实, 丛枝菌根的形成始于根系与AMF之间的信号交流(
辽宁省9种兰科植物根内与根际土壤中真菌群落结构的差异
1
2019
... 上述这些研究中, 本地植物菌根真菌群落组成都是通过变性梯度凝胶电泳(DGGE)(
辽宁省9种兰科植物根内与根际土壤中真菌群落结构的差异
1
2019
... 上述这些研究中, 本地植物菌根真菌群落组成都是通过变性梯度凝胶电泳(DGGE)(
Nitrogen enrichment alters mycorrhizal allocation at five mesic to semiarid grasslands
3
2003
... 外来植物入侵后, 本地植物菌根真菌侵染率有时没有受到显著影响(
... 丛枝菌根内部结构概念图(改自
... A conceptual diagram of the internal structures of arbuscular mycorrhizae in root tissues (adapted from
Resource limitation is a driver of local adaptation in mycorrhizal symbioses
1
2010
... 入侵植物可以通过多种生态机制影响本地植物的菌根真菌(
Evidence of qualitative differences between soil-occupancy effects of invasive vs. native grassland plant species
2
2011
... 入侵植物除了影响本地植物菌根真菌的侵染率、根内和根外菌丝的结构与功能外, 还可能改变其菌根真菌的组成.有研究发现, 外来植物入侵会使本地植物原有的AMF被其他种类的AMF替代(
... 外来植物通过干扰本地植物菌根真菌获得竞争优势是一种重要的入侵机制.尽管如此, 相对于其他入侵机理, 我们对外来植物入侵与本地植物菌根真菌间关系的认识仍然十分匮乏(
近十年中国生物入侵研究进展
1
2012
... 随着全球社会经济活动的日益紧密, 越来越多的植物被带入非原产地环境, 快速扩散成为入侵植物.全球变化背景下, 入侵植物已对当地生物多样性、社会经济以及人类健康造成重大威胁(
近十年中国生物入侵研究进展
1
2012
... 随着全球社会经济活动的日益紧密, 越来越多的植物被带入非原产地环境, 快速扩散成为入侵植物.全球变化背景下, 入侵植物已对当地生物多样性、社会经济以及人类健康造成重大威胁(
Reciprocal rewards stabilize cooperation in the mycorrhizal symbiosis
1
2011
... 上述这些研究中, 本地植物菌根真菌群落组成都是通过变性梯度凝胶电泳(DGGE)(
The effects of arbuscular mycorrhizal (AM) fungal and garlic mustard introductions on native AM fungal diversity
1
2011
... 入侵植物除了影响本地植物菌根真菌的侵染率、根内和根外菌丝的结构与功能外, 还可能改变其菌根真菌的组成.有研究发现, 外来植物入侵会使本地植物原有的AMF被其他种类的AMF替代(
Convergent losses of decay mechanisms and rapid turnover of symbiosis genes in mycorrhizal mutualists
2
2015
... 除了这些被广泛关注的途径, 近年来还发现一类重要的间接途径, 即入侵植物能通过干扰本地物种的共生关系(如植物-传粉昆虫和根系-菌根真菌等)而抑制本地植物的生长(
... 植物与菌根真菌形成共生关系是一个复杂的信号转导过程.目前已经证实, 丛枝菌根的形成始于根系与AMF之间的信号交流(
Evolutionary limits ameliorate the negative impact of an invasive plant
2
2009
... 已有研究证实外来植物的成功入侵受入侵时间的影响(
... )及化感作用(
When two invasion hypotheses are better than one
1
2015
... 随着全球社会经济活动的日益紧密, 越来越多的植物被带入非原产地环境, 快速扩散成为入侵植物.全球变化背景下, 入侵植物已对当地生物多样性、社会经济以及人类健康造成重大威胁(
外来植物入侵对生物多样性的影响及本地生物的进化响应
1
2010
... 已有研究证实外来植物的成功入侵受入侵时间的影响(
外来植物入侵对生物多样性的影响及本地生物的进化响应
1
2010
... 已有研究证实外来植物的成功入侵受入侵时间的影响(
Severe plant invasions can increase mycorrhizal fungal abundance and diversity
2
2013
... 早在21世纪初,
... 外来植物通过干扰本地植物菌根真菌获得竞争优势是一种重要的入侵机制.尽管如此, 相对于其他入侵机理, 我们对外来植物入侵与本地植物菌根真菌间关系的认识仍然十分匮乏(
Facilitation promotes invasions in plant-associated microbial communities
1
2019
... 外来植物入侵不仅会影响本地植物菌根真菌, 也会影响其非菌根真菌(NMF), 包括致病真菌、寄生真菌、一些对植物可能无害的内生真菌以及在极端环境下可以帮助植物生存的有益内生菌(
Changes in soil microbial communities due to biological invasions can reduce allelopathic effects
1
2017
... 已有研究证实外来植物的成功入侵受入侵时间的影响(
Arbuscular mycorrhizal fungi counteract the Janzen- Connell effect of soil pathogens
1
2015
... 外来植物入侵不仅会影响本地植物菌根真菌, 也会影响其非菌根真菌(NMF), 包括致病真菌、寄生真菌、一些对植物可能无害的内生真菌以及在极端环境下可以帮助植物生存的有益内生菌(
Effect of exotic Spartina alterniflora on fungal symbiosis with native plants Phragmites australis and Scirpus mariqueter, and model plants Lolium perenne L. and Trifolium repens
2016
Altered ecosystem carbon and nitrogen cycles by plant invasion, a meta-analysis
3
2008
... 外来植物入侵后, 本地植物菌根真菌侵染率有时没有受到显著影响(
... 入侵植物可以通过多种生态机制影响本地植物的菌根真菌(
... Ecological mechanisms of alien plant invasion
| 生态机制 Mechanism of ecology | 举例 Example | 参考文献 Reference |
|---|---|---|
| 物种竞争 Species competition | 与农作物(如高粱(Sorghum bicolor)、玉米(Zea mays))相比, 入侵植物Parthenium hysterophorus具有快速早期生长的能力. Compared with crops (such as Sorghum bicolor and Zea mays), the invasive plant Parthenium hysterophorus can grow much more rapidly in their early stage. | |
| 比较夏威夷19对入侵植物和本地植物的资源利用效率, 发现入侵植物具有较高的碳同化率、光利用效率、瞬时氮利用效率和瞬时能量利用效率. Comparing the resource utilization efficiency for 19 paired invasive-native plants in Hawaii, invasive plants are observed to have higher carbon assimilation rate, light use efficiency, instantaneous nitrogen and energy use efficiency. | ||
| 比较47对入侵植物和非入侵植物对草食性动物的抵抗能力及养分含量发现, 入侵植物拥有更高的叶氮含量, 受到食草动物的损伤较小. Comparisons among 47 paired invasive and non-invasive plants for leaf herbivore-resistance and nutrient content show that invasive plants have higher leaf nitrogen content and less damage by herbivores. | ||
| 比对125种入侵植物和196种非入侵植物的生理性状发现, 入侵植物在生长率、养分分配及抗逆性更有优势. Comparing physiological traits of 125 invasive with plants 196 non-invasive plants show that invasive plants have more advantages in growth rate, nutrient allocation and stress resistance. | ||
| 外来植物Solanum carolinense具有较强的抗寒能力和无性繁殖能力. Exotic plant, Solanum carolinense, has strong ability for cold resistance and asexual reproduction. | ||
| 气候变暖及氮沉降的增加可以增强入侵植物Solidago canadensis叶片的资源获取能力. Climate warming and increased nitrogen deposition can enhance leaf resource acquisition of an invasive plant, Solidago canadensis. | ||
| 化感物质 Allelochemical | 入侵植物白羊草(Bothriochloa ischaemum)的渗滤液抑制本地植物Andropogon gerardii和Schizachyrium scoparium发芽及生长. The leachate of an invasive plant, Bothriochloa ischaemum, inhibits germination and growth of the native plants, Andropogon gerardii and Schizachyrium scoparium. | |
| 入侵植物水烛(Typha angustifolia)根系分泌的酚类物质抑制了本地植物Bolboschoenus fluviatilis的生长发育. Phenolic compounds secreted by roots of an invasive plant, Typha angustifolia, inhibit growth and development of a native plant, Bolboschoenus fluviatilis. | ||
| Solidago canadensis入侵分泌的化感物质抑制了本地植物莴苣(Lactuca sativa)种子萌发和幼苗生长. Allelochemicals secreted by the invasion of Solidago canadensis inhibit seed germination and seedling growth of a native plant, Lactuca sativa. | ||
| 外来植物Solanum carolinense含有龙葵碱及草酸盐结晶. The exotic plant Solanum carolinense contains solanine and oxalate crystals. | ||
| Parthenium hysterophorus可以分泌帕台单宁、咖啡酸、香草酸等化感物质帮助自身入侵. Parthenium hysterophorus can secrete parthenin, caffeic acid, vanillic acid and other allelochemicals to facilitate its invasion. | ||
| 土壤养分变化 Soil nutrient variation | 氮沉降会增强S. canadensis入侵初期分泌的化感物质对本地植物莴苣种子萌发和幼苗生长的抑制程度. Nitrogen deposition can enhance the inhibition of the allelochemicals secreted by S. canadensis at its initial invasive stage on seed germination and seedling growth of a native plant, Lactuca sativa. | |
| 分析94篇文章发现, 入侵植物增大了植物、土壤和土壤微生物中的碳和氮库大小. Analysis of 94 published papers show that invasive plants increase the size of carbon and nitrogen pools in plants, soils and soil microorganisms. | ||
| 与本地植物Artemisia californica、Salvia mellifera、Salvia apiana、锦鸡儿(Calochortus splendens)和Dichelostemma capitatum相比, 外来植物Centaurea stoebe、Hirschfeldia incana和Bromus madritensis可以通过加快凋落物分解改善土壤养分. Compared with native plants such as Artemisia californica, Salvia mellifera, Salvia apiana, Calochortus splendens and Dichelostemma capitatum, exotic plants, like Centaurea stoebe, Hirschfeldia incana and Bromus madritensis, can increase soil nutrient level by increasing litter decomposition. | ||
| 入侵植物Mikania micrantha分泌的化感物质绿原酸和β-石竹烯加快了土壤有机氮的矿化过程. Allelochemicals secreted by the invasive plant, Mikania micrantha, such as chlorogenic acid and β-caryophyllene accelerate mineralization of soil organic nitrogen. | ||
| 入侵植物通过凋落物改变了根际微生物土壤群落和养分循环, 形成植物-土壤正反馈调节. Invasive plants can form a positive plant-soil feedback via its litter decomposition which could change rhizosphere soil microbial communities and nutrient cycling. | ||
| 杂草Conyza canadensis入侵的土壤中有效氮和有机质含量增加. The content of available nitrogen and organic matter increases in the soil invaded by Conyza canadensis. | ||
| 与非入侵地土壤相比, Eupatorium catarium入侵后土壤氮含量提高4.32 mg·kg-1. Compared with non-invasive soils, soil nitrogen content increases by 4.32 mg·kg-1 after invasion by Eupatorium catarium. | ||
| Ambrosia trifida在入侵地的土壤-植物反馈高于原产地. Ambrosia trifida has a greater plant-soil feedback in invaded ranges than that in its native ranges. |
入侵种加拿大一枝黄花对土壤特性的影响
1
2005
... 入侵植物可以通过多种生态机制影响本地植物的菌根真菌(
入侵种加拿大一枝黄花对土壤特性的影响
1
2005
... 入侵植物可以通过多种生态机制影响本地植物的菌根真菌(
Multiple phosphorus acquisition strategies adopted by fine roots in low-fertility soils in Central Amazonia
2
2019
... 外来植物入侵后, 本地植物菌根真菌侵染率有时没有受到显著影响(
... 外来植物通过干扰本地植物菌根真菌获得竞争优势是一种重要的入侵机制.尽管如此, 相对于其他入侵机理, 我们对外来植物入侵与本地植物菌根真菌间关系的认识仍然十分匮乏(
Mycorrhizal colonization in a successional plant community
2
2019
... 菌根的根内菌丝延伸至根外形成根外菌丝, 根外菌丝直接参与土壤养分的吸收.外来入侵植物在对本地植物菌根真菌侵染率、真菌内部结构产生影响的同时, 也会对根外菌丝产生影响(
... 外来植物通过干扰本地植物菌根真菌获得竞争优势是一种重要的入侵机制.尽管如此, 相对于其他入侵机理, 我们对外来植物入侵与本地植物菌根真菌间关系的认识仍然十分匮乏(
Influence of spotted knapweed (Centaurea maculosa) management treatments on arbuscular mycorrhizae and soil aggregation
1
2004
... 菌根的根内菌丝延伸至根外形成根外菌丝, 根外菌丝直接参与土壤养分的吸收.外来入侵植物在对本地植物菌根真菌侵染率、真菌内部结构产生影响的同时, 也会对根外菌丝产生影响(
Influence of phylogeny on fungal community assembly and ecosystem functioning
1
2007
... 外来植物入侵不仅会影响本地植物菌根真菌, 也会影响其非菌根真菌(NMF), 包括致病真菌、寄生真菌、一些对植物可能无害的内生真菌以及在极端环境下可以帮助植物生存的有益内生菌(
Fungal lipochitooligosaccharide symbiotic signals in arbuscular mycorrhiza
1
2011
... 植物与菌根真菌形成共生关系是一个复杂的信号转导过程.目前已经证实, 丛枝菌根的形成始于根系与AMF之间的信号交流(
Invasive plant management techniques alter arbuscular mycorrhizal fungi
2016
The genome of Laccaria bicolor provides insights into mycorrhizal symbiosis
2
2008
... 菌根的根内菌丝延伸至根外形成根外菌丝, 根外菌丝直接参与土壤养分的吸收.外来入侵植物在对本地植物菌根真菌侵染率、真菌内部结构产生影响的同时, 也会对根外菌丝产生影响(
... 植物与菌根真菌形成共生关系是一个复杂的信号转导过程.目前已经证实, 丛枝菌根的形成始于根系与AMF之间的信号交流(
Ancestral alliances: plant mutualistic symbioses with fungi and bacteria
5
2017
... 除了这些被广泛关注的途径, 近年来还发现一类重要的间接途径, 即入侵植物能通过干扰本地物种的共生关系(如植物-传粉昆虫和根系-菌根真菌等)而抑制本地植物的生长(
... 植物与菌根真菌形成共生关系是一个复杂的信号转导过程.目前已经证实, 丛枝菌根的形成始于根系与AMF之间的信号交流(
... ,
... 植物与菌根真菌共生关系分子模式图(参考
... Conceptual model for the symbiotic relationship between plants and mycorrhizal fungi (adapted from
Disrupting mycorrhizal mutualisms, a potential mechanism by which exotic tamarisk outcompetes native cottonwoods
3
2012
... 入侵植物除了影响本地植物菌根真菌的侵染率、根内和根外菌丝的结构与功能外, 还可能改变其菌根真菌的组成.有研究发现, 外来植物入侵会使本地植物原有的AMF被其他种类的AMF替代(
... 外来植物入侵不仅会影响本地植物菌根真菌, 也会影响其非菌根真菌(NMF), 包括致病真菌、寄生真菌、一些对植物可能无害的内生真菌以及在极端环境下可以帮助植物生存的有益内生菌(
...
Biotic interactions and plant invasions
1
2006
... 除了这些被广泛关注的途径, 近年来还发现一类重要的间接途径, 即入侵植物能通过干扰本地物种的共生关系(如植物-传粉昆虫和根系-菌根真菌等)而抑制本地植物的生长(
The invasive plant species Centaurea maculosa alters arbuscular mycorrhizal fungal communities in the field
1
2006
... 菌根的根内菌丝延伸至根外形成根外菌丝, 根外菌丝直接参与土壤养分的吸收.外来入侵植物在对本地植物菌根真菌侵染率、真菌内部结构产生影响的同时, 也会对根外菌丝产生影响(
The invasive plant, Brassica nigra, degrades local mycorrhizas across a wide geographical landscape
2015
Arbuscular mycorrhiza, the mother of plant root endosymbioses
1
2008
... 除了这些被广泛关注的途径, 近年来还发现一类重要的间接途径, 即入侵植物能通过干扰本地物种的共生关系(如植物-传粉昆虫和根系-菌根真菌等)而抑制本地植物的生长(
Alien plant invasion in the Indian Himalayan Region: state of knowledge and research priorities
1
2019
... 随着全球社会经济活动的日益紧密, 越来越多的植物被带入非原产地环境, 快速扩散成为入侵植物.全球变化背景下, 入侵植物已对当地生物多样性、社会经济以及人类健康造成重大威胁(
Positive plant-soil feedbacks of the invasive Impatiens glandulifera and their effects on above-ground microbial communities
1
2016
... 外来植物入侵后, 本地植物菌根真菌侵染率有时没有受到显著影响(
Soil microbial responses to drought and exotic plants shift carbon metabolism
2
2019
... 入侵植物可以通过多种生态机制影响本地植物的菌根真菌(
... Ecological mechanisms of alien plant invasion
| 生态机制 Mechanism of ecology | 举例 Example | 参考文献 Reference |
|---|---|---|
| 物种竞争 Species competition | 与农作物(如高粱(Sorghum bicolor)、玉米(Zea mays))相比, 入侵植物Parthenium hysterophorus具有快速早期生长的能力. Compared with crops (such as Sorghum bicolor and Zea mays), the invasive plant Parthenium hysterophorus can grow much more rapidly in their early stage. | |
| 比较夏威夷19对入侵植物和本地植物的资源利用效率, 发现入侵植物具有较高的碳同化率、光利用效率、瞬时氮利用效率和瞬时能量利用效率. Comparing the resource utilization efficiency for 19 paired invasive-native plants in Hawaii, invasive plants are observed to have higher carbon assimilation rate, light use efficiency, instantaneous nitrogen and energy use efficiency. | ||
| 比较47对入侵植物和非入侵植物对草食性动物的抵抗能力及养分含量发现, 入侵植物拥有更高的叶氮含量, 受到食草动物的损伤较小. Comparisons among 47 paired invasive and non-invasive plants for leaf herbivore-resistance and nutrient content show that invasive plants have higher leaf nitrogen content and less damage by herbivores. | ||
| 比对125种入侵植物和196种非入侵植物的生理性状发现, 入侵植物在生长率、养分分配及抗逆性更有优势. Comparing physiological traits of 125 invasive with plants 196 non-invasive plants show that invasive plants have more advantages in growth rate, nutrient allocation and stress resistance. | ||
| 外来植物Solanum carolinense具有较强的抗寒能力和无性繁殖能力. Exotic plant, Solanum carolinense, has strong ability for cold resistance and asexual reproduction. | ||
| 气候变暖及氮沉降的增加可以增强入侵植物Solidago canadensis叶片的资源获取能力. Climate warming and increased nitrogen deposition can enhance leaf resource acquisition of an invasive plant, Solidago canadensis. | ||
| 化感物质 Allelochemical | 入侵植物白羊草(Bothriochloa ischaemum)的渗滤液抑制本地植物Andropogon gerardii和Schizachyrium scoparium发芽及生长. The leachate of an invasive plant, Bothriochloa ischaemum, inhibits germination and growth of the native plants, Andropogon gerardii and Schizachyrium scoparium. | |
| 入侵植物水烛(Typha angustifolia)根系分泌的酚类物质抑制了本地植物Bolboschoenus fluviatilis的生长发育. Phenolic compounds secreted by roots of an invasive plant, Typha angustifolia, inhibit growth and development of a native plant, Bolboschoenus fluviatilis. | ||
| Solidago canadensis入侵分泌的化感物质抑制了本地植物莴苣(Lactuca sativa)种子萌发和幼苗生长. Allelochemicals secreted by the invasion of Solidago canadensis inhibit seed germination and seedling growth of a native plant, Lactuca sativa. | ||
| 外来植物Solanum carolinense含有龙葵碱及草酸盐结晶. The exotic plant Solanum carolinense contains solanine and oxalate crystals. | ||
| Parthenium hysterophorus可以分泌帕台单宁、咖啡酸、香草酸等化感物质帮助自身入侵. Parthenium hysterophorus can secrete parthenin, caffeic acid, vanillic acid and other allelochemicals to facilitate its invasion. | ||
| 土壤养分变化 Soil nutrient variation | 氮沉降会增强S. canadensis入侵初期分泌的化感物质对本地植物莴苣种子萌发和幼苗生长的抑制程度. Nitrogen deposition can enhance the inhibition of the allelochemicals secreted by S. canadensis at its initial invasive stage on seed germination and seedling growth of a native plant, Lactuca sativa. | |
| 分析94篇文章发现, 入侵植物增大了植物、土壤和土壤微生物中的碳和氮库大小. Analysis of 94 published papers show that invasive plants increase the size of carbon and nitrogen pools in plants, soils and soil microorganisms. | ||
| 与本地植物Artemisia californica、Salvia mellifera、Salvia apiana、锦鸡儿(Calochortus splendens)和Dichelostemma capitatum相比, 外来植物Centaurea stoebe、Hirschfeldia incana和Bromus madritensis可以通过加快凋落物分解改善土壤养分. Compared with native plants such as Artemisia californica, Salvia mellifera, Salvia apiana, Calochortus splendens and Dichelostemma capitatum, exotic plants, like Centaurea stoebe, Hirschfeldia incana and Bromus madritensis, can increase soil nutrient level by increasing litter decomposition. | ||
| 入侵植物Mikania micrantha分泌的化感物质绿原酸和β-石竹烯加快了土壤有机氮的矿化过程. Allelochemicals secreted by the invasive plant, Mikania micrantha, such as chlorogenic acid and β-caryophyllene accelerate mineralization of soil organic nitrogen. | ||
| 入侵植物通过凋落物改变了根际微生物土壤群落和养分循环, 形成植物-土壤正反馈调节. Invasive plants can form a positive plant-soil feedback via its litter decomposition which could change rhizosphere soil microbial communities and nutrient cycling. | ||
| 杂草Conyza canadensis入侵的土壤中有效氮和有机质含量增加. The content of available nitrogen and organic matter increases in the soil invaded by Conyza canadensis. | ||
| 与非入侵地土壤相比, Eupatorium catarium入侵后土壤氮含量提高4.32 mg·kg-1. Compared with non-invasive soils, soil nitrogen content increases by 4.32 mg·kg-1 after invasion by Eupatorium catarium. | ||
| Ambrosia trifida在入侵地的土壤-植物反馈高于原产地. Ambrosia trifida has a greater plant-soil feedback in invaded ranges than that in its native ranges. |
Mycorrhizal effector PaMiSSP10b alters polyamine biosynthesis in Eucalyptus root cells and promotes root colonization
1
2020
... 植物与菌根真菌形成共生关系是一个复杂的信号转导过程.目前已经证实, 丛枝菌根的形成始于根系与AMF之间的信号交流(
Allelopathic inhibition of germination by Alliaria petiolata (Brassicaceae)
1
2004
... 入侵植物可以通过多种生态机制影响本地植物的菌根真菌(
Mycorrhizal symbioses and plant invasions
1
2009
... 除了这些被广泛关注的途径, 近年来还发现一类重要的间接途径, 即入侵植物能通过干扰本地物种的共生关系(如植物-传粉昆虫和根系-菌根真菌等)而抑制本地植物的生长(
Jack of all trades, master of some? On the role of phenotypic plasticity in plant invasions
1
2006
... 随着全球社会经济活动的日益紧密, 越来越多的植物被带入非原产地环境, 快速扩散成为入侵植物.全球变化背景下, 入侵植物已对当地生物多样性、社会经济以及人类健康造成重大威胁(
Invasion of an annual exotic plant into deciduous forests suppresses arbuscular mycorrhiza symbiosis and reduces performance of sycamore maple saplings
2014
Disrupting ectomycorrhizal symbiosis: indirect effects of an annual invasive plant on growth and survival of beech (Fagus sylvatica) saplings
1
2016
... 已有研究证实外来植物的成功入侵受入侵时间的影响(
Changes in soil diversity and global activities following invasions of the exotic invasive plant, Amaranthus viridis L., decrease the growth of native sahelian Acacia species
2009
Differences in nutrient availability and mycorrhizal infectivity in soils invaded by an exotic plant negatively influence the development of indigenous Acacia species
2012
The inconspicuous gatekeeper: endophytic Serendipita vermifera acts as extended plant protection barrier in the rhizosphere
1
2019
... 外来植物入侵不仅会影响本地植物菌根真菌, 也会影响其非菌根真菌(NMF), 包括致病真菌、寄生真菌、一些对植物可能无害的内生真菌以及在极端环境下可以帮助植物生存的有益内生菌(
Non-random association patterns in a plant-mycorrhizal fungal network reveal host-symbiont specificity
2
2019
... 同种植物常常能与多种菌根真菌共生, 即使是单个根尖也往往被多个特定的菌根真菌类群侵染(
... ).
Plant-soil feedbacks between invasive shrubs and native forest understory species lead to shifts in the abundance of mycorrhizal fungi
2014
Relative phytotoxicity of parthenium weed (Parthenium hysterophorus L.) residues on the seedling growth of a range of Australian native and introduced species
1
2018
... Ecological mechanisms of alien plant invasion
| 生态机制 Mechanism of ecology | 举例 Example | 参考文献 Reference |
|---|---|---|
| 物种竞争 Species competition | 与农作物(如高粱(Sorghum bicolor)、玉米(Zea mays))相比, 入侵植物Parthenium hysterophorus具有快速早期生长的能力. Compared with crops (such as Sorghum bicolor and Zea mays), the invasive plant Parthenium hysterophorus can grow much more rapidly in their early stage. | |
| 比较夏威夷19对入侵植物和本地植物的资源利用效率, 发现入侵植物具有较高的碳同化率、光利用效率、瞬时氮利用效率和瞬时能量利用效率. Comparing the resource utilization efficiency for 19 paired invasive-native plants in Hawaii, invasive plants are observed to have higher carbon assimilation rate, light use efficiency, instantaneous nitrogen and energy use efficiency. | ||
| 比较47对入侵植物和非入侵植物对草食性动物的抵抗能力及养分含量发现, 入侵植物拥有更高的叶氮含量, 受到食草动物的损伤较小. Comparisons among 47 paired invasive and non-invasive plants for leaf herbivore-resistance and nutrient content show that invasive plants have higher leaf nitrogen content and less damage by herbivores. | ||
| 比对125种入侵植物和196种非入侵植物的生理性状发现, 入侵植物在生长率、养分分配及抗逆性更有优势. Comparing physiological traits of 125 invasive with plants 196 non-invasive plants show that invasive plants have more advantages in growth rate, nutrient allocation and stress resistance. | ||
| 外来植物Solanum carolinense具有较强的抗寒能力和无性繁殖能力. Exotic plant, Solanum carolinense, has strong ability for cold resistance and asexual reproduction. | ||
| 气候变暖及氮沉降的增加可以增强入侵植物Solidago canadensis叶片的资源获取能力. Climate warming and increased nitrogen deposition can enhance leaf resource acquisition of an invasive plant, Solidago canadensis. | ||
| 化感物质 Allelochemical | 入侵植物白羊草(Bothriochloa ischaemum)的渗滤液抑制本地植物Andropogon gerardii和Schizachyrium scoparium发芽及生长. The leachate of an invasive plant, Bothriochloa ischaemum, inhibits germination and growth of the native plants, Andropogon gerardii and Schizachyrium scoparium. | |
| 入侵植物水烛(Typha angustifolia)根系分泌的酚类物质抑制了本地植物Bolboschoenus fluviatilis的生长发育. Phenolic compounds secreted by roots of an invasive plant, Typha angustifolia, inhibit growth and development of a native plant, Bolboschoenus fluviatilis. | ||
| Solidago canadensis入侵分泌的化感物质抑制了本地植物莴苣(Lactuca sativa)种子萌发和幼苗生长. Allelochemicals secreted by the invasion of Solidago canadensis inhibit seed germination and seedling growth of a native plant, Lactuca sativa. | ||
| 外来植物Solanum carolinense含有龙葵碱及草酸盐结晶. The exotic plant Solanum carolinense contains solanine and oxalate crystals. | ||
| Parthenium hysterophorus可以分泌帕台单宁、咖啡酸、香草酸等化感物质帮助自身入侵. Parthenium hysterophorus can secrete parthenin, caffeic acid, vanillic acid and other allelochemicals to facilitate its invasion. | ||
| 土壤养分变化 Soil nutrient variation | 氮沉降会增强S. canadensis入侵初期分泌的化感物质对本地植物莴苣种子萌发和幼苗生长的抑制程度. Nitrogen deposition can enhance the inhibition of the allelochemicals secreted by S. canadensis at its initial invasive stage on seed germination and seedling growth of a native plant, Lactuca sativa. | |
| 分析94篇文章发现, 入侵植物增大了植物、土壤和土壤微生物中的碳和氮库大小. Analysis of 94 published papers show that invasive plants increase the size of carbon and nitrogen pools in plants, soils and soil microorganisms. | ||
| 与本地植物Artemisia californica、Salvia mellifera、Salvia apiana、锦鸡儿(Calochortus splendens)和Dichelostemma capitatum相比, 外来植物Centaurea stoebe、Hirschfeldia incana和Bromus madritensis可以通过加快凋落物分解改善土壤养分. Compared with native plants such as Artemisia californica, Salvia mellifera, Salvia apiana, Calochortus splendens and Dichelostemma capitatum, exotic plants, like Centaurea stoebe, Hirschfeldia incana and Bromus madritensis, can increase soil nutrient level by increasing litter decomposition. | ||
| 入侵植物Mikania micrantha分泌的化感物质绿原酸和β-石竹烯加快了土壤有机氮的矿化过程. Allelochemicals secreted by the invasive plant, Mikania micrantha, such as chlorogenic acid and β-caryophyllene accelerate mineralization of soil organic nitrogen. | ||
| 入侵植物通过凋落物改变了根际微生物土壤群落和养分循环, 形成植物-土壤正反馈调节. Invasive plants can form a positive plant-soil feedback via its litter decomposition which could change rhizosphere soil microbial communities and nutrient cycling. | ||
| 杂草Conyza canadensis入侵的土壤中有效氮和有机质含量增加. The content of available nitrogen and organic matter increases in the soil invaded by Conyza canadensis. | ||
| 与非入侵地土壤相比, Eupatorium catarium入侵后土壤氮含量提高4.32 mg·kg-1. Compared with non-invasive soils, soil nitrogen content increases by 4.32 mg·kg-1 after invasion by Eupatorium catarium. | ||
| Ambrosia trifida在入侵地的土壤-植物反馈高于原产地. Ambrosia trifida has a greater plant-soil feedback in invaded ranges than that in its native ranges. |
1
2008
... 外来植物入侵后, 本地植物菌根真菌侵染率有时没有受到显著影响(
Global patterns of plant root colonization intensity by mycorrhizal fungi explained by climate and soil chemistry
1
2015
... 入侵植物可以通过多种生态机制影响本地植物的菌根真菌(
Invasion of Rosa rugosa induced changes in soil nutrients and microbial communities of coastal sand dunes
1
2019
... 外来植物通过干扰本地植物菌根真菌获得竞争优势是一种重要的入侵机制.尽管如此, 相对于其他入侵机理, 我们对外来植物入侵与本地植物菌根真菌间关系的认识仍然十分匮乏(
Climatic controls of decomposition drive the global biogeography of forest-tree symbioses
1
2019
... 除了这些被广泛关注的途径, 近年来还发现一类重要的间接途径, 即入侵植物能通过干扰本地物种的共生关系(如植物-传粉昆虫和根系-菌根真菌等)而抑制本地植物的生长(
Invasive plant suppresses the growth of native tree seedlings by disrupting belowground mutualisms
4
2006
... 除了这些被广泛关注的途径, 近年来还发现一类重要的间接途径, 即入侵植物能通过干扰本地物种的共生关系(如植物-传粉昆虫和根系-菌根真菌等)而抑制本地植物的生长(
... 早在21世纪初,
... 丛枝菌根真菌(AMF)的侵染率, 下降幅度分别是30%、14%和42% (
... ).
入侵豚草与本地植物竞争对丛枝菌根真菌多样性的影响
1
2019
... 外来植物通过干扰本地植物菌根真菌获得竞争优势是一种重要的入侵机制.尽管如此, 相对于其他入侵机理, 我们对外来植物入侵与本地植物菌根真菌间关系的认识仍然十分匮乏(
入侵豚草与本地植物竞争对丛枝菌根真菌多样性的影响
1
2019
... 外来植物通过干扰本地植物菌根真菌获得竞争优势是一种重要的入侵机制.尽管如此, 相对于其他入侵机理, 我们对外来植物入侵与本地植物菌根真菌间关系的认识仍然十分匮乏(
The impact of two non-native plant species on native flora performance: potential implications for habitat restoration
1
2013
... 早在21世纪初,
How mycorrhizal associations drive plant population and community biology
1
2020
... 除了这些被广泛关注的途径, 近年来还发现一类重要的间接途径, 即入侵植物能通过干扰本地物种的共生关系(如植物-传粉昆虫和根系-菌根真菌等)而抑制本地植物的生长(
Root exudate is allelopathic in invaded community but not in native community: field evidence for the novel weapons hypothesis
1
2009
... 入侵植物可以通过多种生态机制影响本地植物的菌根真菌(
Molecular signal communication during arbuscular mycorrhizal formation induces significant transcriptional reprogramming of wheat (Triticum aestivum L.) roots
1
2019
... 植物与菌根真菌形成共生关系是一个复杂的信号转导过程.目前已经证实, 丛枝菌根的形成始于根系与AMF之间的信号交流(
丛枝菌根真菌脂类代谢对共生信号调控的响应和反馈
2
2016
... 菌根的根内菌丝延伸至根外形成根外菌丝, 根外菌丝直接参与土壤养分的吸收.外来入侵植物在对本地植物菌根真菌侵染率、真菌内部结构产生影响的同时, 也会对根外菌丝产生影响(
... 植物与菌根真菌形成共生关系是一个复杂的信号转导过程.目前已经证实, 丛枝菌根的形成始于根系与AMF之间的信号交流(
丛枝菌根真菌脂类代谢对共生信号调控的响应和反馈
2
2016
... 菌根的根内菌丝延伸至根外形成根外菌丝, 根外菌丝直接参与土壤养分的吸收.外来入侵植物在对本地植物菌根真菌侵染率、真菌内部结构产生影响的同时, 也会对根外菌丝产生影响(
... 植物与菌根真菌形成共生关系是一个复杂的信号转导过程.目前已经证实, 丛枝菌根的形成始于根系与AMF之间的信号交流(
Structural diversity across arbuscular mycorrhizal, ectomycorrhizal, and endophytic plant-fungus networks
2
2018
... 同种植物常常能与多种菌根真菌共生, 即使是单个根尖也往往被多个特定的菌根真菌类群侵染(
... 外来植物通过干扰本地植物菌根真菌获得竞争优势是一种重要的入侵机制.尽管如此, 相对于其他入侵机理, 我们对外来植物入侵与本地植物菌根真菌间关系的认识仍然十分匮乏(
Cogongrass (Imperata cylindrica) affects above- and belowground processes in commercial loblolly pine (Pinus taeda) stands
2017
Global exchange and accumulation of non-native plants
1
2015
... 随着全球社会经济活动的日益紧密, 越来越多的植物被带入非原产地环境, 快速扩散成为入侵植物.全球变化背景下, 入侵植物已对当地生物多样性、社会经济以及人类健康造成重大威胁(
A meta-analysis of trait differences between invasive and non-invasive plant species
2
2010
... 入侵植物可以通过多种生态机制影响本地植物的菌根真菌(
... Ecological mechanisms of alien plant invasion
| 生态机制 Mechanism of ecology | 举例 Example | 参考文献 Reference |
|---|---|---|
| 物种竞争 Species competition | 与农作物(如高粱(Sorghum bicolor)、玉米(Zea mays))相比, 入侵植物Parthenium hysterophorus具有快速早期生长的能力. Compared with crops (such as Sorghum bicolor and Zea mays), the invasive plant Parthenium hysterophorus can grow much more rapidly in their early stage. | |
| 比较夏威夷19对入侵植物和本地植物的资源利用效率, 发现入侵植物具有较高的碳同化率、光利用效率、瞬时氮利用效率和瞬时能量利用效率. Comparing the resource utilization efficiency for 19 paired invasive-native plants in Hawaii, invasive plants are observed to have higher carbon assimilation rate, light use efficiency, instantaneous nitrogen and energy use efficiency. | ||
| 比较47对入侵植物和非入侵植物对草食性动物的抵抗能力及养分含量发现, 入侵植物拥有更高的叶氮含量, 受到食草动物的损伤较小. Comparisons among 47 paired invasive and non-invasive plants for leaf herbivore-resistance and nutrient content show that invasive plants have higher leaf nitrogen content and less damage by herbivores. | ||
| 比对125种入侵植物和196种非入侵植物的生理性状发现, 入侵植物在生长率、养分分配及抗逆性更有优势. Comparing physiological traits of 125 invasive with plants 196 non-invasive plants show that invasive plants have more advantages in growth rate, nutrient allocation and stress resistance. | ||
| 外来植物Solanum carolinense具有较强的抗寒能力和无性繁殖能力. Exotic plant, Solanum carolinense, has strong ability for cold resistance and asexual reproduction. | ||
| 气候变暖及氮沉降的增加可以增强入侵植物Solidago canadensis叶片的资源获取能力. Climate warming and increased nitrogen deposition can enhance leaf resource acquisition of an invasive plant, Solidago canadensis. | ||
| 化感物质 Allelochemical | 入侵植物白羊草(Bothriochloa ischaemum)的渗滤液抑制本地植物Andropogon gerardii和Schizachyrium scoparium发芽及生长. The leachate of an invasive plant, Bothriochloa ischaemum, inhibits germination and growth of the native plants, Andropogon gerardii and Schizachyrium scoparium. | |
| 入侵植物水烛(Typha angustifolia)根系分泌的酚类物质抑制了本地植物Bolboschoenus fluviatilis的生长发育. Phenolic compounds secreted by roots of an invasive plant, Typha angustifolia, inhibit growth and development of a native plant, Bolboschoenus fluviatilis. | ||
| Solidago canadensis入侵分泌的化感物质抑制了本地植物莴苣(Lactuca sativa)种子萌发和幼苗生长. Allelochemicals secreted by the invasion of Solidago canadensis inhibit seed germination and seedling growth of a native plant, Lactuca sativa. | ||
| 外来植物Solanum carolinense含有龙葵碱及草酸盐结晶. The exotic plant Solanum carolinense contains solanine and oxalate crystals. | ||
| Parthenium hysterophorus可以分泌帕台单宁、咖啡酸、香草酸等化感物质帮助自身入侵. Parthenium hysterophorus can secrete parthenin, caffeic acid, vanillic acid and other allelochemicals to facilitate its invasion. | ||
| 土壤养分变化 Soil nutrient variation | 氮沉降会增强S. canadensis入侵初期分泌的化感物质对本地植物莴苣种子萌发和幼苗生长的抑制程度. Nitrogen deposition can enhance the inhibition of the allelochemicals secreted by S. canadensis at its initial invasive stage on seed germination and seedling growth of a native plant, Lactuca sativa. | |
| 分析94篇文章发现, 入侵植物增大了植物、土壤和土壤微生物中的碳和氮库大小. Analysis of 94 published papers show that invasive plants increase the size of carbon and nitrogen pools in plants, soils and soil microorganisms. | ||
| 与本地植物Artemisia californica、Salvia mellifera、Salvia apiana、锦鸡儿(Calochortus splendens)和Dichelostemma capitatum相比, 外来植物Centaurea stoebe、Hirschfeldia incana和Bromus madritensis可以通过加快凋落物分解改善土壤养分. Compared with native plants such as Artemisia californica, Salvia mellifera, Salvia apiana, Calochortus splendens and Dichelostemma capitatum, exotic plants, like Centaurea stoebe, Hirschfeldia incana and Bromus madritensis, can increase soil nutrient level by increasing litter decomposition. | ||
| 入侵植物Mikania micrantha分泌的化感物质绿原酸和β-石竹烯加快了土壤有机氮的矿化过程. Allelochemicals secreted by the invasive plant, Mikania micrantha, such as chlorogenic acid and β-caryophyllene accelerate mineralization of soil organic nitrogen. | ||
| 入侵植物通过凋落物改变了根际微生物土壤群落和养分循环, 形成植物-土壤正反馈调节. Invasive plants can form a positive plant-soil feedback via its litter decomposition which could change rhizosphere soil microbial communities and nutrient cycling. | ||
| 杂草Conyza canadensis入侵的土壤中有效氮和有机质含量增加. The content of available nitrogen and organic matter increases in the soil invaded by Conyza canadensis. | ||
| 与非入侵地土壤相比, Eupatorium catarium入侵后土壤氮含量提高4.32 mg·kg-1. Compared with non-invasive soils, soil nitrogen content increases by 4.32 mg·kg-1 after invasion by Eupatorium catarium. | ||
| Ambrosia trifida在入侵地的土壤-植物反馈高于原产地. Ambrosia trifida has a greater plant-soil feedback in invaded ranges than that in its native ranges. |
Mycorrhizal densities decline in association with nonnative plants and contribute to plant invasion
1
2009
... 除了这些被广泛关注的途径, 近年来还发现一类重要的间接途径, 即入侵植物能通过干扰本地物种的共生关系(如植物-传粉昆虫和根系-菌根真菌等)而抑制本地植物的生长(
1
2011
... 随着全球社会经济活动的日益紧密, 越来越多的植物被带入非原产地环境, 快速扩散成为入侵植物.全球变化背景下, 入侵植物已对当地生物多样性、社会经济以及人类健康造成重大威胁(
1
2011
... 随着全球社会经济活动的日益紧密, 越来越多的植物被带入非原产地环境, 快速扩散成为入侵植物.全球变化背景下, 入侵植物已对当地生物多样性、社会经济以及人类健康造成重大威胁(
Effects of ecoregional vulnerability on habitat suitability of invasive alien plants, an assessment using 13 species on a global scale
1
2019
... 随着全球社会经济活动的日益紧密, 越来越多的植物被带入非原产地环境, 快速扩散成为入侵植物.全球变化背景下, 入侵植物已对当地生物多样性、社会经济以及人类健康造成重大威胁(
入侵性杂草北美刺龙葵在北京市的首次发现及防除
3
2020
... 入侵植物可以通过多种生态机制影响本地植物的菌根真菌(
... Ecological mechanisms of alien plant invasion
| 生态机制 Mechanism of ecology | 举例 Example | 参考文献 Reference |
|---|---|---|
| 物种竞争 Species competition | 与农作物(如高粱(Sorghum bicolor)、玉米(Zea mays))相比, 入侵植物Parthenium hysterophorus具有快速早期生长的能力. Compared with crops (such as Sorghum bicolor and Zea mays), the invasive plant Parthenium hysterophorus can grow much more rapidly in their early stage. | |
| 比较夏威夷19对入侵植物和本地植物的资源利用效率, 发现入侵植物具有较高的碳同化率、光利用效率、瞬时氮利用效率和瞬时能量利用效率. Comparing the resource utilization efficiency for 19 paired invasive-native plants in Hawaii, invasive plants are observed to have higher carbon assimilation rate, light use efficiency, instantaneous nitrogen and energy use efficiency. | ||
| 比较47对入侵植物和非入侵植物对草食性动物的抵抗能力及养分含量发现, 入侵植物拥有更高的叶氮含量, 受到食草动物的损伤较小. Comparisons among 47 paired invasive and non-invasive plants for leaf herbivore-resistance and nutrient content show that invasive plants have higher leaf nitrogen content and less damage by herbivores. | ||
| 比对125种入侵植物和196种非入侵植物的生理性状发现, 入侵植物在生长率、养分分配及抗逆性更有优势. Comparing physiological traits of 125 invasive with plants 196 non-invasive plants show that invasive plants have more advantages in growth rate, nutrient allocation and stress resistance. | ||
| 外来植物Solanum carolinense具有较强的抗寒能力和无性繁殖能力. Exotic plant, Solanum carolinense, has strong ability for cold resistance and asexual reproduction. | ||
| 气候变暖及氮沉降的增加可以增强入侵植物Solidago canadensis叶片的资源获取能力. Climate warming and increased nitrogen deposition can enhance leaf resource acquisition of an invasive plant, Solidago canadensis. | ||
| 化感物质 Allelochemical | 入侵植物白羊草(Bothriochloa ischaemum)的渗滤液抑制本地植物Andropogon gerardii和Schizachyrium scoparium发芽及生长. The leachate of an invasive plant, Bothriochloa ischaemum, inhibits germination and growth of the native plants, Andropogon gerardii and Schizachyrium scoparium. | |
| 入侵植物水烛(Typha angustifolia)根系分泌的酚类物质抑制了本地植物Bolboschoenus fluviatilis的生长发育. Phenolic compounds secreted by roots of an invasive plant, Typha angustifolia, inhibit growth and development of a native plant, Bolboschoenus fluviatilis. | ||
| Solidago canadensis入侵分泌的化感物质抑制了本地植物莴苣(Lactuca sativa)种子萌发和幼苗生长. Allelochemicals secreted by the invasion of Solidago canadensis inhibit seed germination and seedling growth of a native plant, Lactuca sativa. | ||
| 外来植物Solanum carolinense含有龙葵碱及草酸盐结晶. The exotic plant Solanum carolinense contains solanine and oxalate crystals. | ||
| Parthenium hysterophorus可以分泌帕台单宁、咖啡酸、香草酸等化感物质帮助自身入侵. Parthenium hysterophorus can secrete parthenin, caffeic acid, vanillic acid and other allelochemicals to facilitate its invasion. | ||
| 土壤养分变化 Soil nutrient variation | 氮沉降会增强S. canadensis入侵初期分泌的化感物质对本地植物莴苣种子萌发和幼苗生长的抑制程度. Nitrogen deposition can enhance the inhibition of the allelochemicals secreted by S. canadensis at its initial invasive stage on seed germination and seedling growth of a native plant, Lactuca sativa. | |
| 分析94篇文章发现, 入侵植物增大了植物、土壤和土壤微生物中的碳和氮库大小. Analysis of 94 published papers show that invasive plants increase the size of carbon and nitrogen pools in plants, soils and soil microorganisms. | ||
| 与本地植物Artemisia californica、Salvia mellifera、Salvia apiana、锦鸡儿(Calochortus splendens)和Dichelostemma capitatum相比, 外来植物Centaurea stoebe、Hirschfeldia incana和Bromus madritensis可以通过加快凋落物分解改善土壤养分. Compared with native plants such as Artemisia californica, Salvia mellifera, Salvia apiana, Calochortus splendens and Dichelostemma capitatum, exotic plants, like Centaurea stoebe, Hirschfeldia incana and Bromus madritensis, can increase soil nutrient level by increasing litter decomposition. | ||
| 入侵植物Mikania micrantha分泌的化感物质绿原酸和β-石竹烯加快了土壤有机氮的矿化过程. Allelochemicals secreted by the invasive plant, Mikania micrantha, such as chlorogenic acid and β-caryophyllene accelerate mineralization of soil organic nitrogen. | ||
| 入侵植物通过凋落物改变了根际微生物土壤群落和养分循环, 形成植物-土壤正反馈调节. Invasive plants can form a positive plant-soil feedback via its litter decomposition which could change rhizosphere soil microbial communities and nutrient cycling. | ||
| 杂草Conyza canadensis入侵的土壤中有效氮和有机质含量增加. The content of available nitrogen and organic matter increases in the soil invaded by Conyza canadensis. | ||
| 与非入侵地土壤相比, Eupatorium catarium入侵后土壤氮含量提高4.32 mg·kg-1. Compared with non-invasive soils, soil nitrogen content increases by 4.32 mg·kg-1 after invasion by Eupatorium catarium. | ||
| Ambrosia trifida在入侵地的土壤-植物反馈高于原产地. Ambrosia trifida has a greater plant-soil feedback in invaded ranges than that in its native ranges. |
...
Parthenium hysterophorus can secrete parthenin, caffeic acid, vanillic acid and other allelochemicals to facilitate its invasion.
Soil nutrient variation
Nitrogen deposition can enhance the inhibition of the allelochemicals secreted by S. canadensis at its initial invasive stage on seed germination and seedling growth of a native plant, Lactuca sativa.
Analysis of 94 published papers show that invasive plants increase the size of carbon and nitrogen pools in plants, soils and soil microorganisms.
Compared with native plants such as Artemisia californica, Salvia mellifera, Salvia apiana, Calochortus splendens and Dichelostemma capitatum, exotic plants, like Centaurea stoebe, Hirschfeldia incana and Bromus madritensis, can increase soil nutrient level by increasing litter decomposition.
Allelochemicals secreted by the invasive plant, Mikania micrantha, such as chlorogenic acid and β-caryophyllene accelerate mineralization of soil organic nitrogen.
Invasive plants can form a positive plant-soil feedback via its litter decomposition which could change rhizosphere soil microbial communities and nutrient cycling.
The content of available nitrogen and organic matter increases in the soil invaded by Conyza canadensis.
Compared with non-invasive soils, soil nitrogen content increases by 4.32 mg·kg-1 after invasion by Eupatorium catarium.
Ambrosia trifida has a greater plant-soil feedback in invaded ranges than that in its native ranges.
入侵性杂草北美刺龙葵在北京市的首次发现及防除
3
2020
... 入侵植物可以通过多种生态机制影响本地植物的菌根真菌(
... Ecological mechanisms of alien plant invasion
| 生态机制 Mechanism of ecology | 举例 Example | 参考文献 Reference |
|---|---|---|
| 物种竞争 Species competition | 与农作物(如高粱(Sorghum bicolor)、玉米(Zea mays))相比, 入侵植物Parthenium hysterophorus具有快速早期生长的能力. Compared with crops (such as Sorghum bicolor and Zea mays), the invasive plant Parthenium hysterophorus can grow much more rapidly in their early stage. | |
| 比较夏威夷19对入侵植物和本地植物的资源利用效率, 发现入侵植物具有较高的碳同化率、光利用效率、瞬时氮利用效率和瞬时能量利用效率. Comparing the resource utilization efficiency for 19 paired invasive-native plants in Hawaii, invasive plants are observed to have higher carbon assimilation rate, light use efficiency, instantaneous nitrogen and energy use efficiency. | ||
| 比较47对入侵植物和非入侵植物对草食性动物的抵抗能力及养分含量发现, 入侵植物拥有更高的叶氮含量, 受到食草动物的损伤较小. Comparisons among 47 paired invasive and non-invasive plants for leaf herbivore-resistance and nutrient content show that invasive plants have higher leaf nitrogen content and less damage by herbivores. | ||
| 比对125种入侵植物和196种非入侵植物的生理性状发现, 入侵植物在生长率、养分分配及抗逆性更有优势. Comparing physiological traits of 125 invasive with plants 196 non-invasive plants show that invasive plants have more advantages in growth rate, nutrient allocation and stress resistance. | ||
| 外来植物Solanum carolinense具有较强的抗寒能力和无性繁殖能力. Exotic plant, Solanum carolinense, has strong ability for cold resistance and asexual reproduction. | ||
| 气候变暖及氮沉降的增加可以增强入侵植物Solidago canadensis叶片的资源获取能力. Climate warming and increased nitrogen deposition can enhance leaf resource acquisition of an invasive plant, Solidago canadensis. | ||
| 化感物质 Allelochemical | 入侵植物白羊草(Bothriochloa ischaemum)的渗滤液抑制本地植物Andropogon gerardii和Schizachyrium scoparium发芽及生长. The leachate of an invasive plant, Bothriochloa ischaemum, inhibits germination and growth of the native plants, Andropogon gerardii and Schizachyrium scoparium. | |
| 入侵植物水烛(Typha angustifolia)根系分泌的酚类物质抑制了本地植物Bolboschoenus fluviatilis的生长发育. Phenolic compounds secreted by roots of an invasive plant, Typha angustifolia, inhibit growth and development of a native plant, Bolboschoenus fluviatilis. | ||
| Solidago canadensis入侵分泌的化感物质抑制了本地植物莴苣(Lactuca sativa)种子萌发和幼苗生长. Allelochemicals secreted by the invasion of Solidago canadensis inhibit seed germination and seedling growth of a native plant, Lactuca sativa. | ||
| 外来植物Solanum carolinense含有龙葵碱及草酸盐结晶. The exotic plant Solanum carolinense contains solanine and oxalate crystals. | ||
| Parthenium hysterophorus可以分泌帕台单宁、咖啡酸、香草酸等化感物质帮助自身入侵. Parthenium hysterophorus can secrete parthenin, caffeic acid, vanillic acid and other allelochemicals to facilitate its invasion. | ||
| 土壤养分变化 Soil nutrient variation | 氮沉降会增强S. canadensis入侵初期分泌的化感物质对本地植物莴苣种子萌发和幼苗生长的抑制程度. Nitrogen deposition can enhance the inhibition of the allelochemicals secreted by S. canadensis at its initial invasive stage on seed germination and seedling growth of a native plant, Lactuca sativa. | |
| 分析94篇文章发现, 入侵植物增大了植物、土壤和土壤微生物中的碳和氮库大小. Analysis of 94 published papers show that invasive plants increase the size of carbon and nitrogen pools in plants, soils and soil microorganisms. | ||
| 与本地植物Artemisia californica、Salvia mellifera、Salvia apiana、锦鸡儿(Calochortus splendens)和Dichelostemma capitatum相比, 外来植物Centaurea stoebe、Hirschfeldia incana和Bromus madritensis可以通过加快凋落物分解改善土壤养分. Compared with native plants such as Artemisia californica, Salvia mellifera, Salvia apiana, Calochortus splendens and Dichelostemma capitatum, exotic plants, like Centaurea stoebe, Hirschfeldia incana and Bromus madritensis, can increase soil nutrient level by increasing litter decomposition. | ||
| 入侵植物Mikania micrantha分泌的化感物质绿原酸和β-石竹烯加快了土壤有机氮的矿化过程. Allelochemicals secreted by the invasive plant, Mikania micrantha, such as chlorogenic acid and β-caryophyllene accelerate mineralization of soil organic nitrogen. | ||
| 入侵植物通过凋落物改变了根际微生物土壤群落和养分循环, 形成植物-土壤正反馈调节. Invasive plants can form a positive plant-soil feedback via its litter decomposition which could change rhizosphere soil microbial communities and nutrient cycling. | ||
| 杂草Conyza canadensis入侵的土壤中有效氮和有机质含量增加. The content of available nitrogen and organic matter increases in the soil invaded by Conyza canadensis. | ||
| 与非入侵地土壤相比, Eupatorium catarium入侵后土壤氮含量提高4.32 mg·kg-1. Compared with non-invasive soils, soil nitrogen content increases by 4.32 mg·kg-1 after invasion by Eupatorium catarium. | ||
| Ambrosia trifida在入侵地的土壤-植物反馈高于原产地. Ambrosia trifida has a greater plant-soil feedback in invaded ranges than that in its native ranges. |
...
Parthenium hysterophorus can secrete parthenin, caffeic acid, vanillic acid and other allelochemicals to facilitate its invasion.
Soil nutrient variation
Nitrogen deposition can enhance the inhibition of the allelochemicals secreted by S. canadensis at its initial invasive stage on seed germination and seedling growth of a native plant, Lactuca sativa.
Analysis of 94 published papers show that invasive plants increase the size of carbon and nitrogen pools in plants, soils and soil microorganisms.
Compared with native plants such as Artemisia californica, Salvia mellifera, Salvia apiana, Calochortus splendens and Dichelostemma capitatum, exotic plants, like Centaurea stoebe, Hirschfeldia incana and Bromus madritensis, can increase soil nutrient level by increasing litter decomposition.
Allelochemicals secreted by the invasive plant, Mikania micrantha, such as chlorogenic acid and β-caryophyllene accelerate mineralization of soil organic nitrogen.
Invasive plants can form a positive plant-soil feedback via its litter decomposition which could change rhizosphere soil microbial communities and nutrient cycling.
The content of available nitrogen and organic matter increases in the soil invaded by Conyza canadensis.
Compared with non-invasive soils, soil nitrogen content increases by 4.32 mg·kg-1 after invasion by Eupatorium catarium.
Ambrosia trifida has a greater plant-soil feedback in invaded ranges than that in its native ranges.
Invasive warm-season grasses reduce mycorrhizal root colonization and biomass production of native prairie grasses
1
2012
... 除了这些被广泛关注的途径, 近年来还发现一类重要的间接途径, 即入侵植物能通过干扰本地物种的共生关系(如植物-传粉昆虫和根系-菌根真菌等)而抑制本地植物的生长(
The invasive plant Alliaria petiolata (garlic mustard) inhibits ectomycorrhizal fungi in its introduced range
2
2008
... 随着全球社会经济活动的日益紧密, 越来越多的植物被带入非原产地环境, 快速扩散成为入侵植物.全球变化背景下, 入侵植物已对当地生物多样性、社会经济以及人类健康造成重大威胁(
... 相对于AMF, 外生菌根真菌(EMF)对外来干扰的抵抗能力更强(
1
2019
... 上述这些研究中, 本地植物菌根真菌群落组成都是通过变性梯度凝胶电泳(DGGE)(
1
2019
... 上述这些研究中, 本地植物菌根真菌群落组成都是通过变性梯度凝胶电泳(DGGE)(
1
2018
... 上述这些研究中, 本地植物菌根真菌群落组成都是通过变性梯度凝胶电泳(DGGE)(
1
2018
... 上述这些研究中, 本地植物菌根真菌群落组成都是通过变性梯度凝胶电泳(DGGE)(
入侵植物与本地植物互作对丛枝菌根真菌AMF侵染率的影响
2019
入侵植物与本地植物互作对丛枝菌根真菌AMF侵染率的影响
2019
尖峰岭热带山地雨林根部真菌-植物互作网络结构特征
2
2019
... 同种植物常常能与多种菌根真菌共生, 即使是单个根尖也往往被多个特定的菌根真菌类群侵染(
... 发现, 干旱情况下真菌共生网络的复杂性和稳定性均有所降低; 在AMF和EMF植物共存的系统中, AMF网络与宿主植物的连接性更强, EMF网络与宿主植物的专一性(共生伙伴偏好性)更强(
尖峰岭热带山地雨林根部真菌-植物互作网络结构特征
2
2019
... 同种植物常常能与多种菌根真菌共生, 即使是单个根尖也往往被多个特定的菌根真菌类群侵染(
... 发现, 干旱情况下真菌共生网络的复杂性和稳定性均有所降低; 在AMF和EMF植物共存的系统中, AMF网络与宿主植物的连接性更强, EMF网络与宿主植物的专一性(共生伙伴偏好性)更强(
Self-reinforcing impacts of plant invasions change over time
1
2013
... 已有研究证实外来植物的成功入侵受入侵时间的影响(
外来入侵植物薇甘菊的2种化感物质对土壤氮循环的影响
1
2020
... Ecological mechanisms of alien plant invasion
| 生态机制 Mechanism of ecology | 举例 Example | 参考文献 Reference |
|---|---|---|
| 物种竞争 Species competition | 与农作物(如高粱(Sorghum bicolor)、玉米(Zea mays))相比, 入侵植物Parthenium hysterophorus具有快速早期生长的能力. Compared with crops (such as Sorghum bicolor and Zea mays), the invasive plant Parthenium hysterophorus can grow much more rapidly in their early stage. | |
| 比较夏威夷19对入侵植物和本地植物的资源利用效率, 发现入侵植物具有较高的碳同化率、光利用效率、瞬时氮利用效率和瞬时能量利用效率. Comparing the resource utilization efficiency for 19 paired invasive-native plants in Hawaii, invasive plants are observed to have higher carbon assimilation rate, light use efficiency, instantaneous nitrogen and energy use efficiency. | ||
| 比较47对入侵植物和非入侵植物对草食性动物的抵抗能力及养分含量发现, 入侵植物拥有更高的叶氮含量, 受到食草动物的损伤较小. Comparisons among 47 paired invasive and non-invasive plants for leaf herbivore-resistance and nutrient content show that invasive plants have higher leaf nitrogen content and less damage by herbivores. | ||
| 比对125种入侵植物和196种非入侵植物的生理性状发现, 入侵植物在生长率、养分分配及抗逆性更有优势. Comparing physiological traits of 125 invasive with plants 196 non-invasive plants show that invasive plants have more advantages in growth rate, nutrient allocation and stress resistance. | ||
| 外来植物Solanum carolinense具有较强的抗寒能力和无性繁殖能力. Exotic plant, Solanum carolinense, has strong ability for cold resistance and asexual reproduction. | ||
| 气候变暖及氮沉降的增加可以增强入侵植物Solidago canadensis叶片的资源获取能力. Climate warming and increased nitrogen deposition can enhance leaf resource acquisition of an invasive plant, Solidago canadensis. | ||
| 化感物质 Allelochemical | 入侵植物白羊草(Bothriochloa ischaemum)的渗滤液抑制本地植物Andropogon gerardii和Schizachyrium scoparium发芽及生长. The leachate of an invasive plant, Bothriochloa ischaemum, inhibits germination and growth of the native plants, Andropogon gerardii and Schizachyrium scoparium. | |
| 入侵植物水烛(Typha angustifolia)根系分泌的酚类物质抑制了本地植物Bolboschoenus fluviatilis的生长发育. Phenolic compounds secreted by roots of an invasive plant, Typha angustifolia, inhibit growth and development of a native plant, Bolboschoenus fluviatilis. | ||
| Solidago canadensis入侵分泌的化感物质抑制了本地植物莴苣(Lactuca sativa)种子萌发和幼苗生长. Allelochemicals secreted by the invasion of Solidago canadensis inhibit seed germination and seedling growth of a native plant, Lactuca sativa. | ||
| 外来植物Solanum carolinense含有龙葵碱及草酸盐结晶. The exotic plant Solanum carolinense contains solanine and oxalate crystals. | ||
| Parthenium hysterophorus可以分泌帕台单宁、咖啡酸、香草酸等化感物质帮助自身入侵. Parthenium hysterophorus can secrete parthenin, caffeic acid, vanillic acid and other allelochemicals to facilitate its invasion. | ||
| 土壤养分变化 Soil nutrient variation | 氮沉降会增强S. canadensis入侵初期分泌的化感物质对本地植物莴苣种子萌发和幼苗生长的抑制程度. Nitrogen deposition can enhance the inhibition of the allelochemicals secreted by S. canadensis at its initial invasive stage on seed germination and seedling growth of a native plant, Lactuca sativa. | |
| 分析94篇文章发现, 入侵植物增大了植物、土壤和土壤微生物中的碳和氮库大小. Analysis of 94 published papers show that invasive plants increase the size of carbon and nitrogen pools in plants, soils and soil microorganisms. | ||
| 与本地植物Artemisia californica、Salvia mellifera、Salvia apiana、锦鸡儿(Calochortus splendens)和Dichelostemma capitatum相比, 外来植物Centaurea stoebe、Hirschfeldia incana和Bromus madritensis可以通过加快凋落物分解改善土壤养分. Compared with native plants such as Artemisia californica, Salvia mellifera, Salvia apiana, Calochortus splendens and Dichelostemma capitatum, exotic plants, like Centaurea stoebe, Hirschfeldia incana and Bromus madritensis, can increase soil nutrient level by increasing litter decomposition. | ||
| 入侵植物Mikania micrantha分泌的化感物质绿原酸和β-石竹烯加快了土壤有机氮的矿化过程. Allelochemicals secreted by the invasive plant, Mikania micrantha, such as chlorogenic acid and β-caryophyllene accelerate mineralization of soil organic nitrogen. | ||
| 入侵植物通过凋落物改变了根际微生物土壤群落和养分循环, 形成植物-土壤正反馈调节. Invasive plants can form a positive plant-soil feedback via its litter decomposition which could change rhizosphere soil microbial communities and nutrient cycling. | ||
| 杂草Conyza canadensis入侵的土壤中有效氮和有机质含量增加. The content of available nitrogen and organic matter increases in the soil invaded by Conyza canadensis. | ||
| 与非入侵地土壤相比, Eupatorium catarium入侵后土壤氮含量提高4.32 mg·kg-1. Compared with non-invasive soils, soil nitrogen content increases by 4.32 mg·kg-1 after invasion by Eupatorium catarium. | ||
| Ambrosia trifida在入侵地的土壤-植物反馈高于原产地. Ambrosia trifida has a greater plant-soil feedback in invaded ranges than that in its native ranges. |
外来入侵植物薇甘菊的2种化感物质对土壤氮循环的影响
1
2020
... Ecological mechanisms of alien plant invasion
| 生态机制 Mechanism of ecology | 举例 Example | 参考文献 Reference |
|---|---|---|
| 物种竞争 Species competition | 与农作物(如高粱(Sorghum bicolor)、玉米(Zea mays))相比, 入侵植物Parthenium hysterophorus具有快速早期生长的能力. Compared with crops (such as Sorghum bicolor and Zea mays), the invasive plant Parthenium hysterophorus can grow much more rapidly in their early stage. | |
| 比较夏威夷19对入侵植物和本地植物的资源利用效率, 发现入侵植物具有较高的碳同化率、光利用效率、瞬时氮利用效率和瞬时能量利用效率. Comparing the resource utilization efficiency for 19 paired invasive-native plants in Hawaii, invasive plants are observed to have higher carbon assimilation rate, light use efficiency, instantaneous nitrogen and energy use efficiency. | ||
| 比较47对入侵植物和非入侵植物对草食性动物的抵抗能力及养分含量发现, 入侵植物拥有更高的叶氮含量, 受到食草动物的损伤较小. Comparisons among 47 paired invasive and non-invasive plants for leaf herbivore-resistance and nutrient content show that invasive plants have higher leaf nitrogen content and less damage by herbivores. | ||
| 比对125种入侵植物和196种非入侵植物的生理性状发现, 入侵植物在生长率、养分分配及抗逆性更有优势. Comparing physiological traits of 125 invasive with plants 196 non-invasive plants show that invasive plants have more advantages in growth rate, nutrient allocation and stress resistance. | ||
| 外来植物Solanum carolinense具有较强的抗寒能力和无性繁殖能力. Exotic plant, Solanum carolinense, has strong ability for cold resistance and asexual reproduction. | ||
| 气候变暖及氮沉降的增加可以增强入侵植物Solidago canadensis叶片的资源获取能力. Climate warming and increased nitrogen deposition can enhance leaf resource acquisition of an invasive plant, Solidago canadensis. | ||
| 化感物质 Allelochemical | 入侵植物白羊草(Bothriochloa ischaemum)的渗滤液抑制本地植物Andropogon gerardii和Schizachyrium scoparium发芽及生长. The leachate of an invasive plant, Bothriochloa ischaemum, inhibits germination and growth of the native plants, Andropogon gerardii and Schizachyrium scoparium. | |
| 入侵植物水烛(Typha angustifolia)根系分泌的酚类物质抑制了本地植物Bolboschoenus fluviatilis的生长发育. Phenolic compounds secreted by roots of an invasive plant, Typha angustifolia, inhibit growth and development of a native plant, Bolboschoenus fluviatilis. | ||
| Solidago canadensis入侵分泌的化感物质抑制了本地植物莴苣(Lactuca sativa)种子萌发和幼苗生长. Allelochemicals secreted by the invasion of Solidago canadensis inhibit seed germination and seedling growth of a native plant, Lactuca sativa. | ||
| 外来植物Solanum carolinense含有龙葵碱及草酸盐结晶. The exotic plant Solanum carolinense contains solanine and oxalate crystals. | ||
| Parthenium hysterophorus可以分泌帕台单宁、咖啡酸、香草酸等化感物质帮助自身入侵. Parthenium hysterophorus can secrete parthenin, caffeic acid, vanillic acid and other allelochemicals to facilitate its invasion. | ||
| 土壤养分变化 Soil nutrient variation | 氮沉降会增强S. canadensis入侵初期分泌的化感物质对本地植物莴苣种子萌发和幼苗生长的抑制程度. Nitrogen deposition can enhance the inhibition of the allelochemicals secreted by S. canadensis at its initial invasive stage on seed germination and seedling growth of a native plant, Lactuca sativa. | |
| 分析94篇文章发现, 入侵植物增大了植物、土壤和土壤微生物中的碳和氮库大小. Analysis of 94 published papers show that invasive plants increase the size of carbon and nitrogen pools in plants, soils and soil microorganisms. | ||
| 与本地植物Artemisia californica、Salvia mellifera、Salvia apiana、锦鸡儿(Calochortus splendens)和Dichelostemma capitatum相比, 外来植物Centaurea stoebe、Hirschfeldia incana和Bromus madritensis可以通过加快凋落物分解改善土壤养分. Compared with native plants such as Artemisia californica, Salvia mellifera, Salvia apiana, Calochortus splendens and Dichelostemma capitatum, exotic plants, like Centaurea stoebe, Hirschfeldia incana and Bromus madritensis, can increase soil nutrient level by increasing litter decomposition. | ||
| 入侵植物Mikania micrantha分泌的化感物质绿原酸和β-石竹烯加快了土壤有机氮的矿化过程. Allelochemicals secreted by the invasive plant, Mikania micrantha, such as chlorogenic acid and β-caryophyllene accelerate mineralization of soil organic nitrogen. | ||
| 入侵植物通过凋落物改变了根际微生物土壤群落和养分循环, 形成植物-土壤正反馈调节. Invasive plants can form a positive plant-soil feedback via its litter decomposition which could change rhizosphere soil microbial communities and nutrient cycling. | ||
| 杂草Conyza canadensis入侵的土壤中有效氮和有机质含量增加. The content of available nitrogen and organic matter increases in the soil invaded by Conyza canadensis. | ||
| 与非入侵地土壤相比, Eupatorium catarium入侵后土壤氮含量提高4.32 mg·kg-1. Compared with non-invasive soils, soil nitrogen content increases by 4.32 mg·kg-1 after invasion by Eupatorium catarium. | ||
| Ambrosia trifida在入侵地的土壤-植物反馈高于原产地. Ambrosia trifida has a greater plant-soil feedback in invaded ranges than that in its native ranges. |
Arbuscular mycorrhizal fungi facilitate growth and competitive ability of an exotic species Flaveria bidentis
2017
AM fungi facilitate the competitive growth of two invasive plant species, Ambrosia artemisiifolia and Bidens pilosa
1
2018
... 外来植物入侵后, 本地植物菌根真菌侵染率有时没有受到显著影响(
外来入侵植物与地下生态系统相互影响的研究进展
1
2009
... 除了这些被广泛关注的途径, 近年来还发现一类重要的间接途径, 即入侵植物能通过干扰本地物种的共生关系(如植物-传粉昆虫和根系-菌根真菌等)而抑制本地植物的生长(
外来入侵植物与地下生态系统相互影响的研究进展
1
2009
... 除了这些被广泛关注的途径, 近年来还发现一类重要的间接途径, 即入侵植物能通过干扰本地物种的共生关系(如植物-传粉昆虫和根系-菌根真菌等)而抑制本地植物的生长(
6
2019
... 尽管入侵植物对本地植物菌根真菌影响的这些差异性被报道, 但是很少有研究者关注这些差异背后的原因.我们推测这可能至少与以下几个方面的原因有关: 1)本地植物和菌根真菌共生关系的强弱.例如, 有两个本地植物A和B, 如果A的菌根真菌侵染率远高于B (或者说A对菌根真菌的依赖性远超过B)(
... )菌根真菌的影响反而较小, 这可能是由于入侵种的化感物质较弱.3)土壤养分的高低.例如,
... 以往研究大多只关注具有单共生关系的本地植物(只受菌根真菌侵染), 而对具有双共生关系的本地植物(根瘤菌和AMF共同侵染, 如豆科植物)的研究很少.我们团队的研究发现, 入侵植物X. strumarium抑制了本地植物苍耳(单共生关系)的AMF侵染率(
... 菌根的根内菌丝延伸至根外形成根外菌丝, 根外菌丝直接参与土壤养分的吸收.外来入侵植物在对本地植物菌根真菌侵染率、真菌内部结构产生影响的同时, 也会对根外菌丝产生影响(
... 入侵植物除了影响本地植物菌根真菌的侵染率、根内和根外菌丝的结构与功能外, 还可能改变其菌根真菌的组成.有研究发现, 外来植物入侵会使本地植物原有的AMF被其他种类的AMF替代(
... 同种植物常常能与多种菌根真菌共生, 即使是单个根尖也往往被多个特定的菌根真菌类群侵染(
6
2019
... 尽管入侵植物对本地植物菌根真菌影响的这些差异性被报道, 但是很少有研究者关注这些差异背后的原因.我们推测这可能至少与以下几个方面的原因有关: 1)本地植物和菌根真菌共生关系的强弱.例如, 有两个本地植物A和B, 如果A的菌根真菌侵染率远高于B (或者说A对菌根真菌的依赖性远超过B)(
... )菌根真菌的影响反而较小, 这可能是由于入侵种的化感物质较弱.3)土壤养分的高低.例如,
... 以往研究大多只关注具有单共生关系的本地植物(只受菌根真菌侵染), 而对具有双共生关系的本地植物(根瘤菌和AMF共同侵染, 如豆科植物)的研究很少.我们团队的研究发现, 入侵植物X. strumarium抑制了本地植物苍耳(单共生关系)的AMF侵染率(
... 菌根的根内菌丝延伸至根外形成根外菌丝, 根外菌丝直接参与土壤养分的吸收.外来入侵植物在对本地植物菌根真菌侵染率、真菌内部结构产生影响的同时, 也会对根外菌丝产生影响(
... 入侵植物除了影响本地植物菌根真菌的侵染率、根内和根外菌丝的结构与功能外, 还可能改变其菌根真菌的组成.有研究发现, 外来植物入侵会使本地植物原有的AMF被其他种类的AMF替代(
... 同种植物常常能与多种菌根真菌共生, 即使是单个根尖也往往被多个特定的菌根真菌类群侵染(
Invasion by the weed Conyza canadensis alters soil nutrient supply and shifts microbiota structure
2
2020
... 入侵植物可以通过多种生态机制影响本地植物的菌根真菌(
... Ecological mechanisms of alien plant invasion
| 生态机制 Mechanism of ecology | 举例 Example | 参考文献 Reference |
|---|---|---|
| 物种竞争 Species competition | 与农作物(如高粱(Sorghum bicolor)、玉米(Zea mays))相比, 入侵植物Parthenium hysterophorus具有快速早期生长的能力. Compared with crops (such as Sorghum bicolor and Zea mays), the invasive plant Parthenium hysterophorus can grow much more rapidly in their early stage. | |
| 比较夏威夷19对入侵植物和本地植物的资源利用效率, 发现入侵植物具有较高的碳同化率、光利用效率、瞬时氮利用效率和瞬时能量利用效率. Comparing the resource utilization efficiency for 19 paired invasive-native plants in Hawaii, invasive plants are observed to have higher carbon assimilation rate, light use efficiency, instantaneous nitrogen and energy use efficiency. | ||
| 比较47对入侵植物和非入侵植物对草食性动物的抵抗能力及养分含量发现, 入侵植物拥有更高的叶氮含量, 受到食草动物的损伤较小. Comparisons among 47 paired invasive and non-invasive plants for leaf herbivore-resistance and nutrient content show that invasive plants have higher leaf nitrogen content and less damage by herbivores. | ||
| 比对125种入侵植物和196种非入侵植物的生理性状发现, 入侵植物在生长率、养分分配及抗逆性更有优势. Comparing physiological traits of 125 invasive with plants 196 non-invasive plants show that invasive plants have more advantages in growth rate, nutrient allocation and stress resistance. | ||
| 外来植物Solanum carolinense具有较强的抗寒能力和无性繁殖能力. Exotic plant, Solanum carolinense, has strong ability for cold resistance and asexual reproduction. | ||
| 气候变暖及氮沉降的增加可以增强入侵植物Solidago canadensis叶片的资源获取能力. Climate warming and increased nitrogen deposition can enhance leaf resource acquisition of an invasive plant, Solidago canadensis. | ||
| 化感物质 Allelochemical | 入侵植物白羊草(Bothriochloa ischaemum)的渗滤液抑制本地植物Andropogon gerardii和Schizachyrium scoparium发芽及生长. The leachate of an invasive plant, Bothriochloa ischaemum, inhibits germination and growth of the native plants, Andropogon gerardii and Schizachyrium scoparium. | |
| 入侵植物水烛(Typha angustifolia)根系分泌的酚类物质抑制了本地植物Bolboschoenus fluviatilis的生长发育. Phenolic compounds secreted by roots of an invasive plant, Typha angustifolia, inhibit growth and development of a native plant, Bolboschoenus fluviatilis. | ||
| Solidago canadensis入侵分泌的化感物质抑制了本地植物莴苣(Lactuca sativa)种子萌发和幼苗生长. Allelochemicals secreted by the invasion of Solidago canadensis inhibit seed germination and seedling growth of a native plant, Lactuca sativa. | ||
| 外来植物Solanum carolinense含有龙葵碱及草酸盐结晶. The exotic plant Solanum carolinense contains solanine and oxalate crystals. | ||
| Parthenium hysterophorus可以分泌帕台单宁、咖啡酸、香草酸等化感物质帮助自身入侵. Parthenium hysterophorus can secrete parthenin, caffeic acid, vanillic acid and other allelochemicals to facilitate its invasion. | ||
| 土壤养分变化 Soil nutrient variation | 氮沉降会增强S. canadensis入侵初期分泌的化感物质对本地植物莴苣种子萌发和幼苗生长的抑制程度. Nitrogen deposition can enhance the inhibition of the allelochemicals secreted by S. canadensis at its initial invasive stage on seed germination and seedling growth of a native plant, Lactuca sativa. | |
| 分析94篇文章发现, 入侵植物增大了植物、土壤和土壤微生物中的碳和氮库大小. Analysis of 94 published papers show that invasive plants increase the size of carbon and nitrogen pools in plants, soils and soil microorganisms. | ||
| 与本地植物Artemisia californica、Salvia mellifera、Salvia apiana、锦鸡儿(Calochortus splendens)和Dichelostemma capitatum相比, 外来植物Centaurea stoebe、Hirschfeldia incana和Bromus madritensis可以通过加快凋落物分解改善土壤养分. Compared with native plants such as Artemisia californica, Salvia mellifera, Salvia apiana, Calochortus splendens and Dichelostemma capitatum, exotic plants, like Centaurea stoebe, Hirschfeldia incana and Bromus madritensis, can increase soil nutrient level by increasing litter decomposition. | ||
| 入侵植物Mikania micrantha分泌的化感物质绿原酸和β-石竹烯加快了土壤有机氮的矿化过程. Allelochemicals secreted by the invasive plant, Mikania micrantha, such as chlorogenic acid and β-caryophyllene accelerate mineralization of soil organic nitrogen. | ||
| 入侵植物通过凋落物改变了根际微生物土壤群落和养分循环, 形成植物-土壤正反馈调节. Invasive plants can form a positive plant-soil feedback via its litter decomposition which could change rhizosphere soil microbial communities and nutrient cycling. | ||
| 杂草Conyza canadensis入侵的土壤中有效氮和有机质含量增加. The content of available nitrogen and organic matter increases in the soil invaded by Conyza canadensis. | ||
| 与非入侵地土壤相比, Eupatorium catarium入侵后土壤氮含量提高4.32 mg·kg-1. Compared with non-invasive soils, soil nitrogen content increases by 4.32 mg·kg-1 after invasion by Eupatorium catarium. | ||
| Ambrosia trifida在入侵地的土壤-植物反馈高于原产地. Ambrosia trifida has a greater plant-soil feedback in invaded ranges than that in its native ranges. |
Signal beyond nutrient, fructose, exuded by an arbuscular mycorrhizal fungus triggers phytate mineralization by a phosphate solubilizing bacterium
2
2018
... 菌根的根内菌丝延伸至根外形成根外菌丝, 根外菌丝直接参与土壤养分的吸收.外来入侵植物在对本地植物菌根真菌侵染率、真菌内部结构产生影响的同时, 也会对根外菌丝产生影响(
... 入侵植物可以通过多种生态机制影响本地植物的菌根真菌(
Invasive plants differentially affect soil biota through litter and rhizosphere pathways: a meta-analysis
4
2018
... 入侵植物除了影响本地植物菌根真菌的侵染率、根内和根外菌丝的结构与功能外, 还可能改变其菌根真菌的组成.有研究发现, 外来植物入侵会使本地植物原有的AMF被其他种类的AMF替代(
... 同种植物常常能与多种菌根真菌共生, 即使是单个根尖也往往被多个特定的菌根真菌类群侵染(
... 入侵植物可以通过多种生态机制影响本地植物的菌根真菌(
... Ecological mechanisms of alien plant invasion
| 生态机制 Mechanism of ecology | 举例 Example | 参考文献 Reference |
|---|---|---|
| 物种竞争 Species competition | 与农作物(如高粱(Sorghum bicolor)、玉米(Zea mays))相比, 入侵植物Parthenium hysterophorus具有快速早期生长的能力. Compared with crops (such as Sorghum bicolor and Zea mays), the invasive plant Parthenium hysterophorus can grow much more rapidly in their early stage. | |
| 比较夏威夷19对入侵植物和本地植物的资源利用效率, 发现入侵植物具有较高的碳同化率、光利用效率、瞬时氮利用效率和瞬时能量利用效率. Comparing the resource utilization efficiency for 19 paired invasive-native plants in Hawaii, invasive plants are observed to have higher carbon assimilation rate, light use efficiency, instantaneous nitrogen and energy use efficiency. | ||
| 比较47对入侵植物和非入侵植物对草食性动物的抵抗能力及养分含量发现, 入侵植物拥有更高的叶氮含量, 受到食草动物的损伤较小. Comparisons among 47 paired invasive and non-invasive plants for leaf herbivore-resistance and nutrient content show that invasive plants have higher leaf nitrogen content and less damage by herbivores. | ||
| 比对125种入侵植物和196种非入侵植物的生理性状发现, 入侵植物在生长率、养分分配及抗逆性更有优势. Comparing physiological traits of 125 invasive with plants 196 non-invasive plants show that invasive plants have more advantages in growth rate, nutrient allocation and stress resistance. | ||
| 外来植物Solanum carolinense具有较强的抗寒能力和无性繁殖能力. Exotic plant, Solanum carolinense, has strong ability for cold resistance and asexual reproduction. | ||
| 气候变暖及氮沉降的增加可以增强入侵植物Solidago canadensis叶片的资源获取能力. Climate warming and increased nitrogen deposition can enhance leaf resource acquisition of an invasive plant, Solidago canadensis. | ||
| 化感物质 Allelochemical | 入侵植物白羊草(Bothriochloa ischaemum)的渗滤液抑制本地植物Andropogon gerardii和Schizachyrium scoparium发芽及生长. The leachate of an invasive plant, Bothriochloa ischaemum, inhibits germination and growth of the native plants, Andropogon gerardii and Schizachyrium scoparium. | |
| 入侵植物水烛(Typha angustifolia)根系分泌的酚类物质抑制了本地植物Bolboschoenus fluviatilis的生长发育. Phenolic compounds secreted by roots of an invasive plant, Typha angustifolia, inhibit growth and development of a native plant, Bolboschoenus fluviatilis. | ||
| Solidago canadensis入侵分泌的化感物质抑制了本地植物莴苣(Lactuca sativa)种子萌发和幼苗生长. Allelochemicals secreted by the invasion of Solidago canadensis inhibit seed germination and seedling growth of a native plant, Lactuca sativa. | ||
| 外来植物Solanum carolinense含有龙葵碱及草酸盐结晶. The exotic plant Solanum carolinense contains solanine and oxalate crystals. | ||
| Parthenium hysterophorus可以分泌帕台单宁、咖啡酸、香草酸等化感物质帮助自身入侵. Parthenium hysterophorus can secrete parthenin, caffeic acid, vanillic acid and other allelochemicals to facilitate its invasion. | ||
| 土壤养分变化 Soil nutrient variation | 氮沉降会增强S. canadensis入侵初期分泌的化感物质对本地植物莴苣种子萌发和幼苗生长的抑制程度. Nitrogen deposition can enhance the inhibition of the allelochemicals secreted by S. canadensis at its initial invasive stage on seed germination and seedling growth of a native plant, Lactuca sativa. | |
| 分析94篇文章发现, 入侵植物增大了植物、土壤和土壤微生物中的碳和氮库大小. Analysis of 94 published papers show that invasive plants increase the size of carbon and nitrogen pools in plants, soils and soil microorganisms. | ||
| 与本地植物Artemisia californica、Salvia mellifera、Salvia apiana、锦鸡儿(Calochortus splendens)和Dichelostemma capitatum相比, 外来植物Centaurea stoebe、Hirschfeldia incana和Bromus madritensis可以通过加快凋落物分解改善土壤养分. Compared with native plants such as Artemisia californica, Salvia mellifera, Salvia apiana, Calochortus splendens and Dichelostemma capitatum, exotic plants, like Centaurea stoebe, Hirschfeldia incana and Bromus madritensis, can increase soil nutrient level by increasing litter decomposition. | ||
| 入侵植物Mikania micrantha分泌的化感物质绿原酸和β-石竹烯加快了土壤有机氮的矿化过程. Allelochemicals secreted by the invasive plant, Mikania micrantha, such as chlorogenic acid and β-caryophyllene accelerate mineralization of soil organic nitrogen. | ||
| 入侵植物通过凋落物改变了根际微生物土壤群落和养分循环, 形成植物-土壤正反馈调节. Invasive plants can form a positive plant-soil feedback via its litter decomposition which could change rhizosphere soil microbial communities and nutrient cycling. | ||
| 杂草Conyza canadensis入侵的土壤中有效氮和有机质含量增加. The content of available nitrogen and organic matter increases in the soil invaded by Conyza canadensis. | ||
| 与非入侵地土壤相比, Eupatorium catarium入侵后土壤氮含量提高4.32 mg·kg-1. Compared with non-invasive soils, soil nitrogen content increases by 4.32 mg·kg-1 after invasion by Eupatorium catarium. | ||
| Ambrosia trifida在入侵地的土壤-植物反馈高于原产地. Ambrosia trifida has a greater plant-soil feedback in invaded ranges than that in its native ranges. |
The receptor kinase CERK1 has dual functions in symbiosis and immunity signalling
1
2015
... 植物与菌根真菌形成共生关系是一个复杂的信号转导过程.目前已经证实, 丛枝菌根的形成始于根系与AMF之间的信号交流(
Plant nitrogen acquisition from inorganic and organic sources via root and mycelia pathways in ectomycorrhizal alpine forests
2019
Ageratina adenophora invasions are associated with microbially mediated differences in biogeochemical cycles
2
2019
... 入侵植物可以通过多种生态机制影响本地植物的菌根真菌(
... Ecological mechanisms of alien plant invasion
| 生态机制 Mechanism of ecology | 举例 Example | 参考文献 Reference |
|---|---|---|
| 物种竞争 Species competition | 与农作物(如高粱(Sorghum bicolor)、玉米(Zea mays))相比, 入侵植物Parthenium hysterophorus具有快速早期生长的能力. Compared with crops (such as Sorghum bicolor and Zea mays), the invasive plant Parthenium hysterophorus can grow much more rapidly in their early stage. | |
| 比较夏威夷19对入侵植物和本地植物的资源利用效率, 发现入侵植物具有较高的碳同化率、光利用效率、瞬时氮利用效率和瞬时能量利用效率. Comparing the resource utilization efficiency for 19 paired invasive-native plants in Hawaii, invasive plants are observed to have higher carbon assimilation rate, light use efficiency, instantaneous nitrogen and energy use efficiency. | ||
| 比较47对入侵植物和非入侵植物对草食性动物的抵抗能力及养分含量发现, 入侵植物拥有更高的叶氮含量, 受到食草动物的损伤较小. Comparisons among 47 paired invasive and non-invasive plants for leaf herbivore-resistance and nutrient content show that invasive plants have higher leaf nitrogen content and less damage by herbivores. | ||
| 比对125种入侵植物和196种非入侵植物的生理性状发现, 入侵植物在生长率、养分分配及抗逆性更有优势. Comparing physiological traits of 125 invasive with plants 196 non-invasive plants show that invasive plants have more advantages in growth rate, nutrient allocation and stress resistance. | ||
| 外来植物Solanum carolinense具有较强的抗寒能力和无性繁殖能力. Exotic plant, Solanum carolinense, has strong ability for cold resistance and asexual reproduction. | ||
| 气候变暖及氮沉降的增加可以增强入侵植物Solidago canadensis叶片的资源获取能力. Climate warming and increased nitrogen deposition can enhance leaf resource acquisition of an invasive plant, Solidago canadensis. | ||
| 化感物质 Allelochemical | 入侵植物白羊草(Bothriochloa ischaemum)的渗滤液抑制本地植物Andropogon gerardii和Schizachyrium scoparium发芽及生长. The leachate of an invasive plant, Bothriochloa ischaemum, inhibits germination and growth of the native plants, Andropogon gerardii and Schizachyrium scoparium. | |
| 入侵植物水烛(Typha angustifolia)根系分泌的酚类物质抑制了本地植物Bolboschoenus fluviatilis的生长发育. Phenolic compounds secreted by roots of an invasive plant, Typha angustifolia, inhibit growth and development of a native plant, Bolboschoenus fluviatilis. | ||
| Solidago canadensis入侵分泌的化感物质抑制了本地植物莴苣(Lactuca sativa)种子萌发和幼苗生长. Allelochemicals secreted by the invasion of Solidago canadensis inhibit seed germination and seedling growth of a native plant, Lactuca sativa. | ||
| 外来植物Solanum carolinense含有龙葵碱及草酸盐结晶. The exotic plant Solanum carolinense contains solanine and oxalate crystals. | ||
| Parthenium hysterophorus可以分泌帕台单宁、咖啡酸、香草酸等化感物质帮助自身入侵. Parthenium hysterophorus can secrete parthenin, caffeic acid, vanillic acid and other allelochemicals to facilitate its invasion. | ||
| 土壤养分变化 Soil nutrient variation | 氮沉降会增强S. canadensis入侵初期分泌的化感物质对本地植物莴苣种子萌发和幼苗生长的抑制程度. Nitrogen deposition can enhance the inhibition of the allelochemicals secreted by S. canadensis at its initial invasive stage on seed germination and seedling growth of a native plant, Lactuca sativa. | |
| 分析94篇文章发现, 入侵植物增大了植物、土壤和土壤微生物中的碳和氮库大小. Analysis of 94 published papers show that invasive plants increase the size of carbon and nitrogen pools in plants, soils and soil microorganisms. | ||
| 与本地植物Artemisia californica、Salvia mellifera、Salvia apiana、锦鸡儿(Calochortus splendens)和Dichelostemma capitatum相比, 外来植物Centaurea stoebe、Hirschfeldia incana和Bromus madritensis可以通过加快凋落物分解改善土壤养分. Compared with native plants such as Artemisia californica, Salvia mellifera, Salvia apiana, Calochortus splendens and Dichelostemma capitatum, exotic plants, like Centaurea stoebe, Hirschfeldia incana and Bromus madritensis, can increase soil nutrient level by increasing litter decomposition. | ||
| 入侵植物Mikania micrantha分泌的化感物质绿原酸和β-石竹烯加快了土壤有机氮的矿化过程. Allelochemicals secreted by the invasive plant, Mikania micrantha, such as chlorogenic acid and β-caryophyllene accelerate mineralization of soil organic nitrogen. | ||
| 入侵植物通过凋落物改变了根际微生物土壤群落和养分循环, 形成植物-土壤正反馈调节. Invasive plants can form a positive plant-soil feedback via its litter decomposition which could change rhizosphere soil microbial communities and nutrient cycling. | ||
| 杂草Conyza canadensis入侵的土壤中有效氮和有机质含量增加. The content of available nitrogen and organic matter increases in the soil invaded by Conyza canadensis. | ||
| 与非入侵地土壤相比, Eupatorium catarium入侵后土壤氮含量提高4.32 mg·kg-1. Compared with non-invasive soils, soil nitrogen content increases by 4.32 mg·kg-1 after invasion by Eupatorium catarium. | ||
| Ambrosia trifida在入侵地的土壤-植物反馈高于原产地. Ambrosia trifida has a greater plant-soil feedback in invaded ranges than that in its native ranges. |
Release from below- and aboveground natural enemies contributes to invasion success of a temperate invader
3
2020
... 随着全球社会经济活动的日益紧密, 越来越多的植物被带入非原产地环境, 快速扩散成为入侵植物.全球变化背景下, 入侵植物已对当地生物多样性、社会经济以及人类健康造成重大威胁(
... 入侵植物可以通过多种生态机制影响本地植物的菌根真菌(
... Ecological mechanisms of alien plant invasion
| 生态机制 Mechanism of ecology | 举例 Example | 参考文献 Reference |
|---|---|---|
| 物种竞争 Species competition | 与农作物(如高粱(Sorghum bicolor)、玉米(Zea mays))相比, 入侵植物Parthenium hysterophorus具有快速早期生长的能力. Compared with crops (such as Sorghum bicolor and Zea mays), the invasive plant Parthenium hysterophorus can grow much more rapidly in their early stage. | |
| 比较夏威夷19对入侵植物和本地植物的资源利用效率, 发现入侵植物具有较高的碳同化率、光利用效率、瞬时氮利用效率和瞬时能量利用效率. Comparing the resource utilization efficiency for 19 paired invasive-native plants in Hawaii, invasive plants are observed to have higher carbon assimilation rate, light use efficiency, instantaneous nitrogen and energy use efficiency. | ||
| 比较47对入侵植物和非入侵植物对草食性动物的抵抗能力及养分含量发现, 入侵植物拥有更高的叶氮含量, 受到食草动物的损伤较小. Comparisons among 47 paired invasive and non-invasive plants for leaf herbivore-resistance and nutrient content show that invasive plants have higher leaf nitrogen content and less damage by herbivores. | ||
| 比对125种入侵植物和196种非入侵植物的生理性状发现, 入侵植物在生长率、养分分配及抗逆性更有优势. Comparing physiological traits of 125 invasive with plants 196 non-invasive plants show that invasive plants have more advantages in growth rate, nutrient allocation and stress resistance. | ||
| 外来植物Solanum carolinense具有较强的抗寒能力和无性繁殖能力. Exotic plant, Solanum carolinense, has strong ability for cold resistance and asexual reproduction. | ||
| 气候变暖及氮沉降的增加可以增强入侵植物Solidago canadensis叶片的资源获取能力. Climate warming and increased nitrogen deposition can enhance leaf resource acquisition of an invasive plant, Solidago canadensis. | ||
| 化感物质 Allelochemical | 入侵植物白羊草(Bothriochloa ischaemum)的渗滤液抑制本地植物Andropogon gerardii和Schizachyrium scoparium发芽及生长. The leachate of an invasive plant, Bothriochloa ischaemum, inhibits germination and growth of the native plants, Andropogon gerardii and Schizachyrium scoparium. | |
| 入侵植物水烛(Typha angustifolia)根系分泌的酚类物质抑制了本地植物Bolboschoenus fluviatilis的生长发育. Phenolic compounds secreted by roots of an invasive plant, Typha angustifolia, inhibit growth and development of a native plant, Bolboschoenus fluviatilis. | ||
| Solidago canadensis入侵分泌的化感物质抑制了本地植物莴苣(Lactuca sativa)种子萌发和幼苗生长. Allelochemicals secreted by the invasion of Solidago canadensis inhibit seed germination and seedling growth of a native plant, Lactuca sativa. | ||
| 外来植物Solanum carolinense含有龙葵碱及草酸盐结晶. The exotic plant Solanum carolinense contains solanine and oxalate crystals. | ||
| Parthenium hysterophorus可以分泌帕台单宁、咖啡酸、香草酸等化感物质帮助自身入侵. Parthenium hysterophorus can secrete parthenin, caffeic acid, vanillic acid and other allelochemicals to facilitate its invasion. | ||
| 土壤养分变化 Soil nutrient variation | 氮沉降会增强S. canadensis入侵初期分泌的化感物质对本地植物莴苣种子萌发和幼苗生长的抑制程度. Nitrogen deposition can enhance the inhibition of the allelochemicals secreted by S. canadensis at its initial invasive stage on seed germination and seedling growth of a native plant, Lactuca sativa. | |
| 分析94篇文章发现, 入侵植物增大了植物、土壤和土壤微生物中的碳和氮库大小. Analysis of 94 published papers show that invasive plants increase the size of carbon and nitrogen pools in plants, soils and soil microorganisms. | ||
| 与本地植物Artemisia californica、Salvia mellifera、Salvia apiana、锦鸡儿(Calochortus splendens)和Dichelostemma capitatum相比, 外来植物Centaurea stoebe、Hirschfeldia incana和Bromus madritensis可以通过加快凋落物分解改善土壤养分. Compared with native plants such as Artemisia californica, Salvia mellifera, Salvia apiana, Calochortus splendens and Dichelostemma capitatum, exotic plants, like Centaurea stoebe, Hirschfeldia incana and Bromus madritensis, can increase soil nutrient level by increasing litter decomposition. | ||
| 入侵植物Mikania micrantha分泌的化感物质绿原酸和β-石竹烯加快了土壤有机氮的矿化过程. Allelochemicals secreted by the invasive plant, Mikania micrantha, such as chlorogenic acid and β-caryophyllene accelerate mineralization of soil organic nitrogen. | ||
| 入侵植物通过凋落物改变了根际微生物土壤群落和养分循环, 形成植物-土壤正反馈调节. Invasive plants can form a positive plant-soil feedback via its litter decomposition which could change rhizosphere soil microbial communities and nutrient cycling. | ||
| 杂草Conyza canadensis入侵的土壤中有效氮和有机质含量增加. The content of available nitrogen and organic matter increases in the soil invaded by Conyza canadensis. | ||
| 与非入侵地土壤相比, Eupatorium catarium入侵后土壤氮含量提高4.32 mg·kg-1. Compared with non-invasive soils, soil nitrogen content increases by 4.32 mg·kg-1 after invasion by Eupatorium catarium. | ||
| Ambrosia trifida在入侵地的土壤-植物反馈高于原产地. Ambrosia trifida has a greater plant-soil feedback in invaded ranges than that in its native ranges. |
Integrating novel chemical weapons and evolutionarily increased competitive ability in success of a tropical invader
2
2015
... 随着全球社会经济活动的日益紧密, 越来越多的植物被带入非原产地环境, 快速扩散成为入侵植物.全球变化背景下, 入侵植物已对当地生物多样性、社会经济以及人类健康造成重大威胁(
... 入侵植物可以通过多种生态机制影响本地植物的菌根真菌(
模拟气候变暖和氮沉降对两种来源加拿大一枝黄花叶性状和性状谱的影响
1
2019
... Ecological mechanisms of alien plant invasion
| 生态机制 Mechanism of ecology | 举例 Example | 参考文献 Reference |
|---|---|---|
| 物种竞争 Species competition | 与农作物(如高粱(Sorghum bicolor)、玉米(Zea mays))相比, 入侵植物Parthenium hysterophorus具有快速早期生长的能力. Compared with crops (such as Sorghum bicolor and Zea mays), the invasive plant Parthenium hysterophorus can grow much more rapidly in their early stage. | |
| 比较夏威夷19对入侵植物和本地植物的资源利用效率, 发现入侵植物具有较高的碳同化率、光利用效率、瞬时氮利用效率和瞬时能量利用效率. Comparing the resource utilization efficiency for 19 paired invasive-native plants in Hawaii, invasive plants are observed to have higher carbon assimilation rate, light use efficiency, instantaneous nitrogen and energy use efficiency. | ||
| 比较47对入侵植物和非入侵植物对草食性动物的抵抗能力及养分含量发现, 入侵植物拥有更高的叶氮含量, 受到食草动物的损伤较小. Comparisons among 47 paired invasive and non-invasive plants for leaf herbivore-resistance and nutrient content show that invasive plants have higher leaf nitrogen content and less damage by herbivores. | ||
| 比对125种入侵植物和196种非入侵植物的生理性状发现, 入侵植物在生长率、养分分配及抗逆性更有优势. Comparing physiological traits of 125 invasive with plants 196 non-invasive plants show that invasive plants have more advantages in growth rate, nutrient allocation and stress resistance. | ||
| 外来植物Solanum carolinense具有较强的抗寒能力和无性繁殖能力. Exotic plant, Solanum carolinense, has strong ability for cold resistance and asexual reproduction. | ||
| 气候变暖及氮沉降的增加可以增强入侵植物Solidago canadensis叶片的资源获取能力. Climate warming and increased nitrogen deposition can enhance leaf resource acquisition of an invasive plant, Solidago canadensis. | ||
| 化感物质 Allelochemical | 入侵植物白羊草(Bothriochloa ischaemum)的渗滤液抑制本地植物Andropogon gerardii和Schizachyrium scoparium发芽及生长. The leachate of an invasive plant, Bothriochloa ischaemum, inhibits germination and growth of the native plants, Andropogon gerardii and Schizachyrium scoparium. | |
| 入侵植物水烛(Typha angustifolia)根系分泌的酚类物质抑制了本地植物Bolboschoenus fluviatilis的生长发育. Phenolic compounds secreted by roots of an invasive plant, Typha angustifolia, inhibit growth and development of a native plant, Bolboschoenus fluviatilis. | ||
| Solidago canadensis入侵分泌的化感物质抑制了本地植物莴苣(Lactuca sativa)种子萌发和幼苗生长. Allelochemicals secreted by the invasion of Solidago canadensis inhibit seed germination and seedling growth of a native plant, Lactuca sativa. | ||
| 外来植物Solanum carolinense含有龙葵碱及草酸盐结晶. The exotic plant Solanum carolinense contains solanine and oxalate crystals. | ||
| Parthenium hysterophorus可以分泌帕台单宁、咖啡酸、香草酸等化感物质帮助自身入侵. Parthenium hysterophorus can secrete parthenin, caffeic acid, vanillic acid and other allelochemicals to facilitate its invasion. | ||
| 土壤养分变化 Soil nutrient variation | 氮沉降会增强S. canadensis入侵初期分泌的化感物质对本地植物莴苣种子萌发和幼苗生长的抑制程度. Nitrogen deposition can enhance the inhibition of the allelochemicals secreted by S. canadensis at its initial invasive stage on seed germination and seedling growth of a native plant, Lactuca sativa. | |
| 分析94篇文章发现, 入侵植物增大了植物、土壤和土壤微生物中的碳和氮库大小. Analysis of 94 published papers show that invasive plants increase the size of carbon and nitrogen pools in plants, soils and soil microorganisms. | ||
| 与本地植物Artemisia californica、Salvia mellifera、Salvia apiana、锦鸡儿(Calochortus splendens)和Dichelostemma capitatum相比, 外来植物Centaurea stoebe、Hirschfeldia incana和Bromus madritensis可以通过加快凋落物分解改善土壤养分. Compared with native plants such as Artemisia californica, Salvia mellifera, Salvia apiana, Calochortus splendens and Dichelostemma capitatum, exotic plants, like Centaurea stoebe, Hirschfeldia incana and Bromus madritensis, can increase soil nutrient level by increasing litter decomposition. | ||
| 入侵植物Mikania micrantha分泌的化感物质绿原酸和β-石竹烯加快了土壤有机氮的矿化过程. Allelochemicals secreted by the invasive plant, Mikania micrantha, such as chlorogenic acid and β-caryophyllene accelerate mineralization of soil organic nitrogen. | ||
| 入侵植物通过凋落物改变了根际微生物土壤群落和养分循环, 形成植物-土壤正反馈调节. Invasive plants can form a positive plant-soil feedback via its litter decomposition which could change rhizosphere soil microbial communities and nutrient cycling. | ||
| 杂草Conyza canadensis入侵的土壤中有效氮和有机质含量增加. The content of available nitrogen and organic matter increases in the soil invaded by Conyza canadensis. | ||
| 与非入侵地土壤相比, Eupatorium catarium入侵后土壤氮含量提高4.32 mg·kg-1. Compared with non-invasive soils, soil nitrogen content increases by 4.32 mg·kg-1 after invasion by Eupatorium catarium. | ||
| Ambrosia trifida在入侵地的土壤-植物反馈高于原产地. Ambrosia trifida has a greater plant-soil feedback in invaded ranges than that in its native ranges. |
模拟气候变暖和氮沉降对两种来源加拿大一枝黄花叶性状和性状谱的影响
1
2019
... Ecological mechanisms of alien plant invasion
| 生态机制 Mechanism of ecology | 举例 Example | 参考文献 Reference |
|---|---|---|
| 物种竞争 Species competition | 与农作物(如高粱(Sorghum bicolor)、玉米(Zea mays))相比, 入侵植物Parthenium hysterophorus具有快速早期生长的能力. Compared with crops (such as Sorghum bicolor and Zea mays), the invasive plant Parthenium hysterophorus can grow much more rapidly in their early stage. | |
| 比较夏威夷19对入侵植物和本地植物的资源利用效率, 发现入侵植物具有较高的碳同化率、光利用效率、瞬时氮利用效率和瞬时能量利用效率. Comparing the resource utilization efficiency for 19 paired invasive-native plants in Hawaii, invasive plants are observed to have higher carbon assimilation rate, light use efficiency, instantaneous nitrogen and energy use efficiency. | ||
| 比较47对入侵植物和非入侵植物对草食性动物的抵抗能力及养分含量发现, 入侵植物拥有更高的叶氮含量, 受到食草动物的损伤较小. Comparisons among 47 paired invasive and non-invasive plants for leaf herbivore-resistance and nutrient content show that invasive plants have higher leaf nitrogen content and less damage by herbivores. | ||
| 比对125种入侵植物和196种非入侵植物的生理性状发现, 入侵植物在生长率、养分分配及抗逆性更有优势. Comparing physiological traits of 125 invasive with plants 196 non-invasive plants show that invasive plants have more advantages in growth rate, nutrient allocation and stress resistance. | ||
| 外来植物Solanum carolinense具有较强的抗寒能力和无性繁殖能力. Exotic plant, Solanum carolinense, has strong ability for cold resistance and asexual reproduction. | ||
| 气候变暖及氮沉降的增加可以增强入侵植物Solidago canadensis叶片的资源获取能力. Climate warming and increased nitrogen deposition can enhance leaf resource acquisition of an invasive plant, Solidago canadensis. | ||
| 化感物质 Allelochemical | 入侵植物白羊草(Bothriochloa ischaemum)的渗滤液抑制本地植物Andropogon gerardii和Schizachyrium scoparium发芽及生长. The leachate of an invasive plant, Bothriochloa ischaemum, inhibits germination and growth of the native plants, Andropogon gerardii and Schizachyrium scoparium. | |
| 入侵植物水烛(Typha angustifolia)根系分泌的酚类物质抑制了本地植物Bolboschoenus fluviatilis的生长发育. Phenolic compounds secreted by roots of an invasive plant, Typha angustifolia, inhibit growth and development of a native plant, Bolboschoenus fluviatilis. | ||
| Solidago canadensis入侵分泌的化感物质抑制了本地植物莴苣(Lactuca sativa)种子萌发和幼苗生长. Allelochemicals secreted by the invasion of Solidago canadensis inhibit seed germination and seedling growth of a native plant, Lactuca sativa. | ||
| 外来植物Solanum carolinense含有龙葵碱及草酸盐结晶. The exotic plant Solanum carolinense contains solanine and oxalate crystals. | ||
| Parthenium hysterophorus可以分泌帕台单宁、咖啡酸、香草酸等化感物质帮助自身入侵. Parthenium hysterophorus can secrete parthenin, caffeic acid, vanillic acid and other allelochemicals to facilitate its invasion. | ||
| 土壤养分变化 Soil nutrient variation | 氮沉降会增强S. canadensis入侵初期分泌的化感物质对本地植物莴苣种子萌发和幼苗生长的抑制程度. Nitrogen deposition can enhance the inhibition of the allelochemicals secreted by S. canadensis at its initial invasive stage on seed germination and seedling growth of a native plant, Lactuca sativa. | |
| 分析94篇文章发现, 入侵植物增大了植物、土壤和土壤微生物中的碳和氮库大小. Analysis of 94 published papers show that invasive plants increase the size of carbon and nitrogen pools in plants, soils and soil microorganisms. | ||
| 与本地植物Artemisia californica、Salvia mellifera、Salvia apiana、锦鸡儿(Calochortus splendens)和Dichelostemma capitatum相比, 外来植物Centaurea stoebe、Hirschfeldia incana和Bromus madritensis可以通过加快凋落物分解改善土壤养分. Compared with native plants such as Artemisia californica, Salvia mellifera, Salvia apiana, Calochortus splendens and Dichelostemma capitatum, exotic plants, like Centaurea stoebe, Hirschfeldia incana and Bromus madritensis, can increase soil nutrient level by increasing litter decomposition. | ||
| 入侵植物Mikania micrantha分泌的化感物质绿原酸和β-石竹烯加快了土壤有机氮的矿化过程. Allelochemicals secreted by the invasive plant, Mikania micrantha, such as chlorogenic acid and β-caryophyllene accelerate mineralization of soil organic nitrogen. | ||
| 入侵植物通过凋落物改变了根际微生物土壤群落和养分循环, 形成植物-土壤正反馈调节. Invasive plants can form a positive plant-soil feedback via its litter decomposition which could change rhizosphere soil microbial communities and nutrient cycling. | ||
| 杂草Conyza canadensis入侵的土壤中有效氮和有机质含量增加. The content of available nitrogen and organic matter increases in the soil invaded by Conyza canadensis. | ||
| 与非入侵地土壤相比, Eupatorium catarium入侵后土壤氮含量提高4.32 mg·kg-1. Compared with non-invasive soils, soil nitrogen content increases by 4.32 mg·kg-1 after invasion by Eupatorium catarium. | ||
| Ambrosia trifida在入侵地的土壤-植物反馈高于原产地. Ambrosia trifida has a greater plant-soil feedback in invaded ranges than that in its native ranges. |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}