Population status and conservation of an extremely small population species Euryodendron excelsum
Xue-Ying WEI1, Yu-Shi YE1, Xi-Po LIN2, Yu-Wen CUI1, Fei-Yan ZENG1, Fa-Guo WANG,1,*1Key Laboratory of Plant Resources Conservation and Sustainable Utilization, Guangdong Provincial Key Laboratory of Applied Botany, South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China and 2Management Department of Ehuangzhang Provincial Nature Reserve, Yangchun, Guangdong 529631, China
Supported by the Project of Protection Department of State Forestry and Grassland Administration Protection and Management Project of Wild Animals and Plants in Guangdong Province
Abstract Aims Euryodendron excelsum, an extremely endangered species of Theaceae, is endemic to China. However, there are few studies on the cause of population structure change of E. excelsum, and its conservation strategy is not perfect. We studied the age structure and dynamics of the E. excelsum population, compared them with the research results in 2007, and analyzed the changes of the E. excelsum population in the last ten years, so as to provide scientific reference for the effective protection and management of the E. excelsum population. Methods Based on field investigation data, the values of parameters such as standard life table, survival curve, killing power and life expectancy curve were established, and the structure status of the E. excelsum population was analyzed as well. The community composition was analyzed using DET’s multiple analysis method. Important findings (1) The analysis of population dynamic indices showed that dynamic indices (Vpi and V′pi) of the wild E. excelsum population were all less than 0. The survival curve of the whole population generally fit a Deevey-III type curve, suggesting a declining population. The number of young individuals was abundant, but impaired development, and hence cannot support the population growth in time. (2) The analysis of survival curve, killing power and life expectancy showed that the sapling, small tree II and middle tree II obstructed the natural population renewal. (3) The Sanjia and Bajia group seedlings of E. excelsum decreased by 39.3% and 71.9%, respectively. The seedling conversion rate of Sanjia group was nearly 1/2 times lower than the Bajia group of E. excelsum. Human interference was the main driver of population decline of E. excelsum. In addition, the conservation strategy that has been implemented is not suitable for the current situation. Keywords:Euryodendron excelsum;population structure;survival curve
PDF (4175KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 魏雪莹, 叶育石, 林喜珀, 崔煜文, 曾飞燕, 王发国. 极小种群植物猪血木的种群现状及保护对策. 植物生态学报, 2020, 44(12): 1236-1246. DOI: 10.17521/cjpe.2020.0059 WEI Xue-Ying, YE Yu-Shi, LIN Xi-Po, CUI Yu-Wen, ZENG Fei-Yan, WANG Fa-Guo. Population status and conservation of an extremely small population species Euryodendron excelsum. Chinese Journal of Plant Ecology, 2020, 44(12): 1236-1246. DOI: 10.17521/cjpe.2020.0059
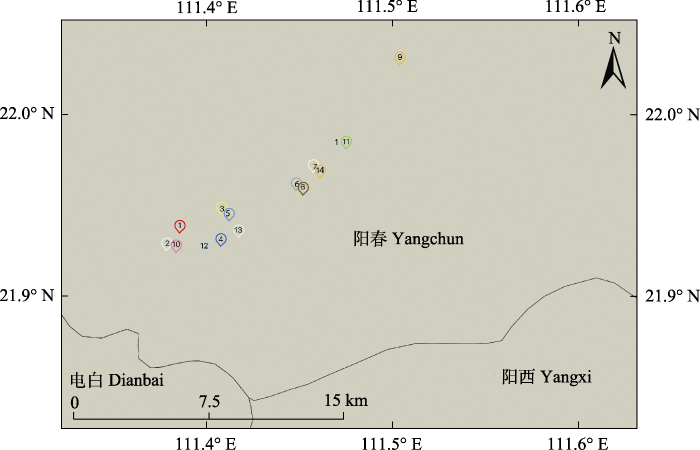
1, Helu Village in Bajia Town; 2, Muli Village in Bajia Town; 3, Chengdong Village in Bajia Town; 4, Village forest of Chengdong Village in Bajia Town; 5, Chongtian Village in Bajia Town; 6, Shanhou of Zhongtian Village in Bajia Town; 7, Shima of Zhongtian Village in Bajia Town; 8, Tangyong of Zhongtian Village in Bajia Town; 9, Banbei Village in Sanjia Town; 10, Shijiao of Gaowu Village in Bajia Town; 11, Chaliao of Lianhe Village in Bajia Town; 12, Sanye of Gaowu Village in Bajia Town; 13, Zhugen of Chengdong Village in Bajia Town; 14, Zhongtian of Chengdong Village in Bajia Town.
Table 1 表1 表1在阳春猪血木分布区域基于实测法调查到的植物种类 Table 1Species recorded with the field measurement method in the distributional range of Yangchun Euryodendron excelsum population
生活型 Life form
类别 Category
物种 Species
德氏多度 Drude abundance
乔木 Arbor
优势种 Dominant species
阴香 Cinnamomum burmannii
cop3
粉箪竹 Bambusa chungii
cop1
卫矛叶蒲桃 Syzygium euonymifolium
sol
龙眼 Dimocarpus longan
sp
伴生种 Companion species
假苹婆 Sterculia lanceolata
sp
车筒竹 Bambusa sinospinosa
cop1
楝 Melia azedarach
sol
破布叶 Microcos paniculata
sp
鹅掌柴 Schefflera heptaphylla
cop1
华润楠 Machilus chinensis
un
杉木 Cunninghamia lanceolata
sol
灌木 Shrub
优势种 Dominant species
九里香 Murraya exotica
sol
九节 Psychotria asiatica
cop2
赪桐 Clerodendrum japonicum
sp
对叶榕 Ficus hispida
cop1
黄椿木姜子 Litsea variabilis
cop1
八角枫 Alangium chinense
sol
伴生种 Companion species
紫玉盘 Uvaria macrophylla
sp
大罗伞树 Ardisia hanceana
sp
小花山小橘 Glycosmis parviflora
sp
大青 Clerodendrum cyrtophyllum
sol
猴耳环 Archidendron clypearia
sp
粗叶榕 Ficus hirta
sol
草本 Herb
优势种 Dominant species
假蒟 Piper sarmentosum
sp
热亚海芋 Alocasia macrorrhizos
cop1
半边旗 Pteris semipinnata
cop1
伴生种 Companion species
火炭母 Polygonum chinense
un
欧洲凤尾蕨 Pteris cretica
sol
淡竹叶 Lophatherum gracile
sol
华南毛蕨 Cyclosorus parasiticus
sol
牛白藤 Hedyotis hedyotidea
sol
玉叶金花 Mussaenda pubescens
sol
cop, 多; sol, 稀少; sp, 不多而分散; un, 单株。 cop, copiosa; sol, solitariae; sp, sparsal; un, unicurn.
A, 八甲镇澄垌村。B, 八甲镇澄垌村竹根。C, 八甲镇澄垌中田村。D, 八甲镇高屋村石角。E, 八甲镇高屋村三叶。F, 八甲镇联合村茶嘹。G, 八甲镇合路村。H, 八家镇木力村。I, 八甲镇中田村山喉。J, 八甲镇中田村师马。K, 八甲镇中田村塘涌。L, 三甲镇湴背村。M, 八甲镇澄垌村。N, 八甲镇冲田村。O, 三甲镇湴背村。 Fig. 2General situation of the habitat of Euryodendron excelsum.
A, Chengdong Village in Bajia Town. B, Zhugen of Chengdong Village in Bajia Town. C, Zhongtian of Chengdong Village in Bajia Town. D, Shijiao of Gaowu Village in Bajia Town. E, Sanye of Gaowu Village in Bajia Town. F, Chaliao of Lianhe Village in Bajia Town. G, Helu Village in Bajia Town. H, Muli Village in Bajia Town. I, Shanhou of Zhongtian Village in Bajia Town. J, Shima of Zhongtian Village in Bajia Town. K, Tangyong of Zhongtian Village in Bajia Town. L, Banbei Village in Sanjia Town. M, Chengdong Village in Bajia Town. N, Chongtian Village in Bajia Town. O, Banbei Village in Sanjia Town.
Table 2 表2 表2猪血木各分布点生境和数量概况 Table 2General situation of habitat and quantity of each locality of Euryodendron excelsum
编号 Number
地点 Place
生境 Habitat
乔木层盖度 Coverage of arbor layer
海拔 Altitude (m)
人为干扰 Human disturbance
干扰原因 Cause of interference
成株 Adult plant
幼树 Sapling
幼苗 Seedling
1
八甲镇合路村 Helu Village in Bajia Town
村边林 Village forest
50
52
弱 Weak
1
0
10
2
八甲镇木力村 Muli Village in Bajia Town
村边林 Village forest
65
68
中 Middle
2
0
3
3
八甲镇澄垌村 Chengdong Village in Bajia Town
田野 Field
45
42
中 Middle
阴蔽 Shading
0
0
10
4
八甲镇澄垌村 Chengdong Village in Bajia Town
村边林 Village forest
60
42
中 Middle
断梢 Branches broken
2
2
20
5
八甲镇澄垌村冲田 Chongtian of Chengdong Village in Bajia Town
村边林 Village forest
70
30
中 Middle
8
0
20
6
八甲镇中田村山喉 Shanhou of Zhongtian Village in Bajia Town
村边林 Village forest
30
72
弱 Weak
1
4
28
7
八甲镇中田村师马 Shima of Zhongtian Village in Bajia Town
村边 Near the village
65
42
强 Strong
栽培 Cultivation
1
3
10
8
八甲镇中田村塘涌 Tangyong of Zhongtian Village in Bajia Town
村边 Village
20
30
强 Strong
断梢 Branches broken
0
2
0
9
三甲镇湴背村 Banbei Village in Sanjia Town
村边风水林 Fengshui woods near village
80
44
强 Strong
盗挖 Stolen
16
5
40
10
八甲镇高屋村石角 Shijiao of Gaowu Village in Bajia Town
村边林 Village forest
60
42
强 Strong
1
0
2
11
八甲镇联合村茶嘹 Chaliao of Lianhe Village in Bajia Town
村边林 Village forest
70
33
中 Middle
断梢 Branches broken
2
3
7
12
八甲镇高屋村三叶 Sanye of Gaowu Village in Bajia Town
村边林 Village forest
75
65
中 Middle
阴蔽 Shading
1
0
8
13
八甲镇澄垌村竹根 Zhugen of Chengdong Village in Bajia Town
屋后风水林 Fengshui woods behind house
80
32
中 Middle
2
0
10
14
八甲镇澄垌村中田 Zhongtian of Chengdong Village in Bajia Town
St, 野生猪血木种群(包括三甲镇和八甲镇); Sa, 八甲镇野生猪血木种群; Ss, 三甲镇野生猪血木种群。I, 幼苗1级, 0 < H ≤ 100.0 cm; II, 幼苗2级, H > 100.0 cm, DBH < 3.0 cm; III, 幼树, 3.0 cm ≤ DBH < 9.0 cm; IV, 小树1级, 9.0 cm ≤ DBH < 15.0 cm; V, 小树2级, 15.0 cm ≤ DBH < 21.0 cm; VI, 中龄树1级, 21.0 cm ≤ DBH < 27.0 cm; VII, 中龄树2级, 27.0 cm ≤ DBH < 33.0 cm; VIII, 成年树1级, 33.0 cm ≤ DBH < 39.0 cm; IX, 成年树2级, 39.0 cm ≤ DBH < 45.0 cm; X, 大树, DBH ≥ 45.0 cm。 Fig. 3Height (H) and diameter at breast height (DBH) class of Euryodendron excelsum population.
St, wild E. excelsum population (including Sanjia Town and Bajia Town); Sa, wild E. excelsum population of Bajia Town; Ss, wild E. excelsum population of Sanjia Town. I, the seedling I stage, 0 < H ≤ 100.0 cm; II, the seedling II stage, H > 100.0 cm, DBH < 3.0 cm; III, the sapling stage, 3.0 cm ≤ DBH < 9.0 cm; IV, the small tree I stage, 9.0 cm ≤ DBH < 15.0 cm; V, the small tree II stage, 15.0 cm ≤ DBH < 21.0 cm; VI, the middle tree I stage, 21.0 cm ≤ DBH < 27.0 cm; VII, the middle tree II stage, 27.0 cm ≤ DBH < 33.0 cm; VIII, the adult tree I stage, 33.0 cm ≤ DBH < 39.0 cm; IX, the adult tree II stage, 39.0 cm ≤ DBH < 45.0 cm; X, the big tree stage, DBH ≥ 45.0 cm.
Table 3 表3 表3阳春猪血木种群标准生命表 Table 3Standard life table of Euryodendron excelsum population in Yangchun City
大小级 Size stage
划分标准 Division standard
ax
lx
dx
qx
Lx
Tx
kx
ex
lnlx
I (St)
0 ≤ H ≤ 100
97
1 000
185.
0.19
1 814
1 855
0.21
1.86
6.91
II (St)
H > 100, DBH < 3.0
79
814
618
0.76
1 010
1 051
1.43
1.29
6.70
III (St)
DBH [3.0, 9.0)
19
196
123
0.63
268
309
1.00
1.58
5.28
IV (St)
DBH [9.0, 15.0)
7
72
-20
-0.29
164
206
-0.25
2.86
4.28
V (St)
DBH [15.0, 21.0)
9
93
30
0.33
154
195
0.41
2.11
4.53
VI (St)
DBH [21.0, 27.0)
6
62
-41
-0.67
164
206
-0.51
3.33
4.12
VII (St)
DBH [27.0, 33.0)
10
103
92
0.90
113
154
2.30
1.50
4.64
VIII (St)
DBH [33.0, 39.0)
1
10
-20
-2.00
41
82
-1.10
8.00
2.33
IX (St)
DBH [39.0, 45.0)
3
31
-10
-0.33
72
113
-0.29
3.67
3.43
X (St)
≥ 45.0
4
41
NA
NA
41
364
3.72
8.83
3.72
I (Sa)
0 ≤ H ≤ 100
80
1 000
453
0.45
1 546
1 587
0.60
1.59
6.91
II (Sa)
H > 100, DBH < 3.0
53
546
402
0.74
690
731
1.33
1.34
6.30
III (Sa)
DBH [3.0, 9.0)
14
144
123
0.86
164
206
1.95
1.43
4.97
IV (Sa)
DBH [9.0, 15.0)
2
21
-10
-0.50
51
92
-0.41
4.50
3.03
V (Sa)
DBH [15.0, 21.0)
3
31
-10
-0.33
72
113
-0.29
3.67
3.43
VI (Sa)
DBH [21.0, 27.0)
4
41
-30
-0.75
113
154
-0.56
3.75
3.72
VII (Sa)
DBH [27.0, 33.0)
7
72
61
0.86
82
123
1.95
1.71
4.28
VIII (Sa)
DBH [33.0, 39.0)
1
10
-20
-2.00
41
82
-1.10
8.00
2.33
IX (Sa)
DBH [39.0, 45.0)
3
31
-10
-0.33
72
113
-0.29
3.67
3.43
X (Sa)
≥ 45.0
4
41
NA
NA
41
364
3.72
8.83
3.72
I (Ss)
0 ≤ H ≤ 100
24
1 000
835
0.31
1 164
1 206
1.80
1.21
6.91
II (Ss)
H > 100, DBH < 3.0
16
165
113
0.84
216
257
1.16
1.56
6.91
III (Ss)
DBH [3.0, 9.0)
5
52
0
0.69
103
144
0.00
2.80
5.11
IV (Ss)
DBH [9.0, 15.0)
5
52
-10
0.00
113
154
-0.18
3.00
3.94
V (Ss)
DBH [15.0, 21.0)
6
62
41
-0.20
82
123
1.10
2.00
3.94
VI (Ss)
DBH [21.0, 27.0)
2
21
-10
0.67
51
92
-0.41
4.50
4.12
VII (Ss)
DBH [27.0, 33.0)
3
31
3.03
ax, 在x年内出现的个体数; dx, 从x到x + 1年间隔阶段标准化死亡数; DBH, 胸径; ex, 进入x年个体的寿命期望值; H, 个体高度; kx, 各级的致死率, kx = lnlx/lnlx + 1; lnlx, 标准化存活数的自然对数值; lx, 在x年内标准存活个体数; Lx, (lx + lx+1)/2; qx, 从x到x + 1年间隔阶段标准化死亡率; Tx, 从x年到超过x年的个体总数。Sa, 八甲镇野生猪血木种群; Ss, 三甲镇野生猪血木; St, 野生猪血木种群(包括三甲镇和八甲镇)。 ax, number of alive organisms at age x; dx, number of dead organisms from age x to x + 1; DBH, diameter at breast height; ex, life expectancy at age x; H, the height of individual; kx, age-specific mortality; lnlx, the natural logarithm of lx; lx, proportion organisms surviving from the beginning of the life table to age x; Lx, number of survivors from age x to x + 1; qx, mortality rate from age x to x + 1; Tx, total number of alive organisms from age x to x + 1. Sa, wild E. excelsum (only including Bajia Town); Ss, wild E. excelsum (only including Sanjia Town); St, wild E. excelsum (including Sanjia Town and Bajia Town).
Sa, 八甲镇野生猪血木种群; Ss, 三甲镇野生猪血木种群; St, 野生猪血木种群(包括三甲镇和八甲镇)。I-X同图3。 Fig. 4Standard survival curve of Euryodendron excelsum population. lx, proportion organisms surviving from the beginning of the life table to age x.
Sa, wild E. excelsum population of Bajia Town; Ss, wild E. excelsum population of Sanjia Town; St, wild E. excelsum population (including Sanjia Town and Bajia Town). I-X see Fig. 3.
Sa-ex, 八甲镇野生猪血木种群进入x年个体的寿命期望值; Sa-kx, 八甲镇野生猪血木种群致死率; Sa-qx, 八甲镇野生猪血木种群标准化死亡率; St-ex, 野生猪血木种群(包括三甲镇和八甲镇)进入x年个体的寿命期望值; St-kx, 野生猪血木种群(包括三甲镇和八甲镇)致死率; St-qx, 野生猪血木种群(包括三甲镇和八甲镇)标准化死亡率。I-X同图3。 Fig. 5Killing power and life expectancy curve of Euryodendron excelsum population.
Sa-ex, life expectancy at age x of wild E. excelsum (only including Bajia Town); Sa-kx, specific mortality of wild E. excelsum (only including Bajia Town); Sa-qx, mortality rate of wild E. excelsum (only including Bajia Town); St-ex, life expectancy at age x of wild E. excelsum (including Sanjia Town and Bajia Town); St-kx, specific mortality of wild E. excelsum (including Sanjia Town and Bajia Town); St-qx, mortality rate of wild E. excelsum (including Sanjia Town and Bajia Town). I-X see Fig. 3.
CaiF, SongYC (1997). A study on the structure and dynamics of Schima superba population on Wuyi Mountain Acta Phytoecologica Sinica, 21, 138-149. [本文引用: 1]
ChenXD (1998). A study on the method of quantitative analysis for plant population and community structural dynamics Acta Ecologica Sinica, 18, 214-217. [本文引用: 1]
ChenYZ, MaXQ, FengLZ, ZhengQR (2006). The population life table and periodic fluctuation of Cinnamomun micranthum, an endangered plant Acta Ecologica Sinica, 26, 4267-4272. [本文引用: 1]
CuiL, SuY (2015). SSR markers for a critically endangered species Euryodendron excelsum and a distantly related species Ternstroemia gymnanthera(Ternstroemiaceae) Biochemical Systematics and Ecology, 58, 288-292. [本文引用: 1]
HeH, WangYH (2001). The existing circumstance and the biological characteristic research of Euryodendron excelsum H. T. Chang, the paticular genera in China Journal of Yunnan University (Natural Sciences Edition), 23(S1), 52-55. [本文引用: 1]
HiltonTC (2000). IUCN Red List of Threatened Species IUCN Publication Service Unit, Cambridge. 21. [本文引用: 1]
KangHJ, ChenZL, LiuP, HaoCY, WeiFM (2007). The population structure and distribution pattern of Emmenopterys henryi in Dapanshan natural reserve of Zhejiang Province Acta Ecologica Sinica, 27, 389-396. [本文引用: 1]
LiXK, SuZM, XiangWS, NingSJ, TangRQ, OuZL, LiRT (2002). Study on the structure and spatial pattern of the endangered plant population of Abies yuanbaoshanensis Acta Ecologica Sinica, 22, 2246-2253. [本文引用: 1]
LiXBY, FuL, WangFG, XingFW (2017). Michelia guangdongensis (Magnoliaceae), an endangered plant species with extremely small population, should be evaluated as CR C2a (i); D Biodiversity Science, 25, 91-93. [本文引用: 1]
LuoXY, TangGD, XuH, ZhuangXY, ZhengWJ, ZengM (2005). Genetic diversity of three endemic and endangered species of the family Theaceae in Guangdong, China Biodiversity Science, 13, 112-121. URL [本文引用: 1] Euryodendron excelsum, Apterosperma oblata, and Camellia changii are three species of the family Theaceae endemic to China. Genetic diversity of the three endangered species was studied using random amplified polymorphic DNA (RAPD). A total of 206-305 bands were generated by 20-24 random primers. Percentages of polymorphic loci(PPL) of the three species were 51.80%, 80.26% and 38.83%, respectively, and coefficients of gene differentiation (GST) were 0.3566, 0.1713, and 0.1242, respectively. The results indicate that Apterosperma oblata, with wider natural distribution, has higher genetic diversity than the other two species with restricted natural distributions. Genetic variation mainly existed within populations; however, population genetic differentiation was obviously higher in Euryodendron excelsumthan in the other two species, which possibly resulted from habitat fragmentation. Based on the different genetic structure, we put forward some conservation strategies, including in situ preservation, establishing a conservation centre for endangered species of the family Theaceae in Guangdong, and developing fast propagation techniques.]]> [ 罗晓莹, 唐光大, 许涵, 庄雪影, 郑文经, 曾明 (2005). 山茶科3种中国特有濒危植物的遗传多样性研究 生物多样性, 13, 112-121.] [本文引用: 1]
RanQ, WeiHY, ZhaoZF, ZhangQZ, LiuJ, GuW (2019). Impact of climate change on the potential distribution and habitat fragmentation of the relict plant Cathaya argyrophylla Chun et Kuang Acta Ecologica Sinica, 39, 2481-2493. [本文引用: 1]
ShenSK, MaHY, LiuXY, WangYH (2007). The endangered causes and preserving strategies for Euryodendron excelsum, a plant endemic to China Ecology and Environment, 16, 1819-1823. [本文引用: 3]
ShenSK, MaHY, WangYH, WangBY, ShenGZ (2008b). The structure and dynamics of natural population of the endangered plant Euryodendron excelsum H. T. Chang Acta Ecologica Sinica, 28, 2404-2412. [本文引用: 5]
[ 申仕康, 马海英, 王跃华, 王博轶, 申国柱 (2008b). 濒危植物猪血木(Euryodendron excelsum H. T. Chang)自然种群结构及动态 生态学报, 28, 2404-2412.] [本文引用: 5]
SuYJ, WangT, SunYF, YeHG (2009). High ISSR variation in 14 surviving individuals of Euryodendron excelsum(Ternstroemiaceae) endemic to China Biochemical Genetics, 47, 56-65. DOI:10.1007/s10528-008-9206-2URLPMID:19096927 [本文引用: 1] Euryodendron excelsum is a critically endangered Ternstroemiaceae species endemic to southern China, with only 14 individuals surviving in Ba Jia Zhen of Yangchun, Guangdong Province. Intersimple sequence repeat (ISSR) markers were used to assess genetic variation and relationships among these individuals. Population genetic parameters were estimated by a Bayesian approach as well as conventional methods. Of the 225 loci generated by 21 primers, 147 (65.33%) were polymorphic. Compared with other species of Theaceae and related families, a high level of genetic variation was identified in E. excelsum (Nei's gene diversity, 0.2458; Shannon's index, 0.3626). An unweighted pair group method with arithmetic mean (UPGMA) dendrogram showed that the 14 individuals were mainly clustered into three groups, a conclusion further supported by principal coordinate analysis. Based on these results, a management and conservation strategy for E. excelsum was proposed.
TaoDL, XuZB, LiX (1985). Experimental study on the effect of dead and living ground covers on the natural regeneration of Pinus koraiensis Sieb. et Zucc. associated tree species Acta Phytoecologica et Geobotanica Sinica, 9, 47-58. [本文引用: 1]
WangT, SuYJ, YeHG, OuyangPY, JiangY, SunYF, ChenGP, DengF, ZhangHD (2005). Genetic differentiation and conservation of 14 surviving individuals of Euryodendron excelsum endemic to China Acta Scientiarum Naturalium Universitatis Sunyatseni, 44, 68-72. [本文引用: 1]
WangXG, WuCZ, HeDJ, LiaoCZ, ChengY, FengL (2004). Life table and spectral analysis of endangered plant Taxus chinensis var. mairei population Journal of Applied Ecology, 15, 201-204. URLPMID:15146623 [本文引用: 1]
WangYH, CaoLM, HeH, HuXL (2002). Cutting propagation of the endangered species Euryodendron excelsum H. T. Chang from Theaceae Journal of Yunnan University (Natural Science Edition), 24, 227-228. [本文引用: 1]
[ 王跃华, 曹丽敏, 何瀚, 胡晓立 (2002). 山茶科濒危植物猪血木(Euryodendron excelsum H. T. Chang)的扦插繁殖 云南大学学报(自然科学版), 24, 227-228.] [本文引用: 1]
WangYH, ShenSK, ShenGZ, WangBY, HuXL, WangM, LiuXY (2016). Pollination biology of endangered Euryodendron excelsum Journal of West China Forestry Science, 36(4), 11-15. [本文引用: 1]
RenZ, ChenL, PengH (2016). Orophea yunnanensis should be listed as a plant species with extremely small population Biodiversity Science, 24, 358-359. [本文引用: 1]
SSR markers for a critically endangered species Euryodendron excelsum and a distantly related species Ternstroemia gymnanthera(Ternstroemiaceae) 1 2015
Development and characterization of genomic microsatellite markers inApterosperma oblata (Theaceae) and cross-amplification in Euryodendron excelsum (Ternstroemiaceae) 1 2015
 ,1,*
,1,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT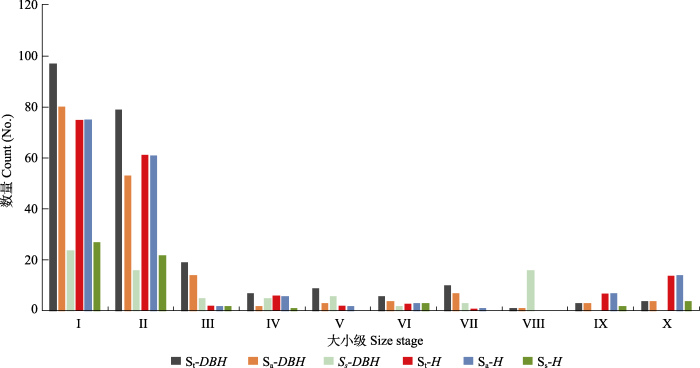 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT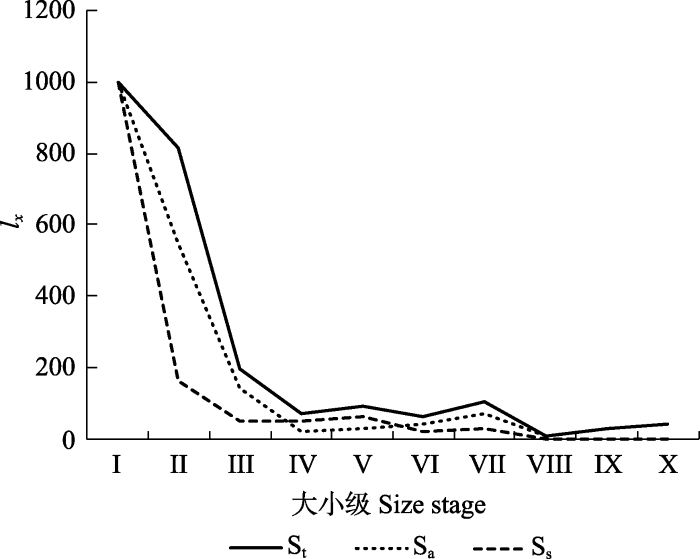 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT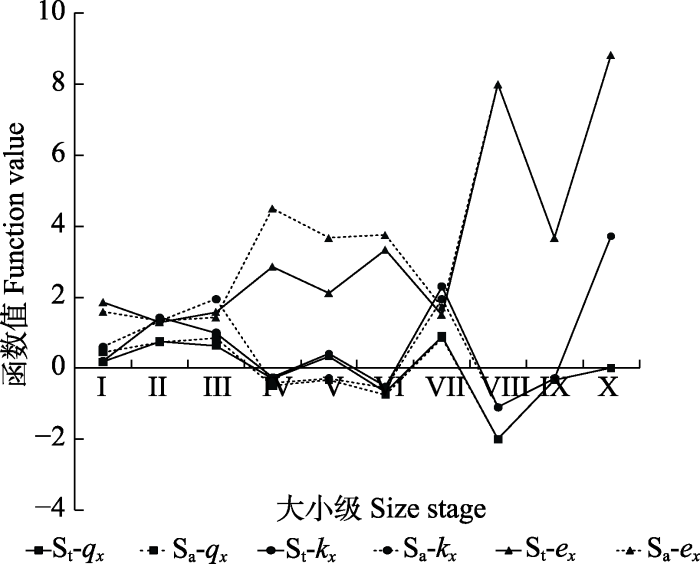 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT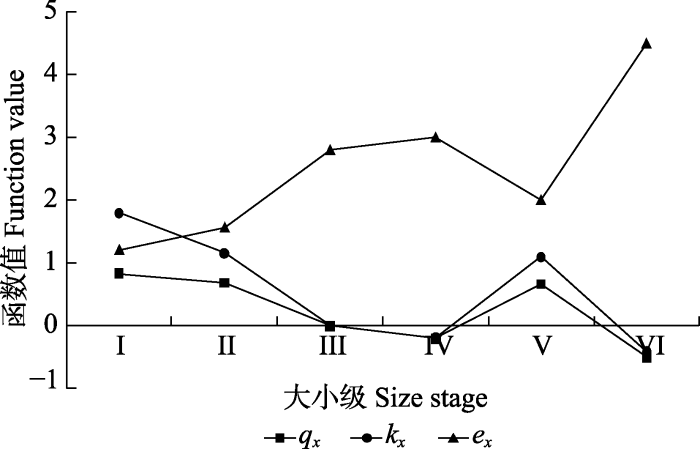 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}