 ,
, ,1,*
,1,*Seed predation and dispersal pattern of Fagaceae species in a fragmented landscape, eastern China
Yu-Chen ZHONG1, Bin WANG2, Zhong-Ping FANG2, Xiao-Zhong XU2, Ming-Jian YU,,1,*通讯作者: *于明坚: ORCID:0000-0001-8060-8427 (fishmj@zju.edu.cn)
编委: 孙书存
责任编辑: 赵航
收稿日期:2020-03-20接受日期:2020-06-17网络出版日期:2021-02-20
| 基金资助: |
Corresponding authors: *(fishmj@zju.edu.cn)
Received:2020-03-20Accepted:2020-06-17Online:2021-02-20
| Fund supported: |

摘要
生境片段化伴随的面积效应和边缘效应, 可改变分散贮食动物的竞争强度、觅食行为以及隐蔽条件, 影响种子捕食和扩散模式。阐明生境片段化对多物种种子捕食和扩散的影响, 对理解片段化生境中的植物更新和生物多样性维持十分重要。该研究在浙江省千岛湖地区的岛屿和大陆上开展了针对6种壳斗科植物的种子捕食和扩散实验, 分析了物种、分散贮食动物相对多度、种子产量、岛屿大小和边缘效应如何共同影响种子命运和种子扩散距离。主要结果: (1)种子命运和扩散距离在物种间存在显著差异; (2)大陆比岛屿有更长的种子留存时间, 小岛种子留存时间最短, 岛屿内部比岛屿边缘有更长的种子留存时间; (3)物种和岛屿大小对种子原地取食率存在交互作用, 白栎(Quercus fabri)种子在大岛上有更高的原地取食率; (4)种子在小岛上有最高的扩散率, 分散贮食动物相对多度对种子扩散后贮藏率有负效应。表明在千岛湖地区, 生境片段化改变了种子捕食和扩散模式, 且面积效应对不同物种的种子捕食和扩散模式产生了不同作用, 从而影响森林群落更新和生物多样性维持。
关键词:
Abstract
Aims Due to the area effect and edge effect, habitat fragmentation can alter competition intensity, foraging behavior and shelter condition of scatter-hoarders, and consequently affect seed predation and dispersal patterns. Though there have been some studies focusing on seed dispersal in fragmented landscapes, few studies have compared the effects of habitat fragmentation on seed predation and dispersal among multiple species.
Methods To explore how species identity, abundance of scatter-hoarders, seed production, island size and edge effect work together to influence seed fates and dispersal distance, we conducted seed predation and dispersal experiments for six Fagaceae species in the island and mainland forests of the Thousand Island Lake region in Zhejiang Province, eastern China.
Important findings (1) There were significant differences in seed fates and dispersal distance among six Fagaceae species. (2) Seeds were removed faster on the islands than on the mainland, and even faster on smaller islands than on larger islands. Further, seeds were removed faster at the island edges than in the interiors. (3) There was a significant interactive effect between island size and species identity on the probability of seeds eaten in situ. More seeds of Quercus fabri were eaten in situ on larger islands. (4) Seeds had the highest probability of removal on small islands; the relative abundance of scatter-hoarders had a negative effect on the probability of seeds cached. These findings implied that habitat fragmentation altered seed predation and dispersal patterns, and that area effect posed species-specific effects on seed predation and dispersal patterns, which would contribute to forest regeneration and biodiversity maintenance.
Keywords:
PDF (3931KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
钟雨辰, 王斌, 方中平, 徐小忠, 于明坚. 片段化景观中壳斗科植物种子捕食和扩散模式. 植物生态学报, 2021, 45(2): 154-162. DOI: 10.17521/cjpe.2020.0079
ZHONG Yu-Chen, WANG Bin, FANG Zhong-Ping, XU Xiao-Zhong, YU Ming-Jian.

随着人类对自然的干扰加大, 生境片段化对森林更新和生物多样性的影响受到越来越多的关注。生境片段化是指连续的生境被分割成多个相互隔离的小斑块(片段)的过程(Wilcove et al., 1986), 其带来的生境面积减小、生境隔离度增加、边缘效应增强等影响着一些重要生态过程(Lindenmayer & Fischer, 2013), 从而干扰生物多样性和生态系统功能维持(Fahrig, 2003; Haddad et al., 2015)。
种子扩散对植物更新具有十分重要的意义(Vander Wall, 2001)。分散贮食动物在种子扩散过程中扮演重要角色(Vander Wall, 1990, 2010; Vander Wall & Beck, 2012; Lichiti et al., 2017), 它们既能扩散种子, 同时也会捕食种子(Vander Wall, 2010; Xiao et al., 2013)。
一些研究认为, 生境片段化带来的生境面积减小和边缘效应是影响动植物关系如种子捕食和扩散的重要因素(Fleury & Galetti, 2006; Mendes et al., 2016; Chen et al., 2017)。生境面积和边缘效应可通过改变分散贮食动物对种子的竞争强度以及隐蔽条件来影响分散贮食动物与种子之间的关系(Andreazzi et al., 2012; Morán-Lópezet al., 2015)。例如, Morán-López等(2015)发现小的森林片段隐蔽条件较差, 鼠类更易受天敌威胁, 且对种子的竞争强度减小, 导致更低的种子扩散率、贮藏率和扩散距离; Andreazzi等(2012)认为小片段更好的光照条件利于种子生产, 而中大型种子捕食者丧失, 导致较低的种子捕食率和较高的存活率; Fleury和Galetti (2006)则发现中等大小片段中种子被捕食概率最高。边缘效应对种子命运的影响同样不一致, Morán-López等(2015)认为森林边缘隐蔽条件较差而被分散贮食动物回避, 导致森林边缘比森林内部有更低的种子贮藏率和扩散距离; Kollmann和Buschor (2003)则发现森林边缘更高比例的灌丛覆盖增加了啮齿动物多度, 导致更高的种子捕食率。另外, 生境片段化还可通过大型兽类的丧失引起营养级联效应, 改变分散贮食动物的数量和行为, 间接影响种子捕食率和扩散率(Kurten, 2013; Galetti et al., 2015; Zeng et al., 2019)。
已有很多研究发现种子大小和种子外壳厚度等性状会影响分散贮食动物对种子的选择(Moore et al., 2007; Vander Wall, 2010; Lichti et al., 2017; Wang & Ives, 2017)。而森林中往往存在多个物种果期重叠的现象, 意味着不同物种种子将受到分散贮食动物觅食选择上的偏好(Lichiti et al., 2014; Wang et al., 2014; Yi & Wang, 2015), 然而很少有研究阐明生境片段化对不同物种种子捕食和扩散模式的影响存在怎样的差异(Chen et al., 2017), 而这对于理解片段化生境中的植物更新和生物多样性维持十分重要。
位于我国亚热带东部的千岛湖, 因水库大坝建设形成了1 000多个陆桥岛屿, 从而形成了由不同面积片段组成的片段化景观。千岛湖地区分布着多种壳斗科植物, 因而是研究生境片段化对动物介导的多物种种子捕食和扩散模式影响的理想平台。本研究利用千岛湖片段化景观这个平台, 以6种壳斗科植物种子及其分散贮食动物为研究对象, 结合分散贮食动物相对多度和种子产量, 探究以下3个问题: (1)物种间存在怎样的种子命运和种子扩散距离差异; (2)生境类型(岛屿/大陆)、片段大小和边缘梯度如何影响种子命运与种子扩散距离; (3)生境类型(岛屿/大陆)、片段大小和边缘梯度对不同物种种子命运和种子扩散距离的影响是否存在差异。
1 材料和方法
1.1 研究地点概况
研究地位于浙江省杭州市淳安县境内的新安江水库(29.37°-29.83° N, 118.57°-119.25° E;图1), 亦称千岛湖, 于1959年修建新安江水电站时形成, 因最高水位108 m高程时拥有1 078个面积>0.25 hm2的陆桥岛屿而得名。水域面积约540 km2, 平均水深为30.44 m, 容积为178.4亿m3。该地区为典型的亚热带季风气候, 温暖湿润, 四季分明, 雨热同期。年平均气温17 ℃, 年最高气温达41.8 ℃, 年最低气温为-7.6 ℃, 年降水量为1 430 mm。土壤主要为红壤、黄壤和岩性土3类, 土层较薄, 保水保肥能力一般。水库建设前, 原有森林基本被破坏(Liu et al., 2020)。如今, 千岛湖岛屿上的森林覆盖率>88.5%, 以次生马尾松(Pinus massoniana)林为主, 林下有檵木(Loropetalum chinense)、短尾越桔(Vaccinium carlesii)、杜鹃(Rhododendron simsii)等小乔木和灌木(Liu et al., 2020)。已有调查表明, 社鼠(Niviventer confucianus)为千岛湖主要鼠类, 且贮食行为为分散贮食, 另还有赤腹松鼠(Callosciurus erythraeus)等分布(Zeng et al., 2019)。图1
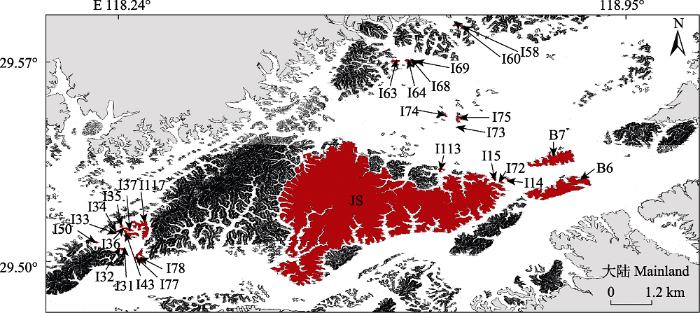 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1千岛湖地区研究岛屿分布。数字和字母表示岛屿号, 岛屿信息见附录I。
Fig. 1Study islands in the Thousand Island Lake region. Numbers and letters represent islands mark. See details in Supplement I.
1.2 研究方法
1.2.1 物种选择及种子采集和标记
白栎(Quercus fabri)、短柄枹(Quercus serrata)、麻栎(Quercus acutissima)、苦槠(Castanopsis sclerophylla)、青冈(Cyclobalanopsis glauca)和石栎(Lithocarpus glaber)是千岛湖地区较常见的6种壳斗科植物, 它们的果期相互重叠(表1)。石栎具有厚且坚硬的种子外壳(Wang et al., 2014, 2016)。于2018年11月在千岛湖地区收集这6个物种的成熟果实。采用浮水法和观察法挑选出健康饱满无虫害的种子(去除了壳斗, 通俗称为种子), 并保证同一物种种内种子大小均一。自然风干后, 每种种子随机取3组各100粒种子测量百粒质量。Table 1
表1
表1千岛湖研究区6种壳斗科植物种子质量(平均值±标准差)和果期
Table 1
| 物种 Species | 种子质量 Seed mass (g) | 果期 Nut-ripening periods |
|---|---|---|
| 短柄枹 Quercus serrata | 0.90 ± 0.02a | 9-12月 Sept.-Dec. |
| 青冈 Cyclobalanopsis glauca | 1.06 ± 0.02a | 9-10月 Sept.-Oct. |
| 苦槠 Castanopsis sclerophylla | 1.64 ± 0.08b | 10-11月 Oct.-Nov. |
| 石栎 Lithocarpus glaber | 1.93 ± 0.07b | 7-12月 July-Dec. |
| 白栎 Quercus fabri | 2.05 ± 0.03b | 10-12月 Oct.-Dec. |
| 麻栎 Quercus acutissima | 7.18 ± 0.34c | 9-12月 Sept.-Dec. |
新窗口打开|下载CSV
利用塑料片标签法(Xiao et al., 2006)标记种子, 在远离种子胚的一端用直径0.5 mm (麻栎1 mm)的钻头钻孔, 用一根长15 cm、直径0.3 mm的铁丝的两头分别拴住种子和一块3.6 cm × 2.5 cm的白色塑料标签牌, 铁丝与标签牌的质量为(0.30 ± 0.02) g。标签牌上写有每粒种子的唯一编号, 编号包含释放点、物种和序号等信息。
1.2.2 实验设置
根据岛屿面积, 本研究从29个植物样岛中选取6个小岛(<0.9 hm2)、3个中岛(0.98-1.5 hm2)和3个大岛(>20 hm2); 另从岛屿周边大陆的次生马尾松林动态监测样地(简称大陆样地)中选取3个样地作为研究地点(图1; 附录I)。每个小岛和大陆样地设置2个种子释放点, 中岛和大岛均设置2个边缘释放点(距边缘<25 m)和2个内部释放点(距边缘>30 m), 释放点之间的距离>25 m。每个释放点放置6个物种种子各10粒, 种子围绕中心随机排列成一个圆圈。在距释放点1 m左右的树干上安置一台红外数码相机(每次触发连拍3张照片和1段15 s的视频), 对准种子释放点, 以观测种子捕食者。于2018年12月中旬释放种子, 并于释放后的第1、2、4、10、22、38天在每个释放点50 m范围(小岛为全岛范围)内进行搜查并记录种子命运, 种子命运分为原地完好(IIS, 完好存留在释放点)、原地取食(EIS, 在释放点被取食)和扩散(R, 扩散到释放点0.5 m以外的区域), 扩散种子又分为扩散后取食(EAR, 离开释放点后被取食)、扩散后贮藏(CAR, 扩散后被完整贮藏在土壤、凋落物中)和丢失(M, 被贮藏在鼠洞深处或超出搜查范围), 同时测量并记录扩散距离(DD)和扩散方向。对于贮藏种子, 我们在距种子0.5 m处插上写有种子信息的白色塑料牌来标记位置, 以便下一次追踪。种子的命运和扩散距离以最后一次追踪到该种子为准。
1.2.3 分散贮食动物相对多度与种子产量估计
统计红外相机独立照片数量(同一个相机同种物种相隔1.5 h以上的照片), 计算出每个岛屿种子释放后的30天内平均每天出现在释放点的分散贮食动物独立照片数量, 经对数转换后作为该岛的分散贮食动物相对多度(徐爱春等, 2014; 陈立军等, 2019)。自2013年, 在29个岛屿和大陆1 hm2样地内共设置250个种子雨收集框, 每月对种子雨收集框内的种子和果实进行一次收集、鉴定和称量, 具体设置方法见南歌(2017)。统计2018年每个岛屿上种子雨收集框内成熟种子和果实质量, 将每m2成熟种子和果实质量作为该岛的种子产量。
1.3 数据分析
对分散贮食动物相对多度、种子产量和岛屿面积进行Spearman相关性检验。采用Cox回归(Wang et al., 2014)分析不同物种、生境类型(大陆/岛屿)、岛屿大小和边缘梯度(边缘/内部)种子留存时间的差异。种子命运百分比的分析则采用基于二项分布族的广义线性混合效应模型(GLMMs), 响应变量为针对所有实验种子计算得到的原地完好率、原地取食率和扩散率, 以及针对扩散且找回种子计算得到的扩散后贮藏率。种子扩散距离的分析采用线性混合效应模型(LMMs)。对种子命运和扩散距离各进行3种分析, 完整模型的固定效应为: (1)物种、岛屿大小、种子产量和分散贮食动物相对多度, 以及物种与其余参数间的相互作用; (2)物种和生境类型(岛屿/大陆)及其相互作用; (3)物种和边缘梯度(边缘/内部)及其相互作用。岛屿或样地号及嵌套在岛屿、样地中的种子释放点作为随机效应。结果只展示具有最低小样本情况下的赤池信息准则值(AICc) (ΔAICc > 2)的最优模型, 模型采用似然比检验(LRTs)进行评估。本研究的所有分析均由R 3.6.1软件和相应软件包完成。Cox回归由survival (Therneau & Grambsch, 2000; Therneau, 2015)软件包完成, 线性混合效应模型和广义线性混合效应模型及模型选择由lme4 (Bates et al., 2015)、MuMIn (Bartoń,2019)和car (Fox & Weisberg, 2019)软件包完成。使用ggplot2 (Wickham, 2016)软件包进行画图。
2 结果和分析
本研究共计使用种子2 520粒, 种子的回收率为62.02% (n = 1 563)。原地完好种子占2.62% (n = 66), 原地取食种子占6.75% (n = 170), 被扩散的种子占90.63% (n = 2 284)。在扩散且找回的种子中, 11.68% (n = 155)被贮藏。种子命运和扩散距离受到物种、岛屿大小和分散贮食动物多度的影响(表2)。Table 2
表2
表2千岛湖地区岛屿种子命运(广义线性混合效应模型)和扩散距离(线性混合效应模型)的最优模型结果
Table 2
| 固定效应 Fixed factor | 原地完好率 IIS (χ2) | 原地取食率 EIS (χ2) | 扩散率 R (χ2) | 扩散后贮藏率 CAR (χ2) | 扩散距离 DD (F) |
|---|---|---|---|---|---|
| 物种 Species | 13.61* | 32.63*** | 40.40*** | 38.17*** | 19.40*** |
| 岛屿大小 Island size | - | 3.42 | 6.05* | 1.99 | 1.49 |
| 分散贮食动物相对多度 ASH | 0.004 | - | - | 5.42* | 1.75 |
| 种子产量 Seed production | - | - | 0.33 | - | - |
| 物种×岛屿大小 Species × Island size | - | 30.40*** | 15.30 | 10.79 | 1.86· |
| 物种×分散贮食动物相对多度 Species ×ASH | 11.55* | - | - | 3.15 | 2.81* |
| 物种×种子产量 Species × Seed production | - | - | 10.52· | - | - |
新窗口打开|下载CSV
实验期间, 通过红外相机监测, 发现鼠类是千岛湖壳斗科种子的主要分散贮食动物, 赤腹松鼠在63号岛屿(占45.37%)以及松鸦(Garrulus glandarius)在58 (占63.64%)、60 (占42.65%)和B7 (占23.53%)号岛屿也是重要的分散贮食动物(附录I)。Spearman相关性检验表明, 分散贮食动物相对多度(r = 0.16, p= 1.00)、种子产量(r = 0.01, p= 1.00)与岛屿面积均不相关。
2.1 种子留存动态
6个物种种子的留存动态存在显著差异(Wald = 21.09, p< 0.001,图2A)。其中, 苦槠种子遭到分散贮食动物捕食和扩散的速率显著大于青冈、石栎、麻栎和短柄枹这4个物种的种子(p< 0.01), 其次为白栎种子, 其留存时间显著短于麻栎和短柄枹的种子(p< 0.05)。相较大陆, 岛屿上的种子更快被分散贮食动物捕食和扩散(Wald = 47.72, p< 0.001,图2B), 其中小岛的速度最快(p< 0.001), 而岛屿边缘的种子比岛屿内部更快遭到分散贮食动物捕食和扩散(Wald = 55.61, p< 0.001,图2C)。图2
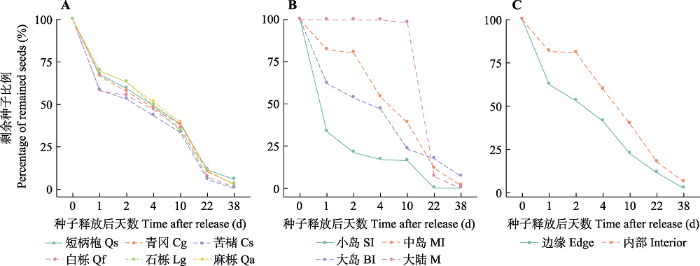 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2千岛湖研究区种子留存动态。A, 不同物种。B, 不同生境类型。C, 边缘梯度。
Fig. 2Survival analysis of experimental seeds in the study area of the Thousand Island Lake region. A, Different species. B, Different habitat types. C, Edge gradients. Cg, Cyclobalanopsis glauca; Cs, Castanopsis sclerophylla; Lg, Lithocarpus glaber; Qa, Quercus acutissima; Qf, Quercus fabri; Qs, Quercus serrata. BI, big island; M, mainland; MI, medium island; SI, small island.
2.2 种子命运
6个物种种子的原地完好率、原地取食率、扩散率和扩散后贮藏率均存在显著差异(表2)。短柄枹和青冈种子有最高的原地完好率、原地取食率和最低的扩散率; 苦槠种子有最低的原地完好率, 白栎种子有最低的扩散后贮藏率; 石栎和麻栎种子有最高的扩散率和扩散后贮藏率以及最低的原地取食率(图3A、3B; 附录II)。图3
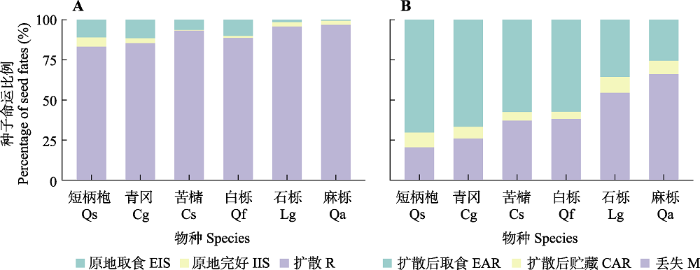 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3千岛湖研究区6个壳斗科物种种子命运比例。A, 释放种子分为原地取食、原地完好和扩散3种情况。B, 扩散种子分为扩散后取食、扩散后贮藏和丢失3种情况。
Fig. 3Seed fates of six Fagaceae species in the study area of the Thousand Island Lake region. A, Released seeds categorized as eaten in situ (EIS), intact in situ (IIS) and removed (R). B, Dispersed seeds categorized as eaten after removal (EAR), cached after removal (CAR) and missing (M). Cg, Cyclobalanopsis glauca; Cs, Castanopsis sclerophylla; Lg, Lithocarpus glaber; Qa, Quercus acutissima; Qf, Quercus fabri; Qs, Quercus serrata.
种子原地取食率在物种与岛屿大小间存在显著的交互作用(表2), 大岛上白栎种子的原地取食率显著高于小岛(z = 2.85, p< 0.01,图4A)。岛屿大小对种子扩散率有显著影响(表2), 小岛的种子扩散率显著高于大岛(z = 2.26, p< 0.05,图4B)。分散贮食动物相对多度对种子的扩散后贮藏率有显著负效应(z = -2.30, p< 0.05,表2)。生境类型和边缘梯度对种子命运无显著影响。
图4
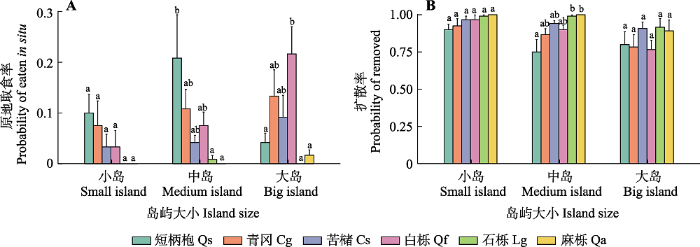 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4千岛湖研究区物种和岛屿大小对种子原地取食率和扩散率的影响(平均值+标准差)。A, 原地取食率。B, 扩散率。不同小写字母表示同一岛屿大小物种间差异显著(p < 0.05)。
Fig. 4Effects of species and island size on seed fates in the study area of the Thousand Island Lake region (mean + SD). A, Probability of seeds eaten in situ. B, Probability of seeds removed. Different lowercase letters indicate significant differences at the 0.05 level among different species for the same island size. Cg, Cyclobalanopsis glauca; Cs, Castanopsis sclerophylla; Lg, Lithocarpus glaber; Qa, Quercus acutissima; Qf, Quercus fabri; Qs, Quercus serrata.
2.3 种子扩散距离
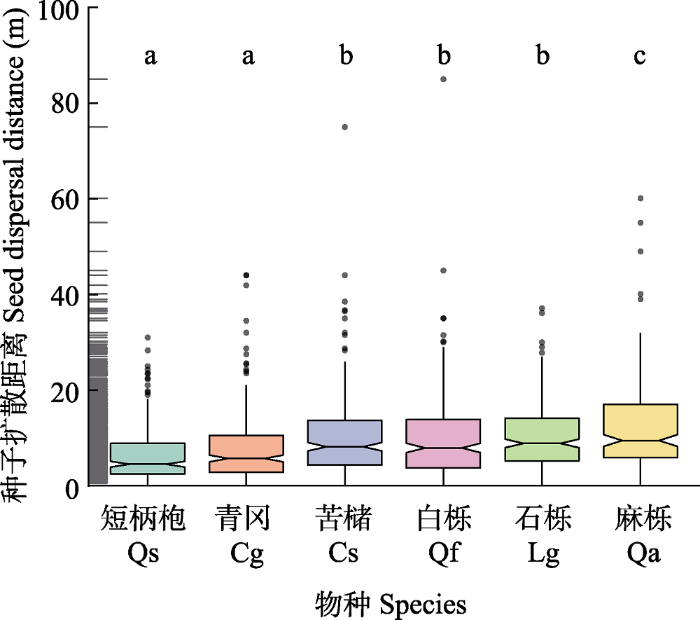
6个物种种子的扩散距离存在显著差异(表2), 扩散距离由大到小依次为麻栎、石栎、苦槠、白栎、青冈和短柄枹(图5)。种子扩散距离在物种与分散贮食动物相对多度间存在显著的交互作用(表2)。苦槠(t= -2.14,p< 0.05)、青冈(t = -2.45,p< 0.05)和麻栎(t = -2.21, p= 0.05)种子的扩散距离随分散贮食动物的增加而减小(图6B)。生境类型和边缘梯度对种子扩散距离无显著影响。图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5千岛湖研究区6个壳斗科物种种子扩散距离。不同小写字母表示物种间差异显著(p < 0.05)。
Fig. 5Seed dispersal distances of six Fagaceae plants in the study area of the Thousand Island Lake region. Different lowercase letters indicate significant difference at the 0.05 level among different species. Cg, Cyclobalanopsis glauca; Cs, Castanopsis sclerophylla; Lg, Lithocarpus glaber; Qa, Quercus acutissima; Qf, Quercus fabri; Qs, Quercus serrata.
图6
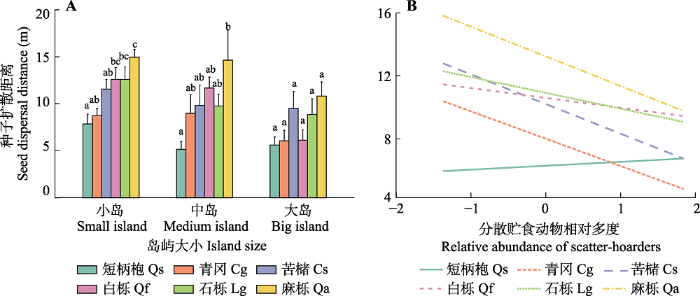 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6千岛湖研究区物种、岛屿大小和分散贮食动物相对多度对种子扩散距离的影响。A, 岛屿大小和6个物种种子扩散距离的关系(平均值+标准差)。B, 分散贮食动物相对多度和6个物种种子扩散距离的关系。不同小写字母表示同一岛屿大小物种间差异显著(p < 0.05)。
Fig. 6Effects of species, island size and relative abundance of scatter-hoarders on seed dispersal distances in the study area of the Thousand Island Lake region. A, Relationships between island size and seed dispersal distances for six species (mean + SD). B, Relationships between relative abundance of scatter-hoarders and seed dispersal distances for six species. Different lowercase letters indicate significant difference at the 0.05 level among different species for the same island size. Cg, Cyclobalanopsis glauca; Cs, Castanopsis sclerophylla; Lg, Lithocarpus glaber; Qa, Quercus acutissima; Qf, Quercus fabri; Qs, Quercus serrata.
3 讨论
分散贮食动物在觅食过程中会权衡种子所含的营养和次生代谢物质以及处理、运输种子需消耗的能量(Wang et al., 2014; Lichti et al., 2017), 导致不同物种种子面临不同的命运。本研究发现6种壳斗科植物种子的捕食和扩散模式存在显著差异(表2), 青冈和短柄枹、苦槠和白栎, 以及石栎和麻栎之间分别具有较为接近的种子捕食和扩散模式(图3; 附录II), 证明多种种子共存的情况下, 分散贮食动物对不同物种种子做出了不同的觅食选择。一些研究发现, 种子大小可反映种子价值, 是决定种子命运和扩散距离的关键因素(Xiao et al., 2005; Wang & Chen, 2009), 种子越大的物种具有越低的种子原地取食率和越大的种子扩散率和扩散距离(Xiao et al., 2005; Moore et al., 2007; Yi & Wang, 2015)。本研究中质量最小的短柄枹和青冈种子具有最高的原地完好率、原地取食率及最小的扩散率和扩散距离, 而质量最大的麻栎种子具有最低的原地取食率和最大的扩散距离的结果(图3, 图5; 附录II)与之一致, 因此推测种子大小在壳斗科植物与分散贮食动物的关系中扮演重要角色。此外, 种子外壳厚度会影响分散贮食动物的觅食偏好。Wang等(2014)发现石栎属种子因坚硬的种子外壳需要更长的处理时间, 给鼠类造成更大的被捕食风险, 导致更低的种子取食率和更高的贮藏率, 本研究中石栎种子的命运(附录II)与之一致。然而, 生境片段化及其造成的面积效应和边缘效应对种子的捕食和扩散产生了一定影响(图2B、2C; 表2)。Zeng等(2019)表明岛屿上大型兽类的丧失导致鼠类数量增加, 岛屿面积的减小间接增强鼠类间的种子竞争。而分散贮食动物在处理一粒种子时需要考虑错失其他种子资源的代价(Lichti et al., 2017), Hopewell等(2008)发现在资源有限且竞争者存在时, 灰松鼠(Sciurus carolinensis)会减少贮藏种子的时间而更频繁地返回种子资源地进行觅食。这解释了种子在岛屿比在大陆、在小岛比在中大岛更快被分散贮食动物光顾, 且小岛比大岛有更高种子扩散率的现象。而岛屿边缘的种子比在岛屿内部更快遭到分散贮食动物光顾, 可能缘于常见种白栎为不耐阴种, 在岛屿边缘有更多分布, 导致分散贮食动物在边缘活动更频繁。
而面积效应对不同物种的种子捕食和扩散模式产生了不同影响。分散贮食动物在小岛和中岛上倾向于原地取食短柄枹这种小种子, 并且质量越大的种子具有越远的扩散距离, 而随着面积增加, 分散贮食动物展现出对白栎种子的取食偏好, 物种间扩散距离的差异也有所减小(图4A、4B, 图6A)。Morán-López等(2018)发现小的生境片段, 啮齿动物面临激烈的种内竞争, 会对种子性状进行更多的选择以确保获得最有价值的食物, 导致大种子被优先选择和贮藏。这解释了小岛和中岛上小种子短柄枹更多被原地取食、大种子麻栎更多被扩散的命运差异。而在大岛上, 分散贮食动物间竞争相对缓和, 不需要为减少错失种子资源的代价而快速扩散种子, 从而有更充分的种子处理时间, 导致质量较大的白栎种子原地取食率增加。而小岛上较大的竞争强度也导致更高的种子偷食率, Moore等(2007)认为扩大贮藏间距能减小偷食成功率, 因此分散贮食动物在小岛上会花费更多能量扩散更有价值的种子, 导致种子较大的麻栎、石栎和白栎在小岛上有较远的扩散距离(图6A)。岛屿大小对不同物种的种子产生了不同影响, 决定了该物种后续的种子萌发和幼苗建成, 而这可能是不同岛屿植物更新和群落构建过程差异的重要原因。
此外, 分散贮食动物多度对种子扩散后贮藏率有显著负效应(表2), 意味着过多分散贮食动物的存在不利于种子的最终存活。对分散贮食动物进行更精确的监测以及增加种子命运追踪时间, 将有助于我们更好地理解生境片段化如何改变分散贮食动物种类和密度从而影响种子的命运。
综上, 本研究在片段化景观中探究了多物种的捕食和扩散模式, 强调了物种和片段大小对种子命运和扩散距离的综合影响, 发现物种是造成种子捕食与扩散模式差异的主要因素, 生境片段化带来的面积效应和边缘效应影响种子留存时间和扩散率, 且不同物种的种子命运和扩散距离受面积效应影响不同, 所获结果为理解生境片段化对植物更新及生物多样性维持的影响提供了新的线索。不过, 因岛屿面积和工作量的限制, 本研究设置的种子释放点数量偏少, 并且没有对不同物种的种子性状进行全方位的测定, 因而未能深入探究生境片段化对不同物种种子捕食和扩散模式影响差异的机制。
致谢
浙江大学曾頔、贵州师范大学金毅和温州大学刘金亮对论文进行修改并提供建设性意见, 浙江大学巫东豪等采集种子雨数据, 赖仲卿和当地村民帮助野外实验, 特此致谢。附录I 千岛湖地区实验岛屿和大陆样地基本信息
Supplement I Basic information of twelve study islands and three mainland study sites in the study area of the Thousand Island Lake regionhttps://www.plant-ecology.com/fileup/1005-264X/PDF/cjpe.2020.0079-S1.pdf
附录II 千岛湖地区6种壳斗科植物的种子命运
Supplement II Seed fates of six Fagaceae species in the study area of the Thousand Island Lake regionhttps://www.plant-ecology.com/fileup/1005-264X/PDF/cjpe.2020.0079-S1.pdf
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1111/j.1744-7429.2011.00782.xURL [本文引用: 3]
[
URL [本文引用: 1]
[本文引用: 1]
DOI:10.17520/biods.2018327URL [本文引用: 1]
DOI:10.17520/biods.2018327 [本文引用: 1]

多度是衡量物种种群数量的参数之一, 多度的动态及其影响因素是种群生态学研究的经典问题。物种相对多度指数(relative abundance index, RAI)作为一种简单、便利的指标, 广泛应用于动物本底清查中。但RAI易受物种自身特征、探测率和环境因素的影响, 需要结合其他物种数量分析方法, 以验证其与种群大小的相关性。随着红外相机技术在野生动物调查中的广泛应用, 用红外相机数据估计动物种群数量的研究越来越多。目前, 基于红外相机数据计算RAI的方法有多种, 不同计算方法和应用范围存在差异, 亟需对现有方法和应用进行梳理。本文综述了根据红外相机数据计算物种相对多度的4种主要方法: (1)拍摄一张有效照片所需要的天数; (2)基于单位调查强度的物种拍摄率; (3)每个位点每天的物种拍摄率; (4)某一物种的照片数占所有物种的比例。总结了我国野生动物监测调查中采用红外相机方法计算RAI的应用现状。国内的研究主要采用第2种和第4种计算方法, 其中约72.5%的研究论文应用第2种计算方法, 而第4种方法一般适用于群落中的物种组成比较。我们建议根据红外相机数据计算RAI时尽量使用第2种计算方法, 这有助于研究或管理人员对不同研究中的物种RAI进行比较分析。
DOI:10.1111/inz2.2017.12.issue-6URL [本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
DOI:10.1146/annurev.ecolsys.34.011802.132419URL [本文引用: 1]
DOI:10.1016/j.biocon.2005.10.049URL [本文引用: 2]
3rd ed. Sage Publications,
[本文引用: 1]
DOI:10.1016/j.gecco.2015.04.008URL [本文引用: 1]
DOI:10.1126/sciadv.1500052URL [本文引用: 1]
DOI:10.1093/beheco/arn095URL [本文引用: 1]
DOI:10.1023/A:1021225728493URL [本文引用: 1]
DOI:10.1016/j.biocon.2013.04.025URL [本文引用: 1]
DOI:10.1111/brv.2017.92.issue-1URL [本文引用: 4]
DOI:10.1890/13-1657.1URL
[本文引用: 1]
DOI:10.1111/jec.v108.2URL [本文引用: 2]
DOI:10.1111/ecog.01592URL [本文引用: 1]
DOI:10.1890/07-0247.1URL [本文引用: 3]
DOI:10.1111/oik.2015.v124.i11URL [本文引用: 3]
DOI:10.1007/s11258-018-0798-2URL [本文引用: 1]
Master degree dissertation,
[本文引用: 1]
硕士学位论文,
[本文引用: 1]
[2020-01-06]. https://CRAN.R-project.org/package=survival.
URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/BF02857850URL [本文引用: 1]
DOI:10.1098/rstb.2009.0205URL [本文引用: 3]
DOI:10.1007/s12229-011-9093-9URL [本文引用: 1]
PMID:19967858 [本文引用: 1]
Scatter-hoarding rodents play an important dispersal role for many large-seeded plants. Seed traits affect their foraging behavior; however, it is difficult to isolate their effects because of the covariance among traits. Here, we used artificial seeds to partition the effects of seed size, tannin content, and nutrient content on scatter-hoarding rodents in a natural pine forest in Northwest Yunnan, China. Apodemus, a common genus of Old World rodents, consistently consumed small seeds in situ but removed medium-sized seeds (1.2-2.5 cm in diameter) and transported bigger seeds farther. Seed nutrient and tannin contents also significantly influenced rodents' behavior, but response varied substantially between years. Rodent behavior did involve some aspects of multivariate optimization. Our results strongly indicate that seed size is a decisive factor for scatter-hoarding rodents in the choice between seed predation and dispersal, while nutrient and tannin content played a less consistent role, possibly responding to confounding factors in the community. This result also has important implications for seed production by trees, which can improve the probability of long-distance dispersal of high-quality offspring by simply making them larger. The ability to tease apart the relative influence of different seed traits on the behavior of predators provides powerful insight into this important coevolutionary dynamic.
DOI:10.1007/s00442-016-3793-0PMID:28000021 [本文引用: 1]
Individual variation in seed size and seed production is high in many plant species. How does this variation affect seed-dispersing animals and, in turn, the fitness of individual plants? In this study, we first surveyed intraspecific variation in seed mass and production in a population of a Chinese white pine, Pinus armandii. For 134 target trees investigated in 2012, there was very high variation in seed size, with mean seed mass varying among trees almost tenfold, from 0.038 to 0.361?g. Furthermore, 30 of the 134 trees produced seeds 2?years later, and for these individuals there was a correlation in seed mass of 0.59 between years, implying consistent differences among individuals. For a subset of 67 trees, we monitored the foraging preferences of scatter-hoarding rodents on a total of 15,301 seeds: 8380 were ignored, 3184 were eaten in situ, 2651 were eaten after being cached, and 395 were successfully dispersed (cached and left intact). At the scale of individual seeds, seed mass affected almost every decision that rodents made to eat, remove, and cache individual seeds. At the level of individual trees, larger seeds had increased probabilities of both predation and successful dispersal: the effects of mean seed size on costs (predation) and benefits (caching) balanced out. Thus, despite seed size affecting rodent decisions, variation among trees in dispersal success associated with mean seed size was small once seeds were harvested. This might explain, at least in part, the maintenance of high variation in mean seed mass among tree individuals.
DOI:10.1111/1749-4877.12086URL [本文引用: 5]
DOI:10.1007/s10342-016-0983-6URL [本文引用: 1]
[本文引用: 1]
//Soul'e ME. Conservation Biology: the Science of Scarcity and Diversity. Sinauer Associates, Sunderland, England.
[本文引用: 1]
DOI:10.1016/j.foreco.2005.10.054URL [本文引用: 1]
DOI:10.1111/jec.2013.101.issue-5URL [本文引用: 1]
DOI:10.1016/j.actao.2005.04.006URL [本文引用: 2]
DOI:10.3724/SP.J.1003.2014.14176URL [本文引用: 1]
DOI:10.3724/SP.J.1003.2014.14176 [本文引用: 1]
作为典型的陆桥岛屿, 千岛湖成为检验栖息地片段化理论的自然研究平台。2011年5月1日至2014年3月31日, 我们在千岛湖32个岛屿和1个大陆对照样点布设了60台红外相机, 对千岛湖体型较大的地栖兽类及其最小监测时长进行了监测和研究。在27,798个相机日的监测中, 共获得动物影像照片23,639张, 照片清晰、可进行物种鉴定的有2,414张, 占照片总数的10.2%; 其中体型较大的地栖兽类独立照片988张, 识别为9个物种: 穿山甲(Manis pentadactyla)、黄麂(Muntiacus reevesi)、野猪(Sus scrofa)、华南兔(Lepus sinensis)、马来豪猪(Hystrix brachyuran)、猪獾(Arctonyx collaris)、鼬獾(Melogale moschata)、花面狸(Paguma larvata)和豹猫(Prionailurus bengalensis), 平均独立照片拍摄率为40.9%。种-面积曲线研究表明, 岛屿上的地栖兽类物种丰富度随着岛屿面积的增大而增大, 曲线的z值为0.27。大岛(>10 ha)中, 最小监测时长随面积增加而增加, 而小岛没有明显趋势; 最小监测时长随隔离度增加而减小, 但关系不显著(d.f. = 20, F = 3.067, P = 0.095), 表明建湖后栖息地的片段化与岛屿化导致了一些对面积或栖息地较为敏感的大型兽类在小岛屿上的局部灭绝。因此, 我们建议对于面积较大的片段化栖息地, 红外相机应保证较长的最小监测时长, 而面积较小的片段化栖息地在监测中应根据隔离度、基质性质、物种种类适时调整调查强度, 以完整反映当地物种实际情况。
DOI:10.1007/s11258-014-0421-0URL [本文引用: 2]
DOI:10.1111/1365-2745.13122 [本文引用: 3]
Habitat loss and fragmentation often leads to defaunation of large-bodied mammals, and their loss could trigger release from top-down control or food resource competition for small mammal seed dispersers, which in turn may affect the effectiveness of seed dispersal by altering the number of dispersed seeds or the manner in which they are dispersed. Although rodents are primary seed dispersers in habitat subjected to defaunation, changes in seed dispersal effectiveness of rodents along mammalian defaunation gradients, and empirical support for mechanisms underlying alteration of this ecological process, are unclear. We assessed the direct and indirect effects of forested area and isolation on seed dispersal effectiveness of rodents on 21 study islands with varying levels of defaunation in the Thousand Island Lake, China. We used camera sampling, live traps and semi-quantitative acorn counts to assess occurrence of large-bodied mammal species, relative abundance of small rodent species and seed crop size respectively. Seed dispersal, post-dispersal seed survival, seedling emergence, and seedling survival were estimated by tracking fates of tagged acorns and by planting acorns in exclosures. Forested area had positive indirect effects on seed dispersal effectiveness through defaunation and rodent competition for acorns, whereas isolation had negative direct and weaker positive indirect effects on seed dispersal effectiveness mediated by loss of large-bodied mammals and rodent competition for acorns. Loss of large-bodied mammals negatively affected seed dispersal effectiveness indirectly by virtue of its impact on rodent competition for acorns. Seed dispersal effectiveness exhibited a unimodal relationship with intensity of rodent competition for acorns, peaking at intermediate levels. Synthesis. Indirect effects of island attributes mediated by defaunation of large-bodied mammals on small or isolated islands appear to drive altered competition for food among rodents and decreased seed dispersal effectiveness. Altered interactions between acorns and their rodent consumers/dispersers can substantially affect oak population demography in the Thousand Island Lake system. More broadly, our findings highlight the importance to the seed dispersal process of multiple interwoven effects between habitat fragmentation and defaunation of large-bodied mammals.
Increased productivity and reduced seed predation favor a large-seeded palm in small Atlantic forest fragments
3
2012
... 一些研究认为, 生境片段化带来的生境面积减小和边缘效应是影响动植物关系如种子捕食和扩散的重要因素(Fleury & Galetti,
... )发现小的森林片段隐蔽条件较差, 鼠类更易受天敌威胁, 且对种子的竞争强度减小, 导致更低的种子扩散率、贮藏率和扩散距离; Andreazzi等(
... 已有很多研究发现种子大小和种子外壳厚度等性状会影响分散贮食动物对种子的选择(Moore et al.,
MuMIn: multi-model inference. R package version 1.43.15
1
2020
... 本研究的所有分析均由R 3.6.1软件和相应软件包完成.Cox回归由survival (Therneau & Grambsch,
Fitting linear mixed-effects models using lme4
1
2015
... 本研究的所有分析均由R 3.6.1软件和相应软件包完成.Cox回归由survival (Therneau & Grambsch,
物种相对多度指数在红外相机数据分析中的应用及局限
1
2019
... 统计红外相机独立照片数量(同一个相机同种物种相隔1.5 h以上的照片), 计算出每个岛屿种子释放后的30天内平均每天出现在释放点的分散贮食动物独立照片数量, 经对数转换后作为该岛的分散贮食动物相对多度(徐爱春等,
物种相对多度指数在红外相机数据分析中的应用及局限
1
2019
... 统计红外相机独立照片数量(同一个相机同种物种相隔1.5 h以上的照片), 计算出每个岛屿种子释放后的30天内平均每天出现在释放点的分散贮食动物独立照片数量, 经对数转换后作为该岛的分散贮食动物相对多度(徐爱春等,
Effects of fragmentation on the seed predation and dispersal by rodents differ among species with different seed size
2
2017
... 一些研究认为, 生境片段化带来的生境面积减小和边缘效应是影响动植物关系如种子捕食和扩散的重要因素(Fleury & Galetti,
... 已有很多研究发现种子大小和种子外壳厚度等性状会影响分散贮食动物对种子的选择(Moore et al.,
2
1998
... 果期参考《浙江植物志》(浙江植物志编委会,
... Nut-ripening periods are referred from the Flora of Zhejiang Province(Editorial Committee of Flora of Zhejiang,
2
1998
... 果期参考《浙江植物志》(浙江植物志编委会,
... Nut-ripening periods are referred from the Flora of Zhejiang Province(Editorial Committee of Flora of Zhejiang,
2
1993
... 果期参考《浙江植物志》(浙江植物志编委会,
... Nut-ripening periods are referred from the Flora of Zhejiang Province(Editorial Committee of Flora of Zhejiang,
2
1993
... 果期参考《浙江植物志》(浙江植物志编委会,
... Nut-ripening periods are referred from the Flora of Zhejiang Province(Editorial Committee of Flora of Zhejiang,
Effects of habitat fragmentation on biodiversity
1
2003
... 随着人类对自然的干扰加大, 生境片段化对森林更新和生物多样性的影响受到越来越多的关注.生境片段化是指连续的生境被分割成多个相互隔离的小斑块(片段)的过程(Wilcove et al.,
Forest fragment size and microhabitat effects on palm seed predation
2
2006
... 一些研究认为, 生境片段化带来的生境面积减小和边缘效应是影响动植物关系如种子捕食和扩散的重要因素(Fleury & Galetti,
... )认为小片段更好的光照条件利于种子生产, 而中大型种子捕食者丧失, 导致较低的种子捕食率和较高的存活率; Fleury和Galetti (
An R Companion to Applied Regression
1
2019
... 本研究的所有分析均由R 3.6.1软件和相应软件包完成.Cox回归由survival (Therneau & Grambsch,
Defaunation of large mammals leads to an increase in seed predation in the Atlantic forests
1
2015
... 一些研究认为, 生境片段化带来的生境面积减小和边缘效应是影响动植物关系如种子捕食和扩散的重要因素(Fleury & Galetti,
Habitat fragmentation and its lasting impact on Earth’s ecosystems
1
2015
... 随着人类对自然的干扰加大, 生境片段化对森林更新和生物多样性的影响受到越来越多的关注.生境片段化是指连续的生境被分割成多个相互隔离的小斑块(片段)的过程(Wilcove et al.,
Effects of competition and food availability on travel time in scatter-hoarding gray squirrels (Sciurus carolinensis)
1
2008
... 然而, 生境片段化及其造成的面积效应和边缘效应对种子的捕食和扩散产生了一定影响(
Edges effects on seed predation by rodents in deciduous forests of northern Switzerland
1
2003
... 一些研究认为, 生境片段化带来的生境面积减小和边缘效应是影响动植物关系如种子捕食和扩散的重要因素(Fleury & Galetti,
Cascading effects of contemporaneous defaunation on tropical forest communities
1
2013
... 一些研究认为, 生境片段化带来的生境面积减小和边缘效应是影响动植物关系如种子捕食和扩散的重要因素(Fleury & Galetti,
Seed fate and decision-making processes in scatter-hoarding rodents
4
2017
... 种子扩散对植物更新具有十分重要的意义(Vander Wall,
... 已有很多研究发现种子大小和种子外壳厚度等性状会影响分散贮食动物对种子的选择(Moore et al.,
... 分散贮食动物在觅食过程中会权衡种子所含的营养和次生代谢物质以及处理、运输种子需消耗的能量(Wang et al.,
... 然而, 生境片段化及其造成的面积效应和边缘效应对种子的捕食和扩散产生了一定影响(
Mast species composition alters seed fate in north American rodent- dispersed hardwoods
2014
1
2013
... 随着人类对自然的干扰加大, 生境片段化对森林更新和生物多样性的影响受到越来越多的关注.生境片段化是指连续的生境被分割成多个相互隔离的小斑块(片段)的过程(Wilcove et al.,
Environmental filtering underpins the island species-area relationship in a subtropical anthropogenic archipelago
2
2020
... 研究地位于浙江省杭州市淳安县境内的新安江水库(29.37°-29.83° N, 118.57°-119.25° E;
... ,
Patch size, shape and edge distance influence seed predation on a palm species in the Atlantic forest
1
2016
... 一些研究认为, 生境片段化带来的生境面积减小和边缘效应是影响动植物关系如种子捕食和扩散的重要因素(Fleury & Galetti,
Determinants of seed removal distance by scatter- hoarding rodents in deciduous forests
3
2007
... 已有很多研究发现种子大小和种子外壳厚度等性状会影响分散贮食动物对种子的选择(Moore et al.,
... 分散贮食动物在觅食过程中会权衡种子所含的营养和次生代谢物质以及处理、运输种子需消耗的能量(Wang et al.,
... 而面积效应对不同物种的种子捕食和扩散模式产生了不同影响.分散贮食动物在小岛和中岛上倾向于原地取食短柄枹这种小种子, 并且质量越大的种子具有越远的扩散距离, 而随着面积增加, 分散贮食动物展现出对白栎种子的取食偏好, 物种间扩散距离的差异也有所减小(
Effects of forest fragmentation on the oak-rodent mutualism
3
2015
... 一些研究认为, 生境片段化带来的生境面积减小和边缘效应是影响动植物关系如种子捕食和扩散的重要因素(Fleury & Galetti,
... ).例如, Morán-López等(
... )则发现中等大小片段中种子被捕食概率最高.边缘效应对种子命运的影响同样不一致, Morán-López等(
Fragmentation modifies seed trait effects on scatter-hoarders’ foraging decisions
1
2018
... 而面积效应对不同物种的种子捕食和扩散模式产生了不同影响.分散贮食动物在小岛和中岛上倾向于原地取食短柄枹这种小种子, 并且质量越大的种子具有越远的扩散距离, 而随着面积增加, 分散贮食动物展现出对白栎种子的取食偏好, 物种间扩散距离的差异也有所减小(
千岛湖库区马尾松林内种子雨和土壤种子库组成及动态
1
2017
... 自2013年, 在29个岛屿和大陆1 hm2样地内共设置250个种子雨收集框, 每月对种子雨收集框内的种子和果实进行一次收集、鉴定和称量, 具体设置方法见南歌(
千岛湖库区马尾松林内种子雨和土壤种子库组成及动态
1
2017
... 自2013年, 在29个岛屿和大陆1 hm2样地内共设置250个种子雨收集框, 每月对种子雨收集框内的种子和果实进行一次收集、鉴定和称量, 具体设置方法见南歌(
A package for survival analysis in S. R package version 2.38
1
2015
... 本研究的所有分析均由R 3.6.1软件和相应软件包完成.Cox回归由survival (Therneau & Grambsch,
1
2000
... 本研究的所有分析均由R 3.6.1软件和相应软件包完成.Cox回归由survival (Therneau & Grambsch,
1
1990
... 种子扩散对植物更新具有十分重要的意义(Vander Wall,
The evolutionary ecology of nut dispersal
1
2001
... 种子扩散对植物更新具有十分重要的意义(Vander Wall,
How plants manipulate the scatter- hoarding behaviour of seed-dispersing animals
3
2010
... 种子扩散对植物更新具有十分重要的意义(Vander Wall,
... ), 它们既能扩散种子, 同时也会捕食种子(Vander Wall,
... 已有很多研究发现种子大小和种子外壳厚度等性状会影响分散贮食动物对种子的选择(Moore et al.,
A comparison of frugivory and scatter-hoarding seed-dispersal syndromes
1
2012
... 种子扩散对植物更新具有十分重要的意义(Vander Wall,
Seed size, more than nutrient or tannin content, affects seed caching behavior of a common genus of Old World rodents
1
2009
... 分散贮食动物在觅食过程中会权衡种子所含的营养和次生代谢物质以及处理、运输种子需消耗的能量(Wang et al.,
Tree-to-tree variation in seed size and its consequences for seed dispersal versus predation by rodents
1
2017
... 已有很多研究发现种子大小和种子外壳厚度等性状会影响分散贮食动物对种子的选择(Moore et al.,
Seed traits and taxonomic relationships determine the occurrence of mutualisms versus seed predation in a tropical forest rodent and seed dispersal system
5
2014
... 已有很多研究发现种子大小和种子外壳厚度等性状会影响分散贮食动物对种子的选择(Moore et al.,
... 白栎(Quercus fabri)、短柄枹(Quercus serrata)、麻栎(Quercus acutissima)、苦槠(Castanopsis sclerophylla)、青冈(Cyclobalanopsis glauca)和石栎(Lithocarpus glaber)是千岛湖地区较常见的6种壳斗科植物, 它们的果期相互重叠(
... 对分散贮食动物相对多度、种子产量和岛屿面积进行Spearman相关性检验.采用Cox回归(Wang et al.,
... 分散贮食动物在觅食过程中会权衡种子所含的营养和次生代谢物质以及处理、运输种子需消耗的能量(Wang et al.,
... ; 附录II)与之一致, 因此推测种子大小在壳斗科植物与分散贮食动物的关系中扮演重要角色.此外, 种子外壳厚度会影响分散贮食动物的觅食偏好.Wang等(
Nutritional and defensive properties of Fagaceae nuts dispersed by animals: a multiple species study
1
2016
... 白栎(Quercus fabri)、短柄枹(Quercus serrata)、麻栎(Quercus acutissima)、苦槠(Castanopsis sclerophylla)、青冈(Cyclobalanopsis glauca)和石栎(Lithocarpus glaber)是千岛湖地区较常见的6种壳斗科植物, 它们的果期相互重叠(
1
2016
... 本研究的所有分析均由R 3.6.1软件和相应软件包完成.Cox回归由survival (Therneau & Grambsch,
Habitat fragmentation in the temperate zone
1
1986
... 随着人类对自然的干扰加大, 生境片段化对森林更新和生物多样性的影响受到越来越多的关注.生境片段化是指连续的生境被分割成多个相互隔离的小斑块(片段)的过程(Wilcove et al.,
Using seed-tagging methods for assessing post-dispersal seed fate in rodent- dispersed trees
1
2006
... 利用塑料片标签法(Xiao et al.,
Long-term seed survival and dispersal dynamics in a rodent-dispersed tree: testing the predator satiation hypothesis and the predator dispersal hypothesis
1
2013
... 种子扩散对植物更新具有十分重要的意义(Vander Wall,
Effects of seed size on dispersal distance in five rodent-dispersed Fagaceous species
2
2005
... 分散贮食动物在觅食过程中会权衡种子所含的营养和次生代谢物质以及处理、运输种子需消耗的能量(Wang et al.,
... ,
千岛湖片段化栖息地地栖哺乳动物的红外相机监测及最小监测时长
1
2014
... 统计红外相机独立照片数量(同一个相机同种物种相隔1.5 h以上的照片), 计算出每个岛屿种子释放后的30天内平均每天出现在释放点的分散贮食动物独立照片数量, 经对数转换后作为该岛的分散贮食动物相对多度(徐爱春等,
千岛湖片段化栖息地地栖哺乳动物的红外相机监测及最小监测时长
1
2014
... 统计红外相机独立照片数量(同一个相机同种物种相隔1.5 h以上的照片), 计算出每个岛屿种子释放后的30天内平均每天出现在释放点的分散贮食动物独立照片数量, 经对数转换后作为该岛的分散贮食动物相对多度(徐爱春等,
Context-dependent seed dispersal determines acorn survival of sympatric oak species
2
2015
... 已有很多研究发现种子大小和种子外壳厚度等性状会影响分散贮食动物对种子的选择(Moore et al.,
... 分散贮食动物在觅食过程中会权衡种子所含的营养和次生代谢物质以及处理、运输种子需消耗的能量(Wang et al.,
Cascading effects of forested area and isolation on seed dispersal effectiveness of rodents on subtropical islands
3
2019
... 一些研究认为, 生境片段化带来的生境面积减小和边缘效应是影响动植物关系如种子捕食和扩散的重要因素(Fleury & Galetti,
... 研究地位于浙江省杭州市淳安县境内的新安江水库(29.37°-29.83° N, 118.57°-119.25° E;
... 然而, 生境片段化及其造成的面积效应和边缘效应对种子的捕食和扩散产生了一定影响(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}