杨克彤
1 , 常海龙
1 , 陈国鹏
, , 1 , * , 俞筱押
2 , 鲜骏仁
3 1 甘肃农业大学林学院, 兰州 7300702 黔南民族师范学院旅游与资源环境学院, 贵州都匀 5580003 四川农业大学环境学院, 成都 611130Stomatal traits of main greening plant species in Lanzhou Ke-Tong YANG
1 , Hai-Long CHANG
1 , Guo-Peng CHEN
, , 1 , * , Xiao-Ya YU
2 , Jun-Ren XIAN
3 1 College of Forestry, Gansu Agricultural University, Lanhou 730070, China2 School of Tourism and Resource Environment, Qiannan Normal College for Nationalities, Duyun, Guizhou 558000, China3 College of Environmental Sciences, Sichuan Agricultural University, Chengdu 611130, China通讯作者: *陈国鹏: ORCID: 0000-0002-3854-8755 (
chgp1986@gmail.com )
编委: 赵长明
责任编辑: 李敏
收稿日期: 2020-07-30
接受日期: 2020-12-28
网络出版日期: 2021-02-20
基金资助: 国家自然科学基金 (31800352 )全国大学生创新创业训练计划项目 (201710733052 )全国大学生创新创业训练计划项目 (202010733027 )
Corresponding authors: *(
chgp1986@gmail.com )
Received: 2020-07-30
Accepted: 2020-12-28
Online: 2021-02-20
Fund supported: National Natural Science Foundation of China (31800352 )College Students’ Innovative Entrepreneurial Training Plan Program (201710733052 )College Students’ Innovative Entrepreneurial Training Plan Program (202010733027 )
摘要 气孔是植物叶片与外界环境进行水汽交换的门户, 利用气孔特征反映植物对环境变化的响应与适应有助于了解干旱区绿化植物的适应策略。于2019年7月通过气孔印迹法对兰州市40种主要绿化植物气孔性状进行观察与测定。采用标准化主轴估计和系统独立比较分析建立气孔性状间的相关关系; 通过计算Blomgerg’s
K 值以检验系统发育信号; 利用聚类分析和主成分分析对气孔性状特征进行功能群划分。结果表明: (1)在所研究的植物中, 气孔性状特征在个体间差异显著, 植物生长型(乔木、小乔木、灌木和草本)会显著影响气孔长度、宽度、开度和密度, 叶习性(落叶和常绿)仅对气孔开度有显著影响; (2)气孔长度与宽度、气孔开度与面积间均为显著的异速生长关系, 气孔密度与面积和开度间为负异速生长关系; (3)系统发育会对气孔性状的相关关系产生显著影响, 虽然气孔性状的系统发育保守性不强(
K < 1), 但气孔开度和气孔开张比具有显著的系统发育信号; 4)依据气孔性状特征可以将绿化植物划分为: 低密度-大面积、高密度-小面积和中等密度-适中面积共3种功能群。结合系统发育和异速生长理论能更好地解释气孔性状变异及适应策略。
关键词: 黄土高原 ;
绿化植物 ;
气孔性状 ;
异速生长 ;
系统发育 Abstract Aims Stomata are the main channels for water and gas exchange between plants and the atmosphere. Stomatal traits reflect plant responses to environmental changes, and thus could help us better understand plant adaptation strategies to arid environments.
Methods In July 2019, six traits for stomata were measured from 40 main greening plant species in Lanzhou, Gansu Province. One-way ANOVA was used to test the traits difference among the growth forms (tree, semi-tree, shrub and herb) and leaf habits (deciduous and evergreen). Standardized major axis estimation and phylogenetically independent contrast analysis were used to compare the allometry relationship between stomatal traits. Then the Blomberg’s
K value was calculated to determine phylogenetic signal. Functional groups were partition by k-means clustering and principal component analysis.
Important findings The stomatal traits varied significantly among greening plant species. Different growth forms (trees, semi-trees, shrubs and herbs) showed significant variations in stomatal length (
SL ), width (
SW ), opening level (
SOL ) and density (
SD ), while deciduous and evergreen plants only differed in
SOL . Significant allometric relations were detected between
SL vs.
SW ,
SOL vs.
SA . We also observed negative allometric relations between
SD vs.
SA and
SD vs.
SOL . Phylogeny significantly influenced the correlations among stomatal traits. Although the phylogenetic conservatism of stomatal traits was not strong (
K < 1),
SOL and stomatal opening ratio (
SOR ) showed significant phylogenetic signals (
p < 0.05). According to the stomatal traits, greening plants can be divided into three functional groups: lower-density with larger area, higher-density with smaller area, or medium- density with moderate area. The combination of phylogeny and allometric scaling relations can provide a better explanation for stomatal trait variations and adaptation strategies.
Keywords: Loess Plateau ;
greening plant ;
stomata ;
allometry ;
phylogeny PDF (1704KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 杨克彤, 常海龙, 陈国鹏, 俞筱押, 鲜骏仁. 兰州市主要绿化植物气孔性状特征. 植物生态学报, 2021, 45(2): 187-196. DOI:
10.17521/cjpe.2020.0257 YANG Ke-Tong, CHANG Hai-Long, CHEN Guo-Peng, YU Xiao-Ya, XIAN Jun-Ren.
Stomatal traits of main greening plant species in Lanzhou .
Chinese Journal of Plant Ecology , 2021, 45(2): 187-196. DOI:
10.17521/cjpe.2020.0257 气孔作为植物叶表皮的特殊结构, 是植物与外界环境进行物质交换的门户(宋璐璐等,
2011 ; 朱燕华等,
2011 ), 也是植物体内水汽逸散的主要通道(金鹰和王传宽,
2015 ; 范嘉智等,
2016 )。气孔易受环境条件的修饰, 其形态特征是植物进化过程中对外界环境长期适应的结果(王碧霞等,
2010 ; 朱燕华等,
2011 ; 温婧雯等,
2018 )。为适应(或响应)复杂多变的环境, 气孔一方面通过调节开闭来应对短期或局域环境变化, 另一方面则协调大小和密度来适应异质生境(Brodribb & Jordan,
2011 ; Drake
et al. ,
2013 ; 金鹰和王传宽,
2015 ; 高冠龙等,
2018 )。利用气孔特征反映植物对环境变化的响应有助于明晰植物应对水气耦合过程的策略。
气孔性状(如气孔密度)的种间变异被认为是植物叶片经济谱的一部分(Loranger & Shipley,
2010 ; 王瑞丽等,
2016 ), 它在维持植物稳定碳收益、调节水力结构、吸收速效养分和信息传递等方面具有重要作用(Mencuccini
et al. ,
2015 ; Fiorin
et al. ,
2016 )。因此, 量化气孔性状在异质生境中的变异程度并解析其应对策略有助于进一步阐明植物对环境的适应机制。然而, 多数研究仅侧重于气孔性状对自然环境(海拔、水分和温度等)和控制试验(光强、氮添加和CO
2 浓度等)的响应并得出了一系列结论, 如: 气孔长度与密度之间为负相关关系(任昱等,
2014 ; 姜星星等,
2018 ), 随海拔升高气孔宽度呈减小趋势(王瑞丽等,
2016 ), 土壤湿度增大会显著降低气孔密度(杨再强等,
2015 ; 夏振华等,
2018 ; 张大才等,
2018 ), 高CO
2 浓度会显著降低气孔开张比(贾剑波等,
2019 )等, 但忽视了气孔自身在发育过程中的优先发育性状及其相互间关系的协变与权衡。
系统发育和环境异质是影响植物性状发生分异的动力来源。气孔主要分布于植物叶片下表皮并参与调控碳水摄入与排出, 在其发育过程中迫于环境压力与生存维持, 气孔往往在多个维度存在权衡(Brodribb & Jordan,
2011 ; Brodribb
et al. ,
2013 ; 丁凌子等,
2014 ; Fiorin
et al. ,
2016 )。虽然气孔数量的增加有助于提高光合效率, 但过多的气孔会加速叶内水汽逸散(潘莹萍和陈亚鹏,
2014 ; 王学芳等,
2014 ; 宋慧清等,
2020 ), 而且气孔的建成需要消耗资源, 这就决定了单位面积的气孔数量存在分布上限。气孔密度与大小共同决定了最大可能气孔导度并影响最大同化速率, 在考虑气孔建成实际资源消耗与光合作用水气协调的过程中, 两个相互矛盾的刚需必然达到最优化(Sack & Scoffoni,
2013 )。气孔面积与开度分别代表着气孔发育与运动, 且气孔发育总是先于气孔运动, 尽管大气孔对于开度的调控不及小气孔灵敏, 但可以获得相对较多的碳收益。就此而言, 气孔发育和运动过程中资源分配不均甚至存在矛盾(Xu & Zhou,
2008 ; Drake
et al. ,
2013 )。在异质生境中, 气孔各性状并非独立变化, 而是通过性状或特征组合的协变与权衡来共同适应环境。有限资源在植物体内的分配与权衡已通过异速生长理论在多个尺度得到证实(陈国鹏等,
2016 ; 段贝贝等,
2016 ; 杨克彤等,
2020 ), 但以往研究着重于相对宏观的尺度如群落、个体、构件与器官, 而且忽视了系统发育对资源权衡的影响, 对微观尺度如气孔性状的权衡机制关注较少。为此, 本文希望通过研究气孔特征并结合系统发育来探讨微观尺度气孔性状间的生长关系, 以进一步揭示微观尺度的权衡过程, 并丰富植物生活史研究。
黄土高原的高温、干旱、强日照和城市内空气污染、土壤重金属危害与CO
2 施肥效应共同驱动着绿化植物的适应策略并使其在外部形态上加以表征(段贝贝等,
2016 )。本研究以兰州市主要绿化植物为研究对象, 测定气孔形态特征并结合系统发育以探究叶片气孔性状的权衡关系, 并试图回答以下问题: (1)气孔特征是否受植物生长型、叶习性及系统发育的影响? (2)气孔长度与宽度、气孔数量和密度与气孔面积之间是否存在权衡? (3)能否根据气孔特征进行物种功能群划分?
1 材料和方法 1.1 研究区概况 兰州市(103.40° E, 36.03° N, 海拔1 500 m)位于黄土高原西北部, 地势西南高东北低, 呈东西狭长带状盆地特征, 属于大陆性很强的温带季风气候。市区地貌以山地丘陵为主, 占72.9%, 沟壑纵横, 地形破碎, 谷深坡陡, 年平均气温为10.3 ℃, 年日照时间2 446 h, 无霜期180天, 年降水量327 mm左右, 主要集中在6-9月(占全年降水量的50%-70%)。土壤以灰钙土为主, 其次为栗钙土和灰褐土, 自然植被稀少, 主要是多年生禾草、旱生灌木和小乔木(段贝贝等,
2016 ; 杨克彤等,
2020 ), 覆盖率低于23%, 其中森林覆盖率仅为6.67%, 低于全国平均水平13.95%, 植被类型单一、结构简单, 是我国生态环境十分脆弱的地区之一。
兰州城市绿地占城区面积的36.2%, 绿化植物的种类以乔木和灌木占绝对优势, 草本植物次之。对市区内的主要绿化区域踏查后, 根据兰州市不同区域人流量、车流量等干扰, 将采样地点选择为生境相对一致的公园。本研究将目标绿化植物确定为栽培时间较长、已基本适应当地气候的乔木13种、小乔木4种、灌木16种和草本7种(附录I)。
1.2 数据测定 于2019年7月选择无风、晴朗的天气, 在公园进行植物叶片采集。于9:00-11:00在选定好的3株标准木四周及中部分别采集10片完整、成熟叶片, 立即置于FAA固定液(70%酒精、38%甲醛、冰醋酸, 体积比为18:1:1)并带回室内, 固定24 h后进行气孔性状测定。首先, 随机取出5片已固定好的叶片用70%酒精冲洗残液, 后用蒸馏水冲洗并拭干水分, 然后在叶片背面中部选择一片避开主脉的区域(约1 cm
2 )涂上无色透明指甲油, 晾干后揭下制成临时装片并观察气孔特征(张大才等,
2018 )。观察、拍照时采用Imager A2生物显微镜(Zeiss, Jena, Germany), 每个装片选取3个镜头, 分别记录视野面积内气孔总数(
SNO )和开张气孔数(
OSN ), 气孔长度(
SL )、宽度(
SW )、面积(
SA )与气孔的孔径开度(
AOL )通过Image J软件计算, 气孔开张比(
SOR ) = (开张气孔数/气孔总数) × 100%, 气孔密度(
SD ) =气孔总数/视野面积(400 μm × 375 μm = 0.15 mm
2 )。鉴于自然界中多数植物的气孔主要分布在叶片下表皮, 故仅以下表皮气孔作为研究对象。气孔保卫细胞的长轴长度(μm)和短轴长度(μm)分别代表气孔长度和宽度, 将气孔开度定义为其孔径开度与宽度的比值, 即气孔开度(
SOL ) = (孔径开度/气孔宽度) × 100%, 介于0-100%之间, 值越大表示开度越大。
1.3 数据分析 将实测气孔性状参数的平均值作为该物种的气孔性状, 采用单因素方差分析检验植物种间、生长型和叶习性对气孔性状参数的影响, 用最小显著差异(
LSD )法对不同生长型和叶习性的气孔性状进行多重比较。将各物种气孔性状均值进行lg转化并做出性状间的散点图, 采用标准化主轴估计方法(SMA)计算性状间的斜率, 并检验斜率与1.0 (或-1.0)的差异。在R 4.0.3软件中使用ape包和picante包进行系统独立比较(PIC)分析, 将产生的去除系统发育信息的数据用最小二乘法(OLS)进行回归分析并比较与斜率1.0的差异, 即可明确系统发育是否会对气孔性状关系产生影响。计算Blomgerg’s
K 统计量以检验系统发育信号, 当
K > 1时, 表示气孔性状具有强的系统发育信号; 当
K < 1时, 表示气孔性状具有弱的系统发育信号。取40种植物气孔性状的平均值进行
K -means均值聚类, 将聚类结果划分为3类功能群并进行主成分分析, 以此探讨气孔性状的整体相似性。统计、计算使用Microsoft Excel 2010软件进行, 作图使用Origin 2019软件, 显著水平均采用
p = 0.05。
2 结果和分析 2.1 绿化植物气孔的基本特征 物种间气孔特征差异显著(
图1 ), 费菜(
Phedimus aizoon )的气孔长度、宽度和面积均最大, 分别为(105.01 ± 1.86) μm、(87.12 ± 0.94) μm和(7 138.82 ± 243.59) μm
2 , 但气孔密度最小((22.22 ± 1.81) No.·mm
-2 ); 刺槐(
Robinia pseudoacacia )的气孔长度和面积均最小, 分别为(18.67 ± 0.39) μm和(182.12 ± 37.93) μm
2 ; 旱柳(
Salix matsudana )的气孔宽度最小((11.97 ± 0.73) μm), 小叶黄杨(
Buxus sinica var.
parvifolia )的气孔开度最大(95.30% ± 7.30%), 栾树(
Koelreuteria paniculata )的气孔开度最小, 为15.24% ± 1.82%, 紫荆(
Cercis chinensis )气孔开张比最小(12.70% ± 2.04%), 山楂(
Crataegus pinnatifida )气孔开张比最大(100% ± 0.22%), 栾树气孔密度最大(344.44 No.·mm
-2 )。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1兰州市主要绿化植物的气孔性状。 Ace. tru., 元宝枫; Aco. cal., 菖蒲; Ail. alt., 臭椿; Amy. per., 碧桃; Ber. thu., 日本小檗; Bux. sin., 小叶黄杨; Car. cat., 山核桃; Cer. chi., 紫荆; Cer. ser., 日本晚樱; Cra. pin., 山楂; Euc. ulm., 杜仲; Euo. jap., 冬青卫矛; Euo. maa., 白杜; For. sus., 连翘; For. vir., 金钟花; Fra. chi., 白蜡树; Gin. bil., 银杏; Hem. Ful., 萱草; Hib. syr., 木槿; Hos. pla., 玉簪; Hyl. spe., 长药八宝; Iri. tec., 鸢尾; Koe. pan., 栾树; Lig. luc., 女贞; Lig. ova., 卵叶女贞; Phe. aiz., 费菜; Pla. gra., 桔梗; Pru. cer., 樱桃李; Pun. gra., 石榴; Pyr. xer., 木梨; Rob. pse., 刺槐; Ros. luc., 亮叶月季; Sal. mat., 旱柳; Sam. wil., 接骨木; Sop. jap., 龙爪槐; Syr. obl., 紫丁香; Syr. vil., 辽东丁香; Ulm. pum., 榆树; Vit. vin., 葡萄; Yul. den., 玉兰。 Fig. 1Stomatal traits of main greening plant species in Lanzhou. Ace. tru., Acer truncatum; Aco. cal., Acorus calamus; Ail. alt., Ailanthus altissima; Amy. per., Amygdalus persica f. duplex; Ber. thu., Berberis thunbergii; Bux. sin., Buxus sinica var. parvifolia; Car. cat., Carya cathayensis; Cer. chi., Cercis chinensis; Cer. ser., Cerasus serrulata var. lannesiana; Cra. pin., Crataegus pinnatifida; Euc. ulm., Eucommia ulmoides; Euo. jap., Euonymus japonicus; Euo. maa., Euonymus maackii; For. sus., Forsythia suspensa; For. vir., Forsythia viridissima; Fra. chi., Fraxinus chinensis; Gin. bil., Ginkgo biloba; Hem. ful., Hemerocallis fulva; Hib. syr., Hibiscus syriacus; Hos. pla., Hosta plantaginea; Hyl. spe., Hylotelephium spectabile; Iri. tec., Iris tectorum; Koe. pan., Koelreuteria paniculata; Lig. luc., Ligustrum lucidum; Lig. ova., Ligustrum ovalifolium; Phe. aiz., Phedimus aizoon; Pla. gra., Platycodon grandiflorus; Pru. cer., Prunus cerasifera; Pun. gra., Punica granatum; Pyr. xer., Pyrus xerophila; Rob. pse., Robinia pseudoacacia; Ros. luc., Rosa lucidissima; Sal. mat., Salix matsudana; Sam. wil., Sambucus williamsii; Sop. jap., Sophora japonica var. Japonica f. pendula; Syr. obl., Syringa oblata; Syr. vil., Syringa villosa subsp. wolfii; Ulm. pum., Ulmus pumila; Vit. Vin., Vitis vinifera; Yul. den., Yulania denudata. 2.2 生长型及叶习性对气孔特征的影响 单因素方差分析与多重比较结果(
图2 )表明, 生长型显著影响植物气孔特征。草本植物气孔长度极显著大于乔木(
p = 0.008), 显著大于小乔木(
p = 0.049), 宽度显著大于乔木(
p = 0.023), 气孔开度显著大于其他生长型植物(
p < 0.05); 乔木气孔密度显著大于草本(
p = 0.028); 气孔面积(
p = 0.216)与开张比在生长型间差异不显著(
p = 0.344); 叶习性仅显著影响气孔开度, 常绿植物叶片气孔开度显著大于落叶植物(
p = 0.043)。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2兰州市主要绿化植物生长型及叶习性对气孔性状的影响(平均值±标准误)。Dec, 落叶植物; Eve, 常绿植物; Her, 草本; Sem, 小乔木; Shr, 灌木; Tre, 乔木。SA, 气孔面积; SD, 气孔密度; SL, 气孔长度; SOL, 气孔开度; SOR, 气孔开张比; SW, 气孔宽度。不同小写字母代表差异显著(p < 0.05)。 Fig. 2Effects of growth forms and leaf habitus on stomatal traits of main greening plant species in Lanzhou (mean ± SE). Dec, deciduous; Eve, evergreen; Her, herb; Sem, semi-tree; Shr, shrub; Tre, tree. SA, stomatal area; SD, stomatal density; SL, stomatal length; SOL, stomatal opening level; SOR, stomatal opening ratio; SW, stomatal width. Different lowercase letters indicate significant differences among growth forms and leaf habitus (p < 0.05). 2.3 气孔的权衡特征 气孔长度与宽度的标准化主轴回归斜率为1.245 (95%
CI = 1.111, 1.395;
CI 为置信区间,
图3A )、
R 2 = 0.879 (
p < 0.01), 斜率与1.0差异显著 (
p < 0.01), 表明气孔长度与宽度之间为异速生长关系, 且气孔宽度的增加速率大于长度; 气孔面积与气孔开度间的回归斜率为0.572 (95%
CI = 0.429, 0.723;
图3B ),
R 2 = 0.210 (
p < 0.05), 斜率与1.0差异显著, 表明气孔面积与开度之间为异速关系, 且气孔的张开速率小于面积的扩展速率; 气孔密度与气孔面积的回归斜率为-1.144 (95%
CI = -1.480, -0.884;
图3C )、
R 2 = 0.371 (
p < 0.01), 表明气孔面积随着气孔密度增大而减小; 气孔密度与气孔开度的回归斜率为-0.654 (95%
CI = -0.881, -0.486;
图3D )、
R 2 = 0.154 (
p < 0.05), 斜率与1.0差异显著, 说明气孔开度随着气孔密度增大而减小。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3兰州市主要绿化植物气孔性状间的异速生长关系。SA, 气孔面积; SD, 气孔密度; SL, 气孔长度; SOL, 气孔开度; SW, 气孔宽度。 Fig. 3Allometric relations of stomatal traits of main greening plant species in Lanzhou. SA, stomatal area; SD, stomatal density; SL, stomatal length; SOL, stomatal opening level; SW, stomatal width. 2.4 气孔性状的系统发育 PIC分析表明, 去除系统发育信息后, 气孔长度与宽度之间的异速关系发生变化, 表现为气孔宽度的增加速率小于长度增加速率, 斜率为0.892, 显著小于1.0 (95%
CI = 0.814-0.970,
图3E ), 其他性状间则无显著的相关关系, 表明气孔性状的相互关系与物种的进化密切相关。系统发育信号检测结果表明, 气孔开度和气孔开张比均具有显著的系统发育信号(
p < 0.05)(
表1 ), 说明这2组性状与物种进化史有关, 但各性状的系统发育信号仍然较弱(
K < 1), 表明各性状的系统发育保守性不强。
Table 1 表1 表1 兰州市主要绿化植物气孔性状的系统发育信号
Table 1
Phylogenetic signals of stomatal traits across main greening plant species in Lanzhou 性状 Trait K p SL 0.298 0.110 SW 0.291 0.140 SOL 0.334 0.023 SA 0.269 0.379 SOR 0.688 0.006 SD 0.281 0.185
SA, 气孔面积; SD, 气孔密度; SL, 气孔长度; SOL, 气孔开度; SOR, 气孔开张比; SW, 气孔宽度; K, Blomberg’ sK 值。 SA, stomatal area; SD, stomatal density; SL, stomatal length; SOL, stomatal opening level; SOR, stomatal opening ratio; SW, stomatal width; K, Blomberg’ sK value. 新窗口打开 |
下载CSV 2.5 基于气孔性状的物种功能群划分 主成分分析表明前2个主成分可以解释气孔性状变异总方差的79.17% (
图4 , 附录II), 反映了气孔性状的大部分信息, PC1轴与气孔长度(
r = 0.51,
表2 )、宽度(
r = 0.50)、面积(
r = 0.50)与开度(
r = 0.34)等性状的相关性较强, PC2轴与气孔开张比(
r = 0.79)和气孔密度(
r = 0.59)的相关性较强。根据气孔参数特征可以将40种植物划分为3类功能群(附录III): 第1类为较大的气孔面积与较低的气孔密度; 第2类为较小的气孔面积与较大的气孔密度, 以及低气孔开度和气孔开张比; 第3类为气孔大小、开度、开张比和密度都相对均衡。
图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4兰州市主要绿化植物基于气孔性状特征的物种功能群划分。SA, 气孔面积; SD, 气孔密度; SL, 气孔长度; SOL, 气孔开度; SOR, 气孔开张比; SW, 气孔宽度。物种缩写同图1。 Fig. 4Classification of plant functional groups based on stomatal traits of main greening plant species in Lanzhou. SA, stomatal area; SD, stomatal density; SL, stomatal length; SOL, stomatal opening level; SOR, stomatal opening ratio; SW, stomatal width. See Fig. 1 for species abbreviation. Table 2 表2 表2 兰州市主要绿化植物气孔性状与主成分的相关性
Table 2
Correlations between stomatal traits of main greening plant species in Lanzhou and principal components 性状 主成分1 主成分2 SL 0.512 63 -0.044 93 SW 0.501 39 -0.010 85 SOL 0.338 33 0.141 74 SA 0.497 11 0.024 11 SOR 0.184 12 0.792 84 SD -0.300 55 0.590 43
SA, 气孔面积; SD, 气孔密度; SL, 气孔长度; SOL, 气孔开度; SOR, 气孔开张比; SW, 气孔宽度。 SA, stomatal area; SD, stomatal density; SL, stomatal length; SOL, stomatal opening level; SOR, stomatal opening ratio; SW, stomatal width. 新窗口打开 |
下载CSV 3 讨论 植物可以通过协调气孔特征以适应干旱生境并谋求最大生长速率。气孔性状的分异规律展现了植物在适应环境过程中的最佳策略, 这种分异是在环境驱动下兼容了个体特征、进化历程、资源权衡等多种因素在内的最优表征。
生长型和叶习性是引起植物性状变异的重要因素(Sack & Scoffoni,
2013 ; Mencuccini
et al. ,
2015 )。本研究中气孔性状在4种生长型和2种叶习性中均表现出显著差异。生长型主要造成局域环境内光照、温度和水分的分配不均, 叶习性则决定了植物的碳收益方式和水力结构。林冠上层高大乔木叶的生境为光照相对较强、温度变幅大而水分不足, 灌木或草本则相反。冠层植物叶片往往接受更多辐射, 需要较多的水气蒸腾以维持叶表温度(吴冰洁等,
2015 ), 常分化出小气孔开度、密集的气孔来应对强光照与高蒸腾强度; 林下植物则发育出大而疏散的气孔与较大的气孔开度, 这有助于叶片排出多余的水分(熊慧等,
2014 ; 王海珍等,
2015 )。相比于阔叶植物, 常绿植物叶片小而厚, 其表皮功能与微观结构也相对复杂(朱燕华等,
2011 ; 熊慧等,
2014 )。除气孔开张比外, 本研究中常绿植物的气孔性状普遍大于落叶植物, 这既有别于湿生物种大而疏散的气孔, 也与旱生植物小而密集的气孔有所区别。以往研究表明, 常绿植物的抗旱与抗栓塞能力强于落叶植物, 而且在叶片建成时更倾向于“缓慢投资-收益”策略(潘莹萍和陈亚鹏,
2014 ; 金鹰和王传宽,
2015 )。由于常绿植物叶片建成初期的碳投资高于落叶植物, 需要在叶片发育后期节约并获取更多的碳来偿还成本。因此, 常绿植物趋向于发育大而紧密的气孔以加速碳同化和积累更多的干物质。
功能各异的群组是稳定占据生态位并差异化利用资源的具象。根据气孔性状对40种植物进行功能群划分, 发现不同群组之间气孔性状差异显著(
图4 )。大气孔植物如山楂、小叶黄杨和费菜分别代表了乡土树种、常绿物种与景天酸代谢植物, 这些植物特殊的适应功能决定了即便在干旱区, 它们依旧能够发育出较大的气孔, 这可能是物种特性决定的(罗丹丹等,
2019 )。植物的小气孔与高气孔密度以及低气孔开度和开张比更具备典型旱生植物的形态, 这可能是对黄土高原干旱生境长期适应的结果(任昱等,
2014 ; 段贝贝等,
2016 ; 胡选萍等,
2016 )。气孔功能群的差异能够高效利用有限空间中的光照与水分等资源, 功能群的组合也维持了垂直梯度上的层级结构, 这有助于维护生境小气候的稳态, 但植物气孔类群分化的驱动因素需进一步研究揭示。
系统发育决定着植物的亲缘关系并印刻着植物进化的信息, 在研究植物性状特征的过程中是否考虑系统发育将显著地影响研究结果(Ackerly,
2009 ; Zhang
et al. ,
2014 )。本研究中气孔性状间存在显著的相关关系, 在去除系统发育信息后, 这种关系大都发生了转变, 仅气孔长度与宽度依旧维持在稳健的正相关关系, 表明气孔长度与宽度间可能存在关联进化(张亚等,
2014 ; 高林浩等,
2020 )。即不论是否考虑系统发育, 气孔长度与宽度间的正相关关系都是存在的, 暗示这种协同关系可能在古老的植物类群中就已经出现。有研究认为, 植物气孔几何形状和间距总是沿着空间上最优的异速生长关系进化(Sack & Scoffoni,
2013 ; Fiorin
et al. ,
2016 ; Sperry
et al. ,
2017 ), 由于气孔发育属于同步型, 在气孔发育过程中长度与宽度总保持在相对稳定的比率(Franks & Farquhar,
2001 ), 这也从一定程度上解释了去除系统发育后, 气孔长度与宽度依旧保持稳健相关关系的原因。此外, 系统发育信号检测结果表明, 气孔性状的发育信号较弱(
K < 1), 表明气孔性状受系统发育影响较小, 但气孔开度和气孔开张比的发育信号显著, 暗示亲缘关系越近的物种气孔开度与开张比越相似(高林浩等,
2020 )。虽然系统发育会影响气孔性状, 但在长时间适应环境的过程中, 系统发育的影响似乎正在被逐渐削弱。
异速生长分析是研究资源优化配置的重要方法。本研究中, 气孔性状间存在显著的异速生长关系, 表明即便在微观尺度, 异速生长理论依旧适用。气孔面积主要由气孔长度与宽度决定, 而气孔密度也会制约气孔面积的扩增。气孔长度与宽度的异速生长关系(
图3A )表明, 宽度生长速率显著大于长度生长速率; 换言之, 气孔发育过程中并非一直倾向于“长”生长, 当宽度生长速率大于长度生长速率时, 气孔逐渐向“圆”发展。圆形气孔具有较小的周长和等周长下最大的气孔面积, 这有助于减小叶表气孔器的物质与能量投资并使其具有相对无损的功能。还有可能气孔发育是随着叶片发育进行的, 在叶片逐渐增大、成熟的过程中, 光合能力的增强促使气孔向着有益于最大光合效益的过程发展。
气孔面积与开度的关系表明, 随着气孔面积逐渐增大, 气孔开度也逐渐增大, 但气孔开度的扩大速率不及面积的增加速率。有研究认为, 气孔越大对气孔孔径调控的能力越弱, 在遭遇干旱胁迫时所丢失的水分就越多(Drake
et al. ,
2013 ; Sperry
et al. ,
2017 ), 虽然较大的气孔开度可以交换更多的碳, 但这种交换存在风险。气孔面积增大时, 保持CO
2 摄入量不变的情况下, 减小气孔开度依旧可以达到等效水平。但是, 大气孔对开度的调节缺乏灵敏性并对应着较低的气体交换速率, 随着气孔开度逐渐增大, 其进一步开张的阻力也越大、耗费能量越多, 它们的慢动力学特性也使大气孔植物在遭遇干旱等环境胁迫时更容易发生功能紊乱(Hetherington & Woodward,
2003 ; 范嘉智等,
2016 ; 胡选萍等,
2016 )。
气孔数量的发育往往先于面积的发育(Xu & Zhou,
2008 )。当气孔密度逐渐增加, 因其空间有限, 气孔面积发育受阻, 气孔开度也随之减小, 较小的气孔开度与较大的气孔密度弥补了碳获取通道的不足。干旱区植物生长的主要限制因子是水分, 较小的气孔面积便于控制气孔的开合, 而较小的气孔开度则有助于减少水分蒸腾, 缓解水分压力(Hetherington & Woodward,
2003 ; Drake
et al. ,
2013 ; Sperry
et al. ,
2017 )。虽然气孔密度增加有助于获取CO
2 物质量和外界信息流, 但气孔密度并非越大越好, 在遵循“单细胞间隔法则”的基础上(宋璐璐等,
2011 ; 任昱等,
2014 ; 王学芳等,
2014 ; 范嘉智等,
2016 ), 高密度的气孔反而造成气孔拥挤与调控失灵, 为灵活调控气孔开合, 减小气孔面积并降低开度更有助于精准调控。
本研究中, 植物生长型、叶习性与系统发育都会对气孔性状产生影响, 为了提高生存适合度, 气孔发展出多种策略适应生境。气孔性状间的异速生长关系反映了物种对异质生境适应及自我调控的能力, 这种权衡与协变关系在多组性状间均有体现。气孔性状功能群是植物为充分利用局域资源发展出的有效模式, 但其驱动因素和机理有待深入研究。
致谢 感谢甘肃农业大学科技创新基金(GSAU- RCZX201708和GSAU-STS-1734)资助。
附录I 兰州市主要绿化植物名录 Supplement I List of main greening plant species in Lanzhou https://www.plant-ecology.com/fileup/1005-264X/PDF/cjpe.2020.0257-S1.pdf 附录II 兰州主要绿化植物气孔功能群特征 Supplement II Stomatal traits of greening plant functional groups in Lanzhou https://www.plant-ecology.com/fileup/1005-264X/PDF/cjpe.2020.0257-S2.pdf 附录III 兰州市主要绿化植物气孔性状的主成分特征值与累计百分比 Supplement III Importance of principal component and cumulative percentage of stomatal traits of main greening species in Lanzhou https://www.plant-ecology.com/fileup/1005-264X/PDF/cjpe.2020.0257-S3.pdf [1] Ackerly D 2009 ). Conservatism and diversification of plant functional traits: evolutionary rates versus phylogenetic signalProceedings of the National Academy of Sciences of the United States of America 19699 -19706 . [本文引用: 1] [2] Brodribb TJ Jordan GJ 2011 ). Water supply and demand remain balanced during leaf acclimation of Nothofagus cunninghamii trees New Phytologist 437 -448 . DOI:
10.1111/nph.2011.192.issue-2 URL [本文引用: 2] [3] Brodribb TJ Jordan GJ Carpenter RJ 2013 ). Unified changes in cell size permit coordinated leaf evolutionNew Phytologist 559 -570 . DOI:
10.1111/nph.2013.199.issue-2 URL [本文引用: 1] [4] Chen GP Zhao WZ He SX Fu X 2016 ). Biomass allocation and allometric relationship in aboveground components of Salix psammophila branches Journal of Desert Research 357 -363 . [本文引用: 1] [ 陈国鹏 , 赵文智 , 何世雄 , 付晓 (2016 ). 沙柳( Salix psammophila )丛生枝生物量最优分配与异速生长 中国沙漠 , 36,357 -363 .] [本文引用: 1] [5] Ding LZ Chen YJ Zhang JL 2014 ). Leaf traits and their associations among liana species in tropical rainforestPlant Science Journal 362 -370 . [本文引用: 1] [ 丁凌子 , 陈亚军 , 张教林 (2014 ). 热带雨林木质藤本植物叶片性状及其关联植物科学学报 , 32,362 -370 .] [本文引用: 1] [6] Drake PL Froend RH Franks PJ 2013 ). Smaller, faster stomata: scaling of stomatal size, rate of response, and stomatal conductanceJournal of Experimental Botany 495 -505 . DOI:
10.1093/jxb/ers347 PMID:
23264516 [本文引用: 4] Maximum and minimum stomatal conductance, as well as stomatal size and rate of response, are known to vary widely across plant species, but the functional relationship between these static and dynamic stomatal properties is unknown. The objective of this study was to test three hypotheses: (i) operating stomatal conductance under standard conditions (g (op)) correlates with minimum stomatal conductance prior to morning light [g (min(dawn))]; (ii) stomatal size (S) is negatively correlated with g (op) and the maximum rate of stomatal opening in response to light, (dg/dt)(max); and (iii) g (op) correlates negatively with instantaneous water-use efficiency (WUE) despite positive correlations with maximum rate of carboxylation (Vc (max)) and light-saturated rate of electron transport (J (max)). Using five closely related species of the genus Banksia, the above variables were measured, and it was found that all three hypotheses were supported by the results. Overall, this indicates that leaves built for higher rates of gas exchange have smaller stomata and faster dynamic characteristics. With the aid of a stomatal control model, it is demonstrated that higher g (op) can potentially expose plants to larger tissue water potential gradients, and that faster stomatal response times can help offset this risk.
[7] Duan BB Zhao CZ Xu T Zheng HL Feng W Han L 2016 ). Correlation analysis between vein density and stomatal traits of Robinia pseudoacacia in different aspects of Beishan Mountain in Lanzhou Chinese Journal of Plant Ecology 1289 -1297 . DOI:
10.17521/cjpe.2016.0215 URL [本文引用: 4] [ 段贝贝 , 赵成章 , 徐婷 , 郑慧玲 , 冯威 , 韩玲 (2016 ). 兰州北山不同坡向刺槐叶脉密度与气孔性状的关联性分析植物生态学报 , 40,1289 -1297 .] DOI:
10.17521/cjpe.2016.0215 [本文引用: 4] 植物叶脉和气孔性状的关系反映了叶片的水力特性, 对认识它们与植物水分利用有关的生理功能间的关系及其调控作用具有重要意义。该文利用GIS (geographic information system)与实验生态学相结合的方法, 采用标准化主轴估计方法, 研究了兰州市北山不同坡向人工林刺槐(Robinia pseudoacacia)叶脉密度与气孔密度、气孔大小的关系。结果表明: 随着坡向由南坡向东坡、西坡和北坡转变, 植被群落的郁闭度、高度和土壤含水量呈逐渐增加的趋势, 刺槐的净光合速率(P<sub>n</sub>)、蒸腾速率(T<sub>r</sub>)、光合有效辐射(PAR)、叶脉密度和气孔密度呈逐渐减小的趋势, 气孔与叶面积呈逐渐增大的趋势; 各个坡向的刺槐叶脉密度与气孔密度呈显著正相关关系, 与气孔大小呈显著负相关关系, 且在南坡达到极显著相关关系。生长在南坡的刺槐具有高的叶脉密度和密而小的气孔, 生长在北坡的刺槐具有低的叶脉密度和疏而大的气孔。不同坡向刺槐叶脉密度与气孔特征间的资源分配模式, 反映了植物在异质性生境中根据其功能需求在自身性状之间进行投资权衡机制的优化。
[8] Fan JZ Wang D Hu YL Jing PP Wang PP Chen JQ 2016 ). Optimal stomatal behavior theory for simulating stomatal conductanceChinese Journal of Plant Ecology 631 -642 . DOI:
10.17521/cjpe.2015.0480 URL [本文引用: 3] [ 范嘉智 , 王丹 , 胡亚林 , 景盼盼 , 王朋朋 , 陈吉泉 (2016 ). 最优气孔行为理论和气孔导度模拟植物生态学报 , 40,631 -642 .] DOI:
10.17521/cjpe.2015.0480 [本文引用: 3] 气孔调节功能是陆地生态系统碳-水耦合过程中最重要的环节。与即时的气孔导度测量相比, 气孔导度斜率能有效地反映气孔导度对CO<sub>2</sub>浓度、饱和水汽压亏缺和光合作用的敏感性, 包含了环境因子对光合作用和临界水分利用效率等的综合影响, 为研究全球变化下陆地生态系统碳-水耦合关系提供了一个简明且综合的框架。气孔导度模型从经验模型、半经验模型发展到机理模型, 经过很多****的改进, 但是模型参数的生物学意义和变化规律还不明确。鉴于气孔导度斜率方面研究的重要性和研究的不足, 为了加强对气孔导度调节规律的认识, 并减少气孔导度模拟的不确定性, 该文主要综述了长期以来国内外关于最优气孔行为理论和气孔导度模拟方面的研究成果, 其中包括广泛使用的气孔导度模型及参数意义, 讨论影响气孔导度斜率的主要因素以及气孔导度机理模型的应用, 并对最优气孔行为理论和气孔导度模拟方面的研究做了简单展望。
[9] Fiorin L Brodribb TJ Anfodillo T 2016 ). Transport efficiency through uniformity: organization of veins and stomata in angiosperm leavesNew Phytologist 216 -227 . DOI:
10.1111/nph.2016.209.issue-1 URL [本文引用: 3] [10] Franks PJ Farquhar GD 2001 ). The effect of exogenous abscisic acid on stomatal development, stomatal mechanics, and leaf gas exchange in Tradescantia virginianaPlant Physiology 935 -942 . DOI:
10.1104/pp.125.2.935 URL [本文引用: 1] [11] Gao GL Feng Q Zhang XY Si JH Yu TF 2018 ). An overview of stomatal and non-stomatal limitations to photosynthesis of plantsArid Zone Research 929 -937 . [本文引用: 1] [ 高冠龙 , 冯起 , 张小由 , 司建华 , 鱼腾飞 (2018 ). 植物叶片光合作用的气孔与非气孔限制研究综述干旱区研究 , 35,929 -937 .] [本文引用: 1] [12] Gao LH Sun H Bai XQ Dai S Fan YW Liu C Wang XP Yin WL 2020 ). Effects of climate and phylogeny on the relationship between specific leaf area and leaf element concentration of trees and shrubs in Changbai Mountain of northeastern ChinaJournal of Beijing Forestry University 2 ),19 -30 . [本文引用: 2] [ 高林浩 , 孙晗 , 白雪卡 , 代爽 , 樊艳文 , 刘超 , 王襄平 , 尹伟伦 (2020 ). 气候、系统发育对长白山乔灌木比叶面积与叶元素含量关系的影响北京林业大学学报 , 42(2 ),19 -30 .] [本文引用: 2] [13] Hetherington AM Woodward FI 2003 ). The role of stomata in sensing and driving environmental changeNature 901 -908 . PMID:
12931178 [本文引用: 2] Stomata, the small pores on the surfaces of leaves and stalks, regulate the flow of gases in and out of leaves and thus plants as a whole. They adapt to local and global changes on all timescales from minutes to millennia. Recent data from diverse fields are establishing their central importance to plant physiology, evolution and global ecology. Stomatal morphology, distribution and behaviour respond to a spectrum of signals, from intracellular signalling to global climatic change. Such concerted adaptation results from a web of control systems, reminiscent of a 'scale-free' network, whose untangling requires integrated approaches beyond those currently used.
[14] Hu XP Ji CJ An LH 2016 ). Leaf epidermis characteristics of the main grassland monocotyledonous plant species on the Tibetan PlateauActa Ecologica Sinica 6465 -6474 . [本文引用: 2] [ 胡选萍 , 吉成均 , 安丽华 (2016 ). 青藏高原草地主要单子叶植物的叶表面特征生态学报 , 36,6465 -6474 .] [本文引用: 2] [15] Jia JB Liu WN Wen SZ Wang ZC 2019 ). Effects of CO2 concentration and soil moisture on leaf stomatal traits and gas exchange parameters of Phoebe bourneiScience of Soil and Water Conservation 1 -7 . [本文引用: 1] [ 贾剑波 , 刘文娜 , 文仕知 , 王忠诚 (2019 ). 水碳控制条件对闽楠叶片气孔特征和气体交换参数的影响中国水土保持科学 , 17,1 -7 .] [本文引用: 1] [16] Jiang XX Zou AL Wang YY Zhou XL Ji CJ 2018 ). Leaf stomatal traits of woody plants and their response to nitrogen addition in typical forests in Eastern ChinaActa Scientiarum Naturalium Universitatis Pekinensis 839 -847 . [本文引用: 1] [ 姜星星 , 邹安龙 , 王媛媛 , 周序力 , 吉成均 (2018 ). 我国东部典型森林木本植物的气孔特征及其对氮添加的响应北京大学学报(自然科学版) , 54,839 -847 .] [本文引用: 1] [17] Jin Y Wang CK 2015 ). Trade-offs between plant leaf hydraulic and economic traitsChinese Journal of Plant Ecology 1021 -1032 . DOI:
10.17521/cjpe.2015.0099 URL [本文引用: 3] [ 金鹰 , 王传宽 (2015 ). 植物叶片水力与经济性状权衡关系的研究进展植物生态学报 , 39,1021 -1032 .] DOI:
10.17521/cjpe.2015.0099 [本文引用: 3] 叶片既是植物光合产物形成的主要场所, 又是整株植物的水力瓶颈、应对灾难性水力失调的安全阀门, 是植物碳水耦合权衡的重要器官。叶经济型谱反映了叶片经济性状“投资-收益”的权衡, 为验证植物进化过程中形成的物种对策提供了适用的理论框架。叶片水力性状变化会影响叶片经济性状及植物存活和生长。因此, 探索植物叶片水力与经济性状的权衡关系, 对建立植物碳-水耦合模型、揭示植物水-碳投资机理、扩展植物性状型谱等均有重要意义。该文首先综述了叶片水力性状、经济性状及两者之间的权衡关系, 分析了叶片导水率与水力脆弱性、失膨点水势、水容、安全阈值等水力性状以及与叶片的形态、结构和气体交换功能性状之间的关系。然后, 从叶片形态、解剖和叶脉网络结构以及气孔功能方面探讨了叶片水力性状与经济性状的调节机制。最后, 提出今后应加强三方面的研究: (1)探索建立植物根-茎-叶水力输导系统的碳-氮-水资源的整株经济型谱, 以揭示植物功能结构耦合、高效固碳用水的生理生态学机制; (2)探索叶片水力安全、水力效率和固碳效率之间的普适性权衡关系, 以深入理解抗旱植物叶片构建的生物物理结构与生理代谢的关系; (3)探索个体水平碳水代谢关系、水分运输与生长速率的耦合, 为代谢推演理论和植物群落尺度预测提供基础。
[18] Loranger J Shipley B 2010 ). Interspecific covariation between stomatal density and other functional leaf traits in a local floraBotany 30 -38 . DOI:
10.1139/B09-103 URL [本文引用: 1] [19] Luo DD Wang CK Jin Y 2019 ). Stomatal regulation of plants in response to drought stressChinese Journal of Applied Ecology 4333 -4343 . [本文引用: 1] [ 罗丹丹 , 王传宽 , 金鹰 (2019 ). 植物应对干旱胁迫的气孔调节应用生态学报 , 30,4333 -4343 .] [本文引用: 1] [20] Mencuccini M Minunno F Salmon Y Martínez-Vilalta J H?ltt? T 2015 ). Coordination of physiological traits involved in drought-induced mortality of woody plantsNew Phytologist 396 -409 . DOI:
10.1111/nph.2015.208.issue-2 URL [本文引用: 2] [21] Pan YP Chen YP 2014 ). Recent advances in leaf hydraulic traitsChinese Journal of Ecology 2834 -2841 . [本文引用: 2] [ 潘莹萍 , 陈亚鹏 (2014 ). 叶片水力性状研究进展生态学杂志 , 33,2834 -2841 .] [本文引用: 2] [22] Ren Y Lu Q Wu B Li YH Xin ZM Yao B 2014 ). Response of leaf of Nitraria tangutorum Bobr stomata characteristics to artificial simulation of rainfall Acta Ecologica Sinica 6101 -6106 . [本文引用: 3] [ 任昱 , 卢琦 , 吴波 , 李永华 , 辛智鸣 , 姚斌 (2014 ). 白刺叶片气孔特征对人工模拟降雨的响应生态学报 , 34,6101 -6106 .] [本文引用: 3] [23] Sack L Scoffoni C 2013 ). Leaf venation: structure, function, development, evolution, ecology and applications in the past, present and futureNew Phytologist 983 -1000 . DOI:
10.1111/nph.2013.198.issue-4 URL [本文引用: 3] [24] Song HQ Ni MY Zhu SD 2020 ). Hydraulic and photosynthetic characteristics differ between co-generic tree and Liana species: a case study of Millettia and Gnetum in tropical forestChinese Journal of Plant Ecology 192 -204 . DOI:
10.17521/cjpe.2019.0304 URL [本文引用: 1] [ 宋慧清 , 倪鸣源 , 朱师丹 (2020 ). 乔木与木质藤本的水力与光合性状的差异: 以热带森林崖豆藤属和买麻藤属为例植物生态学报 , 44,192 -204 .] DOI:
10.17521/cjpe.2019.0304 [本文引用: 1] 木质藤本是热带森林的重要组成部分, 显著影响森林的结构和功能。已有研究发现木质藤本与乔木的水力结构存在显著差异: 木质藤本的缠绕或攀缘茎细小, 但其木质部具有粗大的长导管, 输水效率高, 抗栓塞能力低。为降低基因型差异对比较结果的影响, 该研究选取热带崖豆藤属(Millettia)和买麻藤属(Gnetum)的乔木和木质藤本, 比较同属内不同生长型植物的水力和光合性状的差异, 分析水分传导效率与抗栓塞能力之间以及水力与光合性状之间的相关关系。结果发现: (1)崖豆藤属植物水力性状的种间差异大, 与生活型和需光性有关。耐阴的木质藤本反而具有较低的水分传导效率和较高的抗栓塞能力。(2)买麻藤属植物是裸子植物较为进化的类群(具有导管和阔叶), 其乔木的水分传导效率很低, 但是其木质藤本的水分传导效率高于其他阳生性的被子植物。(3)不论乔木还是木质藤本, 水分传导的有效性与安全性在枝条和叶片水平上均没有显著的权衡关系。(4)与同属乔木相比, 木质藤本的叶片较枝条的抗栓塞能力更强, 在旱季具有更高的最大净光合速率和气孔导度, 支持了木质藤本的“旱季生长优势假说”。该研究揭示了热带木质藤本水力性状的多样性和重要性, 为阐明环境变化对这一重要植物类群的影响, 需要对它们的水力特征进行更广泛的研究。
[25] Song LL Fan JW Wu SH 2011 ). Research advances on changes of leaf traits along an altitude gradientProgress in Geography 1431 -1439 . [本文引用: 2] [ 宋璐璐 , 樊江文 , 吴绍洪 (2011 ). 植物叶片性状沿海拔梯度变化研究进展地理科学进展 , 30,1431 -1439 .] DOI:
10.11820/dlkxjz.2011.11.014 [本文引用: 2] 由于在海拔梯度上各种环境因子表现出连续的梯度性变化,以海拔梯度为平台研究植物叶片性状的适应特征有助于揭示植物对气候变化的响应,进而可以为研究全球变化与陆地生态系统的关系提供研究基础。本文重点分析了植物的叶寿命、比叶面积、叶氮含量、叶绿素含量等叶片结构性状和气孔导度、叶片羧化效率、水分利用效率和叶片δ<sup>13</sup>C等叶片功能性状随海拔梯度的变化规律和特点,探讨了植物叶片性状的研究不足及未来发展方向,以便为国内相关研究的开展提供参考。
[26] Sperry JS Venturas MD Anderegg WRL Mencuccini M MacKay DS Wang YJ Love DM 2017 ). Predicting stomatal responses to the environment from the optimization of photosynthetic gain and hydraulic costPlant, Cell & Environment 816 -830 . [本文引用: 3] [27] Wang BX Zeng YH Wang DY Zhao R Xu X 2010 ). Responses of leaf stomata to environmental stresses in distribution and physiological characteristicsAgricultural Research in the Arid Areas 122 -126 . [本文引用: 1] [ 王碧霞 , 曾永海 , 王大勇 , 赵蓉 , 胥晓 (2010 ). 叶片气孔分布及生理特征对环境胁迫的响应干旱地区农业研究 , 28,122 -126 .] [本文引用: 1] [28] Wang HZ Han L Xu YL Niu JL 2015 ). Model analysis of the stomatal conductance response to light in Populus pruinosa at different temperatures in the Taklimakan desert Ecology and Environmental Sciences 741 -748 . [本文引用: 1] [ 王海珍 , 韩路 , 徐雅丽 , 牛建龙 (2015 ). 不同温度下灰胡杨叶片气孔导度对光强响应的模型分析生态环境学报 , 24,741 -748 .] [本文引用: 1] [29] Wang RL Yu GR He NP Wang QF Zhao N Xu ZW 2016 ). Altitudinal variation in the covariation of stomatal traits with leaf functional traits in Changbai MountainActa Ecologica Sinica 2175 -2184 . [本文引用: 2] [ 王瑞丽 , 于贵瑞 , 何念鹏 , 王秋凤 , 赵宁 , 徐志伟 (2016 ). 气孔特征与叶片功能性状之间关联性沿海拔梯度的变化规律——以长白山为例生态学报 , 36,2175 -2184 .] [本文引用: 2] [30] Wen JW Chen HX Teng YP Zhang SX Wang RL 2018 ). Variation of leaf stomatal traits in Quercus species along the altitudinal gradient in Taibai Mountain, China Acta Ecologica Sinica 6712 -6721 . [本文引用: 1] [ 温婧雯 , 陈昊轩 , 滕一平 , 张硕新 , 王瑞丽 (2018 ). 太白山栎属树种气孔特征沿海拔梯度的变化规律生态学报 , 38,6712 -6721 .] [本文引用: 1] [31] Wu BJ Liu YJ Jiang CD Shi L 2015 ). Effects of stomatal development on leaf temperature during leaf expansionPlant Physiology Journal 119 -126 . [本文引用: 1] [ 吴冰洁 , 刘玉军 , 姜闯道 , 石雷 (2015 ). 叶片生长进程中气孔发育对叶温调节的影响植物生理学报 , 51,119 -126 .] [本文引用: 1] [32] Wang XF Li RY Li XZ Ma FJ Sun BN Wang JY Wang YK 2014 ). Variations in leaf characteristics of three species of angiosperms with changing of altitude in Qilian Mountains and their inland high-altitude patternScience China: Earth Sciences (Chinese version )706 -714 . [本文引用: 2] [ 王学芳 , 李瑞云 , 李孝泽 , 马福军 , 孙柏年 , 吴靖宇 , 汪有奎 (2014 ). 祁连山3种被子植物叶特征随海拔变化及其内陆高海拔模式中国科学: 地球科学(中文版) , 44,706 -714 .] [本文引用: 2] [33] Xia ZH Chen YN Zhu CG Zhou YY Chen XL 2018 ). Stomatal change in leaves of population euphratica under drought stressArid Zone Research 1111 -1117 . [本文引用: 1] [ 夏振华 , 陈亚宁 , 朱成刚 , 周莹莹 , 陈晓林 (2018 ). 干旱胁迫环境下的胡杨叶片气孔变化干旱区研究 , 35,1111 -1117 .] [本文引用: 1] [34] Xiong H Ma CE Li L Zeng H Guo DL 2014 ). Stomatal characteristics of ferns and angiosperms and their responses to changing light intensity at different habitatsChinese Journal of Plant Ecology 868 -877 . DOI:
10.3724/SP.J.1258.2014.00081 URL [本文引用: 2] [ 熊慧 , 马承恩 , 李乐 , 曾辉 , 郭大立 (2014 ). 不同生境条件下蕨类和被子植物的气孔形态特征及其对光强变化的响应植物生态学报 , 38,868 -877 .] DOI:
10.3724/SP.J.1258.2014.00081 [本文引用: 2] 气孔是植物与大气环境进行气体交换的重要通道, 在调控植物碳水平衡方面发挥着重要作用。为探讨生境和植物类型对气孔形态特征的影响以及气孔对光强变化的响应格局在不同植物间和不同生境条件下的变异, 选取开阔生境和林下生境的5种蕨类植物和4种被子植物, 测定了它们的气孔形态特征和气孔导度对光强变化的响应。此外, 还收集了8篇文献中开阔和林下生境的45种蕨类植物和70种被子植物的气孔密度和气孔长度数据, 以增大样本量从而更好地探讨不同生境条件下蕨类和被子植物气孔密度及长度的变异格局, 并通过分析生境和植物类型对气孔形态特征的影响来推测生境和植物类型对气孔响应行为的可能影响。实验结果表明, 与林下植物相比, 开阔环境下的植物气孔密度更大, 气孔长度更小, 气孔对光强降低的响应更敏感; 但植物类型对气孔形态特征的影响以及对气孔响应光强的敏感程度的影响均不显著。对文献数据的分析表明, 生境和植物类型对气孔形态特征均有显著影响。考虑到气孔响应快慢与气孔形态特征密切相关, 与蕨类植物相比, 被子植物小而密的气孔可能为其更快地响应环境变化提供了基础。研究表明生境和植物类型对气孔响应行为均有显著影响。
[35] Xu ZZ Zhou GS 2008 ). Responses of leaf stomatal density to water status and its relationship with photosynthesis in a grassJournal of Experimental Botany 3317 -3325 . DOI:
10.1093/jxb/ern185 URL [本文引用: 2] [36] Yang KT Chen GP Li G Yu XY Zhang K Tang D Zhang WX Guo YJ 2020 ). Trade-off among leaf traits of typical greening tree species in LanzhouChinese Journal of Ecology 1518 -1526 . [本文引用: 2] [ 杨克彤 , 陈国鹏 , 李广 , 俞筱押 , 张凯 , 汤东 , 张文祥 , 郭英杰 (2020 ). 兰州市典型绿化树种叶性状间的权衡关系生态学杂志 , 39,1518 -1526 .] [本文引用: 2] [37] Yang ZQ Tan W Liu ZX Chen YQ 2015 ). Effect of soil water stress on stomatal characters of greenhouse tomato leavesChinese Journal of Ecology 1234 -1240 . [本文引用: 1] [ 杨再强 , 谭文 , 刘朝霞 , 陈艳秋 (2015 ). 土壤水分胁迫对设施番茄叶片气孔特性的影响生态学杂志 , 34,1234 -1240 .] [本文引用: 1] [38] Zhang DC Zhu YH Li SZ 2018 ). Variation in stomatal characteristics of eight plant species along a soil moisture gradient in alpine meadow of the Dongda Mountains in southeast TibetActa Prataculturae Sinica 36 -46 . [本文引用: 2] [ 张大才 , 朱玉怀 , 李双智 (2018 ). 东达山高寒草甸8种植物气孔特征沿土壤水分梯度的变化草业学报 , 27,36 -46 .] [本文引用: 2] [39] Zhang SB Sun M Cao KF Hu H Zhang JL 2014 ). Leaf photosynthetic rate of tropical ferns is evolutionarily linked to water transport capacityPLOS ONE e84682 . DOI: 10.1371/journal.pone.0084682 . DOI:
10.1371/journal.pone.0084682 URL [本文引用: 1] [40] Zhang Y Yang SJ Sun M Cao KF 2014 ). Stomatal traits are evolutionarily associated with vein density in basal angiospermsPlant Science Journal 320 -328 . [本文引用: 1] [ 张亚 , 杨石建 , 孙梅 , 曹坤芳 (2014 ). 基部被子植物气孔性状与叶脉密度的关联进化植物科学学报 , 32,320 -328 .] [本文引用: 1] [41] Zhu YH Kang HZ Liu CJ 2011 ). Affecting factors of plant stomatal traits variability and relevant investigation methodsChinese Journal of Applied Ecology 250 -256 . [本文引用: 3] [ 朱燕华 , 康宏樟 , 刘春江 (2011 ). 植物叶片气孔性状变异的影响因素及研究方法应用生态学报 , 22,250 -256 .] [本文引用: 3] Conservatism and diversification of plant functional traits: evolutionary rates versus phylogenetic signal
1
2009
... 系统发育决定着植物的亲缘关系并印刻着植物进化的信息, 在研究植物性状特征的过程中是否考虑系统发育将显著地影响研究结果(Ackerly,
2009 ; Zhang
et al. ,
2014 ).本研究中气孔性状间存在显著的相关关系, 在去除系统发育信息后, 这种关系大都发生了转变, 仅气孔长度与宽度依旧维持在稳健的正相关关系, 表明气孔长度与宽度间可能存在关联进化(张亚等,
2014 ; 高林浩等,
2020 ).即不论是否考虑系统发育, 气孔长度与宽度间的正相关关系都是存在的, 暗示这种协同关系可能在古老的植物类群中就已经出现.有研究认为, 植物气孔几何形状和间距总是沿着空间上最优的异速生长关系进化(Sack & Scoffoni,
2013 ; Fiorin
et al. ,
2016 ; Sperry
et al. ,
2017 ), 由于气孔发育属于同步型, 在气孔发育过程中长度与宽度总保持在相对稳定的比率(Franks & Farquhar,
2001 ), 这也从一定程度上解释了去除系统发育后, 气孔长度与宽度依旧保持稳健相关关系的原因.此外, 系统发育信号检测结果表明, 气孔性状的发育信号较弱(
K < 1), 表明气孔性状受系统发育影响较小, 但气孔开度和气孔开张比的发育信号显著, 暗示亲缘关系越近的物种气孔开度与开张比越相似(高林浩等,
2020 ).虽然系统发育会影响气孔性状, 但在长时间适应环境的过程中, 系统发育的影响似乎正在被逐渐削弱. ...
Water supply and demand remain balanced during leaf acclimation of
Nothofagus cunninghamii trees
2
2011
... 气孔作为植物叶表皮的特殊结构, 是植物与外界环境进行物质交换的门户(宋璐璐等,
2011 ; 朱燕华等,
2011 ), 也是植物体内水汽逸散的主要通道(金鹰和王传宽,
2015 ; 范嘉智等,
2016 ).气孔易受环境条件的修饰, 其形态特征是植物进化过程中对外界环境长期适应的结果(王碧霞等,
2010 ; 朱燕华等,
2011 ; 温婧雯等,
2018 ).为适应(或响应)复杂多变的环境, 气孔一方面通过调节开闭来应对短期或局域环境变化, 另一方面则协调大小和密度来适应异质生境(Brodribb & Jordan,
2011 ; Drake
et al. ,
2013 ; 金鹰和王传宽,
2015 ; 高冠龙等,
2018 ).利用气孔特征反映植物对环境变化的响应有助于明晰植物应对水气耦合过程的策略. ...
... 系统发育和环境异质是影响植物性状发生分异的动力来源.气孔主要分布于植物叶片下表皮并参与调控碳水摄入与排出, 在其发育过程中迫于环境压力与生存维持, 气孔往往在多个维度存在权衡(Brodribb & Jordan,
2011 ; Brodribb
et al. ,
2013 ; 丁凌子等,
2014 ; Fiorin
et al. ,
2016 ).虽然气孔数量的增加有助于提高光合效率, 但过多的气孔会加速叶内水汽逸散(潘莹萍和陈亚鹏,
2014 ; 王学芳等,
2014 ; 宋慧清等,
2020 ), 而且气孔的建成需要消耗资源, 这就决定了单位面积的气孔数量存在分布上限.气孔密度与大小共同决定了最大可能气孔导度并影响最大同化速率, 在考虑气孔建成实际资源消耗与光合作用水气协调的过程中, 两个相互矛盾的刚需必然达到最优化(Sack & Scoffoni,
2013 ).气孔面积与开度分别代表着气孔发育与运动, 且气孔发育总是先于气孔运动, 尽管大气孔对于开度的调控不及小气孔灵敏, 但可以获得相对较多的碳收益.就此而言, 气孔发育和运动过程中资源分配不均甚至存在矛盾(Xu & Zhou,
2008 ; Drake
et al. ,
2013 ).在异质生境中, 气孔各性状并非独立变化, 而是通过性状或特征组合的协变与权衡来共同适应环境.有限资源在植物体内的分配与权衡已通过异速生长理论在多个尺度得到证实(陈国鹏等,
2016 ; 段贝贝等,
2016 ; 杨克彤等,
2020 ), 但以往研究着重于相对宏观的尺度如群落、个体、构件与器官, 而且忽视了系统发育对资源权衡的影响, 对微观尺度如气孔性状的权衡机制关注较少.为此, 本文希望通过研究气孔特征并结合系统发育来探讨微观尺度气孔性状间的生长关系, 以进一步揭示微观尺度的权衡过程, 并丰富植物生活史研究. ...
Unified changes in cell size permit coordinated leaf evolution
1
2013
... 系统发育和环境异质是影响植物性状发生分异的动力来源.气孔主要分布于植物叶片下表皮并参与调控碳水摄入与排出, 在其发育过程中迫于环境压力与生存维持, 气孔往往在多个维度存在权衡(Brodribb & Jordan,
2011 ; Brodribb
et al. ,
2013 ; 丁凌子等,
2014 ; Fiorin
et al. ,
2016 ).虽然气孔数量的增加有助于提高光合效率, 但过多的气孔会加速叶内水汽逸散(潘莹萍和陈亚鹏,
2014 ; 王学芳等,
2014 ; 宋慧清等,
2020 ), 而且气孔的建成需要消耗资源, 这就决定了单位面积的气孔数量存在分布上限.气孔密度与大小共同决定了最大可能气孔导度并影响最大同化速率, 在考虑气孔建成实际资源消耗与光合作用水气协调的过程中, 两个相互矛盾的刚需必然达到最优化(Sack & Scoffoni,
2013 ).气孔面积与开度分别代表着气孔发育与运动, 且气孔发育总是先于气孔运动, 尽管大气孔对于开度的调控不及小气孔灵敏, 但可以获得相对较多的碳收益.就此而言, 气孔发育和运动过程中资源分配不均甚至存在矛盾(Xu & Zhou,
2008 ; Drake
et al. ,
2013 ).在异质生境中, 气孔各性状并非独立变化, 而是通过性状或特征组合的协变与权衡来共同适应环境.有限资源在植物体内的分配与权衡已通过异速生长理论在多个尺度得到证实(陈国鹏等,
2016 ; 段贝贝等,
2016 ; 杨克彤等,
2020 ), 但以往研究着重于相对宏观的尺度如群落、个体、构件与器官, 而且忽视了系统发育对资源权衡的影响, 对微观尺度如气孔性状的权衡机制关注较少.为此, 本文希望通过研究气孔特征并结合系统发育来探讨微观尺度气孔性状间的生长关系, 以进一步揭示微观尺度的权衡过程, 并丰富植物生活史研究. ...
沙柳(
Salix psammophila )丛生枝生物量最优分配与异速生长
1
2016
... 系统发育和环境异质是影响植物性状发生分异的动力来源.气孔主要分布于植物叶片下表皮并参与调控碳水摄入与排出, 在其发育过程中迫于环境压力与生存维持, 气孔往往在多个维度存在权衡(Brodribb & Jordan,
2011 ; Brodribb
et al. ,
2013 ; 丁凌子等,
2014 ; Fiorin
et al. ,
2016 ).虽然气孔数量的增加有助于提高光合效率, 但过多的气孔会加速叶内水汽逸散(潘莹萍和陈亚鹏,
2014 ; 王学芳等,
2014 ; 宋慧清等,
2020 ), 而且气孔的建成需要消耗资源, 这就决定了单位面积的气孔数量存在分布上限.气孔密度与大小共同决定了最大可能气孔导度并影响最大同化速率, 在考虑气孔建成实际资源消耗与光合作用水气协调的过程中, 两个相互矛盾的刚需必然达到最优化(Sack & Scoffoni,
2013 ).气孔面积与开度分别代表着气孔发育与运动, 且气孔发育总是先于气孔运动, 尽管大气孔对于开度的调控不及小气孔灵敏, 但可以获得相对较多的碳收益.就此而言, 气孔发育和运动过程中资源分配不均甚至存在矛盾(Xu & Zhou,
2008 ; Drake
et al. ,
2013 ).在异质生境中, 气孔各性状并非独立变化, 而是通过性状或特征组合的协变与权衡来共同适应环境.有限资源在植物体内的分配与权衡已通过异速生长理论在多个尺度得到证实(陈国鹏等,
2016 ; 段贝贝等,
2016 ; 杨克彤等,
2020 ), 但以往研究着重于相对宏观的尺度如群落、个体、构件与器官, 而且忽视了系统发育对资源权衡的影响, 对微观尺度如气孔性状的权衡机制关注较少.为此, 本文希望通过研究气孔特征并结合系统发育来探讨微观尺度气孔性状间的生长关系, 以进一步揭示微观尺度的权衡过程, 并丰富植物生活史研究. ...
沙柳(
Salix psammophila )丛生枝生物量最优分配与异速生长
1
2016
... 系统发育和环境异质是影响植物性状发生分异的动力来源.气孔主要分布于植物叶片下表皮并参与调控碳水摄入与排出, 在其发育过程中迫于环境压力与生存维持, 气孔往往在多个维度存在权衡(Brodribb & Jordan,
2011 ; Brodribb
et al. ,
2013 ; 丁凌子等,
2014 ; Fiorin
et al. ,
2016 ).虽然气孔数量的增加有助于提高光合效率, 但过多的气孔会加速叶内水汽逸散(潘莹萍和陈亚鹏,
2014 ; 王学芳等,
2014 ; 宋慧清等,
2020 ), 而且气孔的建成需要消耗资源, 这就决定了单位面积的气孔数量存在分布上限.气孔密度与大小共同决定了最大可能气孔导度并影响最大同化速率, 在考虑气孔建成实际资源消耗与光合作用水气协调的过程中, 两个相互矛盾的刚需必然达到最优化(Sack & Scoffoni,
2013 ).气孔面积与开度分别代表着气孔发育与运动, 且气孔发育总是先于气孔运动, 尽管大气孔对于开度的调控不及小气孔灵敏, 但可以获得相对较多的碳收益.就此而言, 气孔发育和运动过程中资源分配不均甚至存在矛盾(Xu & Zhou,
2008 ; Drake
et al. ,
2013 ).在异质生境中, 气孔各性状并非独立变化, 而是通过性状或特征组合的协变与权衡来共同适应环境.有限资源在植物体内的分配与权衡已通过异速生长理论在多个尺度得到证实(陈国鹏等,
2016 ; 段贝贝等,
2016 ; 杨克彤等,
2020 ), 但以往研究着重于相对宏观的尺度如群落、个体、构件与器官, 而且忽视了系统发育对资源权衡的影响, 对微观尺度如气孔性状的权衡机制关注较少.为此, 本文希望通过研究气孔特征并结合系统发育来探讨微观尺度气孔性状间的生长关系, 以进一步揭示微观尺度的权衡过程, 并丰富植物生活史研究. ...
热带雨林木质藤本植物叶片性状及其关联
1
2014
... 系统发育和环境异质是影响植物性状发生分异的动力来源.气孔主要分布于植物叶片下表皮并参与调控碳水摄入与排出, 在其发育过程中迫于环境压力与生存维持, 气孔往往在多个维度存在权衡(Brodribb & Jordan,
2011 ; Brodribb
et al. ,
2013 ; 丁凌子等,
2014 ; Fiorin
et al. ,
2016 ).虽然气孔数量的增加有助于提高光合效率, 但过多的气孔会加速叶内水汽逸散(潘莹萍和陈亚鹏,
2014 ; 王学芳等,
2014 ; 宋慧清等,
2020 ), 而且气孔的建成需要消耗资源, 这就决定了单位面积的气孔数量存在分布上限.气孔密度与大小共同决定了最大可能气孔导度并影响最大同化速率, 在考虑气孔建成实际资源消耗与光合作用水气协调的过程中, 两个相互矛盾的刚需必然达到最优化(Sack & Scoffoni,
2013 ).气孔面积与开度分别代表着气孔发育与运动, 且气孔发育总是先于气孔运动, 尽管大气孔对于开度的调控不及小气孔灵敏, 但可以获得相对较多的碳收益.就此而言, 气孔发育和运动过程中资源分配不均甚至存在矛盾(Xu & Zhou,
2008 ; Drake
et al. ,
2013 ).在异质生境中, 气孔各性状并非独立变化, 而是通过性状或特征组合的协变与权衡来共同适应环境.有限资源在植物体内的分配与权衡已通过异速生长理论在多个尺度得到证实(陈国鹏等,
2016 ; 段贝贝等,
2016 ; 杨克彤等,
2020 ), 但以往研究着重于相对宏观的尺度如群落、个体、构件与器官, 而且忽视了系统发育对资源权衡的影响, 对微观尺度如气孔性状的权衡机制关注较少.为此, 本文希望通过研究气孔特征并结合系统发育来探讨微观尺度气孔性状间的生长关系, 以进一步揭示微观尺度的权衡过程, 并丰富植物生活史研究. ...
热带雨林木质藤本植物叶片性状及其关联
1
2014
... 系统发育和环境异质是影响植物性状发生分异的动力来源.气孔主要分布于植物叶片下表皮并参与调控碳水摄入与排出, 在其发育过程中迫于环境压力与生存维持, 气孔往往在多个维度存在权衡(Brodribb & Jordan,
2011 ; Brodribb
et al. ,
2013 ; 丁凌子等,
2014 ; Fiorin
et al. ,
2016 ).虽然气孔数量的增加有助于提高光合效率, 但过多的气孔会加速叶内水汽逸散(潘莹萍和陈亚鹏,
2014 ; 王学芳等,
2014 ; 宋慧清等,
2020 ), 而且气孔的建成需要消耗资源, 这就决定了单位面积的气孔数量存在分布上限.气孔密度与大小共同决定了最大可能气孔导度并影响最大同化速率, 在考虑气孔建成实际资源消耗与光合作用水气协调的过程中, 两个相互矛盾的刚需必然达到最优化(Sack & Scoffoni,
2013 ).气孔面积与开度分别代表着气孔发育与运动, 且气孔发育总是先于气孔运动, 尽管大气孔对于开度的调控不及小气孔灵敏, 但可以获得相对较多的碳收益.就此而言, 气孔发育和运动过程中资源分配不均甚至存在矛盾(Xu & Zhou,
2008 ; Drake
et al. ,
2013 ).在异质生境中, 气孔各性状并非独立变化, 而是通过性状或特征组合的协变与权衡来共同适应环境.有限资源在植物体内的分配与权衡已通过异速生长理论在多个尺度得到证实(陈国鹏等,
2016 ; 段贝贝等,
2016 ; 杨克彤等,
2020 ), 但以往研究着重于相对宏观的尺度如群落、个体、构件与器官, 而且忽视了系统发育对资源权衡的影响, 对微观尺度如气孔性状的权衡机制关注较少.为此, 本文希望通过研究气孔特征并结合系统发育来探讨微观尺度气孔性状间的生长关系, 以进一步揭示微观尺度的权衡过程, 并丰富植物生活史研究. ...
Smaller, faster stomata: scaling of stomatal size, rate of response, and stomatal conductance
4
2013
... 气孔作为植物叶表皮的特殊结构, 是植物与外界环境进行物质交换的门户(宋璐璐等,
2011 ; 朱燕华等,
2011 ), 也是植物体内水汽逸散的主要通道(金鹰和王传宽,
2015 ; 范嘉智等,
2016 ).气孔易受环境条件的修饰, 其形态特征是植物进化过程中对外界环境长期适应的结果(王碧霞等,
2010 ; 朱燕华等,
2011 ; 温婧雯等,
2018 ).为适应(或响应)复杂多变的环境, 气孔一方面通过调节开闭来应对短期或局域环境变化, 另一方面则协调大小和密度来适应异质生境(Brodribb & Jordan,
2011 ; Drake
et al. ,
2013 ; 金鹰和王传宽,
2015 ; 高冠龙等,
2018 ).利用气孔特征反映植物对环境变化的响应有助于明晰植物应对水气耦合过程的策略. ...
... 系统发育和环境异质是影响植物性状发生分异的动力来源.气孔主要分布于植物叶片下表皮并参与调控碳水摄入与排出, 在其发育过程中迫于环境压力与生存维持, 气孔往往在多个维度存在权衡(Brodribb & Jordan,
2011 ; Brodribb
et al. ,
2013 ; 丁凌子等,
2014 ; Fiorin
et al. ,
2016 ).虽然气孔数量的增加有助于提高光合效率, 但过多的气孔会加速叶内水汽逸散(潘莹萍和陈亚鹏,
2014 ; 王学芳等,
2014 ; 宋慧清等,
2020 ), 而且气孔的建成需要消耗资源, 这就决定了单位面积的气孔数量存在分布上限.气孔密度与大小共同决定了最大可能气孔导度并影响最大同化速率, 在考虑气孔建成实际资源消耗与光合作用水气协调的过程中, 两个相互矛盾的刚需必然达到最优化(Sack & Scoffoni,
2013 ).气孔面积与开度分别代表着气孔发育与运动, 且气孔发育总是先于气孔运动, 尽管大气孔对于开度的调控不及小气孔灵敏, 但可以获得相对较多的碳收益.就此而言, 气孔发育和运动过程中资源分配不均甚至存在矛盾(Xu & Zhou,
2008 ; Drake
et al. ,
2013 ).在异质生境中, 气孔各性状并非独立变化, 而是通过性状或特征组合的协变与权衡来共同适应环境.有限资源在植物体内的分配与权衡已通过异速生长理论在多个尺度得到证实(陈国鹏等,
2016 ; 段贝贝等,
2016 ; 杨克彤等,
2020 ), 但以往研究着重于相对宏观的尺度如群落、个体、构件与器官, 而且忽视了系统发育对资源权衡的影响, 对微观尺度如气孔性状的权衡机制关注较少.为此, 本文希望通过研究气孔特征并结合系统发育来探讨微观尺度气孔性状间的生长关系, 以进一步揭示微观尺度的权衡过程, 并丰富植物生活史研究. ...
... 气孔面积与开度的关系表明, 随着气孔面积逐渐增大, 气孔开度也逐渐增大, 但气孔开度的扩大速率不及面积的增加速率.有研究认为, 气孔越大对气孔孔径调控的能力越弱, 在遭遇干旱胁迫时所丢失的水分就越多(Drake
et al. ,
2013 ; Sperry
et al. ,
2017 ), 虽然较大的气孔开度可以交换更多的碳, 但这种交换存在风险.气孔面积增大时, 保持CO
2 摄入量不变的情况下, 减小气孔开度依旧可以达到等效水平.但是, 大气孔对开度的调节缺乏灵敏性并对应着较低的气体交换速率, 随着气孔开度逐渐增大, 其进一步开张的阻力也越大、耗费能量越多, 它们的慢动力学特性也使大气孔植物在遭遇干旱等环境胁迫时更容易发生功能紊乱(Hetherington & Woodward,
2003 ; 范嘉智等,
2016 ; 胡选萍等,
2016 ). ...
... 气孔数量的发育往往先于面积的发育(Xu & Zhou,
2008 ).当气孔密度逐渐增加, 因其空间有限, 气孔面积发育受阻, 气孔开度也随之减小, 较小的气孔开度与较大的气孔密度弥补了碳获取通道的不足.干旱区植物生长的主要限制因子是水分, 较小的气孔面积便于控制气孔的开合, 而较小的气孔开度则有助于减少水分蒸腾, 缓解水分压力(Hetherington & Woodward,
2003 ; Drake
et al. ,
2013 ; Sperry
et al. ,
2017 ).虽然气孔密度增加有助于获取CO
2 物质量和外界信息流, 但气孔密度并非越大越好, 在遵循“单细胞间隔法则”的基础上(宋璐璐等,
2011 ; 任昱等,
2014 ; 王学芳等,
2014 ; 范嘉智等,
2016 ), 高密度的气孔反而造成气孔拥挤与调控失灵, 为灵活调控气孔开合, 减小气孔面积并降低开度更有助于精准调控. ...
兰州北山不同坡向刺槐叶脉密度与气孔性状的关联性分析
4
2016
... 系统发育和环境异质是影响植物性状发生分异的动力来源.气孔主要分布于植物叶片下表皮并参与调控碳水摄入与排出, 在其发育过程中迫于环境压力与生存维持, 气孔往往在多个维度存在权衡(Brodribb & Jordan,
2011 ; Brodribb
et al. ,
2013 ; 丁凌子等,
2014 ; Fiorin
et al. ,
2016 ).虽然气孔数量的增加有助于提高光合效率, 但过多的气孔会加速叶内水汽逸散(潘莹萍和陈亚鹏,
2014 ; 王学芳等,
2014 ; 宋慧清等,
2020 ), 而且气孔的建成需要消耗资源, 这就决定了单位面积的气孔数量存在分布上限.气孔密度与大小共同决定了最大可能气孔导度并影响最大同化速率, 在考虑气孔建成实际资源消耗与光合作用水气协调的过程中, 两个相互矛盾的刚需必然达到最优化(Sack & Scoffoni,
2013 ).气孔面积与开度分别代表着气孔发育与运动, 且气孔发育总是先于气孔运动, 尽管大气孔对于开度的调控不及小气孔灵敏, 但可以获得相对较多的碳收益.就此而言, 气孔发育和运动过程中资源分配不均甚至存在矛盾(Xu & Zhou,
2008 ; Drake
et al. ,
2013 ).在异质生境中, 气孔各性状并非独立变化, 而是通过性状或特征组合的协变与权衡来共同适应环境.有限资源在植物体内的分配与权衡已通过异速生长理论在多个尺度得到证实(陈国鹏等,
2016 ; 段贝贝等,
2016 ; 杨克彤等,
2020 ), 但以往研究着重于相对宏观的尺度如群落、个体、构件与器官, 而且忽视了系统发育对资源权衡的影响, 对微观尺度如气孔性状的权衡机制关注较少.为此, 本文希望通过研究气孔特征并结合系统发育来探讨微观尺度气孔性状间的生长关系, 以进一步揭示微观尺度的权衡过程, 并丰富植物生活史研究. ...
... 黄土高原的高温、干旱、强日照和城市内空气污染、土壤重金属危害与CO
2 施肥效应共同驱动着绿化植物的适应策略并使其在外部形态上加以表征(段贝贝等,
2016 ).本研究以兰州市主要绿化植物为研究对象, 测定气孔形态特征并结合系统发育以探究叶片气孔性状的权衡关系, 并试图回答以下问题: (1)气孔特征是否受植物生长型、叶习性及系统发育的影响? (2)气孔长度与宽度、气孔数量和密度与气孔面积之间是否存在权衡? (3)能否根据气孔特征进行物种功能群划分? ...
... 兰州市(103.40° E, 36.03° N, 海拔1 500 m)位于黄土高原西北部, 地势西南高东北低, 呈东西狭长带状盆地特征, 属于大陆性很强的温带季风气候.市区地貌以山地丘陵为主, 占72.9%, 沟壑纵横, 地形破碎, 谷深坡陡, 年平均气温为10.3 ℃, 年日照时间2 446 h, 无霜期180天, 年降水量327 mm左右, 主要集中在6-9月(占全年降水量的50%-70%).土壤以灰钙土为主, 其次为栗钙土和灰褐土, 自然植被稀少, 主要是多年生禾草、旱生灌木和小乔木(段贝贝等,
2016 ; 杨克彤等,
2020 ), 覆盖率低于23%, 其中森林覆盖率仅为6.67%, 低于全国平均水平13.95%, 植被类型单一、结构简单, 是我国生态环境十分脆弱的地区之一. ...
... 功能各异的群组是稳定占据生态位并差异化利用资源的具象.根据气孔性状对40种植物进行功能群划分, 发现不同群组之间气孔性状差异显著(
图4 ).大气孔植物如山楂、小叶黄杨和费菜分别代表了乡土树种、常绿物种与景天酸代谢植物, 这些植物特殊的适应功能决定了即便在干旱区, 它们依旧能够发育出较大的气孔, 这可能是物种特性决定的(罗丹丹等,
2019 ).植物的小气孔与高气孔密度以及低气孔开度和开张比更具备典型旱生植物的形态, 这可能是对黄土高原干旱生境长期适应的结果(任昱等,
2014 ; 段贝贝等,
2016 ; 胡选萍等,
2016 ).气孔功能群的差异能够高效利用有限空间中的光照与水分等资源, 功能群的组合也维持了垂直梯度上的层级结构, 这有助于维护生境小气候的稳态, 但植物气孔类群分化的驱动因素需进一步研究揭示. ...
兰州北山不同坡向刺槐叶脉密度与气孔性状的关联性分析
4
2016
... 系统发育和环境异质是影响植物性状发生分异的动力来源.气孔主要分布于植物叶片下表皮并参与调控碳水摄入与排出, 在其发育过程中迫于环境压力与生存维持, 气孔往往在多个维度存在权衡(Brodribb & Jordan,
2011 ; Brodribb
et al. ,
2013 ; 丁凌子等,
2014 ; Fiorin
et al. ,
2016 ).虽然气孔数量的增加有助于提高光合效率, 但过多的气孔会加速叶内水汽逸散(潘莹萍和陈亚鹏,
2014 ; 王学芳等,
2014 ; 宋慧清等,
2020 ), 而且气孔的建成需要消耗资源, 这就决定了单位面积的气孔数量存在分布上限.气孔密度与大小共同决定了最大可能气孔导度并影响最大同化速率, 在考虑气孔建成实际资源消耗与光合作用水气协调的过程中, 两个相互矛盾的刚需必然达到最优化(Sack & Scoffoni,
2013 ).气孔面积与开度分别代表着气孔发育与运动, 且气孔发育总是先于气孔运动, 尽管大气孔对于开度的调控不及小气孔灵敏, 但可以获得相对较多的碳收益.就此而言, 气孔发育和运动过程中资源分配不均甚至存在矛盾(Xu & Zhou,
2008 ; Drake
et al. ,
2013 ).在异质生境中, 气孔各性状并非独立变化, 而是通过性状或特征组合的协变与权衡来共同适应环境.有限资源在植物体内的分配与权衡已通过异速生长理论在多个尺度得到证实(陈国鹏等,
2016 ; 段贝贝等,
2016 ; 杨克彤等,
2020 ), 但以往研究着重于相对宏观的尺度如群落、个体、构件与器官, 而且忽视了系统发育对资源权衡的影响, 对微观尺度如气孔性状的权衡机制关注较少.为此, 本文希望通过研究气孔特征并结合系统发育来探讨微观尺度气孔性状间的生长关系, 以进一步揭示微观尺度的权衡过程, 并丰富植物生活史研究. ...
... 黄土高原的高温、干旱、强日照和城市内空气污染、土壤重金属危害与CO
2 施肥效应共同驱动着绿化植物的适应策略并使其在外部形态上加以表征(段贝贝等,
2016 ).本研究以兰州市主要绿化植物为研究对象, 测定气孔形态特征并结合系统发育以探究叶片气孔性状的权衡关系, 并试图回答以下问题: (1)气孔特征是否受植物生长型、叶习性及系统发育的影响? (2)气孔长度与宽度、气孔数量和密度与气孔面积之间是否存在权衡? (3)能否根据气孔特征进行物种功能群划分? ...
... 兰州市(103.40° E, 36.03° N, 海拔1 500 m)位于黄土高原西北部, 地势西南高东北低, 呈东西狭长带状盆地特征, 属于大陆性很强的温带季风气候.市区地貌以山地丘陵为主, 占72.9%, 沟壑纵横, 地形破碎, 谷深坡陡, 年平均气温为10.3 ℃, 年日照时间2 446 h, 无霜期180天, 年降水量327 mm左右, 主要集中在6-9月(占全年降水量的50%-70%).土壤以灰钙土为主, 其次为栗钙土和灰褐土, 自然植被稀少, 主要是多年生禾草、旱生灌木和小乔木(段贝贝等,
2016 ; 杨克彤等,
2020 ), 覆盖率低于23%, 其中森林覆盖率仅为6.67%, 低于全国平均水平13.95%, 植被类型单一、结构简单, 是我国生态环境十分脆弱的地区之一. ...
... 功能各异的群组是稳定占据生态位并差异化利用资源的具象.根据气孔性状对40种植物进行功能群划分, 发现不同群组之间气孔性状差异显著(
图4 ).大气孔植物如山楂、小叶黄杨和费菜分别代表了乡土树种、常绿物种与景天酸代谢植物, 这些植物特殊的适应功能决定了即便在干旱区, 它们依旧能够发育出较大的气孔, 这可能是物种特性决定的(罗丹丹等,
2019 ).植物的小气孔与高气孔密度以及低气孔开度和开张比更具备典型旱生植物的形态, 这可能是对黄土高原干旱生境长期适应的结果(任昱等,
2014 ; 段贝贝等,
2016 ; 胡选萍等,
2016 ).气孔功能群的差异能够高效利用有限空间中的光照与水分等资源, 功能群的组合也维持了垂直梯度上的层级结构, 这有助于维护生境小气候的稳态, 但植物气孔类群分化的驱动因素需进一步研究揭示. ...
最优气孔行为理论和气孔导度模拟
3
2016
... 气孔作为植物叶表皮的特殊结构, 是植物与外界环境进行物质交换的门户(宋璐璐等,
2011 ; 朱燕华等,
2011 ), 也是植物体内水汽逸散的主要通道(金鹰和王传宽,
2015 ; 范嘉智等,
2016 ).气孔易受环境条件的修饰, 其形态特征是植物进化过程中对外界环境长期适应的结果(王碧霞等,
2010 ; 朱燕华等,
2011 ; 温婧雯等,
2018 ).为适应(或响应)复杂多变的环境, 气孔一方面通过调节开闭来应对短期或局域环境变化, 另一方面则协调大小和密度来适应异质生境(Brodribb & Jordan,
2011 ; Drake
et al. ,
2013 ; 金鹰和王传宽,
2015 ; 高冠龙等,
2018 ).利用气孔特征反映植物对环境变化的响应有助于明晰植物应对水气耦合过程的策略. ...
... 气孔面积与开度的关系表明, 随着气孔面积逐渐增大, 气孔开度也逐渐增大, 但气孔开度的扩大速率不及面积的增加速率.有研究认为, 气孔越大对气孔孔径调控的能力越弱, 在遭遇干旱胁迫时所丢失的水分就越多(Drake
et al. ,
2013 ; Sperry
et al. ,
2017 ), 虽然较大的气孔开度可以交换更多的碳, 但这种交换存在风险.气孔面积增大时, 保持CO
2 摄入量不变的情况下, 减小气孔开度依旧可以达到等效水平.但是, 大气孔对开度的调节缺乏灵敏性并对应着较低的气体交换速率, 随着气孔开度逐渐增大, 其进一步开张的阻力也越大、耗费能量越多, 它们的慢动力学特性也使大气孔植物在遭遇干旱等环境胁迫时更容易发生功能紊乱(Hetherington & Woodward,
2003 ; 范嘉智等,
2016 ; 胡选萍等,
2016 ). ...
... 气孔数量的发育往往先于面积的发育(Xu & Zhou,
2008 ).当气孔密度逐渐增加, 因其空间有限, 气孔面积发育受阻, 气孔开度也随之减小, 较小的气孔开度与较大的气孔密度弥补了碳获取通道的不足.干旱区植物生长的主要限制因子是水分, 较小的气孔面积便于控制气孔的开合, 而较小的气孔开度则有助于减少水分蒸腾, 缓解水分压力(Hetherington & Woodward,
2003 ; Drake
et al. ,
2013 ; Sperry
et al. ,
2017 ).虽然气孔密度增加有助于获取CO
2 物质量和外界信息流, 但气孔密度并非越大越好, 在遵循“单细胞间隔法则”的基础上(宋璐璐等,
2011 ; 任昱等,
2014 ; 王学芳等,
2014 ; 范嘉智等,
2016 ), 高密度的气孔反而造成气孔拥挤与调控失灵, 为灵活调控气孔开合, 减小气孔面积并降低开度更有助于精准调控. ...
最优气孔行为理论和气孔导度模拟
3
2016
... 气孔作为植物叶表皮的特殊结构, 是植物与外界环境进行物质交换的门户(宋璐璐等,
2011 ; 朱燕华等,
2011 ), 也是植物体内水汽逸散的主要通道(金鹰和王传宽,
2015 ; 范嘉智等,
2016 ).气孔易受环境条件的修饰, 其形态特征是植物进化过程中对外界环境长期适应的结果(王碧霞等,
2010 ; 朱燕华等,
2011 ; 温婧雯等,
2018 ).为适应(或响应)复杂多变的环境, 气孔一方面通过调节开闭来应对短期或局域环境变化, 另一方面则协调大小和密度来适应异质生境(Brodribb & Jordan,
2011 ; Drake
et al. ,
2013 ; 金鹰和王传宽,
2015 ; 高冠龙等,
2018 ).利用气孔特征反映植物对环境变化的响应有助于明晰植物应对水气耦合过程的策略. ...
... 气孔面积与开度的关系表明, 随着气孔面积逐渐增大, 气孔开度也逐渐增大, 但气孔开度的扩大速率不及面积的增加速率.有研究认为, 气孔越大对气孔孔径调控的能力越弱, 在遭遇干旱胁迫时所丢失的水分就越多(Drake
et al. ,
2013 ; Sperry
et al. ,
2017 ), 虽然较大的气孔开度可以交换更多的碳, 但这种交换存在风险.气孔面积增大时, 保持CO
2 摄入量不变的情况下, 减小气孔开度依旧可以达到等效水平.但是, 大气孔对开度的调节缺乏灵敏性并对应着较低的气体交换速率, 随着气孔开度逐渐增大, 其进一步开张的阻力也越大、耗费能量越多, 它们的慢动力学特性也使大气孔植物在遭遇干旱等环境胁迫时更容易发生功能紊乱(Hetherington & Woodward,
2003 ; 范嘉智等,
2016 ; 胡选萍等,
2016 ). ...
... 气孔数量的发育往往先于面积的发育(Xu & Zhou,
2008 ).当气孔密度逐渐增加, 因其空间有限, 气孔面积发育受阻, 气孔开度也随之减小, 较小的气孔开度与较大的气孔密度弥补了碳获取通道的不足.干旱区植物生长的主要限制因子是水分, 较小的气孔面积便于控制气孔的开合, 而较小的气孔开度则有助于减少水分蒸腾, 缓解水分压力(Hetherington & Woodward,
2003 ; Drake
et al. ,
2013 ; Sperry
et al. ,
2017 ).虽然气孔密度增加有助于获取CO
2 物质量和外界信息流, 但气孔密度并非越大越好, 在遵循“单细胞间隔法则”的基础上(宋璐璐等,
2011 ; 任昱等,
2014 ; 王学芳等,
2014 ; 范嘉智等,
2016 ), 高密度的气孔反而造成气孔拥挤与调控失灵, 为灵活调控气孔开合, 减小气孔面积并降低开度更有助于精准调控. ...
Transport efficiency through uniformity: organization of veins and stomata in angiosperm leaves
3
2016
... 气孔性状(如气孔密度)的种间变异被认为是植物叶片经济谱的一部分(Loranger & Shipley,
2010 ; 王瑞丽等,
2016 ), 它在维持植物稳定碳收益、调节水力结构、吸收速效养分和信息传递等方面具有重要作用(Mencuccini
et al. ,
2015 ; Fiorin
et al. ,
2016 ).因此, 量化气孔性状在异质生境中的变异程度并解析其应对策略有助于进一步阐明植物对环境的适应机制.然而, 多数研究仅侧重于气孔性状对自然环境(海拔、水分和温度等)和控制试验(光强、氮添加和CO
2 浓度等)的响应并得出了一系列结论, 如: 气孔长度与密度之间为负相关关系(任昱等,
2014 ; 姜星星等,
2018 ), 随海拔升高气孔宽度呈减小趋势(王瑞丽等,
2016 ), 土壤湿度增大会显著降低气孔密度(杨再强等,
2015 ; 夏振华等,
2018 ; 张大才等,
2018 ), 高CO
2 浓度会显著降低气孔开张比(贾剑波等,
2019 )等, 但忽视了气孔自身在发育过程中的优先发育性状及其相互间关系的协变与权衡. ...
... 系统发育和环境异质是影响植物性状发生分异的动力来源.气孔主要分布于植物叶片下表皮并参与调控碳水摄入与排出, 在其发育过程中迫于环境压力与生存维持, 气孔往往在多个维度存在权衡(Brodribb & Jordan,
2011 ; Brodribb
et al. ,
2013 ; 丁凌子等,
2014 ; Fiorin
et al. ,
2016 ).虽然气孔数量的增加有助于提高光合效率, 但过多的气孔会加速叶内水汽逸散(潘莹萍和陈亚鹏,
2014 ; 王学芳等,
2014 ; 宋慧清等,
2020 ), 而且气孔的建成需要消耗资源, 这就决定了单位面积的气孔数量存在分布上限.气孔密度与大小共同决定了最大可能气孔导度并影响最大同化速率, 在考虑气孔建成实际资源消耗与光合作用水气协调的过程中, 两个相互矛盾的刚需必然达到最优化(Sack & Scoffoni,
2013 ).气孔面积与开度分别代表着气孔发育与运动, 且气孔发育总是先于气孔运动, 尽管大气孔对于开度的调控不及小气孔灵敏, 但可以获得相对较多的碳收益.就此而言, 气孔发育和运动过程中资源分配不均甚至存在矛盾(Xu & Zhou,
2008 ; Drake
et al. ,
2013 ).在异质生境中, 气孔各性状并非独立变化, 而是通过性状或特征组合的协变与权衡来共同适应环境.有限资源在植物体内的分配与权衡已通过异速生长理论在多个尺度得到证实(陈国鹏等,
2016 ; 段贝贝等,
2016 ; 杨克彤等,
2020 ), 但以往研究着重于相对宏观的尺度如群落、个体、构件与器官, 而且忽视了系统发育对资源权衡的影响, 对微观尺度如气孔性状的权衡机制关注较少.为此, 本文希望通过研究气孔特征并结合系统发育来探讨微观尺度气孔性状间的生长关系, 以进一步揭示微观尺度的权衡过程, 并丰富植物生活史研究. ...
... 系统发育决定着植物的亲缘关系并印刻着植物进化的信息, 在研究植物性状特征的过程中是否考虑系统发育将显著地影响研究结果(Ackerly,
2009 ; Zhang
et al. ,
2014 ).本研究中气孔性状间存在显著的相关关系, 在去除系统发育信息后, 这种关系大都发生了转变, 仅气孔长度与宽度依旧维持在稳健的正相关关系, 表明气孔长度与宽度间可能存在关联进化(张亚等,
2014 ; 高林浩等,
2020 ).即不论是否考虑系统发育, 气孔长度与宽度间的正相关关系都是存在的, 暗示这种协同关系可能在古老的植物类群中就已经出现.有研究认为, 植物气孔几何形状和间距总是沿着空间上最优的异速生长关系进化(Sack & Scoffoni,
2013 ; Fiorin
et al. ,
2016 ; Sperry
et al. ,
2017 ), 由于气孔发育属于同步型, 在气孔发育过程中长度与宽度总保持在相对稳定的比率(Franks & Farquhar,
2001 ), 这也从一定程度上解释了去除系统发育后, 气孔长度与宽度依旧保持稳健相关关系的原因.此外, 系统发育信号检测结果表明, 气孔性状的发育信号较弱(
K < 1), 表明气孔性状受系统发育影响较小, 但气孔开度和气孔开张比的发育信号显著, 暗示亲缘关系越近的物种气孔开度与开张比越相似(高林浩等,
2020 ).虽然系统发育会影响气孔性状, 但在长时间适应环境的过程中, 系统发育的影响似乎正在被逐渐削弱. ...
The effect of exogenous abscisic acid on stomatal development, stomatal mechanics, and leaf gas exchange in Tradescantia virginiana
1
2001
... 系统发育决定着植物的亲缘关系并印刻着植物进化的信息, 在研究植物性状特征的过程中是否考虑系统发育将显著地影响研究结果(Ackerly,
2009 ; Zhang
et al. ,
2014 ).本研究中气孔性状间存在显著的相关关系, 在去除系统发育信息后, 这种关系大都发生了转变, 仅气孔长度与宽度依旧维持在稳健的正相关关系, 表明气孔长度与宽度间可能存在关联进化(张亚等,
2014 ; 高林浩等,
2020 ).即不论是否考虑系统发育, 气孔长度与宽度间的正相关关系都是存在的, 暗示这种协同关系可能在古老的植物类群中就已经出现.有研究认为, 植物气孔几何形状和间距总是沿着空间上最优的异速生长关系进化(Sack & Scoffoni,
2013 ; Fiorin
et al. ,
2016 ; Sperry
et al. ,
2017 ), 由于气孔发育属于同步型, 在气孔发育过程中长度与宽度总保持在相对稳定的比率(Franks & Farquhar,
2001 ), 这也从一定程度上解释了去除系统发育后, 气孔长度与宽度依旧保持稳健相关关系的原因.此外, 系统发育信号检测结果表明, 气孔性状的发育信号较弱(
K < 1), 表明气孔性状受系统发育影响较小, 但气孔开度和气孔开张比的发育信号显著, 暗示亲缘关系越近的物种气孔开度与开张比越相似(高林浩等,
2020 ).虽然系统发育会影响气孔性状, 但在长时间适应环境的过程中, 系统发育的影响似乎正在被逐渐削弱. ...
植物叶片光合作用的气孔与非气孔限制研究综述
1
2018
... 气孔作为植物叶表皮的特殊结构, 是植物与外界环境进行物质交换的门户(宋璐璐等,
2011 ; 朱燕华等,
2011 ), 也是植物体内水汽逸散的主要通道(金鹰和王传宽,
2015 ; 范嘉智等,
2016 ).气孔易受环境条件的修饰, 其形态特征是植物进化过程中对外界环境长期适应的结果(王碧霞等,
2010 ; 朱燕华等,
2011 ; 温婧雯等,
2018 ).为适应(或响应)复杂多变的环境, 气孔一方面通过调节开闭来应对短期或局域环境变化, 另一方面则协调大小和密度来适应异质生境(Brodribb & Jordan,
2011 ; Drake
et al. ,
2013 ; 金鹰和王传宽,
2015 ; 高冠龙等,
2018 ).利用气孔特征反映植物对环境变化的响应有助于明晰植物应对水气耦合过程的策略. ...
植物叶片光合作用的气孔与非气孔限制研究综述
1
2018
... 气孔作为植物叶表皮的特殊结构, 是植物与外界环境进行物质交换的门户(宋璐璐等,
2011 ; 朱燕华等,
2011 ), 也是植物体内水汽逸散的主要通道(金鹰和王传宽,
2015 ; 范嘉智等,
2016 ).气孔易受环境条件的修饰, 其形态特征是植物进化过程中对外界环境长期适应的结果(王碧霞等,
2010 ; 朱燕华等,
2011 ; 温婧雯等,
2018 ).为适应(或响应)复杂多变的环境, 气孔一方面通过调节开闭来应对短期或局域环境变化, 另一方面则协调大小和密度来适应异质生境(Brodribb & Jordan,
2011 ; Drake
et al. ,
2013 ; 金鹰和王传宽,
2015 ; 高冠龙等,
2018 ).利用气孔特征反映植物对环境变化的响应有助于明晰植物应对水气耦合过程的策略. ...
气候、系统发育对长白山乔灌木比叶面积与叶元素含量关系的影响
2
2020
... 系统发育决定着植物的亲缘关系并印刻着植物进化的信息, 在研究植物性状特征的过程中是否考虑系统发育将显著地影响研究结果(Ackerly,
2009 ; Zhang
et al. ,
2014 ).本研究中气孔性状间存在显著的相关关系, 在去除系统发育信息后, 这种关系大都发生了转变, 仅气孔长度与宽度依旧维持在稳健的正相关关系, 表明气孔长度与宽度间可能存在关联进化(张亚等,
2014 ; 高林浩等,
2020 ).即不论是否考虑系统发育, 气孔长度与宽度间的正相关关系都是存在的, 暗示这种协同关系可能在古老的植物类群中就已经出现.有研究认为, 植物气孔几何形状和间距总是沿着空间上最优的异速生长关系进化(Sack & Scoffoni,
2013 ; Fiorin
et al. ,
2016 ; Sperry
et al. ,
2017 ), 由于气孔发育属于同步型, 在气孔发育过程中长度与宽度总保持在相对稳定的比率(Franks & Farquhar,
2001 ), 这也从一定程度上解释了去除系统发育后, 气孔长度与宽度依旧保持稳健相关关系的原因.此外, 系统发育信号检测结果表明, 气孔性状的发育信号较弱(
K < 1), 表明气孔性状受系统发育影响较小, 但气孔开度和气孔开张比的发育信号显著, 暗示亲缘关系越近的物种气孔开度与开张比越相似(高林浩等,
2020 ).虽然系统发育会影响气孔性状, 但在长时间适应环境的过程中, 系统发育的影响似乎正在被逐渐削弱. ...
... < 1), 表明气孔性状受系统发育影响较小, 但气孔开度和气孔开张比的发育信号显著, 暗示亲缘关系越近的物种气孔开度与开张比越相似(高林浩等,
2020 ).虽然系统发育会影响气孔性状, 但在长时间适应环境的过程中, 系统发育的影响似乎正在被逐渐削弱. ...
气候、系统发育对长白山乔灌木比叶面积与叶元素含量关系的影响
2
2020
... 系统发育决定着植物的亲缘关系并印刻着植物进化的信息, 在研究植物性状特征的过程中是否考虑系统发育将显著地影响研究结果(Ackerly,
2009 ; Zhang
et al. ,
2014 ).本研究中气孔性状间存在显著的相关关系, 在去除系统发育信息后, 这种关系大都发生了转变, 仅气孔长度与宽度依旧维持在稳健的正相关关系, 表明气孔长度与宽度间可能存在关联进化(张亚等,
2014 ; 高林浩等,
2020 ).即不论是否考虑系统发育, 气孔长度与宽度间的正相关关系都是存在的, 暗示这种协同关系可能在古老的植物类群中就已经出现.有研究认为, 植物气孔几何形状和间距总是沿着空间上最优的异速生长关系进化(Sack & Scoffoni,
2013 ; Fiorin
et al. ,
2016 ; Sperry
et al. ,
2017 ), 由于气孔发育属于同步型, 在气孔发育过程中长度与宽度总保持在相对稳定的比率(Franks & Farquhar,
2001 ), 这也从一定程度上解释了去除系统发育后, 气孔长度与宽度依旧保持稳健相关关系的原因.此外, 系统发育信号检测结果表明, 气孔性状的发育信号较弱(
K < 1), 表明气孔性状受系统发育影响较小, 但气孔开度和气孔开张比的发育信号显著, 暗示亲缘关系越近的物种气孔开度与开张比越相似(高林浩等,
2020 ).虽然系统发育会影响气孔性状, 但在长时间适应环境的过程中, 系统发育的影响似乎正在被逐渐削弱. ...
... < 1), 表明气孔性状受系统发育影响较小, 但气孔开度和气孔开张比的发育信号显著, 暗示亲缘关系越近的物种气孔开度与开张比越相似(高林浩等,
2020 ).虽然系统发育会影响气孔性状, 但在长时间适应环境的过程中, 系统发育的影响似乎正在被逐渐削弱. ...
The role of stomata in sensing and driving environmental change
2
2003
... 气孔面积与开度的关系表明, 随着气孔面积逐渐增大, 气孔开度也逐渐增大, 但气孔开度的扩大速率不及面积的增加速率.有研究认为, 气孔越大对气孔孔径调控的能力越弱, 在遭遇干旱胁迫时所丢失的水分就越多(Drake
et al. ,
2013 ; Sperry
et al. ,
2017 ), 虽然较大的气孔开度可以交换更多的碳, 但这种交换存在风险.气孔面积增大时, 保持CO
2 摄入量不变的情况下, 减小气孔开度依旧可以达到等效水平.但是, 大气孔对开度的调节缺乏灵敏性并对应着较低的气体交换速率, 随着气孔开度逐渐增大, 其进一步开张的阻力也越大、耗费能量越多, 它们的慢动力学特性也使大气孔植物在遭遇干旱等环境胁迫时更容易发生功能紊乱(Hetherington & Woodward,
2003 ; 范嘉智等,
2016 ; 胡选萍等,
2016 ). ...
... 气孔数量的发育往往先于面积的发育(Xu & Zhou,
2008 ).当气孔密度逐渐增加, 因其空间有限, 气孔面积发育受阻, 气孔开度也随之减小, 较小的气孔开度与较大的气孔密度弥补了碳获取通道的不足.干旱区植物生长的主要限制因子是水分, 较小的气孔面积便于控制气孔的开合, 而较小的气孔开度则有助于减少水分蒸腾, 缓解水分压力(Hetherington & Woodward,
2003 ; Drake
et al. ,
2013 ; Sperry
et al. ,
2017 ).虽然气孔密度增加有助于获取CO
2 物质量和外界信息流, 但气孔密度并非越大越好, 在遵循“单细胞间隔法则”的基础上(宋璐璐等,
2011 ; 任昱等,
2014 ; 王学芳等,
2014 ; 范嘉智等,
2016 ), 高密度的气孔反而造成气孔拥挤与调控失灵, 为灵活调控气孔开合, 减小气孔面积并降低开度更有助于精准调控. ...
青藏高原草地主要单子叶植物的叶表面特征
2
2016
... 功能各异的群组是稳定占据生态位并差异化利用资源的具象.根据气孔性状对40种植物进行功能群划分, 发现不同群组之间气孔性状差异显著(
图4 ).大气孔植物如山楂、小叶黄杨和费菜分别代表了乡土树种、常绿物种与景天酸代谢植物, 这些植物特殊的适应功能决定了即便在干旱区, 它们依旧能够发育出较大的气孔, 这可能是物种特性决定的(罗丹丹等,
2019 ).植物的小气孔与高气孔密度以及低气孔开度和开张比更具备典型旱生植物的形态, 这可能是对黄土高原干旱生境长期适应的结果(任昱等,
2014 ; 段贝贝等,
2016 ; 胡选萍等,
2016 ).气孔功能群的差异能够高效利用有限空间中的光照与水分等资源, 功能群的组合也维持了垂直梯度上的层级结构, 这有助于维护生境小气候的稳态, 但植物气孔类群分化的驱动因素需进一步研究揭示. ...
... 气孔面积与开度的关系表明, 随着气孔面积逐渐增大, 气孔开度也逐渐增大, 但气孔开度的扩大速率不及面积的增加速率.有研究认为, 气孔越大对气孔孔径调控的能力越弱, 在遭遇干旱胁迫时所丢失的水分就越多(Drake
et al. ,
2013 ; Sperry
et al. ,
2017 ), 虽然较大的气孔开度可以交换更多的碳, 但这种交换存在风险.气孔面积增大时, 保持CO
2 摄入量不变的情况下, 减小气孔开度依旧可以达到等效水平.但是, 大气孔对开度的调节缺乏灵敏性并对应着较低的气体交换速率, 随着气孔开度逐渐增大, 其进一步开张的阻力也越大、耗费能量越多, 它们的慢动力学特性也使大气孔植物在遭遇干旱等环境胁迫时更容易发生功能紊乱(Hetherington & Woodward,
2003 ; 范嘉智等,
2016 ; 胡选萍等,
2016 ). ...
青藏高原草地主要单子叶植物的叶表面特征
2
2016
... 功能各异的群组是稳定占据生态位并差异化利用资源的具象.根据气孔性状对40种植物进行功能群划分, 发现不同群组之间气孔性状差异显著(
图4 ).大气孔植物如山楂、小叶黄杨和费菜分别代表了乡土树种、常绿物种与景天酸代谢植物, 这些植物特殊的适应功能决定了即便在干旱区, 它们依旧能够发育出较大的气孔, 这可能是物种特性决定的(罗丹丹等,
2019 ).植物的小气孔与高气孔密度以及低气孔开度和开张比更具备典型旱生植物的形态, 这可能是对黄土高原干旱生境长期适应的结果(任昱等,
2014 ; 段贝贝等,
2016 ; 胡选萍等,
2016 ).气孔功能群的差异能够高效利用有限空间中的光照与水分等资源, 功能群的组合也维持了垂直梯度上的层级结构, 这有助于维护生境小气候的稳态, 但植物气孔类群分化的驱动因素需进一步研究揭示. ...
... 气孔面积与开度的关系表明, 随着气孔面积逐渐增大, 气孔开度也逐渐增大, 但气孔开度的扩大速率不及面积的增加速率.有研究认为, 气孔越大对气孔孔径调控的能力越弱, 在遭遇干旱胁迫时所丢失的水分就越多(Drake
et al. ,
2013 ; Sperry
et al. ,
2017 ), 虽然较大的气孔开度可以交换更多的碳, 但这种交换存在风险.气孔面积增大时, 保持CO
2 摄入量不变的情况下, 减小气孔开度依旧可以达到等效水平.但是, 大气孔对开度的调节缺乏灵敏性并对应着较低的气体交换速率, 随着气孔开度逐渐增大, 其进一步开张的阻力也越大、耗费能量越多, 它们的慢动力学特性也使大气孔植物在遭遇干旱等环境胁迫时更容易发生功能紊乱(Hetherington & Woodward,
2003 ; 范嘉智等,
2016 ; 胡选萍等,
2016 ). ...
水碳控制条件对闽楠叶片气孔特征和气体交换参数的影响
1
2019
... 气孔性状(如气孔密度)的种间变异被认为是植物叶片经济谱的一部分(Loranger & Shipley,
2010 ; 王瑞丽等,
2016 ), 它在维持植物稳定碳收益、调节水力结构、吸收速效养分和信息传递等方面具有重要作用(Mencuccini
et al. ,
2015 ; Fiorin
et al. ,
2016 ).因此, 量化气孔性状在异质生境中的变异程度并解析其应对策略有助于进一步阐明植物对环境的适应机制.然而, 多数研究仅侧重于气孔性状对自然环境(海拔、水分和温度等)和控制试验(光强、氮添加和CO
2 浓度等)的响应并得出了一系列结论, 如: 气孔长度与密度之间为负相关关系(任昱等,
2014 ; 姜星星等,
2018 ), 随海拔升高气孔宽度呈减小趋势(王瑞丽等,
2016 ), 土壤湿度增大会显著降低气孔密度(杨再强等,
2015 ; 夏振华等,
2018 ; 张大才等,
2018 ), 高CO
2 浓度会显著降低气孔开张比(贾剑波等,
2019 )等, 但忽视了气孔自身在发育过程中的优先发育性状及其相互间关系的协变与权衡. ...
水碳控制条件对闽楠叶片气孔特征和气体交换参数的影响
1
2019
... 气孔性状(如气孔密度)的种间变异被认为是植物叶片经济谱的一部分(Loranger & Shipley,
2010 ; 王瑞丽等,
2016 ), 它在维持植物稳定碳收益、调节水力结构、吸收速效养分和信息传递等方面具有重要作用(Mencuccini
et al. ,
2015 ; Fiorin
et al. ,
2016 ).因此, 量化气孔性状在异质生境中的变异程度并解析其应对策略有助于进一步阐明植物对环境的适应机制.然而, 多数研究仅侧重于气孔性状对自然环境(海拔、水分和温度等)和控制试验(光强、氮添加和CO
2 浓度等)的响应并得出了一系列结论, 如: 气孔长度与密度之间为负相关关系(任昱等,
2014 ; 姜星星等,
2018 ), 随海拔升高气孔宽度呈减小趋势(王瑞丽等,
2016 ), 土壤湿度增大会显著降低气孔密度(杨再强等,
2015 ; 夏振华等,
2018 ; 张大才等,
2018 ), 高CO
2 浓度会显著降低气孔开张比(贾剑波等,
2019 )等, 但忽视了气孔自身在发育过程中的优先发育性状及其相互间关系的协变与权衡. ...
我国东部典型森林木本植物的气孔特征及其对氮添加的响应
1
2018
... 气孔性状(如气孔密度)的种间变异被认为是植物叶片经济谱的一部分(Loranger & Shipley,
2010 ; 王瑞丽等,
2016 ), 它在维持植物稳定碳收益、调节水力结构、吸收速效养分和信息传递等方面具有重要作用(Mencuccini
et al. ,
2015 ; Fiorin
et al. ,
2016 ).因此, 量化气孔性状在异质生境中的变异程度并解析其应对策略有助于进一步阐明植物对环境的适应机制.然而, 多数研究仅侧重于气孔性状对自然环境(海拔、水分和温度等)和控制试验(光强、氮添加和CO
2 浓度等)的响应并得出了一系列结论, 如: 气孔长度与密度之间为负相关关系(任昱等,
2014 ; 姜星星等,
2018 ), 随海拔升高气孔宽度呈减小趋势(王瑞丽等,
2016 ), 土壤湿度增大会显著降低气孔密度(杨再强等,
2015 ; 夏振华等,
2018 ; 张大才等,
2018 ), 高CO
2 浓度会显著降低气孔开张比(贾剑波等,
2019 )等, 但忽视了气孔自身在发育过程中的优先发育性状及其相互间关系的协变与权衡. ...
我国东部典型森林木本植物的气孔特征及其对氮添加的响应
1
2018
... 气孔性状(如气孔密度)的种间变异被认为是植物叶片经济谱的一部分(Loranger & Shipley,
2010 ; 王瑞丽等,
2016 ), 它在维持植物稳定碳收益、调节水力结构、吸收速效养分和信息传递等方面具有重要作用(Mencuccini
et al. ,
2015 ; Fiorin
et al. ,
2016 ).因此, 量化气孔性状在异质生境中的变异程度并解析其应对策略有助于进一步阐明植物对环境的适应机制.然而, 多数研究仅侧重于气孔性状对自然环境(海拔、水分和温度等)和控制试验(光强、氮添加和CO
2 浓度等)的响应并得出了一系列结论, 如: 气孔长度与密度之间为负相关关系(任昱等,
2014 ; 姜星星等,
2018 ), 随海拔升高气孔宽度呈减小趋势(王瑞丽等,
2016 ), 土壤湿度增大会显著降低气孔密度(杨再强等,
2015 ; 夏振华等,
2018 ; 张大才等,
2018 ), 高CO
2 浓度会显著降低气孔开张比(贾剑波等,
2019 )等, 但忽视了气孔自身在发育过程中的优先发育性状及其相互间关系的协变与权衡. ...
植物叶片水力与经济性状权衡关系的研究进展
3
2015
... 气孔作为植物叶表皮的特殊结构, 是植物与外界环境进行物质交换的门户(宋璐璐等,
2011 ; 朱燕华等,
2011 ), 也是植物体内水汽逸散的主要通道(金鹰和王传宽,
2015 ; 范嘉智等,
2016 ).气孔易受环境条件的修饰, 其形态特征是植物进化过程中对外界环境长期适应的结果(王碧霞等,
2010 ; 朱燕华等,
2011 ; 温婧雯等,
2018 ).为适应(或响应)复杂多变的环境, 气孔一方面通过调节开闭来应对短期或局域环境变化, 另一方面则协调大小和密度来适应异质生境(Brodribb & Jordan,
2011 ; Drake
et al. ,
2013 ; 金鹰和王传宽,
2015 ; 高冠龙等,
2018 ).利用气孔特征反映植物对环境变化的响应有助于明晰植物应对水气耦合过程的策略. ...
... ; 金鹰和王传宽,
2015 ; 高冠龙等,
2018 ).利用气孔特征反映植物对环境变化的响应有助于明晰植物应对水气耦合过程的策略. ...
... 生长型和叶习性是引起植物性状变异的重要因素(Sack & Scoffoni,
2013 ; Mencuccini
et al. ,
2015 ).本研究中气孔性状在4种生长型和2种叶习性中均表现出显著差异.生长型主要造成局域环境内光照、温度和水分的分配不均, 叶习性则决定了植物的碳收益方式和水力结构.林冠上层高大乔木叶的生境为光照相对较强、温度变幅大而水分不足, 灌木或草本则相反.冠层植物叶片往往接受更多辐射, 需要较多的水气蒸腾以维持叶表温度(吴冰洁等,
2015 ), 常分化出小气孔开度、密集的气孔来应对强光照与高蒸腾强度; 林下植物则发育出大而疏散的气孔与较大的气孔开度, 这有助于叶片排出多余的水分(熊慧等,
2014 ; 王海珍等,
2015 ).相比于阔叶植物, 常绿植物叶片小而厚, 其表皮功能与微观结构也相对复杂(朱燕华等,
2011 ; 熊慧等,
2014 ).除气孔开张比外, 本研究中常绿植物的气孔性状普遍大于落叶植物, 这既有别于湿生物种大而疏散的气孔, 也与旱生植物小而密集的气孔有所区别.以往研究表明, 常绿植物的抗旱与抗栓塞能力强于落叶植物, 而且在叶片建成时更倾向于“缓慢投资-收益”策略(潘莹萍和陈亚鹏,
2014 ; 金鹰和王传宽,
2015 ).由于常绿植物叶片建成初期的碳投资高于落叶植物, 需要在叶片发育后期节约并获取更多的碳来偿还成本.因此, 常绿植物趋向于发育大而紧密的气孔以加速碳同化和积累更多的干物质. ...
植物叶片水力与经济性状权衡关系的研究进展
3
2015
... 气孔作为植物叶表皮的特殊结构, 是植物与外界环境进行物质交换的门户(宋璐璐等,
2011 ; 朱燕华等,
2011 ), 也是植物体内水汽逸散的主要通道(金鹰和王传宽,
2015 ; 范嘉智等,
2016 ).气孔易受环境条件的修饰, 其形态特征是植物进化过程中对外界环境长期适应的结果(王碧霞等,
2010 ; 朱燕华等,
2011 ; 温婧雯等,
2018 ).为适应(或响应)复杂多变的环境, 气孔一方面通过调节开闭来应对短期或局域环境变化, 另一方面则协调大小和密度来适应异质生境(Brodribb & Jordan,
2011 ; Drake
et al. ,
2013 ; 金鹰和王传宽,
2015 ; 高冠龙等,
2018 ).利用气孔特征反映植物对环境变化的响应有助于明晰植物应对水气耦合过程的策略. ...
... ; 金鹰和王传宽,
2015 ; 高冠龙等,
2018 ).利用气孔特征反映植物对环境变化的响应有助于明晰植物应对水气耦合过程的策略. ...
... 生长型和叶习性是引起植物性状变异的重要因素(Sack & Scoffoni,
2013 ; Mencuccini
et al. ,
2015 ).本研究中气孔性状在4种生长型和2种叶习性中均表现出显著差异.生长型主要造成局域环境内光照、温度和水分的分配不均, 叶习性则决定了植物的碳收益方式和水力结构.林冠上层高大乔木叶的生境为光照相对较强、温度变幅大而水分不足, 灌木或草本则相反.冠层植物叶片往往接受更多辐射, 需要较多的水气蒸腾以维持叶表温度(吴冰洁等,
2015 ), 常分化出小气孔开度、密集的气孔来应对强光照与高蒸腾强度; 林下植物则发育出大而疏散的气孔与较大的气孔开度, 这有助于叶片排出多余的水分(熊慧等,
2014 ; 王海珍等,
2015 ).相比于阔叶植物, 常绿植物叶片小而厚, 其表皮功能与微观结构也相对复杂(朱燕华等,
2011 ; 熊慧等,
2014 ).除气孔开张比外, 本研究中常绿植物的气孔性状普遍大于落叶植物, 这既有别于湿生物种大而疏散的气孔, 也与旱生植物小而密集的气孔有所区别.以往研究表明, 常绿植物的抗旱与抗栓塞能力强于落叶植物, 而且在叶片建成时更倾向于“缓慢投资-收益”策略(潘莹萍和陈亚鹏,
2014 ; 金鹰和王传宽,
2015 ).由于常绿植物叶片建成初期的碳投资高于落叶植物, 需要在叶片发育后期节约并获取更多的碳来偿还成本.因此, 常绿植物趋向于发育大而紧密的气孔以加速碳同化和积累更多的干物质. ...
Interspecific covariation between stomatal density and other functional leaf traits in a local flora
1
2010
... 气孔性状(如气孔密度)的种间变异被认为是植物叶片经济谱的一部分(Loranger & Shipley,
2010 ; 王瑞丽等,
2016 ), 它在维持植物稳定碳收益、调节水力结构、吸收速效养分和信息传递等方面具有重要作用(Mencuccini
et al. ,
2015 ; Fiorin
et al. ,
2016 ).因此, 量化气孔性状在异质生境中的变异程度并解析其应对策略有助于进一步阐明植物对环境的适应机制.然而, 多数研究仅侧重于气孔性状对自然环境(海拔、水分和温度等)和控制试验(光强、氮添加和CO
2 浓度等)的响应并得出了一系列结论, 如: 气孔长度与密度之间为负相关关系(任昱等,
2014 ; 姜星星等,
2018 ), 随海拔升高气孔宽度呈减小趋势(王瑞丽等,
2016 ), 土壤湿度增大会显著降低气孔密度(杨再强等,
2015 ; 夏振华等,
2018 ; 张大才等,
2018 ), 高CO
2 浓度会显著降低气孔开张比(贾剑波等,
2019 )等, 但忽视了气孔自身在发育过程中的优先发育性状及其相互间关系的协变与权衡. ...
植物应对干旱胁迫的气孔调节
1
2019
... 功能各异的群组是稳定占据生态位并差异化利用资源的具象.根据气孔性状对40种植物进行功能群划分, 发现不同群组之间气孔性状差异显著(
图4 ).大气孔植物如山楂、小叶黄杨和费菜分别代表了乡土树种、常绿物种与景天酸代谢植物, 这些植物特殊的适应功能决定了即便在干旱区, 它们依旧能够发育出较大的气孔, 这可能是物种特性决定的(罗丹丹等,
2019 ).植物的小气孔与高气孔密度以及低气孔开度和开张比更具备典型旱生植物的形态, 这可能是对黄土高原干旱生境长期适应的结果(任昱等,
2014 ; 段贝贝等,
2016 ; 胡选萍等,
2016 ).气孔功能群的差异能够高效利用有限空间中的光照与水分等资源, 功能群的组合也维持了垂直梯度上的层级结构, 这有助于维护生境小气候的稳态, 但植物气孔类群分化的驱动因素需进一步研究揭示. ...
植物应对干旱胁迫的气孔调节
1
2019
... 功能各异的群组是稳定占据生态位并差异化利用资源的具象.根据气孔性状对40种植物进行功能群划分, 发现不同群组之间气孔性状差异显著(
图4 ).大气孔植物如山楂、小叶黄杨和费菜分别代表了乡土树种、常绿物种与景天酸代谢植物, 这些植物特殊的适应功能决定了即便在干旱区, 它们依旧能够发育出较大的气孔, 这可能是物种特性决定的(罗丹丹等,
2019 ).植物的小气孔与高气孔密度以及低气孔开度和开张比更具备典型旱生植物的形态, 这可能是对黄土高原干旱生境长期适应的结果(任昱等,
2014 ; 段贝贝等,
2016 ; 胡选萍等,
2016 ).气孔功能群的差异能够高效利用有限空间中的光照与水分等资源, 功能群的组合也维持了垂直梯度上的层级结构, 这有助于维护生境小气候的稳态, 但植物气孔类群分化的驱动因素需进一步研究揭示. ...
Coordination of physiological traits involved in drought-induced mortality of woody plants
2
2015
... 气孔性状(如气孔密度)的种间变异被认为是植物叶片经济谱的一部分(Loranger & Shipley,
2010 ; 王瑞丽等,
2016 ), 它在维持植物稳定碳收益、调节水力结构、吸收速效养分和信息传递等方面具有重要作用(Mencuccini
et al. ,
2015 ; Fiorin
et al. ,
2016 ).因此, 量化气孔性状在异质生境中的变异程度并解析其应对策略有助于进一步阐明植物对环境的适应机制.然而, 多数研究仅侧重于气孔性状对自然环境(海拔、水分和温度等)和控制试验(光强、氮添加和CO
2 浓度等)的响应并得出了一系列结论, 如: 气孔长度与密度之间为负相关关系(任昱等,
2014 ; 姜星星等,
2018 ), 随海拔升高气孔宽度呈减小趋势(王瑞丽等,
2016 ), 土壤湿度增大会显著降低气孔密度(杨再强等,
2015 ; 夏振华等,
2018 ; 张大才等,
2018 ), 高CO
2 浓度会显著降低气孔开张比(贾剑波等,
2019 )等, 但忽视了气孔自身在发育过程中的优先发育性状及其相互间关系的协变与权衡. ...
... 生长型和叶习性是引起植物性状变异的重要因素(Sack & Scoffoni,
2013 ; Mencuccini
et al. ,
2015 ).本研究中气孔性状在4种生长型和2种叶习性中均表现出显著差异.生长型主要造成局域环境内光照、温度和水分的分配不均, 叶习性则决定了植物的碳收益方式和水力结构.林冠上层高大乔木叶的生境为光照相对较强、温度变幅大而水分不足, 灌木或草本则相反.冠层植物叶片往往接受更多辐射, 需要较多的水气蒸腾以维持叶表温度(吴冰洁等,
2015 ), 常分化出小气孔开度、密集的气孔来应对强光照与高蒸腾强度; 林下植物则发育出大而疏散的气孔与较大的气孔开度, 这有助于叶片排出多余的水分(熊慧等,
2014 ; 王海珍等,
2015 ).相比于阔叶植物, 常绿植物叶片小而厚, 其表皮功能与微观结构也相对复杂(朱燕华等,
2011 ; 熊慧等,
2014 ).除气孔开张比外, 本研究中常绿植物的气孔性状普遍大于落叶植物, 这既有别于湿生物种大而疏散的气孔, 也与旱生植物小而密集的气孔有所区别.以往研究表明, 常绿植物的抗旱与抗栓塞能力强于落叶植物, 而且在叶片建成时更倾向于“缓慢投资-收益”策略(潘莹萍和陈亚鹏,
2014 ; 金鹰和王传宽,
2015 ).由于常绿植物叶片建成初期的碳投资高于落叶植物, 需要在叶片发育后期节约并获取更多的碳来偿还成本.因此, 常绿植物趋向于发育大而紧密的气孔以加速碳同化和积累更多的干物质. ...
叶片水力性状研究进展
2
2014
... 系统发育和环境异质是影响植物性状发生分异的动力来源.气孔主要分布于植物叶片下表皮并参与调控碳水摄入与排出, 在其发育过程中迫于环境压力与生存维持, 气孔往往在多个维度存在权衡(Brodribb & Jordan,
2011 ; Brodribb
et al. ,
2013 ; 丁凌子等,
2014 ; Fiorin
et al. ,
2016 ).虽然气孔数量的增加有助于提高光合效率, 但过多的气孔会加速叶内水汽逸散(潘莹萍和陈亚鹏,
2014 ; 王学芳等,
2014 ; 宋慧清等,
2020 ), 而且气孔的建成需要消耗资源, 这就决定了单位面积的气孔数量存在分布上限.气孔密度与大小共同决定了最大可能气孔导度并影响最大同化速率, 在考虑气孔建成实际资源消耗与光合作用水气协调的过程中, 两个相互矛盾的刚需必然达到最优化(Sack & Scoffoni,
2013 ).气孔面积与开度分别代表着气孔发育与运动, 且气孔发育总是先于气孔运动, 尽管大气孔对于开度的调控不及小气孔灵敏, 但可以获得相对较多的碳收益.就此而言, 气孔发育和运动过程中资源分配不均甚至存在矛盾(Xu & Zhou,
2008 ; Drake
et al. ,
2013 ).在异质生境中, 气孔各性状并非独立变化, 而是通过性状或特征组合的协变与权衡来共同适应环境.有限资源在植物体内的分配与权衡已通过异速生长理论在多个尺度得到证实(陈国鹏等,
2016 ; 段贝贝等,
2016 ; 杨克彤等,
2020 ), 但以往研究着重于相对宏观的尺度如群落、个体、构件与器官, 而且忽视了系统发育对资源权衡的影响, 对微观尺度如气孔性状的权衡机制关注较少.为此, 本文希望通过研究气孔特征并结合系统发育来探讨微观尺度气孔性状间的生长关系, 以进一步揭示微观尺度的权衡过程, 并丰富植物生活史研究. ...
... 生长型和叶习性是引起植物性状变异的重要因素(Sack & Scoffoni,
2013 ; Mencuccini
et al. ,
2015 ).本研究中气孔性状在4种生长型和2种叶习性中均表现出显著差异.生长型主要造成局域环境内光照、温度和水分的分配不均, 叶习性则决定了植物的碳收益方式和水力结构.林冠上层高大乔木叶的生境为光照相对较强、温度变幅大而水分不足, 灌木或草本则相反.冠层植物叶片往往接受更多辐射, 需要较多的水气蒸腾以维持叶表温度(吴冰洁等,
2015 ), 常分化出小气孔开度、密集的气孔来应对强光照与高蒸腾强度; 林下植物则发育出大而疏散的气孔与较大的气孔开度, 这有助于叶片排出多余的水分(熊慧等,
2014 ; 王海珍等,
2015 ).相比于阔叶植物, 常绿植物叶片小而厚, 其表皮功能与微观结构也相对复杂(朱燕华等,
2011 ; 熊慧等,
2014 ).除气孔开张比外, 本研究中常绿植物的气孔性状普遍大于落叶植物, 这既有别于湿生物种大而疏散的气孔, 也与旱生植物小而密集的气孔有所区别.以往研究表明, 常绿植物的抗旱与抗栓塞能力强于落叶植物, 而且在叶片建成时更倾向于“缓慢投资-收益”策略(潘莹萍和陈亚鹏,
2014 ; 金鹰和王传宽,
2015 ).由于常绿植物叶片建成初期的碳投资高于落叶植物, 需要在叶片发育后期节约并获取更多的碳来偿还成本.因此, 常绿植物趋向于发育大而紧密的气孔以加速碳同化和积累更多的干物质. ...
叶片水力性状研究进展
2
2014
... 系统发育和环境异质是影响植物性状发生分异的动力来源.气孔主要分布于植物叶片下表皮并参与调控碳水摄入与排出, 在其发育过程中迫于环境压力与生存维持, 气孔往往在多个维度存在权衡(Brodribb & Jordan,
2011 ; Brodribb
et al. ,
2013 ; 丁凌子等,
2014 ; Fiorin
et al. ,
2016 ).虽然气孔数量的增加有助于提高光合效率, 但过多的气孔会加速叶内水汽逸散(潘莹萍和陈亚鹏,
2014 ; 王学芳等,
2014 ; 宋慧清等,
2020 ), 而且气孔的建成需要消耗资源, 这就决定了单位面积的气孔数量存在分布上限.气孔密度与大小共同决定了最大可能气孔导度并影响最大同化速率, 在考虑气孔建成实际资源消耗与光合作用水气协调的过程中, 两个相互矛盾的刚需必然达到最优化(Sack & Scoffoni,
2013 ).气孔面积与开度分别代表着气孔发育与运动, 且气孔发育总是先于气孔运动, 尽管大气孔对于开度的调控不及小气孔灵敏, 但可以获得相对较多的碳收益.就此而言, 气孔发育和运动过程中资源分配不均甚至存在矛盾(Xu & Zhou,
2008 ; Drake
et al. ,
2013 ).在异质生境中, 气孔各性状并非独立变化, 而是通过性状或特征组合的协变与权衡来共同适应环境.有限资源在植物体内的分配与权衡已通过异速生长理论在多个尺度得到证实(陈国鹏等,
2016 ; 段贝贝等,
2016 ; 杨克彤等,
2020 ), 但以往研究着重于相对宏观的尺度如群落、个体、构件与器官, 而且忽视了系统发育对资源权衡的影响, 对微观尺度如气孔性状的权衡机制关注较少.为此, 本文希望通过研究气孔特征并结合系统发育来探讨微观尺度气孔性状间的生长关系, 以进一步揭示微观尺度的权衡过程, 并丰富植物生活史研究. ...
... 生长型和叶习性是引起植物性状变异的重要因素(Sack & Scoffoni,
2013 ; Mencuccini
et al. ,
2015 ).本研究中气孔性状在4种生长型和2种叶习性中均表现出显著差异.生长型主要造成局域环境内光照、温度和水分的分配不均, 叶习性则决定了植物的碳收益方式和水力结构.林冠上层高大乔木叶的生境为光照相对较强、温度变幅大而水分不足, 灌木或草本则相反.冠层植物叶片往往接受更多辐射, 需要较多的水气蒸腾以维持叶表温度(吴冰洁等,
2015 ), 常分化出小气孔开度、密集的气孔来应对强光照与高蒸腾强度; 林下植物则发育出大而疏散的气孔与较大的气孔开度, 这有助于叶片排出多余的水分(熊慧等,
2014 ; 王海珍等,
2015 ).相比于阔叶植物, 常绿植物叶片小而厚, 其表皮功能与微观结构也相对复杂(朱燕华等,
2011 ; 熊慧等,
2014 ).除气孔开张比外, 本研究中常绿植物的气孔性状普遍大于落叶植物, 这既有别于湿生物种大而疏散的气孔, 也与旱生植物小而密集的气孔有所区别.以往研究表明, 常绿植物的抗旱与抗栓塞能力强于落叶植物, 而且在叶片建成时更倾向于“缓慢投资-收益”策略(潘莹萍和陈亚鹏,
2014 ; 金鹰和王传宽,
2015 ).由于常绿植物叶片建成初期的碳投资高于落叶植物, 需要在叶片发育后期节约并获取更多的碳来偿还成本.因此, 常绿植物趋向于发育大而紧密的气孔以加速碳同化和积累更多的干物质. ...
白刺叶片气孔特征对人工模拟降雨的响应
3
2014
... 气孔性状(如气孔密度)的种间变异被认为是植物叶片经济谱的一部分(Loranger & Shipley,
2010 ; 王瑞丽等,
2016 ), 它在维持植物稳定碳收益、调节水力结构、吸收速效养分和信息传递等方面具有重要作用(Mencuccini
et al. ,
2015 ; Fiorin
et al. ,
2016 ).因此, 量化气孔性状在异质生境中的变异程度并解析其应对策略有助于进一步阐明植物对环境的适应机制.然而, 多数研究仅侧重于气孔性状对自然环境(海拔、水分和温度等)和控制试验(光强、氮添加和CO
2 浓度等)的响应并得出了一系列结论, 如: 气孔长度与密度之间为负相关关系(任昱等,
2014 ; 姜星星等,
2018 ), 随海拔升高气孔宽度呈减小趋势(王瑞丽等,
2016 ), 土壤湿度增大会显著降低气孔密度(杨再强等,
2015 ; 夏振华等,
2018 ; 张大才等,
2018 ), 高CO
2 浓度会显著降低气孔开张比(贾剑波等,
2019 )等, 但忽视了气孔自身在发育过程中的优先发育性状及其相互间关系的协变与权衡. ...
... 功能各异的群组是稳定占据生态位并差异化利用资源的具象.根据气孔性状对40种植物进行功能群划分, 发现不同群组之间气孔性状差异显著(
图4 ).大气孔植物如山楂、小叶黄杨和费菜分别代表了乡土树种、常绿物种与景天酸代谢植物, 这些植物特殊的适应功能决定了即便在干旱区, 它们依旧能够发育出较大的气孔, 这可能是物种特性决定的(罗丹丹等,
2019 ).植物的小气孔与高气孔密度以及低气孔开度和开张比更具备典型旱生植物的形态, 这可能是对黄土高原干旱生境长期适应的结果(任昱等,
2014 ; 段贝贝等,
2016 ; 胡选萍等,
2016 ).气孔功能群的差异能够高效利用有限空间中的光照与水分等资源, 功能群的组合也维持了垂直梯度上的层级结构, 这有助于维护生境小气候的稳态, 但植物气孔类群分化的驱动因素需进一步研究揭示. ...
... 气孔数量的发育往往先于面积的发育(Xu & Zhou,
2008 ).当气孔密度逐渐增加, 因其空间有限, 气孔面积发育受阻, 气孔开度也随之减小, 较小的气孔开度与较大的气孔密度弥补了碳获取通道的不足.干旱区植物生长的主要限制因子是水分, 较小的气孔面积便于控制气孔的开合, 而较小的气孔开度则有助于减少水分蒸腾, 缓解水分压力(Hetherington & Woodward,
2003 ; Drake
et al. ,
2013 ; Sperry
et al. ,
2017 ).虽然气孔密度增加有助于获取CO
2 物质量和外界信息流, 但气孔密度并非越大越好, 在遵循“单细胞间隔法则”的基础上(宋璐璐等,
2011 ; 任昱等,
2014 ; 王学芳等,
2014 ; 范嘉智等,
2016 ), 高密度的气孔反而造成气孔拥挤与调控失灵, 为灵活调控气孔开合, 减小气孔面积并降低开度更有助于精准调控. ...
白刺叶片气孔特征对人工模拟降雨的响应
3
2014
... 气孔性状(如气孔密度)的种间变异被认为是植物叶片经济谱的一部分(Loranger & Shipley,
2010 ; 王瑞丽等,
2016 ), 它在维持植物稳定碳收益、调节水力结构、吸收速效养分和信息传递等方面具有重要作用(Mencuccini
et al. ,
2015 ; Fiorin
et al. ,
2016 ).因此, 量化气孔性状在异质生境中的变异程度并解析其应对策略有助于进一步阐明植物对环境的适应机制.然而, 多数研究仅侧重于气孔性状对自然环境(海拔、水分和温度等)和控制试验(光强、氮添加和CO
2 浓度等)的响应并得出了一系列结论, 如: 气孔长度与密度之间为负相关关系(任昱等,
2014 ; 姜星星等,
2018 ), 随海拔升高气孔宽度呈减小趋势(王瑞丽等,
2016 ), 土壤湿度增大会显著降低气孔密度(杨再强等,
2015 ; 夏振华等,
2018 ; 张大才等,
2018 ), 高CO
2 浓度会显著降低气孔开张比(贾剑波等,
2019 )等, 但忽视了气孔自身在发育过程中的优先发育性状及其相互间关系的协变与权衡. ...
... 功能各异的群组是稳定占据生态位并差异化利用资源的具象.根据气孔性状对40种植物进行功能群划分, 发现不同群组之间气孔性状差异显著(
图4 ).大气孔植物如山楂、小叶黄杨和费菜分别代表了乡土树种、常绿物种与景天酸代谢植物, 这些植物特殊的适应功能决定了即便在干旱区, 它们依旧能够发育出较大的气孔, 这可能是物种特性决定的(罗丹丹等,
2019 ).植物的小气孔与高气孔密度以及低气孔开度和开张比更具备典型旱生植物的形态, 这可能是对黄土高原干旱生境长期适应的结果(任昱等,
2014 ; 段贝贝等,
2016 ; 胡选萍等,
2016 ).气孔功能群的差异能够高效利用有限空间中的光照与水分等资源, 功能群的组合也维持了垂直梯度上的层级结构, 这有助于维护生境小气候的稳态, 但植物气孔类群分化的驱动因素需进一步研究揭示. ...
... 气孔数量的发育往往先于面积的发育(Xu & Zhou,
2008 ).当气孔密度逐渐增加, 因其空间有限, 气孔面积发育受阻, 气孔开度也随之减小, 较小的气孔开度与较大的气孔密度弥补了碳获取通道的不足.干旱区植物生长的主要限制因子是水分, 较小的气孔面积便于控制气孔的开合, 而较小的气孔开度则有助于减少水分蒸腾, 缓解水分压力(Hetherington & Woodward,
2003 ; Drake
et al. ,
2013 ; Sperry
et al. ,
2017 ).虽然气孔密度增加有助于获取CO
2 物质量和外界信息流, 但气孔密度并非越大越好, 在遵循“单细胞间隔法则”的基础上(宋璐璐等,
2011 ; 任昱等,
2014 ; 王学芳等,
2014 ; 范嘉智等,
2016 ), 高密度的气孔反而造成气孔拥挤与调控失灵, 为灵活调控气孔开合, 减小气孔面积并降低开度更有助于精准调控. ...
Leaf venation: structure, function, development, evolution, ecology and applications in the past, present and future
3
2013
... 系统发育和环境异质是影响植物性状发生分异的动力来源.气孔主要分布于植物叶片下表皮并参与调控碳水摄入与排出, 在其发育过程中迫于环境压力与生存维持, 气孔往往在多个维度存在权衡(Brodribb & Jordan,
2011 ; Brodribb
et al. ,
2013 ; 丁凌子等,
2014 ; Fiorin
et al. ,
2016 ).虽然气孔数量的增加有助于提高光合效率, 但过多的气孔会加速叶内水汽逸散(潘莹萍和陈亚鹏,
2014 ; 王学芳等,
2014 ; 宋慧清等,
2020 ), 而且气孔的建成需要消耗资源, 这就决定了单位面积的气孔数量存在分布上限.气孔密度与大小共同决定了最大可能气孔导度并影响最大同化速率, 在考虑气孔建成实际资源消耗与光合作用水气协调的过程中, 两个相互矛盾的刚需必然达到最优化(Sack & Scoffoni,
2013 ).气孔面积与开度分别代表着气孔发育与运动, 且气孔发育总是先于气孔运动, 尽管大气孔对于开度的调控不及小气孔灵敏, 但可以获得相对较多的碳收益.就此而言, 气孔发育和运动过程中资源分配不均甚至存在矛盾(Xu & Zhou,
2008 ; Drake
et al. ,
2013 ).在异质生境中, 气孔各性状并非独立变化, 而是通过性状或特征组合的协变与权衡来共同适应环境.有限资源在植物体内的分配与权衡已通过异速生长理论在多个尺度得到证实(陈国鹏等,
2016 ; 段贝贝等,
2016 ; 杨克彤等,
2020 ), 但以往研究着重于相对宏观的尺度如群落、个体、构件与器官, 而且忽视了系统发育对资源权衡的影响, 对微观尺度如气孔性状的权衡机制关注较少.为此, 本文希望通过研究气孔特征并结合系统发育来探讨微观尺度气孔性状间的生长关系, 以进一步揭示微观尺度的权衡过程, 并丰富植物生活史研究. ...
... 生长型和叶习性是引起植物性状变异的重要因素(Sack & Scoffoni,
2013 ; Mencuccini
et al. ,
2015 ).本研究中气孔性状在4种生长型和2种叶习性中均表现出显著差异.生长型主要造成局域环境内光照、温度和水分的分配不均, 叶习性则决定了植物的碳收益方式和水力结构.林冠上层高大乔木叶的生境为光照相对较强、温度变幅大而水分不足, 灌木或草本则相反.冠层植物叶片往往接受更多辐射, 需要较多的水气蒸腾以维持叶表温度(吴冰洁等,
2015 ), 常分化出小气孔开度、密集的气孔来应对强光照与高蒸腾强度; 林下植物则发育出大而疏散的气孔与较大的气孔开度, 这有助于叶片排出多余的水分(熊慧等,
2014 ; 王海珍等,
2015 ).相比于阔叶植物, 常绿植物叶片小而厚, 其表皮功能与微观结构也相对复杂(朱燕华等,
2011 ; 熊慧等,
2014 ).除气孔开张比外, 本研究中常绿植物的气孔性状普遍大于落叶植物, 这既有别于湿生物种大而疏散的气孔, 也与旱生植物小而密集的气孔有所区别.以往研究表明, 常绿植物的抗旱与抗栓塞能力强于落叶植物, 而且在叶片建成时更倾向于“缓慢投资-收益”策略(潘莹萍和陈亚鹏,
2014 ; 金鹰和王传宽,
2015 ).由于常绿植物叶片建成初期的碳投资高于落叶植物, 需要在叶片发育后期节约并获取更多的碳来偿还成本.因此, 常绿植物趋向于发育大而紧密的气孔以加速碳同化和积累更多的干物质. ...
... 系统发育决定着植物的亲缘关系并印刻着植物进化的信息, 在研究植物性状特征的过程中是否考虑系统发育将显著地影响研究结果(Ackerly,
2009 ; Zhang
et al. ,
2014 ).本研究中气孔性状间存在显著的相关关系, 在去除系统发育信息后, 这种关系大都发生了转变, 仅气孔长度与宽度依旧维持在稳健的正相关关系, 表明气孔长度与宽度间可能存在关联进化(张亚等,
2014 ; 高林浩等,
2020 ).即不论是否考虑系统发育, 气孔长度与宽度间的正相关关系都是存在的, 暗示这种协同关系可能在古老的植物类群中就已经出现.有研究认为, 植物气孔几何形状和间距总是沿着空间上最优的异速生长关系进化(Sack & Scoffoni,
2013 ; Fiorin
et al. ,
2016 ; Sperry
et al. ,
2017 ), 由于气孔发育属于同步型, 在气孔发育过程中长度与宽度总保持在相对稳定的比率(Franks & Farquhar,
2001 ), 这也从一定程度上解释了去除系统发育后, 气孔长度与宽度依旧保持稳健相关关系的原因.此外, 系统发育信号检测结果表明, 气孔性状的发育信号较弱(
K < 1), 表明气孔性状受系统发育影响较小, 但气孔开度和气孔开张比的发育信号显著, 暗示亲缘关系越近的物种气孔开度与开张比越相似(高林浩等,
2020 ).虽然系统发育会影响气孔性状, 但在长时间适应环境的过程中, 系统发育的影响似乎正在被逐渐削弱. ...
乔木与木质藤本的水力与光合性状的差异: 以热带森林崖豆藤属和买麻藤属为例
1
2020
... 系统发育和环境异质是影响植物性状发生分异的动力来源.气孔主要分布于植物叶片下表皮并参与调控碳水摄入与排出, 在其发育过程中迫于环境压力与生存维持, 气孔往往在多个维度存在权衡(Brodribb & Jordan,
2011 ; Brodribb
et al. ,
2013 ; 丁凌子等,
2014 ; Fiorin
et al. ,
2016 ).虽然气孔数量的增加有助于提高光合效率, 但过多的气孔会加速叶内水汽逸散(潘莹萍和陈亚鹏,
2014 ; 王学芳等,
2014 ; 宋慧清等,
2020 ), 而且气孔的建成需要消耗资源, 这就决定了单位面积的气孔数量存在分布上限.气孔密度与大小共同决定了最大可能气孔导度并影响最大同化速率, 在考虑气孔建成实际资源消耗与光合作用水气协调的过程中, 两个相互矛盾的刚需必然达到最优化(Sack & Scoffoni,
2013 ).气孔面积与开度分别代表着气孔发育与运动, 且气孔发育总是先于气孔运动, 尽管大气孔对于开度的调控不及小气孔灵敏, 但可以获得相对较多的碳收益.就此而言, 气孔发育和运动过程中资源分配不均甚至存在矛盾(Xu & Zhou,
2008 ; Drake
et al. ,
2013 ).在异质生境中, 气孔各性状并非独立变化, 而是通过性状或特征组合的协变与权衡来共同适应环境.有限资源在植物体内的分配与权衡已通过异速生长理论在多个尺度得到证实(陈国鹏等,
2016 ; 段贝贝等,
2016 ; 杨克彤等,
2020 ), 但以往研究着重于相对宏观的尺度如群落、个体、构件与器官, 而且忽视了系统发育对资源权衡的影响, 对微观尺度如气孔性状的权衡机制关注较少.为此, 本文希望通过研究气孔特征并结合系统发育来探讨微观尺度气孔性状间的生长关系, 以进一步揭示微观尺度的权衡过程, 并丰富植物生活史研究. ...
乔木与木质藤本的水力与光合性状的差异: 以热带森林崖豆藤属和买麻藤属为例
1
2020
... 系统发育和环境异质是影响植物性状发生分异的动力来源.气孔主要分布于植物叶片下表皮并参与调控碳水摄入与排出, 在其发育过程中迫于环境压力与生存维持, 气孔往往在多个维度存在权衡(Brodribb & Jordan,
2011 ; Brodribb
et al. ,
2013 ; 丁凌子等,
2014 ; Fiorin
et al. ,
2016 ).虽然气孔数量的增加有助于提高光合效率, 但过多的气孔会加速叶内水汽逸散(潘莹萍和陈亚鹏,
2014 ; 王学芳等,
2014 ; 宋慧清等,
2020 ), 而且气孔的建成需要消耗资源, 这就决定了单位面积的气孔数量存在分布上限.气孔密度与大小共同决定了最大可能气孔导度并影响最大同化速率, 在考虑气孔建成实际资源消耗与光合作用水气协调的过程中, 两个相互矛盾的刚需必然达到最优化(Sack & Scoffoni,
2013 ).气孔面积与开度分别代表着气孔发育与运动, 且气孔发育总是先于气孔运动, 尽管大气孔对于开度的调控不及小气孔灵敏, 但可以获得相对较多的碳收益.就此而言, 气孔发育和运动过程中资源分配不均甚至存在矛盾(Xu & Zhou,
2008 ; Drake
et al. ,
2013 ).在异质生境中, 气孔各性状并非独立变化, 而是通过性状或特征组合的协变与权衡来共同适应环境.有限资源在植物体内的分配与权衡已通过异速生长理论在多个尺度得到证实(陈国鹏等,
2016 ; 段贝贝等,
2016 ; 杨克彤等,
2020 ), 但以往研究着重于相对宏观的尺度如群落、个体、构件与器官, 而且忽视了系统发育对资源权衡的影响, 对微观尺度如气孔性状的权衡机制关注较少.为此, 本文希望通过研究气孔特征并结合系统发育来探讨微观尺度气孔性状间的生长关系, 以进一步揭示微观尺度的权衡过程, 并丰富植物生活史研究. ...
植物叶片性状沿海拔梯度变化研究进展
2
2011
... 气孔作为植物叶表皮的特殊结构, 是植物与外界环境进行物质交换的门户(宋璐璐等,
2011 ; 朱燕华等,
2011 ), 也是植物体内水汽逸散的主要通道(金鹰和王传宽,
2015 ; 范嘉智等,
2016 ).气孔易受环境条件的修饰, 其形态特征是植物进化过程中对外界环境长期适应的结果(王碧霞等,
2010 ; 朱燕华等,
2011 ; 温婧雯等,
2018 ).为适应(或响应)复杂多变的环境, 气孔一方面通过调节开闭来应对短期或局域环境变化, 另一方面则协调大小和密度来适应异质生境(Brodribb & Jordan,
2011 ; Drake
et al. ,
2013 ; 金鹰和王传宽,
2015 ; 高冠龙等,
2018 ).利用气孔特征反映植物对环境变化的响应有助于明晰植物应对水气耦合过程的策略. ...
... 气孔数量的发育往往先于面积的发育(Xu & Zhou,
2008 ).当气孔密度逐渐增加, 因其空间有限, 气孔面积发育受阻, 气孔开度也随之减小, 较小的气孔开度与较大的气孔密度弥补了碳获取通道的不足.干旱区植物生长的主要限制因子是水分, 较小的气孔面积便于控制气孔的开合, 而较小的气孔开度则有助于减少水分蒸腾, 缓解水分压力(Hetherington & Woodward,
2003 ; Drake
et al. ,
2013 ; Sperry
et al. ,
2017 ).虽然气孔密度增加有助于获取CO
2 物质量和外界信息流, 但气孔密度并非越大越好, 在遵循“单细胞间隔法则”的基础上(宋璐璐等,
2011 ; 任昱等,
2014 ; 王学芳等,
2014 ; 范嘉智等,
2016 ), 高密度的气孔反而造成气孔拥挤与调控失灵, 为灵活调控气孔开合, 减小气孔面积并降低开度更有助于精准调控. ...
植物叶片性状沿海拔梯度变化研究进展
2
2011
... 气孔作为植物叶表皮的特殊结构, 是植物与外界环境进行物质交换的门户(宋璐璐等,
2011 ; 朱燕华等,
2011 ), 也是植物体内水汽逸散的主要通道(金鹰和王传宽,
2015 ; 范嘉智等,
2016 ).气孔易受环境条件的修饰, 其形态特征是植物进化过程中对外界环境长期适应的结果(王碧霞等,
2010 ; 朱燕华等,
2011 ; 温婧雯等,
2018 ).为适应(或响应)复杂多变的环境, 气孔一方面通过调节开闭来应对短期或局域环境变化, 另一方面则协调大小和密度来适应异质生境(Brodribb & Jordan,
2011 ; Drake
et al. ,
2013 ; 金鹰和王传宽,
2015 ; 高冠龙等,
2018 ).利用气孔特征反映植物对环境变化的响应有助于明晰植物应对水气耦合过程的策略. ...
... 气孔数量的发育往往先于面积的发育(Xu & Zhou,
2008 ).当气孔密度逐渐增加, 因其空间有限, 气孔面积发育受阻, 气孔开度也随之减小, 较小的气孔开度与较大的气孔密度弥补了碳获取通道的不足.干旱区植物生长的主要限制因子是水分, 较小的气孔面积便于控制气孔的开合, 而较小的气孔开度则有助于减少水分蒸腾, 缓解水分压力(Hetherington & Woodward,
2003 ; Drake
et al. ,
2013 ; Sperry
et al. ,
2017 ).虽然气孔密度增加有助于获取CO
2 物质量和外界信息流, 但气孔密度并非越大越好, 在遵循“单细胞间隔法则”的基础上(宋璐璐等,
2011 ; 任昱等,
2014 ; 王学芳等,
2014 ; 范嘉智等,
2016 ), 高密度的气孔反而造成气孔拥挤与调控失灵, 为灵活调控气孔开合, 减小气孔面积并降低开度更有助于精准调控. ...
Predicting stomatal responses to the environment from the optimization of photosynthetic gain and hydraulic cost
3
2017
... 系统发育决定着植物的亲缘关系并印刻着植物进化的信息, 在研究植物性状特征的过程中是否考虑系统发育将显著地影响研究结果(Ackerly,
2009 ; Zhang
et al. ,
2014 ).本研究中气孔性状间存在显著的相关关系, 在去除系统发育信息后, 这种关系大都发生了转变, 仅气孔长度与宽度依旧维持在稳健的正相关关系, 表明气孔长度与宽度间可能存在关联进化(张亚等,
2014 ; 高林浩等,
2020 ).即不论是否考虑系统发育, 气孔长度与宽度间的正相关关系都是存在的, 暗示这种协同关系可能在古老的植物类群中就已经出现.有研究认为, 植物气孔几何形状和间距总是沿着空间上最优的异速生长关系进化(Sack & Scoffoni,
2013 ; Fiorin
et al. ,
2016 ; Sperry
et al. ,
2017 ), 由于气孔发育属于同步型, 在气孔发育过程中长度与宽度总保持在相对稳定的比率(Franks & Farquhar,
2001 ), 这也从一定程度上解释了去除系统发育后, 气孔长度与宽度依旧保持稳健相关关系的原因.此外, 系统发育信号检测结果表明, 气孔性状的发育信号较弱(
K < 1), 表明气孔性状受系统发育影响较小, 但气孔开度和气孔开张比的发育信号显著, 暗示亲缘关系越近的物种气孔开度与开张比越相似(高林浩等,
2020 ).虽然系统发育会影响气孔性状, 但在长时间适应环境的过程中, 系统发育的影响似乎正在被逐渐削弱. ...
... 气孔面积与开度的关系表明, 随着气孔面积逐渐增大, 气孔开度也逐渐增大, 但气孔开度的扩大速率不及面积的增加速率.有研究认为, 气孔越大对气孔孔径调控的能力越弱, 在遭遇干旱胁迫时所丢失的水分就越多(Drake
et al. ,
2013 ; Sperry
et al. ,
2017 ), 虽然较大的气孔开度可以交换更多的碳, 但这种交换存在风险.气孔面积增大时, 保持CO
2 摄入量不变的情况下, 减小气孔开度依旧可以达到等效水平.但是, 大气孔对开度的调节缺乏灵敏性并对应着较低的气体交换速率, 随着气孔开度逐渐增大, 其进一步开张的阻力也越大、耗费能量越多, 它们的慢动力学特性也使大气孔植物在遭遇干旱等环境胁迫时更容易发生功能紊乱(Hetherington & Woodward,
2003 ; 范嘉智等,
2016 ; 胡选萍等,
2016 ). ...
... 气孔数量的发育往往先于面积的发育(Xu & Zhou,
2008 ).当气孔密度逐渐增加, 因其空间有限, 气孔面积发育受阻, 气孔开度也随之减小, 较小的气孔开度与较大的气孔密度弥补了碳获取通道的不足.干旱区植物生长的主要限制因子是水分, 较小的气孔面积便于控制气孔的开合, 而较小的气孔开度则有助于减少水分蒸腾, 缓解水分压力(Hetherington & Woodward,
2003 ; Drake
et al. ,
2013 ; Sperry
et al. ,
2017 ).虽然气孔密度增加有助于获取CO
2 物质量和外界信息流, 但气孔密度并非越大越好, 在遵循“单细胞间隔法则”的基础上(宋璐璐等,
2011 ; 任昱等,
2014 ; 王学芳等,
2014 ; 范嘉智等,
2016 ), 高密度的气孔反而造成气孔拥挤与调控失灵, 为灵活调控气孔开合, 减小气孔面积并降低开度更有助于精准调控. ...
叶片气孔分布及生理特征对环境胁迫的响应
1
2010
... 气孔作为植物叶表皮的特殊结构, 是植物与外界环境进行物质交换的门户(宋璐璐等,
2011 ; 朱燕华等,
2011 ), 也是植物体内水汽逸散的主要通道(金鹰和王传宽,
2015 ; 范嘉智等,
2016 ).气孔易受环境条件的修饰, 其形态特征是植物进化过程中对外界环境长期适应的结果(王碧霞等,
2010 ; 朱燕华等,
2011 ; 温婧雯等,
2018 ).为适应(或响应)复杂多变的环境, 气孔一方面通过调节开闭来应对短期或局域环境变化, 另一方面则协调大小和密度来适应异质生境(Brodribb & Jordan,
2011 ; Drake
et al. ,
2013 ; 金鹰和王传宽,
2015 ; 高冠龙等,
2018 ).利用气孔特征反映植物对环境变化的响应有助于明晰植物应对水气耦合过程的策略. ...
叶片气孔分布及生理特征对环境胁迫的响应
1
2010
... 气孔作为植物叶表皮的特殊结构, 是植物与外界环境进行物质交换的门户(宋璐璐等,
2011 ; 朱燕华等,
2011 ), 也是植物体内水汽逸散的主要通道(金鹰和王传宽,
2015 ; 范嘉智等,
2016 ).气孔易受环境条件的修饰, 其形态特征是植物进化过程中对外界环境长期适应的结果(王碧霞等,
2010 ; 朱燕华等,
2011 ; 温婧雯等,
2018 ).为适应(或响应)复杂多变的环境, 气孔一方面通过调节开闭来应对短期或局域环境变化, 另一方面则协调大小和密度来适应异质生境(Brodribb & Jordan,
2011 ; Drake
et al. ,
2013 ; 金鹰和王传宽,
2015 ; 高冠龙等,
2018 ).利用气孔特征反映植物对环境变化的响应有助于明晰植物应对水气耦合过程的策略. ...
不同温度下灰胡杨叶片气孔导度对光强响应的模型分析
1
2015
... 生长型和叶习性是引起植物性状变异的重要因素(Sack & Scoffoni,
2013 ; Mencuccini
et al. ,
2015 ).本研究中气孔性状在4种生长型和2种叶习性中均表现出显著差异.生长型主要造成局域环境内光照、温度和水分的分配不均, 叶习性则决定了植物的碳收益方式和水力结构.林冠上层高大乔木叶的生境为光照相对较强、温度变幅大而水分不足, 灌木或草本则相反.冠层植物叶片往往接受更多辐射, 需要较多的水气蒸腾以维持叶表温度(吴冰洁等,
2015 ), 常分化出小气孔开度、密集的气孔来应对强光照与高蒸腾强度; 林下植物则发育出大而疏散的气孔与较大的气孔开度, 这有助于叶片排出多余的水分(熊慧等,
2014 ; 王海珍等,
2015 ).相比于阔叶植物, 常绿植物叶片小而厚, 其表皮功能与微观结构也相对复杂(朱燕华等,
2011 ; 熊慧等,
2014 ).除气孔开张比外, 本研究中常绿植物的气孔性状普遍大于落叶植物, 这既有别于湿生物种大而疏散的气孔, 也与旱生植物小而密集的气孔有所区别.以往研究表明, 常绿植物的抗旱与抗栓塞能力强于落叶植物, 而且在叶片建成时更倾向于“缓慢投资-收益”策略(潘莹萍和陈亚鹏,
2014 ; 金鹰和王传宽,
2015 ).由于常绿植物叶片建成初期的碳投资高于落叶植物, 需要在叶片发育后期节约并获取更多的碳来偿还成本.因此, 常绿植物趋向于发育大而紧密的气孔以加速碳同化和积累更多的干物质. ...
不同温度下灰胡杨叶片气孔导度对光强响应的模型分析
1
2015
... 生长型和叶习性是引起植物性状变异的重要因素(Sack & Scoffoni,
2013 ; Mencuccini
et al. ,
2015 ).本研究中气孔性状在4种生长型和2种叶习性中均表现出显著差异.生长型主要造成局域环境内光照、温度和水分的分配不均, 叶习性则决定了植物的碳收益方式和水力结构.林冠上层高大乔木叶的生境为光照相对较强、温度变幅大而水分不足, 灌木或草本则相反.冠层植物叶片往往接受更多辐射, 需要较多的水气蒸腾以维持叶表温度(吴冰洁等,
2015 ), 常分化出小气孔开度、密集的气孔来应对强光照与高蒸腾强度; 林下植物则发育出大而疏散的气孔与较大的气孔开度, 这有助于叶片排出多余的水分(熊慧等,
2014 ; 王海珍等,
2015 ).相比于阔叶植物, 常绿植物叶片小而厚, 其表皮功能与微观结构也相对复杂(朱燕华等,
2011 ; 熊慧等,
2014 ).除气孔开张比外, 本研究中常绿植物的气孔性状普遍大于落叶植物, 这既有别于湿生物种大而疏散的气孔, 也与旱生植物小而密集的气孔有所区别.以往研究表明, 常绿植物的抗旱与抗栓塞能力强于落叶植物, 而且在叶片建成时更倾向于“缓慢投资-收益”策略(潘莹萍和陈亚鹏,
2014 ; 金鹰和王传宽,
2015 ).由于常绿植物叶片建成初期的碳投资高于落叶植物, 需要在叶片发育后期节约并获取更多的碳来偿还成本.因此, 常绿植物趋向于发育大而紧密的气孔以加速碳同化和积累更多的干物质. ...
气孔特征与叶片功能性状之间关联性沿海拔梯度的变化规律——以长白山为例
2
2016
... 气孔性状(如气孔密度)的种间变异被认为是植物叶片经济谱的一部分(Loranger & Shipley,
2010 ; 王瑞丽等,
2016 ), 它在维持植物稳定碳收益、调节水力结构、吸收速效养分和信息传递等方面具有重要作用(Mencuccini
et al. ,
2015 ; Fiorin
et al. ,
2016 ).因此, 量化气孔性状在异质生境中的变异程度并解析其应对策略有助于进一步阐明植物对环境的适应机制.然而, 多数研究仅侧重于气孔性状对自然环境(海拔、水分和温度等)和控制试验(光强、氮添加和CO
2 浓度等)的响应并得出了一系列结论, 如: 气孔长度与密度之间为负相关关系(任昱等,
2014 ; 姜星星等,
2018 ), 随海拔升高气孔宽度呈减小趋势(王瑞丽等,
2016 ), 土壤湿度增大会显著降低气孔密度(杨再强等,
2015 ; 夏振华等,
2018 ; 张大才等,
2018 ), 高CO
2 浓度会显著降低气孔开张比(贾剑波等,
2019 )等, 但忽视了气孔自身在发育过程中的优先发育性状及其相互间关系的协变与权衡. ...
... ), 随海拔升高气孔宽度呈减小趋势(王瑞丽等,
2016 ), 土壤湿度增大会显著降低气孔密度(杨再强等,
2015 ; 夏振华等,
2018 ; 张大才等,
2018 ), 高CO
2 浓度会显著降低气孔开张比(贾剑波等,
2019 )等, 但忽视了气孔自身在发育过程中的优先发育性状及其相互间关系的协变与权衡. ...
气孔特征与叶片功能性状之间关联性沿海拔梯度的变化规律——以长白山为例
2
2016
... 气孔性状(如气孔密度)的种间变异被认为是植物叶片经济谱的一部分(Loranger & Shipley,
2010 ; 王瑞丽等,
2016 ), 它在维持植物稳定碳收益、调节水力结构、吸收速效养分和信息传递等方面具有重要作用(Mencuccini
et al. ,
2015 ; Fiorin
et al. ,
2016 ).因此, 量化气孔性状在异质生境中的变异程度并解析其应对策略有助于进一步阐明植物对环境的适应机制.然而, 多数研究仅侧重于气孔性状对自然环境(海拔、水分和温度等)和控制试验(光强、氮添加和CO
2 浓度等)的响应并得出了一系列结论, 如: 气孔长度与密度之间为负相关关系(任昱等,
2014 ; 姜星星等,
2018 ), 随海拔升高气孔宽度呈减小趋势(王瑞丽等,
2016 ), 土壤湿度增大会显著降低气孔密度(杨再强等,
2015 ; 夏振华等,
2018 ; 张大才等,
2018 ), 高CO
2 浓度会显著降低气孔开张比(贾剑波等,
2019 )等, 但忽视了气孔自身在发育过程中的优先发育性状及其相互间关系的协变与权衡. ...
... ), 随海拔升高气孔宽度呈减小趋势(王瑞丽等,
2016 ), 土壤湿度增大会显著降低气孔密度(杨再强等,
2015 ; 夏振华等,
2018 ; 张大才等,
2018 ), 高CO
2 浓度会显著降低气孔开张比(贾剑波等,
2019 )等, 但忽视了气孔自身在发育过程中的优先发育性状及其相互间关系的协变与权衡. ...
太白山栎属树种气孔特征沿海拔梯度的变化规律
1
2018
... 气孔作为植物叶表皮的特殊结构, 是植物与外界环境进行物质交换的门户(宋璐璐等,
2011 ; 朱燕华等,
2011 ), 也是植物体内水汽逸散的主要通道(金鹰和王传宽,
2015 ; 范嘉智等,
2016 ).气孔易受环境条件的修饰, 其形态特征是植物进化过程中对外界环境长期适应的结果(王碧霞等,
2010 ; 朱燕华等,
2011 ; 温婧雯等,
2018 ).为适应(或响应)复杂多变的环境, 气孔一方面通过调节开闭来应对短期或局域环境变化, 另一方面则协调大小和密度来适应异质生境(Brodribb & Jordan,
2011 ; Drake
et al. ,
2013 ; 金鹰和王传宽,
2015 ; 高冠龙等,
2018 ).利用气孔特征反映植物对环境变化的响应有助于明晰植物应对水气耦合过程的策略. ...
太白山栎属树种气孔特征沿海拔梯度的变化规律
1
2018
... 气孔作为植物叶表皮的特殊结构, 是植物与外界环境进行物质交换的门户(宋璐璐等,
2011 ; 朱燕华等,
2011 ), 也是植物体内水汽逸散的主要通道(金鹰和王传宽,
2015 ; 范嘉智等,
2016 ).气孔易受环境条件的修饰, 其形态特征是植物进化过程中对外界环境长期适应的结果(王碧霞等,
2010 ; 朱燕华等,
2011 ; 温婧雯等,
2018 ).为适应(或响应)复杂多变的环境, 气孔一方面通过调节开闭来应对短期或局域环境变化, 另一方面则协调大小和密度来适应异质生境(Brodribb & Jordan,
2011 ; Drake
et al. ,
2013 ; 金鹰和王传宽,
2015 ; 高冠龙等,
2018 ).利用气孔特征反映植物对环境变化的响应有助于明晰植物应对水气耦合过程的策略. ...
叶片生长进程中气孔发育对叶温调节的影响
1
2015
... 生长型和叶习性是引起植物性状变异的重要因素(Sack & Scoffoni,
2013 ; Mencuccini
et al. ,
2015 ).本研究中气孔性状在4种生长型和2种叶习性中均表现出显著差异.生长型主要造成局域环境内光照、温度和水分的分配不均, 叶习性则决定了植物的碳收益方式和水力结构.林冠上层高大乔木叶的生境为光照相对较强、温度变幅大而水分不足, 灌木或草本则相反.冠层植物叶片往往接受更多辐射, 需要较多的水气蒸腾以维持叶表温度(吴冰洁等,
2015 ), 常分化出小气孔开度、密集的气孔来应对强光照与高蒸腾强度; 林下植物则发育出大而疏散的气孔与较大的气孔开度, 这有助于叶片排出多余的水分(熊慧等,
2014 ; 王海珍等,
2015 ).相比于阔叶植物, 常绿植物叶片小而厚, 其表皮功能与微观结构也相对复杂(朱燕华等,
2011 ; 熊慧等,
2014 ).除气孔开张比外, 本研究中常绿植物的气孔性状普遍大于落叶植物, 这既有别于湿生物种大而疏散的气孔, 也与旱生植物小而密集的气孔有所区别.以往研究表明, 常绿植物的抗旱与抗栓塞能力强于落叶植物, 而且在叶片建成时更倾向于“缓慢投资-收益”策略(潘莹萍和陈亚鹏,
2014 ; 金鹰和王传宽,
2015 ).由于常绿植物叶片建成初期的碳投资高于落叶植物, 需要在叶片发育后期节约并获取更多的碳来偿还成本.因此, 常绿植物趋向于发育大而紧密的气孔以加速碳同化和积累更多的干物质. ...
叶片生长进程中气孔发育对叶温调节的影响
1
2015
... 生长型和叶习性是引起植物性状变异的重要因素(Sack & Scoffoni,
2013 ; Mencuccini
et al. ,
2015 ).本研究中气孔性状在4种生长型和2种叶习性中均表现出显著差异.生长型主要造成局域环境内光照、温度和水分的分配不均, 叶习性则决定了植物的碳收益方式和水力结构.林冠上层高大乔木叶的生境为光照相对较强、温度变幅大而水分不足, 灌木或草本则相反.冠层植物叶片往往接受更多辐射, 需要较多的水气蒸腾以维持叶表温度(吴冰洁等,
2015 ), 常分化出小气孔开度、密集的气孔来应对强光照与高蒸腾强度; 林下植物则发育出大而疏散的气孔与较大的气孔开度, 这有助于叶片排出多余的水分(熊慧等,
2014 ; 王海珍等,
2015 ).相比于阔叶植物, 常绿植物叶片小而厚, 其表皮功能与微观结构也相对复杂(朱燕华等,
2011 ; 熊慧等,
2014 ).除气孔开张比外, 本研究中常绿植物的气孔性状普遍大于落叶植物, 这既有别于湿生物种大而疏散的气孔, 也与旱生植物小而密集的气孔有所区别.以往研究表明, 常绿植物的抗旱与抗栓塞能力强于落叶植物, 而且在叶片建成时更倾向于“缓慢投资-收益”策略(潘莹萍和陈亚鹏,
2014 ; 金鹰和王传宽,
2015 ).由于常绿植物叶片建成初期的碳投资高于落叶植物, 需要在叶片发育后期节约并获取更多的碳来偿还成本.因此, 常绿植物趋向于发育大而紧密的气孔以加速碳同化和积累更多的干物质. ...
祁连山3种被子植物叶特征随海拔变化及其内陆高海拔模式
2
2014
... 系统发育和环境异质是影响植物性状发生分异的动力来源.气孔主要分布于植物叶片下表皮并参与调控碳水摄入与排出, 在其发育过程中迫于环境压力与生存维持, 气孔往往在多个维度存在权衡(Brodribb & Jordan,
2011 ; Brodribb
et al. ,
2013 ; 丁凌子等,
2014 ; Fiorin
et al. ,
2016 ).虽然气孔数量的增加有助于提高光合效率, 但过多的气孔会加速叶内水汽逸散(潘莹萍和陈亚鹏,
2014 ; 王学芳等,
2014 ; 宋慧清等,
2020 ), 而且气孔的建成需要消耗资源, 这就决定了单位面积的气孔数量存在分布上限.气孔密度与大小共同决定了最大可能气孔导度并影响最大同化速率, 在考虑气孔建成实际资源消耗与光合作用水气协调的过程中, 两个相互矛盾的刚需必然达到最优化(Sack & Scoffoni,
2013 ).气孔面积与开度分别代表着气孔发育与运动, 且气孔发育总是先于气孔运动, 尽管大气孔对于开度的调控不及小气孔灵敏, 但可以获得相对较多的碳收益.就此而言, 气孔发育和运动过程中资源分配不均甚至存在矛盾(Xu & Zhou,
2008 ; Drake
et al. ,
2013 ).在异质生境中, 气孔各性状并非独立变化, 而是通过性状或特征组合的协变与权衡来共同适应环境.有限资源在植物体内的分配与权衡已通过异速生长理论在多个尺度得到证实(陈国鹏等,
2016 ; 段贝贝等,
2016 ; 杨克彤等,
2020 ), 但以往研究着重于相对宏观的尺度如群落、个体、构件与器官, 而且忽视了系统发育对资源权衡的影响, 对微观尺度如气孔性状的权衡机制关注较少.为此, 本文希望通过研究气孔特征并结合系统发育来探讨微观尺度气孔性状间的生长关系, 以进一步揭示微观尺度的权衡过程, 并丰富植物生活史研究. ...
... 气孔数量的发育往往先于面积的发育(Xu & Zhou,
2008 ).当气孔密度逐渐增加, 因其空间有限, 气孔面积发育受阻, 气孔开度也随之减小, 较小的气孔开度与较大的气孔密度弥补了碳获取通道的不足.干旱区植物生长的主要限制因子是水分, 较小的气孔面积便于控制气孔的开合, 而较小的气孔开度则有助于减少水分蒸腾, 缓解水分压力(Hetherington & Woodward,
2003 ; Drake
et al. ,
2013 ; Sperry
et al. ,
2017 ).虽然气孔密度增加有助于获取CO
2 物质量和外界信息流, 但气孔密度并非越大越好, 在遵循“单细胞间隔法则”的基础上(宋璐璐等,
2011 ; 任昱等,
2014 ; 王学芳等,
2014 ; 范嘉智等,
2016 ), 高密度的气孔反而造成气孔拥挤与调控失灵, 为灵活调控气孔开合, 减小气孔面积并降低开度更有助于精准调控. ...
祁连山3种被子植物叶特征随海拔变化及其内陆高海拔模式
2
2014
... 系统发育和环境异质是影响植物性状发生分异的动力来源.气孔主要分布于植物叶片下表皮并参与调控碳水摄入与排出, 在其发育过程中迫于环境压力与生存维持, 气孔往往在多个维度存在权衡(Brodribb & Jordan,
2011 ; Brodribb
et al. ,
2013 ; 丁凌子等,
2014 ; Fiorin
et al. ,
2016 ).虽然气孔数量的增加有助于提高光合效率, 但过多的气孔会加速叶内水汽逸散(潘莹萍和陈亚鹏,
2014 ; 王学芳等,
2014 ; 宋慧清等,
2020 ), 而且气孔的建成需要消耗资源, 这就决定了单位面积的气孔数量存在分布上限.气孔密度与大小共同决定了最大可能气孔导度并影响最大同化速率, 在考虑气孔建成实际资源消耗与光合作用水气协调的过程中, 两个相互矛盾的刚需必然达到最优化(Sack & Scoffoni,
2013 ).气孔面积与开度分别代表着气孔发育与运动, 且气孔发育总是先于气孔运动, 尽管大气孔对于开度的调控不及小气孔灵敏, 但可以获得相对较多的碳收益.就此而言, 气孔发育和运动过程中资源分配不均甚至存在矛盾(Xu & Zhou,
2008 ; Drake
et al. ,
2013 ).在异质生境中, 气孔各性状并非独立变化, 而是通过性状或特征组合的协变与权衡来共同适应环境.有限资源在植物体内的分配与权衡已通过异速生长理论在多个尺度得到证实(陈国鹏等,
2016 ; 段贝贝等,
2016 ; 杨克彤等,
2020 ), 但以往研究着重于相对宏观的尺度如群落、个体、构件与器官, 而且忽视了系统发育对资源权衡的影响, 对微观尺度如气孔性状的权衡机制关注较少.为此, 本文希望通过研究气孔特征并结合系统发育来探讨微观尺度气孔性状间的生长关系, 以进一步揭示微观尺度的权衡过程, 并丰富植物生活史研究. ...
... 气孔数量的发育往往先于面积的发育(Xu & Zhou,
2008 ).当气孔密度逐渐增加, 因其空间有限, 气孔面积发育受阻, 气孔开度也随之减小, 较小的气孔开度与较大的气孔密度弥补了碳获取通道的不足.干旱区植物生长的主要限制因子是水分, 较小的气孔面积便于控制气孔的开合, 而较小的气孔开度则有助于减少水分蒸腾, 缓解水分压力(Hetherington & Woodward,
2003 ; Drake
et al. ,
2013 ; Sperry
et al. ,
2017 ).虽然气孔密度增加有助于获取CO
2 物质量和外界信息流, 但气孔密度并非越大越好, 在遵循“单细胞间隔法则”的基础上(宋璐璐等,
2011 ; 任昱等,
2014 ; 王学芳等,
2014 ; 范嘉智等,
2016 ), 高密度的气孔反而造成气孔拥挤与调控失灵, 为灵活调控气孔开合, 减小气孔面积并降低开度更有助于精准调控. ...
干旱胁迫环境下的胡杨叶片气孔变化
1
2018
... 气孔性状(如气孔密度)的种间变异被认为是植物叶片经济谱的一部分(Loranger & Shipley,
2010 ; 王瑞丽等,
2016 ), 它在维持植物稳定碳收益、调节水力结构、吸收速效养分和信息传递等方面具有重要作用(Mencuccini
et al. ,
2015 ; Fiorin
et al. ,
2016 ).因此, 量化气孔性状在异质生境中的变异程度并解析其应对策略有助于进一步阐明植物对环境的适应机制.然而, 多数研究仅侧重于气孔性状对自然环境(海拔、水分和温度等)和控制试验(光强、氮添加和CO
2 浓度等)的响应并得出了一系列结论, 如: 气孔长度与密度之间为负相关关系(任昱等,
2014 ; 姜星星等,
2018 ), 随海拔升高气孔宽度呈减小趋势(王瑞丽等,
2016 ), 土壤湿度增大会显著降低气孔密度(杨再强等,
2015 ; 夏振华等,
2018 ; 张大才等,
2018 ), 高CO
2 浓度会显著降低气孔开张比(贾剑波等,
2019 )等, 但忽视了气孔自身在发育过程中的优先发育性状及其相互间关系的协变与权衡. ...
干旱胁迫环境下的胡杨叶片气孔变化
1
2018
... 气孔性状(如气孔密度)的种间变异被认为是植物叶片经济谱的一部分(Loranger & Shipley,
2010 ; 王瑞丽等,
2016 ), 它在维持植物稳定碳收益、调节水力结构、吸收速效养分和信息传递等方面具有重要作用(Mencuccini
et al. ,
2015 ; Fiorin
et al. ,
2016 ).因此, 量化气孔性状在异质生境中的变异程度并解析其应对策略有助于进一步阐明植物对环境的适应机制.然而, 多数研究仅侧重于气孔性状对自然环境(海拔、水分和温度等)和控制试验(光强、氮添加和CO
2 浓度等)的响应并得出了一系列结论, 如: 气孔长度与密度之间为负相关关系(任昱等,
2014 ; 姜星星等,
2018 ), 随海拔升高气孔宽度呈减小趋势(王瑞丽等,
2016 ), 土壤湿度增大会显著降低气孔密度(杨再强等,
2015 ; 夏振华等,
2018 ; 张大才等,
2018 ), 高CO
2 浓度会显著降低气孔开张比(贾剑波等,
2019 )等, 但忽视了气孔自身在发育过程中的优先发育性状及其相互间关系的协变与权衡. ...
不同生境条件下蕨类和被子植物的气孔形态特征及其对光强变化的响应
2
2014
... 生长型和叶习性是引起植物性状变异的重要因素(Sack & Scoffoni,
2013 ; Mencuccini
et al. ,
2015 ).本研究中气孔性状在4种生长型和2种叶习性中均表现出显著差异.生长型主要造成局域环境内光照、温度和水分的分配不均, 叶习性则决定了植物的碳收益方式和水力结构.林冠上层高大乔木叶的生境为光照相对较强、温度变幅大而水分不足, 灌木或草本则相反.冠层植物叶片往往接受更多辐射, 需要较多的水气蒸腾以维持叶表温度(吴冰洁等,
2015 ), 常分化出小气孔开度、密集的气孔来应对强光照与高蒸腾强度; 林下植物则发育出大而疏散的气孔与较大的气孔开度, 这有助于叶片排出多余的水分(熊慧等,
2014 ; 王海珍等,
2015 ).相比于阔叶植物, 常绿植物叶片小而厚, 其表皮功能与微观结构也相对复杂(朱燕华等,
2011 ; 熊慧等,
2014 ).除气孔开张比外, 本研究中常绿植物的气孔性状普遍大于落叶植物, 这既有别于湿生物种大而疏散的气孔, 也与旱生植物小而密集的气孔有所区别.以往研究表明, 常绿植物的抗旱与抗栓塞能力强于落叶植物, 而且在叶片建成时更倾向于“缓慢投资-收益”策略(潘莹萍和陈亚鹏,
2014 ; 金鹰和王传宽,
2015 ).由于常绿植物叶片建成初期的碳投资高于落叶植物, 需要在叶片发育后期节约并获取更多的碳来偿还成本.因此, 常绿植物趋向于发育大而紧密的气孔以加速碳同化和积累更多的干物质. ...
... ; 熊慧等,
2014 ).除气孔开张比外, 本研究中常绿植物的气孔性状普遍大于落叶植物, 这既有别于湿生物种大而疏散的气孔, 也与旱生植物小而密集的气孔有所区别.以往研究表明, 常绿植物的抗旱与抗栓塞能力强于落叶植物, 而且在叶片建成时更倾向于“缓慢投资-收益”策略(潘莹萍和陈亚鹏,
2014 ; 金鹰和王传宽,
2015 ).由于常绿植物叶片建成初期的碳投资高于落叶植物, 需要在叶片发育后期节约并获取更多的碳来偿还成本.因此, 常绿植物趋向于发育大而紧密的气孔以加速碳同化和积累更多的干物质. ...
不同生境条件下蕨类和被子植物的气孔形态特征及其对光强变化的响应
2
2014
... 生长型和叶习性是引起植物性状变异的重要因素(Sack & Scoffoni,
2013 ; Mencuccini
et al. ,
2015 ).本研究中气孔性状在4种生长型和2种叶习性中均表现出显著差异.生长型主要造成局域环境内光照、温度和水分的分配不均, 叶习性则决定了植物的碳收益方式和水力结构.林冠上层高大乔木叶的生境为光照相对较强、温度变幅大而水分不足, 灌木或草本则相反.冠层植物叶片往往接受更多辐射, 需要较多的水气蒸腾以维持叶表温度(吴冰洁等,
2015 ), 常分化出小气孔开度、密集的气孔来应对强光照与高蒸腾强度; 林下植物则发育出大而疏散的气孔与较大的气孔开度, 这有助于叶片排出多余的水分(熊慧等,
2014 ; 王海珍等,
2015 ).相比于阔叶植物, 常绿植物叶片小而厚, 其表皮功能与微观结构也相对复杂(朱燕华等,
2011 ; 熊慧等,
2014 ).除气孔开张比外, 本研究中常绿植物的气孔性状普遍大于落叶植物, 这既有别于湿生物种大而疏散的气孔, 也与旱生植物小而密集的气孔有所区别.以往研究表明, 常绿植物的抗旱与抗栓塞能力强于落叶植物, 而且在叶片建成时更倾向于“缓慢投资-收益”策略(潘莹萍和陈亚鹏,
2014 ; 金鹰和王传宽,
2015 ).由于常绿植物叶片建成初期的碳投资高于落叶植物, 需要在叶片发育后期节约并获取更多的碳来偿还成本.因此, 常绿植物趋向于发育大而紧密的气孔以加速碳同化和积累更多的干物质. ...
... ; 熊慧等,
2014 ).除气孔开张比外, 本研究中常绿植物的气孔性状普遍大于落叶植物, 这既有别于湿生物种大而疏散的气孔, 也与旱生植物小而密集的气孔有所区别.以往研究表明, 常绿植物的抗旱与抗栓塞能力强于落叶植物, 而且在叶片建成时更倾向于“缓慢投资-收益”策略(潘莹萍和陈亚鹏,
2014 ; 金鹰和王传宽,
2015 ).由于常绿植物叶片建成初期的碳投资高于落叶植物, 需要在叶片发育后期节约并获取更多的碳来偿还成本.因此, 常绿植物趋向于发育大而紧密的气孔以加速碳同化和积累更多的干物质. ...
Responses of leaf stomatal density to water status and its relationship with photosynthesis in a grass
2
2008
... 系统发育和环境异质是影响植物性状发生分异的动力来源.气孔主要分布于植物叶片下表皮并参与调控碳水摄入与排出, 在其发育过程中迫于环境压力与生存维持, 气孔往往在多个维度存在权衡(Brodribb & Jordan,
2011 ; Brodribb
et al. ,
2013 ; 丁凌子等,
2014 ; Fiorin
et al. ,
2016 ).虽然气孔数量的增加有助于提高光合效率, 但过多的气孔会加速叶内水汽逸散(潘莹萍和陈亚鹏,
2014 ; 王学芳等,
2014 ; 宋慧清等,
2020 ), 而且气孔的建成需要消耗资源, 这就决定了单位面积的气孔数量存在分布上限.气孔密度与大小共同决定了最大可能气孔导度并影响最大同化速率, 在考虑气孔建成实际资源消耗与光合作用水气协调的过程中, 两个相互矛盾的刚需必然达到最优化(Sack & Scoffoni,
2013 ).气孔面积与开度分别代表着气孔发育与运动, 且气孔发育总是先于气孔运动, 尽管大气孔对于开度的调控不及小气孔灵敏, 但可以获得相对较多的碳收益.就此而言, 气孔发育和运动过程中资源分配不均甚至存在矛盾(Xu & Zhou,
2008 ; Drake
et al. ,
2013 ).在异质生境中, 气孔各性状并非独立变化, 而是通过性状或特征组合的协变与权衡来共同适应环境.有限资源在植物体内的分配与权衡已通过异速生长理论在多个尺度得到证实(陈国鹏等,
2016 ; 段贝贝等,
2016 ; 杨克彤等,
2020 ), 但以往研究着重于相对宏观的尺度如群落、个体、构件与器官, 而且忽视了系统发育对资源权衡的影响, 对微观尺度如气孔性状的权衡机制关注较少.为此, 本文希望通过研究气孔特征并结合系统发育来探讨微观尺度气孔性状间的生长关系, 以进一步揭示微观尺度的权衡过程, 并丰富植物生活史研究. ...
... 气孔数量的发育往往先于面积的发育(Xu & Zhou,
2008 ).当气孔密度逐渐增加, 因其空间有限, 气孔面积发育受阻, 气孔开度也随之减小, 较小的气孔开度与较大的气孔密度弥补了碳获取通道的不足.干旱区植物生长的主要限制因子是水分, 较小的气孔面积便于控制气孔的开合, 而较小的气孔开度则有助于减少水分蒸腾, 缓解水分压力(Hetherington & Woodward,
2003 ; Drake
et al. ,
2013 ; Sperry
et al. ,
2017 ).虽然气孔密度增加有助于获取CO
2 物质量和外界信息流, 但气孔密度并非越大越好, 在遵循“单细胞间隔法则”的基础上(宋璐璐等,
2011 ; 任昱等,
2014 ; 王学芳等,
2014 ; 范嘉智等,
2016 ), 高密度的气孔反而造成气孔拥挤与调控失灵, 为灵活调控气孔开合, 减小气孔面积并降低开度更有助于精准调控. ...
兰州市典型绿化树种叶性状间的权衡关系
2
2020
... 系统发育和环境异质是影响植物性状发生分异的动力来源.气孔主要分布于植物叶片下表皮并参与调控碳水摄入与排出, 在其发育过程中迫于环境压力与生存维持, 气孔往往在多个维度存在权衡(Brodribb & Jordan,
2011 ; Brodribb
et al. ,
2013 ; 丁凌子等,
2014 ; Fiorin
et al. ,
2016 ).虽然气孔数量的增加有助于提高光合效率, 但过多的气孔会加速叶内水汽逸散(潘莹萍和陈亚鹏,
2014 ; 王学芳等,
2014 ; 宋慧清等,
2020 ), 而且气孔的建成需要消耗资源, 这就决定了单位面积的气孔数量存在分布上限.气孔密度与大小共同决定了最大可能气孔导度并影响最大同化速率, 在考虑气孔建成实际资源消耗与光合作用水气协调的过程中, 两个相互矛盾的刚需必然达到最优化(Sack & Scoffoni,
2013 ).气孔面积与开度分别代表着气孔发育与运动, 且气孔发育总是先于气孔运动, 尽管大气孔对于开度的调控不及小气孔灵敏, 但可以获得相对较多的碳收益.就此而言, 气孔发育和运动过程中资源分配不均甚至存在矛盾(Xu & Zhou,
2008 ; Drake
et al. ,
2013 ).在异质生境中, 气孔各性状并非独立变化, 而是通过性状或特征组合的协变与权衡来共同适应环境.有限资源在植物体内的分配与权衡已通过异速生长理论在多个尺度得到证实(陈国鹏等,
2016 ; 段贝贝等,
2016 ; 杨克彤等,
2020 ), 但以往研究着重于相对宏观的尺度如群落、个体、构件与器官, 而且忽视了系统发育对资源权衡的影响, 对微观尺度如气孔性状的权衡机制关注较少.为此, 本文希望通过研究气孔特征并结合系统发育来探讨微观尺度气孔性状间的生长关系, 以进一步揭示微观尺度的权衡过程, 并丰富植物生活史研究. ...
... 兰州市(103.40° E, 36.03° N, 海拔1 500 m)位于黄土高原西北部, 地势西南高东北低, 呈东西狭长带状盆地特征, 属于大陆性很强的温带季风气候.市区地貌以山地丘陵为主, 占72.9%, 沟壑纵横, 地形破碎, 谷深坡陡, 年平均气温为10.3 ℃, 年日照时间2 446 h, 无霜期180天, 年降水量327 mm左右, 主要集中在6-9月(占全年降水量的50%-70%).土壤以灰钙土为主, 其次为栗钙土和灰褐土, 自然植被稀少, 主要是多年生禾草、旱生灌木和小乔木(段贝贝等,
2016 ; 杨克彤等,
2020 ), 覆盖率低于23%, 其中森林覆盖率仅为6.67%, 低于全国平均水平13.95%, 植被类型单一、结构简单, 是我国生态环境十分脆弱的地区之一. ...
兰州市典型绿化树种叶性状间的权衡关系
2
2020
... 系统发育和环境异质是影响植物性状发生分异的动力来源.气孔主要分布于植物叶片下表皮并参与调控碳水摄入与排出, 在其发育过程中迫于环境压力与生存维持, 气孔往往在多个维度存在权衡(Brodribb & Jordan,
2011 ; Brodribb
et al. ,
2013 ; 丁凌子等,
2014 ; Fiorin
et al. ,
2016 ).虽然气孔数量的增加有助于提高光合效率, 但过多的气孔会加速叶内水汽逸散(潘莹萍和陈亚鹏,
2014 ; 王学芳等,
2014 ; 宋慧清等,
2020 ), 而且气孔的建成需要消耗资源, 这就决定了单位面积的气孔数量存在分布上限.气孔密度与大小共同决定了最大可能气孔导度并影响最大同化速率, 在考虑气孔建成实际资源消耗与光合作用水气协调的过程中, 两个相互矛盾的刚需必然达到最优化(Sack & Scoffoni,
2013 ).气孔面积与开度分别代表着气孔发育与运动, 且气孔发育总是先于气孔运动, 尽管大气孔对于开度的调控不及小气孔灵敏, 但可以获得相对较多的碳收益.就此而言, 气孔发育和运动过程中资源分配不均甚至存在矛盾(Xu & Zhou,
2008 ; Drake
et al. ,
2013 ).在异质生境中, 气孔各性状并非独立变化, 而是通过性状或特征组合的协变与权衡来共同适应环境.有限资源在植物体内的分配与权衡已通过异速生长理论在多个尺度得到证实(陈国鹏等,
2016 ; 段贝贝等,
2016 ; 杨克彤等,
2020 ), 但以往研究着重于相对宏观的尺度如群落、个体、构件与器官, 而且忽视了系统发育对资源权衡的影响, 对微观尺度如气孔性状的权衡机制关注较少.为此, 本文希望通过研究气孔特征并结合系统发育来探讨微观尺度气孔性状间的生长关系, 以进一步揭示微观尺度的权衡过程, 并丰富植物生活史研究. ...
... 兰州市(103.40° E, 36.03° N, 海拔1 500 m)位于黄土高原西北部, 地势西南高东北低, 呈东西狭长带状盆地特征, 属于大陆性很强的温带季风气候.市区地貌以山地丘陵为主, 占72.9%, 沟壑纵横, 地形破碎, 谷深坡陡, 年平均气温为10.3 ℃, 年日照时间2 446 h, 无霜期180天, 年降水量327 mm左右, 主要集中在6-9月(占全年降水量的50%-70%).土壤以灰钙土为主, 其次为栗钙土和灰褐土, 自然植被稀少, 主要是多年生禾草、旱生灌木和小乔木(段贝贝等,
2016 ; 杨克彤等,
2020 ), 覆盖率低于23%, 其中森林覆盖率仅为6.67%, 低于全国平均水平13.95%, 植被类型单一、结构简单, 是我国生态环境十分脆弱的地区之一. ...
土壤水分胁迫对设施番茄叶片气孔特性的影响
1
2015
... 气孔性状(如气孔密度)的种间变异被认为是植物叶片经济谱的一部分(Loranger & Shipley,
2010 ; 王瑞丽等,
2016 ), 它在维持植物稳定碳收益、调节水力结构、吸收速效养分和信息传递等方面具有重要作用(Mencuccini
et al. ,
2015 ; Fiorin
et al. ,
2016 ).因此, 量化气孔性状在异质生境中的变异程度并解析其应对策略有助于进一步阐明植物对环境的适应机制.然而, 多数研究仅侧重于气孔性状对自然环境(海拔、水分和温度等)和控制试验(光强、氮添加和CO
2 浓度等)的响应并得出了一系列结论, 如: 气孔长度与密度之间为负相关关系(任昱等,
2014 ; 姜星星等,
2018 ), 随海拔升高气孔宽度呈减小趋势(王瑞丽等,
2016 ), 土壤湿度增大会显著降低气孔密度(杨再强等,
2015 ; 夏振华等,
2018 ; 张大才等,
2018 ), 高CO
2 浓度会显著降低气孔开张比(贾剑波等,
2019 )等, 但忽视了气孔自身在发育过程中的优先发育性状及其相互间关系的协变与权衡. ...
土壤水分胁迫对设施番茄叶片气孔特性的影响
1
2015
... 气孔性状(如气孔密度)的种间变异被认为是植物叶片经济谱的一部分(Loranger & Shipley,
2010 ; 王瑞丽等,
2016 ), 它在维持植物稳定碳收益、调节水力结构、吸收速效养分和信息传递等方面具有重要作用(Mencuccini
et al. ,
2015 ; Fiorin
et al. ,
2016 ).因此, 量化气孔性状在异质生境中的变异程度并解析其应对策略有助于进一步阐明植物对环境的适应机制.然而, 多数研究仅侧重于气孔性状对自然环境(海拔、水分和温度等)和控制试验(光强、氮添加和CO
2 浓度等)的响应并得出了一系列结论, 如: 气孔长度与密度之间为负相关关系(任昱等,
2014 ; 姜星星等,
2018 ), 随海拔升高气孔宽度呈减小趋势(王瑞丽等,
2016 ), 土壤湿度增大会显著降低气孔密度(杨再强等,
2015 ; 夏振华等,
2018 ; 张大才等,
2018 ), 高CO
2 浓度会显著降低气孔开张比(贾剑波等,
2019 )等, 但忽视了气孔自身在发育过程中的优先发育性状及其相互间关系的协变与权衡. ...
东达山高寒草甸8种植物气孔特征沿土壤水分梯度的变化
2
2018
... 气孔性状(如气孔密度)的种间变异被认为是植物叶片经济谱的一部分(Loranger & Shipley,
2010 ; 王瑞丽等,
2016 ), 它在维持植物稳定碳收益、调节水力结构、吸收速效养分和信息传递等方面具有重要作用(Mencuccini
et al. ,
2015 ; Fiorin
et al. ,
2016 ).因此, 量化气孔性状在异质生境中的变异程度并解析其应对策略有助于进一步阐明植物对环境的适应机制.然而, 多数研究仅侧重于气孔性状对自然环境(海拔、水分和温度等)和控制试验(光强、氮添加和CO
2 浓度等)的响应并得出了一系列结论, 如: 气孔长度与密度之间为负相关关系(任昱等,
2014 ; 姜星星等,
2018 ), 随海拔升高气孔宽度呈减小趋势(王瑞丽等,
2016 ), 土壤湿度增大会显著降低气孔密度(杨再强等,
2015 ; 夏振华等,
2018 ; 张大才等,
2018 ), 高CO
2 浓度会显著降低气孔开张比(贾剑波等,
2019 )等, 但忽视了气孔自身在发育过程中的优先发育性状及其相互间关系的协变与权衡. ...
... 于2019年7月选择无风、晴朗的天气, 在公园进行植物叶片采集.于9:00-11:00在选定好的3株标准木四周及中部分别采集10片完整、成熟叶片, 立即置于FAA固定液(70%酒精、38%甲醛、冰醋酸, 体积比为18:1:1)并带回室内, 固定24 h后进行气孔性状测定.首先, 随机取出5片已固定好的叶片用70%酒精冲洗残液, 后用蒸馏水冲洗并拭干水分, 然后在叶片背面中部选择一片避开主脉的区域(约1 cm
2 )涂上无色透明指甲油, 晾干后揭下制成临时装片并观察气孔特征(张大才等,
2018 ).观察、拍照时采用Imager A2生物显微镜(Zeiss, Jena, Germany), 每个装片选取3个镜头, 分别记录视野面积内气孔总数(
SNO )和开张气孔数(
OSN ), 气孔长度(
SL )、宽度(
SW )、面积(
SA )与气孔的孔径开度(
AOL )通过Image J软件计算, 气孔开张比(
SOR ) = (开张气孔数/气孔总数) × 100%, 气孔密度(
SD ) =气孔总数/视野面积(400 μm × 375 μm = 0.15 mm
2 ).鉴于自然界中多数植物的气孔主要分布在叶片下表皮, 故仅以下表皮气孔作为研究对象.气孔保卫细胞的长轴长度(μm)和短轴长度(μm)分别代表气孔长度和宽度, 将气孔开度定义为其孔径开度与宽度的比值, 即气孔开度(
SOL ) = (孔径开度/气孔宽度) × 100%, 介于0-100%之间, 值越大表示开度越大. ...
东达山高寒草甸8种植物气孔特征沿土壤水分梯度的变化
2
2018
... 气孔性状(如气孔密度)的种间变异被认为是植物叶片经济谱的一部分(Loranger & Shipley,
2010 ; 王瑞丽等,
2016 ), 它在维持植物稳定碳收益、调节水力结构、吸收速效养分和信息传递等方面具有重要作用(Mencuccini
et al. ,
2015 ; Fiorin
et al. ,
2016 ).因此, 量化气孔性状在异质生境中的变异程度并解析其应对策略有助于进一步阐明植物对环境的适应机制.然而, 多数研究仅侧重于气孔性状对自然环境(海拔、水分和温度等)和控制试验(光强、氮添加和CO
2 浓度等)的响应并得出了一系列结论, 如: 气孔长度与密度之间为负相关关系(任昱等,
2014 ; 姜星星等,
2018 ), 随海拔升高气孔宽度呈减小趋势(王瑞丽等,
2016 ), 土壤湿度增大会显著降低气孔密度(杨再强等,
2015 ; 夏振华等,
2018 ; 张大才等,
2018 ), 高CO
2 浓度会显著降低气孔开张比(贾剑波等,
2019 )等, 但忽视了气孔自身在发育过程中的优先发育性状及其相互间关系的协变与权衡. ...
... 于2019年7月选择无风、晴朗的天气, 在公园进行植物叶片采集.于9:00-11:00在选定好的3株标准木四周及中部分别采集10片完整、成熟叶片, 立即置于FAA固定液(70%酒精、38%甲醛、冰醋酸, 体积比为18:1:1)并带回室内, 固定24 h后进行气孔性状测定.首先, 随机取出5片已固定好的叶片用70%酒精冲洗残液, 后用蒸馏水冲洗并拭干水分, 然后在叶片背面中部选择一片避开主脉的区域(约1 cm
2 )涂上无色透明指甲油, 晾干后揭下制成临时装片并观察气孔特征(张大才等,
2018 ).观察、拍照时采用Imager A2生物显微镜(Zeiss, Jena, Germany), 每个装片选取3个镜头, 分别记录视野面积内气孔总数(
SNO )和开张气孔数(
OSN ), 气孔长度(
SL )、宽度(
SW )、面积(
SA )与气孔的孔径开度(
AOL )通过Image J软件计算, 气孔开张比(
SOR ) = (开张气孔数/气孔总数) × 100%, 气孔密度(
SD ) =气孔总数/视野面积(400 μm × 375 μm = 0.15 mm
2 ).鉴于自然界中多数植物的气孔主要分布在叶片下表皮, 故仅以下表皮气孔作为研究对象.气孔保卫细胞的长轴长度(μm)和短轴长度(μm)分别代表气孔长度和宽度, 将气孔开度定义为其孔径开度与宽度的比值, 即气孔开度(
SOL ) = (孔径开度/气孔宽度) × 100%, 介于0-100%之间, 值越大表示开度越大. ...
Leaf photosynthetic rate of tropical ferns is evolutionarily linked to water transport capacity
1
2014
... 系统发育决定着植物的亲缘关系并印刻着植物进化的信息, 在研究植物性状特征的过程中是否考虑系统发育将显著地影响研究结果(Ackerly,
2009 ; Zhang
et al. ,
2014 ).本研究中气孔性状间存在显著的相关关系, 在去除系统发育信息后, 这种关系大都发生了转变, 仅气孔长度与宽度依旧维持在稳健的正相关关系, 表明气孔长度与宽度间可能存在关联进化(张亚等,
2014 ; 高林浩等,
2020 ).即不论是否考虑系统发育, 气孔长度与宽度间的正相关关系都是存在的, 暗示这种协同关系可能在古老的植物类群中就已经出现.有研究认为, 植物气孔几何形状和间距总是沿着空间上最优的异速生长关系进化(Sack & Scoffoni,
2013 ; Fiorin
et al. ,
2016 ; Sperry
et al. ,
2017 ), 由于气孔发育属于同步型, 在气孔发育过程中长度与宽度总保持在相对稳定的比率(Franks & Farquhar,
2001 ), 这也从一定程度上解释了去除系统发育后, 气孔长度与宽度依旧保持稳健相关关系的原因.此外, 系统发育信号检测结果表明, 气孔性状的发育信号较弱(
K < 1), 表明气孔性状受系统发育影响较小, 但气孔开度和气孔开张比的发育信号显著, 暗示亲缘关系越近的物种气孔开度与开张比越相似(高林浩等,
2020 ).虽然系统发育会影响气孔性状, 但在长时间适应环境的过程中, 系统发育的影响似乎正在被逐渐削弱. ...
基部被子植物气孔性状与叶脉密度的关联进化
1
2014
... 系统发育决定着植物的亲缘关系并印刻着植物进化的信息, 在研究植物性状特征的过程中是否考虑系统发育将显著地影响研究结果(Ackerly,
2009 ; Zhang
et al. ,
2014 ).本研究中气孔性状间存在显著的相关关系, 在去除系统发育信息后, 这种关系大都发生了转变, 仅气孔长度与宽度依旧维持在稳健的正相关关系, 表明气孔长度与宽度间可能存在关联进化(张亚等,
2014 ; 高林浩等,
2020 ).即不论是否考虑系统发育, 气孔长度与宽度间的正相关关系都是存在的, 暗示这种协同关系可能在古老的植物类群中就已经出现.有研究认为, 植物气孔几何形状和间距总是沿着空间上最优的异速生长关系进化(Sack & Scoffoni,
2013 ; Fiorin
et al. ,
2016 ; Sperry
et al. ,
2017 ), 由于气孔发育属于同步型, 在气孔发育过程中长度与宽度总保持在相对稳定的比率(Franks & Farquhar,
2001 ), 这也从一定程度上解释了去除系统发育后, 气孔长度与宽度依旧保持稳健相关关系的原因.此外, 系统发育信号检测结果表明, 气孔性状的发育信号较弱(
K < 1), 表明气孔性状受系统发育影响较小, 但气孔开度和气孔开张比的发育信号显著, 暗示亲缘关系越近的物种气孔开度与开张比越相似(高林浩等,
2020 ).虽然系统发育会影响气孔性状, 但在长时间适应环境的过程中, 系统发育的影响似乎正在被逐渐削弱. ...
基部被子植物气孔性状与叶脉密度的关联进化
1
2014
... 系统发育决定着植物的亲缘关系并印刻着植物进化的信息, 在研究植物性状特征的过程中是否考虑系统发育将显著地影响研究结果(Ackerly,
2009 ; Zhang
et al. ,
2014 ).本研究中气孔性状间存在显著的相关关系, 在去除系统发育信息后, 这种关系大都发生了转变, 仅气孔长度与宽度依旧维持在稳健的正相关关系, 表明气孔长度与宽度间可能存在关联进化(张亚等,
2014 ; 高林浩等,
2020 ).即不论是否考虑系统发育, 气孔长度与宽度间的正相关关系都是存在的, 暗示这种协同关系可能在古老的植物类群中就已经出现.有研究认为, 植物气孔几何形状和间距总是沿着空间上最优的异速生长关系进化(Sack & Scoffoni,
2013 ; Fiorin
et al. ,
2016 ; Sperry
et al. ,
2017 ), 由于气孔发育属于同步型, 在气孔发育过程中长度与宽度总保持在相对稳定的比率(Franks & Farquhar,
2001 ), 这也从一定程度上解释了去除系统发育后, 气孔长度与宽度依旧保持稳健相关关系的原因.此外, 系统发育信号检测结果表明, 气孔性状的发育信号较弱(
K < 1), 表明气孔性状受系统发育影响较小, 但气孔开度和气孔开张比的发育信号显著, 暗示亲缘关系越近的物种气孔开度与开张比越相似(高林浩等,
2020 ).虽然系统发育会影响气孔性状, 但在长时间适应环境的过程中, 系统发育的影响似乎正在被逐渐削弱. ...
植物叶片气孔性状变异的影响因素及研究方法
3
2011
... 气孔作为植物叶表皮的特殊结构, 是植物与外界环境进行物质交换的门户(宋璐璐等,
2011 ; 朱燕华等,
2011 ), 也是植物体内水汽逸散的主要通道(金鹰和王传宽,
2015 ; 范嘉智等,
2016 ).气孔易受环境条件的修饰, 其形态特征是植物进化过程中对外界环境长期适应的结果(王碧霞等,
2010 ; 朱燕华等,
2011 ; 温婧雯等,
2018 ).为适应(或响应)复杂多变的环境, 气孔一方面通过调节开闭来应对短期或局域环境变化, 另一方面则协调大小和密度来适应异质生境(Brodribb & Jordan,
2011 ; Drake
et al. ,
2013 ; 金鹰和王传宽,
2015 ; 高冠龙等,
2018 ).利用气孔特征反映植物对环境变化的响应有助于明晰植物应对水气耦合过程的策略. ...
... ; 朱燕华等,
2011 ; 温婧雯等,
2018 ).为适应(或响应)复杂多变的环境, 气孔一方面通过调节开闭来应对短期或局域环境变化, 另一方面则协调大小和密度来适应异质生境(Brodribb & Jordan,
2011 ; Drake
et al. ,
2013 ; 金鹰和王传宽,
2015 ; 高冠龙等,
2018 ).利用气孔特征反映植物对环境变化的响应有助于明晰植物应对水气耦合过程的策略. ...
... 生长型和叶习性是引起植物性状变异的重要因素(Sack & Scoffoni,
2013 ; Mencuccini
et al. ,
2015 ).本研究中气孔性状在4种生长型和2种叶习性中均表现出显著差异.生长型主要造成局域环境内光照、温度和水分的分配不均, 叶习性则决定了植物的碳收益方式和水力结构.林冠上层高大乔木叶的生境为光照相对较强、温度变幅大而水分不足, 灌木或草本则相反.冠层植物叶片往往接受更多辐射, 需要较多的水气蒸腾以维持叶表温度(吴冰洁等,
2015 ), 常分化出小气孔开度、密集的气孔来应对强光照与高蒸腾强度; 林下植物则发育出大而疏散的气孔与较大的气孔开度, 这有助于叶片排出多余的水分(熊慧等,
2014 ; 王海珍等,
2015 ).相比于阔叶植物, 常绿植物叶片小而厚, 其表皮功能与微观结构也相对复杂(朱燕华等,
2011 ; 熊慧等,
2014 ).除气孔开张比外, 本研究中常绿植物的气孔性状普遍大于落叶植物, 这既有别于湿生物种大而疏散的气孔, 也与旱生植物小而密集的气孔有所区别.以往研究表明, 常绿植物的抗旱与抗栓塞能力强于落叶植物, 而且在叶片建成时更倾向于“缓慢投资-收益”策略(潘莹萍和陈亚鹏,
2014 ; 金鹰和王传宽,
2015 ).由于常绿植物叶片建成初期的碳投资高于落叶植物, 需要在叶片发育后期节约并获取更多的碳来偿还成本.因此, 常绿植物趋向于发育大而紧密的气孔以加速碳同化和积累更多的干物质. ...
植物叶片气孔性状变异的影响因素及研究方法
3
2011
... 气孔作为植物叶表皮的特殊结构, 是植物与外界环境进行物质交换的门户(宋璐璐等,
2011 ; 朱燕华等,
2011 ), 也是植物体内水汽逸散的主要通道(金鹰和王传宽,
2015 ; 范嘉智等,
2016 ).气孔易受环境条件的修饰, 其形态特征是植物进化过程中对外界环境长期适应的结果(王碧霞等,
2010 ; 朱燕华等,
2011 ; 温婧雯等,
2018 ).为适应(或响应)复杂多变的环境, 气孔一方面通过调节开闭来应对短期或局域环境变化, 另一方面则协调大小和密度来适应异质生境(Brodribb & Jordan,
2011 ; Drake
et al. ,
2013 ; 金鹰和王传宽,
2015 ; 高冠龙等,
2018 ).利用气孔特征反映植物对环境变化的响应有助于明晰植物应对水气耦合过程的策略. ...
... ; 朱燕华等,
2011 ; 温婧雯等,
2018 ).为适应(或响应)复杂多变的环境, 气孔一方面通过调节开闭来应对短期或局域环境变化, 另一方面则协调大小和密度来适应异质生境(Brodribb & Jordan,
2011 ; Drake
et al. ,
2013 ; 金鹰和王传宽,
2015 ; 高冠龙等,
2018 ).利用气孔特征反映植物对环境变化的响应有助于明晰植物应对水气耦合过程的策略. ...
... 生长型和叶习性是引起植物性状变异的重要因素(Sack & Scoffoni,
2013 ; Mencuccini
et al. ,
2015 ).本研究中气孔性状在4种生长型和2种叶习性中均表现出显著差异.生长型主要造成局域环境内光照、温度和水分的分配不均, 叶习性则决定了植物的碳收益方式和水力结构.林冠上层高大乔木叶的生境为光照相对较强、温度变幅大而水分不足, 灌木或草本则相反.冠层植物叶片往往接受更多辐射, 需要较多的水气蒸腾以维持叶表温度(吴冰洁等,
2015 ), 常分化出小气孔开度、密集的气孔来应对强光照与高蒸腾强度; 林下植物则发育出大而疏散的气孔与较大的气孔开度, 这有助于叶片排出多余的水分(熊慧等,
2014 ; 王海珍等,
2015 ).相比于阔叶植物, 常绿植物叶片小而厚, 其表皮功能与微观结构也相对复杂(朱燕华等,
2011 ; 熊慧等,
2014 ).除气孔开张比外, 本研究中常绿植物的气孔性状普遍大于落叶植物, 这既有别于湿生物种大而疏散的气孔, 也与旱生植物小而密集的气孔有所区别.以往研究表明, 常绿植物的抗旱与抗栓塞能力强于落叶植物, 而且在叶片建成时更倾向于“缓慢投资-收益”策略(潘莹萍和陈亚鹏,
2014 ; 金鹰和王传宽,
2015 ).由于常绿植物叶片建成初期的碳投资高于落叶植物, 需要在叶片发育后期节约并获取更多的碳来偿还成本.因此, 常绿植物趋向于发育大而紧密的气孔以加速碳同化和积累更多的干物质. ...
 ,
, ,1,*, 俞筱押2, 鲜骏仁3
,1,*, 俞筱押2, 鲜骏仁3

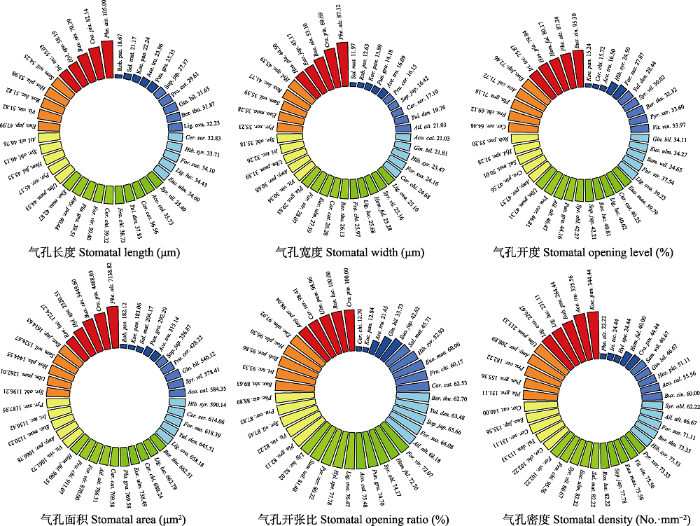 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT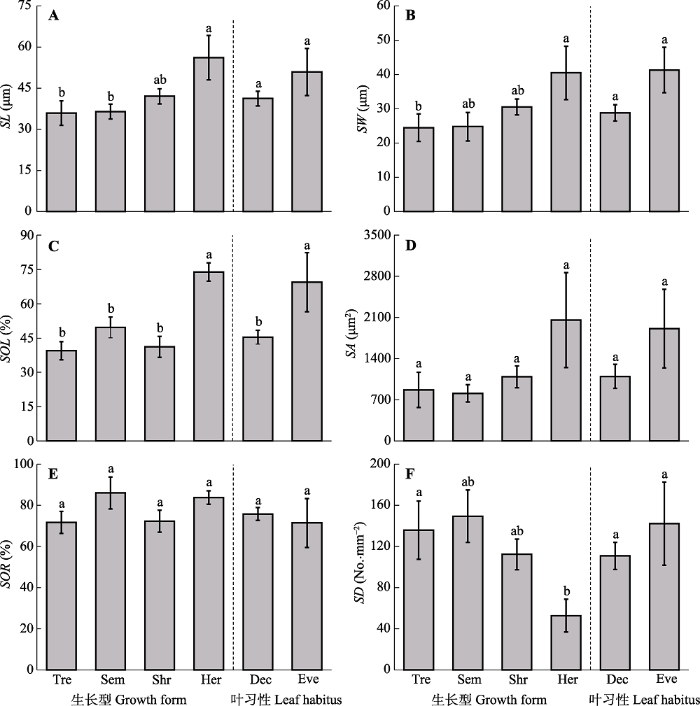 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT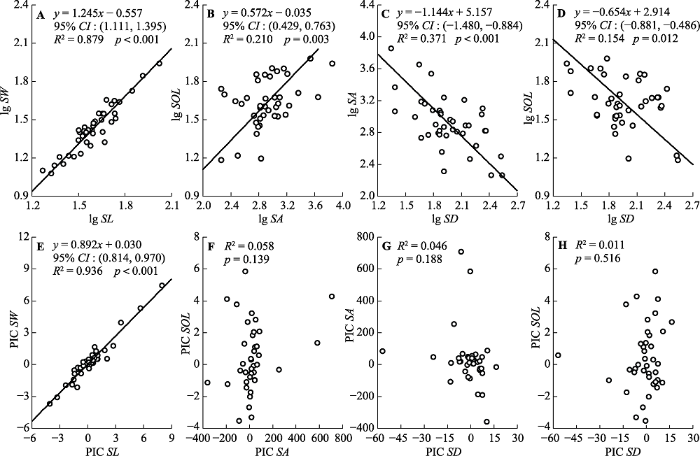 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT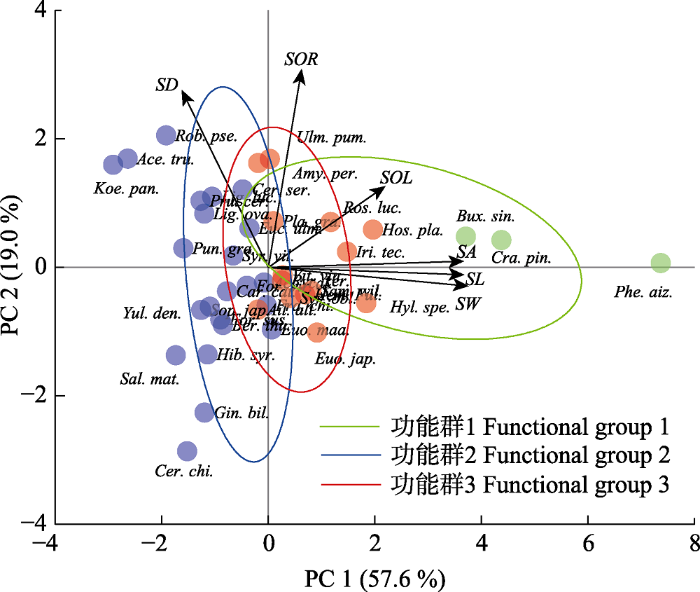 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}