Characteristics and environmental factors controlling methane emission from a Sphagnum bog with different plant cover types in a subalpine area, southwest of Hubei, China
Li MOU, Lin WU,*, Xue-Fei LIU, Xiao-Ling LI, Han WANG, Hao WU, Yu-Rong YU, Sheng-Lan DUBiological Resources Protection and Utilization of the Key Laboratory of Hubei Province, School of Forestry and Horticulture, Hubei Minzu University, Enshi, Hubei 445000, China
Abstract Aims Wetlands are important sources of atmospheric methane (CH4), but there are few reports on the CH4 emissions of subtropical and subalpine wetlands. In particular, the accurate estimation of CH4 emissions from plots with different types of plant cover and the controlling environmental factors are not clear. The objective of this study is to study characteristics and influencing factors of CH4 emission fluxes from a Sphagnum bog with different plant cover types in a subalpine area, southwest of Hubei Province. Methods A Sphagnum bog in the subalpine area in southwest of Hubei Province was selected, and CH4emission fluxes were measured in the Sphagnum bog with three types of plant cover using closed static chamber and gas chromatography method from November 2018 to October 2019. Simultaneously, air temperature, soil temperature of 5 cm depth, and groundwater level were recorded. Important findings (1) Under sunlight, the CH4-C fluxes from the bare land plot (B), the Sphagnum paluster plot (S), and the Polytrichum commune plot (P) varied throughout the year within the following ranges: 0.012- 1.372, 0.022-1.474 and 0.027-3.385 mg·m-2·h-1, respectively; under shading treatment, the variation range of CH4-C flux throughout the year from B, S and P plots were 0.012-1.372, 0.009-1.839 and 0.017-2.484 mg·m-2·h-1, respectively, indicating CH4emission sources for all types. At the same time, for all plant cover types, CH4 emissions under sunlight conditions were slightly higher than those under dark conditions, but the difference was not significant. (2) For all plant cover types, CH4 emissions showed obvious seasonal variations, with the order of summer > autumn > spring > winter; and summer emissions were significantly greater than emissions in other seasons, accounting for about 57% to 84% of the annual cumulative flux. This study found that the flux of CH4 emission was highly related to air temperature and soil temperature of 5 cm depth showing exponential relationships, which indicated that temperature is the main environment factor affecting the temporal variations of CH4 emission from the Sphagnum bog. (3) Plant cover types significantly affected CH4 emissions from theSphagnum bog. The annual average and cumulative CH4 emissions from the three cover types were in the order of: P > S > B. P plot showed significantly higher emission than B plot. This study found a significant correlation between vegetation types and CH4 emissions, indicating that plant cover type is the main influencing factor of the spatial variations of CH4 emissions from the Sphagnum bog. (4) CH4 emissions were not significantly related to the groundwater levels. This study further enriched the mechanisms of CH4 emission in the Sphagnum bog and provided basic data for regional carbon cycling. Keywords:subtropical;vegetation type;marshland;methane emission
PDF (1679KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 牟利, 吴林, 刘雪飞, 李小玲, 王涵, 吴浩, 余玉蓉, 杜胜蓝. 鄂西南亚高山不同覆被类型泥炭藓沼泽湿地甲烷排放特征及其环境影响因子. 植物生态学报, 2021, 45(2): 131-143. DOI: 10.17521/cjpe.2020.0292 MOU Li, WU Lin, LIU Xue-Fei, LI Xiao-Ling, WANG Han, WU Hao, YU Yu-Rong, DU Sheng-Lan. Characteristics and environmental factors controlling methane emission from a Sphagnum bog with different plant cover types in a subalpine area, southwest of Hubei, China . Chinese Journal of Plant Ecology, 2021, 45(2): 131-143. DOI: 10.17521/cjpe.2020.0292
泥炭藓沼泽湿地是一类以泥炭藓属(Sphagnum)植物为优势物种的特殊湿地类型, 其强酸性、贫营养、潮湿和缺氧的环境为甲烷(CH4)生成提供了良好环境(Dedysh, 2002)。研究表明, 北方泥炭藓沼泽湿地全年约排放 46 Tg CH4-C (Gorham, 1991), 是大气CH4的重要来源(Wuebbles & Hayhoe, 2002; Turetsky et al., 2014; Leroy et al., 2019)。但是, 全球泥炭藓沼泽湿地CH4排放量存在很大的变异(Turetsky et al., 2014; Hanson et al., 2016)。例如, 国外泥炭藓沼泽湿地平均CH4-C排放量通常为0.01-3.3 mg·m-2·h-1 (Blodau, 2002); 而我国东北泥炭藓沼泽湿地全年CH4-C排放量为0.026-0.38 mg·m-2·h-1 (孙晓新等, 2009; Miao et al., 2012; 常怡慧等, 2020)。同时, 研究发现贫营养泥炭藓沼泽湿地CH4排放量通常小于富营养泥炭藓沼泽湿地(Brown, 1998; Chasar et al., 2000; B?ckstrandet al., 2010)。甚至, 同一泥炭藓湿地不同微地形间CH4的排放也存在很大的差异(Frenzel & Karofeld, 2000; Beetz et al., 2013; Gill et al., 2017; Zalman et al., 2018)。因此, 对不同地区不同生境泥炭藓沼泽湿地CH4排放特征及其影响因素的研究是准确评估全球泥炭藓沼泽湿地碳循环的关键。
目前, 有关泥炭藓沼泽湿地CH4排放的研究主要集中在北半球纬度较高的温带和寒温带地区(Chasar et al., 2000; Larmola et al., 2010; St?pniewskaet al., 2018)。研究表明, 沼泽湿地内的泥炭藓体内有甲烷氧化细菌, 可氧化从泥炭地产生的CH4, 并为其光合作用提供碳源, 从而减少CH4排放(Raghoebarsing et al., 2005; Kip et al., 2010)。但是, 在我国北方泥炭藓沼泽地中, 去除泥炭藓后CH4的排放反而减少了50% (Meng et al., 2014)。由此可见, 植被对泥炭藓沼泽湿地CH4排放的影响是一个复杂过程, 其通常与温度、水位及其他环境因子发生耦合作用, 进一步影响CH4的排放(Mikkel?et al., 1995; 孙晓新等, 2009; Riutta et al., 2020)。Nyk?nen等(1998)对芬兰泥炭藓沼泽湿地和李艳元等(2017)对神农架大九湖的研究均表明温度和土壤湿度会影响CH4排放; 而中欧有关泥炭藓沼泽湿地的研究表明地下水位对CH4排放的影响更为重要(Urbanováet al., 2013)。还有研究发现, 落叶松-泥炭藓沼泽湿地CH4排放与土壤温度、土壤含水量等环境因子均不显著相关(常怡慧等, 2020)。此外, 部分研究表明光对泥炭藓中微生物氮的分解和CH4的氧化有很强的促进作用, 光照条件下泥炭藓CH4的氧化速率比在黑暗条件下提高5倍, 而黑暗条件下CH4氧化活动较低导致CH4排放量随之升高(Kox et al., 2020)。综上所述, 不同地区影响泥炭藓沼泽湿地CH4排放的关键因素不同。并且, 亚热带地区有关泥炭藓沼泽湿地CH4通量的研究鲜有报道(李艳元等, 2017; Alfadhel et al., 2019; 谌佳伟等, 2020)。
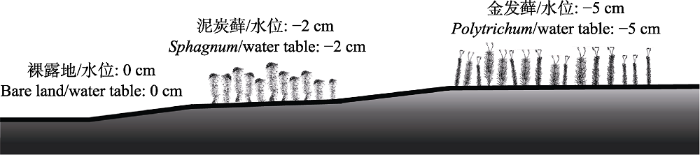
Fig. 2Diurnal changes of methane (CH4) emission fluxes in the Sphagnum bog (mean + SE). ▲, bare land plot; ■, Sphagnum paluster plot; ●, Polytrichum commune plot. The lines represent soil temperature at 5 cm depth, the lines with points represent CH4 emission fluxes, the bars represent air temperature. * represents a significant difference among the three cover types at the same time (p < 0.05), ** represents a significant difference among the three cover types at the same time (p < 0.01).
Table 1 表1 表1植被类型、季节及其交互作用对泥炭藓沼泽湿地CH4排放通量影响的重复测量方差分析结果 Table 1Results of repeated measures ANOVAs on the effects of vegetation type, season, and their interactions on methane emission fluxes of the Sphagnum bog
Fig. 3Seasonal changes in methane emission fluxes in the Sphagnum bog (mean ± SE). B, bare land plot; P, Polytrichum commune plot; S, Sphagnum paluster plot. Different uppercase letters indicate significant difference among the same cover type in different seasons (p < 0.01); different lowercase letters indicate significant difference among the annual methane emissions of three cover types (p < 0.05). * represents a significant difference among the three cover types at the same season (p < 0.05); ** represents a significant difference among the three cover types at the same season (p < 0.01).
Fig. 4Relationship between methane emission fluxes and air temperature in the Sphagnum bog. B, bare land plot; P, Polytrichum commune plot; S, Sphagnum paluster plot.
Fig. 5Relationship between methane emission fluxes and soil temperature of 5 cm depth in the Sphagnum bog. B, bare land plot; P, Polytrichum commune plot; S, Sphagnum paluster plot.
Fig. 6Relationship between methane emission fluxes and water table in the Sphagnum bog. B, bare land plot; P, Polytrichum commune plot; S, Sphagnum paluster plot.
新窗口打开|下载原图ZIP|生成PPT 图7不同地区泥炭藓沼泽湿地CH4排放通量示意图。本综合示意图是根据Nykanen et al.(1998), Beetz et al.(2013), Hanson et al.(2016)及本研究测定的CH4排放绘制的。
Fig. 7Schematic diagram of methane emission flux of the Sphagnum bog in different regions. This comprehensive diagram is based on methane emission flux measured by Nykanen et al.(1998), Beetz et al.(2013), Hanson et al.(2016) and this study.
Table 2 表2 表2不同地区泥炭藓沼泽湿地CH4-C排放对比 Table 2CH4-C emission fluxes from the Sphagnum bog in different regions
Fig. 8Average methane emission fluxes from different cover types of the Sphagnum bogs. B, bare land plot; P, Polytrichum commune plot; S, Sphagnum paluster plot.
本研究区泥炭藓覆被样地CH4排放量大于裸露地, 与国外很多研究结论相反, 这可能与甲烷氧化细菌含量有关。研究表明, 多数泥炭藓属植物体中有甲烷氧化细菌(Raghoebarsing et al., 2005; Larmola et al., 2010), 泥炭藓沼泽湿地CH4排放量要小于裸露地(Waddington & Day, 2007; Korrensalo et al., 2018)。然而, 研究发现不同泥炭藓物种甲烷氧化细菌含量不同(Zalman et al., 2018), 部分地区泥炭藓中甲烷氧化细菌多样性很低(Knoblaunch et al., 2008; 谌佳伟等, 2020)。例如, 神农架大九湖湿地甲烷氧化细菌就很少, 且大九湖湿地环境与本研究区较为接近, 泥炭藓物种也相同(谌佳伟等, 2020)。因此,本研究区泥炭藓中的甲烷氧化细菌含量可能也较少。另外, 研究区泥炭藓覆被样地CH4排放量大于裸露地可能与有机底物及地上生物量的限制有关(孙晓新等, 2009; Yavitt et al., 2012)。研究表明, 裸露地缺少凋落物, 导致土壤微生物生长代谢的主要底物缺失, 同时土壤理化性质被改变, 降低CH4排放量(Strakováet al., 2011); 另一方面, 裸露地水位常年高于或接近地表, 泥炭中少量的CH4在穿过土壤水层时, 增加CH4被氧化的时间, 进一步减少CH4排放量(Kelley et al., 1995)。Meng等(2014)在东北发现去除泥炭藓2年后, CH4排放量降低了50%, 本研究结论与之一致。与之相反, 多数研究表明维管植物入侵, 泥炭藓减少会增加泥炭藓沼泽湿地CH4排放量(Leroy et al., 2019)。本研究中的覆被类型没有考虑维管植物, 但研究结论还是在一定程度上论证了覆被类型对泥炭藓沼泽湿地CH4排放的重要性。同时, 仍有必要研究更多植被类型对泥炭藓沼泽湿地CH4排放的影响, 这将有助于我们更加深刻认识亚热带亚高山泥炭藓湿地碳循环过程。
3.3 影响泥炭藓沼泽湿地CH4排放通量的环境因子
本研究对比分析3种覆被类型泥炭藓沼泽湿地CH4排放量与气温和地下5 cm土壤温度关系, 发现两者均呈显著正相关关系(p <0.05, 图4, 图5), 且沼泽湿地CH4的排放量随着温度增加呈指数增加的趋势, 与以往多数研究(Crill et al., 1988; Bohn et al., 2007; 李艳元等, 2017)结论一致。Gill等(2017)对泥炭藓沼泽湿地不同增温处理的研究表明, 泥炭地对气候变暖很敏感, 全球变暖可能增加泥炭藓湿地CH4排放量; 主要是温度升高和温度波动可能改变泥炭基质的分压差, 增加泥炭地气体沸腾, 从而绕过CH4氧化的富氧区, 促进气体逸出(Beckmann et al., 2004; Waddington et al., 2010; Gill et al., 2017); 另一方面, 温度升高促进沼泽湿地水面蒸发, 间接加速CH4的排放(Saarnio et al., 1997)。因此, 温度是影响本研究区CH4排放的一个关键环境因子。
AlfadhelI, GeJ, SinanY, LiuY (2019). Methane flux and its environmental impact factors in Dajiuhu wetland of Shennongjia Wuhan University Journal of Natural Sciences, 24,455-460. DOI:10.1007/s11859-019-1421-7URL [本文引用: 2]
B?ckstrandK, CrillPM, Jackowicz-Korczy?skiM, MastepanovM, ChristensenTR, BastvikenD (2010). Annual carbon gas budget for a subarctic peatland, Northern Sweden Biogeosciences, 7,95-108. DOI:10.5194/bg-7-95-2010URL [本文引用: 1]
BeckmannM, SheppardSK, LloydD (2004). Mass spectrometric monitoring of gas dynamics in peat monoliths: effects of temperature and diurnal cycles on emissions Atmospheric Environment, 38,6907-6913. DOI:10.1016/j.atmosenv.2004.08.004URL [本文引用: 1]
BeetzS, LiebersbachH, GlatzelS, JurasinskiG, BuczkoU, H?perH (2013). Effects of land use intensity on the full greenhouse gas balance in an Atlantic peat bog. Biogeosciences, 10,1067-1082. DOI:10.5194/bg-10-1067-2013URL [本文引用: 3]
BlodauC (2002). Carbon cycling in peatlands—A review of processes and controls Environmental Reviews, 10,111-134. DOI:10.1139/a02-004URL [本文引用: 1]
BohdálkováL, ?u?íkJ, AaK, B?zekF (2012). Dynamics of methane fluxes from two peat bogs in the Ore Mountains, Czech Republic Plant, Soil and Environment, 59,14-21. DOI:10.17221/PSEURL [本文引用: 1]
BohnTJ, LettenmaierDP, SathulurK, BowlingLC, PodestE, McDonaldKC, FriborgT (2007). Methane emissions from western Siberian wetlands: heterogeneity and sensitivity to climate change Environmental Research Letters, 2,45015. DOI: 10.1088/1748-9326/2/4/045015. DOI:10.1088/1748-9326/2/4/045015URL [本文引用: 1]
BrownD (1998). Gas production from an ombrotrophic bog—Effect of climate change on microbial ecology Climatic Change, 40,277-284. DOI:10.1023/A:1005421004276URL [本文引用: 1]
BuZJ, YangYF, DaiD, WangXW (2005). Age structure and growth pattern of Polytrichum juniperum populations in a mire of Changbai Mountains Chinese Journal of Applied Ecology, 16,44-48. [本文引用: 2]
ChangYH, MuCC, PengWH, HaoL, HanLD (2020). Characteristics of greenhouse gas emissions from seven swamp types in the permafrost region of Daxing'an Mountains, northeast China Acta Ecologica Sinica, 40,2333-2346. [本文引用: 2]
ChasarLS, ChantonJP, GlaserPH, SiegelDI (2000). Methane concentration and stable isotope distribution as evidence of rhizospheric processes: comparison of a fen and bog in the glacial Lake Agassiz Peatland complex Annals of Botany, 86,655-663. DOI:10.1006/anbo.2000.1172URL [本文引用: 2]
ChenJW, GeJW, FengL, ZhouY, GanJ, LiYF, ZhangZQ (2020). Methane flux characteristics and its relationship with soil microbial community composition of Dajiuhu peatland in Shennongjia Earth Science, 45,1082-1092. [本文引用: 9]
CuiHJ, ZhangY, ZhangYF, TianK, XiaoDR, WangK, GuoY, LiLJ (2018). Characteristics of moss bog plant community and interspecific relationships of dominant species in Niangniang Mountain wetland, Guizhou Chinese Journal of Ecology, 37,2619-2626. [本文引用: 2]
FrenzelP, KarofeldE (2000). CH4 emission from a hollow-ridge complex in a raised bog: the role of CH4 production and oxidation Biogeochemistry, 51,91-112. DOI:10.1023/A:1006351118347URL [本文引用: 1]
GillAL, GiassonMA, YuR, FinziAC (2017). Deep peat warming increases surface methane and carbon dioxide emissions in a black spruce-dominated ombrotrophic bog. Global Change Biology, 23,5398-5411. DOI:10.1111/gcb.2017.23.issue-12URL [本文引用: 3]
GorhamE (1991). Northern peatlands: role in the carbon cycle and probable responses to climatic warming Ecological Applications, 1,182-195. DOI:10.2307/1941811URL [本文引用: 1]
HansonPJ, GillAL, XuX, PhillipsJR, WestonDJ, KolkaRK, RiggsJS, HookLA (2016). Intermediate-scale community-level flux of CO2 and CH4 in a Minnesota peatland: putting the SPRUCE project in a global context Biogeochemistry, 129,255-272. DOI:10.1007/s10533-016-0230-8URL [本文引用: 3]
HaoL, MuCC, ChangYH, ShenZQ, HanLD, JiangN, PengWH (2019). Effects of harvest on greenhouse gas emissions from forested swamp during non-growing season in Xiaoxing’an Mountains of China Chinese Journal of Applied Ecology, 30,1713-1725. DOI:10.13287/j.1001-9332.201905.036PMID:31107028 [本文引用: 1] Soil greenhouse gas emission during non-growing season plays an important role in ecosystem carbon and nitrogen cycling in mid and high latitude regions. However, the effects of harvest on greenhouse gas emission during non-growing remain unclear. We measured the fluxes of CO, CH and NO and environmental factors (soil temperature, moisture, soil organic carbon and total nitrogen etc.) during non-growing season from four kinds of forested swamps (Alnus sibirica swamp, Betula platyphylla swamp, Larix gmelinii-Carex schmidti swamp, L. gmelinii-moss swamp) under different harvest disturbances for 10 years, including control (no cutting), 45% selective cutting, clear cutting, by using static chamber technique and gas chromatography in Xiaoxing'an Mountains, Northeast China. The aim of this study was to reveal the effects of harvest on greenhouse gas emission from temperate forested swamp during non-growing season and the main controlling factors. The results showed that the average fluxes of CO, CH and NO from four kinds of swamps distributed in 53.08-81.31 mg·m·h, 0.09-3.07 mg·m·h and 4.07-8.83 μg·m·h, respectively. Clear cutting significantly increased the fluxes of CO, CH, and NO from A. sibirica swamp and L. gmelinii-moss swamp. Selective cutting significantly increased CO fluxes from B. platyphylla swamp and L. gmelinii-moss swamp and decreased CO flux from A. sibirica swamp. Selective cutting significantly decreased CH fluxes from all the four forested swamps and NO flux from Larix gmelinii-Carex schmidti swamp. The CO fluxes from natural forested swamps were strongly influenced by soil temperature, soil organic carbon and C/N. CH fluxes were influenced by soil temperature, soil organic carbon. NO fluxes were affected by air temperature and soil pH. Harvesting increased the correlation between soil CO flux and air temperature, soil moisture and snow depth, the correlation between soil CH flux and air temperature, soil moisture and C/N, as well as the correlation between soil NO flux and soil total nitrogen and C/N. The annual cumulative contribution of CO, CH and NO emission from natural forested swamp during non-growing season were 33.2%-46.5%, 6.3%-9.1% and 61.5%-68.3%, respectively. The clear cutting increased the annual cumulative contribution of CO from B. platyphylla swamp and L. gmelinii-moss swamp and that of NO from other swamps except L. gmelinii-moss swamp. The selective cutting increased the annual cumulative contribution of CO, CH and NO from L. gmelinii-C. schmidti swamp and L. gmelinii-moss swamp, but decreased that from B. platyphylla swamp. The annual cumulative contributions of NO and CO during non-growing season were relatively high from temperate natural forested swamps, and clear cutting further increased their contribution, while the selective cutting just increased that of CH during non-growing season. [ 郝利, 牟长城, 常怡慧, 申忠奇, 韩丽冬, 姜宁, 彭文宏 (2019). 采伐对小兴安岭森林沼泽非生长季土壤温室气体排放的影响 应用生态学报, 30,1713-1725.] PMID:31107028 [本文引用: 1]
HinesME, DuddlestonKN, Rooney-VargaJN, FieldsD, ChantonJP (2008). Uncoupling of acetate degradation from methane formation in Alaskan wetlands: connections to vegetation distribution Global Biogeochemical Cycles, 22, GB2017. DOI: 10.1029/2006GB002903. DOI:10.1029/2006GB002903 [本文引用: 1]
KelleyCA, MartensCS, UsslerIII W (1995). Methane dynamics across a tidally flooded riverbank margin Limnology and Oceanography, 40,1112-1129. DOI:10.4319/lo.1995.40.6.1112URL [本文引用: 1]
KettunenA, KaitalaV, LehtinenA, LohilaA, AlmJ, SilvolaJ, MartikainenPJ (1999). Methane production and oxidation potentials in relation to water table fluctuations in two boreal mires Soil Biology & Biochemistry, 31,1741-1749. DOI:10.1016/S0038-0717(99)00093-0URL [本文引用: 1]
KipN, vanWinden JF, PanY, BodrossyL, ReichartGJ, SmoldersAJP, JettenMSM, DamstéJSS, den Camp HJMO (2010). Global prevalence of methane oxidation by symbiotic bacteria in peat-moss ecosystems Nature Geoscience, 3,617-621. DOI:10.1038/ngeo939URL [本文引用: 1]
KnoblauchC, ZimmermannU, BlumenbergM, MichaelisW, PfeifferEM (2008). Methane turnover and temperature response of methane-oxidizing bacteria in permafrost- affected soils of northeast Siberia Soil Biology & Biochemistry, 40,3004-3013. DOI:10.1016/j.soilbio.2008.08.020URL [本文引用: 1]
KoponenHT, MartikainenPJ (2004). Soil water content and freezing temperature affect freeze-thaw related N2O production in organic soil Nutrient Cycling in Agroecosystems, 69,213-219. DOI:10.1023/B:FRES.0000035172.37839.24URL [本文引用: 1]
KorrensaloA, M?nnist?E, AlekseychikP, MammarellaI, RinneJ, VesalaT, TuittilaES (2018). Small spatial variability in methane emission measured from a wet patterned boreal bog. Biogeosciences, 15,1749-1761. DOI:10.5194/bg-15-1749-2018URL [本文引用: 1]
KoxMAR, vanden Elzen E, LamersLPM, JettenMSM, van Kessel MAHJ (2020). Microbial nitrogen fixation and methane oxidation are strongly enhanced by light in Sphagnum mosses AMB Express, 10, 61. DOI: 10.1186/s13568-020-00994-9. DOI:10.1186/s13568-020-00994-9 [本文引用: 2]
LarmolaT, TuittilaES, TiirolaM, Nyk?nenH, MartikainenPJ, Yrj?l?K, TuomivirtaT, FritzeH (2010). The role of Sphagnum mosses in the methane cycling of a boreal mire Ecology, 91,2356-2365. PMID:20836457 [本文引用: 2] Peatlands are a major natural source of atmospheric methane (CH4). Emissions from Sphagnum-dominated mires are lower than those measured from other mire types. This observation may partly be due to methanotrophic (i.e., methane-consuming) bacteria associated with Sphagnum. Twenty-three of the 41 Sphagnum species in Finland can be found in the peatland at Lakkasuo. To better understand the Sphagnum-methanotroph system, we tested the following hypotheses: (1) all these Sphagnum species support methanotrophic bacteria; (2) water level is the key environmental determinant for differences in methanotrophy across habitats; (3) under dry conditions, Sphagnum species will not host methanotrophic bacteria; and (4) methanotrophs can move from one Sphagnum shoot to another in an aquatic environment. To address hypotheses 1 and 2, we measured the water table and CH4 oxidation for all Sphagnum species at Lakkasuo in 1-5 replicates for each species. Using this systematic approach, we included Sphagnum spp. with narrow and broad ecological tolerances. To estimate the potential contribution of CH4 to moss carbon, we measured the uptake of delta13C supplied as CH4 or as carbon dioxide dissolved in water. To test hypotheses 2-4, we transplanted inactive moss patches to active sites and measured their methanotroph communities before and after transplantation. All 23 Sphagnum species showed methanotrophic activity, confirming hypothesis 1. We found that water level was the key environmental factor regulating methanotrophy in Sphagnum (hypothesis 2). Mosses that previously exhibited no CH4 oxidation became active when transplanted to an environment in which the microbes in the control mosses were actively oxidizing CH4 (hypothesis 4). Newly active transplants possessed a Methylocystis signature also found in the control Sphagnum spp. Inactive transplants also supported a Methylocystis signature in common with active transplants and control mosses, which rejects hypothesis 3. Our results imply a loose symbiosis between Sphagnum spp. and methanotrophic bacteria that accounts for potentially 10-30% of Sphagnum carbon.
LeroyF, GogoS, GuimbaudC, Bernard-JanninL, YinX, BelotG, ShuguangW, Laggoun-DéfargeF (2019). CO2 and CH4budgets and global warming potential modifications in Sphagnum-dominated peat mesocosms invaded by Molinia caerulea Biogeosciences, 16,4085-4095. DOI:10.5194/bg-16-4085-2019URL [本文引用: 2]
LiYL, MiaoJL, HeY (2013). Influence of calculation methods on calculated daily mean temperature Meteorogical Science and Technology, 41(1),88-92. [本文引用: 1]
LiuLF, ChenH, PengCH, ZhuQA, LiBX (2016). CH4 emissions under warming schemes from peatlands of different depths in the Zoige Plateau Chinese Journal of Applied and Environmental Biology, 22,1-7. [本文引用: 1]
MengHN, SongCC, MiaoYQ, MaoR, WangXW (2014). Response of CH4 emissions to moss removal and N addition in boreal peatland of northeast China Biogeosciences, 11,4809-4816. DOI:10.5194/bg-11-4809-2014URL [本文引用: 2]
MiaoY, SongC, SunL, WangX, MengH, MaoR (2012). Growing season methane emission from a boreal peatland in the continuous permafrost zone of Northeast China: effects of active layer depth and vegetation Biogeosciences, 9,4455-4464. DOI:10.5194/bg-9-4455-2012URL [本文引用: 1]
Mikkel?C, SundhI, SvenssonBH, NilssonM (1995). Diurnal variation in methane emission in relation to the water table, soil temperature, climate and vegetation cover in a Swedish acid mire Biogeochemistry, 28,93-114. DOI:10.1007/BF02180679URL [本文引用: 1]
Nyk?nenH, AlmJ, SilvolaJ, TolonenK, MartikainenPJ (1998). Methane fluxes on boreal peatlands of different fertility and the effect of long-term experimental lowering of the water table on flux rates Global Biogeochemical Cycles, 12,53-69. DOI:10.1029/97GB02732URL [本文引用: 5]
RaghoebarsingAA, SmoldersAJP, SchmidMC, RijpstraWIC, Wolters-ArtsM, DerksenJ, JettenMSM, SchoutenS, SinningheDamsté JS, LamersLPM, RoelofsJGM, OpDen Camp HJM, StrousM (2005). Methanotrophic symbionts provide carbon for photosynthesis in peat bogs Nature, 436,1153-1156. PMID:16121180 [本文引用: 3] Wetlands are the largest natural source of atmospheric methane, the second most important greenhouse gas. Methane flux to the atmosphere depends strongly on the climate; however, by far the largest part of the methane formed in wetland ecosystems is recycled and does not reach the atmosphere. The biogeochemical controls on the efficient oxidation of methane are still poorly understood. Here we show that submerged Sphagnum mosses, the dominant plants in some of these habitats, consume methane through symbiosis with partly endophytic methanotrophic bacteria, leading to highly effective in situ methane recycling. Molecular probes revealed the presence of the bacteria in the hyaline cells of the plant and on stem leaves. Incubation with (13)C-methane showed rapid in situ oxidation by these bacteria to carbon dioxide, which was subsequently fixed by Sphagnum, as shown by incorporation of (13)C-methane into plant sterols. In this way, methane acts as a significant (10-15%) carbon source for Sphagnum. The symbiosis explains both the efficient recycling of methane and the high organic carbon burial in these wetland ecosystems.
RaskH, SchoenauJ, AndersonD (2002). Factors influencing methane flux from a boreal forest wetland in Saskatchewan, Canada Soil Biology & Biochemistry, 34,435-443. DOI:10.1016/S0038-0717(01)00197-3URL [本文引用: 1]
RiuttaT, KorrensaloA, LaineAM, LaineJ, TuittilaES (2020). Interacting effects of vegetation components and water level on methane dynamics in a boreal fen. Biogeosciences, 17,727-740. DOI:10.5194/bg-17-727-2020URL [本文引用: 1]
SaarnioS, AlmJ, SilvolaJ, LohilaA, Nyk?nenH, MartikainenPJ (1997). Seasonal variation in CH4 emissions and production and oxidation potentials at microsites on an oligotrophic pine fen. Oecologia, 110,414-422. DOI:10.1007/s004420050176PMID:28307231 [本文引用: 1] ?Temporal and spatial variation in CH emissions was studied at hummock, Eriophorum lawn, flark and Carex lawn microsites in an oligotrophic pine fen over the growing season using a static chamber method, and CH production and oxidation potentials in peat profiles from hummock and flark were determined in laboratory incubation experiments. Emissions were lowest in the hummocks, and decreased with increasing hummock height, while in the lawns and flarks they increased with increasing sedge cover. Statistical response functions with water table and peat temperature as independent variables were calculated in order to reconstruct seasonal CH emissions by reference to the time series for peat temperature and water table specific to each microsite type. Mean CH emissions in the whole area in the snow-free period of 1993, weighted in terms of the proportions of the microsites, were 1.7 mol CH m. Potential CH production and oxidation rates were very low in the hummocks rising above the groundwater table, but were relatively similar when expressed per dry weight of peat both in the hummocks and flarks below the water table. The CH production potential increased in autumn at both microsites and CH oxidation potential seemed to decrease. The decrease in temperature in autumn certainly reduced in situ decomposition processes, possibly leaving unused substrates in the peat, which would explain the increase in CH production potential.
ShannonRD, WhiteJR, LawsonJE, GilmourBS (1996). Methane efflux from emergent vegetation in peatlands Journal of Ecology, 84,239-246. DOI:10.2307/2261359URL [本文引用: 1]
St?pniewskaZ, GorajW, Ku?niarA, Szafranek-NakoniecznaA, BanachA, GórskiA, PytlakA, UrbanD (2018). Methane oxidation by endophytic bacteria inhabiting Sphagnum sp. and some vascular plants Wetlands, 38,411-422. DOI:10.1007/s13157-017-0984-3URL [本文引用: 1]
StrakováP, NiemiRM, FreemanC, PeltoniemiK, TobermanH, HeiskanenI, FritzeH, LaihoR (2011). Litter type affects the activity of aerobic decomposers in a boreal peatland more than site nutrient and water table regimes Biogeosciences, 8,2741-2755. DOI:10.5194/bg-8-2741-2011URL [本文引用: 1]
SunXX, MuCC, ShiLY, ChengW, LiuX, WuYX, FengDJ (2009). Methane emission from forested swamps in Xiaoxing’an Mountains, northeastern China Chinese Journal of Plant Ecology, 33,535-545. [本文引用: 5]
TuretskyMR, KotowskaA, BubierJ, DiseNB, CrillP, HornibrookERC, MinkkinenK, MooreTR, Myers-SmithIH, Nyk?nenH, OlefeldtD, RinneJ, SaarnioS, ShurpaliN, TuittilaES, et al. (2014). A synthesis of methane emissions from 71 northern, temperate, and subtropical wetlands Global Change Biology, 20,2183-2197. DOI:10.1111/gcb.12580PMID:24777536 [本文引用: 2] Wetlands are the largest natural source of atmospheric methane. Here, we assess controls on methane flux using a database of approximately 19 000 instantaneous measurements from 71 wetland sites located across subtropical, temperate, and northern high latitude regions. Our analyses confirm general controls on wetland methane emissions from soil temperature, water table, and vegetation, but also show that these relationships are modified depending on wetland type (bog, fen, or swamp), region (subarctic to temperate), and disturbance. Fen methane flux was more sensitive to vegetation and less sensitive to temperature than bog or swamp fluxes. The optimal water table for methane flux was consistently below the peat surface in bogs, close to the peat surface in poor fens, and above the peat surface in rich fens. However, the largest flux in bogs occurred when dry 30-day averaged antecedent conditions were followed by wet conditions, while in fens and swamps, the largest flux occurred when both 30-day averaged antecedent and current conditions were wet. Drained wetlands exhibited distinct characteristics, e.g. the absence of large flux following wet and warm conditions, suggesting that the same functional relationships between methane flux and environmental conditions cannot be used across pristine and disturbed wetlands. Together, our results suggest that water table and temperature are dominant controls on methane flux in pristine bogs and swamps, while other processes, such as vascular transport in pristine fens, have the potential to partially override the effect of these controls in other wetland types. Because wetland types vary in methane emissions and have distinct controls, these ecosystems need to be considered separately to yield reliable estimates of global wetland methane release.? 2014 John Wiley & Sons Ltd.
TuretskyMR, TreatCC, WaldropMP, WaddingtonJM, HardenJW, McguireAD (2008). Short-term response of methane fluxes and methanogen activity to water table and soil warming manipulations in an Alaskan peatland Journal of Geophysical Research, 113,G00A10. DOI: 10.1029/2007JG000496. DOI:10.1029/2007JG000496 [本文引用: 1]
UrbanováZ, BártaJ, PicekT (2013). Methane emissions and methanogenic archaea on pristine, drained and restored mountain peatlands, central Europe Ecosystems, 16,664-677. DOI:10.1007/s10021-013-9637-4URL [本文引用: 1]
WaddingtonJM, DaySM (2007). Methane emissions from a peatland following restoration Journal of Geophysical Research: Biogeosciences, 112,G03018. DOI: 10.1029/2007JG000400. DOI:10.1029/2007JG000400 [本文引用: 1]
WaddingtonJM, KellnerE, StrackM, PriceJS (2010). Differential peat deformation, compressibility, and water storage between peatland microforms: implications for ecosystem function and development Water Resources Research, 46,W07538. DOI: 10.1029/2009WR008802. DOI:10.1029/2009WR008802 [本文引用: 1]
WangH, WuL, XueD, LiuXF, HongL, MouL, LiXL (2020). Distribution and environmental characteristics of Sphagnum peat bogs in Taishanmiao in Enshi City, Hubei Province Wetland Science, 18,266-274. [本文引用: 3]
WangXL, ZhangH, YaoZS, ZhengXH, ZhangSQ (2016). A preliminary study on methane emission from a seasonal- freezing alpine peat wetland during a non-growing period Climatic and Environmental Research, 21,282-292. [本文引用: 1]
WangYY, YangZF, YuT, WenYB, XiaXQ, BaiRJ (2011). Contrastive studies on different interpolation methods in soil carbon storage calculation in Da'an City, Jilin Province Carsologica Sinica, 30,479-486. [本文引用: 1]
WangZX, LeiY, LiuSX, FangYP, ManJS, PengZL, ZhangL, MaGL (2005). One subalpine Sphagnum wetland being discovered in Qizimei Mountains Nature Reserve, Hubei Journal of Central China Normal University(Natural Sciences), 39,387-388. [本文引用: 1]
XiaoSH, ZhangZR, QinDH, HeDJ, LiuJS, WuJQ, CaiCT, YouWB, ShenYQ, JianLY (2016). Community composition of four types of Sphagnum wetlands and its relationship with environmental factors in Tianbaoyan National Nature Reserve Chinese Journal of Applied and Environmental Biology, 22,631-638. [本文引用: 1]
YavittJB, YashiroE, Cadillo-QuirozH, ZinderSH (2012). Methanogen diversity and community composition in peatlands of the central to northern Appalachian Mountain region, North America Biogeochemistry, 109,117-131. DOI:10.1007/s10533-011-9644-5URL [本文引用: 1]
ZalmanC, KellerJK, TfailyM, KoltonM, Pfeifer-MeisterL, WilsonRM, LinX, ChantonJ, KostkaJE, GillA, FinziA, HoppleAM, BohannanBJM, BridghamSD (2018). Small differences in ombrotrophy control regional-scale variation in methane cycling among Sphagnum-dominated peatlands Biogeochemistry, 139,155-177. DOI:10.1007/s10533-018-0460-zURL [本文引用: 2]
ZhanQF (2006). Mosses' Stem Structure and Application of Stem Structure on Brachythecium taxonomy Master degree dissertation, East China Normal University, Shanghai. [本文引用: 1]
ZhangCF, ShengLX, GongC, HeCG, ZhangJ (2018). Effects of freeze-thaw cycles on soil microbial biomass carbon and carbon emissions from wetland soils, Northeast China Chinese Journal of Ecology, 37,304-311. [本文引用: 1]
Schematic diagram of methane emission flux of the Sphagnum bog in different regions. This comprehensive diagram is based on methane emission flux measured by Nykanen <i>et al.</i>(<xref ref-type="bibr" rid="b38">1998</xref>), Beetz <i>et al.</i>(<xref ref-type="bibr" rid="b4">2013</xref>), Hanson <i>et al.</i>(<xref ref-type="bibr" rid="b20">2016</xref>) and this study. Fig. 7 10.17521/cjpe.2020.0292.T0002
不同地区泥炭藓沼泽湿地CH4-C排放对比 ...
... (2013), Hanson et al.(2016) and this study. Fig. 7 10.17521/cjpe.2020.0292.T0002
不同地区泥炭藓沼泽湿地CH4-C排放对比 ...
Carbon cycling in peatlands—A review of processes and controls 1 2002
... 泥炭藓沼泽湿地是一类以泥炭藓属(Sphagnum)植物为优势物种的特殊湿地类型, 其强酸性、贫营养、潮湿和缺氧的环境为甲烷(CH4)生成提供了良好环境(Dedysh, 2002).研究表明, 北方泥炭藓沼泽湿地全年约排放 46 Tg CH4-C (Gorham, 1991), 是大气CH4的重要来源(Wuebbles & Hayhoe, 2002; Turetsky et al., 2014; Leroy et al., 2019).但是, 全球泥炭藓沼泽湿地CH4排放量存在很大的变异(Turetsky et al., 2014; Hanson et al., 2016).例如, 国外泥炭藓沼泽湿地平均CH4-C排放量通常为0.01-3.3 mg·m-2·h-1 (Blodau, 2002); 而我国东北泥炭藓沼泽湿地全年CH4-C排放量为0.026-0.38 mg·m-2·h-1 (孙晓新等, 2009; Miao et al., 2012; 常怡慧等, 2020).同时, 研究发现贫营养泥炭藓沼泽湿地CH4排放量通常小于富营养泥炭藓沼泽湿地(Brown, 1998; Chasar et al., 2000; B?ckstrandet al., 2010).甚至, 同一泥炭藓湿地不同微地形间CH4的排放也存在很大的差异(Frenzel & Karofeld, 2000; Beetz et al., 2013; Gill et al., 2017; Zalman et al., 2018).因此, 对不同地区不同生境泥炭藓沼泽湿地CH4排放特征及其影响因素的研究是准确评估全球泥炭藓沼泽湿地碳循环的关键. ...
Dynamics of methane fluxes from two peat bogs in the Ore Mountains, Czech Republic 1 2012
... 泥炭藓沼泽湿地是一类以泥炭藓属(Sphagnum)植物为优势物种的特殊湿地类型, 其强酸性、贫营养、潮湿和缺氧的环境为甲烷(CH4)生成提供了良好环境(Dedysh, 2002).研究表明, 北方泥炭藓沼泽湿地全年约排放 46 Tg CH4-C (Gorham, 1991), 是大气CH4的重要来源(Wuebbles & Hayhoe, 2002; Turetsky et al., 2014; Leroy et al., 2019).但是, 全球泥炭藓沼泽湿地CH4排放量存在很大的变异(Turetsky et al., 2014; Hanson et al., 2016).例如, 国外泥炭藓沼泽湿地平均CH4-C排放量通常为0.01-3.3 mg·m-2·h-1 (Blodau, 2002); 而我国东北泥炭藓沼泽湿地全年CH4-C排放量为0.026-0.38 mg·m-2·h-1 (孙晓新等, 2009; Miao et al., 2012; 常怡慧等, 2020).同时, 研究发现贫营养泥炭藓沼泽湿地CH4排放量通常小于富营养泥炭藓沼泽湿地(Brown, 1998; Chasar et al., 2000; B?ckstrandet al., 2010).甚至, 同一泥炭藓湿地不同微地形间CH4的排放也存在很大的差异(Frenzel & Karofeld, 2000; Beetz et al., 2013; Gill et al., 2017; Zalman et al., 2018).因此, 对不同地区不同生境泥炭藓沼泽湿地CH4排放特征及其影响因素的研究是准确评估全球泥炭藓沼泽湿地碳循环的关键. ...
... 目前, 有关泥炭藓沼泽湿地CH4排放的研究主要集中在北半球纬度较高的温带和寒温带地区(Chasar et al., 2000; Larmola et al., 2010; St?pniewskaet al., 2018).研究表明, 沼泽湿地内的泥炭藓体内有甲烷氧化细菌, 可氧化从泥炭地产生的CH4, 并为其光合作用提供碳源, 从而减少CH4排放(Raghoebarsing et al., 2005; Kip et al., 2010).但是, 在我国北方泥炭藓沼泽地中, 去除泥炭藓后CH4的排放反而减少了50% (Meng et al., 2014).由此可见, 植被对泥炭藓沼泽湿地CH4排放的影响是一个复杂过程, 其通常与温度、水位及其他环境因子发生耦合作用, 进一步影响CH4的排放(Mikkel?et al., 1995; 孙晓新等, 2009; Riutta et al., 2020).Nyk?nen等(1998)对芬兰泥炭藓沼泽湿地和李艳元等(2017)对神农架大九湖的研究均表明温度和土壤湿度会影响CH4排放; 而中欧有关泥炭藓沼泽湿地的研究表明地下水位对CH4排放的影响更为重要(Urbanováet al., 2013).还有研究发现, 落叶松-泥炭藓沼泽湿地CH4排放与土壤温度、土壤含水量等环境因子均不显著相关(常怡慧等, 2020).此外, 部分研究表明光对泥炭藓中微生物氮的分解和CH4的氧化有很强的促进作用, 光照条件下泥炭藓CH4的氧化速率比在黑暗条件下提高5倍, 而黑暗条件下CH4氧化活动较低导致CH4排放量随之升高(Kox et al., 2020).综上所述, 不同地区影响泥炭藓沼泽湿地CH4排放的关键因素不同.并且, 亚热带地区有关泥炭藓沼泽湿地CH4通量的研究鲜有报道(李艳元等, 2017; Alfadhel et al., 2019; 谌佳伟等, 2020). ...
大兴安岭永久冻土区7种沼泽类型土壤温室气体排放特征 2 2020
... 泥炭藓沼泽湿地是一类以泥炭藓属(Sphagnum)植物为优势物种的特殊湿地类型, 其强酸性、贫营养、潮湿和缺氧的环境为甲烷(CH4)生成提供了良好环境(Dedysh, 2002).研究表明, 北方泥炭藓沼泽湿地全年约排放 46 Tg CH4-C (Gorham, 1991), 是大气CH4的重要来源(Wuebbles & Hayhoe, 2002; Turetsky et al., 2014; Leroy et al., 2019).但是, 全球泥炭藓沼泽湿地CH4排放量存在很大的变异(Turetsky et al., 2014; Hanson et al., 2016).例如, 国外泥炭藓沼泽湿地平均CH4-C排放量通常为0.01-3.3 mg·m-2·h-1 (Blodau, 2002); 而我国东北泥炭藓沼泽湿地全年CH4-C排放量为0.026-0.38 mg·m-2·h-1 (孙晓新等, 2009; Miao et al., 2012; 常怡慧等, 2020).同时, 研究发现贫营养泥炭藓沼泽湿地CH4排放量通常小于富营养泥炭藓沼泽湿地(Brown, 1998; Chasar et al., 2000; B?ckstrandet al., 2010).甚至, 同一泥炭藓湿地不同微地形间CH4的排放也存在很大的差异(Frenzel & Karofeld, 2000; Beetz et al., 2013; Gill et al., 2017; Zalman et al., 2018).因此, 对不同地区不同生境泥炭藓沼泽湿地CH4排放特征及其影响因素的研究是准确评估全球泥炭藓沼泽湿地碳循环的关键. ...
... 目前, 有关泥炭藓沼泽湿地CH4排放的研究主要集中在北半球纬度较高的温带和寒温带地区(Chasar et al., 2000; Larmola et al., 2010; St?pniewskaet al., 2018).研究表明, 沼泽湿地内的泥炭藓体内有甲烷氧化细菌, 可氧化从泥炭地产生的CH4, 并为其光合作用提供碳源, 从而减少CH4排放(Raghoebarsing et al., 2005; Kip et al., 2010).但是, 在我国北方泥炭藓沼泽地中, 去除泥炭藓后CH4的排放反而减少了50% (Meng et al., 2014).由此可见, 植被对泥炭藓沼泽湿地CH4排放的影响是一个复杂过程, 其通常与温度、水位及其他环境因子发生耦合作用, 进一步影响CH4的排放(Mikkel?et al., 1995; 孙晓新等, 2009; Riutta et al., 2020).Nyk?nen等(1998)对芬兰泥炭藓沼泽湿地和李艳元等(2017)对神农架大九湖的研究均表明温度和土壤湿度会影响CH4排放; 而中欧有关泥炭藓沼泽湿地的研究表明地下水位对CH4排放的影响更为重要(Urbanováet al., 2013).还有研究发现, 落叶松-泥炭藓沼泽湿地CH4排放与土壤温度、土壤含水量等环境因子均不显著相关(常怡慧等, 2020).此外, 部分研究表明光对泥炭藓中微生物氮的分解和CH4的氧化有很强的促进作用, 光照条件下泥炭藓CH4的氧化速率比在黑暗条件下提高5倍, 而黑暗条件下CH4氧化活动较低导致CH4排放量随之升高(Kox et al., 2020).综上所述, 不同地区影响泥炭藓沼泽湿地CH4排放的关键因素不同.并且, 亚热带地区有关泥炭藓沼泽湿地CH4通量的研究鲜有报道(李艳元等, 2017; Alfadhel et al., 2019; 谌佳伟等, 2020). ...
Methane concentration and stable isotope distribution as evidence of rhizospheric processes: comparison of a fen and bog in the glacial Lake Agassiz Peatland complex 2 2000
... 泥炭藓沼泽湿地是一类以泥炭藓属(Sphagnum)植物为优势物种的特殊湿地类型, 其强酸性、贫营养、潮湿和缺氧的环境为甲烷(CH4)生成提供了良好环境(Dedysh, 2002).研究表明, 北方泥炭藓沼泽湿地全年约排放 46 Tg CH4-C (Gorham, 1991), 是大气CH4的重要来源(Wuebbles & Hayhoe, 2002; Turetsky et al., 2014; Leroy et al., 2019).但是, 全球泥炭藓沼泽湿地CH4排放量存在很大的变异(Turetsky et al., 2014; Hanson et al., 2016).例如, 国外泥炭藓沼泽湿地平均CH4-C排放量通常为0.01-3.3 mg·m-2·h-1 (Blodau, 2002); 而我国东北泥炭藓沼泽湿地全年CH4-C排放量为0.026-0.38 mg·m-2·h-1 (孙晓新等, 2009; Miao et al., 2012; 常怡慧等, 2020).同时, 研究发现贫营养泥炭藓沼泽湿地CH4排放量通常小于富营养泥炭藓沼泽湿地(Brown, 1998; Chasar et al., 2000; B?ckstrandet al., 2010).甚至, 同一泥炭藓湿地不同微地形间CH4的排放也存在很大的差异(Frenzel & Karofeld, 2000; Beetz et al., 2013; Gill et al., 2017; Zalman et al., 2018).因此, 对不同地区不同生境泥炭藓沼泽湿地CH4排放特征及其影响因素的研究是准确评估全球泥炭藓沼泽湿地碳循环的关键. ...
... 目前, 有关泥炭藓沼泽湿地CH4排放的研究主要集中在北半球纬度较高的温带和寒温带地区(Chasar et al., 2000; Larmola et al., 2010; St?pniewskaet al., 2018).研究表明, 沼泽湿地内的泥炭藓体内有甲烷氧化细菌, 可氧化从泥炭地产生的CH4, 并为其光合作用提供碳源, 从而减少CH4排放(Raghoebarsing et al., 2005; Kip et al., 2010).但是, 在我国北方泥炭藓沼泽地中, 去除泥炭藓后CH4的排放反而减少了50% (Meng et al., 2014).由此可见, 植被对泥炭藓沼泽湿地CH4排放的影响是一个复杂过程, 其通常与温度、水位及其他环境因子发生耦合作用, 进一步影响CH4的排放(Mikkel?et al., 1995; 孙晓新等, 2009; Riutta et al., 2020).Nyk?nen等(1998)对芬兰泥炭藓沼泽湿地和李艳元等(2017)对神农架大九湖的研究均表明温度和土壤湿度会影响CH4排放; 而中欧有关泥炭藓沼泽湿地的研究表明地下水位对CH4排放的影响更为重要(Urbanováet al., 2013).还有研究发现, 落叶松-泥炭藓沼泽湿地CH4排放与土壤温度、土壤含水量等环境因子均不显著相关(常怡慧等, 2020).此外, 部分研究表明光对泥炭藓中微生物氮的分解和CH4的氧化有很强的促进作用, 光照条件下泥炭藓CH4的氧化速率比在黑暗条件下提高5倍, 而黑暗条件下CH4氧化活动较低导致CH4排放量随之升高(Kox et al., 2020).综上所述, 不同地区影响泥炭藓沼泽湿地CH4排放的关键因素不同.并且, 亚热带地区有关泥炭藓沼泽湿地CH4通量的研究鲜有报道(李艳元等, 2017; Alfadhel et al., 2019; 谌佳伟等, 2020). ...
神农架大九湖泥炭湿地甲烷通量特征及其与土壤微生物群落组成的关系 9 2020
... 目前, 有关泥炭藓沼泽湿地CH4排放的研究主要集中在北半球纬度较高的温带和寒温带地区(Chasar et al., 2000; Larmola et al., 2010; St?pniewskaet al., 2018).研究表明, 沼泽湿地内的泥炭藓体内有甲烷氧化细菌, 可氧化从泥炭地产生的CH4, 并为其光合作用提供碳源, 从而减少CH4排放(Raghoebarsing et al., 2005; Kip et al., 2010).但是, 在我国北方泥炭藓沼泽地中, 去除泥炭藓后CH4的排放反而减少了50% (Meng et al., 2014).由此可见, 植被对泥炭藓沼泽湿地CH4排放的影响是一个复杂过程, 其通常与温度、水位及其他环境因子发生耦合作用, 进一步影响CH4的排放(Mikkel?et al., 1995; 孙晓新等, 2009; Riutta et al., 2020).Nyk?nen等(1998)对芬兰泥炭藓沼泽湿地和李艳元等(2017)对神农架大九湖的研究均表明温度和土壤湿度会影响CH4排放; 而中欧有关泥炭藓沼泽湿地的研究表明地下水位对CH4排放的影响更为重要(Urbanováet al., 2013).还有研究发现, 落叶松-泥炭藓沼泽湿地CH4排放与土壤温度、土壤含水量等环境因子均不显著相关(常怡慧等, 2020).此外, 部分研究表明光对泥炭藓中微生物氮的分解和CH4的氧化有很强的促进作用, 光照条件下泥炭藓CH4的氧化速率比在黑暗条件下提高5倍, 而黑暗条件下CH4氧化活动较低导致CH4排放量随之升高(Kox et al., 2020).综上所述, 不同地区影响泥炭藓沼泽湿地CH4排放的关键因素不同.并且, 亚热带地区有关泥炭藓沼泽湿地CH4通量的研究鲜有报道(李艳元等, 2017; Alfadhel et al., 2019; 谌佳伟等, 2020). ...
Schematic diagram of methane emission flux of the Sphagnum bog in different regions. This comprehensive diagram is based on methane emission flux measured by Nykanen <i>et al.</i>(<xref ref-type="bibr" rid="b38">1998</xref>), Beetz <i>et al.</i>(<xref ref-type="bibr" rid="b4">2013</xref>), Hanson <i>et al.</i>(<xref ref-type="bibr" rid="b20">2016</xref>) and this study. Fig. 7 10.17521/cjpe.2020.0292.T0002
不同地区泥炭藓沼泽湿地CH4-C排放对比 ...
... (2016) and this study. Fig. 7 10.17521/cjpe.2020.0292.T0002
Response of CH4 emissions to moss removal and N addition in boreal peatland of northeast China 2 2014
... 目前, 有关泥炭藓沼泽湿地CH4排放的研究主要集中在北半球纬度较高的温带和寒温带地区(Chasar et al., 2000; Larmola et al., 2010; St?pniewskaet al., 2018).研究表明, 沼泽湿地内的泥炭藓体内有甲烷氧化细菌, 可氧化从泥炭地产生的CH4, 并为其光合作用提供碳源, 从而减少CH4排放(Raghoebarsing et al., 2005; Kip et al., 2010).但是, 在我国北方泥炭藓沼泽地中, 去除泥炭藓后CH4的排放反而减少了50% (Meng et al., 2014).由此可见, 植被对泥炭藓沼泽湿地CH4排放的影响是一个复杂过程, 其通常与温度、水位及其他环境因子发生耦合作用, 进一步影响CH4的排放(Mikkel?et al., 1995; 孙晓新等, 2009; Riutta et al., 2020).Nyk?nen等(1998)对芬兰泥炭藓沼泽湿地和李艳元等(2017)对神农架大九湖的研究均表明温度和土壤湿度会影响CH4排放; 而中欧有关泥炭藓沼泽湿地的研究表明地下水位对CH4排放的影响更为重要(Urbanováet al., 2013).还有研究发现, 落叶松-泥炭藓沼泽湿地CH4排放与土壤温度、土壤含水量等环境因子均不显著相关(常怡慧等, 2020).此外, 部分研究表明光对泥炭藓中微生物氮的分解和CH4的氧化有很强的促进作用, 光照条件下泥炭藓CH4的氧化速率比在黑暗条件下提高5倍, 而黑暗条件下CH4氧化活动较低导致CH4排放量随之升高(Kox et al., 2020).综上所述, 不同地区影响泥炭藓沼泽湿地CH4排放的关键因素不同.并且, 亚热带地区有关泥炭藓沼泽湿地CH4通量的研究鲜有报道(李艳元等, 2017; Alfadhel et al., 2019; 谌佳伟等, 2020). ...
... 本研究区泥炭藓覆被样地CH4排放量大于裸露地, 与国外很多研究结论相反, 这可能与甲烷氧化细菌含量有关.研究表明, 多数泥炭藓属植物体中有甲烷氧化细菌(Raghoebarsing et al., 2005; Larmola et al., 2010), 泥炭藓沼泽湿地CH4排放量要小于裸露地(Waddington & Day, 2007; Korrensalo et al., 2018).然而, 研究发现不同泥炭藓物种甲烷氧化细菌含量不同(Zalman et al., 2018), 部分地区泥炭藓中甲烷氧化细菌多样性很低(Knoblaunch et al., 2008; 谌佳伟等, 2020).例如, 神农架大九湖湿地甲烷氧化细菌就很少, 且大九湖湿地环境与本研究区较为接近, 泥炭藓物种也相同(谌佳伟等, 2020).因此,本研究区泥炭藓中的甲烷氧化细菌含量可能也较少.另外, 研究区泥炭藓覆被样地CH4排放量大于裸露地可能与有机底物及地上生物量的限制有关(孙晓新等, 2009; Yavitt et al., 2012).研究表明, 裸露地缺少凋落物, 导致土壤微生物生长代谢的主要底物缺失, 同时土壤理化性质被改变, 降低CH4排放量(Strakováet al., 2011); 另一方面, 裸露地水位常年高于或接近地表, 泥炭中少量的CH4在穿过土壤水层时, 增加CH4被氧化的时间, 进一步减少CH4排放量(Kelley et al., 1995).Meng等(2014)在东北发现去除泥炭藓2年后, CH4排放量降低了50%, 本研究结论与之一致.与之相反, 多数研究表明维管植物入侵, 泥炭藓减少会增加泥炭藓沼泽湿地CH4排放量(Leroy et al., 2019).本研究中的覆被类型没有考虑维管植物, 但研究结论还是在一定程度上论证了覆被类型对泥炭藓沼泽湿地CH4排放的重要性.同时, 仍有必要研究更多植被类型对泥炭藓沼泽湿地CH4排放的影响, 这将有助于我们更加深刻认识亚热带亚高山泥炭藓湿地碳循环过程. ...
Growing season methane emission from a boreal peatland in the continuous permafrost zone of Northeast China: effects of active layer depth and vegetation 1 2012
... 泥炭藓沼泽湿地是一类以泥炭藓属(Sphagnum)植物为优势物种的特殊湿地类型, 其强酸性、贫营养、潮湿和缺氧的环境为甲烷(CH4)生成提供了良好环境(Dedysh, 2002).研究表明, 北方泥炭藓沼泽湿地全年约排放 46 Tg CH4-C (Gorham, 1991), 是大气CH4的重要来源(Wuebbles & Hayhoe, 2002; Turetsky et al., 2014; Leroy et al., 2019).但是, 全球泥炭藓沼泽湿地CH4排放量存在很大的变异(Turetsky et al., 2014; Hanson et al., 2016).例如, 国外泥炭藓沼泽湿地平均CH4-C排放量通常为0.01-3.3 mg·m-2·h-1 (Blodau, 2002); 而我国东北泥炭藓沼泽湿地全年CH4-C排放量为0.026-0.38 mg·m-2·h-1 (孙晓新等, 2009; Miao et al., 2012; 常怡慧等, 2020).同时, 研究发现贫营养泥炭藓沼泽湿地CH4排放量通常小于富营养泥炭藓沼泽湿地(Brown, 1998; Chasar et al., 2000; B?ckstrandet al., 2010).甚至, 同一泥炭藓湿地不同微地形间CH4的排放也存在很大的差异(Frenzel & Karofeld, 2000; Beetz et al., 2013; Gill et al., 2017; Zalman et al., 2018).因此, 对不同地区不同生境泥炭藓沼泽湿地CH4排放特征及其影响因素的研究是准确评估全球泥炭藓沼泽湿地碳循环的关键. ...
Diurnal variation in methane emission in relation to the water table, soil temperature, climate and vegetation cover in a Swedish acid mire 1 1995
... 目前, 有关泥炭藓沼泽湿地CH4排放的研究主要集中在北半球纬度较高的温带和寒温带地区(Chasar et al., 2000; Larmola et al., 2010; St?pniewskaet al., 2018).研究表明, 沼泽湿地内的泥炭藓体内有甲烷氧化细菌, 可氧化从泥炭地产生的CH4, 并为其光合作用提供碳源, 从而减少CH4排放(Raghoebarsing et al., 2005; Kip et al., 2010).但是, 在我国北方泥炭藓沼泽地中, 去除泥炭藓后CH4的排放反而减少了50% (Meng et al., 2014).由此可见, 植被对泥炭藓沼泽湿地CH4排放的影响是一个复杂过程, 其通常与温度、水位及其他环境因子发生耦合作用, 进一步影响CH4的排放(Mikkel?et al., 1995; 孙晓新等, 2009; Riutta et al., 2020).Nyk?nen等(1998)对芬兰泥炭藓沼泽湿地和李艳元等(2017)对神农架大九湖的研究均表明温度和土壤湿度会影响CH4排放; 而中欧有关泥炭藓沼泽湿地的研究表明地下水位对CH4排放的影响更为重要(Urbanováet al., 2013).还有研究发现, 落叶松-泥炭藓沼泽湿地CH4排放与土壤温度、土壤含水量等环境因子均不显著相关(常怡慧等, 2020).此外, 部分研究表明光对泥炭藓中微生物氮的分解和CH4的氧化有很强的促进作用, 光照条件下泥炭藓CH4的氧化速率比在黑暗条件下提高5倍, 而黑暗条件下CH4氧化活动较低导致CH4排放量随之升高(Kox et al., 2020).综上所述, 不同地区影响泥炭藓沼泽湿地CH4排放的关键因素不同.并且, 亚热带地区有关泥炭藓沼泽湿地CH4通量的研究鲜有报道(李艳元等, 2017; Alfadhel et al., 2019; 谌佳伟等, 2020). ...
Methane fluxes on boreal peatlands of different fertility and the effect of long-term experimental lowering of the water table on flux rates 5 1998
... 目前, 有关泥炭藓沼泽湿地CH4排放的研究主要集中在北半球纬度较高的温带和寒温带地区(Chasar et al., 2000; Larmola et al., 2010; St?pniewskaet al., 2018).研究表明, 沼泽湿地内的泥炭藓体内有甲烷氧化细菌, 可氧化从泥炭地产生的CH4, 并为其光合作用提供碳源, 从而减少CH4排放(Raghoebarsing et al., 2005; Kip et al., 2010).但是, 在我国北方泥炭藓沼泽地中, 去除泥炭藓后CH4的排放反而减少了50% (Meng et al., 2014).由此可见, 植被对泥炭藓沼泽湿地CH4排放的影响是一个复杂过程, 其通常与温度、水位及其他环境因子发生耦合作用, 进一步影响CH4的排放(Mikkel?et al., 1995; 孙晓新等, 2009; Riutta et al., 2020).Nyk?nen等(1998)对芬兰泥炭藓沼泽湿地和李艳元等(2017)对神农架大九湖的研究均表明温度和土壤湿度会影响CH4排放; 而中欧有关泥炭藓沼泽湿地的研究表明地下水位对CH4排放的影响更为重要(Urbanováet al., 2013).还有研究发现, 落叶松-泥炭藓沼泽湿地CH4排放与土壤温度、土壤含水量等环境因子均不显著相关(常怡慧等, 2020).此外, 部分研究表明光对泥炭藓中微生物氮的分解和CH4的氧化有很强的促进作用, 光照条件下泥炭藓CH4的氧化速率比在黑暗条件下提高5倍, 而黑暗条件下CH4氧化活动较低导致CH4排放量随之升高(Kox et al., 2020).综上所述, 不同地区影响泥炭藓沼泽湿地CH4排放的关键因素不同.并且, 亚热带地区有关泥炭藓沼泽湿地CH4通量的研究鲜有报道(李艳元等, 2017; Alfadhel et al., 2019; 谌佳伟等, 2020). ...
Schematic diagram of methane emission flux of the Sphagnum bog in different regions. This comprehensive diagram is based on methane emission flux measured by Nykanen <i>et al.</i>(<xref ref-type="bibr" rid="b38">1998</xref>), Beetz <i>et al.</i>(<xref ref-type="bibr" rid="b4">2013</xref>), Hanson <i>et al.</i>(<xref ref-type="bibr" rid="b20">2016</xref>) and this study. Fig. 7 10.17521/cjpe.2020.0292.T0002
不同地区泥炭藓沼泽湿地CH4-C排放对比 ...
... (1998), Beetz et al.(2013), Hanson et al.(2016) and this study. Fig. 7 10.17521/cjpe.2020.0292.T0002
不同地区泥炭藓沼泽湿地CH4-C排放对比 ...
... CH4-C emission fluxes from the Sphagnum bog in different regions Table 2
Methane emissions and methanogenic archaea on pristine, drained and restored mountain peatlands, central Europe 1 2013
... 目前, 有关泥炭藓沼泽湿地CH4排放的研究主要集中在北半球纬度较高的温带和寒温带地区(Chasar et al., 2000; Larmola et al., 2010; St?pniewskaet al., 2018).研究表明, 沼泽湿地内的泥炭藓体内有甲烷氧化细菌, 可氧化从泥炭地产生的CH4, 并为其光合作用提供碳源, 从而减少CH4排放(Raghoebarsing et al., 2005; Kip et al., 2010).但是, 在我国北方泥炭藓沼泽地中, 去除泥炭藓后CH4的排放反而减少了50% (Meng et al., 2014).由此可见, 植被对泥炭藓沼泽湿地CH4排放的影响是一个复杂过程, 其通常与温度、水位及其他环境因子发生耦合作用, 进一步影响CH4的排放(Mikkel?et al., 1995; 孙晓新等, 2009; Riutta et al., 2020).Nyk?nen等(1998)对芬兰泥炭藓沼泽湿地和李艳元等(2017)对神农架大九湖的研究均表明温度和土壤湿度会影响CH4排放; 而中欧有关泥炭藓沼泽湿地的研究表明地下水位对CH4排放的影响更为重要(Urbanováet al., 2013).还有研究发现, 落叶松-泥炭藓沼泽湿地CH4排放与土壤温度、土壤含水量等环境因子均不显著相关(常怡慧等, 2020).此外, 部分研究表明光对泥炭藓中微生物氮的分解和CH4的氧化有很强的促进作用, 光照条件下泥炭藓CH4的氧化速率比在黑暗条件下提高5倍, 而黑暗条件下CH4氧化活动较低导致CH4排放量随之升高(Kox et al., 2020).综上所述, 不同地区影响泥炭藓沼泽湿地CH4排放的关键因素不同.并且, 亚热带地区有关泥炭藓沼泽湿地CH4通量的研究鲜有报道(李艳元等, 2017; Alfadhel et al., 2019; 谌佳伟等, 2020). ...
Methane emissions from a peatland following restoration 1 2007
... 本研究区泥炭藓覆被样地CH4排放量大于裸露地, 与国外很多研究结论相反, 这可能与甲烷氧化细菌含量有关.研究表明, 多数泥炭藓属植物体中有甲烷氧化细菌(Raghoebarsing et al., 2005; Larmola et al., 2010), 泥炭藓沼泽湿地CH4排放量要小于裸露地(Waddington & Day, 2007; Korrensalo et al., 2018).然而, 研究发现不同泥炭藓物种甲烷氧化细菌含量不同(Zalman et al., 2018), 部分地区泥炭藓中甲烷氧化细菌多样性很低(Knoblaunch et al., 2008; 谌佳伟等, 2020).例如, 神农架大九湖湿地甲烷氧化细菌就很少, 且大九湖湿地环境与本研究区较为接近, 泥炭藓物种也相同(谌佳伟等, 2020).因此,本研究区泥炭藓中的甲烷氧化细菌含量可能也较少.另外, 研究区泥炭藓覆被样地CH4排放量大于裸露地可能与有机底物及地上生物量的限制有关(孙晓新等, 2009; Yavitt et al., 2012).研究表明, 裸露地缺少凋落物, 导致土壤微生物生长代谢的主要底物缺失, 同时土壤理化性质被改变, 降低CH4排放量(Strakováet al., 2011); 另一方面, 裸露地水位常年高于或接近地表, 泥炭中少量的CH4在穿过土壤水层时, 增加CH4被氧化的时间, 进一步减少CH4排放量(Kelley et al., 1995).Meng等(2014)在东北发现去除泥炭藓2年后, CH4排放量降低了50%, 本研究结论与之一致.与之相反, 多数研究表明维管植物入侵, 泥炭藓减少会增加泥炭藓沼泽湿地CH4排放量(Leroy et al., 2019).本研究中的覆被类型没有考虑维管植物, 但研究结论还是在一定程度上论证了覆被类型对泥炭藓沼泽湿地CH4排放的重要性.同时, 仍有必要研究更多植被类型对泥炭藓沼泽湿地CH4排放的影响, 这将有助于我们更加深刻认识亚热带亚高山泥炭藓湿地碳循环过程. ...
Differential peat deformation, compressibility, and water storage between peatland microforms: implications for ecosystem function and development 1 2010
... 本研究对比分析3种覆被类型泥炭藓沼泽湿地CH4排放量与气温和地下5 cm土壤温度关系, 发现两者均呈显著正相关关系(p <0.05, 图4, 图5), 且沼泽湿地CH4的排放量随着温度增加呈指数增加的趋势, 与以往多数研究(Crill et al., 1988; Bohn et al., 2007; 李艳元等, 2017)结论一致.Gill等(2017)对泥炭藓沼泽湿地不同增温处理的研究表明, 泥炭地对气候变暖很敏感, 全球变暖可能增加泥炭藓湿地CH4排放量; 主要是温度升高和温度波动可能改变泥炭基质的分压差, 增加泥炭地气体沸腾, 从而绕过CH4氧化的富氧区, 促进气体逸出(Beckmann et al., 2004; Waddington et al., 2010; Gill et al., 2017); 另一方面, 温度升高促进沼泽湿地水面蒸发, 间接加速CH4的排放(Saarnio et al., 1997).因此, 温度是影响本研究区CH4排放的一个关键环境因子. ...
 ,*, 刘雪飞, 李小玲, 王涵, 吴浩, 余玉蓉, 杜胜蓝
,*, 刘雪飞, 李小玲, 王涵, 吴浩, 余玉蓉, 杜胜蓝

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT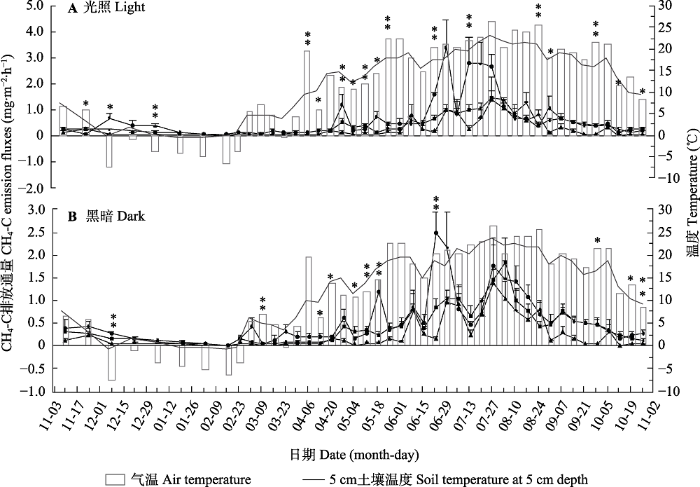 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT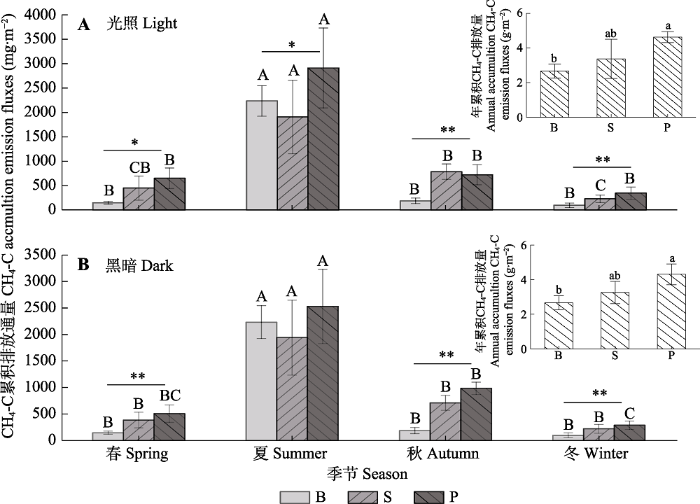 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT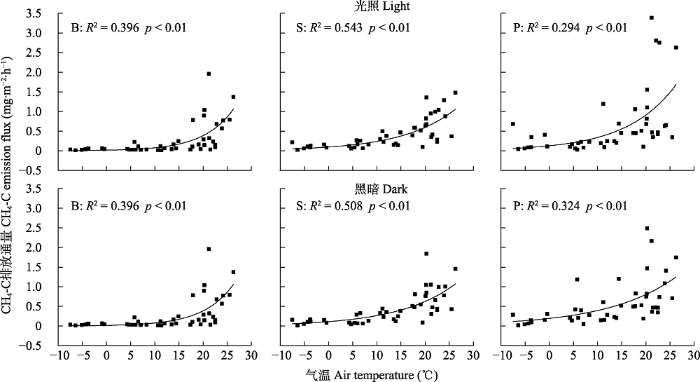 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT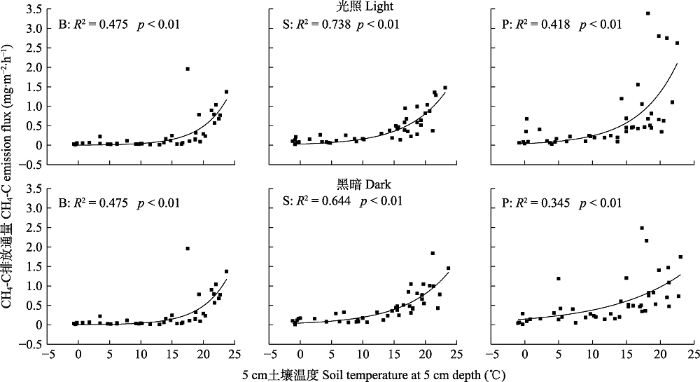 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT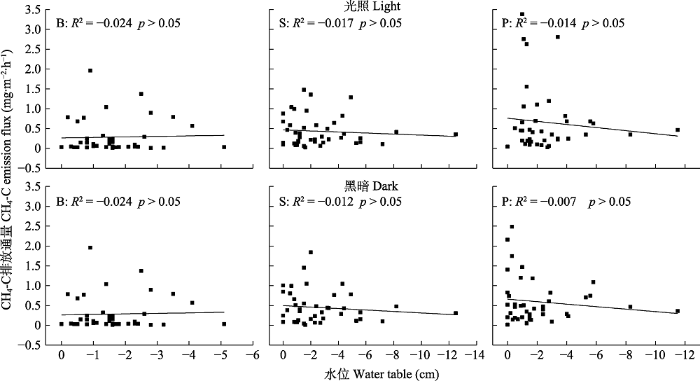 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT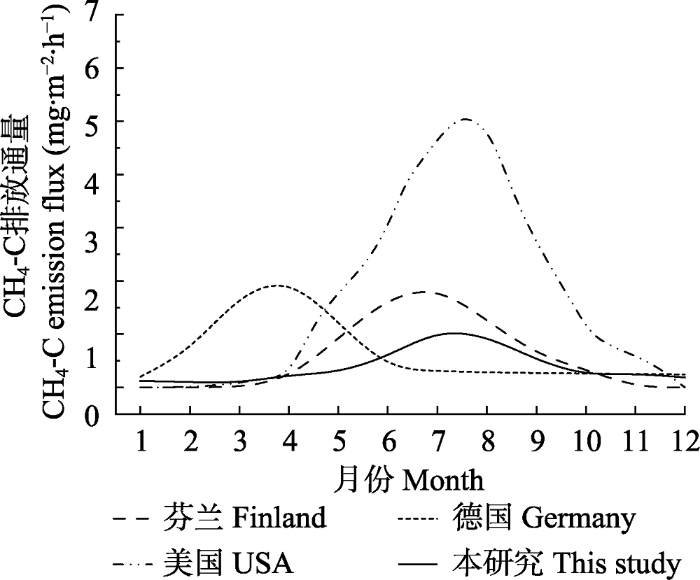 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT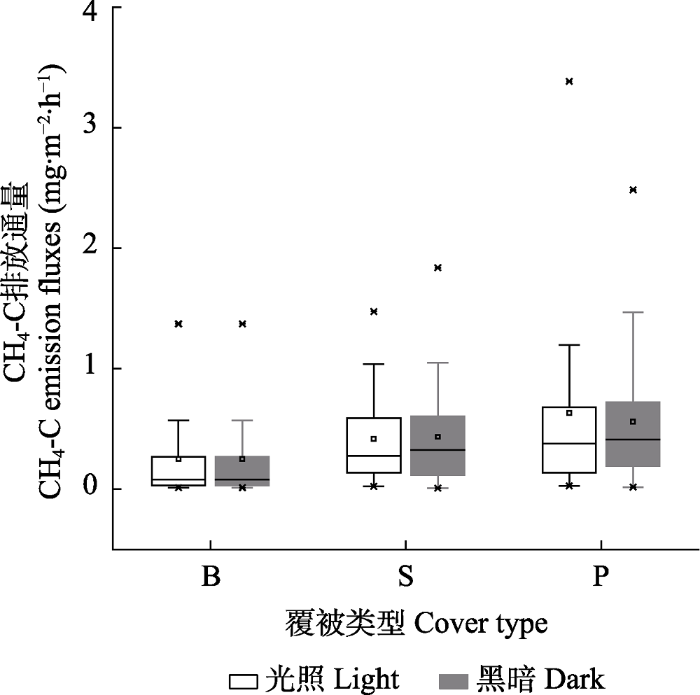 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}