 ,1,2,*, 胡宝清1,2
,1,2,*, 胡宝清1,2Effects of long-term human disturbances on soil microbial diversity and community structure in a karst grassland ecosystem of northwestern Guangxi, China
Guang-Ting PEI1,2, Jian-Fei SUN1,2, Tong-Xin HE,1,2,*, Bao-Qing HU1,2通讯作者: *(htx@nnnu.edu.cn)
编委: 张朝晖
责任编辑: 李敏
收稿日期:2020-09-21接受日期:2020-11-24网络出版日期:2021-01-20
| 基金资助: |
Corresponding authors: *(htx@nnnu.edu.cn)
Received:2020-09-21Accepted:2020-11-24Online:2021-01-20
| Fund supported: |

摘要
人为干扰是喀斯特草地正向演替的主要障碍因素, 从土壤微生物角度探讨草地对干扰的响应可为喀斯特生态恢复与科学利用提供依据。该研究依托中国科学院环江喀斯特生态系统观测站长期干扰监测样地, 通过高通量测序测定3个干扰模式(火烧、刈割、刈割除根)和1个对照(封育)处理的土壤细菌、真菌α多样性和群落结构, 并分析它们与环境因子(坡位、土壤理化性质)的关系, 揭示人为干扰对桂西北喀斯特草地土壤微生物的影响及机制。结果显示: 1)对于α多样性, 在中坡和下坡, 火烧均显著降低了真菌Chao1指数, 刈割均显著降低了细菌的香农指数和谱系多样性指数; 刈割除根在中、下坡分别显著降低了真菌Chao1指数和细菌香农指数。2)对于群落结构, 火烧、刈割和刈割除根在中、下坡均显著降低了酸杆菌门细菌相对丰度, 火烧在下坡使子囊菌门真菌相对丰度从74.49%显著降低至34.72%。3)冗余分析显示, 微生物生物量碳含量解释细菌和真菌α多样性变化的29.8%和26.8%以及细菌群落结构变化的31.7%; 细根生物量解释细菌α多样性和真菌群落结构变化的13.9%和10.3%。可见, 长期人为干扰对土壤微生物α多样性产生负面影响, 并显著改变微生物群落结构, 影响程度因干扰方式、微生物种类不同而异, 且受坡位调控。土壤微生物生物量碳含量和细根生物量的减少是人为干扰影响土壤微生物多样性和群落结构的重要因素。α多样性及子囊菌的减少将不利于土壤生态系统稳定性的维持, 酸杆菌的减少将不利于土壤有机质降解和铁循环。因此, 长期火烧、刈割等人为干扰将导致喀斯特草地生态系统功能发生退化。
关键词:
Abstract
Aims Human disturbance is one of the main obstacles to the forward succession of karst grassland, exploring the response of grassland to disturbance in terms of soil microorganism can provide the basis for the restoration and rational utilization of karst land. Our objective was to study the effects of different human disturbances on soil microorganisms and the underlying mechanisms in a karst grassland ecosystem of northwestern Guangxi, China.
Methods Three patterns of disturbances (burning, mowing, and mowing plus root removal) and one control treatment (enclosure) were conducted at the long-term monitoring plots in the Huanjiang Observation and Research Station for Karst Ecosystems, Chinese Academy of Sciences. We analyzed the changes of soil microbial diversity and community structure by high-throughput sequencing, and determined their relationships with environmental factors (slope position, soil physicochemical properties).
Important findings 1) For α diversity, at both middle and lower slope positions, the burning treatment significantly reduced the fungal Chao1 index, while the mowing treatment significantly reduced the bacterial Shannon index and Pedigree diversity index. However, the mowing plus root removal treatment significantly reduced the fungal Chao1 index and the bacterial Shannon index, respectively, at middle and lower slope positions. 2) For microbial community structure, burning, mowing and mowing plus root removal treatments significantly reduced the relative abundance of Acidobacteria at both middle and lower slope positions, while the fire treatment significantly reduced the relative abundance of Ascomycota from 74.49% to 34.72% at the lower slope position. 3) Redundancy analysis showed that soil microbial biomass carbon explained 29.8% and 26.8% of the changes of bacterial and fungal α diversity, respectively, and 31.7% of the changes of bacterial community structure. Root biomass explained 13.9% and 10.3% of the changes of bacterial α diversity and fungal community structure, respectively. In conclusion, the three studied human disturbances have significantly negative influence on soil microbial α diversity as well as having a significant change in and changed community structure, and the degree of influence varied among the pattern of disturbances and the type of microorganisms. Moreover, the effects were also regulated by slope position. Long-term human disturbances mainly affected the diversity and structure of soil microbial communities by changing soil microbial biomass carbon and root biomass. The decreases of α diversity and Ascomycota will not be conducive to the maintenance of soil ecosystem stability, and the decrease of Acidobacteria will not facilitate to soil organic matter degradation and iron cycling. Therefore, the long-term human disturbances such as burning and mowing will induce the functional degradation of grassland ecosystem.
Keywords:
PDF (1382KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
裴广廷, 孙建飞, 贺同鑫, 胡宝清. 长期人为干扰对桂西北喀斯特草地土壤微生物多样性及群落结构的影响. 植物生态学报, 2021, 45(1): 74-84. DOI: 10.17521/cjpe.2020.0316
PEI Guang-Ting, SUN Jian-Fei, HE Tong-Xin, HU Bao-Qing.

中国的喀斯特面积居于世界首位, 其裸露面积约为90.7万km2, 约占国土总面积的10% (宋同清等, 2014)。西南地区是我国喀斯特最为集中分布区, 是典型的生态环境脆弱区(滕秋梅等, 2020), 其生态环境容量小, 人地矛盾尖锐, 对人类活动的影响十分敏感, 不合理的人类活动极易导致水土流失, 引发石漠化等生态问题(黄玙璠等, 2020)。火烧、刈割和放牧是喀斯特地区较为常见的人为干扰方式, 对石漠化恢复初期草地生态系统的稳定和演替产生严重威胁(陈香碧等, 2012; 许留兴等, 2015)。
土壤微生物是土壤养分循环的驱动者, 它控制着土壤有机质的分解与形成、碳氮循环等过程(Fontúrbelet al., 2012), 在生态系统功能和服务中发挥着重要作用(Philippot et al., 2013)。已有研究表明, 生物多样性越高, 生态系统越可能趋于稳定(Finlay et al., 1997; Maron et al., 2018; Morriss et al., 2020); 相反, 生物多样性的减少将不利于生态系统的可持续发展(Naeem et al., 2000)。细菌和真菌是微生物群落中的主要成员, 两者中一些特定的类群能分别完成有机质分解、硝化和反硝化等生态过程(Bugg et al., 2011; Philippot et al., 2013), 因此人为干扰下微生物群落结构的改变将影响生态系统过程与功能的正常进行。因此, 研究土壤微生物多样性及群落结构对人为干扰的响应具有重要的生态学意义。
土壤微生物群落对环境条件的改变较为敏感, 温度、水分、pH、养分等理化性质与微生物的生长和代谢紧密相关(陶玉柱和邸雪颖, 2013)。有研究发现, 火烧可提高或降低土壤含水量、pH和根系生物量等土壤性质进而刺激或抑制微生物群落多样性(Vázquezet al., 1993; Huffman & Madritch, 2018)。刈割和放牧则主要通过减少地上植被有机质输入来改变微生物多样性和群落结构(谭红妍, 2015)。由于干扰作用的机制不同, 三者对微生物多样性和群落结构的影响也可能不尽相同。另外, 微生物对人为干扰的响应还可能与植被类型、地形等环境因子相关(Huffman & Madritch, 2018)。在西南喀斯特地区, 关于火烧、刈割、放牧等人为干扰方式方面的研究主要集中在地上植被部分(徐丽丽等, 2008; 向凯旋, 2019), 而关于人为干扰对地下土壤细菌和真菌多样性及其群落结构的影响还少有研究, 其驱动机制是什么, 也尚未清楚。
本研究以桂西北喀斯特草地生态系统为研究对象, 设置火烧、刈割、刈割除根3种不同的人为干扰方式, 采用高通量测序技术(基于16S rRNA和ITS基因分析), 通过测定细菌和真菌多样性指数及群落结构的变化, 并分析它们与环境因子之间的关系, 探究不同干扰方式对土壤微生物群落的影响及其内在机制, 以期为喀斯特生态系统的恢复与合理利用提供科学依据。
1 材料和方法
1.1 研究区概况
长期干扰监测样地位于桂西北环江毛南族自治县中国科学院环江喀斯特生态系统观测研究站的木连综合试验示范区(24.72°-24.73° N, 108.30°- 108.32° E)(陈洪松等,2012)。研究区属于中亚热带季风气候, 年降水量为1 389 mm, 干湿季节明显(雨季: 4-9月, 旱季: 10-3月), 70%以上降水集中在雨季; 年平均气温为19.9 ℃, 极端高温38.7 ℃, 极端低温-5.2 ℃, 全年无霜期为300-330天(刘艳等, 2014)。地貌类型为典型的喀斯特峰丛洼地, 土壤类型为白云岩发育的棕色石灰土, 沿坡向下土层平均厚度由10-30 cm增加到50-80 cm (陈洪松等, 2012; 刘艳等, 2014)。1985年之前研究区生态系统经历着频繁的火烧、刈割和放牧, 1985年研究区大部分居民外迁, 退化生态系统得以逐渐恢复, 植被类型以荒草地和稀疏灌丛为主(陈洪松等, 2012)。1.2 样地设置和样品采集
2004年底, 选择土壤和植被较为均一且坡向、坡度条件基本相同的一面东南方向山坡, 鉴于喀斯特山坡中下部被当地居民利用率较高(陈洪松等, 2012; 刘艳等, 2014), 按照表1在山坡中下部建立火烧、刈割、刈割除根和封育(对照) 4种处理模式。Table 1
表1
表1桂西北喀斯特草地生态系统样地基本特征
Table 1
| 干扰模式 Disturbance type | 坡向 Slope aspect | 平均坡度 Mean slope angle (°) | 处理方法 Treatment process | 植被现状 Vegetation status |
|---|---|---|---|---|
| 封育 Enclosure | SE | 33.0 | 保留原始植被, 作为对照 Control plot with native vegetation | 以灌木为主, 并伴有大量草本分布 (由于自然恢复多年, 植被类型已由草本逐渐转变为灌木), 主要植被为黄荆、楤木、火棘、紫穗槐、小花梾木等。 The vegetation are dominated by shrubs and accompanied by numberous herbs (due to many years after natural recovery, the vegetation type has gradually changed from herbs to shrubs), and the dominant species are Vitex negundo, Aralia chinensis, Pyracantha fortuneana, Amorpha fruticosa,and Swida parviflora,etc. |
| 火烧 Burning | SE | 33.7 | 每年一月火烧一次 Burning in January of each year | 中坡位以草本为主, 下坡位以草灌为主, 主要植被为白茅、蔓生莠竹、毛桐苗、紫穗槐苗等。 The middle slope position are dominated by herbs and the lower slope position are dominated by herb-shrub plants. The dominant species are Imperata cylindrical, Microstegium vagans, Mallotus barbatus,and Amorpha fruticosa, etc. |
| 刈割 Mowing | SE | 34.5 | 每年一月砍伐地上植被, 搬移出样地, 不去除植物根系 Cutting aboveground plants without root removal in January of each year | 中坡位以草本为主, 下坡位以草灌为主, 主要植被为白茅、蔓生莠竹、紫穗槐苗等。 The middle slope position are dominated by herbs and the lower slope position are dominated by herb-shrub plants. The dominant species are Imperata cylindrical, Microstegium vagans, Amorpha fruticosa,etc. |
| 刈割除根 Mowing plus root removal | SE | 33.5 | 每年一月砍伐地上植被, 搬移出样地, 且去除植物根系 Cutting aboveground plants and removing roots in January of each year | 以草本为主, 主要植被为白茅、蔓生莠竹等。 The vegetation are dominated by herbs, and the dominant species are Imperata cylindrical, Microstegium vagans,etc. |
新窗口打开|下载CSV
2017年6月(长期干扰处理已有12年), 分别在每种处理模式下的中坡和下坡位置随机建立3个5 m × 10 m的样方, 共计24个样方(4个处理×2个坡位×3个重复)。为了减少喀斯特土壤分布高异质性的影响, 每个样方内按“S”形5点取样法采集0-10 cm表层土壤, 充分混合成一份土壤样品, 累计土壤采样点数为120个, 最终获取土壤样品数24份; 根钻(直径7 cm)随机采集每个样方内3个植物根系样品, 充分混合成一份根系样品, 累计根系采样点数为72个, 最终获取根系样品数24份(贺同鑫等, 2020)。
1.3 样品处理
土壤样品过2 mm筛, 挑出植物根系和石砾, 分成3份。一部分用于测定土壤干基含水量、微生物生物量碳(MBC)和微生物生物量氮(MBN)含量; 另一部分保存于-20 ℃, 用于高通量测序分析。最后一部分风干, 用于测定土壤pH、有机碳(SOC)和总氮(TN)含量。根系样品用去离子水洗净附着的土壤, 将活的≤2 mm细根挑出, 65 ℃烘干, 称质量, 计算植物细根生物量。
1.4 土壤理化性质的测定
土壤含水量采用烘干称质量法(105 ℃, 24 h)测定; 土壤pH采用电极电位法(土水质量比为1:2.5)测定(李伟峰等, 2017; 李阳等, 2019)。土壤SOC含量采用酸洗法(去除无机碳)测定(贺同鑫等, 2020)。
土壤TN含量采用元素分析仪(Isoprime vario ISOTOPE cube, Elementar, Frankfurt, Germany)测定(贺同鑫等, 2020)。
土壤MBC和MBN含量采用氯仿熏蒸-K2SO4浸提法测定(Vance et al., 1987), 最后用TOC分析仪(multi N/C 2100, Analytik Jena GmbH, Jena, Germany)测定总有机碳和总有机氮含量, 以计算微生物生物量碳、氮含量。
1.5 高通量测序分析
土壤DNA的提取、PCR扩增和HiSeq测序均由北京诺禾致源科技股份有限公司(www.novogene. com)完成。基因组DNA的提取采用CTAB或SDS法。PCR扩增的引物包括16S V4区515F-806R, 18S V4区528F-706R, 18S V9区1380F-1510R, ITS1区ITS1FITS2, ITS2区ITS3-ITS4; PCR反应体系为Phusion Master Mix (2×) 15 μL, Primer (2 μmol·mol-1) 3 μL, gDNA (1 ng·μL -1) 10 μL, H 2O 2 μL; 反应程序为98 ℃预变性1 min, 30个循环(98 ℃, 10 s; 50 ℃, 30 s; 72 ℃, 30 s)。最后使用HiSeq测序, 测序得到原始序列(Raw Data), 再进行拼接、过滤, 得到有效序列(Clean Data), 根据97%相似水平, 对序列进行聚类得到操作分类单(OTU)。1.6 数据统计和分析
微生物生物量碳、氮含量计算公式:式中, M(C,N)为微生物生物量碳或氮含量(mg·kg-1); ?E(C,N)为熏蒸处理减去未熏蒸处理有机碳或氮含量(mg·kg-1); k(C,N)为换算系数, 微生物生物量碳和氮分别取值0.45和0.54 (Brookes et al., 1985; Vance et al., 1987)。
使用R语言4.2中的‘vegan’包, 基于OTU和进化树计算微生物的α多样性指数: Chao1指数、香农指数、谱系多样性指数。Chao1指数代表微生物的物种丰富度, 香农指数代表微生物的物种均匀度, 谱系多样性指数代表微生物的遗传多样性。用非度量多维尺度分析(NMDS)分析不同干扰方式对微生物群落结构的影响; 并采用基于Bray-Curtis距离的多元方差分析检验不同处理和不同坡位之间是否存在显著差异。用冗余分析(RDA)分析微生物多样性和群落结构与土壤理化性质之间的关系。用单因素方差分析和最小显著差异(LSD)分析土壤微生物多样性在不同干扰方式之间是否存在显著差异。方差分析在SPSS 18.0中完成。
2 实验结果
2.1 不同干扰方式对土壤理化性质的影响
相比封育处理, 火烧、刈割、刈割除根3种干扰方式对土壤含水量、pH和SOC含量均无显著影响(p > 0.05, 表2)。刈割在中坡位使土壤TN含量显著增加了21.96% (p < 0.05, 表2), 其他干扰处理对土壤TN含量均无显著影响。然而, 无论是在中坡位还是在下坡位, 3种干扰方式均显著降低了土壤MBC含量(p < 0.05, 表2), 且火烧降低幅度最大(在中下坡位分别比封育降低了73.56%和72.73%), 但是它们对土壤MBN含量无显著影响。同样的, 3种干扰方式均降低了土壤细根生物量, 火烧在下坡位使细根生物量显著降低了62.81% (p < 0.05, 表2)。Table 2
表2
表2不同干扰方式对桂西北喀斯特草地生态系统土壤理化性质的影响(平均值±标准误)
Table 2
| 土壤性质 Soil property | 中坡位 Middle slope position | 下坡位 Lower slope position | ||||||
|---|---|---|---|---|---|---|---|---|
| 封育 Enclosure | 火烧 Burning | 刈割 Mowing | 刈割除根 Mowing plus root removal | 封育 Enclosure | 火烧 Burning | 刈割 Mowing | 刈割除根 Mowing plus root removal | |
| 含水量 Moisture (%) | 40.49 ± 1.71 a | 42.32 ± 0.44 a | 42.99 ± 2.12 a | 38.08 ± 1.28 a | 48.41 ± 1.78 a | 46.10 ± 2.84 a | 44.22 ± 4.98 a | 46.57 ± 2.54 a |
| pH | 7.81 ± 0.04 a | 7.88 ± 0.07 a | 7.93 ± 0.05 a | 7.98 ± 0.05 a | 7.69 ± 0.10 a | 7.73 ± 0.04 a | 7.90 ± 0.08 a | 7.82 ± 0.03 a |
| 有机碳含量 Soil organic carbon content (g·kg-1) | 53.89 ± 3.26 ab | 62.31 ± 3.02 a | 62.67 ± 4.06 a | 50.61 ± 1.34 b | 65.24 ± 4.84 a | 47.11 ± 5.83 a | 61.69 ± 8.92 a | 58.70 ± 4.31 a |
| 总氮含量 Total nitrogen content (g·kg-1) | 3.78 ± 0.19 b | 4.44 ± 0.31 ab | 4.61 ± 0.17 a | 4.00 ± 0.13 ab | 5.88 ± 0.59 a | 4.80 ± 0.49 a | 4.89 ± 0.89 a | 5.09 ± 0.35 a |
| 碳氮比 Carbon-nitrogen ratio | 14.29 ± 0.86 a | 14.07 ± 0.35 a | 13.56 ± 0.48 a | 12.68 ± 0.59 a | 11.14 ± 0.34 b | 9.77 ± 0.34 c | 12.78 ± 0.46 a | 11.54 ± 0.34 b |
| 微生物生物量碳含量 Microbial biomass carbon content (g·kg-1) | 0.87 ± 0.08 a | 0.23 ± 0.02 b | 0.24 ± 0.03 b | 0.44 ± 0.11 b | 1.00 ± 0.04 a | 0.27 ± 0.04 b | 0.48 ± 0.09 b | 0.42 ± 0.14 b |
| 微生物生物量氮含量 Microbial biomass nitrogen content (g·kg-1) | 0.25 ± 0.02 a | 0.19 ± 0.02 a | 0.18 ± 0.04 a | 0.22 ± 0.03 a | 0.25 ± 0.04 ab | 0.20 ± 0.03 b | 0.34 ± 0.02 a | 0.20 ± 0.07 ab |
| 细根生物量 Fine root biomass (g·m-2) | 127.82 ± 16.23 a | 102.61 ± 5.59 a | 85.37 ± 22.03 a | 68.14 ± 25.27 a | 132.79 ± 39.01 a | 49.38 ± 12.04 b | 126.51 ± 11.93 a | 77.42 ± 3.75 ab |
新窗口打开|下载CSV
2.2 不同干扰方式对土壤微生物多样性的影响
总体上, 相比封育处理, 火烧、刈割和刈割除根均使微生物α多样性指数出现下降趋势(图1)。具体表现为: 火烧在中坡位和下坡位分别使真菌Chao1指数显著降低了3.91%和7.05% (p < 0.05, 图1)。刈割在中坡位和下坡位分别使细菌香农指数显著降低了6.53%和6.50% (p < 0.05, 图1), 也分别使细菌谱系多样性指数显著降低了13.88%和14.83% (p < 0.05, 图1), 且在下坡位使真菌Chao1指数显著下降了42.97%。刈割除根在下坡位使细菌香农指数显著降低了7.73% (p < 0.05, 图1), 在中坡位使真菌Chao1指数显著降低了4.57% (p < 0.05, 图1)。图1
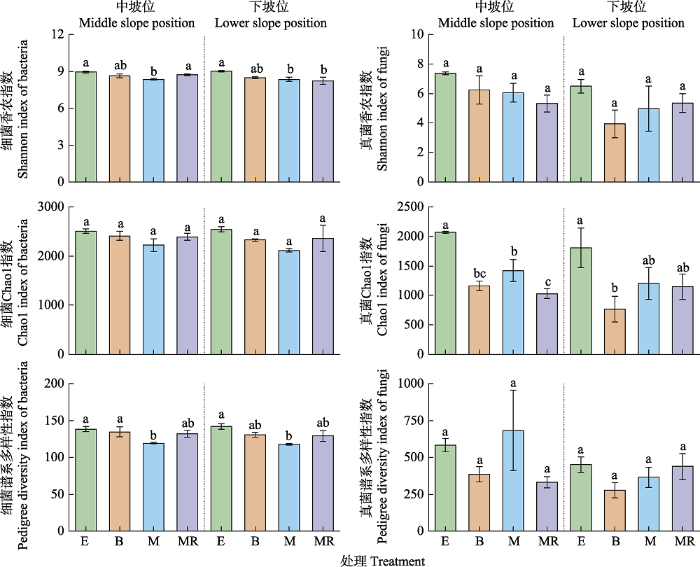 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1不同干扰方式对桂西北喀斯特草地生态系统土壤微生物多样性的影响(平均值±标准误)。B, 火烧; E, 封育; M, 刈割; MR, 刈割除根。不同小写字母表示不同处理之间差异显著(p < 0.05)。
Fig. 1Effects of different disturbances on soil microbial diversity in a karst grassland ecosystem of northwestern Guangxi, China (mean ± SE). B, burning; E, enclosure; M, mowing; MR, mowing plus root removal. Different lowercase letters indicate significant differences among different treatments (p < 0.05).
2.3 不同干扰方式对土壤微生物群落结构的影响
非度量多维尺度分析和多元方差分析的结果显示, 细菌群落结构受干扰方式显著影响(p < 0.01, 图2A), 具体表现为: 火烧、刈割和刈割除根与封育处理之间的细菌群落结构差异显著(p = 0.003-0.004, 表3), 火烧与刈割除根之间无显著差异(p = 0.153, 表3)。真菌群落结构不仅受干扰方式显著影响(p < 0.01, 图2B), 且受坡位显著影响(p < 0.05, 图2B), 具体表现为: 相比封育处理, 火烧、刈割和刈割除根与封育处理之间的真菌群落结构差异显著(p = 0.002-0.003, 表3), 火烧与刈割以及刈割除根之间无显著差异(p = 0.138, p = 0.061, 表3)。图2
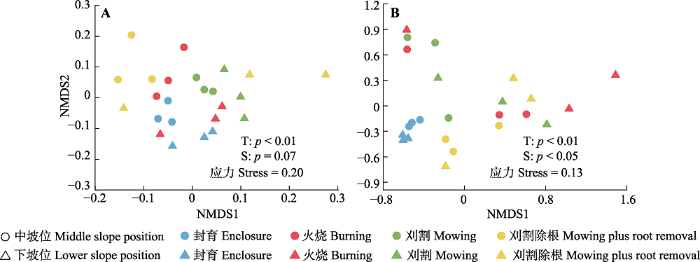 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2不同干扰方式对桂西北喀斯特草地生态系统土壤微生物群落结构影响的非度量多维尺度分析。A, 细菌群落。B, 真菌群落。S, 坡位; T, 处理。
Fig. 2Non-metric multidimensional scale analysis for the effects of different disturbances on soil microbial community structures in a karst grassland ecosystem of northwestern Guangxi, China. A, Bacterial community. B, Fungal community. S, slope position; T, treatment.
Table 3
表3
表3不同干扰方式对土壤微生物群落结构影响的多元方差分析结果
Table 3
| 干扰方式 Disturbance | 细菌 Bacteria | 真菌 Fungi | |||
|---|---|---|---|---|---|
| F | p | F | p | ||
| 封育-火烧 Enclosure-Burning | 11.10 | 0.003 | 3.28 | 0.002 | |
| 封育-刈割 Enclosure-Mowing | 11.10 | 0.004 | 2.00 | 0.002 | |
| 封育-刈割除根 Enclosure-Mowing plus root removal | 5.82 | 0.004 | 3.19 | 0.003 | |
| 火烧-刈割 Burning-Mowing | 2.16 | 0.004 | 1.30 | 0.138 | |
| 火烧-刈割除根 Burning-Mowing | 1.46 | 0.153 | 1.61 | 0.061 | |
| 刈割-刈割除根 Mowing-Mowing plus root removal | 2.94 | 0.003 | 1.68 | 0.036 | |
| 中坡位-下坡位 Middle slope position-Lower slope position | 1.90 | 0.070 | 2.17 | 0.003 | |
新窗口打开|下载CSV
细菌群落主要以变形菌门(Proteobacteria)、放线菌门(Actinobacteria)和酸杆菌门(Acidobacteria)为主, 它们的相对丰度分别为30.60%-42.53%、25.71%- 35.92%和9.82%-17.53% (图3)。火烧、刈割和刈割除根处理在中、下坡位均显著降低了酸杆菌门细菌的相对丰度, 在中坡位它们分别使酸杆菌门细菌的相对丰度从15.05%减少至10.56% (p < 0.05)、从15.05%减少至10.97% (p < 0.05), 以及从15.05%减少至11.30% (p < 0.05), 在下坡位分别使酸杆菌门细菌从17.53%减少至11.63% (p < 0.05)、从17.53%减少至11.81% (p < 0.05), 以及从17.53%减少至9.82% (p < 0.05)( 图3)。
图3
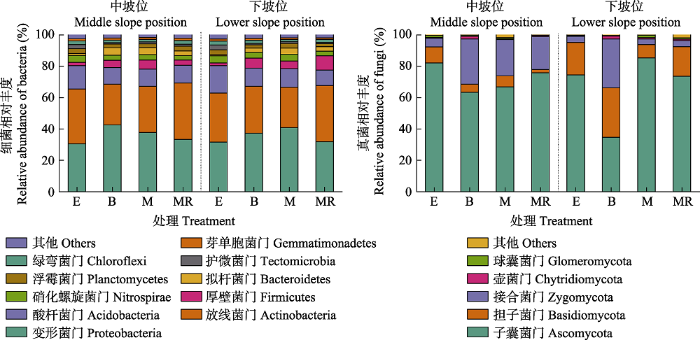 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3不同干扰方式对桂西北喀斯特草地生态系统土壤微生物群落门水平相对丰度的影响。B, 火烧; E, 封育; M, 刈割; MR, 刈割除根。
Fig. 3Effects of different disturbances on the relative abundance of microbial communities at phylum level in a karst grassland ecosystem of northwestern Guangxi, China. B, burning; E, enclosure; M, mowing; MR, mowing plus root removal.
真菌群落主要以子囊菌门(Ascomycota)、担子菌门(Basidiomycota)和接合菌门(Zygomycota)为主, 它们的相对丰度分别为34.72%-85.44%、2.16%- 31.52%和3.51%-31.23% (图3)。相比封育处理, 在中坡位刈割除根使担子菌门真菌的相对丰度从10.23%降低至2.16% (p < 0.05), 在下坡位火烧使子囊菌门真菌的相对丰度从74.49%降低至34.72% (p < 0.05)( 图3)。
2.4 不同干扰方式下土壤微生物多样性和群落结构与土壤理化性质的相关关系
冗余分析的结果显示, 土壤MBC含量与香农指数、Chao1指数和谱系多样性指数呈正相关关系(图4A、4C), 土壤MBC含量能解释细菌多样性变化的29.80% (p = 0.012, 表4), 能解释真菌多样性变化的26.80% (p = 0.008, 表4); 土壤中细根生物量(FRB)也与香农指数、Chao1指数和谱系多样性指数呈正相关关系(图4A、4C), 能解释细菌多样性变化的13.9% (p = 0.038, 表4)。图4
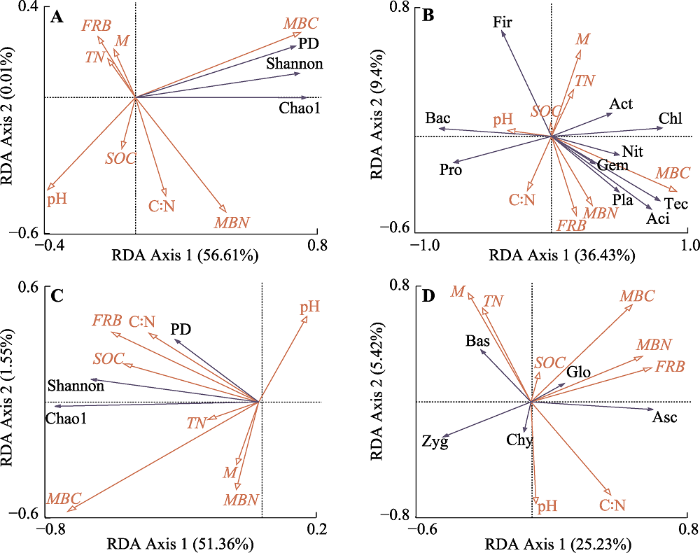 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4桂西北喀斯特草地生态系统土壤性质与土壤微生物多样性和群落结构的冗余分析。A, 细菌多样性。B, 细菌群落结构。C, 真菌多样性。D, 真菌群落结构。C:N, 碳氮比; FRB, 细根生物量; M, 含水量; MBC, 微生物生物量碳含量; MBN, 微生物生物量碳含量; SOC, 有机碳含量; TN, 总氮含量。Chao1, Chao1指数; PD, 谱系多样性指数; Shannon, 香农指数。Aci, 酸杆菌门; Act, 放线菌门; Asc, 子囊菌门; Bac, 拟杆菌门; Bas, 担子菌门; Chl, 绿弯菌门; Chy, 壶菌门; Fir, 后壁菌门; Gem, 牙单胞菌门; Glo, 球囊菌门; Nit, 硝化螺旋菌门; Pla, 浮霉菌门; Pro, 变形菌门; Tec, 护微菌门; Zyg, 接合菌门。
Fig. 4RDA analysis for the relationship between soil properties and microbial diversity and community in a karst grassland ecosystem of northwestern Guangxi, China. A, bacterial diversity. B, bacterial community structure. C, fungal diversity. D, fungal community structure. C:N, carbon-nitrogen ratio; FRB, Fine root biomass; M, moisture; MBC, microbial biomass carbon content; MBN, microbial biomass nitrogen content; SOC, soil organic carbon content; TN, total nitrogen content. Chao1, Chao1 index; PD, Pedigree diversity index; Shannon, Shannon index. Aci, Acidobacteria; Act, Actinobacteria; Asc, Ascomycota; Bac, Bacteroidetes; Bas, Basidiomycota; Chl, Chloroflexi; Chy, Chytridiomycota; Fir, Firmicutes; Gem, Gemmatimonadetes; Glo, Glomeromycota; Nit, Nitrospirae; Pla, Planctomycetes; Pro, Proteobacteria; Tec, Tectomicrobia; Zyg, Zygomycota.
Table 4
表4
表4冗余分析中土壤性质对微生物多样性和群落结构变化的解释率
Table 4
| 土壤性质 Soil property | α多样性 α diversity | 群落结构 Community structure | ||||||
|---|---|---|---|---|---|---|---|---|
| 细菌 Bacteria | 真菌 Fungi | 细菌 Bacteria | 真菌 Fungi | |||||
| E (%) | p | E (%) | p | E (%) | p | E (%) | p | |
| 含水量 Moisture (%) | 1.90 | 0.408 | 0.40 | 0.832 | 4.50 | 0.202 | 7.00 | 0.17 |
| pH | 4.50 | 0.188 | 0.30 | 0.852 | 2.60 | 0.37 | 0.60 | 0.868 |
| 有机碳含量 Soil organic carbon content (g·kg-1) | 0.20 | 0.82 | 5.10 | 0.188 | 2.50 | 0.402 | 0.10 | 0.972 |
| 总氮含量 Total nitrogen content (g·kg-1) | 0.80 | 0.572 | 1.40 | 0.536 | 1.00 | 0.728 | 4.10 | 0.46 |
| 碳氮比 Carbon-nitrogen ratio | 5.30 | 0.166 | 10.90 | 0.072 | 0.50 | 0.854 | 1.00 | 0.784 |
| 微生物生物量碳含量 Microbial biomass carbon content (g·kg-1) | 29.80 | 0.012 | 26.80 | 0.008 | 31.70 | 0.002 | 1.60 | 0.744 |
| 微生物生物量氮含量 Microbial biomass nitrogen content (g·kg-1) | 0.20 | 0.792 | 6.30 | 0.176 | 3.10 | 0.322 | 6.10 | 0.23 |
| 细根生物量 Fine root biomass (g·m-2) | 13.90 | 0.038 | 1.80 | 0.474 | 3.40 | 0.292 | 10.30 | 0.088 |
新窗口打开|下载CSV
土壤MBC含量与硝化螺旋菌门(Nitrospirae)、护微菌门(Tectomicrobia)、酸杆菌门、浮霉菌门(Planctomycetes)、芽单胞菌门(Gemmatimonadetes)、绿弯菌门(Chloroflexi)、放线菌门呈正相关关系(图4B), 它能解释细菌群落结构变化的31.70% (p = 0.002, 表4)。土壤中细根生物量(FRB)与子囊菌门、球囊菌门(Glomeromycota)呈正相关关系(图4D), 能解释真菌群落结构变化的10.30% (p = 0.088, 表4)。
3 讨论
3.1 不同干扰方式对土壤微生物多样性的影响
总体上, 火烧、刈割、刈割除根均在不同程度上降低了喀斯特草地土壤微生物多样性, 这种影响因不同干扰方式、不同微生物(细菌和真菌)类型而不同(图1)。比如, 火烧显著降低了真菌的Chao1指数, 但是不影响细菌α多样性(图1), 说明真菌α多样性对火烧干扰的响应比细菌更为敏感。有研究表明, 土壤细菌比真菌更耐热(Bollen,1969; 陶玉柱和邸雪颖, 2013), 细菌的致死温度比真菌要高出40 ℃ (Choromanska & DeLuca, 2002), 且细菌的耐高温种类远高于真菌(Bárcenas-Moreno & B??th,2009)。因此, 火烧引起的高温容易导致真菌细胞的溶解和死亡, 进而可能改变真菌群落的微生物量和多样性(Hart et al., 2005)。D’Ascoli等(2005)的研究也发现, 火烧对真菌群落的影响大于细菌群落。另外, 由于植物根系相比微生物的热致死温度更低(陶玉柱和邸雪颖, 2013), 且地下植物根系与真菌菌根存在共生关系, 火烧导致的植物根系死亡也可能是影响真菌多样性的重要因素。本研究结果显示, 火烧干扰显著降低了土壤细根生物量(表2), 支持了这个结论。相比火烧, 刈割显著降低了细菌的香农指数和谱系多样性指数且在下坡位显著降低了真菌的Chao1指数, 而刈割除根在中、下坡位分别显著降低了细菌香农指数和真菌Chao1指数(图1), 说明刈割和刈割除根对细菌和真菌α多样性都存在显著影响。刈割干扰的显著特征是把地上植物收割走, 刈割除根则连同地上植物和地下根系都收割走, 两者均直接导致有机生物量输入减少进而可能影响微生物多样性。本研究发现, 刈割和刈割除根处理确实在不同程度上减少了细根生物量(表2), 且细根生物量的减少能解释细菌α多样性变化的13.9% (表4)。由于根系与微生物的生长或活性均存在紧密联系(Ohtonen & V?re, 1998), 特别是根际效应(Grayston, 2000), 因此无论是火烧、刈割还是刈割除根干扰引起根系生物量的减少都将不利于微生物多样性的维持。
以前的研究表明, 土壤含水量和pH是土壤微生物多样性的主要影响因素(Xu et al., 2004; 韩丛丛等, 2014)。土壤含水量对维持微生物正常代谢至关重要, 土壤含水量适量时土壤微生物量和多样性达到最大(韩丛丛等, 2014), 但是过多或过少都会抑制土壤微生物活性(Xu et al., 2004)。pH指示土壤酸碱度, 它可以通过改变微生物细胞膜的通透性和稳定性及微生物的酶活性来影响微生物的多样性(H?gberg & Read,2006; 韩丛丛等, 2014)。然而, 在我们的研究中, 人为干扰没有显著影响土壤含水量和pH (表2), 这与先前的一些研究结果(Vázquezet al., 1993; Abril et al., 2005)并不一致。另外我们发现, 人为干扰下显著减少土壤MBC含量(表2), MBC含量的减少能分别解释细菌和真菌α多样性变化的29.80%和26.8% (图4; 表4), 这表明土壤MBC含量也是人为干扰影响微生物α多样性的重要因素。最近发表的一项研究发现, 氮添加干扰下微生物α多样性与MBC含量也存在显著的正相关关系(Wanget al., 2020)。
3.2 不同干扰方式对土壤微生物群落结构的影响
本研究发现, 喀斯特草地土壤细菌群落主要以变形菌门、放线菌门和酸杆菌门为主, 而真菌群落主要以子囊菌门、担子菌门和接合菌门为主(图3)。火烧、刈割、刈割除根对细菌和真菌的群落结构均存在显著影响(图2; 表3)。对于细菌群落, 相比封育处理, 火烧、刈割和刈割除根3种人为干扰方式均显著降低了酸杆菌门细菌的相对丰度(图3)。有研究表明, 酸杆菌门细菌在生态系统功能中的多个方面发挥着重要作用: 第一, 酸杆菌门细菌不仅具有降解植物残体多聚物的功能, 如Telmatobacter bradus被发现能够降解纤维素(Pankratov et al., 2012; 王光华等, 2016); 酸杆菌门细菌还能参与单碳化合物的代谢(王光华等, 2016), 如Pankratov等(2008)研究发现往土壤中添加甲醇能促进酸杆菌门细菌的生长。第二, 酸杆菌门细菌在生态系统中参与铁循环, 如Geothrix fermentans和Acidobacterium capsulatum都被证明是具有异化铁还原能力的酸杆菌门细菌(Coates et al., 1999; Lu et al., 2010; 王光华等, 2016)。因此, 在喀斯特草地生态系统中, 人为干扰引起酸杆菌门细菌的减少将不利于土壤中有机质的降解和铁的循环。对于真菌群落, 相比封育处理, 刈割除根显著降低了担子菌门真菌的相对丰度, 而火烧显著降低了子囊菌门真菌的相对丰度(图3)。有研究表明, 担子菌门真菌在生态系统功能特别是碳循环过程中发挥着重要作用, 因为担子门真菌(特别是Agaricomycetes)能产生胞外水解酶和氧化酶参与木质纤维素降解(Sergentani et al., 2016)。Bastida等(2016)研究发现土壤溶解有机碳(DOC)含量随担子菌门真菌相对丰度的增加而增加, 进一步证实了担子菌门真菌在土壤碳循环中的重要性。另外, 有研究表明, 子囊菌门真菌能增强土壤抗侵蚀能力, 在土壤的稳定性方面发挥重要作用(Challacombe et al., 2019), 这个特征在易发生水土流失的喀斯特生态系统中尤为重要。因此, 由火烧干扰引起的子囊菌门真菌的减少将可能削弱喀斯特草地生态系统土壤的抗侵蚀能力, 从而降低其稳定性。
尽管土壤水分、pH、有机质含量等土壤理化性质被认为是影响土壤微生物群落结构的主要因素(李娜等, 2012), 但是本研究发现人为干扰下的微生物群落结构与土壤含水量、pH、碳氮含量并不存在显著的相关性(图4; 表4), MBC含量和细根生物量的改变才是影响土壤微生物群落结构变化的主要因素, 人为干扰引起MBC含量的减少能解释细菌群落结构变化的31.70%, 而细根生物量的减少能解释真菌群落结构变化的10.30% (图4; 表4)。因此, 在用模型预测未来人为干扰下喀斯特草地土壤微生物群落结构变化的方向和强度时, MBC含量和细根生物量应被考虑为重要的调控因素。
4 结论
火烧、刈割、刈割除根对土壤微生物α多样性产生负面影响, 并显著改变微生物群落结构, 其影响与改变程度因干扰方式、微生物类型(细菌、真菌)不同而异, 且受坡位调控。火烧、刈割、刈割除根导致α多样性显著降低及火烧导致子囊菌门真菌相对丰度显著减少, 将不利于喀斯特草地土壤生态系统稳定性的维持; 火烧、刈割、刈割除根导致酸杆菌门细菌相对丰度显著减少, 将不利于土壤有机质的降解和铁元素的循环。在本研究区域, 土壤微生物生物量碳含量和细根生物量的减少是人为干扰影响土壤微生物多样性和群落结构的重要因素。致谢
感谢中国科学院亚热带农业生态研究所张伟和刘坤平两位老师在样地调查和选择方面给予的帮助和建议。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1016/j.foreco.2004.11.014URL [本文引用: 1]
DOI:10.1016/j.soilbio.2009.09.010URL [本文引用: 1]
DOI:10.1111/mec.13783PMID:27481114 [本文引用: 1]

Biogeochemical processes and ecosystemic functions are mostly driven by soil microbial communities. However, most methods focus on evaluating the total microbial community and fail to discriminate its active fraction which is linked to soil functionality. Precisely, the activity of the microbial community is strongly limited by the availability of organic carbon (C) in soils under arid and semi-arid climate. Here, we provide a complementary genomic and metaproteomic approach to investigate the relationships between the diversity of the total community, the active diversity and ecosystem functionality across a dissolved organic carbon (DOC) gradient in southeast Spain. DOC correlated with the ecosystem multifunctionality index composed by soil respiration, enzyme activities (urease, alkaline phosphatase and β-glucosidase) and microbial biomass (phospholipid fatty acids, PLFA). This study highlights that the active diversity (determined by metaprotoemics) but not the diversity of the whole microbial community (evaluated by amplicon gene sequencing) is related to the availability of organic C and it is also connected to the ecosystem multifunctionality index. We reveal that DOC shapes the activities of bacterial and fungal populations in Mediterranean semi-arid soils and determines the compartmentalization of functional niches. For instance, Rhizobales thrived at high-DOC sites probably fuelled by metabolism of one-C compounds. Moreover, the analysis of proteins involved in the transport and metabolism of carbohydrates revealed that Ascomycota and Basidiomycota occupied different nutritional niches. The functional mechanisms for niche specialization were not constant across the DOC gradient.? 2016 John Wiley & Sons Ltd.
DOI:10.1007/BF02137211URL [本文引用: 1]
DOI:10.1016/0038-0717(85)90143-9URL [本文引用: 1]
DOI:10.1016/j.copbio.2010.10.009URL [本文引用: 1]
DOI:10.1186/s12864-019-6358-xPMID:31830917 [本文引用: 1]
The dominant fungi in arid grasslands and shrublands are members of the Ascomycota phylum. Ascomycota fungi are important drivers in carbon and nitrogen cycling in arid ecosystems. These fungi play roles in soil stability, plant biomass decomposition, and endophytic interactions with plants. They may also form symbiotic associations with biocrust components or be latent saprotrophs or pathogens that live on plant tissues. However, their functional potential in arid soils, where organic matter, nutrients and water are very low or only periodically available, is poorly characterized.Five Ascomycota fungi were isolated from different soil crust microhabitats and rhizosphere soils around the native bunchgrass Pleuraphis jamesii in an arid grassland near Moab, UT, USA. Putative genera were Coniochaeta, isolated from lichen biocrust, Embellisia from cyanobacteria biocrust, Chaetomium from below lichen biocrust, Phoma from a moss microhabitat, and Aspergillus from the soil. The fungi were grown in replicate cultures on different carbon sources (chitin, native bunchgrass or pine wood) relevant to plant biomass and soil carbon sources. Secretomes produced by the fungi on each substrate were characterized. Results demonstrate that these fungi likely interact with primary producers (biocrust or plants) by secreting a wide range of proteins that facilitate symbiotic associations. Each of the fungal isolates secreted enzymes that degrade plant biomass, small secreted effector proteins, and proteins involved in either beneficial plant interactions or virulence. Aspergillus and Phoma expressed more plant biomass degrading enzymes when grown in grass- and pine-containing cultures than in chitin. Coniochaeta and Embellisia expressed similar numbers of these enzymes under all conditions, while Chaetomium secreted more of these enzymes in grass-containing cultures.This study of Ascomycota genomes and secretomes provides important insights about the lifestyles and the roles that Ascomycota fungi likely play in arid grassland, ecosystems. However, the exact nature of those interactions, whether any or all of the isolates are true endophytes, latent saprotrophs or opportunistic phytopathogens, will be the topic of future studies.
[本文引用: 4]
[本文引用: 4]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/S0038-0717(01)00180-8URL [本文引用: 1]
DOI:10.1099/00207713-49-4-1615URL [本文引用: 1]
DOI:10.1071/WF05032URL [本文引用: 1]
DOI:10.2307/3546587URL [本文引用: 1]
DOI:10.1016/j.geoderma.2012.01.037URL [本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 3]
DOI:10.1016/j.foreco.2005.08.012URL [本文引用: 1]
DOI:10.5846/stxb202006231627 [本文引用: 5]
DOI:10.5846/stxb202006231627 [本文引用: 5]
DOI:10.1016/j.tree.2006.06.004URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s00374-018-1322-5URL [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.17521/cjpe.2018.0245URLPMID:6693763B-4886-460B-A562-05055DEFAC55 [本文引用: 1]
DOI:10.17521/cjpe.2018.0245PMID:6693763B-4886-460B-A562-05055DEFAC55 [本文引用: 1]
由全球变化和工农业生产引发的大气氮沉降增加已经对生态系统结构和功能产生了不可忽视的影响, 但是氮沉降的组成成分存在多种形态, 不同形态的氮对生态系统的结构与功能的影响是否有差异目前还不清楚。因此, 该研究选择内蒙古草甸草原开展不同形态和不同水平的外源氮添加试验, 每年添加5种不同形态的氮肥, 包括: 尿素、碳酸氢铵、硝酸铵、硫酸铵、缓释尿素, 添加量分别为: 0 (N0)、2 (N2)、5 (N5)、10 (N10)、20 (N20)及50 (N50) g·m <sup>-2</sup>·a <sup>-1</sup>, 均为纯氮添加量。通过野外原位取土、室内控制温度和水分(25 ℃和60%田间持水量)的培养试验测定土壤净氮矿化(mg·kg <sup>-1</sup>·h <sup>-1</sup>)潜力、土壤微生物呼吸(μg·g <sup>-1</sup>·h <sup>-1</sup>)潜力、土壤微生物生物量碳(氮)(mg·kg <sup>-1</sup>)的潜力以及土壤碳(g·kg <sup>-1</sup>)、氮(g·kg <sup>-1</sup>)、磷(g·kg <sup>-1</sup>)含量等指标, 研究添加不同形态和不同水平的氮对土壤净氮矿化潜力的影响。试验结果表明: (1)短期内不同形态、不同水平的氮添加改变了土壤中无机氮的含量、铵态氮和硝态氮的累积量, 并且表现出铵态氮肥的促进作用比硝态氮肥更加显著, 铵态氮的累积显著提高了土壤净氮矿化潜力, 短期铵态氮和硝态氮的累积可增加微生物和植物对有效氮的快速固持; (2)不同形态、不同水平氮添加导致土壤微生物活性发生改变, 包括土壤微生物生物量碳(MBC)含量、微生物生物量氮(MBN)含量及其碳氮比(MBC:MBN), 并且在低水平氮添加下显著增强土壤微生物的呼吸速率, 高水平氮添加显著降低微生物呼吸速率和呼吸熵; (3)不同形态、不同水平氮添加短期内对土壤含水量、土壤有机碳含量、土壤全磷含量、土壤全氮含量无显著影响, 但是高水平氮添加不仅提高了速效磷的含量, 而且导致土壤迅速酸化。室内培养净氮矿化潜力的结果进一步验证了内蒙古草甸草原受氮限制, 添加中低水平的氮可以通过提高土壤微生物的活性而增加该地区草原土壤的净氮矿化潜力, 从而提高草地生产力。
[本文引用: 5]
[本文引用: 5]
DOI:10.1128/AEM.01931-10URL [本文引用: 1]
DOI:10.1128/aem.02738-17 [本文引用: 1]
DOI:10.1098/rstb.2019.0244URL [本文引用: 1]
PMID:10693803 [本文引用: 1]
Producers, such as plants and algae, acquire nutrients from inorganic sources that are supplied primarily by decomposers whereas decomposers, mostly fungi and bacteria, acquire carbon from organic sources that are supplied primarily by producers. This producer-decomposer co-dependency is important in governing ecosystem processes, which implies that the impacts of declining biodiversity on ecosystem functioning should be strongly influenced by this process. Here we show, by simultaneously manipulating producer (green algal) and decomposer (heterotrophic bacterial) diversity in freshwater microcosms, that algal biomass production varies considerably among microcosms (0.0-0.67 mg ml(-1)), but that neither algal nor bacterial diversity by itself can explain this variation. Instead, production is a joint function of both algal and bacterial diversity. Furthermore, the range in algal production in microscosms in which bacterial diversity was manipulated was nearly double (1.82 times) that of microcosms in which bacterial diversity was not manipulated. Measures of organic carbon use by bacteria in these microcosms indicate that carbon usage is the mechanism responsible for these results. Because both producer and microbial diversity respond to disturbance and habitat modification, the main causes of biodiversity loss, these results suggest that ecosystem response to changing biodiversity is likely to be more complex than other studies have shown.
PMID:9852512 [本文引用: 1]
Forest soil ecology was studied in Fennoscandinavian dry Scots pine forests grazed by reindeer to varying extents (ungrazed, lichen-dominated-sites; grazed sites; and bryophyte-dominated sites). We hypothesized that the productivity parameters of the site (i.e., tree growth and soil nutrient concentrations), the vegetation composition, and the microbial activities are directly correlated. Since the productivity of the lichen-dominated ecosystem is low, microbial activities are assumed to be naturally low. Grazing was expected to decrease both the amount of Scots pine fine roots and the soil microbial activities. Several variables on the characteristics of the soil microbial community, Scots pine fine roots, soil nutrients, and tree growth were studied in relation to vegetation composition by using non-metric multidimensional scaling (NMDS). Basal respiration (Bas), metabolic quotient of the microbial community (qCO2), and pine fine root parameters increased toward the ungrazed, nutrient-poor, lichen-dominated sites, which were grouped at one end of the first axis in the NMDS ordination. Soil nutrient and tree growth parameters and thickness of the humus layer increased toward bryophyte-dominated sites, which were grouped at the other end of the first axis in the ordination. The grazed sites fell between them. These were characterized by lower Bas and qCO2 values and longer lag, compared to ungrazed lichen- or bryophyte-dominated sites, probably due to decreased carbon input and microclimatic change (the soil without lichen carpet is exposed to direct sunlight and wind). Microbial biomass (Cmic), fungal biomass (ergosterol concentration), and the specific growth rate (μCO2) were not related to vegetation ordination. The high fine root production is the most plausible explanation for the high microbial activities at nutrient-poor, lichen-dominated sites, which produce qualitatively poor and slowly decomposing litter, as fine roots secrete considerable amounts of organic substances. At bryophyte-dominated sites, the higher soil nutrient concentrations and the higher production of easily decomposable substrates are likely to maintain the microbial activities.
DOI:10.1099/ijs.0.029629-0PMID:21460138 [本文引用: 1]
A gram-negative, facultatively anaerobic, chemo-organotrophic, non-pigmented, slow-growing bacterium was isolated from acidic Sphagnum peat and designated strain TPB6017(T). Cells of this strain were long rods that multiplied by normal cell division and were motile by means of a single flagellum. Cells grew under reduced oxygen tension and under anoxic conditions and were able to ferment sugars and several polysaccharides, including amorphous and crystalline cellulose. Strain TPB6017(T) was a psychrotolerant acidophile capable of growth between pH 3.0 and 7.5 (optimum 4.5-5.0) and at 4-35 °C (optimum 20-28 °C). It was extremely sensitive to salt stress; growth was inhibited at NaCl concentrations above 0.1?% (w/v). The major fatty acids were iso-C(15?:?0) and iso-C(17?:?1)ω9c; the polar lipids were phosphatidylethanolamine and a number of phospholipids and aminophospholipids with an unknown structure. The quinone was MK-8. The DNA G+C content was 57.6 mol%. Comparative 16S rRNA gene sequence analysis revealed that strain TPB6017(T) was a member of subdivision 1 of the phylum Acidobacteria and belonged to a phylogenetic lineage defined by the acidophilic aerobic chemo-organotroph Acidobacterium capsulatum (92.3?% sequence similarity). However, cell morphology, type of flagellation, the absence of pigment, differences in fatty acid and polar lipid composition, possession of a cellulolytic capability, inability to grow under fully oxic conditions and good growth in anoxic conditions distinguished strain TPB6017(T) from A. capsulatum. Therefore, it is proposed that strain TPB6017(T) represents a novel acidobacterium species in a new genus, Telmatobacter bradus gen. nov., sp. nov.; strain TPB6017(T) (?=?DSM 23630(T)?=?VKM B-2570(T)) is the type strain.
DOI:10.1038/ismej.2008.7URL [本文引用: 1]
DOI:10.1038/ismej.2013.34URL [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/0038-0717(87)90052-6URL [本文引用: 2]
DOI:10.1111/fem.1993.13.issue-2URL [本文引用: 2]
DOI:10.1016/j.soilbio.2019.107687URL [本文引用: 1]
[本文引用: 3]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
DOI:10.1029/2004gb002281 [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
The effect of fire and overgrazing, disturbes on soil carbon balance in the dry Chaco forest
1
2005
... 以前的研究表明, 土壤含水量和pH是土壤微生物多样性的主要影响因素(Xu et al.,
Bacterial and fungal growth in soil heated at different temperatures to simulate a range of fire intensities
1
2009
... 总体上, 火烧、刈割、刈割除根均在不同程度上降低了喀斯特草地土壤微生物多样性, 这种影响因不同干扰方式、不同微生物(细菌和真菌)类型而不同(
The active microbial diversity drives ecosystem multifunctionality and is physiologically related to carbon availability in Mediterranean semi-arid soils
1
2016
... 对于真菌群落, 相比封育处理, 刈割除根显著降低了担子菌门真菌的相对丰度, 而火烧显著降低了子囊菌门真菌的相对丰度(
The selective effect of heat treatment on the microflora of a greenhouse soil
1
1969
... 总体上, 火烧、刈割、刈割除根均在不同程度上降低了喀斯特草地土壤微生物多样性, 这种影响因不同干扰方式、不同微生物(细菌和真菌)类型而不同(
Chloroform fumigation and the release of soil nitrogen: the effects of fumigation time and temperature
1
1985
... 式中, M(C,N)为微生物生物量碳或氮含量(mg·kg-1); ?E(C,N)为熏蒸处理减去未熏蒸处理有机碳或氮含量(mg·kg-1); k(C,N)为换算系数, 微生物生物量碳和氮分别取值0.45和0.54 (Brookes et al.,
The emerging role for bacteria in lignin degradation and bio-product formation
1
2011
... 土壤微生物是土壤养分循环的驱动者, 它控制着土壤有机质的分解与形成、碳氮循环等过程(Fontúrbelet al.,
Genomes and secretomes of Ascomycota fungi reveal diverse functions in plant biomass decomposition and pathogenesis
1
2019
... 对于真菌群落, 相比封育处理, 刈割除根显著降低了担子菌门真菌的相对丰度, 而火烧显著降低了子囊菌门真菌的相对丰度(
桂西北喀斯特峰丛不同土地利用方式坡面产流产沙特征
4
2012
... 长期干扰监测样地位于桂西北环江毛南族自治县中国科学院环江喀斯特生态系统观测研究站的木连综合试验示范区(24.72°-24.73° N, 108.30°- 108.32° E)(陈洪松等,
... ).地貌类型为典型的喀斯特峰丛洼地, 土壤类型为白云岩发育的棕色石灰土, 沿坡向下土层平均厚度由10-30 cm增加到50-80 cm (陈洪松等,
... ).1985年之前研究区生态系统经历着频繁的火烧、刈割和放牧, 1985年研究区大部分居民外迁, 退化生态系统得以逐渐恢复, 植被类型以荒草地和稀疏灌丛为主(陈洪松等,
... 2004年底, 选择土壤和植被较为均一且坡向、坡度条件基本相同的一面东南方向山坡, 鉴于喀斯特山坡中下部被当地居民利用率较高(陈洪松等,
桂西北喀斯特峰丛不同土地利用方式坡面产流产沙特征
4
2012
... 长期干扰监测样地位于桂西北环江毛南族自治县中国科学院环江喀斯特生态系统观测研究站的木连综合试验示范区(24.72°-24.73° N, 108.30°- 108.32° E)(陈洪松等,
... ).地貌类型为典型的喀斯特峰丛洼地, 土壤类型为白云岩发育的棕色石灰土, 沿坡向下土层平均厚度由10-30 cm增加到50-80 cm (陈洪松等,
... ).1985年之前研究区生态系统经历着频繁的火烧、刈割和放牧, 1985年研究区大部分居民外迁, 退化生态系统得以逐渐恢复, 植被类型以荒草地和稀疏灌丛为主(陈洪松等,
... 2004年底, 选择土壤和植被较为均一且坡向、坡度条件基本相同的一面东南方向山坡, 鉴于喀斯特山坡中下部被当地居民利用率较高(陈洪松等,
不同干扰方式对喀斯特生态系统土壤细菌优势类群——变形菌群落的影响
1
2012
... 中国的喀斯特面积居于世界首位, 其裸露面积约为90.7万km2, 约占国土总面积的10% (宋同清等,
不同干扰方式对喀斯特生态系统土壤细菌优势类群——变形菌群落的影响
1
2012
... 中国的喀斯特面积居于世界首位, 其裸露面积约为90.7万km2, 约占国土总面积的10% (宋同清等,
Microbial activity and nitrogen mineralization in forest mineral soils following heating: evaluation of post-fire effects
1
2002
... 总体上, 火烧、刈割、刈割除根均在不同程度上降低了喀斯特草地土壤微生物多样性, 这种影响因不同干扰方式、不同微生物(细菌和真菌)类型而不同(
Geothrix fermentans gen. nov., sp. nov., a novel Fe(III)-reducing bacterium from a hydrocarbon-contaminated aquifer
1
1999
... 本研究发现, 喀斯特草地土壤细菌群落主要以变形菌门、放线菌门和酸杆菌门为主, 而真菌群落主要以子囊菌门、担子菌门和接合菌门为主(
Functional diversity of the microbial community in Mediterranean maquis soils as affected by fires
1
2005
... 总体上, 火烧、刈割、刈割除根均在不同程度上降低了喀斯特草地土壤微生物多样性, 这种影响因不同干扰方式、不同微生物(细菌和真菌)类型而不同(
Microbial diversity and ecosystem function
1
1997
... 土壤微生物是土壤养分循环的驱动者, 它控制着土壤有机质的分解与形成、碳氮循环等过程(Fontúrbelet al.,
Effects of an experimental fire and post-fire stabilization treatments on soil microbial communities
1
2012
... 土壤微生物是土壤养分循环的驱动者, 它控制着土壤有机质的分解与形成、碳氮循环等过程(Fontúrbelet al.,
Rhizodeposition and its impact on microbial community structure and function in trees
1
2000
... 相比火烧, 刈割显著降低了细菌的香农指数和谱系多样性指数且在下坡位显著降低了真菌的Chao1指数, 而刈割除根在中、下坡位分别显著降低了细菌香农指数和真菌Chao1指数(
草地土壤微生物多样性影响因子
3
2014
... 以前的研究表明, 土壤含水量和pH是土壤微生物多样性的主要影响因素(Xu et al.,
... ).土壤含水量对维持微生物正常代谢至关重要, 土壤含水量适量时土壤微生物量和多样性达到最大(韩丛丛等,
... ; 韩丛丛等,
草地土壤微生物多样性影响因子
3
2014
... 以前的研究表明, 土壤含水量和pH是土壤微生物多样性的主要影响因素(Xu et al.,
... ).土壤含水量对维持微生物正常代谢至关重要, 土壤含水量适量时土壤微生物量和多样性达到最大(韩丛丛等,
... ; 韩丛丛等,
Post-fire vegetative dynamics as drivers of microbial community structure and function in forest soils
1
2005
... 总体上, 火烧、刈割、刈割除根均在不同程度上降低了喀斯特草地土壤微生物多样性, 这种影响因不同干扰方式、不同微生物(细菌和真菌)类型而不同(
植被恢复十年喀斯特坡地细根对土壤碳氮存留与可利用性的影响
5
2020
... 引用自刘艳等(
... Table data were referenced from Liu et al. (
... 2017年6月(长期干扰处理已有12年), 分别在每种处理模式下的中坡和下坡位置随机建立3个5 m × 10 m的样方, 共计24个样方(4个处理×2个坡位×3个重复).为了减少喀斯特土壤分布高异质性的影响, 每个样方内按“S”形5点取样法采集0-10 cm表层土壤, 充分混合成一份土壤样品, 累计土壤采样点数为120个, 最终获取土壤样品数24份; 根钻(直径7 cm)随机采集每个样方内3个植物根系样品, 充分混合成一份根系样品, 累计根系采样点数为72个, 最终获取根系样品数24份(贺同鑫等,
... 土壤SOC含量采用酸洗法(去除无机碳)测定(贺同鑫等,
... 土壤TN含量采用元素分析仪(Isoprime vario ISOTOPE cube, Elementar, Frankfurt, Germany)测定(贺同鑫等,
植被恢复十年喀斯特坡地细根对土壤碳氮存留与可利用性的影响
5
2020
... 引用自刘艳等(
... Table data were referenced from Liu et al. (
... 2017年6月(长期干扰处理已有12年), 分别在每种处理模式下的中坡和下坡位置随机建立3个5 m × 10 m的样方, 共计24个样方(4个处理×2个坡位×3个重复).为了减少喀斯特土壤分布高异质性的影响, 每个样方内按“S”形5点取样法采集0-10 cm表层土壤, 充分混合成一份土壤样品, 累计土壤采样点数为120个, 最终获取土壤样品数24份; 根钻(直径7 cm)随机采集每个样方内3个植物根系样品, 充分混合成一份根系样品, 累计根系采样点数为72个, 最终获取根系样品数24份(贺同鑫等,
... 土壤SOC含量采用酸洗法(去除无机碳)测定(贺同鑫等,
... 土壤TN含量采用元素分析仪(Isoprime vario ISOTOPE cube, Elementar, Frankfurt, Germany)测定(贺同鑫等,
Towards a more plant physiological perspective on soil ecology
1
2006
... 以前的研究表明, 土壤含水量和pH是土壤微生物多样性的主要影响因素(Xu et al.,
喀斯特山区不同草地土壤养分与酶活性特征
1
2020
... 中国的喀斯特面积居于世界首位, 其裸露面积约为90.7万km2, 约占国土总面积的10% (宋同清等,
喀斯特山区不同草地土壤养分与酶活性特征
1
2020
... 中国的喀斯特面积居于世界首位, 其裸露面积约为90.7万km2, 约占国土总面积的10% (宋同清等,
Soil microbial response following wildfires in thermic oak-pine forests
2
2018
... 土壤微生物群落对环境条件的改变较为敏感, 温度、水分、pH、养分等理化性质与微生物的生长和代谢紧密相关(陶玉柱和邸雪颖,
... ).由于干扰作用的机制不同, 三者对微生物多样性和群落结构的影响也可能不尽相同.另外, 微生物对人为干扰的响应还可能与植被类型、地形等环境因子相关(Huffman & Madritch,
土壤微生物群落结构影响因素的探讨
1
2012
... 尽管土壤水分、pH、有机质含量等土壤理化性质被认为是影响土壤微生物群落结构的主要因素(李娜等,
土壤微生物群落结构影响因素的探讨
1
2012
... 尽管土壤水分、pH、有机质含量等土壤理化性质被认为是影响土壤微生物群落结构的主要因素(李娜等,
近30 a江西省农田土壤pH时空变化及其与酸雨和施肥量间关系
1
2017
... 土壤含水量采用烘干称质量法(105 ℃, 24 h)测定; 土壤pH采用电极电位法(土水质量比为1:2.5)测定(李伟峰等,
近30 a江西省农田土壤pH时空变化及其与酸雨和施肥量间关系
1
2017
... 土壤含水量采用烘干称质量法(105 ℃, 24 h)测定; 土壤pH采用电极电位法(土水质量比为1:2.5)测定(李伟峰等,
不同形态和水平的氮添加对内蒙古草甸草原土壤净氮矿化潜力的影响
1
2019
... 土壤含水量采用烘干称质量法(105 ℃, 24 h)测定; 土壤pH采用电极电位法(土水质量比为1:2.5)测定(李伟峰等,
不同形态和水平的氮添加对内蒙古草甸草原土壤净氮矿化潜力的影响
1
2019
... 土壤含水量采用烘干称质量法(105 ℃, 24 h)测定; 土壤pH采用电极电位法(土水质量比为1:2.5)测定(李伟峰等,
喀斯特峰丛洼地不同土地利用方式土壤肥力特征
5
2014
... 长期干扰监测样地位于桂西北环江毛南族自治县中国科学院环江喀斯特生态系统观测研究站的木连综合试验示范区(24.72°-24.73° N, 108.30°- 108.32° E)(陈洪松等,
... ; 刘艳等,
... 2004年底, 选择土壤和植被较为均一且坡向、坡度条件基本相同的一面东南方向山坡, 鉴于喀斯特山坡中下部被当地居民利用率较高(陈洪松等,
... 引用自刘艳等(
... Table data were referenced from Liu et al. (
喀斯特峰丛洼地不同土地利用方式土壤肥力特征
5
2014
... 长期干扰监测样地位于桂西北环江毛南族自治县中国科学院环江喀斯特生态系统观测研究站的木连综合试验示范区(24.72°-24.73° N, 108.30°- 108.32° E)(陈洪松等,
... ; 刘艳等,
... 2004年底, 选择土壤和植被较为均一且坡向、坡度条件基本相同的一面东南方向山坡, 鉴于喀斯特山坡中下部被当地居民利用率较高(陈洪松等,
... 引用自刘艳等(
... Table data were referenced from Liu et al. (
Ecophysiology of Fe-cycling bacteria in acidic sediments
1
2010
... 本研究发现, 喀斯特草地土壤细菌群落主要以变形菌门、放线菌门和酸杆菌门为主, 而真菌群落主要以子囊菌门、担子菌门和接合菌门为主(
High microbial diversity promotes soil ecosystem functioning
1
2018
... 土壤微生物是土壤养分循环的驱动者, 它控制着土壤有机质的分解与形成、碳氮循环等过程(Fontúrbelet al.,
Linking microbial communities to ecosystem functions: What we can learn from genotype-phenotype mapping in organisms
1
2020
... 土壤微生物是土壤养分循环的驱动者, 它控制着土壤有机质的分解与形成、碳氮循环等过程(Fontúrbelet al.,
Producer-decomposer co-dependency influences biodiversity effects
1
2000
... 土壤微生物是土壤养分循环的驱动者, 它控制着土壤有机质的分解与形成、碳氮循环等过程(Fontúrbelet al.,
Vegetation composition determines microbial activities in a boreal forest soil
1
1998
... 相比火烧, 刈割显著降低了细菌的香农指数和谱系多样性指数且在下坡位显著降低了真菌的Chao1指数, 而刈割除根在中、下坡位分别显著降低了细菌香农指数和真菌Chao1指数(
Telmatobacter bradus gen. nov., sp. nov., a cellulolytic facultative anaerobe from subdivision 1 of the Acidobacteria, and emended description of Acidobacterium capsulatum Kishimoto et al. 1991
1
2012
... 本研究发现, 喀斯特草地土壤细菌群落主要以变形菌门、放线菌门和酸杆菌门为主, 而真菌群落主要以子囊菌门、担子菌门和接合菌门为主(
Substrate-induced growth and isolation of Acidobacteria from acidic Sphagnum peat
1
2008
... 本研究发现, 喀斯特草地土壤细菌群落主要以变形菌门、放线菌门和酸杆菌门为主, 而真菌群落主要以子囊菌门、担子菌门和接合菌门为主(
Loss in microbial diversity affects nitrogen cycling in soil
2
2013
... 土壤微生物是土壤养分循环的驱动者, 它控制着土壤有机质的分解与形成、碳氮循环等过程(Fontúrbelet al.,
... ,
Lignocellulose degradation potential of Basidiomycota from Thrace (NE Greece)
1
2016
... 对于真菌群落, 相比封育处理, 刈割除根显著降低了担子菌门真菌的相对丰度, 而火烧显著降低了子囊菌门真菌的相对丰度(
中国西南喀斯特石漠化时空演变特征、发生机制与调控对策
1
2014
... 中国的喀斯特面积居于世界首位, 其裸露面积约为90.7万km2, 约占国土总面积的10% (宋同清等,
中国西南喀斯特石漠化时空演变特征、发生机制与调控对策
1
2014
... 中国的喀斯特面积居于世界首位, 其裸露面积约为90.7万km2, 约占国土总面积的10% (宋同清等,
1
2015
... 土壤微生物群落对环境条件的改变较为敏感, 温度、水分、pH、养分等理化性质与微生物的生长和代谢紧密相关(陶玉柱和邸雪颖,
1
2015
... 土壤微生物群落对环境条件的改变较为敏感, 温度、水分、pH、养分等理化性质与微生物的生长和代谢紧密相关(陶玉柱和邸雪颖,
火对森林土壤微生物群落的干扰作用及其机制研究进展
3
2013
... 土壤微生物群落对环境条件的改变较为敏感, 温度、水分、pH、养分等理化性质与微生物的生长和代谢紧密相关(陶玉柱和邸雪颖,
... 总体上, 火烧、刈割、刈割除根均在不同程度上降低了喀斯特草地土壤微生物多样性, 这种影响因不同干扰方式、不同微生物(细菌和真菌)类型而不同(
... )的研究也发现, 火烧对真菌群落的影响大于细菌群落.另外, 由于植物根系相比微生物的热致死温度更低(陶玉柱和邸雪颖,
火对森林土壤微生物群落的干扰作用及其机制研究进展
3
2013
... 土壤微生物群落对环境条件的改变较为敏感, 温度、水分、pH、养分等理化性质与微生物的生长和代谢紧密相关(陶玉柱和邸雪颖,
... 总体上, 火烧、刈割、刈割除根均在不同程度上降低了喀斯特草地土壤微生物多样性, 这种影响因不同干扰方式、不同微生物(细菌和真菌)类型而不同(
... )的研究也发现, 火烧对真菌群落的影响大于细菌群落.另外, 由于植物根系相比微生物的热致死温度更低(陶玉柱和邸雪颖,
桂北喀斯特山区不同植被类型土壤碳库管理指数的变化特征
1
2020
... 中国的喀斯特面积居于世界首位, 其裸露面积约为90.7万km2, 约占国土总面积的10% (宋同清等,
桂北喀斯特山区不同植被类型土壤碳库管理指数的变化特征
1
2020
... 中国的喀斯特面积居于世界首位, 其裸露面积约为90.7万km2, 约占国土总面积的10% (宋同清等,
An extraction method for measuring soil microbial biomass C.
2
1987
... 土壤MBC和MBN含量采用氯仿熏蒸-K2SO4浸提法测定(Vance et al.,
... 式中, M(C,N)为微生物生物量碳或氮含量(mg·kg-1); ?E(C,N)为熏蒸处理减去未熏蒸处理有机碳或氮含量(mg·kg-1); k(C,N)为换算系数, 微生物生物量碳和氮分别取值0.45和0.54 (Brookes et al.,
Soil microbial populations after wildfire
2
1993
... 土壤微生物群落对环境条件的改变较为敏感, 温度、水分、pH、养分等理化性质与微生物的生长和代谢紧密相关(陶玉柱和邸雪颖,
... 以前的研究表明, 土壤含水量和pH是土壤微生物多样性的主要影响因素(Xu et al.,
Stabilization of microbial residues in soil organic matter after two years of decomposition
1
2020
... 以前的研究表明, 土壤含水量和pH是土壤微生物多样性的主要影响因素(Xu et al.,
土壤酸杆菌门细菌生态学研究进展
3
2016
... 本研究发现, 喀斯特草地土壤细菌群落主要以变形菌门、放线菌门和酸杆菌门为主, 而真菌群落主要以子囊菌门、担子菌门和接合菌门为主(
... ); 酸杆菌门细菌还能参与单碳化合物的代谢(王光华等,
... ; 王光华等,
土壤酸杆菌门细菌生态学研究进展
3
2016
... 本研究发现, 喀斯特草地土壤细菌群落主要以变形菌门、放线菌门和酸杆菌门为主, 而真菌群落主要以子囊菌门、担子菌门和接合菌门为主(
... ); 酸杆菌门细菌还能参与单碳化合物的代谢(王光华等,
... ; 王光华等,
1
2019
... 土壤微生物群落对环境条件的改变较为敏感, 温度、水分、pH、养分等理化性质与微生物的生长和代谢紧密相关(陶玉柱和邸雪颖,
1
2019
... 土壤微生物群落对环境条件的改变较为敏感, 温度、水分、pH、养分等理化性质与微生物的生长和代谢紧密相关(陶玉柱和邸雪颖,
How soil moisture, rain pulses, and growth alter the response of ecosystem respiration to temperature
2
2004
... 以前的研究表明, 土壤含水量和pH是土壤微生物多样性的主要影响因素(Xu et al.,
... ,
不同人为干扰方式对桂西北喀斯特草丛群落土壤种子库组成与分布的影响
1
2008
... 土壤微生物群落对环境条件的改变较为敏感, 温度、水分、pH、养分等理化性质与微生物的生长和代谢紧密相关(陶玉柱和邸雪颖,
不同人为干扰方式对桂西北喀斯特草丛群落土壤种子库组成与分布的影响
1
2008
... 土壤微生物群落对环境条件的改变较为敏感, 温度、水分、pH、养分等理化性质与微生物的生长和代谢紧密相关(陶玉柱和邸雪颖,
西南喀斯特地区草地生态系统面临的问题及对策
1
2015
... 中国的喀斯特面积居于世界首位, 其裸露面积约为90.7万km2, 约占国土总面积的10% (宋同清等,
西南喀斯特地区草地生态系统面临的问题及对策
1
2015
... 中国的喀斯特面积居于世界首位, 其裸露面积约为90.7万km2, 约占国土总面积的10% (宋同清等,

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}