Responses of soil extracellular enzyme activities to carbon input alteration and warming in a subtropical evergreen broad-leaved forest
Shan-Shan LIU1,2,5, Wen-Jun ZHOU,1,2,5,*, Lu-Hui KUANG3,5, Zhan-Feng LIU3,5, Qing-Hai SONG1,2,5, Yun- Tong LIU1,2,5, Yi-Ping ZHANG1,2,5, Zhi-Yun LU1,4, Li-Qing SHA,1,2,5,*1CAS Key Laboratory of Tropical Forest Ecology, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Mengla, Yunnan 666303, China 2Center for Plant Ecology, Core Botanical Gardens, Chinese Academy of Sciences, Mengla, Yunnan 666303, China 3South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China 4Ailaoshan Station for Subtropical Forest Ecosystem Studies, Jingdong, Yunnan 676209, China; and 5University of Chinese Academy of Sciences, Beijing 100049, China 5University of Chinese Academy of Sciences, Beijing 100049, China
National Natural Science Foundation of China(31870467) National Natural Science Foundation of China(U1602234) CAS 135 Project(2017XTBG-F01) CAS 135 Project(2017XTBG-T01)
Abstract Aims The objective was to investigate the responses of soil extracellular enzyme activities to carbon input alteration and warming in a subtropical evergreen broad-leaved forest of Ailao Mountain, Yunnan, southwest China. Methods This study was based on two soil depths (0-5 and 5-10 cm) for four treatments under a long-term soil warming experiment in a subtropical evergreen broad-leaved forest of Ailao Mountain, Yunnan, southwest China. Potential activities of β-glucosidase (BG), polyphenol oxidase (POX), peroxidase (PER), β-1,4-N-acetylglucosaminidase (NAG) and acid phosphatase (AP) and their stoichiometric ratios were measured. Soil physical and chemical properties were also analyzed. Important findings The results showed that in the control treatment, activities of all enzymes except POX decreased significantly with soil depth. Compared with the control treatment, long-term litter removal significantly reduced AP and BG activities at 0-5 cm soil depth, but had no significant effect on NAG, PER and POX activities at both 0-5 and 5-10 cm soil depths. Long-term root removal significantly reduced BG activity at 0-5 cm soil depth, while increased PER activity at both soil depths. Long-term root removal and warming treatment significantly reduced AP and BG activities at 0-5 cm soil depth, but had no significant effect on activities of other enzymes at both soil depths. The results of redundancy analysis showed that soil water content and NH4+-N content were likely important factors driving the changes soil enzyme activities among treatments. This research provides critical information on the activities of soil enzymes related to carbon, nitrogen and phosphorus cycling in response to global change in this subtropical forest ecosystem. Keywords:soil extracellular enzyme activity;ecological stoichiometry;litter removal;root removal;warming
PDF (1307KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 刘珊杉, 周文君, 况露辉, 刘占锋, 宋清海, 刘运通, 张一平, 鲁志云, 沙丽清. 亚热带常绿阔叶林土壤胞外酶活性对碳输入变化及增温的响应. 植物生态学报, 2020, 44(12): 1262-1272. DOI: 10.17521/cjpe.2020.0310 LIU Shan-Shan, ZHOU Wen-Jun, KUANG Lu-Hui, LIU Zhan-Feng, SONG Qing-Hai, LIU Yun- Tong, ZHANG Yi-Ping, LU Zhi-Yun, SHA Li-Qing. Responses of soil extracellular enzyme activities to carbon input alteration and warming in a subtropical evergreen broad-leaved forest. Chinese Journal of Plant Ecology, 2020, 44(12): 1262-1272. DOI: 10.17521/cjpe.2020.0310
利用哀牢山亚热带常绿阔叶林模拟增温实验平台, 林下随机设置20个90 cm × 90 cm × 50 cm (长×宽×高)的呼吸箱(每个呼吸箱覆盖面积为0.81 m2), 分别进行4种处理: 对照(CK)、切根(TR)、切根并增温(TR+W)、去凋(LR), 每种处理分别设置5个重复(Wu et al., 2014), 土壤酶相关实验均在呼吸箱内采样。各处理样方面积均为0.81 m2, 对照不做实验处理。
土壤含水量(SWC)采用烘干称质量法测定(张学礼等, 2005)。土壤溶解有机碳(DOC)、溶解有机氮(DON)和溶解总氮(DN)含量用TOC/TN分析仪(Liqui TOC II, Elementar Analyzer system GmbH, Frankfurt, Germany)测定, 方法为Pt-催化高温煅烧法(680 ℃)(Zhou et al., 2019)。土壤硝态氮(NO3--N)和铵态氮(NH4+-N)含量使用连续流动分析仪(Skalar San++, Breda, the Netherlands)测定(Jones & Willett, 2006)。土壤矿质氮(Mineral-N)是以离子态存在于土壤中的氮, 以土壤NH4+-N及NO3--N含量相加进行计算。
Table 3 表3 表3不同处理下亚热带常绿阔叶林土壤基本理化性质(平均值±标准偏差, n = 5) Table 3Basic physical and chemical properties of soil in the subtropical evergreen broad-leaved forest under different treatments (mean ± SD, n = 5)
CK, 对照处理; LR, 去除地表凋落物处理; TR, 切根处理; TR+W, 切根+增温处理。不同小写字母表示相同深度不同处理间差异显著(p < 0.05), 不同大写字母表示相同处理不同深度土壤之间差异显著(p < 0.05)。 CK, control; LR, litter removal; TR, root removal; TR+W, root removal + warming. Different lowercase letters indicate significant differences among different treatments at the same soil depth (p < 0.05). Different uppercase letters indicate significant differences among different soil depths in the same treatment (p < 0.05).
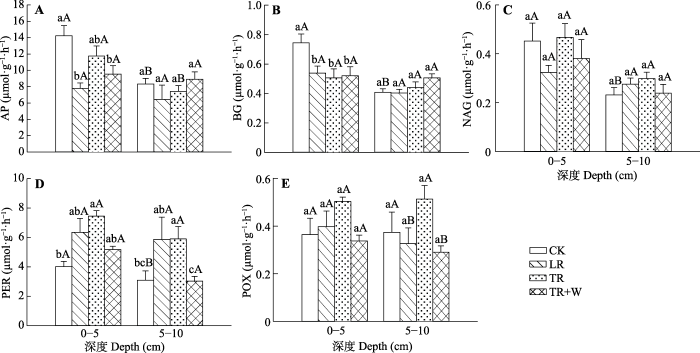
CK, 对照处理; LR, 去除地表凋落物处理; TR, 切根处理; TR+W, 切根+增温处理。不同小写字母表示相同深度不同处理间差异显著(p < 0.05), 不同大写字母表示相同处理不同深度土壤之间差异显著(p < 0.05)。AP, 酸性磷酸酶; BG, β-葡萄糖苷酶; NAG, β-1,4-N-乙酰氨基葡萄糖苷酶; PER, 过氧化物酶; POX, 多酚氧化酶。 Fig. 1Changes of enzyme activities under different treatments and depths in the subtropical evergreen broad-leaved forest (mean + SD, n = 5).
CK, control; LR, litter removal; TR, root removal; TR+W, root removal + warming. Different lowercase letters indicate significant differences among different treatments at the same soil depth (p < 0.05). Different uppercase letters indicate significant differences among different soil depths in the same treatment (p < 0.05). AP, acid phosphatase; BG, β-glucosidase; NAG, β-1,4-N- acetylglucosaminidase; PER, peroxidase; POX, polyphenol oxidase.
CK, 对照处理; LR, 去除地表凋落物处理; TR, 切根处理; TR+W, 切根+增温处理。不同小写字母表示相同深度不同处理间差异显著(p < 0.05), 不同大写字母表示相同处理不同深度土壤之间差异显著(p < 0.05)。AP, 酸性磷酸酶; BG, β-葡萄糖苷酶; NAG, β-1,4-N-乙酰氨基葡萄糖苷酶。 Fig. 2Stoichiometric ratios of soil enzymes at different treatments and depths in the subtropical evergreen broad-leaved forest (mean + SD, n = 5).
CK, control; LR, litter removal; TR, root removal; TR+W, root removal + warming. Different lowercase letters indicate significant differences among different treatments at the same soil depth (p < 0.05). Different uppercase letters indicate significant differences among different soil depths in the same treatment (p < 0.05). AP, acid phosphatase; BG, β-glucosidase; NAG, β-1,4-N-acetylglucosaminidase.
图3
新窗口打开|下载原图ZIP|生成PPT 图3亚热带常绿阔叶林0-5 (A、B、C)和5-10 cm (D、E、F)土层BG、NAG和AP活性关系的标准主轴回归分析(n = 20)。
CK, 对照处理; LR, 去除地表凋落物处理; TR, 切根处理; TR+W, 切根+增温处理。AP, 酸性磷酸酶; BG, β-葡萄糖苷酶; NAG, β-1,4-N-乙酰氨基葡萄糖苷酶。 Fig. 3Standardized major axis regressions of the log-transformed soil BG, NAG and AP activities in 0-5 (A, B, C) and 5-10 cm (D, E, F) soil layers in the subtropical evergreen broad-leaved forest (n = 20).
AllisonSD, VitousekPM (2004). Extracellular enzyme activities and carbon chemistry as drivers of tropical plant litter decomposition Biotropica, 36, 285-296. [本文引用: 1]
AllisonSD, WeintraubMN, GartnerTB, WaldropMP (2010). Evolutionary-economic principles as regulators of soil enzyme production and ecosystem function//Shukla G, Varma A. Soil Enzymology Springer-Verlag, Berlin. 229-243. [本文引用: 1]
BaldrianP (2009). Ectomycorrhizal fungi and their enzymes in soils: Is there enough evidence for their role as facultative soil saprotrophs? Oecologia, 161, 657-660. URLPMID:19685081 [本文引用: 1]
BrzostekER, GrecoA, DrakeJE, FinziAC (2013). Root carbon inputs to the rhizosphere stimulate extracellular enzyme activity and increase nitrogen availability in temperate forest soils Biogeochemistry, 115, 65-76. [本文引用: 1]
ChaerGM, MyroldDD, BottomleyPJ (2009). A soil quality index based on the equilibrium between soil organic matter and biochemical properties of undisturbed coniferous forest soils of the Pacific Northwest Soil Biology & Biochemistry, 41, 822-830. [本文引用: 1]
ChenJ, ElsgaardL, vanGroenigen KJ, OlesenJE, LiangZ, JiangY, LaerkePE, ZhangYF, LuoYQ, HungateBA, SinsabaughRL, J?rgensenU (2020). Soil carbon loss with warming: new evidence from carbon-degrading enzymes Global Change Biology, 26, 1944-1952. [本文引用: 1]
ClevelandCC, LiptzinD (2007). C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass? Biogeochemistry, 85, 235-252. [本文引用: 1]
CrowSE, LajthaK, FilleyTR, SwanstonCW, BowdenRD, CaldwellBA (2009). Sources of plant-derived carbon and stability of organic matter in soil: implications for global change Global Change Biology, 15, 2003-2019. [本文引用: 1]
FeketeI, KotroczóZ, VargaC, NagyPT, VárbíróG, BowdenRD, TóthJA, LajthaK (2014). Alterations in forest detritus inputs influence soil carbon concentration and soil respiration in a Central-European deciduous forest Soil Biology & Biochemistry, 74, 106-114. [本文引用: 1]
GeXG, XiaoWF, ZengLX, HuangZL, ZhouBZ, SchaubM, LiMH (2017). Relationships between soil-litter interface enzyme activities and decomposition in Pinus massoniana plantations in China Journal of Soils and Sediments, 17, 996-1008. [本文引用: 1]
GianfredaL (2015). Enzymes of importance to rhizosphere processes Journal of Soil Science and Plant Nutrition, 15, 283-306. [本文引用: 1]
HarringtonRA, FownesJH, VitousekPM (2001). Production and resource use efficiencies in N- and P-limited tropical forests: a comparison of responses to long-term fertilization Ecosystems, 4, 646-657. [本文引用: 1]
HillBH, ElonenCM, JichaTM, KolkaRK, LehtoLLP, SebestyenSD, Seifert-MonsonLR (2014). Ecoenzymatic stoichiometry and microbial processing of organic matter in northern bogs and fens reveals a common P-limitation between peatland types Biogeochemistry, 120, 203-224. [本文引用: 1]
HuR, WangXP, XuJS, ZhangYF, PanYX, SuX (2020). Themechanism of soil nitrogen transformation under different biocrusts to warming and reduced precipitation: from microbial functional genes to enzyme activity Science of the Total Environment, 722, 137849. DOI: 10.1016/j.scitotenv. 2020.137849. [本文引用: 1]
IPCC (2013). Climate Change 2013: the Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge. [本文引用: 1]
JiangMH, NiMY, ZhouJC, ChenYM, YangYS (2018). Effects of warming and precipitation reduction on soil enzyme activity in a young Cunning-hamia lanceolata plantation Chinese Journal of Ecology, 37, 3210-3219. [本文引用: 1]
JonesDL, WillettVB (2006). Experimental evaluation of methods to quantify Dissolved organic nitrogen (DON) and Dissolved organic carbon (DOC) in soil Soil Biology & Biochemistry, 38, 991-999. [本文引用: 1]
KangH, LeeD (2005). Inhibition of extracellular enzyme activities in a forest soil by additions of inorganic nitrogen Communications in Soil Science and Plant Analysis, 36, 2129-2135. [本文引用: 2]
KarhuK, AuffretMD, DungaitJAJ, HopkinsDW, ProsserJI, SinghBK, SubkeJA, WookeyPA, AgrenGI, SebastiàMT, GouriveauF, BergkvistG, MeirP, NottinghamAT, SalinasN, HartleyIP (2014). Temperature sensitivity of soil respiration rates enhanced by microbial community response Nature, 513, 81-84. URLPMID:25186902
LiGL, KimS, HanSH, ChangHN, DuDL, SonY (2018). Precipitation affects soil microbial and extracellular enzymatic responses to warming Soil Biology & Biochemistry, 120, 212-221. [本文引用: 1]
LuM, ZhouXH, YangQ, LiH, LouYQ, FangCM, ChenJK, YangX, LiB (2013). Responses of ecosystem carbon cycle to experimental warming: a meta-analysis Ecology, 94, 726-738. URLPMID:23687898 [本文引用: 1]
LuZY (2016). Effects of Increasing Temperature and Root Cutting on Soil Nitrogen Mineralization in Evergreen Broad-leaved Forest of Ailao Mountain Master degree dissertation, University of Chinese Academy of Sciences, Beijing. [本文引用: 2]
LuoYQ, SherryR, ZhouXH, WanSQ (2009). Terrestrial carbon- cycle feedback to climate warming: experimental evidence on plant regulation and impacts of biofuel feedstock harvest Global Change Biology: Bioenergy, 1, 62-74. [本文引用: 1]
MartensDA, JohansonJB, FrankenbergerWT JR (1992). Production and persistence of soil enzymes with repeated addition of organic residues Soil Science, 153, 53-61. [本文引用: 1]
McDanielMD, KayeJP, KayeMW (2013). Increased temperature and precipitation had limited effects on soil extracellular enzyme activities in a post-harvest forest Soil Biology & Biochemistry, 56, 90-98. [本文引用: 1]
MichalakA (2006). Phenolic compounds and their antioxidant activity in plants growing under heavy metal stress Polish Journal of Environmental Studies, 15, 523-530. [本文引用: 1]
MoorheadDL, SinsabaughRL, HillBH, WeintraubMN (2016). Vector analysis of ecoenzyme activities reveal constraints on coupled C, N and P dynamics Soil Biology & Biochemistry, 93, 1-7. [本文引用: 1]
PengXQ, WangW (2016). Stoichiometry of soil extracellular enzyme activity along a climatic transect in temperate grasslands of Northern China Soil Biology & Biochemistry, 98, 74-84. [本文引用: 1]
Piotrowska-D?ugoszA, CharzyńskiP (2015). The impact of the soil sealing degree on microbial biomass, enzymatic activity, and physicochemical properties in the Ekranic Technosols of Toruń (Poland) Journal of Soils and Sediments, 15, 47-59. [本文引用: 1]
RenCJ, ZhaoFZ, ShiZ, ChenJ, HanXH, YangGH, FengYZ, RenGX (2017). Differential responses of soil microbial biomass and carbon-degrading enzyme activities to altered precipitation Soil Biology & Biochemistry, 115, 1-10. [本文引用: 1]
RustadL, CampbellJ, MarionG, NorbyR, MitchellM, HartleyA, CornelissenJ, GurevitchJ, GCTE-NEWS (2001). A meta-analysis of the response of soil respiration, net nitrogen mineralization, and aboveground plant growth to experimental ecosystem warming Oecologia, 126, 543-562. DOI:10.1007/s004420000544URLPMID:28547240 [本文引用: 1] Climate change due to greenhouse gas emissions is predicted to raise the mean global temperature by 1.0-3.5 degrees C in the next 50-100 years. The direct and indirect effects of this potential increase in temperature on terrestrial ecosystems and ecosystem processes are likely to be complex and highly varied in time and space. The Global Change and Terrestrial Ecosystems core project of the International Geosphere-Biosphere Programme has recently launched a Network of Ecosystem Warming Studies, the goals of which are to integrate and foster research on ecosystem-level effects of rising temperature. In this paper, we use meta-analysis to synthesize data on the response of soil respiration, net N mineralization, and aboveground plant productivity to experimental ecosystem warming at 32 research sites representing four broadly defined biomes, including high (latitude or altitude) tundra, low tundra, grassland, and forest. Warming methods included electrical heat-resistance ground cables, greenhouses, vented and unvented field chambers, overhead infrared lamps, and passive night-time warming. Although results from individual sites showed considerable variation in response to warming, results from the meta-analysis showed that, across all sites and years, 2-9 years of experimental warming in the range 0.3-6.0 degrees C significantly increased soil respiration rates by 20% (with a 95% confidence interval of 18-22%), net N mineralization rates by 46% (with a 95% confidence interval of 30-64%), and plant productivity by 19% (with a 95% confidence interval of 15-23%). The response of soil respiration to warming was generally larger in forested ecosystems compared to low tundra and grassland ecosystems, and the response of plant productivity was generally larger in low tundra ecosystems than in forest and grassland ecosystems. With the exception of aboveground plant productivity, which showed a greater positive response to warming in colder ecosystems, the magnitude of the response of these three processes to experimental warming was not generally significantly related to the geographic, climatic, or environmental variables evaluated in this analysis. This underscores the need to understand the relative importance of specific factors (such as temperature, moisture, site quality, vegetation type, successional status, land-use history, etc.) at different spatial and temporal scales, and suggests that we should be cautious in
SardansJ, Pe?uelasJ, EstiarteM (2008). Changes in soil enzymes related to C and N cycle and in soil C and N content under prolonged warming and drought in a Mediterranean shrubland Applied Soil Ecology, 39, 223-235. [本文引用: 2]
SeoJ, JangI, JungJY, LeeYK, KangH (2015). Warming and increased precipitation enhance phenol oxidase activity in soil while warming induces drought stress in vegetation of an Arctic ecosystem Geoderma, 259- 260, 347-353. [本文引用: 1]
SinsabaughRL, CarreiroMM, RepertDA (2002). Allocation of extracellular enzymatic activity in relation to litter composition, N deposition, and mass loss Biogeochemistry, 60, 1-24.
SinsabaughRL, FollstadShah JJ (2012). Ecoenzymatic stoichiometry and ecological theory Annual Review of Ecology, Evolution, and Systematics, 43, 313-343. [本文引用: 1]
SinsabaughRL, HillBH, FollstadShah JJ (2009). Ecoenzymatic stoichiometry of microbial organic nutrient acquisition in soil and sediment Nature, 462, 795-798. URLPMID:20010687 [本文引用: 3]
SinsabaughRL, LauberCL, WeintraubMN, AhmedB, AllisonSD, CrenshawC, ContostaAR, CusackD, FreyS, GalloME, GartnerTB, HobbieSE, HollandK, KeelerBL, PowersJS, StursovaM, Takacs-VesbachC, WaldropMP, WallensteinMD, ZakDR, ZeglinLH (2008). Stoichiometry of soil enzyme activity at global scale Ecology Letters, 11, 1252-1264. DOI:10.1111/j.1461-0248.2008.01245.xURLPMID:18823393 [本文引用: 3] Extracellular enzymes are the proximate agents of organic matter decomposition and measures of these activities can be used as indicators of microbial nutrient demand. We conducted a global-scale meta-analysis of the seven-most widely measured soil enzyme activities, using data from 40 ecosystems. The activities of beta-1,4-glucosidase, cellobiohydrolase, beta-1,4-N-acetylglucosaminidase and phosphatase g(-1) soil increased with organic matter concentration; leucine aminopeptidase, phenol oxidase and peroxidase activities showed no relationship. All activities were significantly related to soil pH. Specific activities, i.e. activity g(-1) soil organic matter, also varied in relation to soil pH for all enzymes. Relationships with mean annual temperature (MAT) and precipitation (MAP) were generally weak. For hydrolases, ratios of specific C, N and P acquisition activities converged on 1 : 1 : 1 but across ecosystems, the ratio of C : P acquisition was inversely related to MAP and MAT while the ratio of C : N acquisition increased with MAP. Oxidative activities were more variable than hydrolytic activities and increased with soil pH. Our analyses indicate that the enzymatic potential for hydrolyzing the labile components of soil organic matter is tied to substrate availability, soil pH and the stoichiometry of microbial nutrient demand. The enzymatic potential for oxidizing the recalcitrant fractions of soil organic material, which is a proximate control on soil organic matter accumulation, is most strongly related to soil pH. These trends provide insight into the biogeochemical processes that create global patterns in ecological stoichiometry and organic matter storage.
SpohnM, KuzyakovY (2014). Spatial and temporal dynamics of hotspots of enzyme activity in soil as affected by living and dead roots—A soil zymography analysis Plant and Soil, 379, 67-77. [本文引用: 1]
SteinwegJM, DukesJS, PaulEA, WallensteinMD (2013). Microbial responses to multi-factor climate change: effects on soil enzymes Frontiers in Microbiology, 4, 146. DOI: 10.3389/fmicb.2013.00146. DOI:10.3389/fmicb.2013.00146URLPMID:23781218 [本文引用: 1] The activities of extracellular enzymes, the proximate agents of decomposition in soils, are known to depend strongly on temperature, but less is known about how they respond to changes in precipitation patterns, and the interaction of these two components of climate change. Both enzyme production and turnover can be affected by changes in temperature and soil moisture, thus it is difficult to predict how enzyme pool size may respond to altered climate. Soils from the Boston-Area Climate Experiment (BACE), which is located in an old field (on abandoned farmland), were used to examine how climate variables affect enzyme activities and microbial biomass carbon (MBC) in different seasons and in soils exposed to a combination of three levels of precipitation treatments (ambient, 150% of ambient during growing season, and 50% of ambient year-round) and four levels of warming treatments (unwarmed to ~4 degrees C above ambient) over the course of a year. Warming, precipitation and season had very little effect on potential enzyme activity. Most models assume that enzyme dynamics follow microbial biomass, because enzyme production should be directly controlled by the size and activity of microbial biomass. We observed differences among seasons and treatments in mass-specific potential enzyme activity, suggesting that this assumption is invalid. In June 2009, mass-specific potential enzyme activity, using chloroform fumigation-extraction MBC, increased with temperature, peaking under medium warming and then declining under the highest warming. This finding suggests that either enzyme production increased with temperature or turnover rates decreased. Increased maintenance costs associated with warming may have resulted in increased mass-specific enzyme activities due to increased nutrient demand. Our research suggests that allocation of resources to enzyme production could be affected by climate-induced changes in microbial efficiency and maintenance costs.
SteinwegJM, DukesJS, WallensteinMD (2012). Modeling the effects of temperature and moisture on soil enzyme activity: linking laboratory assays to continuous field data Soil Biology & Biochemistry, 55, 85-92. [本文引用: 1]
StoneMM, DeForestJL, PlanteAF (2014). Changes in extracellular enzyme activity and microbial community structure with soil depth at the Luquillo Critical Zone Observatory Soil Biology & Biochemistry, 75, 237-247. [本文引用: 1]
StoneMM, WeissMS, GoodaleCL, AdamsMB, FernandezIJ, GermanDP, AllisonSD (2012). Temperature sensitivity of soil enzyme kinetics under N-fertilization in two temperate forests Global Change Biology, 18, 1173-1184. [本文引用: 1]
TanZH, ZhangYP, LiangNS, HsiaYJ, ZhangYJ, ZhouGY, LiYL, JuangJY, ChuHS, YanJH, YuGR, SunXM, SongQH, CaoKF, SchaeferDA, LiuYH (2012). An observational study of the carbon-sink strength of East Asian subtropical evergreen forests Environmental Research Letters, 7, 044017. DOI: 10.1088/1748-9326/7/4/044017. [本文引用: 1]
VeresZ, KotroczóZ, FeketeI, TóthJA, LajthaK, TownsendK, TóthmérészB (2015). Soil extracellular enzyme activities are sensitive indicators of detrital inputs and carbon availability Applied Soil Ecology, 92, 18-23. [本文引用: 1]
WangSP, DuanJC, XuGP, WangYF, ZhangZH, RuiYC, LuoCY, XuB, ZhuXX, ChangXF, CuiXY, NiuHS, ZhaoXQ, WangWY (2012). Effects of warming and grazing on soil N availability, species composition, and ANPP in an alpine meadow Ecology, 93, 2365-2376. URLPMID:23236908 [本文引用: 1]
WangXC, LuQ (2006). Beta-glucosidase activity in paddy soils of the Taihu Lake region, China Pedosphere, 16, 118-124. [本文引用: 2]
WaringBG, WeintraubSR, SinsabaughRL (2014). Ecoenzymatic stoichiometry of microbial nutrient acquisition in tropical soils Biogeochemistry, 117, 101-113. [本文引用: 2]
WartonDI, DuursmaRA, FalsterDS, TaskinenS (2012). Smatr 3—An R package for estimation and inference about allometric lines Methods in Ecology and Evolution, 3, 257-259.
WeiCC, LiuXF, LinCF, LiXF, LiY, ZhengYX (2018). Response of soil enzyme activities to litter input changes in two secondary Castanopsis carlessii forests in subtropical China Chinese Journal of Plant Ecology, 42, 692-702. [本文引用: 3]
WeintraubSR, WiederWR, ClevelandCC, TownsendAR (2013). Organic matter inputs shift soil enzyme activity and allocation patterns in a wet tropical forest Biogeochemistry, 114, 313-326. [本文引用: 1]
WuCS, LiangNS, ShaLQ, XuXL, ZhangYP, LuHZ, SongL, SongQH, XieYN (2016). Heterotrophic respiration does not acclimate to continuous warming in a subtropical forest Scientific Reports, 6, 21561. DOI: 10.1038/srep21561. URLPMID:26900028 [本文引用: 3]
WuCS, ZhangYP, XuXL, ShaLQ, YouGY, LiuYH, XieYN (2014). Influence of interactions between litter decomposition and rhizosphere activity on soil respiration and on the temperature sensitivity in a subtropical montane forest in SW China Plant and Soil, 381, 215-224. [本文引用: 3]
XiaoW, ChenX, JingX, ZhuB (2018). A meta-analysis of soil extracellular enzyme activities in response to global change Soil Biology & Biochemistry, 123, 21-32. [本文引用: 1]
YangWQ, WangKY (2004). Advances in forest soil enzymology Scientia Silvae Sinicae, 40, 152-159. [本文引用: 1]
ZhengWG, XueL, XuPB, LiangLL, FengHF (2011). Soil response to litter in a Pinus caribaea woodland Journal of South China Agricultural University, 32, 120-123. [本文引用: 1]
ZhouLG, LiuYT, ZhangYP, ShaLQ, SongQH, ZhouWJ, BalasubramanianD, PalingamoorthyG, GaoJB, LinYX, LiJ, ZhouRW, ZarMyo ST, TangXH, ZhangJ, ZhangP, WangSS, GraceJ (2019). Soil respiration after six years of continuous drought stress in the tropical rainforest in Southwest China Soil Biology & Biochemistry, 138, 107564. DOI: 10.1016/j.soilbio.2019.107564. [本文引用: 1]
ZhouXQ, ChenCR, WangYF, XuZH, HanHY, LiLH, WanSQ (2013). Warming and increased precipitation have differential effects on soil extracellular enzyme activities in a temperate grassland Science of the Total Environment, 444, 552-558. [本文引用: 1]
ZhuH (2016). Discussion on the origin of mid-montane wet evergreen broad-leaved forest in Yunnan Plant Science Journal, 34, 715-723. [本文引用: 2]
Root carbon inputs to the rhizosphere stimulate extracellular enzyme activity and increase nitrogen availability in temperate forest soils 1 2013
... 由于土壤酶主要来自土壤微生物及植物根系分泌的具有催化作用的蛋白质, 因此根系分泌物是土壤酶的重要来源(杨万勤和王开运, 2004).同时在森林生态系统中, 凋落物是土壤有机质的重要来源, 凋落物输入量的改变会造成土壤微生物及其代谢方式的改变, 进而影响土壤胞外酶活性, 故碳源输入方式的改变可对土壤胞外酶活性产生影响.森林土壤中与碳、氮、磷循环相关的胞外酶活性对土壤有机质的输入较为敏感.Weintraub等(2013)发现, 减少凋落物输入后, 热带森林土壤磷限制性增强, 与磷循环相关的酶活性增强; 魏翠翠等(2018)研究发现凋落物加倍和凋落物去除均降低了亚热带森林酸性磷酸酶、β-葡萄糖苷酶和多酚氧化酶的活性; Ge等(2017)对亚热带马尾松林研究发现, 凋落物输入量与纤维素酶、脲酶和多酚氧化酶活性呈显著线性相关关系, 并证明土壤温度、凋落物含水量和凋落物氮含量是影响酶活性的主要因素.由此可见, 凋落物输入变化会引起微生物碳源和养分源变化, 进而影响土壤胞外酶活性(Allison & Vitousek, 2004).根系分泌物作为土壤酶的重要来源, 也发挥着重要作用, 研究表明根际土壤中有机碳含量较高, 根际碳的输入可以刺激根际微生物活动和胞外酶的活性, 因此根际土壤的酶活性普遍高于整体(Brzostek et al., 2013; Gianfreda, 2015).切根通常会导致土壤有机碳含量的下降(Crow et al., 2009; Fekete et al., 2014), 随之影响土壤酶活性.Spohn和Kuzyakov (2014)研究表明切根后纤维素酶和几丁质酶活性均升高, 但根系与胞外酶活性的相互影响机制还缺乏进一步研究. ...
A soil quality index based on the equilibrium between soil organic matter and biochemical properties of undisturbed coniferous forest soils of the Pacific Northwest 1 2009
Soil carbon loss with warming: new evidence from carbon-degrading enzymes 1 2020
... 土壤酶是来自土壤微生物、植物和动物的生物活性物质, 是专一生物化学反应的生物催化剂, 作为土壤生物地球化学过程的积极参与者, 在森林生态系统的物质循环和能量流动过程中扮演着重要的角色(Sardans et al., 2008).土壤中以游离态存在或与土壤有机质及矿质组分结合的酶称为土壤胞外酶.当温度、降水量和底物输入产生变化时, 土壤胞外酶活性将会发生相应改变(Seo et al., 2015; Chen et al., 2020; Hu et al., 2020), 表现为促进或抑制土壤养分、有机物分解, 进而影响陆地生态系统的物质循环过程(Karhu et al., 2014).因此, 气候变化可能对陆地生态系统土壤胞外酶活性产生复杂的影响, 而探讨各环境因子的变化对土壤胞外酶活性的影响是预测未来气候变化对森林生态系统过程影响的关键(Xiao et al., 2018). ...
C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass? 1 2007
... 由于微生物生物量C:N:P具有一定的稳定性(Cleveland & Liptzin, 2007), 因此土壤酶生态化学计量比也表现为相对稳定(Sinsabaugh et al., 2008).该研究中酶C:P和N:P远低于全球主要陆地生态系统土壤酶C:P和N:P的平均值0.62和0.44, 而酶C:N则略高于平均值1.14 (Sinsabaugh et al., 2009), 说明AP活性远远高于BG和NAG活性, 基于全球尺度上BG、NAG和AP对数转化后比值约为1:1:1 (Sinsabaugh et al., 2008).本研究中两个土层比值大体相同, 都表现为lg (AP)比值偏大, 说明该地区受磷限制.Allison等(2010)研究表明当微生物的生长受到某元素限制时, 会增加与该元素相关胞外酶的活性, 因此该地土壤AP活性较高, 与Waring等(2014)的研究结果一致.另有研究表明丰富的降水也会增加表层土壤中磷的淋溶, 这会进一步降低磷的有效性(Waring et al., 2014), 本研究样地年降水量可达1 800 mm以上, 且集中在5-10月, 属于典型的湿性常绿阔叶林(朱华, 2016), 土壤磷淋溶强烈, 本地土壤AP活性偏高和降水量大联系紧密. ...
Sources of plant-derived carbon and stability of organic matter in soil: implications for global change 1 2009
... 由于土壤酶主要来自土壤微生物及植物根系分泌的具有催化作用的蛋白质, 因此根系分泌物是土壤酶的重要来源(杨万勤和王开运, 2004).同时在森林生态系统中, 凋落物是土壤有机质的重要来源, 凋落物输入量的改变会造成土壤微生物及其代谢方式的改变, 进而影响土壤胞外酶活性, 故碳源输入方式的改变可对土壤胞外酶活性产生影响.森林土壤中与碳、氮、磷循环相关的胞外酶活性对土壤有机质的输入较为敏感.Weintraub等(2013)发现, 减少凋落物输入后, 热带森林土壤磷限制性增强, 与磷循环相关的酶活性增强; 魏翠翠等(2018)研究发现凋落物加倍和凋落物去除均降低了亚热带森林酸性磷酸酶、β-葡萄糖苷酶和多酚氧化酶的活性; Ge等(2017)对亚热带马尾松林研究发现, 凋落物输入量与纤维素酶、脲酶和多酚氧化酶活性呈显著线性相关关系, 并证明土壤温度、凋落物含水量和凋落物氮含量是影响酶活性的主要因素.由此可见, 凋落物输入变化会引起微生物碳源和养分源变化, 进而影响土壤胞外酶活性(Allison & Vitousek, 2004).根系分泌物作为土壤酶的重要来源, 也发挥着重要作用, 研究表明根际土壤中有机碳含量较高, 根际碳的输入可以刺激根际微生物活动和胞外酶的活性, 因此根际土壤的酶活性普遍高于整体(Brzostek et al., 2013; Gianfreda, 2015).切根通常会导致土壤有机碳含量的下降(Crow et al., 2009; Fekete et al., 2014), 随之影响土壤酶活性.Spohn和Kuzyakov (2014)研究表明切根后纤维素酶和几丁质酶活性均升高, 但根系与胞外酶活性的相互影响机制还缺乏进一步研究. ...
Alterations in forest detritus inputs influence soil carbon concentration and soil respiration in a Central-European deciduous forest 1 2014
... 由于土壤酶主要来自土壤微生物及植物根系分泌的具有催化作用的蛋白质, 因此根系分泌物是土壤酶的重要来源(杨万勤和王开运, 2004).同时在森林生态系统中, 凋落物是土壤有机质的重要来源, 凋落物输入量的改变会造成土壤微生物及其代谢方式的改变, 进而影响土壤胞外酶活性, 故碳源输入方式的改变可对土壤胞外酶活性产生影响.森林土壤中与碳、氮、磷循环相关的胞外酶活性对土壤有机质的输入较为敏感.Weintraub等(2013)发现, 减少凋落物输入后, 热带森林土壤磷限制性增强, 与磷循环相关的酶活性增强; 魏翠翠等(2018)研究发现凋落物加倍和凋落物去除均降低了亚热带森林酸性磷酸酶、β-葡萄糖苷酶和多酚氧化酶的活性; Ge等(2017)对亚热带马尾松林研究发现, 凋落物输入量与纤维素酶、脲酶和多酚氧化酶活性呈显著线性相关关系, 并证明土壤温度、凋落物含水量和凋落物氮含量是影响酶活性的主要因素.由此可见, 凋落物输入变化会引起微生物碳源和养分源变化, 进而影响土壤胞外酶活性(Allison & Vitousek, 2004).根系分泌物作为土壤酶的重要来源, 也发挥着重要作用, 研究表明根际土壤中有机碳含量较高, 根际碳的输入可以刺激根际微生物活动和胞外酶的活性, 因此根际土壤的酶活性普遍高于整体(Brzostek et al., 2013; Gianfreda, 2015).切根通常会导致土壤有机碳含量的下降(Crow et al., 2009; Fekete et al., 2014), 随之影响土壤酶活性.Spohn和Kuzyakov (2014)研究表明切根后纤维素酶和几丁质酶活性均升高, 但根系与胞外酶活性的相互影响机制还缺乏进一步研究. ...
Relationships between soil-litter interface enzyme activities and decomposition in Pinus massoniana plantations in China 1 2017
... 由于土壤酶主要来自土壤微生物及植物根系分泌的具有催化作用的蛋白质, 因此根系分泌物是土壤酶的重要来源(杨万勤和王开运, 2004).同时在森林生态系统中, 凋落物是土壤有机质的重要来源, 凋落物输入量的改变会造成土壤微生物及其代谢方式的改变, 进而影响土壤胞外酶活性, 故碳源输入方式的改变可对土壤胞外酶活性产生影响.森林土壤中与碳、氮、磷循环相关的胞外酶活性对土壤有机质的输入较为敏感.Weintraub等(2013)发现, 减少凋落物输入后, 热带森林土壤磷限制性增强, 与磷循环相关的酶活性增强; 魏翠翠等(2018)研究发现凋落物加倍和凋落物去除均降低了亚热带森林酸性磷酸酶、β-葡萄糖苷酶和多酚氧化酶的活性; Ge等(2017)对亚热带马尾松林研究发现, 凋落物输入量与纤维素酶、脲酶和多酚氧化酶活性呈显著线性相关关系, 并证明土壤温度、凋落物含水量和凋落物氮含量是影响酶活性的主要因素.由此可见, 凋落物输入变化会引起微生物碳源和养分源变化, 进而影响土壤胞外酶活性(Allison & Vitousek, 2004).根系分泌物作为土壤酶的重要来源, 也发挥着重要作用, 研究表明根际土壤中有机碳含量较高, 根际碳的输入可以刺激根际微生物活动和胞外酶的活性, 因此根际土壤的酶活性普遍高于整体(Brzostek et al., 2013; Gianfreda, 2015).切根通常会导致土壤有机碳含量的下降(Crow et al., 2009; Fekete et al., 2014), 随之影响土壤酶活性.Spohn和Kuzyakov (2014)研究表明切根后纤维素酶和几丁质酶活性均升高, 但根系与胞外酶活性的相互影响机制还缺乏进一步研究. ...
Enzymes of importance to rhizosphere processes 1 2015
... 由于土壤酶主要来自土壤微生物及植物根系分泌的具有催化作用的蛋白质, 因此根系分泌物是土壤酶的重要来源(杨万勤和王开运, 2004).同时在森林生态系统中, 凋落物是土壤有机质的重要来源, 凋落物输入量的改变会造成土壤微生物及其代谢方式的改变, 进而影响土壤胞外酶活性, 故碳源输入方式的改变可对土壤胞外酶活性产生影响.森林土壤中与碳、氮、磷循环相关的胞外酶活性对土壤有机质的输入较为敏感.Weintraub等(2013)发现, 减少凋落物输入后, 热带森林土壤磷限制性增强, 与磷循环相关的酶活性增强; 魏翠翠等(2018)研究发现凋落物加倍和凋落物去除均降低了亚热带森林酸性磷酸酶、β-葡萄糖苷酶和多酚氧化酶的活性; Ge等(2017)对亚热带马尾松林研究发现, 凋落物输入量与纤维素酶、脲酶和多酚氧化酶活性呈显著线性相关关系, 并证明土壤温度、凋落物含水量和凋落物氮含量是影响酶活性的主要因素.由此可见, 凋落物输入变化会引起微生物碳源和养分源变化, 进而影响土壤胞外酶活性(Allison & Vitousek, 2004).根系分泌物作为土壤酶的重要来源, 也发挥着重要作用, 研究表明根际土壤中有机碳含量较高, 根际碳的输入可以刺激根际微生物活动和胞外酶的活性, 因此根际土壤的酶活性普遍高于整体(Brzostek et al., 2013; Gianfreda, 2015).切根通常会导致土壤有机碳含量的下降(Crow et al., 2009; Fekete et al., 2014), 随之影响土壤酶活性.Spohn和Kuzyakov (2014)研究表明切根后纤维素酶和几丁质酶活性均升高, 但根系与胞外酶活性的相互影响机制还缺乏进一步研究. ...
Production and resource use efficiencies in N- and P-limited tropical forests: a comparison of responses to long-term fertilization 1 2001
Ecoenzymatic stoichiometry and microbial processing of organic matter in northern bogs and fens reveals a common P-limitation between peatland types 1 2014
Themechanism of soil nitrogen transformation under different biocrusts to warming and reduced precipitation: from microbial functional genes to enzyme activity 1 2020
... 土壤酶是来自土壤微生物、植物和动物的生物活性物质, 是专一生物化学反应的生物催化剂, 作为土壤生物地球化学过程的积极参与者, 在森林生态系统的物质循环和能量流动过程中扮演着重要的角色(Sardans et al., 2008).土壤中以游离态存在或与土壤有机质及矿质组分结合的酶称为土壤胞外酶.当温度、降水量和底物输入产生变化时, 土壤胞外酶活性将会发生相应改变(Seo et al., 2015; Chen et al., 2020; Hu et al., 2020), 表现为促进或抑制土壤养分、有机物分解, 进而影响陆地生态系统的物质循环过程(Karhu et al., 2014).因此, 气候变化可能对陆地生态系统土壤胞外酶活性产生复杂的影响, 而探讨各环境因子的变化对土壤胞外酶活性的影响是预测未来气候变化对森林生态系统过程影响的关键(Xiao et al., 2018). ...
1 2013
... 工业革命以来, 全球平均气温呈上升趋势, 预计21世纪全球气温将增加0.3-4.8 ℃ (IPCC, 2013).温度是土壤酶活性的重要调控因子(Rustad et al., 2001; Kang & Lee, 2005; Luo et al., 2009), 直接影响了土壤酶活性, 进而影响土壤呼吸、土壤有机物分解等生化过程(Lu et al., 2013).McDaniel等(2013)对砍伐后的森林土壤进行模拟增温实验, 发现增温显著降低了土壤中β-葡萄糖苷酶、β-1,4-N-乙酰氨基葡萄糖苷酶和多酚氧化酶活性.但亦有研究表明, 短期增温和长期增温均未显著改变土壤胞外酶的活性(Wang et al., 2012).此外, 除土壤温度变化外, 土壤水分能直接影响土壤中的生化反应强度和土壤胞外酶活性.相关研究表明, 土壤水分与土壤胞外酶活性之间有显著的正相关关系(Ren et al., 2017; Li et al., 2018).Zhou等(2013)对温带草原的研究发现增加降雨会提高土壤碱性磷酸酶和脲酶的活性, 说明适量的增加土壤水分能够提升土壤胞外酶活性. ...
Experimental evaluation of methods to quantify Dissolved organic nitrogen (DON) and Dissolved organic carbon (DOC) in soil 1 2006
... 土壤含水量(SWC)采用烘干称质量法测定(张学礼等, 2005).土壤溶解有机碳(DOC)、溶解有机氮(DON)和溶解总氮(DN)含量用TOC/TN分析仪(Liqui TOC II, Elementar Analyzer system GmbH, Frankfurt, Germany)测定, 方法为Pt-催化高温煅烧法(680 ℃)(Zhou et al., 2019).土壤硝态氮(NO3--N)和铵态氮(NH4+-N)含量使用连续流动分析仪(Skalar San++, Breda, the Netherlands)测定(Jones & Willett, 2006).土壤矿质氮(Mineral-N)是以离子态存在于土壤中的氮, 以土壤NH4+-N及NO3--N含量相加进行计算. ...
Inhibition of extracellular enzyme activities in a forest soil by additions of inorganic nitrogen 2 2005
... 土壤酶是来自土壤微生物、植物和动物的生物活性物质, 是专一生物化学反应的生物催化剂, 作为土壤生物地球化学过程的积极参与者, 在森林生态系统的物质循环和能量流动过程中扮演着重要的角色(Sardans et al., 2008).土壤中以游离态存在或与土壤有机质及矿质组分结合的酶称为土壤胞外酶.当温度、降水量和底物输入产生变化时, 土壤胞外酶活性将会发生相应改变(Seo et al., 2015; Chen et al., 2020; Hu et al., 2020), 表现为促进或抑制土壤养分、有机物分解, 进而影响陆地生态系统的物质循环过程(Karhu et al., 2014).因此, 气候变化可能对陆地生态系统土壤胞外酶活性产生复杂的影响, 而探讨各环境因子的变化对土壤胞外酶活性的影响是预测未来气候变化对森林生态系统过程影响的关键(Xiao et al., 2018). ...
... 工业革命以来, 全球平均气温呈上升趋势, 预计21世纪全球气温将增加0.3-4.8 ℃ (IPCC, 2013).温度是土壤酶活性的重要调控因子(Rustad et al., 2001; Kang & Lee, 2005; Luo et al., 2009), 直接影响了土壤酶活性, 进而影响土壤呼吸、土壤有机物分解等生化过程(Lu et al., 2013).McDaniel等(2013)对砍伐后的森林土壤进行模拟增温实验, 发现增温显著降低了土壤中β-葡萄糖苷酶、β-1,4-N-乙酰氨基葡萄糖苷酶和多酚氧化酶活性.但亦有研究表明, 短期增温和长期增温均未显著改变土壤胞外酶的活性(Wang et al., 2012).此外, 除土壤温度变化外, 土壤水分能直接影响土壤中的生化反应强度和土壤胞外酶活性.相关研究表明, 土壤水分与土壤胞外酶活性之间有显著的正相关关系(Ren et al., 2017; Li et al., 2018).Zhou等(2013)对温带草原的研究发现增加降雨会提高土壤碱性磷酸酶和脲酶的活性, 说明适量的增加土壤水分能够提升土壤胞外酶活性. ...
Temperature sensitivity of soil respiration rates enhanced by microbial community response 2014
Precipitation affects soil microbial and extracellular enzymatic responses to warming 1 2018
... 工业革命以来, 全球平均气温呈上升趋势, 预计21世纪全球气温将增加0.3-4.8 ℃ (IPCC, 2013).温度是土壤酶活性的重要调控因子(Rustad et al., 2001; Kang & Lee, 2005; Luo et al., 2009), 直接影响了土壤酶活性, 进而影响土壤呼吸、土壤有机物分解等生化过程(Lu et al., 2013).McDaniel等(2013)对砍伐后的森林土壤进行模拟增温实验, 发现增温显著降低了土壤中β-葡萄糖苷酶、β-1,4-N-乙酰氨基葡萄糖苷酶和多酚氧化酶活性.但亦有研究表明, 短期增温和长期增温均未显著改变土壤胞外酶的活性(Wang et al., 2012).此外, 除土壤温度变化外, 土壤水分能直接影响土壤中的生化反应强度和土壤胞外酶活性.相关研究表明, 土壤水分与土壤胞外酶活性之间有显著的正相关关系(Ren et al., 2017; Li et al., 2018).Zhou等(2013)对温带草原的研究发现增加降雨会提高土壤碱性磷酸酶和脲酶的活性, 说明适量的增加土壤水分能够提升土壤胞外酶活性. ...
Responses of ecosystem carbon cycle to experimental warming: a meta-analysis 1 2013
... 工业革命以来, 全球平均气温呈上升趋势, 预计21世纪全球气温将增加0.3-4.8 ℃ (IPCC, 2013).温度是土壤酶活性的重要调控因子(Rustad et al., 2001; Kang & Lee, 2005; Luo et al., 2009), 直接影响了土壤酶活性, 进而影响土壤呼吸、土壤有机物分解等生化过程(Lu et al., 2013).McDaniel等(2013)对砍伐后的森林土壤进行模拟增温实验, 发现增温显著降低了土壤中β-葡萄糖苷酶、β-1,4-N-乙酰氨基葡萄糖苷酶和多酚氧化酶活性.但亦有研究表明, 短期增温和长期增温均未显著改变土壤胞外酶的活性(Wang et al., 2012).此外, 除土壤温度变化外, 土壤水分能直接影响土壤中的生化反应强度和土壤胞外酶活性.相关研究表明, 土壤水分与土壤胞外酶活性之间有显著的正相关关系(Ren et al., 2017; Li et al., 2018).Zhou等(2013)对温带草原的研究发现增加降雨会提高土壤碱性磷酸酶和脲酶的活性, 说明适量的增加土壤水分能够提升土壤胞外酶活性. ...
... 研究样地位于景东县太忠镇徐家坝的中国科学院哀牢山亚热带森林生态系统研究站(24.53° N, 101.02° E, 海拔2 480 m).研究区域属亚热带山地常绿阔叶林潮湿气候带, 干湿季分明, 年平均气温11.3 ℃, 年降水量1 881.5 mm, 降水主要集中在5-10月.区域内土壤为山地黄棕壤, 质地为砂壤土, 植物种类丰富、群落类型多样、垂直带谱完整、过渡性特征明显(朱华, 2016), 表层土壤理化性质见表2 (鲁志云, 2016). ...
Terrestrial carbon- cycle feedback to climate warming: experimental evidence on plant regulation and impacts of biofuel feedstock harvest 1 2009
... 工业革命以来, 全球平均气温呈上升趋势, 预计21世纪全球气温将增加0.3-4.8 ℃ (IPCC, 2013).温度是土壤酶活性的重要调控因子(Rustad et al., 2001; Kang & Lee, 2005; Luo et al., 2009), 直接影响了土壤酶活性, 进而影响土壤呼吸、土壤有机物分解等生化过程(Lu et al., 2013).McDaniel等(2013)对砍伐后的森林土壤进行模拟增温实验, 发现增温显著降低了土壤中β-葡萄糖苷酶、β-1,4-N-乙酰氨基葡萄糖苷酶和多酚氧化酶活性.但亦有研究表明, 短期增温和长期增温均未显著改变土壤胞外酶的活性(Wang et al., 2012).此外, 除土壤温度变化外, 土壤水分能直接影响土壤中的生化反应强度和土壤胞外酶活性.相关研究表明, 土壤水分与土壤胞外酶活性之间有显著的正相关关系(Ren et al., 2017; Li et al., 2018).Zhou等(2013)对温带草原的研究发现增加降雨会提高土壤碱性磷酸酶和脲酶的活性, 说明适量的增加土壤水分能够提升土壤胞外酶活性. ...
Production and persistence of soil enzymes with repeated addition of organic residues 1 1992
Increased temperature and precipitation had limited effects on soil extracellular enzyme activities in a post-harvest forest 1 2013
... 工业革命以来, 全球平均气温呈上升趋势, 预计21世纪全球气温将增加0.3-4.8 ℃ (IPCC, 2013).温度是土壤酶活性的重要调控因子(Rustad et al., 2001; Kang & Lee, 2005; Luo et al., 2009), 直接影响了土壤酶活性, 进而影响土壤呼吸、土壤有机物分解等生化过程(Lu et al., 2013).McDaniel等(2013)对砍伐后的森林土壤进行模拟增温实验, 发现增温显著降低了土壤中β-葡萄糖苷酶、β-1,4-N-乙酰氨基葡萄糖苷酶和多酚氧化酶活性.但亦有研究表明, 短期增温和长期增温均未显著改变土壤胞外酶的活性(Wang et al., 2012).此外, 除土壤温度变化外, 土壤水分能直接影响土壤中的生化反应强度和土壤胞外酶活性.相关研究表明, 土壤水分与土壤胞外酶活性之间有显著的正相关关系(Ren et al., 2017; Li et al., 2018).Zhou等(2013)对温带草原的研究发现增加降雨会提高土壤碱性磷酸酶和脲酶的活性, 说明适量的增加土壤水分能够提升土壤胞外酶活性. ...
Phenolic compounds and their antioxidant activity in plants growing under heavy metal stress 1 2006
The impact of the soil sealing degree on microbial biomass, enzymatic activity, and physicochemical properties in the Ekranic Technosols of Toruń (Poland) 1 2015
Differential responses of soil microbial biomass and carbon-degrading enzyme activities to altered precipitation 1 2017
... 工业革命以来, 全球平均气温呈上升趋势, 预计21世纪全球气温将增加0.3-4.8 ℃ (IPCC, 2013).温度是土壤酶活性的重要调控因子(Rustad et al., 2001; Kang & Lee, 2005; Luo et al., 2009), 直接影响了土壤酶活性, 进而影响土壤呼吸、土壤有机物分解等生化过程(Lu et al., 2013).McDaniel等(2013)对砍伐后的森林土壤进行模拟增温实验, 发现增温显著降低了土壤中β-葡萄糖苷酶、β-1,4-N-乙酰氨基葡萄糖苷酶和多酚氧化酶活性.但亦有研究表明, 短期增温和长期增温均未显著改变土壤胞外酶的活性(Wang et al., 2012).此外, 除土壤温度变化外, 土壤水分能直接影响土壤中的生化反应强度和土壤胞外酶活性.相关研究表明, 土壤水分与土壤胞外酶活性之间有显著的正相关关系(Ren et al., 2017; Li et al., 2018).Zhou等(2013)对温带草原的研究发现增加降雨会提高土壤碱性磷酸酶和脲酶的活性, 说明适量的增加土壤水分能够提升土壤胞外酶活性. ...
A meta-analysis of the response of soil respiration, net nitrogen mineralization, and aboveground plant growth to experimental ecosystem warming 1 2001
... 工业革命以来, 全球平均气温呈上升趋势, 预计21世纪全球气温将增加0.3-4.8 ℃ (IPCC, 2013).温度是土壤酶活性的重要调控因子(Rustad et al., 2001; Kang & Lee, 2005; Luo et al., 2009), 直接影响了土壤酶活性, 进而影响土壤呼吸、土壤有机物分解等生化过程(Lu et al., 2013).McDaniel等(2013)对砍伐后的森林土壤进行模拟增温实验, 发现增温显著降低了土壤中β-葡萄糖苷酶、β-1,4-N-乙酰氨基葡萄糖苷酶和多酚氧化酶活性.但亦有研究表明, 短期增温和长期增温均未显著改变土壤胞外酶的活性(Wang et al., 2012).此外, 除土壤温度变化外, 土壤水分能直接影响土壤中的生化反应强度和土壤胞外酶活性.相关研究表明, 土壤水分与土壤胞外酶活性之间有显著的正相关关系(Ren et al., 2017; Li et al., 2018).Zhou等(2013)对温带草原的研究发现增加降雨会提高土壤碱性磷酸酶和脲酶的活性, 说明适量的增加土壤水分能够提升土壤胞外酶活性. ...
Changes in soil enzymes related to C and N cycle and in soil C and N content under prolonged warming and drought in a Mediterranean shrubland 2 2008
... 土壤酶是来自土壤微生物、植物和动物的生物活性物质, 是专一生物化学反应的生物催化剂, 作为土壤生物地球化学过程的积极参与者, 在森林生态系统的物质循环和能量流动过程中扮演着重要的角色(Sardans et al., 2008).土壤中以游离态存在或与土壤有机质及矿质组分结合的酶称为土壤胞外酶.当温度、降水量和底物输入产生变化时, 土壤胞外酶活性将会发生相应改变(Seo et al., 2015; Chen et al., 2020; Hu et al., 2020), 表现为促进或抑制土壤养分、有机物分解, 进而影响陆地生态系统的物质循环过程(Karhu et al., 2014).因此, 气候变化可能对陆地生态系统土壤胞外酶活性产生复杂的影响, 而探讨各环境因子的变化对土壤胞外酶活性的影响是预测未来气候变化对森林生态系统过程影响的关键(Xiao et al., 2018). ...
Warming and increased precipitation enhance phenol oxidase activity in soil while warming induces drought stress in vegetation of an Arctic ecosystem 1 2015
... 土壤酶是来自土壤微生物、植物和动物的生物活性物质, 是专一生物化学反应的生物催化剂, 作为土壤生物地球化学过程的积极参与者, 在森林生态系统的物质循环和能量流动过程中扮演着重要的角色(Sardans et al., 2008).土壤中以游离态存在或与土壤有机质及矿质组分结合的酶称为土壤胞外酶.当温度、降水量和底物输入产生变化时, 土壤胞外酶活性将会发生相应改变(Seo et al., 2015; Chen et al., 2020; Hu et al., 2020), 表现为促进或抑制土壤养分、有机物分解, 进而影响陆地生态系统的物质循环过程(Karhu et al., 2014).因此, 气候变化可能对陆地生态系统土壤胞外酶活性产生复杂的影响, 而探讨各环境因子的变化对土壤胞外酶活性的影响是预测未来气候变化对森林生态系统过程影响的关键(Xiao et al., 2018). ...
Allocation of extracellular enzymatic activity in relation to litter composition, N deposition, and mass loss 2002
Ecoenzymatic stoichiometry and ecological theory 1 2012
Effects of warming and grazing on soil N availability, species composition, and ANPP in an alpine meadow 1 2012
... 工业革命以来, 全球平均气温呈上升趋势, 预计21世纪全球气温将增加0.3-4.8 ℃ (IPCC, 2013).温度是土壤酶活性的重要调控因子(Rustad et al., 2001; Kang & Lee, 2005; Luo et al., 2009), 直接影响了土壤酶活性, 进而影响土壤呼吸、土壤有机物分解等生化过程(Lu et al., 2013).McDaniel等(2013)对砍伐后的森林土壤进行模拟增温实验, 发现增温显著降低了土壤中β-葡萄糖苷酶、β-1,4-N-乙酰氨基葡萄糖苷酶和多酚氧化酶活性.但亦有研究表明, 短期增温和长期增温均未显著改变土壤胞外酶的活性(Wang et al., 2012).此外, 除土壤温度变化外, 土壤水分能直接影响土壤中的生化反应强度和土壤胞外酶活性.相关研究表明, 土壤水分与土壤胞外酶活性之间有显著的正相关关系(Ren et al., 2017; Li et al., 2018).Zhou等(2013)对温带草原的研究发现增加降雨会提高土壤碱性磷酸酶和脲酶的活性, 说明适量的增加土壤水分能够提升土壤胞外酶活性. ...
Beta-glucosidase activity in paddy soils of the Taihu Lake region, China 2 2006
Influence of interactions between litter decomposition and rhizosphere activity on soil respiration and on the temperature sensitivity in a subtropical montane forest in SW China 3 2014
... 利用哀牢山亚热带常绿阔叶林模拟增温实验平台, 林下随机设置20个90 cm × 90 cm × 50 cm (长×宽×高)的呼吸箱(每个呼吸箱覆盖面积为0.81 m2), 分别进行4种处理: 对照(CK)、切根(TR)、切根并增温(TR+W)、去凋(LR), 每种处理分别设置5个重复(Wu et al., 2014), 土壤酶相关实验均在呼吸箱内采样.各处理样方面积均为0.81 m2, 对照不做实验处理. ...
A meta-analysis of soil extracellular enzyme activities in response to global change 1 2018
... 土壤酶是来自土壤微生物、植物和动物的生物活性物质, 是专一生物化学反应的生物催化剂, 作为土壤生物地球化学过程的积极参与者, 在森林生态系统的物质循环和能量流动过程中扮演着重要的角色(Sardans et al., 2008).土壤中以游离态存在或与土壤有机质及矿质组分结合的酶称为土壤胞外酶.当温度、降水量和底物输入产生变化时, 土壤胞外酶活性将会发生相应改变(Seo et al., 2015; Chen et al., 2020; Hu et al., 2020), 表现为促进或抑制土壤养分、有机物分解, 进而影响陆地生态系统的物质循环过程(Karhu et al., 2014).因此, 气候变化可能对陆地生态系统土壤胞外酶活性产生复杂的影响, 而探讨各环境因子的变化对土壤胞外酶活性的影响是预测未来气候变化对森林生态系统过程影响的关键(Xiao et al., 2018). ...
森林土壤酶的研究进展 1 2004
... 由于土壤酶主要来自土壤微生物及植物根系分泌的具有催化作用的蛋白质, 因此根系分泌物是土壤酶的重要来源(杨万勤和王开运, 2004).同时在森林生态系统中, 凋落物是土壤有机质的重要来源, 凋落物输入量的改变会造成土壤微生物及其代谢方式的改变, 进而影响土壤胞外酶活性, 故碳源输入方式的改变可对土壤胞外酶活性产生影响.森林土壤中与碳、氮、磷循环相关的胞外酶活性对土壤有机质的输入较为敏感.Weintraub等(2013)发现, 减少凋落物输入后, 热带森林土壤磷限制性增强, 与磷循环相关的酶活性增强; 魏翠翠等(2018)研究发现凋落物加倍和凋落物去除均降低了亚热带森林酸性磷酸酶、β-葡萄糖苷酶和多酚氧化酶的活性; Ge等(2017)对亚热带马尾松林研究发现, 凋落物输入量与纤维素酶、脲酶和多酚氧化酶活性呈显著线性相关关系, 并证明土壤温度、凋落物含水量和凋落物氮含量是影响酶活性的主要因素.由此可见, 凋落物输入变化会引起微生物碳源和养分源变化, 进而影响土壤胞外酶活性(Allison & Vitousek, 2004).根系分泌物作为土壤酶的重要来源, 也发挥着重要作用, 研究表明根际土壤中有机碳含量较高, 根际碳的输入可以刺激根际微生物活动和胞外酶的活性, 因此根际土壤的酶活性普遍高于整体(Brzostek et al., 2013; Gianfreda, 2015).切根通常会导致土壤有机碳含量的下降(Crow et al., 2009; Fekete et al., 2014), 随之影响土壤酶活性.Spohn和Kuzyakov (2014)研究表明切根后纤维素酶和几丁质酶活性均升高, 但根系与胞外酶活性的相互影响机制还缺乏进一步研究. ...
森林土壤酶的研究进展 1 2004
... 由于土壤酶主要来自土壤微生物及植物根系分泌的具有催化作用的蛋白质, 因此根系分泌物是土壤酶的重要来源(杨万勤和王开运, 2004).同时在森林生态系统中, 凋落物是土壤有机质的重要来源, 凋落物输入量的改变会造成土壤微生物及其代谢方式的改变, 进而影响土壤胞外酶活性, 故碳源输入方式的改变可对土壤胞外酶活性产生影响.森林土壤中与碳、氮、磷循环相关的胞外酶活性对土壤有机质的输入较为敏感.Weintraub等(2013)发现, 减少凋落物输入后, 热带森林土壤磷限制性增强, 与磷循环相关的酶活性增强; 魏翠翠等(2018)研究发现凋落物加倍和凋落物去除均降低了亚热带森林酸性磷酸酶、β-葡萄糖苷酶和多酚氧化酶的活性; Ge等(2017)对亚热带马尾松林研究发现, 凋落物输入量与纤维素酶、脲酶和多酚氧化酶活性呈显著线性相关关系, 并证明土壤温度、凋落物含水量和凋落物氮含量是影响酶活性的主要因素.由此可见, 凋落物输入变化会引起微生物碳源和养分源变化, 进而影响土壤胞外酶活性(Allison & Vitousek, 2004).根系分泌物作为土壤酶的重要来源, 也发挥着重要作用, 研究表明根际土壤中有机碳含量较高, 根际碳的输入可以刺激根际微生物活动和胞外酶的活性, 因此根际土壤的酶活性普遍高于整体(Brzostek et al., 2013; Gianfreda, 2015).切根通常会导致土壤有机碳含量的下降(Crow et al., 2009; Fekete et al., 2014), 随之影响土壤酶活性.Spohn和Kuzyakov (2014)研究表明切根后纤维素酶和几丁质酶活性均升高, 但根系与胞外酶活性的相互影响机制还缺乏进一步研究. ...
土壤含水量测定方法研究进展 1 2005
... 土壤含水量(SWC)采用烘干称质量法测定(张学礼等, 2005).土壤溶解有机碳(DOC)、溶解有机氮(DON)和溶解总氮(DN)含量用TOC/TN分析仪(Liqui TOC II, Elementar Analyzer system GmbH, Frankfurt, Germany)测定, 方法为Pt-催化高温煅烧法(680 ℃)(Zhou et al., 2019).土壤硝态氮(NO3--N)和铵态氮(NH4+-N)含量使用连续流动分析仪(Skalar San++, Breda, the Netherlands)测定(Jones & Willett, 2006).土壤矿质氮(Mineral-N)是以离子态存在于土壤中的氮, 以土壤NH4+-N及NO3--N含量相加进行计算. ...
土壤含水量测定方法研究进展 1 2005
... 土壤含水量(SWC)采用烘干称质量法测定(张学礼等, 2005).土壤溶解有机碳(DOC)、溶解有机氮(DON)和溶解总氮(DN)含量用TOC/TN分析仪(Liqui TOC II, Elementar Analyzer system GmbH, Frankfurt, Germany)测定, 方法为Pt-催化高温煅烧法(680 ℃)(Zhou et al., 2019).土壤硝态氮(NO3--N)和铵态氮(NH4+-N)含量使用连续流动分析仪(Skalar San++, Breda, the Netherlands)测定(Jones & Willett, 2006).土壤矿质氮(Mineral-N)是以离子态存在于土壤中的氮, 以土壤NH4+-N及NO3--N含量相加进行计算. ...
Soil respiration after six years of continuous drought stress in the tropical rainforest in Southwest China 1 2019
... 土壤含水量(SWC)采用烘干称质量法测定(张学礼等, 2005).土壤溶解有机碳(DOC)、溶解有机氮(DON)和溶解总氮(DN)含量用TOC/TN分析仪(Liqui TOC II, Elementar Analyzer system GmbH, Frankfurt, Germany)测定, 方法为Pt-催化高温煅烧法(680 ℃)(Zhou et al., 2019).土壤硝态氮(NO3--N)和铵态氮(NH4+-N)含量使用连续流动分析仪(Skalar San++, Breda, the Netherlands)测定(Jones & Willett, 2006).土壤矿质氮(Mineral-N)是以离子态存在于土壤中的氮, 以土壤NH4+-N及NO3--N含量相加进行计算. ...
Warming and increased precipitation have differential effects on soil extracellular enzyme activities in a temperate grassland 1 2013
... 工业革命以来, 全球平均气温呈上升趋势, 预计21世纪全球气温将增加0.3-4.8 ℃ (IPCC, 2013).温度是土壤酶活性的重要调控因子(Rustad et al., 2001; Kang & Lee, 2005; Luo et al., 2009), 直接影响了土壤酶活性, 进而影响土壤呼吸、土壤有机物分解等生化过程(Lu et al., 2013).McDaniel等(2013)对砍伐后的森林土壤进行模拟增温实验, 发现增温显著降低了土壤中β-葡萄糖苷酶、β-1,4-N-乙酰氨基葡萄糖苷酶和多酚氧化酶活性.但亦有研究表明, 短期增温和长期增温均未显著改变土壤胞外酶的活性(Wang et al., 2012).此外, 除土壤温度变化外, 土壤水分能直接影响土壤中的生化反应强度和土壤胞外酶活性.相关研究表明, 土壤水分与土壤胞外酶活性之间有显著的正相关关系(Ren et al., 2017; Li et al., 2018).Zhou等(2013)对温带草原的研究发现增加降雨会提高土壤碱性磷酸酶和脲酶的活性, 说明适量的增加土壤水分能够提升土壤胞外酶活性. ...
云南中山湿性常绿阔叶林起源的探讨 2 2016
... 研究样地位于景东县太忠镇徐家坝的中国科学院哀牢山亚热带森林生态系统研究站(24.53° N, 101.02° E, 海拔2 480 m).研究区域属亚热带山地常绿阔叶林潮湿气候带, 干湿季分明, 年平均气温11.3 ℃, 年降水量1 881.5 mm, 降水主要集中在5-10月.区域内土壤为山地黄棕壤, 质地为砂壤土, 植物种类丰富、群落类型多样、垂直带谱完整、过渡性特征明显(朱华, 2016), 表层土壤理化性质见表2 (鲁志云, 2016). ...
... 由于微生物生物量C:N:P具有一定的稳定性(Cleveland & Liptzin, 2007), 因此土壤酶生态化学计量比也表现为相对稳定(Sinsabaugh et al., 2008).该研究中酶C:P和N:P远低于全球主要陆地生态系统土壤酶C:P和N:P的平均值0.62和0.44, 而酶C:N则略高于平均值1.14 (Sinsabaugh et al., 2009), 说明AP活性远远高于BG和NAG活性, 基于全球尺度上BG、NAG和AP对数转化后比值约为1:1:1 (Sinsabaugh et al., 2008).本研究中两个土层比值大体相同, 都表现为lg (AP)比值偏大, 说明该地区受磷限制.Allison等(2010)研究表明当微生物的生长受到某元素限制时, 会增加与该元素相关胞外酶的活性, 因此该地土壤AP活性较高, 与Waring等(2014)的研究结果一致.另有研究表明丰富的降水也会增加表层土壤中磷的淋溶, 这会进一步降低磷的有效性(Waring et al., 2014), 本研究样地年降水量可达1 800 mm以上, 且集中在5-10月, 属于典型的湿性常绿阔叶林(朱华, 2016), 土壤磷淋溶强烈, 本地土壤AP活性偏高和降水量大联系紧密. ...
云南中山湿性常绿阔叶林起源的探讨 2 2016
... 研究样地位于景东县太忠镇徐家坝的中国科学院哀牢山亚热带森林生态系统研究站(24.53° N, 101.02° E, 海拔2 480 m).研究区域属亚热带山地常绿阔叶林潮湿气候带, 干湿季分明, 年平均气温11.3 ℃, 年降水量1 881.5 mm, 降水主要集中在5-10月.区域内土壤为山地黄棕壤, 质地为砂壤土, 植物种类丰富、群落类型多样、垂直带谱完整、过渡性特征明显(朱华, 2016), 表层土壤理化性质见表2 (鲁志云, 2016). ...
... 由于微生物生物量C:N:P具有一定的稳定性(Cleveland & Liptzin, 2007), 因此土壤酶生态化学计量比也表现为相对稳定(Sinsabaugh et al., 2008).该研究中酶C:P和N:P远低于全球主要陆地生态系统土壤酶C:P和N:P的平均值0.62和0.44, 而酶C:N则略高于平均值1.14 (Sinsabaugh et al., 2009), 说明AP活性远远高于BG和NAG活性, 基于全球尺度上BG、NAG和AP对数转化后比值约为1:1:1 (Sinsabaugh et al., 2008).本研究中两个土层比值大体相同, 都表现为lg (AP)比值偏大, 说明该地区受磷限制.Allison等(2010)研究表明当微生物的生长受到某元素限制时, 会增加与该元素相关胞外酶的活性, 因此该地土壤AP活性较高, 与Waring等(2014)的研究结果一致.另有研究表明丰富的降水也会增加表层土壤中磷的淋溶, 这会进一步降低磷的有效性(Waring et al., 2014), 本研究样地年降水量可达1 800 mm以上, 且集中在5-10月, 属于典型的湿性常绿阔叶林(朱华, 2016), 土壤磷淋溶强烈, 本地土壤AP活性偏高和降水量大联系紧密. ...
 ,1,2,5,*, 况露辉3,5, 刘占锋3,5, 宋清海1,2,5, 刘运通1,2,5, 张一平1,2,5, 鲁志云1,4, 沙丽清
,1,2,5,*, 况露辉3,5, 刘占锋3,5, 宋清海1,2,5, 刘运通1,2,5, 张一平1,2,5, 鲁志云1,4, 沙丽清
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT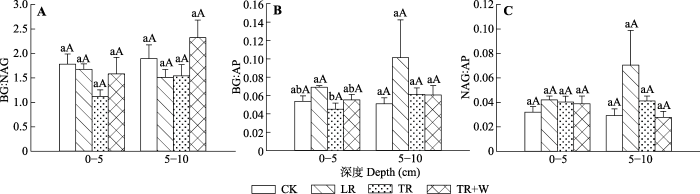 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT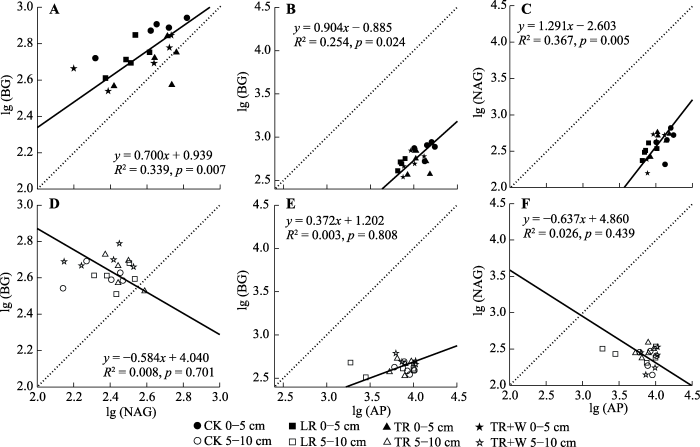 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT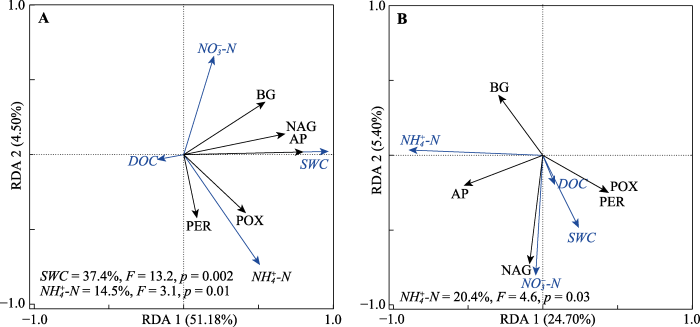 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}