吴建波, 王小丹
, * 中国科学院申扎高寒草原与湿地生态系统观测试验站, 山地表生过程与生态调控重点实验室, 中国科学院、水利部成都山地灾害与环境研究所, 成都, 610041 Analyzing leaf anatomical structure of dominant species Stipa purpurea adapting to alpine and drought environment at Qingzang Plateau Jian-Bo WU, Xiao-Dan WANG
, * Xainzha Alpine Steppe and Wetland Ecosystem Observation and Experiment Station, Key Laboratory of Mountain Environment Evolution and Regulation, Institute of Mountain Hazard and Environment, Chinese Academy of Sciences, Chengdu 610041, China 通讯作者: * (
wxd@imde.ac.cn )
编委: 储诚进
责任编辑: 李敏
收稿日期: 2020-09-25
接受日期: 2021-01-14
网络出版日期: 2021-03-20
基金资助: 第二次青藏高原综合科学考察研究 (SQ2019QZKK2004 )国家重点研发计划 (2016YFC0502002 )国家自然科学基金 (41401072 )
Corresponding authors: * (
wxd@imde.ac.cn )
Received: 2020-09-25
Accepted: 2021-01-14
Online: 2021-03-20
Fund supported: Second Comprehensive Scientific Investigation of the Qinghai-Tibet Plateau (SQ2019QZKK2004 )National Key R&D Program of China (2016YFC0502002 )National Natural Science Foundation of China (41401072 )
摘要 随着气候变暖, 高寒草原分布面积逐步增加, 高寒草原植物如何适应高寒干旱环境的研究还比较缺乏。该研究通过分析高寒草原优势种紫花针茅(
Stipa purpurea )不同地理种群叶片解剖结构特征差异及其与气候因子的相关性, 阐明紫花针茅叶片适应高寒环境的策略, 为理解高寒植物对高寒干旱胁迫环境的适应机制提供科学依据。在青藏高原不同地理位置选择8个紫花针茅种群, 选择成熟健康叶片用卡诺氏固定液固定, 将固定好的叶片带回实验室进行石蜡切片和染色, 用显微镜观察叶片结构, 并用数码相机拍摄, 然后用软件Image-pro plus 6对叶片结构进行测量。结果显示: 紫花针茅叶片普遍具有较厚的角质层, 可减少水分散失和抵御较强的辐射; 不同地理种群紫花针茅叶片解剖结构在厚壁细胞厚度、叶片厚度、导管直径、主脉导管腔面积/主脉维管束面积和维管束面积/叶横切面积等特征上存在较大差异, 以适应不同区域的生境。Pearson相关性和聚类分析结果表明紫花针茅叶片解剖结构与气候因子密切相关; 主成分和冗余分析结果表明在干旱区域紫花针茅叶片解剖结构主要受到蒸发量的影响, 而在相对湿润区域紫花针茅叶片解剖结构主要受生长季降水量、湿润系数和年降水量/年蒸发量影响。综上所述, 紫花针茅通过增加厚壁细胞减少水分散失, 同时增加导管直径、主脉导管面积/主脉维管束面积和维管束面积/叶横切面积等输水组织面积适应高寒干旱气候。
关键词: 高寒草原 ;
紫花针茅 ;
叶片解剖结构 ;
气候因子 ;
适应机制 Abstract Aims The anatomical structure of plant leaves could be a direct reflection of the plant’s mechanism of response and adaptation to climate change. The distribution of alpine steppe has increased due to climatic warming over the Qingzang Plateau.
Stipa purpurea is the dominant species of alpine steppe. However, few studies have been conducted on the adaptation mechanisms of alpine steppe plants. In this study, we analyzed the characteristics of leaf anatomical structure among different populations of
S. purpurea and the relationship between these characteristics and climatic factors over the Qingzang Plateau. The ultimate aim was to identify the mechanism by which
S. purpurea acclimatizes to alpine environments.
Methods Leaves of
S. purpurea were collected from eight sites on the Qingzang Plateau and fixed using FAA fixative solution. Then, the leaves were paraffin wax sectioned and double-stained. Samples were then observed with a microscope and photographed with a digital camera. The area and thickness of leaf anatomical structure were measured with the soft (Image-pro plus 6.0).
Important findings Leaves of
S. purpurea generally had thick cuticles, which were able to reduce water loss and radiation exposure. From the results of one-way analysis of variance, there were significant differences among the eight populations in collenchymatous cell thickness, vessel diameter, bundle catheter cavity area/bundle of the main vascular area, and vascular area/leaf cross-sectional area, which were beneficial characteristics for
S. purpurea in adapting to the local environment. The characteristics of leaf anatomical structure were significantly correlated with environmental factors via Pearson’s analysis and cluster analysis. The results from principal component analysis and redundancy analysis showed that the anatomical structures in arid regions were mainly affected by annual evaporation, and those in semi-humid regions were mainly affected by the average precipitation humidity index and annual precipitation/annual evaporation during the growing season. In conclusion, to adapt to the alpine and arid environment,
S. purpurea has reduced water loss by simultaneously increasing its collenchymatous cell thickness and water-conducting tissue area (vessel diameter, bundle catheter cavity area/bundle of main vascular area, and vascular bundle area/leaf cross-sectional area).
Keywords: alpine steppe ;
Stipa purpurea ;
leaf anatomical structure ;
climate factor ;
adapted mechanism PDF (1287KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 吴建波, 王小丹. 高寒草原优势种紫花针茅叶片解剖结构对青藏高原高寒干旱环境适应性分析. 植物生态学报, 2021, 45(3): 265-273. DOI:
10.17521/cjpe.2020.0322 WU Jian-Bo, WANG Xiao-Dan.
Analyzing leaf anatomical structure of dominant species Stipa purpurea adapting to alpine and drought environment at Qingzang Plateau .
Chinese Journal of Plant Ecology , 2021, 45(3): 265-273. DOI:
10.17521/cjpe.2020.0322 叶片解剖结构特征是植物长期适应环境以及自身进化形成的, 体现了植物对环境的适应策略(Hetherington & Woodward,
2003 ), 而且叶片解剖结构在不同区域的变异特征可以直接反映植物对不同生长环境的适应策略(Wright
et al .,
2001 ; 钟悦鸣等,
2017 )。例如中间锦鸡儿(
Caragana davazamcii )随着平均年降水量的减少, 角质层厚度有所增加, 栅栏组织分化逐渐加强, 叶脉的机械组织分化逐渐加强(杨明博,
2007 ); 长梗扁桃(
Amygdalus pedunculata )生长在干旱地区的叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的叶片栅栏组织排列疏松, 叶厚度较薄(郭改改等,
2013 ); 青藏高原的菊科植物叶片厚度、上下表皮厚度、上下角质层厚度、栅栏细胞系数均随海拔升高而增加(孙会婷等,
2016 ); 蒙古莸(
Caryopteris mongholica )叶片解剖结构随着年平均气温升高, 叶厚度、栅栏组织厚度呈增大趋势(纪若璇等,
2020 )。因此, 叶片解剖结构可以体现植物对环境因子的适应性进化, 开展不同区域植物叶片解剖结构变异特征的研究, 对阐明植物适应气候变化策略具有重要意义。
紫花针茅(
Stipa purpurea )作为高寒草原的优势种, 对维持群落稳定性具有重要作用(Wu & Wang,
2019 )。气候变暖导致青藏高原的西部地区变暖变干, 东部地区变暖变湿(谭春萍等,
2010 ; 陈德亮等,
2015 ), 进而改变青藏高原草地生态系统的空间格局, 高山嵩草(
Kobresia pygmaea )群落向紫花针茅群落退化(王谋等,
2005 ), 紫花针茅在青藏高原的分布范围随着温度升高将继续扩大(陈德亮等,
2015 ; Ma & Sun,
2018 )。紫花针茅分布范围增加说明其具有对干旱生境较强的适应能力, 然而其如何适应高寒干旱气候变化的研究还相对薄弱(胡建莹等,
2008 ; 胡梦瑶等,
2012 ; 李雄等,
2015 ; 王常顺和汪诗平,
2015 )。对不同区域紫花针茅叶片解剖结构的分析, 可以直接反映紫花针茅对青藏高原气候变化的适应策略。因此, 本研究通过对紫花针茅在青藏高原分布区域进行采样, 分析其叶片解剖结构特征变异规律及其如何对环境因子响应, 以期为阐明高寒草原植物对气候变化适应机制提供科学依据。
1 材料和方法 1.1 研究地点 选择羌塘高寒草原区域自东至西按降水梯度分别选择班戈县、尼玛县、改则县、革吉县和噶尔县, 以及可可西里高寒草原区域五道梁地区, 同时选择了高寒草甸和高寒草原交错带区域当雄县和安多县, 各样地植被类型和气候特征见
表1 。
Table 1 表1 表1 青藏高原高寒草原各样地环境特征
Table 1
Environmental characters of sample sites at alpine steppe of Qingzang Plateau 样点 地理位置 海拔 草原类型 优势种 ≥0 ℃积温 年降水量 MAP, mm) 年平均气温 MAT, ℃) 年蒸发量 安多 32.24° N 4 746 高寒草甸 高山嵩草 Kobresia pygmaea 869 439 -2.8 1 725 当雄 32.25° N 4 228 高寒草原化草甸 高山嵩草 K. pygmaea Stipa purpurea 1 536 456 1.3 1 710 五道梁 32.26° N 4 576 高寒草原 紫花针茅 S. purpurea 683 259 -5.1 1 421 班戈 31.36° N 4 682 高寒草原 紫花针茅 S. purpurea 1 048 321 -0.8 1 994 尼玛 31.78° N 4 554 高寒草原 紫花针茅 S. purpurea 1 453 200 -0.4 2 200 改则 31.20° N 4 477 高寒草原 紫花针茅 S. purpurea 1 453 170 0.1 2 255 革吉 31.20° N 4 502 高寒草原 紫花针茅 S. purpurea 1 566 120 0.4 2 420 噶尔 32.46° N 4 300 高寒荒漠 紫花针茅 S. purpurea 1 573 72 0.7 2 696
新窗口打开 |
下载CSV 1.2 样品采集和处理 2018年8月在各样地内取样(
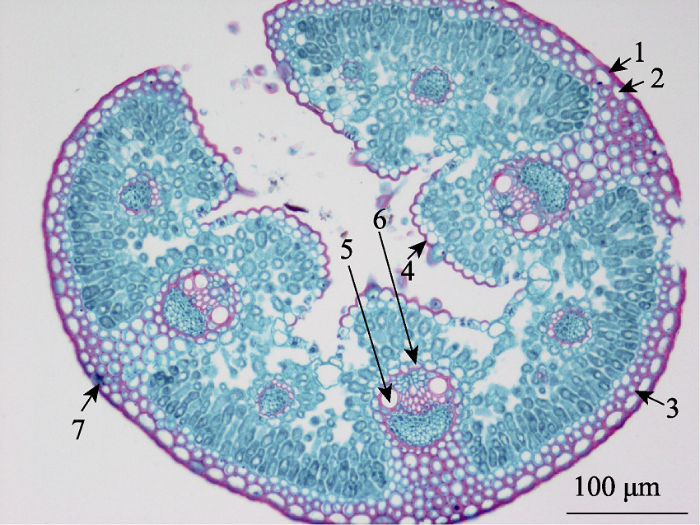
表1 )。在每个样地选取典型样地, 然后在样地内随机选取5株紫花针茅, 每株随机选取2片已完全伸展的新鲜叶片, 上面无虫咬和病菌感染, 将紫花针茅叶片剪下后, 立即用卡诺氏固定液(FAA)固定, 并用注射器将叶片空气排空。将固定好的叶片带回实验室进行石蜡切片和番红-固绿双重染色, 在光学显微镜下对叶片组织结构特征进行观察和拍照, 然后用Image-pro plus 6软件对叶片组织结构进行测量, 获得叶片组织结构特征数据。根据叶片组织结构特征分为叶片保护组织(外表皮、角质层、内表皮)、维管组织(导管直径、主脉导管腔面积、主脉维管束面积和维管束面积)和其他特征(厚角细胞、叶片厚度、气孔宽度和叶片横切面积)(
图1 )。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1青藏高原高寒草原紫花针茅叶片解剖结构。1, 角质层; 2, 外表皮; 3, 厚角组织; 4, 内表皮; 5, 导管; 6, 主维管束; 7, 气孔。 Fig. 1Folia characters of Stipa purpurea in paraffin section at alpine steppe of Qingzang Plateau. 1, cuticle; 2, outer epidermis; 3, collenchymatous cell; 4, inner epidermis; 5, vessel; 6, mainly vascular bundle; 7, stoma. 1.3 气象数据 气象数据来自对全国气象台站观测数据插值。气候指标包括: 年降水量、年平均气温、生长季平均气温、生长季降水量、7月平均气温、7月降水量、≥0 ℃积温、年蒸发量。湿润指数采用谢良尼诺夫公式根据中国实际情况修改进行计算(孟猛等,
2004 )。干旱半干旱区与半湿润区按照胡梦瑶等(
2012 )标准划分。
1.4 统计分析 叶片组织结构特征差异采用单因素方差分析法分析。叶片特征与气候因子相关性采用Pearson相关性分析(两侧检验)。采用聚类分析不同地理种群紫花针茅叶片组织结构相似性, 并应用主成分分析影响紫花针茅分布的主要组织结构特征。采用冗余分析(RDA)进一步分析环境变量、叶片组织结构特征和紫花针茅分布的关系。以上分析采用SPSS 21和Canoco 5进行。
2 结果 2.1 不同地理种群紫花针茅叶片解剖结构特征 在不同地理气候条件下, 紫花针茅叶片解剖结构特征表现出对生境的适应性(
表2 ;
图2 )。在保护组织方面, 安多紫花针茅种群叶片外表皮厚度和内表皮厚度显著高于其他地理种群, 而叶片外表皮厚度和内表皮厚度在其他地理种群间不存在显著差异; 叶片角质层厚度在各个种群都不存在显著差异。在维管组织方面, 革吉、噶尔和安多种群叶片导管直径显著高于其他地理种群, 五道梁种群叶片导管直径最小; 改则、革吉和噶尔种群叶片主脉导管腔面积与主脉维管束面积比显著高于其他地理种群; 五道梁种群叶片主脉导管腔面积与主脉维管束面积比最小; 安多和当雄种群叶片的维管束面积与叶横切面积比显著小于其他地理种群。安多种群叶片不存在厚角细胞, 但是在五道梁和当雄种群叶片中厚角细胞厚度有所增加, 在班戈、尼玛、改则、革吉和噶尔种群叶片中厚角细胞厚度最大。安多种群叶片厚度最大, 噶尔和革吉种群叶片厚度次之, 五道梁、当雄、班戈和改则种群叶片厚度最小; 气孔宽度变化趋势与叶片厚度一致。
Table 2 表2 表2 青藏高原高寒草原不同地理种群紫花针茅叶片组织结构特征(平均值±标准误)
Table 2
Leaf characters of Stipa purpurea at different population at alpine steppe of Qingzang Plateau (mean ± SE) 高寒草甸 高寒草原 安多 当雄 五道梁 班戈 尼玛 改则 革吉 噶尔 角质层厚度 4.75 ± 0.95 5.22 ± 0.46 5.85 ± 0.38 5.51 ± 0.74 4.36 ± 0.15 5.18 ± 0.40 4.99 ± 0.81 5.25 ± 0.62 外表皮厚度 12.16 ± 2.31 a 8.93 ± 0.40 b 9.51 ± 0.34 b 7.63 ± 0.51 b 9.76 ± 1.53 b 7.36 ± 0.80 b 8.89 ± 0.59 b 8.08 ± 0.71 b 内表皮厚度 18.82 ± 3.07 a 7.80 ± 0.45 b 6.95 ± 0.33 b 5.46 ± 0.33 b 6.35 ± 0.45 b 6.13 ± 0.58 b 7.83 ± 0.77 b 8.49 ± 1.00 b 厚角细胞厚度 0 5.16 ± 0.82 b 4.92 ± 0.22 b 9.18 ± 2.12 a 13.32 ± 1.35 a 11.67 ± 1.72 a 10.66 ± 0.76 a 13.97 ± 0.78 a 气孔宽度 28.33 ± 8.82 a 12.42 ± 1.73 bc 10.06 ± 0.60 c 9.97 ± 1.22 c 12.35 ± 0.50 bc 13.55 ± 1.14 bc 15.37 ± 1.45 b 15.62 ± 1.05 b 叶片厚度 269.89 ± 19.67 a 179.12 ± 16.49 c 170.74 ± 2.60 c 173.46 ± 2.69 c 196.55 ± 11.10bc 190.09 ± 19.18 bc 246.28 ± 31.81 b 212.33 ± 2.64 b 导管直径 16.04 ± 2.09 a 12.13 ± 1.13 b 9.73 ± 0.42 c 12.83 ± 1.36 b 11.82 ± 1.13 b 12.73 ± 1.34 b 21.15 ± 2.84 a 18.36 ± 2.23 a 主脉导管腔面积/主脉维管束面积 0.11 ± 0.00 b 0.11 ± 0.01 b 0.07 ± 0.00 c 0.11 ± 0.02 b 0.11 ± 0.00 b 0.15 ± 0.02 a 0.16 ± 0.03 a 0.16 ± 0.02 a 维管束面积/叶横切面积Vascular area/leaf cross sectional area 0.04 ± 0.01 b 0.05 ± 0.01 b 0.07 ± 0.01 a 0.08 ± 0.01 a 0.07 ± 0.01 a 0.08 ± 0.01 a 0.08 ± 0.01 a 0.08 ± 0.01 a
同行不同小写字母表示差异显著(p < 0.05)。 Different lowercase letters of the same row indicate significance at the 0.05 level. 新窗口打开 |
下载CSV 图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2青藏高原高寒草原不同地理种群紫花针茅叶片解剖结构。 Fig. 2Leaf characters of Stipa purpurea in different populations at alpine steppe of Qingzang Plateau. 2.2 不同地理种群紫花针茅叶片解剖结构与气候因子相关性分析 紫花针茅叶片解剖结构特征受到气候因子显著的影响。Pearson相关性分析结果表明导管直径与生长季降水量和7月降水量呈显著的负相关关系; 厚角细胞厚度与年降水量和年降水量/年蒸发量呈显著负相关关系, 与年蒸发量呈显著的正相关关系; 主脉导管腔面积/主脉维管束面积与年蒸发量呈显著的正相关关系, 而与年降水量、年降水量/年蒸发量、湿润指数、生长季降水量和7月降水量呈显著的负相关关系; 维管束面积/叶横切面积与年平均气温、年蒸发量、生长季平均气温和7月平均气温呈显著的正相关关系(
表3 )。
Table 3 表3 表3 青藏高原高寒草原紫花针茅叶结构特征与气候因子的相关性
Table 3
Correlations between climatic factors and leaf structures of Stipa purpurea at alpine steppe of Qingzang Plateau 外表皮厚度 内表皮厚度 角质层厚度 气孔宽度 叶片厚度 导管直径 厚角细胞厚度 主脉导管腔面积/ 维管束面积/ 年降水量 0.58 0.42 0.30 0.21 0.01 -0.55 -0.78* -0.87** -0.61 年平均气温 -0.34 -0.39 0.24 -0.31 -0.12 0.15 0.50 0.21 0.77* ≥0 ℃积温 0.11 0.29 -0.56 0.50 0.56 0.64 0.34 0.67 0.33 年蒸发量 -0.43 -0.26 -0.21 -0.05 0.16 0.61 0.76* 0.79* 0.76* 年降水量/年蒸发量 0.59 0.44 0.32 0.21 0.00 -0.55 -0.83* -0.87** -0.71* 湿润指数 0.29 0.07 0.56 -0.18 -0.31 -0.69 -0.65 -0.91** -0.50 生长季平均气温 -0.50 -0.39 0.19 -0.24 -0.01 0.51 0.57 0.64 0.81* 生长季降水量 0.52 0.28 0.05 0.10 -0.16 -0.75* -0.51 -0.90** -0.53 7月平均气温 -0.42 -0.27 0.07 -0.10 0.12 0.59 0.55 0.70 0.79* 7月降水量 0.50 0.28 0.05 0.11 -0.15 -0.73* -0.48 -0.87** -0.47
*, 在0.05水平(双侧)上显著相关; **, 在0.01水平(双侧)上显著相关。 *, difference is significant at the 0.05 level (two-tailed); **, difference is significant at the 0.01 level (two-tailed). 新窗口打开 |
下载CSV 2.3 不同地理种群紫花针茅叶片解剖结构主成分分析和聚类分析 主成分分析结果表明前2个主成分累计方差贡献率达到了83.00% (
表3 ), 其中保护组织外表皮、角质层和厚角细胞厚度在第一主成分得分最高, 输水组织导管直径和主脉导管腔面积/主脉维管束面积在第二主成分得分最高(
表4 )。聚类分析结果表明8个地理种群分为2个分支, 其中安多种群单独为1个分支, 其他7个种群聚为一个分支, 然后又分为3个分支, 班戈、尼玛和改则聚为一个分支, 五道梁和当雄种群聚为一分支, 革吉和噶尔聚为一个分支(
图3 )。
Table 4 表4 表4 主成分分析特征根与方差贡献率
Table 4
Eigenvalues and extraction squared loading of principal component analysis 成分 特征根植 变异系数 累计值 1 4.80 53.32 53.32 2 2.67 29.68 83.00 3 0.94 10.42 93.42 4 0.28 3.08 96.50 5 0.22 2.44 98.94 6 0.08 0.86 99.80 7 0.02 0.20 100.00 8 0.00 0.00 100.00
新窗口打开 |
下载CSV Table 5 表5 表5 青藏高原高寒草原紫花针茅叶片解剖结构特征主成分分析
Table 5
Principal component analysis of leaf structures of Stipa purpurea at alpine steppe of Qingzang Plateau 第一主成分 第二主成分 外表皮厚度 0.47 -0.05 角质层厚度 Cuticle thickness 0.45 0.19 厚角细胞厚度 0.41 0.32 气孔宽度 Stoma width 0.08 -0.39 导管直径 Vessel diameter 0.04 0.55 主脉导管腔面积/主脉维管束面积 -0.16 0.55 内表皮厚度 -0.19 -0.21 叶片厚度 Leaf thickness -0.39 0.14 维管束面积/叶横切面积 -0.44 0.19
新窗口打开 |
下载CSV 图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3青藏高原高寒草原紫花针茅叶片解剖结构特征的聚类分析。 Fig. 3Hierarchical clustering of leaf structures of Stipa purpurea at alpine steppe of Qingzang Plateau. 2.4 不同地理种群紫花针茅叶片解剖结构与环境因子冗余分析 RDA结果表明: 第一和第二排序轴累计解释率分别为59.52%和34.19% (
图4 ), RDA前2个排序轴保留了土壤酶活性数据总方差的93.71%。冗余分析结果表明气候因子年蒸发量、生长季降水量、湿润指数和年降水量/年蒸发量对紫花针茅叶片解剖结构影响较大(
图4 )。其中年蒸发量对噶尔、革吉、改则、尼玛和班戈紫花针茅叶片的厚角细胞厚度、导管直径和主脉导管腔面积/主脉维管束面积影响较大(
图4 ); 生长季降水量、湿润指数、年降水量和年降水量/年蒸发量对安多、五道梁和当雄紫花针茅叶片的角质层厚度和外表皮厚度影响较大(
图4 )。
图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4青藏高原高寒草原气候因子与不同地理种群紫花针茅叶片解剖特征冗余分析(RDA)。 Fig. 4Redundancy analysis (RAD) between climatic factors and leaf structures of Stipa purpurea at alpine steppe of Qingzang Plateau. 3 讨论 3.1 不同地理种群紫花针茅叶片解剖结构特征的共性和变异 植物与其生长的环境是统一的整体, 环境对植物的长期作用影响植物叶片解剖结构。在本研究中, 不同地理种群紫花针茅叶片解剖结构特征既存在相似性, 也存在较大变异性。角质层厚度在不同地理种群紫花针茅叶片结构特征中不存在显著差异, 这是因为角质层不仅可以反射强光, 还可以减少叶片水分蒸散, 有利于植物适应高原环境(李正理,
1981 ; Holmes & Keiller,
2002 ; 何涛等,
2007 ), 体现了不同地理种群紫花针茅叶片结构对高海拔生境的趋同适应进化。但是, 生境的不同导致紫花针茅叶片结构解剖特征存在较大差异, 例如紫花针茅叶片厚角细胞厚度随着干旱程度的增加而增加, 厚角细胞从安多紫花针茅叶片解剖结构中不存在到噶尔紫花针茅叶片中厚度逐渐增加, 而增加厚壁细胞厚度可减少水分蒸散损失(康萨如拉等,
2013 )。紫花针茅叶片解剖结构输水组织在不同区域也存在较大差异, 导管直径、主脉导管腔面积/主脉维管束面积和维管束面积/叶横切面积随着降水量的减少而增加, 这利于增加水分运输能力, 适应干旱环境(Cox & Moorc,
2010 ; 康萨如拉等,
2013 ; 寇萌等,
2019 )。因此, 不同地理种群紫花针茅叶片既有相似的解剖结构特征适应高海拔生境, 也有不同的叶片解剖结构适应区域生境。
3.2 紫花针茅叶片解剖结构与气候因素的关系 高寒草原优势种紫花针茅在青藏高原分布范围较广, 其叶片解剖结构与其生境相适应。Pearson相关性分析和聚类分析结果表明紫花针茅叶片解剖结构与气候因子密切相关(康萨如拉等,
2013 ), 例如五道梁和当雄2个地理种群距离较远, 但其叶片解剖结构比较相似。进一步的主成分和冗余分析结果表明, 与水分相关的气候因子对不同区域紫花针茅叶片解剖结构影响较大(胡选萍等,
2016 ), 即在噶尔、革吉和改则等干旱区域, 年蒸发量是影响厚角细胞厚度、导管直径和主脉导管腔面积/主脉维管束面积的主要因子, 而在相对湿润区域年降水量和生长季降水量等气候因子是影响保护组织(外表皮厚度和角质层厚度)的主要因子。紫花针茅叶片解剖结构未受到低温影响, 这可能是由于紫花针茅分布在高海拔区域, 相较于其他区域植物, 其已经适应了低温环境(Ma
et al .,
2012 ; 李全发等,
2013 )。
3.3 紫花针茅叶片解剖结构对高寒干旱环境适应机制 紫花针茅作为高寒草原的优势种, 其叶片解剖结构对高寒干旱环境具有较好的适应性。紫花针茅叶片外表皮普遍具有较厚角质层, 而在特别干旱区域(噶尔), 紫花针茅内表皮上也有角质层形成(
图2 H), 这有利于减少水分散失, 适应干旱环境(邓彦斌等,
1998 ; 蔡永立和宋永昌,
2001 )。同时紫花针茅叶片通过增加厚角细胞厚度, 减少水分散失(
图2 ), 这与内蒙古短花针茅适应干旱环境的叶片解剖结构 (康萨如拉等,
2013 )相一致。为了适应干旱环境, 紫花针茅在改变保护组织的同时, 增加输水组织面积比例(导管直径、主脉导管面积/主脉维管束面积和维管束面积/叶横切面积), 以增强水分运输适应干旱环境(康萨如拉等,
2013 ; Buckley
et al .,
2015 ; 寇萌等,
2019 )。
4 结论 不同地理种群紫花针茅叶片解剖结构既有相似之处, 但也存在较大的变异。与水分相关的气候因子是引起叶片解剖结构变异的主要因素。紫花针茅叶片解剖结构对高寒干旱环境表现出较强的适应性, 紫花针茅通过增加厚角细胞厚度, 减少水分散失; 同时增加导管直径和输水组织在叶片结构中的面积比例, 提高输水能力以适应高原干旱环境。
[1] Buckley TN John GP Scoffoni C Sack L 2015 ). How does leaf anatomy influence water transport outside the xylemPlant Physiology 1616 -1635 . DOI:
10.1104/pp.15.00731 PMID:
26084922 [本文引用: 1] Leaves are arguably the most complex and important physicobiological systems in the ecosphere. Yet, water transport outside the leaf xylem remains poorly understood, despite its impacts on stomatal function and photosynthesis. We applied anatomical measurements from 14 diverse species to a novel model of water flow in an areole (the smallest region bounded by minor veins) to predict the impact of anatomical variation across species on outside-xylem hydraulic conductance (Kox). Several predictions verified previous correlational studies: (1) vein length per unit area is the strongest anatomical determinant of Kox, due to effects on hydraulic pathlength and bundle sheath (BS) surface area; (2) palisade mesophyll remains well hydrated in hypostomatous species, which may benefit photosynthesis, (3) BS extensions enhance Kox; and (4) the upper and lower epidermis are hydraulically sequestered from one another despite their proximity. Our findings also provided novel insights: (5) the BS contributes a minority of outside-xylem resistance; (6) vapor transport contributes up to two-thirds of Kox; (7) Kox is strongly enhanced by the proximity of veins to lower epidermis; and (8) Kox is strongly influenced by spongy mesophyll anatomy, decreasing with protoplast size and increasing with airspace fraction and cell wall thickness. Correlations between anatomy and Kox across species sometimes diverged from predicted causal effects, demonstrating the need for integrative models to resolve causation. For example, (9) Kox was enhanced far more in heterobaric species than predicted by their having BS extensions. Our approach provides detailed insights into the role of anatomical variation in leaf function. ? 2015 American Society of Plant Biologists. All Rights Reserved.
[2] Cai YL Song YC 2001 ). Adaptive ecology of lianas in Tiantong evergreen broad-leaved forest, Zhejiang, China. I. Leaf anatomical charactersActa Phytoecologica Sinica 90 -98 . [本文引用: 1] [蔡永立 , 宋永昌 (2001 ). 浙江天童常绿阔叶林藤本植物的适应生态学: I. 叶片解剖特征的比较植物生态学报 , 25, 90 -98 .] [本文引用: 1] [3] Chen DL Xu BQ Yao TD Guo ZT Cui P Chen FH Zhang RH Zhang XZ Zhang YL Fan J Hou ZQ Zhang TH 2015 ). Assessment of past, present and future environmental changes on the Tibetan PlateauChinese Science Bulletin 3025 -3035 . [本文引用: 2] [陈德亮 , 徐柏青 , 姚檀栋 , 郭正堂 , 崔鹏 , 陈发虎 , 张人禾 , 张宪洲 , 张镱锂 , 樊杰 , 侯增谦 , 张天华 (2015 ). 青藏高原环境变化科学评估: 过去、现在与未来科学通报 , 60, 3025 -3035 .] [本文引用: 2] [4] Cox CB Moorc PD 2010 ). Biogeography: an Ecological and Evolutionary Approach [本文引用: 1] [5] Deng YB Jiang YC Liu J 1998 ). The xeromophic and saline mophic structure of leaves and assimilating branches in ten Chenopodiacea species in XinjiangActa Phytoecologica Sinica 164 -170 . [本文引用: 1] [邓彦斌 , 姜彦成 , 刘健 (1998 ). 新疆10种藜科植物叶片和同化枝的旱生和盐生结构的研究植物生态学报 , 22, 164 -170 .] [本文引用: 1] 本文应用扫描电镜和光学显微镜对生长在新疆荒漠地区10种藜科植物中亚滨藜(Atriplex centralasiatica),心叶驼绒藜(Ceratoides ewersmanniana),驼绒藜(Ceratoides latens),盐节木(Halocnemum strobilaceum),盐穗木(Halostachys caspica),梭梭(Haloxylon ammodendron),圆叶盐爪爪(Kalidium schrenkianum),绒藜(Londesia eriantha),费尔干猪毛菜(Salsola ferganica),浆果猪毛菜(Salsola foliosa)的叶和同化枝进行了形态解剖学研究。结果表明,它们是通过以下结构来适应旱生和盐生环境的:叶片及角质膜厚,气孔器下陷,具表皮毛;栅栏组织发达,多为等面叶;部分植物叶片退化成鳞片状,而由同化枝执行光合功能;多数植物叶片和同化枝内部具有粘液和含晶细胞,贮水组织发达。根据盐分是否排出体外,又划分出聚盐和泌盐植物。在泌盐植物中,盐腺具有单细胞和多细胞及分泌孔类型,并对其聚盐和泌盐机理作了初步探讨。
[6] Guo GG Feng B Ma BL Zhang YL Guo CH Jing ZB 2013 ). Leaf anatomical structures of different regional Amygdalus pedunculata Pall. and their drought resistance analysisActa Botanica Boreali-Occidentalia Sinica 720 -728 . [本文引用: 1] [郭改改 , 封斌 , 麻保林 , 张应龙 , 郭春会 , 井赵斌 (2013 ). 不同区域长柄扁桃叶片解剖结构及其抗旱性分析西北植物学报 , 33, 720 -728 .] [本文引用: 1] [7] He T Wu XM Jia JF 2007 ). Research advances in morphology and anatomy of alpine plants growing in the Qinghai- Tibet Plateau and their adaptations to environmentsActa Ecologica Sinica 2574 -2583 . [本文引用: 1] [何涛 , 吴学明 , 贾敬芬 (2007 ). 青藏高原高山植物的形态和解剖结构及其对环境的适应性研究进展生态学报 , 27, 2574 -2583 .] [本文引用: 1] [8] Hetherington AM Woodward FI 2003 ). The role of stomata in sensing and driving environmental changeNature 901 -908 . PMID:
12931178 [本文引用: 1] Stomata, the small pores on the surfaces of leaves and stalks, regulate the flow of gases in and out of leaves and thus plants as a whole. They adapt to local and global changes on all timescales from minutes to millennia. Recent data from diverse fields are establishing their central importance to plant physiology, evolution and global ecology. Stomatal morphology, distribution and behaviour respond to a spectrum of signals, from intracellular signalling to global climatic change. Such concerted adaptation results from a web of control systems, reminiscent of a 'scale-free' network, whose untangling requires integrated approaches beyond those currently used.
[9] Holmes MG Keiller DR 2002 ). Effects of pubescence and waxes on the reflectance of leaves in the ultraviolet and photosynthetic wavebands: a comparison of a range of speciesPlant, Cell & Environment 85 -93 . [本文引用: 1] [10] Hu JY Guo K Dong M 2008 ). Variation of leaf structure of two dominant species in alpine grassland and the relationship between leaf structure and ecological factorsJournal of Plant Ecology Chinese Version ), 32, 370 -378 . [本文引用: 1] [胡建莹 , 郭柯 , 董鸣 (2008 ). 高寒草原优势种叶片结构变化与生态因子的关系植物生态学报 , 32, 370 -378 .] DOI:
10.3773/j.issn.1005-264x.2008.02.014 [本文引用: 1] 为了揭示青藏高原高寒草原优势物种青藏苔草(Carex moorcroftii)和紫花针茅(Stipa purpurea)对高原特殊环境的适应性,该文研究了 它们的叶片结构在自然环境梯度下的变化以及这些变化与生态因子之间的关系。结果表明:这两个物种叶片的大多数结构特征在各个样地间变 化显著,其结构特征与环境因子间存在显著的相关关系。紫花针茅叶肉细胞大小随土壤有效K含量的增高而减小,下表皮细胞厚度和韧皮部面积 随生长季云盖度的增高而增加,单一导管半径和导管平均面积随生长季月均湿度的增加而增大;青藏苔草上表皮细胞厚度随生长季月均最低温 的降低而增厚,泡状细胞厚度(径向直径)随大陆度的增强而增加,上表皮细胞大小随土壤pH值的增大而增大,导管总数和韧皮部面积随土壤 速效P含量的增高而增加。青藏苔草的保护组织、光合组织以及综合指标变异系数明显大于紫花针茅,仅维管组织指标变异系数小于紫花针茅。
[11] Hu MY Zhang L Luo TX Shen W 2012 ). Variations in leaf functional traits of Stipa purpurea along a rainfall gradient in Xizang, ChinaChinese Journal of Plant Ecology 136 -143 . DOI:
10.3724/SP.J.1258.2012.00136 URL [本文引用: 2] [胡梦瑶 , 张林 , 罗天祥 , 沈维 (2012 ). 西藏紫花针茅叶功能性状沿降水梯度的变化植物生态学报 , 36, 136 -143 .] DOI:
10.3724/SP.J.1258.2012.00136 [本文引用: 2] 植物叶功能性状与环境因子的关系是近10年来植物生态学的研究热点。该文以广泛分布于青藏高原干旱、半干旱草地的优势植物种紫花针茅(Stipa purpurea)为研究对象, 沿降水梯度(69–479 mm)系统测定了日土、改则、珠峰、当雄和纳木错5个调查地点紫花针茅比叶面积(SLA)、单位重量和单位面积叶氮含量(N<sub>mass</sub>, N<sub>area</sub>)、叶密度和厚度等叶功能性状以及土壤全氮含量等因子, 试图验证干旱胁迫地区同一物种内SLA-N<sub>mass</sub>关系沿降水梯度的策略位移现象是否具有普遍性, 并对是否出现策略位移现象提出可能的解释。研究结果表明: 1) SLA和N<sub>mass</sub>与生长季温度和降水以及土壤全氮含量均没有显著关系, SLA与N<sub>mass</sub>的关系在干旱半干旱区(年降水/蒸发比 0.11)之间并没有出现典型的位移现象; 2)叶密度是决定半湿润区SLA变化的主导因子, 而叶厚度则是干旱半干旱区SLA变化的控制因子, 两者与SLA均呈负相关, 随着温度增加或降水减少, 叶厚度增加而叶密度降低, 导致SLA随温度和降水变化不明显; 3)半湿润区的叶密度增加引起N<sub>area</sub>增加, 而干旱半干旱区的叶厚度增加并没有造成N<sub>area</sub>的显著变化, 导致N<sub>area</sub>沿降水梯度没有显著变化; 4)紫花针茅地上生物量与N<sub>area</sub>具有显著正相关关系, 表明N<sub>area</sub>的增加有助于提高植被生产力。结果表明, 在干旱胁迫下, 植物通过增加叶厚度来维持不变的N<sub>area</sub>可能有助于保持与较湿润地区相似的光合生产和水分利用效率。叶厚度和叶密度对比叶面积的相对影响在干旱半干旱区与半湿润区之间发生转变, 这为进一步检测高寒草地植被的水分限制阈值提供了新思路。
[12] Hu XP Ji CJ An LH 2016 ). Leaf epidermis characteristics of the main grassland monocotyledonous plant species on the Tibetan PlateauActa Ecologica Sinica 6465 -6474 . [本文引用: 1] [胡选萍 , 吉成均 , 安丽华 (2016 ). 青藏高原草地主要单子叶植物的叶表面特征生态学报 , 36, 6465 -6474 .] [本文引用: 1] [13] Ji RX Yu X Chang Y Shen C Bai XQ Xia XL Yin WL Liu C 2020 ). Geographical provenance variation of leaf anatomical structure of Caryopteris mongholica and its significance in response to environmental changesChinese Joumal of Plant Ecology 277 -286 . [本文引用: 1] [纪若璇 , 于笑 , 常远 , 沈超 , 白雪卡 , 夏新莉 , 尹伟伦 , 刘超 (2020 ). 蒙古莸叶片解剖结构的地理种源变异及其对环境变化响应的意义植物生态学报 , 44, 277 -286 .] DOI:
10.17521/cjpe.2019.0117 [本文引用: 1] 长期受到生长环境影响而形成的遗传变异对植物生长发育有着显著的影响。叶片是植物对环境变化最敏感的器官, 了解叶片解剖结构在不同环境中产生的适应性变异是探索植物对环境适应的基础。同质园试验是研究遗传与环境因素对植物生长代谢等影响的一种有效方法, 该研究利用同质园试验排除了环境梯度的影响, 通过常规石蜡切片、多重比较、相关性分析、一般线性模型分析等方法, 对7个不同种源地的蒙古莸(Caryopteris mongholica)叶片解剖结构及其影响因素进行了定量比较。结果表明, 7个种源地的蒙古莸叶片均为等面叶, 无海绵组织分化, 其上表皮细胞较下表皮细胞厚, 上栅栏组织较下栅栏组织厚; 叶片各解剖结构参数间存在显著的自相关性, 不同种源叶片解剖结构存在显著差异: 随种源地年平均气温升高, 叶厚度、栅栏组织厚度呈增大趋势, 其中, 最西南部的阿左旗种源蒙古莸叶片的上下栅栏组织、叶厚度及叶片结构紧密度值均最大, 表现出明显的抗旱特征。种源地经纬度、气温、降水等对解剖结构指标有显著的影响, 其解释程度为34.09%-81.43%。同质园试验说明, 种源地气候差异驱动的遗传变异是引起不同种源叶片解剖结构差异的重要因素。
[14] Kang S Niu JM Zhang Q Chen LP 2013 ). Anatomical structure of Stipa breviflora leaves and its relationship with environmental factorsActa Prataculturae Sinica 77 -86 . [本文引用: 5] [康萨如拉 , 牛建明 , 张庆 , 陈丽萍 (2013 ). 短花针茅叶片解剖结构及与气候因子的关系草业学报 , 22, 77 -86 .] [本文引用: 5] [15] Kou M Yin QL Jiao JY 2019 ). Leaf anatomical structures and acclimation of ten monocotyledons in the Hilly-Gullied Loess Plateau regionActa Botanica Boreal-Occidentalia Sinica 102 -109 . [本文引用: 2] [寇萌 , 尹秋龙 , 焦菊英 (2019 ). 黄土丘陵沟壑区10种单子叶植物叶片解剖结构及环境适应性西北植物学报 , 39, 102 -109 .] [本文引用: 2] [16] Li QF Wang BJ An LH Ji CJ 2013 ). Leaf anatomical characteristics of the plants of grasslands in the Tibetan PlateauActa Ecologica Sinica 2062 -2070 . DOI:
10.5846/stxb URL [本文引用: 1] [李全发 , 王宝娟 , 安丽华 , 吉成均 (2013 ). 青藏高原草地植物叶解剖特征生态学报 , 33, 2062 -2070 .] [本文引用: 1] [17] Li X Yang SH Yang YQ Yin X Sun XD Yang YP 2015 ). Comparative physiological and molecular analyses of intraspecific differences of Stipa purpurea (Poaceae) response to droughtPlant Diversity and Resources 439 -452 . [本文引用: 1] [李雄 , 杨时海 , 杨云强 , 尹欣 , 孙旭东 , 杨永平 (2015 ). 不同居群紫花针茅响应干旱胁迫的生理和分子差异分析植物分类与资源学报 , 37, 439 -452 .] [本文引用: 1] [18] Li ZL 1981 ). Morphology and structure of drought vegetationBiology of Journal 9 -12 . [本文引用: 1] [李正理 (1981 ). 旱生植物的形态和结构生物学通报 , 16, 9 -12 .] [本文引用: 1] [19] Ma B Sun J 2018 ). Predicting the distribution of Stipa purpurea across the Tibetan Plateau via the MaxEnt modelBMC Ecology 1 -12 . DOI:
10.1186/s12898-018-0157-0 URL [本文引用: 1] [20] Ma JJ Ji CJ Han M Zhang FT Yan XD Hu D Zeng H He JS 2012 ). Comparative analyses of leaf anatomy of dicotyledonous species in Tibetan and Inner Mongolian grasslandsScience China, Life Science 68 -79 . DOI:
10.1007/s11427-012-4268-0 URL [本文引用: 1] [21] Meng M Ni J Zhang ZG 2004 ). Aridity index and its applications in geo-ecological studyActa Phytoecologica Sinica 853 -861 . [本文引用: 1] [孟猛 , 倪健 , 张治国 (2004 ). 地理生态学的干燥度指数及其应用评述植物生态学报 , 28, 853 -861 .] DOI:
10.17521/cjpe.2004.0111 [本文引用: 1] 干燥度指数(Aridity index,AI,该文特指气候干燥度)是表征一个地区干湿程度的指标,在地理学和生态学研究中长期应用,近来成为全球变化研究中经常涉及到的气候指标之一,尤其是气候变化和干旱化、荒漠化等研究。该文列举了国内外目前存在的22种干燥度指数,并对目前常用的8种指数进行了详细描述和分析,包括各自的原理、计算方法和在生态学与地理学研究中的应用等。结合其应用和中国的实际状况,分析了各自的优缺点,认为修改后的谢氏干燥度、de Martonne干燥度和Holdridge可能蒸散率(等同于某种意义上的干燥度)计算方法简单实用,有着明确的物理学和生态学意义,与我国的实际情况对应性较强,适合于中国气候变化及其相关的干旱化、荒漠化等方面的研究。
[22] Sun HT Jiang S Liu JM Cuo YJ Shen GS Gu S 2016 ). Structure and ecological adaptability of the leaves of three Asteracae species different altitudes on the Qinghai-Tibet PlateauActa Ecologica Sinica 1559 -1570 . [本文引用: 1] [孙会婷 , 江莎 , 刘婧敏 , 郭亚娇 , 沈广爽 , 古松 (2016 ). 青藏高原不同海拔3种菊科植物时片结构变化及其生态适应性生态学报 , 36, 1559 -1570 .] [本文引用: 1] [23] Tan CP Yang JP Mi R 2010 ). Analysis of the climatic change characteristics in the Southern Tibetan Plateau from 1971 to 2007Journal of Glaciology and Geocryology 1111 -1120 . [本文引用: 1] [谭春萍 , 杨建平 , 米睿 (2010 ). 1971-2007年青藏高原南部气候变化特征分析冰川冻土 , 32, 1111 -1120 .] [本文引用: 1] [24] Wang CS Wang SP 2015 ). A review of research on responses of leaf traits to climate changeChinese Journal of Plant Ecology 206 -216 . DOI:
10.17521/cjpe.2015.0020 URL [本文引用: 1] [王常顺 , 汪诗平 (2015 ). 植物叶片性状对气候变化的响应研究进展植物生态学报 , 39, 206 -216 .] DOI:
10.17521/cjpe.2015.0020 [本文引用: 1] 叶片性状反映了植物对环境的高度适应能力及其在复杂生境下的自我调控能力。叶片性状如何响应和适应气候变化是植物适应性研究的重点内容。该文系统综述了叶片大小、比叶质量、叶片氮含量、碳同位素等指标对气候变化响应的最新研究结果。不同叶片性状对气候变化的响应结果存在差异, 所指示的生态学含义也有所不同。单一叶片性状不能全面地反映植物对气候变化的响应; 不同尺度的研究(如环境的修饰或筛选作用的研究)还存在很多不确定性。高寒地区的研究工作相对缺乏。该文有助于理解植物与气候之间的相互关系、植物对气候变化的响应与适应对策, 对了解植物演化、预测植物在未来气候变化条件下的变化特征具有一定意义。
[25] Wang M Li Y Hang RQ Li YL 2005 ). The effects of climate warming on the alpine vegetation of the Qinghai-Tibetan Plateau hinterlandActa Ecologica Sinica 1275 -1281 . [本文引用: 1] [王谋 , 李勇 , 黄润秋 , 李亚林 (2005 ). 气候变暖对青藏高原腹地高寒植被的影响生态学报 , 25, 1275 -1281 .] [本文引用: 1] [26] Wright IJ Reich PB Westoby M 2001 ). Strategy shifts in leaf physiology, structure and nutrient content between species of high- and 1ow-rainfall and high- and low-nutrient habitatsFunctional Ecology 423 -434 . DOI:
10.1046/j.0269-8463.2001.00542.x URL [本文引用: 1] [27] Wu JB Wang XD 2019 ). Temporal stability of aboveground net primary production in northern Tibet alpine steppe in response to nitrogen additionJournal of Mountain Science 2679 -2686 . DOI:
10.1007/s11629-018-5135-7 URL [本文引用: 1] [28] Yang MB Yang J Yang JY Liang N Qing H 2007 ). Changes of characteristics of the leaf epidermis and genetic diversity of Caragana davazamcii in different habitats in Erdos Plateau, ChinaJournal of Plant Ecology Chinese Version ), 31, 1181 -1189 . [本文引用: 1] [杨明博 , 杨劼 , 杨九艳 , 梁娜 , 清华 (2007 ). 鄂尔多斯高原不同生境条件下中间锦鸡儿植物叶片表皮特征及遗传多样性变化分析植物生态学报 , 31, 1181 -1189 .] DOI:
10.17521/cjpe.2007.0147 [本文引用: 1] 该研究在鄂尔多斯高原从东到西按照降雨量逐渐减少的梯度选取了5个地区,对中间锦鸡儿(Caragana davazamcii)叶片的表皮结构进行了比较 观察,发现不同生境条件下的中间锦鸡儿由于环境的长期作用表皮表现出一定的差异性,主要表现在随着多年平均降雨量的减少叶片表皮毛密 度、气孔密度、气孔指数均有所增加,气孔深陷程度逐渐加深,角质层由光滑变得粗糙,但并不是线性增加,因此本研究对不同生境的中间锦 鸡儿植物叶片做了遗传多样性分析,发现随着生 境由东向西的变化,中间锦鸡儿种群遗传多样性有增加的趋势,与表皮形态特征变化一致, 说明中间锦鸡儿呈现明显的表现型可塑性,可塑性反应比较高。
[29] Zhong YM Dong FY Wang WJ Wang JM Li JW Wu B Jia XH 2017 ). Anatomical characteristics and adaptability plasticity of Populus euphratica in different habitatsJournal of Beijing Forestry University 10 ), 53 -61 . [本文引用: 1] [钟悦鸣 , 董芳宇 , 王文娟 , 王健铭 , 李景文 , 吴波 , 贾晓红 (2017 ). 不同生境胡杨叶片解剖特征及其适应可塑性北京林业大学学报 , 39(10 ), 53 -61 .] [本文引用: 1] How does leaf anatomy influence water transport outside the xylem
1
2015
... 紫花针茅作为高寒草原的优势种, 其叶片解剖结构对高寒干旱环境具有较好的适应性.紫花针茅叶片外表皮普遍具有较厚角质层, 而在特别干旱区域(噶尔), 紫花针茅内表皮上也有角质层形成(
图2 H), 这有利于减少水分散失, 适应干旱环境(邓彦斌等,
1998 ; 蔡永立和宋永昌,
2001 ).同时紫花针茅叶片通过增加厚角细胞厚度, 减少水分散失(
图2 ), 这与内蒙古短花针茅适应干旱环境的叶片解剖结构 (康萨如拉等,
2013 )相一致.为了适应干旱环境, 紫花针茅在改变保护组织的同时, 增加输水组织面积比例(导管直径、主脉导管面积/主脉维管束面积和维管束面积/叶横切面积), 以增强水分运输适应干旱环境(康萨如拉等,
2013 ; Buckley
et al .,
2015 ; 寇萌等,
2019 ). ...
浙江天童常绿阔叶林藤本植物的适应生态学: I. 叶片解剖特征的比较
1
2001
... 紫花针茅作为高寒草原的优势种, 其叶片解剖结构对高寒干旱环境具有较好的适应性.紫花针茅叶片外表皮普遍具有较厚角质层, 而在特别干旱区域(噶尔), 紫花针茅内表皮上也有角质层形成(
图2 H), 这有利于减少水分散失, 适应干旱环境(邓彦斌等,
1998 ; 蔡永立和宋永昌,
2001 ).同时紫花针茅叶片通过增加厚角细胞厚度, 减少水分散失(
图2 ), 这与内蒙古短花针茅适应干旱环境的叶片解剖结构 (康萨如拉等,
2013 )相一致.为了适应干旱环境, 紫花针茅在改变保护组织的同时, 增加输水组织面积比例(导管直径、主脉导管面积/主脉维管束面积和维管束面积/叶横切面积), 以增强水分运输适应干旱环境(康萨如拉等,
2013 ; Buckley
et al .,
2015 ; 寇萌等,
2019 ). ...
浙江天童常绿阔叶林藤本植物的适应生态学: I. 叶片解剖特征的比较
1
2001
... 紫花针茅作为高寒草原的优势种, 其叶片解剖结构对高寒干旱环境具有较好的适应性.紫花针茅叶片外表皮普遍具有较厚角质层, 而在特别干旱区域(噶尔), 紫花针茅内表皮上也有角质层形成(
图2 H), 这有利于减少水分散失, 适应干旱环境(邓彦斌等,
1998 ; 蔡永立和宋永昌,
2001 ).同时紫花针茅叶片通过增加厚角细胞厚度, 减少水分散失(
图2 ), 这与内蒙古短花针茅适应干旱环境的叶片解剖结构 (康萨如拉等,
2013 )相一致.为了适应干旱环境, 紫花针茅在改变保护组织的同时, 增加输水组织面积比例(导管直径、主脉导管面积/主脉维管束面积和维管束面积/叶横切面积), 以增强水分运输适应干旱环境(康萨如拉等,
2013 ; Buckley
et al .,
2015 ; 寇萌等,
2019 ). ...
青藏高原环境变化科学评估: 过去、现在与未来
2
2015
... 紫花针茅(
Stipa purpurea )作为高寒草原的优势种, 对维持群落稳定性具有重要作用(Wu & Wang,
2019 ).气候变暖导致青藏高原的西部地区变暖变干, 东部地区变暖变湿(谭春萍等,
2010 ; 陈德亮等,
2015 ), 进而改变青藏高原草地生态系统的空间格局, 高山嵩草(
Kobresia pygmaea )群落向紫花针茅群落退化(王谋等,
2005 ), 紫花针茅在青藏高原的分布范围随着温度升高将继续扩大(陈德亮等,
2015 ; Ma & Sun,
2018 ).紫花针茅分布范围增加说明其具有对干旱生境较强的适应能力, 然而其如何适应高寒干旱气候变化的研究还相对薄弱(胡建莹等,
2008 ; 胡梦瑶等,
2012 ; 李雄等,
2015 ; 王常顺和汪诗平,
2015 ).对不同区域紫花针茅叶片解剖结构的分析, 可以直接反映紫花针茅对青藏高原气候变化的适应策略.因此, 本研究通过对紫花针茅在青藏高原分布区域进行采样, 分析其叶片解剖结构特征变异规律及其如何对环境因子响应, 以期为阐明高寒草原植物对气候变化适应机制提供科学依据. ...
... ), 紫花针茅在青藏高原的分布范围随着温度升高将继续扩大(陈德亮等,
2015 ; Ma & Sun,
2018 ).紫花针茅分布范围增加说明其具有对干旱生境较强的适应能力, 然而其如何适应高寒干旱气候变化的研究还相对薄弱(胡建莹等,
2008 ; 胡梦瑶等,
2012 ; 李雄等,
2015 ; 王常顺和汪诗平,
2015 ).对不同区域紫花针茅叶片解剖结构的分析, 可以直接反映紫花针茅对青藏高原气候变化的适应策略.因此, 本研究通过对紫花针茅在青藏高原分布区域进行采样, 分析其叶片解剖结构特征变异规律及其如何对环境因子响应, 以期为阐明高寒草原植物对气候变化适应机制提供科学依据. ...
青藏高原环境变化科学评估: 过去、现在与未来
2
2015
... 紫花针茅(
Stipa purpurea )作为高寒草原的优势种, 对维持群落稳定性具有重要作用(Wu & Wang,
2019 ).气候变暖导致青藏高原的西部地区变暖变干, 东部地区变暖变湿(谭春萍等,
2010 ; 陈德亮等,
2015 ), 进而改变青藏高原草地生态系统的空间格局, 高山嵩草(
Kobresia pygmaea )群落向紫花针茅群落退化(王谋等,
2005 ), 紫花针茅在青藏高原的分布范围随着温度升高将继续扩大(陈德亮等,
2015 ; Ma & Sun,
2018 ).紫花针茅分布范围增加说明其具有对干旱生境较强的适应能力, 然而其如何适应高寒干旱气候变化的研究还相对薄弱(胡建莹等,
2008 ; 胡梦瑶等,
2012 ; 李雄等,
2015 ; 王常顺和汪诗平,
2015 ).对不同区域紫花针茅叶片解剖结构的分析, 可以直接反映紫花针茅对青藏高原气候变化的适应策略.因此, 本研究通过对紫花针茅在青藏高原分布区域进行采样, 分析其叶片解剖结构特征变异规律及其如何对环境因子响应, 以期为阐明高寒草原植物对气候变化适应机制提供科学依据. ...
... ), 紫花针茅在青藏高原的分布范围随着温度升高将继续扩大(陈德亮等,
2015 ; Ma & Sun,
2018 ).紫花针茅分布范围增加说明其具有对干旱生境较强的适应能力, 然而其如何适应高寒干旱气候变化的研究还相对薄弱(胡建莹等,
2008 ; 胡梦瑶等,
2012 ; 李雄等,
2015 ; 王常顺和汪诗平,
2015 ).对不同区域紫花针茅叶片解剖结构的分析, 可以直接反映紫花针茅对青藏高原气候变化的适应策略.因此, 本研究通过对紫花针茅在青藏高原分布区域进行采样, 分析其叶片解剖结构特征变异规律及其如何对环境因子响应, 以期为阐明高寒草原植物对气候变化适应机制提供科学依据. ...
1
2010
... 植物与其生长的环境是统一的整体, 环境对植物的长期作用影响植物叶片解剖结构.在本研究中, 不同地理种群紫花针茅叶片解剖结构特征既存在相似性, 也存在较大变异性.角质层厚度在不同地理种群紫花针茅叶片结构特征中不存在显著差异, 这是因为角质层不仅可以反射强光, 还可以减少叶片水分蒸散, 有利于植物适应高原环境(李正理,
1981 ; Holmes & Keiller,
2002 ; 何涛等,
2007 ), 体现了不同地理种群紫花针茅叶片结构对高海拔生境的趋同适应进化.但是, 生境的不同导致紫花针茅叶片结构解剖特征存在较大差异, 例如紫花针茅叶片厚角细胞厚度随着干旱程度的增加而增加, 厚角细胞从安多紫花针茅叶片解剖结构中不存在到噶尔紫花针茅叶片中厚度逐渐增加, 而增加厚壁细胞厚度可减少水分蒸散损失(康萨如拉等,
2013 ).紫花针茅叶片解剖结构输水组织在不同区域也存在较大差异, 导管直径、主脉导管腔面积/主脉维管束面积和维管束面积/叶横切面积随着降水量的减少而增加, 这利于增加水分运输能力, 适应干旱环境(Cox & Moorc,
2010 ; 康萨如拉等,
2013 ; 寇萌等,
2019 ).因此, 不同地理种群紫花针茅叶片既有相似的解剖结构特征适应高海拔生境, 也有不同的叶片解剖结构适应区域生境. ...
新疆10种藜科植物叶片和同化枝的旱生和盐生结构的研究
1
1998
... 紫花针茅作为高寒草原的优势种, 其叶片解剖结构对高寒干旱环境具有较好的适应性.紫花针茅叶片外表皮普遍具有较厚角质层, 而在特别干旱区域(噶尔), 紫花针茅内表皮上也有角质层形成(
图2 H), 这有利于减少水分散失, 适应干旱环境(邓彦斌等,
1998 ; 蔡永立和宋永昌,
2001 ).同时紫花针茅叶片通过增加厚角细胞厚度, 减少水分散失(
图2 ), 这与内蒙古短花针茅适应干旱环境的叶片解剖结构 (康萨如拉等,
2013 )相一致.为了适应干旱环境, 紫花针茅在改变保护组织的同时, 增加输水组织面积比例(导管直径、主脉导管面积/主脉维管束面积和维管束面积/叶横切面积), 以增强水分运输适应干旱环境(康萨如拉等,
2013 ; Buckley
et al .,
2015 ; 寇萌等,
2019 ). ...
新疆10种藜科植物叶片和同化枝的旱生和盐生结构的研究
1
1998
... 紫花针茅作为高寒草原的优势种, 其叶片解剖结构对高寒干旱环境具有较好的适应性.紫花针茅叶片外表皮普遍具有较厚角质层, 而在特别干旱区域(噶尔), 紫花针茅内表皮上也有角质层形成(
图2 H), 这有利于减少水分散失, 适应干旱环境(邓彦斌等,
1998 ; 蔡永立和宋永昌,
2001 ).同时紫花针茅叶片通过增加厚角细胞厚度, 减少水分散失(
图2 ), 这与内蒙古短花针茅适应干旱环境的叶片解剖结构 (康萨如拉等,
2013 )相一致.为了适应干旱环境, 紫花针茅在改变保护组织的同时, 增加输水组织面积比例(导管直径、主脉导管面积/主脉维管束面积和维管束面积/叶横切面积), 以增强水分运输适应干旱环境(康萨如拉等,
2013 ; Buckley
et al .,
2015 ; 寇萌等,
2019 ). ...
不同区域长柄扁桃叶片解剖结构及其抗旱性分析
1
2013
... 叶片解剖结构特征是植物长期适应环境以及自身进化形成的, 体现了植物对环境的适应策略(Hetherington & Woodward,
2003 ), 而且叶片解剖结构在不同区域的变异特征可以直接反映植物对不同生长环境的适应策略(Wright
et al .,
2001 ; 钟悦鸣等,
2017 ).例如中间锦鸡儿(
Caragana davazamcii )随着平均年降水量的减少, 角质层厚度有所增加, 栅栏组织分化逐渐加强, 叶脉的机械组织分化逐渐加强(杨明博,
2007 ); 长梗扁桃(
Amygdalus pedunculata )生长在干旱地区的叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的叶片栅栏组织排列疏松, 叶厚度较薄(郭改改等,
2013 ); 青藏高原的菊科植物叶片厚度、上下表皮厚度、上下角质层厚度、栅栏细胞系数均随海拔升高而增加(孙会婷等,
2016 ); 蒙古莸(
Caryopteris mongholica )叶片解剖结构随着年平均气温升高, 叶厚度、栅栏组织厚度呈增大趋势(纪若璇等,
2020 ).因此, 叶片解剖结构可以体现植物对环境因子的适应性进化, 开展不同区域植物叶片解剖结构变异特征的研究, 对阐明植物适应气候变化策略具有重要意义. ...
不同区域长柄扁桃叶片解剖结构及其抗旱性分析
1
2013
... 叶片解剖结构特征是植物长期适应环境以及自身进化形成的, 体现了植物对环境的适应策略(Hetherington & Woodward,
2003 ), 而且叶片解剖结构在不同区域的变异特征可以直接反映植物对不同生长环境的适应策略(Wright
et al .,
2001 ; 钟悦鸣等,
2017 ).例如中间锦鸡儿(
Caragana davazamcii )随着平均年降水量的减少, 角质层厚度有所增加, 栅栏组织分化逐渐加强, 叶脉的机械组织分化逐渐加强(杨明博,
2007 ); 长梗扁桃(
Amygdalus pedunculata )生长在干旱地区的叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的叶片栅栏组织排列疏松, 叶厚度较薄(郭改改等,
2013 ); 青藏高原的菊科植物叶片厚度、上下表皮厚度、上下角质层厚度、栅栏细胞系数均随海拔升高而增加(孙会婷等,
2016 ); 蒙古莸(
Caryopteris mongholica )叶片解剖结构随着年平均气温升高, 叶厚度、栅栏组织厚度呈增大趋势(纪若璇等,
2020 ).因此, 叶片解剖结构可以体现植物对环境因子的适应性进化, 开展不同区域植物叶片解剖结构变异特征的研究, 对阐明植物适应气候变化策略具有重要意义. ...
青藏高原高山植物的形态和解剖结构及其对环境的适应性研究进展
1
2007
... 植物与其生长的环境是统一的整体, 环境对植物的长期作用影响植物叶片解剖结构.在本研究中, 不同地理种群紫花针茅叶片解剖结构特征既存在相似性, 也存在较大变异性.角质层厚度在不同地理种群紫花针茅叶片结构特征中不存在显著差异, 这是因为角质层不仅可以反射强光, 还可以减少叶片水分蒸散, 有利于植物适应高原环境(李正理,
1981 ; Holmes & Keiller,
2002 ; 何涛等,
2007 ), 体现了不同地理种群紫花针茅叶片结构对高海拔生境的趋同适应进化.但是, 生境的不同导致紫花针茅叶片结构解剖特征存在较大差异, 例如紫花针茅叶片厚角细胞厚度随着干旱程度的增加而增加, 厚角细胞从安多紫花针茅叶片解剖结构中不存在到噶尔紫花针茅叶片中厚度逐渐增加, 而增加厚壁细胞厚度可减少水分蒸散损失(康萨如拉等,
2013 ).紫花针茅叶片解剖结构输水组织在不同区域也存在较大差异, 导管直径、主脉导管腔面积/主脉维管束面积和维管束面积/叶横切面积随着降水量的减少而增加, 这利于增加水分运输能力, 适应干旱环境(Cox & Moorc,
2010 ; 康萨如拉等,
2013 ; 寇萌等,
2019 ).因此, 不同地理种群紫花针茅叶片既有相似的解剖结构特征适应高海拔生境, 也有不同的叶片解剖结构适应区域生境. ...
青藏高原高山植物的形态和解剖结构及其对环境的适应性研究进展
1
2007
... 植物与其生长的环境是统一的整体, 环境对植物的长期作用影响植物叶片解剖结构.在本研究中, 不同地理种群紫花针茅叶片解剖结构特征既存在相似性, 也存在较大变异性.角质层厚度在不同地理种群紫花针茅叶片结构特征中不存在显著差异, 这是因为角质层不仅可以反射强光, 还可以减少叶片水分蒸散, 有利于植物适应高原环境(李正理,
1981 ; Holmes & Keiller,
2002 ; 何涛等,
2007 ), 体现了不同地理种群紫花针茅叶片结构对高海拔生境的趋同适应进化.但是, 生境的不同导致紫花针茅叶片结构解剖特征存在较大差异, 例如紫花针茅叶片厚角细胞厚度随着干旱程度的增加而增加, 厚角细胞从安多紫花针茅叶片解剖结构中不存在到噶尔紫花针茅叶片中厚度逐渐增加, 而增加厚壁细胞厚度可减少水分蒸散损失(康萨如拉等,
2013 ).紫花针茅叶片解剖结构输水组织在不同区域也存在较大差异, 导管直径、主脉导管腔面积/主脉维管束面积和维管束面积/叶横切面积随着降水量的减少而增加, 这利于增加水分运输能力, 适应干旱环境(Cox & Moorc,
2010 ; 康萨如拉等,
2013 ; 寇萌等,
2019 ).因此, 不同地理种群紫花针茅叶片既有相似的解剖结构特征适应高海拔生境, 也有不同的叶片解剖结构适应区域生境. ...
The role of stomata in sensing and driving environmental change
1
2003
... 叶片解剖结构特征是植物长期适应环境以及自身进化形成的, 体现了植物对环境的适应策略(Hetherington & Woodward,
2003 ), 而且叶片解剖结构在不同区域的变异特征可以直接反映植物对不同生长环境的适应策略(Wright
et al .,
2001 ; 钟悦鸣等,
2017 ).例如中间锦鸡儿(
Caragana davazamcii )随着平均年降水量的减少, 角质层厚度有所增加, 栅栏组织分化逐渐加强, 叶脉的机械组织分化逐渐加强(杨明博,
2007 ); 长梗扁桃(
Amygdalus pedunculata )生长在干旱地区的叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的叶片栅栏组织排列疏松, 叶厚度较薄(郭改改等,
2013 ); 青藏高原的菊科植物叶片厚度、上下表皮厚度、上下角质层厚度、栅栏细胞系数均随海拔升高而增加(孙会婷等,
2016 ); 蒙古莸(
Caryopteris mongholica )叶片解剖结构随着年平均气温升高, 叶厚度、栅栏组织厚度呈增大趋势(纪若璇等,
2020 ).因此, 叶片解剖结构可以体现植物对环境因子的适应性进化, 开展不同区域植物叶片解剖结构变异特征的研究, 对阐明植物适应气候变化策略具有重要意义. ...
Effects of pubescence and waxes on the reflectance of leaves in the ultraviolet and photosynthetic wavebands: a comparison of a range of species
1
2002
... 植物与其生长的环境是统一的整体, 环境对植物的长期作用影响植物叶片解剖结构.在本研究中, 不同地理种群紫花针茅叶片解剖结构特征既存在相似性, 也存在较大变异性.角质层厚度在不同地理种群紫花针茅叶片结构特征中不存在显著差异, 这是因为角质层不仅可以反射强光, 还可以减少叶片水分蒸散, 有利于植物适应高原环境(李正理,
1981 ; Holmes & Keiller,
2002 ; 何涛等,
2007 ), 体现了不同地理种群紫花针茅叶片结构对高海拔生境的趋同适应进化.但是, 生境的不同导致紫花针茅叶片结构解剖特征存在较大差异, 例如紫花针茅叶片厚角细胞厚度随着干旱程度的增加而增加, 厚角细胞从安多紫花针茅叶片解剖结构中不存在到噶尔紫花针茅叶片中厚度逐渐增加, 而增加厚壁细胞厚度可减少水分蒸散损失(康萨如拉等,
2013 ).紫花针茅叶片解剖结构输水组织在不同区域也存在较大差异, 导管直径、主脉导管腔面积/主脉维管束面积和维管束面积/叶横切面积随着降水量的减少而增加, 这利于增加水分运输能力, 适应干旱环境(Cox & Moorc,
2010 ; 康萨如拉等,
2013 ; 寇萌等,
2019 ).因此, 不同地理种群紫花针茅叶片既有相似的解剖结构特征适应高海拔生境, 也有不同的叶片解剖结构适应区域生境. ...
高寒草原优势种叶片结构变化与生态因子的关系
1
2008
... 紫花针茅(
Stipa purpurea )作为高寒草原的优势种, 对维持群落稳定性具有重要作用(Wu & Wang,
2019 ).气候变暖导致青藏高原的西部地区变暖变干, 东部地区变暖变湿(谭春萍等,
2010 ; 陈德亮等,
2015 ), 进而改变青藏高原草地生态系统的空间格局, 高山嵩草(
Kobresia pygmaea )群落向紫花针茅群落退化(王谋等,
2005 ), 紫花针茅在青藏高原的分布范围随着温度升高将继续扩大(陈德亮等,
2015 ; Ma & Sun,
2018 ).紫花针茅分布范围增加说明其具有对干旱生境较强的适应能力, 然而其如何适应高寒干旱气候变化的研究还相对薄弱(胡建莹等,
2008 ; 胡梦瑶等,
2012 ; 李雄等,
2015 ; 王常顺和汪诗平,
2015 ).对不同区域紫花针茅叶片解剖结构的分析, 可以直接反映紫花针茅对青藏高原气候变化的适应策略.因此, 本研究通过对紫花针茅在青藏高原分布区域进行采样, 分析其叶片解剖结构特征变异规律及其如何对环境因子响应, 以期为阐明高寒草原植物对气候变化适应机制提供科学依据. ...
高寒草原优势种叶片结构变化与生态因子的关系
1
2008
... 紫花针茅(
Stipa purpurea )作为高寒草原的优势种, 对维持群落稳定性具有重要作用(Wu & Wang,
2019 ).气候变暖导致青藏高原的西部地区变暖变干, 东部地区变暖变湿(谭春萍等,
2010 ; 陈德亮等,
2015 ), 进而改变青藏高原草地生态系统的空间格局, 高山嵩草(
Kobresia pygmaea )群落向紫花针茅群落退化(王谋等,
2005 ), 紫花针茅在青藏高原的分布范围随着温度升高将继续扩大(陈德亮等,
2015 ; Ma & Sun,
2018 ).紫花针茅分布范围增加说明其具有对干旱生境较强的适应能力, 然而其如何适应高寒干旱气候变化的研究还相对薄弱(胡建莹等,
2008 ; 胡梦瑶等,
2012 ; 李雄等,
2015 ; 王常顺和汪诗平,
2015 ).对不同区域紫花针茅叶片解剖结构的分析, 可以直接反映紫花针茅对青藏高原气候变化的适应策略.因此, 本研究通过对紫花针茅在青藏高原分布区域进行采样, 分析其叶片解剖结构特征变异规律及其如何对环境因子响应, 以期为阐明高寒草原植物对气候变化适应机制提供科学依据. ...
西藏紫花针茅叶功能性状沿降水梯度的变化
2
2012
... 紫花针茅(
Stipa purpurea )作为高寒草原的优势种, 对维持群落稳定性具有重要作用(Wu & Wang,
2019 ).气候变暖导致青藏高原的西部地区变暖变干, 东部地区变暖变湿(谭春萍等,
2010 ; 陈德亮等,
2015 ), 进而改变青藏高原草地生态系统的空间格局, 高山嵩草(
Kobresia pygmaea )群落向紫花针茅群落退化(王谋等,
2005 ), 紫花针茅在青藏高原的分布范围随着温度升高将继续扩大(陈德亮等,
2015 ; Ma & Sun,
2018 ).紫花针茅分布范围增加说明其具有对干旱生境较强的适应能力, 然而其如何适应高寒干旱气候变化的研究还相对薄弱(胡建莹等,
2008 ; 胡梦瑶等,
2012 ; 李雄等,
2015 ; 王常顺和汪诗平,
2015 ).对不同区域紫花针茅叶片解剖结构的分析, 可以直接反映紫花针茅对青藏高原气候变化的适应策略.因此, 本研究通过对紫花针茅在青藏高原分布区域进行采样, 分析其叶片解剖结构特征变异规律及其如何对环境因子响应, 以期为阐明高寒草原植物对气候变化适应机制提供科学依据. ...
... 气象数据来自对全国气象台站观测数据插值.气候指标包括: 年降水量、年平均气温、生长季平均气温、生长季降水量、7月平均气温、7月降水量、≥0 ℃积温、年蒸发量.湿润指数采用谢良尼诺夫公式根据中国实际情况修改进行计算(孟猛等,
2004 ).干旱半干旱区与半湿润区按照胡梦瑶等(
2012 )标准划分. ...
西藏紫花针茅叶功能性状沿降水梯度的变化
2
2012
... 紫花针茅(
Stipa purpurea )作为高寒草原的优势种, 对维持群落稳定性具有重要作用(Wu & Wang,
2019 ).气候变暖导致青藏高原的西部地区变暖变干, 东部地区变暖变湿(谭春萍等,
2010 ; 陈德亮等,
2015 ), 进而改变青藏高原草地生态系统的空间格局, 高山嵩草(
Kobresia pygmaea )群落向紫花针茅群落退化(王谋等,
2005 ), 紫花针茅在青藏高原的分布范围随着温度升高将继续扩大(陈德亮等,
2015 ; Ma & Sun,
2018 ).紫花针茅分布范围增加说明其具有对干旱生境较强的适应能力, 然而其如何适应高寒干旱气候变化的研究还相对薄弱(胡建莹等,
2008 ; 胡梦瑶等,
2012 ; 李雄等,
2015 ; 王常顺和汪诗平,
2015 ).对不同区域紫花针茅叶片解剖结构的分析, 可以直接反映紫花针茅对青藏高原气候变化的适应策略.因此, 本研究通过对紫花针茅在青藏高原分布区域进行采样, 分析其叶片解剖结构特征变异规律及其如何对环境因子响应, 以期为阐明高寒草原植物对气候变化适应机制提供科学依据. ...
... 气象数据来自对全国气象台站观测数据插值.气候指标包括: 年降水量、年平均气温、生长季平均气温、生长季降水量、7月平均气温、7月降水量、≥0 ℃积温、年蒸发量.湿润指数采用谢良尼诺夫公式根据中国实际情况修改进行计算(孟猛等,
2004 ).干旱半干旱区与半湿润区按照胡梦瑶等(
2012 )标准划分. ...
青藏高原草地主要单子叶植物的叶表面特征
1
2016
... 高寒草原优势种紫花针茅在青藏高原分布范围较广, 其叶片解剖结构与其生境相适应.Pearson相关性分析和聚类分析结果表明紫花针茅叶片解剖结构与气候因子密切相关(康萨如拉等,
2013 ), 例如五道梁和当雄2个地理种群距离较远, 但其叶片解剖结构比较相似.进一步的主成分和冗余分析结果表明, 与水分相关的气候因子对不同区域紫花针茅叶片解剖结构影响较大(胡选萍等,
2016 ), 即在噶尔、革吉和改则等干旱区域, 年蒸发量是影响厚角细胞厚度、导管直径和主脉导管腔面积/主脉维管束面积的主要因子, 而在相对湿润区域年降水量和生长季降水量等气候因子是影响保护组织(外表皮厚度和角质层厚度)的主要因子.紫花针茅叶片解剖结构未受到低温影响, 这可能是由于紫花针茅分布在高海拔区域, 相较于其他区域植物, 其已经适应了低温环境(Ma
et al .,
2012 ; 李全发等,
2013 ). ...
青藏高原草地主要单子叶植物的叶表面特征
1
2016
... 高寒草原优势种紫花针茅在青藏高原分布范围较广, 其叶片解剖结构与其生境相适应.Pearson相关性分析和聚类分析结果表明紫花针茅叶片解剖结构与气候因子密切相关(康萨如拉等,
2013 ), 例如五道梁和当雄2个地理种群距离较远, 但其叶片解剖结构比较相似.进一步的主成分和冗余分析结果表明, 与水分相关的气候因子对不同区域紫花针茅叶片解剖结构影响较大(胡选萍等,
2016 ), 即在噶尔、革吉和改则等干旱区域, 年蒸发量是影响厚角细胞厚度、导管直径和主脉导管腔面积/主脉维管束面积的主要因子, 而在相对湿润区域年降水量和生长季降水量等气候因子是影响保护组织(外表皮厚度和角质层厚度)的主要因子.紫花针茅叶片解剖结构未受到低温影响, 这可能是由于紫花针茅分布在高海拔区域, 相较于其他区域植物, 其已经适应了低温环境(Ma
et al .,
2012 ; 李全发等,
2013 ). ...
蒙古莸叶片解剖结构的地理种源变异及其对环境变化响应的意义
1
2020
... 叶片解剖结构特征是植物长期适应环境以及自身进化形成的, 体现了植物对环境的适应策略(Hetherington & Woodward,
2003 ), 而且叶片解剖结构在不同区域的变异特征可以直接反映植物对不同生长环境的适应策略(Wright
et al .,
2001 ; 钟悦鸣等,
2017 ).例如中间锦鸡儿(
Caragana davazamcii )随着平均年降水量的减少, 角质层厚度有所增加, 栅栏组织分化逐渐加强, 叶脉的机械组织分化逐渐加强(杨明博,
2007 ); 长梗扁桃(
Amygdalus pedunculata )生长在干旱地区的叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的叶片栅栏组织排列疏松, 叶厚度较薄(郭改改等,
2013 ); 青藏高原的菊科植物叶片厚度、上下表皮厚度、上下角质层厚度、栅栏细胞系数均随海拔升高而增加(孙会婷等,
2016 ); 蒙古莸(
Caryopteris mongholica )叶片解剖结构随着年平均气温升高, 叶厚度、栅栏组织厚度呈增大趋势(纪若璇等,
2020 ).因此, 叶片解剖结构可以体现植物对环境因子的适应性进化, 开展不同区域植物叶片解剖结构变异特征的研究, 对阐明植物适应气候变化策略具有重要意义. ...
蒙古莸叶片解剖结构的地理种源变异及其对环境变化响应的意义
1
2020
... 叶片解剖结构特征是植物长期适应环境以及自身进化形成的, 体现了植物对环境的适应策略(Hetherington & Woodward,
2003 ), 而且叶片解剖结构在不同区域的变异特征可以直接反映植物对不同生长环境的适应策略(Wright
et al .,
2001 ; 钟悦鸣等,
2017 ).例如中间锦鸡儿(
Caragana davazamcii )随着平均年降水量的减少, 角质层厚度有所增加, 栅栏组织分化逐渐加强, 叶脉的机械组织分化逐渐加强(杨明博,
2007 ); 长梗扁桃(
Amygdalus pedunculata )生长在干旱地区的叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的叶片栅栏组织排列疏松, 叶厚度较薄(郭改改等,
2013 ); 青藏高原的菊科植物叶片厚度、上下表皮厚度、上下角质层厚度、栅栏细胞系数均随海拔升高而增加(孙会婷等,
2016 ); 蒙古莸(
Caryopteris mongholica )叶片解剖结构随着年平均气温升高, 叶厚度、栅栏组织厚度呈增大趋势(纪若璇等,
2020 ).因此, 叶片解剖结构可以体现植物对环境因子的适应性进化, 开展不同区域植物叶片解剖结构变异特征的研究, 对阐明植物适应气候变化策略具有重要意义. ...
短花针茅叶片解剖结构及与气候因子的关系
5
2013
... 植物与其生长的环境是统一的整体, 环境对植物的长期作用影响植物叶片解剖结构.在本研究中, 不同地理种群紫花针茅叶片解剖结构特征既存在相似性, 也存在较大变异性.角质层厚度在不同地理种群紫花针茅叶片结构特征中不存在显著差异, 这是因为角质层不仅可以反射强光, 还可以减少叶片水分蒸散, 有利于植物适应高原环境(李正理,
1981 ; Holmes & Keiller,
2002 ; 何涛等,
2007 ), 体现了不同地理种群紫花针茅叶片结构对高海拔生境的趋同适应进化.但是, 生境的不同导致紫花针茅叶片结构解剖特征存在较大差异, 例如紫花针茅叶片厚角细胞厚度随着干旱程度的增加而增加, 厚角细胞从安多紫花针茅叶片解剖结构中不存在到噶尔紫花针茅叶片中厚度逐渐增加, 而增加厚壁细胞厚度可减少水分蒸散损失(康萨如拉等,
2013 ).紫花针茅叶片解剖结构输水组织在不同区域也存在较大差异, 导管直径、主脉导管腔面积/主脉维管束面积和维管束面积/叶横切面积随着降水量的减少而增加, 这利于增加水分运输能力, 适应干旱环境(Cox & Moorc,
2010 ; 康萨如拉等,
2013 ; 寇萌等,
2019 ).因此, 不同地理种群紫花针茅叶片既有相似的解剖结构特征适应高海拔生境, 也有不同的叶片解剖结构适应区域生境. ...
... ; 康萨如拉等,
2013 ; 寇萌等,
2019 ).因此, 不同地理种群紫花针茅叶片既有相似的解剖结构特征适应高海拔生境, 也有不同的叶片解剖结构适应区域生境. ...
... 高寒草原优势种紫花针茅在青藏高原分布范围较广, 其叶片解剖结构与其生境相适应.Pearson相关性分析和聚类分析结果表明紫花针茅叶片解剖结构与气候因子密切相关(康萨如拉等,
2013 ), 例如五道梁和当雄2个地理种群距离较远, 但其叶片解剖结构比较相似.进一步的主成分和冗余分析结果表明, 与水分相关的气候因子对不同区域紫花针茅叶片解剖结构影响较大(胡选萍等,
2016 ), 即在噶尔、革吉和改则等干旱区域, 年蒸发量是影响厚角细胞厚度、导管直径和主脉导管腔面积/主脉维管束面积的主要因子, 而在相对湿润区域年降水量和生长季降水量等气候因子是影响保护组织(外表皮厚度和角质层厚度)的主要因子.紫花针茅叶片解剖结构未受到低温影响, 这可能是由于紫花针茅分布在高海拔区域, 相较于其他区域植物, 其已经适应了低温环境(Ma
et al .,
2012 ; 李全发等,
2013 ). ...
... 紫花针茅作为高寒草原的优势种, 其叶片解剖结构对高寒干旱环境具有较好的适应性.紫花针茅叶片外表皮普遍具有较厚角质层, 而在特别干旱区域(噶尔), 紫花针茅内表皮上也有角质层形成(
图2 H), 这有利于减少水分散失, 适应干旱环境(邓彦斌等,
1998 ; 蔡永立和宋永昌,
2001 ).同时紫花针茅叶片通过增加厚角细胞厚度, 减少水分散失(
图2 ), 这与内蒙古短花针茅适应干旱环境的叶片解剖结构 (康萨如拉等,
2013 )相一致.为了适应干旱环境, 紫花针茅在改变保护组织的同时, 增加输水组织面积比例(导管直径、主脉导管面积/主脉维管束面积和维管束面积/叶横切面积), 以增强水分运输适应干旱环境(康萨如拉等,
2013 ; Buckley
et al .,
2015 ; 寇萌等,
2019 ). ...
... )相一致.为了适应干旱环境, 紫花针茅在改变保护组织的同时, 增加输水组织面积比例(导管直径、主脉导管面积/主脉维管束面积和维管束面积/叶横切面积), 以增强水分运输适应干旱环境(康萨如拉等,
2013 ; Buckley
et al .,
2015 ; 寇萌等,
2019 ). ...
短花针茅叶片解剖结构及与气候因子的关系
5
2013
... 植物与其生长的环境是统一的整体, 环境对植物的长期作用影响植物叶片解剖结构.在本研究中, 不同地理种群紫花针茅叶片解剖结构特征既存在相似性, 也存在较大变异性.角质层厚度在不同地理种群紫花针茅叶片结构特征中不存在显著差异, 这是因为角质层不仅可以反射强光, 还可以减少叶片水分蒸散, 有利于植物适应高原环境(李正理,
1981 ; Holmes & Keiller,
2002 ; 何涛等,
2007 ), 体现了不同地理种群紫花针茅叶片结构对高海拔生境的趋同适应进化.但是, 生境的不同导致紫花针茅叶片结构解剖特征存在较大差异, 例如紫花针茅叶片厚角细胞厚度随着干旱程度的增加而增加, 厚角细胞从安多紫花针茅叶片解剖结构中不存在到噶尔紫花针茅叶片中厚度逐渐增加, 而增加厚壁细胞厚度可减少水分蒸散损失(康萨如拉等,
2013 ).紫花针茅叶片解剖结构输水组织在不同区域也存在较大差异, 导管直径、主脉导管腔面积/主脉维管束面积和维管束面积/叶横切面积随着降水量的减少而增加, 这利于增加水分运输能力, 适应干旱环境(Cox & Moorc,
2010 ; 康萨如拉等,
2013 ; 寇萌等,
2019 ).因此, 不同地理种群紫花针茅叶片既有相似的解剖结构特征适应高海拔生境, 也有不同的叶片解剖结构适应区域生境. ...
... ; 康萨如拉等,
2013 ; 寇萌等,
2019 ).因此, 不同地理种群紫花针茅叶片既有相似的解剖结构特征适应高海拔生境, 也有不同的叶片解剖结构适应区域生境. ...
... 高寒草原优势种紫花针茅在青藏高原分布范围较广, 其叶片解剖结构与其生境相适应.Pearson相关性分析和聚类分析结果表明紫花针茅叶片解剖结构与气候因子密切相关(康萨如拉等,
2013 ), 例如五道梁和当雄2个地理种群距离较远, 但其叶片解剖结构比较相似.进一步的主成分和冗余分析结果表明, 与水分相关的气候因子对不同区域紫花针茅叶片解剖结构影响较大(胡选萍等,
2016 ), 即在噶尔、革吉和改则等干旱区域, 年蒸发量是影响厚角细胞厚度、导管直径和主脉导管腔面积/主脉维管束面积的主要因子, 而在相对湿润区域年降水量和生长季降水量等气候因子是影响保护组织(外表皮厚度和角质层厚度)的主要因子.紫花针茅叶片解剖结构未受到低温影响, 这可能是由于紫花针茅分布在高海拔区域, 相较于其他区域植物, 其已经适应了低温环境(Ma
et al .,
2012 ; 李全发等,
2013 ). ...
... 紫花针茅作为高寒草原的优势种, 其叶片解剖结构对高寒干旱环境具有较好的适应性.紫花针茅叶片外表皮普遍具有较厚角质层, 而在特别干旱区域(噶尔), 紫花针茅内表皮上也有角质层形成(
图2 H), 这有利于减少水分散失, 适应干旱环境(邓彦斌等,
1998 ; 蔡永立和宋永昌,
2001 ).同时紫花针茅叶片通过增加厚角细胞厚度, 减少水分散失(
图2 ), 这与内蒙古短花针茅适应干旱环境的叶片解剖结构 (康萨如拉等,
2013 )相一致.为了适应干旱环境, 紫花针茅在改变保护组织的同时, 增加输水组织面积比例(导管直径、主脉导管面积/主脉维管束面积和维管束面积/叶横切面积), 以增强水分运输适应干旱环境(康萨如拉等,
2013 ; Buckley
et al .,
2015 ; 寇萌等,
2019 ). ...
... )相一致.为了适应干旱环境, 紫花针茅在改变保护组织的同时, 增加输水组织面积比例(导管直径、主脉导管面积/主脉维管束面积和维管束面积/叶横切面积), 以增强水分运输适应干旱环境(康萨如拉等,
2013 ; Buckley
et al .,
2015 ; 寇萌等,
2019 ). ...
黄土丘陵沟壑区10种单子叶植物叶片解剖结构及环境适应性
2
2019
... 植物与其生长的环境是统一的整体, 环境对植物的长期作用影响植物叶片解剖结构.在本研究中, 不同地理种群紫花针茅叶片解剖结构特征既存在相似性, 也存在较大变异性.角质层厚度在不同地理种群紫花针茅叶片结构特征中不存在显著差异, 这是因为角质层不仅可以反射强光, 还可以减少叶片水分蒸散, 有利于植物适应高原环境(李正理,
1981 ; Holmes & Keiller,
2002 ; 何涛等,
2007 ), 体现了不同地理种群紫花针茅叶片结构对高海拔生境的趋同适应进化.但是, 生境的不同导致紫花针茅叶片结构解剖特征存在较大差异, 例如紫花针茅叶片厚角细胞厚度随着干旱程度的增加而增加, 厚角细胞从安多紫花针茅叶片解剖结构中不存在到噶尔紫花针茅叶片中厚度逐渐增加, 而增加厚壁细胞厚度可减少水分蒸散损失(康萨如拉等,
2013 ).紫花针茅叶片解剖结构输水组织在不同区域也存在较大差异, 导管直径、主脉导管腔面积/主脉维管束面积和维管束面积/叶横切面积随着降水量的减少而增加, 这利于增加水分运输能力, 适应干旱环境(Cox & Moorc,
2010 ; 康萨如拉等,
2013 ; 寇萌等,
2019 ).因此, 不同地理种群紫花针茅叶片既有相似的解剖结构特征适应高海拔生境, 也有不同的叶片解剖结构适应区域生境. ...
... 紫花针茅作为高寒草原的优势种, 其叶片解剖结构对高寒干旱环境具有较好的适应性.紫花针茅叶片外表皮普遍具有较厚角质层, 而在特别干旱区域(噶尔), 紫花针茅内表皮上也有角质层形成(
图2 H), 这有利于减少水分散失, 适应干旱环境(邓彦斌等,
1998 ; 蔡永立和宋永昌,
2001 ).同时紫花针茅叶片通过增加厚角细胞厚度, 减少水分散失(
图2 ), 这与内蒙古短花针茅适应干旱环境的叶片解剖结构 (康萨如拉等,
2013 )相一致.为了适应干旱环境, 紫花针茅在改变保护组织的同时, 增加输水组织面积比例(导管直径、主脉导管面积/主脉维管束面积和维管束面积/叶横切面积), 以增强水分运输适应干旱环境(康萨如拉等,
2013 ; Buckley
et al .,
2015 ; 寇萌等,
2019 ). ...
黄土丘陵沟壑区10种单子叶植物叶片解剖结构及环境适应性
2
2019
... 植物与其生长的环境是统一的整体, 环境对植物的长期作用影响植物叶片解剖结构.在本研究中, 不同地理种群紫花针茅叶片解剖结构特征既存在相似性, 也存在较大变异性.角质层厚度在不同地理种群紫花针茅叶片结构特征中不存在显著差异, 这是因为角质层不仅可以反射强光, 还可以减少叶片水分蒸散, 有利于植物适应高原环境(李正理,
1981 ; Holmes & Keiller,
2002 ; 何涛等,
2007 ), 体现了不同地理种群紫花针茅叶片结构对高海拔生境的趋同适应进化.但是, 生境的不同导致紫花针茅叶片结构解剖特征存在较大差异, 例如紫花针茅叶片厚角细胞厚度随着干旱程度的增加而增加, 厚角细胞从安多紫花针茅叶片解剖结构中不存在到噶尔紫花针茅叶片中厚度逐渐增加, 而增加厚壁细胞厚度可减少水分蒸散损失(康萨如拉等,
2013 ).紫花针茅叶片解剖结构输水组织在不同区域也存在较大差异, 导管直径、主脉导管腔面积/主脉维管束面积和维管束面积/叶横切面积随着降水量的减少而增加, 这利于增加水分运输能力, 适应干旱环境(Cox & Moorc,
2010 ; 康萨如拉等,
2013 ; 寇萌等,
2019 ).因此, 不同地理种群紫花针茅叶片既有相似的解剖结构特征适应高海拔生境, 也有不同的叶片解剖结构适应区域生境. ...
... 紫花针茅作为高寒草原的优势种, 其叶片解剖结构对高寒干旱环境具有较好的适应性.紫花针茅叶片外表皮普遍具有较厚角质层, 而在特别干旱区域(噶尔), 紫花针茅内表皮上也有角质层形成(
图2 H), 这有利于减少水分散失, 适应干旱环境(邓彦斌等,
1998 ; 蔡永立和宋永昌,
2001 ).同时紫花针茅叶片通过增加厚角细胞厚度, 减少水分散失(
图2 ), 这与内蒙古短花针茅适应干旱环境的叶片解剖结构 (康萨如拉等,
2013 )相一致.为了适应干旱环境, 紫花针茅在改变保护组织的同时, 增加输水组织面积比例(导管直径、主脉导管面积/主脉维管束面积和维管束面积/叶横切面积), 以增强水分运输适应干旱环境(康萨如拉等,
2013 ; Buckley
et al .,
2015 ; 寇萌等,
2019 ). ...
青藏高原草地植物叶解剖特征
1
2013
... 高寒草原优势种紫花针茅在青藏高原分布范围较广, 其叶片解剖结构与其生境相适应.Pearson相关性分析和聚类分析结果表明紫花针茅叶片解剖结构与气候因子密切相关(康萨如拉等,
2013 ), 例如五道梁和当雄2个地理种群距离较远, 但其叶片解剖结构比较相似.进一步的主成分和冗余分析结果表明, 与水分相关的气候因子对不同区域紫花针茅叶片解剖结构影响较大(胡选萍等,
2016 ), 即在噶尔、革吉和改则等干旱区域, 年蒸发量是影响厚角细胞厚度、导管直径和主脉导管腔面积/主脉维管束面积的主要因子, 而在相对湿润区域年降水量和生长季降水量等气候因子是影响保护组织(外表皮厚度和角质层厚度)的主要因子.紫花针茅叶片解剖结构未受到低温影响, 这可能是由于紫花针茅分布在高海拔区域, 相较于其他区域植物, 其已经适应了低温环境(Ma
et al .,
2012 ; 李全发等,
2013 ). ...
青藏高原草地植物叶解剖特征
1
2013
... 高寒草原优势种紫花针茅在青藏高原分布范围较广, 其叶片解剖结构与其生境相适应.Pearson相关性分析和聚类分析结果表明紫花针茅叶片解剖结构与气候因子密切相关(康萨如拉等,
2013 ), 例如五道梁和当雄2个地理种群距离较远, 但其叶片解剖结构比较相似.进一步的主成分和冗余分析结果表明, 与水分相关的气候因子对不同区域紫花针茅叶片解剖结构影响较大(胡选萍等,
2016 ), 即在噶尔、革吉和改则等干旱区域, 年蒸发量是影响厚角细胞厚度、导管直径和主脉导管腔面积/主脉维管束面积的主要因子, 而在相对湿润区域年降水量和生长季降水量等气候因子是影响保护组织(外表皮厚度和角质层厚度)的主要因子.紫花针茅叶片解剖结构未受到低温影响, 这可能是由于紫花针茅分布在高海拔区域, 相较于其他区域植物, 其已经适应了低温环境(Ma
et al .,
2012 ; 李全发等,
2013 ). ...
不同居群紫花针茅响应干旱胁迫的生理和分子差异分析
1
2015
... 紫花针茅(
Stipa purpurea )作为高寒草原的优势种, 对维持群落稳定性具有重要作用(Wu & Wang,
2019 ).气候变暖导致青藏高原的西部地区变暖变干, 东部地区变暖变湿(谭春萍等,
2010 ; 陈德亮等,
2015 ), 进而改变青藏高原草地生态系统的空间格局, 高山嵩草(
Kobresia pygmaea )群落向紫花针茅群落退化(王谋等,
2005 ), 紫花针茅在青藏高原的分布范围随着温度升高将继续扩大(陈德亮等,
2015 ; Ma & Sun,
2018 ).紫花针茅分布范围增加说明其具有对干旱生境较强的适应能力, 然而其如何适应高寒干旱气候变化的研究还相对薄弱(胡建莹等,
2008 ; 胡梦瑶等,
2012 ; 李雄等,
2015 ; 王常顺和汪诗平,
2015 ).对不同区域紫花针茅叶片解剖结构的分析, 可以直接反映紫花针茅对青藏高原气候变化的适应策略.因此, 本研究通过对紫花针茅在青藏高原分布区域进行采样, 分析其叶片解剖结构特征变异规律及其如何对环境因子响应, 以期为阐明高寒草原植物对气候变化适应机制提供科学依据. ...
不同居群紫花针茅响应干旱胁迫的生理和分子差异分析
1
2015
... 紫花针茅(
Stipa purpurea )作为高寒草原的优势种, 对维持群落稳定性具有重要作用(Wu & Wang,
2019 ).气候变暖导致青藏高原的西部地区变暖变干, 东部地区变暖变湿(谭春萍等,
2010 ; 陈德亮等,
2015 ), 进而改变青藏高原草地生态系统的空间格局, 高山嵩草(
Kobresia pygmaea )群落向紫花针茅群落退化(王谋等,
2005 ), 紫花针茅在青藏高原的分布范围随着温度升高将继续扩大(陈德亮等,
2015 ; Ma & Sun,
2018 ).紫花针茅分布范围增加说明其具有对干旱生境较强的适应能力, 然而其如何适应高寒干旱气候变化的研究还相对薄弱(胡建莹等,
2008 ; 胡梦瑶等,
2012 ; 李雄等,
2015 ; 王常顺和汪诗平,
2015 ).对不同区域紫花针茅叶片解剖结构的分析, 可以直接反映紫花针茅对青藏高原气候变化的适应策略.因此, 本研究通过对紫花针茅在青藏高原分布区域进行采样, 分析其叶片解剖结构特征变异规律及其如何对环境因子响应, 以期为阐明高寒草原植物对气候变化适应机制提供科学依据. ...
旱生植物的形态和结构
1
1981
... 植物与其生长的环境是统一的整体, 环境对植物的长期作用影响植物叶片解剖结构.在本研究中, 不同地理种群紫花针茅叶片解剖结构特征既存在相似性, 也存在较大变异性.角质层厚度在不同地理种群紫花针茅叶片结构特征中不存在显著差异, 这是因为角质层不仅可以反射强光, 还可以减少叶片水分蒸散, 有利于植物适应高原环境(李正理,
1981 ; Holmes & Keiller,
2002 ; 何涛等,
2007 ), 体现了不同地理种群紫花针茅叶片结构对高海拔生境的趋同适应进化.但是, 生境的不同导致紫花针茅叶片结构解剖特征存在较大差异, 例如紫花针茅叶片厚角细胞厚度随着干旱程度的增加而增加, 厚角细胞从安多紫花针茅叶片解剖结构中不存在到噶尔紫花针茅叶片中厚度逐渐增加, 而增加厚壁细胞厚度可减少水分蒸散损失(康萨如拉等,
2013 ).紫花针茅叶片解剖结构输水组织在不同区域也存在较大差异, 导管直径、主脉导管腔面积/主脉维管束面积和维管束面积/叶横切面积随着降水量的减少而增加, 这利于增加水分运输能力, 适应干旱环境(Cox & Moorc,
2010 ; 康萨如拉等,
2013 ; 寇萌等,
2019 ).因此, 不同地理种群紫花针茅叶片既有相似的解剖结构特征适应高海拔生境, 也有不同的叶片解剖结构适应区域生境. ...
旱生植物的形态和结构
1
1981
... 植物与其生长的环境是统一的整体, 环境对植物的长期作用影响植物叶片解剖结构.在本研究中, 不同地理种群紫花针茅叶片解剖结构特征既存在相似性, 也存在较大变异性.角质层厚度在不同地理种群紫花针茅叶片结构特征中不存在显著差异, 这是因为角质层不仅可以反射强光, 还可以减少叶片水分蒸散, 有利于植物适应高原环境(李正理,
1981 ; Holmes & Keiller,
2002 ; 何涛等,
2007 ), 体现了不同地理种群紫花针茅叶片结构对高海拔生境的趋同适应进化.但是, 生境的不同导致紫花针茅叶片结构解剖特征存在较大差异, 例如紫花针茅叶片厚角细胞厚度随着干旱程度的增加而增加, 厚角细胞从安多紫花针茅叶片解剖结构中不存在到噶尔紫花针茅叶片中厚度逐渐增加, 而增加厚壁细胞厚度可减少水分蒸散损失(康萨如拉等,
2013 ).紫花针茅叶片解剖结构输水组织在不同区域也存在较大差异, 导管直径、主脉导管腔面积/主脉维管束面积和维管束面积/叶横切面积随着降水量的减少而增加, 这利于增加水分运输能力, 适应干旱环境(Cox & Moorc,
2010 ; 康萨如拉等,
2013 ; 寇萌等,
2019 ).因此, 不同地理种群紫花针茅叶片既有相似的解剖结构特征适应高海拔生境, 也有不同的叶片解剖结构适应区域生境. ...
Predicting the distribution of
Stipa purpurea across the Tibetan Plateau via the MaxEnt model
1
2018
... 紫花针茅(
Stipa purpurea )作为高寒草原的优势种, 对维持群落稳定性具有重要作用(Wu & Wang,
2019 ).气候变暖导致青藏高原的西部地区变暖变干, 东部地区变暖变湿(谭春萍等,
2010 ; 陈德亮等,
2015 ), 进而改变青藏高原草地生态系统的空间格局, 高山嵩草(
Kobresia pygmaea )群落向紫花针茅群落退化(王谋等,
2005 ), 紫花针茅在青藏高原的分布范围随着温度升高将继续扩大(陈德亮等,
2015 ; Ma & Sun,
2018 ).紫花针茅分布范围增加说明其具有对干旱生境较强的适应能力, 然而其如何适应高寒干旱气候变化的研究还相对薄弱(胡建莹等,
2008 ; 胡梦瑶等,
2012 ; 李雄等,
2015 ; 王常顺和汪诗平,
2015 ).对不同区域紫花针茅叶片解剖结构的分析, 可以直接反映紫花针茅对青藏高原气候变化的适应策略.因此, 本研究通过对紫花针茅在青藏高原分布区域进行采样, 分析其叶片解剖结构特征变异规律及其如何对环境因子响应, 以期为阐明高寒草原植物对气候变化适应机制提供科学依据. ...
Comparative analyses of leaf anatomy of dicotyledonous species in Tibetan and Inner Mongolian grasslands
1
2012
... 高寒草原优势种紫花针茅在青藏高原分布范围较广, 其叶片解剖结构与其生境相适应.Pearson相关性分析和聚类分析结果表明紫花针茅叶片解剖结构与气候因子密切相关(康萨如拉等,
2013 ), 例如五道梁和当雄2个地理种群距离较远, 但其叶片解剖结构比较相似.进一步的主成分和冗余分析结果表明, 与水分相关的气候因子对不同区域紫花针茅叶片解剖结构影响较大(胡选萍等,
2016 ), 即在噶尔、革吉和改则等干旱区域, 年蒸发量是影响厚角细胞厚度、导管直径和主脉导管腔面积/主脉维管束面积的主要因子, 而在相对湿润区域年降水量和生长季降水量等气候因子是影响保护组织(外表皮厚度和角质层厚度)的主要因子.紫花针茅叶片解剖结构未受到低温影响, 这可能是由于紫花针茅分布在高海拔区域, 相较于其他区域植物, 其已经适应了低温环境(Ma
et al .,
2012 ; 李全发等,
2013 ). ...
地理生态学的干燥度指数及其应用评述
1
2004
... 气象数据来自对全国气象台站观测数据插值.气候指标包括: 年降水量、年平均气温、生长季平均气温、生长季降水量、7月平均气温、7月降水量、≥0 ℃积温、年蒸发量.湿润指数采用谢良尼诺夫公式根据中国实际情况修改进行计算(孟猛等,
2004 ).干旱半干旱区与半湿润区按照胡梦瑶等(
2012 )标准划分. ...
地理生态学的干燥度指数及其应用评述
1
2004
... 气象数据来自对全国气象台站观测数据插值.气候指标包括: 年降水量、年平均气温、生长季平均气温、生长季降水量、7月平均气温、7月降水量、≥0 ℃积温、年蒸发量.湿润指数采用谢良尼诺夫公式根据中国实际情况修改进行计算(孟猛等,
2004 ).干旱半干旱区与半湿润区按照胡梦瑶等(
2012 )标准划分. ...
青藏高原不同海拔3种菊科植物时片结构变化及其生态适应性
1
2016
... 叶片解剖结构特征是植物长期适应环境以及自身进化形成的, 体现了植物对环境的适应策略(Hetherington & Woodward,
2003 ), 而且叶片解剖结构在不同区域的变异特征可以直接反映植物对不同生长环境的适应策略(Wright
et al .,
2001 ; 钟悦鸣等,
2017 ).例如中间锦鸡儿(
Caragana davazamcii )随着平均年降水量的减少, 角质层厚度有所增加, 栅栏组织分化逐渐加强, 叶脉的机械组织分化逐渐加强(杨明博,
2007 ); 长梗扁桃(
Amygdalus pedunculata )生长在干旱地区的叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的叶片栅栏组织排列疏松, 叶厚度较薄(郭改改等,
2013 ); 青藏高原的菊科植物叶片厚度、上下表皮厚度、上下角质层厚度、栅栏细胞系数均随海拔升高而增加(孙会婷等,
2016 ); 蒙古莸(
Caryopteris mongholica )叶片解剖结构随着年平均气温升高, 叶厚度、栅栏组织厚度呈增大趋势(纪若璇等,
2020 ).因此, 叶片解剖结构可以体现植物对环境因子的适应性进化, 开展不同区域植物叶片解剖结构变异特征的研究, 对阐明植物适应气候变化策略具有重要意义. ...
青藏高原不同海拔3种菊科植物时片结构变化及其生态适应性
1
2016
... 叶片解剖结构特征是植物长期适应环境以及自身进化形成的, 体现了植物对环境的适应策略(Hetherington & Woodward,
2003 ), 而且叶片解剖结构在不同区域的变异特征可以直接反映植物对不同生长环境的适应策略(Wright
et al .,
2001 ; 钟悦鸣等,
2017 ).例如中间锦鸡儿(
Caragana davazamcii )随着平均年降水量的减少, 角质层厚度有所增加, 栅栏组织分化逐渐加强, 叶脉的机械组织分化逐渐加强(杨明博,
2007 ); 长梗扁桃(
Amygdalus pedunculata )生长在干旱地区的叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的叶片栅栏组织排列疏松, 叶厚度较薄(郭改改等,
2013 ); 青藏高原的菊科植物叶片厚度、上下表皮厚度、上下角质层厚度、栅栏细胞系数均随海拔升高而增加(孙会婷等,
2016 ); 蒙古莸(
Caryopteris mongholica )叶片解剖结构随着年平均气温升高, 叶厚度、栅栏组织厚度呈增大趋势(纪若璇等,
2020 ).因此, 叶片解剖结构可以体现植物对环境因子的适应性进化, 开展不同区域植物叶片解剖结构变异特征的研究, 对阐明植物适应气候变化策略具有重要意义. ...
1971-2007年青藏高原南部气候变化特征分析
1
2010
... 紫花针茅(
Stipa purpurea )作为高寒草原的优势种, 对维持群落稳定性具有重要作用(Wu & Wang,
2019 ).气候变暖导致青藏高原的西部地区变暖变干, 东部地区变暖变湿(谭春萍等,
2010 ; 陈德亮等,
2015 ), 进而改变青藏高原草地生态系统的空间格局, 高山嵩草(
Kobresia pygmaea )群落向紫花针茅群落退化(王谋等,
2005 ), 紫花针茅在青藏高原的分布范围随着温度升高将继续扩大(陈德亮等,
2015 ; Ma & Sun,
2018 ).紫花针茅分布范围增加说明其具有对干旱生境较强的适应能力, 然而其如何适应高寒干旱气候变化的研究还相对薄弱(胡建莹等,
2008 ; 胡梦瑶等,
2012 ; 李雄等,
2015 ; 王常顺和汪诗平,
2015 ).对不同区域紫花针茅叶片解剖结构的分析, 可以直接反映紫花针茅对青藏高原气候变化的适应策略.因此, 本研究通过对紫花针茅在青藏高原分布区域进行采样, 分析其叶片解剖结构特征变异规律及其如何对环境因子响应, 以期为阐明高寒草原植物对气候变化适应机制提供科学依据. ...
1971-2007年青藏高原南部气候变化特征分析
1
2010
... 紫花针茅(
Stipa purpurea )作为高寒草原的优势种, 对维持群落稳定性具有重要作用(Wu & Wang,
2019 ).气候变暖导致青藏高原的西部地区变暖变干, 东部地区变暖变湿(谭春萍等,
2010 ; 陈德亮等,
2015 ), 进而改变青藏高原草地生态系统的空间格局, 高山嵩草(
Kobresia pygmaea )群落向紫花针茅群落退化(王谋等,
2005 ), 紫花针茅在青藏高原的分布范围随着温度升高将继续扩大(陈德亮等,
2015 ; Ma & Sun,
2018 ).紫花针茅分布范围增加说明其具有对干旱生境较强的适应能力, 然而其如何适应高寒干旱气候变化的研究还相对薄弱(胡建莹等,
2008 ; 胡梦瑶等,
2012 ; 李雄等,
2015 ; 王常顺和汪诗平,
2015 ).对不同区域紫花针茅叶片解剖结构的分析, 可以直接反映紫花针茅对青藏高原气候变化的适应策略.因此, 本研究通过对紫花针茅在青藏高原分布区域进行采样, 分析其叶片解剖结构特征变异规律及其如何对环境因子响应, 以期为阐明高寒草原植物对气候变化适应机制提供科学依据. ...
植物叶片性状对气候变化的响应研究进展
1
2015
... 紫花针茅(
Stipa purpurea )作为高寒草原的优势种, 对维持群落稳定性具有重要作用(Wu & Wang,
2019 ).气候变暖导致青藏高原的西部地区变暖变干, 东部地区变暖变湿(谭春萍等,
2010 ; 陈德亮等,
2015 ), 进而改变青藏高原草地生态系统的空间格局, 高山嵩草(
Kobresia pygmaea )群落向紫花针茅群落退化(王谋等,
2005 ), 紫花针茅在青藏高原的分布范围随着温度升高将继续扩大(陈德亮等,
2015 ; Ma & Sun,
2018 ).紫花针茅分布范围增加说明其具有对干旱生境较强的适应能力, 然而其如何适应高寒干旱气候变化的研究还相对薄弱(胡建莹等,
2008 ; 胡梦瑶等,
2012 ; 李雄等,
2015 ; 王常顺和汪诗平,
2015 ).对不同区域紫花针茅叶片解剖结构的分析, 可以直接反映紫花针茅对青藏高原气候变化的适应策略.因此, 本研究通过对紫花针茅在青藏高原分布区域进行采样, 分析其叶片解剖结构特征变异规律及其如何对环境因子响应, 以期为阐明高寒草原植物对气候变化适应机制提供科学依据. ...
植物叶片性状对气候变化的响应研究进展
1
2015
... 紫花针茅(
Stipa purpurea )作为高寒草原的优势种, 对维持群落稳定性具有重要作用(Wu & Wang,
2019 ).气候变暖导致青藏高原的西部地区变暖变干, 东部地区变暖变湿(谭春萍等,
2010 ; 陈德亮等,
2015 ), 进而改变青藏高原草地生态系统的空间格局, 高山嵩草(
Kobresia pygmaea )群落向紫花针茅群落退化(王谋等,
2005 ), 紫花针茅在青藏高原的分布范围随着温度升高将继续扩大(陈德亮等,
2015 ; Ma & Sun,
2018 ).紫花针茅分布范围增加说明其具有对干旱生境较强的适应能力, 然而其如何适应高寒干旱气候变化的研究还相对薄弱(胡建莹等,
2008 ; 胡梦瑶等,
2012 ; 李雄等,
2015 ; 王常顺和汪诗平,
2015 ).对不同区域紫花针茅叶片解剖结构的分析, 可以直接反映紫花针茅对青藏高原气候变化的适应策略.因此, 本研究通过对紫花针茅在青藏高原分布区域进行采样, 分析其叶片解剖结构特征变异规律及其如何对环境因子响应, 以期为阐明高寒草原植物对气候变化适应机制提供科学依据. ...
气候变暖对青藏高原腹地高寒植被的影响
1
2005
... 紫花针茅(
Stipa purpurea )作为高寒草原的优势种, 对维持群落稳定性具有重要作用(Wu & Wang,
2019 ).气候变暖导致青藏高原的西部地区变暖变干, 东部地区变暖变湿(谭春萍等,
2010 ; 陈德亮等,
2015 ), 进而改变青藏高原草地生态系统的空间格局, 高山嵩草(
Kobresia pygmaea )群落向紫花针茅群落退化(王谋等,
2005 ), 紫花针茅在青藏高原的分布范围随着温度升高将继续扩大(陈德亮等,
2015 ; Ma & Sun,
2018 ).紫花针茅分布范围增加说明其具有对干旱生境较强的适应能力, 然而其如何适应高寒干旱气候变化的研究还相对薄弱(胡建莹等,
2008 ; 胡梦瑶等,
2012 ; 李雄等,
2015 ; 王常顺和汪诗平,
2015 ).对不同区域紫花针茅叶片解剖结构的分析, 可以直接反映紫花针茅对青藏高原气候变化的适应策略.因此, 本研究通过对紫花针茅在青藏高原分布区域进行采样, 分析其叶片解剖结构特征变异规律及其如何对环境因子响应, 以期为阐明高寒草原植物对气候变化适应机制提供科学依据. ...
气候变暖对青藏高原腹地高寒植被的影响
1
2005
... 紫花针茅(
Stipa purpurea )作为高寒草原的优势种, 对维持群落稳定性具有重要作用(Wu & Wang,
2019 ).气候变暖导致青藏高原的西部地区变暖变干, 东部地区变暖变湿(谭春萍等,
2010 ; 陈德亮等,
2015 ), 进而改变青藏高原草地生态系统的空间格局, 高山嵩草(
Kobresia pygmaea )群落向紫花针茅群落退化(王谋等,
2005 ), 紫花针茅在青藏高原的分布范围随着温度升高将继续扩大(陈德亮等,
2015 ; Ma & Sun,
2018 ).紫花针茅分布范围增加说明其具有对干旱生境较强的适应能力, 然而其如何适应高寒干旱气候变化的研究还相对薄弱(胡建莹等,
2008 ; 胡梦瑶等,
2012 ; 李雄等,
2015 ; 王常顺和汪诗平,
2015 ).对不同区域紫花针茅叶片解剖结构的分析, 可以直接反映紫花针茅对青藏高原气候变化的适应策略.因此, 本研究通过对紫花针茅在青藏高原分布区域进行采样, 分析其叶片解剖结构特征变异规律及其如何对环境因子响应, 以期为阐明高寒草原植物对气候变化适应机制提供科学依据. ...
Strategy shifts in leaf physiology, structure and nutrient content between species of high- and 1ow-rainfall and high- and low-nutrient habitats
1
2001
... 叶片解剖结构特征是植物长期适应环境以及自身进化形成的, 体现了植物对环境的适应策略(Hetherington & Woodward,
2003 ), 而且叶片解剖结构在不同区域的变异特征可以直接反映植物对不同生长环境的适应策略(Wright
et al .,
2001 ; 钟悦鸣等,
2017 ).例如中间锦鸡儿(
Caragana davazamcii )随着平均年降水量的减少, 角质层厚度有所增加, 栅栏组织分化逐渐加强, 叶脉的机械组织分化逐渐加强(杨明博,
2007 ); 长梗扁桃(
Amygdalus pedunculata )生长在干旱地区的叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的叶片栅栏组织排列疏松, 叶厚度较薄(郭改改等,
2013 ); 青藏高原的菊科植物叶片厚度、上下表皮厚度、上下角质层厚度、栅栏细胞系数均随海拔升高而增加(孙会婷等,
2016 ); 蒙古莸(
Caryopteris mongholica )叶片解剖结构随着年平均气温升高, 叶厚度、栅栏组织厚度呈增大趋势(纪若璇等,
2020 ).因此, 叶片解剖结构可以体现植物对环境因子的适应性进化, 开展不同区域植物叶片解剖结构变异特征的研究, 对阐明植物适应气候变化策略具有重要意义. ...
Temporal stability of aboveground net primary production in northern Tibet alpine steppe in response to nitrogen addition
1
2019
... 紫花针茅(
Stipa purpurea )作为高寒草原的优势种, 对维持群落稳定性具有重要作用(Wu & Wang,
2019 ).气候变暖导致青藏高原的西部地区变暖变干, 东部地区变暖变湿(谭春萍等,
2010 ; 陈德亮等,
2015 ), 进而改变青藏高原草地生态系统的空间格局, 高山嵩草(
Kobresia pygmaea )群落向紫花针茅群落退化(王谋等,
2005 ), 紫花针茅在青藏高原的分布范围随着温度升高将继续扩大(陈德亮等,
2015 ; Ma & Sun,
2018 ).紫花针茅分布范围增加说明其具有对干旱生境较强的适应能力, 然而其如何适应高寒干旱气候变化的研究还相对薄弱(胡建莹等,
2008 ; 胡梦瑶等,
2012 ; 李雄等,
2015 ; 王常顺和汪诗平,
2015 ).对不同区域紫花针茅叶片解剖结构的分析, 可以直接反映紫花针茅对青藏高原气候变化的适应策略.因此, 本研究通过对紫花针茅在青藏高原分布区域进行采样, 分析其叶片解剖结构特征变异规律及其如何对环境因子响应, 以期为阐明高寒草原植物对气候变化适应机制提供科学依据. ...
鄂尔多斯高原不同生境条件下中间锦鸡儿植物叶片表皮特征及遗传多样性变化分析
1
2007
... 叶片解剖结构特征是植物长期适应环境以及自身进化形成的, 体现了植物对环境的适应策略(Hetherington & Woodward,
2003 ), 而且叶片解剖结构在不同区域的变异特征可以直接反映植物对不同生长环境的适应策略(Wright
et al .,
2001 ; 钟悦鸣等,
2017 ).例如中间锦鸡儿(
Caragana davazamcii )随着平均年降水量的减少, 角质层厚度有所增加, 栅栏组织分化逐渐加强, 叶脉的机械组织分化逐渐加强(杨明博,
2007 ); 长梗扁桃(
Amygdalus pedunculata )生长在干旱地区的叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的叶片栅栏组织排列疏松, 叶厚度较薄(郭改改等,
2013 ); 青藏高原的菊科植物叶片厚度、上下表皮厚度、上下角质层厚度、栅栏细胞系数均随海拔升高而增加(孙会婷等,
2016 ); 蒙古莸(
Caryopteris mongholica )叶片解剖结构随着年平均气温升高, 叶厚度、栅栏组织厚度呈增大趋势(纪若璇等,
2020 ).因此, 叶片解剖结构可以体现植物对环境因子的适应性进化, 开展不同区域植物叶片解剖结构变异特征的研究, 对阐明植物适应气候变化策略具有重要意义. ...
鄂尔多斯高原不同生境条件下中间锦鸡儿植物叶片表皮特征及遗传多样性变化分析
1
2007
... 叶片解剖结构特征是植物长期适应环境以及自身进化形成的, 体现了植物对环境的适应策略(Hetherington & Woodward,
2003 ), 而且叶片解剖结构在不同区域的变异特征可以直接反映植物对不同生长环境的适应策略(Wright
et al .,
2001 ; 钟悦鸣等,
2017 ).例如中间锦鸡儿(
Caragana davazamcii )随着平均年降水量的减少, 角质层厚度有所增加, 栅栏组织分化逐渐加强, 叶脉的机械组织分化逐渐加强(杨明博,
2007 ); 长梗扁桃(
Amygdalus pedunculata )生长在干旱地区的叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的叶片栅栏组织排列疏松, 叶厚度较薄(郭改改等,
2013 ); 青藏高原的菊科植物叶片厚度、上下表皮厚度、上下角质层厚度、栅栏细胞系数均随海拔升高而增加(孙会婷等,
2016 ); 蒙古莸(
Caryopteris mongholica )叶片解剖结构随着年平均气温升高, 叶厚度、栅栏组织厚度呈增大趋势(纪若璇等,
2020 ).因此, 叶片解剖结构可以体现植物对环境因子的适应性进化, 开展不同区域植物叶片解剖结构变异特征的研究, 对阐明植物适应气候变化策略具有重要意义. ...
不同生境胡杨叶片解剖特征及其适应可塑性
1
2017
... 叶片解剖结构特征是植物长期适应环境以及自身进化形成的, 体现了植物对环境的适应策略(Hetherington & Woodward,
2003 ), 而且叶片解剖结构在不同区域的变异特征可以直接反映植物对不同生长环境的适应策略(Wright
et al .,
2001 ; 钟悦鸣等,
2017 ).例如中间锦鸡儿(
Caragana davazamcii )随着平均年降水量的减少, 角质层厚度有所增加, 栅栏组织分化逐渐加强, 叶脉的机械组织分化逐渐加强(杨明博,
2007 ); 长梗扁桃(
Amygdalus pedunculata )生长在干旱地区的叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的叶片栅栏组织排列疏松, 叶厚度较薄(郭改改等,
2013 ); 青藏高原的菊科植物叶片厚度、上下表皮厚度、上下角质层厚度、栅栏细胞系数均随海拔升高而增加(孙会婷等,
2016 ); 蒙古莸(
Caryopteris mongholica )叶片解剖结构随着年平均气温升高, 叶厚度、栅栏组织厚度呈增大趋势(纪若璇等,
2020 ).因此, 叶片解剖结构可以体现植物对环境因子的适应性进化, 开展不同区域植物叶片解剖结构变异特征的研究, 对阐明植物适应气候变化策略具有重要意义. ...
不同生境胡杨叶片解剖特征及其适应可塑性
1
2017
... 叶片解剖结构特征是植物长期适应环境以及自身进化形成的, 体现了植物对环境的适应策略(Hetherington & Woodward,
2003 ), 而且叶片解剖结构在不同区域的变异特征可以直接反映植物对不同生长环境的适应策略(Wright
et al .,
2001 ; 钟悦鸣等,
2017 ).例如中间锦鸡儿(
Caragana davazamcii )随着平均年降水量的减少, 角质层厚度有所增加, 栅栏组织分化逐渐加强, 叶脉的机械组织分化逐渐加强(杨明博,
2007 ); 长梗扁桃(
Amygdalus pedunculata )生长在干旱地区的叶片较厚, 茸毛、蜡质等覆盖物增加, 而生长于年降水量充沛地区的叶片栅栏组织排列疏松, 叶厚度较薄(郭改改等,
2013 ); 青藏高原的菊科植物叶片厚度、上下表皮厚度、上下角质层厚度、栅栏细胞系数均随海拔升高而增加(孙会婷等,
2016 ); 蒙古莸(
Caryopteris mongholica )叶片解剖结构随着年平均气温升高, 叶厚度、栅栏组织厚度呈增大趋势(纪若璇等,
2020 ).因此, 叶片解剖结构可以体现植物对环境因子的适应性进化, 开展不同区域植物叶片解剖结构变异特征的研究, 对阐明植物适应气候变化策略具有重要意义. ...
 ,*
,*

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT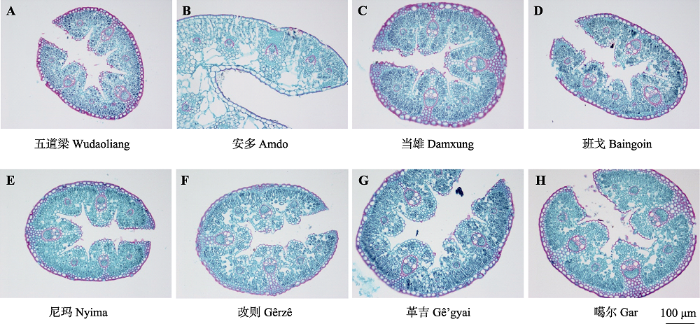 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT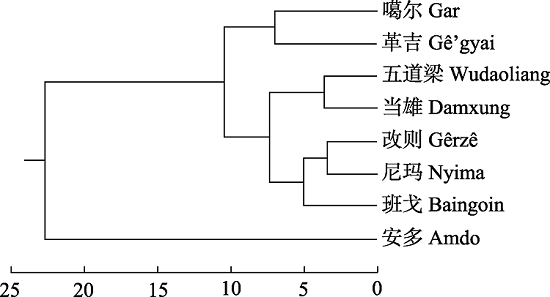 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT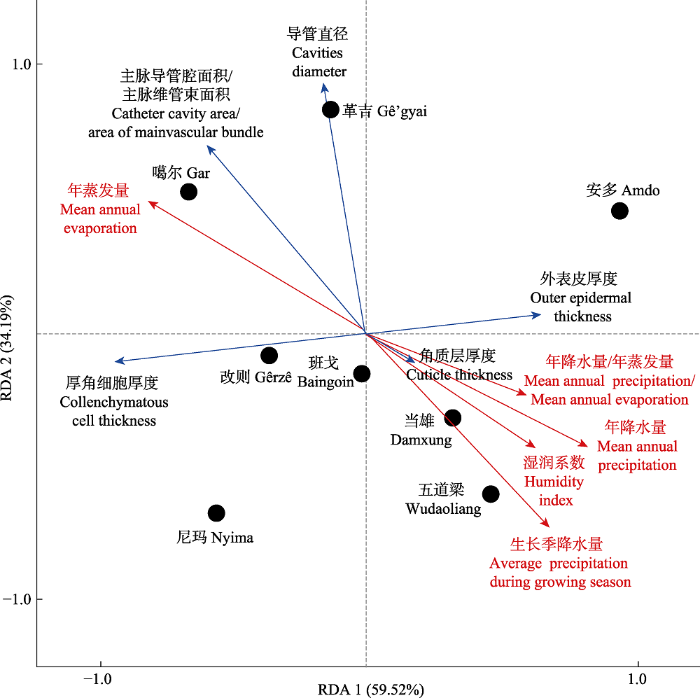 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}