Abstract Aims The mixed broadleaved-Korean pine (Pinus koraiensis) forest is a zonal vegetation type in Northeast China. Clarifying the influencing factors of productivity is of great significance for understanding the maintenance mechanism of productivity in temperate forests. Methods The study was conducted based on survey data of a 9 hm2 permanent plot in a typical mixed broadleaved- Korean pine forest in the Xiaoxing’an Mountains. Census data of 2005 and 2015 for all individuals with diameter at breast height >6.5 cm within each 30 m × 30 m sub-plot were used to calculate canopy structural complexity, species diversity and stand basal area. We analyzed the direct and indirect factors affecting forest productivity using structural equation model and data on topography and soil physicochemical properties in each sub-plot. Important findings Both the canopy structural complexity and the species diversity were positively related to productivity, with canopy structural complexity having a significantly greater effect than species diversity. The effects of canopy structural complexity on productivity consisted of two attributes: the vertical stratification and the plasticity. The vertical stratification played a significant role in driving the changes in forest productivity, while plasticity had no significant effect. The stand basal area was positively related to productivity, but with less effect than the canopy structural complexity. Both the canopy structural complexity and the species diversity had indirect effects on productivity via stand basal area. However, the environmental factor representative of slope and soil total phosphorus played differential roles on productivity for varying canopy structral complexity, and it was significantly and negatively correlated with productivity after removing the effects of canopy vertical stratification. In short, in a typical mixed broadleaved-Korean pine forest, the canopy structural complexity is more crucial for explaining the diversity-productivity relationship than species diversity, and the roles of other biotic and abiotic factors in productivity should not be ignored. Keywords:biodiversity;canopy structural complexity;species evenness;productivity;mixed broadleaved- Korean pine forest
PDF (1344KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 哈努拉?塔斯肯, 蔡慧颖, 金光泽. 树冠结构对典型阔叶红松林生产力的影响. 植物生态学报, 2021, 45(1): 38-50. DOI: 10.17521/cjpe.2020.0176 Hanula TASIKEN, CAI Hui-Ying, JIN Guang-Ze. Effects of canopy structure on productivity in a typical mixed broadleaved-Korean pine forest. Chinese Journal of Plant Ecology, 2021, 45(1): 38-50. DOI: 10.17521/cjpe.2020.0176
生物多样性-生产力关系(BPR)是理解全球物种灭绝危机及其对生态系统功能影响的基础, 对于准确评估和有效保护生物多样性至关重要(Zhang et al., 2012; Liang et al., 2016; Yuan et al., 2018)。早期的BPR研究多集中于分析物种多样性的作用, 对生产力的解释和预测能力较低(Grace et al., 2016; Zeller et al., 2018)。近年来, 随着对生物多样性不同维度(物种、功能、谱系和结构多样性)认识的加深和扩展, 生态学家们基于树冠结构在垂直和水平方向上的差异提出林分结构多样性的衍生多样性——树冠结构复杂性。树冠结构复杂性作为综合评价森林冠层对光照时空分配利用情况的重要参数, 可以较全面地描述森林的林分结构特征(Sapijanskaset al., 2014), 其值越高说明林内个体对立地空间的利用率越高, 对林分中光照等资源的利用更全面, 进而具有更高的森林生产力(Ali et al., 2019)。
树冠结构复杂性可分两部分来分别解释树冠结构在垂直与水平方向上的作用(Pretzsch, 2014; Jucker et al., 2015)。一是树冠垂直分层: 当树木的叶片分布在冠层的不同高度剖面时, 森林冠层能够最大限度地截留光线, 从而对森林生产力产生积极的促进作用(Williams et al., 2017)。共存树种对光照的分层截留以及对郁闭环境的生理适应, 均有利于树冠垂直分层(Sterba et al., 2019)。二是树冠可塑性: 树木的大小和形状在生长过程中具有较强的可塑性, 可以形成镶嵌的树冠结构, 覆盖大部分或所有的树冠空间(Purves et al., 2007)。树冠横向发展, 如展叶、分枝和树冠形状的变化, 均有助于减少林分中激烈的种内、种间竞争, 进而促进森林生态系统功能优化(Jucker et al., 2015; Fotis et al., 2018)。树冠结构复杂性被系统地提出之后, 该研究领域的成果迅速增加, 大量实验证明树冠结构复杂性可以提高森林生态系统的生产力, 但是树冠垂直分层和树冠可塑性二者的作用强度和内在作用机制还有争议(Pretzsch, 2014; Fahey et al., 2015)。部分****认为多林层林分具有更高的生产潜能, 林冠垂直分层是诸多森林生物量积累的重要驱动因素(Hardiman et al., 2013; D?nescuet al., 2016)。然而, 这与Bayer等(2013)和Jucker等(2015)的研究相悖, 他们通过分离垂直和水平方向上的树冠结构复杂性发现, 可塑性较强的树冠结构才是影响生产力的重要因素, 树冠垂直分层反而没有明显作用。不同地区和林龄中树冠结构复杂性对生产力的贡献相异(Bohn & Huth, 2017), 因此仍需更多系统的研究来阐明树冠垂直分层和树冠可塑性的相对重要性。此外, 林分中较高的物种丰富度, 往往会形成较高的树冠结构复杂性, 从而间接地促进森林生产力。然而, 物种多样性对生产力的这种间接影响尚未得到完全证实(Forrester, 2014; Sapijanskas et al., 2014)。
通常, 树木生长是多种因素共同作用的结果, 在研究多样性-生产力关系时考虑气候、地形、土壤养分和群落结构等多个生物和非生物因素的作用十分必要, 有利于更好地理解多驱动因素对天然林多样性和生态系统功能的影响(Ali et al., 2016; Yuan et al., 2018)。区域尺度上, 林分胸高断面积是影响生产力的重要因子。多数研究以单位面积内所有个体的胸高断面积之和作为木本植物地上生物量的衡量指标, 探讨其与生产力的相关关系(郭屹立等, 2016)。林分胸高断面积越大, 森林生产潜力越高(Muth & Bazzaz, 2003)。此外, 物种多样性和树冠结构也会通过影响林分胸高断面积, 间接地驱动森林生产力(D?nescuet al., 2016)。地形与土壤物理化学属性互相联系、互相制约, 是导致不同尺度环境异质性的重要因素, 地形的变化可以引起水、肥、气、热的重新分配, 进而影响植物的分布和生长(Williams et al., 2017)。如通常在地势平坦地区, 土壤有机质和土壤水分较高, 树木生长速度较快, 对森林生产力存在正向的影响(Zheng et al., 2019); 但较高的土壤养分可利用性, 也会加剧种内、种间的竞争, 导致树木较高的死亡率和周转率(Quesada et al., 2012)。综上, 生物因素和非生物因素对生产力的影响仍需要进一步验证。
本文参考有关树冠结构复杂性的研究, 采用胸径>6.5 cm的个体进行计算, 即舍弃复查期间容易消失的小径级树木, 保证目标树处于冠层, 使结果更具生态学意义(Faheyet al., 2015; Jucker et al., 2015)。本研究预分析了不同尺度下树冠结构复杂性和物种多样性与生产力关系(附录I), 其中30 m × 30 m尺度下模型赤池信息准则(AIC)最低, 故主要研究30 m × 30 m空间尺度下的多样性-生产力关系。
Supplement I 附录I 附录I不同尺度下生产力最优结构方程模型拟合结果 Supplement IResults of the best-fit structural equation models (SEMs) of productivity at varying scales
尺度 Scale
比较拟合指数 CFI
标准均方根残差 SRMR
渐进残差均方和平方根 RMSEA
赤池信息准则 AIC
p
10 m × 10 m
1
0.006
0
12 400.938
0.653
20 m × 20 m
0.99
0.026
0.065
3 105.839
0.118
30 m × 30 m
1
0.020
0
1 308.250
0.720
AIC,akaike information criterion; CFI,comparative fit index; RMSEA, root mean square error of approximation; SRMR, standardized root mean square residual.
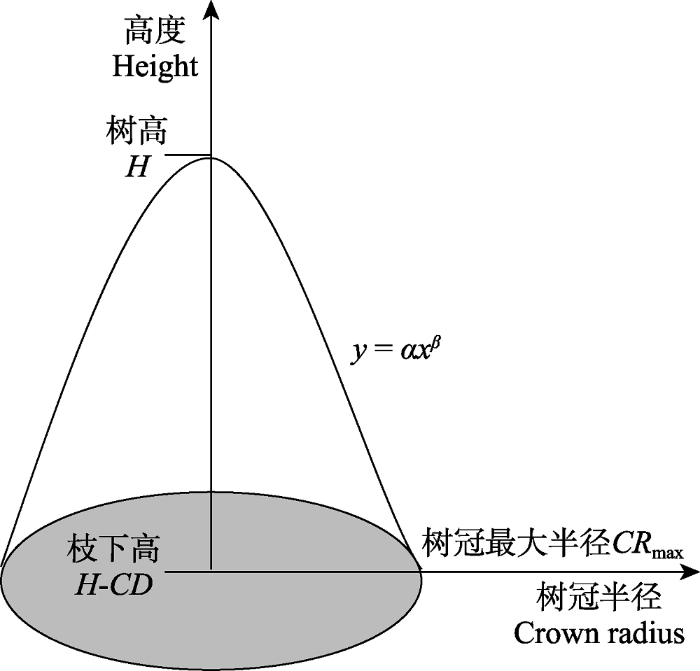
Fig. 1Schematic illustration of crown volume of a tree. CRmax, the maximum of radius of crown; H, tree height; H-CD, under branch height. The shape of the crown is determined by equation y = αx β, and β is the shape parameter which determines the curvature of the crown.
1.3.3 树冠结构复杂性的计算
树冠结构复杂性是由冠层镶嵌指数测算, 冠层镶嵌指数被定义为树木潜在的可利用地上空间的比例, 而这些空间实际由冠层占据(Jucker et al., 2015)。
Table 2 表2 表2典型阔叶红松林样地地形和土壤物理化学性质状况统计表 Table 2Summary of topographic factors and soil physical and chemical properties in a typical mixed broadleaved-Korean pine forest plot
环境因子 Environmental factor
平均值 Mean
标准差 SD
变异系数 CV
范围 Range
海拔 Elevation (m)
463.22
18.07
0.04
427.93-499.07
坡度 Slope (°)
15.30
4.67
0.31
8.10-33.40
坡向 Aspect (sin)
0.02
0.72
36.00
-0.997-0.994
坡向 Aspect (cos)
-0.07
0.70
-10.00
-0.999-0.999
凹凸度 Convex
0.06
1.05
17.50
-1.55-4.45
土壤速效磷含量 Soil available phosphorus content (mg·kg-1)
8.83
6.04
0.68
3.53-43.11
土壤速效钾含量 Soil available potassium content (mg·kg-1)
332.61
52.69
0.16
200.87-485.34
土壤速效氮含量 Soil available nitrogen content (mg·kg-1)
1 103.60
147.95
0.13
666.56-1 450.17
土壤有机碳含量 Soil organic carbon content (g·kg-1)
75.75
29.71
0.39
37.63-186.29
土壤全氮含量 Soil total nitrogen content (g·kg-1)
8.40
1.46
0.17
5.22-11.70
土壤全磷含量 Soil total phosphorus content (g·kg-1)
0.82
0.13
0.16
0.44-1.13
土壤pH Soil pH
5.77
0.15
0.03
5.47-6.19
土壤容重 Soil bulk density (g·cm-3)
0.62
0.09
0.15
0.43-0.85
体积含水率 Soil volumetric moisture content (%)
30.35
5.37
0.18
15.44-44.06
质量含水率 Soil mass moisture content (g·g-1)
0.92
0.22
0.24
0.55-1.40
CV, coefficient of variation;SD, standard deviation.
Fig. 2A conceptual model of hypothesized causal pathways linking the environmental factor, species diversity, canopy structural complexity, stand basal area, and productivity in a mixed broadleaved-Korean pine forest. The environmental factor is a latent variable consisting of soil physical and chemical properties and topography.
Supplement II 附录II 附录II典型阔叶红松林与生产力建立多重回归模型中各变量的相对重要性结果 Supplement IIResults of the relative importance of each variable in multiple regression model with productivity in typical mixed broadleaved-Korean pine forest
Table 3 表3 表3典型阔叶红松林最优结构方程模型检验多样性与生产力关系的结果 Table 3Results of the best-fit structural equation models (SEMs) testing the relationships between forest productivity and diversity in a typical mixed broadleaved-Korean pine forest
模型 Model
CFI
SRMR
RMSEA
AIC
p
树冠结构复杂性 Canopy structural complexity
1
0.020
0
1 308.25
0.720
树冠垂直分层 Canopy vertical stratification
1
0.019
0
1 295.63
0.568
树冠可塑性 Canopy plasticity
1
0.021
0
1 346.06
0.755
AIC, 赤池信息准则; CFI, 比较拟合指数; p,显著性概率值; RMSEA, 渐进残差均方和平方根; SRMR,标准均方根残差。 AIC, akaike information criterion; CFI,comparative fit index; p,probability of significance; RMSEA, root mean square error of approximation; SRMR, standardized root mean square residual.
新窗口打开|下载原图ZIP|生成PPT 图3阔叶红松林树冠结构复杂性、均匀度指数、林分胸高断面积、环境因素(土壤全磷(P)和坡度结合的潜变量)与生产力之间的结构方程模型。A, 考虑树冠结构复杂性的模型。B, 考虑树冠垂直分层的模型。C, 考虑树冠可塑性的模型。图中实线表示作用路径显著, 虚线表示作用路径不显著。箭头旁边的值是每个路径的标准化回归系数和显著性。R2表示由组合自变量解释的因变量的总变化。*, p < 0.05; **, p < 0.01; ***, p < 0.001。
Fig. 3Structural equation models (SEMs) for linking multivariate relationships among canopy structural complexity, evenness index, stand basal area, environmental factor (a latent variable of soil total phosphorus (P) and slope), and productivity in typical mixed broadleaved-Korean pine forest. A, Considering canopy structural complexity. B, Considering canopy vertical stratification. C, Considering canopy plasticity in the structural equation model. Solid lines indicate significant paths, and dash lines indicate insignificant paths. Standardized regression coefficients and significance are shown next to the arrow for each path. R2 indicates the total variation in a dependent variable explained by the combined independent variables. *, p < 0.05; **, p < 0.01; ***, p < 0.001.
Table 4 表4 表4均匀度指数、树冠结构复杂性、林分胸高断面积和环境因素对典型阔叶红松林生产力的直接、间接和总效应 Table 4Direct, indirect, and total standardized effects of evenness index (EI), canopy structural complexity (CSC), stand basal area (SBA) and environmental factor on productivity in a typical mixed broadleaved-Korean pine forest
Table 5 表5 表5均匀度指数、树冠垂直分层、林分胸高断面积和环境因素对典型阔叶红松林生产力的直接、间接和总效应 Table 5Direct, indirect, and total standardized effects of evenness index (EI), canopy vertical stratification (CVS), stand basal area (SBA) and environmental factor on productivity in a typical mixed broadleaved-Korean pine forest
新窗口打开|下载原图ZIP|生成PPT 图4典型阔叶红松林生产力(对数转换值)与各个解释变量的双变量关系。p < 0.05表示拟合关系显著; p > 0.05时拟合关系不显著, 无回归直线。
Fig. 4Bivariate relationships between productivity (ln transformed) and explanatory variables in a typical mixed broadleaved- Korean pine forest. Fitted regression is significant at p < 0.05 and the relationships without fitted lines are insignificant at p > 0.05.
Table 6 表6 表6均匀度指数、树冠可塑性、林分胸高断面积和环境因素对典型阔叶红松林生产力的直接、间接和总效应 Table 6Direct, indirect, and total standardized effects of evenness index (EI), canopy plasticity (CP), stand basal area (SBA) and environmental factors on productivity in a typical mixed broadleaved-Korean pine forest
AliA, LinSL, HeJK, KongFM, YuJH, JiangHS (2019). Tree crown complementarity links positive functional diversity and aboveground biomass along large-scale ecological gradients in tropical forests Science of the Total Environment, 656,45-54. DOI:10.1016/j.scitotenv.2018.11.342URL [本文引用: 2]
AliA, YanER, ChenHYH, ChangSX, ZhaoYT, YangXD, XuMS (2016). Stand structural diversity rather than species diversity enhances aboveground carbon storage in secondary subtropical forests in Eastern China Biogeosciences, 13,4627-4635. DOI:10.5194/bg-13-4627-2016URL [本文引用: 2]
BayerD, SeifertS, PretzschH (2013). Structural crown properties of Norway spruce (Picea abies L. Karst.) and European beech (Fagus sylvatica L.) in mixed versus pure stands revealed by terrestrial laser scanning Trees, 27, 1035- 1047. [本文引用: 2]
BohnFJ, HuthA (2017). The importance of forest structure to biodiversity-productivity relationships Royal Society Open Science, 4,160521. DOI: 10.1098/rsos.160521. DOI:10.1098/rsos.160521URL [本文引用: 2]
ChaveJ, ConditR, LaoS, CaspersenJP, FosterRB, HubbellSP (2003). Spatial and temporal variation of biomass in a tropical forest: results from a large census plot in Panama Journal of Ecology, 91,240-252. DOI:10.1046/j.1365-2745.2003.00757.xURL [本文引用: 1]
ChenCG, ZhuJF (1989). A Handbook for Main Tree Species Biomass in Northeast China. China Forestry Press, Beijing. [本文引用: 1]
D?nescuA, AlbrechtAT, BauhusJ (2016). Structural diversity promotes productivity of mixed, uneven-aged forests in southwestern Germany Oecologia, 182,319-333. DOI:10.1007/s00442-016-3623-4PMID:27059713 [本文引用: 5] Forest diversity-productivity relationships have been intensively investigated in recent decades. However, few studies have considered the interplay between species and structural diversity in driving productivity. We analyzed these factors using data from 52 permanent plots in southwestern Germany with more than 53,000 repeated tree measurements. We used basal area increment as a proxy for productivity and hypothesized that: (1) structural diversity would increase tree and stand productivity, (2) diversity-productivity relationships would be weaker for species diversity than for structural diversity, and (3) species diversity would also indirectly impact stand productivity via changes in size structure. We measured diversity using distance-independent indices. We fitted separate linear mixed-effects models for fir, spruce and beech at the tree level, whereas at the stand level we pooled all available data. We tested our third hypothesis using structural equation modeling. Structural and species diversity acted as direct and independent drivers of stand productivity, with structural diversity being a slightly better predictor. Structural diversity, but not species diversity, had a significant, albeit asymmetric, effect on tree productivity. The functioning of structurally diverse, mixed forests is influenced by both structural and species diversity. These sources of trait diversity contribute to increased vertical stratification and crown plasticity, which in turn diminish competitive interferences and lead to more densely packed canopies per unit area. Our research highlights the positive effects of species diversity and structural diversity on forest productivity and ecosystem dynamics.
FaheyRT, FotisAT, WoodsKD (2015). Quantifying canopy complexity and effects on productivity and resilience in late-successional hemlock-hardwood forests Ecological Applications, 25,834-847. DOI:10.1890/14-1012.1URL [本文引用: 4]
ForresterDI (2014). The spatial and temporal dynamics of species interactions in mixed-species forests: from pattern to process Forest Ecology and Management, 312,282-292. DOI:10.1016/j.foreco.2013.10.003URL [本文引用: 1]
FotisAT, MorinTH, FaheyRT, HardimanBS, BohrerG, CurtisPS (2018). Forest structure in space and time: biotic and abiotic determinants of canopy complexity and their effects on net primary productivity Agricultural and Forest Meteorology, 250-251,181-191. [本文引用: 1]
GuoYL, WangB, XiangWS, DingT, LuSH, HuangFZ, WenSJ, LiDX, HeYL, LiXK (2016). Responses of spatial pattern of woody plants’ basal area to topographic factors in a tropical karst seasonal rainforest in Nonggang, Guangxi, southern China Biodiversity Science, 24,30-39. DOI:10.17520/biods.2015207URLPMID:fc0a8d36-df63-4e0f-a8f8-4819a4b8a13e [本文引用: 1]
HardimanBS, GoughCM, HalperinA, HofmeisterKL, NaveLE, BohrerG, CurtisPS (2013). Maintaining high rates of carbon storage in old forests: a mechanism linking canopy structure to forest function Forest Ecology and Management, 298,111-119. DOI:10.1016/j.foreco.2013.02.031URL [本文引用: 3]
HillebrandH, BennettDM, CadotteMW (2008). Consequences of dominance: a review of evenness effects on local and regional ecosystem processes Ecology, 89,1510-1520. PMID:18589516 [本文引用: 1] The composition of communities is strongly altered by anthropogenic manipulations of biogeochemical cycles, abiotic conditions, and trophic structure in all major ecosystems. Whereas the effects of species loss on ecosystem processes have received broad attention, the consequences of altered species dominance for emergent properties of communities and ecosystems are poorly investigated. Here we propose a framework guiding our understanding of how dominance affects species interactions within communities, processes within ecosystems, and dynamics on regional scales. Dominance (or the complementary term, evenness) reflects the distribution of traits in a community, which in turn affects the strength and sign of both intraspecifc and interspecific interactions. Consequently, dominance also mediates the effect of such interactions on species coexistence. We review the evidence for the fact that dominance directly affects ecosystem functions such as process rates via species identity (the dominant trait) and evenness (the frequency distribution of traits), and indirectly alters the relationship between process rates and species richness. Dominance also influences the temporal and spatial variability of aggregate community properties and compositional stability (invasibility). Finally, we propose that dominance affects regional species coexistence by altering metacommunity dynamics. Local dominance leads to high beta diversity, and rare species can persist because of source-sink dynamics, but anthropogenically induced environmental changes result in regional dominance and low beta diversity, reducing regional coexistence. Given the rapid anthropogenic alterations of dominance in many ecosystems and the strong implications of these changes, dominance should be considered explicitly in the analysis of consequences of altered biodiversity.
HowardAL (2013). Handbook of structural equation modeling Structural Equation Modeling: a Multidisciplinary Journal, 20,354-360. [本文引用: 1]
JuckerT, BouriaudO, CoomesDA (2015). Crown plasticity enables trees to optimize canopy packing in mixed-species forests Functional Ecology, 29,1078-1086. DOI:10.1111/fec.2015.29.issue-8URL [本文引用: 6]
KluteA (1986). Methods of Soil Analysis: Part 1—Physical and Mineralogical Methods. 2nd ed. American Society of Agronomy, Soil Science Society of America, Madison, USA. [本文引用: 1]
LegendreP, MiXC, RenHB, MaKP, YuMJ, SunIF, HeFL (2009). Partitioning beta diversity in a subtropical broad- leaved forest of China Ecology, 90,663-674. PMID:19341137 [本文引用: 1] The classical environmental control model assumes that species distribution is determined by the spatial variation of underlying habitat conditions. This niche-based model has recently been challenged by the neutral theory of biodiversity which assumes that ecological drift is a key process regulating species coexistence. Understanding the mechanisms that maintain biodiversity in communities critically depends on our ability to decompose the variation of diversity into the contributions of different processes affecting it. Here we investigated the effects of pure habitat, pure spatial, and spatially structured habitat processes on the distributions of species richness and species composition in a recently established 24-ha stem-mapping plot in the subtropical evergreen broad-leaved forest of Gutianshan National Nature Reserve in East China. We used the new spatial analysis method of principal coordinates of neighbor matrices (PCNM) to disentangle the contributions of these processes. The results showed that (1) habitat and space jointly explained approximately 53% of the variation in richness and approximately 65% of the variation in species composition, depending on the scale (sampling unit size); (2) tree diversity (richness and composition) in the Gutianshan forest was dominantly controlled by spatially structured habitat (24%) and habitat-independent spatial component (29%); the spatially independent habitat contributed a negligible effect (6%); (3) distributions of richness and species composition were strongly affected by altitude and terrain convexity, while the effects of slope and aspect were weak; (4) the spatial distribution of diversity in the forest was dominated by broad-scaled spatial variation; (5) environmental control on the one hand and unexplained spatial variation on the other (unmeasured environmental variables and neutral processes) corresponded to spatial structures with different scales in the Gutianshan forest plot; and (6) five habitat types were recognized; a few species were statistically significant indicators of three of these habitats, whereas two habitats had no significant indicator species. The results suggest that the diversity of the forest is equally governed by environmental control (30%) and neutral processes (29%). In the fine-scale analysis (10 x 10 m cells), neutral processes dominated (43%) over environmental control (20%).
LiangJ, CrowtherTW, PicardN, WiserS, ZhouM, AlbertiG, SchulzeE-D, DavidMcGuire A, BozzatoF, PretzschH, deMiguel S, PaquetteA, HéraultB, Scherer-LorenzenM, BarrettCB, et al. (2016). Positive biodiversity-productivity relationship predominant in global forests Science, 354,aaf8957. DOI: 10.1126/science.aaf8957. DOI:10.1126/science.aaf8957URL [本文引用: 2]
LinF, ComitaLS, WangXG, BaiXJ, YuanZQ, XingDL, HaoZQ (2014). The contribution of understory light availability and biotic neighborhood to seedling survival in secondary versus old-growth temperate forest Plant Ecology, 215,795-807. DOI:10.1007/s11258-014-0332-0URL [本文引用: 1]
MuthCC, BazzazFA (2003). Tree canopy displacement and neighborhood interactions Canadian Journal of Forest Research, 33,1323-1330. DOI:10.1139/x03-045URL [本文引用: 3]
PretzschH (2014). Canopy space filling and tree crown morphology in mixed-species stands compared with monocultures Forest Ecology and Management, 327,251-264. DOI:10.1016/j.foreco.2014.04.027URL [本文引用: 3]
PurvesDW, LichsteinJW, PacalaSW (2007). Crown plasticity and competition for canopy space: a new spatially implicit model parameterized for 250 North American tree species PLOS ONE, 2,e870. DOI: 10.1371/journal.pone.0000870. DOI:10.1371/journal.pone.0000870URL [本文引用: 3]
QuesadaCA, PhillipsOL, SchwarzM, CzimczikCI, BakerTR, Pati?oS, FyllasNM, HodnettMG, HerreraR, AlmeidaS, AlvarezDávila E, ArnethA, ArroyoL, ChaoKJ, DezzeoN, et al. (2012). Basin-wide variations in Amazon forest structure and function are mediated by both soils and climate Biogeosciences, 9,2203-2246. DOI:10.5194/bg-9-2203-2012URL [本文引用: 1]
RissanenK, Martin-GuayMO, Riopel-BouvierAS, PaquetteA (2019). Light interception in experimental forests affected by tree diversity and structural complexity of dominant canopy Agricultural and Forest Meteorology, 278,107655. DOI: 10.1016/j.agrformet.2019.107655. DOI:10.1016/j.agrformet.2019.107655URL [本文引用: 1]
RosseelY (2012). lavaan: an R package for structural equation modeling Journal of Statistical Software, 48,2. DOI: 10.18637/jss.v048.i02. DOI:10.18637/jss.v048.i02 [本文引用: 1]
SapijanskasJ, PaquetteA, PotvinC, KunertN, LoreauM (2014). Tropical tree diversity enhances light capture through crown plasticity and spatial and temporal niche differences Ecology, 95,2479-2492. DOI:10.1890/13-1366.1URL [本文引用: 2]
SeidelD, LeuschnerC, MüllerA, KrauseB (2011). Crown plasticity in mixed forests—Quantifying asymmetry as a measure of competition using terrestrial laser scanning Forest Ecology and Management, 261,2123-2132. DOI:10.1016/j.foreco.2011.03.008URL [本文引用: 1]
SterbaH, DirnbergerG, RitterT (2019). Vertical distribution of leaf area of European larch (Larix decidua Mill.) and Norway spruce (Picea abies L. Karst.) in pure and mixed stand Forests, 10,570. DOI: 10.3390/f10070570. DOI:10.3390/f10070570URL [本文引用: 1]
To?goM, PerotT, CourbaudB, CastagneyrolB, GégoutJC, LonguetaudF, JactelH, ValletP (2018). Difference in shade tolerance drives the mixture effect on oak productivity Journal of Ecology, 106,1073-1082. DOI:10.1111/jec.2018.106.issue-3URL [本文引用: 1]
VockenhuberEA, ScherberC, LangenbruchC, Mei?nerM, SeidelD, TscharntkeT (2011). Tree diversity and environmental context predict herb species richness and cover in Germany?s largest connected deciduous forest Perspectives in Plant Ecology, Evolution and Systematics, 13,111-119. DOI:10.1016/j.ppees.2011.02.004URL [本文引用: 1]
WangYQ (1995). The Mixed Broadleaved-Korean Pine Forest. Northeast Forestry University Press, Harbin. [本文引用: 2]
WenPY, JinGZ (2019). Effects of topography on species diversity in a typical mixed broadleaved-Korean pine forest Acta Ecologica Sinica, 39,945-956. [本文引用: 1]
YuanZ, AliA, WangS, GazolA, FreckletonR, WangX, LinF, YeJ, ZhouL, HaoZ, LoreauM (2018). Abiotic and biotic determinants of coarse woody productivity in temperate mixed forests Science of the Total Environment, 630,422-431. DOI:10.1016/j.scitotenv.2018.02.125URL [本文引用: 3]
ZellerL, LiangJ, PretzschH (2018). Tree species richness enhances stand productivity while stand structure can have opposite effects, based on forest inventory data from Germany and the United States of America Forest Ecosystems, 5,4. DOI: 10.1186/s40663-017-0127-6. DOI:10.1186/s40663-017-0127-6URL [本文引用: 1]
ZhangY, ChenHYH, ReichPB (2012). Forest productivity increases with evenness, species richness and trait variation: a global meta-analysis Journal of Ecology, 100,742-749. DOI:10.1111/j.1365-2745.2011.01944.xURL [本文引用: 2]
ZhengLT, ChenHYH, YanER (2019). Tree species diversity promotes litterfall productivity through crown complementarity in subtropical forests Journal of Ecology, 107,1852-1861. DOI:10.1111/jec.2019.107.issue-4URL [本文引用: 3]
ZhuY, CaiHY, JiangF, JinGZ (2017). Variation of the biotic neighbourhood and topographic effects on tree survival in an old-growth temperate forest Journal of Vegetation Science, 28,1166-1177. DOI:10.1111/jvs.2017.28.issue-6URL [本文引用: 1]
Tree crown complementarity links positive functional diversity and aboveground biomass along large-scale ecological gradients in tropical forests 2 2019
... 生物多样性-生产力关系(BPR)是理解全球物种灭绝危机及其对生态系统功能影响的基础, 对于准确评估和有效保护生物多样性至关重要(Zhang et al., 2012; Liang et al., 2016; Yuan et al., 2018).早期的BPR研究多集中于分析物种多样性的作用, 对生产力的解释和预测能力较低(Grace et al., 2016; Zeller et al., 2018).近年来, 随着对生物多样性不同维度(物种、功能、谱系和结构多样性)认识的加深和扩展, 生态学家们基于树冠结构在垂直和水平方向上的差异提出林分结构多样性的衍生多样性——树冠结构复杂性.树冠结构复杂性作为综合评价森林冠层对光照时空分配利用情况的重要参数, 可以较全面地描述森林的林分结构特征(Sapijanskaset al., 2014), 其值越高说明林内个体对立地空间的利用率越高, 对林分中光照等资源的利用更全面, 进而具有更高的森林生产力(Ali et al., 2019). ...
Structural crown properties of Norway spruce (Picea abies L. Karst.) and European beech (Fagus sylvatica L.) in mixed versus pure stands revealed by terrestrial laser scanning 2 2013
... 树冠结构复杂性可分两部分来分别解释树冠结构在垂直与水平方向上的作用(Pretzsch, 2014; Jucker et al., 2015).一是树冠垂直分层: 当树木的叶片分布在冠层的不同高度剖面时, 森林冠层能够最大限度地截留光线, 从而对森林生产力产生积极的促进作用(Williams et al., 2017).共存树种对光照的分层截留以及对郁闭环境的生理适应, 均有利于树冠垂直分层(Sterba et al., 2019).二是树冠可塑性: 树木的大小和形状在生长过程中具有较强的可塑性, 可以形成镶嵌的树冠结构, 覆盖大部分或所有的树冠空间(Purves et al., 2007).树冠横向发展, 如展叶、分枝和树冠形状的变化, 均有助于减少林分中激烈的种内、种间竞争, 进而促进森林生态系统功能优化(Jucker et al., 2015; Fotis et al., 2018).树冠结构复杂性被系统地提出之后, 该研究领域的成果迅速增加, 大量实验证明树冠结构复杂性可以提高森林生态系统的生产力, 但是树冠垂直分层和树冠可塑性二者的作用强度和内在作用机制还有争议(Pretzsch, 2014; Fahey et al., 2015).部分****认为多林层林分具有更高的生产潜能, 林冠垂直分层是诸多森林生物量积累的重要驱动因素(Hardiman et al., 2013; D?nescuet al., 2016).然而, 这与Bayer等(2013)和Jucker等(2015)的研究相悖, 他们通过分离垂直和水平方向上的树冠结构复杂性发现, 可塑性较强的树冠结构才是影响生产力的重要因素, 树冠垂直分层反而没有明显作用.不同地区和林龄中树冠结构复杂性对生产力的贡献相异(Bohn & Huth, 2017), 因此仍需更多系统的研究来阐明树冠垂直分层和树冠可塑性的相对重要性.此外, 林分中较高的物种丰富度, 往往会形成较高的树冠结构复杂性, 从而间接地促进森林生产力.然而, 物种多样性对生产力的这种间接影响尚未得到完全证实(Forrester, 2014; Sapijanskas et al., 2014). ...
Tree species richness enhances stand productivity while stand structure can have opposite effects, based on forest inventory data from Germany and the United States of America 1 2018
... 生物多样性-生产力关系(BPR)是理解全球物种灭绝危机及其对生态系统功能影响的基础, 对于准确评估和有效保护生物多样性至关重要(Zhang et al., 2012; Liang et al., 2016; Yuan et al., 2018).早期的BPR研究多集中于分析物种多样性的作用, 对生产力的解释和预测能力较低(Grace et al., 2016; Zeller et al., 2018).近年来, 随着对生物多样性不同维度(物种、功能、谱系和结构多样性)认识的加深和扩展, 生态学家们基于树冠结构在垂直和水平方向上的差异提出林分结构多样性的衍生多样性——树冠结构复杂性.树冠结构复杂性作为综合评价森林冠层对光照时空分配利用情况的重要参数, 可以较全面地描述森林的林分结构特征(Sapijanskaset al., 2014), 其值越高说明林内个体对立地空间的利用率越高, 对林分中光照等资源的利用更全面, 进而具有更高的森林生产力(Ali et al., 2019). ...
Forest productivity increases with evenness, species richness and trait variation: a global meta-analysis 2 2012
... 生物多样性-生产力关系(BPR)是理解全球物种灭绝危机及其对生态系统功能影响的基础, 对于准确评估和有效保护生物多样性至关重要(Zhang et al., 2012; Liang et al., 2016; Yuan et al., 2018).早期的BPR研究多集中于分析物种多样性的作用, 对生产力的解释和预测能力较低(Grace et al., 2016; Zeller et al., 2018).近年来, 随着对生物多样性不同维度(物种、功能、谱系和结构多样性)认识的加深和扩展, 生态学家们基于树冠结构在垂直和水平方向上的差异提出林分结构多样性的衍生多样性——树冠结构复杂性.树冠结构复杂性作为综合评价森林冠层对光照时空分配利用情况的重要参数, 可以较全面地描述森林的林分结构特征(Sapijanskaset al., 2014), 其值越高说明林内个体对立地空间的利用率越高, 对林分中光照等资源的利用更全面, 进而具有更高的森林生产力(Ali et al., 2019). ...
 ,
, ,1,3,*
,1,3,*

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT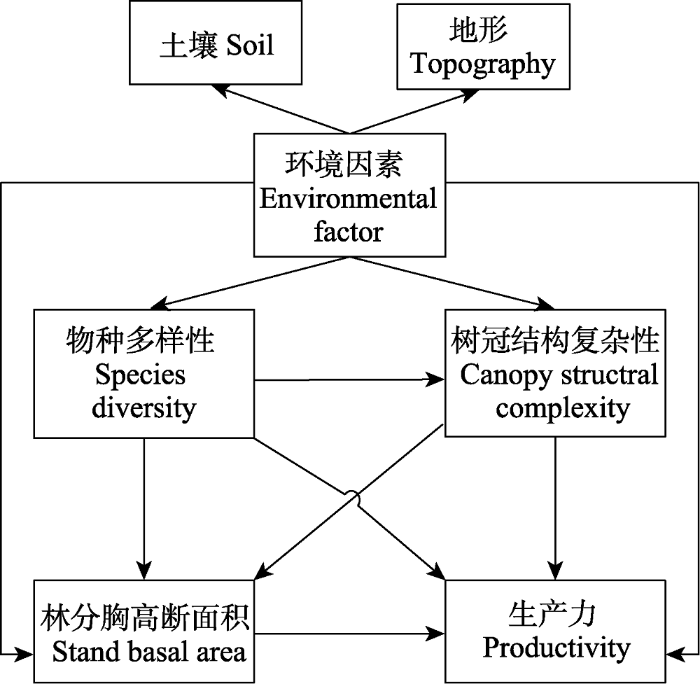 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT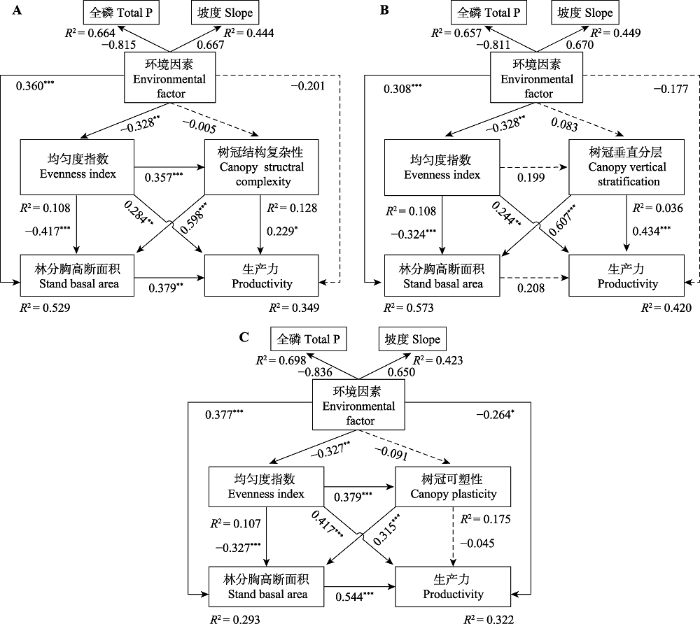 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT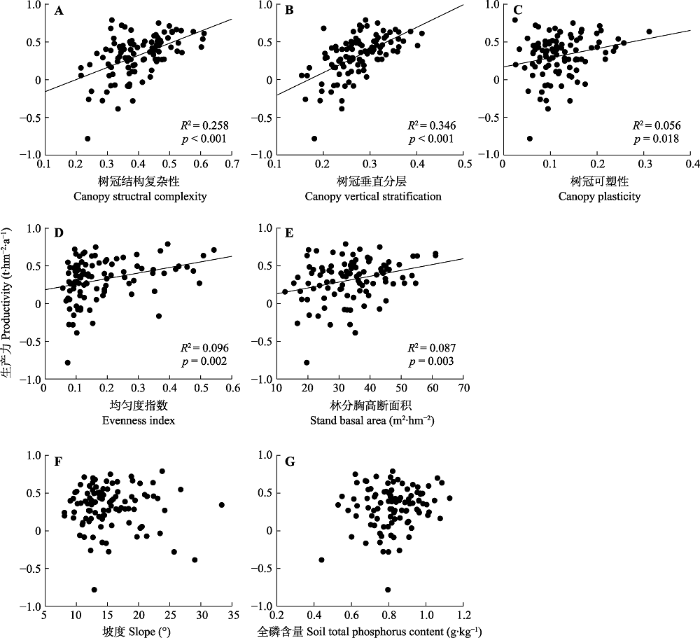 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}