王雪梅
1 , 2 , 闫帮国
3 , 史亮涛
3 , 刘刚才
, 2 , * 1 绵阳师范学院资源环境工程学院, 四川绵阳 6210002 中国科学院·水利部成都山地灾害与环境研究所, 成都 6100413 云南省农业科学院热区生态农业研究所, 云南元谋 651300Different responses of biomass allocation and leaf traits of Dodonaea viscosa to concentrations of nitrogen and phosphorus Xue-Mei WANG
1 , 2 , Bang-Guo YAN
3 , Liang-Tao SHI
3 , Gang-Cai LIU
, 2 , * 1 School of Resource and Environmental Engineering, Mianyang Normal University, Mianyang, Sichuan 621000, China2 Institute of Mountain Hazards and Environment, Ministry of Water Resources, Chinese Academy of Sciences, Chengdu 610041, China; and 3Institute of Tropical Eco-Agriculture, Yunnan Academy of Agricultural Sciences, Yuanmou, Yunnan 651300, China3 Institute of Tropical Eco-Agriculture, Yunnan Academy of Agricultural Sciences, Yuanmou, Yunnan 651300, China通讯作者: *(
liugc@imde.ac.cn )
编委: 石培礼
责任编辑: 李敏
收稿日期: 2020-06-18
接受日期: 2020-09-25
网络出版日期: 2020-12-20
Corresponding authors: *(
liugc@imde.ac.cn )
Received: 2020-06-18
Accepted: 2020-09-25
Online: 2020-12-20
Fund supported: National Natural Science Foundation of China (31460127 )
摘要 调整叶性状和生物量分配格局是植物适应环境变化的主要途径, 研究车桑子(
Dodonaea viscosa )幼苗生物量分配与叶性状对氮磷浓度的响应对认识车桑子在氮磷浓度变化下的适应策略具有重要意义。该研究通过砂培法, 测定不同氮浓度(3、5、15、30 mmol·L
-1 )与不同磷浓度(0.25、0.5、1、2 mmol·L
-1 )下车桑子幼苗的生长、生物量分配、叶性状的响应特征及其相互关系。结果表明: 高浓度氮(30 mmol·L
-1 )促进了车桑子幼苗生长、叶片氮含量和生物量积累, 其余氮添加条件(3、5、15 mmol·L
-1 )下车桑子幼苗各性状无显著差异, 但相比高氮水平, 其生物量积累和叶片氮含量显著降低, 根冠比和氮利用效率显著增加。随着磷添加浓度的增加, 车桑子幼苗生物量显著增加, 低磷条件(0.25、0.5 mmol·L
-1 )限制了车桑子幼苗生长和生物量积累, 其根冠比和磷利用效率均没有发生显著变化, 但比叶面积和叶/茎生物量比例显著增加, 叶干物质含量显著降低。氮处理下, 叶片氮含量与根冠比显著负相关; 磷处理下, 叶片氮含量与比叶面积显著正相关。同时, 氮处理下, 车桑子幼苗株高、基径、总生物量等生长性状均与根冠比显著负相关, 与叶片氮含量显著正相关, 表明根冠比和叶片氮含量的调整在车桑子适应氮限制中发挥重要作用; 而磷处理下, 株高、基径、总生物量与比叶面积显著负相关, 与叶干物质含量显著正相关, 表明叶片结构性状的调整在车桑子适应低磷环境中具有重要意义。该研究表明, 车桑子幼苗生物量分配和叶性状及性状间的权衡策略对氮、磷的响应具有明显差异性, 在今后的研究中, 应关注氮和磷对植物性状影响的差异性。
关键词: 车桑子 ;
氮浓度 ;
磷浓度 ;
干热河谷 ;
养分限制 ;
适应策略 Abstract Aims The adjustment of leaf traits and biomass allocation is an important way for plants to adapt to environmental changes. Revealing the responses of biomass allocation and leaf traits of
Dodonaea viscosa seedlings to nitrogen and phosphorus concentrations, is crucial to understand the adaptation strategies of
D. viscosa under the changes of nitrogen and phosphorus.
Methods Seedlings of
D. viscosa were planted under nitrogen concentrations (3, 5, 15, 30 mmol·L
-1 ) and phosphorus concentrations (0.25, 0.5, 1, 2 mmol·L
-1 ) by sand culture. Plant height, base diameter, biomass allocation, leaf traits and their correlations were quantified.
Important findings The results showed that high nitrogen concentration (30 mmol·L
-1 ) increased the height, diameter, leaf nitrogen concentration, and biomass accumulation of
D. viscosa, and there were no significant differences of the traits under other concentrations (3, 5, 15 mmol·L
-1 ). Compared with the high nitrogen level, other treatments significantly reduced the biomass accumulation and leaf nitrogen concentration, and significantly increased the root:shoot biomass ratio and nitrogen utilization efficiency. With the increase of phosphorus concentration, the biomass of
D. viscosa increased significantly. Low phosphorus concentrations (0.25, 0.5 mmol·L
-1 ) significantly constrained the growth of
D. viscosa, and the root:shoot biomass ratio and phosphorus utilization efficiency did not change significantly. Low phosphorus conditions increased the specific leaf area and the leaf:stem biomass ratio, and decreased leaf dry matter content significantly. Under the nitrogen treatment, leaf nitrogen concentration was negatively correlated with the root:shoot biomass ratio, while under phosphorus treatment, leaf nitrogen concentration was positively correlated with specific leaf area. Height, diameter and total biomass of
D. viscosa were negatively correlated with the root:shoot biomass ratio, and positively correlated with leaf nitrogen concentration under the condition of nitrogen treatment, indicating that the adjustment of root:shoot biomass ratio and leaf nitrogen concentration played an important role in adapting to nitrogen limitation. However, under the condition of phosphorus treatment, height, diameter and total biomass were negatively correlated with specific leaf area, and positively correlated with leaf dry matter content, indicating that the adjustment of leaf structural traits was of great significance in adapting to changes of phosphorus. Our findings suggest that the biomass allocation, leaf traits and their relationships responded differently to changes in nitrogen and phosphorus, and the effects of nitrogen or phosphorus on plant traits should be discriminated in the future.
Keywords: Dodonaea viscosa ;
nitrogen concentration ;
phosphorus concentration ;
dry-hot valley ;
nutrient limitation ;
adaptive strategy PDF (1353KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 王雪梅, 闫帮国, 史亮涛, 刘刚才. 车桑子幼苗生物量分配与叶性状对氮磷浓度的响应差异. 植物生态学报, 2020, 44(12): 1247-1261. DOI:
10.17521/cjpe.2020.0199 WANG Xue-Mei, YAN Bang-Guo, SHI Liang-Tao, LIU Gang-Cai.
Different responses of biomass allocation and leaf traits of Dodonaea viscosa to concentrations of nitrogen and phosphorus .
Chinese Journal of Plant Ecology , 2020, 44(12): 1247-1261. DOI:
10.17521/cjpe.2020.0199 氮和磷是构成生命体生物膜、蛋白质、核酸、磷脂和ATP等物质的重要成分, 因此是植物生命过程中必需的营养元素, 但植物生长普遍会受到氮磷养分的限制(
Yan et al ., 2015 )。氮、磷的限制格局和驱动因素各不相同。通常认为, 演替早期的生态系统主要受到氮的限制, 而演替后期的生态系统主要受到磷的限制(
Menge et al ., 2012 )。淋溶、挥发、反硝化作用和微生物对氮有限的固定作用等都可引起植物氮的缺乏(
Vitousek & Howarth, 1991 )。相比之下, 土壤物理屏蔽、低磷母岩、闭蓄态转化和人类活动(增加其他资源特别是氮的供给)等是引起植物生长过程中磷限制的主要原因(
Vitousek et al ., 2010 )。从植物个体的适应性出发, 充分利用植物自身对养分限制的适应性, 对提高植物生产力具有重要的意义。因此, 植物性状对氮、磷的响应特征对生态系统生产力和生态系统功能具有重要影响(
Wang et al ., 2018 ;
Xu et al ., 2018 ;
Zhang et al ., 2019 )。
在养分有效性发生变化时, 植物会通过生物量分配和叶片功能性状的调整来适应养分变化。按照最优分配理论, 植物会将更多的生物量分配至能够获取限制资源的器官(
Mccarthy & Enquist, 2007 )。在养分有效性较低的情况下通常会通过增加根生物量分配和根冠比来获取更多的养分(
Poorter et al ., 2012 )。此外, 养分贫瘠的条件下, 植物会增加地上部分叶生物量的比例(相对于茎)(
Yan et al ., 2016 ), 但这方面的研究还相对较少。另一方面, 叶片功能性状的调整与资源获取和利用密切相关, 其形态结构和营养元素含量等均会随环境中氮磷条件的改变而发生适应性变化(
Qi et al ., 2009 )。比如, 较低的养分有效性通常会降低比叶面积, 增加叶干物质含量和养分利用效率, 而随着养分有效性的增加, 叶片养分含量显著增加(
Ordo?ez et al ., 2009 ;
林婉奇等, 2019 )。然而, 不同物种对氮磷添加的响应具有物种特异性(
Han et al ., 2013 ;
Mayor et al ., 2014 )。在氮限制条件下, 具有更高根长和根生物量比例的物种占据优势; 相反, 在磷限制条件下, 植物根长不能解释物种间的竞争优势, 菌根菌丝长度、根系分泌物、磷酸酶活性等相关性状起着重要作用(
Venterink & Güsewell, 2010 ;
Minden & Venterink, 2019 ), 这说明植物性状对氮和磷可能具有不同的响应特征, 包括生物量分配、叶片性状或养分利用效率等多个植物性状。由于不同的性状在适应氮限制和磷限制中表现出不同的变化趋势, 植物可能通过调整不同的性状来响应氮变化或磷变化, 但植物性状对氮、磷的响应是否表现出差异并不十分清楚。
此外, 如上所述, 植物种内性状具有极大的变异性, 这被认为是植物性状在适应环境中的可塑性决定的。另一方面物种之间性状上也存在极大的差异, 并形成一个叶经济型谱。叶经济型谱理论认为植物叶片在其形态结构、化学组分和生理功能等性状之间存在着相互依存、相互关联的内在关系(
Wright et al ., 2004 ;
于鸿莹等, 2014 )。多种功能性状之间关系紧密, 不同性状的组合可体现出植物对不同环境的适应策略(
熊静等, 2019 )。那么, 在环境梯度如养分梯度下植物种内性状的可塑性及其关联性是否与种间的这种经济型谱变化趋势一致? 例如, 经济型谱中比叶面积与叶片氮含量显著正相关, 那么在氮浓度梯度中植物种内的比叶面积与叶片氮含量是否会产生类似的关系? 植物种内性状的可塑性调整在研究植物适应策略方面有着重要作用。目前,环境变化对叶性状和生物量分配的表型可塑性方面的研究较多, 然而, 在氮磷变化条件下, 植物在生物量分配和叶性状的调整上是否存在如同经济型谱理论的相互变化规律? 这方面的证据还比较有限。开展生物量分配和叶性状对氮磷浓度的响应研究, 有助于全面理解和揭示植物对养分有效性变化的响应与适应机制。
车桑子(
Dodonaea viscosa )又名坡柳、明油子, 具有耐干旱、耐贫瘠和适应性强等特性, 是我国干热河谷区的优势物种, 也是该区植被恢复工程中的常见植物, 具有重要的生态价值。关于车桑子的研究, 目前重点研究了车桑子对干旱胁迫的响应特征(
李吉跃等, 2003 ;
陈少瑜等, 2006 )。但除了干旱胁迫之外, 养分胁迫成为干旱胁迫的次生效应(
da Silva et al ., 2011 )。加之该区土壤侵蚀严重, 干热河谷土壤(燥红土)普遍存在缺氮少磷的特点(
张燕平等, 2005 ), 严重影响了干热河谷植物的生长(
闫帮国等, 2012 )。然而, 关于车桑子性状对氮磷限制的响应目前尚不清楚。因此, 本研究以车桑子幼苗为研究对象, 采用砂培的方式进行试验, 研究车桑子幼苗生长特征、生物量分配、叶片功能性状及其相互关系对氮磷梯度的响应特征, 从生物量分配和叶片功能性状的角度探讨车桑子对低氮和低磷条件的适应性, 并验证以下假说: (1)车桑子幼苗生物量分配和叶片功能性状对氮变化和磷变化的响应具有差异性; (2)生物量分配和叶性状对氮磷浓度响应存在协同或权衡关系。
1 材料和方法 1.1 试验材料 试验在中国科学院成都山地灾害与环境研究所元谋干热河谷沟蚀崩塌观测研究站(101.82° E, 25.85° N)温室大棚内进行。试验前, 在干热河谷典型区域选择自然植被受人类干扰相对较小、性状优良的车桑子母树采集成熟饱满的种子。采集区位于元谋县黄瓜园镇苴林村附近, 属于典型的干热河谷气候区, 土壤类型为燥红土, 表层土的碱解氮、铵态氮、硝态氮和有效磷含量分别为41.28、3.09、8.57和4.90 mg·kg
-1 。该区年平均气温21.5 ℃, 蒸发量大于3 500 mm, 年降水量小于700 mm。其中6-10月为雨季, 期间降水量占全年的90%以上, 而干季较长, 达6-7个月(11月至翌年5月)。将采集的种子装于布袋, 带回实验室自然风干保存。试验基质为经过高温和高压(121 ℃、103 kPa, 15 min)灭菌后的洁净河沙(直径< 2 mm)。栽培容器为塑料盆钵, 其规格为: 上口径×下口径×高= 26 cm × 16 cm × 24 cm, 将等体积的沙子装入各盆钵。挑选饱满、健康的车桑子种子, 用浓硫酸破除种子休眠(98%浓H
2 SO
4 , 10 min)后均匀播种于盆钵中, 播种后在表面再覆盖一层沙子, 约3-5 cm。试验前期, 为了保证车桑子能够正常出苗和生长, 并避免养分浓度过高对车桑子幼苗的毒害作用, 所有盆钵定期等量添加稀释5倍后的Hoagland营养液。待车桑子出苗长出4-5片真叶时, 间苗至3株/盆, 雨季来临时(6月初)开始进行氮磷养分添加处理。
1.2 试验设计 本试验采用氮和磷的单因素设计, 分别包括4个氮浓度和4个磷浓度。由于盆栽实验中植物根系伸展空间有限, 按照野外土壤实际养分含量设置浓度梯度下植物很难获得充分生长。因此, 本试验参照前人的相关研究(
Yu et al ., 2011 ;
Yan et al ., 2015 ), 以Hoagland营养液(15 mmol·L
-1 N、1 mmol·L
-1 P)为基础进行氮磷养分添加。Hoagland营养液已经比较成熟, 溶液的pH较为稳定, 其养分配制考虑了植物需求, 养分浓度对大多数植物都很适合, 被广泛应用于水培和砂培实验中(
Hoagland & Arnon, 1938 )。由于河沙中没有养分, 不添加额外养分植物将无法生长, 因此, 本研究没有设置不添加的对照处理。结合浓度梯度的可控性, 4个氮浓度分别为3、5、15、30 mmol·L
-1 , 记为N1、N2、N3、N4, 其中, N4为高浓度氮水平, N3为中等浓度氮水平, N1和N2为低浓度氮水平, 通过添加NH
4 NO
3 进行调整; 4个磷浓度分别为0.25、0.5、1、2 mmol·L
-1 , 记为P1、P2、P3、P4, 其中, P4为高浓度磷水平, P3为中等浓度磷水平, P1和P2为低浓度磷水平, 通过添加KH
2 PO
4 进行调整。各氮、磷水平的设置及营养液的氮磷比如
表1 所示。其中, N3和P3处理一致, 因此, 试验共计7个处理, 每个处理5个重复。除氮、磷浓度不同外, 其余钾、钙、镁含量及微量元素含量均与全量Hoagland营养液一致, 配好后用H
2 SO
4 或NaOH调整溶液pH为6。由于钾、钙、镁的添加均为硫酸盐物质, 通过H
2 SO
4 调节pH对物质的平衡影响较小, 而钠离子为植物非必需元素, 对养分需求的平衡不会造成影响, 同时营养液中所添加的物质主要为中性物质, 不同营养液中pH差异较小, 所需H
2 SO
4 或NaOH的量是极小的, 因此, 这些影响基本可以忽略, 试验处理中不存在离子不平衡问题。由于砂培基质对营养液的保持能力较低, 试验处理期间, 每隔2天浇1次营养液, 每次营养液添加量为200 mL, 以保持养分浓度尽量维持在设定水平。同时, 将所有盆钵放在遮雨大棚内, 可避免降水对养分的淋失。
Table 1 表1 表1 各氮磷浓度梯度下营养液的氮磷浓度和氮磷比值
Table 1
N and P concentrations and the N:P ratio in different nitrogen and phosphorus concentration gradients 氮、磷水平 氮浓度-1 ) 磷浓度-1 ) 氮磷比 N1 3 1 3 N2 5 1 5 N3 15 1 15 N4 30 1 30 P1 15 0.25 60 P2 15 0.5 30 P3 15 1 15 P4 15 2 7.5
新窗口打开 |
下载CSV 除营养液添加当天外, 其余时间各盆钵每天浇等量的自来水, 以保证植株不受干旱胁迫的影响。此外, 由于试验前不清楚车桑子对氮磷养分的吸收情况, 添加营养液5次后用2 L自来水淋洗1次花盆, 以避免盐基离子积累对车桑子幼苗的毒害作用(
Yu et al ., 2011 )。试验处理时间为一个生长季, 在雨季结束时(11月)停止试验。
1.3 取样和测定方法 试验末期, 用直尺量取每株植株的高度, 并用精度为0.05 mm的游标卡尺测量主干基径。同时, 每盆从植株中部随机选取6片充分展开的叶片作为代表测量叶面积、叶干物质含量(
LDMC )和比叶面积(
SLA )。将各组叶片插入水中, 在5 ℃的黑暗环境中储藏12 h后, 将叶片取出, 在1/10 000的电子天平上称质量(饱和鲜质量)。然后将离体叶片用扫描仪扫描, 扫描图片用Image J数字化计算后获得叶面积。最后将叶片放入65 ℃烘箱内烘干48 h后取出称质量(干质量), 根据公式计算
LDMC 和
SLA 。
LDMC (g·g
-1 ) =叶片干质量/叶片饱和鲜质量
SLA (cm
2 ·g
-1 ) =叶片面积/叶片干质量
采集每盆植株中部的叶片, 将叶片烘干、称质量和粉碎, 测定其氮磷含量。全氮含量采用H
2 SO
4 -H
2 O
2 消煮-凯氏蒸馏法测定, 全磷含量采用H
2 SO
4 -H
2 O
2 消煮-钼锑抗比色法测定。养分利用效率可用单位养分吸收量所产生的干物质量大小来表示。因此, 氮(磷)利用效率=总生物量/氮(磷)吸收量(
冷华妮等, 2009 )。
植株收获时, 将各盆中植株的根、茎、叶分开取样, 冲洗干净。随后立即放入65 ℃烘箱中烘干至恒质量, 用电子天平分别称量每盆中根、茎、叶的干质量, 并计算各处理条件下车桑子幼苗的地下生物量/地上生物量的比例(根冠比)和叶/茎生物量比例。
1.4 数据处理 为了进一步明确车桑子叶片的氮磷营养特征, 通过化学计量内稳性模型计算车桑子叶片氮、磷化学计量的内稳性指数。该模型指数公式为:
Y =
cX 1/ H 式中,
Y 是车桑子幼苗叶片氮、磷含量及N:P,
X 是营养液中对应的氮、磷浓度及对应的N:P,
c 为常数,
H 为植物的内稳性指数(
Yu et al ., 2011 )。
用Excel软件对数据进行初步整理和统计分析后, 用SPSS 19.0统计软件分别对不同氮浓度和不同磷浓度进行单因素方差分析, 并用最小显著差法(
LSD )对不同处理间的差异性进行多重比较, 用Pearson相关分析法分析各性状之间的关联特征, 显著性水平设定为
α = 0.05。
2 结果和分析 2.1 不同氮磷浓度下车桑子幼苗生长特征 不同的氮和磷浓度处理均极显著影响车桑子株高和基径生长(
p < 0.001)(
图1 )。相比中等浓度氮水平(N3), 高浓度氮水平(N4)处理的株高和基径分别显著提高了53.8%和37.0%, 但低、中浓度氮水平(N1、N2、N3)处理的株高和基径均没有显著的差异(
p > 0.05)。随磷添加浓度的增加, 车桑子幼苗的株高和基径显著增加。高浓度磷处理(P4)比中等浓度磷处理(P3)的株高和基径分别显著提高了40.4%和30.8%, 同时, 低浓度磷处理(P1、P2)显著降低了车桑子幼苗的株高和基径, 说明低磷处理较低的磷浓度限制了车桑子幼苗的生长。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1不同氮浓度和磷浓度对车桑子幼苗株高和基径的影响(平均值±标准误差)。 N1-N4, 氮浓度分别为3、5、15、30 mmol·L
-1 ; P1-P4, 磷浓度分别为0.25、0.5、1、2 mmol·L
-1 。不同小写字母表示不同氮处理或不同磷处理下差异显著(
p < 0.05)。
Fig. 1Effects of different nitrogen and phosphorus concentrations on height and base diameter of Dodonaea viscosa seedlings (mean ± SE ). N1-N4, the nitrogen concentrations were 3, 5, 15 and 30 mmol·L
-1 , respectively; P1-P4, the phosphorus concentrations were 0.25, 0.5, 1 and 2 mmol·L
-1 , respectively. Different lowercase letters indicate significant differences (
p < 0.05) among different nitrogen treatments or different phosphorus treatments.
不同氮浓度处理下车桑子幼苗的根、茎和叶生物量差异显著(
p < 0.05)(
图2 )。与上述株高和基径的结果一致, N4处理显著提高了车桑子幼苗的根、茎和叶生物量, 其总生物量是N3处理的2倍, 但低、中浓度氮处理下(N1、N2、N3)各生物量均无显著差异。同样地, 随着磷浓度的增加, 车桑子根、茎和叶生物量均显著增加, 即P4 > P3 > P2 > P1, P4处理的车桑子总生物量比P3处理增加了1倍。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2不同氮浓度和磷浓度对车桑子幼苗生物量的影响(平均值±标准误差)。 N1-N4, 氮浓度分别为3、5、15、30 mmol·L
-1 ; P1-P4, 磷浓度分别为0.25、0.5、1、2 mmol·L
-1 。不同小写字母表示不同氮处理或不同磷处理下差异显著(
p < 0.05)。
Fig. 2Effects of different nitrogen and phosphorus concentrations on biomass of Dodonaea viscosa seedlings (mean ± SE ). N1-N4, the nitrogen concentrations were 3, 5, 15 and 30 mmol·L
-1 , respectively; P1-P4, the phosphorus concentrations were 0.25, 0.5, 1 and 2 mmol·L
-1 , respectively. Different lowercase letters indicate significant differences (
p < 0.05) among different nitrogen treatments or different phosphorus treatments.
2.2 不同氮磷浓度下车桑子幼苗生物量分配特征 氮浓度处理显著影响了车桑子幼苗的根冠比(
p < 0.01)。高浓度氮水平(N4)显著降低了根冠比, 低浓度和中等浓度的根冠比无显著差异。然而, 各磷浓度处理下车桑子幼苗的根冠比无显著差异(
图3A 、3B)。不同的氮浓度和磷浓度对车桑子幼苗的叶/茎生物量分配比例均造成了显著影响, 叶/茎生物量比例随氮浓度和磷浓度的增加而降低(
图3C 、3D)。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3不同氮浓度和磷浓度对车桑子幼苗根冠比和叶/茎生物量比例的影响(平均值±标准误差)。 N1-N4, 氮浓度分别为3、5、15、30 mmol·L
-1 ; P1-P4, 磷浓度分别为0.25、0.5、1、2 mmol·L
-1 。不同小写字母表示不同氮处理或不同磷处理下差异显著(
p < 0.05)。
Fig. 3Effects of different nitrogen and phosphorus concentrations on the root:shoot biomass ratio and the leaf:stem biomass ratio of Dodonaea viscosa seedlings (mean ± SE ). N1-N4, the nitrogen concentrations were 3, 5, 15 and 30 mmol·L
-1 , respectively; P1-P4, the phosphorus concentrations were 0.25, 0.5, 1 and 2 mmol·L
-1 , respectively. Different lowercase letters indicate significant differences (
p < 0.05) among different nitrogen treatments or different phosphorus treatments.
2.3 不同氮磷浓度下车桑子幼苗叶片性状 不同的氮磷浓度显著影响了车桑子叶面积(
p < 0.05)(
图4 )。N4处理的叶面积比N3处理显著提高了33.4%, 但低浓度和中等浓度氮处理的叶面积没有显著性差异(
p > 0.05)。各氮浓度处理条件下,
LDMC 和
SLA 没有显著差异(
p > 0.05)。磷浓度处理对叶面积、
LDMC 和
SLA 影响显著(
p < 0.001)。随着磷浓度的增加, 叶面积和
LDMC 显著增加, 而
SLA 显著降低。
图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4不同氮浓度和磷浓度对车桑子幼苗叶面积、叶干物质含量和比叶面积的影响(平均值±标准误差)。 N1-N4, 氮浓度分别为3、5、15、30 mmol·L
-1 ; P1-P4, 磷浓度分别为0.25、0.5、1、2 mmol·L
-1 。不同小写字母表示不同氮处理或不同磷处理下差异显著(
p < 0.05)。
Fig. 4Effects of different nitrogen and phosphorus concentrations on leaf area, leaf dry mass content and specific leaf area of Dodonaea viscosa seedlings (mean ± SE ). N1-N4, the nitrogen concentrations were 3, 5, 15 and 30 mmol·L
-1 , respectively; P1-P4, the phosphorus concentrations were 0.25, 0.5, 1 and 2 mmol·L
-1 , respectively. Different lowercase letters indicate significant differences (
p < 0.05) among different nitrogen treatments or different phosphorus treatments.
N4处理显著提高了车桑子幼苗叶片氮含量(
p < 0.05), 其叶片氮含量比N3处理显著增加了24.5%, 但低浓度和中等浓度氮处理的叶片氮含量则无显著差异。不同磷浓度下, 叶片氮含量差异显著(
p < 0.05), 其中, 低浓度磷水平(P1)的叶片氮含量显著高于其他处理。不同氮处理和不同磷处理下, 车桑子幼苗的叶片磷含量均没有显著性差异(
p > 0.05), 同时, 车桑子幼苗叶片N:P也没有显著性差异(
图5 )。
图5 新窗口打开 |
下载原图ZIP |
生成PPT 图5不同氮浓度和磷浓度对车桑子幼苗叶片氮含量、磷含量和叶片氮磷比的影响(平均值±标准误差)。 N1-N4, 氮浓度分别为3、5、15、30 mmol·L
-1 ; P1-P4, 磷浓度分别为0.25、0.5、1、2 mmol·L
-1 。不同小写字母表示不同氮处理或不同磷处理下差异显著(
p < 0.05)。
Fig. 5Effects of different nitrogen and phosphorus concentrations on leaf N concentration, leaf P concentration and leaf N:P of Dodonaea viscosa seedlings (mean ± SE ). N1-N4, the nitrogen concentrations were 3, 5, 15 and 30 mmol·L
-1 , respectively; P1-P4, the phosphorus concentrations were 0.25, 0.5, 1 and 2 mmol·L
-1 , respectively. Different lowercase letters indicate significant differences (
p < 0.05) among different nitrogen treatments or different phosphorus treatments.
不同处理下叶片磷含量和叶片氮磷比均无显著性差异, 因此, 进一步分析了氮处理和磷处理条件下车桑子幼苗的叶片内稳性特征。用内稳性模型
Y =
cX 1/ H 模拟发现, 不同氮浓度处理下, 叶片氮含量和叶片N:P的内稳性指数分别为6.86和7.31, 叶片N:P的内稳性大于叶片氮的内稳性(
图6 )。而不同磷浓度处理下, 叶片磷含量、叶片N:P与营养液的磷浓度、营养液N:P并没有得到显著的幂函数关系。
图6 新窗口打开 |
下载原图ZIP |
生成PPT 图6氮处理条件下车桑子幼苗叶片氮含量与营养液氮浓度、叶片氮磷比与营养液氮磷比的关系。 H, 内稳性指数。
Fig. 6Relationships between leaf N concentration of Dodonaea viscosa seedlings and N concentration of the nutrient solution, leaf N:P of Dodonaea viscosa seedlings and N:P of the nutrient solution under different nitrogen treatments. H, homeostatic regulation coefficients.
随氮浓度的增加, 氮利用效率显著降低。不同磷浓度处理也造成了氮利用效率的显著差异(
p < 0.01), P1处理的氮利用效率显著低于其他处理。各氮浓度和磷浓度条件下, 磷利用效率无显著差异(
p > 0.05)。总的来看, 氮利用效率随营养液N:P的增加呈显著降低的趋势, 同时, 在营养液N:P为30时, P2处理的氮利用效率高于N4处理。而磷利用效率随营养液N:P的增加并没有呈现出明显的规律性变化(
图7 )。
图7 新窗口打开 |
下载原图ZIP |
生成PPT 图7各营养液氮磷比条件下车桑子幼苗叶片氮利用效率和磷利用效率(平均值±标准误差)。 Fig. 7Leaf N utilization efficiency and leaf P utilization efficiency of Dodonaea viscosa seedlings under different N:P ratios of the nutrient solution (mean ± SE ). 2.4 各性状间的相关特征 从
表2 可知, 氮磷处理体现出较大差异的是: 磷处理下车桑子株高、基径和生物量均与
LDMC 显著正相关, 而与
SLA 和叶片氮含量显著负相关; 相反, 氮处理下这3项生物生长指标均与根冠比显著负相关, 与叶片氮含量显著正相关, 同时叶片氮含量与根冠比显著负相关。此外, 磷处理下叶茎比与根冠比显著负相关, 叶片氮含量与
SLA 显著正相关, 与
LDMC 显著负相关。无论在氮处理还是磷处理条件下, 车桑子株高、基径和生物量3项生长性状之间呈显著正相关关系,
LDMC 和
SLA 呈极显著负相关关系。同时, 叶片磷含量和N:P均呈显著负相关关系, 说明叶片N:P的变化主要受到磷的影响。但氮处理下, 叶片氮含量和N:P显著正相关, 说明氮处理下N:P的变化也受到氮的影响, 叶片氮磷含量共同调节着车桑子叶片的N:P。
Table 2 表2 表2 氮和磷处理下车桑子幼苗各性状间的相关性
Table 2
Correlations among growth characteristics, biomass allocation and leaf traits of Dodonaea viscosa under nitrogen and phosphorus treatments 处理 指标 生长性状 生物量分配 叶功能性状 株高 基径 生物量 根冠比 叶茎比 叶干物质LDMC 比叶面积SLA 叶片氮含量 叶片磷含量 氮磷比 氮处理 株高 Plant height 1 0.917*** 0.915*** -0.848*** -0.477* 0.404 -0.249 0.840*** 0.180 0.464 基径 Base diameter 1 0.955*** -0.705** -0.380 0.366 -0.205 0.886*** 0.301 0.436 生物量 Biomass 1 -0.773*** -0.282 0.403 -0.269 0.917*** 0.340 0.421 根冠比 1 0.187 -0.339 0.227 -0.720** -0.032 -0.518* 叶茎比 1 -0.206 0.223 -0.234 0.092 -0.217 叶干物质含量 LDMC 1 -0.902*** 0.271 -0.087 0.316 比叶面积 SLA 1 -0.121 0.285 -0.370 叶片氮含量 1 0.282 0.535* 叶片磷含量 1 -0.636** 氮磷比 N:P 1 磷处理Phosphorus treatment 株高 Plant height 1 0.971*** 0.953*** -0.201 -0.404 0.939*** -0.880*** -0.702** -0.175 -0.254 基径 Base diameter 1 0.951*** -0.058 -0.452* 0.908*** -0.862*** -0.649** -0.154 -0.228 生物量 Biomass 1 -0.099 -0.426 0.919*** -0.882*** -0.622* -0.131 -0.244 根冠比 1 -0.528* -0.221 0.073 0.425 -0.026 0.265 叶茎比 1 -0.281 0.331 0.074 0.255 -0.186 叶干物质含量 LDMC 1 -0.945*** -0.765** -0.293 -0.169 比叶面积 SLA 1 0.653** 0.350 0.050 叶片氮含量 1 0.216 0.385 叶片磷含量 1 -0.797*** 氮磷比 N:P 1
LMDC, leaf dry mass content; SLA, specific leaf area. *, p < 0.05; **, p < 0.01; ***, p < 0.001.新窗口打开 |
下载CSV 3 讨论 3.1 车桑子幼苗生长和生物量分配对氮磷浓度的响应特征 幼苗是植物生活史过程中比较脆弱的一个阶段, 也是对环境变化最为敏感的一个阶段, 车桑子幼苗的死亡率较高, 在车桑子种植过程中应加强苗期管护(
张琴等, 2016 )。幼苗生长对植物群落的补员起着关键作用(
李秋艳和赵文智, 2006 )。因此, 本文基于车桑子幼苗的研究对车桑子在植被恢复中的应用具有一定的参考价值。
在植物养分限制的判断中, 若一种有效态元素的添加引起了植物生化过程速率或生物量的增加, 则意味着存在这种养分的限制(
Vitousek et al ., 2010 ;
Sullivan et al ., 2014 )。本研究表明, 高浓度氮水平显著促进了车桑子的生长和生物量积累, 但车桑子生长和生物量积累在低浓度和中等浓度氮水平下没有显著差异。野外条件下, 车桑子生长区的土壤有效氮(硝态氮与铵态氮之和)含量大约为11.7 mg·kg
-1 , 考虑到土壤中水分含量在10%左右, 也就是每kg土壤约含0.1 L水, 那么土壤中的有效氮含量约为117 mg·L
-1 , 介于本研究设置的N3 (210 mg·L
-1 )和N2 (70 mg·L
-1 )水平之间。因此, 本研究中, N1-N3处理下车桑子的生长没有显著差异, 可能是由于干热河谷长期的氮限制使得植物形成了对低氮环境的一系列形态与生理适应性。干旱生态系统氮限制尤为普遍(
Yahdjian et al ., 2011 )。已有研究表明, 氮是限制金沙江干旱河谷退化草地系统恢复的主要因素(
张彦东等, 2004 )。同时, 我们的结果表明, 车桑子主要通过增加对氮的吸收和利用来适应低氮环境。一方面, 车桑子在低氮条件下, 其根冠比显著增加可促进氮的吸收; 另一方面, 吸收的氮的利用效率又显著提高。根冠比的增加和养分利用效率的提高均是植物适应养分限制的主要机制(
冷华妮等, 2009 ;
Venterink & Güsewell, 2010 )。然而, 由于本研究中养分添加只持续了一个生长季, 试验时间较短, 鉴于车桑子幼苗长期对氮限制的适应, 对试验中的低浓度氮水平可能还不太敏感, 通过根冠比的调整和养分利用效率的提高即可保持其正常的生长和生物量积累, 但在长期的低浓度氮水平下车桑子生长和生物量积累可能会受到限制。
在各磷浓度处理下, 车桑子生长和生物量积累随磷浓度的增加而显著增加。野外条件下, 车桑子生长区的土壤有效磷含量为4.90 mg·kg
-1 , 同理换算出来为1.58 mmol·L
-1 , 高于本研究的P3水平, 但低于P4水平。车桑子对野外磷水平的适应进化在一定程度上可以解释本研究低浓度和中等浓度磷处理对车桑子生长的限制作用。然而, 低浓度磷水平下车桑子根冠比并没有显著增加, 表明车桑子不会通过调节根冠比来适应低磷环境, 这与低氮处理结果不同。与我们的假设(1)一致, 车桑子生物量分配对氮变化和磷变化的响应具有差异性。Venterink和Güsewell (2010)认为, 磷限制条件下, 由于根系生长需要消耗的能量较多, 而且生长过程中伴随着细胞分裂, 需要大量的DNA和RNA合成(这两种物质中含有大量的磷), 需要更多的磷。因此, 植物不会大量投资根系来吸收磷离子, 而是通过其他机制促进磷的吸收, 比如磷酸酶活性(
Minden & Venterink, 2019 )、根系分泌物、钙离子吸收(
Devau et al ., 2010 )等。本研究中, 低浓度磷水平下叶/茎生物量比例显著增加, 表明低磷处理增加了地上部分叶生物量的分配。由于茎的生长也需要较多的磷, 在缺磷时, 植物为了保住最关键的功能, 可能会增加叶片投入以维持光合作用, 而减少茎的支出。研究表明, 叶片氮与光合作用直接相关, 氮是光合作用的首要影响因子, 而叶片磷对光合作用的影响较弱(
Reich et al ., 2009 ), 这也反映了车桑子通过维持叶片生长以提高在低磷条件下的适应能力。
需要说明的是, 本研究是基于室内砂培实验条件下得出的结果, 砂培条件和野外自然环境存在较大差异, 该试验结果与野外环境的试验结果是否一致并不确定。研究表明, 丛枝菌根真菌对植物养分吸收尤其是磷的吸收具有重大的贡献(
Smith et al ., 2004 ;
Smith & Smith, 2011 ), 并且, 干热河谷自然植被的根际土壤中大多有丰富的丛枝菌根真菌(
Li et al ., 2010 )。在这种条件下, 我们推测, 车桑子可能已经形成了相关的内部信号调控机制, 主要依赖菌根真菌吸收磷, 而减少了自身对低磷的生理生长调节能力, 使得根系分配不会对磷缺乏形成响应, 这也进一步解释了低磷条件下根冠比没有发生显著增加。而本研究中, 砂培条件缺乏菌根真菌的侵染, 车桑子自身对低磷的适应能力较差, 因此, 低磷水平限制了车桑子生长。在野外土壤环境条件下, 车桑子对氮磷有效性变化的响应特征是否与本研究不同, 尚需进一步验证。
3.2 车桑子幼苗叶片性状对氮磷浓度的响应特征 SLA 和
LDMC 是叶片功能性状中最主要的两个指标, 被认为是指示植物养分利用策略的关键性指标(
Jager et al ., 2015 )。与我们的假设(1)一致, 叶片功能性状对氮变化和磷变化的响应具有差异性。随磷浓度的增加,
SLA 显著降低,
LDMC 显著增加, 而氮处理下却没有相似的结果。前人研究表明,
SLA 随养分的减少而降低, 较低的
SLA 和较高的
LDMC 有利于植物适应资源贫瘠的环境(
Jager et al ., 2015 )。然而, 我们的研究中, 高浓度磷水平降低了
SLA, 增加了
LDMC; 相反, 低浓度磷水平增加了
SLA, 降低了
LDMC 。高磷水平下低
SLA 、高
LDMC 表明车桑子采取养分的有效保存策略, 低磷水平下高
SLA 、低
LDMC 可提高对光资源的获取能力, 表明车桑子采取快速生长的策略, 该研究结果与
周光霞等(2016) 的研究相似。然而, 低磷水平下车桑子生长依然受到严重的限制。
林婉奇等(2019) 也发现磷添加处理减少了
SLA, 但显著促进了幼苗生长。叶面积和
SLA 呈负相关关系, 叶面积增加可提高植物的光获取能力, 弥补因
SLA 减小而导致的碳摄取效率的损失(
陈延松等, 2019 )。因此, 高磷水平下叶面积的增加对促进车桑子生长和生物量积累具有重要作用。
本研究中, 不同的氮浓度和磷浓度处理显著影响了车桑子叶片氮含量, 但叶片磷含量和叶片N:P并未受到显著影响。由于植物具有保持自身化学组成相对稳定的能力, 环境中的养分变化对植物氮磷元素的影响也取决于植物化学计量内稳性的强弱(
Persson et al ., 2010 )。植物内稳性特征可用内稳性指数
H 进行界定, 根据内稳性指数可将植物分为四类:
H > 4为稳态型; 2 <
H < 4为弱稳态型; 4/3 <
H < 2为弱敏感型;
H < 4/3为敏感型(
Persson et al ., 2010 )。本研究中, 不同氮浓度处理下, 叶片氮含量和叶片N:P的
H 分别为6.86和7.31, 均大于4, 呈稳态型, 并且叶片N:P的内稳性大于氮的内稳性。其他研究也表明, 氮磷比的内稳性大于氮和磷的内稳性(
Yu et al ., 2011 ;
宁志英等, 2019 )。这说明, 植物对其自身内稳性的调节主要体现在N:P而不是单一的元素含量上。然而, 不同磷浓度处理下, 叶片磷含量、叶片N:P与营养液的磷浓度和营养液N:P并没有显著的幂函数关系, 其内稳性指数计算结果为负值。
Xing等(2015) 用1/
H 来衡量内稳性的强弱, 也测出内稳性指数为负值, 并将其归为严格内稳型。这说明磷处理条件下, 车桑子叶片磷含量和N:P具有极强的内稳性。较高的内稳性意味着养分元素变化小, 表明具有保守的养分利用策略, 也就更容易适应贫瘠环境而成为优势种(
Yu et al ., 2010 )。不同生长期植物内稳性不同, 内稳性随着植物生长发育逐渐增强(
Yu et al ., 2011 )。本研究中车桑子幼苗已表现出较高的内稳性, 因此, 这可能是促进车桑子成为干热河谷优势植物的原因之一。此外, 所有处理条件下, 车桑子的叶片N:P均在30左右波动。按照叶片临界N:P对限制元素的判断理论, 车桑子生长受到磷限制(
Güsewell, 2004 ;
Sullivan et al ., 2014 ), 这和氮磷添加对车桑子生长和生物量积累的结果是不一致的。与前人的研究观点(
Drenovsky & Richards, 2004 ;
Zhang et al ., 2004 )比较一致, 我们的结果也表明, 在物种水平上, 用叶片N:P来指示植物所受到的养分限制类型是不可靠的。
车桑子叶片氮利用效率随营养液N:P的增加而降低。研究表明, 氮磷养分供应量对植物生长起着主要作用, 而除此之外, 氮磷供给比也影响着植物生长(
Güsewell & Bollens, 2003 )。本研究中, N4 (30 mmol·L
-1 N、1 mmol·L
-1 P)和P2 (15 mmol·L
-1 N、0.5 mmol·L
-1 P)处理的氮磷供给比均为30, 但P2处理的车桑子生长和生物量积累显著低于N4处理, 而氮利用效率显著高于N4处理。车桑子其他生长指标、生物量分配、叶片性状和磷利用效率比较一致, 随营养液氮磷比的变化并没有发生规律性变化。因此, 我们的结果表明, 氮磷的供应量而不是单纯的氮磷供给比对植物性状和生长的影响更大。
3.3 车桑子幼苗各性状间的相关性 通过前面的分析已经发现, 叶片功能性状在不同的氮浓度和磷浓度处理下具有差异性响应, 那么, 这种差异性响应是否会改变性状间的相关性, 进而改变植物的权衡策略? 结果表明, 不同的氮处理和磷处理下, 车桑子叶片功能性状间的相关性既表现出一定的一致性, 也有一定的差异性。不论是在氮处理还是磷处理下,
LDMC 和
SLA 呈显著负相关关系, 与其他研究结果(
安慧, 2012 )一致, 即: 随着
LDMC 的增加, 植物叶片含水率降低, 叶片组织密度增加, 从而导致
SLA 降低。这说明
LDMC 和
SLA 之间的相关性较为稳定, 具有普遍性意义。同时, 两种处理条件下, 车桑子叶片磷含量与叶片N:P呈极显著负相关
关系, 可见车桑子叶片磷含量的变化是驱动N:P变化的主要因子。
吴统贵等(2010) 和
宋彦涛等(2012) 的研究也表明, 叶片N:P主要受到磷的驱动, N:P的变化受控于叶片磷含量的变化。然而, 氮、磷处理条件下, 车桑子叶片氮和磷含量并没有显著正相关关系, 这可能是由于氮、磷养分不同的内稳性导致的。
黄端等(2017) 也发现叶片氮、磷含量之间无显著的正相关关系。此外, 本研究中, 磷处理下叶片氮含量和
SLA 显著正相关, 而氮处理下却没有这种相关性, 这与以往的研究结果不一致。以往大量的研究发现植物叶片
SLA 与叶片氮含量显著正相关, 二者的关联是植物叶经济型谱的重要组成部分(
于鸿莹等, 2014 ;
熊静等, 2019 )。以往的数据基本是从多个物种之间发现的, 也就是
SLA 与叶片氮含量正相关是种间变异的结果,而本研究是通过设置氮浓度梯度, 是基于环境中氮浓度变化对单个物种内
SLA 与叶片氮含量影响后的结果。在氮有效性增加的情况下, 叶片氮含量的响应更直接更迅速, 植物吸收氮后叶片氮含量显著增加, 而
SLA 的响应相对滞后, 与
Wright和Sutton-Grier (2012) 在氮添加条件下的结果是一致的, 这也反映了长期的氮限制使车桑子在较高的氮水平下短期内依然保持相对保守的氮利用策略。因此, 氮添加条件下车桑子叶片氮含量与
SLA 不相关甚至呈现出较弱的负相关关系。本研究结果表明, 环境中养分驱动下单个物种内叶片的性状变化可能并不完全符合经济型谱理论, 而会受到氮、磷养分有效性的影响, 符合叶经济型谱形成的环境因子主导假说(
赖小红等, 2019 )。此外, 植物功能性状和生物量分配之间存在着某种共轭的变化(
高景等, 2016 )。本研究中, 氮处理下, 车桑子根冠比与叶片氮含量、N:P显著负相关, 磷处理下却没有显著的相关性。以上结果支持我们假设(2)的观点, 车桑子生物量分配与叶性状间存在着一定的协同和权衡关系, 但是车桑子在面临氮、磷有效性的变化时具有不同的响应策略。
从车桑子生长特征与生物量分配、叶性状的相关性来看, 氮处理条件下, 株高、基径、生物量等生长性状均与根冠比呈显著负相关关系, 与叶片氮含量显著正相关, 但生长性状与叶片氮含量的相关性更强, 表明叶片氮含量的调整对车桑子适应低氮具有重要意义; 而磷处理下, 株高、基径、生物量与
SLA 、叶片氮含量显著负相关, 与
LDMC 显著正相关, 表明叶片功能性状的调整在车桑子适应低磷环境中发挥重要作用。
周俊宏等(2015) 针对紫楠(
Phoebe sheareri )幼苗的研究表明, 相比叶性状的调整, 紫楠幼苗主要通过生物量分配比例的改变来适应不同生境的变化。我们的研究与之相反, 无论氮处理还是磷处理条件下, 相比生物量分配, 叶片性状的调整是车桑子对养分有效性变化更为主要的适应策略。生物量分配的变化也是一系列植物性状对环境变化适应的结果(
Mccarthy & Enquist, 2007 )。因此, 叶性状的调整是车桑子对养分变化更为主要的适应策略。我们的结果表明, 车桑子在响应氮、磷限制时会采用不同的适应策略, 表征其适应策略的性状及性状间的权衡策略亦有所差异。然而, 本研究是基于短期试验条件下的单因素控制试验, 由于氮、磷之间存在交互作用, 不同组合的氮磷添加对车桑子幼苗生长性状的影响值得进一步研究。
4 结论 以往的研究主要关注养分有效性对植物生理生长的影响, 但忽略了植物在氮有效性和磷有效性变化时的响应差异。本研究发现, 车桑子生物量分配和叶片功能性状对氮变化和磷变化的可塑性响应具有差异性。车桑子幼苗主要通过叶片氮含量和根冠比的调整适应氮有效性的变化, 在低浓度氮水平下, 通过增加根冠比和提高氮利用效率, 能够在低浓度氮水平下保持稳定生长。而在磷处理条件下, 车桑子幼苗主要通过叶/茎生物量比、
SLA 和
LDMC 的调整适应磷有效性的变化, 但低浓度磷水平限制了车桑子幼苗的生长和生物量积累。相比于生物量分配, 叶性状与车桑子生长的相关性更强, 叶性状的响应是车桑子适应氮变化和磷变化更为主要的策略。同时, 车桑子生物量分配和叶片功能性状之间存在着一定的相互变化关系, 但这种相关关系受到氮磷养分的影响。本研究从生物量分配和叶片功能性状的角度, 系统研究了车桑子生物量分配和叶性状及其相互关系对氮、磷浓度的响应特征, 这对更全面地揭示植物对养分有效性变化的适应与权衡具有重要意义。另外, 由于氮浓度和磷浓度下植物性状趋异分化, 未来在探讨养分有效性对植物生长的影响时, 应明确说明是氮养分有效性还是磷养分有效性。
致谢 感谢绵阳师范学院生态安全与保护四川省重点实验室开放基金项目(ESP1902)资助。感谢云南省农业科学院热区生态农业研究所何光熊、李建查等人在实验过程中的无私帮助。
[1] An H 2012 ). Effects of grazing disturbance on leaf traits and their interrelationships of plants in desert steppeChinese Journal of Applied Ecology 2991 -2996 . URL PMID:
23431780 [本文引用: 1] [ 安慧 (2012 ). 放牧干扰对荒漠草原植物叶性状及其相互关系的影响应用生态学报 , 23, 2991 -2996 .] PMID:
23431780 [本文引用: 1] [2] Chen SY Lang NJ Jia LQ Wu LY 2006 ). Effects of drought stress on lipid peroxidation and activity of defense enzymes of Dodonaea viscosa, Leucaena leucocephala and Tephrosia candida seedlingsBulletin of Botanical Research 88 -92 . [本文引用: 1] [ 陈少瑜 , 郎南军 , 贾利强 , 吴丽圆 (2006 ). 干旱胁迫对坡柳等抗旱树种幼苗膜脂过氧化及保护酶活性的影响植物研究 , 26, 88 -92 .] [本文引用: 1] [3] Chen YS Li LL Zhou SB Ou ZL 2019 ). The differentiation of plant functional traits in genus Lysimachia LChinese Journal of Ecology 1653 -1661 . [本文引用: 1] [ 陈延松 , 李玲玲 , 周守标 , 欧祖兰 (2019 ). 珍珠菜属植物功能性状的趋异分化生态学杂志 , 38, 1653 -1661 .] [本文引用: 1] [4] da Silva EC Nogueira RJMC da Silva MA de Albuquerque MB 2011 ). Drought stress and plant nutritionPlant Stress 32 -41 . [本文引用: 1] [5] Devau N Le Cadre E Hinsinger P Gérard F 2010 ). A mechanistic model for understanding root-induced chemical changes controlling phosphorus availabilityAnnals of Botany 1183 -1197 . URL PMID:
20495198 [本文引用: 1] [6] Drenovsky RE Richards JH 2004 ). Critical N:P values: predicting nutrient deficiencies in desert shrublandsPlant and Soil 59 -69 . [本文引用: 1] [7] Gao J Wang JN Xu B Xie Y He JD Wu Y 2016 ). Plant leaf traits, height and biomass partitioning in typical ephemerals under different levels of snow cover thickness in an alpine meadowChinese Journal of Plant Ecology 775 -787 . [本文引用: 1] [ 高景 , 王金牛 , 徐波 , 谢雨 , 贺俊东 , 吴彦 (2016 ). 不同雪被厚度下典型高山草地早春植物叶片性状、株高及生物量分配的研究植物生态学报 , 40, 775 -787 .] [本文引用: 1] [8] Güsewell S 2004 ). N:P ratios in terrestrial plants: variation and functional significanceNew Phytologist 243 -266 . [本文引用: 1] [9] Güsewell S Bollens U 2003 ). Composition of plant species mixtures grown at various N:P ratios and levels of nutrient supplyBasic and Applied Ecology 453 -466 . [本文引用: 1] [10] Han X Tsunekawa A Tsubo M Shao H 2013 ). Responses of plant-soil properties to increasing N deposition and implications for large-scale eco-restoration in the semiarid grassland of the northern Loess Plateau, ChinaEcological Engineering 1 -9 . [本文引用: 1] [11] Hoagland DR Arnon DI 1938 ). The Water Culture Method for Growing Plants Without Soil California Agricultural Experiment Station , 347, 1 -37 . [2020 -06-18]. https:// www.researchgate.net/file.PostFileLoader.html?id=54aefd7ed4c118b6358b45db&assetKey=AS%3A273668901408776%401442259158553. URL [本文引用: 1] [12] Huang D Wang DM Ren Y Qin YB Wu LC 2017 ). Responses of leaf traits to submergence stress and analysis of the economic spectrum of plant species in an aquatic- terrestrial ecotone, the Li RiverActa Ecologica Sinica 750 -759 . [本文引用: 1] [ 黄端 , 王冬梅 , 任远 , 覃云斌 , 吴林川 (2017 ). 漓江水陆交错带植物叶性状对水淹胁迫的响应及经济谱分析生态学报 , 37, 750 -759 .] [本文引用: 1] [13] Jager MM Richardson SJ Bellingham PJ Clearwater MJ Laughlin DC 2015 ). Soil fertility induces coordinated responses of multiple independent functional traitsJournal of Ecology 374 -385 . [本文引用: 2] [14] Lai XH Wang HY Zhong YH Lin L Li MY 2019 ). The impact of short-term NO2 pollution on leaf functional traits of nine landscape plant speciesActa Ecologica Sinica 8058 -8067 . [本文引用: 1] [ 赖小红 , 王海洋 , 钟雨航 , 林立 , 李名扬 (2019 ). 人工控制条件下9种园林植物叶功能性状对短期NO2 污染的响应生态学报 , 39, 8058 -8067 .] [本文引用: 1] [15] Leng HN Chen YT Rao LB Duan HP 2009 ). Effects of N rate on seedling growth and uptake and utilization of nitrogen and phosphorus in different provenances of Liquidambar formosana HanceJournal of Soil and Water Conservation 79 -84 . [本文引用: 2] [ 冷华妮 , 陈益泰 , 饶龙兵 , 段红平 (2009 ). 供氮水平对枫香种源幼苗生长及氮、磷吸收利用的影响水土保持学报 , 23, 79 -84 .] [本文引用: 2] [16] Li JY Jia LQ Lang NJ Chen SY Wu LY 2003 ). Characteristics of photosynthesis of Dodonaea viscosa in the dry-hot valley of Jinsha RiverJournal of Beijing Forestry University 5 ), 20 -24 . [本文引用: 1] [ 李吉跃 , 贾利强 , 郎南军 , 陈少瑜 , 吴丽圆 (2003 ). 金沙江干热河谷坡柳的光合特性北京林业大学学报 , 25(5 ), 20 -24 .] [本文引用: 1] [17] Li QY Zhao WZ 2006 ). Seedling emergence and growth responses of five desert species to sand burial depthActa Ecologica Sinica 1802 -1808 . [本文引用: 1] [ 李秋艳 , 赵文智 (2006 ). 五种荒漠植物幼苗出土及生长对沙埋深度的响应生态学报 , 26, 1802 -1808 .] [本文引用: 1] [18] Li T Li LF Sha T Zhang HB Zhao ZW 2010 ). Molecular diversity of arbuscular mycorrhizal fungi associated with two dominant xerophytes in a valley-type savanna, southwest ChinaApplied Soil Ecology 61 -66 . [本文引用: 1] [19] Lin WQ Cai JH Xue L 2019 ). Responses of Cinnamomum camphora seedling growth and leaf traits to additions of nitrogen and phosphorous under different planting densitiesActa Ecologica Sinica 6738 -6744 . [本文引用: 2] [ 林婉奇 , 蔡金桓 , 薛立 (2019 ). 不同密度樟树(Cinnamomum camphora )幼苗生长和叶片性状对氮磷添加的响应生态学报 , 39, 6738 -6744 .] [本文引用: 2] [20] Mayor JR Wright SJ Turner BL 2014 ). Species-specific responses of foliar nutrients to long-term nitrogen and phosphorus additions in a lowland tropical forestJournal of Ecology 36 -44 . [本文引用: 1] [21] Mccarthy MC Enquist BJ 2007 ). Consistency between an allometric approach and optimal partitioning theory in global patterns of plant biomass allocationFunctional Ecology 713 -720 . [本文引用: 2] [22] Menge DNL Hedin LO Pacala SW 2012 ). Nitrogen and phosphorus limitation over long-term ecosystem development in terrestrial ecosystemsPLOS ONE e42045 . DOI: 10. 1371/journal.pone.0042045 . URL PMID:
22870281 [本文引用: 1] [23] Minden V Venterink HO 2019 ). Plant traits and species interactions along gradients of N, P and K availabilitiesFunctional Ecology 1611 -1626 . [本文引用: 2] [24] Ning ZY Li YL Yang HL Zhang ZQ 2019 ). Nitrogen and phosphorus stoichiometric homoeostasis in leaves of dominant sand-fixing shrubs in Horqin Sandy Land, ChinaChinese Journal of Plant Ecology 46 -54 . [本文引用: 1] [ 宁志英 , 李玉霖 , 杨红玲 , 张子谦 (2019 ). 科尔沁沙地优势固沙灌木叶片氮磷化学计量内稳性植物生态学报 , 43, 46 -54 .] [本文引用: 1] [25] Ordo?ez JC van Bodegom PM Witte JPM Wright IJ Reich PB Aerts R 2009 ). A global study of relationships between leaf traits, climate and soil measures of nutrient fertilityGlobal Ecology and Biogeography 137 -149 . [本文引用: 1] [26] Persson J Fink P Goto A Hood JM Jonas J Kato S 2010 ). To be or not to be what you eat: regulation of stoichiometric homeostasis among autotrophs and heterotrophsOikos 741 -751 . [本文引用: 2] [27] Poorter H Niklas KJ Reich PB Oleksyn J Poot P Mommer L 2012 ). Biomass allocation to leaves, stems and roots: meta-analyses of interspecific variation and environmental controlNew Phytologist 30 -50 . [本文引用: 1] [28] Qi J Ma KM Zhang YX 2009 ). Leaf-trait relationships of Quercus liaotungensis along an altitudinal gradient in Dongling Mountain, BeijingEcological Research 1243 -1250 . [本文引用: 1] [29] Reich PB Oleksyn J Wright IJ 2009 ). Leaf phosphorus influences the photosynthesis-nitrogen relation: a cross- biome analysis of 314 speciesOecologia 207 -212 . DOI:
10.1007/s00442-009-1291-3 URL PMID:
19212782 [本文引用: 1] The ecophysiological linkage of leaf phosphorus (P) to photosynthetic capacity (A (max)) and to the A (max)-nitrogen relation remains poorly understood. To address this issue we compiled published and unpublished field data for mass-based A (max), nitrogen (N) and P (n = 517 observations) from 314 species at 42 sites in 14 countries. Data were from four biomes: arctic, cold temperate, subtropical (including Mediterranean), and tropical. We asked whether plants with low P levels have low A (max), a shallower slope of the A (max)-N relationship, and whether these patterns have a geographic signature. On average, leaf P was substantially lower in the two warmer than in the two colder biomes, with the reverse true for N:P ratios. The evidence indicates that the response of A (max) to leaf N is constrained by low leaf P. Using a full factorial model for all data, A (max) was related to leaf N, but not to leaf P on its own, with a significant leaf N x leaf P interaction indicating that the response of A (max) to N increased with increasing leaf P. This was also found in analyses using one value per species per site, or by comparing only angiosperms or only woody plants. Additionally, the slope of the A (max)-N relationship was higher in the colder arctic and temperate than warmer tropical and subtropical biomes. Sorting data into low, medium, and high leaf P groupings also showed that the A (max)-N slope increases with leaf P. These analyses support claims that in P-limited ecosystems the A (max)-N relationship may be constrained by low P, and are consistent with laboratory studies that show P-deficient plants have limited ribulose-1,5-bisphosphate regeneration, a likely mechanism for the P influence upon the A (max)-N relation.
[30] Smith SE Smith FA 2011 ). Roles of arbuscular mycorrhizas in plant nutrition and growth: new paradigms from cellular to ecosystem scalesAnnual Review of Plant Biology 227 -250 . [本文引用: 1] [31] Smith SE Smith FA Jakobsen I 2004 ). Functional diversity in arbuscular mycorrhizal (AM) symbioses: the contribution of the mycorrhizal P uptake pathway is not correlated with mycorrhizal responses in growth or total P uptakeNew Phytologist 511 -524 . [本文引用: 1] [32] Song YT Zhou DW Li Q Wang P Huang YX 2012 ). Leaf nitrogen and phosphorus stoichiometry in 80 herbaceous plant species of Songnen grassland in Northeast ChinaChinese Journal of Plant Ecology 222 -230 . [ 宋彦涛 , 周道玮 , 李强 , 王平 , 黄迎新 (2012 ). 松嫩草地80种草本植物叶片氮磷化学计量特征植物生态学报 , 36, 222 -230 .] [33] Sullivan BW Alvarez-Clare S Castle SC Porder S Reed SC Schreeg L Townsend AR Cleveland CC 2014 ). Assessing nutrient limitation in complex forested ecosystems: alternatives to large-scale fertilization experimentsEcology 668 -681 . DOI:
10.1890/13-0825.1 URL PMID:
24804451 [本文引用: 2] Quantifying nutrient limitation of primary productivity is a fundamental task of terrestrial ecosystem ecology, but in a high carbon dioxide environment it is even more critical that we understand potential nutrient constraints on plant growth. Ecologists often manipulate nutrients with fertilizer to assess nutrient limitation, yet for a variety of reasons, nutrient fertilization experiments are either impractical or incapable of resolving ecosystem responses to some global changes. The challenges of conducting large, in situ fertilization experiments are magnified in forests, especially the high-diversity forests common throughout the lowland tropics. A number of methods, including fertilization experiments, could be seen as tools in a toolbox that ecologists may use to attempt to assess nutrient limitation, but there has been no compilation or synthetic discussion of those methods in the literature. Here, we group these methods into one of three categories (indicators of soil nutrient supply, organismal indicators of nutrient limitation, and lab-based experiments and nutrient depletions), and discuss some of the strengths and limitations of each. Next, using a case study, we compare nutrient limitation assessed using these methods to results obtained using large-scale fertilizations across the Hawaiian Archipelago. We then explore the application of these methods in high-diversity tropical forests. In the end, we suggest that, although no single method is likely to predict nutrient limitation in all ecosystems and at all scales, by simultaneously utilizing a number of the methods we describe, investigators may begin to understand nutrient limitation in complex and diverse ecosystems such as tropical forests. In combination, these methods represent our best hope for understanding nutrient constraints on the global carbon cycle, especially in tropical forest ecosystems.
[34] Venterink HO Güsewell S 2010 ). Competitive interactions between two meadow grasses under nitrogen and phosphorus limitationFunctional Ecology 877 -886 . [本文引用: 3] [35] Vitousek PM Howarth RW 1991 ). Nitrogen limitation on land and in the sea: How can it occur?Biogeochemistry 87 -115 . [本文引用: 1] [36] Vitousek PM Porder S Houlton BZ Chadwick OA 2010 ). Terrestrial phosphorus limitation: mechanisms, implications, and nitrogen-phosphorus interactionsEcological Applications 5 -15 . URL PMID:
20349827 [本文引用: 2] [37] Wang FP Shi GX Nicholas O Yao BQ Ji MF Wang WY Ma Z Zhou HK Zhao XQ 2018 ). Ecosystem nitrogen retention is regulated by plant community trait interactions with nutrient status in an alpine meadowJournal of Ecology 1570 -1581 . [本文引用: 1] [38] Wright IJ Reich PB Westoby M Ackerly DD Baruch Z Bongers F Cavender-Bares J Chapin T Cornelissen JHC Diemer M Flexas J Garnier E Groom PK Gulias J Hikosaka K et al 2004 ). The worldwide leaf economics spectrumNature 821 -827 . URL PMID:
15103368 [本文引用: 1] [39] Wright JP Sutton-Grier A 2012 ). Does the leaf economic spectrum hold within local species pools across varying environmental conditions?Functional Ecology 1390 -1398 . [本文引用: 1] [40] Wu TG Wu M Liu L Xiao JH 2010 ). Seasonal variations of leaf nitrogen and phosphorus stoichiometry of three herbaceous species in Hangzhou Bay coastal wetlands, ChinaChinese Journal of Plant Ecology 23 -28 . [本文引用: 1] [ 吴统贵 , 吴明 , 刘丽 , 萧江华 (2010 ). 杭州湾滨海湿地3种草本植物叶片N、P化学计量学的季节变化植物生态学报 , 34, 23 -28 .] [本文引用: 1] [41] Xing W Wu HP Shi Q Hao BB Liu H Wang ZX Liu GH 2015 ). Multielement stoichiometry of submerged macrophytes across Yunnan plateau lakes (China)Scientific Reports 10186 . DOI: 10.1038/srep10186 . URL PMID:
25970822 [本文引用: 1] [42] Xiong J Xing WL Yu MK Cheng XR 2019 ). Variations in leaf traits of native and introduced tree species in Cunninghamia lanceolata plantationsActa Ecologica Sinica 1897 -1907 . [本文引用: 2] [ 熊静 , 邢文黎 , 虞木奎 , 成向荣 (2019 ). 杉木人工林下原生与引入树种叶性状变异特征生态学报 , 39, 1897 -1907 .] [本文引用: 2] [43] Xu ZW Li MH Zimmermann NE Li SP Li H Ren HY Sun H Han XG Jiang Y Jiang L 2018 ). Plant functional diversity modulates global environmental change effects on grassland productivityJournal of Ecology 1941 -1951 . [本文引用: 1] [44] Yahdjian L Gherardi L Sala OE 2011 ). Nitrogen limitation in arid-subhumid ecosystems: a meta-analysis of fertilization studiesJournal of Arid Environments 675 -680 . [本文引用: 1] [45] Yan BG He GX Li JC Ji ZH 2012 ). Scaling relationships and stoichiometry of plant leaf biogenic elements from the arid-hot valley of Jinsha River, ChinaChinese Journal of Plant Ecology 1136 -1144 . [本文引用: 1] [ 闫帮国 , 何光熊 , 李纪潮 , 纪中华 (2012 ). 金沙江干热河谷地区植物叶片中各生源要素的化学计量特征以及异速增长关系植物生态学报 , 36, 1136 -1144 .] [本文引用: 1] [46] Yan BG Ji ZH Fan B Wang XM He GX Shi LT Liu GC 2016 ). Plants adapted to nutrient limitation allocate less biomass into stems in an arid-hot grasslandNew Phytologist 1232 -1240 . [本文引用: 1] [47] Yan ZB Kim N Han WX Guo YL Han TS Du EZ Fang JY 2015 ). Effects of nitrogen and phosphorus supply on growth rate, leaf stoichiometry, and nutrient resorption of Arabidopsis thaliana Plant and Soil 147 -155 . [本文引用: 2] [48] Yu HY Chen YT Xu ZZ Zhou GS 2014 ). Analysis of relationships among leaf functional traits and economics spectrum of plant species in the desert steppe of Nei MongolChinese Journal of Plant Ecology 1029 -1040 . [本文引用: 2] [ 于鸿莹 , 陈莹婷 , 许振柱 , 周广胜 (2014 ). 内蒙古荒漠草原植物叶片功能性状关系及其经济谱分析植物生态学报 , 38, 1029 -1040 .] [本文引用: 2] [49] Yu Q Chen QS Elser JJ He NP Wu HH Zhang GM Wu JG Bai YF Han XG 2010 ). Linking stoichiometric homoeostasis with ecosystem structure, functioning and stabilityEcology Letters 1390 -1399 . URL PMID:
20849443 [本文引用: 1] [50] Yu Q Elser JJ He NP Wu HH Chen QS Zhang GM Han XG 2011 ). Stoichiometric homeostasis of vascular plants in the Inner Mongolia grasslandOecologia 1 -10 . URL PMID:
21221646 [本文引用: 5] [51] Zhang DY Peng YF Li F Yang GB Wang J Yu JC Zhou GY Yang YH 2019 ). Trait identity and functional diversity co-drive response of ecosystem productivity to nitrogen enrichmentJournal of Ecology 2402 -2414 . [本文引用: 1] [52] Zhang LX Bai YF Han XG 2004 ). Differential responses of N:P stoichiometry of Leymus chinensis and Carex korshinskyi to N additions in a steppe ecosystem in Nei MongolActa Botanica Sinica 259 -270 . [本文引用: 1] [53] Zhang Q Liu LW Li JQ Wang CH 2016 ). Characteristics of Dodonaea viscosa population regenerated in abandoned cropland of Yunnan dry-hot valleysSouthwest China Journal of Agricultural Sciences 2234 -2238 . [本文引用: 1] [ 张琴 , 刘利文 , 李俊清 , 王传华 (2016 ). 云南干热河谷区裸地植被恢复过程中车桑子的种群特征西南农业学报 , 29, 2234 -2238 .] [本文引用: 1] [54] Zhang YD Shen YX Liu WY 2004 ). Fertilization effects of N, P on a grass community at the dryvalley of Jinsha RiverBulletin of Botanical Research 59 -64 . [本文引用: 1] [ 张彦东 , 沈有信 , 刘文耀 (2004 ). 金沙江干旱河谷退化草地群落对氮磷施肥的反应植物研究 , 24, 59 -64 .] [本文引用: 1] [55] Zhang YP Zhao FX Liu XX Zheng YX He B Wei Y 2005 ). Relationship between growth increment of Azadirachta indica and site condition in hot and arid valleyForest Research 74 -79 . [本文引用: 1] [ 张燕平 , 赵粉侠 , 刘秀贤 , 郑益兴 , 贺斌 , 韦宇 (2005 ). 干热河谷印楝生长与立地条件关系林业科学研究 , 18, 74 -79 .] [本文引用: 1] [56] Zhou GX Huang LX Zang XW Wei X Ye WH Shen H 2016 ). Effects of habitat heterogeneity on community functional diversity of Dinghu Mountain evergreen broad- leaved forestGuihaia 127 -136 . [本文引用: 1] [ 周光霞 , 黄立新 , 臧晓蔚 , 韦霄 , 叶万辉 , 沈浩 (2016 ). 生境异质性对鼎湖山常绿阔叶林群落功能多样性的影响广西植物 , 36, 127 -136 .] [本文引用: 1] [57] Zhou JH Cheng XR Yu MK Zhuge JJ 2015 ). Responses of growth, leaf traits and biomass allocation of Phoebe sheareri seedlings to different habitats in forest gapJournal of Northeast Forestry University 12 ), 21 -24 . [本文引用: 1] [ 周俊宏 , 成向荣 , 虞木奎 , 诸葛建军 (2015 ). 紫楠幼苗生长、叶性状和生物量分配对林窗不同生境的响应东北林业大学学报 , 43(12 ), 21 -24 .] [本文引用: 1] 放牧干扰对荒漠草原植物叶性状及其相互关系的影响
1
2012
... 通过前面的分析已经发现, 叶片功能性状在不同的氮浓度和磷浓度处理下具有差异性响应, 那么, 这种差异性响应是否会改变性状间的相关性, 进而改变植物的权衡策略? 结果表明, 不同的氮处理和磷处理下, 车桑子叶片功能性状间的相关性既表现出一定的一致性, 也有一定的差异性.不论是在氮处理还是磷处理下,
LDMC 和
SLA 呈显著负相关关系, 与其他研究结果(
安慧, 2012 )一致, 即: 随着
LDMC 的增加, 植物叶片含水率降低, 叶片组织密度增加, 从而导致
SLA 降低.这说明
LDMC 和
SLA 之间的相关性较为稳定, 具有普遍性意义.同时, 两种处理条件下, 车桑子叶片磷含量与叶片N:P呈极显著负相关 ...
放牧干扰对荒漠草原植物叶性状及其相互关系的影响
1
2012
... 通过前面的分析已经发现, 叶片功能性状在不同的氮浓度和磷浓度处理下具有差异性响应, 那么, 这种差异性响应是否会改变性状间的相关性, 进而改变植物的权衡策略? 结果表明, 不同的氮处理和磷处理下, 车桑子叶片功能性状间的相关性既表现出一定的一致性, 也有一定的差异性.不论是在氮处理还是磷处理下,
LDMC 和
SLA 呈显著负相关关系, 与其他研究结果(
安慧, 2012 )一致, 即: 随着
LDMC 的增加, 植物叶片含水率降低, 叶片组织密度增加, 从而导致
SLA 降低.这说明
LDMC 和
SLA 之间的相关性较为稳定, 具有普遍性意义.同时, 两种处理条件下, 车桑子叶片磷含量与叶片N:P呈极显著负相关 ...
干旱胁迫对坡柳等抗旱树种幼苗膜脂过氧化及保护酶活性的影响
1
2006
... 车桑子(
Dodonaea viscosa )又名坡柳、明油子, 具有耐干旱、耐贫瘠和适应性强等特性, 是我国干热河谷区的优势物种, 也是该区植被恢复工程中的常见植物, 具有重要的生态价值.关于车桑子的研究, 目前重点研究了车桑子对干旱胁迫的响应特征(
李吉跃等, 2003 ;
陈少瑜等, 2006 ).但除了干旱胁迫之外, 养分胁迫成为干旱胁迫的次生效应(
da Silva et al ., 2011 ).加之该区土壤侵蚀严重, 干热河谷土壤(燥红土)普遍存在缺氮少磷的特点(
张燕平等, 2005 ), 严重影响了干热河谷植物的生长(
闫帮国等, 2012 ).然而, 关于车桑子性状对氮磷限制的响应目前尚不清楚.因此, 本研究以车桑子幼苗为研究对象, 采用砂培的方式进行试验, 研究车桑子幼苗生长特征、生物量分配、叶片功能性状及其相互关系对氮磷梯度的响应特征, 从生物量分配和叶片功能性状的角度探讨车桑子对低氮和低磷条件的适应性, 并验证以下假说: (1)车桑子幼苗生物量分配和叶片功能性状对氮变化和磷变化的响应具有差异性; (2)生物量分配和叶性状对氮磷浓度响应存在协同或权衡关系. ...
干旱胁迫对坡柳等抗旱树种幼苗膜脂过氧化及保护酶活性的影响
1
2006
... 车桑子(
Dodonaea viscosa )又名坡柳、明油子, 具有耐干旱、耐贫瘠和适应性强等特性, 是我国干热河谷区的优势物种, 也是该区植被恢复工程中的常见植物, 具有重要的生态价值.关于车桑子的研究, 目前重点研究了车桑子对干旱胁迫的响应特征(
李吉跃等, 2003 ;
陈少瑜等, 2006 ).但除了干旱胁迫之外, 养分胁迫成为干旱胁迫的次生效应(
da Silva et al ., 2011 ).加之该区土壤侵蚀严重, 干热河谷土壤(燥红土)普遍存在缺氮少磷的特点(
张燕平等, 2005 ), 严重影响了干热河谷植物的生长(
闫帮国等, 2012 ).然而, 关于车桑子性状对氮磷限制的响应目前尚不清楚.因此, 本研究以车桑子幼苗为研究对象, 采用砂培的方式进行试验, 研究车桑子幼苗生长特征、生物量分配、叶片功能性状及其相互关系对氮磷梯度的响应特征, 从生物量分配和叶片功能性状的角度探讨车桑子对低氮和低磷条件的适应性, 并验证以下假说: (1)车桑子幼苗生物量分配和叶片功能性状对氮变化和磷变化的响应具有差异性; (2)生物量分配和叶性状对氮磷浓度响应存在协同或权衡关系. ...
珍珠菜属植物功能性状的趋异分化
1
2019
...
SLA 和
LDMC 是叶片功能性状中最主要的两个指标, 被认为是指示植物养分利用策略的关键性指标(
Jager et al ., 2015 ).与我们的假设(1)一致, 叶片功能性状对氮变化和磷变化的响应具有差异性.随磷浓度的增加,
SLA 显著降低,
LDMC 显著增加, 而氮处理下却没有相似的结果.前人研究表明,
SLA 随养分的减少而降低, 较低的
SLA 和较高的
LDMC 有利于植物适应资源贫瘠的环境(
Jager et al ., 2015 ).然而, 我们的研究中, 高浓度磷水平降低了
SLA, 增加了
LDMC; 相反, 低浓度磷水平增加了
SLA, 降低了
LDMC .高磷水平下低
SLA 、高
LDMC 表明车桑子采取养分的有效保存策略, 低磷水平下高
SLA 、低
LDMC 可提高对光资源的获取能力, 表明车桑子采取快速生长的策略, 该研究结果与
周光霞等(2016) 的研究相似.然而, 低磷水平下车桑子生长依然受到严重的限制.
林婉奇等(2019) 也发现磷添加处理减少了
SLA, 但显著促进了幼苗生长.叶面积和
SLA 呈负相关关系, 叶面积增加可提高植物的光获取能力, 弥补因
SLA 减小而导致的碳摄取效率的损失(
陈延松等, 2019 ).因此, 高磷水平下叶面积的增加对促进车桑子生长和生物量积累具有重要作用. ...
珍珠菜属植物功能性状的趋异分化
1
2019
...
SLA 和
LDMC 是叶片功能性状中最主要的两个指标, 被认为是指示植物养分利用策略的关键性指标(
Jager et al ., 2015 ).与我们的假设(1)一致, 叶片功能性状对氮变化和磷变化的响应具有差异性.随磷浓度的增加,
SLA 显著降低,
LDMC 显著增加, 而氮处理下却没有相似的结果.前人研究表明,
SLA 随养分的减少而降低, 较低的
SLA 和较高的
LDMC 有利于植物适应资源贫瘠的环境(
Jager et al ., 2015 ).然而, 我们的研究中, 高浓度磷水平降低了
SLA, 增加了
LDMC; 相反, 低浓度磷水平增加了
SLA, 降低了
LDMC .高磷水平下低
SLA 、高
LDMC 表明车桑子采取养分的有效保存策略, 低磷水平下高
SLA 、低
LDMC 可提高对光资源的获取能力, 表明车桑子采取快速生长的策略, 该研究结果与
周光霞等(2016) 的研究相似.然而, 低磷水平下车桑子生长依然受到严重的限制.
林婉奇等(2019) 也发现磷添加处理减少了
SLA, 但显著促进了幼苗生长.叶面积和
SLA 呈负相关关系, 叶面积增加可提高植物的光获取能力, 弥补因
SLA 减小而导致的碳摄取效率的损失(
陈延松等, 2019 ).因此, 高磷水平下叶面积的增加对促进车桑子生长和生物量积累具有重要作用. ...
Drought stress and plant nutrition
1
2011
... 车桑子(
Dodonaea viscosa )又名坡柳、明油子, 具有耐干旱、耐贫瘠和适应性强等特性, 是我国干热河谷区的优势物种, 也是该区植被恢复工程中的常见植物, 具有重要的生态价值.关于车桑子的研究, 目前重点研究了车桑子对干旱胁迫的响应特征(
李吉跃等, 2003 ;
陈少瑜等, 2006 ).但除了干旱胁迫之外, 养分胁迫成为干旱胁迫的次生效应(
da Silva et al ., 2011 ).加之该区土壤侵蚀严重, 干热河谷土壤(燥红土)普遍存在缺氮少磷的特点(
张燕平等, 2005 ), 严重影响了干热河谷植物的生长(
闫帮国等, 2012 ).然而, 关于车桑子性状对氮磷限制的响应目前尚不清楚.因此, 本研究以车桑子幼苗为研究对象, 采用砂培的方式进行试验, 研究车桑子幼苗生长特征、生物量分配、叶片功能性状及其相互关系对氮磷梯度的响应特征, 从生物量分配和叶片功能性状的角度探讨车桑子对低氮和低磷条件的适应性, 并验证以下假说: (1)车桑子幼苗生物量分配和叶片功能性状对氮变化和磷变化的响应具有差异性; (2)生物量分配和叶性状对氮磷浓度响应存在协同或权衡关系. ...
A mechanistic model for understanding root-induced chemical changes controlling phosphorus availability
1
2010
... 在各磷浓度处理下, 车桑子生长和生物量积累随磷浓度的增加而显著增加.野外条件下, 车桑子生长区的土壤有效磷含量为4.90 mg·kg
-1 , 同理换算出来为1.58 mmol·L
-1 , 高于本研究的P3水平, 但低于P4水平.车桑子对野外磷水平的适应进化在一定程度上可以解释本研究低浓度和中等浓度磷处理对车桑子生长的限制作用.然而, 低浓度磷水平下车桑子根冠比并没有显著增加, 表明车桑子不会通过调节根冠比来适应低磷环境, 这与低氮处理结果不同.与我们的假设(1)一致, 车桑子生物量分配对氮变化和磷变化的响应具有差异性.Venterink和Güsewell (2010)认为, 磷限制条件下, 由于根系生长需要消耗的能量较多, 而且生长过程中伴随着细胞分裂, 需要大量的DNA和RNA合成(这两种物质中含有大量的磷), 需要更多的磷.因此, 植物不会大量投资根系来吸收磷离子, 而是通过其他机制促进磷的吸收, 比如磷酸酶活性(
Minden & Venterink, 2019 )、根系分泌物、钙离子吸收(
Devau et al ., 2010 )等.本研究中, 低浓度磷水平下叶/茎生物量比例显著增加, 表明低磷处理增加了地上部分叶生物量的分配.由于茎的生长也需要较多的磷, 在缺磷时, 植物为了保住最关键的功能, 可能会增加叶片投入以维持光合作用, 而减少茎的支出.研究表明, 叶片氮与光合作用直接相关, 氮是光合作用的首要影响因子, 而叶片磷对光合作用的影响较弱(
Reich et al ., 2009 ), 这也反映了车桑子通过维持叶片生长以提高在低磷条件下的适应能力. ...
Critical N:P values: predicting nutrient deficiencies in desert shrublands
1
2004
... 本研究中, 不同的氮浓度和磷浓度处理显著影响了车桑子叶片氮含量, 但叶片磷含量和叶片N:P并未受到显著影响.由于植物具有保持自身化学组成相对稳定的能力, 环境中的养分变化对植物氮磷元素的影响也取决于植物化学计量内稳性的强弱(
Persson et al ., 2010 ).植物内稳性特征可用内稳性指数
H 进行界定, 根据内稳性指数可将植物分为四类:
H > 4为稳态型; 2 <
H < 4为弱稳态型; 4/3 <
H < 2为弱敏感型;
H < 4/3为敏感型(
Persson et al ., 2010 ).本研究中, 不同氮浓度处理下, 叶片氮含量和叶片N:P的
H 分别为6.86和7.31, 均大于4, 呈稳态型, 并且叶片N:P的内稳性大于氮的内稳性.其他研究也表明, 氮磷比的内稳性大于氮和磷的内稳性(
Yu et al ., 2011 ;
宁志英等, 2019 ).这说明, 植物对其自身内稳性的调节主要体现在N:P而不是单一的元素含量上.然而, 不同磷浓度处理下, 叶片磷含量、叶片N:P与营养液的磷浓度和营养液N:P并没有显著的幂函数关系, 其内稳性指数计算结果为负值.
Xing等(2015) 用1/
H 来衡量内稳性的强弱, 也测出内稳性指数为负值, 并将其归为严格内稳型.这说明磷处理条件下, 车桑子叶片磷含量和N:P具有极强的内稳性.较高的内稳性意味着养分元素变化小, 表明具有保守的养分利用策略, 也就更容易适应贫瘠环境而成为优势种(
Yu et al ., 2010 ).不同生长期植物内稳性不同, 内稳性随着植物生长发育逐渐增强(
Yu et al ., 2011 ).本研究中车桑子幼苗已表现出较高的内稳性, 因此, 这可能是促进车桑子成为干热河谷优势植物的原因之一.此外, 所有处理条件下, 车桑子的叶片N:P均在30左右波动.按照叶片临界N:P对限制元素的判断理论, 车桑子生长受到磷限制(
Güsewell, 2004 ;
Sullivan et al ., 2014 ), 这和氮磷添加对车桑子生长和生物量积累的结果是不一致的.与前人的研究观点(
Drenovsky & Richards, 2004 ;
Zhang et al ., 2004 )比较一致, 我们的结果也表明, 在物种水平上, 用叶片N:P来指示植物所受到的养分限制类型是不可靠的. ...
不同雪被厚度下典型高山草地早春植物叶片性状、株高及生物量分配的研究
1
2016
... 关系, 可见车桑子叶片磷含量的变化是驱动N:P变化的主要因子.
吴统贵等(2010) 和
宋彦涛等(2012) 的研究也表明, 叶片N:P主要受到磷的驱动, N:P的变化受控于叶片磷含量的变化.然而, 氮、磷处理条件下, 车桑子叶片氮和磷含量并没有显著正相关关系, 这可能是由于氮、磷养分不同的内稳性导致的.
黄端等(2017) 也发现叶片氮、磷含量之间无显著的正相关关系.此外, 本研究中, 磷处理下叶片氮含量和
SLA 显著正相关, 而氮处理下却没有这种相关性, 这与以往的研究结果不一致.以往大量的研究发现植物叶片
SLA 与叶片氮含量显著正相关, 二者的关联是植物叶经济型谱的重要组成部分(
于鸿莹等, 2014 ;
熊静等, 2019 ).以往的数据基本是从多个物种之间发现的, 也就是
SLA 与叶片氮含量正相关是种间变异的结果,而本研究是通过设置氮浓度梯度, 是基于环境中氮浓度变化对单个物种内
SLA 与叶片氮含量影响后的结果.在氮有效性增加的情况下, 叶片氮含量的响应更直接更迅速, 植物吸收氮后叶片氮含量显著增加, 而
SLA 的响应相对滞后, 与
Wright和Sutton-Grier (2012) 在氮添加条件下的结果是一致的, 这也反映了长期的氮限制使车桑子在较高的氮水平下短期内依然保持相对保守的氮利用策略.因此, 氮添加条件下车桑子叶片氮含量与
SLA 不相关甚至呈现出较弱的负相关关系.本研究结果表明, 环境中养分驱动下单个物种内叶片的性状变化可能并不完全符合经济型谱理论, 而会受到氮、磷养分有效性的影响, 符合叶经济型谱形成的环境因子主导假说(
赖小红等, 2019 ).此外, 植物功能性状和生物量分配之间存在着某种共轭的变化(
高景等, 2016 ).本研究中, 氮处理下, 车桑子根冠比与叶片氮含量、N:P显著负相关, 磷处理下却没有显著的相关性.以上结果支持我们假设(2)的观点, 车桑子生物量分配与叶性状间存在着一定的协同和权衡关系, 但是车桑子在面临氮、磷有效性的变化时具有不同的响应策略. ...
不同雪被厚度下典型高山草地早春植物叶片性状、株高及生物量分配的研究
1
2016
... 关系, 可见车桑子叶片磷含量的变化是驱动N:P变化的主要因子.
吴统贵等(2010) 和
宋彦涛等(2012) 的研究也表明, 叶片N:P主要受到磷的驱动, N:P的变化受控于叶片磷含量的变化.然而, 氮、磷处理条件下, 车桑子叶片氮和磷含量并没有显著正相关关系, 这可能是由于氮、磷养分不同的内稳性导致的.
黄端等(2017) 也发现叶片氮、磷含量之间无显著的正相关关系.此外, 本研究中, 磷处理下叶片氮含量和
SLA 显著正相关, 而氮处理下却没有这种相关性, 这与以往的研究结果不一致.以往大量的研究发现植物叶片
SLA 与叶片氮含量显著正相关, 二者的关联是植物叶经济型谱的重要组成部分(
于鸿莹等, 2014 ;
熊静等, 2019 ).以往的数据基本是从多个物种之间发现的, 也就是
SLA 与叶片氮含量正相关是种间变异的结果,而本研究是通过设置氮浓度梯度, 是基于环境中氮浓度变化对单个物种内
SLA 与叶片氮含量影响后的结果.在氮有效性增加的情况下, 叶片氮含量的响应更直接更迅速, 植物吸收氮后叶片氮含量显著增加, 而
SLA 的响应相对滞后, 与
Wright和Sutton-Grier (2012) 在氮添加条件下的结果是一致的, 这也反映了长期的氮限制使车桑子在较高的氮水平下短期内依然保持相对保守的氮利用策略.因此, 氮添加条件下车桑子叶片氮含量与
SLA 不相关甚至呈现出较弱的负相关关系.本研究结果表明, 环境中养分驱动下单个物种内叶片的性状变化可能并不完全符合经济型谱理论, 而会受到氮、磷养分有效性的影响, 符合叶经济型谱形成的环境因子主导假说(
赖小红等, 2019 ).此外, 植物功能性状和生物量分配之间存在着某种共轭的变化(
高景等, 2016 ).本研究中, 氮处理下, 车桑子根冠比与叶片氮含量、N:P显著负相关, 磷处理下却没有显著的相关性.以上结果支持我们假设(2)的观点, 车桑子生物量分配与叶性状间存在着一定的协同和权衡关系, 但是车桑子在面临氮、磷有效性的变化时具有不同的响应策略. ...
N:P ratios in terrestrial plants: variation and functional significance
1
2004
... 本研究中, 不同的氮浓度和磷浓度处理显著影响了车桑子叶片氮含量, 但叶片磷含量和叶片N:P并未受到显著影响.由于植物具有保持自身化学组成相对稳定的能力, 环境中的养分变化对植物氮磷元素的影响也取决于植物化学计量内稳性的强弱(
Persson et al ., 2010 ).植物内稳性特征可用内稳性指数
H 进行界定, 根据内稳性指数可将植物分为四类:
H > 4为稳态型; 2 <
H < 4为弱稳态型; 4/3 <
H < 2为弱敏感型;
H < 4/3为敏感型(
Persson et al ., 2010 ).本研究中, 不同氮浓度处理下, 叶片氮含量和叶片N:P的
H 分别为6.86和7.31, 均大于4, 呈稳态型, 并且叶片N:P的内稳性大于氮的内稳性.其他研究也表明, 氮磷比的内稳性大于氮和磷的内稳性(
Yu et al ., 2011 ;
宁志英等, 2019 ).这说明, 植物对其自身内稳性的调节主要体现在N:P而不是单一的元素含量上.然而, 不同磷浓度处理下, 叶片磷含量、叶片N:P与营养液的磷浓度和营养液N:P并没有显著的幂函数关系, 其内稳性指数计算结果为负值.
Xing等(2015) 用1/
H 来衡量内稳性的强弱, 也测出内稳性指数为负值, 并将其归为严格内稳型.这说明磷处理条件下, 车桑子叶片磷含量和N:P具有极强的内稳性.较高的内稳性意味着养分元素变化小, 表明具有保守的养分利用策略, 也就更容易适应贫瘠环境而成为优势种(
Yu et al ., 2010 ).不同生长期植物内稳性不同, 内稳性随着植物生长发育逐渐增强(
Yu et al ., 2011 ).本研究中车桑子幼苗已表现出较高的内稳性, 因此, 这可能是促进车桑子成为干热河谷优势植物的原因之一.此外, 所有处理条件下, 车桑子的叶片N:P均在30左右波动.按照叶片临界N:P对限制元素的判断理论, 车桑子生长受到磷限制(
Güsewell, 2004 ;
Sullivan et al ., 2014 ), 这和氮磷添加对车桑子生长和生物量积累的结果是不一致的.与前人的研究观点(
Drenovsky & Richards, 2004 ;
Zhang et al ., 2004 )比较一致, 我们的结果也表明, 在物种水平上, 用叶片N:P来指示植物所受到的养分限制类型是不可靠的. ...
Composition of plant species mixtures grown at various N:P ratios and levels of nutrient supply
1
2003
... 车桑子叶片氮利用效率随营养液N:P的增加而降低.研究表明, 氮磷养分供应量对植物生长起着主要作用, 而除此之外, 氮磷供给比也影响着植物生长(
Güsewell & Bollens, 2003 ).本研究中, N4 (30 mmol·L
-1 N、1 mmol·L
-1 P)和P2 (15 mmol·L
-1 N、0.5 mmol·L
-1 P)处理的氮磷供给比均为30, 但P2处理的车桑子生长和生物量积累显著低于N4处理, 而氮利用效率显著高于N4处理.车桑子其他生长指标、生物量分配、叶片性状和磷利用效率比较一致, 随营养液氮磷比的变化并没有发生规律性变化.因此, 我们的结果表明, 氮磷的供应量而不是单纯的氮磷供给比对植物性状和生长的影响更大. ...
Responses of plant-soil properties to increasing N deposition and implications for large-scale eco-restoration in the semiarid grassland of the northern Loess Plateau, China
1
2013
... 在养分有效性发生变化时, 植物会通过生物量分配和叶片功能性状的调整来适应养分变化.按照最优分配理论, 植物会将更多的生物量分配至能够获取限制资源的器官(
Mccarthy & Enquist, 2007 ).在养分有效性较低的情况下通常会通过增加根生物量分配和根冠比来获取更多的养分(
Poorter et al ., 2012 ).此外, 养分贫瘠的条件下, 植物会增加地上部分叶生物量的比例(相对于茎)(
Yan et al ., 2016 ), 但这方面的研究还相对较少.另一方面, 叶片功能性状的调整与资源获取和利用密切相关, 其形态结构和营养元素含量等均会随环境中氮磷条件的改变而发生适应性变化(
Qi et al ., 2009 ).比如, 较低的养分有效性通常会降低比叶面积, 增加叶干物质含量和养分利用效率, 而随着养分有效性的增加, 叶片养分含量显著增加(
Ordo?ez et al ., 2009 ;
林婉奇等, 2019 ).然而, 不同物种对氮磷添加的响应具有物种特异性(
Han et al ., 2013 ;
Mayor et al ., 2014 ).在氮限制条件下, 具有更高根长和根生物量比例的物种占据优势; 相反, 在磷限制条件下, 植物根长不能解释物种间的竞争优势, 菌根菌丝长度、根系分泌物、磷酸酶活性等相关性状起着重要作用(
Venterink & Güsewell, 2010 ;
Minden & Venterink, 2019 ), 这说明植物性状对氮和磷可能具有不同的响应特征, 包括生物量分配、叶片性状或养分利用效率等多个植物性状.由于不同的性状在适应氮限制和磷限制中表现出不同的变化趋势, 植物可能通过调整不同的性状来响应氮变化或磷变化, 但植物性状对氮、磷的响应是否表现出差异并不十分清楚. ...
The Water Culture Method for Growing Plants Without Soil 1
2020
... 本试验采用氮和磷的单因素设计, 分别包括4个氮浓度和4个磷浓度.由于盆栽实验中植物根系伸展空间有限, 按照野外土壤实际养分含量设置浓度梯度下植物很难获得充分生长.因此, 本试验参照前人的相关研究(
Yu et al ., 2011 ;
Yan et al ., 2015 ), 以Hoagland营养液(15 mmol·L
-1 N、1 mmol·L
-1 P)为基础进行氮磷养分添加.Hoagland营养液已经比较成熟, 溶液的pH较为稳定, 其养分配制考虑了植物需求, 养分浓度对大多数植物都很适合, 被广泛应用于水培和砂培实验中(
Hoagland & Arnon, 1938 ).由于河沙中没有养分, 不添加额外养分植物将无法生长, 因此, 本研究没有设置不添加的对照处理.结合浓度梯度的可控性, 4个氮浓度分别为3、5、15、30 mmol·L
-1 , 记为N1、N2、N3、N4, 其中, N4为高浓度氮水平, N3为中等浓度氮水平, N1和N2为低浓度氮水平, 通过添加NH
4 NO
3 进行调整; 4个磷浓度分别为0.25、0.5、1、2 mmol·L
-1 , 记为P1、P2、P3、P4, 其中, P4为高浓度磷水平, P3为中等浓度磷水平, P1和P2为低浓度磷水平, 通过添加KH
2 PO
4 进行调整.各氮、磷水平的设置及营养液的氮磷比如
表1 所示.其中, N3和P3处理一致, 因此, 试验共计7个处理, 每个处理5个重复.除氮、磷浓度不同外, 其余钾、钙、镁含量及微量元素含量均与全量Hoagland营养液一致, 配好后用H
2 SO
4 或NaOH调整溶液pH为6.由于钾、钙、镁的添加均为硫酸盐物质, 通过H
2 SO
4 调节pH对物质的平衡影响较小, 而钠离子为植物非必需元素, 对养分需求的平衡不会造成影响, 同时营养液中所添加的物质主要为中性物质, 不同营养液中pH差异较小, 所需H
2 SO
4 或NaOH的量是极小的, 因此, 这些影响基本可以忽略, 试验处理中不存在离子不平衡问题.由于砂培基质对营养液的保持能力较低, 试验处理期间, 每隔2天浇1次营养液, 每次营养液添加量为200 mL, 以保持养分浓度尽量维持在设定水平.同时, 将所有盆钵放在遮雨大棚内, 可避免降水对养分的淋失. ...
漓江水陆交错带植物叶性状对水淹胁迫的响应及经济谱分析
1
2017
... 关系, 可见车桑子叶片磷含量的变化是驱动N:P变化的主要因子.
吴统贵等(2010) 和
宋彦涛等(2012) 的研究也表明, 叶片N:P主要受到磷的驱动, N:P的变化受控于叶片磷含量的变化.然而, 氮、磷处理条件下, 车桑子叶片氮和磷含量并没有显著正相关关系, 这可能是由于氮、磷养分不同的内稳性导致的.
黄端等(2017) 也发现叶片氮、磷含量之间无显著的正相关关系.此外, 本研究中, 磷处理下叶片氮含量和
SLA 显著正相关, 而氮处理下却没有这种相关性, 这与以往的研究结果不一致.以往大量的研究发现植物叶片
SLA 与叶片氮含量显著正相关, 二者的关联是植物叶经济型谱的重要组成部分(
于鸿莹等, 2014 ;
熊静等, 2019 ).以往的数据基本是从多个物种之间发现的, 也就是
SLA 与叶片氮含量正相关是种间变异的结果,而本研究是通过设置氮浓度梯度, 是基于环境中氮浓度变化对单个物种内
SLA 与叶片氮含量影响后的结果.在氮有效性增加的情况下, 叶片氮含量的响应更直接更迅速, 植物吸收氮后叶片氮含量显著增加, 而
SLA 的响应相对滞后, 与
Wright和Sutton-Grier (2012) 在氮添加条件下的结果是一致的, 这也反映了长期的氮限制使车桑子在较高的氮水平下短期内依然保持相对保守的氮利用策略.因此, 氮添加条件下车桑子叶片氮含量与
SLA 不相关甚至呈现出较弱的负相关关系.本研究结果表明, 环境中养分驱动下单个物种内叶片的性状变化可能并不完全符合经济型谱理论, 而会受到氮、磷养分有效性的影响, 符合叶经济型谱形成的环境因子主导假说(
赖小红等, 2019 ).此外, 植物功能性状和生物量分配之间存在着某种共轭的变化(
高景等, 2016 ).本研究中, 氮处理下, 车桑子根冠比与叶片氮含量、N:P显著负相关, 磷处理下却没有显著的相关性.以上结果支持我们假设(2)的观点, 车桑子生物量分配与叶性状间存在着一定的协同和权衡关系, 但是车桑子在面临氮、磷有效性的变化时具有不同的响应策略. ...
漓江水陆交错带植物叶性状对水淹胁迫的响应及经济谱分析
1
2017
... 关系, 可见车桑子叶片磷含量的变化是驱动N:P变化的主要因子.
吴统贵等(2010) 和
宋彦涛等(2012) 的研究也表明, 叶片N:P主要受到磷的驱动, N:P的变化受控于叶片磷含量的变化.然而, 氮、磷处理条件下, 车桑子叶片氮和磷含量并没有显著正相关关系, 这可能是由于氮、磷养分不同的内稳性导致的.
黄端等(2017) 也发现叶片氮、磷含量之间无显著的正相关关系.此外, 本研究中, 磷处理下叶片氮含量和
SLA 显著正相关, 而氮处理下却没有这种相关性, 这与以往的研究结果不一致.以往大量的研究发现植物叶片
SLA 与叶片氮含量显著正相关, 二者的关联是植物叶经济型谱的重要组成部分(
于鸿莹等, 2014 ;
熊静等, 2019 ).以往的数据基本是从多个物种之间发现的, 也就是
SLA 与叶片氮含量正相关是种间变异的结果,而本研究是通过设置氮浓度梯度, 是基于环境中氮浓度变化对单个物种内
SLA 与叶片氮含量影响后的结果.在氮有效性增加的情况下, 叶片氮含量的响应更直接更迅速, 植物吸收氮后叶片氮含量显著增加, 而
SLA 的响应相对滞后, 与
Wright和Sutton-Grier (2012) 在氮添加条件下的结果是一致的, 这也反映了长期的氮限制使车桑子在较高的氮水平下短期内依然保持相对保守的氮利用策略.因此, 氮添加条件下车桑子叶片氮含量与
SLA 不相关甚至呈现出较弱的负相关关系.本研究结果表明, 环境中养分驱动下单个物种内叶片的性状变化可能并不完全符合经济型谱理论, 而会受到氮、磷养分有效性的影响, 符合叶经济型谱形成的环境因子主导假说(
赖小红等, 2019 ).此外, 植物功能性状和生物量分配之间存在着某种共轭的变化(
高景等, 2016 ).本研究中, 氮处理下, 车桑子根冠比与叶片氮含量、N:P显著负相关, 磷处理下却没有显著的相关性.以上结果支持我们假设(2)的观点, 车桑子生物量分配与叶性状间存在着一定的协同和权衡关系, 但是车桑子在面临氮、磷有效性的变化时具有不同的响应策略. ...
Soil fertility induces coordinated responses of multiple independent functional traits
2
2015
...
SLA 和
LDMC 是叶片功能性状中最主要的两个指标, 被认为是指示植物养分利用策略的关键性指标(
Jager et al ., 2015 ).与我们的假设(1)一致, 叶片功能性状对氮变化和磷变化的响应具有差异性.随磷浓度的增加,
SLA 显著降低,
LDMC 显著增加, 而氮处理下却没有相似的结果.前人研究表明,
SLA 随养分的减少而降低, 较低的
SLA 和较高的
LDMC 有利于植物适应资源贫瘠的环境(
Jager et al ., 2015 ).然而, 我们的研究中, 高浓度磷水平降低了
SLA, 增加了
LDMC; 相反, 低浓度磷水平增加了
SLA, 降低了
LDMC .高磷水平下低
SLA 、高
LDMC 表明车桑子采取养分的有效保存策略, 低磷水平下高
SLA 、低
LDMC 可提高对光资源的获取能力, 表明车桑子采取快速生长的策略, 该研究结果与
周光霞等(2016) 的研究相似.然而, 低磷水平下车桑子生长依然受到严重的限制.
林婉奇等(2019) 也发现磷添加处理减少了
SLA, 但显著促进了幼苗生长.叶面积和
SLA 呈负相关关系, 叶面积增加可提高植物的光获取能力, 弥补因
SLA 减小而导致的碳摄取效率的损失(
陈延松等, 2019 ).因此, 高磷水平下叶面积的增加对促进车桑子生长和生物量积累具有重要作用. ...
... 有利于植物适应资源贫瘠的环境(
Jager et al ., 2015 ).然而, 我们的研究中, 高浓度磷水平降低了
SLA, 增加了
LDMC; 相反, 低浓度磷水平增加了
SLA, 降低了
LDMC .高磷水平下低
SLA 、高
LDMC 表明车桑子采取养分的有效保存策略, 低磷水平下高
SLA 、低
LDMC 可提高对光资源的获取能力, 表明车桑子采取快速生长的策略, 该研究结果与
周光霞等(2016) 的研究相似.然而, 低磷水平下车桑子生长依然受到严重的限制.
林婉奇等(2019) 也发现磷添加处理减少了
SLA, 但显著促进了幼苗生长.叶面积和
SLA 呈负相关关系, 叶面积增加可提高植物的光获取能力, 弥补因
SLA 减小而导致的碳摄取效率的损失(
陈延松等, 2019 ).因此, 高磷水平下叶面积的增加对促进车桑子生长和生物量积累具有重要作用. ...
人工控制条件下9种园林植物叶功能性状对短期NO
2 污染的响应
1
2019
... 关系, 可见车桑子叶片磷含量的变化是驱动N:P变化的主要因子.
吴统贵等(2010) 和
宋彦涛等(2012) 的研究也表明, 叶片N:P主要受到磷的驱动, N:P的变化受控于叶片磷含量的变化.然而, 氮、磷处理条件下, 车桑子叶片氮和磷含量并没有显著正相关关系, 这可能是由于氮、磷养分不同的内稳性导致的.
黄端等(2017) 也发现叶片氮、磷含量之间无显著的正相关关系.此外, 本研究中, 磷处理下叶片氮含量和
SLA 显著正相关, 而氮处理下却没有这种相关性, 这与以往的研究结果不一致.以往大量的研究发现植物叶片
SLA 与叶片氮含量显著正相关, 二者的关联是植物叶经济型谱的重要组成部分(
于鸿莹等, 2014 ;
熊静等, 2019 ).以往的数据基本是从多个物种之间发现的, 也就是
SLA 与叶片氮含量正相关是种间变异的结果,而本研究是通过设置氮浓度梯度, 是基于环境中氮浓度变化对单个物种内
SLA 与叶片氮含量影响后的结果.在氮有效性增加的情况下, 叶片氮含量的响应更直接更迅速, 植物吸收氮后叶片氮含量显著增加, 而
SLA 的响应相对滞后, 与
Wright和Sutton-Grier (2012) 在氮添加条件下的结果是一致的, 这也反映了长期的氮限制使车桑子在较高的氮水平下短期内依然保持相对保守的氮利用策略.因此, 氮添加条件下车桑子叶片氮含量与
SLA 不相关甚至呈现出较弱的负相关关系.本研究结果表明, 环境中养分驱动下单个物种内叶片的性状变化可能并不完全符合经济型谱理论, 而会受到氮、磷养分有效性的影响, 符合叶经济型谱形成的环境因子主导假说(
赖小红等, 2019 ).此外, 植物功能性状和生物量分配之间存在着某种共轭的变化(
高景等, 2016 ).本研究中, 氮处理下, 车桑子根冠比与叶片氮含量、N:P显著负相关, 磷处理下却没有显著的相关性.以上结果支持我们假设(2)的观点, 车桑子生物量分配与叶性状间存在着一定的协同和权衡关系, 但是车桑子在面临氮、磷有效性的变化时具有不同的响应策略. ...
人工控制条件下9种园林植物叶功能性状对短期NO
2 污染的响应
1
2019
... 关系, 可见车桑子叶片磷含量的变化是驱动N:P变化的主要因子.
吴统贵等(2010) 和
宋彦涛等(2012) 的研究也表明, 叶片N:P主要受到磷的驱动, N:P的变化受控于叶片磷含量的变化.然而, 氮、磷处理条件下, 车桑子叶片氮和磷含量并没有显著正相关关系, 这可能是由于氮、磷养分不同的内稳性导致的.
黄端等(2017) 也发现叶片氮、磷含量之间无显著的正相关关系.此外, 本研究中, 磷处理下叶片氮含量和
SLA 显著正相关, 而氮处理下却没有这种相关性, 这与以往的研究结果不一致.以往大量的研究发现植物叶片
SLA 与叶片氮含量显著正相关, 二者的关联是植物叶经济型谱的重要组成部分(
于鸿莹等, 2014 ;
熊静等, 2019 ).以往的数据基本是从多个物种之间发现的, 也就是
SLA 与叶片氮含量正相关是种间变异的结果,而本研究是通过设置氮浓度梯度, 是基于环境中氮浓度变化对单个物种内
SLA 与叶片氮含量影响后的结果.在氮有效性增加的情况下, 叶片氮含量的响应更直接更迅速, 植物吸收氮后叶片氮含量显著增加, 而
SLA 的响应相对滞后, 与
Wright和Sutton-Grier (2012) 在氮添加条件下的结果是一致的, 这也反映了长期的氮限制使车桑子在较高的氮水平下短期内依然保持相对保守的氮利用策略.因此, 氮添加条件下车桑子叶片氮含量与
SLA 不相关甚至呈现出较弱的负相关关系.本研究结果表明, 环境中养分驱动下单个物种内叶片的性状变化可能并不完全符合经济型谱理论, 而会受到氮、磷养分有效性的影响, 符合叶经济型谱形成的环境因子主导假说(
赖小红等, 2019 ).此外, 植物功能性状和生物量分配之间存在着某种共轭的变化(
高景等, 2016 ).本研究中, 氮处理下, 车桑子根冠比与叶片氮含量、N:P显著负相关, 磷处理下却没有显著的相关性.以上结果支持我们假设(2)的观点, 车桑子生物量分配与叶性状间存在着一定的协同和权衡关系, 但是车桑子在面临氮、磷有效性的变化时具有不同的响应策略. ...
供氮水平对枫香种源幼苗生长及氮、磷吸收利用的影响
2
2009
... 采集每盆植株中部的叶片, 将叶片烘干、称质量和粉碎, 测定其氮磷含量.全氮含量采用H
2 SO
4 -H
2 O
2 消煮-凯氏蒸馏法测定, 全磷含量采用H
2 SO
4 -H
2 O
2 消煮-钼锑抗比色法测定.养分利用效率可用单位养分吸收量所产生的干物质量大小来表示.因此, 氮(磷)利用效率=总生物量/氮(磷)吸收量(
冷华妮等, 2009 ). ...
... 在植物养分限制的判断中, 若一种有效态元素的添加引起了植物生化过程速率或生物量的增加, 则意味着存在这种养分的限制(
Vitousek et al ., 2010 ;
Sullivan et al ., 2014 ).本研究表明, 高浓度氮水平显著促进了车桑子的生长和生物量积累, 但车桑子生长和生物量积累在低浓度和中等浓度氮水平下没有显著差异.野外条件下, 车桑子生长区的土壤有效氮(硝态氮与铵态氮之和)含量大约为11.7 mg·kg
-1 , 考虑到土壤中水分含量在10%左右, 也就是每kg土壤约含0.1 L水, 那么土壤中的有效氮含量约为117 mg·L
-1 , 介于本研究设置的N3 (210 mg·L
-1 )和N2 (70 mg·L
-1 )水平之间.因此, 本研究中, N1-N3处理下车桑子的生长没有显著差异, 可能是由于干热河谷长期的氮限制使得植物形成了对低氮环境的一系列形态与生理适应性.干旱生态系统氮限制尤为普遍(
Yahdjian et al ., 2011 ).已有研究表明, 氮是限制金沙江干旱河谷退化草地系统恢复的主要因素(
张彦东等, 2004 ).同时, 我们的结果表明, 车桑子主要通过增加对氮的吸收和利用来适应低氮环境.一方面, 车桑子在低氮条件下, 其根冠比显著增加可促进氮的吸收; 另一方面, 吸收的氮的利用效率又显著提高.根冠比的增加和养分利用效率的提高均是植物适应养分限制的主要机制(
冷华妮等, 2009 ;
Venterink & Güsewell, 2010 ).然而, 由于本研究中养分添加只持续了一个生长季, 试验时间较短, 鉴于车桑子幼苗长期对氮限制的适应, 对试验中的低浓度氮水平可能还不太敏感, 通过根冠比的调整和养分利用效率的提高即可保持其正常的生长和生物量积累, 但在长期的低浓度氮水平下车桑子生长和生物量积累可能会受到限制. ...
供氮水平对枫香种源幼苗生长及氮、磷吸收利用的影响
2
2009
... 采集每盆植株中部的叶片, 将叶片烘干、称质量和粉碎, 测定其氮磷含量.全氮含量采用H
2 SO
4 -H
2 O
2 消煮-凯氏蒸馏法测定, 全磷含量采用H
2 SO
4 -H
2 O
2 消煮-钼锑抗比色法测定.养分利用效率可用单位养分吸收量所产生的干物质量大小来表示.因此, 氮(磷)利用效率=总生物量/氮(磷)吸收量(
冷华妮等, 2009 ). ...
... 在植物养分限制的判断中, 若一种有效态元素的添加引起了植物生化过程速率或生物量的增加, 则意味着存在这种养分的限制(
Vitousek et al ., 2010 ;
Sullivan et al ., 2014 ).本研究表明, 高浓度氮水平显著促进了车桑子的生长和生物量积累, 但车桑子生长和生物量积累在低浓度和中等浓度氮水平下没有显著差异.野外条件下, 车桑子生长区的土壤有效氮(硝态氮与铵态氮之和)含量大约为11.7 mg·kg
-1 , 考虑到土壤中水分含量在10%左右, 也就是每kg土壤约含0.1 L水, 那么土壤中的有效氮含量约为117 mg·L
-1 , 介于本研究设置的N3 (210 mg·L
-1 )和N2 (70 mg·L
-1 )水平之间.因此, 本研究中, N1-N3处理下车桑子的生长没有显著差异, 可能是由于干热河谷长期的氮限制使得植物形成了对低氮环境的一系列形态与生理适应性.干旱生态系统氮限制尤为普遍(
Yahdjian et al ., 2011 ).已有研究表明, 氮是限制金沙江干旱河谷退化草地系统恢复的主要因素(
张彦东等, 2004 ).同时, 我们的结果表明, 车桑子主要通过增加对氮的吸收和利用来适应低氮环境.一方面, 车桑子在低氮条件下, 其根冠比显著增加可促进氮的吸收; 另一方面, 吸收的氮的利用效率又显著提高.根冠比的增加和养分利用效率的提高均是植物适应养分限制的主要机制(
冷华妮等, 2009 ;
Venterink & Güsewell, 2010 ).然而, 由于本研究中养分添加只持续了一个生长季, 试验时间较短, 鉴于车桑子幼苗长期对氮限制的适应, 对试验中的低浓度氮水平可能还不太敏感, 通过根冠比的调整和养分利用效率的提高即可保持其正常的生长和生物量积累, 但在长期的低浓度氮水平下车桑子生长和生物量积累可能会受到限制. ...
金沙江干热河谷坡柳的光合特性
1
2003
... 车桑子(
Dodonaea viscosa )又名坡柳、明油子, 具有耐干旱、耐贫瘠和适应性强等特性, 是我国干热河谷区的优势物种, 也是该区植被恢复工程中的常见植物, 具有重要的生态价值.关于车桑子的研究, 目前重点研究了车桑子对干旱胁迫的响应特征(
李吉跃等, 2003 ;
陈少瑜等, 2006 ).但除了干旱胁迫之外, 养分胁迫成为干旱胁迫的次生效应(
da Silva et al ., 2011 ).加之该区土壤侵蚀严重, 干热河谷土壤(燥红土)普遍存在缺氮少磷的特点(
张燕平等, 2005 ), 严重影响了干热河谷植物的生长(
闫帮国等, 2012 ).然而, 关于车桑子性状对氮磷限制的响应目前尚不清楚.因此, 本研究以车桑子幼苗为研究对象, 采用砂培的方式进行试验, 研究车桑子幼苗生长特征、生物量分配、叶片功能性状及其相互关系对氮磷梯度的响应特征, 从生物量分配和叶片功能性状的角度探讨车桑子对低氮和低磷条件的适应性, 并验证以下假说: (1)车桑子幼苗生物量分配和叶片功能性状对氮变化和磷变化的响应具有差异性; (2)生物量分配和叶性状对氮磷浓度响应存在协同或权衡关系. ...
金沙江干热河谷坡柳的光合特性
1
2003
... 车桑子(
Dodonaea viscosa )又名坡柳、明油子, 具有耐干旱、耐贫瘠和适应性强等特性, 是我国干热河谷区的优势物种, 也是该区植被恢复工程中的常见植物, 具有重要的生态价值.关于车桑子的研究, 目前重点研究了车桑子对干旱胁迫的响应特征(
李吉跃等, 2003 ;
陈少瑜等, 2006 ).但除了干旱胁迫之外, 养分胁迫成为干旱胁迫的次生效应(
da Silva et al ., 2011 ).加之该区土壤侵蚀严重, 干热河谷土壤(燥红土)普遍存在缺氮少磷的特点(
张燕平等, 2005 ), 严重影响了干热河谷植物的生长(
闫帮国等, 2012 ).然而, 关于车桑子性状对氮磷限制的响应目前尚不清楚.因此, 本研究以车桑子幼苗为研究对象, 采用砂培的方式进行试验, 研究车桑子幼苗生长特征、生物量分配、叶片功能性状及其相互关系对氮磷梯度的响应特征, 从生物量分配和叶片功能性状的角度探讨车桑子对低氮和低磷条件的适应性, 并验证以下假说: (1)车桑子幼苗生物量分配和叶片功能性状对氮变化和磷变化的响应具有差异性; (2)生物量分配和叶性状对氮磷浓度响应存在协同或权衡关系. ...
五种荒漠植物幼苗出土及生长对沙埋深度的响应
1
2006
... 幼苗是植物生活史过程中比较脆弱的一个阶段, 也是对环境变化最为敏感的一个阶段, 车桑子幼苗的死亡率较高, 在车桑子种植过程中应加强苗期管护(
张琴等, 2016 ).幼苗生长对植物群落的补员起着关键作用(
李秋艳和赵文智, 2006 ).因此, 本文基于车桑子幼苗的研究对车桑子在植被恢复中的应用具有一定的参考价值. ...
五种荒漠植物幼苗出土及生长对沙埋深度的响应
1
2006
... 幼苗是植物生活史过程中比较脆弱的一个阶段, 也是对环境变化最为敏感的一个阶段, 车桑子幼苗的死亡率较高, 在车桑子种植过程中应加强苗期管护(
张琴等, 2016 ).幼苗生长对植物群落的补员起着关键作用(
李秋艳和赵文智, 2006 ).因此, 本文基于车桑子幼苗的研究对车桑子在植被恢复中的应用具有一定的参考价值. ...
Molecular diversity of arbuscular mycorrhizal fungi associated with two dominant xerophytes in a valley-type savanna, southwest China
1
2010
... 需要说明的是, 本研究是基于室内砂培实验条件下得出的结果, 砂培条件和野外自然环境存在较大差异, 该试验结果与野外环境的试验结果是否一致并不确定.研究表明, 丛枝菌根真菌对植物养分吸收尤其是磷的吸收具有重大的贡献(
Smith et al ., 2004 ;
Smith & Smith, 2011 ), 并且, 干热河谷自然植被的根际土壤中大多有丰富的丛枝菌根真菌(
Li et al ., 2010 ).在这种条件下, 我们推测, 车桑子可能已经形成了相关的内部信号调控机制, 主要依赖菌根真菌吸收磷, 而减少了自身对低磷的生理生长调节能力, 使得根系分配不会对磷缺乏形成响应, 这也进一步解释了低磷条件下根冠比没有发生显著增加.而本研究中, 砂培条件缺乏菌根真菌的侵染, 车桑子自身对低磷的适应能力较差, 因此, 低磷水平限制了车桑子生长.在野外土壤环境条件下, 车桑子对氮磷有效性变化的响应特征是否与本研究不同, 尚需进一步验证. ...
不同密度樟树(
Cinnamomum camphora )幼苗生长和叶片性状对氮磷添加的响应
2
2019
... 在养分有效性发生变化时, 植物会通过生物量分配和叶片功能性状的调整来适应养分变化.按照最优分配理论, 植物会将更多的生物量分配至能够获取限制资源的器官(
Mccarthy & Enquist, 2007 ).在养分有效性较低的情况下通常会通过增加根生物量分配和根冠比来获取更多的养分(
Poorter et al ., 2012 ).此外, 养分贫瘠的条件下, 植物会增加地上部分叶生物量的比例(相对于茎)(
Yan et al ., 2016 ), 但这方面的研究还相对较少.另一方面, 叶片功能性状的调整与资源获取和利用密切相关, 其形态结构和营养元素含量等均会随环境中氮磷条件的改变而发生适应性变化(
Qi et al ., 2009 ).比如, 较低的养分有效性通常会降低比叶面积, 增加叶干物质含量和养分利用效率, 而随着养分有效性的增加, 叶片养分含量显著增加(
Ordo?ez et al ., 2009 ;
林婉奇等, 2019 ).然而, 不同物种对氮磷添加的响应具有物种特异性(
Han et al ., 2013 ;
Mayor et al ., 2014 ).在氮限制条件下, 具有更高根长和根生物量比例的物种占据优势; 相反, 在磷限制条件下, 植物根长不能解释物种间的竞争优势, 菌根菌丝长度、根系分泌物、磷酸酶活性等相关性状起着重要作用(
Venterink & Güsewell, 2010 ;
Minden & Venterink, 2019 ), 这说明植物性状对氮和磷可能具有不同的响应特征, 包括生物量分配、叶片性状或养分利用效率等多个植物性状.由于不同的性状在适应氮限制和磷限制中表现出不同的变化趋势, 植物可能通过调整不同的性状来响应氮变化或磷变化, 但植物性状对氮、磷的响应是否表现出差异并不十分清楚. ...
...
SLA 和
LDMC 是叶片功能性状中最主要的两个指标, 被认为是指示植物养分利用策略的关键性指标(
Jager et al ., 2015 ).与我们的假设(1)一致, 叶片功能性状对氮变化和磷变化的响应具有差异性.随磷浓度的增加,
SLA 显著降低,
LDMC 显著增加, 而氮处理下却没有相似的结果.前人研究表明,
SLA 随养分的减少而降低, 较低的
SLA 和较高的
LDMC 有利于植物适应资源贫瘠的环境(
Jager et al ., 2015 ).然而, 我们的研究中, 高浓度磷水平降低了
SLA, 增加了
LDMC; 相反, 低浓度磷水平增加了
SLA, 降低了
LDMC .高磷水平下低
SLA 、高
LDMC 表明车桑子采取养分的有效保存策略, 低磷水平下高
SLA 、低
LDMC 可提高对光资源的获取能力, 表明车桑子采取快速生长的策略, 该研究结果与
周光霞等(2016) 的研究相似.然而, 低磷水平下车桑子生长依然受到严重的限制.
林婉奇等(2019) 也发现磷添加处理减少了
SLA, 但显著促进了幼苗生长.叶面积和
SLA 呈负相关关系, 叶面积增加可提高植物的光获取能力, 弥补因
SLA 减小而导致的碳摄取效率的损失(
陈延松等, 2019 ).因此, 高磷水平下叶面积的增加对促进车桑子生长和生物量积累具有重要作用. ...
不同密度樟树(
Cinnamomum camphora )幼苗生长和叶片性状对氮磷添加的响应
2
2019
... 在养分有效性发生变化时, 植物会通过生物量分配和叶片功能性状的调整来适应养分变化.按照最优分配理论, 植物会将更多的生物量分配至能够获取限制资源的器官(
Mccarthy & Enquist, 2007 ).在养分有效性较低的情况下通常会通过增加根生物量分配和根冠比来获取更多的养分(
Poorter et al ., 2012 ).此外, 养分贫瘠的条件下, 植物会增加地上部分叶生物量的比例(相对于茎)(
Yan et al ., 2016 ), 但这方面的研究还相对较少.另一方面, 叶片功能性状的调整与资源获取和利用密切相关, 其形态结构和营养元素含量等均会随环境中氮磷条件的改变而发生适应性变化(
Qi et al ., 2009 ).比如, 较低的养分有效性通常会降低比叶面积, 增加叶干物质含量和养分利用效率, 而随着养分有效性的增加, 叶片养分含量显著增加(
Ordo?ez et al ., 2009 ;
林婉奇等, 2019 ).然而, 不同物种对氮磷添加的响应具有物种特异性(
Han et al ., 2013 ;
Mayor et al ., 2014 ).在氮限制条件下, 具有更高根长和根生物量比例的物种占据优势; 相反, 在磷限制条件下, 植物根长不能解释物种间的竞争优势, 菌根菌丝长度、根系分泌物、磷酸酶活性等相关性状起着重要作用(
Venterink & Güsewell, 2010 ;
Minden & Venterink, 2019 ), 这说明植物性状对氮和磷可能具有不同的响应特征, 包括生物量分配、叶片性状或养分利用效率等多个植物性状.由于不同的性状在适应氮限制和磷限制中表现出不同的变化趋势, 植物可能通过调整不同的性状来响应氮变化或磷变化, 但植物性状对氮、磷的响应是否表现出差异并不十分清楚. ...
...
SLA 和
LDMC 是叶片功能性状中最主要的两个指标, 被认为是指示植物养分利用策略的关键性指标(
Jager et al ., 2015 ).与我们的假设(1)一致, 叶片功能性状对氮变化和磷变化的响应具有差异性.随磷浓度的增加,
SLA 显著降低,
LDMC 显著增加, 而氮处理下却没有相似的结果.前人研究表明,
SLA 随养分的减少而降低, 较低的
SLA 和较高的
LDMC 有利于植物适应资源贫瘠的环境(
Jager et al ., 2015 ).然而, 我们的研究中, 高浓度磷水平降低了
SLA, 增加了
LDMC; 相反, 低浓度磷水平增加了
SLA, 降低了
LDMC .高磷水平下低
SLA 、高
LDMC 表明车桑子采取养分的有效保存策略, 低磷水平下高
SLA 、低
LDMC 可提高对光资源的获取能力, 表明车桑子采取快速生长的策略, 该研究结果与
周光霞等(2016) 的研究相似.然而, 低磷水平下车桑子生长依然受到严重的限制.
林婉奇等(2019) 也发现磷添加处理减少了
SLA, 但显著促进了幼苗生长.叶面积和
SLA 呈负相关关系, 叶面积增加可提高植物的光获取能力, 弥补因
SLA 减小而导致的碳摄取效率的损失(
陈延松等, 2019 ).因此, 高磷水平下叶面积的增加对促进车桑子生长和生物量积累具有重要作用. ...
Species-specific responses of foliar nutrients to long-term nitrogen and phosphorus additions in a lowland tropical forest
1
2014
... 在养分有效性发生变化时, 植物会通过生物量分配和叶片功能性状的调整来适应养分变化.按照最优分配理论, 植物会将更多的生物量分配至能够获取限制资源的器官(
Mccarthy & Enquist, 2007 ).在养分有效性较低的情况下通常会通过增加根生物量分配和根冠比来获取更多的养分(
Poorter et al ., 2012 ).此外, 养分贫瘠的条件下, 植物会增加地上部分叶生物量的比例(相对于茎)(
Yan et al ., 2016 ), 但这方面的研究还相对较少.另一方面, 叶片功能性状的调整与资源获取和利用密切相关, 其形态结构和营养元素含量等均会随环境中氮磷条件的改变而发生适应性变化(
Qi et al ., 2009 ).比如, 较低的养分有效性通常会降低比叶面积, 增加叶干物质含量和养分利用效率, 而随着养分有效性的增加, 叶片养分含量显著增加(
Ordo?ez et al ., 2009 ;
林婉奇等, 2019 ).然而, 不同物种对氮磷添加的响应具有物种特异性(
Han et al ., 2013 ;
Mayor et al ., 2014 ).在氮限制条件下, 具有更高根长和根生物量比例的物种占据优势; 相反, 在磷限制条件下, 植物根长不能解释物种间的竞争优势, 菌根菌丝长度、根系分泌物、磷酸酶活性等相关性状起着重要作用(
Venterink & Güsewell, 2010 ;
Minden & Venterink, 2019 ), 这说明植物性状对氮和磷可能具有不同的响应特征, 包括生物量分配、叶片性状或养分利用效率等多个植物性状.由于不同的性状在适应氮限制和磷限制中表现出不同的变化趋势, 植物可能通过调整不同的性状来响应氮变化或磷变化, 但植物性状对氮、磷的响应是否表现出差异并不十分清楚. ...
Consistency between an allometric approach and optimal partitioning theory in global patterns of plant biomass allocation
2
2007
... 在养分有效性发生变化时, 植物会通过生物量分配和叶片功能性状的调整来适应养分变化.按照最优分配理论, 植物会将更多的生物量分配至能够获取限制资源的器官(
Mccarthy & Enquist, 2007 ).在养分有效性较低的情况下通常会通过增加根生物量分配和根冠比来获取更多的养分(
Poorter et al ., 2012 ).此外, 养分贫瘠的条件下, 植物会增加地上部分叶生物量的比例(相对于茎)(
Yan et al ., 2016 ), 但这方面的研究还相对较少.另一方面, 叶片功能性状的调整与资源获取和利用密切相关, 其形态结构和营养元素含量等均会随环境中氮磷条件的改变而发生适应性变化(
Qi et al ., 2009 ).比如, 较低的养分有效性通常会降低比叶面积, 增加叶干物质含量和养分利用效率, 而随着养分有效性的增加, 叶片养分含量显著增加(
Ordo?ez et al ., 2009 ;
林婉奇等, 2019 ).然而, 不同物种对氮磷添加的响应具有物种特异性(
Han et al ., 2013 ;
Mayor et al ., 2014 ).在氮限制条件下, 具有更高根长和根生物量比例的物种占据优势; 相反, 在磷限制条件下, 植物根长不能解释物种间的竞争优势, 菌根菌丝长度、根系分泌物、磷酸酶活性等相关性状起着重要作用(
Venterink & Güsewell, 2010 ;
Minden & Venterink, 2019 ), 这说明植物性状对氮和磷可能具有不同的响应特征, 包括生物量分配、叶片性状或养分利用效率等多个植物性状.由于不同的性状在适应氮限制和磷限制中表现出不同的变化趋势, 植物可能通过调整不同的性状来响应氮变化或磷变化, 但植物性状对氮、磷的响应是否表现出差异并不十分清楚. ...
... 从车桑子生长特征与生物量分配、叶性状的相关性来看, 氮处理条件下, 株高、基径、生物量等生长性状均与根冠比呈显著负相关关系, 与叶片氮含量显著正相关, 但生长性状与叶片氮含量的相关性更强, 表明叶片氮含量的调整对车桑子适应低氮具有重要意义; 而磷处理下, 株高、基径、生物量与
SLA 、叶片氮含量显著负相关, 与
LDMC 显著正相关, 表明叶片功能性状的调整在车桑子适应低磷环境中发挥重要作用.
周俊宏等(2015) 针对紫楠(
Phoebe sheareri )幼苗的研究表明, 相比叶性状的调整, 紫楠幼苗主要通过生物量分配比例的改变来适应不同生境的变化.我们的研究与之相反, 无论氮处理还是磷处理条件下, 相比生物量分配, 叶片性状的调整是车桑子对养分有效性变化更为主要的适应策略.生物量分配的变化也是一系列植物性状对环境变化适应的结果(
Mccarthy & Enquist, 2007 ).因此, 叶性状的调整是车桑子对养分变化更为主要的适应策略.我们的结果表明, 车桑子在响应氮、磷限制时会采用不同的适应策略, 表征其适应策略的性状及性状间的权衡策略亦有所差异.然而, 本研究是基于短期试验条件下的单因素控制试验, 由于氮、磷之间存在交互作用, 不同组合的氮磷添加对车桑子幼苗生长性状的影响值得进一步研究. ...
Nitrogen and phosphorus limitation over long-term ecosystem development in terrestrial ecosystems
1
2012
... 氮和磷是构成生命体生物膜、蛋白质、核酸、磷脂和ATP等物质的重要成分, 因此是植物生命过程中必需的营养元素, 但植物生长普遍会受到氮磷养分的限制(
Yan et al ., 2015 ).氮、磷的限制格局和驱动因素各不相同.通常认为, 演替早期的生态系统主要受到氮的限制, 而演替后期的生态系统主要受到磷的限制(
Menge et al ., 2012 ).淋溶、挥发、反硝化作用和微生物对氮有限的固定作用等都可引起植物氮的缺乏(
Vitousek & Howarth, 1991 ).相比之下, 土壤物理屏蔽、低磷母岩、闭蓄态转化和人类活动(增加其他资源特别是氮的供给)等是引起植物生长过程中磷限制的主要原因(
Vitousek et al ., 2010 ).从植物个体的适应性出发, 充分利用植物自身对养分限制的适应性, 对提高植物生产力具有重要的意义.因此, 植物性状对氮、磷的响应特征对生态系统生产力和生态系统功能具有重要影响(
Wang et al ., 2018 ;
Xu et al ., 2018 ;
Zhang et al ., 2019 ). ...
Plant traits and species interactions along gradients of N, P and K availabilities
2
2019
... 在养分有效性发生变化时, 植物会通过生物量分配和叶片功能性状的调整来适应养分变化.按照最优分配理论, 植物会将更多的生物量分配至能够获取限制资源的器官(
Mccarthy & Enquist, 2007 ).在养分有效性较低的情况下通常会通过增加根生物量分配和根冠比来获取更多的养分(
Poorter et al ., 2012 ).此外, 养分贫瘠的条件下, 植物会增加地上部分叶生物量的比例(相对于茎)(
Yan et al ., 2016 ), 但这方面的研究还相对较少.另一方面, 叶片功能性状的调整与资源获取和利用密切相关, 其形态结构和营养元素含量等均会随环境中氮磷条件的改变而发生适应性变化(
Qi et al ., 2009 ).比如, 较低的养分有效性通常会降低比叶面积, 增加叶干物质含量和养分利用效率, 而随着养分有效性的增加, 叶片养分含量显著增加(
Ordo?ez et al ., 2009 ;
林婉奇等, 2019 ).然而, 不同物种对氮磷添加的响应具有物种特异性(
Han et al ., 2013 ;
Mayor et al ., 2014 ).在氮限制条件下, 具有更高根长和根生物量比例的物种占据优势; 相反, 在磷限制条件下, 植物根长不能解释物种间的竞争优势, 菌根菌丝长度、根系分泌物、磷酸酶活性等相关性状起着重要作用(
Venterink & Güsewell, 2010 ;
Minden & Venterink, 2019 ), 这说明植物性状对氮和磷可能具有不同的响应特征, 包括生物量分配、叶片性状或养分利用效率等多个植物性状.由于不同的性状在适应氮限制和磷限制中表现出不同的变化趋势, 植物可能通过调整不同的性状来响应氮变化或磷变化, 但植物性状对氮、磷的响应是否表现出差异并不十分清楚. ...
... 在各磷浓度处理下, 车桑子生长和生物量积累随磷浓度的增加而显著增加.野外条件下, 车桑子生长区的土壤有效磷含量为4.90 mg·kg
-1 , 同理换算出来为1.58 mmol·L
-1 , 高于本研究的P3水平, 但低于P4水平.车桑子对野外磷水平的适应进化在一定程度上可以解释本研究低浓度和中等浓度磷处理对车桑子生长的限制作用.然而, 低浓度磷水平下车桑子根冠比并没有显著增加, 表明车桑子不会通过调节根冠比来适应低磷环境, 这与低氮处理结果不同.与我们的假设(1)一致, 车桑子生物量分配对氮变化和磷变化的响应具有差异性.Venterink和Güsewell (2010)认为, 磷限制条件下, 由于根系生长需要消耗的能量较多, 而且生长过程中伴随着细胞分裂, 需要大量的DNA和RNA合成(这两种物质中含有大量的磷), 需要更多的磷.因此, 植物不会大量投资根系来吸收磷离子, 而是通过其他机制促进磷的吸收, 比如磷酸酶活性(
Minden & Venterink, 2019 )、根系分泌物、钙离子吸收(
Devau et al ., 2010 )等.本研究中, 低浓度磷水平下叶/茎生物量比例显著增加, 表明低磷处理增加了地上部分叶生物量的分配.由于茎的生长也需要较多的磷, 在缺磷时, 植物为了保住最关键的功能, 可能会增加叶片投入以维持光合作用, 而减少茎的支出.研究表明, 叶片氮与光合作用直接相关, 氮是光合作用的首要影响因子, 而叶片磷对光合作用的影响较弱(
Reich et al ., 2009 ), 这也反映了车桑子通过维持叶片生长以提高在低磷条件下的适应能力. ...
科尔沁沙地优势固沙灌木叶片氮磷化学计量内稳性
1
2019
... 本研究中, 不同的氮浓度和磷浓度处理显著影响了车桑子叶片氮含量, 但叶片磷含量和叶片N:P并未受到显著影响.由于植物具有保持自身化学组成相对稳定的能力, 环境中的养分变化对植物氮磷元素的影响也取决于植物化学计量内稳性的强弱(
Persson et al ., 2010 ).植物内稳性特征可用内稳性指数
H 进行界定, 根据内稳性指数可将植物分为四类:
H > 4为稳态型; 2 <
H < 4为弱稳态型; 4/3 <
H < 2为弱敏感型;
H < 4/3为敏感型(
Persson et al ., 2010 ).本研究中, 不同氮浓度处理下, 叶片氮含量和叶片N:P的
H 分别为6.86和7.31, 均大于4, 呈稳态型, 并且叶片N:P的内稳性大于氮的内稳性.其他研究也表明, 氮磷比的内稳性大于氮和磷的内稳性(
Yu et al ., 2011 ;
宁志英等, 2019 ).这说明, 植物对其自身内稳性的调节主要体现在N:P而不是单一的元素含量上.然而, 不同磷浓度处理下, 叶片磷含量、叶片N:P与营养液的磷浓度和营养液N:P并没有显著的幂函数关系, 其内稳性指数计算结果为负值.
Xing等(2015) 用1/
H 来衡量内稳性的强弱, 也测出内稳性指数为负值, 并将其归为严格内稳型.这说明磷处理条件下, 车桑子叶片磷含量和N:P具有极强的内稳性.较高的内稳性意味着养分元素变化小, 表明具有保守的养分利用策略, 也就更容易适应贫瘠环境而成为优势种(
Yu et al ., 2010 ).不同生长期植物内稳性不同, 内稳性随着植物生长发育逐渐增强(
Yu et al ., 2011 ).本研究中车桑子幼苗已表现出较高的内稳性, 因此, 这可能是促进车桑子成为干热河谷优势植物的原因之一.此外, 所有处理条件下, 车桑子的叶片N:P均在30左右波动.按照叶片临界N:P对限制元素的判断理论, 车桑子生长受到磷限制(
Güsewell, 2004 ;
Sullivan et al ., 2014 ), 这和氮磷添加对车桑子生长和生物量积累的结果是不一致的.与前人的研究观点(
Drenovsky & Richards, 2004 ;
Zhang et al ., 2004 )比较一致, 我们的结果也表明, 在物种水平上, 用叶片N:P来指示植物所受到的养分限制类型是不可靠的. ...
科尔沁沙地优势固沙灌木叶片氮磷化学计量内稳性
1
2019
... 本研究中, 不同的氮浓度和磷浓度处理显著影响了车桑子叶片氮含量, 但叶片磷含量和叶片N:P并未受到显著影响.由于植物具有保持自身化学组成相对稳定的能力, 环境中的养分变化对植物氮磷元素的影响也取决于植物化学计量内稳性的强弱(
Persson et al ., 2010 ).植物内稳性特征可用内稳性指数
H 进行界定, 根据内稳性指数可将植物分为四类:
H > 4为稳态型; 2 <
H < 4为弱稳态型; 4/3 <
H < 2为弱敏感型;
H < 4/3为敏感型(
Persson et al ., 2010 ).本研究中, 不同氮浓度处理下, 叶片氮含量和叶片N:P的
H 分别为6.86和7.31, 均大于4, 呈稳态型, 并且叶片N:P的内稳性大于氮的内稳性.其他研究也表明, 氮磷比的内稳性大于氮和磷的内稳性(
Yu et al ., 2011 ;
宁志英等, 2019 ).这说明, 植物对其自身内稳性的调节主要体现在N:P而不是单一的元素含量上.然而, 不同磷浓度处理下, 叶片磷含量、叶片N:P与营养液的磷浓度和营养液N:P并没有显著的幂函数关系, 其内稳性指数计算结果为负值.
Xing等(2015) 用1/
H 来衡量内稳性的强弱, 也测出内稳性指数为负值, 并将其归为严格内稳型.这说明磷处理条件下, 车桑子叶片磷含量和N:P具有极强的内稳性.较高的内稳性意味着养分元素变化小, 表明具有保守的养分利用策略, 也就更容易适应贫瘠环境而成为优势种(
Yu et al ., 2010 ).不同生长期植物内稳性不同, 内稳性随着植物生长发育逐渐增强(
Yu et al ., 2011 ).本研究中车桑子幼苗已表现出较高的内稳性, 因此, 这可能是促进车桑子成为干热河谷优势植物的原因之一.此外, 所有处理条件下, 车桑子的叶片N:P均在30左右波动.按照叶片临界N:P对限制元素的判断理论, 车桑子生长受到磷限制(
Güsewell, 2004 ;
Sullivan et al ., 2014 ), 这和氮磷添加对车桑子生长和生物量积累的结果是不一致的.与前人的研究观点(
Drenovsky & Richards, 2004 ;
Zhang et al ., 2004 )比较一致, 我们的结果也表明, 在物种水平上, 用叶片N:P来指示植物所受到的养分限制类型是不可靠的. ...
A global study of relationships between leaf traits, climate and soil measures of nutrient fertility
1
2009
... 在养分有效性发生变化时, 植物会通过生物量分配和叶片功能性状的调整来适应养分变化.按照最优分配理论, 植物会将更多的生物量分配至能够获取限制资源的器官(
Mccarthy & Enquist, 2007 ).在养分有效性较低的情况下通常会通过增加根生物量分配和根冠比来获取更多的养分(
Poorter et al ., 2012 ).此外, 养分贫瘠的条件下, 植物会增加地上部分叶生物量的比例(相对于茎)(
Yan et al ., 2016 ), 但这方面的研究还相对较少.另一方面, 叶片功能性状的调整与资源获取和利用密切相关, 其形态结构和营养元素含量等均会随环境中氮磷条件的改变而发生适应性变化(
Qi et al ., 2009 ).比如, 较低的养分有效性通常会降低比叶面积, 增加叶干物质含量和养分利用效率, 而随着养分有效性的增加, 叶片养分含量显著增加(
Ordo?ez et al ., 2009 ;
林婉奇等, 2019 ).然而, 不同物种对氮磷添加的响应具有物种特异性(
Han et al ., 2013 ;
Mayor et al ., 2014 ).在氮限制条件下, 具有更高根长和根生物量比例的物种占据优势; 相反, 在磷限制条件下, 植物根长不能解释物种间的竞争优势, 菌根菌丝长度、根系分泌物、磷酸酶活性等相关性状起着重要作用(
Venterink & Güsewell, 2010 ;
Minden & Venterink, 2019 ), 这说明植物性状对氮和磷可能具有不同的响应特征, 包括生物量分配、叶片性状或养分利用效率等多个植物性状.由于不同的性状在适应氮限制和磷限制中表现出不同的变化趋势, 植物可能通过调整不同的性状来响应氮变化或磷变化, 但植物性状对氮、磷的响应是否表现出差异并不十分清楚. ...
To be or not to be what you eat: regulation of stoichiometric homeostasis among autotrophs and heterotrophs
2
2010
... 本研究中, 不同的氮浓度和磷浓度处理显著影响了车桑子叶片氮含量, 但叶片磷含量和叶片N:P并未受到显著影响.由于植物具有保持自身化学组成相对稳定的能力, 环境中的养分变化对植物氮磷元素的影响也取决于植物化学计量内稳性的强弱(
Persson et al ., 2010 ).植物内稳性特征可用内稳性指数
H 进行界定, 根据内稳性指数可将植物分为四类:
H > 4为稳态型; 2 <
H < 4为弱稳态型; 4/3 <
H < 2为弱敏感型;
H < 4/3为敏感型(
Persson et al ., 2010 ).本研究中, 不同氮浓度处理下, 叶片氮含量和叶片N:P的
H 分别为6.86和7.31, 均大于4, 呈稳态型, 并且叶片N:P的内稳性大于氮的内稳性.其他研究也表明, 氮磷比的内稳性大于氮和磷的内稳性(
Yu et al ., 2011 ;
宁志英等, 2019 ).这说明, 植物对其自身内稳性的调节主要体现在N:P而不是单一的元素含量上.然而, 不同磷浓度处理下, 叶片磷含量、叶片N:P与营养液的磷浓度和营养液N:P并没有显著的幂函数关系, 其内稳性指数计算结果为负值.
Xing等(2015) 用1/
H 来衡量内稳性的强弱, 也测出内稳性指数为负值, 并将其归为严格内稳型.这说明磷处理条件下, 车桑子叶片磷含量和N:P具有极强的内稳性.较高的内稳性意味着养分元素变化小, 表明具有保守的养分利用策略, 也就更容易适应贫瘠环境而成为优势种(
Yu et al ., 2010 ).不同生长期植物内稳性不同, 内稳性随着植物生长发育逐渐增强(
Yu et al ., 2011 ).本研究中车桑子幼苗已表现出较高的内稳性, 因此, 这可能是促进车桑子成为干热河谷优势植物的原因之一.此外, 所有处理条件下, 车桑子的叶片N:P均在30左右波动.按照叶片临界N:P对限制元素的判断理论, 车桑子生长受到磷限制(
Güsewell, 2004 ;
Sullivan et al ., 2014 ), 这和氮磷添加对车桑子生长和生物量积累的结果是不一致的.与前人的研究观点(
Drenovsky & Richards, 2004 ;
Zhang et al ., 2004 )比较一致, 我们的结果也表明, 在物种水平上, 用叶片N:P来指示植物所受到的养分限制类型是不可靠的. ...
... < 4/3为敏感型(
Persson et al ., 2010 ).本研究中, 不同氮浓度处理下, 叶片氮含量和叶片N:P的
H 分别为6.86和7.31, 均大于4, 呈稳态型, 并且叶片N:P的内稳性大于氮的内稳性.其他研究也表明, 氮磷比的内稳性大于氮和磷的内稳性(
Yu et al ., 2011 ;
宁志英等, 2019 ).这说明, 植物对其自身内稳性的调节主要体现在N:P而不是单一的元素含量上.然而, 不同磷浓度处理下, 叶片磷含量、叶片N:P与营养液的磷浓度和营养液N:P并没有显著的幂函数关系, 其内稳性指数计算结果为负值.
Xing等(2015) 用1/
H 来衡量内稳性的强弱, 也测出内稳性指数为负值, 并将其归为严格内稳型.这说明磷处理条件下, 车桑子叶片磷含量和N:P具有极强的内稳性.较高的内稳性意味着养分元素变化小, 表明具有保守的养分利用策略, 也就更容易适应贫瘠环境而成为优势种(
Yu et al ., 2010 ).不同生长期植物内稳性不同, 内稳性随着植物生长发育逐渐增强(
Yu et al ., 2011 ).本研究中车桑子幼苗已表现出较高的内稳性, 因此, 这可能是促进车桑子成为干热河谷优势植物的原因之一.此外, 所有处理条件下, 车桑子的叶片N:P均在30左右波动.按照叶片临界N:P对限制元素的判断理论, 车桑子生长受到磷限制(
Güsewell, 2004 ;
Sullivan et al ., 2014 ), 这和氮磷添加对车桑子生长和生物量积累的结果是不一致的.与前人的研究观点(
Drenovsky & Richards, 2004 ;
Zhang et al ., 2004 )比较一致, 我们的结果也表明, 在物种水平上, 用叶片N:P来指示植物所受到的养分限制类型是不可靠的. ...
Biomass allocation to leaves, stems and roots: meta-analyses of interspecific variation and environmental control
1
2012
... 在养分有效性发生变化时, 植物会通过生物量分配和叶片功能性状的调整来适应养分变化.按照最优分配理论, 植物会将更多的生物量分配至能够获取限制资源的器官(
Mccarthy & Enquist, 2007 ).在养分有效性较低的情况下通常会通过增加根生物量分配和根冠比来获取更多的养分(
Poorter et al ., 2012 ).此外, 养分贫瘠的条件下, 植物会增加地上部分叶生物量的比例(相对于茎)(
Yan et al ., 2016 ), 但这方面的研究还相对较少.另一方面, 叶片功能性状的调整与资源获取和利用密切相关, 其形态结构和营养元素含量等均会随环境中氮磷条件的改变而发生适应性变化(
Qi et al ., 2009 ).比如, 较低的养分有效性通常会降低比叶面积, 增加叶干物质含量和养分利用效率, 而随着养分有效性的增加, 叶片养分含量显著增加(
Ordo?ez et al ., 2009 ;
林婉奇等, 2019 ).然而, 不同物种对氮磷添加的响应具有物种特异性(
Han et al ., 2013 ;
Mayor et al ., 2014 ).在氮限制条件下, 具有更高根长和根生物量比例的物种占据优势; 相反, 在磷限制条件下, 植物根长不能解释物种间的竞争优势, 菌根菌丝长度、根系分泌物、磷酸酶活性等相关性状起着重要作用(
Venterink & Güsewell, 2010 ;
Minden & Venterink, 2019 ), 这说明植物性状对氮和磷可能具有不同的响应特征, 包括生物量分配、叶片性状或养分利用效率等多个植物性状.由于不同的性状在适应氮限制和磷限制中表现出不同的变化趋势, 植物可能通过调整不同的性状来响应氮变化或磷变化, 但植物性状对氮、磷的响应是否表现出差异并不十分清楚. ...
Leaf-trait relationships of
Quercus liaotungensis along an altitudinal gradient in Dongling Mountain, Beijing
1
2009
... 在养分有效性发生变化时, 植物会通过生物量分配和叶片功能性状的调整来适应养分变化.按照最优分配理论, 植物会将更多的生物量分配至能够获取限制资源的器官(
Mccarthy & Enquist, 2007 ).在养分有效性较低的情况下通常会通过增加根生物量分配和根冠比来获取更多的养分(
Poorter et al ., 2012 ).此外, 养分贫瘠的条件下, 植物会增加地上部分叶生物量的比例(相对于茎)(
Yan et al ., 2016 ), 但这方面的研究还相对较少.另一方面, 叶片功能性状的调整与资源获取和利用密切相关, 其形态结构和营养元素含量等均会随环境中氮磷条件的改变而发生适应性变化(
Qi et al ., 2009 ).比如, 较低的养分有效性通常会降低比叶面积, 增加叶干物质含量和养分利用效率, 而随着养分有效性的增加, 叶片养分含量显著增加(
Ordo?ez et al ., 2009 ;
林婉奇等, 2019 ).然而, 不同物种对氮磷添加的响应具有物种特异性(
Han et al ., 2013 ;
Mayor et al ., 2014 ).在氮限制条件下, 具有更高根长和根生物量比例的物种占据优势; 相反, 在磷限制条件下, 植物根长不能解释物种间的竞争优势, 菌根菌丝长度、根系分泌物、磷酸酶活性等相关性状起着重要作用(
Venterink & Güsewell, 2010 ;
Minden & Venterink, 2019 ), 这说明植物性状对氮和磷可能具有不同的响应特征, 包括生物量分配、叶片性状或养分利用效率等多个植物性状.由于不同的性状在适应氮限制和磷限制中表现出不同的变化趋势, 植物可能通过调整不同的性状来响应氮变化或磷变化, 但植物性状对氮、磷的响应是否表现出差异并不十分清楚. ...
Leaf phosphorus influences the photosynthesis-nitrogen relation: a cross- biome analysis of 314 species
1
2009
... 在各磷浓度处理下, 车桑子生长和生物量积累随磷浓度的增加而显著增加.野外条件下, 车桑子生长区的土壤有效磷含量为4.90 mg·kg
-1 , 同理换算出来为1.58 mmol·L
-1 , 高于本研究的P3水平, 但低于P4水平.车桑子对野外磷水平的适应进化在一定程度上可以解释本研究低浓度和中等浓度磷处理对车桑子生长的限制作用.然而, 低浓度磷水平下车桑子根冠比并没有显著增加, 表明车桑子不会通过调节根冠比来适应低磷环境, 这与低氮处理结果不同.与我们的假设(1)一致, 车桑子生物量分配对氮变化和磷变化的响应具有差异性.Venterink和Güsewell (2010)认为, 磷限制条件下, 由于根系生长需要消耗的能量较多, 而且生长过程中伴随着细胞分裂, 需要大量的DNA和RNA合成(这两种物质中含有大量的磷), 需要更多的磷.因此, 植物不会大量投资根系来吸收磷离子, 而是通过其他机制促进磷的吸收, 比如磷酸酶活性(
Minden & Venterink, 2019 )、根系分泌物、钙离子吸收(
Devau et al ., 2010 )等.本研究中, 低浓度磷水平下叶/茎生物量比例显著增加, 表明低磷处理增加了地上部分叶生物量的分配.由于茎的生长也需要较多的磷, 在缺磷时, 植物为了保住最关键的功能, 可能会增加叶片投入以维持光合作用, 而减少茎的支出.研究表明, 叶片氮与光合作用直接相关, 氮是光合作用的首要影响因子, 而叶片磷对光合作用的影响较弱(
Reich et al ., 2009 ), 这也反映了车桑子通过维持叶片生长以提高在低磷条件下的适应能力. ...
Roles of arbuscular mycorrhizas in plant nutrition and growth: new paradigms from cellular to ecosystem scales
1
2011
... 需要说明的是, 本研究是基于室内砂培实验条件下得出的结果, 砂培条件和野外自然环境存在较大差异, 该试验结果与野外环境的试验结果是否一致并不确定.研究表明, 丛枝菌根真菌对植物养分吸收尤其是磷的吸收具有重大的贡献(
Smith et al ., 2004 ;
Smith & Smith, 2011 ), 并且, 干热河谷自然植被的根际土壤中大多有丰富的丛枝菌根真菌(
Li et al ., 2010 ).在这种条件下, 我们推测, 车桑子可能已经形成了相关的内部信号调控机制, 主要依赖菌根真菌吸收磷, 而减少了自身对低磷的生理生长调节能力, 使得根系分配不会对磷缺乏形成响应, 这也进一步解释了低磷条件下根冠比没有发生显著增加.而本研究中, 砂培条件缺乏菌根真菌的侵染, 车桑子自身对低磷的适应能力较差, 因此, 低磷水平限制了车桑子生长.在野外土壤环境条件下, 车桑子对氮磷有效性变化的响应特征是否与本研究不同, 尚需进一步验证. ...
Functional diversity in arbuscular mycorrhizal (AM) symbioses: the contribution of the mycorrhizal P uptake pathway is not correlated with mycorrhizal responses in growth or total P uptake
1
2004
... 需要说明的是, 本研究是基于室内砂培实验条件下得出的结果, 砂培条件和野外自然环境存在较大差异, 该试验结果与野外环境的试验结果是否一致并不确定.研究表明, 丛枝菌根真菌对植物养分吸收尤其是磷的吸收具有重大的贡献(
Smith et al ., 2004 ;
Smith & Smith, 2011 ), 并且, 干热河谷自然植被的根际土壤中大多有丰富的丛枝菌根真菌(
Li et al ., 2010 ).在这种条件下, 我们推测, 车桑子可能已经形成了相关的内部信号调控机制, 主要依赖菌根真菌吸收磷, 而减少了自身对低磷的生理生长调节能力, 使得根系分配不会对磷缺乏形成响应, 这也进一步解释了低磷条件下根冠比没有发生显著增加.而本研究中, 砂培条件缺乏菌根真菌的侵染, 车桑子自身对低磷的适应能力较差, 因此, 低磷水平限制了车桑子生长.在野外土壤环境条件下, 车桑子对氮磷有效性变化的响应特征是否与本研究不同, 尚需进一步验证. ...
松嫩草地80种草本植物叶片氮磷化学计量特征
2012
松嫩草地80种草本植物叶片氮磷化学计量特征
2012
Assessing nutrient limitation in complex forested ecosystems: alternatives to large-scale fertilization experiments
2
2014
... 在植物养分限制的判断中, 若一种有效态元素的添加引起了植物生化过程速率或生物量的增加, 则意味着存在这种养分的限制(
Vitousek et al ., 2010 ;
Sullivan et al ., 2014 ).本研究表明, 高浓度氮水平显著促进了车桑子的生长和生物量积累, 但车桑子生长和生物量积累在低浓度和中等浓度氮水平下没有显著差异.野外条件下, 车桑子生长区的土壤有效氮(硝态氮与铵态氮之和)含量大约为11.7 mg·kg
-1 , 考虑到土壤中水分含量在10%左右, 也就是每kg土壤约含0.1 L水, 那么土壤中的有效氮含量约为117 mg·L
-1 , 介于本研究设置的N3 (210 mg·L
-1 )和N2 (70 mg·L
-1 )水平之间.因此, 本研究中, N1-N3处理下车桑子的生长没有显著差异, 可能是由于干热河谷长期的氮限制使得植物形成了对低氮环境的一系列形态与生理适应性.干旱生态系统氮限制尤为普遍(
Yahdjian et al ., 2011 ).已有研究表明, 氮是限制金沙江干旱河谷退化草地系统恢复的主要因素(
张彦东等, 2004 ).同时, 我们的结果表明, 车桑子主要通过增加对氮的吸收和利用来适应低氮环境.一方面, 车桑子在低氮条件下, 其根冠比显著增加可促进氮的吸收; 另一方面, 吸收的氮的利用效率又显著提高.根冠比的增加和养分利用效率的提高均是植物适应养分限制的主要机制(
冷华妮等, 2009 ;
Venterink & Güsewell, 2010 ).然而, 由于本研究中养分添加只持续了一个生长季, 试验时间较短, 鉴于车桑子幼苗长期对氮限制的适应, 对试验中的低浓度氮水平可能还不太敏感, 通过根冠比的调整和养分利用效率的提高即可保持其正常的生长和生物量积累, 但在长期的低浓度氮水平下车桑子生长和生物量积累可能会受到限制. ...
... 本研究中, 不同的氮浓度和磷浓度处理显著影响了车桑子叶片氮含量, 但叶片磷含量和叶片N:P并未受到显著影响.由于植物具有保持自身化学组成相对稳定的能力, 环境中的养分变化对植物氮磷元素的影响也取决于植物化学计量内稳性的强弱(
Persson et al ., 2010 ).植物内稳性特征可用内稳性指数
H 进行界定, 根据内稳性指数可将植物分为四类:
H > 4为稳态型; 2 <
H < 4为弱稳态型; 4/3 <
H < 2为弱敏感型;
H < 4/3为敏感型(
Persson et al ., 2010 ).本研究中, 不同氮浓度处理下, 叶片氮含量和叶片N:P的
H 分别为6.86和7.31, 均大于4, 呈稳态型, 并且叶片N:P的内稳性大于氮的内稳性.其他研究也表明, 氮磷比的内稳性大于氮和磷的内稳性(
Yu et al ., 2011 ;
宁志英等, 2019 ).这说明, 植物对其自身内稳性的调节主要体现在N:P而不是单一的元素含量上.然而, 不同磷浓度处理下, 叶片磷含量、叶片N:P与营养液的磷浓度和营养液N:P并没有显著的幂函数关系, 其内稳性指数计算结果为负值.
Xing等(2015) 用1/
H 来衡量内稳性的强弱, 也测出内稳性指数为负值, 并将其归为严格内稳型.这说明磷处理条件下, 车桑子叶片磷含量和N:P具有极强的内稳性.较高的内稳性意味着养分元素变化小, 表明具有保守的养分利用策略, 也就更容易适应贫瘠环境而成为优势种(
Yu et al ., 2010 ).不同生长期植物内稳性不同, 内稳性随着植物生长发育逐渐增强(
Yu et al ., 2011 ).本研究中车桑子幼苗已表现出较高的内稳性, 因此, 这可能是促进车桑子成为干热河谷优势植物的原因之一.此外, 所有处理条件下, 车桑子的叶片N:P均在30左右波动.按照叶片临界N:P对限制元素的判断理论, 车桑子生长受到磷限制(
Güsewell, 2004 ;
Sullivan et al ., 2014 ), 这和氮磷添加对车桑子生长和生物量积累的结果是不一致的.与前人的研究观点(
Drenovsky & Richards, 2004 ;
Zhang et al ., 2004 )比较一致, 我们的结果也表明, 在物种水平上, 用叶片N:P来指示植物所受到的养分限制类型是不可靠的. ...
Competitive interactions between two meadow grasses under nitrogen and phosphorus limitation
3
2010
... 在养分有效性发生变化时, 植物会通过生物量分配和叶片功能性状的调整来适应养分变化.按照最优分配理论, 植物会将更多的生物量分配至能够获取限制资源的器官(
Mccarthy & Enquist, 2007 ).在养分有效性较低的情况下通常会通过增加根生物量分配和根冠比来获取更多的养分(
Poorter et al ., 2012 ).此外, 养分贫瘠的条件下, 植物会增加地上部分叶生物量的比例(相对于茎)(
Yan et al ., 2016 ), 但这方面的研究还相对较少.另一方面, 叶片功能性状的调整与资源获取和利用密切相关, 其形态结构和营养元素含量等均会随环境中氮磷条件的改变而发生适应性变化(
Qi et al ., 2009 ).比如, 较低的养分有效性通常会降低比叶面积, 增加叶干物质含量和养分利用效率, 而随着养分有效性的增加, 叶片养分含量显著增加(
Ordo?ez et al ., 2009 ;
林婉奇等, 2019 ).然而, 不同物种对氮磷添加的响应具有物种特异性(
Han et al ., 2013 ;
Mayor et al ., 2014 ).在氮限制条件下, 具有更高根长和根生物量比例的物种占据优势; 相反, 在磷限制条件下, 植物根长不能解释物种间的竞争优势, 菌根菌丝长度、根系分泌物、磷酸酶活性等相关性状起着重要作用(
Venterink & Güsewell, 2010 ;
Minden & Venterink, 2019 ), 这说明植物性状对氮和磷可能具有不同的响应特征, 包括生物量分配、叶片性状或养分利用效率等多个植物性状.由于不同的性状在适应氮限制和磷限制中表现出不同的变化趋势, 植物可能通过调整不同的性状来响应氮变化或磷变化, 但植物性状对氮、磷的响应是否表现出差异并不十分清楚. ...
... 在植物养分限制的判断中, 若一种有效态元素的添加引起了植物生化过程速率或生物量的增加, 则意味着存在这种养分的限制(
Vitousek et al ., 2010 ;
Sullivan et al ., 2014 ).本研究表明, 高浓度氮水平显著促进了车桑子的生长和生物量积累, 但车桑子生长和生物量积累在低浓度和中等浓度氮水平下没有显著差异.野外条件下, 车桑子生长区的土壤有效氮(硝态氮与铵态氮之和)含量大约为11.7 mg·kg
-1 , 考虑到土壤中水分含量在10%左右, 也就是每kg土壤约含0.1 L水, 那么土壤中的有效氮含量约为117 mg·L
-1 , 介于本研究设置的N3 (210 mg·L
-1 )和N2 (70 mg·L
-1 )水平之间.因此, 本研究中, N1-N3处理下车桑子的生长没有显著差异, 可能是由于干热河谷长期的氮限制使得植物形成了对低氮环境的一系列形态与生理适应性.干旱生态系统氮限制尤为普遍(
Yahdjian et al ., 2011 ).已有研究表明, 氮是限制金沙江干旱河谷退化草地系统恢复的主要因素(
张彦东等, 2004 ).同时, 我们的结果表明, 车桑子主要通过增加对氮的吸收和利用来适应低氮环境.一方面, 车桑子在低氮条件下, 其根冠比显著增加可促进氮的吸收; 另一方面, 吸收的氮的利用效率又显著提高.根冠比的增加和养分利用效率的提高均是植物适应养分限制的主要机制(
冷华妮等, 2009 ;
Venterink & Güsewell, 2010 ).然而, 由于本研究中养分添加只持续了一个生长季, 试验时间较短, 鉴于车桑子幼苗长期对氮限制的适应, 对试验中的低浓度氮水平可能还不太敏感, 通过根冠比的调整和养分利用效率的提高即可保持其正常的生长和生物量积累, 但在长期的低浓度氮水平下车桑子生长和生物量积累可能会受到限制. ...
... 关系, 可见车桑子叶片磷含量的变化是驱动N:P变化的主要因子.
吴统贵等(2010) 和
宋彦涛等(2012) 的研究也表明, 叶片N:P主要受到磷的驱动, N:P的变化受控于叶片磷含量的变化.然而, 氮、磷处理条件下, 车桑子叶片氮和磷含量并没有显著正相关关系, 这可能是由于氮、磷养分不同的内稳性导致的.
黄端等(2017) 也发现叶片氮、磷含量之间无显著的正相关关系.此外, 本研究中, 磷处理下叶片氮含量和
SLA 显著正相关, 而氮处理下却没有这种相关性, 这与以往的研究结果不一致.以往大量的研究发现植物叶片
SLA 与叶片氮含量显著正相关, 二者的关联是植物叶经济型谱的重要组成部分(
于鸿莹等, 2014 ;
熊静等, 2019 ).以往的数据基本是从多个物种之间发现的, 也就是
SLA 与叶片氮含量正相关是种间变异的结果,而本研究是通过设置氮浓度梯度, 是基于环境中氮浓度变化对单个物种内
SLA 与叶片氮含量影响后的结果.在氮有效性增加的情况下, 叶片氮含量的响应更直接更迅速, 植物吸收氮后叶片氮含量显著增加, 而
SLA 的响应相对滞后, 与
Wright和Sutton-Grier (2012) 在氮添加条件下的结果是一致的, 这也反映了长期的氮限制使车桑子在较高的氮水平下短期内依然保持相对保守的氮利用策略.因此, 氮添加条件下车桑子叶片氮含量与
SLA 不相关甚至呈现出较弱的负相关关系.本研究结果表明, 环境中养分驱动下单个物种内叶片的性状变化可能并不完全符合经济型谱理论, 而会受到氮、磷养分有效性的影响, 符合叶经济型谱形成的环境因子主导假说(
赖小红等, 2019 ).此外, 植物功能性状和生物量分配之间存在着某种共轭的变化(
高景等, 2016 ).本研究中, 氮处理下, 车桑子根冠比与叶片氮含量、N:P显著负相关, 磷处理下却没有显著的相关性.以上结果支持我们假设(2)的观点, 车桑子生物量分配与叶性状间存在着一定的协同和权衡关系, 但是车桑子在面临氮、磷有效性的变化时具有不同的响应策略. ...
Nitrogen limitation on land and in the sea: How can it occur?
1
1991
... 氮和磷是构成生命体生物膜、蛋白质、核酸、磷脂和ATP等物质的重要成分, 因此是植物生命过程中必需的营养元素, 但植物生长普遍会受到氮磷养分的限制(
Yan et al ., 2015 ).氮、磷的限制格局和驱动因素各不相同.通常认为, 演替早期的生态系统主要受到氮的限制, 而演替后期的生态系统主要受到磷的限制(
Menge et al ., 2012 ).淋溶、挥发、反硝化作用和微生物对氮有限的固定作用等都可引起植物氮的缺乏(
Vitousek & Howarth, 1991 ).相比之下, 土壤物理屏蔽、低磷母岩、闭蓄态转化和人类活动(增加其他资源特别是氮的供给)等是引起植物生长过程中磷限制的主要原因(
Vitousek et al ., 2010 ).从植物个体的适应性出发, 充分利用植物自身对养分限制的适应性, 对提高植物生产力具有重要的意义.因此, 植物性状对氮、磷的响应特征对生态系统生产力和生态系统功能具有重要影响(
Wang et al ., 2018 ;
Xu et al ., 2018 ;
Zhang et al ., 2019 ). ...
Terrestrial phosphorus limitation: mechanisms, implications, and nitrogen-phosphorus interactions
2
2010
... 氮和磷是构成生命体生物膜、蛋白质、核酸、磷脂和ATP等物质的重要成分, 因此是植物生命过程中必需的营养元素, 但植物生长普遍会受到氮磷养分的限制(
Yan et al ., 2015 ).氮、磷的限制格局和驱动因素各不相同.通常认为, 演替早期的生态系统主要受到氮的限制, 而演替后期的生态系统主要受到磷的限制(
Menge et al ., 2012 ).淋溶、挥发、反硝化作用和微生物对氮有限的固定作用等都可引起植物氮的缺乏(
Vitousek & Howarth, 1991 ).相比之下, 土壤物理屏蔽、低磷母岩、闭蓄态转化和人类活动(增加其他资源特别是氮的供给)等是引起植物生长过程中磷限制的主要原因(
Vitousek et al ., 2010 ).从植物个体的适应性出发, 充分利用植物自身对养分限制的适应性, 对提高植物生产力具有重要的意义.因此, 植物性状对氮、磷的响应特征对生态系统生产力和生态系统功能具有重要影响(
Wang et al ., 2018 ;
Xu et al ., 2018 ;
Zhang et al ., 2019 ). ...
... 在植物养分限制的判断中, 若一种有效态元素的添加引起了植物生化过程速率或生物量的增加, 则意味着存在这种养分的限制(
Vitousek et al ., 2010 ;
Sullivan et al ., 2014 ).本研究表明, 高浓度氮水平显著促进了车桑子的生长和生物量积累, 但车桑子生长和生物量积累在低浓度和中等浓度氮水平下没有显著差异.野外条件下, 车桑子生长区的土壤有效氮(硝态氮与铵态氮之和)含量大约为11.7 mg·kg
-1 , 考虑到土壤中水分含量在10%左右, 也就是每kg土壤约含0.1 L水, 那么土壤中的有效氮含量约为117 mg·L
-1 , 介于本研究设置的N3 (210 mg·L
-1 )和N2 (70 mg·L
-1 )水平之间.因此, 本研究中, N1-N3处理下车桑子的生长没有显著差异, 可能是由于干热河谷长期的氮限制使得植物形成了对低氮环境的一系列形态与生理适应性.干旱生态系统氮限制尤为普遍(
Yahdjian et al ., 2011 ).已有研究表明, 氮是限制金沙江干旱河谷退化草地系统恢复的主要因素(
张彦东等, 2004 ).同时, 我们的结果表明, 车桑子主要通过增加对氮的吸收和利用来适应低氮环境.一方面, 车桑子在低氮条件下, 其根冠比显著增加可促进氮的吸收; 另一方面, 吸收的氮的利用效率又显著提高.根冠比的增加和养分利用效率的提高均是植物适应养分限制的主要机制(
冷华妮等, 2009 ;
Venterink & Güsewell, 2010 ).然而, 由于本研究中养分添加只持续了一个生长季, 试验时间较短, 鉴于车桑子幼苗长期对氮限制的适应, 对试验中的低浓度氮水平可能还不太敏感, 通过根冠比的调整和养分利用效率的提高即可保持其正常的生长和生物量积累, 但在长期的低浓度氮水平下车桑子生长和生物量积累可能会受到限制. ...
Ecosystem nitrogen retention is regulated by plant community trait interactions with nutrient status in an alpine meadow
1
2018
... 氮和磷是构成生命体生物膜、蛋白质、核酸、磷脂和ATP等物质的重要成分, 因此是植物生命过程中必需的营养元素, 但植物生长普遍会受到氮磷养分的限制(
Yan et al ., 2015 ).氮、磷的限制格局和驱动因素各不相同.通常认为, 演替早期的生态系统主要受到氮的限制, 而演替后期的生态系统主要受到磷的限制(
Menge et al ., 2012 ).淋溶、挥发、反硝化作用和微生物对氮有限的固定作用等都可引起植物氮的缺乏(
Vitousek & Howarth, 1991 ).相比之下, 土壤物理屏蔽、低磷母岩、闭蓄态转化和人类活动(增加其他资源特别是氮的供给)等是引起植物生长过程中磷限制的主要原因(
Vitousek et al ., 2010 ).从植物个体的适应性出发, 充分利用植物自身对养分限制的适应性, 对提高植物生产力具有重要的意义.因此, 植物性状对氮、磷的响应特征对生态系统生产力和生态系统功能具有重要影响(
Wang et al ., 2018 ;
Xu et al ., 2018 ;
Zhang et al ., 2019 ). ...
The worldwide leaf economics spectrum
1
2004
... 此外, 如上所述, 植物种内性状具有极大的变异性, 这被认为是植物性状在适应环境中的可塑性决定的.另一方面物种之间性状上也存在极大的差异, 并形成一个叶经济型谱.叶经济型谱理论认为植物叶片在其形态结构、化学组分和生理功能等性状之间存在着相互依存、相互关联的内在关系(
Wright et al ., 2004 ;
于鸿莹等, 2014 ).多种功能性状之间关系紧密, 不同性状的组合可体现出植物对不同环境的适应策略(
熊静等, 2019 ).那么, 在环境梯度如养分梯度下植物种内性状的可塑性及其关联性是否与种间的这种经济型谱变化趋势一致? 例如, 经济型谱中比叶面积与叶片氮含量显著正相关, 那么在氮浓度梯度中植物种内的比叶面积与叶片氮含量是否会产生类似的关系? 植物种内性状的可塑性调整在研究植物适应策略方面有着重要作用.目前,环境变化对叶性状和生物量分配的表型可塑性方面的研究较多, 然而, 在氮磷变化条件下, 植物在生物量分配和叶性状的调整上是否存在如同经济型谱理论的相互变化规律? 这方面的证据还比较有限.开展生物量分配和叶性状对氮磷浓度的响应研究, 有助于全面理解和揭示植物对养分有效性变化的响应与适应机制. ...
Does the leaf economic spectrum hold within local species pools across varying environmental conditions?
1
2012
... 关系, 可见车桑子叶片磷含量的变化是驱动N:P变化的主要因子.
吴统贵等(2010) 和
宋彦涛等(2012) 的研究也表明, 叶片N:P主要受到磷的驱动, N:P的变化受控于叶片磷含量的变化.然而, 氮、磷处理条件下, 车桑子叶片氮和磷含量并没有显著正相关关系, 这可能是由于氮、磷养分不同的内稳性导致的.
黄端等(2017) 也发现叶片氮、磷含量之间无显著的正相关关系.此外, 本研究中, 磷处理下叶片氮含量和
SLA 显著正相关, 而氮处理下却没有这种相关性, 这与以往的研究结果不一致.以往大量的研究发现植物叶片
SLA 与叶片氮含量显著正相关, 二者的关联是植物叶经济型谱的重要组成部分(
于鸿莹等, 2014 ;
熊静等, 2019 ).以往的数据基本是从多个物种之间发现的, 也就是
SLA 与叶片氮含量正相关是种间变异的结果,而本研究是通过设置氮浓度梯度, 是基于环境中氮浓度变化对单个物种内
SLA 与叶片氮含量影响后的结果.在氮有效性增加的情况下, 叶片氮含量的响应更直接更迅速, 植物吸收氮后叶片氮含量显著增加, 而
SLA 的响应相对滞后, 与
Wright和Sutton-Grier (2012) 在氮添加条件下的结果是一致的, 这也反映了长期的氮限制使车桑子在较高的氮水平下短期内依然保持相对保守的氮利用策略.因此, 氮添加条件下车桑子叶片氮含量与
SLA 不相关甚至呈现出较弱的负相关关系.本研究结果表明, 环境中养分驱动下单个物种内叶片的性状变化可能并不完全符合经济型谱理论, 而会受到氮、磷养分有效性的影响, 符合叶经济型谱形成的环境因子主导假说(
赖小红等, 2019 ).此外, 植物功能性状和生物量分配之间存在着某种共轭的变化(
高景等, 2016 ).本研究中, 氮处理下, 车桑子根冠比与叶片氮含量、N:P显著负相关, 磷处理下却没有显著的相关性.以上结果支持我们假设(2)的观点, 车桑子生物量分配与叶性状间存在着一定的协同和权衡关系, 但是车桑子在面临氮、磷有效性的变化时具有不同的响应策略. ...
杭州湾滨海湿地3种草本植物叶片N、P化学计量学的季节变化
1
2010
... 关系, 可见车桑子叶片磷含量的变化是驱动N:P变化的主要因子.
吴统贵等(2010) 和
宋彦涛等(2012) 的研究也表明, 叶片N:P主要受到磷的驱动, N:P的变化受控于叶片磷含量的变化.然而, 氮、磷处理条件下, 车桑子叶片氮和磷含量并没有显著正相关关系, 这可能是由于氮、磷养分不同的内稳性导致的.
黄端等(2017) 也发现叶片氮、磷含量之间无显著的正相关关系.此外, 本研究中, 磷处理下叶片氮含量和
SLA 显著正相关, 而氮处理下却没有这种相关性, 这与以往的研究结果不一致.以往大量的研究发现植物叶片
SLA 与叶片氮含量显著正相关, 二者的关联是植物叶经济型谱的重要组成部分(
于鸿莹等, 2014 ;
熊静等, 2019 ).以往的数据基本是从多个物种之间发现的, 也就是
SLA 与叶片氮含量正相关是种间变异的结果,而本研究是通过设置氮浓度梯度, 是基于环境中氮浓度变化对单个物种内
SLA 与叶片氮含量影响后的结果.在氮有效性增加的情况下, 叶片氮含量的响应更直接更迅速, 植物吸收氮后叶片氮含量显著增加, 而
SLA 的响应相对滞后, 与
Wright和Sutton-Grier (2012) 在氮添加条件下的结果是一致的, 这也反映了长期的氮限制使车桑子在较高的氮水平下短期内依然保持相对保守的氮利用策略.因此, 氮添加条件下车桑子叶片氮含量与
SLA 不相关甚至呈现出较弱的负相关关系.本研究结果表明, 环境中养分驱动下单个物种内叶片的性状变化可能并不完全符合经济型谱理论, 而会受到氮、磷养分有效性的影响, 符合叶经济型谱形成的环境因子主导假说(
赖小红等, 2019 ).此外, 植物功能性状和生物量分配之间存在着某种共轭的变化(
高景等, 2016 ).本研究中, 氮处理下, 车桑子根冠比与叶片氮含量、N:P显著负相关, 磷处理下却没有显著的相关性.以上结果支持我们假设(2)的观点, 车桑子生物量分配与叶性状间存在着一定的协同和权衡关系, 但是车桑子在面临氮、磷有效性的变化时具有不同的响应策略. ...
杭州湾滨海湿地3种草本植物叶片N、P化学计量学的季节变化
1
2010
... 关系, 可见车桑子叶片磷含量的变化是驱动N:P变化的主要因子.
吴统贵等(2010) 和
宋彦涛等(2012) 的研究也表明, 叶片N:P主要受到磷的驱动, N:P的变化受控于叶片磷含量的变化.然而, 氮、磷处理条件下, 车桑子叶片氮和磷含量并没有显著正相关关系, 这可能是由于氮、磷养分不同的内稳性导致的.
黄端等(2017) 也发现叶片氮、磷含量之间无显著的正相关关系.此外, 本研究中, 磷处理下叶片氮含量和
SLA 显著正相关, 而氮处理下却没有这种相关性, 这与以往的研究结果不一致.以往大量的研究发现植物叶片
SLA 与叶片氮含量显著正相关, 二者的关联是植物叶经济型谱的重要组成部分(
于鸿莹等, 2014 ;
熊静等, 2019 ).以往的数据基本是从多个物种之间发现的, 也就是
SLA 与叶片氮含量正相关是种间变异的结果,而本研究是通过设置氮浓度梯度, 是基于环境中氮浓度变化对单个物种内
SLA 与叶片氮含量影响后的结果.在氮有效性增加的情况下, 叶片氮含量的响应更直接更迅速, 植物吸收氮后叶片氮含量显著增加, 而
SLA 的响应相对滞后, 与
Wright和Sutton-Grier (2012) 在氮添加条件下的结果是一致的, 这也反映了长期的氮限制使车桑子在较高的氮水平下短期内依然保持相对保守的氮利用策略.因此, 氮添加条件下车桑子叶片氮含量与
SLA 不相关甚至呈现出较弱的负相关关系.本研究结果表明, 环境中养分驱动下单个物种内叶片的性状变化可能并不完全符合经济型谱理论, 而会受到氮、磷养分有效性的影响, 符合叶经济型谱形成的环境因子主导假说(
赖小红等, 2019 ).此外, 植物功能性状和生物量分配之间存在着某种共轭的变化(
高景等, 2016 ).本研究中, 氮处理下, 车桑子根冠比与叶片氮含量、N:P显著负相关, 磷处理下却没有显著的相关性.以上结果支持我们假设(2)的观点, 车桑子生物量分配与叶性状间存在着一定的协同和权衡关系, 但是车桑子在面临氮、磷有效性的变化时具有不同的响应策略. ...
Multielement stoichiometry of submerged macrophytes across Yunnan plateau lakes (China)
1
2015
... 本研究中, 不同的氮浓度和磷浓度处理显著影响了车桑子叶片氮含量, 但叶片磷含量和叶片N:P并未受到显著影响.由于植物具有保持自身化学组成相对稳定的能力, 环境中的养分变化对植物氮磷元素的影响也取决于植物化学计量内稳性的强弱(
Persson et al ., 2010 ).植物内稳性特征可用内稳性指数
H 进行界定, 根据内稳性指数可将植物分为四类:
H > 4为稳态型; 2 <
H < 4为弱稳态型; 4/3 <
H < 2为弱敏感型;
H < 4/3为敏感型(
Persson et al ., 2010 ).本研究中, 不同氮浓度处理下, 叶片氮含量和叶片N:P的
H 分别为6.86和7.31, 均大于4, 呈稳态型, 并且叶片N:P的内稳性大于氮的内稳性.其他研究也表明, 氮磷比的内稳性大于氮和磷的内稳性(
Yu et al ., 2011 ;
宁志英等, 2019 ).这说明, 植物对其自身内稳性的调节主要体现在N:P而不是单一的元素含量上.然而, 不同磷浓度处理下, 叶片磷含量、叶片N:P与营养液的磷浓度和营养液N:P并没有显著的幂函数关系, 其内稳性指数计算结果为负值.
Xing等(2015) 用1/
H 来衡量内稳性的强弱, 也测出内稳性指数为负值, 并将其归为严格内稳型.这说明磷处理条件下, 车桑子叶片磷含量和N:P具有极强的内稳性.较高的内稳性意味着养分元素变化小, 表明具有保守的养分利用策略, 也就更容易适应贫瘠环境而成为优势种(
Yu et al ., 2010 ).不同生长期植物内稳性不同, 内稳性随着植物生长发育逐渐增强(
Yu et al ., 2011 ).本研究中车桑子幼苗已表现出较高的内稳性, 因此, 这可能是促进车桑子成为干热河谷优势植物的原因之一.此外, 所有处理条件下, 车桑子的叶片N:P均在30左右波动.按照叶片临界N:P对限制元素的判断理论, 车桑子生长受到磷限制(
Güsewell, 2004 ;
Sullivan et al ., 2014 ), 这和氮磷添加对车桑子生长和生物量积累的结果是不一致的.与前人的研究观点(
Drenovsky & Richards, 2004 ;
Zhang et al ., 2004 )比较一致, 我们的结果也表明, 在物种水平上, 用叶片N:P来指示植物所受到的养分限制类型是不可靠的. ...
杉木人工林下原生与引入树种叶性状变异特征
2
2019
... 此外, 如上所述, 植物种内性状具有极大的变异性, 这被认为是植物性状在适应环境中的可塑性决定的.另一方面物种之间性状上也存在极大的差异, 并形成一个叶经济型谱.叶经济型谱理论认为植物叶片在其形态结构、化学组分和生理功能等性状之间存在着相互依存、相互关联的内在关系(
Wright et al ., 2004 ;
于鸿莹等, 2014 ).多种功能性状之间关系紧密, 不同性状的组合可体现出植物对不同环境的适应策略(
熊静等, 2019 ).那么, 在环境梯度如养分梯度下植物种内性状的可塑性及其关联性是否与种间的这种经济型谱变化趋势一致? 例如, 经济型谱中比叶面积与叶片氮含量显著正相关, 那么在氮浓度梯度中植物种内的比叶面积与叶片氮含量是否会产生类似的关系? 植物种内性状的可塑性调整在研究植物适应策略方面有着重要作用.目前,环境变化对叶性状和生物量分配的表型可塑性方面的研究较多, 然而, 在氮磷变化条件下, 植物在生物量分配和叶性状的调整上是否存在如同经济型谱理论的相互变化规律? 这方面的证据还比较有限.开展生物量分配和叶性状对氮磷浓度的响应研究, 有助于全面理解和揭示植物对养分有效性变化的响应与适应机制. ...
... 关系, 可见车桑子叶片磷含量的变化是驱动N:P变化的主要因子.
吴统贵等(2010) 和
宋彦涛等(2012) 的研究也表明, 叶片N:P主要受到磷的驱动, N:P的变化受控于叶片磷含量的变化.然而, 氮、磷处理条件下, 车桑子叶片氮和磷含量并没有显著正相关关系, 这可能是由于氮、磷养分不同的内稳性导致的.
黄端等(2017) 也发现叶片氮、磷含量之间无显著的正相关关系.此外, 本研究中, 磷处理下叶片氮含量和
SLA 显著正相关, 而氮处理下却没有这种相关性, 这与以往的研究结果不一致.以往大量的研究发现植物叶片
SLA 与叶片氮含量显著正相关, 二者的关联是植物叶经济型谱的重要组成部分(
于鸿莹等, 2014 ;
熊静等, 2019 ).以往的数据基本是从多个物种之间发现的, 也就是
SLA 与叶片氮含量正相关是种间变异的结果,而本研究是通过设置氮浓度梯度, 是基于环境中氮浓度变化对单个物种内
SLA 与叶片氮含量影响后的结果.在氮有效性增加的情况下, 叶片氮含量的响应更直接更迅速, 植物吸收氮后叶片氮含量显著增加, 而
SLA 的响应相对滞后, 与
Wright和Sutton-Grier (2012) 在氮添加条件下的结果是一致的, 这也反映了长期的氮限制使车桑子在较高的氮水平下短期内依然保持相对保守的氮利用策略.因此, 氮添加条件下车桑子叶片氮含量与
SLA 不相关甚至呈现出较弱的负相关关系.本研究结果表明, 环境中养分驱动下单个物种内叶片的性状变化可能并不完全符合经济型谱理论, 而会受到氮、磷养分有效性的影响, 符合叶经济型谱形成的环境因子主导假说(
赖小红等, 2019 ).此外, 植物功能性状和生物量分配之间存在着某种共轭的变化(
高景等, 2016 ).本研究中, 氮处理下, 车桑子根冠比与叶片氮含量、N:P显著负相关, 磷处理下却没有显著的相关性.以上结果支持我们假设(2)的观点, 车桑子生物量分配与叶性状间存在着一定的协同和权衡关系, 但是车桑子在面临氮、磷有效性的变化时具有不同的响应策略. ...
杉木人工林下原生与引入树种叶性状变异特征
2
2019
... 此外, 如上所述, 植物种内性状具有极大的变异性, 这被认为是植物性状在适应环境中的可塑性决定的.另一方面物种之间性状上也存在极大的差异, 并形成一个叶经济型谱.叶经济型谱理论认为植物叶片在其形态结构、化学组分和生理功能等性状之间存在着相互依存、相互关联的内在关系(
Wright et al ., 2004 ;
于鸿莹等, 2014 ).多种功能性状之间关系紧密, 不同性状的组合可体现出植物对不同环境的适应策略(
熊静等, 2019 ).那么, 在环境梯度如养分梯度下植物种内性状的可塑性及其关联性是否与种间的这种经济型谱变化趋势一致? 例如, 经济型谱中比叶面积与叶片氮含量显著正相关, 那么在氮浓度梯度中植物种内的比叶面积与叶片氮含量是否会产生类似的关系? 植物种内性状的可塑性调整在研究植物适应策略方面有着重要作用.目前,环境变化对叶性状和生物量分配的表型可塑性方面的研究较多, 然而, 在氮磷变化条件下, 植物在生物量分配和叶性状的调整上是否存在如同经济型谱理论的相互变化规律? 这方面的证据还比较有限.开展生物量分配和叶性状对氮磷浓度的响应研究, 有助于全面理解和揭示植物对养分有效性变化的响应与适应机制. ...
... 关系, 可见车桑子叶片磷含量的变化是驱动N:P变化的主要因子.
吴统贵等(2010) 和
宋彦涛等(2012) 的研究也表明, 叶片N:P主要受到磷的驱动, N:P的变化受控于叶片磷含量的变化.然而, 氮、磷处理条件下, 车桑子叶片氮和磷含量并没有显著正相关关系, 这可能是由于氮、磷养分不同的内稳性导致的.
黄端等(2017) 也发现叶片氮、磷含量之间无显著的正相关关系.此外, 本研究中, 磷处理下叶片氮含量和
SLA 显著正相关, 而氮处理下却没有这种相关性, 这与以往的研究结果不一致.以往大量的研究发现植物叶片
SLA 与叶片氮含量显著正相关, 二者的关联是植物叶经济型谱的重要组成部分(
于鸿莹等, 2014 ;
熊静等, 2019 ).以往的数据基本是从多个物种之间发现的, 也就是
SLA 与叶片氮含量正相关是种间变异的结果,而本研究是通过设置氮浓度梯度, 是基于环境中氮浓度变化对单个物种内
SLA 与叶片氮含量影响后的结果.在氮有效性增加的情况下, 叶片氮含量的响应更直接更迅速, 植物吸收氮后叶片氮含量显著增加, 而
SLA 的响应相对滞后, 与
Wright和Sutton-Grier (2012) 在氮添加条件下的结果是一致的, 这也反映了长期的氮限制使车桑子在较高的氮水平下短期内依然保持相对保守的氮利用策略.因此, 氮添加条件下车桑子叶片氮含量与
SLA 不相关甚至呈现出较弱的负相关关系.本研究结果表明, 环境中养分驱动下单个物种内叶片的性状变化可能并不完全符合经济型谱理论, 而会受到氮、磷养分有效性的影响, 符合叶经济型谱形成的环境因子主导假说(
赖小红等, 2019 ).此外, 植物功能性状和生物量分配之间存在着某种共轭的变化(
高景等, 2016 ).本研究中, 氮处理下, 车桑子根冠比与叶片氮含量、N:P显著负相关, 磷处理下却没有显著的相关性.以上结果支持我们假设(2)的观点, 车桑子生物量分配与叶性状间存在着一定的协同和权衡关系, 但是车桑子在面临氮、磷有效性的变化时具有不同的响应策略. ...
Plant functional diversity modulates global environmental change effects on grassland productivity
1
2018
... 氮和磷是构成生命体生物膜、蛋白质、核酸、磷脂和ATP等物质的重要成分, 因此是植物生命过程中必需的营养元素, 但植物生长普遍会受到氮磷养分的限制(
Yan et al ., 2015 ).氮、磷的限制格局和驱动因素各不相同.通常认为, 演替早期的生态系统主要受到氮的限制, 而演替后期的生态系统主要受到磷的限制(
Menge et al ., 2012 ).淋溶、挥发、反硝化作用和微生物对氮有限的固定作用等都可引起植物氮的缺乏(
Vitousek & Howarth, 1991 ).相比之下, 土壤物理屏蔽、低磷母岩、闭蓄态转化和人类活动(增加其他资源特别是氮的供给)等是引起植物生长过程中磷限制的主要原因(
Vitousek et al ., 2010 ).从植物个体的适应性出发, 充分利用植物自身对养分限制的适应性, 对提高植物生产力具有重要的意义.因此, 植物性状对氮、磷的响应特征对生态系统生产力和生态系统功能具有重要影响(
Wang et al ., 2018 ;
Xu et al ., 2018 ;
Zhang et al ., 2019 ). ...
Nitrogen limitation in arid-subhumid ecosystems: a meta-analysis of fertilization studies
1
2011
... 在植物养分限制的判断中, 若一种有效态元素的添加引起了植物生化过程速率或生物量的增加, 则意味着存在这种养分的限制(
Vitousek et al ., 2010 ;
Sullivan et al ., 2014 ).本研究表明, 高浓度氮水平显著促进了车桑子的生长和生物量积累, 但车桑子生长和生物量积累在低浓度和中等浓度氮水平下没有显著差异.野外条件下, 车桑子生长区的土壤有效氮(硝态氮与铵态氮之和)含量大约为11.7 mg·kg
-1 , 考虑到土壤中水分含量在10%左右, 也就是每kg土壤约含0.1 L水, 那么土壤中的有效氮含量约为117 mg·L
-1 , 介于本研究设置的N3 (210 mg·L
-1 )和N2 (70 mg·L
-1 )水平之间.因此, 本研究中, N1-N3处理下车桑子的生长没有显著差异, 可能是由于干热河谷长期的氮限制使得植物形成了对低氮环境的一系列形态与生理适应性.干旱生态系统氮限制尤为普遍(
Yahdjian et al ., 2011 ).已有研究表明, 氮是限制金沙江干旱河谷退化草地系统恢复的主要因素(
张彦东等, 2004 ).同时, 我们的结果表明, 车桑子主要通过增加对氮的吸收和利用来适应低氮环境.一方面, 车桑子在低氮条件下, 其根冠比显著增加可促进氮的吸收; 另一方面, 吸收的氮的利用效率又显著提高.根冠比的增加和养分利用效率的提高均是植物适应养分限制的主要机制(
冷华妮等, 2009 ;
Venterink & Güsewell, 2010 ).然而, 由于本研究中养分添加只持续了一个生长季, 试验时间较短, 鉴于车桑子幼苗长期对氮限制的适应, 对试验中的低浓度氮水平可能还不太敏感, 通过根冠比的调整和养分利用效率的提高即可保持其正常的生长和生物量积累, 但在长期的低浓度氮水平下车桑子生长和生物量积累可能会受到限制. ...
金沙江干热河谷地区植物叶片中各生源要素的化学计量特征以及异速增长关系
1
2012
... 车桑子(
Dodonaea viscosa )又名坡柳、明油子, 具有耐干旱、耐贫瘠和适应性强等特性, 是我国干热河谷区的优势物种, 也是该区植被恢复工程中的常见植物, 具有重要的生态价值.关于车桑子的研究, 目前重点研究了车桑子对干旱胁迫的响应特征(
李吉跃等, 2003 ;
陈少瑜等, 2006 ).但除了干旱胁迫之外, 养分胁迫成为干旱胁迫的次生效应(
da Silva et al ., 2011 ).加之该区土壤侵蚀严重, 干热河谷土壤(燥红土)普遍存在缺氮少磷的特点(
张燕平等, 2005 ), 严重影响了干热河谷植物的生长(
闫帮国等, 2012 ).然而, 关于车桑子性状对氮磷限制的响应目前尚不清楚.因此, 本研究以车桑子幼苗为研究对象, 采用砂培的方式进行试验, 研究车桑子幼苗生长特征、生物量分配、叶片功能性状及其相互关系对氮磷梯度的响应特征, 从生物量分配和叶片功能性状的角度探讨车桑子对低氮和低磷条件的适应性, 并验证以下假说: (1)车桑子幼苗生物量分配和叶片功能性状对氮变化和磷变化的响应具有差异性; (2)生物量分配和叶性状对氮磷浓度响应存在协同或权衡关系. ...
金沙江干热河谷地区植物叶片中各生源要素的化学计量特征以及异速增长关系
1
2012
... 车桑子(
Dodonaea viscosa )又名坡柳、明油子, 具有耐干旱、耐贫瘠和适应性强等特性, 是我国干热河谷区的优势物种, 也是该区植被恢复工程中的常见植物, 具有重要的生态价值.关于车桑子的研究, 目前重点研究了车桑子对干旱胁迫的响应特征(
李吉跃等, 2003 ;
陈少瑜等, 2006 ).但除了干旱胁迫之外, 养分胁迫成为干旱胁迫的次生效应(
da Silva et al ., 2011 ).加之该区土壤侵蚀严重, 干热河谷土壤(燥红土)普遍存在缺氮少磷的特点(
张燕平等, 2005 ), 严重影响了干热河谷植物的生长(
闫帮国等, 2012 ).然而, 关于车桑子性状对氮磷限制的响应目前尚不清楚.因此, 本研究以车桑子幼苗为研究对象, 采用砂培的方式进行试验, 研究车桑子幼苗生长特征、生物量分配、叶片功能性状及其相互关系对氮磷梯度的响应特征, 从生物量分配和叶片功能性状的角度探讨车桑子对低氮和低磷条件的适应性, 并验证以下假说: (1)车桑子幼苗生物量分配和叶片功能性状对氮变化和磷变化的响应具有差异性; (2)生物量分配和叶性状对氮磷浓度响应存在协同或权衡关系. ...
Plants adapted to nutrient limitation allocate less biomass into stems in an arid-hot grassland
1
2016
... 在养分有效性发生变化时, 植物会通过生物量分配和叶片功能性状的调整来适应养分变化.按照最优分配理论, 植物会将更多的生物量分配至能够获取限制资源的器官(
Mccarthy & Enquist, 2007 ).在养分有效性较低的情况下通常会通过增加根生物量分配和根冠比来获取更多的养分(
Poorter et al ., 2012 ).此外, 养分贫瘠的条件下, 植物会增加地上部分叶生物量的比例(相对于茎)(
Yan et al ., 2016 ), 但这方面的研究还相对较少.另一方面, 叶片功能性状的调整与资源获取和利用密切相关, 其形态结构和营养元素含量等均会随环境中氮磷条件的改变而发生适应性变化(
Qi et al ., 2009 ).比如, 较低的养分有效性通常会降低比叶面积, 增加叶干物质含量和养分利用效率, 而随着养分有效性的增加, 叶片养分含量显著增加(
Ordo?ez et al ., 2009 ;
林婉奇等, 2019 ).然而, 不同物种对氮磷添加的响应具有物种特异性(
Han et al ., 2013 ;
Mayor et al ., 2014 ).在氮限制条件下, 具有更高根长和根生物量比例的物种占据优势; 相反, 在磷限制条件下, 植物根长不能解释物种间的竞争优势, 菌根菌丝长度、根系分泌物、磷酸酶活性等相关性状起着重要作用(
Venterink & Güsewell, 2010 ;
Minden & Venterink, 2019 ), 这说明植物性状对氮和磷可能具有不同的响应特征, 包括生物量分配、叶片性状或养分利用效率等多个植物性状.由于不同的性状在适应氮限制和磷限制中表现出不同的变化趋势, 植物可能通过调整不同的性状来响应氮变化或磷变化, 但植物性状对氮、磷的响应是否表现出差异并不十分清楚. ...
Effects of nitrogen and phosphorus supply on growth rate, leaf stoichiometry, and nutrient resorption of
Arabidopsis thaliana 2
2015
... 氮和磷是构成生命体生物膜、蛋白质、核酸、磷脂和ATP等物质的重要成分, 因此是植物生命过程中必需的营养元素, 但植物生长普遍会受到氮磷养分的限制(
Yan et al ., 2015 ).氮、磷的限制格局和驱动因素各不相同.通常认为, 演替早期的生态系统主要受到氮的限制, 而演替后期的生态系统主要受到磷的限制(
Menge et al ., 2012 ).淋溶、挥发、反硝化作用和微生物对氮有限的固定作用等都可引起植物氮的缺乏(
Vitousek & Howarth, 1991 ).相比之下, 土壤物理屏蔽、低磷母岩、闭蓄态转化和人类活动(增加其他资源特别是氮的供给)等是引起植物生长过程中磷限制的主要原因(
Vitousek et al ., 2010 ).从植物个体的适应性出发, 充分利用植物自身对养分限制的适应性, 对提高植物生产力具有重要的意义.因此, 植物性状对氮、磷的响应特征对生态系统生产力和生态系统功能具有重要影响(
Wang et al ., 2018 ;
Xu et al ., 2018 ;
Zhang et al ., 2019 ). ...
... 本试验采用氮和磷的单因素设计, 分别包括4个氮浓度和4个磷浓度.由于盆栽实验中植物根系伸展空间有限, 按照野外土壤实际养分含量设置浓度梯度下植物很难获得充分生长.因此, 本试验参照前人的相关研究(
Yu et al ., 2011 ;
Yan et al ., 2015 ), 以Hoagland营养液(15 mmol·L
-1 N、1 mmol·L
-1 P)为基础进行氮磷养分添加.Hoagland营养液已经比较成熟, 溶液的pH较为稳定, 其养分配制考虑了植物需求, 养分浓度对大多数植物都很适合, 被广泛应用于水培和砂培实验中(
Hoagland & Arnon, 1938 ).由于河沙中没有养分, 不添加额外养分植物将无法生长, 因此, 本研究没有设置不添加的对照处理.结合浓度梯度的可控性, 4个氮浓度分别为3、5、15、30 mmol·L
-1 , 记为N1、N2、N3、N4, 其中, N4为高浓度氮水平, N3为中等浓度氮水平, N1和N2为低浓度氮水平, 通过添加NH
4 NO
3 进行调整; 4个磷浓度分别为0.25、0.5、1、2 mmol·L
-1 , 记为P1、P2、P3、P4, 其中, P4为高浓度磷水平, P3为中等浓度磷水平, P1和P2为低浓度磷水平, 通过添加KH
2 PO
4 进行调整.各氮、磷水平的设置及营养液的氮磷比如
表1 所示.其中, N3和P3处理一致, 因此, 试验共计7个处理, 每个处理5个重复.除氮、磷浓度不同外, 其余钾、钙、镁含量及微量元素含量均与全量Hoagland营养液一致, 配好后用H
2 SO
4 或NaOH调整溶液pH为6.由于钾、钙、镁的添加均为硫酸盐物质, 通过H
2 SO
4 调节pH对物质的平衡影响较小, 而钠离子为植物非必需元素, 对养分需求的平衡不会造成影响, 同时营养液中所添加的物质主要为中性物质, 不同营养液中pH差异较小, 所需H
2 SO
4 或NaOH的量是极小的, 因此, 这些影响基本可以忽略, 试验处理中不存在离子不平衡问题.由于砂培基质对营养液的保持能力较低, 试验处理期间, 每隔2天浇1次营养液, 每次营养液添加量为200 mL, 以保持养分浓度尽量维持在设定水平.同时, 将所有盆钵放在遮雨大棚内, 可避免降水对养分的淋失. ...
内蒙古荒漠草原植物叶片功能性状关系及其经济谱分析
2
2014
... 此外, 如上所述, 植物种内性状具有极大的变异性, 这被认为是植物性状在适应环境中的可塑性决定的.另一方面物种之间性状上也存在极大的差异, 并形成一个叶经济型谱.叶经济型谱理论认为植物叶片在其形态结构、化学组分和生理功能等性状之间存在着相互依存、相互关联的内在关系(
Wright et al ., 2004 ;
于鸿莹等, 2014 ).多种功能性状之间关系紧密, 不同性状的组合可体现出植物对不同环境的适应策略(
熊静等, 2019 ).那么, 在环境梯度如养分梯度下植物种内性状的可塑性及其关联性是否与种间的这种经济型谱变化趋势一致? 例如, 经济型谱中比叶面积与叶片氮含量显著正相关, 那么在氮浓度梯度中植物种内的比叶面积与叶片氮含量是否会产生类似的关系? 植物种内性状的可塑性调整在研究植物适应策略方面有着重要作用.目前,环境变化对叶性状和生物量分配的表型可塑性方面的研究较多, 然而, 在氮磷变化条件下, 植物在生物量分配和叶性状的调整上是否存在如同经济型谱理论的相互变化规律? 这方面的证据还比较有限.开展生物量分配和叶性状对氮磷浓度的响应研究, 有助于全面理解和揭示植物对养分有效性变化的响应与适应机制. ...
... 关系, 可见车桑子叶片磷含量的变化是驱动N:P变化的主要因子.
吴统贵等(2010) 和
宋彦涛等(2012) 的研究也表明, 叶片N:P主要受到磷的驱动, N:P的变化受控于叶片磷含量的变化.然而, 氮、磷处理条件下, 车桑子叶片氮和磷含量并没有显著正相关关系, 这可能是由于氮、磷养分不同的内稳性导致的.
黄端等(2017) 也发现叶片氮、磷含量之间无显著的正相关关系.此外, 本研究中, 磷处理下叶片氮含量和
SLA 显著正相关, 而氮处理下却没有这种相关性, 这与以往的研究结果不一致.以往大量的研究发现植物叶片
SLA 与叶片氮含量显著正相关, 二者的关联是植物叶经济型谱的重要组成部分(
于鸿莹等, 2014 ;
熊静等, 2019 ).以往的数据基本是从多个物种之间发现的, 也就是
SLA 与叶片氮含量正相关是种间变异的结果,而本研究是通过设置氮浓度梯度, 是基于环境中氮浓度变化对单个物种内
SLA 与叶片氮含量影响后的结果.在氮有效性增加的情况下, 叶片氮含量的响应更直接更迅速, 植物吸收氮后叶片氮含量显著增加, 而
SLA 的响应相对滞后, 与
Wright和Sutton-Grier (2012) 在氮添加条件下的结果是一致的, 这也反映了长期的氮限制使车桑子在较高的氮水平下短期内依然保持相对保守的氮利用策略.因此, 氮添加条件下车桑子叶片氮含量与
SLA 不相关甚至呈现出较弱的负相关关系.本研究结果表明, 环境中养分驱动下单个物种内叶片的性状变化可能并不完全符合经济型谱理论, 而会受到氮、磷养分有效性的影响, 符合叶经济型谱形成的环境因子主导假说(
赖小红等, 2019 ).此外, 植物功能性状和生物量分配之间存在着某种共轭的变化(
高景等, 2016 ).本研究中, 氮处理下, 车桑子根冠比与叶片氮含量、N:P显著负相关, 磷处理下却没有显著的相关性.以上结果支持我们假设(2)的观点, 车桑子生物量分配与叶性状间存在着一定的协同和权衡关系, 但是车桑子在面临氮、磷有效性的变化时具有不同的响应策略. ...
内蒙古荒漠草原植物叶片功能性状关系及其经济谱分析
2
2014
... 此外, 如上所述, 植物种内性状具有极大的变异性, 这被认为是植物性状在适应环境中的可塑性决定的.另一方面物种之间性状上也存在极大的差异, 并形成一个叶经济型谱.叶经济型谱理论认为植物叶片在其形态结构、化学组分和生理功能等性状之间存在着相互依存、相互关联的内在关系(
Wright et al ., 2004 ;
于鸿莹等, 2014 ).多种功能性状之间关系紧密, 不同性状的组合可体现出植物对不同环境的适应策略(
熊静等, 2019 ).那么, 在环境梯度如养分梯度下植物种内性状的可塑性及其关联性是否与种间的这种经济型谱变化趋势一致? 例如, 经济型谱中比叶面积与叶片氮含量显著正相关, 那么在氮浓度梯度中植物种内的比叶面积与叶片氮含量是否会产生类似的关系? 植物种内性状的可塑性调整在研究植物适应策略方面有着重要作用.目前,环境变化对叶性状和生物量分配的表型可塑性方面的研究较多, 然而, 在氮磷变化条件下, 植物在生物量分配和叶性状的调整上是否存在如同经济型谱理论的相互变化规律? 这方面的证据还比较有限.开展生物量分配和叶性状对氮磷浓度的响应研究, 有助于全面理解和揭示植物对养分有效性变化的响应与适应机制. ...
... 关系, 可见车桑子叶片磷含量的变化是驱动N:P变化的主要因子.
吴统贵等(2010) 和
宋彦涛等(2012) 的研究也表明, 叶片N:P主要受到磷的驱动, N:P的变化受控于叶片磷含量的变化.然而, 氮、磷处理条件下, 车桑子叶片氮和磷含量并没有显著正相关关系, 这可能是由于氮、磷养分不同的内稳性导致的.
黄端等(2017) 也发现叶片氮、磷含量之间无显著的正相关关系.此外, 本研究中, 磷处理下叶片氮含量和
SLA 显著正相关, 而氮处理下却没有这种相关性, 这与以往的研究结果不一致.以往大量的研究发现植物叶片
SLA 与叶片氮含量显著正相关, 二者的关联是植物叶经济型谱的重要组成部分(
于鸿莹等, 2014 ;
熊静等, 2019 ).以往的数据基本是从多个物种之间发现的, 也就是
SLA 与叶片氮含量正相关是种间变异的结果,而本研究是通过设置氮浓度梯度, 是基于环境中氮浓度变化对单个物种内
SLA 与叶片氮含量影响后的结果.在氮有效性增加的情况下, 叶片氮含量的响应更直接更迅速, 植物吸收氮后叶片氮含量显著增加, 而
SLA 的响应相对滞后, 与
Wright和Sutton-Grier (2012) 在氮添加条件下的结果是一致的, 这也反映了长期的氮限制使车桑子在较高的氮水平下短期内依然保持相对保守的氮利用策略.因此, 氮添加条件下车桑子叶片氮含量与
SLA 不相关甚至呈现出较弱的负相关关系.本研究结果表明, 环境中养分驱动下单个物种内叶片的性状变化可能并不完全符合经济型谱理论, 而会受到氮、磷养分有效性的影响, 符合叶经济型谱形成的环境因子主导假说(
赖小红等, 2019 ).此外, 植物功能性状和生物量分配之间存在着某种共轭的变化(
高景等, 2016 ).本研究中, 氮处理下, 车桑子根冠比与叶片氮含量、N:P显著负相关, 磷处理下却没有显著的相关性.以上结果支持我们假设(2)的观点, 车桑子生物量分配与叶性状间存在着一定的协同和权衡关系, 但是车桑子在面临氮、磷有效性的变化时具有不同的响应策略. ...
Linking stoichiometric homoeostasis with ecosystem structure, functioning and stability
1
2010
... 本研究中, 不同的氮浓度和磷浓度处理显著影响了车桑子叶片氮含量, 但叶片磷含量和叶片N:P并未受到显著影响.由于植物具有保持自身化学组成相对稳定的能力, 环境中的养分变化对植物氮磷元素的影响也取决于植物化学计量内稳性的强弱(
Persson et al ., 2010 ).植物内稳性特征可用内稳性指数
H 进行界定, 根据内稳性指数可将植物分为四类:
H > 4为稳态型; 2 <
H < 4为弱稳态型; 4/3 <
H < 2为弱敏感型;
H < 4/3为敏感型(
Persson et al ., 2010 ).本研究中, 不同氮浓度处理下, 叶片氮含量和叶片N:P的
H 分别为6.86和7.31, 均大于4, 呈稳态型, 并且叶片N:P的内稳性大于氮的内稳性.其他研究也表明, 氮磷比的内稳性大于氮和磷的内稳性(
Yu et al ., 2011 ;
宁志英等, 2019 ).这说明, 植物对其自身内稳性的调节主要体现在N:P而不是单一的元素含量上.然而, 不同磷浓度处理下, 叶片磷含量、叶片N:P与营养液的磷浓度和营养液N:P并没有显著的幂函数关系, 其内稳性指数计算结果为负值.
Xing等(2015) 用1/
H 来衡量内稳性的强弱, 也测出内稳性指数为负值, 并将其归为严格内稳型.这说明磷处理条件下, 车桑子叶片磷含量和N:P具有极强的内稳性.较高的内稳性意味着养分元素变化小, 表明具有保守的养分利用策略, 也就更容易适应贫瘠环境而成为优势种(
Yu et al ., 2010 ).不同生长期植物内稳性不同, 内稳性随着植物生长发育逐渐增强(
Yu et al ., 2011 ).本研究中车桑子幼苗已表现出较高的内稳性, 因此, 这可能是促进车桑子成为干热河谷优势植物的原因之一.此外, 所有处理条件下, 车桑子的叶片N:P均在30左右波动.按照叶片临界N:P对限制元素的判断理论, 车桑子生长受到磷限制(
Güsewell, 2004 ;
Sullivan et al ., 2014 ), 这和氮磷添加对车桑子生长和生物量积累的结果是不一致的.与前人的研究观点(
Drenovsky & Richards, 2004 ;
Zhang et al ., 2004 )比较一致, 我们的结果也表明, 在物种水平上, 用叶片N:P来指示植物所受到的养分限制类型是不可靠的. ...
Stoichiometric homeostasis of vascular plants in the Inner Mongolia grassland
5
2011
... 本试验采用氮和磷的单因素设计, 分别包括4个氮浓度和4个磷浓度.由于盆栽实验中植物根系伸展空间有限, 按照野外土壤实际养分含量设置浓度梯度下植物很难获得充分生长.因此, 本试验参照前人的相关研究(
Yu et al ., 2011 ;
Yan et al ., 2015 ), 以Hoagland营养液(15 mmol·L
-1 N、1 mmol·L
-1 P)为基础进行氮磷养分添加.Hoagland营养液已经比较成熟, 溶液的pH较为稳定, 其养分配制考虑了植物需求, 养分浓度对大多数植物都很适合, 被广泛应用于水培和砂培实验中(
Hoagland & Arnon, 1938 ).由于河沙中没有养分, 不添加额外养分植物将无法生长, 因此, 本研究没有设置不添加的对照处理.结合浓度梯度的可控性, 4个氮浓度分别为3、5、15、30 mmol·L
-1 , 记为N1、N2、N3、N4, 其中, N4为高浓度氮水平, N3为中等浓度氮水平, N1和N2为低浓度氮水平, 通过添加NH
4 NO
3 进行调整; 4个磷浓度分别为0.25、0.5、1、2 mmol·L
-1 , 记为P1、P2、P3、P4, 其中, P4为高浓度磷水平, P3为中等浓度磷水平, P1和P2为低浓度磷水平, 通过添加KH
2 PO
4 进行调整.各氮、磷水平的设置及营养液的氮磷比如
表1 所示.其中, N3和P3处理一致, 因此, 试验共计7个处理, 每个处理5个重复.除氮、磷浓度不同外, 其余钾、钙、镁含量及微量元素含量均与全量Hoagland营养液一致, 配好后用H
2 SO
4 或NaOH调整溶液pH为6.由于钾、钙、镁的添加均为硫酸盐物质, 通过H
2 SO
4 调节pH对物质的平衡影响较小, 而钠离子为植物非必需元素, 对养分需求的平衡不会造成影响, 同时营养液中所添加的物质主要为中性物质, 不同营养液中pH差异较小, 所需H
2 SO
4 或NaOH的量是极小的, 因此, 这些影响基本可以忽略, 试验处理中不存在离子不平衡问题.由于砂培基质对营养液的保持能力较低, 试验处理期间, 每隔2天浇1次营养液, 每次营养液添加量为200 mL, 以保持养分浓度尽量维持在设定水平.同时, 将所有盆钵放在遮雨大棚内, 可避免降水对养分的淋失. ...
... 除营养液添加当天外, 其余时间各盆钵每天浇等量的自来水, 以保证植株不受干旱胁迫的影响.此外, 由于试验前不清楚车桑子对氮磷养分的吸收情况, 添加营养液5次后用2 L自来水淋洗1次花盆, 以避免盐基离子积累对车桑子幼苗的毒害作用(
Yu et al ., 2011 ).试验处理时间为一个生长季, 在雨季结束时(11月)停止试验. ...
... 式中,
Y 是车桑子幼苗叶片氮、磷含量及N:P,
X 是营养液中对应的氮、磷浓度及对应的N:P,
c 为常数,
H 为植物的内稳性指数(
Yu et al ., 2011 ). ...
... 本研究中, 不同的氮浓度和磷浓度处理显著影响了车桑子叶片氮含量, 但叶片磷含量和叶片N:P并未受到显著影响.由于植物具有保持自身化学组成相对稳定的能力, 环境中的养分变化对植物氮磷元素的影响也取决于植物化学计量内稳性的强弱(
Persson et al ., 2010 ).植物内稳性特征可用内稳性指数
H 进行界定, 根据内稳性指数可将植物分为四类:
H > 4为稳态型; 2 <
H < 4为弱稳态型; 4/3 <
H < 2为弱敏感型;
H < 4/3为敏感型(
Persson et al ., 2010 ).本研究中, 不同氮浓度处理下, 叶片氮含量和叶片N:P的
H 分别为6.86和7.31, 均大于4, 呈稳态型, 并且叶片N:P的内稳性大于氮的内稳性.其他研究也表明, 氮磷比的内稳性大于氮和磷的内稳性(
Yu et al ., 2011 ;
宁志英等, 2019 ).这说明, 植物对其自身内稳性的调节主要体现在N:P而不是单一的元素含量上.然而, 不同磷浓度处理下, 叶片磷含量、叶片N:P与营养液的磷浓度和营养液N:P并没有显著的幂函数关系, 其内稳性指数计算结果为负值.
Xing等(2015) 用1/
H 来衡量内稳性的强弱, 也测出内稳性指数为负值, 并将其归为严格内稳型.这说明磷处理条件下, 车桑子叶片磷含量和N:P具有极强的内稳性.较高的内稳性意味着养分元素变化小, 表明具有保守的养分利用策略, 也就更容易适应贫瘠环境而成为优势种(
Yu et al ., 2010 ).不同生长期植物内稳性不同, 内稳性随着植物生长发育逐渐增强(
Yu et al ., 2011 ).本研究中车桑子幼苗已表现出较高的内稳性, 因此, 这可能是促进车桑子成为干热河谷优势植物的原因之一.此外, 所有处理条件下, 车桑子的叶片N:P均在30左右波动.按照叶片临界N:P对限制元素的判断理论, 车桑子生长受到磷限制(
Güsewell, 2004 ;
Sullivan et al ., 2014 ), 这和氮磷添加对车桑子生长和生物量积累的结果是不一致的.与前人的研究观点(
Drenovsky & Richards, 2004 ;
Zhang et al ., 2004 )比较一致, 我们的结果也表明, 在物种水平上, 用叶片N:P来指示植物所受到的养分限制类型是不可靠的. ...
... ).不同生长期植物内稳性不同, 内稳性随着植物生长发育逐渐增强(
Yu et al ., 2011 ).本研究中车桑子幼苗已表现出较高的内稳性, 因此, 这可能是促进车桑子成为干热河谷优势植物的原因之一.此外, 所有处理条件下, 车桑子的叶片N:P均在30左右波动.按照叶片临界N:P对限制元素的判断理论, 车桑子生长受到磷限制(
Güsewell, 2004 ;
Sullivan et al ., 2014 ), 这和氮磷添加对车桑子生长和生物量积累的结果是不一致的.与前人的研究观点(
Drenovsky & Richards, 2004 ;
Zhang et al ., 2004 )比较一致, 我们的结果也表明, 在物种水平上, 用叶片N:P来指示植物所受到的养分限制类型是不可靠的. ...
Trait identity and functional diversity co-drive response of ecosystem productivity to nitrogen enrichment
1
2019
... 氮和磷是构成生命体生物膜、蛋白质、核酸、磷脂和ATP等物质的重要成分, 因此是植物生命过程中必需的营养元素, 但植物生长普遍会受到氮磷养分的限制(
Yan et al ., 2015 ).氮、磷的限制格局和驱动因素各不相同.通常认为, 演替早期的生态系统主要受到氮的限制, 而演替后期的生态系统主要受到磷的限制(
Menge et al ., 2012 ).淋溶、挥发、反硝化作用和微生物对氮有限的固定作用等都可引起植物氮的缺乏(
Vitousek & Howarth, 1991 ).相比之下, 土壤物理屏蔽、低磷母岩、闭蓄态转化和人类活动(增加其他资源特别是氮的供给)等是引起植物生长过程中磷限制的主要原因(
Vitousek et al ., 2010 ).从植物个体的适应性出发, 充分利用植物自身对养分限制的适应性, 对提高植物生产力具有重要的意义.因此, 植物性状对氮、磷的响应特征对生态系统生产力和生态系统功能具有重要影响(
Wang et al ., 2018 ;
Xu et al ., 2018 ;
Zhang et al ., 2019 ). ...
Differential responses of N:P stoichiometry of
Leymus chinensis and
Carex korshinskyi to N additions in a steppe ecosystem in Nei Mongol
1
2004
... 本研究中, 不同的氮浓度和磷浓度处理显著影响了车桑子叶片氮含量, 但叶片磷含量和叶片N:P并未受到显著影响.由于植物具有保持自身化学组成相对稳定的能力, 环境中的养分变化对植物氮磷元素的影响也取决于植物化学计量内稳性的强弱(
Persson et al ., 2010 ).植物内稳性特征可用内稳性指数
H 进行界定, 根据内稳性指数可将植物分为四类:
H > 4为稳态型; 2 <
H < 4为弱稳态型; 4/3 <
H < 2为弱敏感型;
H < 4/3为敏感型(
Persson et al ., 2010 ).本研究中, 不同氮浓度处理下, 叶片氮含量和叶片N:P的
H 分别为6.86和7.31, 均大于4, 呈稳态型, 并且叶片N:P的内稳性大于氮的内稳性.其他研究也表明, 氮磷比的内稳性大于氮和磷的内稳性(
Yu et al ., 2011 ;
宁志英等, 2019 ).这说明, 植物对其自身内稳性的调节主要体现在N:P而不是单一的元素含量上.然而, 不同磷浓度处理下, 叶片磷含量、叶片N:P与营养液的磷浓度和营养液N:P并没有显著的幂函数关系, 其内稳性指数计算结果为负值.
Xing等(2015) 用1/
H 来衡量内稳性的强弱, 也测出内稳性指数为负值, 并将其归为严格内稳型.这说明磷处理条件下, 车桑子叶片磷含量和N:P具有极强的内稳性.较高的内稳性意味着养分元素变化小, 表明具有保守的养分利用策略, 也就更容易适应贫瘠环境而成为优势种(
Yu et al ., 2010 ).不同生长期植物内稳性不同, 内稳性随着植物生长发育逐渐增强(
Yu et al ., 2011 ).本研究中车桑子幼苗已表现出较高的内稳性, 因此, 这可能是促进车桑子成为干热河谷优势植物的原因之一.此外, 所有处理条件下, 车桑子的叶片N:P均在30左右波动.按照叶片临界N:P对限制元素的判断理论, 车桑子生长受到磷限制(
Güsewell, 2004 ;
Sullivan et al ., 2014 ), 这和氮磷添加对车桑子生长和生物量积累的结果是不一致的.与前人的研究观点(
Drenovsky & Richards, 2004 ;
Zhang et al ., 2004 )比较一致, 我们的结果也表明, 在物种水平上, 用叶片N:P来指示植物所受到的养分限制类型是不可靠的. ...
云南干热河谷区裸地植被恢复过程中车桑子的种群特征
1
2016
... 幼苗是植物生活史过程中比较脆弱的一个阶段, 也是对环境变化最为敏感的一个阶段, 车桑子幼苗的死亡率较高, 在车桑子种植过程中应加强苗期管护(
张琴等, 2016 ).幼苗生长对植物群落的补员起着关键作用(
李秋艳和赵文智, 2006 ).因此, 本文基于车桑子幼苗的研究对车桑子在植被恢复中的应用具有一定的参考价值. ...
云南干热河谷区裸地植被恢复过程中车桑子的种群特征
1
2016
... 幼苗是植物生活史过程中比较脆弱的一个阶段, 也是对环境变化最为敏感的一个阶段, 车桑子幼苗的死亡率较高, 在车桑子种植过程中应加强苗期管护(
张琴等, 2016 ).幼苗生长对植物群落的补员起着关键作用(
李秋艳和赵文智, 2006 ).因此, 本文基于车桑子幼苗的研究对车桑子在植被恢复中的应用具有一定的参考价值. ...
金沙江干旱河谷退化草地群落对氮磷施肥的反应
1
2004
... 在植物养分限制的判断中, 若一种有效态元素的添加引起了植物生化过程速率或生物量的增加, 则意味着存在这种养分的限制(
Vitousek et al ., 2010 ;
Sullivan et al ., 2014 ).本研究表明, 高浓度氮水平显著促进了车桑子的生长和生物量积累, 但车桑子生长和生物量积累在低浓度和中等浓度氮水平下没有显著差异.野外条件下, 车桑子生长区的土壤有效氮(硝态氮与铵态氮之和)含量大约为11.7 mg·kg
-1 , 考虑到土壤中水分含量在10%左右, 也就是每kg土壤约含0.1 L水, 那么土壤中的有效氮含量约为117 mg·L
-1 , 介于本研究设置的N3 (210 mg·L
-1 )和N2 (70 mg·L
-1 )水平之间.因此, 本研究中, N1-N3处理下车桑子的生长没有显著差异, 可能是由于干热河谷长期的氮限制使得植物形成了对低氮环境的一系列形态与生理适应性.干旱生态系统氮限制尤为普遍(
Yahdjian et al ., 2011 ).已有研究表明, 氮是限制金沙江干旱河谷退化草地系统恢复的主要因素(
张彦东等, 2004 ).同时, 我们的结果表明, 车桑子主要通过增加对氮的吸收和利用来适应低氮环境.一方面, 车桑子在低氮条件下, 其根冠比显著增加可促进氮的吸收; 另一方面, 吸收的氮的利用效率又显著提高.根冠比的增加和养分利用效率的提高均是植物适应养分限制的主要机制(
冷华妮等, 2009 ;
Venterink & Güsewell, 2010 ).然而, 由于本研究中养分添加只持续了一个生长季, 试验时间较短, 鉴于车桑子幼苗长期对氮限制的适应, 对试验中的低浓度氮水平可能还不太敏感, 通过根冠比的调整和养分利用效率的提高即可保持其正常的生长和生物量积累, 但在长期的低浓度氮水平下车桑子生长和生物量积累可能会受到限制. ...
金沙江干旱河谷退化草地群落对氮磷施肥的反应
1
2004
... 在植物养分限制的判断中, 若一种有效态元素的添加引起了植物生化过程速率或生物量的增加, 则意味着存在这种养分的限制(
Vitousek et al ., 2010 ;
Sullivan et al ., 2014 ).本研究表明, 高浓度氮水平显著促进了车桑子的生长和生物量积累, 但车桑子生长和生物量积累在低浓度和中等浓度氮水平下没有显著差异.野外条件下, 车桑子生长区的土壤有效氮(硝态氮与铵态氮之和)含量大约为11.7 mg·kg
-1 , 考虑到土壤中水分含量在10%左右, 也就是每kg土壤约含0.1 L水, 那么土壤中的有效氮含量约为117 mg·L
-1 , 介于本研究设置的N3 (210 mg·L
-1 )和N2 (70 mg·L
-1 )水平之间.因此, 本研究中, N1-N3处理下车桑子的生长没有显著差异, 可能是由于干热河谷长期的氮限制使得植物形成了对低氮环境的一系列形态与生理适应性.干旱生态系统氮限制尤为普遍(
Yahdjian et al ., 2011 ).已有研究表明, 氮是限制金沙江干旱河谷退化草地系统恢复的主要因素(
张彦东等, 2004 ).同时, 我们的结果表明, 车桑子主要通过增加对氮的吸收和利用来适应低氮环境.一方面, 车桑子在低氮条件下, 其根冠比显著增加可促进氮的吸收; 另一方面, 吸收的氮的利用效率又显著提高.根冠比的增加和养分利用效率的提高均是植物适应养分限制的主要机制(
冷华妮等, 2009 ;
Venterink & Güsewell, 2010 ).然而, 由于本研究中养分添加只持续了一个生长季, 试验时间较短, 鉴于车桑子幼苗长期对氮限制的适应, 对试验中的低浓度氮水平可能还不太敏感, 通过根冠比的调整和养分利用效率的提高即可保持其正常的生长和生物量积累, 但在长期的低浓度氮水平下车桑子生长和生物量积累可能会受到限制. ...
干热河谷印楝生长与立地条件关系
1
2005
... 车桑子(
Dodonaea viscosa )又名坡柳、明油子, 具有耐干旱、耐贫瘠和适应性强等特性, 是我国干热河谷区的优势物种, 也是该区植被恢复工程中的常见植物, 具有重要的生态价值.关于车桑子的研究, 目前重点研究了车桑子对干旱胁迫的响应特征(
李吉跃等, 2003 ;
陈少瑜等, 2006 ).但除了干旱胁迫之外, 养分胁迫成为干旱胁迫的次生效应(
da Silva et al ., 2011 ).加之该区土壤侵蚀严重, 干热河谷土壤(燥红土)普遍存在缺氮少磷的特点(
张燕平等, 2005 ), 严重影响了干热河谷植物的生长(
闫帮国等, 2012 ).然而, 关于车桑子性状对氮磷限制的响应目前尚不清楚.因此, 本研究以车桑子幼苗为研究对象, 采用砂培的方式进行试验, 研究车桑子幼苗生长特征、生物量分配、叶片功能性状及其相互关系对氮磷梯度的响应特征, 从生物量分配和叶片功能性状的角度探讨车桑子对低氮和低磷条件的适应性, 并验证以下假说: (1)车桑子幼苗生物量分配和叶片功能性状对氮变化和磷变化的响应具有差异性; (2)生物量分配和叶性状对氮磷浓度响应存在协同或权衡关系. ...
干热河谷印楝生长与立地条件关系
1
2005
... 车桑子(
Dodonaea viscosa )又名坡柳、明油子, 具有耐干旱、耐贫瘠和适应性强等特性, 是我国干热河谷区的优势物种, 也是该区植被恢复工程中的常见植物, 具有重要的生态价值.关于车桑子的研究, 目前重点研究了车桑子对干旱胁迫的响应特征(
李吉跃等, 2003 ;
陈少瑜等, 2006 ).但除了干旱胁迫之外, 养分胁迫成为干旱胁迫的次生效应(
da Silva et al ., 2011 ).加之该区土壤侵蚀严重, 干热河谷土壤(燥红土)普遍存在缺氮少磷的特点(
张燕平等, 2005 ), 严重影响了干热河谷植物的生长(
闫帮国等, 2012 ).然而, 关于车桑子性状对氮磷限制的响应目前尚不清楚.因此, 本研究以车桑子幼苗为研究对象, 采用砂培的方式进行试验, 研究车桑子幼苗生长特征、生物量分配、叶片功能性状及其相互关系对氮磷梯度的响应特征, 从生物量分配和叶片功能性状的角度探讨车桑子对低氮和低磷条件的适应性, 并验证以下假说: (1)车桑子幼苗生物量分配和叶片功能性状对氮变化和磷变化的响应具有差异性; (2)生物量分配和叶性状对氮磷浓度响应存在协同或权衡关系. ...
生境异质性对鼎湖山常绿阔叶林群落功能多样性的影响
1
2016
...
SLA 和
LDMC 是叶片功能性状中最主要的两个指标, 被认为是指示植物养分利用策略的关键性指标(
Jager et al ., 2015 ).与我们的假设(1)一致, 叶片功能性状对氮变化和磷变化的响应具有差异性.随磷浓度的增加,
SLA 显著降低,
LDMC 显著增加, 而氮处理下却没有相似的结果.前人研究表明,
SLA 随养分的减少而降低, 较低的
SLA 和较高的
LDMC 有利于植物适应资源贫瘠的环境(
Jager et al ., 2015 ).然而, 我们的研究中, 高浓度磷水平降低了
SLA, 增加了
LDMC; 相反, 低浓度磷水平增加了
SLA, 降低了
LDMC .高磷水平下低
SLA 、高
LDMC 表明车桑子采取养分的有效保存策略, 低磷水平下高
SLA 、低
LDMC 可提高对光资源的获取能力, 表明车桑子采取快速生长的策略, 该研究结果与
周光霞等(2016) 的研究相似.然而, 低磷水平下车桑子生长依然受到严重的限制.
林婉奇等(2019) 也发现磷添加处理减少了
SLA, 但显著促进了幼苗生长.叶面积和
SLA 呈负相关关系, 叶面积增加可提高植物的光获取能力, 弥补因
SLA 减小而导致的碳摄取效率的损失(
陈延松等, 2019 ).因此, 高磷水平下叶面积的增加对促进车桑子生长和生物量积累具有重要作用. ...
生境异质性对鼎湖山常绿阔叶林群落功能多样性的影响
1
2016
...
SLA 和
LDMC 是叶片功能性状中最主要的两个指标, 被认为是指示植物养分利用策略的关键性指标(
Jager et al ., 2015 ).与我们的假设(1)一致, 叶片功能性状对氮变化和磷变化的响应具有差异性.随磷浓度的增加,
SLA 显著降低,
LDMC 显著增加, 而氮处理下却没有相似的结果.前人研究表明,
SLA 随养分的减少而降低, 较低的
SLA 和较高的
LDMC 有利于植物适应资源贫瘠的环境(
Jager et al ., 2015 ).然而, 我们的研究中, 高浓度磷水平降低了
SLA, 增加了
LDMC; 相反, 低浓度磷水平增加了
SLA, 降低了
LDMC .高磷水平下低
SLA 、高
LDMC 表明车桑子采取养分的有效保存策略, 低磷水平下高
SLA 、低
LDMC 可提高对光资源的获取能力, 表明车桑子采取快速生长的策略, 该研究结果与
周光霞等(2016) 的研究相似.然而, 低磷水平下车桑子生长依然受到严重的限制.
林婉奇等(2019) 也发现磷添加处理减少了
SLA, 但显著促进了幼苗生长.叶面积和
SLA 呈负相关关系, 叶面积增加可提高植物的光获取能力, 弥补因
SLA 减小而导致的碳摄取效率的损失(
陈延松等, 2019 ).因此, 高磷水平下叶面积的增加对促进车桑子生长和生物量积累具有重要作用. ...
紫楠幼苗生长、叶性状和生物量分配对林窗不同生境的响应
1
2015
... 从车桑子生长特征与生物量分配、叶性状的相关性来看, 氮处理条件下, 株高、基径、生物量等生长性状均与根冠比呈显著负相关关系, 与叶片氮含量显著正相关, 但生长性状与叶片氮含量的相关性更强, 表明叶片氮含量的调整对车桑子适应低氮具有重要意义; 而磷处理下, 株高、基径、生物量与
SLA 、叶片氮含量显著负相关, 与
LDMC 显著正相关, 表明叶片功能性状的调整在车桑子适应低磷环境中发挥重要作用.
周俊宏等(2015) 针对紫楠(
Phoebe sheareri )幼苗的研究表明, 相比叶性状的调整, 紫楠幼苗主要通过生物量分配比例的改变来适应不同生境的变化.我们的研究与之相反, 无论氮处理还是磷处理条件下, 相比生物量分配, 叶片性状的调整是车桑子对养分有效性变化更为主要的适应策略.生物量分配的变化也是一系列植物性状对环境变化适应的结果(
Mccarthy & Enquist, 2007 ).因此, 叶性状的调整是车桑子对养分变化更为主要的适应策略.我们的结果表明, 车桑子在响应氮、磷限制时会采用不同的适应策略, 表征其适应策略的性状及性状间的权衡策略亦有所差异.然而, 本研究是基于短期试验条件下的单因素控制试验, 由于氮、磷之间存在交互作用, 不同组合的氮磷添加对车桑子幼苗生长性状的影响值得进一步研究. ...
紫楠幼苗生长、叶性状和生物量分配对林窗不同生境的响应
1
2015
... 从车桑子生长特征与生物量分配、叶性状的相关性来看, 氮处理条件下, 株高、基径、生物量等生长性状均与根冠比呈显著负相关关系, 与叶片氮含量显著正相关, 但生长性状与叶片氮含量的相关性更强, 表明叶片氮含量的调整对车桑子适应低氮具有重要意义; 而磷处理下, 株高、基径、生物量与
SLA 、叶片氮含量显著负相关, 与
LDMC 显著正相关, 表明叶片功能性状的调整在车桑子适应低磷环境中发挥重要作用.
周俊宏等(2015) 针对紫楠(
Phoebe sheareri )幼苗的研究表明, 相比叶性状的调整, 紫楠幼苗主要通过生物量分配比例的改变来适应不同生境的变化.我们的研究与之相反, 无论氮处理还是磷处理条件下, 相比生物量分配, 叶片性状的调整是车桑子对养分有效性变化更为主要的适应策略.生物量分配的变化也是一系列植物性状对环境变化适应的结果(
Mccarthy & Enquist, 2007 ).因此, 叶性状的调整是车桑子对养分变化更为主要的适应策略.我们的结果表明, 车桑子在响应氮、磷限制时会采用不同的适应策略, 表征其适应策略的性状及性状间的权衡策略亦有所差异.然而, 本研究是基于短期试验条件下的单因素控制试验, 由于氮、磷之间存在交互作用, 不同组合的氮磷添加对车桑子幼苗生长性状的影响值得进一步研究. ...
 ,2,*
,2,*
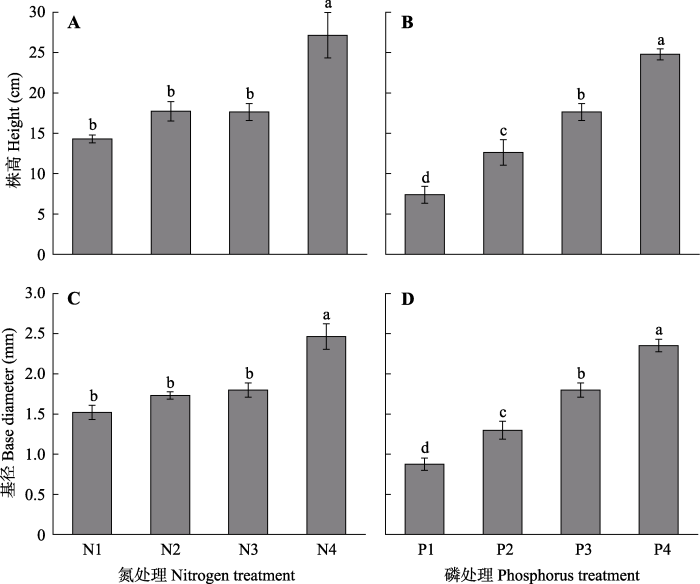 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT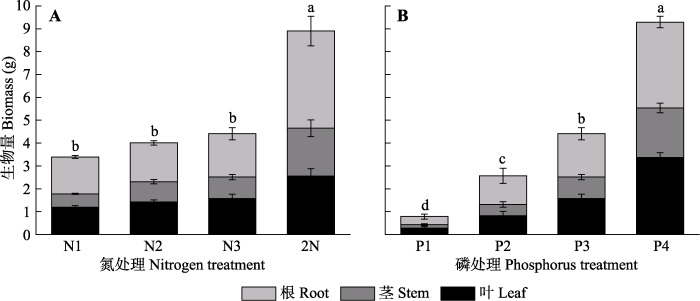 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT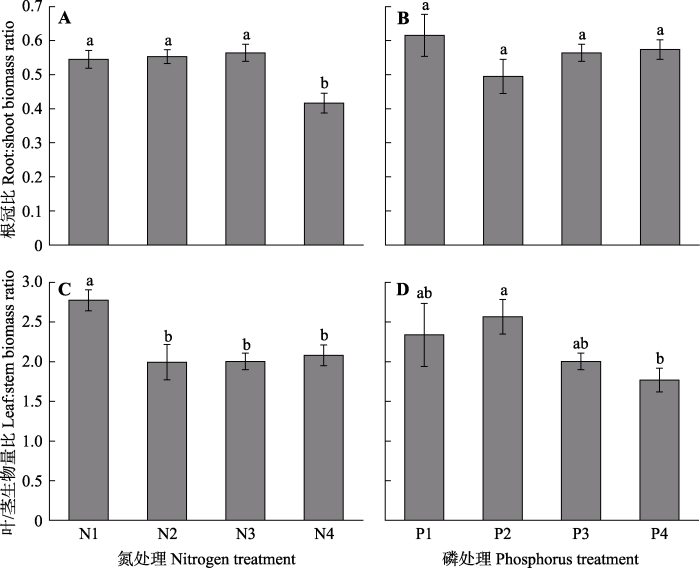 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT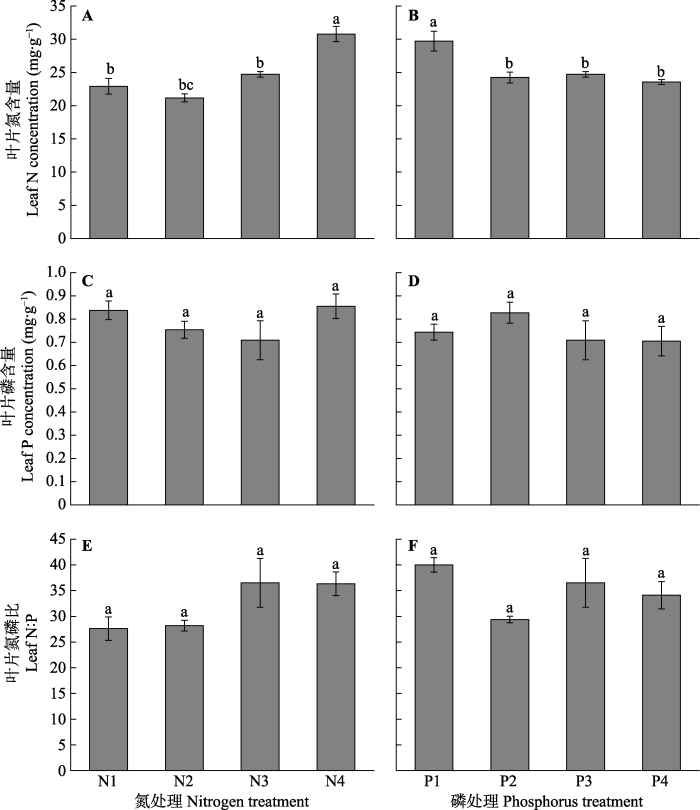 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT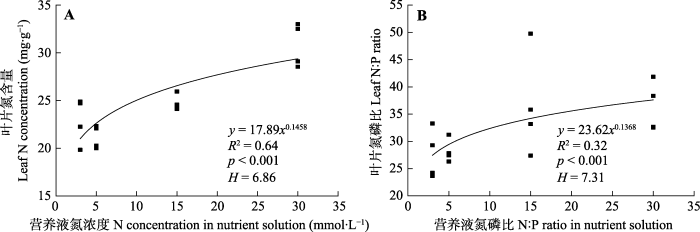 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT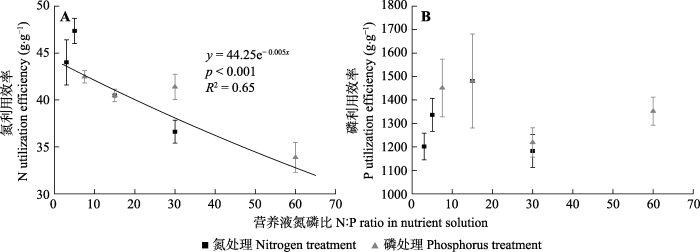 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}