陈国鹏
, , 1 , * , 杨克彤
1 , 王立
1 , 王飞
2 , 曹秀文
2 , 陈林生
3 1 甘肃农业大学林学院, 兰州 7300702 甘肃省白龙江林业管理局林业科学研究所, 兰州 7300703 甘肃省白龙江林业管理局生态监测和林业调查规划院, 兰州 730070Allometric relations for biomass partitioning of seven alpine Rhododendron species in south of Gansu Guo-Peng CHEN
, , 1 , * , Ke-Tong YANG
1 , Li WANG
1 , Fei WANG
2 , Xiu-Wen CAO
2 , Lin-Sheng CHEN
3 1 College of Forestry, Gansu Agricultural University, Lanzhou 730070, China2 Institute of Forestry, Bailongjiang Forestry Administration, Lanzhou 730070, China3 Institute of Ecological Monitoring and Forestry Survey and Planning, Bailongjiang Forestry Administration, Lanzhou 730070, China通讯作者: *陈国鹏:ORCID:0000-0002-3854-8755,
chgp1986@gmail.com 编委: 黄建国(特邀)
责任编辑: 李敏
收稿日期: 2020-04-24
接受日期: 2020-08-22
网络出版日期: 2020-10-20
基金资助: 国家自然科学基金 (31800352 )甘肃省高等学校科研项目 (2017A-032 )
Corresponding authors: *ORCID:0000-0002-3854-8755,
chgp1986@gmail.com Received: 2020-04-24
Accepted: 2020-08-22
Online: 2020-10-20
Fund supported: National Natural Science Foundation of China (31800352 )Institutions of Higher Learning Scientific Research Project in Gansu Province (2017A-032 )
摘要 生物量分配模式影响着植物个体生长和繁殖到整个群落的质量和能量流动等所有层次的功能, 揭示高寒灌丛的生物量分配模式不仅可以掌握植物的生活史策略, 而且对理解灌丛碳汇不确定性具有重要意义。该研究以甘肃南部高山-亚高山区的常绿灌丛——杜鹃(
Rhododendron spp.)灌丛的7个典型种为对象, 采用全株收获法研究了不同物种个体水平上各器官生物量的分配比例和异速生长关系。结果表明: 7种高寒杜鹃根、茎、叶生物量的分配平均比例为35.57%、45.61%和18.83%, 各器官生物量分配比例的物种差异显著; 7种高寒杜鹃的叶与茎、叶与根、茎与根以及地上生物量与地下生物量之间既有异速生长关系, 也有等速生长关系, 异速生长指数不完全支持生态代谢理论和小个体等速生长理论的参考值; 各器官异速生长关系的物种差异显著。结合最优分配理论和异速生长理论能更好地解释陇南山地7种高寒杜鹃生物量的变异及适应机制。
关键词: 高寒灌丛 ;
生物量 ;
最优分配 ;
异速生长 ;
杜鹃 Abstract Aims Biomass-partitioning patterns influences the growth and reproduction of individual plant, and the mass and energy flow of a plant community. Revealing the biomass-partitioning patterns of alpine shrubs can help understand their life-cycle strategies and the uncertainty of shrub carbon sink.
Methods In this study, the biomass-partitioning ratio and allometric relations of each organ at individual level were analyzed in seven typical evergreen shrub species in
Rhododendron in alpine-subalpine of southern Gansu using whole-plant harvesting method.
Important findings The results showed that the average fractions of biomass allocation in root, stem and leave for seven
Rhododendron species were 35.57%, 45.61% and 18.83%, respectively, with significant differences in fractions of biomass allocation of each among species. There were allometric relations and isometric relations between leaves vs stems, leaves vs roots, stems vs roots, and aboveground biomass vs underground biomass for all species. However, allometric scale did not fully support the reference values of metabolic scaling theory and isometric scaling theory of small plants. There were significant allometric relationships in leaf mass, stem mass and root mass among species. The combination of the optimal partitioning theory and allelotropic theory help better understand the biomass variation and adaptation mechanism of
Rhododendron species in mountainous areas in southern Gansu.
Keywords: alpine shrub ;
biomass ;
optimal partitioning ;
allometric growth ;
Rhododendron PDF (1496KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 陈国鹏, 杨克彤, 王立, 王飞, 曹秀文, 陈林生. 甘肃南部7种高寒杜鹃生物量分配的异速生长关系. 植物生态学报, 2020, 44(10): 1040-1049. DOI:
10.17521/cjpe.2020.0119 CHEN Guo-Peng, YANG Ke-Tong, WANG Li, WANG Fei, CAO Xiu-Wen, CHEN Lin-Sheng.
Allometric relations for biomass partitioning of seven alpine Rhododendron species in south of Gansu .
Chinese Journal of Plant Ecology , 2020, 44(10): 1040-1049. DOI:
10.17521/cjpe.2020.0119 生物量在不同器官间的分配是植物生态学研究的核心问题(
West et al .,1999 ;
Enquist & Niklas, 2002 ;
Poorter et al ., 2012 )。生物量分配模式不仅有助于从易测量的地上生物量估算地下生物量, 还体现着植物不同的生态策略(
Weiner, 2004 ), 反映出光合产物对植物不同器官(地上不同器官之间, 地上器官和地下器官之间)的投资以适应环境条件(
Niklas, 2006 ;
Poorter & Sack, 2012 ;
闫帮国等, 2016 )。最优分配假说认为, 植物倾向于将资源优先分配到能获得限制性资源的器官, 以帮助植物获取更多的受限资源, 从而维持最大生长速率(
Bloom et al ., 1985 ;
McCarthy & Enquist, 2007 ), 这意味着, 根据植物生长的环境, 如果光照受限, 生物量将倾向于叶和枝分配; 如果营养或水分受限, 生物量将倾向于根系分配(
Gleeson & Tilman, 1990 )。但植物生物量分配模式存在个体发育漂变(
Poorter & Sack, 2012 ;
Poorter et al ., 2015 ;
王杨等, 2017 ), 其分配比例会随植株的大小变化而变化(
Poorter et al ., 2015 ;
陈国鹏等, 2016 ), 使得最优分配假说的使用受到了限制, 而异速生长理论能较好地解决这一问题(
Weiner, 2004 ;
Gargaglione et al ., 2010 ), 且已被越来越多地应用到生物量分配研究中, 有****也曾对两种方法的优劣进行了比较(
McCarthy & Enquist, 2007 ;
Poorter & Sack, 2012 ), 更赞同将两种方法结合应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 )。
异速生长理论认为, 生物量的分配主要由植物的大小决定, 并可用幂函数
Y 1 =
βY 2 α 表示, 该方程经过对数转换后, 可线性化为log(
Y 1 ) = log(
β )
+ α log(
Y 2 ), 其中
Y 1 和
Y 2 是相互依赖的变量(如茎生物量和根生物量), log(
β )是异速生长常数,
α 是异速生长指数(
Niklas & Enquist, 2002a ,
2002b )。当
α = 1时, 模型反映的是等速生长关系; 当
α ≠ 1时, 模型反映的是异速生长关系(
Niklas & Enquist, 2002b )。生态代谢理论认为, 全球尺度上植物个体生物量具有恒定的异速生长指数, 其地上、地下生物量间均表现为异速生长关系, 叶生物量与茎生物量或地下生物量间的异速生长指数为0.75 (
West et al ., 1997 ;
Enquist & Niklas, 2002 ;
Niklas & Enquist, 2002 ), 但后续研究又证明小个体植物在叶生物量、茎生物量和地下生物量间也可能为等速关系(
Niklas, 2005 ;
Cheng et al ., 2014 ;
陈国鹏和赵文智, 2016 ),
Poorter等(2015) 进一步分析个体大小影响时发现, 植物叶与茎、叶与根、茎与根等生物量间的异速生长指数随个体增大而减小。尽管基于生态代谢理论的异速生长指数在不同尺度上得到了检验(
Cheng & Niklas, 2007 ), 但是该理论以关注种间的变异为主, 跨物种的恒定关系不一定适用于种内, 对物种作用的考虑不足(
Müller et al ., 2000 ;
Cheng et al ., 2014 ), 尤其对一些极端生境的物种关注较少也限制着该理论的普适性。
理论上来说, 极端环境通过长期的筛选作用, 其适生物种响应资源约束时的生物量权衡策略应具有相似性(
Reich et al ., 2014 )。如, 在高寒山地, 高海拔、寒冷、短生长季和低气压等环境约束可能使植物的资源权衡具有独特表现(
高巧等, 2014 ;
左有璐等, 2014 ;
刘尊驰等, 2015 ), 植物会增加根生物量、降低叶生物量比例来适应较冷的环境(
徐波等, 2013 )。但目前的相关研究材料以常绿裸子植物或落叶被子植物为主(
Reich et al ., 2014 ;
Poorter et al ., 2015 ), 对高寒区生长的常绿被子植物的生物量权衡机制缺乏认知, 而常绿植物与落叶植物叶生物量分配策略存在差别, 常绿植物在叶片上投入的较多, 因此叶生物量分配比例高于落叶植物(
Poorter et al ., 2015 )。
广布于我国青藏高原及其边缘山地的杜鹃(
Rhododendron spp.)常绿灌丛是亚高山暗针叶林向高山草甸过渡的群落类型, 在维持高山-亚高山生态系统稳定等方面具有关键作用(
钟泽兵等, 2014 ;
杨路存等, 2018 )。本研究以甘肃南部高山-亚高山区常见的7种杜鹃为材料, 通过全株收获法测定根、茎和叶的生物量, 采用标准化主轴法估计检验各器官间的异速生长关系, 并进行种间差异比较, 以期了解高寒杜鹃的生物量权衡策略, 为进一步认识高寒植物规避风险、提高适应性提供理论基础。
1 材料和方法 1.1 研究区概况 白龙江林业管理局的舟曲生态建设局(103.95°- 104.70° E, 33.24°-33.90° N, 1 173-4 536 m), 处在秦岭西翼与岷山山脉交汇地带, 属西秦岭地质构造带南部陇南山地。研究区年降水量951.0 mm, 年蒸发量918.8 mm, 相对湿度为82.0%, 干燥度0.52, 年平均气温4.3 ℃, 全年无霜期96.7天, 日平均气温≥5 ℃的日数为178天, 年积温1 994.6 ℃。植被分布的垂直带谱明显, 从低到高依次为干旱河谷灌丛、亚高山次生林、亚高山暗针叶林和高山杜鹃灌丛。杜鹃灌丛建群种有陇蜀杜鹃(
Rhododendron przewalskii )、黄毛杜鹃(
R. rufum )、头花杜鹃(
R. capitatum )和山光杜鹃(
R. oreodoxa )等, 优势种有美容杜鹃(
R. calophytum )、太白杜鹃(
R. taibaiense )和麻花杜鹃(
R. maculiferum )等。
1.2 研究方法 在对生态局资源干事访谈和初步踏查后, 选定可及性强的扎尕梁、洋布梁和人命池沟等处杜鹃灌丛为研究对象, 在杜鹃灌丛中布设5 m × 5 m的样方(样方信息见
表1 ), 开展群落学调查, 并根据基径分布特征, 粗选每种杜鹃灌木的分级标准木, 详细测量标准木的基径、株高和冠幅后用全株收获法带回实验室烘干测定根、茎、叶生物量。
Table 1 表1 表1 甘肃南部7种高寒杜鹃的样方信息
Table 1
Information for samples of seven alpine Rhododendron species in south of Gansu 编号No. 纬度 经度 海拔 坡向 坡度 地点 优势种 盖度 密度 -2 ) 平均基径BD 平均株高PH 1 33.72° 104.09° 3 488 东北 43 洋布梁 黄毛杜鹃 R. rufum 100 2.36 55.32 256.95 2 33.75° 104.15° 2 810 东 28 烧哈沟脑 美容杜鹃、山光杜鹃、岷江冷杉、太白杜鹃、橿子栎、峨眉蔷薇 R. calophytum, R. oreodoxa, Abies faxoniana, R. taibaiense, Quercus baronii, Rosa omeiensis 65 0.45 71.34 259.13 3 33.00° 104.14° 2 766 西北 33 人命池沟 美容杜鹃、山光杜鹃、岷江冷杉、糙皮桦Rhododendron. calophytum, R. oreodoxa, Abies faxoniana, Betula utilis 55 4.05 19.67 208.15 4 33.71° 104.05° 3 488 东南 12 八小号 黄毛杜鹃、头花杜鹃、高山柳 R. rufum, R. capitatum, Salix cupularis 95 4.84 15.51 52.26 5 33.71° 104.05° 3 494 东南 9 八小号 陇蜀杜鹃、头花杜鹃、金露梅、高山柳R. przewalskii, R. capitatum, Potentilla fruticosa, Salix cupularis 95 9.04 10.68 107.79 6 33.69° 104.05° 3 517 东 East 44 青砂梁 陇蜀杜鹃、头花杜鹃R. przewalskii, R. capitatum 100 12.34 13.32 56.32 7 33.34° 104.50° 3 693 东 East 36 扎尕梁 陇蜀杜鹃、头花杜鹃R. przewalskii, R. capitatum 66 8.58 8.72 38.42 8 33.35° 104.50° 3 605 东 East 38 扎尕梁 陇蜀杜鹃、头花杜鹃、高山柳R. przewalskii, R. capitatum, Salix cupularis 88 7.66 22.35 106.58 9 33.36° 104.52° 3 151 北 North 15 扎尕梁 岷江冷杉、山光杜鹃、 麻花杜鹃Abies faxoniana, R. oreodoxa, 60 3.65 35.57 183.36
BD, basal diameter; PH, plant height新窗口打开 |
下载CSV 1.3 数据分析 用单因素方差分析和Tukey’s HSD多重比较分析根、茎和叶生物量分配比例的种间差异。采用标准主轴回归(SMA)在软件Standardized Major Axis Tests and Routines (SMATR)中计算不同器官生物量间的异速生长指数
α 和常数log(
β ), 若异速生长指数
α 与1差异显著, 则表明两变量间为异速生长关系; 若差异不显著, 则为等速生长关系; 检验不同物种间SMA斜率
α 是否存在异质性, 当斜率异质时, 进行多重比较; 当斜率同质时, 进一步采用Wald显著性检验分析组间是否存在
y 轴截距和SMA斜率方向上的变化, 并进行多重比较。
2 结果 2.1 生物量分配特征 7种杜鹃根、茎和叶间生物量的分配比例在种内存在差异(
图1A -1G), 单因素方差分析表明, 根、茎和叶生物量分配比例的种间差异显著(
图1H ,
p < 0.05)。多重比较(
表2 )发现: 根生物量分配比例的种间变异较小, 仅陇蜀杜鹃显著小于黄毛杜鹃、山光杜鹃和麻花杜鹃, 其余种间差异不显著; 叶生物量分配比例的种间变异次之, 美容杜鹃显著大于其他物种, 山光杜鹃显著大于陇蜀杜鹃、头花杜鹃和麻花杜鹃, 头花杜鹃显著大于麻花杜鹃; 茎生物量分配比例的种间变异最大, 美容杜鹃显著小于其他物种, 山光杜鹃显著小于4个物种, 麻花杜鹃、黄毛杜鹃和太白杜鹃显著小于陇蜀杜鹃。7种杜鹃根、茎、叶生物量的平均分配比例分别为35.57%、45.61%和18.83%。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1甘肃南部7种高寒杜鹃生物量分配特征。A, 黄毛杜鹃。B, 陇蜀杜鹃。C, 美容杜鹃。D, 山光杜鹃。E, 太白杜鹃。F, 头花杜鹃。G, 麻花杜鹃。H, 所有物种。P LB , 叶生物量所占比例。P SB , 茎生物量所占比例。P RB , 根生物量所占比例。 Fig. 1Biomass allocation for seven alpine Rhododendron species in south of Gansu. A, R. rufum. B, R. przewalskii. C, R. calophytum. D, R. oreodoxa. E, R. taibaiense. F, R. capitatum. G, R. maculiferum. H, Rhododendron spp. P LB , percent of leaf biomass. P SB , percent of stem biomass. P RB , percent of root biomass. Table 2 表2 表2 甘肃南部7种高寒杜鹃不同器官生物量分配的多重比较
Table 2
Multiple comparison of the biomass allocation of different organs for seven alpine Rhododendron species in south of Gansu 种 根生物量 Root biomass 茎生物量 Stem biomass 叶生物量 Leaf biomass A B C D E F A B C D E F A B C D E F B * * ns C ns ns * * * * D ns * ns * * * * ns * E ns ns ns ns ns * * * ns ns * ns F ns ns ns ns ns ns ns * * ns ns ns * * ns G ns * ns ns ns ns ns * * ns ns ns ns ns * * * ns
A, 黄毛杜鹃; B, 陇蜀杜鹃; C, 美容杜鹃; D, 山光杜鹃; E, 太白杜鹃; F, 头花杜鹃; G, 麻花杜鹃。*, p < 0.05; **, p < 0.01; ns, p > 0.05。 A, R. rufum; B, R. przewalskii; C, R. calophytum; D, R. oreodoxa; E, R. taibaiense; F, R. capitatum; G, R. maculiferum . *, p < 0.05; **, p < 0.01; ns, p > 0.05. 新窗口打开 |
下载CSV 2.2 异速生长关系 2.2.1 叶生物量与茎生物量的异速生长关系
7种杜鹃叶生物量与茎生物量间的异速生长指数介于0.584-1.079 (
图2 ), 以山光杜鹃的最小, 头花杜鹃的最大, 斜率的种间差异显著(
表3 ); 基于95%置信区间重叠部分和方差分析表明, 头花杜鹃(95%
CI = 0.995-1.170,
CI 为置信区间)和麻花杜鹃(95%
CI = 0.833-1.203)的异速生长指数与1.0无显著差异, 其余5种杜鹃的异速生长指数显著小于1.0 (
p < 0.01)。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2甘肃南部7种高寒杜鹃叶生物量(B L )与茎生物量(B S )的回归关系。A, 黄毛杜鹃。B, 陇蜀杜鹃。C, 美容杜鹃。D, 山光杜鹃。E, 太白杜鹃。F, 头花杜鹃。G, 麻花杜鹃。H, 所有物种。CI, 置信区间。 Fig. 2Regression relationships between the leaf biomass (B L ) and stem biomass (B S ) for seven alpine Rhododendron species in south of Gansu. A, R. rufum. B, R. przewalskii. C, R. calophytum. D, R. oreodoxa. E, R. taibaiense. F, R. capitatum. G, R. maculiferum. H, Rhododendron spp. CI, confidence interval. Table 3 表3 表3 甘肃南部7种高寒杜鹃地上地下生物量异速生长指数的多重比较
Table 3
Multiple comparison of allometric slopes between above- and below-ground biomass for seven alpine Rhododendron species in south of Gansu 树种 logB L vs. logB S logB L vs. logB R logB S vs. logB R logB A vs. logB R A B C D E F A B C D E F A B C D E F A B C D E F B ns ** ** ** C ns ns ** ** ** ** ** ** D * * ns ** ** ns * ** ** ** ** ** E ** * ** ** ns ** ** ** ** ** ns * ** ** ** ns F ** ** ** ** * ** ns ** ** ** ns ** ** ns ** ns ** ** ** ** G ** * ** ** ns ns ** ns ** ** ** ** ** ns ** ** ** ** ** ns ** ** ** **
B A , 地上生物量; B L , 叶生物量; B R , 根生物量; B S , 茎生物量。A, 黄毛杜鹃; B, 陇蜀杜鹃; C, 美容杜鹃; D, 山光杜鹃; E, 太白杜鹃; F, 头花杜鹃; G, 麻花杜鹃。*, p < 0.05; **, p < 0.01; ns, p > 0.05。B A , aboveground biomass; B L , leaf biomass; B R , root biomass; B S , stem biomass. A, R. rufum; B, R. przewalskii; C, R. calophytum; D, R. oreodoxa; E, R. taibaiense; F, R. capitatum; G, R. maculiferum. *, p < 0.05; **, p < 0.01; ns, p > 0.05.新窗口打开 |
下载CSV 2.2.2 叶生物量与根生物量的异速生长关系
叶生物量与根生物量的异速生长指数中(
图3A -3G), 美容杜鹃的0.543为最小、麻花杜鹃的1.625最大; 黄毛杜鹃、美容杜鹃、山光杜鹃和太白杜鹃的异速生长指数显著小于1.0, 其余3种显著大于1.0。7种杜鹃无共有斜率, 多重比较表明(
表3 ), 黄毛杜鹃与太白杜鹃(
p = 0.652)、陇蜀杜鹃与头花杜鹃(
p = 0.296)和麻花杜鹃(
p = 0.141)、美容杜鹃与山光杜鹃(
p = 0.475)的斜率无显著差异, 其余物种间斜率差异极显著(
p < 0.01)。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3甘肃南部7种高寒杜鹃叶生物量(B L )与根生物量(B R )的回归关系。A, 黄毛杜鹃。B, 陇蜀杜鹃。C, 美容杜鹃。D, 山光杜鹃。E, 太白杜鹃。F, 头花杜鹃。G, 麻花杜鹃。H, 所有物种。CI, 置信区间。 Fig. 3Regression relationships between the leaf biomass (B L ) and root biomass (B R ) for seven alpine Rhododendron species in south of Gansu. A, R. rufum. B, R. przewalskii. C, R. calophytum. D, R. oreodoxa. E, R. taibaiense. F, R. capitatum. G, R. maculiferum. H, Rhododendron spp. CI, confidence interval. 2.2.3 茎生物量与根生物量的异速生长关系
茎生物量与根生物量间的异速生长指数中(
图4A -4G): 陇蜀杜鹃、麻花杜鹃和黄毛杜鹃的显著大于1.0 (
p < 0.001); 头花杜鹃和山光杜鹃的与理论值1.0无显著差异(
p = 0.06 (头花杜鹃)、0.907 (山光杜鹃)); 太白杜鹃和美容杜鹃的显著小于1.0 (
p < 0.05)。多重比较表明(
表3 ), 除太白杜鹃与美容杜鹃(
p = 0.285)、头花杜鹃与黄毛杜鹃(
p = 0.289)和山光杜鹃(
p = 0.192)、麻花杜鹃与陇蜀杜鹃(
p = 0.577)间的差异不显著外, 其余各物种间异速生长指数差异显著(
p < 0.05)。
图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4甘肃南部7种高寒杜鹃茎生物量(B S )与根生物量(B R )的回归关系。A, 黄毛杜鹃。B, 陇蜀杜鹃。C, 美容杜鹃。D, 山光杜鹃。E, 太白杜鹃。F, 头花杜鹃。G, 麻花杜鹃。H, 所有物种。CI, 置信区间。 Fig. 4Regression relationships between the stem biomass (B S ) and root biomass (B R ) for seven alpine Rhododendron species in south of Gansu. A, R. rufum. B, R. przewalskii. C, R. calophytum. D, R. oreodoxa. E, R. taibaiense. F, R. capitatum. G, R. maculiferum. H, Rhododendron spp. CI, confidence interval. 2.2.4 地上生物量与根生物量的异速生长关系
地上生物量与根生物量异速生长指数的种间差异也很明显(
图5 ;
表3 ), 其中美容杜鹃、山光杜鹃和太白杜鹃的显著小于1.0 (
p < 0.01), 黄毛杜鹃和头花杜鹃的与1.0无显著差异(
p = 0.191 (黄毛杜鹃)、0.053 (头花杜鹃)), 麻花杜鹃和陇蜀杜鹃的显著大于1.0 (
p < 0.01); 除黄毛杜鹃与头花杜鹃(
p = 0.973)、陇蜀杜鹃与麻花杜鹃(
p = 0.860)、山光杜鹃与太白杜鹃(
p = 0.717)的异速生长指数不显著外, 其余物种间的差异极显著(
p < 0.01)。
图5 新窗口打开 |
下载原图ZIP |
生成PPT 图5甘肃南部7种高寒杜鹃地上生物量(B A )与根生物量(B R )的回归关系。A, 黄毛杜鹃。B, 陇蜀杜鹃。C, 美容杜鹃。D, 山光杜鹃。E, 太白杜鹃。F, 头花杜鹃。G, 麻花杜鹃。H, 所有物种。CI, 置信区间。 Fig. 5Regression relationships of aboveground biomass (B A ) and root biomass (B R ) for seven alpine Rhododendron species in south of Gansu. A, R. rufum. B, R. przewalskii. C, R. calophytum. D, R. oreodoxa. E, R. taibaiense. F, R. capitatum. G, R. maculiferum. H, Rhododendron spp. CI, confidence interval. 3 讨论 尽管异速生长分析不受个体大小等因素干扰, 可以较好地检验不同器官生长发育协调性, 并确定这种协调性在物种间的变化或对环境的响应, 但在一些生态模型的构建研究中, 叶、茎和根分配比例的实际值要比器官间的协调性重要(
Poorter & Sack, 2012 ), 考虑到不同分析方法在研究生物量分配中的局限性, 将异速生长法与最优分配假说结合已在许多研究中得到了应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ;
陈国鹏等, 2016 )。本研究也结合最优分配假说与异速生长分析来研究甘肃南部高山-亚高山区杜鹃地上、地下器官生物量的分配特征, 结果表明7种高寒常绿杜鹃根、茎、叶生物量分配比例和异速生长关系受物种影响显著, 高寒杜鹃对特殊生境适应过程中的生物量分配模式有许多相似之处, 而物种差异又导致相似生境中生物量分配模式的分异。
树木对温度适应性策略的假设认为, 在寒冷的气候环境下, 根的分配比例较大、叶的分配比例较小(
Reich et al ., 2014 ), 本研究中, 7种杜鹃根生物量分配比例高达35.57%, 个别物种甚至超过40% (
图1 ), 高于全球绝大多数植物(
Poorter et al ., 2015 ), 也高于我国亚热带山地杜鹃灌丛根系生物量的分配比例(
张蔷等, 2017 ), 与一些高寒植被(
高巧等, 2014 ;
钟泽兵等, 2014 ;
杨路存等, 2018 ;
左有璐等, 2018 )相似, 低速率的养分循环和受限的土壤溶液运动驱动着高山、亚高山地区的低营养供应(
魏晶等, 2004 ;
Luo et al ., 2012 ;
Reich et al ., 2014 ), 这使得高寒杜鹃需提高根生物量分配比例以增强从寒冷土壤中吸收养分, 生物量向根系中的倾向分配支持最优分配假说。7种杜鹃叶生物量分配比例低于全球绝大多数植物(
Poorter et al ., 2015 ), 其适应机制也与一些高寒植物(
Reich et al ., 2014 )相似, 如
钟泽兵等(2014) 对青海玉树海拔3 715-4 296 m的5种建群灌木的研究发现叶生物量分配比例不足10%。甘肃南部高山-亚高山区的杜鹃会通过减少资源向叶片的分配, 保持常绿, 以较低的叶片周转率来适应寒冷环境。随着植株叶投资盈利能力的下降, 叶生物量将饱和, 投资会逐渐转向茎和根(
Poorter et al ., 2015 ), 而为增强植株的机械支撑, 必须增加茎的生物量投资(
Dybzinski et al ., 2011 )。本研究中7种高寒杜鹃茎生物量分配平均比例为45.61%, 接近于全球数据库平均值(
Poorter et al ., 2015 )。同时, 相比于叶和根, 茎生物量分配比例的种间差异最大(可达21%), 且多组物种间的差异显著(
表2 ), 这与
Poorter等(2015) 对全球1 200种植物整合分析的结果具有一致性, 茎不仅是植株的支撑、运输体系, 还在光竞争时向四周伸展小枝扩展空域, 而小枝的扩张又带来了高寒区防风的压力, 功能的多样性可能导致茎生物量分配的可塑性。各物种分配的差异还可能与茎构型有关, 如黄毛杜鹃和陇蜀杜鹃多为丛生, 茎枝扭曲, 其茎生物量分配比例的变异范围较大; 而美容杜鹃和山光杜鹃主干通直, 其茎生物量分配比例的变幅较小(
图1 )。
异速生长理论预测小个体植物叶、茎、根器官生长可能为等速关系(
Enquist & Niklas, 2002 ;
Weiner, 2004 ;
Niklas, 2006 ), 这种基于种间变异建立的跨物种关系已在种内变异的研究中得到了较多应用(
Gargaglione et al ., 2010 ;
Cheng et al ., 2014 ;
陈国鹏等, 2016 ;
王杨等, 2017 ), 如
Cheng等(2014) 发现南亚热带生长的5种苗木叶、茎和根生物量的积累为等速或近似等速生长关系。本研究中, 7种高寒杜鹃叶与茎、叶与根以及茎与根之间既有异速生长表现, 也有等速生长表现(
图2 -5), 与荒漠植物(
杨昊天等, 2013 ;
陶冶和张元明, 2014 )、干热河谷草本植物(
闫帮国等, 2016 )的异速生长关系具有相似性, 不同于川西高寒草甸优势植物(
左有璐等, 2018 ), 并不完全支持小个体器官间等速生长关系的预测, 也不完全支持生态代谢理论的预测, 这可能是由于植物在适应特殊环境的过程中, 不同物种间具有很大的功能群差异, 这些功能群差异可能是不同器官生物量间异速生长关系分化的主要因素(
Poorter et al ., 2015 ;
闫帮国等, 2016 ), 而基于跨物种建立的生态代谢理论、小个体等速生长理论忽略了物种间的功能群差异。
物种是影响植物器官生物量分配异速生长关系的重要因素(
Poorter et al ., 2012 ;
闫帮国等, 2016 ;
张蔷等, 2017 ), 基于大样本的研究则发现生物量分配比例与物种的系统发育有关, 如裸子植物叶片上的投入更多(
Reich et al ., 2014 ;
Poorter et al ., 2015 )。本研究中7种杜鹃隶属于同科同属, 其生物量分配比例及异速生长关系的种间差异显著(
表2 ,
表3 ), 陇蜀杜鹃根生物量分配比例较低, 美容杜鹃叶生物量分配比例较高而茎生物量分配比例较低, 黄毛杜鹃、头花杜鹃地上生物量与地下生物量呈等速生长, 陇蜀杜鹃、麻花杜鹃地上生物量的积累速率大于地下, 其余3种地上生物量积累速率小于地下(
图5 ), 而叶与茎、叶与根、茎与根间的异速生长关系的物种差异更为复杂多样, 如黄毛杜鹃、陇蜀杜鹃、美容杜鹃和山光杜鹃的叶与茎, 所有物种的叶与根, 美容杜鹃、太白杜鹃、黄毛杜鹃、陇蜀杜鹃和麻花杜鹃的茎与根为异速生长关系, 而太白杜鹃、头花杜鹃和麻花杜鹃的叶与茎, 山光杜鹃和头花杜鹃的茎与根为等速生长关系(
图2 -4)。这也说明不同的物种可能在进化过程中会通过生态位分化而占有不同的环境, 在本研究区域, 头花杜鹃主要分布于海拔3 500 m以上并形成单优群落或与陇蜀杜鹃形成共生群落; 陇蜀杜鹃、黄毛杜鹃在海拔3 000-3 700 m间有分布, 但在3 200 m以上才形成单优群落或群落优势种; 山光杜鹃和麻花杜鹃海拔分布范围跨度大(2 300-3 500 m), 但呈群落优势种的仅见3 000 m处个别的冷杉皆伐更新迹地,
Veresoglou和 Pe?uelas (2019) 分析欧亚大陆植物生物量时发现分配的变异强度与物种分布范围有密切的关系, 但杜鹃分布范围与生物量分配特征间是否存在相关关系, 尚需持续研究。
本研究以甘肃南部高山-亚高山区7种同科同属的杜鹃为材料, 结合最优分配假说和异速生长理论, 开展了高寒植物根、茎和叶生物量分配研究, 对高寒植物的生活史策略认知具有重要意义。研究结果表明, 根、茎、叶生物量的分配平均比例为45.61%、35.57%和18.83%, 根和叶生物量的分配比例体现着高寒植物的适应策略; 7种高寒杜鹃的叶与茎、叶与根、茎与根以及地上生物量与地下生物量之间既有异速生长关系, 也有等速生长关系, 异速生长指数不完全支持生态代谢理论和小个体等速生长理论的参考值; 各器官异速生长关系的物种差异显著。并认为结合最优分配理论和异速生长理论能更好地解释7种高寒杜鹃生物量的变异及适应机制。
致谢 感谢甘肃农业大学科技创新基金(GSAU-RCZX201708、GAU-XKJS-2018-097和GSAU-STS-1734)资助。
[1] Bloom AJ Chapin III FS Mooney HA 1985 ). Resource limitation in plants-an economic analogyAnnual Review of Ecology and Systematics, 363 -392 . [本文引用: 1] [2] Chen GP Zhao WZ 2016 ). Effects of age classes on metabolic exponents of Salix psammophila branchesChinese Journal of Applied Ecology, 1870 -1876 . [本文引用: 3] [ 陈国鹏 , 赵文智 (2016 ). 沙柳丛生枝代谢指数的龄级效应应用生态学报 , 27, 1870 -1876 .] [本文引用: 3] [3] Chen GP Zhao WZ He SX Fu X 2016 ). Biomass allocation and allometric relationship in aboveground components of Salix psammophila branchesJournal of Desert Research, 357 -363 . [本文引用: 1] [ 陈国鹏 , 赵文智 , 何世雄 , 付晓 (2016 ). 沙柳(Salix psammophila )丛生枝生物量最优分配与异速生长中国沙漠 , 36, 357 -363 .] [本文引用: 1] [4] Cheng DL Ma YZ Zhong QL Xu WF 2014 ). Allometric scaling relationship between above- and below-ground biomass within and across five woody seedlingsEcology and Evolution, 3968 -3977 . [本文引用: 4] [5] Cheng DL Niklas KJ 2007 ). Above- and below-ground biomass relationships across 1534 forested communitiesAnnals of Botany, 95 -102 . [本文引用: 1] [6] Dybzinski R Farrior C Wolf A Reich PB Pacala SW 2011 ). Evolutionarily stable strategy carbon allocation to foliage, wood, and fine roots in trees competing for light and nitrogen: an analytically tractable, individual-based model and quantitative comparisons to dataThe American Naturalist, 153 -166 . [本文引用: 1] [7] Enquist BJ Niklas KJ 2002 ). Global allocation rules for patterns of biomass partitioning in seed plantsScience, 1517 -1520 . [本文引用: 3] [8] Gao Q Yang XC Yin CY Liu Q 2014 ). Estimation of biomass allocation and carbon density in alpine dwarf shrubs in Garzê Zangzu Autonomous Prefecture of Sichuan Province, ChinaChinese Journal of Plant Ecology, 355 -365 . [本文引用: 2] [ 高巧 , 阳小成 , 尹春英 , 刘庆 (2014 ). 四川省甘孜藏族自治州高寒矮灌丛生物量分配及其碳密度的估算植物生态学报 , 38, 355 -365 .] [本文引用: 2] [9] Gargaglione V Peri PL Rubio G 2010 ). Allometric relations for biomass partitioning of Nothofagus antarctica trees of different crown classes over a site quality gradientForest Ecology and Management, 1118 -1126 . [本文引用: 2] [10] Gleeson SK Tilman D 1990 ). Allocation and the transient dynamics of succession on poor soilsEcology, 1144 -1155 . [本文引用: 1] [11] Liu ZC Liu HF Zhao D Luo N Sun YY Hao XR Liu T 2015 ). Influence of altitude and difference of different- sized individuals on reproductive allocation in Salsola affinis C. A. Mey. and Salsola nitraria PallActa Ecologica Sinica, 5957 -5965 . [本文引用: 1] [ 刘尊驰 , 刘华峰 , 赵丹 , 罗宁 , 孙园园 , 郝晓冉 , 刘彤 (2015 ). 紫翅猪毛菜、钠猪毛菜不同个体大小繁殖分配差异及随海拔的变化生态学报 , 35, 5957 -5965 .] [本文引用: 1] [12] Luo YJ Wang XK Zhang XQ Booth TH Lu F 2012 ). Root: shoot ratios across China’s forests: forest type and climatic effectsForest Ecology and Management, 19 -25 . [本文引用: 1] [13] McCarthy MC Enquist BJ 2007 ). Consistency between an allometric approach and optimal partitioning theory in global patterns of plant biomass allocationFunctional Ecology, 713 -720 . [本文引用: 2] [14] Müller I Schmid B Weiner J 2000 ). The effect of nutrient availability on biomass allocation patterns in 27 species of herbaceous plantsPerspectives in Plant Ecology, Evolution and Systematics, 115 -127 . [本文引用: 1] [15] Niklas KJ 2005 ). Modelling below- and above-ground biomass for non-woody and woody plantsAnnals of Botany, 315 -321 . [本文引用: 1] [16] Niklas KJ 2006 ). A phyletic perspective on the allometry of plant biomass-partitioning patterns and functionally equivalent organ-categoriesNew Phytologis t, 27 -40 . [本文引用: 2] [17] Niklas KJ Enquist BJ 2002 a). Canonical rules for plant organ biomass partitioning and annual allocationAmerican Journal of Botany, 812 -819 . [本文引用: 1] [18] Niklas KJ Enquist BJ 2002 b). On the vegetative biomass partitioning of seed plant leaves, stems, and rootsThe American Naturalist, 482 -497 . [本文引用: 3] [19] Poorter H Jagodzinski AM Ruiz-Peinado R Kuyah S Luo YJ Oleksyn J Usoltsev VA Buckley TN Reich PB Sack L 2015 ). How does biomass distribution change with size and differ among species? An analysis for 1200 plant species from five continentsNew Phytologist, 736 -749 . [本文引用: 12] [20] Poorter H Niklas KJ Reich PB Oleksyn J Poot P Mommer L 2012 ). Biomass allocation to leaves, stems and roots: meta-analyses of interspecific variation and environmental controlNew Phytologist, 30 -50 . [本文引用: 4] [21] Poorter H Sack L 2012 ). Pitfalls and possibilities in the analysis of biomass allocation patterns in plantsFrontiers in Plant Science, 259 -269 . [本文引用: 6] [22] Reich PB Luo YJ Bradford JB Poorter H Perry CH Oleksyn J 2014 ). Temperature drives global patterns in forest biomass distribution in leaves, stems, and rootsProceedings of the National Academy of Sciences of the United States of America, 13721 -13726 . [本文引用: 6] [23] Tao Y Zhang YM 2014 ). Biomass allocation patterns and allometric relationships of six ephemeroid species in Junggar Basin, ChinaActa Prataculturae Sinica, 38 -48 . [本文引用: 1] [ 陶冶 , 张元明 (2014 ). 准噶尔荒漠6种类短命植物生物量分配与异速生长关系草业学报 , 23, 38 -48 .] [本文引用: 1] [24] Veresoglou SD Pe?uelas J 2019 ). Variance in biomass- allocation fractions is explained by distribution in European treesNew Phytologist, 1352 -1363 . [本文引用: 1] [25] Wang Y Xu WT Xiong GM Li JX Zhao CM Lu ZJ Li YL Xie ZQ 2017 ). Biomass allocation patterns of Loropetalum chinense Chinese Journal of Plant Ecology, 105 -114 . [本文引用: 2] [ 王杨 , 徐文婷 , 熊高明 , 李家湘 , 赵常明 , 卢志军 , 李跃林 , 谢宗强 (2017 ). 檵木生物量分配特征植物生态学报 , 41, 105 -114 .] [本文引用: 2] [26] Wei J Wu G Deng HB 2004 ). Vegetation biomass distribution characteristics of alpine tundra ecosystem in Changbai MountainsChinese Journal of Applied Ecology, 1999 -2004 . [本文引用: 1] [ 魏晶 , 吴钢 , 邓红兵 (2004 ). 长白山高山冻原植被生物量的分布规律应用生态学报 , 15, 1999 -2004 .] [本文引用: 1] [27] Weiner J 2004 ). Allocation, plasticity and allometry in plantsPerspectives in Plant Ecology, Evolution and Systematics, 207 -215 . [本文引用: 3] [28] West GB Brown JH Enquist BJ 1997 ). A general model for the origin of allometric scaling laws in biologyScience, 122 -126 . [本文引用: 1] [29] West GB Brown JH Enquist BJ 1999 ). A general model for the structure and allometry of plant vascular systemsNature, 664 -667 . DOI:
10.1038/23251 URL [本文引用: 1] [30] Xu B Wang JN Shi FS Gao J Wu N 2013 ). Adaptation of biomass allocation patterns of wild Fritillaria unibracteata to alpine environment in the eastern Qinghai-Xizang PlateauChinese Journal of Plant Ecology, 187 -196 . [本文引用: 1] [ 徐波 , 王金牛 , 石福孙 , 高景 , 吴宁 (2013 ). 青藏高原东缘野生暗紫贝母生物量分配格局对高山生态环境的适应植物生态学报 , 37, 187 -196 .] [本文引用: 1] [31] Yan BG Fan B He GX Shi LT Pan ZX Li JC Yue XW Liu GC 2016 ). Biomass allocations and their response to environmental factors for grass species in an arid-hot valleyChinese Journal of Applied Ecology, 3173 -3181 . [本文引用: 4] [ 闫帮国 , 樊博 , 何光熊 , 史亮涛 , 潘志贤 , 李建查 , 岳学文 , 刘刚才 (2016 ). 干热河谷草本植物生物量分配及其对环境因子的响应应用生态学报 , 27, 3173 -3181 .] [本文引用: 4] [32] Yang HT Li XR Liu LC Jia RL Wang ZR Li XJ Li G 2013 ). Biomass allocation patterns of four shrubs in desert grasslandJournal of Desert Research, 1340 -1348 . [本文引用: 3] [ 杨昊天 , 李新荣 , 刘立超 , 贾荣亮 , 王增如 , 李小军 , 李刚 (2013 ). 荒漠草地4种灌木生物量分配特征中国沙漠 , 33, 1340 -1348 .] [本文引用: 3] [33] Yang LC Zhao YH Xu WH Zhou GY 2018 ). Species diversity, biomass, and their relationship in the alpine shrubberies of Qinghai ProvinceActa Ecologica Sinica, 309 -315 . [本文引用: 2] [ 杨路存 , 赵玉红 , 徐文华 , 周国英 (2018 ). 青海省高寒灌丛物种多样性、生物量及其关系生态学报 , 38, 309 -315 .] [本文引用: 2] [34] Zhang Q Li JX Xu WT Xiong GM Xie ZQ 2017 ). Estimation of biomass allocation and carbon density of Rhododendron simsii shrubland in the subtropical mountainous areas of ChinaChinese Journal of Plant Ecology, 43 -52 . [本文引用: 2] [ 张蔷 , 李家湘 , 徐文婷 , 熊高明 , 谢宗强 (2017 ). 中国亚热带山地杜鹃灌丛生物量分配及其碳密度估算植物生态学报 , 41, 43 -52 .] [本文引用: 2] [35] Zhong ZB Yang LC Liu HC Song WZ Li F Zhou GY 2014 ). The main shrubs aboveground biomass and effect factors in Yushu, Qinghai, ChinaMountain Research, 678 -684 . [本文引用: 3] [ 钟泽兵 , 杨路存 , 刘何春 , 宋文珠 , 李璠 , 周国英 (2014 ). 青海玉树地区主要灌丛类型地上生物量及其影响因素山地学报 , 32, 678 -684 .] [本文引用: 3] [36] Zuo YL Wang ZM Xi XQ Xiang S Sun SC 2018 ). Plant biomass allocation strategies of the dominant species in an alpine meadow of northwestern Sichuan, ChinaChinese Journal of Applied Environmental Biology, 1195 -1203 . [本文引用: 3] [ 左有璐 , 王振孟 , 习新强 , 向双 , 孙书存 (2018 ). 川西北高寒草甸优势植物生物量分配对策应用与环境生物学报 , 24, 1195 -1203 .] [本文引用: 3] Resource limitation in plants-an economic analogy
1
1985
... 生物量在不同器官间的分配是植物生态学研究的核心问题(
West et al .,1999 ;
Enquist & Niklas, 2002 ;
Poorter et al ., 2012 ).生物量分配模式不仅有助于从易测量的地上生物量估算地下生物量, 还体现着植物不同的生态策略(
Weiner, 2004 ), 反映出光合产物对植物不同器官(地上不同器官之间, 地上器官和地下器官之间)的投资以适应环境条件(
Niklas, 2006 ;
Poorter & Sack, 2012 ;
闫帮国等, 2016 ).最优分配假说认为, 植物倾向于将资源优先分配到能获得限制性资源的器官, 以帮助植物获取更多的受限资源, 从而维持最大生长速率(
Bloom et al ., 1985 ;
McCarthy & Enquist, 2007 ), 这意味着, 根据植物生长的环境, 如果光照受限, 生物量将倾向于叶和枝分配; 如果营养或水分受限, 生物量将倾向于根系分配(
Gleeson & Tilman, 1990 ).但植物生物量分配模式存在个体发育漂变(
Poorter & Sack, 2012 ;
Poorter et al ., 2015 ;
王杨等, 2017 ), 其分配比例会随植株的大小变化而变化(
Poorter et al ., 2015 ;
陈国鹏等, 2016 ), 使得最优分配假说的使用受到了限制, 而异速生长理论能较好地解决这一问题(
Weiner, 2004 ;
Gargaglione et al ., 2010 ), 且已被越来越多地应用到生物量分配研究中, 有****也曾对两种方法的优劣进行了比较(
McCarthy & Enquist, 2007 ;
Poorter & Sack, 2012 ), 更赞同将两种方法结合应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ). ...
沙柳丛生枝代谢指数的龄级效应
3
2016
... 生物量在不同器官间的分配是植物生态学研究的核心问题(
West et al .,1999 ;
Enquist & Niklas, 2002 ;
Poorter et al ., 2012 ).生物量分配模式不仅有助于从易测量的地上生物量估算地下生物量, 还体现着植物不同的生态策略(
Weiner, 2004 ), 反映出光合产物对植物不同器官(地上不同器官之间, 地上器官和地下器官之间)的投资以适应环境条件(
Niklas, 2006 ;
Poorter & Sack, 2012 ;
闫帮国等, 2016 ).最优分配假说认为, 植物倾向于将资源优先分配到能获得限制性资源的器官, 以帮助植物获取更多的受限资源, 从而维持最大生长速率(
Bloom et al ., 1985 ;
McCarthy & Enquist, 2007 ), 这意味着, 根据植物生长的环境, 如果光照受限, 生物量将倾向于叶和枝分配; 如果营养或水分受限, 生物量将倾向于根系分配(
Gleeson & Tilman, 1990 ).但植物生物量分配模式存在个体发育漂变(
Poorter & Sack, 2012 ;
Poorter et al ., 2015 ;
王杨等, 2017 ), 其分配比例会随植株的大小变化而变化(
Poorter et al ., 2015 ;
陈国鹏等, 2016 ), 使得最优分配假说的使用受到了限制, 而异速生长理论能较好地解决这一问题(
Weiner, 2004 ;
Gargaglione et al ., 2010 ), 且已被越来越多地应用到生物量分配研究中, 有****也曾对两种方法的优劣进行了比较(
McCarthy & Enquist, 2007 ;
Poorter & Sack, 2012 ), 更赞同将两种方法结合应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ). ...
... 尽管异速生长分析不受个体大小等因素干扰, 可以较好地检验不同器官生长发育协调性, 并确定这种协调性在物种间的变化或对环境的响应, 但在一些生态模型的构建研究中, 叶、茎和根分配比例的实际值要比器官间的协调性重要(
Poorter & Sack, 2012 ), 考虑到不同分析方法在研究生物量分配中的局限性, 将异速生长法与最优分配假说结合已在许多研究中得到了应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ;
陈国鹏等, 2016 ).本研究也结合最优分配假说与异速生长分析来研究甘肃南部高山-亚高山区杜鹃地上、地下器官生物量的分配特征, 结果表明7种高寒常绿杜鹃根、茎、叶生物量分配比例和异速生长关系受物种影响显著, 高寒杜鹃对特殊生境适应过程中的生物量分配模式有许多相似之处, 而物种差异又导致相似生境中生物量分配模式的分异. ...
... 异速生长理论预测小个体植物叶、茎、根器官生长可能为等速关系(
Enquist & Niklas, 2002 ;
Weiner, 2004 ;
Niklas, 2006 ), 这种基于种间变异建立的跨物种关系已在种内变异的研究中得到了较多应用(
Gargaglione et al ., 2010 ;
Cheng et al ., 2014 ;
陈国鹏等, 2016 ;
王杨等, 2017 ), 如
Cheng等(2014) 发现南亚热带生长的5种苗木叶、茎和根生物量的积累为等速或近似等速生长关系.本研究中, 7种高寒杜鹃叶与茎、叶与根以及茎与根之间既有异速生长表现, 也有等速生长表现(
图2 -5), 与荒漠植物(
杨昊天等, 2013 ;
陶冶和张元明, 2014 )、干热河谷草本植物(
闫帮国等, 2016 )的异速生长关系具有相似性, 不同于川西高寒草甸优势植物(
左有璐等, 2018 ), 并不完全支持小个体器官间等速生长关系的预测, 也不完全支持生态代谢理论的预测, 这可能是由于植物在适应特殊环境的过程中, 不同物种间具有很大的功能群差异, 这些功能群差异可能是不同器官生物量间异速生长关系分化的主要因素(
Poorter et al ., 2015 ;
闫帮国等, 2016 ), 而基于跨物种建立的生态代谢理论、小个体等速生长理论忽略了物种间的功能群差异. ...
沙柳丛生枝代谢指数的龄级效应
3
2016
... 生物量在不同器官间的分配是植物生态学研究的核心问题(
West et al .,1999 ;
Enquist & Niklas, 2002 ;
Poorter et al ., 2012 ).生物量分配模式不仅有助于从易测量的地上生物量估算地下生物量, 还体现着植物不同的生态策略(
Weiner, 2004 ), 反映出光合产物对植物不同器官(地上不同器官之间, 地上器官和地下器官之间)的投资以适应环境条件(
Niklas, 2006 ;
Poorter & Sack, 2012 ;
闫帮国等, 2016 ).最优分配假说认为, 植物倾向于将资源优先分配到能获得限制性资源的器官, 以帮助植物获取更多的受限资源, 从而维持最大生长速率(
Bloom et al ., 1985 ;
McCarthy & Enquist, 2007 ), 这意味着, 根据植物生长的环境, 如果光照受限, 生物量将倾向于叶和枝分配; 如果营养或水分受限, 生物量将倾向于根系分配(
Gleeson & Tilman, 1990 ).但植物生物量分配模式存在个体发育漂变(
Poorter & Sack, 2012 ;
Poorter et al ., 2015 ;
王杨等, 2017 ), 其分配比例会随植株的大小变化而变化(
Poorter et al ., 2015 ;
陈国鹏等, 2016 ), 使得最优分配假说的使用受到了限制, 而异速生长理论能较好地解决这一问题(
Weiner, 2004 ;
Gargaglione et al ., 2010 ), 且已被越来越多地应用到生物量分配研究中, 有****也曾对两种方法的优劣进行了比较(
McCarthy & Enquist, 2007 ;
Poorter & Sack, 2012 ), 更赞同将两种方法结合应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ). ...
... 尽管异速生长分析不受个体大小等因素干扰, 可以较好地检验不同器官生长发育协调性, 并确定这种协调性在物种间的变化或对环境的响应, 但在一些生态模型的构建研究中, 叶、茎和根分配比例的实际值要比器官间的协调性重要(
Poorter & Sack, 2012 ), 考虑到不同分析方法在研究生物量分配中的局限性, 将异速生长法与最优分配假说结合已在许多研究中得到了应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ;
陈国鹏等, 2016 ).本研究也结合最优分配假说与异速生长分析来研究甘肃南部高山-亚高山区杜鹃地上、地下器官生物量的分配特征, 结果表明7种高寒常绿杜鹃根、茎、叶生物量分配比例和异速生长关系受物种影响显著, 高寒杜鹃对特殊生境适应过程中的生物量分配模式有许多相似之处, 而物种差异又导致相似生境中生物量分配模式的分异. ...
... 异速生长理论预测小个体植物叶、茎、根器官生长可能为等速关系(
Enquist & Niklas, 2002 ;
Weiner, 2004 ;
Niklas, 2006 ), 这种基于种间变异建立的跨物种关系已在种内变异的研究中得到了较多应用(
Gargaglione et al ., 2010 ;
Cheng et al ., 2014 ;
陈国鹏等, 2016 ;
王杨等, 2017 ), 如
Cheng等(2014) 发现南亚热带生长的5种苗木叶、茎和根生物量的积累为等速或近似等速生长关系.本研究中, 7种高寒杜鹃叶与茎、叶与根以及茎与根之间既有异速生长表现, 也有等速生长表现(
图2 -5), 与荒漠植物(
杨昊天等, 2013 ;
陶冶和张元明, 2014 )、干热河谷草本植物(
闫帮国等, 2016 )的异速生长关系具有相似性, 不同于川西高寒草甸优势植物(
左有璐等, 2018 ), 并不完全支持小个体器官间等速生长关系的预测, 也不完全支持生态代谢理论的预测, 这可能是由于植物在适应特殊环境的过程中, 不同物种间具有很大的功能群差异, 这些功能群差异可能是不同器官生物量间异速生长关系分化的主要因素(
Poorter et al ., 2015 ;
闫帮国等, 2016 ), 而基于跨物种建立的生态代谢理论、小个体等速生长理论忽略了物种间的功能群差异. ...
沙柳(
Salix psammophila )丛生枝生物量最优分配与异速生长
1
2016
... 异速生长理论认为, 生物量的分配主要由植物的大小决定, 并可用幂函数
Y 1 =
βY 2 α 表示, 该方程经过对数转换后, 可线性化为log(
Y 1 ) = log(
β )
+ α log(
Y 2 ), 其中
Y 1 和
Y 2 是相互依赖的变量(如茎生物量和根生物量), log(
β )是异速生长常数,
α 是异速生长指数(
Niklas & Enquist, 2002a ,
2002b ).当
α = 1时, 模型反映的是等速生长关系; 当
α ≠ 1时, 模型反映的是异速生长关系(
Niklas & Enquist, 2002b ).生态代谢理论认为, 全球尺度上植物个体生物量具有恒定的异速生长指数, 其地上、地下生物量间均表现为异速生长关系, 叶生物量与茎生物量或地下生物量间的异速生长指数为0.75 (
West et al ., 1997 ;
Enquist & Niklas, 2002 ;
Niklas & Enquist, 2002 ), 但后续研究又证明小个体植物在叶生物量、茎生物量和地下生物量间也可能为等速关系(
Niklas, 2005 ;
Cheng et al ., 2014 ;
陈国鹏和赵文智, 2016 ),
Poorter等(2015) 进一步分析个体大小影响时发现, 植物叶与茎、叶与根、茎与根等生物量间的异速生长指数随个体增大而减小.尽管基于生态代谢理论的异速生长指数在不同尺度上得到了检验(
Cheng & Niklas, 2007 ), 但是该理论以关注种间的变异为主, 跨物种的恒定关系不一定适用于种内, 对物种作用的考虑不足(
Müller et al ., 2000 ;
Cheng et al ., 2014 ), 尤其对一些极端生境的物种关注较少也限制着该理论的普适性. ...
沙柳(
Salix psammophila )丛生枝生物量最优分配与异速生长
1
2016
... 异速生长理论认为, 生物量的分配主要由植物的大小决定, 并可用幂函数
Y 1 =
βY 2 α 表示, 该方程经过对数转换后, 可线性化为log(
Y 1 ) = log(
β )
+ α log(
Y 2 ), 其中
Y 1 和
Y 2 是相互依赖的变量(如茎生物量和根生物量), log(
β )是异速生长常数,
α 是异速生长指数(
Niklas & Enquist, 2002a ,
2002b ).当
α = 1时, 模型反映的是等速生长关系; 当
α ≠ 1时, 模型反映的是异速生长关系(
Niklas & Enquist, 2002b ).生态代谢理论认为, 全球尺度上植物个体生物量具有恒定的异速生长指数, 其地上、地下生物量间均表现为异速生长关系, 叶生物量与茎生物量或地下生物量间的异速生长指数为0.75 (
West et al ., 1997 ;
Enquist & Niklas, 2002 ;
Niklas & Enquist, 2002 ), 但后续研究又证明小个体植物在叶生物量、茎生物量和地下生物量间也可能为等速关系(
Niklas, 2005 ;
Cheng et al ., 2014 ;
陈国鹏和赵文智, 2016 ),
Poorter等(2015) 进一步分析个体大小影响时发现, 植物叶与茎、叶与根、茎与根等生物量间的异速生长指数随个体增大而减小.尽管基于生态代谢理论的异速生长指数在不同尺度上得到了检验(
Cheng & Niklas, 2007 ), 但是该理论以关注种间的变异为主, 跨物种的恒定关系不一定适用于种内, 对物种作用的考虑不足(
Müller et al ., 2000 ;
Cheng et al ., 2014 ), 尤其对一些极端生境的物种关注较少也限制着该理论的普适性. ...
Allometric scaling relationship between above- and below-ground biomass within and across five woody seedlings
4
2014
... 异速生长理论认为, 生物量的分配主要由植物的大小决定, 并可用幂函数
Y 1 =
βY 2 α 表示, 该方程经过对数转换后, 可线性化为log(
Y 1 ) = log(
β )
+ α log(
Y 2 ), 其中
Y 1 和
Y 2 是相互依赖的变量(如茎生物量和根生物量), log(
β )是异速生长常数,
α 是异速生长指数(
Niklas & Enquist, 2002a ,
2002b ).当
α = 1时, 模型反映的是等速生长关系; 当
α ≠ 1时, 模型反映的是异速生长关系(
Niklas & Enquist, 2002b ).生态代谢理论认为, 全球尺度上植物个体生物量具有恒定的异速生长指数, 其地上、地下生物量间均表现为异速生长关系, 叶生物量与茎生物量或地下生物量间的异速生长指数为0.75 (
West et al ., 1997 ;
Enquist & Niklas, 2002 ;
Niklas & Enquist, 2002 ), 但后续研究又证明小个体植物在叶生物量、茎生物量和地下生物量间也可能为等速关系(
Niklas, 2005 ;
Cheng et al ., 2014 ;
陈国鹏和赵文智, 2016 ),
Poorter等(2015) 进一步分析个体大小影响时发现, 植物叶与茎、叶与根、茎与根等生物量间的异速生长指数随个体增大而减小.尽管基于生态代谢理论的异速生长指数在不同尺度上得到了检验(
Cheng & Niklas, 2007 ), 但是该理论以关注种间的变异为主, 跨物种的恒定关系不一定适用于种内, 对物种作用的考虑不足(
Müller et al ., 2000 ;
Cheng et al ., 2014 ), 尤其对一些极端生境的物种关注较少也限制着该理论的普适性. ...
... ;
Cheng et al ., 2014 ), 尤其对一些极端生境的物种关注较少也限制着该理论的普适性. ...
... 异速生长理论预测小个体植物叶、茎、根器官生长可能为等速关系(
Enquist & Niklas, 2002 ;
Weiner, 2004 ;
Niklas, 2006 ), 这种基于种间变异建立的跨物种关系已在种内变异的研究中得到了较多应用(
Gargaglione et al ., 2010 ;
Cheng et al ., 2014 ;
陈国鹏等, 2016 ;
王杨等, 2017 ), 如
Cheng等(2014) 发现南亚热带生长的5种苗木叶、茎和根生物量的积累为等速或近似等速生长关系.本研究中, 7种高寒杜鹃叶与茎、叶与根以及茎与根之间既有异速生长表现, 也有等速生长表现(
图2 -5), 与荒漠植物(
杨昊天等, 2013 ;
陶冶和张元明, 2014 )、干热河谷草本植物(
闫帮国等, 2016 )的异速生长关系具有相似性, 不同于川西高寒草甸优势植物(
左有璐等, 2018 ), 并不完全支持小个体器官间等速生长关系的预测, 也不完全支持生态代谢理论的预测, 这可能是由于植物在适应特殊环境的过程中, 不同物种间具有很大的功能群差异, 这些功能群差异可能是不同器官生物量间异速生长关系分化的主要因素(
Poorter et al ., 2015 ;
闫帮国等, 2016 ), 而基于跨物种建立的生态代谢理论、小个体等速生长理论忽略了物种间的功能群差异. ...
... ), 如
Cheng等(2014) 发现南亚热带生长的5种苗木叶、茎和根生物量的积累为等速或近似等速生长关系.本研究中, 7种高寒杜鹃叶与茎、叶与根以及茎与根之间既有异速生长表现, 也有等速生长表现(
图2 -5), 与荒漠植物(
杨昊天等, 2013 ;
陶冶和张元明, 2014 )、干热河谷草本植物(
闫帮国等, 2016 )的异速生长关系具有相似性, 不同于川西高寒草甸优势植物(
左有璐等, 2018 ), 并不完全支持小个体器官间等速生长关系的预测, 也不完全支持生态代谢理论的预测, 这可能是由于植物在适应特殊环境的过程中, 不同物种间具有很大的功能群差异, 这些功能群差异可能是不同器官生物量间异速生长关系分化的主要因素(
Poorter et al ., 2015 ;
闫帮国等, 2016 ), 而基于跨物种建立的生态代谢理论、小个体等速生长理论忽略了物种间的功能群差异. ...
Above- and below-ground biomass relationships across 1534 forested communities
1
2007
... 异速生长理论认为, 生物量的分配主要由植物的大小决定, 并可用幂函数
Y 1 =
βY 2 α 表示, 该方程经过对数转换后, 可线性化为log(
Y 1 ) = log(
β )
+ α log(
Y 2 ), 其中
Y 1 和
Y 2 是相互依赖的变量(如茎生物量和根生物量), log(
β )是异速生长常数,
α 是异速生长指数(
Niklas & Enquist, 2002a ,
2002b ).当
α = 1时, 模型反映的是等速生长关系; 当
α ≠ 1时, 模型反映的是异速生长关系(
Niklas & Enquist, 2002b ).生态代谢理论认为, 全球尺度上植物个体生物量具有恒定的异速生长指数, 其地上、地下生物量间均表现为异速生长关系, 叶生物量与茎生物量或地下生物量间的异速生长指数为0.75 (
West et al ., 1997 ;
Enquist & Niklas, 2002 ;
Niklas & Enquist, 2002 ), 但后续研究又证明小个体植物在叶生物量、茎生物量和地下生物量间也可能为等速关系(
Niklas, 2005 ;
Cheng et al ., 2014 ;
陈国鹏和赵文智, 2016 ),
Poorter等(2015) 进一步分析个体大小影响时发现, 植物叶与茎、叶与根、茎与根等生物量间的异速生长指数随个体增大而减小.尽管基于生态代谢理论的异速生长指数在不同尺度上得到了检验(
Cheng & Niklas, 2007 ), 但是该理论以关注种间的变异为主, 跨物种的恒定关系不一定适用于种内, 对物种作用的考虑不足(
Müller et al ., 2000 ;
Cheng et al ., 2014 ), 尤其对一些极端生境的物种关注较少也限制着该理论的普适性. ...
Evolutionarily stable strategy carbon allocation to foliage, wood, and fine roots in trees competing for light and nitrogen: an analytically tractable, individual-based model and quantitative comparisons to data
1
2011
... 树木对温度适应性策略的假设认为, 在寒冷的气候环境下, 根的分配比例较大、叶的分配比例较小(
Reich et al ., 2014 ), 本研究中, 7种杜鹃根生物量分配比例高达35.57%, 个别物种甚至超过40% (
图1 ), 高于全球绝大多数植物(
Poorter et al ., 2015 ), 也高于我国亚热带山地杜鹃灌丛根系生物量的分配比例(
张蔷等, 2017 ), 与一些高寒植被(
高巧等, 2014 ;
钟泽兵等, 2014 ;
杨路存等, 2018 ;
左有璐等, 2018 )相似, 低速率的养分循环和受限的土壤溶液运动驱动着高山、亚高山地区的低营养供应(
魏晶等, 2004 ;
Luo et al ., 2012 ;
Reich et al ., 2014 ), 这使得高寒杜鹃需提高根生物量分配比例以增强从寒冷土壤中吸收养分, 生物量向根系中的倾向分配支持最优分配假说.7种杜鹃叶生物量分配比例低于全球绝大多数植物(
Poorter et al ., 2015 ), 其适应机制也与一些高寒植物(
Reich et al ., 2014 )相似, 如
钟泽兵等(2014) 对青海玉树海拔3 715-4 296 m的5种建群灌木的研究发现叶生物量分配比例不足10%.甘肃南部高山-亚高山区的杜鹃会通过减少资源向叶片的分配, 保持常绿, 以较低的叶片周转率来适应寒冷环境.随着植株叶投资盈利能力的下降, 叶生物量将饱和, 投资会逐渐转向茎和根(
Poorter et al ., 2015 ), 而为增强植株的机械支撑, 必须增加茎的生物量投资(
Dybzinski et al ., 2011 ).本研究中7种高寒杜鹃茎生物量分配平均比例为45.61%, 接近于全球数据库平均值(
Poorter et al ., 2015 ).同时, 相比于叶和根, 茎生物量分配比例的种间差异最大(可达21%), 且多组物种间的差异显著(
表2 ), 这与
Poorter等(2015) 对全球1 200种植物整合分析的结果具有一致性, 茎不仅是植株的支撑、运输体系, 还在光竞争时向四周伸展小枝扩展空域, 而小枝的扩张又带来了高寒区防风的压力, 功能的多样性可能导致茎生物量分配的可塑性.各物种分配的差异还可能与茎构型有关, 如黄毛杜鹃和陇蜀杜鹃多为丛生, 茎枝扭曲, 其茎生物量分配比例的变异范围较大; 而美容杜鹃和山光杜鹃主干通直, 其茎生物量分配比例的变幅较小(
图1 ). ...
Global allocation rules for patterns of biomass partitioning in seed plants
3
2002
... 生物量在不同器官间的分配是植物生态学研究的核心问题(
West et al .,1999 ;
Enquist & Niklas, 2002 ;
Poorter et al ., 2012 ).生物量分配模式不仅有助于从易测量的地上生物量估算地下生物量, 还体现着植物不同的生态策略(
Weiner, 2004 ), 反映出光合产物对植物不同器官(地上不同器官之间, 地上器官和地下器官之间)的投资以适应环境条件(
Niklas, 2006 ;
Poorter & Sack, 2012 ;
闫帮国等, 2016 ).最优分配假说认为, 植物倾向于将资源优先分配到能获得限制性资源的器官, 以帮助植物获取更多的受限资源, 从而维持最大生长速率(
Bloom et al ., 1985 ;
McCarthy & Enquist, 2007 ), 这意味着, 根据植物生长的环境, 如果光照受限, 生物量将倾向于叶和枝分配; 如果营养或水分受限, 生物量将倾向于根系分配(
Gleeson & Tilman, 1990 ).但植物生物量分配模式存在个体发育漂变(
Poorter & Sack, 2012 ;
Poorter et al ., 2015 ;
王杨等, 2017 ), 其分配比例会随植株的大小变化而变化(
Poorter et al ., 2015 ;
陈国鹏等, 2016 ), 使得最优分配假说的使用受到了限制, 而异速生长理论能较好地解决这一问题(
Weiner, 2004 ;
Gargaglione et al ., 2010 ), 且已被越来越多地应用到生物量分配研究中, 有****也曾对两种方法的优劣进行了比较(
McCarthy & Enquist, 2007 ;
Poorter & Sack, 2012 ), 更赞同将两种方法结合应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ). ...
... 异速生长理论认为, 生物量的分配主要由植物的大小决定, 并可用幂函数
Y 1 =
βY 2 α 表示, 该方程经过对数转换后, 可线性化为log(
Y 1 ) = log(
β )
+ α log(
Y 2 ), 其中
Y 1 和
Y 2 是相互依赖的变量(如茎生物量和根生物量), log(
β )是异速生长常数,
α 是异速生长指数(
Niklas & Enquist, 2002a ,
2002b ).当
α = 1时, 模型反映的是等速生长关系; 当
α ≠ 1时, 模型反映的是异速生长关系(
Niklas & Enquist, 2002b ).生态代谢理论认为, 全球尺度上植物个体生物量具有恒定的异速生长指数, 其地上、地下生物量间均表现为异速生长关系, 叶生物量与茎生物量或地下生物量间的异速生长指数为0.75 (
West et al ., 1997 ;
Enquist & Niklas, 2002 ;
Niklas & Enquist, 2002 ), 但后续研究又证明小个体植物在叶生物量、茎生物量和地下生物量间也可能为等速关系(
Niklas, 2005 ;
Cheng et al ., 2014 ;
陈国鹏和赵文智, 2016 ),
Poorter等(2015) 进一步分析个体大小影响时发现, 植物叶与茎、叶与根、茎与根等生物量间的异速生长指数随个体增大而减小.尽管基于生态代谢理论的异速生长指数在不同尺度上得到了检验(
Cheng & Niklas, 2007 ), 但是该理论以关注种间的变异为主, 跨物种的恒定关系不一定适用于种内, 对物种作用的考虑不足(
Müller et al ., 2000 ;
Cheng et al ., 2014 ), 尤其对一些极端生境的物种关注较少也限制着该理论的普适性. ...
... 异速生长理论预测小个体植物叶、茎、根器官生长可能为等速关系(
Enquist & Niklas, 2002 ;
Weiner, 2004 ;
Niklas, 2006 ), 这种基于种间变异建立的跨物种关系已在种内变异的研究中得到了较多应用(
Gargaglione et al ., 2010 ;
Cheng et al ., 2014 ;
陈国鹏等, 2016 ;
王杨等, 2017 ), 如
Cheng等(2014) 发现南亚热带生长的5种苗木叶、茎和根生物量的积累为等速或近似等速生长关系.本研究中, 7种高寒杜鹃叶与茎、叶与根以及茎与根之间既有异速生长表现, 也有等速生长表现(
图2 -5), 与荒漠植物(
杨昊天等, 2013 ;
陶冶和张元明, 2014 )、干热河谷草本植物(
闫帮国等, 2016 )的异速生长关系具有相似性, 不同于川西高寒草甸优势植物(
左有璐等, 2018 ), 并不完全支持小个体器官间等速生长关系的预测, 也不完全支持生态代谢理论的预测, 这可能是由于植物在适应特殊环境的过程中, 不同物种间具有很大的功能群差异, 这些功能群差异可能是不同器官生物量间异速生长关系分化的主要因素(
Poorter et al ., 2015 ;
闫帮国等, 2016 ), 而基于跨物种建立的生态代谢理论、小个体等速生长理论忽略了物种间的功能群差异. ...
四川省甘孜藏族自治州高寒矮灌丛生物量分配及其碳密度的估算
2
2014
... 理论上来说, 极端环境通过长期的筛选作用, 其适生物种响应资源约束时的生物量权衡策略应具有相似性(
Reich et al ., 2014 ).如, 在高寒山地, 高海拔、寒冷、短生长季和低气压等环境约束可能使植物的资源权衡具有独特表现(
高巧等, 2014 ;
左有璐等, 2014 ;
刘尊驰等, 2015 ), 植物会增加根生物量、降低叶生物量比例来适应较冷的环境(
徐波等, 2013 ).但目前的相关研究材料以常绿裸子植物或落叶被子植物为主(
Reich et al ., 2014 ;
Poorter et al ., 2015 ), 对高寒区生长的常绿被子植物的生物量权衡机制缺乏认知, 而常绿植物与落叶植物叶生物量分配策略存在差别, 常绿植物在叶片上投入的较多, 因此叶生物量分配比例高于落叶植物(
Poorter et al ., 2015 ). ...
... 树木对温度适应性策略的假设认为, 在寒冷的气候环境下, 根的分配比例较大、叶的分配比例较小(
Reich et al ., 2014 ), 本研究中, 7种杜鹃根生物量分配比例高达35.57%, 个别物种甚至超过40% (
图1 ), 高于全球绝大多数植物(
Poorter et al ., 2015 ), 也高于我国亚热带山地杜鹃灌丛根系生物量的分配比例(
张蔷等, 2017 ), 与一些高寒植被(
高巧等, 2014 ;
钟泽兵等, 2014 ;
杨路存等, 2018 ;
左有璐等, 2018 )相似, 低速率的养分循环和受限的土壤溶液运动驱动着高山、亚高山地区的低营养供应(
魏晶等, 2004 ;
Luo et al ., 2012 ;
Reich et al ., 2014 ), 这使得高寒杜鹃需提高根生物量分配比例以增强从寒冷土壤中吸收养分, 生物量向根系中的倾向分配支持最优分配假说.7种杜鹃叶生物量分配比例低于全球绝大多数植物(
Poorter et al ., 2015 ), 其适应机制也与一些高寒植物(
Reich et al ., 2014 )相似, 如
钟泽兵等(2014) 对青海玉树海拔3 715-4 296 m的5种建群灌木的研究发现叶生物量分配比例不足10%.甘肃南部高山-亚高山区的杜鹃会通过减少资源向叶片的分配, 保持常绿, 以较低的叶片周转率来适应寒冷环境.随着植株叶投资盈利能力的下降, 叶生物量将饱和, 投资会逐渐转向茎和根(
Poorter et al ., 2015 ), 而为增强植株的机械支撑, 必须增加茎的生物量投资(
Dybzinski et al ., 2011 ).本研究中7种高寒杜鹃茎生物量分配平均比例为45.61%, 接近于全球数据库平均值(
Poorter et al ., 2015 ).同时, 相比于叶和根, 茎生物量分配比例的种间差异最大(可达21%), 且多组物种间的差异显著(
表2 ), 这与
Poorter等(2015) 对全球1 200种植物整合分析的结果具有一致性, 茎不仅是植株的支撑、运输体系, 还在光竞争时向四周伸展小枝扩展空域, 而小枝的扩张又带来了高寒区防风的压力, 功能的多样性可能导致茎生物量分配的可塑性.各物种分配的差异还可能与茎构型有关, 如黄毛杜鹃和陇蜀杜鹃多为丛生, 茎枝扭曲, 其茎生物量分配比例的变异范围较大; 而美容杜鹃和山光杜鹃主干通直, 其茎生物量分配比例的变幅较小(
图1 ). ...
四川省甘孜藏族自治州高寒矮灌丛生物量分配及其碳密度的估算
2
2014
... 理论上来说, 极端环境通过长期的筛选作用, 其适生物种响应资源约束时的生物量权衡策略应具有相似性(
Reich et al ., 2014 ).如, 在高寒山地, 高海拔、寒冷、短生长季和低气压等环境约束可能使植物的资源权衡具有独特表现(
高巧等, 2014 ;
左有璐等, 2014 ;
刘尊驰等, 2015 ), 植物会增加根生物量、降低叶生物量比例来适应较冷的环境(
徐波等, 2013 ).但目前的相关研究材料以常绿裸子植物或落叶被子植物为主(
Reich et al ., 2014 ;
Poorter et al ., 2015 ), 对高寒区生长的常绿被子植物的生物量权衡机制缺乏认知, 而常绿植物与落叶植物叶生物量分配策略存在差别, 常绿植物在叶片上投入的较多, 因此叶生物量分配比例高于落叶植物(
Poorter et al ., 2015 ). ...
... 树木对温度适应性策略的假设认为, 在寒冷的气候环境下, 根的分配比例较大、叶的分配比例较小(
Reich et al ., 2014 ), 本研究中, 7种杜鹃根生物量分配比例高达35.57%, 个别物种甚至超过40% (
图1 ), 高于全球绝大多数植物(
Poorter et al ., 2015 ), 也高于我国亚热带山地杜鹃灌丛根系生物量的分配比例(
张蔷等, 2017 ), 与一些高寒植被(
高巧等, 2014 ;
钟泽兵等, 2014 ;
杨路存等, 2018 ;
左有璐等, 2018 )相似, 低速率的养分循环和受限的土壤溶液运动驱动着高山、亚高山地区的低营养供应(
魏晶等, 2004 ;
Luo et al ., 2012 ;
Reich et al ., 2014 ), 这使得高寒杜鹃需提高根生物量分配比例以增强从寒冷土壤中吸收养分, 生物量向根系中的倾向分配支持最优分配假说.7种杜鹃叶生物量分配比例低于全球绝大多数植物(
Poorter et al ., 2015 ), 其适应机制也与一些高寒植物(
Reich et al ., 2014 )相似, 如
钟泽兵等(2014) 对青海玉树海拔3 715-4 296 m的5种建群灌木的研究发现叶生物量分配比例不足10%.甘肃南部高山-亚高山区的杜鹃会通过减少资源向叶片的分配, 保持常绿, 以较低的叶片周转率来适应寒冷环境.随着植株叶投资盈利能力的下降, 叶生物量将饱和, 投资会逐渐转向茎和根(
Poorter et al ., 2015 ), 而为增强植株的机械支撑, 必须增加茎的生物量投资(
Dybzinski et al ., 2011 ).本研究中7种高寒杜鹃茎生物量分配平均比例为45.61%, 接近于全球数据库平均值(
Poorter et al ., 2015 ).同时, 相比于叶和根, 茎生物量分配比例的种间差异最大(可达21%), 且多组物种间的差异显著(
表2 ), 这与
Poorter等(2015) 对全球1 200种植物整合分析的结果具有一致性, 茎不仅是植株的支撑、运输体系, 还在光竞争时向四周伸展小枝扩展空域, 而小枝的扩张又带来了高寒区防风的压力, 功能的多样性可能导致茎生物量分配的可塑性.各物种分配的差异还可能与茎构型有关, 如黄毛杜鹃和陇蜀杜鹃多为丛生, 茎枝扭曲, 其茎生物量分配比例的变异范围较大; 而美容杜鹃和山光杜鹃主干通直, 其茎生物量分配比例的变幅较小(
图1 ). ...
Allometric relations for biomass partitioning of
Nothofagus antarctica trees of different crown classes over a site quality gradient
2
2010
... 生物量在不同器官间的分配是植物生态学研究的核心问题(
West et al .,1999 ;
Enquist & Niklas, 2002 ;
Poorter et al ., 2012 ).生物量分配模式不仅有助于从易测量的地上生物量估算地下生物量, 还体现着植物不同的生态策略(
Weiner, 2004 ), 反映出光合产物对植物不同器官(地上不同器官之间, 地上器官和地下器官之间)的投资以适应环境条件(
Niklas, 2006 ;
Poorter & Sack, 2012 ;
闫帮国等, 2016 ).最优分配假说认为, 植物倾向于将资源优先分配到能获得限制性资源的器官, 以帮助植物获取更多的受限资源, 从而维持最大生长速率(
Bloom et al ., 1985 ;
McCarthy & Enquist, 2007 ), 这意味着, 根据植物生长的环境, 如果光照受限, 生物量将倾向于叶和枝分配; 如果营养或水分受限, 生物量将倾向于根系分配(
Gleeson & Tilman, 1990 ).但植物生物量分配模式存在个体发育漂变(
Poorter & Sack, 2012 ;
Poorter et al ., 2015 ;
王杨等, 2017 ), 其分配比例会随植株的大小变化而变化(
Poorter et al ., 2015 ;
陈国鹏等, 2016 ), 使得最优分配假说的使用受到了限制, 而异速生长理论能较好地解决这一问题(
Weiner, 2004 ;
Gargaglione et al ., 2010 ), 且已被越来越多地应用到生物量分配研究中, 有****也曾对两种方法的优劣进行了比较(
McCarthy & Enquist, 2007 ;
Poorter & Sack, 2012 ), 更赞同将两种方法结合应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ). ...
... 异速生长理论预测小个体植物叶、茎、根器官生长可能为等速关系(
Enquist & Niklas, 2002 ;
Weiner, 2004 ;
Niklas, 2006 ), 这种基于种间变异建立的跨物种关系已在种内变异的研究中得到了较多应用(
Gargaglione et al ., 2010 ;
Cheng et al ., 2014 ;
陈国鹏等, 2016 ;
王杨等, 2017 ), 如
Cheng等(2014) 发现南亚热带生长的5种苗木叶、茎和根生物量的积累为等速或近似等速生长关系.本研究中, 7种高寒杜鹃叶与茎、叶与根以及茎与根之间既有异速生长表现, 也有等速生长表现(
图2 -5), 与荒漠植物(
杨昊天等, 2013 ;
陶冶和张元明, 2014 )、干热河谷草本植物(
闫帮国等, 2016 )的异速生长关系具有相似性, 不同于川西高寒草甸优势植物(
左有璐等, 2018 ), 并不完全支持小个体器官间等速生长关系的预测, 也不完全支持生态代谢理论的预测, 这可能是由于植物在适应特殊环境的过程中, 不同物种间具有很大的功能群差异, 这些功能群差异可能是不同器官生物量间异速生长关系分化的主要因素(
Poorter et al ., 2015 ;
闫帮国等, 2016 ), 而基于跨物种建立的生态代谢理论、小个体等速生长理论忽略了物种间的功能群差异. ...
Allocation and the transient dynamics of succession on poor soils
1
1990
... 生物量在不同器官间的分配是植物生态学研究的核心问题(
West et al .,1999 ;
Enquist & Niklas, 2002 ;
Poorter et al ., 2012 ).生物量分配模式不仅有助于从易测量的地上生物量估算地下生物量, 还体现着植物不同的生态策略(
Weiner, 2004 ), 反映出光合产物对植物不同器官(地上不同器官之间, 地上器官和地下器官之间)的投资以适应环境条件(
Niklas, 2006 ;
Poorter & Sack, 2012 ;
闫帮国等, 2016 ).最优分配假说认为, 植物倾向于将资源优先分配到能获得限制性资源的器官, 以帮助植物获取更多的受限资源, 从而维持最大生长速率(
Bloom et al ., 1985 ;
McCarthy & Enquist, 2007 ), 这意味着, 根据植物生长的环境, 如果光照受限, 生物量将倾向于叶和枝分配; 如果营养或水分受限, 生物量将倾向于根系分配(
Gleeson & Tilman, 1990 ).但植物生物量分配模式存在个体发育漂变(
Poorter & Sack, 2012 ;
Poorter et al ., 2015 ;
王杨等, 2017 ), 其分配比例会随植株的大小变化而变化(
Poorter et al ., 2015 ;
陈国鹏等, 2016 ), 使得最优分配假说的使用受到了限制, 而异速生长理论能较好地解决这一问题(
Weiner, 2004 ;
Gargaglione et al ., 2010 ), 且已被越来越多地应用到生物量分配研究中, 有****也曾对两种方法的优劣进行了比较(
McCarthy & Enquist, 2007 ;
Poorter & Sack, 2012 ), 更赞同将两种方法结合应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ). ...
紫翅猪毛菜、钠猪毛菜不同个体大小繁殖分配差异及随海拔的变化
1
2015
... 理论上来说, 极端环境通过长期的筛选作用, 其适生物种响应资源约束时的生物量权衡策略应具有相似性(
Reich et al ., 2014 ).如, 在高寒山地, 高海拔、寒冷、短生长季和低气压等环境约束可能使植物的资源权衡具有独特表现(
高巧等, 2014 ;
左有璐等, 2014 ;
刘尊驰等, 2015 ), 植物会增加根生物量、降低叶生物量比例来适应较冷的环境(
徐波等, 2013 ).但目前的相关研究材料以常绿裸子植物或落叶被子植物为主(
Reich et al ., 2014 ;
Poorter et al ., 2015 ), 对高寒区生长的常绿被子植物的生物量权衡机制缺乏认知, 而常绿植物与落叶植物叶生物量分配策略存在差别, 常绿植物在叶片上投入的较多, 因此叶生物量分配比例高于落叶植物(
Poorter et al ., 2015 ). ...
紫翅猪毛菜、钠猪毛菜不同个体大小繁殖分配差异及随海拔的变化
1
2015
... 理论上来说, 极端环境通过长期的筛选作用, 其适生物种响应资源约束时的生物量权衡策略应具有相似性(
Reich et al ., 2014 ).如, 在高寒山地, 高海拔、寒冷、短生长季和低气压等环境约束可能使植物的资源权衡具有独特表现(
高巧等, 2014 ;
左有璐等, 2014 ;
刘尊驰等, 2015 ), 植物会增加根生物量、降低叶生物量比例来适应较冷的环境(
徐波等, 2013 ).但目前的相关研究材料以常绿裸子植物或落叶被子植物为主(
Reich et al ., 2014 ;
Poorter et al ., 2015 ), 对高寒区生长的常绿被子植物的生物量权衡机制缺乏认知, 而常绿植物与落叶植物叶生物量分配策略存在差别, 常绿植物在叶片上投入的较多, 因此叶生物量分配比例高于落叶植物(
Poorter et al ., 2015 ). ...
Root: shoot ratios across China’s forests: forest type and climatic effects
1
2012
... 树木对温度适应性策略的假设认为, 在寒冷的气候环境下, 根的分配比例较大、叶的分配比例较小(
Reich et al ., 2014 ), 本研究中, 7种杜鹃根生物量分配比例高达35.57%, 个别物种甚至超过40% (
图1 ), 高于全球绝大多数植物(
Poorter et al ., 2015 ), 也高于我国亚热带山地杜鹃灌丛根系生物量的分配比例(
张蔷等, 2017 ), 与一些高寒植被(
高巧等, 2014 ;
钟泽兵等, 2014 ;
杨路存等, 2018 ;
左有璐等, 2018 )相似, 低速率的养分循环和受限的土壤溶液运动驱动着高山、亚高山地区的低营养供应(
魏晶等, 2004 ;
Luo et al ., 2012 ;
Reich et al ., 2014 ), 这使得高寒杜鹃需提高根生物量分配比例以增强从寒冷土壤中吸收养分, 生物量向根系中的倾向分配支持最优分配假说.7种杜鹃叶生物量分配比例低于全球绝大多数植物(
Poorter et al ., 2015 ), 其适应机制也与一些高寒植物(
Reich et al ., 2014 )相似, 如
钟泽兵等(2014) 对青海玉树海拔3 715-4 296 m的5种建群灌木的研究发现叶生物量分配比例不足10%.甘肃南部高山-亚高山区的杜鹃会通过减少资源向叶片的分配, 保持常绿, 以较低的叶片周转率来适应寒冷环境.随着植株叶投资盈利能力的下降, 叶生物量将饱和, 投资会逐渐转向茎和根(
Poorter et al ., 2015 ), 而为增强植株的机械支撑, 必须增加茎的生物量投资(
Dybzinski et al ., 2011 ).本研究中7种高寒杜鹃茎生物量分配平均比例为45.61%, 接近于全球数据库平均值(
Poorter et al ., 2015 ).同时, 相比于叶和根, 茎生物量分配比例的种间差异最大(可达21%), 且多组物种间的差异显著(
表2 ), 这与
Poorter等(2015) 对全球1 200种植物整合分析的结果具有一致性, 茎不仅是植株的支撑、运输体系, 还在光竞争时向四周伸展小枝扩展空域, 而小枝的扩张又带来了高寒区防风的压力, 功能的多样性可能导致茎生物量分配的可塑性.各物种分配的差异还可能与茎构型有关, 如黄毛杜鹃和陇蜀杜鹃多为丛生, 茎枝扭曲, 其茎生物量分配比例的变异范围较大; 而美容杜鹃和山光杜鹃主干通直, 其茎生物量分配比例的变幅较小(
图1 ). ...
Consistency between an allometric approach and optimal partitioning theory in global patterns of plant biomass allocation
2
2007
... 生物量在不同器官间的分配是植物生态学研究的核心问题(
West et al .,1999 ;
Enquist & Niklas, 2002 ;
Poorter et al ., 2012 ).生物量分配模式不仅有助于从易测量的地上生物量估算地下生物量, 还体现着植物不同的生态策略(
Weiner, 2004 ), 反映出光合产物对植物不同器官(地上不同器官之间, 地上器官和地下器官之间)的投资以适应环境条件(
Niklas, 2006 ;
Poorter & Sack, 2012 ;
闫帮国等, 2016 ).最优分配假说认为, 植物倾向于将资源优先分配到能获得限制性资源的器官, 以帮助植物获取更多的受限资源, 从而维持最大生长速率(
Bloom et al ., 1985 ;
McCarthy & Enquist, 2007 ), 这意味着, 根据植物生长的环境, 如果光照受限, 生物量将倾向于叶和枝分配; 如果营养或水分受限, 生物量将倾向于根系分配(
Gleeson & Tilman, 1990 ).但植物生物量分配模式存在个体发育漂变(
Poorter & Sack, 2012 ;
Poorter et al ., 2015 ;
王杨等, 2017 ), 其分配比例会随植株的大小变化而变化(
Poorter et al ., 2015 ;
陈国鹏等, 2016 ), 使得最优分配假说的使用受到了限制, 而异速生长理论能较好地解决这一问题(
Weiner, 2004 ;
Gargaglione et al ., 2010 ), 且已被越来越多地应用到生物量分配研究中, 有****也曾对两种方法的优劣进行了比较(
McCarthy & Enquist, 2007 ;
Poorter & Sack, 2012 ), 更赞同将两种方法结合应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ). ...
... ), 且已被越来越多地应用到生物量分配研究中, 有****也曾对两种方法的优劣进行了比较(
McCarthy & Enquist, 2007 ;
Poorter & Sack, 2012 ), 更赞同将两种方法结合应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ). ...
The effect of nutrient availability on biomass allocation patterns in 27 species of herbaceous plants
1
2000
... 异速生长理论认为, 生物量的分配主要由植物的大小决定, 并可用幂函数
Y 1 =
βY 2 α 表示, 该方程经过对数转换后, 可线性化为log(
Y 1 ) = log(
β )
+ α log(
Y 2 ), 其中
Y 1 和
Y 2 是相互依赖的变量(如茎生物量和根生物量), log(
β )是异速生长常数,
α 是异速生长指数(
Niklas & Enquist, 2002a ,
2002b ).当
α = 1时, 模型反映的是等速生长关系; 当
α ≠ 1时, 模型反映的是异速生长关系(
Niklas & Enquist, 2002b ).生态代谢理论认为, 全球尺度上植物个体生物量具有恒定的异速生长指数, 其地上、地下生物量间均表现为异速生长关系, 叶生物量与茎生物量或地下生物量间的异速生长指数为0.75 (
West et al ., 1997 ;
Enquist & Niklas, 2002 ;
Niklas & Enquist, 2002 ), 但后续研究又证明小个体植物在叶生物量、茎生物量和地下生物量间也可能为等速关系(
Niklas, 2005 ;
Cheng et al ., 2014 ;
陈国鹏和赵文智, 2016 ),
Poorter等(2015) 进一步分析个体大小影响时发现, 植物叶与茎、叶与根、茎与根等生物量间的异速生长指数随个体增大而减小.尽管基于生态代谢理论的异速生长指数在不同尺度上得到了检验(
Cheng & Niklas, 2007 ), 但是该理论以关注种间的变异为主, 跨物种的恒定关系不一定适用于种内, 对物种作用的考虑不足(
Müller et al ., 2000 ;
Cheng et al ., 2014 ), 尤其对一些极端生境的物种关注较少也限制着该理论的普适性. ...
Modelling below- and above-ground biomass for non-woody and woody plants
1
2005
... 异速生长理论认为, 生物量的分配主要由植物的大小决定, 并可用幂函数
Y 1 =
βY 2 α 表示, 该方程经过对数转换后, 可线性化为log(
Y 1 ) = log(
β )
+ α log(
Y 2 ), 其中
Y 1 和
Y 2 是相互依赖的变量(如茎生物量和根生物量), log(
β )是异速生长常数,
α 是异速生长指数(
Niklas & Enquist, 2002a ,
2002b ).当
α = 1时, 模型反映的是等速生长关系; 当
α ≠ 1时, 模型反映的是异速生长关系(
Niklas & Enquist, 2002b ).生态代谢理论认为, 全球尺度上植物个体生物量具有恒定的异速生长指数, 其地上、地下生物量间均表现为异速生长关系, 叶生物量与茎生物量或地下生物量间的异速生长指数为0.75 (
West et al ., 1997 ;
Enquist & Niklas, 2002 ;
Niklas & Enquist, 2002 ), 但后续研究又证明小个体植物在叶生物量、茎生物量和地下生物量间也可能为等速关系(
Niklas, 2005 ;
Cheng et al ., 2014 ;
陈国鹏和赵文智, 2016 ),
Poorter等(2015) 进一步分析个体大小影响时发现, 植物叶与茎、叶与根、茎与根等生物量间的异速生长指数随个体增大而减小.尽管基于生态代谢理论的异速生长指数在不同尺度上得到了检验(
Cheng & Niklas, 2007 ), 但是该理论以关注种间的变异为主, 跨物种的恒定关系不一定适用于种内, 对物种作用的考虑不足(
Müller et al ., 2000 ;
Cheng et al ., 2014 ), 尤其对一些极端生境的物种关注较少也限制着该理论的普适性. ...
A phyletic perspective on the allometry of plant biomass-partitioning patterns and functionally equivalent organ-categories
2
2006
... 生物量在不同器官间的分配是植物生态学研究的核心问题(
West et al .,1999 ;
Enquist & Niklas, 2002 ;
Poorter et al ., 2012 ).生物量分配模式不仅有助于从易测量的地上生物量估算地下生物量, 还体现着植物不同的生态策略(
Weiner, 2004 ), 反映出光合产物对植物不同器官(地上不同器官之间, 地上器官和地下器官之间)的投资以适应环境条件(
Niklas, 2006 ;
Poorter & Sack, 2012 ;
闫帮国等, 2016 ).最优分配假说认为, 植物倾向于将资源优先分配到能获得限制性资源的器官, 以帮助植物获取更多的受限资源, 从而维持最大生长速率(
Bloom et al ., 1985 ;
McCarthy & Enquist, 2007 ), 这意味着, 根据植物生长的环境, 如果光照受限, 生物量将倾向于叶和枝分配; 如果营养或水分受限, 生物量将倾向于根系分配(
Gleeson & Tilman, 1990 ).但植物生物量分配模式存在个体发育漂变(
Poorter & Sack, 2012 ;
Poorter et al ., 2015 ;
王杨等, 2017 ), 其分配比例会随植株的大小变化而变化(
Poorter et al ., 2015 ;
陈国鹏等, 2016 ), 使得最优分配假说的使用受到了限制, 而异速生长理论能较好地解决这一问题(
Weiner, 2004 ;
Gargaglione et al ., 2010 ), 且已被越来越多地应用到生物量分配研究中, 有****也曾对两种方法的优劣进行了比较(
McCarthy & Enquist, 2007 ;
Poorter & Sack, 2012 ), 更赞同将两种方法结合应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ). ...
... 异速生长理论预测小个体植物叶、茎、根器官生长可能为等速关系(
Enquist & Niklas, 2002 ;
Weiner, 2004 ;
Niklas, 2006 ), 这种基于种间变异建立的跨物种关系已在种内变异的研究中得到了较多应用(
Gargaglione et al ., 2010 ;
Cheng et al ., 2014 ;
陈国鹏等, 2016 ;
王杨等, 2017 ), 如
Cheng等(2014) 发现南亚热带生长的5种苗木叶、茎和根生物量的积累为等速或近似等速生长关系.本研究中, 7种高寒杜鹃叶与茎、叶与根以及茎与根之间既有异速生长表现, 也有等速生长表现(
图2 -5), 与荒漠植物(
杨昊天等, 2013 ;
陶冶和张元明, 2014 )、干热河谷草本植物(
闫帮国等, 2016 )的异速生长关系具有相似性, 不同于川西高寒草甸优势植物(
左有璐等, 2018 ), 并不完全支持小个体器官间等速生长关系的预测, 也不完全支持生态代谢理论的预测, 这可能是由于植物在适应特殊环境的过程中, 不同物种间具有很大的功能群差异, 这些功能群差异可能是不同器官生物量间异速生长关系分化的主要因素(
Poorter et al ., 2015 ;
闫帮国等, 2016 ), 而基于跨物种建立的生态代谢理论、小个体等速生长理论忽略了物种间的功能群差异. ...
Canonical rules for plant organ biomass partitioning and annual allocation
1
2002
... 异速生长理论认为, 生物量的分配主要由植物的大小决定, 并可用幂函数
Y 1 =
βY 2 α 表示, 该方程经过对数转换后, 可线性化为log(
Y 1 ) = log(
β )
+ α log(
Y 2 ), 其中
Y 1 和
Y 2 是相互依赖的变量(如茎生物量和根生物量), log(
β )是异速生长常数,
α 是异速生长指数(
Niklas & Enquist, 2002a ,
2002b ).当
α = 1时, 模型反映的是等速生长关系; 当
α ≠ 1时, 模型反映的是异速生长关系(
Niklas & Enquist, 2002b ).生态代谢理论认为, 全球尺度上植物个体生物量具有恒定的异速生长指数, 其地上、地下生物量间均表现为异速生长关系, 叶生物量与茎生物量或地下生物量间的异速生长指数为0.75 (
West et al ., 1997 ;
Enquist & Niklas, 2002 ;
Niklas & Enquist, 2002 ), 但后续研究又证明小个体植物在叶生物量、茎生物量和地下生物量间也可能为等速关系(
Niklas, 2005 ;
Cheng et al ., 2014 ;
陈国鹏和赵文智, 2016 ),
Poorter等(2015) 进一步分析个体大小影响时发现, 植物叶与茎、叶与根、茎与根等生物量间的异速生长指数随个体增大而减小.尽管基于生态代谢理论的异速生长指数在不同尺度上得到了检验(
Cheng & Niklas, 2007 ), 但是该理论以关注种间的变异为主, 跨物种的恒定关系不一定适用于种内, 对物种作用的考虑不足(
Müller et al ., 2000 ;
Cheng et al ., 2014 ), 尤其对一些极端生境的物种关注较少也限制着该理论的普适性. ...
On the vegetative biomass partitioning of seed plant leaves, stems, and roots
3
2002
... 异速生长理论认为, 生物量的分配主要由植物的大小决定, 并可用幂函数
Y 1 =
βY 2 α 表示, 该方程经过对数转换后, 可线性化为log(
Y 1 ) = log(
β )
+ α log(
Y 2 ), 其中
Y 1 和
Y 2 是相互依赖的变量(如茎生物量和根生物量), log(
β )是异速生长常数,
α 是异速生长指数(
Niklas & Enquist, 2002a ,
2002b ).当
α = 1时, 模型反映的是等速生长关系; 当
α ≠ 1时, 模型反映的是异速生长关系(
Niklas & Enquist, 2002b ).生态代谢理论认为, 全球尺度上植物个体生物量具有恒定的异速生长指数, 其地上、地下生物量间均表现为异速生长关系, 叶生物量与茎生物量或地下生物量间的异速生长指数为0.75 (
West et al ., 1997 ;
Enquist & Niklas, 2002 ;
Niklas & Enquist, 2002 ), 但后续研究又证明小个体植物在叶生物量、茎生物量和地下生物量间也可能为等速关系(
Niklas, 2005 ;
Cheng et al ., 2014 ;
陈国鹏和赵文智, 2016 ),
Poorter等(2015) 进一步分析个体大小影响时发现, 植物叶与茎、叶与根、茎与根等生物量间的异速生长指数随个体增大而减小.尽管基于生态代谢理论的异速生长指数在不同尺度上得到了检验(
Cheng & Niklas, 2007 ), 但是该理论以关注种间的变异为主, 跨物种的恒定关系不一定适用于种内, 对物种作用的考虑不足(
Müller et al ., 2000 ;
Cheng et al ., 2014 ), 尤其对一些极端生境的物种关注较少也限制着该理论的普适性. ...
... ≠ 1时, 模型反映的是异速生长关系(
Niklas & Enquist, 2002b ).生态代谢理论认为, 全球尺度上植物个体生物量具有恒定的异速生长指数, 其地上、地下生物量间均表现为异速生长关系, 叶生物量与茎生物量或地下生物量间的异速生长指数为0.75 (
West et al ., 1997 ;
Enquist & Niklas, 2002 ;
Niklas & Enquist, 2002 ), 但后续研究又证明小个体植物在叶生物量、茎生物量和地下生物量间也可能为等速关系(
Niklas, 2005 ;
Cheng et al ., 2014 ;
陈国鹏和赵文智, 2016 ),
Poorter等(2015) 进一步分析个体大小影响时发现, 植物叶与茎、叶与根、茎与根等生物量间的异速生长指数随个体增大而减小.尽管基于生态代谢理论的异速生长指数在不同尺度上得到了检验(
Cheng & Niklas, 2007 ), 但是该理论以关注种间的变异为主, 跨物种的恒定关系不一定适用于种内, 对物种作用的考虑不足(
Müller et al ., 2000 ;
Cheng et al ., 2014 ), 尤其对一些极端生境的物种关注较少也限制着该理论的普适性. ...
... ;
Niklas & Enquist, 2002 ), 但后续研究又证明小个体植物在叶生物量、茎生物量和地下生物量间也可能为等速关系(
Niklas, 2005 ;
Cheng et al ., 2014 ;
陈国鹏和赵文智, 2016 ),
Poorter等(2015) 进一步分析个体大小影响时发现, 植物叶与茎、叶与根、茎与根等生物量间的异速生长指数随个体增大而减小.尽管基于生态代谢理论的异速生长指数在不同尺度上得到了检验(
Cheng & Niklas, 2007 ), 但是该理论以关注种间的变异为主, 跨物种的恒定关系不一定适用于种内, 对物种作用的考虑不足(
Müller et al ., 2000 ;
Cheng et al ., 2014 ), 尤其对一些极端生境的物种关注较少也限制着该理论的普适性. ...
How does biomass distribution change with size and differ among species? An analysis for 1200 plant species from five continents
12
2015
... 生物量在不同器官间的分配是植物生态学研究的核心问题(
West et al .,1999 ;
Enquist & Niklas, 2002 ;
Poorter et al ., 2012 ).生物量分配模式不仅有助于从易测量的地上生物量估算地下生物量, 还体现着植物不同的生态策略(
Weiner, 2004 ), 反映出光合产物对植物不同器官(地上不同器官之间, 地上器官和地下器官之间)的投资以适应环境条件(
Niklas, 2006 ;
Poorter & Sack, 2012 ;
闫帮国等, 2016 ).最优分配假说认为, 植物倾向于将资源优先分配到能获得限制性资源的器官, 以帮助植物获取更多的受限资源, 从而维持最大生长速率(
Bloom et al ., 1985 ;
McCarthy & Enquist, 2007 ), 这意味着, 根据植物生长的环境, 如果光照受限, 生物量将倾向于叶和枝分配; 如果营养或水分受限, 生物量将倾向于根系分配(
Gleeson & Tilman, 1990 ).但植物生物量分配模式存在个体发育漂变(
Poorter & Sack, 2012 ;
Poorter et al ., 2015 ;
王杨等, 2017 ), 其分配比例会随植株的大小变化而变化(
Poorter et al ., 2015 ;
陈国鹏等, 2016 ), 使得最优分配假说的使用受到了限制, 而异速生长理论能较好地解决这一问题(
Weiner, 2004 ;
Gargaglione et al ., 2010 ), 且已被越来越多地应用到生物量分配研究中, 有****也曾对两种方法的优劣进行了比较(
McCarthy & Enquist, 2007 ;
Poorter & Sack, 2012 ), 更赞同将两种方法结合应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ). ...
... ), 其分配比例会随植株的大小变化而变化(
Poorter et al ., 2015 ;
陈国鹏等, 2016 ), 使得最优分配假说的使用受到了限制, 而异速生长理论能较好地解决这一问题(
Weiner, 2004 ;
Gargaglione et al ., 2010 ), 且已被越来越多地应用到生物量分配研究中, 有****也曾对两种方法的优劣进行了比较(
McCarthy & Enquist, 2007 ;
Poorter & Sack, 2012 ), 更赞同将两种方法结合应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ). ...
... 异速生长理论认为, 生物量的分配主要由植物的大小决定, 并可用幂函数
Y 1 =
βY 2 α 表示, 该方程经过对数转换后, 可线性化为log(
Y 1 ) = log(
β )
+ α log(
Y 2 ), 其中
Y 1 和
Y 2 是相互依赖的变量(如茎生物量和根生物量), log(
β )是异速生长常数,
α 是异速生长指数(
Niklas & Enquist, 2002a ,
2002b ).当
α = 1时, 模型反映的是等速生长关系; 当
α ≠ 1时, 模型反映的是异速生长关系(
Niklas & Enquist, 2002b ).生态代谢理论认为, 全球尺度上植物个体生物量具有恒定的异速生长指数, 其地上、地下生物量间均表现为异速生长关系, 叶生物量与茎生物量或地下生物量间的异速生长指数为0.75 (
West et al ., 1997 ;
Enquist & Niklas, 2002 ;
Niklas & Enquist, 2002 ), 但后续研究又证明小个体植物在叶生物量、茎生物量和地下生物量间也可能为等速关系(
Niklas, 2005 ;
Cheng et al ., 2014 ;
陈国鹏和赵文智, 2016 ),
Poorter等(2015) 进一步分析个体大小影响时发现, 植物叶与茎、叶与根、茎与根等生物量间的异速生长指数随个体增大而减小.尽管基于生态代谢理论的异速生长指数在不同尺度上得到了检验(
Cheng & Niklas, 2007 ), 但是该理论以关注种间的变异为主, 跨物种的恒定关系不一定适用于种内, 对物种作用的考虑不足(
Müller et al ., 2000 ;
Cheng et al ., 2014 ), 尤其对一些极端生境的物种关注较少也限制着该理论的普适性. ...
... 理论上来说, 极端环境通过长期的筛选作用, 其适生物种响应资源约束时的生物量权衡策略应具有相似性(
Reich et al ., 2014 ).如, 在高寒山地, 高海拔、寒冷、短生长季和低气压等环境约束可能使植物的资源权衡具有独特表现(
高巧等, 2014 ;
左有璐等, 2014 ;
刘尊驰等, 2015 ), 植物会增加根生物量、降低叶生物量比例来适应较冷的环境(
徐波等, 2013 ).但目前的相关研究材料以常绿裸子植物或落叶被子植物为主(
Reich et al ., 2014 ;
Poorter et al ., 2015 ), 对高寒区生长的常绿被子植物的生物量权衡机制缺乏认知, 而常绿植物与落叶植物叶生物量分配策略存在差别, 常绿植物在叶片上投入的较多, 因此叶生物量分配比例高于落叶植物(
Poorter et al ., 2015 ). ...
... ), 对高寒区生长的常绿被子植物的生物量权衡机制缺乏认知, 而常绿植物与落叶植物叶生物量分配策略存在差别, 常绿植物在叶片上投入的较多, 因此叶生物量分配比例高于落叶植物(
Poorter et al ., 2015 ). ...
... 树木对温度适应性策略的假设认为, 在寒冷的气候环境下, 根的分配比例较大、叶的分配比例较小(
Reich et al ., 2014 ), 本研究中, 7种杜鹃根生物量分配比例高达35.57%, 个别物种甚至超过40% (
图1 ), 高于全球绝大多数植物(
Poorter et al ., 2015 ), 也高于我国亚热带山地杜鹃灌丛根系生物量的分配比例(
张蔷等, 2017 ), 与一些高寒植被(
高巧等, 2014 ;
钟泽兵等, 2014 ;
杨路存等, 2018 ;
左有璐等, 2018 )相似, 低速率的养分循环和受限的土壤溶液运动驱动着高山、亚高山地区的低营养供应(
魏晶等, 2004 ;
Luo et al ., 2012 ;
Reich et al ., 2014 ), 这使得高寒杜鹃需提高根生物量分配比例以增强从寒冷土壤中吸收养分, 生物量向根系中的倾向分配支持最优分配假说.7种杜鹃叶生物量分配比例低于全球绝大多数植物(
Poorter et al ., 2015 ), 其适应机制也与一些高寒植物(
Reich et al ., 2014 )相似, 如
钟泽兵等(2014) 对青海玉树海拔3 715-4 296 m的5种建群灌木的研究发现叶生物量分配比例不足10%.甘肃南部高山-亚高山区的杜鹃会通过减少资源向叶片的分配, 保持常绿, 以较低的叶片周转率来适应寒冷环境.随着植株叶投资盈利能力的下降, 叶生物量将饱和, 投资会逐渐转向茎和根(
Poorter et al ., 2015 ), 而为增强植株的机械支撑, 必须增加茎的生物量投资(
Dybzinski et al ., 2011 ).本研究中7种高寒杜鹃茎生物量分配平均比例为45.61%, 接近于全球数据库平均值(
Poorter et al ., 2015 ).同时, 相比于叶和根, 茎生物量分配比例的种间差异最大(可达21%), 且多组物种间的差异显著(
表2 ), 这与
Poorter等(2015) 对全球1 200种植物整合分析的结果具有一致性, 茎不仅是植株的支撑、运输体系, 还在光竞争时向四周伸展小枝扩展空域, 而小枝的扩张又带来了高寒区防风的压力, 功能的多样性可能导致茎生物量分配的可塑性.各物种分配的差异还可能与茎构型有关, 如黄毛杜鹃和陇蜀杜鹃多为丛生, 茎枝扭曲, 其茎生物量分配比例的变异范围较大; 而美容杜鹃和山光杜鹃主干通直, 其茎生物量分配比例的变幅较小(
图1 ). ...
... ), 这使得高寒杜鹃需提高根生物量分配比例以增强从寒冷土壤中吸收养分, 生物量向根系中的倾向分配支持最优分配假说.7种杜鹃叶生物量分配比例低于全球绝大多数植物(
Poorter et al ., 2015 ), 其适应机制也与一些高寒植物(
Reich et al ., 2014 )相似, 如
钟泽兵等(2014) 对青海玉树海拔3 715-4 296 m的5种建群灌木的研究发现叶生物量分配比例不足10%.甘肃南部高山-亚高山区的杜鹃会通过减少资源向叶片的分配, 保持常绿, 以较低的叶片周转率来适应寒冷环境.随着植株叶投资盈利能力的下降, 叶生物量将饱和, 投资会逐渐转向茎和根(
Poorter et al ., 2015 ), 而为增强植株的机械支撑, 必须增加茎的生物量投资(
Dybzinski et al ., 2011 ).本研究中7种高寒杜鹃茎生物量分配平均比例为45.61%, 接近于全球数据库平均值(
Poorter et al ., 2015 ).同时, 相比于叶和根, 茎生物量分配比例的种间差异最大(可达21%), 且多组物种间的差异显著(
表2 ), 这与
Poorter等(2015) 对全球1 200种植物整合分析的结果具有一致性, 茎不仅是植株的支撑、运输体系, 还在光竞争时向四周伸展小枝扩展空域, 而小枝的扩张又带来了高寒区防风的压力, 功能的多样性可能导致茎生物量分配的可塑性.各物种分配的差异还可能与茎构型有关, 如黄毛杜鹃和陇蜀杜鹃多为丛生, 茎枝扭曲, 其茎生物量分配比例的变异范围较大; 而美容杜鹃和山光杜鹃主干通直, 其茎生物量分配比例的变幅较小(
图1 ). ...
... 对青海玉树海拔3 715-4 296 m的5种建群灌木的研究发现叶生物量分配比例不足10%.甘肃南部高山-亚高山区的杜鹃会通过减少资源向叶片的分配, 保持常绿, 以较低的叶片周转率来适应寒冷环境.随着植株叶投资盈利能力的下降, 叶生物量将饱和, 投资会逐渐转向茎和根(
Poorter et al ., 2015 ), 而为增强植株的机械支撑, 必须增加茎的生物量投资(
Dybzinski et al ., 2011 ).本研究中7种高寒杜鹃茎生物量分配平均比例为45.61%, 接近于全球数据库平均值(
Poorter et al ., 2015 ).同时, 相比于叶和根, 茎生物量分配比例的种间差异最大(可达21%), 且多组物种间的差异显著(
表2 ), 这与
Poorter等(2015) 对全球1 200种植物整合分析的结果具有一致性, 茎不仅是植株的支撑、运输体系, 还在光竞争时向四周伸展小枝扩展空域, 而小枝的扩张又带来了高寒区防风的压力, 功能的多样性可能导致茎生物量分配的可塑性.各物种分配的差异还可能与茎构型有关, 如黄毛杜鹃和陇蜀杜鹃多为丛生, 茎枝扭曲, 其茎生物量分配比例的变异范围较大; 而美容杜鹃和山光杜鹃主干通直, 其茎生物量分配比例的变幅较小(
图1 ). ...
... ).本研究中7种高寒杜鹃茎生物量分配平均比例为45.61%, 接近于全球数据库平均值(
Poorter et al ., 2015 ).同时, 相比于叶和根, 茎生物量分配比例的种间差异最大(可达21%), 且多组物种间的差异显著(
表2 ), 这与
Poorter等(2015) 对全球1 200种植物整合分析的结果具有一致性, 茎不仅是植株的支撑、运输体系, 还在光竞争时向四周伸展小枝扩展空域, 而小枝的扩张又带来了高寒区防风的压力, 功能的多样性可能导致茎生物量分配的可塑性.各物种分配的差异还可能与茎构型有关, 如黄毛杜鹃和陇蜀杜鹃多为丛生, 茎枝扭曲, 其茎生物量分配比例的变异范围较大; 而美容杜鹃和山光杜鹃主干通直, 其茎生物量分配比例的变幅较小(
图1 ). ...
... ), 这与
Poorter等(2015) 对全球1 200种植物整合分析的结果具有一致性, 茎不仅是植株的支撑、运输体系, 还在光竞争时向四周伸展小枝扩展空域, 而小枝的扩张又带来了高寒区防风的压力, 功能的多样性可能导致茎生物量分配的可塑性.各物种分配的差异还可能与茎构型有关, 如黄毛杜鹃和陇蜀杜鹃多为丛生, 茎枝扭曲, 其茎生物量分配比例的变异范围较大; 而美容杜鹃和山光杜鹃主干通直, 其茎生物量分配比例的变幅较小(
图1 ). ...
... 异速生长理论预测小个体植物叶、茎、根器官生长可能为等速关系(
Enquist & Niklas, 2002 ;
Weiner, 2004 ;
Niklas, 2006 ), 这种基于种间变异建立的跨物种关系已在种内变异的研究中得到了较多应用(
Gargaglione et al ., 2010 ;
Cheng et al ., 2014 ;
陈国鹏等, 2016 ;
王杨等, 2017 ), 如
Cheng等(2014) 发现南亚热带生长的5种苗木叶、茎和根生物量的积累为等速或近似等速生长关系.本研究中, 7种高寒杜鹃叶与茎、叶与根以及茎与根之间既有异速生长表现, 也有等速生长表现(
图2 -5), 与荒漠植物(
杨昊天等, 2013 ;
陶冶和张元明, 2014 )、干热河谷草本植物(
闫帮国等, 2016 )的异速生长关系具有相似性, 不同于川西高寒草甸优势植物(
左有璐等, 2018 ), 并不完全支持小个体器官间等速生长关系的预测, 也不完全支持生态代谢理论的预测, 这可能是由于植物在适应特殊环境的过程中, 不同物种间具有很大的功能群差异, 这些功能群差异可能是不同器官生物量间异速生长关系分化的主要因素(
Poorter et al ., 2015 ;
闫帮国等, 2016 ), 而基于跨物种建立的生态代谢理论、小个体等速生长理论忽略了物种间的功能群差异. ...
... 物种是影响植物器官生物量分配异速生长关系的重要因素(
Poorter et al ., 2012 ;
闫帮国等, 2016 ;
张蔷等, 2017 ), 基于大样本的研究则发现生物量分配比例与物种的系统发育有关, 如裸子植物叶片上的投入更多(
Reich et al ., 2014 ;
Poorter et al ., 2015 ).本研究中7种杜鹃隶属于同科同属, 其生物量分配比例及异速生长关系的种间差异显著(
表2 ,
表3 ), 陇蜀杜鹃根生物量分配比例较低, 美容杜鹃叶生物量分配比例较高而茎生物量分配比例较低, 黄毛杜鹃、头花杜鹃地上生物量与地下生物量呈等速生长, 陇蜀杜鹃、麻花杜鹃地上生物量的积累速率大于地下, 其余3种地上生物量积累速率小于地下(
图5 ), 而叶与茎、叶与根、茎与根间的异速生长关系的物种差异更为复杂多样, 如黄毛杜鹃、陇蜀杜鹃、美容杜鹃和山光杜鹃的叶与茎, 所有物种的叶与根, 美容杜鹃、太白杜鹃、黄毛杜鹃、陇蜀杜鹃和麻花杜鹃的茎与根为异速生长关系, 而太白杜鹃、头花杜鹃和麻花杜鹃的叶与茎, 山光杜鹃和头花杜鹃的茎与根为等速生长关系(
图2 -4).这也说明不同的物种可能在进化过程中会通过生态位分化而占有不同的环境, 在本研究区域, 头花杜鹃主要分布于海拔3 500 m以上并形成单优群落或与陇蜀杜鹃形成共生群落; 陇蜀杜鹃、黄毛杜鹃在海拔3 000-3 700 m间有分布, 但在3 200 m以上才形成单优群落或群落优势种; 山光杜鹃和麻花杜鹃海拔分布范围跨度大(2 300-3 500 m), 但呈群落优势种的仅见3 000 m处个别的冷杉皆伐更新迹地,
Veresoglou和 Pe?uelas (2019) 分析欧亚大陆植物生物量时发现分配的变异强度与物种分布范围有密切的关系, 但杜鹃分布范围与生物量分配特征间是否存在相关关系, 尚需持续研究. ...
Biomass allocation to leaves, stems and roots: meta-analyses of interspecific variation and environmental control
4
2012
... 生物量在不同器官间的分配是植物生态学研究的核心问题(
West et al .,1999 ;
Enquist & Niklas, 2002 ;
Poorter et al ., 2012 ).生物量分配模式不仅有助于从易测量的地上生物量估算地下生物量, 还体现着植物不同的生态策略(
Weiner, 2004 ), 反映出光合产物对植物不同器官(地上不同器官之间, 地上器官和地下器官之间)的投资以适应环境条件(
Niklas, 2006 ;
Poorter & Sack, 2012 ;
闫帮国等, 2016 ).最优分配假说认为, 植物倾向于将资源优先分配到能获得限制性资源的器官, 以帮助植物获取更多的受限资源, 从而维持最大生长速率(
Bloom et al ., 1985 ;
McCarthy & Enquist, 2007 ), 这意味着, 根据植物生长的环境, 如果光照受限, 生物量将倾向于叶和枝分配; 如果营养或水分受限, 生物量将倾向于根系分配(
Gleeson & Tilman, 1990 ).但植物生物量分配模式存在个体发育漂变(
Poorter & Sack, 2012 ;
Poorter et al ., 2015 ;
王杨等, 2017 ), 其分配比例会随植株的大小变化而变化(
Poorter et al ., 2015 ;
陈国鹏等, 2016 ), 使得最优分配假说的使用受到了限制, 而异速生长理论能较好地解决这一问题(
Weiner, 2004 ;
Gargaglione et al ., 2010 ), 且已被越来越多地应用到生物量分配研究中, 有****也曾对两种方法的优劣进行了比较(
McCarthy & Enquist, 2007 ;
Poorter & Sack, 2012 ), 更赞同将两种方法结合应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ). ...
... ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ). ...
... 尽管异速生长分析不受个体大小等因素干扰, 可以较好地检验不同器官生长发育协调性, 并确定这种协调性在物种间的变化或对环境的响应, 但在一些生态模型的构建研究中, 叶、茎和根分配比例的实际值要比器官间的协调性重要(
Poorter & Sack, 2012 ), 考虑到不同分析方法在研究生物量分配中的局限性, 将异速生长法与最优分配假说结合已在许多研究中得到了应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ;
陈国鹏等, 2016 ).本研究也结合最优分配假说与异速生长分析来研究甘肃南部高山-亚高山区杜鹃地上、地下器官生物量的分配特征, 结果表明7种高寒常绿杜鹃根、茎、叶生物量分配比例和异速生长关系受物种影响显著, 高寒杜鹃对特殊生境适应过程中的生物量分配模式有许多相似之处, 而物种差异又导致相似生境中生物量分配模式的分异. ...
... 物种是影响植物器官生物量分配异速生长关系的重要因素(
Poorter et al ., 2012 ;
闫帮国等, 2016 ;
张蔷等, 2017 ), 基于大样本的研究则发现生物量分配比例与物种的系统发育有关, 如裸子植物叶片上的投入更多(
Reich et al ., 2014 ;
Poorter et al ., 2015 ).本研究中7种杜鹃隶属于同科同属, 其生物量分配比例及异速生长关系的种间差异显著(
表2 ,
表3 ), 陇蜀杜鹃根生物量分配比例较低, 美容杜鹃叶生物量分配比例较高而茎生物量分配比例较低, 黄毛杜鹃、头花杜鹃地上生物量与地下生物量呈等速生长, 陇蜀杜鹃、麻花杜鹃地上生物量的积累速率大于地下, 其余3种地上生物量积累速率小于地下(
图5 ), 而叶与茎、叶与根、茎与根间的异速生长关系的物种差异更为复杂多样, 如黄毛杜鹃、陇蜀杜鹃、美容杜鹃和山光杜鹃的叶与茎, 所有物种的叶与根, 美容杜鹃、太白杜鹃、黄毛杜鹃、陇蜀杜鹃和麻花杜鹃的茎与根为异速生长关系, 而太白杜鹃、头花杜鹃和麻花杜鹃的叶与茎, 山光杜鹃和头花杜鹃的茎与根为等速生长关系(
图2 -4).这也说明不同的物种可能在进化过程中会通过生态位分化而占有不同的环境, 在本研究区域, 头花杜鹃主要分布于海拔3 500 m以上并形成单优群落或与陇蜀杜鹃形成共生群落; 陇蜀杜鹃、黄毛杜鹃在海拔3 000-3 700 m间有分布, 但在3 200 m以上才形成单优群落或群落优势种; 山光杜鹃和麻花杜鹃海拔分布范围跨度大(2 300-3 500 m), 但呈群落优势种的仅见3 000 m处个别的冷杉皆伐更新迹地,
Veresoglou和 Pe?uelas (2019) 分析欧亚大陆植物生物量时发现分配的变异强度与物种分布范围有密切的关系, 但杜鹃分布范围与生物量分配特征间是否存在相关关系, 尚需持续研究. ...
Pitfalls and possibilities in the analysis of biomass allocation patterns in plants
6
2012
... 生物量在不同器官间的分配是植物生态学研究的核心问题(
West et al .,1999 ;
Enquist & Niklas, 2002 ;
Poorter et al ., 2012 ).生物量分配模式不仅有助于从易测量的地上生物量估算地下生物量, 还体现着植物不同的生态策略(
Weiner, 2004 ), 反映出光合产物对植物不同器官(地上不同器官之间, 地上器官和地下器官之间)的投资以适应环境条件(
Niklas, 2006 ;
Poorter & Sack, 2012 ;
闫帮国等, 2016 ).最优分配假说认为, 植物倾向于将资源优先分配到能获得限制性资源的器官, 以帮助植物获取更多的受限资源, 从而维持最大生长速率(
Bloom et al ., 1985 ;
McCarthy & Enquist, 2007 ), 这意味着, 根据植物生长的环境, 如果光照受限, 生物量将倾向于叶和枝分配; 如果营养或水分受限, 生物量将倾向于根系分配(
Gleeson & Tilman, 1990 ).但植物生物量分配模式存在个体发育漂变(
Poorter & Sack, 2012 ;
Poorter et al ., 2015 ;
王杨等, 2017 ), 其分配比例会随植株的大小变化而变化(
Poorter et al ., 2015 ;
陈国鹏等, 2016 ), 使得最优分配假说的使用受到了限制, 而异速生长理论能较好地解决这一问题(
Weiner, 2004 ;
Gargaglione et al ., 2010 ), 且已被越来越多地应用到生物量分配研究中, 有****也曾对两种方法的优劣进行了比较(
McCarthy & Enquist, 2007 ;
Poorter & Sack, 2012 ), 更赞同将两种方法结合应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ). ...
... ).但植物生物量分配模式存在个体发育漂变(
Poorter & Sack, 2012 ;
Poorter et al ., 2015 ;
王杨等, 2017 ), 其分配比例会随植株的大小变化而变化(
Poorter et al ., 2015 ;
陈国鹏等, 2016 ), 使得最优分配假说的使用受到了限制, 而异速生长理论能较好地解决这一问题(
Weiner, 2004 ;
Gargaglione et al ., 2010 ), 且已被越来越多地应用到生物量分配研究中, 有****也曾对两种方法的优劣进行了比较(
McCarthy & Enquist, 2007 ;
Poorter & Sack, 2012 ), 更赞同将两种方法结合应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ). ...
... ;
Poorter & Sack, 2012 ), 更赞同将两种方法结合应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ). ...
... ), 更赞同将两种方法结合应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ). ...
... 尽管异速生长分析不受个体大小等因素干扰, 可以较好地检验不同器官生长发育协调性, 并确定这种协调性在物种间的变化或对环境的响应, 但在一些生态模型的构建研究中, 叶、茎和根分配比例的实际值要比器官间的协调性重要(
Poorter & Sack, 2012 ), 考虑到不同分析方法在研究生物量分配中的局限性, 将异速生长法与最优分配假说结合已在许多研究中得到了应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ;
陈国鹏等, 2016 ).本研究也结合最优分配假说与异速生长分析来研究甘肃南部高山-亚高山区杜鹃地上、地下器官生物量的分配特征, 结果表明7种高寒常绿杜鹃根、茎、叶生物量分配比例和异速生长关系受物种影响显著, 高寒杜鹃对特殊生境适应过程中的生物量分配模式有许多相似之处, 而物种差异又导致相似生境中生物量分配模式的分异. ...
... ), 考虑到不同分析方法在研究生物量分配中的局限性, 将异速生长法与最优分配假说结合已在许多研究中得到了应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ;
陈国鹏等, 2016 ).本研究也结合最优分配假说与异速生长分析来研究甘肃南部高山-亚高山区杜鹃地上、地下器官生物量的分配特征, 结果表明7种高寒常绿杜鹃根、茎、叶生物量分配比例和异速生长关系受物种影响显著, 高寒杜鹃对特殊生境适应过程中的生物量分配模式有许多相似之处, 而物种差异又导致相似生境中生物量分配模式的分异. ...
Temperature drives global patterns in forest biomass distribution in leaves, stems, and roots
6
2014
... 理论上来说, 极端环境通过长期的筛选作用, 其适生物种响应资源约束时的生物量权衡策略应具有相似性(
Reich et al ., 2014 ).如, 在高寒山地, 高海拔、寒冷、短生长季和低气压等环境约束可能使植物的资源权衡具有独特表现(
高巧等, 2014 ;
左有璐等, 2014 ;
刘尊驰等, 2015 ), 植物会增加根生物量、降低叶生物量比例来适应较冷的环境(
徐波等, 2013 ).但目前的相关研究材料以常绿裸子植物或落叶被子植物为主(
Reich et al ., 2014 ;
Poorter et al ., 2015 ), 对高寒区生长的常绿被子植物的生物量权衡机制缺乏认知, 而常绿植物与落叶植物叶生物量分配策略存在差别, 常绿植物在叶片上投入的较多, 因此叶生物量分配比例高于落叶植物(
Poorter et al ., 2015 ). ...
... ).但目前的相关研究材料以常绿裸子植物或落叶被子植物为主(
Reich et al ., 2014 ;
Poorter et al ., 2015 ), 对高寒区生长的常绿被子植物的生物量权衡机制缺乏认知, 而常绿植物与落叶植物叶生物量分配策略存在差别, 常绿植物在叶片上投入的较多, 因此叶生物量分配比例高于落叶植物(
Poorter et al ., 2015 ). ...
... 树木对温度适应性策略的假设认为, 在寒冷的气候环境下, 根的分配比例较大、叶的分配比例较小(
Reich et al ., 2014 ), 本研究中, 7种杜鹃根生物量分配比例高达35.57%, 个别物种甚至超过40% (
图1 ), 高于全球绝大多数植物(
Poorter et al ., 2015 ), 也高于我国亚热带山地杜鹃灌丛根系生物量的分配比例(
张蔷等, 2017 ), 与一些高寒植被(
高巧等, 2014 ;
钟泽兵等, 2014 ;
杨路存等, 2018 ;
左有璐等, 2018 )相似, 低速率的养分循环和受限的土壤溶液运动驱动着高山、亚高山地区的低营养供应(
魏晶等, 2004 ;
Luo et al ., 2012 ;
Reich et al ., 2014 ), 这使得高寒杜鹃需提高根生物量分配比例以增强从寒冷土壤中吸收养分, 生物量向根系中的倾向分配支持最优分配假说.7种杜鹃叶生物量分配比例低于全球绝大多数植物(
Poorter et al ., 2015 ), 其适应机制也与一些高寒植物(
Reich et al ., 2014 )相似, 如
钟泽兵等(2014) 对青海玉树海拔3 715-4 296 m的5种建群灌木的研究发现叶生物量分配比例不足10%.甘肃南部高山-亚高山区的杜鹃会通过减少资源向叶片的分配, 保持常绿, 以较低的叶片周转率来适应寒冷环境.随着植株叶投资盈利能力的下降, 叶生物量将饱和, 投资会逐渐转向茎和根(
Poorter et al ., 2015 ), 而为增强植株的机械支撑, 必须增加茎的生物量投资(
Dybzinski et al ., 2011 ).本研究中7种高寒杜鹃茎生物量分配平均比例为45.61%, 接近于全球数据库平均值(
Poorter et al ., 2015 ).同时, 相比于叶和根, 茎生物量分配比例的种间差异最大(可达21%), 且多组物种间的差异显著(
表2 ), 这与
Poorter等(2015) 对全球1 200种植物整合分析的结果具有一致性, 茎不仅是植株的支撑、运输体系, 还在光竞争时向四周伸展小枝扩展空域, 而小枝的扩张又带来了高寒区防风的压力, 功能的多样性可能导致茎生物量分配的可塑性.各物种分配的差异还可能与茎构型有关, 如黄毛杜鹃和陇蜀杜鹃多为丛生, 茎枝扭曲, 其茎生物量分配比例的变异范围较大; 而美容杜鹃和山光杜鹃主干通直, 其茎生物量分配比例的变幅较小(
图1 ). ...
... ;
Reich et al ., 2014 ), 这使得高寒杜鹃需提高根生物量分配比例以增强从寒冷土壤中吸收养分, 生物量向根系中的倾向分配支持最优分配假说.7种杜鹃叶生物量分配比例低于全球绝大多数植物(
Poorter et al ., 2015 ), 其适应机制也与一些高寒植物(
Reich et al ., 2014 )相似, 如
钟泽兵等(2014) 对青海玉树海拔3 715-4 296 m的5种建群灌木的研究发现叶生物量分配比例不足10%.甘肃南部高山-亚高山区的杜鹃会通过减少资源向叶片的分配, 保持常绿, 以较低的叶片周转率来适应寒冷环境.随着植株叶投资盈利能力的下降, 叶生物量将饱和, 投资会逐渐转向茎和根(
Poorter et al ., 2015 ), 而为增强植株的机械支撑, 必须增加茎的生物量投资(
Dybzinski et al ., 2011 ).本研究中7种高寒杜鹃茎生物量分配平均比例为45.61%, 接近于全球数据库平均值(
Poorter et al ., 2015 ).同时, 相比于叶和根, 茎生物量分配比例的种间差异最大(可达21%), 且多组物种间的差异显著(
表2 ), 这与
Poorter等(2015) 对全球1 200种植物整合分析的结果具有一致性, 茎不仅是植株的支撑、运输体系, 还在光竞争时向四周伸展小枝扩展空域, 而小枝的扩张又带来了高寒区防风的压力, 功能的多样性可能导致茎生物量分配的可塑性.各物种分配的差异还可能与茎构型有关, 如黄毛杜鹃和陇蜀杜鹃多为丛生, 茎枝扭曲, 其茎生物量分配比例的变异范围较大; 而美容杜鹃和山光杜鹃主干通直, 其茎生物量分配比例的变幅较小(
图1 ). ...
... ), 其适应机制也与一些高寒植物(
Reich et al ., 2014 )相似, 如
钟泽兵等(2014) 对青海玉树海拔3 715-4 296 m的5种建群灌木的研究发现叶生物量分配比例不足10%.甘肃南部高山-亚高山区的杜鹃会通过减少资源向叶片的分配, 保持常绿, 以较低的叶片周转率来适应寒冷环境.随着植株叶投资盈利能力的下降, 叶生物量将饱和, 投资会逐渐转向茎和根(
Poorter et al ., 2015 ), 而为增强植株的机械支撑, 必须增加茎的生物量投资(
Dybzinski et al ., 2011 ).本研究中7种高寒杜鹃茎生物量分配平均比例为45.61%, 接近于全球数据库平均值(
Poorter et al ., 2015 ).同时, 相比于叶和根, 茎生物量分配比例的种间差异最大(可达21%), 且多组物种间的差异显著(
表2 ), 这与
Poorter等(2015) 对全球1 200种植物整合分析的结果具有一致性, 茎不仅是植株的支撑、运输体系, 还在光竞争时向四周伸展小枝扩展空域, 而小枝的扩张又带来了高寒区防风的压力, 功能的多样性可能导致茎生物量分配的可塑性.各物种分配的差异还可能与茎构型有关, 如黄毛杜鹃和陇蜀杜鹃多为丛生, 茎枝扭曲, 其茎生物量分配比例的变异范围较大; 而美容杜鹃和山光杜鹃主干通直, 其茎生物量分配比例的变幅较小(
图1 ). ...
... 物种是影响植物器官生物量分配异速生长关系的重要因素(
Poorter et al ., 2012 ;
闫帮国等, 2016 ;
张蔷等, 2017 ), 基于大样本的研究则发现生物量分配比例与物种的系统发育有关, 如裸子植物叶片上的投入更多(
Reich et al ., 2014 ;
Poorter et al ., 2015 ).本研究中7种杜鹃隶属于同科同属, 其生物量分配比例及异速生长关系的种间差异显著(
表2 ,
表3 ), 陇蜀杜鹃根生物量分配比例较低, 美容杜鹃叶生物量分配比例较高而茎生物量分配比例较低, 黄毛杜鹃、头花杜鹃地上生物量与地下生物量呈等速生长, 陇蜀杜鹃、麻花杜鹃地上生物量的积累速率大于地下, 其余3种地上生物量积累速率小于地下(
图5 ), 而叶与茎、叶与根、茎与根间的异速生长关系的物种差异更为复杂多样, 如黄毛杜鹃、陇蜀杜鹃、美容杜鹃和山光杜鹃的叶与茎, 所有物种的叶与根, 美容杜鹃、太白杜鹃、黄毛杜鹃、陇蜀杜鹃和麻花杜鹃的茎与根为异速生长关系, 而太白杜鹃、头花杜鹃和麻花杜鹃的叶与茎, 山光杜鹃和头花杜鹃的茎与根为等速生长关系(
图2 -4).这也说明不同的物种可能在进化过程中会通过生态位分化而占有不同的环境, 在本研究区域, 头花杜鹃主要分布于海拔3 500 m以上并形成单优群落或与陇蜀杜鹃形成共生群落; 陇蜀杜鹃、黄毛杜鹃在海拔3 000-3 700 m间有分布, 但在3 200 m以上才形成单优群落或群落优势种; 山光杜鹃和麻花杜鹃海拔分布范围跨度大(2 300-3 500 m), 但呈群落优势种的仅见3 000 m处个别的冷杉皆伐更新迹地,
Veresoglou和 Pe?uelas (2019) 分析欧亚大陆植物生物量时发现分配的变异强度与物种分布范围有密切的关系, 但杜鹃分布范围与生物量分配特征间是否存在相关关系, 尚需持续研究. ...
准噶尔荒漠6种类短命植物生物量分配与异速生长关系
1
2014
... 异速生长理论预测小个体植物叶、茎、根器官生长可能为等速关系(
Enquist & Niklas, 2002 ;
Weiner, 2004 ;
Niklas, 2006 ), 这种基于种间变异建立的跨物种关系已在种内变异的研究中得到了较多应用(
Gargaglione et al ., 2010 ;
Cheng et al ., 2014 ;
陈国鹏等, 2016 ;
王杨等, 2017 ), 如
Cheng等(2014) 发现南亚热带生长的5种苗木叶、茎和根生物量的积累为等速或近似等速生长关系.本研究中, 7种高寒杜鹃叶与茎、叶与根以及茎与根之间既有异速生长表现, 也有等速生长表现(
图2 -5), 与荒漠植物(
杨昊天等, 2013 ;
陶冶和张元明, 2014 )、干热河谷草本植物(
闫帮国等, 2016 )的异速生长关系具有相似性, 不同于川西高寒草甸优势植物(
左有璐等, 2018 ), 并不完全支持小个体器官间等速生长关系的预测, 也不完全支持生态代谢理论的预测, 这可能是由于植物在适应特殊环境的过程中, 不同物种间具有很大的功能群差异, 这些功能群差异可能是不同器官生物量间异速生长关系分化的主要因素(
Poorter et al ., 2015 ;
闫帮国等, 2016 ), 而基于跨物种建立的生态代谢理论、小个体等速生长理论忽略了物种间的功能群差异. ...
准噶尔荒漠6种类短命植物生物量分配与异速生长关系
1
2014
... 异速生长理论预测小个体植物叶、茎、根器官生长可能为等速关系(
Enquist & Niklas, 2002 ;
Weiner, 2004 ;
Niklas, 2006 ), 这种基于种间变异建立的跨物种关系已在种内变异的研究中得到了较多应用(
Gargaglione et al ., 2010 ;
Cheng et al ., 2014 ;
陈国鹏等, 2016 ;
王杨等, 2017 ), 如
Cheng等(2014) 发现南亚热带生长的5种苗木叶、茎和根生物量的积累为等速或近似等速生长关系.本研究中, 7种高寒杜鹃叶与茎、叶与根以及茎与根之间既有异速生长表现, 也有等速生长表现(
图2 -5), 与荒漠植物(
杨昊天等, 2013 ;
陶冶和张元明, 2014 )、干热河谷草本植物(
闫帮国等, 2016 )的异速生长关系具有相似性, 不同于川西高寒草甸优势植物(
左有璐等, 2018 ), 并不完全支持小个体器官间等速生长关系的预测, 也不完全支持生态代谢理论的预测, 这可能是由于植物在适应特殊环境的过程中, 不同物种间具有很大的功能群差异, 这些功能群差异可能是不同器官生物量间异速生长关系分化的主要因素(
Poorter et al ., 2015 ;
闫帮国等, 2016 ), 而基于跨物种建立的生态代谢理论、小个体等速生长理论忽略了物种间的功能群差异. ...
Variance in biomass- allocation fractions is explained by distribution in European trees
1
2019
... 物种是影响植物器官生物量分配异速生长关系的重要因素(
Poorter et al ., 2012 ;
闫帮国等, 2016 ;
张蔷等, 2017 ), 基于大样本的研究则发现生物量分配比例与物种的系统发育有关, 如裸子植物叶片上的投入更多(
Reich et al ., 2014 ;
Poorter et al ., 2015 ).本研究中7种杜鹃隶属于同科同属, 其生物量分配比例及异速生长关系的种间差异显著(
表2 ,
表3 ), 陇蜀杜鹃根生物量分配比例较低, 美容杜鹃叶生物量分配比例较高而茎生物量分配比例较低, 黄毛杜鹃、头花杜鹃地上生物量与地下生物量呈等速生长, 陇蜀杜鹃、麻花杜鹃地上生物量的积累速率大于地下, 其余3种地上生物量积累速率小于地下(
图5 ), 而叶与茎、叶与根、茎与根间的异速生长关系的物种差异更为复杂多样, 如黄毛杜鹃、陇蜀杜鹃、美容杜鹃和山光杜鹃的叶与茎, 所有物种的叶与根, 美容杜鹃、太白杜鹃、黄毛杜鹃、陇蜀杜鹃和麻花杜鹃的茎与根为异速生长关系, 而太白杜鹃、头花杜鹃和麻花杜鹃的叶与茎, 山光杜鹃和头花杜鹃的茎与根为等速生长关系(
图2 -4).这也说明不同的物种可能在进化过程中会通过生态位分化而占有不同的环境, 在本研究区域, 头花杜鹃主要分布于海拔3 500 m以上并形成单优群落或与陇蜀杜鹃形成共生群落; 陇蜀杜鹃、黄毛杜鹃在海拔3 000-3 700 m间有分布, 但在3 200 m以上才形成单优群落或群落优势种; 山光杜鹃和麻花杜鹃海拔分布范围跨度大(2 300-3 500 m), 但呈群落优势种的仅见3 000 m处个别的冷杉皆伐更新迹地,
Veresoglou和 Pe?uelas (2019) 分析欧亚大陆植物生物量时发现分配的变异强度与物种分布范围有密切的关系, 但杜鹃分布范围与生物量分配特征间是否存在相关关系, 尚需持续研究. ...
檵木生物量分配特征
2
2017
... 生物量在不同器官间的分配是植物生态学研究的核心问题(
West et al .,1999 ;
Enquist & Niklas, 2002 ;
Poorter et al ., 2012 ).生物量分配模式不仅有助于从易测量的地上生物量估算地下生物量, 还体现着植物不同的生态策略(
Weiner, 2004 ), 反映出光合产物对植物不同器官(地上不同器官之间, 地上器官和地下器官之间)的投资以适应环境条件(
Niklas, 2006 ;
Poorter & Sack, 2012 ;
闫帮国等, 2016 ).最优分配假说认为, 植物倾向于将资源优先分配到能获得限制性资源的器官, 以帮助植物获取更多的受限资源, 从而维持最大生长速率(
Bloom et al ., 1985 ;
McCarthy & Enquist, 2007 ), 这意味着, 根据植物生长的环境, 如果光照受限, 生物量将倾向于叶和枝分配; 如果营养或水分受限, 生物量将倾向于根系分配(
Gleeson & Tilman, 1990 ).但植物生物量分配模式存在个体发育漂变(
Poorter & Sack, 2012 ;
Poorter et al ., 2015 ;
王杨等, 2017 ), 其分配比例会随植株的大小变化而变化(
Poorter et al ., 2015 ;
陈国鹏等, 2016 ), 使得最优分配假说的使用受到了限制, 而异速生长理论能较好地解决这一问题(
Weiner, 2004 ;
Gargaglione et al ., 2010 ), 且已被越来越多地应用到生物量分配研究中, 有****也曾对两种方法的优劣进行了比较(
McCarthy & Enquist, 2007 ;
Poorter & Sack, 2012 ), 更赞同将两种方法结合应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ). ...
... 异速生长理论预测小个体植物叶、茎、根器官生长可能为等速关系(
Enquist & Niklas, 2002 ;
Weiner, 2004 ;
Niklas, 2006 ), 这种基于种间变异建立的跨物种关系已在种内变异的研究中得到了较多应用(
Gargaglione et al ., 2010 ;
Cheng et al ., 2014 ;
陈国鹏等, 2016 ;
王杨等, 2017 ), 如
Cheng等(2014) 发现南亚热带生长的5种苗木叶、茎和根生物量的积累为等速或近似等速生长关系.本研究中, 7种高寒杜鹃叶与茎、叶与根以及茎与根之间既有异速生长表现, 也有等速生长表现(
图2 -5), 与荒漠植物(
杨昊天等, 2013 ;
陶冶和张元明, 2014 )、干热河谷草本植物(
闫帮国等, 2016 )的异速生长关系具有相似性, 不同于川西高寒草甸优势植物(
左有璐等, 2018 ), 并不完全支持小个体器官间等速生长关系的预测, 也不完全支持生态代谢理论的预测, 这可能是由于植物在适应特殊环境的过程中, 不同物种间具有很大的功能群差异, 这些功能群差异可能是不同器官生物量间异速生长关系分化的主要因素(
Poorter et al ., 2015 ;
闫帮国等, 2016 ), 而基于跨物种建立的生态代谢理论、小个体等速生长理论忽略了物种间的功能群差异. ...
檵木生物量分配特征
2
2017
... 生物量在不同器官间的分配是植物生态学研究的核心问题(
West et al .,1999 ;
Enquist & Niklas, 2002 ;
Poorter et al ., 2012 ).生物量分配模式不仅有助于从易测量的地上生物量估算地下生物量, 还体现着植物不同的生态策略(
Weiner, 2004 ), 反映出光合产物对植物不同器官(地上不同器官之间, 地上器官和地下器官之间)的投资以适应环境条件(
Niklas, 2006 ;
Poorter & Sack, 2012 ;
闫帮国等, 2016 ).最优分配假说认为, 植物倾向于将资源优先分配到能获得限制性资源的器官, 以帮助植物获取更多的受限资源, 从而维持最大生长速率(
Bloom et al ., 1985 ;
McCarthy & Enquist, 2007 ), 这意味着, 根据植物生长的环境, 如果光照受限, 生物量将倾向于叶和枝分配; 如果营养或水分受限, 生物量将倾向于根系分配(
Gleeson & Tilman, 1990 ).但植物生物量分配模式存在个体发育漂变(
Poorter & Sack, 2012 ;
Poorter et al ., 2015 ;
王杨等, 2017 ), 其分配比例会随植株的大小变化而变化(
Poorter et al ., 2015 ;
陈国鹏等, 2016 ), 使得最优分配假说的使用受到了限制, 而异速生长理论能较好地解决这一问题(
Weiner, 2004 ;
Gargaglione et al ., 2010 ), 且已被越来越多地应用到生物量分配研究中, 有****也曾对两种方法的优劣进行了比较(
McCarthy & Enquist, 2007 ;
Poorter & Sack, 2012 ), 更赞同将两种方法结合应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ). ...
... 异速生长理论预测小个体植物叶、茎、根器官生长可能为等速关系(
Enquist & Niklas, 2002 ;
Weiner, 2004 ;
Niklas, 2006 ), 这种基于种间变异建立的跨物种关系已在种内变异的研究中得到了较多应用(
Gargaglione et al ., 2010 ;
Cheng et al ., 2014 ;
陈国鹏等, 2016 ;
王杨等, 2017 ), 如
Cheng等(2014) 发现南亚热带生长的5种苗木叶、茎和根生物量的积累为等速或近似等速生长关系.本研究中, 7种高寒杜鹃叶与茎、叶与根以及茎与根之间既有异速生长表现, 也有等速生长表现(
图2 -5), 与荒漠植物(
杨昊天等, 2013 ;
陶冶和张元明, 2014 )、干热河谷草本植物(
闫帮国等, 2016 )的异速生长关系具有相似性, 不同于川西高寒草甸优势植物(
左有璐等, 2018 ), 并不完全支持小个体器官间等速生长关系的预测, 也不完全支持生态代谢理论的预测, 这可能是由于植物在适应特殊环境的过程中, 不同物种间具有很大的功能群差异, 这些功能群差异可能是不同器官生物量间异速生长关系分化的主要因素(
Poorter et al ., 2015 ;
闫帮国等, 2016 ), 而基于跨物种建立的生态代谢理论、小个体等速生长理论忽略了物种间的功能群差异. ...
长白山高山冻原植被生物量的分布规律
1
2004
... 树木对温度适应性策略的假设认为, 在寒冷的气候环境下, 根的分配比例较大、叶的分配比例较小(
Reich et al ., 2014 ), 本研究中, 7种杜鹃根生物量分配比例高达35.57%, 个别物种甚至超过40% (
图1 ), 高于全球绝大多数植物(
Poorter et al ., 2015 ), 也高于我国亚热带山地杜鹃灌丛根系生物量的分配比例(
张蔷等, 2017 ), 与一些高寒植被(
高巧等, 2014 ;
钟泽兵等, 2014 ;
杨路存等, 2018 ;
左有璐等, 2018 )相似, 低速率的养分循环和受限的土壤溶液运动驱动着高山、亚高山地区的低营养供应(
魏晶等, 2004 ;
Luo et al ., 2012 ;
Reich et al ., 2014 ), 这使得高寒杜鹃需提高根生物量分配比例以增强从寒冷土壤中吸收养分, 生物量向根系中的倾向分配支持最优分配假说.7种杜鹃叶生物量分配比例低于全球绝大多数植物(
Poorter et al ., 2015 ), 其适应机制也与一些高寒植物(
Reich et al ., 2014 )相似, 如
钟泽兵等(2014) 对青海玉树海拔3 715-4 296 m的5种建群灌木的研究发现叶生物量分配比例不足10%.甘肃南部高山-亚高山区的杜鹃会通过减少资源向叶片的分配, 保持常绿, 以较低的叶片周转率来适应寒冷环境.随着植株叶投资盈利能力的下降, 叶生物量将饱和, 投资会逐渐转向茎和根(
Poorter et al ., 2015 ), 而为增强植株的机械支撑, 必须增加茎的生物量投资(
Dybzinski et al ., 2011 ).本研究中7种高寒杜鹃茎生物量分配平均比例为45.61%, 接近于全球数据库平均值(
Poorter et al ., 2015 ).同时, 相比于叶和根, 茎生物量分配比例的种间差异最大(可达21%), 且多组物种间的差异显著(
表2 ), 这与
Poorter等(2015) 对全球1 200种植物整合分析的结果具有一致性, 茎不仅是植株的支撑、运输体系, 还在光竞争时向四周伸展小枝扩展空域, 而小枝的扩张又带来了高寒区防风的压力, 功能的多样性可能导致茎生物量分配的可塑性.各物种分配的差异还可能与茎构型有关, 如黄毛杜鹃和陇蜀杜鹃多为丛生, 茎枝扭曲, 其茎生物量分配比例的变异范围较大; 而美容杜鹃和山光杜鹃主干通直, 其茎生物量分配比例的变幅较小(
图1 ). ...
长白山高山冻原植被生物量的分布规律
1
2004
... 树木对温度适应性策略的假设认为, 在寒冷的气候环境下, 根的分配比例较大、叶的分配比例较小(
Reich et al ., 2014 ), 本研究中, 7种杜鹃根生物量分配比例高达35.57%, 个别物种甚至超过40% (
图1 ), 高于全球绝大多数植物(
Poorter et al ., 2015 ), 也高于我国亚热带山地杜鹃灌丛根系生物量的分配比例(
张蔷等, 2017 ), 与一些高寒植被(
高巧等, 2014 ;
钟泽兵等, 2014 ;
杨路存等, 2018 ;
左有璐等, 2018 )相似, 低速率的养分循环和受限的土壤溶液运动驱动着高山、亚高山地区的低营养供应(
魏晶等, 2004 ;
Luo et al ., 2012 ;
Reich et al ., 2014 ), 这使得高寒杜鹃需提高根生物量分配比例以增强从寒冷土壤中吸收养分, 生物量向根系中的倾向分配支持最优分配假说.7种杜鹃叶生物量分配比例低于全球绝大多数植物(
Poorter et al ., 2015 ), 其适应机制也与一些高寒植物(
Reich et al ., 2014 )相似, 如
钟泽兵等(2014) 对青海玉树海拔3 715-4 296 m的5种建群灌木的研究发现叶生物量分配比例不足10%.甘肃南部高山-亚高山区的杜鹃会通过减少资源向叶片的分配, 保持常绿, 以较低的叶片周转率来适应寒冷环境.随着植株叶投资盈利能力的下降, 叶生物量将饱和, 投资会逐渐转向茎和根(
Poorter et al ., 2015 ), 而为增强植株的机械支撑, 必须增加茎的生物量投资(
Dybzinski et al ., 2011 ).本研究中7种高寒杜鹃茎生物量分配平均比例为45.61%, 接近于全球数据库平均值(
Poorter et al ., 2015 ).同时, 相比于叶和根, 茎生物量分配比例的种间差异最大(可达21%), 且多组物种间的差异显著(
表2 ), 这与
Poorter等(2015) 对全球1 200种植物整合分析的结果具有一致性, 茎不仅是植株的支撑、运输体系, 还在光竞争时向四周伸展小枝扩展空域, 而小枝的扩张又带来了高寒区防风的压力, 功能的多样性可能导致茎生物量分配的可塑性.各物种分配的差异还可能与茎构型有关, 如黄毛杜鹃和陇蜀杜鹃多为丛生, 茎枝扭曲, 其茎生物量分配比例的变异范围较大; 而美容杜鹃和山光杜鹃主干通直, 其茎生物量分配比例的变幅较小(
图1 ). ...
Allocation, plasticity and allometry in plants
3
2004
... 生物量在不同器官间的分配是植物生态学研究的核心问题(
West et al .,1999 ;
Enquist & Niklas, 2002 ;
Poorter et al ., 2012 ).生物量分配模式不仅有助于从易测量的地上生物量估算地下生物量, 还体现着植物不同的生态策略(
Weiner, 2004 ), 反映出光合产物对植物不同器官(地上不同器官之间, 地上器官和地下器官之间)的投资以适应环境条件(
Niklas, 2006 ;
Poorter & Sack, 2012 ;
闫帮国等, 2016 ).最优分配假说认为, 植物倾向于将资源优先分配到能获得限制性资源的器官, 以帮助植物获取更多的受限资源, 从而维持最大生长速率(
Bloom et al ., 1985 ;
McCarthy & Enquist, 2007 ), 这意味着, 根据植物生长的环境, 如果光照受限, 生物量将倾向于叶和枝分配; 如果营养或水分受限, 生物量将倾向于根系分配(
Gleeson & Tilman, 1990 ).但植物生物量分配模式存在个体发育漂变(
Poorter & Sack, 2012 ;
Poorter et al ., 2015 ;
王杨等, 2017 ), 其分配比例会随植株的大小变化而变化(
Poorter et al ., 2015 ;
陈国鹏等, 2016 ), 使得最优分配假说的使用受到了限制, 而异速生长理论能较好地解决这一问题(
Weiner, 2004 ;
Gargaglione et al ., 2010 ), 且已被越来越多地应用到生物量分配研究中, 有****也曾对两种方法的优劣进行了比较(
McCarthy & Enquist, 2007 ;
Poorter & Sack, 2012 ), 更赞同将两种方法结合应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ). ...
... ), 使得最优分配假说的使用受到了限制, 而异速生长理论能较好地解决这一问题(
Weiner, 2004 ;
Gargaglione et al ., 2010 ), 且已被越来越多地应用到生物量分配研究中, 有****也曾对两种方法的优劣进行了比较(
McCarthy & Enquist, 2007 ;
Poorter & Sack, 2012 ), 更赞同将两种方法结合应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ). ...
... 异速生长理论预测小个体植物叶、茎、根器官生长可能为等速关系(
Enquist & Niklas, 2002 ;
Weiner, 2004 ;
Niklas, 2006 ), 这种基于种间变异建立的跨物种关系已在种内变异的研究中得到了较多应用(
Gargaglione et al ., 2010 ;
Cheng et al ., 2014 ;
陈国鹏等, 2016 ;
王杨等, 2017 ), 如
Cheng等(2014) 发现南亚热带生长的5种苗木叶、茎和根生物量的积累为等速或近似等速生长关系.本研究中, 7种高寒杜鹃叶与茎、叶与根以及茎与根之间既有异速生长表现, 也有等速生长表现(
图2 -5), 与荒漠植物(
杨昊天等, 2013 ;
陶冶和张元明, 2014 )、干热河谷草本植物(
闫帮国等, 2016 )的异速生长关系具有相似性, 不同于川西高寒草甸优势植物(
左有璐等, 2018 ), 并不完全支持小个体器官间等速生长关系的预测, 也不完全支持生态代谢理论的预测, 这可能是由于植物在适应特殊环境的过程中, 不同物种间具有很大的功能群差异, 这些功能群差异可能是不同器官生物量间异速生长关系分化的主要因素(
Poorter et al ., 2015 ;
闫帮国等, 2016 ), 而基于跨物种建立的生态代谢理论、小个体等速生长理论忽略了物种间的功能群差异. ...
A general model for the origin of allometric scaling laws in biology
1
1997
... 异速生长理论认为, 生物量的分配主要由植物的大小决定, 并可用幂函数
Y 1 =
βY 2 α 表示, 该方程经过对数转换后, 可线性化为log(
Y 1 ) = log(
β )
+ α log(
Y 2 ), 其中
Y 1 和
Y 2 是相互依赖的变量(如茎生物量和根生物量), log(
β )是异速生长常数,
α 是异速生长指数(
Niklas & Enquist, 2002a ,
2002b ).当
α = 1时, 模型反映的是等速生长关系; 当
α ≠ 1时, 模型反映的是异速生长关系(
Niklas & Enquist, 2002b ).生态代谢理论认为, 全球尺度上植物个体生物量具有恒定的异速生长指数, 其地上、地下生物量间均表现为异速生长关系, 叶生物量与茎生物量或地下生物量间的异速生长指数为0.75 (
West et al ., 1997 ;
Enquist & Niklas, 2002 ;
Niklas & Enquist, 2002 ), 但后续研究又证明小个体植物在叶生物量、茎生物量和地下生物量间也可能为等速关系(
Niklas, 2005 ;
Cheng et al ., 2014 ;
陈国鹏和赵文智, 2016 ),
Poorter等(2015) 进一步分析个体大小影响时发现, 植物叶与茎、叶与根、茎与根等生物量间的异速生长指数随个体增大而减小.尽管基于生态代谢理论的异速生长指数在不同尺度上得到了检验(
Cheng & Niklas, 2007 ), 但是该理论以关注种间的变异为主, 跨物种的恒定关系不一定适用于种内, 对物种作用的考虑不足(
Müller et al ., 2000 ;
Cheng et al ., 2014 ), 尤其对一些极端生境的物种关注较少也限制着该理论的普适性. ...
A general model for the structure and allometry of plant vascular systems
1
1999
... 生物量在不同器官间的分配是植物生态学研究的核心问题(
West et al .,1999 ;
Enquist & Niklas, 2002 ;
Poorter et al ., 2012 ).生物量分配模式不仅有助于从易测量的地上生物量估算地下生物量, 还体现着植物不同的生态策略(
Weiner, 2004 ), 反映出光合产物对植物不同器官(地上不同器官之间, 地上器官和地下器官之间)的投资以适应环境条件(
Niklas, 2006 ;
Poorter & Sack, 2012 ;
闫帮国等, 2016 ).最优分配假说认为, 植物倾向于将资源优先分配到能获得限制性资源的器官, 以帮助植物获取更多的受限资源, 从而维持最大生长速率(
Bloom et al ., 1985 ;
McCarthy & Enquist, 2007 ), 这意味着, 根据植物生长的环境, 如果光照受限, 生物量将倾向于叶和枝分配; 如果营养或水分受限, 生物量将倾向于根系分配(
Gleeson & Tilman, 1990 ).但植物生物量分配模式存在个体发育漂变(
Poorter & Sack, 2012 ;
Poorter et al ., 2015 ;
王杨等, 2017 ), 其分配比例会随植株的大小变化而变化(
Poorter et al ., 2015 ;
陈国鹏等, 2016 ), 使得最优分配假说的使用受到了限制, 而异速生长理论能较好地解决这一问题(
Weiner, 2004 ;
Gargaglione et al ., 2010 ), 且已被越来越多地应用到生物量分配研究中, 有****也曾对两种方法的优劣进行了比较(
McCarthy & Enquist, 2007 ;
Poorter & Sack, 2012 ), 更赞同将两种方法结合应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ). ...
青藏高原东缘野生暗紫贝母生物量分配格局对高山生态环境的适应
1
2013
... 理论上来说, 极端环境通过长期的筛选作用, 其适生物种响应资源约束时的生物量权衡策略应具有相似性(
Reich et al ., 2014 ).如, 在高寒山地, 高海拔、寒冷、短生长季和低气压等环境约束可能使植物的资源权衡具有独特表现(
高巧等, 2014 ;
左有璐等, 2014 ;
刘尊驰等, 2015 ), 植物会增加根生物量、降低叶生物量比例来适应较冷的环境(
徐波等, 2013 ).但目前的相关研究材料以常绿裸子植物或落叶被子植物为主(
Reich et al ., 2014 ;
Poorter et al ., 2015 ), 对高寒区生长的常绿被子植物的生物量权衡机制缺乏认知, 而常绿植物与落叶植物叶生物量分配策略存在差别, 常绿植物在叶片上投入的较多, 因此叶生物量分配比例高于落叶植物(
Poorter et al ., 2015 ). ...
青藏高原东缘野生暗紫贝母生物量分配格局对高山生态环境的适应
1
2013
... 理论上来说, 极端环境通过长期的筛选作用, 其适生物种响应资源约束时的生物量权衡策略应具有相似性(
Reich et al ., 2014 ).如, 在高寒山地, 高海拔、寒冷、短生长季和低气压等环境约束可能使植物的资源权衡具有独特表现(
高巧等, 2014 ;
左有璐等, 2014 ;
刘尊驰等, 2015 ), 植物会增加根生物量、降低叶生物量比例来适应较冷的环境(
徐波等, 2013 ).但目前的相关研究材料以常绿裸子植物或落叶被子植物为主(
Reich et al ., 2014 ;
Poorter et al ., 2015 ), 对高寒区生长的常绿被子植物的生物量权衡机制缺乏认知, 而常绿植物与落叶植物叶生物量分配策略存在差别, 常绿植物在叶片上投入的较多, 因此叶生物量分配比例高于落叶植物(
Poorter et al ., 2015 ). ...
干热河谷草本植物生物量分配及其对环境因子的响应
4
2016
... 生物量在不同器官间的分配是植物生态学研究的核心问题(
West et al .,1999 ;
Enquist & Niklas, 2002 ;
Poorter et al ., 2012 ).生物量分配模式不仅有助于从易测量的地上生物量估算地下生物量, 还体现着植物不同的生态策略(
Weiner, 2004 ), 反映出光合产物对植物不同器官(地上不同器官之间, 地上器官和地下器官之间)的投资以适应环境条件(
Niklas, 2006 ;
Poorter & Sack, 2012 ;
闫帮国等, 2016 ).最优分配假说认为, 植物倾向于将资源优先分配到能获得限制性资源的器官, 以帮助植物获取更多的受限资源, 从而维持最大生长速率(
Bloom et al ., 1985 ;
McCarthy & Enquist, 2007 ), 这意味着, 根据植物生长的环境, 如果光照受限, 生物量将倾向于叶和枝分配; 如果营养或水分受限, 生物量将倾向于根系分配(
Gleeson & Tilman, 1990 ).但植物生物量分配模式存在个体发育漂变(
Poorter & Sack, 2012 ;
Poorter et al ., 2015 ;
王杨等, 2017 ), 其分配比例会随植株的大小变化而变化(
Poorter et al ., 2015 ;
陈国鹏等, 2016 ), 使得最优分配假说的使用受到了限制, 而异速生长理论能较好地解决这一问题(
Weiner, 2004 ;
Gargaglione et al ., 2010 ), 且已被越来越多地应用到生物量分配研究中, 有****也曾对两种方法的优劣进行了比较(
McCarthy & Enquist, 2007 ;
Poorter & Sack, 2012 ), 更赞同将两种方法结合应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ). ...
... 异速生长理论预测小个体植物叶、茎、根器官生长可能为等速关系(
Enquist & Niklas, 2002 ;
Weiner, 2004 ;
Niklas, 2006 ), 这种基于种间变异建立的跨物种关系已在种内变异的研究中得到了较多应用(
Gargaglione et al ., 2010 ;
Cheng et al ., 2014 ;
陈国鹏等, 2016 ;
王杨等, 2017 ), 如
Cheng等(2014) 发现南亚热带生长的5种苗木叶、茎和根生物量的积累为等速或近似等速生长关系.本研究中, 7种高寒杜鹃叶与茎、叶与根以及茎与根之间既有异速生长表现, 也有等速生长表现(
图2 -5), 与荒漠植物(
杨昊天等, 2013 ;
陶冶和张元明, 2014 )、干热河谷草本植物(
闫帮国等, 2016 )的异速生长关系具有相似性, 不同于川西高寒草甸优势植物(
左有璐等, 2018 ), 并不完全支持小个体器官间等速生长关系的预测, 也不完全支持生态代谢理论的预测, 这可能是由于植物在适应特殊环境的过程中, 不同物种间具有很大的功能群差异, 这些功能群差异可能是不同器官生物量间异速生长关系分化的主要因素(
Poorter et al ., 2015 ;
闫帮国等, 2016 ), 而基于跨物种建立的生态代谢理论、小个体等速生长理论忽略了物种间的功能群差异. ...
... ;
闫帮国等, 2016 ), 而基于跨物种建立的生态代谢理论、小个体等速生长理论忽略了物种间的功能群差异. ...
... 物种是影响植物器官生物量分配异速生长关系的重要因素(
Poorter et al ., 2012 ;
闫帮国等, 2016 ;
张蔷等, 2017 ), 基于大样本的研究则发现生物量分配比例与物种的系统发育有关, 如裸子植物叶片上的投入更多(
Reich et al ., 2014 ;
Poorter et al ., 2015 ).本研究中7种杜鹃隶属于同科同属, 其生物量分配比例及异速生长关系的种间差异显著(
表2 ,
表3 ), 陇蜀杜鹃根生物量分配比例较低, 美容杜鹃叶生物量分配比例较高而茎生物量分配比例较低, 黄毛杜鹃、头花杜鹃地上生物量与地下生物量呈等速生长, 陇蜀杜鹃、麻花杜鹃地上生物量的积累速率大于地下, 其余3种地上生物量积累速率小于地下(
图5 ), 而叶与茎、叶与根、茎与根间的异速生长关系的物种差异更为复杂多样, 如黄毛杜鹃、陇蜀杜鹃、美容杜鹃和山光杜鹃的叶与茎, 所有物种的叶与根, 美容杜鹃、太白杜鹃、黄毛杜鹃、陇蜀杜鹃和麻花杜鹃的茎与根为异速生长关系, 而太白杜鹃、头花杜鹃和麻花杜鹃的叶与茎, 山光杜鹃和头花杜鹃的茎与根为等速生长关系(
图2 -4).这也说明不同的物种可能在进化过程中会通过生态位分化而占有不同的环境, 在本研究区域, 头花杜鹃主要分布于海拔3 500 m以上并形成单优群落或与陇蜀杜鹃形成共生群落; 陇蜀杜鹃、黄毛杜鹃在海拔3 000-3 700 m间有分布, 但在3 200 m以上才形成单优群落或群落优势种; 山光杜鹃和麻花杜鹃海拔分布范围跨度大(2 300-3 500 m), 但呈群落优势种的仅见3 000 m处个别的冷杉皆伐更新迹地,
Veresoglou和 Pe?uelas (2019) 分析欧亚大陆植物生物量时发现分配的变异强度与物种分布范围有密切的关系, 但杜鹃分布范围与生物量分配特征间是否存在相关关系, 尚需持续研究. ...
干热河谷草本植物生物量分配及其对环境因子的响应
4
2016
... 生物量在不同器官间的分配是植物生态学研究的核心问题(
West et al .,1999 ;
Enquist & Niklas, 2002 ;
Poorter et al ., 2012 ).生物量分配模式不仅有助于从易测量的地上生物量估算地下生物量, 还体现着植物不同的生态策略(
Weiner, 2004 ), 反映出光合产物对植物不同器官(地上不同器官之间, 地上器官和地下器官之间)的投资以适应环境条件(
Niklas, 2006 ;
Poorter & Sack, 2012 ;
闫帮国等, 2016 ).最优分配假说认为, 植物倾向于将资源优先分配到能获得限制性资源的器官, 以帮助植物获取更多的受限资源, 从而维持最大生长速率(
Bloom et al ., 1985 ;
McCarthy & Enquist, 2007 ), 这意味着, 根据植物生长的环境, 如果光照受限, 生物量将倾向于叶和枝分配; 如果营养或水分受限, 生物量将倾向于根系分配(
Gleeson & Tilman, 1990 ).但植物生物量分配模式存在个体发育漂变(
Poorter & Sack, 2012 ;
Poorter et al ., 2015 ;
王杨等, 2017 ), 其分配比例会随植株的大小变化而变化(
Poorter et al ., 2015 ;
陈国鹏等, 2016 ), 使得最优分配假说的使用受到了限制, 而异速生长理论能较好地解决这一问题(
Weiner, 2004 ;
Gargaglione et al ., 2010 ), 且已被越来越多地应用到生物量分配研究中, 有****也曾对两种方法的优劣进行了比较(
McCarthy & Enquist, 2007 ;
Poorter & Sack, 2012 ), 更赞同将两种方法结合应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ). ...
... 异速生长理论预测小个体植物叶、茎、根器官生长可能为等速关系(
Enquist & Niklas, 2002 ;
Weiner, 2004 ;
Niklas, 2006 ), 这种基于种间变异建立的跨物种关系已在种内变异的研究中得到了较多应用(
Gargaglione et al ., 2010 ;
Cheng et al ., 2014 ;
陈国鹏等, 2016 ;
王杨等, 2017 ), 如
Cheng等(2014) 发现南亚热带生长的5种苗木叶、茎和根生物量的积累为等速或近似等速生长关系.本研究中, 7种高寒杜鹃叶与茎、叶与根以及茎与根之间既有异速生长表现, 也有等速生长表现(
图2 -5), 与荒漠植物(
杨昊天等, 2013 ;
陶冶和张元明, 2014 )、干热河谷草本植物(
闫帮国等, 2016 )的异速生长关系具有相似性, 不同于川西高寒草甸优势植物(
左有璐等, 2018 ), 并不完全支持小个体器官间等速生长关系的预测, 也不完全支持生态代谢理论的预测, 这可能是由于植物在适应特殊环境的过程中, 不同物种间具有很大的功能群差异, 这些功能群差异可能是不同器官生物量间异速生长关系分化的主要因素(
Poorter et al ., 2015 ;
闫帮国等, 2016 ), 而基于跨物种建立的生态代谢理论、小个体等速生长理论忽略了物种间的功能群差异. ...
... ;
闫帮国等, 2016 ), 而基于跨物种建立的生态代谢理论、小个体等速生长理论忽略了物种间的功能群差异. ...
... 物种是影响植物器官生物量分配异速生长关系的重要因素(
Poorter et al ., 2012 ;
闫帮国等, 2016 ;
张蔷等, 2017 ), 基于大样本的研究则发现生物量分配比例与物种的系统发育有关, 如裸子植物叶片上的投入更多(
Reich et al ., 2014 ;
Poorter et al ., 2015 ).本研究中7种杜鹃隶属于同科同属, 其生物量分配比例及异速生长关系的种间差异显著(
表2 ,
表3 ), 陇蜀杜鹃根生物量分配比例较低, 美容杜鹃叶生物量分配比例较高而茎生物量分配比例较低, 黄毛杜鹃、头花杜鹃地上生物量与地下生物量呈等速生长, 陇蜀杜鹃、麻花杜鹃地上生物量的积累速率大于地下, 其余3种地上生物量积累速率小于地下(
图5 ), 而叶与茎、叶与根、茎与根间的异速生长关系的物种差异更为复杂多样, 如黄毛杜鹃、陇蜀杜鹃、美容杜鹃和山光杜鹃的叶与茎, 所有物种的叶与根, 美容杜鹃、太白杜鹃、黄毛杜鹃、陇蜀杜鹃和麻花杜鹃的茎与根为异速生长关系, 而太白杜鹃、头花杜鹃和麻花杜鹃的叶与茎, 山光杜鹃和头花杜鹃的茎与根为等速生长关系(
图2 -4).这也说明不同的物种可能在进化过程中会通过生态位分化而占有不同的环境, 在本研究区域, 头花杜鹃主要分布于海拔3 500 m以上并形成单优群落或与陇蜀杜鹃形成共生群落; 陇蜀杜鹃、黄毛杜鹃在海拔3 000-3 700 m间有分布, 但在3 200 m以上才形成单优群落或群落优势种; 山光杜鹃和麻花杜鹃海拔分布范围跨度大(2 300-3 500 m), 但呈群落优势种的仅见3 000 m处个别的冷杉皆伐更新迹地,
Veresoglou和 Pe?uelas (2019) 分析欧亚大陆植物生物量时发现分配的变异强度与物种分布范围有密切的关系, 但杜鹃分布范围与生物量分配特征间是否存在相关关系, 尚需持续研究. ...
荒漠草地4种灌木生物量分配特征
3
2013
... 生物量在不同器官间的分配是植物生态学研究的核心问题(
West et al .,1999 ;
Enquist & Niklas, 2002 ;
Poorter et al ., 2012 ).生物量分配模式不仅有助于从易测量的地上生物量估算地下生物量, 还体现着植物不同的生态策略(
Weiner, 2004 ), 反映出光合产物对植物不同器官(地上不同器官之间, 地上器官和地下器官之间)的投资以适应环境条件(
Niklas, 2006 ;
Poorter & Sack, 2012 ;
闫帮国等, 2016 ).最优分配假说认为, 植物倾向于将资源优先分配到能获得限制性资源的器官, 以帮助植物获取更多的受限资源, 从而维持最大生长速率(
Bloom et al ., 1985 ;
McCarthy & Enquist, 2007 ), 这意味着, 根据植物生长的环境, 如果光照受限, 生物量将倾向于叶和枝分配; 如果营养或水分受限, 生物量将倾向于根系分配(
Gleeson & Tilman, 1990 ).但植物生物量分配模式存在个体发育漂变(
Poorter & Sack, 2012 ;
Poorter et al ., 2015 ;
王杨等, 2017 ), 其分配比例会随植株的大小变化而变化(
Poorter et al ., 2015 ;
陈国鹏等, 2016 ), 使得最优分配假说的使用受到了限制, 而异速生长理论能较好地解决这一问题(
Weiner, 2004 ;
Gargaglione et al ., 2010 ), 且已被越来越多地应用到生物量分配研究中, 有****也曾对两种方法的优劣进行了比较(
McCarthy & Enquist, 2007 ;
Poorter & Sack, 2012 ), 更赞同将两种方法结合应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ). ...
... 尽管异速生长分析不受个体大小等因素干扰, 可以较好地检验不同器官生长发育协调性, 并确定这种协调性在物种间的变化或对环境的响应, 但在一些生态模型的构建研究中, 叶、茎和根分配比例的实际值要比器官间的协调性重要(
Poorter & Sack, 2012 ), 考虑到不同分析方法在研究生物量分配中的局限性, 将异速生长法与最优分配假说结合已在许多研究中得到了应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ;
陈国鹏等, 2016 ).本研究也结合最优分配假说与异速生长分析来研究甘肃南部高山-亚高山区杜鹃地上、地下器官生物量的分配特征, 结果表明7种高寒常绿杜鹃根、茎、叶生物量分配比例和异速生长关系受物种影响显著, 高寒杜鹃对特殊生境适应过程中的生物量分配模式有许多相似之处, 而物种差异又导致相似生境中生物量分配模式的分异. ...
... 异速生长理论预测小个体植物叶、茎、根器官生长可能为等速关系(
Enquist & Niklas, 2002 ;
Weiner, 2004 ;
Niklas, 2006 ), 这种基于种间变异建立的跨物种关系已在种内变异的研究中得到了较多应用(
Gargaglione et al ., 2010 ;
Cheng et al ., 2014 ;
陈国鹏等, 2016 ;
王杨等, 2017 ), 如
Cheng等(2014) 发现南亚热带生长的5种苗木叶、茎和根生物量的积累为等速或近似等速生长关系.本研究中, 7种高寒杜鹃叶与茎、叶与根以及茎与根之间既有异速生长表现, 也有等速生长表现(
图2 -5), 与荒漠植物(
杨昊天等, 2013 ;
陶冶和张元明, 2014 )、干热河谷草本植物(
闫帮国等, 2016 )的异速生长关系具有相似性, 不同于川西高寒草甸优势植物(
左有璐等, 2018 ), 并不完全支持小个体器官间等速生长关系的预测, 也不完全支持生态代谢理论的预测, 这可能是由于植物在适应特殊环境的过程中, 不同物种间具有很大的功能群差异, 这些功能群差异可能是不同器官生物量间异速生长关系分化的主要因素(
Poorter et al ., 2015 ;
闫帮国等, 2016 ), 而基于跨物种建立的生态代谢理论、小个体等速生长理论忽略了物种间的功能群差异. ...
荒漠草地4种灌木生物量分配特征
3
2013
... 生物量在不同器官间的分配是植物生态学研究的核心问题(
West et al .,1999 ;
Enquist & Niklas, 2002 ;
Poorter et al ., 2012 ).生物量分配模式不仅有助于从易测量的地上生物量估算地下生物量, 还体现着植物不同的生态策略(
Weiner, 2004 ), 反映出光合产物对植物不同器官(地上不同器官之间, 地上器官和地下器官之间)的投资以适应环境条件(
Niklas, 2006 ;
Poorter & Sack, 2012 ;
闫帮国等, 2016 ).最优分配假说认为, 植物倾向于将资源优先分配到能获得限制性资源的器官, 以帮助植物获取更多的受限资源, 从而维持最大生长速率(
Bloom et al ., 1985 ;
McCarthy & Enquist, 2007 ), 这意味着, 根据植物生长的环境, 如果光照受限, 生物量将倾向于叶和枝分配; 如果营养或水分受限, 生物量将倾向于根系分配(
Gleeson & Tilman, 1990 ).但植物生物量分配模式存在个体发育漂变(
Poorter & Sack, 2012 ;
Poorter et al ., 2015 ;
王杨等, 2017 ), 其分配比例会随植株的大小变化而变化(
Poorter et al ., 2015 ;
陈国鹏等, 2016 ), 使得最优分配假说的使用受到了限制, 而异速生长理论能较好地解决这一问题(
Weiner, 2004 ;
Gargaglione et al ., 2010 ), 且已被越来越多地应用到生物量分配研究中, 有****也曾对两种方法的优劣进行了比较(
McCarthy & Enquist, 2007 ;
Poorter & Sack, 2012 ), 更赞同将两种方法结合应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ). ...
... 尽管异速生长分析不受个体大小等因素干扰, 可以较好地检验不同器官生长发育协调性, 并确定这种协调性在物种间的变化或对环境的响应, 但在一些生态模型的构建研究中, 叶、茎和根分配比例的实际值要比器官间的协调性重要(
Poorter & Sack, 2012 ), 考虑到不同分析方法在研究生物量分配中的局限性, 将异速生长法与最优分配假说结合已在许多研究中得到了应用(
Poorter & Sack, 2012 ;
Poorter et al ., 2012 ;
杨昊天等, 2013 ;
陈国鹏等, 2016 ).本研究也结合最优分配假说与异速生长分析来研究甘肃南部高山-亚高山区杜鹃地上、地下器官生物量的分配特征, 结果表明7种高寒常绿杜鹃根、茎、叶生物量分配比例和异速生长关系受物种影响显著, 高寒杜鹃对特殊生境适应过程中的生物量分配模式有许多相似之处, 而物种差异又导致相似生境中生物量分配模式的分异. ...
... 异速生长理论预测小个体植物叶、茎、根器官生长可能为等速关系(
Enquist & Niklas, 2002 ;
Weiner, 2004 ;
Niklas, 2006 ), 这种基于种间变异建立的跨物种关系已在种内变异的研究中得到了较多应用(
Gargaglione et al ., 2010 ;
Cheng et al ., 2014 ;
陈国鹏等, 2016 ;
王杨等, 2017 ), 如
Cheng等(2014) 发现南亚热带生长的5种苗木叶、茎和根生物量的积累为等速或近似等速生长关系.本研究中, 7种高寒杜鹃叶与茎、叶与根以及茎与根之间既有异速生长表现, 也有等速生长表现(
图2 -5), 与荒漠植物(
杨昊天等, 2013 ;
陶冶和张元明, 2014 )、干热河谷草本植物(
闫帮国等, 2016 )的异速生长关系具有相似性, 不同于川西高寒草甸优势植物(
左有璐等, 2018 ), 并不完全支持小个体器官间等速生长关系的预测, 也不完全支持生态代谢理论的预测, 这可能是由于植物在适应特殊环境的过程中, 不同物种间具有很大的功能群差异, 这些功能群差异可能是不同器官生物量间异速生长关系分化的主要因素(
Poorter et al ., 2015 ;
闫帮国等, 2016 ), 而基于跨物种建立的生态代谢理论、小个体等速生长理论忽略了物种间的功能群差异. ...
青海省高寒灌丛物种多样性、生物量及其关系
2
2018
... 广布于我国青藏高原及其边缘山地的杜鹃(
Rhododendron spp.)常绿灌丛是亚高山暗针叶林向高山草甸过渡的群落类型, 在维持高山-亚高山生态系统稳定等方面具有关键作用(
钟泽兵等, 2014 ;
杨路存等, 2018 ).本研究以甘肃南部高山-亚高山区常见的7种杜鹃为材料, 通过全株收获法测定根、茎和叶的生物量, 采用标准化主轴法估计检验各器官间的异速生长关系, 并进行种间差异比较, 以期了解高寒杜鹃的生物量权衡策略, 为进一步认识高寒植物规避风险、提高适应性提供理论基础. ...
... 树木对温度适应性策略的假设认为, 在寒冷的气候环境下, 根的分配比例较大、叶的分配比例较小(
Reich et al ., 2014 ), 本研究中, 7种杜鹃根生物量分配比例高达35.57%, 个别物种甚至超过40% (
图1 ), 高于全球绝大多数植物(
Poorter et al ., 2015 ), 也高于我国亚热带山地杜鹃灌丛根系生物量的分配比例(
张蔷等, 2017 ), 与一些高寒植被(
高巧等, 2014 ;
钟泽兵等, 2014 ;
杨路存等, 2018 ;
左有璐等, 2018 )相似, 低速率的养分循环和受限的土壤溶液运动驱动着高山、亚高山地区的低营养供应(
魏晶等, 2004 ;
Luo et al ., 2012 ;
Reich et al ., 2014 ), 这使得高寒杜鹃需提高根生物量分配比例以增强从寒冷土壤中吸收养分, 生物量向根系中的倾向分配支持最优分配假说.7种杜鹃叶生物量分配比例低于全球绝大多数植物(
Poorter et al ., 2015 ), 其适应机制也与一些高寒植物(
Reich et al ., 2014 )相似, 如
钟泽兵等(2014) 对青海玉树海拔3 715-4 296 m的5种建群灌木的研究发现叶生物量分配比例不足10%.甘肃南部高山-亚高山区的杜鹃会通过减少资源向叶片的分配, 保持常绿, 以较低的叶片周转率来适应寒冷环境.随着植株叶投资盈利能力的下降, 叶生物量将饱和, 投资会逐渐转向茎和根(
Poorter et al ., 2015 ), 而为增强植株的机械支撑, 必须增加茎的生物量投资(
Dybzinski et al ., 2011 ).本研究中7种高寒杜鹃茎生物量分配平均比例为45.61%, 接近于全球数据库平均值(
Poorter et al ., 2015 ).同时, 相比于叶和根, 茎生物量分配比例的种间差异最大(可达21%), 且多组物种间的差异显著(
表2 ), 这与
Poorter等(2015) 对全球1 200种植物整合分析的结果具有一致性, 茎不仅是植株的支撑、运输体系, 还在光竞争时向四周伸展小枝扩展空域, 而小枝的扩张又带来了高寒区防风的压力, 功能的多样性可能导致茎生物量分配的可塑性.各物种分配的差异还可能与茎构型有关, 如黄毛杜鹃和陇蜀杜鹃多为丛生, 茎枝扭曲, 其茎生物量分配比例的变异范围较大; 而美容杜鹃和山光杜鹃主干通直, 其茎生物量分配比例的变幅较小(
图1 ). ...
青海省高寒灌丛物种多样性、生物量及其关系
2
2018
... 广布于我国青藏高原及其边缘山地的杜鹃(
Rhododendron spp.)常绿灌丛是亚高山暗针叶林向高山草甸过渡的群落类型, 在维持高山-亚高山生态系统稳定等方面具有关键作用(
钟泽兵等, 2014 ;
杨路存等, 2018 ).本研究以甘肃南部高山-亚高山区常见的7种杜鹃为材料, 通过全株收获法测定根、茎和叶的生物量, 采用标准化主轴法估计检验各器官间的异速生长关系, 并进行种间差异比较, 以期了解高寒杜鹃的生物量权衡策略, 为进一步认识高寒植物规避风险、提高适应性提供理论基础. ...
... 树木对温度适应性策略的假设认为, 在寒冷的气候环境下, 根的分配比例较大、叶的分配比例较小(
Reich et al ., 2014 ), 本研究中, 7种杜鹃根生物量分配比例高达35.57%, 个别物种甚至超过40% (
图1 ), 高于全球绝大多数植物(
Poorter et al ., 2015 ), 也高于我国亚热带山地杜鹃灌丛根系生物量的分配比例(
张蔷等, 2017 ), 与一些高寒植被(
高巧等, 2014 ;
钟泽兵等, 2014 ;
杨路存等, 2018 ;
左有璐等, 2018 )相似, 低速率的养分循环和受限的土壤溶液运动驱动着高山、亚高山地区的低营养供应(
魏晶等, 2004 ;
Luo et al ., 2012 ;
Reich et al ., 2014 ), 这使得高寒杜鹃需提高根生物量分配比例以增强从寒冷土壤中吸收养分, 生物量向根系中的倾向分配支持最优分配假说.7种杜鹃叶生物量分配比例低于全球绝大多数植物(
Poorter et al ., 2015 ), 其适应机制也与一些高寒植物(
Reich et al ., 2014 )相似, 如
钟泽兵等(2014) 对青海玉树海拔3 715-4 296 m的5种建群灌木的研究发现叶生物量分配比例不足10%.甘肃南部高山-亚高山区的杜鹃会通过减少资源向叶片的分配, 保持常绿, 以较低的叶片周转率来适应寒冷环境.随着植株叶投资盈利能力的下降, 叶生物量将饱和, 投资会逐渐转向茎和根(
Poorter et al ., 2015 ), 而为增强植株的机械支撑, 必须增加茎的生物量投资(
Dybzinski et al ., 2011 ).本研究中7种高寒杜鹃茎生物量分配平均比例为45.61%, 接近于全球数据库平均值(
Poorter et al ., 2015 ).同时, 相比于叶和根, 茎生物量分配比例的种间差异最大(可达21%), 且多组物种间的差异显著(
表2 ), 这与
Poorter等(2015) 对全球1 200种植物整合分析的结果具有一致性, 茎不仅是植株的支撑、运输体系, 还在光竞争时向四周伸展小枝扩展空域, 而小枝的扩张又带来了高寒区防风的压力, 功能的多样性可能导致茎生物量分配的可塑性.各物种分配的差异还可能与茎构型有关, 如黄毛杜鹃和陇蜀杜鹃多为丛生, 茎枝扭曲, 其茎生物量分配比例的变异范围较大; 而美容杜鹃和山光杜鹃主干通直, 其茎生物量分配比例的变幅较小(
图1 ). ...
中国亚热带山地杜鹃灌丛生物量分配及其碳密度估算
2
2017
... 树木对温度适应性策略的假设认为, 在寒冷的气候环境下, 根的分配比例较大、叶的分配比例较小(
Reich et al ., 2014 ), 本研究中, 7种杜鹃根生物量分配比例高达35.57%, 个别物种甚至超过40% (
图1 ), 高于全球绝大多数植物(
Poorter et al ., 2015 ), 也高于我国亚热带山地杜鹃灌丛根系生物量的分配比例(
张蔷等, 2017 ), 与一些高寒植被(
高巧等, 2014 ;
钟泽兵等, 2014 ;
杨路存等, 2018 ;
左有璐等, 2018 )相似, 低速率的养分循环和受限的土壤溶液运动驱动着高山、亚高山地区的低营养供应(
魏晶等, 2004 ;
Luo et al ., 2012 ;
Reich et al ., 2014 ), 这使得高寒杜鹃需提高根生物量分配比例以增强从寒冷土壤中吸收养分, 生物量向根系中的倾向分配支持最优分配假说.7种杜鹃叶生物量分配比例低于全球绝大多数植物(
Poorter et al ., 2015 ), 其适应机制也与一些高寒植物(
Reich et al ., 2014 )相似, 如
钟泽兵等(2014) 对青海玉树海拔3 715-4 296 m的5种建群灌木的研究发现叶生物量分配比例不足10%.甘肃南部高山-亚高山区的杜鹃会通过减少资源向叶片的分配, 保持常绿, 以较低的叶片周转率来适应寒冷环境.随着植株叶投资盈利能力的下降, 叶生物量将饱和, 投资会逐渐转向茎和根(
Poorter et al ., 2015 ), 而为增强植株的机械支撑, 必须增加茎的生物量投资(
Dybzinski et al ., 2011 ).本研究中7种高寒杜鹃茎生物量分配平均比例为45.61%, 接近于全球数据库平均值(
Poorter et al ., 2015 ).同时, 相比于叶和根, 茎生物量分配比例的种间差异最大(可达21%), 且多组物种间的差异显著(
表2 ), 这与
Poorter等(2015) 对全球1 200种植物整合分析的结果具有一致性, 茎不仅是植株的支撑、运输体系, 还在光竞争时向四周伸展小枝扩展空域, 而小枝的扩张又带来了高寒区防风的压力, 功能的多样性可能导致茎生物量分配的可塑性.各物种分配的差异还可能与茎构型有关, 如黄毛杜鹃和陇蜀杜鹃多为丛生, 茎枝扭曲, 其茎生物量分配比例的变异范围较大; 而美容杜鹃和山光杜鹃主干通直, 其茎生物量分配比例的变幅较小(
图1 ). ...
... 物种是影响植物器官生物量分配异速生长关系的重要因素(
Poorter et al ., 2012 ;
闫帮国等, 2016 ;
张蔷等, 2017 ), 基于大样本的研究则发现生物量分配比例与物种的系统发育有关, 如裸子植物叶片上的投入更多(
Reich et al ., 2014 ;
Poorter et al ., 2015 ).本研究中7种杜鹃隶属于同科同属, 其生物量分配比例及异速生长关系的种间差异显著(
表2 ,
表3 ), 陇蜀杜鹃根生物量分配比例较低, 美容杜鹃叶生物量分配比例较高而茎生物量分配比例较低, 黄毛杜鹃、头花杜鹃地上生物量与地下生物量呈等速生长, 陇蜀杜鹃、麻花杜鹃地上生物量的积累速率大于地下, 其余3种地上生物量积累速率小于地下(
图5 ), 而叶与茎、叶与根、茎与根间的异速生长关系的物种差异更为复杂多样, 如黄毛杜鹃、陇蜀杜鹃、美容杜鹃和山光杜鹃的叶与茎, 所有物种的叶与根, 美容杜鹃、太白杜鹃、黄毛杜鹃、陇蜀杜鹃和麻花杜鹃的茎与根为异速生长关系, 而太白杜鹃、头花杜鹃和麻花杜鹃的叶与茎, 山光杜鹃和头花杜鹃的茎与根为等速生长关系(
图2 -4).这也说明不同的物种可能在进化过程中会通过生态位分化而占有不同的环境, 在本研究区域, 头花杜鹃主要分布于海拔3 500 m以上并形成单优群落或与陇蜀杜鹃形成共生群落; 陇蜀杜鹃、黄毛杜鹃在海拔3 000-3 700 m间有分布, 但在3 200 m以上才形成单优群落或群落优势种; 山光杜鹃和麻花杜鹃海拔分布范围跨度大(2 300-3 500 m), 但呈群落优势种的仅见3 000 m处个别的冷杉皆伐更新迹地,
Veresoglou和 Pe?uelas (2019) 分析欧亚大陆植物生物量时发现分配的变异强度与物种分布范围有密切的关系, 但杜鹃分布范围与生物量分配特征间是否存在相关关系, 尚需持续研究. ...
中国亚热带山地杜鹃灌丛生物量分配及其碳密度估算
2
2017
... 树木对温度适应性策略的假设认为, 在寒冷的气候环境下, 根的分配比例较大、叶的分配比例较小(
Reich et al ., 2014 ), 本研究中, 7种杜鹃根生物量分配比例高达35.57%, 个别物种甚至超过40% (
图1 ), 高于全球绝大多数植物(
Poorter et al ., 2015 ), 也高于我国亚热带山地杜鹃灌丛根系生物量的分配比例(
张蔷等, 2017 ), 与一些高寒植被(
高巧等, 2014 ;
钟泽兵等, 2014 ;
杨路存等, 2018 ;
左有璐等, 2018 )相似, 低速率的养分循环和受限的土壤溶液运动驱动着高山、亚高山地区的低营养供应(
魏晶等, 2004 ;
Luo et al ., 2012 ;
Reich et al ., 2014 ), 这使得高寒杜鹃需提高根生物量分配比例以增强从寒冷土壤中吸收养分, 生物量向根系中的倾向分配支持最优分配假说.7种杜鹃叶生物量分配比例低于全球绝大多数植物(
Poorter et al ., 2015 ), 其适应机制也与一些高寒植物(
Reich et al ., 2014 )相似, 如
钟泽兵等(2014) 对青海玉树海拔3 715-4 296 m的5种建群灌木的研究发现叶生物量分配比例不足10%.甘肃南部高山-亚高山区的杜鹃会通过减少资源向叶片的分配, 保持常绿, 以较低的叶片周转率来适应寒冷环境.随着植株叶投资盈利能力的下降, 叶生物量将饱和, 投资会逐渐转向茎和根(
Poorter et al ., 2015 ), 而为增强植株的机械支撑, 必须增加茎的生物量投资(
Dybzinski et al ., 2011 ).本研究中7种高寒杜鹃茎生物量分配平均比例为45.61%, 接近于全球数据库平均值(
Poorter et al ., 2015 ).同时, 相比于叶和根, 茎生物量分配比例的种间差异最大(可达21%), 且多组物种间的差异显著(
表2 ), 这与
Poorter等(2015) 对全球1 200种植物整合分析的结果具有一致性, 茎不仅是植株的支撑、运输体系, 还在光竞争时向四周伸展小枝扩展空域, 而小枝的扩张又带来了高寒区防风的压力, 功能的多样性可能导致茎生物量分配的可塑性.各物种分配的差异还可能与茎构型有关, 如黄毛杜鹃和陇蜀杜鹃多为丛生, 茎枝扭曲, 其茎生物量分配比例的变异范围较大; 而美容杜鹃和山光杜鹃主干通直, 其茎生物量分配比例的变幅较小(
图1 ). ...
... 物种是影响植物器官生物量分配异速生长关系的重要因素(
Poorter et al ., 2012 ;
闫帮国等, 2016 ;
张蔷等, 2017 ), 基于大样本的研究则发现生物量分配比例与物种的系统发育有关, 如裸子植物叶片上的投入更多(
Reich et al ., 2014 ;
Poorter et al ., 2015 ).本研究中7种杜鹃隶属于同科同属, 其生物量分配比例及异速生长关系的种间差异显著(
表2 ,
表3 ), 陇蜀杜鹃根生物量分配比例较低, 美容杜鹃叶生物量分配比例较高而茎生物量分配比例较低, 黄毛杜鹃、头花杜鹃地上生物量与地下生物量呈等速生长, 陇蜀杜鹃、麻花杜鹃地上生物量的积累速率大于地下, 其余3种地上生物量积累速率小于地下(
图5 ), 而叶与茎、叶与根、茎与根间的异速生长关系的物种差异更为复杂多样, 如黄毛杜鹃、陇蜀杜鹃、美容杜鹃和山光杜鹃的叶与茎, 所有物种的叶与根, 美容杜鹃、太白杜鹃、黄毛杜鹃、陇蜀杜鹃和麻花杜鹃的茎与根为异速生长关系, 而太白杜鹃、头花杜鹃和麻花杜鹃的叶与茎, 山光杜鹃和头花杜鹃的茎与根为等速生长关系(
图2 -4).这也说明不同的物种可能在进化过程中会通过生态位分化而占有不同的环境, 在本研究区域, 头花杜鹃主要分布于海拔3 500 m以上并形成单优群落或与陇蜀杜鹃形成共生群落; 陇蜀杜鹃、黄毛杜鹃在海拔3 000-3 700 m间有分布, 但在3 200 m以上才形成单优群落或群落优势种; 山光杜鹃和麻花杜鹃海拔分布范围跨度大(2 300-3 500 m), 但呈群落优势种的仅见3 000 m处个别的冷杉皆伐更新迹地,
Veresoglou和 Pe?uelas (2019) 分析欧亚大陆植物生物量时发现分配的变异强度与物种分布范围有密切的关系, 但杜鹃分布范围与生物量分配特征间是否存在相关关系, 尚需持续研究. ...
青海玉树地区主要灌丛类型地上生物量及其影响因素
3
2014
... 广布于我国青藏高原及其边缘山地的杜鹃(
Rhododendron spp.)常绿灌丛是亚高山暗针叶林向高山草甸过渡的群落类型, 在维持高山-亚高山生态系统稳定等方面具有关键作用(
钟泽兵等, 2014 ;
杨路存等, 2018 ).本研究以甘肃南部高山-亚高山区常见的7种杜鹃为材料, 通过全株收获法测定根、茎和叶的生物量, 采用标准化主轴法估计检验各器官间的异速生长关系, 并进行种间差异比较, 以期了解高寒杜鹃的生物量权衡策略, 为进一步认识高寒植物规避风险、提高适应性提供理论基础. ...
... 树木对温度适应性策略的假设认为, 在寒冷的气候环境下, 根的分配比例较大、叶的分配比例较小(
Reich et al ., 2014 ), 本研究中, 7种杜鹃根生物量分配比例高达35.57%, 个别物种甚至超过40% (
图1 ), 高于全球绝大多数植物(
Poorter et al ., 2015 ), 也高于我国亚热带山地杜鹃灌丛根系生物量的分配比例(
张蔷等, 2017 ), 与一些高寒植被(
高巧等, 2014 ;
钟泽兵等, 2014 ;
杨路存等, 2018 ;
左有璐等, 2018 )相似, 低速率的养分循环和受限的土壤溶液运动驱动着高山、亚高山地区的低营养供应(
魏晶等, 2004 ;
Luo et al ., 2012 ;
Reich et al ., 2014 ), 这使得高寒杜鹃需提高根生物量分配比例以增强从寒冷土壤中吸收养分, 生物量向根系中的倾向分配支持最优分配假说.7种杜鹃叶生物量分配比例低于全球绝大多数植物(
Poorter et al ., 2015 ), 其适应机制也与一些高寒植物(
Reich et al ., 2014 )相似, 如
钟泽兵等(2014) 对青海玉树海拔3 715-4 296 m的5种建群灌木的研究发现叶生物量分配比例不足10%.甘肃南部高山-亚高山区的杜鹃会通过减少资源向叶片的分配, 保持常绿, 以较低的叶片周转率来适应寒冷环境.随着植株叶投资盈利能力的下降, 叶生物量将饱和, 投资会逐渐转向茎和根(
Poorter et al ., 2015 ), 而为增强植株的机械支撑, 必须增加茎的生物量投资(
Dybzinski et al ., 2011 ).本研究中7种高寒杜鹃茎生物量分配平均比例为45.61%, 接近于全球数据库平均值(
Poorter et al ., 2015 ).同时, 相比于叶和根, 茎生物量分配比例的种间差异最大(可达21%), 且多组物种间的差异显著(
表2 ), 这与
Poorter等(2015) 对全球1 200种植物整合分析的结果具有一致性, 茎不仅是植株的支撑、运输体系, 还在光竞争时向四周伸展小枝扩展空域, 而小枝的扩张又带来了高寒区防风的压力, 功能的多样性可能导致茎生物量分配的可塑性.各物种分配的差异还可能与茎构型有关, 如黄毛杜鹃和陇蜀杜鹃多为丛生, 茎枝扭曲, 其茎生物量分配比例的变异范围较大; 而美容杜鹃和山光杜鹃主干通直, 其茎生物量分配比例的变幅较小(
图1 ). ...
... )相似, 如
钟泽兵等(2014) 对青海玉树海拔3 715-4 296 m的5种建群灌木的研究发现叶生物量分配比例不足10%.甘肃南部高山-亚高山区的杜鹃会通过减少资源向叶片的分配, 保持常绿, 以较低的叶片周转率来适应寒冷环境.随着植株叶投资盈利能力的下降, 叶生物量将饱和, 投资会逐渐转向茎和根(
Poorter et al ., 2015 ), 而为增强植株的机械支撑, 必须增加茎的生物量投资(
Dybzinski et al ., 2011 ).本研究中7种高寒杜鹃茎生物量分配平均比例为45.61%, 接近于全球数据库平均值(
Poorter et al ., 2015 ).同时, 相比于叶和根, 茎生物量分配比例的种间差异最大(可达21%), 且多组物种间的差异显著(
表2 ), 这与
Poorter等(2015) 对全球1 200种植物整合分析的结果具有一致性, 茎不仅是植株的支撑、运输体系, 还在光竞争时向四周伸展小枝扩展空域, 而小枝的扩张又带来了高寒区防风的压力, 功能的多样性可能导致茎生物量分配的可塑性.各物种分配的差异还可能与茎构型有关, 如黄毛杜鹃和陇蜀杜鹃多为丛生, 茎枝扭曲, 其茎生物量分配比例的变异范围较大; 而美容杜鹃和山光杜鹃主干通直, 其茎生物量分配比例的变幅较小(
图1 ). ...
青海玉树地区主要灌丛类型地上生物量及其影响因素
3
2014
... 广布于我国青藏高原及其边缘山地的杜鹃(
Rhododendron spp.)常绿灌丛是亚高山暗针叶林向高山草甸过渡的群落类型, 在维持高山-亚高山生态系统稳定等方面具有关键作用(
钟泽兵等, 2014 ;
杨路存等, 2018 ).本研究以甘肃南部高山-亚高山区常见的7种杜鹃为材料, 通过全株收获法测定根、茎和叶的生物量, 采用标准化主轴法估计检验各器官间的异速生长关系, 并进行种间差异比较, 以期了解高寒杜鹃的生物量权衡策略, 为进一步认识高寒植物规避风险、提高适应性提供理论基础. ...
... 树木对温度适应性策略的假设认为, 在寒冷的气候环境下, 根的分配比例较大、叶的分配比例较小(
Reich et al ., 2014 ), 本研究中, 7种杜鹃根生物量分配比例高达35.57%, 个别物种甚至超过40% (
图1 ), 高于全球绝大多数植物(
Poorter et al ., 2015 ), 也高于我国亚热带山地杜鹃灌丛根系生物量的分配比例(
张蔷等, 2017 ), 与一些高寒植被(
高巧等, 2014 ;
钟泽兵等, 2014 ;
杨路存等, 2018 ;
左有璐等, 2018 )相似, 低速率的养分循环和受限的土壤溶液运动驱动着高山、亚高山地区的低营养供应(
魏晶等, 2004 ;
Luo et al ., 2012 ;
Reich et al ., 2014 ), 这使得高寒杜鹃需提高根生物量分配比例以增强从寒冷土壤中吸收养分, 生物量向根系中的倾向分配支持最优分配假说.7种杜鹃叶生物量分配比例低于全球绝大多数植物(
Poorter et al ., 2015 ), 其适应机制也与一些高寒植物(
Reich et al ., 2014 )相似, 如
钟泽兵等(2014) 对青海玉树海拔3 715-4 296 m的5种建群灌木的研究发现叶生物量分配比例不足10%.甘肃南部高山-亚高山区的杜鹃会通过减少资源向叶片的分配, 保持常绿, 以较低的叶片周转率来适应寒冷环境.随着植株叶投资盈利能力的下降, 叶生物量将饱和, 投资会逐渐转向茎和根(
Poorter et al ., 2015 ), 而为增强植株的机械支撑, 必须增加茎的生物量投资(
Dybzinski et al ., 2011 ).本研究中7种高寒杜鹃茎生物量分配平均比例为45.61%, 接近于全球数据库平均值(
Poorter et al ., 2015 ).同时, 相比于叶和根, 茎生物量分配比例的种间差异最大(可达21%), 且多组物种间的差异显著(
表2 ), 这与
Poorter等(2015) 对全球1 200种植物整合分析的结果具有一致性, 茎不仅是植株的支撑、运输体系, 还在光竞争时向四周伸展小枝扩展空域, 而小枝的扩张又带来了高寒区防风的压力, 功能的多样性可能导致茎生物量分配的可塑性.各物种分配的差异还可能与茎构型有关, 如黄毛杜鹃和陇蜀杜鹃多为丛生, 茎枝扭曲, 其茎生物量分配比例的变异范围较大; 而美容杜鹃和山光杜鹃主干通直, 其茎生物量分配比例的变幅较小(
图1 ). ...
... )相似, 如
钟泽兵等(2014) 对青海玉树海拔3 715-4 296 m的5种建群灌木的研究发现叶生物量分配比例不足10%.甘肃南部高山-亚高山区的杜鹃会通过减少资源向叶片的分配, 保持常绿, 以较低的叶片周转率来适应寒冷环境.随着植株叶投资盈利能力的下降, 叶生物量将饱和, 投资会逐渐转向茎和根(
Poorter et al ., 2015 ), 而为增强植株的机械支撑, 必须增加茎的生物量投资(
Dybzinski et al ., 2011 ).本研究中7种高寒杜鹃茎生物量分配平均比例为45.61%, 接近于全球数据库平均值(
Poorter et al ., 2015 ).同时, 相比于叶和根, 茎生物量分配比例的种间差异最大(可达21%), 且多组物种间的差异显著(
表2 ), 这与
Poorter等(2015) 对全球1 200种植物整合分析的结果具有一致性, 茎不仅是植株的支撑、运输体系, 还在光竞争时向四周伸展小枝扩展空域, 而小枝的扩张又带来了高寒区防风的压力, 功能的多样性可能导致茎生物量分配的可塑性.各物种分配的差异还可能与茎构型有关, 如黄毛杜鹃和陇蜀杜鹃多为丛生, 茎枝扭曲, 其茎生物量分配比例的变异范围较大; 而美容杜鹃和山光杜鹃主干通直, 其茎生物量分配比例的变幅较小(
图1 ). ...
川西北高寒草甸优势植物生物量分配对策
3
2018
... 理论上来说, 极端环境通过长期的筛选作用, 其适生物种响应资源约束时的生物量权衡策略应具有相似性(
Reich et al ., 2014 ).如, 在高寒山地, 高海拔、寒冷、短生长季和低气压等环境约束可能使植物的资源权衡具有独特表现(
高巧等, 2014 ;
左有璐等, 2014 ;
刘尊驰等, 2015 ), 植物会增加根生物量、降低叶生物量比例来适应较冷的环境(
徐波等, 2013 ).但目前的相关研究材料以常绿裸子植物或落叶被子植物为主(
Reich et al ., 2014 ;
Poorter et al ., 2015 ), 对高寒区生长的常绿被子植物的生物量权衡机制缺乏认知, 而常绿植物与落叶植物叶生物量分配策略存在差别, 常绿植物在叶片上投入的较多, 因此叶生物量分配比例高于落叶植物(
Poorter et al ., 2015 ). ...
... 树木对温度适应性策略的假设认为, 在寒冷的气候环境下, 根的分配比例较大、叶的分配比例较小(
Reich et al ., 2014 ), 本研究中, 7种杜鹃根生物量分配比例高达35.57%, 个别物种甚至超过40% (
图1 ), 高于全球绝大多数植物(
Poorter et al ., 2015 ), 也高于我国亚热带山地杜鹃灌丛根系生物量的分配比例(
张蔷等, 2017 ), 与一些高寒植被(
高巧等, 2014 ;
钟泽兵等, 2014 ;
杨路存等, 2018 ;
左有璐等, 2018 )相似, 低速率的养分循环和受限的土壤溶液运动驱动着高山、亚高山地区的低营养供应(
魏晶等, 2004 ;
Luo et al ., 2012 ;
Reich et al ., 2014 ), 这使得高寒杜鹃需提高根生物量分配比例以增强从寒冷土壤中吸收养分, 生物量向根系中的倾向分配支持最优分配假说.7种杜鹃叶生物量分配比例低于全球绝大多数植物(
Poorter et al ., 2015 ), 其适应机制也与一些高寒植物(
Reich et al ., 2014 )相似, 如
钟泽兵等(2014) 对青海玉树海拔3 715-4 296 m的5种建群灌木的研究发现叶生物量分配比例不足10%.甘肃南部高山-亚高山区的杜鹃会通过减少资源向叶片的分配, 保持常绿, 以较低的叶片周转率来适应寒冷环境.随着植株叶投资盈利能力的下降, 叶生物量将饱和, 投资会逐渐转向茎和根(
Poorter et al ., 2015 ), 而为增强植株的机械支撑, 必须增加茎的生物量投资(
Dybzinski et al ., 2011 ).本研究中7种高寒杜鹃茎生物量分配平均比例为45.61%, 接近于全球数据库平均值(
Poorter et al ., 2015 ).同时, 相比于叶和根, 茎生物量分配比例的种间差异最大(可达21%), 且多组物种间的差异显著(
表2 ), 这与
Poorter等(2015) 对全球1 200种植物整合分析的结果具有一致性, 茎不仅是植株的支撑、运输体系, 还在光竞争时向四周伸展小枝扩展空域, 而小枝的扩张又带来了高寒区防风的压力, 功能的多样性可能导致茎生物量分配的可塑性.各物种分配的差异还可能与茎构型有关, 如黄毛杜鹃和陇蜀杜鹃多为丛生, 茎枝扭曲, 其茎生物量分配比例的变异范围较大; 而美容杜鹃和山光杜鹃主干通直, 其茎生物量分配比例的变幅较小(
图1 ). ...
... 异速生长理论预测小个体植物叶、茎、根器官生长可能为等速关系(
Enquist & Niklas, 2002 ;
Weiner, 2004 ;
Niklas, 2006 ), 这种基于种间变异建立的跨物种关系已在种内变异的研究中得到了较多应用(
Gargaglione et al ., 2010 ;
Cheng et al ., 2014 ;
陈国鹏等, 2016 ;
王杨等, 2017 ), 如
Cheng等(2014) 发现南亚热带生长的5种苗木叶、茎和根生物量的积累为等速或近似等速生长关系.本研究中, 7种高寒杜鹃叶与茎、叶与根以及茎与根之间既有异速生长表现, 也有等速生长表现(
图2 -5), 与荒漠植物(
杨昊天等, 2013 ;
陶冶和张元明, 2014 )、干热河谷草本植物(
闫帮国等, 2016 )的异速生长关系具有相似性, 不同于川西高寒草甸优势植物(
左有璐等, 2018 ), 并不完全支持小个体器官间等速生长关系的预测, 也不完全支持生态代谢理论的预测, 这可能是由于植物在适应特殊环境的过程中, 不同物种间具有很大的功能群差异, 这些功能群差异可能是不同器官生物量间异速生长关系分化的主要因素(
Poorter et al ., 2015 ;
闫帮国等, 2016 ), 而基于跨物种建立的生态代谢理论、小个体等速生长理论忽略了物种间的功能群差异. ...
川西北高寒草甸优势植物生物量分配对策
3
2018
... 理论上来说, 极端环境通过长期的筛选作用, 其适生物种响应资源约束时的生物量权衡策略应具有相似性(
Reich et al ., 2014 ).如, 在高寒山地, 高海拔、寒冷、短生长季和低气压等环境约束可能使植物的资源权衡具有独特表现(
高巧等, 2014 ;
左有璐等, 2014 ;
刘尊驰等, 2015 ), 植物会增加根生物量、降低叶生物量比例来适应较冷的环境(
徐波等, 2013 ).但目前的相关研究材料以常绿裸子植物或落叶被子植物为主(
Reich et al ., 2014 ;
Poorter et al ., 2015 ), 对高寒区生长的常绿被子植物的生物量权衡机制缺乏认知, 而常绿植物与落叶植物叶生物量分配策略存在差别, 常绿植物在叶片上投入的较多, 因此叶生物量分配比例高于落叶植物(
Poorter et al ., 2015 ). ...
... 树木对温度适应性策略的假设认为, 在寒冷的气候环境下, 根的分配比例较大、叶的分配比例较小(
Reich et al ., 2014 ), 本研究中, 7种杜鹃根生物量分配比例高达35.57%, 个别物种甚至超过40% (
图1 ), 高于全球绝大多数植物(
Poorter et al ., 2015 ), 也高于我国亚热带山地杜鹃灌丛根系生物量的分配比例(
张蔷等, 2017 ), 与一些高寒植被(
高巧等, 2014 ;
钟泽兵等, 2014 ;
杨路存等, 2018 ;
左有璐等, 2018 )相似, 低速率的养分循环和受限的土壤溶液运动驱动着高山、亚高山地区的低营养供应(
魏晶等, 2004 ;
Luo et al ., 2012 ;
Reich et al ., 2014 ), 这使得高寒杜鹃需提高根生物量分配比例以增强从寒冷土壤中吸收养分, 生物量向根系中的倾向分配支持最优分配假说.7种杜鹃叶生物量分配比例低于全球绝大多数植物(
Poorter et al ., 2015 ), 其适应机制也与一些高寒植物(
Reich et al ., 2014 )相似, 如
钟泽兵等(2014) 对青海玉树海拔3 715-4 296 m的5种建群灌木的研究发现叶生物量分配比例不足10%.甘肃南部高山-亚高山区的杜鹃会通过减少资源向叶片的分配, 保持常绿, 以较低的叶片周转率来适应寒冷环境.随着植株叶投资盈利能力的下降, 叶生物量将饱和, 投资会逐渐转向茎和根(
Poorter et al ., 2015 ), 而为增强植株的机械支撑, 必须增加茎的生物量投资(
Dybzinski et al ., 2011 ).本研究中7种高寒杜鹃茎生物量分配平均比例为45.61%, 接近于全球数据库平均值(
Poorter et al ., 2015 ).同时, 相比于叶和根, 茎生物量分配比例的种间差异最大(可达21%), 且多组物种间的差异显著(
表2 ), 这与
Poorter等(2015) 对全球1 200种植物整合分析的结果具有一致性, 茎不仅是植株的支撑、运输体系, 还在光竞争时向四周伸展小枝扩展空域, 而小枝的扩张又带来了高寒区防风的压力, 功能的多样性可能导致茎生物量分配的可塑性.各物种分配的差异还可能与茎构型有关, 如黄毛杜鹃和陇蜀杜鹃多为丛生, 茎枝扭曲, 其茎生物量分配比例的变异范围较大; 而美容杜鹃和山光杜鹃主干通直, 其茎生物量分配比例的变幅较小(
图1 ). ...
... 异速生长理论预测小个体植物叶、茎、根器官生长可能为等速关系(
Enquist & Niklas, 2002 ;
Weiner, 2004 ;
Niklas, 2006 ), 这种基于种间变异建立的跨物种关系已在种内变异的研究中得到了较多应用(
Gargaglione et al ., 2010 ;
Cheng et al ., 2014 ;
陈国鹏等, 2016 ;
王杨等, 2017 ), 如
Cheng等(2014) 发现南亚热带生长的5种苗木叶、茎和根生物量的积累为等速或近似等速生长关系.本研究中, 7种高寒杜鹃叶与茎、叶与根以及茎与根之间既有异速生长表现, 也有等速生长表现(
图2 -5), 与荒漠植物(
杨昊天等, 2013 ;
陶冶和张元明, 2014 )、干热河谷草本植物(
闫帮国等, 2016 )的异速生长关系具有相似性, 不同于川西高寒草甸优势植物(
左有璐等, 2018 ), 并不完全支持小个体器官间等速生长关系的预测, 也不完全支持生态代谢理论的预测, 这可能是由于植物在适应特殊环境的过程中, 不同物种间具有很大的功能群差异, 这些功能群差异可能是不同器官生物量间异速生长关系分化的主要因素(
Poorter et al ., 2015 ;
闫帮国等, 2016 ), 而基于跨物种建立的生态代谢理论、小个体等速生长理论忽略了物种间的功能群差异. ...
 ,
, ,1,*, 杨克彤1, 王立1, 王飞2, 曹秀文2, 陈林生3
,1,*, 杨克彤1, 王立1, 王飞2, 曹秀文2, 陈林生3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT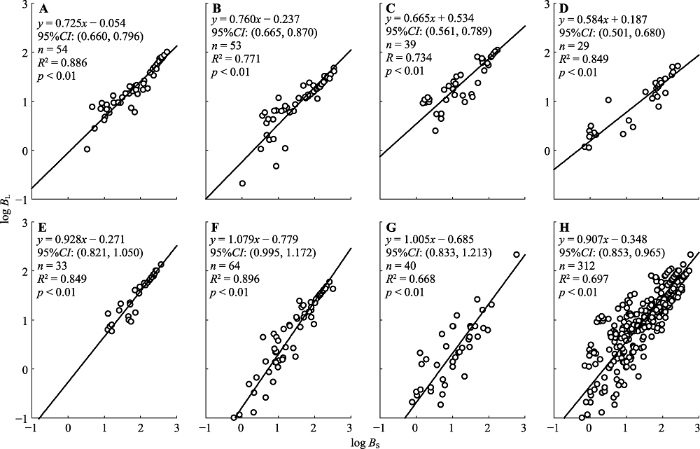 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT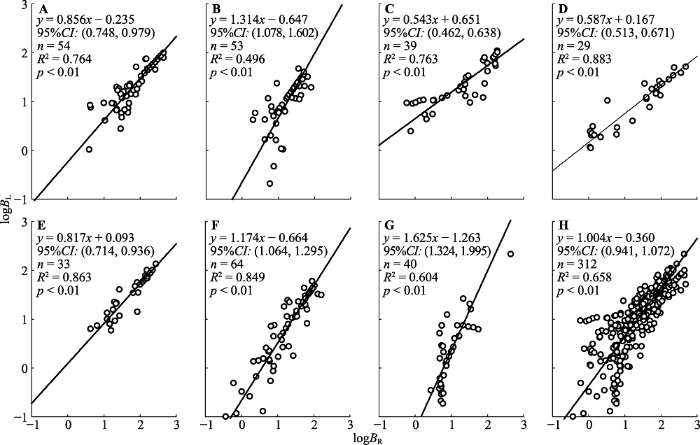 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT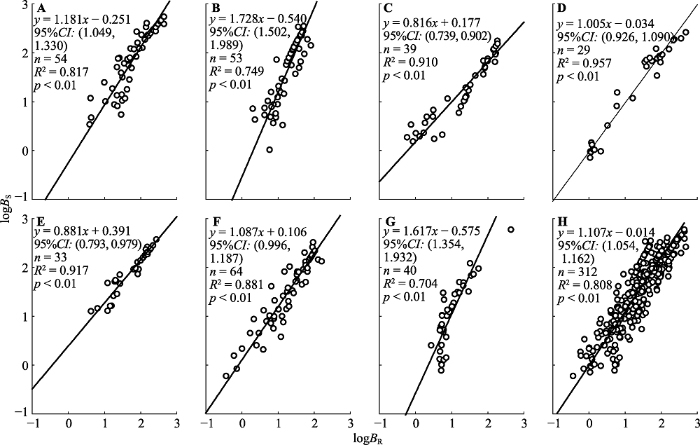 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT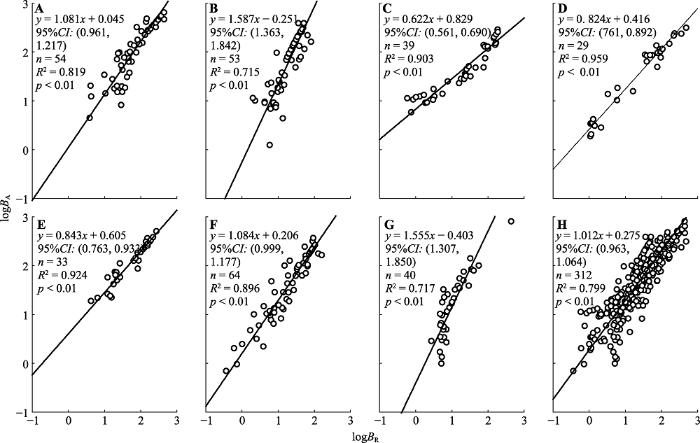 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}