Mechanism of the trade-off between biological nitrogen fixation and phosphorus acquisition strategies of herbaceous legumes under nitrogen and phosphorus addition
Qiang LI,*, Ying-Xin HUANG, Dao-Wei ZHOU, Shan CONGNortheast Institute of Geography and Agroecology, Chinese Academy of Sciences, Jilin Provincial Key Laboratory of Grassland Farming, Changchun 130102, China
Abstract Aims Nitrogen fixation of herbaceous legumes is not only an important natural nitrogen (N) input to terrestrial ecosystems, but also determines the economy and sustainability of grassland production. This study aimed to determine the underlying physiological and ecological mechanisms of the interaction between N and phosphorus (P) on biological N fixation rate of legumes. Methods In a pot experiment, eight species of herbaceous legumes were separately grown in soils with four treatments including no fertiliser, N addition, P addition, and both N and P (NP) addition. Plant biomass and nutrients concentrations, root carbohydrate concentration, pH in rhizosphere, citric concentration in in rhizosphere, avaiable P concentration in rhizosphere, root nodule biomass, P concentration in root nodule, and N fixation rate of these legume plants were examined. Important findings Depending on legume species, N addition significantly increased relative rhizosphere P mobilization, but reduced investment in root biomass and the concentration of non-structural carbohydrate (NSC) in roots. Averaged results of N addition and NP addition treatments indicated that N addition caused 27%-36% decline in nodule biomass and 20%-33% decline in biological N fixation rate for the studied eight legume species. By contrast, P addition significantly promoted root development and NSC accumulation associated with decreasing relative rhizosphere P mobilization. Consequently, P addition increased the biological N fixation rate of the eight legume species by 45%-69% and 0-47% with and without N fertilization, respectively. We concluded that N addition reduced biological N fixation rate via reducing root biomass and root NSC concentration and increasing rhizosphere P mobilization; P addition helped to improve soil N-P balance and promote root growth and NSC accumulation, which can alleviate the inhibition of biological N fixation by N fertilization. Keywords:nitrogen fixation;phosphorus mobilization;legume forage;rhizosphere;root nodule;symbiotic
PDF (770KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 李强, 黄迎新, 周道玮, 丛山. 土壤氮磷添加下豆科草本植物生物固氮与磷获取策略的权衡机制. 植物生态学报, 2021, 45(3): 286-297. DOI: 10.17521/cjpe.2020.0241 LI Qiang, HUANG Ying-Xin, ZHOU Dao-Wei, CONG Shan. Mechanism of the trade-off between biological nitrogen fixation and phosphorus acquisition strategies of herbaceous legumes under nitrogen and phosphorus addition. Chinese Journal of Plant Ecology, 2021, 45(3): 286-297. DOI: 10.17521/cjpe.2020.0241
生物固氮是豆科草本植物的重要氮(N)吸收途径, 豆科草本植物的生物固氮率不仅决定其同化大气氮的总量(Augusto et al., 2013), 还通过改变土壤氮的相对消耗影响豆科草本植物生长的氮经济(Li et al., 2016a, 2020)。外源氮输入和生物固氮的积累增加了土壤中氮的有效性(Galloway, 1998; Vitousek et al., 2013)。然而, 增加的土壤氮有效性会潜在地降低豆科草本植物的生物固氮率(Salvagiotti et al., 2008; Nyfeler et al., 2011), 进而阻碍豆科草本植物对生态系统的进一步固氮输入。以往的研究表明, 豆科植物能够根据土壤氮肥力状况调整在固定大气氮和获取土壤氮方面的投资策略(Vitousek et al., 2002; West et al., 2005)。这些研究认为生物固氮的代谢成本一般高于土壤氮吸收, 因此高土壤氮有效性下豆科植物倾向于增加根系对土壤氮的吸收, 但抑制代谢上高成本的根瘤发育和生物固氮(West et al., 2005)。这一观点暗示当氮有效性增加, 豆科植物生物固氮被抑制将成为普遍现象。然而, 其他研究发现, 外源氮的输入并不总是下调豆科植物生物固氮率, 这意味着一些其他因素可能会改变氮富集对生物固氮率的影响(Reed et al., 2007; Zheng et al., 2019)。
磷(P)对植物生长和代谢具有关键调控作用(White & Hammond, 2008; Png et al., 2017)。因为根瘤发育和生物固氮过程的高磷消耗, 豆科植物对磷的供应尤为敏感(Batterman et al., 2013; Divito & Sadras, 2014)。研究表明磷添加能够提高豆科植物根瘤发育和生物固氮率(Augusto et al., 2013; Maistry et al., 2013), 尤其在磷资源限制环境中(Betencourt et al., 2012)。尽管氮、磷在土壤中有不同的生物地球化学循环过程, 增加的氮供应很可能改变土壤内在的氮-磷平衡(Elser et al., 2010; Pe?uelaset al., 2012), 提高植物磷需求, 驱动土壤磷的消耗(Güsewell,2004; Png et al., 2017), 并最终加剧植物磷匮乏。基于此, 氮增加下, 改变的土壤氮磷供给平衡很可能成为驱动豆科植物生物固氮率下降的额外机制。在这种情景下, 土壤中磷肥的状态能够潜在改变氮富集对豆科植物生物固氮的抑制作用。先前的研究发现当土壤磷有效性较低时, 氮增加抑制根系结瘤, 但当土壤磷有效性较高时, 氮增加能够促进根系结瘤(Gates & Wilson, 1974), 相似的结果也存在于其他研究中(Gentili & Huss-Danell, 2003)。这些研究已经表明氮、磷在调控豆科植物固氮行为中的交互作用。然而, 对于氮-磷平衡的改变如何驱动植物生理变化, 并最终改变豆科植物根系结瘤和生物固氮率, 我们仍旧缺乏深入理解。
磷限制影响豆科植物根瘤细胞的构建, 并影响生物固氮过程中的能量供应(Sulieman et al., 2013; Divito & Sadras, 2014)。然而, 并不清楚氮增加导致的相对磷匮乏是否已经经由降低植物和根瘤磷吸收来抑制根系结瘤和生物固氮率。此外, 在磷资源限制下, 许多陆地植物, 尤其豆科植物已经发展了物种特异性的适应机制, 诸如强化植物-菌根互作、刺激根际磷活化等来满足植物磷需求(Nuruzzaman, 2006; Zhou et al., 2009)。进一步的实验证据表明上述磷获取策略的实现极大依赖于根系形态和生理的改变(Nuruzzaman, 2006; Betencourt et al., 2012; Png et al., 2017)。然而, 氮增加是否激发了这些磷获取策略, 并借此改变豆科植物根系生理, 导致生物固氮率的下降不得而知。
Table 1 表1 表1氮、磷添加及其交互作用影响的土壤有效氮、磷含量和有效氮磷比例(平均值±标准误) Table 1Effect of nitrogen (N) and phosphorus (P) addition and their interaction on available N and P concentration and available N:P ratio in bulk soil (mean ± SE)
处理 Treatment
有效氮 Available N (mg·kg-1)
有效磷 Available P (mg·kg-1)
N:P
Control
37.05 ± 0.50
5.08 ± 0.07 a
7.31 ± 0.13 c
N
72.02 ± 1.15 a
5.11 ± 0.05 a
14.12 ± 0.25 a
P
37.65 ± 0.64 b
9.36 ± 0.11 b
4.03 ± 0.07 d
NP
71.91 ± 1.02 a
9.18 ± 0.09 b
7.84 ± 0.38 b
Control, 无肥料添加; N, 氮添加; P, 磷添加; NP, 氮磷耦合添加。不同小写字母代表施肥处理间差异显著(p < 0.05)。 Control, no fertiliser addition; N, nitrogen addition; P, phosphorus addition; NP, coupling nitrogen and phosphorus addition. Different lowercase letters indicate significant differences among fertilization treatments (p < 0.05).
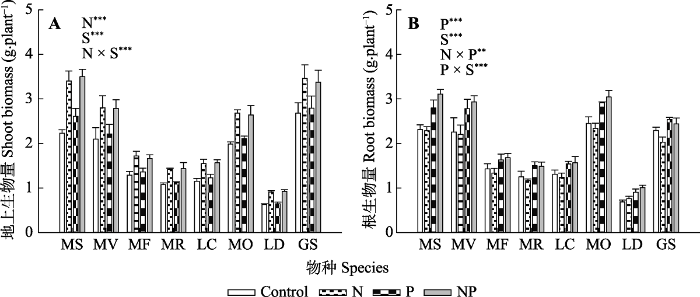
新窗口打开|下载原图ZIP|生成PPT 图1氮、磷添加及其交互作用对不同豆科草本植物地上生物量(A)和根生物量(B)(平均值±标准误)的影响。Control, 无养分添加; N, 氮添加; P, 磷添加; NP, 氮磷耦合添加; S, 物种。GS, 野大豆; LC, 百脉根; LD, 兴安胡枝子; MF, 黄花苜蓿; MO, 草木樨; MR, 花苜蓿; MS, 紫花苜蓿; MV, 杂花苜蓿。星号表示经一般线性模型统计分析得因子效应具有显著性。***, p < 0.001; **, p < 0.01; *, p < 0.05。
Fig. 1Influence of nitrogen (N) and phosphorus (P) addition and their interaction on shoot biomass (A) and root biomass (B)(mean ± SE) of different herbaceous legumes. Control, no nutrient addition; N, N addition; P, P addition; NP, coupled addition of N and P; S, species. GS, Glycine soja; LC, Lotus corniculatus; LD, Lespedeza daurica; MF, Medicago falcata; MO, Melilotus officinalis; MR, Medicago ruthenica; MS, Medicago sativa; MV, Medicago varia. Asterisks indicate that factor effect was significant by general linear model analysis. ***, p < 0.001; **, p < 0.01; *, p < 0.05.
图2
新窗口打开|下载原图ZIP|生成PPT 图2氮、磷添加及其交互作用对不同豆科草本植物氮含量(A)、磷含量(B)和氮磷比(C)(平均值±标准误)的影响。Control, 无养分添加; N, 氮添加; P, 磷添加; NP, 氮磷耦合添加; S, 物种。GS, 野大豆; LC, 百脉根; LD, 兴安胡枝子; MF, 黄花苜蓿; MO, 草木樨; MR, 花苜蓿; MS, 紫花苜蓿; MV, 杂花苜蓿。星号表示经一般线性模型统计分析得因子效应具有显著性。***, p < 0.001; **, p < 0.01; *, p < 0.05。
Fig. 2Influence of nitrogen (N) and phosphorus (P) addition and their interaction on plant N concentration (A), plant P concentration (B) and plant N:P ratio (C)(mean ± SE) of different herbaceous legumes. Control, no nutrient addition; N, N addition; P, P addition; NP, coupled addition of N and P; S, species. GS, Glycine soja; LC, Lotus corniculatus; LD, Lespedeza daurica; MF, Medicago falcata; MO, Melilotus officinalis; MR, Medicago ruthenica; MS, Medicago sativa; MV, Medicago varia. Asterisks indicate that factor effect was significant by general linear model analysis. ***, p < 0.001; **, p < 0.01; *, p < 0.05.
新窗口打开|下载原图ZIP|生成PPT 图3氮、磷添加及其交互作用对不同豆科草本植物根际有效磷含量(A)和根际磷活化效应(B)(平均值±标准误)的影响。Control, 无养分添加; N, 氮添加; P, 磷添加; NP, 氮磷耦合添加; S, 物种。GS, 野大豆; LC, 百脉根; LD, 兴安胡枝子; MF, 黄花苜蓿; MO, 草木樨; MR, 花苜蓿; MS, 紫花苜蓿; MV, 杂花苜蓿。星号表示经一般线性模型统计分析得因子效应具有显著性。***, p < 0.001; **, p < 0.01; *, p < 0.05。
Fig. 3Influence of nitrogen (N) and phosphorus (P) addition and their interaction on available P concentration in rhizosphere (A) and P mobilization in rhizosphere (B)(mean ± SE) of different herbaceous legumes. Control, no nutrient addition; N, N addition; P, P addition; NP, coupled addition of N and P; S, species. GS, Glycine soja; LC, Lotus corniculatus; LD, Lespedeza daurica; MF, Medicago falcata; MO, Melilotus officinalis; MR, Medicago ruthenica; MS, Medicago sativa; MV, Medicago varia. Asterisks indicate that factor effect was significant by general linear model analysis. ***, p < 0.001; **, p < 0.01; *, p < 0.05.
图4
新窗口打开|下载原图ZIP|生成PPT 图4氮、磷添加及其交互作用对不同豆科草本植物根系非结构性碳水化合物(NSC)含量(A)、根系的菌根(AM)侵染率(B)、根际pH (C)和根际柠檬酸含量(D)(平均值±标准误)的影响。Control, 无养分添加; N, 氮添加; P, 磷添加; NP, 氮磷耦合添加; S, 物种。GS, 野大豆; LC, 百脉根; LD, 兴安胡枝子; MF, 黄花苜蓿; MO, 草木樨; MR, 花苜蓿; MS, 紫花苜蓿; MV, 杂花苜蓿。星号表示经一般线性模型统计分析得因子效应具有显著性。***, p < 0.001; **, p < 0.01; *, p < 0.05。
Fig. 4Influence of nitrogen (N) and phosphorus (P) addition and their interaction on non-structure carbohydrate (NSC) concentration in root (A), arbuscular mycorrhizal (AM) colonization rate of root (B), pH in rhizosphere (C) and citric concentration in rhizosphere (D)(mean ± SE) of different herbaceous legumes. Control, no nutrient addition; N, N addition; P, P addition; NP, coupled addition of N and P; S, species. GS, Glycine soja; LC, Lotus corniculatus; LD, Lespedeza daurica; MF, Medicago falcata; MO, Melilotus officinalis; MR, Medicago ruthenica; MS, Medicago sativa; MV, Medicago varia. Asterisks indicate that factor effect was significant by general linear model analysis. ***, p < 0.001; **, p < 0.01; *, p < 0.05.
Fig. 5Correlation relationships between mean root biomass and arbuscular mycorrhizal (AM) colonization rate of root (A), soil pH in rhizosphere (B), citric concentration in rhizosphere (C), and between mean non-structure carbohydrate (NSC) concentration in root and AM colonization rate of root (D), soil pH in rhizosphere (E), citric concentration in rhizosphere (F)(mean ± SE) following different nutrient addition treatments on eight herbaceous legumes.
新窗口打开|下载原图ZIP|生成PPT 图6氮、磷添加及其交互作用对不同豆科草本植物根瘤数目(A)、根瘤生物量(B)、根瘤磷含量(C)和生物固氮率(BNF)(D)(平均值±标准误)的影响。Control, 无养分添加; N, 氮添加; P, 磷添加; NP, 氮磷耦合添加; S, 物种。GS, 野大豆; LC, 百脉根; LD, 兴安胡枝子; MF, 黄花苜蓿; MO, 草木樨; MR, 花苜蓿; MS, 紫花苜蓿; MV, 杂花苜蓿。星号表示经一般线性模型统计分析得因子效应具有显著性。***, p < 0.001; **, p < 0.01; *, p < 0.05。
Fig. 6Influence of nitrogen (N) and phosphorus (P) addition and their interaction on root nodule number (A), root nodule biomass (B), root nodule P concentration (C) and biological N fixation rate (BNF)(D)(mean ± SE) of different herbaceous legumes. Control, no nutrient addition; N, N addition; P, P addition; NP, coupled addition of N and P; S, species. GS, Glycine soja; LC, Lotus corniculatus; LD, Lespedeza daurica; MF, Medicago falcata; MO, Melilotus officinalis; MR, Medicago ruthenica; MS, Medicago sativa; MV, Medicago varia. Asterisks indicate that factor effect was significant by general linear model analysis. ***, p < 0.001; **, p < 0.01; *, p < 0.05.
Fig. 8Correlation relationships between phosphorus (P) mobilization effect in rhizosphere and biological nitrogen fixation rate of legume (BNF)(mean ± SE) following different nutrient addition treatments on eight herbaceous legumes.
3 讨论
3.1 根系生物量和根部非结构性碳水化合物含量的下降导致豆科草本植物固氮抑制
生物固氮和吸收土壤氮是豆科植物两个可供选择的氮获取策略(Ledgard et al., 2001)。如果排除其他环境限制, 土壤氮资源的状态是豆科植物氮获取策略选择的关键决定因素(West et al., 2005)。氮添加提高了土壤氮资源有效性, 可能降低豆科植物为获取大气氮而对根和根瘤发育的投资。再者, 增加的土壤氮有效性可能将豆科草本植物间竞争的压力从地下转移到地上(Farrer & Suding, 2016), 诱使植物投资更多资源到地上生物量用以捕获光, 并因此限制根系发育(Aerts et al., 1991)。本研究中, 对比无肥料添加, 氮添加提高了地上生物量, 并小幅降低根生物量, 已经表明了氮添加下豆科草本植物生物量分配的转移。同时, 我们的研究也发现氮添加降低了豆科草本植物根部非结构性碳水化合物的含量, 间接地表明土壤氮资源的增加可能改变了光合产物的分配, 促使豆科草本植物将更多的光合产物转移并储存在地上, 并导致根部碳水化合物营养储存的下降。
本实验中所用土壤磷含量较低, 可能已经限制了豆科草本植物的生长(Betencourt et al., 2012), 这一推断能够从控制盆中已经较高的根际磷活化效应(图3B)得以佐证。氮添加显著提高了豆科草本植物生物量和组织氮含量, 因为植物生长和代谢过程对氮、磷元素的匹配需求, 氮添加将潜在提高植物对磷的需要(White & Hammond, 2008; Png et al., 2017)。然而, 当前的研究发现氮添加并未改变土壤中磷的供给。而这种氮磷供应的不平衡并未导致豆科草本植物降低植物和根瘤的磷含量, 反而激发了豆科草本植物根际磷的活化和吸收, 以满足增加的磷需求(图4)(Nuruzzaman et al., 2006; Betencourt et al., 2012)。因为植物的养分吸收主要发生在根际(Hinsinger, 1998), 植物物种已经进化发展了一系列策略来提高根际磷吸收。已知的策略包括加强根系-菌根互作以提高根部磷吸收的范围和面积(Smith & Read, 2008); 或通过根际土壤酸化刺激钙结合无机磷的活化, 以增加根际有效磷数量, 这种作用在中性或类似于本研究的碱性土壤中可能尤为重要(Richardson et al., 2009; Betencourt et al., 2012)。此外, 植物根部能够分泌一些磷活化有机物, 如柠檬酸等有机酸, 这些磷活化物质也能提高根际磷有效性(Nuruzzaman et al., 2006)。当前研究发现氮添加下, 根际土壤pH显著下降, 而根部菌根侵染率明显增加, 这些结果表明氮添加已经激发了豆科草本植物多途径的磷截获策略。众所周知, 菌根的共生依赖于根部提供碳水化合物营养(Soper et al., 2019)。此外, 根际土壤pH的下降一般得益于根呼吸和根际微生物呼吸的增加, 而增加的根呼吸和根际微生物呼吸提高了根内及其分泌到根际的碳水化合物的消耗(Hinsinger et al., 2003; Nuruzzaman et al., 2006)。因此, 通过菌根共生和根际磷活化的途径来获取更多磷可能潜在降低根生物量和根碳水化合物的积累(图5)。本研究中根生物量、根碳水化合物和豆科草本植物生物固氮率间存在正相关关系, 表明氮添加下, 豆科草本植物为提高磷吸收, 会抑制其生物固氮率。
最近的meta分析发现, 氮增加对生物固氮的作用随生态系统类型、地理位置、气候条件等的变化而发生改变(Zheng et al., 2019)。实际上这些大尺度环境因子可能已经通过改变土壤养分背景而调控了氮增加对生物固氮的影响。先前的研究发现氮增加有时并不下调豆科植物生物固氮率, 并将其归因为该生态系统具有较低的土壤氮水平, 导致该生态系统即使在氮增加情境下也处于持续的氮限制状态(Reed et al., 2007)。通过本研究, 我们进一步指出土壤磷有效性调控N:P平衡, 对氮增加下的豆科草本植物生物固氮率具有重要调节作用(Gentili & Huss-Danell, 2003)。实际上, 在陆地生态系统中, 磷对植物生长的限制可能远比之前认定的要普遍, 尤其在人类诱导的不平衡的氮、磷输入情景下(Mahowald et al., 2008; Vitousek et al., 2010; Li et al., 2016b), 由此磷对豆科植物生物固氮的调控功能将变得越来越重要。
3.3 磷添加缓解氮富集对豆科草本植物生物固氮的抑制作用
磷添加能够促进豆科植物生长和生物固氮(Augusto et al., 2013; Maistry et al., 2013), 主要有两个原因。首先, 磷是固氮豆科植物及其根瘤细胞构建的主要元素, 并且是生物固氮过程中关键的能量驱动要素(Sulieman et al., 2013; Divito & Sadras, 2014)。此外, 磷的添加也促进豆科植物生长, 并相应增加其对氮的需求, 由此刺激根瘤生物量和生物固氮的增加以满足氮需要(Batterman et al., 2013)。本研究结果证实磷添加显著提高豆科草本植物生物固氮率。进一步地, 本研究阐述了磷添加促进豆科草本植物固氮的一个新机制, 即磷添加降低豆科草本植物根系-菌根互作和根际磷活化, 因此促进根生物量和根碳水化合物的累积, 间接提高生物固氮率。
更大的氮有效性提高植物磷需求, 增加了土壤磷对豆科草本植物生长和固氮的限制(Vitousek et al., 2010; Png et al., 2017)。因此, 氮富集下补充磷资源对缓解生物固氮的磷限制尤为重要。这一推断被我们的结果所支持, 因为本研究发现氮添加情境下补充磷对豆科草本植物生物固氮率有更大的提升。目前, 由于不成比例的氮磷输入(N >> P), 全球变化和农业活动正在改变许多生态系统的氮磷平衡(Vitousek et al., 2010)。根据我们的研究, 这样的氮富集可能导致豆科植物生物固氮发生不同程度的改变, 其变化程度取决于土壤磷状态。生态系统中磷有效性越低, 氮添加对生物固氮的抑制效应越大。因此, 在高土壤N:P或者高氮沉降速率的环境下, 为增加豆科草本植物生物固氮, 补充磷是明智的选择。本研究中, 平均所有豆科物种, 对比无营养添加, 氮和磷协同添加并未改变豆科草本植物生物固氮率(对照: 41% vs. 氮磷添加: 43%), 这一结果表明3.5 g·m-2的磷添加能够有效缓解14 g·m-2的氮添加对实验豆科草本植物生物固氮的抑制效应。这一结果对本研究区豆科草场的优化管理具有重要指导意义。
AertsR, BootRGA, van der Aart PJM (1991). The relation between above- and belowground biomass allocation patterns and competitive ability Oecologia, 87, 551-559. DOI:10.1007/BF00320419PMID:28313698 [本文引用: 1] In a 2-year experiment, the evergreen shrubsErica tetralix andCalluna vulgaris (dominant on nutrient-poor heathland soils) and the perennial deciduous grassMolinia caerulea (dominant on nutrient-rich heathland soils) were grown in replacement series in a factorial combination of four competition types (no competition, only aboveground competition, only belowground competition, full competition) and two levels of nutrient supply (no nutrients and 10 g N+2 g P+10 g K m yr). Both in the unfertilized and in the fertilized treatmentsMolinia allocated about twice as much biomass to its root system than didErica andCalluna. In all three species the relative amount of biomass allocated to the roots was lower at high than at low nutrient supply. The relative decrease was larger forMolinia than forErica andCalluna. In the fertilized monocultures biomass of all three species exceeded that in the unfertilized series.Molinia showed the greatest biomass increase. In the unfertilized series no effects of interspecific competition on the biomass of each species were observed in either of the competition treatments. In the fertilized mixtures where only belowground competition was possibleMolinia increased its biomass at the expense of bothErica andCalluna. When only aboveground competition was possible no effects of interspecific competition on the biomass of the competing species were observed. However, in contrast with the evergreens,Molinia responded by positioning its leaf layers relatively higher in the canopy. The effects of full competition were similar to those of only belowground competition, so in the fertilized series belowground competition determined the outcome of competition. The high competitive ability ofMolinia at high nutrient supply can be attributed to the combination of (1) a high potential productivity, (2) a high percentage biomass allocation to the roots, (3) an extensive root system exploiting a large soil volume, and (4) plasticity in the spatial arrangement of leaf layers over its tall canopy. In the species under study the allocation patterns entailed no apparent trade-off between the abilities to compete for above- and belowground resources. This study suggests that this trade-off can be overcome by: (1) plasticity in the spatial arrangement of leaf layers and roots, and (2) compensatory phenotypic and species-specific differences in specific leaf area and specific root length.
AugustoL, DelerueF, Gallet-BudynekA, AchatDL (2013). Global assessment of limitation to symbiotic nitrogen fixation by phosphorus availability in terrestrial ecosystems using a meta-analysis approach Global Biogeochemical Cycles, 27, 804-815. DOI:10.1002/gbc.20069URL [本文引用: 3]
BaiY, WuJ, ClarkCM, NaeemS, PanQ, HuangJ, ZhangL, HanXG (2010). Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: evidence from Inner Mongolia grasslands Global Change Biology, 16, 358-372. DOI:10.1111/(ISSN)1365-2486URL [本文引用: 1]
BattermanSA, HedinLO, vanBreugel M, RansijnJ, CravenDJ, HallJS (2013). Key role of symbiotic dinitrogen fixation in tropical forest secondary succession Nature, 502, 224-227. DOI:10.1038/nature12525PMID:24037375 [本文引用: 2] Forests contribute a significant portion of the land carbon sink, but their ability to sequester CO2 may be constrained by nitrogen, a major plant-limiting nutrient. Many tropical forests possess tree species capable of fixing atmospheric dinitrogen (N2), but it is unclear whether this functional group can supply the nitrogen needed as forests recover from disturbance or previous land use, or expand in response to rising CO2 (refs 6, 8). Here we identify a powerful feedback mechanism in which N2 fixation can overcome ecosystem-scale deficiencies in nitrogen that emerge during periods of rapid biomass accumulation in tropical forests. Over a 300-year chronosequence in Panama, N2-fixing tree species accumulated carbon up to nine times faster per individual than their non-fixing neighbours (greatest difference in youngest forests), and showed species-specific differences in the amount and timing of fixation. As a result of fast growth and high fixation, fixers provided a large fraction of the nitrogen needed to support net forest growth (50,000?kg carbon per hectare) in the first 12?years. A key element of ecosystem functional diversity was ensured by the presence of different N2-fixing tree species across the entire forest age sequence. These findings show that symbiotic N2 fixation can have a central role in nitrogen cycling during tropical forest stand development, with potentially important implications for the ability of tropical forests to sequester CO2.
BaziramakengaR, SimardRR, LerouxGD (1995). Determination of organic acids in soil extracts by ion chromatography Soil Biology & Biochemistry, 27, 349-356. DOI:10.1016/0038-0717(94)00178-4URL [本文引用: 1]
BetencourtE, DuputelM, ColombB, DesclauxD, HinsingerP (2012). Intercropping promotes the ability of durum wheat and chickpea to increase rhizosphere phosphorus availability in a low P soil Soil Biology & Biochemistry, 46, 181-190. DOI:10.1016/j.soilbio.2011.11.015URL [本文引用: 5]
BuysseJ, MerckxR (1993). An improved colorimetric method to quantify sugar content of plant tissue Journal of Experimental Botany, 44, 1627-1629. DOI:10.1093/jxb/44.10.1627URL [本文引用: 1]
DivitoGA, SadrasVO (2014). How do phosphorus, potassium and sulphur affect plant growth and biological nitrogen fixation in crop and pasture legumes? A meta-analysis Field Crops Research, 156, 161-171. DOI:10.1016/j.fcr.2013.11.004URL [本文引用: 3]
FarrerEC, SudingKN (2016). Teasing apart plant community responses to N enrichment: the roles of resource limitation, competition and soil microbes Ecology Letters, 19, 1287-1296. DOI:10.1111/ele.12665PMID:27531674 [本文引用: 1] Although ecologists have documented the effects of nitrogen enrichment on productivity, diversity and species composition, we know little about the relative importance of the mechanisms driving these effects. We propose that distinct aspects of environmental change associated with N enrichment (resource limitation, asymmetric competition, and interactions with soil microbes) drive different aspects of plant response. We test this in greenhouse mesocosms, experimentally manipulating each factor across three ecosystems: tallgrass prairie, alpine tundra and desert grassland. We found that resource limitation controlled productivity responses to N enrichment in all systems. Asymmetric competition was responsible for diversity declines in two systems. Plant community composition was impacted by both asymmetric competition and altered soil microbes, with some contributions from resource limitation. Results suggest there may be generality in the mechanisms of plant community change with N enrichment. Understanding these links can help us better predict N response across a wide range of ecosystems.? 2016 John Wiley & Sons Ltd/CNRS.
GatesCT, WilsonJR (1974). The interaction of nitrogen and phosphorus on the growth, nutrient status and nodulation of Stylosanthes humilis H.B.K. (townsville stylo) Plant and Soil, 41, 325-333. DOI:10.1007/BF00017260URL [本文引用: 1]
GentiliF, Huss-DanellK (2003). Local and systemic effects of phosphorus and nitrogen on nodulation and nodule function in Alnus incana Journal of Experimental Botany, 54, 2757-2767. DOI:10.1093/jxb/erg311URL [本文引用: 2]
GüsewellS (2004). N:P ratios in terrestrial plants: variation and functional significance New Phytologist, 164, 243-266. DOI:10.1111/nph.2004.164.issue-2URL [本文引用: 1]
HinsingerP (1998). How do plant roots acquire mineral nutrients? Chemical processes involved in the rhizosphere Advances in Agronomy, 64, 255-265. [本文引用: 2]
HinsingerP, PlassardC, TangC, JaillardB (2003). Origins of root-mediated pH changes in the rhizosphere and their responses to environmental constraints: a review Plant and Soil, 248, 43-59. DOI:10.1023/A:1022371130939URL [本文引用: 1]
LedgardSF, SprosenMS, PennoJW, RajendramGS (2001). Nitrogen fixation by white clover in pastures grazed by dairy cows: temporal variation and effects of nitrogen fertilization Plant and Soil, 229, 177-187. DOI:10.1023/A:1004833804002URL [本文引用: 1]
LedgardSF, SteeleKW (1992). Biological nitrogen fixation in mixed legume/grass pastures Plant and Soil, 141, 137-153. DOI:10.1007/BF00011314URL [本文引用: 1]
LiQ, SongY, LiG, YuP, WangP, ZhouD (2015). Grass-legume mixtures impact soil N, species recruitment, and productivity in temperate steppe grassland Plant and Soil, 394, 271-285. DOI:10.1007/s11104-015-2525-2URL [本文引用: 1]
LiQ, YuP, LiG, ZhouD (2016a). Grass-legume ratio can change soil carbon and nitrogen storage in a temperate steppe grassland Soil and Tillage Research, 157, 23-31. DOI:10.1016/j.still.2015.08.021URL [本文引用: 1]
LiQ, ZhangH, HuangY, ZhouD (2020). Forage nitrogen yield and soil nitrogen in artificial grasslands with varied Medicago seedling proportion Archives of Agronomy and Soil Science, 66, 110-125. DOI:10.1080/03650340.2019.1600675URL [本文引用: 1]
LiY, NiuS, YuG (2016b). Aggravated phosphorus limitation on biomass production under increasing nitrogen loading: a meta-analysis Global Change Biology, 22, 934-943. DOI:10.1111/gcb.13125URL [本文引用: 1]
MahowaldN, JickellsTD, BakerAR, ArtaxoP, Benitez-NelsonCR, BergamettiG, BondTC, ChenY, CohenDD, HerutB, KubilayN, LosnoR, LuoC, MaenhautW, McGeeKA, OkinGS, SiefertRL, TsukudaS (2008). Global distribution of atmospheric phosphorus sources, concentrations and deposition rates, and anthropogenic impacts Global Biogeochemical Cycles, 22, GB4026. DOI: 10.1029/2008GB003240. DOI:10.1029/2008GB003240 [本文引用: 1]
MaistryPM, CramerMD, ChimphangoSBM (2013). N and P colimitation of N2-fixing and N-supplied fynbos legumes from the Cape Floristic Region Plant and Soil, 373, 217-228. DOI:10.1007/s11104-013-1793-yURL [本文引用: 2]
MillerRH, KeeneyDR (1982). Methods of Soil Analysis, Part 2. Chemical and Microbiological Properties. 2nd ed. American Society of Agronomy, Madison, USA. [本文引用: 1]
NuruzzamanM, LambersH, BollandMDA, VeneklaasEJ (2006). Distribution of carboxylates and acid phosphatase and depletion of different phosphorus fractions in the rhizosphere of a cereal and three grain legumes Plant and Soil, 281, 109-120. DOI:10.1007/s11104-005-3936-2URL [本文引用: 6]
NyfelerD, Huguenin-ElieO, SuterM, FrossardE, LüscherA (2011). Grass-legume mixtures can yield more nitrogen than legume pure stands due to mutual stimulation of nitrogen uptake from symbiotic and non-symbiotic sources Agriculture, Ecosystem & Environment, 140, 155-163. DOI:10.1016/j.agee.2010.11.022URL [本文引用: 1]
OlsenSR, ColeCV, WatanabeSS, DeanLA (1954). Estimation of Available Phosphorus in Soil by Extraction with Sodium Bicarbonate. United States Department of Agriculture , Washington DC. [本文引用: 1]
Pe?uelasJ, SardansJ, Rivas-UbachA, JanssensIA (2012). The human-induced imbalance between C, N and P in Earth?s life system Global Change Biology, 18, 3-6. DOI:10.1111/j.1365-2486.2011.02568.xURL [本文引用: 1]
PngGK, TurnerBL, AlbornozFE, HayesPE, LambersH, LalibertéE (2017). Greater root phosphatase activity in nitrogen-fixing rhizobial but not actinorhizal plants with declining phosphorus availability Journal of Ecology, 105, 1246-1255. DOI:10.1111/jec.2017.105.issue-5URL [本文引用: 5]
ReedSC, ClevelandCC, TownsendAR (2007). Controls over leaf litter and soil nitrogen fixation in two lowland tropical rain forests Biotropica, 39, 585-592. DOI:10.1111/btp.2007.39.issue-5URL [本文引用: 2]
RichardsonAE, BareaJM, McNeillAM, Prigent-CombaretC (2009). Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms Plant and Soil, 321, 305-339. DOI:10.1007/s11104-009-9895-2URL [本文引用: 1]
SalvagiottiF, CassmanKG, SpechtJE, WaltersDT, WeissA, DobermannA (2008). Nitrogen uptake, fixation and response to fertilizer N in soybeans: a review Field Crops Research, 108, 1-13. DOI:10.1016/j.fcr.2008.03.001URL [本文引用: 1]
SchadeJD, KyleM, HobbieSE, FaganWF, ElserJJ (2003). Stoichiometric tracking of soil nutrients by a desert insect herbivore Ecology Letters, 6, 96-101. DOI:10.1046/j.1461-0248.2003.00409.xURL [本文引用: 1]
SongYT, ZhouDW, LiQ, WangP, HuangYX (2012). Leaf nitrogen and phosphorus stoichiometry in 80 herbaceous plant species of Songnen grassland in Northeast China Chinese Journal of Plant Ecology, 36, 222-230. DOI:10.3724/SP.J.1258.2012.00222URL [本文引用: 1]
SoperFM, NastoMK, OsborneBB, ClevelandCC (2019). Nitrogen fixation and foliar nitrogen do not predict phosphorus acquisition strategies in tropical trees Journal of Ecology, 107, 118-126. DOI:10.1111/jec.2019.107.issue-1URL [本文引用: 1]
SparksDL, PageAL, HelmkePA, LoeppertRH, SoltanpourPN, TabatabaiMA, JohnstonCT, SumnerME (1996). Methods of Soil Analysis. Part 3. Chemical Methods Soil Science Society of America, Madison, USA. [本文引用: 1]
SuliemanS, vanHa C, SchulzeJ, PhanTran LS (2013). Growth and nodulation of symbiotic Medicago truncatula at different levels of phosphorus availability Journal of Experimental Botany, 64, 2701-2712. DOI:10.1093/jxb/ert122PMID:23682114 [本文引用: 2] Medicago truncatula is an important model plant for characterization of P deficiency on leguminous plants at the physiological and molecular levels. Growth optimization of this plant with regard to P supply is the first essential step for elucidation of the role of P in regulation of nodulation. Hence, a study was carried out to address the growth pattern of M. truncatula hydroponically grown at different gradual increases in P levels. The findings revealed that M. truncatula had a narrow P regime, with an optimum P level (12 μM P) which is relatively close to the concentration that induces P toxicity. The accumulated P concentration (2.7 mg g(-1) dry matter), which is normal for other crops and legumes, adversely affected the growth of M. truncatula plants. Under P deficiency, M. truncatula showed a higher symbiotic efficiency with Sinorhizobium meliloti 2011 in comparison with S. meliloti 102F51, partially as a result of higher electron allocation to N2 versus H(+). The total composition of free amino acids in the phloem was significantly affected by P deprivation. This pattern was found to be almost exclusively the result of the increase in the asparagine level, suggesting that asparagine might be the shoot-derived signal that translocates to the nodules and exerts the down-regulation of nitrogenase activity. Additionally, P deprivation was found to have a strong influence on the contents of the nodule carbon metabolites. While levels of sucrose and succinate tended to decrease, a higher accumulation of malate was observed. These findings have provided evidence that N2 fixation of M. truncatula is mediated through an N feedback mechanism which is closely related to nodule carbon metabolism.
UnkovichM, HerridgeD, PeoplesM, CadischG, BoddeyB, GillerK, AlvesB, ChalkP (2008). Measuring Plant- associated Nitrogen Fixation in Agricultural Systems Australian Centre for International Agricultural Research, Canberra, Australia. [本文引用: 2]
VitousekPM, CassmanK, ClevelandC, CrewsT, FieldCB, GrimmNB, HowarthRW, MarinoR, MartinelliL, RastetterEB, SprentJI (2002). Towards an ecological understanding of biological nitrogen fixation //Boyer EW, Howarth RW. The Nitrogen Cycle at Regional to Global Scales. Kluwer Academic Publishers, Dordrecht, the Netherlands.1-45. [本文引用: 1]
VitousekPM, MengeDNL, ReedSC, ClevelandCC (2013). Biological nitrogen fixation: rates, patterns and ecological controls in terrestrial ecosystems Philosophical Transactions of the Royal Society B: Biological Sciences, 368, 20130119. DOI: 10.1098/rstb.2013.0119. DOI:10.1098/rstb.2013.0119URL [本文引用: 1]
WestJB, HilleRisLambersJ, LeeTD, HobbieSE, ReichPB (2005). Legume species identity and soil nitrogen supply determine symbiotic nitrogen-fixation responses to elevated atmospheric [CO2] New Phytologist, 167, 523-530. DOI:10.1111/nph.2005.167.issue-2URL [本文引用: 3]
WhitePJ, HammondJP (2008). Phosphorus nutrition of terrestrial plants //White PJ, Hammond JP. The Ecophysiology of Plant-Phosphorus Interactions. Springer, Dordrecht, the Netherlands. 51-81. [本文引用: 2]
WuN, LiZ, LiuH, TangM (2015). Influence of arbuscular mycorrhiza on photosynthesis and water status of Populus cathayana Rehder males and females under salt stress Acta Physiol Plant, 37, 183. DOI:10.1007/s11738-015-1932-6URL [本文引用: 1]
ZhanS, WangY, ZhuZ, LiW, BaiY (2017). Nitrogen enrichment alters plant N:P stoichiometry and intensifies phosphorus limitation in a steppe ecosystem Environmental and Experimental Botany, 134, 21-32. DOI:10.1016/j.envexpbot.2016.10.014URL [本文引用: 2]
ZhengM, ZhouZ, LuoY, ZhaoP, MoJ (2019). Global pattern and controls of biological nitrogen fixation under nutrient enrichment: a meta-analysis Global Change Biology, 25, 3018-3030. DOI:10.1111/gcb.v25.9URL [本文引用: 2]
ZhouLL, CaoJ, ZhangFS, LiL (2009). Rhizosphere acidification of faba bean, soybean and maize Science of the Total Environment, 407, 4356-4362. DOI:10.1016/j.scitotenv.2009.02.006URL [本文引用: 1]
ZouCM, WangYQ, LiuY, ZhangXH, TangS (2015). Responses of photosynthesis and growth to weak light regime in four legume species Chinese Journal of Plant Ecology, 39, 909-916. DOI:10.17521/cjpe.2015.0087URL [本文引用: 1]
... 生物固氮和吸收土壤氮是豆科植物两个可供选择的氮获取策略(Ledgard et al., 2001).如果排除其他环境限制, 土壤氮资源的状态是豆科植物氮获取策略选择的关键决定因素(West et al., 2005).氮添加提高了土壤氮资源有效性, 可能降低豆科植物为获取大气氮而对根和根瘤发育的投资.再者, 增加的土壤氮有效性可能将豆科草本植物间竞争的压力从地下转移到地上(Farrer & Suding, 2016), 诱使植物投资更多资源到地上生物量用以捕获光, 并因此限制根系发育(Aerts et al., 1991).本研究中, 对比无肥料添加, 氮添加提高了地上生物量, 并小幅降低根生物量, 已经表明了氮添加下豆科草本植物生物量分配的转移.同时, 我们的研究也发现氮添加降低了豆科草本植物根部非结构性碳水化合物的含量, 间接地表明土壤氮资源的增加可能改变了光合产物的分配, 促使豆科草本植物将更多的光合产物转移并储存在地上, 并导致根部碳水化合物营养储存的下降. ...
Global assessment of limitation to symbiotic nitrogen fixation by phosphorus availability in terrestrial ecosystems using a meta-analysis approach 3 2013
... 生物固氮是豆科草本植物的重要氮(N)吸收途径, 豆科草本植物的生物固氮率不仅决定其同化大气氮的总量(Augusto et al., 2013), 还通过改变土壤氮的相对消耗影响豆科草本植物生长的氮经济(Li et al., 2016a, 2020).外源氮输入和生物固氮的积累增加了土壤中氮的有效性(Galloway, 1998; Vitousek et al., 2013).然而, 增加的土壤氮有效性会潜在地降低豆科草本植物的生物固氮率(Salvagiotti et al., 2008; Nyfeler et al., 2011), 进而阻碍豆科草本植物对生态系统的进一步固氮输入.以往的研究表明, 豆科植物能够根据土壤氮肥力状况调整在固定大气氮和获取土壤氮方面的投资策略(Vitousek et al., 2002; West et al., 2005).这些研究认为生物固氮的代谢成本一般高于土壤氮吸收, 因此高土壤氮有效性下豆科植物倾向于增加根系对土壤氮的吸收, 但抑制代谢上高成本的根瘤发育和生物固氮(West et al., 2005).这一观点暗示当氮有效性增加, 豆科植物生物固氮被抑制将成为普遍现象.然而, 其他研究发现, 外源氮的输入并不总是下调豆科植物生物固氮率, 这意味着一些其他因素可能会改变氮富集对生物固氮率的影响(Reed et al., 2007; Zheng et al., 2019). ...
... 磷(P)对植物生长和代谢具有关键调控作用(White & Hammond, 2008; Png et al., 2017).因为根瘤发育和生物固氮过程的高磷消耗, 豆科植物对磷的供应尤为敏感(Batterman et al., 2013; Divito & Sadras, 2014).研究表明磷添加能够提高豆科植物根瘤发育和生物固氮率(Augusto et al., 2013; Maistry et al., 2013), 尤其在磷资源限制环境中(Betencourt et al., 2012).尽管氮、磷在土壤中有不同的生物地球化学循环过程, 增加的氮供应很可能改变土壤内在的氮-磷平衡(Elser et al., 2010; Pe?uelaset al., 2012), 提高植物磷需求, 驱动土壤磷的消耗(Güsewell,2004; Png et al., 2017), 并最终加剧植物磷匮乏.基于此, 氮增加下, 改变的土壤氮磷供给平衡很可能成为驱动豆科植物生物固氮率下降的额外机制.在这种情景下, 土壤中磷肥的状态能够潜在改变氮富集对豆科植物生物固氮的抑制作用.先前的研究发现当土壤磷有效性较低时, 氮增加抑制根系结瘤, 但当土壤磷有效性较高时, 氮增加能够促进根系结瘤(Gates & Wilson, 1974), 相似的结果也存在于其他研究中(Gentili & Huss-Danell, 2003).这些研究已经表明氮、磷在调控豆科植物固氮行为中的交互作用.然而, 对于氮-磷平衡的改变如何驱动植物生理变化, 并最终改变豆科植物根系结瘤和生物固氮率, 我们仍旧缺乏深入理解. ...
... 磷添加能够促进豆科植物生长和生物固氮(Augusto et al., 2013; Maistry et al., 2013), 主要有两个原因.首先, 磷是固氮豆科植物及其根瘤细胞构建的主要元素, 并且是生物固氮过程中关键的能量驱动要素(Sulieman et al., 2013; Divito & Sadras, 2014).此外, 磷的添加也促进豆科植物生长, 并相应增加其对氮的需求, 由此刺激根瘤生物量和生物固氮的增加以满足氮需要(Batterman et al., 2013).本研究结果证实磷添加显著提高豆科草本植物生物固氮率.进一步地, 本研究阐述了磷添加促进豆科草本植物固氮的一个新机制, 即磷添加降低豆科草本植物根系-菌根互作和根际磷活化, 因此促进根生物量和根碳水化合物的累积, 间接提高生物固氮率. ...
Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: evidence from Inner Mongolia grasslands 1 2010
How do phosphorus, potassium and sulphur affect plant growth and biological nitrogen fixation in crop and pasture legumes? A meta-analysis 3 2014
... 磷(P)对植物生长和代谢具有关键调控作用(White & Hammond, 2008; Png et al., 2017).因为根瘤发育和生物固氮过程的高磷消耗, 豆科植物对磷的供应尤为敏感(Batterman et al., 2013; Divito & Sadras, 2014).研究表明磷添加能够提高豆科植物根瘤发育和生物固氮率(Augusto et al., 2013; Maistry et al., 2013), 尤其在磷资源限制环境中(Betencourt et al., 2012).尽管氮、磷在土壤中有不同的生物地球化学循环过程, 增加的氮供应很可能改变土壤内在的氮-磷平衡(Elser et al., 2010; Pe?uelaset al., 2012), 提高植物磷需求, 驱动土壤磷的消耗(Güsewell,2004; Png et al., 2017), 并最终加剧植物磷匮乏.基于此, 氮增加下, 改变的土壤氮磷供给平衡很可能成为驱动豆科植物生物固氮率下降的额外机制.在这种情景下, 土壤中磷肥的状态能够潜在改变氮富集对豆科植物生物固氮的抑制作用.先前的研究发现当土壤磷有效性较低时, 氮增加抑制根系结瘤, 但当土壤磷有效性较高时, 氮增加能够促进根系结瘤(Gates & Wilson, 1974), 相似的结果也存在于其他研究中(Gentili & Huss-Danell, 2003).这些研究已经表明氮、磷在调控豆科植物固氮行为中的交互作用.然而, 对于氮-磷平衡的改变如何驱动植物生理变化, 并最终改变豆科植物根系结瘤和生物固氮率, 我们仍旧缺乏深入理解. ...
... 磷限制影响豆科植物根瘤细胞的构建, 并影响生物固氮过程中的能量供应(Sulieman et al., 2013; Divito & Sadras, 2014).然而, 并不清楚氮增加导致的相对磷匮乏是否已经经由降低植物和根瘤磷吸收来抑制根系结瘤和生物固氮率.此外, 在磷资源限制下, 许多陆地植物, 尤其豆科植物已经发展了物种特异性的适应机制, 诸如强化植物-菌根互作、刺激根际磷活化等来满足植物磷需求(Nuruzzaman, 2006; Zhou et al., 2009).进一步的实验证据表明上述磷获取策略的实现极大依赖于根系形态和生理的改变(Nuruzzaman, 2006; Betencourt et al., 2012; Png et al., 2017).然而, 氮增加是否激发了这些磷获取策略, 并借此改变豆科植物根系生理, 导致生物固氮率的下降不得而知. ...
... 磷添加能够促进豆科植物生长和生物固氮(Augusto et al., 2013; Maistry et al., 2013), 主要有两个原因.首先, 磷是固氮豆科植物及其根瘤细胞构建的主要元素, 并且是生物固氮过程中关键的能量驱动要素(Sulieman et al., 2013; Divito & Sadras, 2014).此外, 磷的添加也促进豆科植物生长, 并相应增加其对氮的需求, 由此刺激根瘤生物量和生物固氮的增加以满足氮需要(Batterman et al., 2013).本研究结果证实磷添加显著提高豆科草本植物生物固氮率.进一步地, 本研究阐述了磷添加促进豆科草本植物固氮的一个新机制, 即磷添加降低豆科草本植物根系-菌根互作和根际磷活化, 因此促进根生物量和根碳水化合物的累积, 间接提高生物固氮率. ...
Teasing apart plant community responses to N enrichment: the roles of resource limitation, competition and soil microbes 1 2016
... 生物固氮和吸收土壤氮是豆科植物两个可供选择的氮获取策略(Ledgard et al., 2001).如果排除其他环境限制, 土壤氮资源的状态是豆科植物氮获取策略选择的关键决定因素(West et al., 2005).氮添加提高了土壤氮资源有效性, 可能降低豆科植物为获取大气氮而对根和根瘤发育的投资.再者, 增加的土壤氮有效性可能将豆科草本植物间竞争的压力从地下转移到地上(Farrer & Suding, 2016), 诱使植物投资更多资源到地上生物量用以捕获光, 并因此限制根系发育(Aerts et al., 1991).本研究中, 对比无肥料添加, 氮添加提高了地上生物量, 并小幅降低根生物量, 已经表明了氮添加下豆科草本植物生物量分配的转移.同时, 我们的研究也发现氮添加降低了豆科草本植物根部非结构性碳水化合物的含量, 间接地表明土壤氮资源的增加可能改变了光合产物的分配, 促使豆科草本植物将更多的光合产物转移并储存在地上, 并导致根部碳水化合物营养储存的下降. ...
The global nitrogen cycle: changes and consequences 1 1998
... 生物固氮是豆科草本植物的重要氮(N)吸收途径, 豆科草本植物的生物固氮率不仅决定其同化大气氮的总量(Augusto et al., 2013), 还通过改变土壤氮的相对消耗影响豆科草本植物生长的氮经济(Li et al., 2016a, 2020).外源氮输入和生物固氮的积累增加了土壤中氮的有效性(Galloway, 1998; Vitousek et al., 2013).然而, 增加的土壤氮有效性会潜在地降低豆科草本植物的生物固氮率(Salvagiotti et al., 2008; Nyfeler et al., 2011), 进而阻碍豆科草本植物对生态系统的进一步固氮输入.以往的研究表明, 豆科植物能够根据土壤氮肥力状况调整在固定大气氮和获取土壤氮方面的投资策略(Vitousek et al., 2002; West et al., 2005).这些研究认为生物固氮的代谢成本一般高于土壤氮吸收, 因此高土壤氮有效性下豆科植物倾向于增加根系对土壤氮的吸收, 但抑制代谢上高成本的根瘤发育和生物固氮(West et al., 2005).这一观点暗示当氮有效性增加, 豆科植物生物固氮被抑制将成为普遍现象.然而, 其他研究发现, 外源氮的输入并不总是下调豆科植物生物固氮率, 这意味着一些其他因素可能会改变氮富集对生物固氮率的影响(Reed et al., 2007; Zheng et al., 2019). ...
The interaction of nitrogen and phosphorus on the growth, nutrient status and nodulation of Stylosanthes humilis H.B.K. (townsville stylo) 1 1974
... 磷(P)对植物生长和代谢具有关键调控作用(White & Hammond, 2008; Png et al., 2017).因为根瘤发育和生物固氮过程的高磷消耗, 豆科植物对磷的供应尤为敏感(Batterman et al., 2013; Divito & Sadras, 2014).研究表明磷添加能够提高豆科植物根瘤发育和生物固氮率(Augusto et al., 2013; Maistry et al., 2013), 尤其在磷资源限制环境中(Betencourt et al., 2012).尽管氮、磷在土壤中有不同的生物地球化学循环过程, 增加的氮供应很可能改变土壤内在的氮-磷平衡(Elser et al., 2010; Pe?uelaset al., 2012), 提高植物磷需求, 驱动土壤磷的消耗(Güsewell,2004; Png et al., 2017), 并最终加剧植物磷匮乏.基于此, 氮增加下, 改变的土壤氮磷供给平衡很可能成为驱动豆科植物生物固氮率下降的额外机制.在这种情景下, 土壤中磷肥的状态能够潜在改变氮富集对豆科植物生物固氮的抑制作用.先前的研究发现当土壤磷有效性较低时, 氮增加抑制根系结瘤, 但当土壤磷有效性较高时, 氮增加能够促进根系结瘤(Gates & Wilson, 1974), 相似的结果也存在于其他研究中(Gentili & Huss-Danell, 2003).这些研究已经表明氮、磷在调控豆科植物固氮行为中的交互作用.然而, 对于氮-磷平衡的改变如何驱动植物生理变化, 并最终改变豆科植物根系结瘤和生物固氮率, 我们仍旧缺乏深入理解. ...
Local and systemic effects of phosphorus and nitrogen on nodulation and nodule function in Alnus incana 2 2003
... 磷(P)对植物生长和代谢具有关键调控作用(White & Hammond, 2008; Png et al., 2017).因为根瘤发育和生物固氮过程的高磷消耗, 豆科植物对磷的供应尤为敏感(Batterman et al., 2013; Divito & Sadras, 2014).研究表明磷添加能够提高豆科植物根瘤发育和生物固氮率(Augusto et al., 2013; Maistry et al., 2013), 尤其在磷资源限制环境中(Betencourt et al., 2012).尽管氮、磷在土壤中有不同的生物地球化学循环过程, 增加的氮供应很可能改变土壤内在的氮-磷平衡(Elser et al., 2010; Pe?uelaset al., 2012), 提高植物磷需求, 驱动土壤磷的消耗(Güsewell,2004; Png et al., 2017), 并最终加剧植物磷匮乏.基于此, 氮增加下, 改变的土壤氮磷供给平衡很可能成为驱动豆科植物生物固氮率下降的额外机制.在这种情景下, 土壤中磷肥的状态能够潜在改变氮富集对豆科植物生物固氮的抑制作用.先前的研究发现当土壤磷有效性较低时, 氮增加抑制根系结瘤, 但当土壤磷有效性较高时, 氮增加能够促进根系结瘤(Gates & Wilson, 1974), 相似的结果也存在于其他研究中(Gentili & Huss-Danell, 2003).这些研究已经表明氮、磷在调控豆科植物固氮行为中的交互作用.然而, 对于氮-磷平衡的改变如何驱动植物生理变化, 并最终改变豆科植物根系结瘤和生物固氮率, 我们仍旧缺乏深入理解. ...
... 最近的meta分析发现, 氮增加对生物固氮的作用随生态系统类型、地理位置、气候条件等的变化而发生改变(Zheng et al., 2019).实际上这些大尺度环境因子可能已经通过改变土壤养分背景而调控了氮增加对生物固氮的影响.先前的研究发现氮增加有时并不下调豆科植物生物固氮率, 并将其归因为该生态系统具有较低的土壤氮水平, 导致该生态系统即使在氮增加情境下也处于持续的氮限制状态(Reed et al., 2007).通过本研究, 我们进一步指出土壤磷有效性调控N:P平衡, 对氮增加下的豆科草本植物生物固氮率具有重要调节作用(Gentili & Huss-Danell, 2003).实际上, 在陆地生态系统中, 磷对植物生长的限制可能远比之前认定的要普遍, 尤其在人类诱导的不平衡的氮、磷输入情景下(Mahowald et al., 2008; Vitousek et al., 2010; Li et al., 2016b), 由此磷对豆科植物生物固氮的调控功能将变得越来越重要. ...
N:P ratios in terrestrial plants: variation and functional significance 1 2004
... 磷(P)对植物生长和代谢具有关键调控作用(White & Hammond, 2008; Png et al., 2017).因为根瘤发育和生物固氮过程的高磷消耗, 豆科植物对磷的供应尤为敏感(Batterman et al., 2013; Divito & Sadras, 2014).研究表明磷添加能够提高豆科植物根瘤发育和生物固氮率(Augusto et al., 2013; Maistry et al., 2013), 尤其在磷资源限制环境中(Betencourt et al., 2012).尽管氮、磷在土壤中有不同的生物地球化学循环过程, 增加的氮供应很可能改变土壤内在的氮-磷平衡(Elser et al., 2010; Pe?uelaset al., 2012), 提高植物磷需求, 驱动土壤磷的消耗(Güsewell,2004; Png et al., 2017), 并最终加剧植物磷匮乏.基于此, 氮增加下, 改变的土壤氮磷供给平衡很可能成为驱动豆科植物生物固氮率下降的额外机制.在这种情景下, 土壤中磷肥的状态能够潜在改变氮富集对豆科植物生物固氮的抑制作用.先前的研究发现当土壤磷有效性较低时, 氮增加抑制根系结瘤, 但当土壤磷有效性较高时, 氮增加能够促进根系结瘤(Gates & Wilson, 1974), 相似的结果也存在于其他研究中(Gentili & Huss-Danell, 2003).这些研究已经表明氮、磷在调控豆科植物固氮行为中的交互作用.然而, 对于氮-磷平衡的改变如何驱动植物生理变化, 并最终改变豆科植物根系结瘤和生物固氮率, 我们仍旧缺乏深入理解. ...
How do plant roots acquire mineral nutrients? Chemical processes involved in the rhizosphere 2 1998
Grass-legume ratio can change soil carbon and nitrogen storage in a temperate steppe grassland 1 2016a
... 生物固氮是豆科草本植物的重要氮(N)吸收途径, 豆科草本植物的生物固氮率不仅决定其同化大气氮的总量(Augusto et al., 2013), 还通过改变土壤氮的相对消耗影响豆科草本植物生长的氮经济(Li et al., 2016a, 2020).外源氮输入和生物固氮的积累增加了土壤中氮的有效性(Galloway, 1998; Vitousek et al., 2013).然而, 增加的土壤氮有效性会潜在地降低豆科草本植物的生物固氮率(Salvagiotti et al., 2008; Nyfeler et al., 2011), 进而阻碍豆科草本植物对生态系统的进一步固氮输入.以往的研究表明, 豆科植物能够根据土壤氮肥力状况调整在固定大气氮和获取土壤氮方面的投资策略(Vitousek et al., 2002; West et al., 2005).这些研究认为生物固氮的代谢成本一般高于土壤氮吸收, 因此高土壤氮有效性下豆科植物倾向于增加根系对土壤氮的吸收, 但抑制代谢上高成本的根瘤发育和生物固氮(West et al., 2005).这一观点暗示当氮有效性增加, 豆科植物生物固氮被抑制将成为普遍现象.然而, 其他研究发现, 外源氮的输入并不总是下调豆科植物生物固氮率, 这意味着一些其他因素可能会改变氮富集对生物固氮率的影响(Reed et al., 2007; Zheng et al., 2019). ...
Forage nitrogen yield and soil nitrogen in artificial grasslands with varied Medicago seedling proportion 1 2020
... 生物固氮是豆科草本植物的重要氮(N)吸收途径, 豆科草本植物的生物固氮率不仅决定其同化大气氮的总量(Augusto et al., 2013), 还通过改变土壤氮的相对消耗影响豆科草本植物生长的氮经济(Li et al., 2016a, 2020).外源氮输入和生物固氮的积累增加了土壤中氮的有效性(Galloway, 1998; Vitousek et al., 2013).然而, 增加的土壤氮有效性会潜在地降低豆科草本植物的生物固氮率(Salvagiotti et al., 2008; Nyfeler et al., 2011), 进而阻碍豆科草本植物对生态系统的进一步固氮输入.以往的研究表明, 豆科植物能够根据土壤氮肥力状况调整在固定大气氮和获取土壤氮方面的投资策略(Vitousek et al., 2002; West et al., 2005).这些研究认为生物固氮的代谢成本一般高于土壤氮吸收, 因此高土壤氮有效性下豆科植物倾向于增加根系对土壤氮的吸收, 但抑制代谢上高成本的根瘤发育和生物固氮(West et al., 2005).这一观点暗示当氮有效性增加, 豆科植物生物固氮被抑制将成为普遍现象.然而, 其他研究发现, 外源氮的输入并不总是下调豆科植物生物固氮率, 这意味着一些其他因素可能会改变氮富集对生物固氮率的影响(Reed et al., 2007; Zheng et al., 2019). ...
Aggravated phosphorus limitation on biomass production under increasing nitrogen loading: a meta-analysis 1 2016b
... 最近的meta分析发现, 氮增加对生物固氮的作用随生态系统类型、地理位置、气候条件等的变化而发生改变(Zheng et al., 2019).实际上这些大尺度环境因子可能已经通过改变土壤养分背景而调控了氮增加对生物固氮的影响.先前的研究发现氮增加有时并不下调豆科植物生物固氮率, 并将其归因为该生态系统具有较低的土壤氮水平, 导致该生态系统即使在氮增加情境下也处于持续的氮限制状态(Reed et al., 2007).通过本研究, 我们进一步指出土壤磷有效性调控N:P平衡, 对氮增加下的豆科草本植物生物固氮率具有重要调节作用(Gentili & Huss-Danell, 2003).实际上, 在陆地生态系统中, 磷对植物生长的限制可能远比之前认定的要普遍, 尤其在人类诱导的不平衡的氮、磷输入情景下(Mahowald et al., 2008; Vitousek et al., 2010; Li et al., 2016b), 由此磷对豆科植物生物固氮的调控功能将变得越来越重要. ...
Global distribution of atmospheric phosphorus sources, concentrations and deposition rates, and anthropogenic impacts 1 2008
... 最近的meta分析发现, 氮增加对生物固氮的作用随生态系统类型、地理位置、气候条件等的变化而发生改变(Zheng et al., 2019).实际上这些大尺度环境因子可能已经通过改变土壤养分背景而调控了氮增加对生物固氮的影响.先前的研究发现氮增加有时并不下调豆科植物生物固氮率, 并将其归因为该生态系统具有较低的土壤氮水平, 导致该生态系统即使在氮增加情境下也处于持续的氮限制状态(Reed et al., 2007).通过本研究, 我们进一步指出土壤磷有效性调控N:P平衡, 对氮增加下的豆科草本植物生物固氮率具有重要调节作用(Gentili & Huss-Danell, 2003).实际上, 在陆地生态系统中, 磷对植物生长的限制可能远比之前认定的要普遍, 尤其在人类诱导的不平衡的氮、磷输入情景下(Mahowald et al., 2008; Vitousek et al., 2010; Li et al., 2016b), 由此磷对豆科植物生物固氮的调控功能将变得越来越重要. ...
N and P colimitation of N2-fixing and N-supplied fynbos legumes from the Cape Floristic Region 2 2013
... 磷(P)对植物生长和代谢具有关键调控作用(White & Hammond, 2008; Png et al., 2017).因为根瘤发育和生物固氮过程的高磷消耗, 豆科植物对磷的供应尤为敏感(Batterman et al., 2013; Divito & Sadras, 2014).研究表明磷添加能够提高豆科植物根瘤发育和生物固氮率(Augusto et al., 2013; Maistry et al., 2013), 尤其在磷资源限制环境中(Betencourt et al., 2012).尽管氮、磷在土壤中有不同的生物地球化学循环过程, 增加的氮供应很可能改变土壤内在的氮-磷平衡(Elser et al., 2010; Pe?uelaset al., 2012), 提高植物磷需求, 驱动土壤磷的消耗(Güsewell,2004; Png et al., 2017), 并最终加剧植物磷匮乏.基于此, 氮增加下, 改变的土壤氮磷供给平衡很可能成为驱动豆科植物生物固氮率下降的额外机制.在这种情景下, 土壤中磷肥的状态能够潜在改变氮富集对豆科植物生物固氮的抑制作用.先前的研究发现当土壤磷有效性较低时, 氮增加抑制根系结瘤, 但当土壤磷有效性较高时, 氮增加能够促进根系结瘤(Gates & Wilson, 1974), 相似的结果也存在于其他研究中(Gentili & Huss-Danell, 2003).这些研究已经表明氮、磷在调控豆科植物固氮行为中的交互作用.然而, 对于氮-磷平衡的改变如何驱动植物生理变化, 并最终改变豆科植物根系结瘤和生物固氮率, 我们仍旧缺乏深入理解. ...
... 磷添加能够促进豆科植物生长和生物固氮(Augusto et al., 2013; Maistry et al., 2013), 主要有两个原因.首先, 磷是固氮豆科植物及其根瘤细胞构建的主要元素, 并且是生物固氮过程中关键的能量驱动要素(Sulieman et al., 2013; Divito & Sadras, 2014).此外, 磷的添加也促进豆科植物生长, 并相应增加其对氮的需求, 由此刺激根瘤生物量和生物固氮的增加以满足氮需要(Batterman et al., 2013).本研究结果证实磷添加显著提高豆科草本植物生物固氮率.进一步地, 本研究阐述了磷添加促进豆科草本植物固氮的一个新机制, 即磷添加降低豆科草本植物根系-菌根互作和根际磷活化, 因此促进根生物量和根碳水化合物的累积, 间接提高生物固氮率. ...
Distribution of carboxylates and acid phosphatase and depletion of different phosphorus fractions in the rhizosphere of a cereal and three grain legumes 6 2006
... 磷限制影响豆科植物根瘤细胞的构建, 并影响生物固氮过程中的能量供应(Sulieman et al., 2013; Divito & Sadras, 2014).然而, 并不清楚氮增加导致的相对磷匮乏是否已经经由降低植物和根瘤磷吸收来抑制根系结瘤和生物固氮率.此外, 在磷资源限制下, 许多陆地植物, 尤其豆科植物已经发展了物种特异性的适应机制, 诸如强化植物-菌根互作、刺激根际磷活化等来满足植物磷需求(Nuruzzaman, 2006; Zhou et al., 2009).进一步的实验证据表明上述磷获取策略的实现极大依赖于根系形态和生理的改变(Nuruzzaman, 2006; Betencourt et al., 2012; Png et al., 2017).然而, 氮增加是否激发了这些磷获取策略, 并借此改变豆科植物根系生理, 导致生物固氮率的下降不得而知. ...
... ).进一步的实验证据表明上述磷获取策略的实现极大依赖于根系形态和生理的改变(Nuruzzaman, 2006; Betencourt et al., 2012; Png et al., 2017).然而, 氮增加是否激发了这些磷获取策略, 并借此改变豆科植物根系生理, 导致生物固氮率的下降不得而知. ...
Grass-legume mixtures can yield more nitrogen than legume pure stands due to mutual stimulation of nitrogen uptake from symbiotic and non-symbiotic sources 1 2011
... 生物固氮是豆科草本植物的重要氮(N)吸收途径, 豆科草本植物的生物固氮率不仅决定其同化大气氮的总量(Augusto et al., 2013), 还通过改变土壤氮的相对消耗影响豆科草本植物生长的氮经济(Li et al., 2016a, 2020).外源氮输入和生物固氮的积累增加了土壤中氮的有效性(Galloway, 1998; Vitousek et al., 2013).然而, 增加的土壤氮有效性会潜在地降低豆科草本植物的生物固氮率(Salvagiotti et al., 2008; Nyfeler et al., 2011), 进而阻碍豆科草本植物对生态系统的进一步固氮输入.以往的研究表明, 豆科植物能够根据土壤氮肥力状况调整在固定大气氮和获取土壤氮方面的投资策略(Vitousek et al., 2002; West et al., 2005).这些研究认为生物固氮的代谢成本一般高于土壤氮吸收, 因此高土壤氮有效性下豆科植物倾向于增加根系对土壤氮的吸收, 但抑制代谢上高成本的根瘤发育和生物固氮(West et al., 2005).这一观点暗示当氮有效性增加, 豆科植物生物固氮被抑制将成为普遍现象.然而, 其他研究发现, 外源氮的输入并不总是下调豆科植物生物固氮率, 这意味着一些其他因素可能会改变氮富集对生物固氮率的影响(Reed et al., 2007; Zheng et al., 2019). ...
... 磷限制影响豆科植物根瘤细胞的构建, 并影响生物固氮过程中的能量供应(Sulieman et al., 2013; Divito & Sadras, 2014).然而, 并不清楚氮增加导致的相对磷匮乏是否已经经由降低植物和根瘤磷吸收来抑制根系结瘤和生物固氮率.此外, 在磷资源限制下, 许多陆地植物, 尤其豆科植物已经发展了物种特异性的适应机制, 诸如强化植物-菌根互作、刺激根际磷活化等来满足植物磷需求(Nuruzzaman, 2006; Zhou et al., 2009).进一步的实验证据表明上述磷获取策略的实现极大依赖于根系形态和生理的改变(Nuruzzaman, 2006; Betencourt et al., 2012; Png et al., 2017).然而, 氮增加是否激发了这些磷获取策略, 并借此改变豆科植物根系生理, 导致生物固氮率的下降不得而知. ...
... 本实验中所用土壤磷含量较低, 可能已经限制了豆科草本植物的生长(Betencourt et al., 2012), 这一推断能够从控制盆中已经较高的根际磷活化效应(图3B)得以佐证.氮添加显著提高了豆科草本植物生物量和组织氮含量, 因为植物生长和代谢过程对氮、磷元素的匹配需求, 氮添加将潜在提高植物对磷的需要(White & Hammond, 2008; Png et al., 2017).然而, 当前的研究发现氮添加并未改变土壤中磷的供给.而这种氮磷供应的不平衡并未导致豆科草本植物降低植物和根瘤的磷含量, 反而激发了豆科草本植物根际磷的活化和吸收, 以满足增加的磷需求(图4)(Nuruzzaman et al., 2006; Betencourt et al., 2012).因为植物的养分吸收主要发生在根际(Hinsinger, 1998), 植物物种已经进化发展了一系列策略来提高根际磷吸收.已知的策略包括加强根系-菌根互作以提高根部磷吸收的范围和面积(Smith & Read, 2008); 或通过根际土壤酸化刺激钙结合无机磷的活化, 以增加根际有效磷数量, 这种作用在中性或类似于本研究的碱性土壤中可能尤为重要(Richardson et al., 2009; Betencourt et al., 2012).此外, 植物根部能够分泌一些磷活化有机物, 如柠檬酸等有机酸, 这些磷活化物质也能提高根际磷有效性(Nuruzzaman et al., 2006).当前研究发现氮添加下, 根际土壤pH显著下降, 而根部菌根侵染率明显增加, 这些结果表明氮添加已经激发了豆科草本植物多途径的磷截获策略.众所周知, 菌根的共生依赖于根部提供碳水化合物营养(Soper et al., 2019).此外, 根际土壤pH的下降一般得益于根呼吸和根际微生物呼吸的增加, 而增加的根呼吸和根际微生物呼吸提高了根内及其分泌到根际的碳水化合物的消耗(Hinsinger et al., 2003; Nuruzzaman et al., 2006).因此, 通过菌根共生和根际磷活化的途径来获取更多磷可能潜在降低根生物量和根碳水化合物的积累(图5).本研究中根生物量、根碳水化合物和豆科草本植物生物固氮率间存在正相关关系, 表明氮添加下, 豆科草本植物为提高磷吸收, 会抑制其生物固氮率. ...
... 更大的氮有效性提高植物磷需求, 增加了土壤磷对豆科草本植物生长和固氮的限制(Vitousek et al., 2010; Png et al., 2017).因此, 氮富集下补充磷资源对缓解生物固氮的磷限制尤为重要.这一推断被我们的结果所支持, 因为本研究发现氮添加情境下补充磷对豆科草本植物生物固氮率有更大的提升.目前, 由于不成比例的氮磷输入(N >> P), 全球变化和农业活动正在改变许多生态系统的氮磷平衡(Vitousek et al., 2010).根据我们的研究, 这样的氮富集可能导致豆科植物生物固氮发生不同程度的改变, 其变化程度取决于土壤磷状态.生态系统中磷有效性越低, 氮添加对生物固氮的抑制效应越大.因此, 在高土壤N:P或者高氮沉降速率的环境下, 为增加豆科草本植物生物固氮, 补充磷是明智的选择.本研究中, 平均所有豆科物种, 对比无营养添加, 氮和磷协同添加并未改变豆科草本植物生物固氮率(对照: 41% vs. 氮磷添加: 43%), 这一结果表明3.5 g·m-2的磷添加能够有效缓解14 g·m-2的氮添加对实验豆科草本植物生物固氮的抑制效应.这一结果对本研究区豆科草场的优化管理具有重要指导意义. ...
Controls over leaf litter and soil nitrogen fixation in two lowland tropical rain forests 2 2007
... 生物固氮是豆科草本植物的重要氮(N)吸收途径, 豆科草本植物的生物固氮率不仅决定其同化大气氮的总量(Augusto et al., 2013), 还通过改变土壤氮的相对消耗影响豆科草本植物生长的氮经济(Li et al., 2016a, 2020).外源氮输入和生物固氮的积累增加了土壤中氮的有效性(Galloway, 1998; Vitousek et al., 2013).然而, 增加的土壤氮有效性会潜在地降低豆科草本植物的生物固氮率(Salvagiotti et al., 2008; Nyfeler et al., 2011), 进而阻碍豆科草本植物对生态系统的进一步固氮输入.以往的研究表明, 豆科植物能够根据土壤氮肥力状况调整在固定大气氮和获取土壤氮方面的投资策略(Vitousek et al., 2002; West et al., 2005).这些研究认为生物固氮的代谢成本一般高于土壤氮吸收, 因此高土壤氮有效性下豆科植物倾向于增加根系对土壤氮的吸收, 但抑制代谢上高成本的根瘤发育和生物固氮(West et al., 2005).这一观点暗示当氮有效性增加, 豆科植物生物固氮被抑制将成为普遍现象.然而, 其他研究发现, 外源氮的输入并不总是下调豆科植物生物固氮率, 这意味着一些其他因素可能会改变氮富集对生物固氮率的影响(Reed et al., 2007; Zheng et al., 2019). ...
... 最近的meta分析发现, 氮增加对生物固氮的作用随生态系统类型、地理位置、气候条件等的变化而发生改变(Zheng et al., 2019).实际上这些大尺度环境因子可能已经通过改变土壤养分背景而调控了氮增加对生物固氮的影响.先前的研究发现氮增加有时并不下调豆科植物生物固氮率, 并将其归因为该生态系统具有较低的土壤氮水平, 导致该生态系统即使在氮增加情境下也处于持续的氮限制状态(Reed et al., 2007).通过本研究, 我们进一步指出土壤磷有效性调控N:P平衡, 对氮增加下的豆科草本植物生物固氮率具有重要调节作用(Gentili & Huss-Danell, 2003).实际上, 在陆地生态系统中, 磷对植物生长的限制可能远比之前认定的要普遍, 尤其在人类诱导的不平衡的氮、磷输入情景下(Mahowald et al., 2008; Vitousek et al., 2010; Li et al., 2016b), 由此磷对豆科植物生物固氮的调控功能将变得越来越重要. ...
Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms 1 2009
... 本实验中所用土壤磷含量较低, 可能已经限制了豆科草本植物的生长(Betencourt et al., 2012), 这一推断能够从控制盆中已经较高的根际磷活化效应(图3B)得以佐证.氮添加显著提高了豆科草本植物生物量和组织氮含量, 因为植物生长和代谢过程对氮、磷元素的匹配需求, 氮添加将潜在提高植物对磷的需要(White & Hammond, 2008; Png et al., 2017).然而, 当前的研究发现氮添加并未改变土壤中磷的供给.而这种氮磷供应的不平衡并未导致豆科草本植物降低植物和根瘤的磷含量, 反而激发了豆科草本植物根际磷的活化和吸收, 以满足增加的磷需求(图4)(Nuruzzaman et al., 2006; Betencourt et al., 2012).因为植物的养分吸收主要发生在根际(Hinsinger, 1998), 植物物种已经进化发展了一系列策略来提高根际磷吸收.已知的策略包括加强根系-菌根互作以提高根部磷吸收的范围和面积(Smith & Read, 2008); 或通过根际土壤酸化刺激钙结合无机磷的活化, 以增加根际有效磷数量, 这种作用在中性或类似于本研究的碱性土壤中可能尤为重要(Richardson et al., 2009; Betencourt et al., 2012).此外, 植物根部能够分泌一些磷活化有机物, 如柠檬酸等有机酸, 这些磷活化物质也能提高根际磷有效性(Nuruzzaman et al., 2006).当前研究发现氮添加下, 根际土壤pH显著下降, 而根部菌根侵染率明显增加, 这些结果表明氮添加已经激发了豆科草本植物多途径的磷截获策略.众所周知, 菌根的共生依赖于根部提供碳水化合物营养(Soper et al., 2019).此外, 根际土壤pH的下降一般得益于根呼吸和根际微生物呼吸的增加, 而增加的根呼吸和根际微生物呼吸提高了根内及其分泌到根际的碳水化合物的消耗(Hinsinger et al., 2003; Nuruzzaman et al., 2006).因此, 通过菌根共生和根际磷活化的途径来获取更多磷可能潜在降低根生物量和根碳水化合物的积累(图5).本研究中根生物量、根碳水化合物和豆科草本植物生物固氮率间存在正相关关系, 表明氮添加下, 豆科草本植物为提高磷吸收, 会抑制其生物固氮率. ...
Nitrogen uptake, fixation and response to fertilizer N in soybeans: a review 1 2008
... 生物固氮是豆科草本植物的重要氮(N)吸收途径, 豆科草本植物的生物固氮率不仅决定其同化大气氮的总量(Augusto et al., 2013), 还通过改变土壤氮的相对消耗影响豆科草本植物生长的氮经济(Li et al., 2016a, 2020).外源氮输入和生物固氮的积累增加了土壤中氮的有效性(Galloway, 1998; Vitousek et al., 2013).然而, 增加的土壤氮有效性会潜在地降低豆科草本植物的生物固氮率(Salvagiotti et al., 2008; Nyfeler et al., 2011), 进而阻碍豆科草本植物对生态系统的进一步固氮输入.以往的研究表明, 豆科植物能够根据土壤氮肥力状况调整在固定大气氮和获取土壤氮方面的投资策略(Vitousek et al., 2002; West et al., 2005).这些研究认为生物固氮的代谢成本一般高于土壤氮吸收, 因此高土壤氮有效性下豆科植物倾向于增加根系对土壤氮的吸收, 但抑制代谢上高成本的根瘤发育和生物固氮(West et al., 2005).这一观点暗示当氮有效性增加, 豆科植物生物固氮被抑制将成为普遍现象.然而, 其他研究发现, 外源氮的输入并不总是下调豆科植物生物固氮率, 这意味着一些其他因素可能会改变氮富集对生物固氮率的影响(Reed et al., 2007; Zheng et al., 2019). ...
Stoichiometric tracking of soil nutrients by a desert insect herbivore 1 2003
Growth and nodulation of symbiotic Medicago truncatula at different levels of phosphorus availability 2 2013
... 磷限制影响豆科植物根瘤细胞的构建, 并影响生物固氮过程中的能量供应(Sulieman et al., 2013; Divito & Sadras, 2014).然而, 并不清楚氮增加导致的相对磷匮乏是否已经经由降低植物和根瘤磷吸收来抑制根系结瘤和生物固氮率.此外, 在磷资源限制下, 许多陆地植物, 尤其豆科植物已经发展了物种特异性的适应机制, 诸如强化植物-菌根互作、刺激根际磷活化等来满足植物磷需求(Nuruzzaman, 2006; Zhou et al., 2009).进一步的实验证据表明上述磷获取策略的实现极大依赖于根系形态和生理的改变(Nuruzzaman, 2006; Betencourt et al., 2012; Png et al., 2017).然而, 氮增加是否激发了这些磷获取策略, 并借此改变豆科植物根系生理, 导致生物固氮率的下降不得而知. ...
... 磷添加能够促进豆科植物生长和生物固氮(Augusto et al., 2013; Maistry et al., 2013), 主要有两个原因.首先, 磷是固氮豆科植物及其根瘤细胞构建的主要元素, 并且是生物固氮过程中关键的能量驱动要素(Sulieman et al., 2013; Divito & Sadras, 2014).此外, 磷的添加也促进豆科植物生长, 并相应增加其对氮的需求, 由此刺激根瘤生物量和生物固氮的增加以满足氮需要(Batterman et al., 2013).本研究结果证实磷添加显著提高豆科草本植物生物固氮率.进一步地, 本研究阐述了磷添加促进豆科草本植物固氮的一个新机制, 即磷添加降低豆科草本植物根系-菌根互作和根际磷活化, 因此促进根生物量和根碳水化合物的累积, 间接提高生物固氮率. ...
Measuring Plant- associated Nitrogen Fixation in Agricultural Systems 2 2008
... 豆科草本植物组织中生物固氮率(BNF,%)采用如下公式计算(Unkovich et al., 2008): ...
... 式中,${{\delta }^{15}}$N表示植物组织相对大气的15N原子丰度变化.B代表豆科草本植物组织完全依赖大气氮生长时的${{\delta }^{\text{15}}}\text{N}$.本研究采用整株植物材料来评估生物固氮, 这时整株植株的15N丰度应与大气一致, 所以B等于0 (Unkovich et al., 2008).根际磷活化效应定义为根际土壤有效磷含量与非根际土壤有效磷含量的比值(Zhan et al., 2017). ...
Towards an ecological understanding of biological nitrogen fixation 1 2002
... 生物固氮是豆科草本植物的重要氮(N)吸收途径, 豆科草本植物的生物固氮率不仅决定其同化大气氮的总量(Augusto et al., 2013), 还通过改变土壤氮的相对消耗影响豆科草本植物生长的氮经济(Li et al., 2016a, 2020).外源氮输入和生物固氮的积累增加了土壤中氮的有效性(Galloway, 1998; Vitousek et al., 2013).然而, 增加的土壤氮有效性会潜在地降低豆科草本植物的生物固氮率(Salvagiotti et al., 2008; Nyfeler et al., 2011), 进而阻碍豆科草本植物对生态系统的进一步固氮输入.以往的研究表明, 豆科植物能够根据土壤氮肥力状况调整在固定大气氮和获取土壤氮方面的投资策略(Vitousek et al., 2002; West et al., 2005).这些研究认为生物固氮的代谢成本一般高于土壤氮吸收, 因此高土壤氮有效性下豆科植物倾向于增加根系对土壤氮的吸收, 但抑制代谢上高成本的根瘤发育和生物固氮(West et al., 2005).这一观点暗示当氮有效性增加, 豆科植物生物固氮被抑制将成为普遍现象.然而, 其他研究发现, 外源氮的输入并不总是下调豆科植物生物固氮率, 这意味着一些其他因素可能会改变氮富集对生物固氮率的影响(Reed et al., 2007; Zheng et al., 2019). ...
Biological nitrogen fixation: rates, patterns and ecological controls in terrestrial ecosystems 1 2013
... 生物固氮是豆科草本植物的重要氮(N)吸收途径, 豆科草本植物的生物固氮率不仅决定其同化大气氮的总量(Augusto et al., 2013), 还通过改变土壤氮的相对消耗影响豆科草本植物生长的氮经济(Li et al., 2016a, 2020).外源氮输入和生物固氮的积累增加了土壤中氮的有效性(Galloway, 1998; Vitousek et al., 2013).然而, 增加的土壤氮有效性会潜在地降低豆科草本植物的生物固氮率(Salvagiotti et al., 2008; Nyfeler et al., 2011), 进而阻碍豆科草本植物对生态系统的进一步固氮输入.以往的研究表明, 豆科植物能够根据土壤氮肥力状况调整在固定大气氮和获取土壤氮方面的投资策略(Vitousek et al., 2002; West et al., 2005).这些研究认为生物固氮的代谢成本一般高于土壤氮吸收, 因此高土壤氮有效性下豆科植物倾向于增加根系对土壤氮的吸收, 但抑制代谢上高成本的根瘤发育和生物固氮(West et al., 2005).这一观点暗示当氮有效性增加, 豆科植物生物固氮被抑制将成为普遍现象.然而, 其他研究发现, 外源氮的输入并不总是下调豆科植物生物固氮率, 这意味着一些其他因素可能会改变氮富集对生物固氮率的影响(Reed et al., 2007; Zheng et al., 2019). ...
Terrestrial phosphorus limitation: mechanisms, implications, and nitrogen-phosphorus interactions 3 2010
... 最近的meta分析发现, 氮增加对生物固氮的作用随生态系统类型、地理位置、气候条件等的变化而发生改变(Zheng et al., 2019).实际上这些大尺度环境因子可能已经通过改变土壤养分背景而调控了氮增加对生物固氮的影响.先前的研究发现氮增加有时并不下调豆科植物生物固氮率, 并将其归因为该生态系统具有较低的土壤氮水平, 导致该生态系统即使在氮增加情境下也处于持续的氮限制状态(Reed et al., 2007).通过本研究, 我们进一步指出土壤磷有效性调控N:P平衡, 对氮增加下的豆科草本植物生物固氮率具有重要调节作用(Gentili & Huss-Danell, 2003).实际上, 在陆地生态系统中, 磷对植物生长的限制可能远比之前认定的要普遍, 尤其在人类诱导的不平衡的氮、磷输入情景下(Mahowald et al., 2008; Vitousek et al., 2010; Li et al., 2016b), 由此磷对豆科植物生物固氮的调控功能将变得越来越重要. ...
... 更大的氮有效性提高植物磷需求, 增加了土壤磷对豆科草本植物生长和固氮的限制(Vitousek et al., 2010; Png et al., 2017).因此, 氮富集下补充磷资源对缓解生物固氮的磷限制尤为重要.这一推断被我们的结果所支持, 因为本研究发现氮添加情境下补充磷对豆科草本植物生物固氮率有更大的提升.目前, 由于不成比例的氮磷输入(N >> P), 全球变化和农业活动正在改变许多生态系统的氮磷平衡(Vitousek et al., 2010).根据我们的研究, 这样的氮富集可能导致豆科植物生物固氮发生不同程度的改变, 其变化程度取决于土壤磷状态.生态系统中磷有效性越低, 氮添加对生物固氮的抑制效应越大.因此, 在高土壤N:P或者高氮沉降速率的环境下, 为增加豆科草本植物生物固氮, 补充磷是明智的选择.本研究中, 平均所有豆科物种, 对比无营养添加, 氮和磷协同添加并未改变豆科草本植物生物固氮率(对照: 41% vs. 氮磷添加: 43%), 这一结果表明3.5 g·m-2的磷添加能够有效缓解14 g·m-2的氮添加对实验豆科草本植物生物固氮的抑制效应.这一结果对本研究区豆科草场的优化管理具有重要指导意义. ...
Legume species identity and soil nitrogen supply determine symbiotic nitrogen-fixation responses to elevated atmospheric [CO2] 3 2005
... 生物固氮是豆科草本植物的重要氮(N)吸收途径, 豆科草本植物的生物固氮率不仅决定其同化大气氮的总量(Augusto et al., 2013), 还通过改变土壤氮的相对消耗影响豆科草本植物生长的氮经济(Li et al., 2016a, 2020).外源氮输入和生物固氮的积累增加了土壤中氮的有效性(Galloway, 1998; Vitousek et al., 2013).然而, 增加的土壤氮有效性会潜在地降低豆科草本植物的生物固氮率(Salvagiotti et al., 2008; Nyfeler et al., 2011), 进而阻碍豆科草本植物对生态系统的进一步固氮输入.以往的研究表明, 豆科植物能够根据土壤氮肥力状况调整在固定大气氮和获取土壤氮方面的投资策略(Vitousek et al., 2002; West et al., 2005).这些研究认为生物固氮的代谢成本一般高于土壤氮吸收, 因此高土壤氮有效性下豆科植物倾向于增加根系对土壤氮的吸收, 但抑制代谢上高成本的根瘤发育和生物固氮(West et al., 2005).这一观点暗示当氮有效性增加, 豆科植物生物固氮被抑制将成为普遍现象.然而, 其他研究发现, 外源氮的输入并不总是下调豆科植物生物固氮率, 这意味着一些其他因素可能会改变氮富集对生物固氮率的影响(Reed et al., 2007; Zheng et al., 2019). ...
... ., 2005).这一观点暗示当氮有效性增加, 豆科植物生物固氮被抑制将成为普遍现象.然而, 其他研究发现, 外源氮的输入并不总是下调豆科植物生物固氮率, 这意味着一些其他因素可能会改变氮富集对生物固氮率的影响(Reed et al., 2007; Zheng et al., 2019). ...
... 生物固氮和吸收土壤氮是豆科植物两个可供选择的氮获取策略(Ledgard et al., 2001).如果排除其他环境限制, 土壤氮资源的状态是豆科植物氮获取策略选择的关键决定因素(West et al., 2005).氮添加提高了土壤氮资源有效性, 可能降低豆科植物为获取大气氮而对根和根瘤发育的投资.再者, 增加的土壤氮有效性可能将豆科草本植物间竞争的压力从地下转移到地上(Farrer & Suding, 2016), 诱使植物投资更多资源到地上生物量用以捕获光, 并因此限制根系发育(Aerts et al., 1991).本研究中, 对比无肥料添加, 氮添加提高了地上生物量, 并小幅降低根生物量, 已经表明了氮添加下豆科草本植物生物量分配的转移.同时, 我们的研究也发现氮添加降低了豆科草本植物根部非结构性碳水化合物的含量, 间接地表明土壤氮资源的增加可能改变了光合产物的分配, 促使豆科草本植物将更多的光合产物转移并储存在地上, 并导致根部碳水化合物营养储存的下降. ...
Phosphorus nutrition of terrestrial plants 2 2008
... 磷(P)对植物生长和代谢具有关键调控作用(White & Hammond, 2008; Png et al., 2017).因为根瘤发育和生物固氮过程的高磷消耗, 豆科植物对磷的供应尤为敏感(Batterman et al., 2013; Divito & Sadras, 2014).研究表明磷添加能够提高豆科植物根瘤发育和生物固氮率(Augusto et al., 2013; Maistry et al., 2013), 尤其在磷资源限制环境中(Betencourt et al., 2012).尽管氮、磷在土壤中有不同的生物地球化学循环过程, 增加的氮供应很可能改变土壤内在的氮-磷平衡(Elser et al., 2010; Pe?uelaset al., 2012), 提高植物磷需求, 驱动土壤磷的消耗(Güsewell,2004; Png et al., 2017), 并最终加剧植物磷匮乏.基于此, 氮增加下, 改变的土壤氮磷供给平衡很可能成为驱动豆科植物生物固氮率下降的额外机制.在这种情景下, 土壤中磷肥的状态能够潜在改变氮富集对豆科植物生物固氮的抑制作用.先前的研究发现当土壤磷有效性较低时, 氮增加抑制根系结瘤, 但当土壤磷有效性较高时, 氮增加能够促进根系结瘤(Gates & Wilson, 1974), 相似的结果也存在于其他研究中(Gentili & Huss-Danell, 2003).这些研究已经表明氮、磷在调控豆科植物固氮行为中的交互作用.然而, 对于氮-磷平衡的改变如何驱动植物生理变化, 并最终改变豆科植物根系结瘤和生物固氮率, 我们仍旧缺乏深入理解. ...
... 本实验中所用土壤磷含量较低, 可能已经限制了豆科草本植物的生长(Betencourt et al., 2012), 这一推断能够从控制盆中已经较高的根际磷活化效应(图3B)得以佐证.氮添加显著提高了豆科草本植物生物量和组织氮含量, 因为植物生长和代谢过程对氮、磷元素的匹配需求, 氮添加将潜在提高植物对磷的需要(White & Hammond, 2008; Png et al., 2017).然而, 当前的研究发现氮添加并未改变土壤中磷的供给.而这种氮磷供应的不平衡并未导致豆科草本植物降低植物和根瘤的磷含量, 反而激发了豆科草本植物根际磷的活化和吸收, 以满足增加的磷需求(图4)(Nuruzzaman et al., 2006; Betencourt et al., 2012).因为植物的养分吸收主要发生在根际(Hinsinger, 1998), 植物物种已经进化发展了一系列策略来提高根际磷吸收.已知的策略包括加强根系-菌根互作以提高根部磷吸收的范围和面积(Smith & Read, 2008); 或通过根际土壤酸化刺激钙结合无机磷的活化, 以增加根际有效磷数量, 这种作用在中性或类似于本研究的碱性土壤中可能尤为重要(Richardson et al., 2009; Betencourt et al., 2012).此外, 植物根部能够分泌一些磷活化有机物, 如柠檬酸等有机酸, 这些磷活化物质也能提高根际磷有效性(Nuruzzaman et al., 2006).当前研究发现氮添加下, 根际土壤pH显著下降, 而根部菌根侵染率明显增加, 这些结果表明氮添加已经激发了豆科草本植物多途径的磷截获策略.众所周知, 菌根的共生依赖于根部提供碳水化合物营养(Soper et al., 2019).此外, 根际土壤pH的下降一般得益于根呼吸和根际微生物呼吸的增加, 而增加的根呼吸和根际微生物呼吸提高了根内及其分泌到根际的碳水化合物的消耗(Hinsinger et al., 2003; Nuruzzaman et al., 2006).因此, 通过菌根共生和根际磷活化的途径来获取更多磷可能潜在降低根生物量和根碳水化合物的积累(图5).本研究中根生物量、根碳水化合物和豆科草本植物生物固氮率间存在正相关关系, 表明氮添加下, 豆科草本植物为提高磷吸收, 会抑制其生物固氮率. ...
Influence of arbuscular mycorrhiza on photosynthesis and water status of Populus cathayana Rehder males and females under salt stress 1 2015
... 式中,${{\delta }^{15}}$N表示植物组织相对大气的15N原子丰度变化.B代表豆科草本植物组织完全依赖大气氮生长时的${{\delta }^{\text{15}}}\text{N}$.本研究采用整株植物材料来评估生物固氮, 这时整株植株的15N丰度应与大气一致, 所以B等于0 (Unkovich et al., 2008).根际磷活化效应定义为根际土壤有效磷含量与非根际土壤有效磷含量的比值(Zhan et al., 2017). ...
Global pattern and controls of biological nitrogen fixation under nutrient enrichment: a meta-analysis 2 2019
... 生物固氮是豆科草本植物的重要氮(N)吸收途径, 豆科草本植物的生物固氮率不仅决定其同化大气氮的总量(Augusto et al., 2013), 还通过改变土壤氮的相对消耗影响豆科草本植物生长的氮经济(Li et al., 2016a, 2020).外源氮输入和生物固氮的积累增加了土壤中氮的有效性(Galloway, 1998; Vitousek et al., 2013).然而, 增加的土壤氮有效性会潜在地降低豆科草本植物的生物固氮率(Salvagiotti et al., 2008; Nyfeler et al., 2011), 进而阻碍豆科草本植物对生态系统的进一步固氮输入.以往的研究表明, 豆科植物能够根据土壤氮肥力状况调整在固定大气氮和获取土壤氮方面的投资策略(Vitousek et al., 2002; West et al., 2005).这些研究认为生物固氮的代谢成本一般高于土壤氮吸收, 因此高土壤氮有效性下豆科植物倾向于增加根系对土壤氮的吸收, 但抑制代谢上高成本的根瘤发育和生物固氮(West et al., 2005).这一观点暗示当氮有效性增加, 豆科植物生物固氮被抑制将成为普遍现象.然而, 其他研究发现, 外源氮的输入并不总是下调豆科植物生物固氮率, 这意味着一些其他因素可能会改变氮富集对生物固氮率的影响(Reed et al., 2007; Zheng et al., 2019). ...
... 最近的meta分析发现, 氮增加对生物固氮的作用随生态系统类型、地理位置、气候条件等的变化而发生改变(Zheng et al., 2019).实际上这些大尺度环境因子可能已经通过改变土壤养分背景而调控了氮增加对生物固氮的影响.先前的研究发现氮增加有时并不下调豆科植物生物固氮率, 并将其归因为该生态系统具有较低的土壤氮水平, 导致该生态系统即使在氮增加情境下也处于持续的氮限制状态(Reed et al., 2007).通过本研究, 我们进一步指出土壤磷有效性调控N:P平衡, 对氮增加下的豆科草本植物生物固氮率具有重要调节作用(Gentili & Huss-Danell, 2003).实际上, 在陆地生态系统中, 磷对植物生长的限制可能远比之前认定的要普遍, 尤其在人类诱导的不平衡的氮、磷输入情景下(Mahowald et al., 2008; Vitousek et al., 2010; Li et al., 2016b), 由此磷对豆科植物生物固氮的调控功能将变得越来越重要. ...
Rhizosphere acidification of faba bean, soybean and maize 1 2009
... 磷限制影响豆科植物根瘤细胞的构建, 并影响生物固氮过程中的能量供应(Sulieman et al., 2013; Divito & Sadras, 2014).然而, 并不清楚氮增加导致的相对磷匮乏是否已经经由降低植物和根瘤磷吸收来抑制根系结瘤和生物固氮率.此外, 在磷资源限制下, 许多陆地植物, 尤其豆科植物已经发展了物种特异性的适应机制, 诸如强化植物-菌根互作、刺激根际磷活化等来满足植物磷需求(Nuruzzaman, 2006; Zhou et al., 2009).进一步的实验证据表明上述磷获取策略的实现极大依赖于根系形态和生理的改变(Nuruzzaman, 2006; Betencourt et al., 2012; Png et al., 2017).然而, 氮增加是否激发了这些磷获取策略, 并借此改变豆科植物根系生理, 导致生物固氮率的下降不得而知. ...
 ,*, 黄迎新, 周道玮, 丛山
,*, 黄迎新, 周道玮, 丛山

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT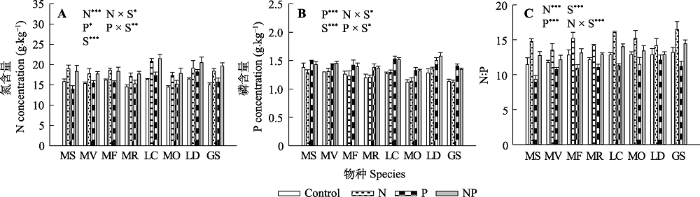 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT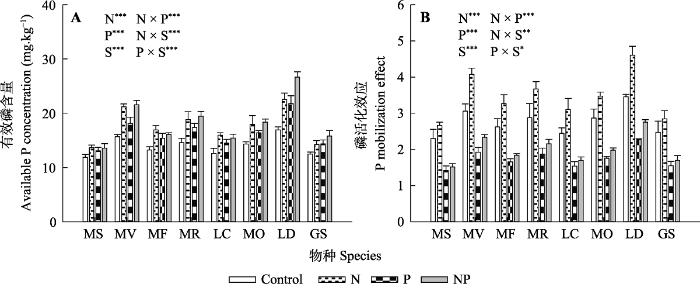 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT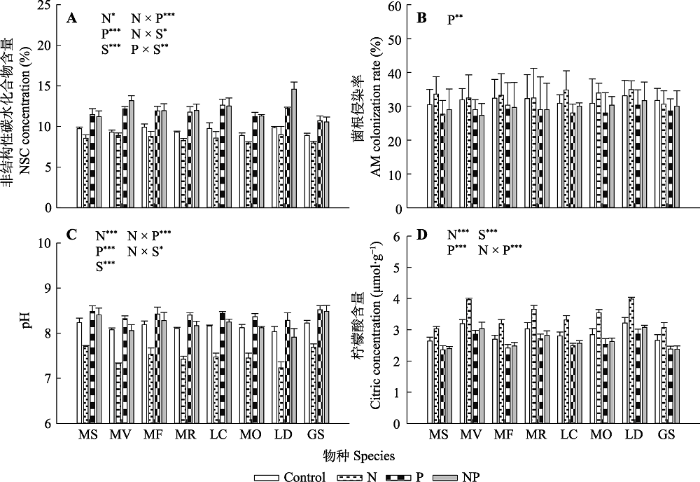 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT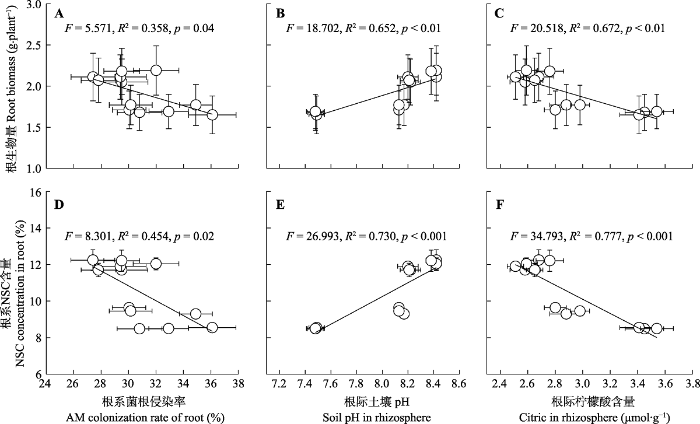 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT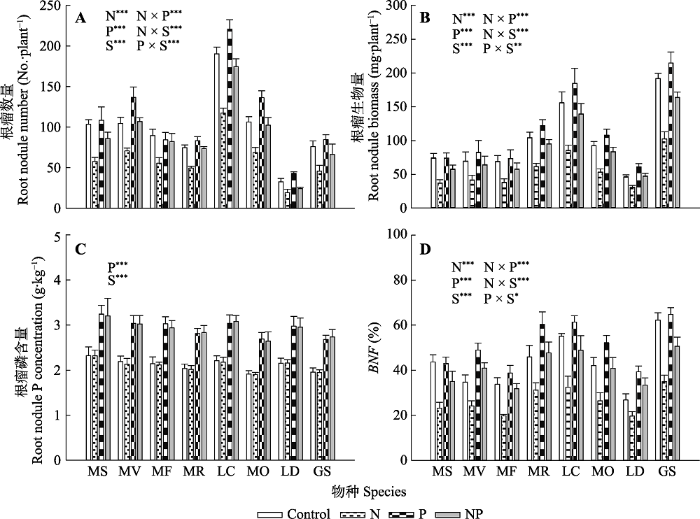 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT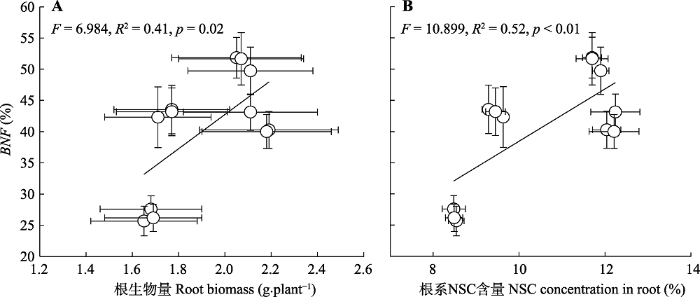 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT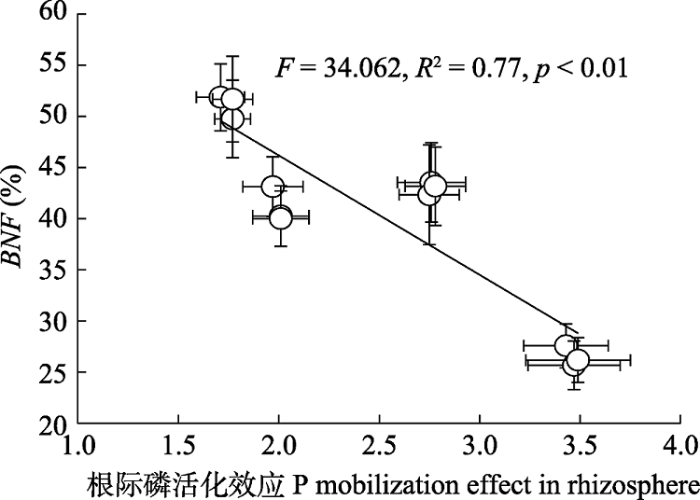 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}