朱湾湾
1 , 2 , 王攀
2 , 许艺馨
2 , 李春环
2 , 余海龙
2 , 黄菊莹
, , 1 , * 1 宁夏大学生态环境学院, 西北土地退化与生态恢复国家重点实验室培育基地, 西北退化生态系统恢复与重建教育部重点实验室, 银川 7500212 宁夏大学地理科学与规划学院, 银川 750021Soil enzyme activities and their influencing factors in a desert steppe of northwestern China under changing precipitation regimes and nitrogen addition Wan-Wan ZHU
1 , 2 , Pan WANG
2 , Yi-Xin XU
2 , Chun-Huan LI
2 , Hai-Long YU
2 , Ju-Ying HUANG
, , 1 , * 1 School of Ecology and Environmental Sciences, Ningxia University, Breeding Base for State Key Laboratory of Land Degradation and Ecological Restoration in Northwestern China, Key Laboratory for Restoration and Reconstruction of Degraded Ecosystems in Northwestern China of Ministry of Education, Yinchuan 750021, China2 School of Geography Science and Planning, Ningxia University, Yinchuan 750021, China通讯作者: * 黄菊莹: ORCID:0000-0002-1351-7282 (
juyinghuang@163.com )
编委: 程晓莉
责任编辑: 赵航
收稿日期: 2020-08-3
接受日期: 2020-12-22
网络出版日期: 2021-03-20
基金资助: 宁夏高等学校一流学科建设(生态学)项目 (NXYLXK2017B06 )国家自然科学基金 (31760144 )国家自然科学基金 (41961001 )
Corresponding authors: * (
juyinghuang@163.com )
Received: 2020-08-3
Accepted: 2020-12-22
Online: 2021-03-20
Fund supported: Project of First-Class Discipline Construction (Ecology) for Ningxia Higher Education (NXYLXK2017B06 )National Natural Science Foundation of China (31760144 )National Natural Science Foundation of China (41961001 )
摘要 土壤酶主要由植物根系和微生物分泌产生, 参与有机质降解和元素循环等重要过程。研究降水量变化和氮(N)添加下土壤酶活性及其与植物群落组成以及微生物活动间的联系, 可为深入理解全球变化背景下植被-土壤系统中元素的循环与转化机制提供科学依据。该研究基于2017年在宁夏荒漠草原设立的降水量变化(减少50%、减少30%、对照、增加30%以及增加50%)和N添加(0和5 g·m
-2 ·a
-1 )的野外试验, 研究了2018-2019年土壤蔗糖酶、脲酶和磷酸酶活性的变化, 分析了其与植物群落组成、微生物生态化学计量特征的关系。结果表明: 与减少降水量相比, 增加降水量对3种酶活性的影响较大, 但其效应与N添加以及年份存在交互作用。2018年增加降水量对3种酶活性的影响缺乏明显的规律性。2019年增加降水量不同程度地提高了3种酶活性。N添加对3种酶活性影响较小(尤其2019年); 草木樨状黄耆(
Astragalus melilotoides )生物量与脲酶和磷酸酶活性负相关。糙隐子草(
Cleistogenes squarrosa )生物量与3种酶活性正相关。除Patrick丰富度指数外, 植物群落多样性指数普遍与3种酶活性负相关; 对酶活性影响较大的因子包括土壤pH、土壤全磷(P)含量和微生物生物量碳(C):N:P。因此, 短期内降水量变化及N添加对荒漠草原土壤酶的影响较小(尤其在减少降水量条件下); 降水量增加及N添加通过提高植物生物量、改变植物多样性、调节微生物生物量元素平衡以及增强土壤P有效性, 直接影响着土壤酶活性。鉴于土壤酶种类的多样化和功能的复杂性, 今后还需结合多种酶活性的长期变化规律, 深入分析全球变化对酶活性的影响机制。
关键词: 干旱半干旱区 ;
降水格局改变 ;
大气氮沉降增加 ;
微生物生态化学计量特征 ;
植物群落多样性 Abstract Aims Soil enzymes, which are mainly produced by plant roots and soil microbes, involve in the organic matter degradation and element cycling and other key processes in plant-soil systems. Study on the relationships between soil enzyme activity and plant community composition and microbial activity under changing precipitation pattern and increasing nitrogen (N) deposition can provide a new insight for evaluating the influencing mechanism of global change on the biogeochemical cycling in plant-soil systems.
Methods Based on a field experiment involving five precipitation treatments (50% reduction, 30% reduction, natural precipitation, 30% increase, and 50% increase) and two N addition treatments (0 and 5 g·m
-2 ·a
-1 ) conducted in a desert steppe of Ningxia since 2017, the changes of soil enzyme activities (sucrase, urease, and phosphatase) were studied and their relationships with plant community composition and microbial ecological stoichiometry were analyzed in 2018 and 2019.
Important findings Compared with decreasing precipitation, increasing precipitation had greater impacts on the three enzyme activities, but its effects were interacted with N addition and sampling year. Increasing precipitation had no significant impacts on the three enzyme activities in 2018, but enhanced them in 2019. By contrast, N addition had less influences on the three enzyme activities, especially in 2019. The biomass of
Astragalus melilotoides was negatively correlated with urease and phosphatase activities, while the biomass of
Cleistogenes squarrosa had positive correlation with the three enzyme activities. Except the Patrick richness index, plant community diversity indices were generally negatively correlated with the three enzyme activities. Soil enzyme activities were more greatly affected by soil pH, soil total phosphorus (P), and microbial biomass carbon (C):N:P. Therefore, short-term precipitation change and N addition have little effects on the soil enzymes in the studied desert steppe (especially under reducing precipitation); increasing precipitation and N addition could pose direct influences on soil enzyme activities by increasing plant biomass, changing plant diversity, regulating microbial biomass ecological stoichiometry, and enhancing soil P availability. Given the diversity and functional complexity of soil enzymes, it is necessary to deeply analyze the influencing mechanism of global change on enzyme activities by measuring the long-term responses of various enzyme activities.
Keywords: arid and semi-arid region ;
changing precipitation regimes ;
increasing atmospheric nitrogen deposition ;
microbial ecological stoichiometry ;
plant community diversity PDF (1543KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 朱湾湾, 王攀, 许艺馨, 李春环, 余海龙, 黄菊莹. 降水量变化与氮添加下荒漠草原土壤酶活性及其影响因素. 植物生态学报, 2021, 45(3): 309-320. DOI:
10.17521/cjpe.2020.0264 ZHU Wan-Wan, WANG Pan, XU Yi-Xin, LI Chun-Huan, YU Hai-Long, HUANG Ju-Ying.
Soil enzyme activities and their influencing factors in a desert steppe of northwestern China under changing precipitation regimes and nitrogen addition .
Chinese Journal of Plant Ecology , 2021, 45(3): 309-320. DOI:
10.17521/cjpe.2020.0264 降水格局改变和大气氮(N)沉降增加是全球变化的两个重要表现(IPCC, 2013)。全球变暖背景下, 降水量在全球范围内呈增加趋势, 同时表现出时空分配不均和极端事件频发等特征(IPCC, 2013)。据报道, 我国西北地区的西部生态区降水量呈增加趋势, 东部生态区降水量有所降低(高继卿等,
2015 )。此外, 人类活动导致全球大气N沉降速率持续增加(Kanakidou
et al .,
2016 )。就中国而言, 虽然N沉降速率在一些南方省市有所下降, 但在西北部分区域呈增加趋势(Yu
et al .,
2017 ,
2019 )。西北干旱半干旱区生态系统稳定性差, 对环境变化敏感, 是我国典型的生态脆弱区之一。作为该区的主要限制因子, 降水和N调控着土壤资源有效性, 影响着植物生长和微生物活动等关键生态过程, 与植被-土壤系统的稳定性密切相关。目前, 国内已有较多研究探讨了降水和N添加对该脆弱生态区的影响, 尚缺乏二者交互作用的相关报道。鉴于降水在介导N沉降效应中发挥着重要作用(Lü
et al .,
2012 ), 因此有必要针对西北干旱半干旱区典型生态系统开展二者交互效应的研究。
土壤酶主要由植物根系和微生物分泌产生, 是土壤中元素生物地球化学循环的催化剂(马文文等,
2014 )。一方面, 土壤酶活性能够反映土壤元素(碳(C)、N、磷(P)等)转化和物质循环的速率(Burns
et al .,
2013 ), 并通过对土壤养分状况的影响改变植物养分策略。同时, 土壤酶能够实现植物营养元素和有机质的循环转化, 对土壤微环境的元素平衡和转化具有重要影响(张美曼等,
2020 )。另一方面, 土壤酶活性可表征微生物的养分需求、反映微生物在环境波动时维持养分平衡的应对策略(闫钟清等,
2017a )。土壤酶活性主要受土壤养分可利用性(李艳红等,
2020 )、植被类型(李静等,
2013 )和微生物活性(闫钟清等,
2017a )等因素的影响。降水和N添加通过调控土壤水分和养分有效性, 改变植物资源利用策略, 从而直接影响到植被结构(张金凤和许雨晴, 2016)和微生物活性(王杰等,
2014a ), 进而改变土壤酶活性(许华等,
2018 )。因此, 揭示降水量变化和N添加下土壤酶活性与植物和微生物的联系, 可为评价全球变化背景下植被-土壤系统功能提供新思路。
荒漠草原是我国西北干旱半干旱区主要的草原生态系统类型, 其植被结构简单、生态系统脆弱。宁夏荒漠草原主要分布于毛乌素沙地西南缘, 是区域重要的生态屏障。该生态系统降水量少、蒸发量大、N沉降临界负荷低, 降水和N是其植被生长和微生物活动的主要限制因子, 因此对降水格局改变和N沉降增加较为敏感(段雷等,
2002 ; 杨蓉和赵多平,
2018 )。那么, 降水量和N添加及其交互作用如何影响荒漠草原土壤酶活性, 降水量改变和N添加下土壤酶活性与植物群落组成以及微生物活动存在怎样的联系等问题, 都值得我们进行深入探讨。为此, 本项目组于2017年在宁夏荒漠草原设立了降水量改变及N添加的裂区试验, 研究了2018-2019年生长季旺盛期土壤酶活性的变化特点, 分析了其与植物群落组成以及微生物生物量C:N:P生态化学计量特征(其值高低与微生物活动的方向密切相关)的关系, 为充分认识降水量变化和N添加对土壤酶活性的影响机制提供基础数据, 并为深入理解全球变化背景下西北脆弱生态区的元素生物地球化学循环提供理论依据。
1 材料和方法 1.1 研究区概况 试验样地设立于宁夏盐池县柳杨堡乡杨寨子村围栏草地内(37.80° N, 107.45° E), 海拔约为1 367 m。该草地自1998年开始围封, 属于黄土高原向鄂尔多斯台地的过渡地带, 具有典型的温带大陆性气候: 年蒸发量为2 131.8 mm; 年降水量为289.4 mm。降水季节分配不均, 多集中在生长季的5-9月; 年平均气温为7.7 ℃, 1月和7月平均气温分别为-8.9和22.5 ℃。土壤类型为灰钙土, 土壤质地为沙壤土, 且土壤pH偏高, 呈碱性。研究区土壤基本理化性质见
表1 。植物群落结构简单, 植被组成以一年生或多年生草本为主, 优势物种包括猪毛蒿(
Artemisia scoparia )、牛枝子(
Lespedeza potaninii )和草木樨状黄耆(
Astragalus melilotoides )等, 常见物种包括白草(
Pennisetum centrasiaticum )、针茅(
Stipa capillata )和糙隐子草(
Cleistogenes squarrosa )等。
Table 1 表1 表1 降水和氮(N)添加处理前试验样地0-20 cm土壤理化性质(平均值±标准误)
Table 1
Soil physical and chemical properties in 0-20 cm layer of experimental field before precipitation and nitrogen (N) addition treatments (mean ± SE) pH 有机碳含量 -1 ) 全氮含量 -1 ) 全磷含量 -1 ) NH4 + -N含量 -1 ) NO3 - -N含量 -1 ) 速效磷含量 -1 ) 8.85 ± 0.01 3.70 ± 0.11 0.48 ± 0.00 0.34 ± 0.01 1.83 ± 0.07 9.96 ± 0.59 2.55 ± 0.29
新窗口打开 |
下载CSV 1.2 试验设计 于2017年9月, 在围栏草地内选择30 m × 50 m的区域作为降水量和N添加处理的试验样地。降水增减量依据近几十年来我国西北地区的西部生态区降水量增加而东部生态区降水量减少的趋势进行设置, 同时参考了国内同类研究方法(Xu
et al .,
2018 )。氮肥施用量依据区域N沉降水平(顾峰雪等,
2016 )和N饱和阈值(Tian
et al .,
2016 )进行设置。试验设计采用随机区组的裂区设计, 主区为降水量, 副区为N添加。降水量处理包括减少50% (W1)、减少30% (W2)、对照(自然降水量, W3)、增加30% (W4)和增加50% (W5)。N添加处理包括0 g·m
-2 ·a
-1 (N0)和5 g·m
-2 ·a
-1 (N5)。试验包含10个处理, 每个处理3次重复, 共计30个小区(
表2 )。每个主区面积为8 m × 8 m, 每个副区面积为4 m × 8 m。各主区之间设置1.5 m的缓冲带。各主区的两个副区间垂直下埋1 m宽的塑料布, 上埋20 cm宽的彩钢板, 用来防止试验干扰。
Table 2 表2 表2 试验处理及其代码
Table 2
Experimental treatments and their abbreviations 试验处理 降水量减少50% 降水量减少30% 自然降水量 降水量增加30% 降水量增加50% 0 g N·m-2 ·a-1 W1N0 W2N0 W3N0 W4N0 W5N0 5 g N·m-2 ·a-1 W1N5 W2N5 W3N5 W4N5 W5N5
新窗口打开 |
下载CSV 降水量减少处理采用自制的减雨架进行全年遮雨。减雨架上端离地约1.8 m, 用5 cm宽的瓦面状高透光性聚氯乙烯板搭建分别相当于50%和30%小区面积的遮雨面。降水量增加处理采用人工喷灌装置实现。喷水时间为每年5-8月的每月月初和月中。选用分析纯NH
4 NO
3 (34% N)进行N添加处理。施N时间为每年5-8月的每月月初。详细的试验处理方法见朱湾湾等(
2019 )。
1.3 样品采集与测定 分别于2018和2019年8月下旬, 采用内径为5 cm的土钻收集每个小区0-20 cm土壤样品。每个小区内随机取3钻, 混匀后分为两部分。一部分装入铝盒中, 采用称质量法进行含水量的测定。另一部分过2 mm标准筛后再分装为两部分: 一部分置于冰箱中4 ℃下冷藏保存, 用于蔗糖酶活性、脲酶活性、磷酸酶活性、pH、电导率、NH
4 + -N、NO
3 - -N、速效P含量、微生物生物量的测定; 另一部分自然风干后, 用于有机C、全N和全P含量的测定。
各指标的测定中, 蔗糖酶活性采用3,5二硝基水杨酸比色法, 脲酶活性采用苯酚钠-次氯酸钠比色法, 磷酸酶活性采用对硝基苯磷酸盐法(刘红梅等,
2018 ), 有机C含量采用重铬酸钾容量法-外加热法, 全N含量采用凯氏定氮法, 全P含量采用钼锑抗比色法, pH采用酸度计法, 电导率采用便携式电导率仪, NH
4 + -N和NO
3 - -N含量采用连续流动分析仪(Auto Analyzer 3, SEAL Analytical GmbH, Hanau, Germany), 速效P含量采用0.5 mol·L
-1 NaHCO
3 法, 微生物生物量C含量采用氯仿熏蒸-K
2 SO
4 浸提-碳分析仪器法, 微生物生物量N含量采用氯仿熏蒸-K
2 SO
4 提取-流动注射氮分析仪器法, 微生物生物量P含量采用氯仿熏蒸-K
2 SO
4 提取-Pi测定-外加Pi矫正法。
同期, 在每个小区内随机选取1 m × 1 m的小样方进行植被群落调查。调查指标包括物种数和物种高度等。调查结束后, 齐平地面剪下地上部分, 按牛枝子、草木樨状黄耆、白草、针茅、糙隐子草及其他物种归类分装于纸袋中带回实验室烘干(65 ℃, 48 h)称质量。群落生物量为所有物种生物量之和。植物群落多样性采用Patrick丰富度指数、Shannon- Wiener多样性指数、Simpson优势度指数和Pielou均匀度指数等指标来表征, 具体计算方法见朱湾湾等(
2019 )。2018年植物生物量、群落多样性以及土壤理化性质引自朱湾湾等(
2019 )。2019年植物生物量、群落多样性以及土壤理化性质的结果见附录I-IV。
1.4 数据分析 采用SigmaPlot 12.0进行图的绘制。采用SPSS 19.0进行数据的统计分析: 采用K-S检验进行数据的正态分布检验; 采用三因素方差分析法分析年份、降水量、N添加及其交互作用对土壤酶活性的影响; 采用单因素方差分析进行相同N添加水平下各指标在不同降水量处理间, 以及相同降水量下各指标在不同N处理间的差异分析。若方差为齐性, 选用最小显著性差异(
LSD )法, 否则选用Games-Howell法; 由于植物生物量能够影响土壤酶活性(杨万勤和王开运,
2002 ), 故采用线性回归方程拟合土壤酶活性与植物种群生物量以及群落多样性之间的关系。采用Canoco 5.0进行土壤酶活性与微生物生物量C:N:P生态化学计量特征及其他环境因子对应关系的冗余分析(RDA)。分析前, 先对数据进行对数转换, 以减少数据间差异。以全部环境因子作为解释变量, 以土壤酶活性作为响应变量, 依据解释变量前项选择, 剔除影响力较小的解释变量后进行RDA, 并通过蒙特卡洛置换检验得出每个环境因子的条件效应。
2 结果和分析 2.1 降水量及N添加对土壤酶活性的影响 年份、降水量处理、N添加以及年份与降水量的交互作用对蔗糖酶和脲酶活性有极显著影响(
表3 ,
p < 0.01)。降水量与N添加以及年份、降水量与N添加对脲酶活性有显著的交互作用(
p < 0.05)。年份、降水量以及二者与N添加的交互作用对磷酸酶活性有极显著影响(
p < 0.01)。
Table 3 表3 表3 年份(α)、降水量(β)、氮添加(γ)及其交互作用对土壤酶活性的影响
Table 3
Effects of sampling year (α), precipitation (β), nitrogen addition (γ), and their interactions on soil enzyme activities 差异来源 α β γ α × β α × γ β × γ α × β × γ SA 25.571** 4.421** 24.767** 5.802** 0.179 0.433 0.472 UA 385.536** 5.662** 8.980** 4.755** 0.563 3.530* 2.959* PA 186.338** 7.070** 3.716 16.133 1.207 1.143 3.945**
表中数据为F 值。SA、UA和PA分别代表蔗糖酶、脲酶和磷酸酶活性。*和**分别代表显著性水平小于0.05和0.01。 Data in the table are F values. SA, UA, and PA represent sucrase, urease, and phosphatase activities, respectively. * and ** indicate that the significant levels are less than 0.05 and 0.01, respectively. 新窗口打开 |
下载CSV 2018年, 与减少降水量相比, 增加降水量对3种酶活性影响较大, 且其影响程度与N添加有关(
图1 ): 在0 g·m
-2 ·a
-1 的N添加水平下, 与自然降水量相比, 仅增加30%降水量显著影响了脲酶活性(
p < 0.05), 其他处理对3种酶活性无显著影响(
p > 0.05); 在5 g·m
-2 ·a
-1 的N添加水平下, 与自然降水量相比, 增减30%和50%降水量对蔗糖酶活性无显著影响(
p > 0.05), 而增减30%和50%降水量均显著降低了脲酶活性, 增减50%降水量均显著降低了磷酸酶活性(
p < 0.05); 两个N添加水平间, 自然降水量以及增加50%降水量条件下N添加显著提高了蔗糖酶活性(
p < 0.05), 自然降水量以及增减50%降水量条件下N添加显著提高了脲酶活性(
p < 0.05), 减少50%降水量条件下N添加显著降低了磷酸酶活性(
p < 0.05)。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图12018和2019年降水量及氮(N)添加对土壤酶活性的影响(平均值±标准误)。W1、W2、W3、W4和W5分别代表降水量减少50%、减少30%、自然降水、增加30%和增加50%。N0和N5分别代表0和5 g·m-2 ·a-1 的N添加水平。不同小写字母表示相同N添加水平下降水量处理间各指标存在显著性差异(p < 0.05)。*表示相同降水量下N添加处理间各指标存在显著性差异(p < 0.05)。 Fig. 1Effects of precipitation and nitrogen addition on soil enzyme activities in 2018 and 2019 (mean ± SE). W1, W2, W3, W4, and W5 represent 50% reduction in precipitation, 30% reduction in precipitation, natural precipitation (control), 30% increase in precipitation, 50% increase in precipitation, respectively. N0 and N5 represent 0 and 5 g·m -2 ·a-1 of nitrogen addition level, respectively. Different lowercase letters indicate significant difference (p < 0.05) of indices among precipitation treatments under the same nitrogen addition level. * indicates significant difference (p < 0.05) of indices among nitrogen addition treatments under the same precipitation. 2019年, 与自然降水量相比, 减少降水量对3种酶活性无显著影响(
p > 0.05), 增加降水量不同程度地提高了3种酶活性, 但其效应在N添加处理间差异较小(
图1 ): 在0 g·m
-2 ·a
-1 的N添加水平下, 与自然降水量相比, 增加30%和50%降水量均显著提高了蔗糖酶活性(
p < 0.05), 增加30%降水量显著提高了脲酶活性(
p < 0.05); 在5 g·m
-2 ·a
-1 的N添加水平下, 与自然降水量相比, 增加30%降水量显著增加了脲酶活性(
p < 0.05), 增加30%和50%降水量均显著增加了磷酸酶活性(
p < 0.05); 两个N添加水平间, 自然降水量条件下N添加显著提高了蔗糖酶活性(
p < 0.05), 减少30%和50%降水量条件下N添加显著提高了脲酶活性(
p < 0.05)。
2.2 土壤酶活性与植物群落组成的关系 图2 中, 蔗糖酶活性随着白草以及糙隐子草生物量的增加而增加(
p < 0.05); 脲酶活性随着草木樨状黄耆生物量的增加而减少, 随着糙隐子草生物量的增加而增加(
p < 0.05); 磷酸酶活性随着草木樨状黄耆生物量的增加而减少, 随着糙隐子草生物量的增加而增加(
p < 0.05)。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2土壤酶活性与植物生物量的关系。A、B、C、D和E分别表示牛枝子、草木樨状黄耆、白草、针茅和糙隐子草的种群生物量。F代表群落生物量。SA、UA和PA分别代表蔗糖酶、脲酶和磷酸酶活性。*、**和***分别表示p < 0.05、p < 0.01和p < 0.001。 Fig. 2Relationships between soil enzyme activities and plant biomass. A, B, C, D and E represent the population biomass of Lespedeza potaninii, Astragalus melilotoides, Pennisetum centrasiaticum, Stipa capillata, Cleistogenes squarrosa, respectively. F represents the community biomass. SA, UA, and PA represent sucrase, urease, and phosphatase activities, respectively. *, ** and *** indicate p < 0.05, p < 0.01 and p < 0.001, respectively. 如
图3 所示, 蔗糖酶活性随着Patrick丰富度指数的增加而增加、随着Pielou均匀度指数以及Simpson优势度指数的增加而减小(
p < 0.05); 脲酶活性随着Shannon-Wiener多样性指数以及Pielou均匀度指数的增加而减小(
p < 0.05); 磷酸酶活性随着Shannon-Wiener多样性指数、Pielou均匀度指数以及Simpson优势度指数的增加而减小(
p < 0.05)。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3荒漠草原土壤酶活性与植物群落多样性的关系。SA、UA和PA分别代表蔗糖酶、脲酶和磷酸酶活性。*、**和***分别表示p < 0.05、p < 0.01和p < 0.001。 Fig. 3Relationships between soil enzyme activities and plant community diversity in desert steppe. SA, UA, and PA represent sucrase, urease, and phosphatase activities, respectively. *, ** and *** indicate p < 0.05, p < 0.01 and p < 0.001, respectively. 2.3 土壤酶活性与微生物生态化学计量特征及其他环境因子的冗余分析 两个典范轴累计方差解释率为64%。对土壤酶活性影响显著的因子包括土壤pH、土壤全P含量、微生物生物量C:P、微生物生物量C:N、土壤N:P、微生物生物量C、微生物生物量P含量、土壤C:P、土壤NH
4 + -N、微生物生物量N和土壤NO
3 - -N含量(
表4 ;
图4 )。其中, 蔗糖酶活性与土壤pH、土壤全P、微生物生物量P、微生物生物量N以及土壤NO
3 - -N含量呈较强的正相关关系, 与微生物生物量C:P、微生物生物量C:N、土壤N:P、微生物生物量C含量、土壤C:P以及土壤NH
4 + -N含量呈较强的负相关关系; 脲酶和磷酸酶活性与以上因子的关系均与蔗糖酶活性的相反。
Table 4 表4 表4 冗余分析中各环境因子的条件效应
Table 4
Conditional effects of environmental factors in redundancy analysis 指标 pH TP C:Pm C:Nm N:Ps MBC MBP C:Ps NH4 + -N MBN NO3 - -N N:Pm SWC SOC TN EC AP C:Ns F 53.7 37.3 28.4 28.0 27.2 22.4 20.1 18.4 17.9 16.4 5.9 2.5 1.1 1.0 0.9 0.4 0.3 0.2 p 0.002 0.002 0.002 0.002 0.002 0.002 0.002 0.002 0.002 0.002 0.016 0.110 0.266 0.318 0.356 0.638 0.676 0.776
SOC 、TN 、TP 、C:Ns 、C:Ps 和N:Ps 分别代表土壤有机碳(C)、全氮(N)、全磷(P)含量、C:N、C:P和N:P。MBC 、MBN 、MBP 、C:Nm 、C:Pm 和N:Pm 分别代表微生物生物量C、N、P含量、C:N、C:P和N:P。NH4 + -N、NO3 - -N、AP 、EC 和SWC 分别代表土壤NH4 + -N、NO3 - -N、速效P含量、电导率和含水量。 SOC, TN, TP, C:Ns , C:Ps and N:Ps represent soil organic carbon (C), total nitrogen (N), total phosphorus (P) content, C:N, C:P and N:P, respectively. MBC, MBN, MBP, C:Nm , C:Pm and N:Pm represent microbial biomass C, N, P content, C:N, C:P and N:P, respectively. NH4 + -N, NO3 - -N, AP, EC and SWC represent soil ammonium nitrogen, nitrate nitrogen, available P content, conductivity, and water content. 新窗口打开 |
下载CSV 图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4土壤酶活性与微生物生态化学计量特征及其他环境因子的冗余分析(RDA)。SA、UA和PA分别代表蔗糖酶、脲酶和磷酸酶活性。TP、C:Ps 和N:Ps 分别代表土壤全P含量、C:P和N:P。MBC、MBN、MBP、C:Nm 和C:Pm 分别代表微生物生物量C、N、P含量, C:N和C:P。NH4 + -N、NO3 - -N分别代表土壤NH4 + -N、NO3 - -N含量。 Fig. 4Redundancy analysis (RDA) of soil enzyme activities and microbial ecological stoichiometry and other environmental factors. SA, UA, and PA represent sucrase, urease, and phosphatase activities, respectively. TP, C:Ps and N:Ps represent soil total phosphorus content, C:P and N:P, respectively. MBC, MBN, MBP, C:Nm and C:Pm represent microbial biomass carbon, nitrogen, phosphorus content, C:N and C:P, respectively. NH4 + -N, NO3 - -N represent soil ammonium nitrogen, nitrate nitrogen content, respectively. 3 讨论 3.1 土壤酶活性对降水量及N添加的响应 本研究中, 减少降水量对蔗糖酶、脲酶和磷酸酶活性的影响较小, 增加降水量不同程度地影响3种酶的活性。这可能是由于研究区长期处于低水平的土壤水分环境下, 植物和微生物对干旱具有高的适应性(胡小文等,
2004 ; 朱义族等,
2019 ), 因而减少降水量对其生长繁殖以及酶的分泌影响较小。相比之下, 在缺水环境下, 增加降水量提高了土壤含水量(附录III), 从而加速凋落物淋溶、促进表层凋落物和腐殖质中可溶性物质进入土壤(豆鹏鹏等,
2018 ), 使微生物可利用底物增加, 微生物活性相应增强, 进而刺激微生物向环境中释放酶(Schimel
et al .,
2007 ), 即微生物分泌的酶随之增加。此外, 本研究发现增加降水量对土壤酶活性的影响与N添加存在交互作用, 与其他研究结果(王杰等,
2014a )相似。这表明水分有利于N肥效的释放, 二者的交互作用促进了植物生长(白春利等,
2013 )、凋落物积累以及微生物活动(王杰等,
2014b ), 进而刺激土壤酶的分泌、导致土壤酶活性增强(闫钟清等,
2017b )。
3.2 土壤酶活性与植物群落组成的关系 本研究中, 降水量和N添加下脲酶和磷酸酶活性与草木樨状黄耆(豆科植物)生物量呈负线性关系, 而蔗糖酶、脲酶和磷酸酶活性均与糙隐子草(禾本科植物)生物量呈正线性关系, 与针对高寒草甸的研究结果(王长庭等,
2010 )存在差异。首先, 土壤酶活性与植物生物量之间的关系因植物种不同而异。这是由于不同植物根系的发达程度以及分泌物存在差异, 进而影响土壤酶活性(王小平等,
2018 )。豆科植物与根瘤菌的共生作用可使其在相对干旱的条件下快速生长(张仁懿等,
2014 )。这不仅消耗了土壤养分, 而且抑制了微生物活性以及酶活性。相比之下, 降水量增加和N添加显著地刺激了禾本科植物的生长(附录I; 朱湾湾等,
2019 ), 其自身的营养物质归还土壤后可为土壤酶的合成提供丰富的底物(乔文静等,
2018 )。其次, 降水量增加及N添加下植物生物量增加(附录I; 朱湾湾等,
2019 ), 提高了凋落物C和养分向土壤中的归还量(Zhong
et al .,
2017 ), 从而促进了蔗糖酶、脲酶和磷酸酶的产生。
研究表明, 土壤酶活性与植物群落多样性亦存在不同程度的相关性(王长庭等,
2010 ; 乔文静等,
2018 )。降水量和N添加及其交互作用改变了植物群落多样性(附录II; 朱湾湾等,
2019 ), 因而可能会对土壤酶活性产生重要影响。本研究中, 蔗糖酶活性随着Patrick丰富度指数增加而增加, 即蔗糖酶活性与植物物种数存在正线性关系。这可能是由于降水量增加和N添加下, 随着植物物种数增多, 土壤酶作用的底物多样性增加, 从而增强了蔗糖酶活性, 与对江河源区人工草地的研究结果(王启兰等,
2010 )一致; 脲酶和磷酸酶活性均与Shannon-Wiener多样性指数以及Pielou均匀度指数存在负线性关系。这可能是由于降水量和N添加下, 植物群落多样性和均匀度的改变直接影响到土壤微环境(李长斌等,
2016 ), 并通过对植物根系和微生物活性的影响, 间接影响到脲酶和磷酸酶活性。由于植物群落特征对土壤酶活性的影响存在时间滞后效应(乔文静等,
2018 ), 因此还需通过长期的试验观测深入探讨土壤酶活性与植物群落多样性之间的联系。
3.3 土壤酶活性与微生物生态化学计量特征的关系 除植物群落组成外, 土壤酶活性还受到土壤因子的综合影响(王启兰等,
2010 ; 李静等,
2013 )。本研究中, 与其他因子相比, 微生物生态化学计量特征对3种酶活性的影响较大。蔗糖酶活性与微生物生物量C含量、C:N和C:P呈负相关关系。其可能原因在于降水和N添加在一定程度上提高了土壤N和P的有效性(涂利华等,
2009 ), 限制了C的有效性, 微生物生物量C含量、C:N和C:P呈降低趋势(附录IV), 从而刺激微生物分泌较多的C获取酶以促进有机C矿化(Rappe-George
et al .,
2017 )。脲酶活性与微生物生物量C:N呈正相关关系, 与微生物生物量N含量呈负相关关系。这可能是由于持续增加的降水量加速了土壤N淋溶, 导致土壤N含量降低(附录III), 即N受限性增强, 进而提高土壤脲酶活性以促进有机N矿化(涂利华等,
2009 ; 刘星等,
2015 )。此外, 磷酸酶活性与微生物生物量C:P呈正相关关系, 与微生物生物量P含量呈负相关关系。结合2019年磷酸酶的变化特点(
图1 ), 以上结果表明降水量变化和N添加下微生物较多地受到土壤P有效性的限制。随着微生物对P的需求量增加, 相关水解酶活性相应增强(Steenbergh
et al .,
2011 )。
此外, 对土壤酶活性影响显著的土壤因子还包括pH、全P含量、N:P、C:P、NH
4 + -N以及NO
3 - -N含量。土壤pH是影响土壤酶活性的重要因素(苏鑫等,
2020 ), 能够调控有机大分子物质的生物降解和矿化过程(万忠梅和宋长春,
2009 )。本研究中, 脲酶和磷酸酶活性均与土壤pH负相关, 意味着研究区土壤较高的pH (附录III)抑制了二者活性, 与对森林土壤的研究结果(吴际友等,
2010 )一致。脲酶活性与土壤N:P以及NH
4 + -N含量正相关。这表明虽然N添加提高了土壤N有效性, 但逐渐增加的降水量导致土壤N淋溶作用加强, 植物和微生物对N的竞争增强, 进而导致脲酶分泌量增多(刘星等,
2015 )。另外, 磷酸酶活性与土壤C:P和N:P正相关, 与全P含量负相关, 与其他研究结果(Allison
et al .,
2007 )相似。这可能是当土壤中P供给不足时, 植物会分泌磷酸酶以促进土壤有机P的矿化, 生成可供植物吸收的无机态P (附录III), 导致土壤全P含量降低(Deng
et al .,
2017 )。总之, 土壤pH和养分有效性通过调控养分固定和矿化等过程, 以正、负效应的形式对土壤酶活性进行调节和驱动。
4 结论 综上, 与减少降水量相比, 增加降水量对蔗糖酶、脲酶和磷酸酶活性影响较大, 但其效应与N添加以及年份存在交互作用; 与其他植物种相比, 草木樨状黄耆和糙隐子草生物量与3种酶活性存在较强的线性关系。Patrick丰富度指数与蔗糖酶活性存在正的线性关系, Shannon-Wiener多样性指数、Pielou均匀度指数以及Simpson优势度指数均与3种酶活性存在不同程度的负相关关系; 对土壤酶活性影响较大的因子包括土壤pH、土壤全P含量、微生物生物量C:P、微生物生物量C:N、土壤N:P、微生物生物量C和P含量。因此, 两年降水量及N添加下荒漠草原土壤酶活性无明显的变化规律, 同时降水量和N添加通过改变植物群落组成、微生物生物量C:N:P生态化学计量特征以及土壤P有效性间接影响植被-土壤系统中元素循环, 进而改变酶活性。
本研究为连续两年降水量变化和N添加下的试验观测结果。由于短期的试验结果易受到降水和温度等气候因子年际波动的影响, 因此有必要通过长期的原位试验深入探讨土壤酶活性与植物、微生物之间的联系。此外, 鉴于土壤酶种类的多样化和功能的复杂性以及土壤酶活性对于生态系统C、N、P循环的重要指示意义, 今后还需结合鲜土中多种酶活性的变化规律, 系统分析全球变化对酶活性的影响机制。
附录 附录I 2019年降水量和氮添加对植物生物量的影响
Supplement I Effects of precipitation and nitrogen addition on plant biomass in 2019
https://www.plant-ecology.com/fileup/1005-264X/PDF/cjpe.2020.0264-S1.pdf 附录II 2019年降水量和氮添加对植物群落多样性的影响
Supplement II Effects of precipitation and nitrogen addition on plant community diversity in 2019
https://www.plant-ecology.com/fileup/1005-264X/PDF/cjpe.2020.0264-S2.pdf 附录III 2019年降水量和氮添加对土壤理化性质的影响
Supplement III Effects of precipitation and nitrogen addition on physical and chemical properties of soil in 2019
https://www.plant-ecology.com/fileup/1005-264X/PDF/cjpe.2020.0264-S3.pdf 附录IV 2019年降水量和氮添加对微生物生态化学计量特征的影响
Supplement IV Effects of precipitation and nitrogen addition on microbial ecological stoichiometry in 2019
https://www.plant-ecology.com/fileup/1005-264X/PDF/cjpe.2020.0264-S4.pdf [1] Allison VJ Condron LM Peltzer DA Richardson SJ Turner BL 2007 ). Changes in enzyme activities and soil microbial community composition along carbon and nutrient gradients at the Franz Josef chronosequence, New ZealandSoil Biology & Biochemistry 1770 -1781 . DOI:
10.1016/j.soilbio.2007.02.006 URL [本文引用: 1] [2] Bai CL 2013 ). Effects of addition of nitrogen and water on plant community characteristics of Stipa breviflora desert steppeChinese Journal of Grassland 2 ), 69 -75 . [本文引用: 1] [白春利 , 阿拉塔 , 陈海军 , 单玉梅 , 额尔敦花 , 王明玖 (2013 ). 氮素和水分添加对短花针茅荒漠草原植物群落特征的影响中国草地学报 , 35(2 ), 69 -75 .] [本文引用: 1] [3] Burns RG DeForest JL Marxsen J Sinsabaugh RL Stromberger ME Wallenstein MD Weintraub MN Zoppini A 2013 ). Soil enzymes in a changing environment: current knowledge and future directionsSoil Biology & Biochemistry 216 -234 . DOI:
10.1016/j.soilbio.2012.11.009 URL [本文引用: 1] [4] Deng Q Hui DF Dennis S Reddy KC 2017 ). Responses of terrestrial ecosystem phosphorus cycling to nitrogen addition: a meta-analysisGlobal Ecology and Biogeography 713 -728 . DOI:
10.1111/geb.2017.26.issue-6 URL [本文引用: 1] [5] Dou PP Wang F Ma Y Pang M Mi XC Ma KP Lin DM 2018 ). Response of litter carbon, nitrogen and phosphorus to simulated leachingChinese Science Bulletin 3114 -3123 . [本文引用: 1] [豆鹏鹏 , 王芳 , 马瑜 , 庞梅 , 米湘成 , 马克平 , 林敦梅 (2018 ). 叶凋落物碳、氮和磷元素对模拟淋溶的响应科学通报 , 63, 3114 -3123 .] [本文引用: 1] [6] Duan L Hao JM Xie SD Zhou ZP 2002 ). Estimating critical loads of sulfur and nitrogen for Chinese soils by steady state methodEnvironmental Science 2 ), 7 -12 . [本文引用: 1] [段雷 , 郝吉明 , 谢绍东 , 周中平 (2002 ). 用稳态法确定中国土壤的硫沉降和氮沉降临界负荷环境科学 , 23(2 ), 7 -12 .] [本文引用: 1] [7] Gao JQ Yang XG Dong CY Li KN 2015 ). Precipitation resource changed characteristics in arid and humid regions in Northern China with climate changesTransactions of the Chinese Society of Agricultural Engineering 12 ), 99 -110 . [本文引用: 1] [高继卿 , 杨晓光 , 董朝阳 , 李克南 (2015 ). 气候变化背景下中国北方干湿区降水资源变化特征分析农业工程学报 , 31(12 ), 99 -110 .] [本文引用: 1] [8] Gu FX Huang M Zhang YD Yan HM Li J Guo R Zhong XL 2016 ). Modeling the temporal-spatial patterns of atmospheric nitrogen deposition in China during 1961-2010Acta Ecologica Sinica 3591 -3600 . [本文引用: 1] [顾峰雪 , 黄玫 , 张远东 , 闫慧敏 , 李洁 , 郭瑞 , 钟秀丽 (2016 ). 1961-2010年中国区域氮沉降时空格局模拟研究生态学报 , 36, 3591 -3600 .] [本文引用: 1] [9] Hu XW Wang YR Wu YP 2004 ). Research progress on eco-physiological responses of desert grassland plants to drought conditionsActa Prataculturae Sinica 3 ), 9 -15 . [本文引用: 1] [胡小文 , 王彦荣 , 武艳培 (2004 ). 荒漠草原植物抗旱生理生态学研究进展草业学报 , 13(3 ), 9 -15 .] [本文引用: 1] [10] IPCC Intergovernmental Panel on Climate Change (2013 ). Summary for policymakers/climate change: the physical science basisContribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change [11] Kanakidou M Myriokefalitakis S Daskalakis N Fanourgakis G Nenes A Baker AR Tsigaridis K Mihalopoulos N 2016 ). Past, present and future atmospheric nitrogen depositionJournal of the Atmospheric Sciences 2039 -2047 . DOI:
10.1175/JAS-D-15-0278.1 URL [本文引用: 1] [12] Li CB Peng YF Zhao DZ Ning Y Zhou GY 2016 ). Effects of precipitation change and nitrogen addition on community structure and plant diversity in an alpine steppe on the Qinghai-Tibetan PlateauResearch of Soil and Water Conservation 6 ), 185 -191 . [本文引用: 1] [李长斌 , 彭云峰 , 赵殿智 , 宁祎 , 周国英 (2016 ). 降水变化和氮素添加对青藏高原高寒草原群落结构和物种多样性的影响水土保持研究 , 23(6 ), 185 -191 .] [本文引用: 1] [13] Li J Yu XY Tang M 2013 ). Effects of different plants on soil microbial biomass and enzyme activities in Zhifangou watershed of Loess PlateauActa Botanica Boreali- Occidentalia Sinica 387 -393 . [本文引用: 2] [李静 , 蔚晓燕 , 唐明 (2013 ). 黄土高原纸坊沟流域不同植物对土壤微生物生物量和土壤酶活性的影响西北植物学报 , 33, 387 -393 .] [本文引用: 2] [14] Li YH Zhu HQ Fang LZ Li FD 2020 ). Soil enzyme activity characteristics and impact factors under plant communities of the Ebinur Lake wetlandActa Ecologica Sinica 549 -559 . [本文引用: 1] [李艳红 , 朱海强 , 方丽章 , 李发东 (2020 ). 艾比湖湿地植物群落土壤酶活性特征及影响因素生态学报 , 40, 549 -559 .] [本文引用: 1] [15] Liu HM Zhou GF Li J Wang LL Wang H Yang DL 2018 ). Effects of nitrogen deposition on soil enzyme activities of Stipa baicalensis steppeEcology and Environmental Sciences 1387 -1394 . [本文引用: 1] [刘红梅 , 周广帆 , 李洁 , 王丽丽 , 王慧 , 杨殿林 (2018 ). 氮沉降对贝加尔针茅草原土壤酶活性的影响生态环境学报 , 27, 1387 -1394 .] [本文引用: 1] [16] Liu X Wang JS Zhao XH 2015 ). Effects of simulated nitrogen deposition on the soil enzyme activities in a Pinus tabulaeformis forest at the Taiyue MountainActa Ecologica Sinica 4613 -4624 . [本文引用: 2] [刘星 , 汪金松 , 赵秀海 (2015 ). 模拟氮沉降对太岳山油松林土壤酶活性的影响生态学报 , 35, 4613 -4624 .] [本文引用: 2] [17] Lü XT Kong DL Pan QM Simmons ME Han XG 2012 ). Nitrogen and water availability interact to affect leaf stoichiometry in a semi-arid grasslandOecologia 301 -310 . DOI:
10.1007/s00442-011-2097-7 URL [本文引用: 1] [18] Ma WW Yao T Jin P Wang GJ Zhang YX 2014 ). Characteristics of microorganisms and enzyme activity under two plant communities in desert steppeJournal of Desert Research 176 -183 . [本文引用: 1] [马文文 , 姚拓 , 靳鹏 , 王国基 , 张玉霞 (2014 ). 荒漠草原2种植物群落土壤微生物及土壤酶特征中国沙漠 , 34, 176 -183 .] [本文引用: 1] [19] Qiao WJ Dai YY Zhang W Fu SY Yang GH Han XH Chen ZX Gao DX Feng YZ 2018 ). Relationship between the vegetation community and soil nutrient and enzyme activity during the restoration of abandoned land in the loess hilly regionEnvironmental Science 5687 -5698 . [本文引用: 3] [乔文静 , 戴银月 , 张伟 , 付淑月 , 杨改河 , 韩新辉 , 陈正兴 , 高德新 , 冯永忠 (2018 ). 黄土丘陵区撂荒恢复过程中植物群落组成与土壤养分及酶活性变化的关系环境科学 , 39, 5687 -5698 .] [本文引用: 3] [20] Rappe-George MO Choma M ?apek P B?rjesson G Ka?tovská E ?antr??ková H G?rden?s AI 2017 ). Indications that long-term nitrogen loading limits carbon resources for soil microbesSoil Biology & Biochemistry 310 -321 . DOI:
10.1016/j.soilbio.2017.07.015 URL [本文引用: 1] [21] Schimel J Balser TC Wallenstein M 2007 ). Microbial stress-response physiology and its implications for ecosystem functionEcology 1386 -1394 . PMID:
17601131 [本文引用: 1] Microorganisms have a variety of evolutionary adaptations and physiological acclimation mechanisms that allow them to survive and remain active in the face of environmental stress. Physiological responses to stress have costs at the organismal level that can result in altered ecosystem-level C, energy, and nutrient flows. These large-scale impacts result from direct effects on active microbes' physiology and by controlling the composition of the active microbial community. We first consider some general aspects of how microbes experience environmental stresses and how they respond to them. We then discuss the impacts of two important ecosystem-level stressors, drought and freezing, on microbial physiology and community composition. Even when microbial community response to stress is limited, the physiological costs imposed on soil microbes are large enough that they may cause large shifts in the allocation and fate of C and N. For example, for microbes to synthesize the osmolytes they need to survive a single drought episode they may consume up to 5% of total annual net primary production in grassland ecosystems, while acclimating to freezing conditions switches Arctic tundra soils from immobilizing N during the growing season to mineralizing it during the winter. We suggest that more effectively integrating microbial ecology into ecosystem ecology will require a more complete integration of microbial physiological ecology, population biology, and process ecology.
[22] Steenbergh AK Bodelier PLE Hoogveld HL Slomp CP Laanbroek HJ 2011 ). Phosphatases relieve carbon limitation of microbial activity in Baltic Sea sediments along a redox-gradientLimnology and Oceanography 2018 -2026 . DOI:
10.4319/lo.2011.56.6.2018 URL [本文引用: 1] [23] Su X Lu M Feng CC Guo YL Yue ZH 2020 ). Responses of seasonal dynamic of soil enzyme activity to soil salinity in Songnen saline-alkali grasslandChinese Journal of Grassland 1 ), 127 -134 . [本文引用: 1] [苏鑫 , 卢嫚 , 冯程程 , 郭迎岚 , 岳中辉 (2020 ). 松嫩草地土壤酶活性对土壤盐分季节动态的响应中国草地学报 , 42(1 ), 127 -134 .] [本文引用: 1] [24] Tian DS Wang H Sun J Niu SL 2016 ). Global evidence on nitrogen saturation of terrestrial ecosystem net primary productivityEnvironmental Research Letters 024012 . DOI: 10.1088/1748-9326/11/2/024012 . DOI:
10.1088/1748-9326/11/2/024012 URL [本文引用: 1] [25] Tu LH Hu TX Zhang J Li RH Dai HZ Luo SH Xiang YB Huang LH 2009 ). Soil enzyme activities in a Pleioblastus amurus plantation in Rainy Area of West China under simulated nitrogen depositionChinese Journal of Applied Ecology 2943 -2948 . [本文引用: 2] [涂利华 , 胡庭兴 , 张健 , 李仁洪 , 戴洪忠 , 雒守华 , 向元彬 , 黄立华 (2009 ). 华西雨屏区苦竹林土壤酶活性对模拟氮沉降的响应应用生态学报 , 20, 2943 -2948 .] [本文引用: 2] [26] Wan ZM Song CC 2009 ). Advance on response of soil enzyme activity to ecological environmentChinese Journal of Soil Science 951 -956 . [本文引用: 1] [万忠梅 , 宋长春 (2009 ). 土壤酶活性对生态环境的响应研究进展土壤通报 , 40, 951 -956 .] [本文引用: 1] [27] Wang CT Long RJ Wang GX Liu W Wang QL Zhang L Wu PF 2010 ). Relationship between plant communities, characters, soil physical and chemical properties, and soil microbiology in alpine meadowsActa Prataculturae Sinica 6 ), 25 -34 . [本文引用: 2] [王长庭 , 龙瑞军 , 王根绪 , 刘伟 , 王启兰 , 张莉 , 吴鹏飞 (2010 ). 高寒草甸群落地表植被特征与土壤理化性状、土壤微生物之间的相关性研究草业学报 , 19(6 ), 25 -34 .] [本文引用: 2] [28] Wang J Li G Xiu WM Song XL Zhao JN Yang DL 2014a ). Effects of nitrogen and water on soil enzyme activity and soil microbial biomass in stipa baicalensis steppe, Inner Mongolia of North ChinaJournal of Agricultural Resources and Environment 237 -245 . [本文引用: 2] [王杰 , 李刚 , 修伟明 , 宋晓龙 , 赵建宁 , 杨殿林 (2014a ). 氮素和水分对贝加尔针茅草原土壤酶活性和微生物量碳氮的影响农业资源与环境学报 , 31, 237 -245 .] [本文引用: 2] [29] Wang J Li G Xiu WM Zhao JN Wang H Yang DL 2014b ). Responses of soil microbial functional diversity to nitrogen and water input in Stipa baicalensis steppe, Inner Mongolia, Northern ChinaActa Prataculturae Sinica 343 -350 . [本文引用: 1] [王杰 , 李刚 , 修伟明 , 赵建宁 , 王慧 , 杨殿林 (2014b ). 贝加尔针茅草原土壤微生物功能多样性对氮素和水分添加的响应草业学报 , 23, 343 -350 .] [本文引用: 1] [30] Wang QL Wang CT Liu W Cao GM Long RJ 2010 ). Changes in plant communities and soil enzyme activities of artificial grasslands in headwater areas of the Yangtze and Yellow riversChinese Journal of Applied and Environmental Biology 662 -666 . [本文引用: 2] [王启兰 , 王长庭 , 刘伟 , 曹广民 , 龙瑞军 (2010 ). 江河源区人工草地植物群落和土壤酶活性变化应用与环境生物学报 , 16, 662 -666 .] [本文引用: 2] [31] Wang XP Xiao X Tang TW Li YX Xiao J 2018 ). Seasonal changes of the input of root exudates and its driving characteristics of rhizosphere microbe in a Cercidiphyllum japonicum Sieb. plantationBulletin of Botanical Research 47 -55 . [本文引用: 1] [王小平 , 肖肖 , 唐天文 , 黎云祥 , 肖娟 (2018 ). 连香树人工林根系分泌物输入季节性变化及其驱动的根际微生物特性研究植物研究 , 38, 47 -55 .] [本文引用: 1] [32] Wu JY Ye DB Wang XJ 2010 ). Soil enzyme activity and its correlation with soil physical and chemical properties in suburban forests in Changsha CityJournal of Northeast Forestry University 3 ), 97 -99 . [本文引用: 1] [吴际友 , 叶道碧 , 王旭军 (2010 ). 长沙市城郊森林土壤酶活性及其与土壤理化性质的相关性东北林业大学学报 , 38(3 ), 97 -99 .] [本文引用: 1] [33] Xu H He MZ Sun Y 2018 ). Response of soil enzyme activities to precipitation regulation in arid desert areasJournal of Lanzhou University Natural Sciences ), 54, 790 -797 . [本文引用: 2] [许华 , 何明珠 , 孙岩 (2018 ). 干旱荒漠区土壤酶活性对降水调控的响应兰州大学学报(自然科学版) , 54, 790 -797 .] [本文引用: 2] [34] Xu ZW Li MH Zimmermann NE Li SP Li H Ren HY Sun H Han XG Jiang Y Jiang L 2018 ). Plant functional diversity modulates global environmental change effects on grassland productivityJournal of Ecology 1941 -1951 . DOI:
10.1111/jec.2018.106.issue-5 URL [35] Yan ZQ Qi YC Li SJ Dong YS Peng Q He YL Li ZL 2017a ). Soil microorganisms and enzyme activity of grassland ecosystem affected by changes in precipitation pattern and increase in nitrogen deposition—A reviewMicrobiology China 1481 -1490 . [本文引用: 2] [闫钟清 , 齐玉春 , 李素俭 , 董云社 , 彭琴 , 贺云龙 , 李兆林 (2017a ). 降水和氮沉降增加对草地土壤微生物与酶活性的影响研究进展微生物学通报 , 44, 1481 -1490 .] [本文引用: 2] [36] Yan ZQ Qi YC Peng Q Dong YS Guo SF He YL Wang LQ Li ZL 2017b ). Effects of increased precipitation and nitrogen deposition on soil enzyme activitiesActa Ecologica Sinica 3019 -3027 . [本文引用: 1] [闫钟清 , 齐玉春 , 彭琴 , 董云社 , 郭树芳 , 贺云龙 , 王丽芹 , 李兆林 (2017b ). 降水和氮沉降增加对草地土壤酶活性的影响生态学报 , 37, 3019 -3027 .] [本文引用: 1] [37] Yang R Zhao DP 2018 ). Characteristics of seasonal variation of precipitation in northwest vulnerable ecotone under the background of climate warming in the past 54 yearsResearch of Soil and Water Conservation 6 ), 85 -93 . [本文引用: 1] [杨蓉 , 赵多平 (2018 ). 气候变暖背景下西北脆弱带近54年降水季节变化特征水土保持研究 , 25(6 ), 85 -93 .] [本文引用: 1] [38] Yang WQ Wang KY 2002 ). Advances on soil enzymologyChinese Journal of Applied and Environmental Biology 564 -570 . [本文引用: 1] [杨万勤 , 王开运 (2002 ). 土壤酶研究动态与展望应用与环境生物学报 , 8, 564 -570 .] [本文引用: 1] [39] Yu G Jia Y He N Zhu J Chen Z Wang Q Piao S Liu X He H Guo X Wen Z Li P Ding G Goulding K 2019 ). Stabilization of atmospheric nitrogen deposition in China over the past decadeNature Geoscience 424 -429 . DOI:
10.1038/s41561-019-0352-4 URL [本文引用: 1] [40] Yu HL He NP Wang QF Zhu JX Gao Y Zhang YH Jia YL Yu GR 2017 ). Development of atmospheric acid deposition in China from the 1990s to the 2010sEnvironmental Pollution 182 -190 . DOI:
10.1016/j.envpol.2017.08.014 URL [本文引用: 1] [41] Zhang JF Xu YQ 2016 ). Responses of plant biomass and net primary production to nitrogen fertilization and increased precipitation in re-grassed croplands in Duolun County of Inner Mongolia, ChinaChinese Journal of Eco-Agriculture 192 -200 . [张金凤 , 徐雨晴 (2016 ). 水氮添加对内蒙古多伦县退耕还草地生物量、生产力及其分配的影响中国生态农业学报 , 24, 192 -200 .] [42] Zhang MM Fan SH Guan FY Yan YJ Yin ZX Huang LY 2020 ). Study on soil microbial biomass and enzyme activities in mixed forest of bamboo and broad-leaved treesSoils 97 -105 . [本文引用: 1] [张美曼 , 范少辉 , 官凤英 , 晏颖杰 , 尹子旭 , 黄兰鹰 (2020 ). 竹阔混交林土壤微生物生物量及酶活性特征研究土壤 , 52, 97 -105 .] [本文引用: 1] [43] Zhang RY Xu DH Chen LY Wang G 2014 ). Plant N status in the alpine grassland of the Qinghai-Tibet Plateau: base on the N:P stoichiometryEnvironmental Science 1131 -1137 . [本文引用: 1] [张仁懿 , 徐当会 , 陈凌云 , 王刚 (2014 ). 基于N:P化学计量特征的高寒草甸植物养分状况研究环境科学 , 35, 1131 -1137 .] [本文引用: 1] [44] Zhong Y Yan WM Wang RW Shangguan ZP 2017 ). Differential responses of litter decomposition to nutrient addition and soil water availability with long-term vegetation recoveryBiology and Fertility of Soils 939 -949 . DOI:
10.1007/s00374-017-1242-9 URL [本文引用: 1] [45] Zhu WW Wang P Fan J Niu YB Yu HL Huang JY 2019 ). Effects of precipitation and N addition on soil C:N:P ecological stoichiometry and plant community composition in a desert steppe of Ningxia, northwestern ChinaActa Prataculturae Sinica 9 ), 33 -44 . [本文引用: 6] [朱湾湾 , 王攀 , 樊瑾 , 牛玉斌 , 余海龙 , 黄菊莹 (2019 ). 降水量及N添加对宁夏荒漠草原土壤C:N:P生态化学计量特征和植被群落组成的影响草业学报 , 28(9 ), 33 -44 .] [本文引用: 6] [46] Zhu YZ Li YY Han JG Yao HY 2019 ). Effects of changes in water status on soil microbes and their response mechanism: a reviewChinese Journal of Applied Ecology 4323 -4332 . [本文引用: 1] [朱义族 , 李雅颖 , 韩继刚 , 姚槐应 (2019 ). 水分条件变化对土壤微生物的影响及其响应机制研究进展应用生态学报 , 30, 4323 -4332 .] [本文引用: 1] Changes in enzyme activities and soil microbial community composition along carbon and nutrient gradients at the Franz Josef chronosequence, New Zealand
1
2007
... 此外, 对土壤酶活性影响显著的土壤因子还包括pH、全P含量、N:P、C:P、NH
4 + -N以及NO
3 - -N含量.土壤pH是影响土壤酶活性的重要因素(苏鑫等,
2020 ), 能够调控有机大分子物质的生物降解和矿化过程(万忠梅和宋长春,
2009 ).本研究中, 脲酶和磷酸酶活性均与土壤pH负相关, 意味着研究区土壤较高的pH (附录III)抑制了二者活性, 与对森林土壤的研究结果(吴际友等,
2010 )一致.脲酶活性与土壤N:P以及NH
4 + -N含量正相关.这表明虽然N添加提高了土壤N有效性, 但逐渐增加的降水量导致土壤N淋溶作用加强, 植物和微生物对N的竞争增强, 进而导致脲酶分泌量增多(刘星等,
2015 ).另外, 磷酸酶活性与土壤C:P和N:P正相关, 与全P含量负相关, 与其他研究结果(Allison
et al .,
2007 )相似.这可能是当土壤中P供给不足时, 植物会分泌磷酸酶以促进土壤有机P的矿化, 生成可供植物吸收的无机态P (附录III), 导致土壤全P含量降低(Deng
et al .,
2017 ).总之, 土壤pH和养分有效性通过调控养分固定和矿化等过程, 以正、负效应的形式对土壤酶活性进行调节和驱动. ...
氮素和水分添加对短花针茅荒漠草原植物群落特征的影响
1
2013
... 本研究中, 减少降水量对蔗糖酶、脲酶和磷酸酶活性的影响较小, 增加降水量不同程度地影响3种酶的活性.这可能是由于研究区长期处于低水平的土壤水分环境下, 植物和微生物对干旱具有高的适应性(胡小文等,
2004 ; 朱义族等,
2019 ), 因而减少降水量对其生长繁殖以及酶的分泌影响较小.相比之下, 在缺水环境下, 增加降水量提高了土壤含水量(附录III), 从而加速凋落物淋溶、促进表层凋落物和腐殖质中可溶性物质进入土壤(豆鹏鹏等,
2018 ), 使微生物可利用底物增加, 微生物活性相应增强, 进而刺激微生物向环境中释放酶(Schimel
et al .,
2007 ), 即微生物分泌的酶随之增加.此外, 本研究发现增加降水量对土壤酶活性的影响与N添加存在交互作用, 与其他研究结果(王杰等,
2014a )相似.这表明水分有利于N肥效的释放, 二者的交互作用促进了植物生长(白春利等,
2013 )、凋落物积累以及微生物活动(王杰等,
2014b ), 进而刺激土壤酶的分泌、导致土壤酶活性增强(闫钟清等,
2017b ). ...
氮素和水分添加对短花针茅荒漠草原植物群落特征的影响
1
2013
... 本研究中, 减少降水量对蔗糖酶、脲酶和磷酸酶活性的影响较小, 增加降水量不同程度地影响3种酶的活性.这可能是由于研究区长期处于低水平的土壤水分环境下, 植物和微生物对干旱具有高的适应性(胡小文等,
2004 ; 朱义族等,
2019 ), 因而减少降水量对其生长繁殖以及酶的分泌影响较小.相比之下, 在缺水环境下, 增加降水量提高了土壤含水量(附录III), 从而加速凋落物淋溶、促进表层凋落物和腐殖质中可溶性物质进入土壤(豆鹏鹏等,
2018 ), 使微生物可利用底物增加, 微生物活性相应增强, 进而刺激微生物向环境中释放酶(Schimel
et al .,
2007 ), 即微生物分泌的酶随之增加.此外, 本研究发现增加降水量对土壤酶活性的影响与N添加存在交互作用, 与其他研究结果(王杰等,
2014a )相似.这表明水分有利于N肥效的释放, 二者的交互作用促进了植物生长(白春利等,
2013 )、凋落物积累以及微生物活动(王杰等,
2014b ), 进而刺激土壤酶的分泌、导致土壤酶活性增强(闫钟清等,
2017b ). ...
Soil enzymes in a changing environment: current knowledge and future directions
1
2013
... 土壤酶主要由植物根系和微生物分泌产生, 是土壤中元素生物地球化学循环的催化剂(马文文等,
2014 ).一方面, 土壤酶活性能够反映土壤元素(碳(C)、N、磷(P)等)转化和物质循环的速率(Burns
et al .,
2013 ), 并通过对土壤养分状况的影响改变植物养分策略.同时, 土壤酶能够实现植物营养元素和有机质的循环转化, 对土壤微环境的元素平衡和转化具有重要影响(张美曼等,
2020 ).另一方面, 土壤酶活性可表征微生物的养分需求、反映微生物在环境波动时维持养分平衡的应对策略(闫钟清等,
2017a ).土壤酶活性主要受土壤养分可利用性(李艳红等,
2020 )、植被类型(李静等,
2013 )和微生物活性(闫钟清等,
2017a )等因素的影响.降水和N添加通过调控土壤水分和养分有效性, 改变植物资源利用策略, 从而直接影响到植被结构(张金凤和许雨晴, 2016)和微生物活性(王杰等,
2014a ), 进而改变土壤酶活性(许华等,
2018 ).因此, 揭示降水量变化和N添加下土壤酶活性与植物和微生物的联系, 可为评价全球变化背景下植被-土壤系统功能提供新思路. ...
Responses of terrestrial ecosystem phosphorus cycling to nitrogen addition: a meta-analysis
1
2017
... 此外, 对土壤酶活性影响显著的土壤因子还包括pH、全P含量、N:P、C:P、NH
4 + -N以及NO
3 - -N含量.土壤pH是影响土壤酶活性的重要因素(苏鑫等,
2020 ), 能够调控有机大分子物质的生物降解和矿化过程(万忠梅和宋长春,
2009 ).本研究中, 脲酶和磷酸酶活性均与土壤pH负相关, 意味着研究区土壤较高的pH (附录III)抑制了二者活性, 与对森林土壤的研究结果(吴际友等,
2010 )一致.脲酶活性与土壤N:P以及NH
4 + -N含量正相关.这表明虽然N添加提高了土壤N有效性, 但逐渐增加的降水量导致土壤N淋溶作用加强, 植物和微生物对N的竞争增强, 进而导致脲酶分泌量增多(刘星等,
2015 ).另外, 磷酸酶活性与土壤C:P和N:P正相关, 与全P含量负相关, 与其他研究结果(Allison
et al .,
2007 )相似.这可能是当土壤中P供给不足时, 植物会分泌磷酸酶以促进土壤有机P的矿化, 生成可供植物吸收的无机态P (附录III), 导致土壤全P含量降低(Deng
et al .,
2017 ).总之, 土壤pH和养分有效性通过调控养分固定和矿化等过程, 以正、负效应的形式对土壤酶活性进行调节和驱动. ...
叶凋落物碳、氮和磷元素对模拟淋溶的响应
1
2018
... 本研究中, 减少降水量对蔗糖酶、脲酶和磷酸酶活性的影响较小, 增加降水量不同程度地影响3种酶的活性.这可能是由于研究区长期处于低水平的土壤水分环境下, 植物和微生物对干旱具有高的适应性(胡小文等,
2004 ; 朱义族等,
2019 ), 因而减少降水量对其生长繁殖以及酶的分泌影响较小.相比之下, 在缺水环境下, 增加降水量提高了土壤含水量(附录III), 从而加速凋落物淋溶、促进表层凋落物和腐殖质中可溶性物质进入土壤(豆鹏鹏等,
2018 ), 使微生物可利用底物增加, 微生物活性相应增强, 进而刺激微生物向环境中释放酶(Schimel
et al .,
2007 ), 即微生物分泌的酶随之增加.此外, 本研究发现增加降水量对土壤酶活性的影响与N添加存在交互作用, 与其他研究结果(王杰等,
2014a )相似.这表明水分有利于N肥效的释放, 二者的交互作用促进了植物生长(白春利等,
2013 )、凋落物积累以及微生物活动(王杰等,
2014b ), 进而刺激土壤酶的分泌、导致土壤酶活性增强(闫钟清等,
2017b ). ...
叶凋落物碳、氮和磷元素对模拟淋溶的响应
1
2018
... 本研究中, 减少降水量对蔗糖酶、脲酶和磷酸酶活性的影响较小, 增加降水量不同程度地影响3种酶的活性.这可能是由于研究区长期处于低水平的土壤水分环境下, 植物和微生物对干旱具有高的适应性(胡小文等,
2004 ; 朱义族等,
2019 ), 因而减少降水量对其生长繁殖以及酶的分泌影响较小.相比之下, 在缺水环境下, 增加降水量提高了土壤含水量(附录III), 从而加速凋落物淋溶、促进表层凋落物和腐殖质中可溶性物质进入土壤(豆鹏鹏等,
2018 ), 使微生物可利用底物增加, 微生物活性相应增强, 进而刺激微生物向环境中释放酶(Schimel
et al .,
2007 ), 即微生物分泌的酶随之增加.此外, 本研究发现增加降水量对土壤酶活性的影响与N添加存在交互作用, 与其他研究结果(王杰等,
2014a )相似.这表明水分有利于N肥效的释放, 二者的交互作用促进了植物生长(白春利等,
2013 )、凋落物积累以及微生物活动(王杰等,
2014b ), 进而刺激土壤酶的分泌、导致土壤酶活性增强(闫钟清等,
2017b ). ...
用稳态法确定中国土壤的硫沉降和氮沉降临界负荷
1
2002
... 荒漠草原是我国西北干旱半干旱区主要的草原生态系统类型, 其植被结构简单、生态系统脆弱.宁夏荒漠草原主要分布于毛乌素沙地西南缘, 是区域重要的生态屏障.该生态系统降水量少、蒸发量大、N沉降临界负荷低, 降水和N是其植被生长和微生物活动的主要限制因子, 因此对降水格局改变和N沉降增加较为敏感(段雷等,
2002 ; 杨蓉和赵多平,
2018 ).那么, 降水量和N添加及其交互作用如何影响荒漠草原土壤酶活性, 降水量改变和N添加下土壤酶活性与植物群落组成以及微生物活动存在怎样的联系等问题, 都值得我们进行深入探讨.为此, 本项目组于2017年在宁夏荒漠草原设立了降水量改变及N添加的裂区试验, 研究了2018-2019年生长季旺盛期土壤酶活性的变化特点, 分析了其与植物群落组成以及微生物生物量C:N:P生态化学计量特征(其值高低与微生物活动的方向密切相关)的关系, 为充分认识降水量变化和N添加对土壤酶活性的影响机制提供基础数据, 并为深入理解全球变化背景下西北脆弱生态区的元素生物地球化学循环提供理论依据. ...
用稳态法确定中国土壤的硫沉降和氮沉降临界负荷
1
2002
... 荒漠草原是我国西北干旱半干旱区主要的草原生态系统类型, 其植被结构简单、生态系统脆弱.宁夏荒漠草原主要分布于毛乌素沙地西南缘, 是区域重要的生态屏障.该生态系统降水量少、蒸发量大、N沉降临界负荷低, 降水和N是其植被生长和微生物活动的主要限制因子, 因此对降水格局改变和N沉降增加较为敏感(段雷等,
2002 ; 杨蓉和赵多平,
2018 ).那么, 降水量和N添加及其交互作用如何影响荒漠草原土壤酶活性, 降水量改变和N添加下土壤酶活性与植物群落组成以及微生物活动存在怎样的联系等问题, 都值得我们进行深入探讨.为此, 本项目组于2017年在宁夏荒漠草原设立了降水量改变及N添加的裂区试验, 研究了2018-2019年生长季旺盛期土壤酶活性的变化特点, 分析了其与植物群落组成以及微生物生物量C:N:P生态化学计量特征(其值高低与微生物活动的方向密切相关)的关系, 为充分认识降水量变化和N添加对土壤酶活性的影响机制提供基础数据, 并为深入理解全球变化背景下西北脆弱生态区的元素生物地球化学循环提供理论依据. ...
气候变化背景下中国北方干湿区降水资源变化特征分析
1
2015
... 降水格局改变和大气氮(N)沉降增加是全球变化的两个重要表现(IPCC, 2013).全球变暖背景下, 降水量在全球范围内呈增加趋势, 同时表现出时空分配不均和极端事件频发等特征(IPCC, 2013).据报道, 我国西北地区的西部生态区降水量呈增加趋势, 东部生态区降水量有所降低(高继卿等,
2015 ).此外, 人类活动导致全球大气N沉降速率持续增加(Kanakidou
et al .,
2016 ).就中国而言, 虽然N沉降速率在一些南方省市有所下降, 但在西北部分区域呈增加趋势(Yu
et al .,
2017 ,
2019 ).西北干旱半干旱区生态系统稳定性差, 对环境变化敏感, 是我国典型的生态脆弱区之一.作为该区的主要限制因子, 降水和N调控着土壤资源有效性, 影响着植物生长和微生物活动等关键生态过程, 与植被-土壤系统的稳定性密切相关.目前, 国内已有较多研究探讨了降水和N添加对该脆弱生态区的影响, 尚缺乏二者交互作用的相关报道.鉴于降水在介导N沉降效应中发挥着重要作用(Lü
et al .,
2012 ), 因此有必要针对西北干旱半干旱区典型生态系统开展二者交互效应的研究. ...
气候变化背景下中国北方干湿区降水资源变化特征分析
1
2015
... 降水格局改变和大气氮(N)沉降增加是全球变化的两个重要表现(IPCC, 2013).全球变暖背景下, 降水量在全球范围内呈增加趋势, 同时表现出时空分配不均和极端事件频发等特征(IPCC, 2013).据报道, 我国西北地区的西部生态区降水量呈增加趋势, 东部生态区降水量有所降低(高继卿等,
2015 ).此外, 人类活动导致全球大气N沉降速率持续增加(Kanakidou
et al .,
2016 ).就中国而言, 虽然N沉降速率在一些南方省市有所下降, 但在西北部分区域呈增加趋势(Yu
et al .,
2017 ,
2019 ).西北干旱半干旱区生态系统稳定性差, 对环境变化敏感, 是我国典型的生态脆弱区之一.作为该区的主要限制因子, 降水和N调控着土壤资源有效性, 影响着植物生长和微生物活动等关键生态过程, 与植被-土壤系统的稳定性密切相关.目前, 国内已有较多研究探讨了降水和N添加对该脆弱生态区的影响, 尚缺乏二者交互作用的相关报道.鉴于降水在介导N沉降效应中发挥着重要作用(Lü
et al .,
2012 ), 因此有必要针对西北干旱半干旱区典型生态系统开展二者交互效应的研究. ...
1961-2010年中国区域氮沉降时空格局模拟研究
1
2016
... 于2017年9月, 在围栏草地内选择30 m × 50 m的区域作为降水量和N添加处理的试验样地.降水增减量依据近几十年来我国西北地区的西部生态区降水量增加而东部生态区降水量减少的趋势进行设置, 同时参考了国内同类研究方法(Xu
et al .,
2018 ).氮肥施用量依据区域N沉降水平(顾峰雪等,
2016 )和N饱和阈值(Tian
et al .,
2016 )进行设置.试验设计采用随机区组的裂区设计, 主区为降水量, 副区为N添加.降水量处理包括减少50% (W1)、减少30% (W2)、对照(自然降水量, W3)、增加30% (W4)和增加50% (W5).N添加处理包括0 g·m
-2 ·a
-1 (N0)和5 g·m
-2 ·a
-1 (N5).试验包含10个处理, 每个处理3次重复, 共计30个小区(
表2 ).每个主区面积为8 m × 8 m, 每个副区面积为4 m × 8 m.各主区之间设置1.5 m的缓冲带.各主区的两个副区间垂直下埋1 m宽的塑料布, 上埋20 cm宽的彩钢板, 用来防止试验干扰. ...
1961-2010年中国区域氮沉降时空格局模拟研究
1
2016
... 于2017年9月, 在围栏草地内选择30 m × 50 m的区域作为降水量和N添加处理的试验样地.降水增减量依据近几十年来我国西北地区的西部生态区降水量增加而东部生态区降水量减少的趋势进行设置, 同时参考了国内同类研究方法(Xu
et al .,
2018 ).氮肥施用量依据区域N沉降水平(顾峰雪等,
2016 )和N饱和阈值(Tian
et al .,
2016 )进行设置.试验设计采用随机区组的裂区设计, 主区为降水量, 副区为N添加.降水量处理包括减少50% (W1)、减少30% (W2)、对照(自然降水量, W3)、增加30% (W4)和增加50% (W5).N添加处理包括0 g·m
-2 ·a
-1 (N0)和5 g·m
-2 ·a
-1 (N5).试验包含10个处理, 每个处理3次重复, 共计30个小区(
表2 ).每个主区面积为8 m × 8 m, 每个副区面积为4 m × 8 m.各主区之间设置1.5 m的缓冲带.各主区的两个副区间垂直下埋1 m宽的塑料布, 上埋20 cm宽的彩钢板, 用来防止试验干扰. ...
荒漠草原植物抗旱生理生态学研究进展
1
2004
... 本研究中, 减少降水量对蔗糖酶、脲酶和磷酸酶活性的影响较小, 增加降水量不同程度地影响3种酶的活性.这可能是由于研究区长期处于低水平的土壤水分环境下, 植物和微生物对干旱具有高的适应性(胡小文等,
2004 ; 朱义族等,
2019 ), 因而减少降水量对其生长繁殖以及酶的分泌影响较小.相比之下, 在缺水环境下, 增加降水量提高了土壤含水量(附录III), 从而加速凋落物淋溶、促进表层凋落物和腐殖质中可溶性物质进入土壤(豆鹏鹏等,
2018 ), 使微生物可利用底物增加, 微生物活性相应增强, 进而刺激微生物向环境中释放酶(Schimel
et al .,
2007 ), 即微生物分泌的酶随之增加.此外, 本研究发现增加降水量对土壤酶活性的影响与N添加存在交互作用, 与其他研究结果(王杰等,
2014a )相似.这表明水分有利于N肥效的释放, 二者的交互作用促进了植物生长(白春利等,
2013 )、凋落物积累以及微生物活动(王杰等,
2014b ), 进而刺激土壤酶的分泌、导致土壤酶活性增强(闫钟清等,
2017b ). ...
荒漠草原植物抗旱生理生态学研究进展
1
2004
... 本研究中, 减少降水量对蔗糖酶、脲酶和磷酸酶活性的影响较小, 增加降水量不同程度地影响3种酶的活性.这可能是由于研究区长期处于低水平的土壤水分环境下, 植物和微生物对干旱具有高的适应性(胡小文等,
2004 ; 朱义族等,
2019 ), 因而减少降水量对其生长繁殖以及酶的分泌影响较小.相比之下, 在缺水环境下, 增加降水量提高了土壤含水量(附录III), 从而加速凋落物淋溶、促进表层凋落物和腐殖质中可溶性物质进入土壤(豆鹏鹏等,
2018 ), 使微生物可利用底物增加, 微生物活性相应增强, 进而刺激微生物向环境中释放酶(Schimel
et al .,
2007 ), 即微生物分泌的酶随之增加.此外, 本研究发现增加降水量对土壤酶活性的影响与N添加存在交互作用, 与其他研究结果(王杰等,
2014a )相似.这表明水分有利于N肥效的释放, 二者的交互作用促进了植物生长(白春利等,
2013 )、凋落物积累以及微生物活动(王杰等,
2014b ), 进而刺激土壤酶的分泌、导致土壤酶活性增强(闫钟清等,
2017b ). ...
Summary for policymakers/climate change: the physical science basis
2013
Past, present and future atmospheric nitrogen deposition
1
2016
... 降水格局改变和大气氮(N)沉降增加是全球变化的两个重要表现(IPCC, 2013).全球变暖背景下, 降水量在全球范围内呈增加趋势, 同时表现出时空分配不均和极端事件频发等特征(IPCC, 2013).据报道, 我国西北地区的西部生态区降水量呈增加趋势, 东部生态区降水量有所降低(高继卿等,
2015 ).此外, 人类活动导致全球大气N沉降速率持续增加(Kanakidou
et al .,
2016 ).就中国而言, 虽然N沉降速率在一些南方省市有所下降, 但在西北部分区域呈增加趋势(Yu
et al .,
2017 ,
2019 ).西北干旱半干旱区生态系统稳定性差, 对环境变化敏感, 是我国典型的生态脆弱区之一.作为该区的主要限制因子, 降水和N调控着土壤资源有效性, 影响着植物生长和微生物活动等关键生态过程, 与植被-土壤系统的稳定性密切相关.目前, 国内已有较多研究探讨了降水和N添加对该脆弱生态区的影响, 尚缺乏二者交互作用的相关报道.鉴于降水在介导N沉降效应中发挥着重要作用(Lü
et al .,
2012 ), 因此有必要针对西北干旱半干旱区典型生态系统开展二者交互效应的研究. ...
降水变化和氮素添加对青藏高原高寒草原群落结构和物种多样性的影响
1
2016
... 研究表明, 土壤酶活性与植物群落多样性亦存在不同程度的相关性(王长庭等,
2010 ; 乔文静等,
2018 ).降水量和N添加及其交互作用改变了植物群落多样性(附录II; 朱湾湾等,
2019 ), 因而可能会对土壤酶活性产生重要影响.本研究中, 蔗糖酶活性随着Patrick丰富度指数增加而增加, 即蔗糖酶活性与植物物种数存在正线性关系.这可能是由于降水量增加和N添加下, 随着植物物种数增多, 土壤酶作用的底物多样性增加, 从而增强了蔗糖酶活性, 与对江河源区人工草地的研究结果(王启兰等,
2010 )一致; 脲酶和磷酸酶活性均与Shannon-Wiener多样性指数以及Pielou均匀度指数存在负线性关系.这可能是由于降水量和N添加下, 植物群落多样性和均匀度的改变直接影响到土壤微环境(李长斌等,
2016 ), 并通过对植物根系和微生物活性的影响, 间接影响到脲酶和磷酸酶活性.由于植物群落特征对土壤酶活性的影响存在时间滞后效应(乔文静等,
2018 ), 因此还需通过长期的试验观测深入探讨土壤酶活性与植物群落多样性之间的联系. ...
降水变化和氮素添加对青藏高原高寒草原群落结构和物种多样性的影响
1
2016
... 研究表明, 土壤酶活性与植物群落多样性亦存在不同程度的相关性(王长庭等,
2010 ; 乔文静等,
2018 ).降水量和N添加及其交互作用改变了植物群落多样性(附录II; 朱湾湾等,
2019 ), 因而可能会对土壤酶活性产生重要影响.本研究中, 蔗糖酶活性随着Patrick丰富度指数增加而增加, 即蔗糖酶活性与植物物种数存在正线性关系.这可能是由于降水量增加和N添加下, 随着植物物种数增多, 土壤酶作用的底物多样性增加, 从而增强了蔗糖酶活性, 与对江河源区人工草地的研究结果(王启兰等,
2010 )一致; 脲酶和磷酸酶活性均与Shannon-Wiener多样性指数以及Pielou均匀度指数存在负线性关系.这可能是由于降水量和N添加下, 植物群落多样性和均匀度的改变直接影响到土壤微环境(李长斌等,
2016 ), 并通过对植物根系和微生物活性的影响, 间接影响到脲酶和磷酸酶活性.由于植物群落特征对土壤酶活性的影响存在时间滞后效应(乔文静等,
2018 ), 因此还需通过长期的试验观测深入探讨土壤酶活性与植物群落多样性之间的联系. ...
黄土高原纸坊沟流域不同植物对土壤微生物生物量和土壤酶活性的影响
2
2013
... 土壤酶主要由植物根系和微生物分泌产生, 是土壤中元素生物地球化学循环的催化剂(马文文等,
2014 ).一方面, 土壤酶活性能够反映土壤元素(碳(C)、N、磷(P)等)转化和物质循环的速率(Burns
et al .,
2013 ), 并通过对土壤养分状况的影响改变植物养分策略.同时, 土壤酶能够实现植物营养元素和有机质的循环转化, 对土壤微环境的元素平衡和转化具有重要影响(张美曼等,
2020 ).另一方面, 土壤酶活性可表征微生物的养分需求、反映微生物在环境波动时维持养分平衡的应对策略(闫钟清等,
2017a ).土壤酶活性主要受土壤养分可利用性(李艳红等,
2020 )、植被类型(李静等,
2013 )和微生物活性(闫钟清等,
2017a )等因素的影响.降水和N添加通过调控土壤水分和养分有效性, 改变植物资源利用策略, 从而直接影响到植被结构(张金凤和许雨晴, 2016)和微生物活性(王杰等,
2014a ), 进而改变土壤酶活性(许华等,
2018 ).因此, 揭示降水量变化和N添加下土壤酶活性与植物和微生物的联系, 可为评价全球变化背景下植被-土壤系统功能提供新思路. ...
... 除植物群落组成外, 土壤酶活性还受到土壤因子的综合影响(王启兰等,
2010 ; 李静等,
2013 ).本研究中, 与其他因子相比, 微生物生态化学计量特征对3种酶活性的影响较大.蔗糖酶活性与微生物生物量C含量、C:N和C:P呈负相关关系.其可能原因在于降水和N添加在一定程度上提高了土壤N和P的有效性(涂利华等,
2009 ), 限制了C的有效性, 微生物生物量C含量、C:N和C:P呈降低趋势(附录IV), 从而刺激微生物分泌较多的C获取酶以促进有机C矿化(Rappe-George
et al .,
2017 ).脲酶活性与微生物生物量C:N呈正相关关系, 与微生物生物量N含量呈负相关关系.这可能是由于持续增加的降水量加速了土壤N淋溶, 导致土壤N含量降低(附录III), 即N受限性增强, 进而提高土壤脲酶活性以促进有机N矿化(涂利华等,
2009 ; 刘星等,
2015 ).此外, 磷酸酶活性与微生物生物量C:P呈正相关关系, 与微生物生物量P含量呈负相关关系.结合2019年磷酸酶的变化特点(
图1 ), 以上结果表明降水量变化和N添加下微生物较多地受到土壤P有效性的限制.随着微生物对P的需求量增加, 相关水解酶活性相应增强(Steenbergh
et al .,
2011 ). ...
黄土高原纸坊沟流域不同植物对土壤微生物生物量和土壤酶活性的影响
2
2013
... 土壤酶主要由植物根系和微生物分泌产生, 是土壤中元素生物地球化学循环的催化剂(马文文等,
2014 ).一方面, 土壤酶活性能够反映土壤元素(碳(C)、N、磷(P)等)转化和物质循环的速率(Burns
et al .,
2013 ), 并通过对土壤养分状况的影响改变植物养分策略.同时, 土壤酶能够实现植物营养元素和有机质的循环转化, 对土壤微环境的元素平衡和转化具有重要影响(张美曼等,
2020 ).另一方面, 土壤酶活性可表征微生物的养分需求、反映微生物在环境波动时维持养分平衡的应对策略(闫钟清等,
2017a ).土壤酶活性主要受土壤养分可利用性(李艳红等,
2020 )、植被类型(李静等,
2013 )和微生物活性(闫钟清等,
2017a )等因素的影响.降水和N添加通过调控土壤水分和养分有效性, 改变植物资源利用策略, 从而直接影响到植被结构(张金凤和许雨晴, 2016)和微生物活性(王杰等,
2014a ), 进而改变土壤酶活性(许华等,
2018 ).因此, 揭示降水量变化和N添加下土壤酶活性与植物和微生物的联系, 可为评价全球变化背景下植被-土壤系统功能提供新思路. ...
... 除植物群落组成外, 土壤酶活性还受到土壤因子的综合影响(王启兰等,
2010 ; 李静等,
2013 ).本研究中, 与其他因子相比, 微生物生态化学计量特征对3种酶活性的影响较大.蔗糖酶活性与微生物生物量C含量、C:N和C:P呈负相关关系.其可能原因在于降水和N添加在一定程度上提高了土壤N和P的有效性(涂利华等,
2009 ), 限制了C的有效性, 微生物生物量C含量、C:N和C:P呈降低趋势(附录IV), 从而刺激微生物分泌较多的C获取酶以促进有机C矿化(Rappe-George
et al .,
2017 ).脲酶活性与微生物生物量C:N呈正相关关系, 与微生物生物量N含量呈负相关关系.这可能是由于持续增加的降水量加速了土壤N淋溶, 导致土壤N含量降低(附录III), 即N受限性增强, 进而提高土壤脲酶活性以促进有机N矿化(涂利华等,
2009 ; 刘星等,
2015 ).此外, 磷酸酶活性与微生物生物量C:P呈正相关关系, 与微生物生物量P含量呈负相关关系.结合2019年磷酸酶的变化特点(
图1 ), 以上结果表明降水量变化和N添加下微生物较多地受到土壤P有效性的限制.随着微生物对P的需求量增加, 相关水解酶活性相应增强(Steenbergh
et al .,
2011 ). ...
艾比湖湿地植物群落土壤酶活性特征及影响因素
1
2020
... 土壤酶主要由植物根系和微生物分泌产生, 是土壤中元素生物地球化学循环的催化剂(马文文等,
2014 ).一方面, 土壤酶活性能够反映土壤元素(碳(C)、N、磷(P)等)转化和物质循环的速率(Burns
et al .,
2013 ), 并通过对土壤养分状况的影响改变植物养分策略.同时, 土壤酶能够实现植物营养元素和有机质的循环转化, 对土壤微环境的元素平衡和转化具有重要影响(张美曼等,
2020 ).另一方面, 土壤酶活性可表征微生物的养分需求、反映微生物在环境波动时维持养分平衡的应对策略(闫钟清等,
2017a ).土壤酶活性主要受土壤养分可利用性(李艳红等,
2020 )、植被类型(李静等,
2013 )和微生物活性(闫钟清等,
2017a )等因素的影响.降水和N添加通过调控土壤水分和养分有效性, 改变植物资源利用策略, 从而直接影响到植被结构(张金凤和许雨晴, 2016)和微生物活性(王杰等,
2014a ), 进而改变土壤酶活性(许华等,
2018 ).因此, 揭示降水量变化和N添加下土壤酶活性与植物和微生物的联系, 可为评价全球变化背景下植被-土壤系统功能提供新思路. ...
艾比湖湿地植物群落土壤酶活性特征及影响因素
1
2020
... 土壤酶主要由植物根系和微生物分泌产生, 是土壤中元素生物地球化学循环的催化剂(马文文等,
2014 ).一方面, 土壤酶活性能够反映土壤元素(碳(C)、N、磷(P)等)转化和物质循环的速率(Burns
et al .,
2013 ), 并通过对土壤养分状况的影响改变植物养分策略.同时, 土壤酶能够实现植物营养元素和有机质的循环转化, 对土壤微环境的元素平衡和转化具有重要影响(张美曼等,
2020 ).另一方面, 土壤酶活性可表征微生物的养分需求、反映微生物在环境波动时维持养分平衡的应对策略(闫钟清等,
2017a ).土壤酶活性主要受土壤养分可利用性(李艳红等,
2020 )、植被类型(李静等,
2013 )和微生物活性(闫钟清等,
2017a )等因素的影响.降水和N添加通过调控土壤水分和养分有效性, 改变植物资源利用策略, 从而直接影响到植被结构(张金凤和许雨晴, 2016)和微生物活性(王杰等,
2014a ), 进而改变土壤酶活性(许华等,
2018 ).因此, 揭示降水量变化和N添加下土壤酶活性与植物和微生物的联系, 可为评价全球变化背景下植被-土壤系统功能提供新思路. ...
氮沉降对贝加尔针茅草原土壤酶活性的影响
1
2018
... 各指标的测定中, 蔗糖酶活性采用3,5二硝基水杨酸比色法, 脲酶活性采用苯酚钠-次氯酸钠比色法, 磷酸酶活性采用对硝基苯磷酸盐法(刘红梅等,
2018 ), 有机C含量采用重铬酸钾容量法-外加热法, 全N含量采用凯氏定氮法, 全P含量采用钼锑抗比色法, pH采用酸度计法, 电导率采用便携式电导率仪, NH
4 + -N和NO
3 - -N含量采用连续流动分析仪(Auto Analyzer 3, SEAL Analytical GmbH, Hanau, Germany), 速效P含量采用0.5 mol·L
-1 NaHCO
3 法, 微生物生物量C含量采用氯仿熏蒸-K
2 SO
4 浸提-碳分析仪器法, 微生物生物量N含量采用氯仿熏蒸-K
2 SO
4 提取-流动注射氮分析仪器法, 微生物生物量P含量采用氯仿熏蒸-K
2 SO
4 提取-Pi测定-外加Pi矫正法. ...
氮沉降对贝加尔针茅草原土壤酶活性的影响
1
2018
... 各指标的测定中, 蔗糖酶活性采用3,5二硝基水杨酸比色法, 脲酶活性采用苯酚钠-次氯酸钠比色法, 磷酸酶活性采用对硝基苯磷酸盐法(刘红梅等,
2018 ), 有机C含量采用重铬酸钾容量法-外加热法, 全N含量采用凯氏定氮法, 全P含量采用钼锑抗比色法, pH采用酸度计法, 电导率采用便携式电导率仪, NH
4 + -N和NO
3 - -N含量采用连续流动分析仪(Auto Analyzer 3, SEAL Analytical GmbH, Hanau, Germany), 速效P含量采用0.5 mol·L
-1 NaHCO
3 法, 微生物生物量C含量采用氯仿熏蒸-K
2 SO
4 浸提-碳分析仪器法, 微生物生物量N含量采用氯仿熏蒸-K
2 SO
4 提取-流动注射氮分析仪器法, 微生物生物量P含量采用氯仿熏蒸-K
2 SO
4 提取-Pi测定-外加Pi矫正法. ...
模拟氮沉降对太岳山油松林土壤酶活性的影响
2
2015
... 除植物群落组成外, 土壤酶活性还受到土壤因子的综合影响(王启兰等,
2010 ; 李静等,
2013 ).本研究中, 与其他因子相比, 微生物生态化学计量特征对3种酶活性的影响较大.蔗糖酶活性与微生物生物量C含量、C:N和C:P呈负相关关系.其可能原因在于降水和N添加在一定程度上提高了土壤N和P的有效性(涂利华等,
2009 ), 限制了C的有效性, 微生物生物量C含量、C:N和C:P呈降低趋势(附录IV), 从而刺激微生物分泌较多的C获取酶以促进有机C矿化(Rappe-George
et al .,
2017 ).脲酶活性与微生物生物量C:N呈正相关关系, 与微生物生物量N含量呈负相关关系.这可能是由于持续增加的降水量加速了土壤N淋溶, 导致土壤N含量降低(附录III), 即N受限性增强, 进而提高土壤脲酶活性以促进有机N矿化(涂利华等,
2009 ; 刘星等,
2015 ).此外, 磷酸酶活性与微生物生物量C:P呈正相关关系, 与微生物生物量P含量呈负相关关系.结合2019年磷酸酶的变化特点(
图1 ), 以上结果表明降水量变化和N添加下微生物较多地受到土壤P有效性的限制.随着微生物对P的需求量增加, 相关水解酶活性相应增强(Steenbergh
et al .,
2011 ). ...
... 此外, 对土壤酶活性影响显著的土壤因子还包括pH、全P含量、N:P、C:P、NH
4 + -N以及NO
3 - -N含量.土壤pH是影响土壤酶活性的重要因素(苏鑫等,
2020 ), 能够调控有机大分子物质的生物降解和矿化过程(万忠梅和宋长春,
2009 ).本研究中, 脲酶和磷酸酶活性均与土壤pH负相关, 意味着研究区土壤较高的pH (附录III)抑制了二者活性, 与对森林土壤的研究结果(吴际友等,
2010 )一致.脲酶活性与土壤N:P以及NH
4 + -N含量正相关.这表明虽然N添加提高了土壤N有效性, 但逐渐增加的降水量导致土壤N淋溶作用加强, 植物和微生物对N的竞争增强, 进而导致脲酶分泌量增多(刘星等,
2015 ).另外, 磷酸酶活性与土壤C:P和N:P正相关, 与全P含量负相关, 与其他研究结果(Allison
et al .,
2007 )相似.这可能是当土壤中P供给不足时, 植物会分泌磷酸酶以促进土壤有机P的矿化, 生成可供植物吸收的无机态P (附录III), 导致土壤全P含量降低(Deng
et al .,
2017 ).总之, 土壤pH和养分有效性通过调控养分固定和矿化等过程, 以正、负效应的形式对土壤酶活性进行调节和驱动. ...
模拟氮沉降对太岳山油松林土壤酶活性的影响
2
2015
... 除植物群落组成外, 土壤酶活性还受到土壤因子的综合影响(王启兰等,
2010 ; 李静等,
2013 ).本研究中, 与其他因子相比, 微生物生态化学计量特征对3种酶活性的影响较大.蔗糖酶活性与微生物生物量C含量、C:N和C:P呈负相关关系.其可能原因在于降水和N添加在一定程度上提高了土壤N和P的有效性(涂利华等,
2009 ), 限制了C的有效性, 微生物生物量C含量、C:N和C:P呈降低趋势(附录IV), 从而刺激微生物分泌较多的C获取酶以促进有机C矿化(Rappe-George
et al .,
2017 ).脲酶活性与微生物生物量C:N呈正相关关系, 与微生物生物量N含量呈负相关关系.这可能是由于持续增加的降水量加速了土壤N淋溶, 导致土壤N含量降低(附录III), 即N受限性增强, 进而提高土壤脲酶活性以促进有机N矿化(涂利华等,
2009 ; 刘星等,
2015 ).此外, 磷酸酶活性与微生物生物量C:P呈正相关关系, 与微生物生物量P含量呈负相关关系.结合2019年磷酸酶的变化特点(
图1 ), 以上结果表明降水量变化和N添加下微生物较多地受到土壤P有效性的限制.随着微生物对P的需求量增加, 相关水解酶活性相应增强(Steenbergh
et al .,
2011 ). ...
... 此外, 对土壤酶活性影响显著的土壤因子还包括pH、全P含量、N:P、C:P、NH
4 + -N以及NO
3 - -N含量.土壤pH是影响土壤酶活性的重要因素(苏鑫等,
2020 ), 能够调控有机大分子物质的生物降解和矿化过程(万忠梅和宋长春,
2009 ).本研究中, 脲酶和磷酸酶活性均与土壤pH负相关, 意味着研究区土壤较高的pH (附录III)抑制了二者活性, 与对森林土壤的研究结果(吴际友等,
2010 )一致.脲酶活性与土壤N:P以及NH
4 + -N含量正相关.这表明虽然N添加提高了土壤N有效性, 但逐渐增加的降水量导致土壤N淋溶作用加强, 植物和微生物对N的竞争增强, 进而导致脲酶分泌量增多(刘星等,
2015 ).另外, 磷酸酶活性与土壤C:P和N:P正相关, 与全P含量负相关, 与其他研究结果(Allison
et al .,
2007 )相似.这可能是当土壤中P供给不足时, 植物会分泌磷酸酶以促进土壤有机P的矿化, 生成可供植物吸收的无机态P (附录III), 导致土壤全P含量降低(Deng
et al .,
2017 ).总之, 土壤pH和养分有效性通过调控养分固定和矿化等过程, 以正、负效应的形式对土壤酶活性进行调节和驱动. ...
Nitrogen and water availability interact to affect leaf stoichiometry in a semi-arid grassland
1
2012
... 降水格局改变和大气氮(N)沉降增加是全球变化的两个重要表现(IPCC, 2013).全球变暖背景下, 降水量在全球范围内呈增加趋势, 同时表现出时空分配不均和极端事件频发等特征(IPCC, 2013).据报道, 我国西北地区的西部生态区降水量呈增加趋势, 东部生态区降水量有所降低(高继卿等,
2015 ).此外, 人类活动导致全球大气N沉降速率持续增加(Kanakidou
et al .,
2016 ).就中国而言, 虽然N沉降速率在一些南方省市有所下降, 但在西北部分区域呈增加趋势(Yu
et al .,
2017 ,
2019 ).西北干旱半干旱区生态系统稳定性差, 对环境变化敏感, 是我国典型的生态脆弱区之一.作为该区的主要限制因子, 降水和N调控着土壤资源有效性, 影响着植物生长和微生物活动等关键生态过程, 与植被-土壤系统的稳定性密切相关.目前, 国内已有较多研究探讨了降水和N添加对该脆弱生态区的影响, 尚缺乏二者交互作用的相关报道.鉴于降水在介导N沉降效应中发挥着重要作用(Lü
et al .,
2012 ), 因此有必要针对西北干旱半干旱区典型生态系统开展二者交互效应的研究. ...
荒漠草原2种植物群落土壤微生物及土壤酶特征
1
2014
... 土壤酶主要由植物根系和微生物分泌产生, 是土壤中元素生物地球化学循环的催化剂(马文文等,
2014 ).一方面, 土壤酶活性能够反映土壤元素(碳(C)、N、磷(P)等)转化和物质循环的速率(Burns
et al .,
2013 ), 并通过对土壤养分状况的影响改变植物养分策略.同时, 土壤酶能够实现植物营养元素和有机质的循环转化, 对土壤微环境的元素平衡和转化具有重要影响(张美曼等,
2020 ).另一方面, 土壤酶活性可表征微生物的养分需求、反映微生物在环境波动时维持养分平衡的应对策略(闫钟清等,
2017a ).土壤酶活性主要受土壤养分可利用性(李艳红等,
2020 )、植被类型(李静等,
2013 )和微生物活性(闫钟清等,
2017a )等因素的影响.降水和N添加通过调控土壤水分和养分有效性, 改变植物资源利用策略, 从而直接影响到植被结构(张金凤和许雨晴, 2016)和微生物活性(王杰等,
2014a ), 进而改变土壤酶活性(许华等,
2018 ).因此, 揭示降水量变化和N添加下土壤酶活性与植物和微生物的联系, 可为评价全球变化背景下植被-土壤系统功能提供新思路. ...
荒漠草原2种植物群落土壤微生物及土壤酶特征
1
2014
... 土壤酶主要由植物根系和微生物分泌产生, 是土壤中元素生物地球化学循环的催化剂(马文文等,
2014 ).一方面, 土壤酶活性能够反映土壤元素(碳(C)、N、磷(P)等)转化和物质循环的速率(Burns
et al .,
2013 ), 并通过对土壤养分状况的影响改变植物养分策略.同时, 土壤酶能够实现植物营养元素和有机质的循环转化, 对土壤微环境的元素平衡和转化具有重要影响(张美曼等,
2020 ).另一方面, 土壤酶活性可表征微生物的养分需求、反映微生物在环境波动时维持养分平衡的应对策略(闫钟清等,
2017a ).土壤酶活性主要受土壤养分可利用性(李艳红等,
2020 )、植被类型(李静等,
2013 )和微生物活性(闫钟清等,
2017a )等因素的影响.降水和N添加通过调控土壤水分和养分有效性, 改变植物资源利用策略, 从而直接影响到植被结构(张金凤和许雨晴, 2016)和微生物活性(王杰等,
2014a ), 进而改变土壤酶活性(许华等,
2018 ).因此, 揭示降水量变化和N添加下土壤酶活性与植物和微生物的联系, 可为评价全球变化背景下植被-土壤系统功能提供新思路. ...
黄土丘陵区撂荒恢复过程中植物群落组成与土壤养分及酶活性变化的关系
3
2018
... 本研究中, 降水量和N添加下脲酶和磷酸酶活性与草木樨状黄耆(豆科植物)生物量呈负线性关系, 而蔗糖酶、脲酶和磷酸酶活性均与糙隐子草(禾本科植物)生物量呈正线性关系, 与针对高寒草甸的研究结果(王长庭等,
2010 )存在差异.首先, 土壤酶活性与植物生物量之间的关系因植物种不同而异.这是由于不同植物根系的发达程度以及分泌物存在差异, 进而影响土壤酶活性(王小平等,
2018 ).豆科植物与根瘤菌的共生作用可使其在相对干旱的条件下快速生长(张仁懿等,
2014 ).这不仅消耗了土壤养分, 而且抑制了微生物活性以及酶活性.相比之下, 降水量增加和N添加显著地刺激了禾本科植物的生长(附录I; 朱湾湾等,
2019 ), 其自身的营养物质归还土壤后可为土壤酶的合成提供丰富的底物(乔文静等,
2018 ).其次, 降水量增加及N添加下植物生物量增加(附录I; 朱湾湾等,
2019 ), 提高了凋落物C和养分向土壤中的归还量(Zhong
et al .,
2017 ), 从而促进了蔗糖酶、脲酶和磷酸酶的产生. ...
... 研究表明, 土壤酶活性与植物群落多样性亦存在不同程度的相关性(王长庭等,
2010 ; 乔文静等,
2018 ).降水量和N添加及其交互作用改变了植物群落多样性(附录II; 朱湾湾等,
2019 ), 因而可能会对土壤酶活性产生重要影响.本研究中, 蔗糖酶活性随着Patrick丰富度指数增加而增加, 即蔗糖酶活性与植物物种数存在正线性关系.这可能是由于降水量增加和N添加下, 随着植物物种数增多, 土壤酶作用的底物多样性增加, 从而增强了蔗糖酶活性, 与对江河源区人工草地的研究结果(王启兰等,
2010 )一致; 脲酶和磷酸酶活性均与Shannon-Wiener多样性指数以及Pielou均匀度指数存在负线性关系.这可能是由于降水量和N添加下, 植物群落多样性和均匀度的改变直接影响到土壤微环境(李长斌等,
2016 ), 并通过对植物根系和微生物活性的影响, 间接影响到脲酶和磷酸酶活性.由于植物群落特征对土壤酶活性的影响存在时间滞后效应(乔文静等,
2018 ), 因此还需通过长期的试验观测深入探讨土壤酶活性与植物群落多样性之间的联系. ...
... ), 并通过对植物根系和微生物活性的影响, 间接影响到脲酶和磷酸酶活性.由于植物群落特征对土壤酶活性的影响存在时间滞后效应(乔文静等,
2018 ), 因此还需通过长期的试验观测深入探讨土壤酶活性与植物群落多样性之间的联系. ...
黄土丘陵区撂荒恢复过程中植物群落组成与土壤养分及酶活性变化的关系
3
2018
... 本研究中, 降水量和N添加下脲酶和磷酸酶活性与草木樨状黄耆(豆科植物)生物量呈负线性关系, 而蔗糖酶、脲酶和磷酸酶活性均与糙隐子草(禾本科植物)生物量呈正线性关系, 与针对高寒草甸的研究结果(王长庭等,
2010 )存在差异.首先, 土壤酶活性与植物生物量之间的关系因植物种不同而异.这是由于不同植物根系的发达程度以及分泌物存在差异, 进而影响土壤酶活性(王小平等,
2018 ).豆科植物与根瘤菌的共生作用可使其在相对干旱的条件下快速生长(张仁懿等,
2014 ).这不仅消耗了土壤养分, 而且抑制了微生物活性以及酶活性.相比之下, 降水量增加和N添加显著地刺激了禾本科植物的生长(附录I; 朱湾湾等,
2019 ), 其自身的营养物质归还土壤后可为土壤酶的合成提供丰富的底物(乔文静等,
2018 ).其次, 降水量增加及N添加下植物生物量增加(附录I; 朱湾湾等,
2019 ), 提高了凋落物C和养分向土壤中的归还量(Zhong
et al .,
2017 ), 从而促进了蔗糖酶、脲酶和磷酸酶的产生. ...
... 研究表明, 土壤酶活性与植物群落多样性亦存在不同程度的相关性(王长庭等,
2010 ; 乔文静等,
2018 ).降水量和N添加及其交互作用改变了植物群落多样性(附录II; 朱湾湾等,
2019 ), 因而可能会对土壤酶活性产生重要影响.本研究中, 蔗糖酶活性随着Patrick丰富度指数增加而增加, 即蔗糖酶活性与植物物种数存在正线性关系.这可能是由于降水量增加和N添加下, 随着植物物种数增多, 土壤酶作用的底物多样性增加, 从而增强了蔗糖酶活性, 与对江河源区人工草地的研究结果(王启兰等,
2010 )一致; 脲酶和磷酸酶活性均与Shannon-Wiener多样性指数以及Pielou均匀度指数存在负线性关系.这可能是由于降水量和N添加下, 植物群落多样性和均匀度的改变直接影响到土壤微环境(李长斌等,
2016 ), 并通过对植物根系和微生物活性的影响, 间接影响到脲酶和磷酸酶活性.由于植物群落特征对土壤酶活性的影响存在时间滞后效应(乔文静等,
2018 ), 因此还需通过长期的试验观测深入探讨土壤酶活性与植物群落多样性之间的联系. ...
... ), 并通过对植物根系和微生物活性的影响, 间接影响到脲酶和磷酸酶活性.由于植物群落特征对土壤酶活性的影响存在时间滞后效应(乔文静等,
2018 ), 因此还需通过长期的试验观测深入探讨土壤酶活性与植物群落多样性之间的联系. ...
Indications that long-term nitrogen loading limits carbon resources for soil microbes
1
2017
... 除植物群落组成外, 土壤酶活性还受到土壤因子的综合影响(王启兰等,
2010 ; 李静等,
2013 ).本研究中, 与其他因子相比, 微生物生态化学计量特征对3种酶活性的影响较大.蔗糖酶活性与微生物生物量C含量、C:N和C:P呈负相关关系.其可能原因在于降水和N添加在一定程度上提高了土壤N和P的有效性(涂利华等,
2009 ), 限制了C的有效性, 微生物生物量C含量、C:N和C:P呈降低趋势(附录IV), 从而刺激微生物分泌较多的C获取酶以促进有机C矿化(Rappe-George
et al .,
2017 ).脲酶活性与微生物生物量C:N呈正相关关系, 与微生物生物量N含量呈负相关关系.这可能是由于持续增加的降水量加速了土壤N淋溶, 导致土壤N含量降低(附录III), 即N受限性增强, 进而提高土壤脲酶活性以促进有机N矿化(涂利华等,
2009 ; 刘星等,
2015 ).此外, 磷酸酶活性与微生物生物量C:P呈正相关关系, 与微生物生物量P含量呈负相关关系.结合2019年磷酸酶的变化特点(
图1 ), 以上结果表明降水量变化和N添加下微生物较多地受到土壤P有效性的限制.随着微生物对P的需求量增加, 相关水解酶活性相应增强(Steenbergh
et al .,
2011 ). ...
Microbial stress-response physiology and its implications for ecosystem function
1
2007
... 本研究中, 减少降水量对蔗糖酶、脲酶和磷酸酶活性的影响较小, 增加降水量不同程度地影响3种酶的活性.这可能是由于研究区长期处于低水平的土壤水分环境下, 植物和微生物对干旱具有高的适应性(胡小文等,
2004 ; 朱义族等,
2019 ), 因而减少降水量对其生长繁殖以及酶的分泌影响较小.相比之下, 在缺水环境下, 增加降水量提高了土壤含水量(附录III), 从而加速凋落物淋溶、促进表层凋落物和腐殖质中可溶性物质进入土壤(豆鹏鹏等,
2018 ), 使微生物可利用底物增加, 微生物活性相应增强, 进而刺激微生物向环境中释放酶(Schimel
et al .,
2007 ), 即微生物分泌的酶随之增加.此外, 本研究发现增加降水量对土壤酶活性的影响与N添加存在交互作用, 与其他研究结果(王杰等,
2014a )相似.这表明水分有利于N肥效的释放, 二者的交互作用促进了植物生长(白春利等,
2013 )、凋落物积累以及微生物活动(王杰等,
2014b ), 进而刺激土壤酶的分泌、导致土壤酶活性增强(闫钟清等,
2017b ). ...
Phosphatases relieve carbon limitation of microbial activity in Baltic Sea sediments along a redox-gradient
1
2011
... 除植物群落组成外, 土壤酶活性还受到土壤因子的综合影响(王启兰等,
2010 ; 李静等,
2013 ).本研究中, 与其他因子相比, 微生物生态化学计量特征对3种酶活性的影响较大.蔗糖酶活性与微生物生物量C含量、C:N和C:P呈负相关关系.其可能原因在于降水和N添加在一定程度上提高了土壤N和P的有效性(涂利华等,
2009 ), 限制了C的有效性, 微生物生物量C含量、C:N和C:P呈降低趋势(附录IV), 从而刺激微生物分泌较多的C获取酶以促进有机C矿化(Rappe-George
et al .,
2017 ).脲酶活性与微生物生物量C:N呈正相关关系, 与微生物生物量N含量呈负相关关系.这可能是由于持续增加的降水量加速了土壤N淋溶, 导致土壤N含量降低(附录III), 即N受限性增强, 进而提高土壤脲酶活性以促进有机N矿化(涂利华等,
2009 ; 刘星等,
2015 ).此外, 磷酸酶活性与微生物生物量C:P呈正相关关系, 与微生物生物量P含量呈负相关关系.结合2019年磷酸酶的变化特点(
图1 ), 以上结果表明降水量变化和N添加下微生物较多地受到土壤P有效性的限制.随着微生物对P的需求量增加, 相关水解酶活性相应增强(Steenbergh
et al .,
2011 ). ...
松嫩草地土壤酶活性对土壤盐分季节动态的响应
1
2020
... 此外, 对土壤酶活性影响显著的土壤因子还包括pH、全P含量、N:P、C:P、NH
4 + -N以及NO
3 - -N含量.土壤pH是影响土壤酶活性的重要因素(苏鑫等,
2020 ), 能够调控有机大分子物质的生物降解和矿化过程(万忠梅和宋长春,
2009 ).本研究中, 脲酶和磷酸酶活性均与土壤pH负相关, 意味着研究区土壤较高的pH (附录III)抑制了二者活性, 与对森林土壤的研究结果(吴际友等,
2010 )一致.脲酶活性与土壤N:P以及NH
4 + -N含量正相关.这表明虽然N添加提高了土壤N有效性, 但逐渐增加的降水量导致土壤N淋溶作用加强, 植物和微生物对N的竞争增强, 进而导致脲酶分泌量增多(刘星等,
2015 ).另外, 磷酸酶活性与土壤C:P和N:P正相关, 与全P含量负相关, 与其他研究结果(Allison
et al .,
2007 )相似.这可能是当土壤中P供给不足时, 植物会分泌磷酸酶以促进土壤有机P的矿化, 生成可供植物吸收的无机态P (附录III), 导致土壤全P含量降低(Deng
et al .,
2017 ).总之, 土壤pH和养分有效性通过调控养分固定和矿化等过程, 以正、负效应的形式对土壤酶活性进行调节和驱动. ...
松嫩草地土壤酶活性对土壤盐分季节动态的响应
1
2020
... 此外, 对土壤酶活性影响显著的土壤因子还包括pH、全P含量、N:P、C:P、NH
4 + -N以及NO
3 - -N含量.土壤pH是影响土壤酶活性的重要因素(苏鑫等,
2020 ), 能够调控有机大分子物质的生物降解和矿化过程(万忠梅和宋长春,
2009 ).本研究中, 脲酶和磷酸酶活性均与土壤pH负相关, 意味着研究区土壤较高的pH (附录III)抑制了二者活性, 与对森林土壤的研究结果(吴际友等,
2010 )一致.脲酶活性与土壤N:P以及NH
4 + -N含量正相关.这表明虽然N添加提高了土壤N有效性, 但逐渐增加的降水量导致土壤N淋溶作用加强, 植物和微生物对N的竞争增强, 进而导致脲酶分泌量增多(刘星等,
2015 ).另外, 磷酸酶活性与土壤C:P和N:P正相关, 与全P含量负相关, 与其他研究结果(Allison
et al .,
2007 )相似.这可能是当土壤中P供给不足时, 植物会分泌磷酸酶以促进土壤有机P的矿化, 生成可供植物吸收的无机态P (附录III), 导致土壤全P含量降低(Deng
et al .,
2017 ).总之, 土壤pH和养分有效性通过调控养分固定和矿化等过程, 以正、负效应的形式对土壤酶活性进行调节和驱动. ...
Global evidence on nitrogen saturation of terrestrial ecosystem net primary productivity
1
2016
... 于2017年9月, 在围栏草地内选择30 m × 50 m的区域作为降水量和N添加处理的试验样地.降水增减量依据近几十年来我国西北地区的西部生态区降水量增加而东部生态区降水量减少的趋势进行设置, 同时参考了国内同类研究方法(Xu
et al .,
2018 ).氮肥施用量依据区域N沉降水平(顾峰雪等,
2016 )和N饱和阈值(Tian
et al .,
2016 )进行设置.试验设计采用随机区组的裂区设计, 主区为降水量, 副区为N添加.降水量处理包括减少50% (W1)、减少30% (W2)、对照(自然降水量, W3)、增加30% (W4)和增加50% (W5).N添加处理包括0 g·m
-2 ·a
-1 (N0)和5 g·m
-2 ·a
-1 (N5).试验包含10个处理, 每个处理3次重复, 共计30个小区(
表2 ).每个主区面积为8 m × 8 m, 每个副区面积为4 m × 8 m.各主区之间设置1.5 m的缓冲带.各主区的两个副区间垂直下埋1 m宽的塑料布, 上埋20 cm宽的彩钢板, 用来防止试验干扰. ...
华西雨屏区苦竹林土壤酶活性对模拟氮沉降的响应
2
2009
... 除植物群落组成外, 土壤酶活性还受到土壤因子的综合影响(王启兰等,
2010 ; 李静等,
2013 ).本研究中, 与其他因子相比, 微生物生态化学计量特征对3种酶活性的影响较大.蔗糖酶活性与微生物生物量C含量、C:N和C:P呈负相关关系.其可能原因在于降水和N添加在一定程度上提高了土壤N和P的有效性(涂利华等,
2009 ), 限制了C的有效性, 微生物生物量C含量、C:N和C:P呈降低趋势(附录IV), 从而刺激微生物分泌较多的C获取酶以促进有机C矿化(Rappe-George
et al .,
2017 ).脲酶活性与微生物生物量C:N呈正相关关系, 与微生物生物量N含量呈负相关关系.这可能是由于持续增加的降水量加速了土壤N淋溶, 导致土壤N含量降低(附录III), 即N受限性增强, 进而提高土壤脲酶活性以促进有机N矿化(涂利华等,
2009 ; 刘星等,
2015 ).此外, 磷酸酶活性与微生物生物量C:P呈正相关关系, 与微生物生物量P含量呈负相关关系.结合2019年磷酸酶的变化特点(
图1 ), 以上结果表明降水量变化和N添加下微生物较多地受到土壤P有效性的限制.随着微生物对P的需求量增加, 相关水解酶活性相应增强(Steenbergh
et al .,
2011 ). ...
... ).脲酶活性与微生物生物量C:N呈正相关关系, 与微生物生物量N含量呈负相关关系.这可能是由于持续增加的降水量加速了土壤N淋溶, 导致土壤N含量降低(附录III), 即N受限性增强, 进而提高土壤脲酶活性以促进有机N矿化(涂利华等,
2009 ; 刘星等,
2015 ).此外, 磷酸酶活性与微生物生物量C:P呈正相关关系, 与微生物生物量P含量呈负相关关系.结合2019年磷酸酶的变化特点(
图1 ), 以上结果表明降水量变化和N添加下微生物较多地受到土壤P有效性的限制.随着微生物对P的需求量增加, 相关水解酶活性相应增强(Steenbergh
et al .,
2011 ). ...
华西雨屏区苦竹林土壤酶活性对模拟氮沉降的响应
2
2009
... 除植物群落组成外, 土壤酶活性还受到土壤因子的综合影响(王启兰等,
2010 ; 李静等,
2013 ).本研究中, 与其他因子相比, 微生物生态化学计量特征对3种酶活性的影响较大.蔗糖酶活性与微生物生物量C含量、C:N和C:P呈负相关关系.其可能原因在于降水和N添加在一定程度上提高了土壤N和P的有效性(涂利华等,
2009 ), 限制了C的有效性, 微生物生物量C含量、C:N和C:P呈降低趋势(附录IV), 从而刺激微生物分泌较多的C获取酶以促进有机C矿化(Rappe-George
et al .,
2017 ).脲酶活性与微生物生物量C:N呈正相关关系, 与微生物生物量N含量呈负相关关系.这可能是由于持续增加的降水量加速了土壤N淋溶, 导致土壤N含量降低(附录III), 即N受限性增强, 进而提高土壤脲酶活性以促进有机N矿化(涂利华等,
2009 ; 刘星等,
2015 ).此外, 磷酸酶活性与微生物生物量C:P呈正相关关系, 与微生物生物量P含量呈负相关关系.结合2019年磷酸酶的变化特点(
图1 ), 以上结果表明降水量变化和N添加下微生物较多地受到土壤P有效性的限制.随着微生物对P的需求量增加, 相关水解酶活性相应增强(Steenbergh
et al .,
2011 ). ...
... ).脲酶活性与微生物生物量C:N呈正相关关系, 与微生物生物量N含量呈负相关关系.这可能是由于持续增加的降水量加速了土壤N淋溶, 导致土壤N含量降低(附录III), 即N受限性增强, 进而提高土壤脲酶活性以促进有机N矿化(涂利华等,
2009 ; 刘星等,
2015 ).此外, 磷酸酶活性与微生物生物量C:P呈正相关关系, 与微生物生物量P含量呈负相关关系.结合2019年磷酸酶的变化特点(
图1 ), 以上结果表明降水量变化和N添加下微生物较多地受到土壤P有效性的限制.随着微生物对P的需求量增加, 相关水解酶活性相应增强(Steenbergh
et al .,
2011 ). ...
土壤酶活性对生态环境的响应研究进展
1
2009
... 此外, 对土壤酶活性影响显著的土壤因子还包括pH、全P含量、N:P、C:P、NH
4 + -N以及NO
3 - -N含量.土壤pH是影响土壤酶活性的重要因素(苏鑫等,
2020 ), 能够调控有机大分子物质的生物降解和矿化过程(万忠梅和宋长春,
2009 ).本研究中, 脲酶和磷酸酶活性均与土壤pH负相关, 意味着研究区土壤较高的pH (附录III)抑制了二者活性, 与对森林土壤的研究结果(吴际友等,
2010 )一致.脲酶活性与土壤N:P以及NH
4 + -N含量正相关.这表明虽然N添加提高了土壤N有效性, 但逐渐增加的降水量导致土壤N淋溶作用加强, 植物和微生物对N的竞争增强, 进而导致脲酶分泌量增多(刘星等,
2015 ).另外, 磷酸酶活性与土壤C:P和N:P正相关, 与全P含量负相关, 与其他研究结果(Allison
et al .,
2007 )相似.这可能是当土壤中P供给不足时, 植物会分泌磷酸酶以促进土壤有机P的矿化, 生成可供植物吸收的无机态P (附录III), 导致土壤全P含量降低(Deng
et al .,
2017 ).总之, 土壤pH和养分有效性通过调控养分固定和矿化等过程, 以正、负效应的形式对土壤酶活性进行调节和驱动. ...
土壤酶活性对生态环境的响应研究进展
1
2009
... 此外, 对土壤酶活性影响显著的土壤因子还包括pH、全P含量、N:P、C:P、NH
4 + -N以及NO
3 - -N含量.土壤pH是影响土壤酶活性的重要因素(苏鑫等,
2020 ), 能够调控有机大分子物质的生物降解和矿化过程(万忠梅和宋长春,
2009 ).本研究中, 脲酶和磷酸酶活性均与土壤pH负相关, 意味着研究区土壤较高的pH (附录III)抑制了二者活性, 与对森林土壤的研究结果(吴际友等,
2010 )一致.脲酶活性与土壤N:P以及NH
4 + -N含量正相关.这表明虽然N添加提高了土壤N有效性, 但逐渐增加的降水量导致土壤N淋溶作用加强, 植物和微生物对N的竞争增强, 进而导致脲酶分泌量增多(刘星等,
2015 ).另外, 磷酸酶活性与土壤C:P和N:P正相关, 与全P含量负相关, 与其他研究结果(Allison
et al .,
2007 )相似.这可能是当土壤中P供给不足时, 植物会分泌磷酸酶以促进土壤有机P的矿化, 生成可供植物吸收的无机态P (附录III), 导致土壤全P含量降低(Deng
et al .,
2017 ).总之, 土壤pH和养分有效性通过调控养分固定和矿化等过程, 以正、负效应的形式对土壤酶活性进行调节和驱动. ...
高寒草甸群落地表植被特征与土壤理化性状、土壤微生物之间的相关性研究
2
2010
... 本研究中, 降水量和N添加下脲酶和磷酸酶活性与草木樨状黄耆(豆科植物)生物量呈负线性关系, 而蔗糖酶、脲酶和磷酸酶活性均与糙隐子草(禾本科植物)生物量呈正线性关系, 与针对高寒草甸的研究结果(王长庭等,
2010 )存在差异.首先, 土壤酶活性与植物生物量之间的关系因植物种不同而异.这是由于不同植物根系的发达程度以及分泌物存在差异, 进而影响土壤酶活性(王小平等,
2018 ).豆科植物与根瘤菌的共生作用可使其在相对干旱的条件下快速生长(张仁懿等,
2014 ).这不仅消耗了土壤养分, 而且抑制了微生物活性以及酶活性.相比之下, 降水量增加和N添加显著地刺激了禾本科植物的生长(附录I; 朱湾湾等,
2019 ), 其自身的营养物质归还土壤后可为土壤酶的合成提供丰富的底物(乔文静等,
2018 ).其次, 降水量增加及N添加下植物生物量增加(附录I; 朱湾湾等,
2019 ), 提高了凋落物C和养分向土壤中的归还量(Zhong
et al .,
2017 ), 从而促进了蔗糖酶、脲酶和磷酸酶的产生. ...
... 研究表明, 土壤酶活性与植物群落多样性亦存在不同程度的相关性(王长庭等,
2010 ; 乔文静等,
2018 ).降水量和N添加及其交互作用改变了植物群落多样性(附录II; 朱湾湾等,
2019 ), 因而可能会对土壤酶活性产生重要影响.本研究中, 蔗糖酶活性随着Patrick丰富度指数增加而增加, 即蔗糖酶活性与植物物种数存在正线性关系.这可能是由于降水量增加和N添加下, 随着植物物种数增多, 土壤酶作用的底物多样性增加, 从而增强了蔗糖酶活性, 与对江河源区人工草地的研究结果(王启兰等,
2010 )一致; 脲酶和磷酸酶活性均与Shannon-Wiener多样性指数以及Pielou均匀度指数存在负线性关系.这可能是由于降水量和N添加下, 植物群落多样性和均匀度的改变直接影响到土壤微环境(李长斌等,
2016 ), 并通过对植物根系和微生物活性的影响, 间接影响到脲酶和磷酸酶活性.由于植物群落特征对土壤酶活性的影响存在时间滞后效应(乔文静等,
2018 ), 因此还需通过长期的试验观测深入探讨土壤酶活性与植物群落多样性之间的联系. ...
高寒草甸群落地表植被特征与土壤理化性状、土壤微生物之间的相关性研究
2
2010
... 本研究中, 降水量和N添加下脲酶和磷酸酶活性与草木樨状黄耆(豆科植物)生物量呈负线性关系, 而蔗糖酶、脲酶和磷酸酶活性均与糙隐子草(禾本科植物)生物量呈正线性关系, 与针对高寒草甸的研究结果(王长庭等,
2010 )存在差异.首先, 土壤酶活性与植物生物量之间的关系因植物种不同而异.这是由于不同植物根系的发达程度以及分泌物存在差异, 进而影响土壤酶活性(王小平等,
2018 ).豆科植物与根瘤菌的共生作用可使其在相对干旱的条件下快速生长(张仁懿等,
2014 ).这不仅消耗了土壤养分, 而且抑制了微生物活性以及酶活性.相比之下, 降水量增加和N添加显著地刺激了禾本科植物的生长(附录I; 朱湾湾等,
2019 ), 其自身的营养物质归还土壤后可为土壤酶的合成提供丰富的底物(乔文静等,
2018 ).其次, 降水量增加及N添加下植物生物量增加(附录I; 朱湾湾等,
2019 ), 提高了凋落物C和养分向土壤中的归还量(Zhong
et al .,
2017 ), 从而促进了蔗糖酶、脲酶和磷酸酶的产生. ...
... 研究表明, 土壤酶活性与植物群落多样性亦存在不同程度的相关性(王长庭等,
2010 ; 乔文静等,
2018 ).降水量和N添加及其交互作用改变了植物群落多样性(附录II; 朱湾湾等,
2019 ), 因而可能会对土壤酶活性产生重要影响.本研究中, 蔗糖酶活性随着Patrick丰富度指数增加而增加, 即蔗糖酶活性与植物物种数存在正线性关系.这可能是由于降水量增加和N添加下, 随着植物物种数增多, 土壤酶作用的底物多样性增加, 从而增强了蔗糖酶活性, 与对江河源区人工草地的研究结果(王启兰等,
2010 )一致; 脲酶和磷酸酶活性均与Shannon-Wiener多样性指数以及Pielou均匀度指数存在负线性关系.这可能是由于降水量和N添加下, 植物群落多样性和均匀度的改变直接影响到土壤微环境(李长斌等,
2016 ), 并通过对植物根系和微生物活性的影响, 间接影响到脲酶和磷酸酶活性.由于植物群落特征对土壤酶活性的影响存在时间滞后效应(乔文静等,
2018 ), 因此还需通过长期的试验观测深入探讨土壤酶活性与植物群落多样性之间的联系. ...
氮素和水分对贝加尔针茅草原土壤酶活性和微生物量碳氮的影响
2
2014a
... 土壤酶主要由植物根系和微生物分泌产生, 是土壤中元素生物地球化学循环的催化剂(马文文等,
2014 ).一方面, 土壤酶活性能够反映土壤元素(碳(C)、N、磷(P)等)转化和物质循环的速率(Burns
et al .,
2013 ), 并通过对土壤养分状况的影响改变植物养分策略.同时, 土壤酶能够实现植物营养元素和有机质的循环转化, 对土壤微环境的元素平衡和转化具有重要影响(张美曼等,
2020 ).另一方面, 土壤酶活性可表征微生物的养分需求、反映微生物在环境波动时维持养分平衡的应对策略(闫钟清等,
2017a ).土壤酶活性主要受土壤养分可利用性(李艳红等,
2020 )、植被类型(李静等,
2013 )和微生物活性(闫钟清等,
2017a )等因素的影响.降水和N添加通过调控土壤水分和养分有效性, 改变植物资源利用策略, 从而直接影响到植被结构(张金凤和许雨晴, 2016)和微生物活性(王杰等,
2014a ), 进而改变土壤酶活性(许华等,
2018 ).因此, 揭示降水量变化和N添加下土壤酶活性与植物和微生物的联系, 可为评价全球变化背景下植被-土壤系统功能提供新思路. ...
... 本研究中, 减少降水量对蔗糖酶、脲酶和磷酸酶活性的影响较小, 增加降水量不同程度地影响3种酶的活性.这可能是由于研究区长期处于低水平的土壤水分环境下, 植物和微生物对干旱具有高的适应性(胡小文等,
2004 ; 朱义族等,
2019 ), 因而减少降水量对其生长繁殖以及酶的分泌影响较小.相比之下, 在缺水环境下, 增加降水量提高了土壤含水量(附录III), 从而加速凋落物淋溶、促进表层凋落物和腐殖质中可溶性物质进入土壤(豆鹏鹏等,
2018 ), 使微生物可利用底物增加, 微生物活性相应增强, 进而刺激微生物向环境中释放酶(Schimel
et al .,
2007 ), 即微生物分泌的酶随之增加.此外, 本研究发现增加降水量对土壤酶活性的影响与N添加存在交互作用, 与其他研究结果(王杰等,
2014a )相似.这表明水分有利于N肥效的释放, 二者的交互作用促进了植物生长(白春利等,
2013 )、凋落物积累以及微生物活动(王杰等,
2014b ), 进而刺激土壤酶的分泌、导致土壤酶活性增强(闫钟清等,
2017b ). ...
氮素和水分对贝加尔针茅草原土壤酶活性和微生物量碳氮的影响
2
2014a
... 土壤酶主要由植物根系和微生物分泌产生, 是土壤中元素生物地球化学循环的催化剂(马文文等,
2014 ).一方面, 土壤酶活性能够反映土壤元素(碳(C)、N、磷(P)等)转化和物质循环的速率(Burns
et al .,
2013 ), 并通过对土壤养分状况的影响改变植物养分策略.同时, 土壤酶能够实现植物营养元素和有机质的循环转化, 对土壤微环境的元素平衡和转化具有重要影响(张美曼等,
2020 ).另一方面, 土壤酶活性可表征微生物的养分需求、反映微生物在环境波动时维持养分平衡的应对策略(闫钟清等,
2017a ).土壤酶活性主要受土壤养分可利用性(李艳红等,
2020 )、植被类型(李静等,
2013 )和微生物活性(闫钟清等,
2017a )等因素的影响.降水和N添加通过调控土壤水分和养分有效性, 改变植物资源利用策略, 从而直接影响到植被结构(张金凤和许雨晴, 2016)和微生物活性(王杰等,
2014a ), 进而改变土壤酶活性(许华等,
2018 ).因此, 揭示降水量变化和N添加下土壤酶活性与植物和微生物的联系, 可为评价全球变化背景下植被-土壤系统功能提供新思路. ...
... 本研究中, 减少降水量对蔗糖酶、脲酶和磷酸酶活性的影响较小, 增加降水量不同程度地影响3种酶的活性.这可能是由于研究区长期处于低水平的土壤水分环境下, 植物和微生物对干旱具有高的适应性(胡小文等,
2004 ; 朱义族等,
2019 ), 因而减少降水量对其生长繁殖以及酶的分泌影响较小.相比之下, 在缺水环境下, 增加降水量提高了土壤含水量(附录III), 从而加速凋落物淋溶、促进表层凋落物和腐殖质中可溶性物质进入土壤(豆鹏鹏等,
2018 ), 使微生物可利用底物增加, 微生物活性相应增强, 进而刺激微生物向环境中释放酶(Schimel
et al .,
2007 ), 即微生物分泌的酶随之增加.此外, 本研究发现增加降水量对土壤酶活性的影响与N添加存在交互作用, 与其他研究结果(王杰等,
2014a )相似.这表明水分有利于N肥效的释放, 二者的交互作用促进了植物生长(白春利等,
2013 )、凋落物积累以及微生物活动(王杰等,
2014b ), 进而刺激土壤酶的分泌、导致土壤酶活性增强(闫钟清等,
2017b ). ...
贝加尔针茅草原土壤微生物功能多样性对氮素和水分添加的响应
1
2014b
... 本研究中, 减少降水量对蔗糖酶、脲酶和磷酸酶活性的影响较小, 增加降水量不同程度地影响3种酶的活性.这可能是由于研究区长期处于低水平的土壤水分环境下, 植物和微生物对干旱具有高的适应性(胡小文等,
2004 ; 朱义族等,
2019 ), 因而减少降水量对其生长繁殖以及酶的分泌影响较小.相比之下, 在缺水环境下, 增加降水量提高了土壤含水量(附录III), 从而加速凋落物淋溶、促进表层凋落物和腐殖质中可溶性物质进入土壤(豆鹏鹏等,
2018 ), 使微生物可利用底物增加, 微生物活性相应增强, 进而刺激微生物向环境中释放酶(Schimel
et al .,
2007 ), 即微生物分泌的酶随之增加.此外, 本研究发现增加降水量对土壤酶活性的影响与N添加存在交互作用, 与其他研究结果(王杰等,
2014a )相似.这表明水分有利于N肥效的释放, 二者的交互作用促进了植物生长(白春利等,
2013 )、凋落物积累以及微生物活动(王杰等,
2014b ), 进而刺激土壤酶的分泌、导致土壤酶活性增强(闫钟清等,
2017b ). ...
贝加尔针茅草原土壤微生物功能多样性对氮素和水分添加的响应
1
2014b
... 本研究中, 减少降水量对蔗糖酶、脲酶和磷酸酶活性的影响较小, 增加降水量不同程度地影响3种酶的活性.这可能是由于研究区长期处于低水平的土壤水分环境下, 植物和微生物对干旱具有高的适应性(胡小文等,
2004 ; 朱义族等,
2019 ), 因而减少降水量对其生长繁殖以及酶的分泌影响较小.相比之下, 在缺水环境下, 增加降水量提高了土壤含水量(附录III), 从而加速凋落物淋溶、促进表层凋落物和腐殖质中可溶性物质进入土壤(豆鹏鹏等,
2018 ), 使微生物可利用底物增加, 微生物活性相应增强, 进而刺激微生物向环境中释放酶(Schimel
et al .,
2007 ), 即微生物分泌的酶随之增加.此外, 本研究发现增加降水量对土壤酶活性的影响与N添加存在交互作用, 与其他研究结果(王杰等,
2014a )相似.这表明水分有利于N肥效的释放, 二者的交互作用促进了植物生长(白春利等,
2013 )、凋落物积累以及微生物活动(王杰等,
2014b ), 进而刺激土壤酶的分泌、导致土壤酶活性增强(闫钟清等,
2017b ). ...
江河源区人工草地植物群落和土壤酶活性变化
2
2010
... 研究表明, 土壤酶活性与植物群落多样性亦存在不同程度的相关性(王长庭等,
2010 ; 乔文静等,
2018 ).降水量和N添加及其交互作用改变了植物群落多样性(附录II; 朱湾湾等,
2019 ), 因而可能会对土壤酶活性产生重要影响.本研究中, 蔗糖酶活性随着Patrick丰富度指数增加而增加, 即蔗糖酶活性与植物物种数存在正线性关系.这可能是由于降水量增加和N添加下, 随着植物物种数增多, 土壤酶作用的底物多样性增加, 从而增强了蔗糖酶活性, 与对江河源区人工草地的研究结果(王启兰等,
2010 )一致; 脲酶和磷酸酶活性均与Shannon-Wiener多样性指数以及Pielou均匀度指数存在负线性关系.这可能是由于降水量和N添加下, 植物群落多样性和均匀度的改变直接影响到土壤微环境(李长斌等,
2016 ), 并通过对植物根系和微生物活性的影响, 间接影响到脲酶和磷酸酶活性.由于植物群落特征对土壤酶活性的影响存在时间滞后效应(乔文静等,
2018 ), 因此还需通过长期的试验观测深入探讨土壤酶活性与植物群落多样性之间的联系. ...
... 除植物群落组成外, 土壤酶活性还受到土壤因子的综合影响(王启兰等,
2010 ; 李静等,
2013 ).本研究中, 与其他因子相比, 微生物生态化学计量特征对3种酶活性的影响较大.蔗糖酶活性与微生物生物量C含量、C:N和C:P呈负相关关系.其可能原因在于降水和N添加在一定程度上提高了土壤N和P的有效性(涂利华等,
2009 ), 限制了C的有效性, 微生物生物量C含量、C:N和C:P呈降低趋势(附录IV), 从而刺激微生物分泌较多的C获取酶以促进有机C矿化(Rappe-George
et al .,
2017 ).脲酶活性与微生物生物量C:N呈正相关关系, 与微生物生物量N含量呈负相关关系.这可能是由于持续增加的降水量加速了土壤N淋溶, 导致土壤N含量降低(附录III), 即N受限性增强, 进而提高土壤脲酶活性以促进有机N矿化(涂利华等,
2009 ; 刘星等,
2015 ).此外, 磷酸酶活性与微生物生物量C:P呈正相关关系, 与微生物生物量P含量呈负相关关系.结合2019年磷酸酶的变化特点(
图1 ), 以上结果表明降水量变化和N添加下微生物较多地受到土壤P有效性的限制.随着微生物对P的需求量增加, 相关水解酶活性相应增强(Steenbergh
et al .,
2011 ). ...
江河源区人工草地植物群落和土壤酶活性变化
2
2010
... 研究表明, 土壤酶活性与植物群落多样性亦存在不同程度的相关性(王长庭等,
2010 ; 乔文静等,
2018 ).降水量和N添加及其交互作用改变了植物群落多样性(附录II; 朱湾湾等,
2019 ), 因而可能会对土壤酶活性产生重要影响.本研究中, 蔗糖酶活性随着Patrick丰富度指数增加而增加, 即蔗糖酶活性与植物物种数存在正线性关系.这可能是由于降水量增加和N添加下, 随着植物物种数增多, 土壤酶作用的底物多样性增加, 从而增强了蔗糖酶活性, 与对江河源区人工草地的研究结果(王启兰等,
2010 )一致; 脲酶和磷酸酶活性均与Shannon-Wiener多样性指数以及Pielou均匀度指数存在负线性关系.这可能是由于降水量和N添加下, 植物群落多样性和均匀度的改变直接影响到土壤微环境(李长斌等,
2016 ), 并通过对植物根系和微生物活性的影响, 间接影响到脲酶和磷酸酶活性.由于植物群落特征对土壤酶活性的影响存在时间滞后效应(乔文静等,
2018 ), 因此还需通过长期的试验观测深入探讨土壤酶活性与植物群落多样性之间的联系. ...
... 除植物群落组成外, 土壤酶活性还受到土壤因子的综合影响(王启兰等,
2010 ; 李静等,
2013 ).本研究中, 与其他因子相比, 微生物生态化学计量特征对3种酶活性的影响较大.蔗糖酶活性与微生物生物量C含量、C:N和C:P呈负相关关系.其可能原因在于降水和N添加在一定程度上提高了土壤N和P的有效性(涂利华等,
2009 ), 限制了C的有效性, 微生物生物量C含量、C:N和C:P呈降低趋势(附录IV), 从而刺激微生物分泌较多的C获取酶以促进有机C矿化(Rappe-George
et al .,
2017 ).脲酶活性与微生物生物量C:N呈正相关关系, 与微生物生物量N含量呈负相关关系.这可能是由于持续增加的降水量加速了土壤N淋溶, 导致土壤N含量降低(附录III), 即N受限性增强, 进而提高土壤脲酶活性以促进有机N矿化(涂利华等,
2009 ; 刘星等,
2015 ).此外, 磷酸酶活性与微生物生物量C:P呈正相关关系, 与微生物生物量P含量呈负相关关系.结合2019年磷酸酶的变化特点(
图1 ), 以上结果表明降水量变化和N添加下微生物较多地受到土壤P有效性的限制.随着微生物对P的需求量增加, 相关水解酶活性相应增强(Steenbergh
et al .,
2011 ). ...
连香树人工林根系分泌物输入季节性变化及其驱动的根际微生物特性研究
1
2018
... 本研究中, 降水量和N添加下脲酶和磷酸酶活性与草木樨状黄耆(豆科植物)生物量呈负线性关系, 而蔗糖酶、脲酶和磷酸酶活性均与糙隐子草(禾本科植物)生物量呈正线性关系, 与针对高寒草甸的研究结果(王长庭等,
2010 )存在差异.首先, 土壤酶活性与植物生物量之间的关系因植物种不同而异.这是由于不同植物根系的发达程度以及分泌物存在差异, 进而影响土壤酶活性(王小平等,
2018 ).豆科植物与根瘤菌的共生作用可使其在相对干旱的条件下快速生长(张仁懿等,
2014 ).这不仅消耗了土壤养分, 而且抑制了微生物活性以及酶活性.相比之下, 降水量增加和N添加显著地刺激了禾本科植物的生长(附录I; 朱湾湾等,
2019 ), 其自身的营养物质归还土壤后可为土壤酶的合成提供丰富的底物(乔文静等,
2018 ).其次, 降水量增加及N添加下植物生物量增加(附录I; 朱湾湾等,
2019 ), 提高了凋落物C和养分向土壤中的归还量(Zhong
et al .,
2017 ), 从而促进了蔗糖酶、脲酶和磷酸酶的产生. ...
连香树人工林根系分泌物输入季节性变化及其驱动的根际微生物特性研究
1
2018
... 本研究中, 降水量和N添加下脲酶和磷酸酶活性与草木樨状黄耆(豆科植物)生物量呈负线性关系, 而蔗糖酶、脲酶和磷酸酶活性均与糙隐子草(禾本科植物)生物量呈正线性关系, 与针对高寒草甸的研究结果(王长庭等,
2010 )存在差异.首先, 土壤酶活性与植物生物量之间的关系因植物种不同而异.这是由于不同植物根系的发达程度以及分泌物存在差异, 进而影响土壤酶活性(王小平等,
2018 ).豆科植物与根瘤菌的共生作用可使其在相对干旱的条件下快速生长(张仁懿等,
2014 ).这不仅消耗了土壤养分, 而且抑制了微生物活性以及酶活性.相比之下, 降水量增加和N添加显著地刺激了禾本科植物的生长(附录I; 朱湾湾等,
2019 ), 其自身的营养物质归还土壤后可为土壤酶的合成提供丰富的底物(乔文静等,
2018 ).其次, 降水量增加及N添加下植物生物量增加(附录I; 朱湾湾等,
2019 ), 提高了凋落物C和养分向土壤中的归还量(Zhong
et al .,
2017 ), 从而促进了蔗糖酶、脲酶和磷酸酶的产生. ...
长沙市城郊森林土壤酶活性及其与土壤理化性质的相关性
1
2010
... 此外, 对土壤酶活性影响显著的土壤因子还包括pH、全P含量、N:P、C:P、NH
4 + -N以及NO
3 - -N含量.土壤pH是影响土壤酶活性的重要因素(苏鑫等,
2020 ), 能够调控有机大分子物质的生物降解和矿化过程(万忠梅和宋长春,
2009 ).本研究中, 脲酶和磷酸酶活性均与土壤pH负相关, 意味着研究区土壤较高的pH (附录III)抑制了二者活性, 与对森林土壤的研究结果(吴际友等,
2010 )一致.脲酶活性与土壤N:P以及NH
4 + -N含量正相关.这表明虽然N添加提高了土壤N有效性, 但逐渐增加的降水量导致土壤N淋溶作用加强, 植物和微生物对N的竞争增强, 进而导致脲酶分泌量增多(刘星等,
2015 ).另外, 磷酸酶活性与土壤C:P和N:P正相关, 与全P含量负相关, 与其他研究结果(Allison
et al .,
2007 )相似.这可能是当土壤中P供给不足时, 植物会分泌磷酸酶以促进土壤有机P的矿化, 生成可供植物吸收的无机态P (附录III), 导致土壤全P含量降低(Deng
et al .,
2017 ).总之, 土壤pH和养分有效性通过调控养分固定和矿化等过程, 以正、负效应的形式对土壤酶活性进行调节和驱动. ...
长沙市城郊森林土壤酶活性及其与土壤理化性质的相关性
1
2010
... 此外, 对土壤酶活性影响显著的土壤因子还包括pH、全P含量、N:P、C:P、NH
4 + -N以及NO
3 - -N含量.土壤pH是影响土壤酶活性的重要因素(苏鑫等,
2020 ), 能够调控有机大分子物质的生物降解和矿化过程(万忠梅和宋长春,
2009 ).本研究中, 脲酶和磷酸酶活性均与土壤pH负相关, 意味着研究区土壤较高的pH (附录III)抑制了二者活性, 与对森林土壤的研究结果(吴际友等,
2010 )一致.脲酶活性与土壤N:P以及NH
4 + -N含量正相关.这表明虽然N添加提高了土壤N有效性, 但逐渐增加的降水量导致土壤N淋溶作用加强, 植物和微生物对N的竞争增强, 进而导致脲酶分泌量增多(刘星等,
2015 ).另外, 磷酸酶活性与土壤C:P和N:P正相关, 与全P含量负相关, 与其他研究结果(Allison
et al .,
2007 )相似.这可能是当土壤中P供给不足时, 植物会分泌磷酸酶以促进土壤有机P的矿化, 生成可供植物吸收的无机态P (附录III), 导致土壤全P含量降低(Deng
et al .,
2017 ).总之, 土壤pH和养分有效性通过调控养分固定和矿化等过程, 以正、负效应的形式对土壤酶活性进行调节和驱动. ...
干旱荒漠区土壤酶活性对降水调控的响应
2
2018
... 土壤酶主要由植物根系和微生物分泌产生, 是土壤中元素生物地球化学循环的催化剂(马文文等,
2014 ).一方面, 土壤酶活性能够反映土壤元素(碳(C)、N、磷(P)等)转化和物质循环的速率(Burns
et al .,
2013 ), 并通过对土壤养分状况的影响改变植物养分策略.同时, 土壤酶能够实现植物营养元素和有机质的循环转化, 对土壤微环境的元素平衡和转化具有重要影响(张美曼等,
2020 ).另一方面, 土壤酶活性可表征微生物的养分需求、反映微生物在环境波动时维持养分平衡的应对策略(闫钟清等,
2017a ).土壤酶活性主要受土壤养分可利用性(李艳红等,
2020 )、植被类型(李静等,
2013 )和微生物活性(闫钟清等,
2017a )等因素的影响.降水和N添加通过调控土壤水分和养分有效性, 改变植物资源利用策略, 从而直接影响到植被结构(张金凤和许雨晴, 2016)和微生物活性(王杰等,
2014a ), 进而改变土壤酶活性(许华等,
2018 ).因此, 揭示降水量变化和N添加下土壤酶活性与植物和微生物的联系, 可为评价全球变化背景下植被-土壤系统功能提供新思路. ...
... 于2017年9月, 在围栏草地内选择30 m × 50 m的区域作为降水量和N添加处理的试验样地.降水增减量依据近几十年来我国西北地区的西部生态区降水量增加而东部生态区降水量减少的趋势进行设置, 同时参考了国内同类研究方法(Xu
et al .,
2018 ).氮肥施用量依据区域N沉降水平(顾峰雪等,
2016 )和N饱和阈值(Tian
et al .,
2016 )进行设置.试验设计采用随机区组的裂区设计, 主区为降水量, 副区为N添加.降水量处理包括减少50% (W1)、减少30% (W2)、对照(自然降水量, W3)、增加30% (W4)和增加50% (W5).N添加处理包括0 g·m
-2 ·a
-1 (N0)和5 g·m
-2 ·a
-1 (N5).试验包含10个处理, 每个处理3次重复, 共计30个小区(
表2 ).每个主区面积为8 m × 8 m, 每个副区面积为4 m × 8 m.各主区之间设置1.5 m的缓冲带.各主区的两个副区间垂直下埋1 m宽的塑料布, 上埋20 cm宽的彩钢板, 用来防止试验干扰. ...
干旱荒漠区土壤酶活性对降水调控的响应
2
2018
... 土壤酶主要由植物根系和微生物分泌产生, 是土壤中元素生物地球化学循环的催化剂(马文文等,
2014 ).一方面, 土壤酶活性能够反映土壤元素(碳(C)、N、磷(P)等)转化和物质循环的速率(Burns
et al .,
2013 ), 并通过对土壤养分状况的影响改变植物养分策略.同时, 土壤酶能够实现植物营养元素和有机质的循环转化, 对土壤微环境的元素平衡和转化具有重要影响(张美曼等,
2020 ).另一方面, 土壤酶活性可表征微生物的养分需求、反映微生物在环境波动时维持养分平衡的应对策略(闫钟清等,
2017a ).土壤酶活性主要受土壤养分可利用性(李艳红等,
2020 )、植被类型(李静等,
2013 )和微生物活性(闫钟清等,
2017a )等因素的影响.降水和N添加通过调控土壤水分和养分有效性, 改变植物资源利用策略, 从而直接影响到植被结构(张金凤和许雨晴, 2016)和微生物活性(王杰等,
2014a ), 进而改变土壤酶活性(许华等,
2018 ).因此, 揭示降水量变化和N添加下土壤酶活性与植物和微生物的联系, 可为评价全球变化背景下植被-土壤系统功能提供新思路. ...
... 于2017年9月, 在围栏草地内选择30 m × 50 m的区域作为降水量和N添加处理的试验样地.降水增减量依据近几十年来我国西北地区的西部生态区降水量增加而东部生态区降水量减少的趋势进行设置, 同时参考了国内同类研究方法(Xu
et al .,
2018 ).氮肥施用量依据区域N沉降水平(顾峰雪等,
2016 )和N饱和阈值(Tian
et al .,
2016 )进行设置.试验设计采用随机区组的裂区设计, 主区为降水量, 副区为N添加.降水量处理包括减少50% (W1)、减少30% (W2)、对照(自然降水量, W3)、增加30% (W4)和增加50% (W5).N添加处理包括0 g·m
-2 ·a
-1 (N0)和5 g·m
-2 ·a
-1 (N5).试验包含10个处理, 每个处理3次重复, 共计30个小区(
表2 ).每个主区面积为8 m × 8 m, 每个副区面积为4 m × 8 m.各主区之间设置1.5 m的缓冲带.各主区的两个副区间垂直下埋1 m宽的塑料布, 上埋20 cm宽的彩钢板, 用来防止试验干扰. ...
Plant functional diversity modulates global environmental change effects on grassland productivity
2018
降水和氮沉降增加对草地土壤微生物与酶活性的影响研究进展
2
2017a
... 土壤酶主要由植物根系和微生物分泌产生, 是土壤中元素生物地球化学循环的催化剂(马文文等,
2014 ).一方面, 土壤酶活性能够反映土壤元素(碳(C)、N、磷(P)等)转化和物质循环的速率(Burns
et al .,
2013 ), 并通过对土壤养分状况的影响改变植物养分策略.同时, 土壤酶能够实现植物营养元素和有机质的循环转化, 对土壤微环境的元素平衡和转化具有重要影响(张美曼等,
2020 ).另一方面, 土壤酶活性可表征微生物的养分需求、反映微生物在环境波动时维持养分平衡的应对策略(闫钟清等,
2017a ).土壤酶活性主要受土壤养分可利用性(李艳红等,
2020 )、植被类型(李静等,
2013 )和微生物活性(闫钟清等,
2017a )等因素的影响.降水和N添加通过调控土壤水分和养分有效性, 改变植物资源利用策略, 从而直接影响到植被结构(张金凤和许雨晴, 2016)和微生物活性(王杰等,
2014a ), 进而改变土壤酶活性(许华等,
2018 ).因此, 揭示降水量变化和N添加下土壤酶活性与植物和微生物的联系, 可为评价全球变化背景下植被-土壤系统功能提供新思路. ...
... )和微生物活性(闫钟清等,
2017a )等因素的影响.降水和N添加通过调控土壤水分和养分有效性, 改变植物资源利用策略, 从而直接影响到植被结构(张金凤和许雨晴, 2016)和微生物活性(王杰等,
2014a ), 进而改变土壤酶活性(许华等,
2018 ).因此, 揭示降水量变化和N添加下土壤酶活性与植物和微生物的联系, 可为评价全球变化背景下植被-土壤系统功能提供新思路. ...
降水和氮沉降增加对草地土壤微生物与酶活性的影响研究进展
2
2017a
... 土壤酶主要由植物根系和微生物分泌产生, 是土壤中元素生物地球化学循环的催化剂(马文文等,
2014 ).一方面, 土壤酶活性能够反映土壤元素(碳(C)、N、磷(P)等)转化和物质循环的速率(Burns
et al .,
2013 ), 并通过对土壤养分状况的影响改变植物养分策略.同时, 土壤酶能够实现植物营养元素和有机质的循环转化, 对土壤微环境的元素平衡和转化具有重要影响(张美曼等,
2020 ).另一方面, 土壤酶活性可表征微生物的养分需求、反映微生物在环境波动时维持养分平衡的应对策略(闫钟清等,
2017a ).土壤酶活性主要受土壤养分可利用性(李艳红等,
2020 )、植被类型(李静等,
2013 )和微生物活性(闫钟清等,
2017a )等因素的影响.降水和N添加通过调控土壤水分和养分有效性, 改变植物资源利用策略, 从而直接影响到植被结构(张金凤和许雨晴, 2016)和微生物活性(王杰等,
2014a ), 进而改变土壤酶活性(许华等,
2018 ).因此, 揭示降水量变化和N添加下土壤酶活性与植物和微生物的联系, 可为评价全球变化背景下植被-土壤系统功能提供新思路. ...
... )和微生物活性(闫钟清等,
2017a )等因素的影响.降水和N添加通过调控土壤水分和养分有效性, 改变植物资源利用策略, 从而直接影响到植被结构(张金凤和许雨晴, 2016)和微生物活性(王杰等,
2014a ), 进而改变土壤酶活性(许华等,
2018 ).因此, 揭示降水量变化和N添加下土壤酶活性与植物和微生物的联系, 可为评价全球变化背景下植被-土壤系统功能提供新思路. ...
降水和氮沉降增加对草地土壤酶活性的影响
1
2017b
... 本研究中, 减少降水量对蔗糖酶、脲酶和磷酸酶活性的影响较小, 增加降水量不同程度地影响3种酶的活性.这可能是由于研究区长期处于低水平的土壤水分环境下, 植物和微生物对干旱具有高的适应性(胡小文等,
2004 ; 朱义族等,
2019 ), 因而减少降水量对其生长繁殖以及酶的分泌影响较小.相比之下, 在缺水环境下, 增加降水量提高了土壤含水量(附录III), 从而加速凋落物淋溶、促进表层凋落物和腐殖质中可溶性物质进入土壤(豆鹏鹏等,
2018 ), 使微生物可利用底物增加, 微生物活性相应增强, 进而刺激微生物向环境中释放酶(Schimel
et al .,
2007 ), 即微生物分泌的酶随之增加.此外, 本研究发现增加降水量对土壤酶活性的影响与N添加存在交互作用, 与其他研究结果(王杰等,
2014a )相似.这表明水分有利于N肥效的释放, 二者的交互作用促进了植物生长(白春利等,
2013 )、凋落物积累以及微生物活动(王杰等,
2014b ), 进而刺激土壤酶的分泌、导致土壤酶活性增强(闫钟清等,
2017b ). ...
降水和氮沉降增加对草地土壤酶活性的影响
1
2017b
... 本研究中, 减少降水量对蔗糖酶、脲酶和磷酸酶活性的影响较小, 增加降水量不同程度地影响3种酶的活性.这可能是由于研究区长期处于低水平的土壤水分环境下, 植物和微生物对干旱具有高的适应性(胡小文等,
2004 ; 朱义族等,
2019 ), 因而减少降水量对其生长繁殖以及酶的分泌影响较小.相比之下, 在缺水环境下, 增加降水量提高了土壤含水量(附录III), 从而加速凋落物淋溶、促进表层凋落物和腐殖质中可溶性物质进入土壤(豆鹏鹏等,
2018 ), 使微生物可利用底物增加, 微生物活性相应增强, 进而刺激微生物向环境中释放酶(Schimel
et al .,
2007 ), 即微生物分泌的酶随之增加.此外, 本研究发现增加降水量对土壤酶活性的影响与N添加存在交互作用, 与其他研究结果(王杰等,
2014a )相似.这表明水分有利于N肥效的释放, 二者的交互作用促进了植物生长(白春利等,
2013 )、凋落物积累以及微生物活动(王杰等,
2014b ), 进而刺激土壤酶的分泌、导致土壤酶活性增强(闫钟清等,
2017b ). ...
气候变暖背景下西北脆弱带近54年降水季节变化特征
1
2018
... 荒漠草原是我国西北干旱半干旱区主要的草原生态系统类型, 其植被结构简单、生态系统脆弱.宁夏荒漠草原主要分布于毛乌素沙地西南缘, 是区域重要的生态屏障.该生态系统降水量少、蒸发量大、N沉降临界负荷低, 降水和N是其植被生长和微生物活动的主要限制因子, 因此对降水格局改变和N沉降增加较为敏感(段雷等,
2002 ; 杨蓉和赵多平,
2018 ).那么, 降水量和N添加及其交互作用如何影响荒漠草原土壤酶活性, 降水量改变和N添加下土壤酶活性与植物群落组成以及微生物活动存在怎样的联系等问题, 都值得我们进行深入探讨.为此, 本项目组于2017年在宁夏荒漠草原设立了降水量改变及N添加的裂区试验, 研究了2018-2019年生长季旺盛期土壤酶活性的变化特点, 分析了其与植物群落组成以及微生物生物量C:N:P生态化学计量特征(其值高低与微生物活动的方向密切相关)的关系, 为充分认识降水量变化和N添加对土壤酶活性的影响机制提供基础数据, 并为深入理解全球变化背景下西北脆弱生态区的元素生物地球化学循环提供理论依据. ...
气候变暖背景下西北脆弱带近54年降水季节变化特征
1
2018
... 荒漠草原是我国西北干旱半干旱区主要的草原生态系统类型, 其植被结构简单、生态系统脆弱.宁夏荒漠草原主要分布于毛乌素沙地西南缘, 是区域重要的生态屏障.该生态系统降水量少、蒸发量大、N沉降临界负荷低, 降水和N是其植被生长和微生物活动的主要限制因子, 因此对降水格局改变和N沉降增加较为敏感(段雷等,
2002 ; 杨蓉和赵多平,
2018 ).那么, 降水量和N添加及其交互作用如何影响荒漠草原土壤酶活性, 降水量改变和N添加下土壤酶活性与植物群落组成以及微生物活动存在怎样的联系等问题, 都值得我们进行深入探讨.为此, 本项目组于2017年在宁夏荒漠草原设立了降水量改变及N添加的裂区试验, 研究了2018-2019年生长季旺盛期土壤酶活性的变化特点, 分析了其与植物群落组成以及微生物生物量C:N:P生态化学计量特征(其值高低与微生物活动的方向密切相关)的关系, 为充分认识降水量变化和N添加对土壤酶活性的影响机制提供基础数据, 并为深入理解全球变化背景下西北脆弱生态区的元素生物地球化学循环提供理论依据. ...
土壤酶研究动态与展望
1
2002
... 采用SigmaPlot 12.0进行图的绘制.采用SPSS 19.0进行数据的统计分析: 采用K-S检验进行数据的正态分布检验; 采用三因素方差分析法分析年份、降水量、N添加及其交互作用对土壤酶活性的影响; 采用单因素方差分析进行相同N添加水平下各指标在不同降水量处理间, 以及相同降水量下各指标在不同N处理间的差异分析.若方差为齐性, 选用最小显著性差异(
LSD )法, 否则选用Games-Howell法; 由于植物生物量能够影响土壤酶活性(杨万勤和王开运,
2002 ), 故采用线性回归方程拟合土壤酶活性与植物种群生物量以及群落多样性之间的关系.采用Canoco 5.0进行土壤酶活性与微生物生物量C:N:P生态化学计量特征及其他环境因子对应关系的冗余分析(RDA).分析前, 先对数据进行对数转换, 以减少数据间差异.以全部环境因子作为解释变量, 以土壤酶活性作为响应变量, 依据解释变量前项选择, 剔除影响力较小的解释变量后进行RDA, 并通过蒙特卡洛置换检验得出每个环境因子的条件效应. ...
土壤酶研究动态与展望
1
2002
... 采用SigmaPlot 12.0进行图的绘制.采用SPSS 19.0进行数据的统计分析: 采用K-S检验进行数据的正态分布检验; 采用三因素方差分析法分析年份、降水量、N添加及其交互作用对土壤酶活性的影响; 采用单因素方差分析进行相同N添加水平下各指标在不同降水量处理间, 以及相同降水量下各指标在不同N处理间的差异分析.若方差为齐性, 选用最小显著性差异(
LSD )法, 否则选用Games-Howell法; 由于植物生物量能够影响土壤酶活性(杨万勤和王开运,
2002 ), 故采用线性回归方程拟合土壤酶活性与植物种群生物量以及群落多样性之间的关系.采用Canoco 5.0进行土壤酶活性与微生物生物量C:N:P生态化学计量特征及其他环境因子对应关系的冗余分析(RDA).分析前, 先对数据进行对数转换, 以减少数据间差异.以全部环境因子作为解释变量, 以土壤酶活性作为响应变量, 依据解释变量前项选择, 剔除影响力较小的解释变量后进行RDA, 并通过蒙特卡洛置换检验得出每个环境因子的条件效应. ...
Stabilization of atmospheric nitrogen deposition in China over the past decade
1
2019
... 降水格局改变和大气氮(N)沉降增加是全球变化的两个重要表现(IPCC, 2013).全球变暖背景下, 降水量在全球范围内呈增加趋势, 同时表现出时空分配不均和极端事件频发等特征(IPCC, 2013).据报道, 我国西北地区的西部生态区降水量呈增加趋势, 东部生态区降水量有所降低(高继卿等,
2015 ).此外, 人类活动导致全球大气N沉降速率持续增加(Kanakidou
et al .,
2016 ).就中国而言, 虽然N沉降速率在一些南方省市有所下降, 但在西北部分区域呈增加趋势(Yu
et al .,
2017 ,
2019 ).西北干旱半干旱区生态系统稳定性差, 对环境变化敏感, 是我国典型的生态脆弱区之一.作为该区的主要限制因子, 降水和N调控着土壤资源有效性, 影响着植物生长和微生物活动等关键生态过程, 与植被-土壤系统的稳定性密切相关.目前, 国内已有较多研究探讨了降水和N添加对该脆弱生态区的影响, 尚缺乏二者交互作用的相关报道.鉴于降水在介导N沉降效应中发挥着重要作用(Lü
et al .,
2012 ), 因此有必要针对西北干旱半干旱区典型生态系统开展二者交互效应的研究. ...
Development of atmospheric acid deposition in China from the 1990s to the 2010s
1
2017
... 降水格局改变和大气氮(N)沉降增加是全球变化的两个重要表现(IPCC, 2013).全球变暖背景下, 降水量在全球范围内呈增加趋势, 同时表现出时空分配不均和极端事件频发等特征(IPCC, 2013).据报道, 我国西北地区的西部生态区降水量呈增加趋势, 东部生态区降水量有所降低(高继卿等,
2015 ).此外, 人类活动导致全球大气N沉降速率持续增加(Kanakidou
et al .,
2016 ).就中国而言, 虽然N沉降速率在一些南方省市有所下降, 但在西北部分区域呈增加趋势(Yu
et al .,
2017 ,
2019 ).西北干旱半干旱区生态系统稳定性差, 对环境变化敏感, 是我国典型的生态脆弱区之一.作为该区的主要限制因子, 降水和N调控着土壤资源有效性, 影响着植物生长和微生物活动等关键生态过程, 与植被-土壤系统的稳定性密切相关.目前, 国内已有较多研究探讨了降水和N添加对该脆弱生态区的影响, 尚缺乏二者交互作用的相关报道.鉴于降水在介导N沉降效应中发挥着重要作用(Lü
et al .,
2012 ), 因此有必要针对西北干旱半干旱区典型生态系统开展二者交互效应的研究. ...
水氮添加对内蒙古多伦县退耕还草地生物量、生产力及其分配的影响
2016
水氮添加对内蒙古多伦县退耕还草地生物量、生产力及其分配的影响
2016
竹阔混交林土壤微生物生物量及酶活性特征研究
1
2020
... 土壤酶主要由植物根系和微生物分泌产生, 是土壤中元素生物地球化学循环的催化剂(马文文等,
2014 ).一方面, 土壤酶活性能够反映土壤元素(碳(C)、N、磷(P)等)转化和物质循环的速率(Burns
et al .,
2013 ), 并通过对土壤养分状况的影响改变植物养分策略.同时, 土壤酶能够实现植物营养元素和有机质的循环转化, 对土壤微环境的元素平衡和转化具有重要影响(张美曼等,
2020 ).另一方面, 土壤酶活性可表征微生物的养分需求、反映微生物在环境波动时维持养分平衡的应对策略(闫钟清等,
2017a ).土壤酶活性主要受土壤养分可利用性(李艳红等,
2020 )、植被类型(李静等,
2013 )和微生物活性(闫钟清等,
2017a )等因素的影响.降水和N添加通过调控土壤水分和养分有效性, 改变植物资源利用策略, 从而直接影响到植被结构(张金凤和许雨晴, 2016)和微生物活性(王杰等,
2014a ), 进而改变土壤酶活性(许华等,
2018 ).因此, 揭示降水量变化和N添加下土壤酶活性与植物和微生物的联系, 可为评价全球变化背景下植被-土壤系统功能提供新思路. ...
竹阔混交林土壤微生物生物量及酶活性特征研究
1
2020
... 土壤酶主要由植物根系和微生物分泌产生, 是土壤中元素生物地球化学循环的催化剂(马文文等,
2014 ).一方面, 土壤酶活性能够反映土壤元素(碳(C)、N、磷(P)等)转化和物质循环的速率(Burns
et al .,
2013 ), 并通过对土壤养分状况的影响改变植物养分策略.同时, 土壤酶能够实现植物营养元素和有机质的循环转化, 对土壤微环境的元素平衡和转化具有重要影响(张美曼等,
2020 ).另一方面, 土壤酶活性可表征微生物的养分需求、反映微生物在环境波动时维持养分平衡的应对策略(闫钟清等,
2017a ).土壤酶活性主要受土壤养分可利用性(李艳红等,
2020 )、植被类型(李静等,
2013 )和微生物活性(闫钟清等,
2017a )等因素的影响.降水和N添加通过调控土壤水分和养分有效性, 改变植物资源利用策略, 从而直接影响到植被结构(张金凤和许雨晴, 2016)和微生物活性(王杰等,
2014a ), 进而改变土壤酶活性(许华等,
2018 ).因此, 揭示降水量变化和N添加下土壤酶活性与植物和微生物的联系, 可为评价全球变化背景下植被-土壤系统功能提供新思路. ...
基于N:P化学计量特征的高寒草甸植物养分状况研究
1
2014
... 本研究中, 降水量和N添加下脲酶和磷酸酶活性与草木樨状黄耆(豆科植物)生物量呈负线性关系, 而蔗糖酶、脲酶和磷酸酶活性均与糙隐子草(禾本科植物)生物量呈正线性关系, 与针对高寒草甸的研究结果(王长庭等,
2010 )存在差异.首先, 土壤酶活性与植物生物量之间的关系因植物种不同而异.这是由于不同植物根系的发达程度以及分泌物存在差异, 进而影响土壤酶活性(王小平等,
2018 ).豆科植物与根瘤菌的共生作用可使其在相对干旱的条件下快速生长(张仁懿等,
2014 ).这不仅消耗了土壤养分, 而且抑制了微生物活性以及酶活性.相比之下, 降水量增加和N添加显著地刺激了禾本科植物的生长(附录I; 朱湾湾等,
2019 ), 其自身的营养物质归还土壤后可为土壤酶的合成提供丰富的底物(乔文静等,
2018 ).其次, 降水量增加及N添加下植物生物量增加(附录I; 朱湾湾等,
2019 ), 提高了凋落物C和养分向土壤中的归还量(Zhong
et al .,
2017 ), 从而促进了蔗糖酶、脲酶和磷酸酶的产生. ...
基于N:P化学计量特征的高寒草甸植物养分状况研究
1
2014
... 本研究中, 降水量和N添加下脲酶和磷酸酶活性与草木樨状黄耆(豆科植物)生物量呈负线性关系, 而蔗糖酶、脲酶和磷酸酶活性均与糙隐子草(禾本科植物)生物量呈正线性关系, 与针对高寒草甸的研究结果(王长庭等,
2010 )存在差异.首先, 土壤酶活性与植物生物量之间的关系因植物种不同而异.这是由于不同植物根系的发达程度以及分泌物存在差异, 进而影响土壤酶活性(王小平等,
2018 ).豆科植物与根瘤菌的共生作用可使其在相对干旱的条件下快速生长(张仁懿等,
2014 ).这不仅消耗了土壤养分, 而且抑制了微生物活性以及酶活性.相比之下, 降水量增加和N添加显著地刺激了禾本科植物的生长(附录I; 朱湾湾等,
2019 ), 其自身的营养物质归还土壤后可为土壤酶的合成提供丰富的底物(乔文静等,
2018 ).其次, 降水量增加及N添加下植物生物量增加(附录I; 朱湾湾等,
2019 ), 提高了凋落物C和养分向土壤中的归还量(Zhong
et al .,
2017 ), 从而促进了蔗糖酶、脲酶和磷酸酶的产生. ...
Differential responses of litter decomposition to nutrient addition and soil water availability with long-term vegetation recovery
1
2017
... 本研究中, 降水量和N添加下脲酶和磷酸酶活性与草木樨状黄耆(豆科植物)生物量呈负线性关系, 而蔗糖酶、脲酶和磷酸酶活性均与糙隐子草(禾本科植物)生物量呈正线性关系, 与针对高寒草甸的研究结果(王长庭等,
2010 )存在差异.首先, 土壤酶活性与植物生物量之间的关系因植物种不同而异.这是由于不同植物根系的发达程度以及分泌物存在差异, 进而影响土壤酶活性(王小平等,
2018 ).豆科植物与根瘤菌的共生作用可使其在相对干旱的条件下快速生长(张仁懿等,
2014 ).这不仅消耗了土壤养分, 而且抑制了微生物活性以及酶活性.相比之下, 降水量增加和N添加显著地刺激了禾本科植物的生长(附录I; 朱湾湾等,
2019 ), 其自身的营养物质归还土壤后可为土壤酶的合成提供丰富的底物(乔文静等,
2018 ).其次, 降水量增加及N添加下植物生物量增加(附录I; 朱湾湾等,
2019 ), 提高了凋落物C和养分向土壤中的归还量(Zhong
et al .,
2017 ), 从而促进了蔗糖酶、脲酶和磷酸酶的产生. ...
降水量及N添加对宁夏荒漠草原土壤C:N:P生态化学计量特征和植被群落组成的影响
6
2019
... 降水量减少处理采用自制的减雨架进行全年遮雨.减雨架上端离地约1.8 m, 用5 cm宽的瓦面状高透光性聚氯乙烯板搭建分别相当于50%和30%小区面积的遮雨面.降水量增加处理采用人工喷灌装置实现.喷水时间为每年5-8月的每月月初和月中.选用分析纯NH
4 NO
3 (34% N)进行N添加处理.施N时间为每年5-8月的每月月初.详细的试验处理方法见朱湾湾等(
2019 ). ...
... 同期, 在每个小区内随机选取1 m × 1 m的小样方进行植被群落调查.调查指标包括物种数和物种高度等.调查结束后, 齐平地面剪下地上部分, 按牛枝子、草木樨状黄耆、白草、针茅、糙隐子草及其他物种归类分装于纸袋中带回实验室烘干(65 ℃, 48 h)称质量.群落生物量为所有物种生物量之和.植物群落多样性采用Patrick丰富度指数、Shannon- Wiener多样性指数、Simpson优势度指数和Pielou均匀度指数等指标来表征, 具体计算方法见朱湾湾等(
2019 ).2018年植物生物量、群落多样性以及土壤理化性质引自朱湾湾等(
2019 ).2019年植物生物量、群落多样性以及土壤理化性质的结果见附录I-IV. ...
... ).2018年植物生物量、群落多样性以及土壤理化性质引自朱湾湾等(
2019 ).2019年植物生物量、群落多样性以及土壤理化性质的结果见附录I-IV. ...
... 本研究中, 降水量和N添加下脲酶和磷酸酶活性与草木樨状黄耆(豆科植物)生物量呈负线性关系, 而蔗糖酶、脲酶和磷酸酶活性均与糙隐子草(禾本科植物)生物量呈正线性关系, 与针对高寒草甸的研究结果(王长庭等,
2010 )存在差异.首先, 土壤酶活性与植物生物量之间的关系因植物种不同而异.这是由于不同植物根系的发达程度以及分泌物存在差异, 进而影响土壤酶活性(王小平等,
2018 ).豆科植物与根瘤菌的共生作用可使其在相对干旱的条件下快速生长(张仁懿等,
2014 ).这不仅消耗了土壤养分, 而且抑制了微生物活性以及酶活性.相比之下, 降水量增加和N添加显著地刺激了禾本科植物的生长(附录I; 朱湾湾等,
2019 ), 其自身的营养物质归还土壤后可为土壤酶的合成提供丰富的底物(乔文静等,
2018 ).其次, 降水量增加及N添加下植物生物量增加(附录I; 朱湾湾等,
2019 ), 提高了凋落物C和养分向土壤中的归还量(Zhong
et al .,
2017 ), 从而促进了蔗糖酶、脲酶和磷酸酶的产生. ...
... ).其次, 降水量增加及N添加下植物生物量增加(附录I; 朱湾湾等,
2019 ), 提高了凋落物C和养分向土壤中的归还量(Zhong
et al .,
2017 ), 从而促进了蔗糖酶、脲酶和磷酸酶的产生. ...
... 研究表明, 土壤酶活性与植物群落多样性亦存在不同程度的相关性(王长庭等,
2010 ; 乔文静等,
2018 ).降水量和N添加及其交互作用改变了植物群落多样性(附录II; 朱湾湾等,
2019 ), 因而可能会对土壤酶活性产生重要影响.本研究中, 蔗糖酶活性随着Patrick丰富度指数增加而增加, 即蔗糖酶活性与植物物种数存在正线性关系.这可能是由于降水量增加和N添加下, 随着植物物种数增多, 土壤酶作用的底物多样性增加, 从而增强了蔗糖酶活性, 与对江河源区人工草地的研究结果(王启兰等,
2010 )一致; 脲酶和磷酸酶活性均与Shannon-Wiener多样性指数以及Pielou均匀度指数存在负线性关系.这可能是由于降水量和N添加下, 植物群落多样性和均匀度的改变直接影响到土壤微环境(李长斌等,
2016 ), 并通过对植物根系和微生物活性的影响, 间接影响到脲酶和磷酸酶活性.由于植物群落特征对土壤酶活性的影响存在时间滞后效应(乔文静等,
2018 ), 因此还需通过长期的试验观测深入探讨土壤酶活性与植物群落多样性之间的联系. ...
降水量及N添加对宁夏荒漠草原土壤C:N:P生态化学计量特征和植被群落组成的影响
6
2019
... 降水量减少处理采用自制的减雨架进行全年遮雨.减雨架上端离地约1.8 m, 用5 cm宽的瓦面状高透光性聚氯乙烯板搭建分别相当于50%和30%小区面积的遮雨面.降水量增加处理采用人工喷灌装置实现.喷水时间为每年5-8月的每月月初和月中.选用分析纯NH
4 NO
3 (34% N)进行N添加处理.施N时间为每年5-8月的每月月初.详细的试验处理方法见朱湾湾等(
2019 ). ...
... 同期, 在每个小区内随机选取1 m × 1 m的小样方进行植被群落调查.调查指标包括物种数和物种高度等.调查结束后, 齐平地面剪下地上部分, 按牛枝子、草木樨状黄耆、白草、针茅、糙隐子草及其他物种归类分装于纸袋中带回实验室烘干(65 ℃, 48 h)称质量.群落生物量为所有物种生物量之和.植物群落多样性采用Patrick丰富度指数、Shannon- Wiener多样性指数、Simpson优势度指数和Pielou均匀度指数等指标来表征, 具体计算方法见朱湾湾等(
2019 ).2018年植物生物量、群落多样性以及土壤理化性质引自朱湾湾等(
2019 ).2019年植物生物量、群落多样性以及土壤理化性质的结果见附录I-IV. ...
... ).2018年植物生物量、群落多样性以及土壤理化性质引自朱湾湾等(
2019 ).2019年植物生物量、群落多样性以及土壤理化性质的结果见附录I-IV. ...
... 本研究中, 降水量和N添加下脲酶和磷酸酶活性与草木樨状黄耆(豆科植物)生物量呈负线性关系, 而蔗糖酶、脲酶和磷酸酶活性均与糙隐子草(禾本科植物)生物量呈正线性关系, 与针对高寒草甸的研究结果(王长庭等,
2010 )存在差异.首先, 土壤酶活性与植物生物量之间的关系因植物种不同而异.这是由于不同植物根系的发达程度以及分泌物存在差异, 进而影响土壤酶活性(王小平等,
2018 ).豆科植物与根瘤菌的共生作用可使其在相对干旱的条件下快速生长(张仁懿等,
2014 ).这不仅消耗了土壤养分, 而且抑制了微生物活性以及酶活性.相比之下, 降水量增加和N添加显著地刺激了禾本科植物的生长(附录I; 朱湾湾等,
2019 ), 其自身的营养物质归还土壤后可为土壤酶的合成提供丰富的底物(乔文静等,
2018 ).其次, 降水量增加及N添加下植物生物量增加(附录I; 朱湾湾等,
2019 ), 提高了凋落物C和养分向土壤中的归还量(Zhong
et al .,
2017 ), 从而促进了蔗糖酶、脲酶和磷酸酶的产生. ...
... ).其次, 降水量增加及N添加下植物生物量增加(附录I; 朱湾湾等,
2019 ), 提高了凋落物C和养分向土壤中的归还量(Zhong
et al .,
2017 ), 从而促进了蔗糖酶、脲酶和磷酸酶的产生. ...
... 研究表明, 土壤酶活性与植物群落多样性亦存在不同程度的相关性(王长庭等,
2010 ; 乔文静等,
2018 ).降水量和N添加及其交互作用改变了植物群落多样性(附录II; 朱湾湾等,
2019 ), 因而可能会对土壤酶活性产生重要影响.本研究中, 蔗糖酶活性随着Patrick丰富度指数增加而增加, 即蔗糖酶活性与植物物种数存在正线性关系.这可能是由于降水量增加和N添加下, 随着植物物种数增多, 土壤酶作用的底物多样性增加, 从而增强了蔗糖酶活性, 与对江河源区人工草地的研究结果(王启兰等,
2010 )一致; 脲酶和磷酸酶活性均与Shannon-Wiener多样性指数以及Pielou均匀度指数存在负线性关系.这可能是由于降水量和N添加下, 植物群落多样性和均匀度的改变直接影响到土壤微环境(李长斌等,
2016 ), 并通过对植物根系和微生物活性的影响, 间接影响到脲酶和磷酸酶活性.由于植物群落特征对土壤酶活性的影响存在时间滞后效应(乔文静等,
2018 ), 因此还需通过长期的试验观测深入探讨土壤酶活性与植物群落多样性之间的联系. ...
水分条件变化对土壤微生物的影响及其响应机制研究进展
1
2019
... 本研究中, 减少降水量对蔗糖酶、脲酶和磷酸酶活性的影响较小, 增加降水量不同程度地影响3种酶的活性.这可能是由于研究区长期处于低水平的土壤水分环境下, 植物和微生物对干旱具有高的适应性(胡小文等,
2004 ; 朱义族等,
2019 ), 因而减少降水量对其生长繁殖以及酶的分泌影响较小.相比之下, 在缺水环境下, 增加降水量提高了土壤含水量(附录III), 从而加速凋落物淋溶、促进表层凋落物和腐殖质中可溶性物质进入土壤(豆鹏鹏等,
2018 ), 使微生物可利用底物增加, 微生物活性相应增强, 进而刺激微生物向环境中释放酶(Schimel
et al .,
2007 ), 即微生物分泌的酶随之增加.此外, 本研究发现增加降水量对土壤酶活性的影响与N添加存在交互作用, 与其他研究结果(王杰等,
2014a )相似.这表明水分有利于N肥效的释放, 二者的交互作用促进了植物生长(白春利等,
2013 )、凋落物积累以及微生物活动(王杰等,
2014b ), 进而刺激土壤酶的分泌、导致土壤酶活性增强(闫钟清等,
2017b ). ...
水分条件变化对土壤微生物的影响及其响应机制研究进展
1
2019
... 本研究中, 减少降水量对蔗糖酶、脲酶和磷酸酶活性的影响较小, 增加降水量不同程度地影响3种酶的活性.这可能是由于研究区长期处于低水平的土壤水分环境下, 植物和微生物对干旱具有高的适应性(胡小文等,
2004 ; 朱义族等,
2019 ), 因而减少降水量对其生长繁殖以及酶的分泌影响较小.相比之下, 在缺水环境下, 增加降水量提高了土壤含水量(附录III), 从而加速凋落物淋溶、促进表层凋落物和腐殖质中可溶性物质进入土壤(豆鹏鹏等,
2018 ), 使微生物可利用底物增加, 微生物活性相应增强, 进而刺激微生物向环境中释放酶(Schimel
et al .,
2007 ), 即微生物分泌的酶随之增加.此外, 本研究发现增加降水量对土壤酶活性的影响与N添加存在交互作用, 与其他研究结果(王杰等,
2014a )相似.这表明水分有利于N肥效的释放, 二者的交互作用促进了植物生长(白春利等,
2013 )、凋落物积累以及微生物活动(王杰等,
2014b ), 进而刺激土壤酶的分泌、导致土壤酶活性增强(闫钟清等,
2017b ). ...
 ,
, ,1,*
,1,*

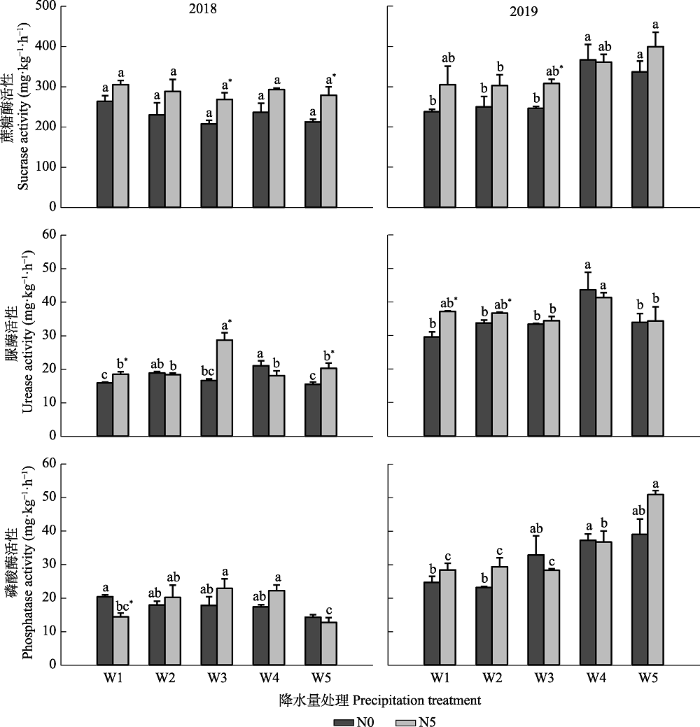 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT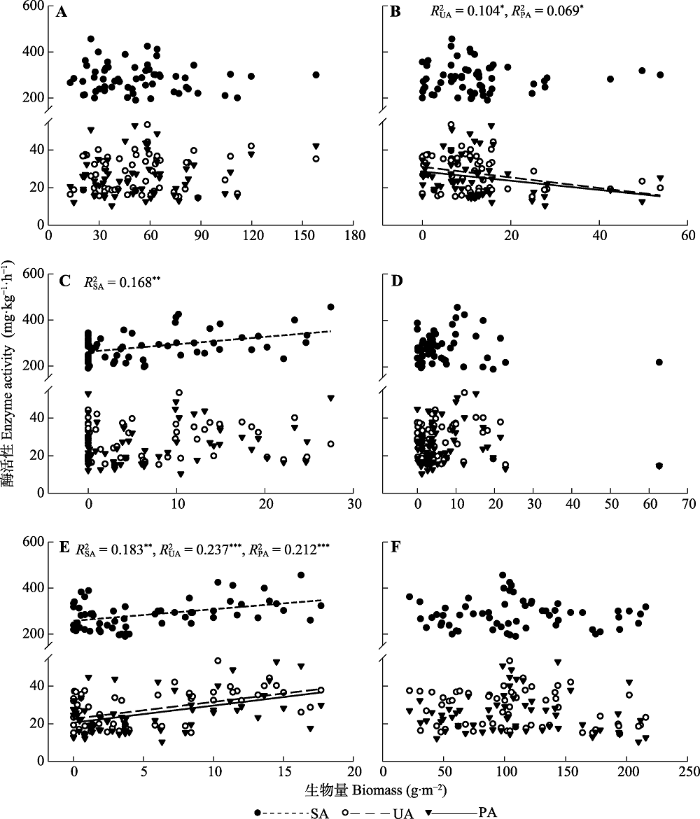 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT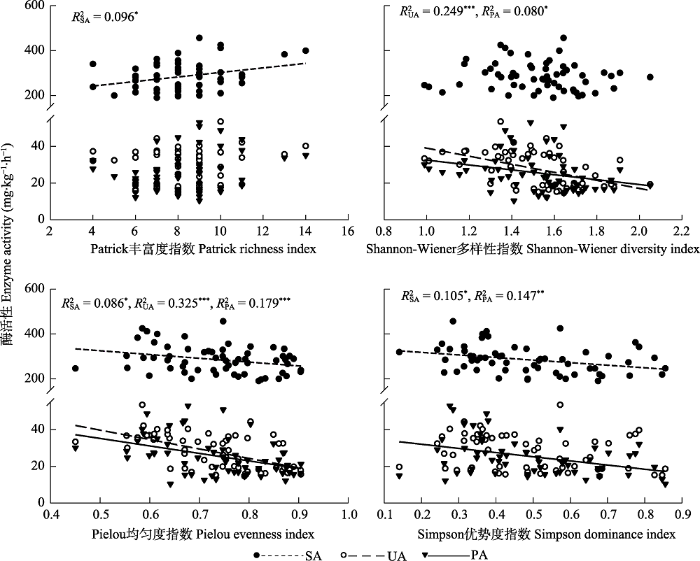 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT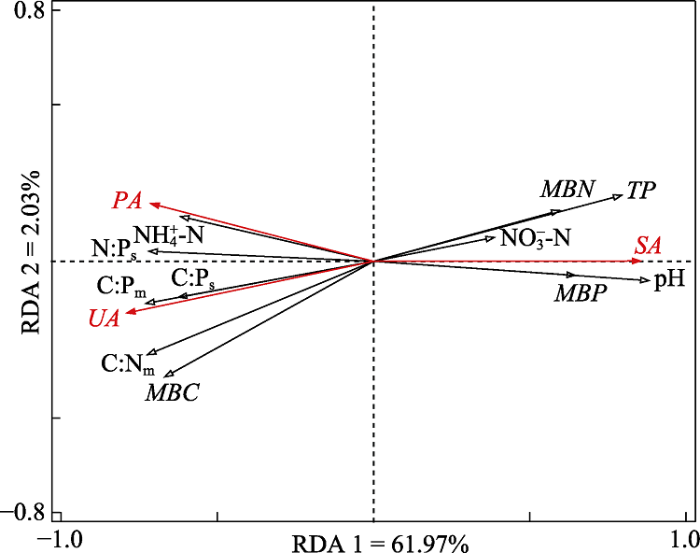 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}